Организмы дрожжей продуцируют ряд белков, которые функционируют вне клетки. Такие белки называют секретируемыми белками. Указанные секретируемые белки сначала экспрессируются внутри клетки в виде предшественника или преформы, содержащей последовательность препептида, обеспечивающего эффективное направление (транслокацию) экспрессированного продукта через мембрану эндоплазматического ретикулума (ЭР). Препептид, обычно называемый сигнальным пептидом, как правило, отщепляется от желаемого продукта в ходе транслокации. После входа в секреторный путь белок транспортируется к аппарату Гольджи. Из аппарата Гольджи белок может следовать разными путями, которые ведут в такие компартменты, как клеточная вакуоль или клеточная мембрана, или он может быть направлен из клетки для секреции во внешнюю среду (Pfeffer et al. (1987) Ann. Rev. Biochem. 56: 829-852).
Инсулин является полипептидным гормоном, секретируемым β-клетками поджелудочной железы, и состоит из двух полипептидных цепей А и В, которые связаны двумя межцепочечными дисульфидными мостиками. Кроме того, особенностью А-цепи является наличие одного внутрицепочечного дисульфидного мостика.
Гормон синтезируется в виде одноцепочечного предшественника проинсулина (препроинсулина), состоящего из препептида длиной 24 аминокислоты, за которым следует проинсулин, содержащий 86 аминокислот в следующем порядке: препептид - В-Arg Arg - С - Lys Arg - А, в котором С означает соединительный пептид из 31 аминокислоты. Arg-Arg и Lys-Агд представляют собой расщепляемые сайты для отщепления соединительного пептида от А- и В-цепей.
Для получения инсулина человека в микроорганизмах использовали три основных способа. В двух способах применяли Escherichia coli либо в случае экспрессии крупного слитого белка в цитоплазме (Frank et al. (1981) in Peptides: Proceedings of the 7th American Peptide Chemistry Symposium (Rich and Gross, eds.), Pierce Chemical Co., Rockford, IL. p.729-739), либо в случае использования сигнального пептида, чтобы обеспечить возможность секреции в периплазматическое пространство (Chan et al. (1981) PNAS 78: 5401-5404). В третьем способе использовали Saccharomyces cerevisiae, чтобы секретировать предшественник инсулина в среду (Thim et al. (1986) PNAS 83: 6766-6770). В предшествующем уровне техники сообщается об ограниченном количестве предшественников инсулина, которые экспрессируются либо в Е. coli, либо в Saccharomyces cerevisiae, смотри патент США 5962267, заявки WO 95/16708, ЕР 0055945, ЕР 0163529, ЕР 0347845 и ЕР 0741188.
Круговой дихроизм (КД) используют для того, чтобы определить стабильность белка и относительные стабильности молекул. КД, наблюдаемый ниже 240 нм, обусловлен амидным хромофором пептида, и его можно использовать для оценки вторичной структуры белка (Johnson (1988) Ann. Rev. Biophys. Chem. 17: 145-166). Спектр инсулина характеризуется минимумами при 220 и 209 нм, отрицательно-положительным переходом вблизи 203 нм и максимумом при 195 нм. При денатурации отрицательная полоса спектра КД в пределах 240-218 нм постепенно уменьшается, что согласуется с потерей упорядоченной вторичной структуры, которая сопутствует разворачиванию белка. Поэтому стабильность укладки предшественника инсулина можно количественно проанализировать, измеряя потерю вторичной структуры как функцию добавленного денатурирующего агента, например гидрохлорида гуанидина (GuHCl) (смотри, например, Расе (1975) CRC Crit. Rev. Biochem. 3: 1-43).
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Особенностью данного изобретения являются новые соединительные пептиды (С-пептиды), которые дают повышенный выход продукции и/или повышенную стабильность молекул предшественников инсулина и молекул аналогов предшественников инсулина при экспрессии в трансформированном микроорганизме, в частности, в дрожжах. Такие предшественники инсулина или аналоги предшественников инсулина затем можно превратить в инсулин или аналоги инсулина в ходе одной или нескольких подходящих, хорошо известных стадий превращения.
Соединительные пептиды согласно данному изобретению содержат, по меньшей мере, один остаток ароматической аминокислоты Phe, Trp или Tyr и, как правило, будут короче, чем природный С-пептид человека, который, включая фланкирующие сайты расщепления из двух основных аминокислот, состоят из 35 аминокислот. Таким образом, новые соединительные пептиды, как правило, будут длиной не более 15 аминокислотных остатков, и предпочтительно не более 9 аминокислотных остатков. Обычно новые соединительные пептиды будут длиной до 7 или до 5 аминокислотных остатков и предпочтительно не более 4 аминокислотных остатков в длину.
Как и в природной молекуле инсулина человека соединительный пептид будет содержать сайт расщепления на его С- и N-конце, обеспечивающий отщепление in vitro соединительного пептида от А- и В-цепей. Такими сайтами расщепления могут быть любые подходящие сайты расщепления, известные в данной области, например Met, отщепляемый бромцианом; отдельный остаток основной аминокислоты или пара остатков основных аминокислот (Lys или Arg), отщепляемые трипсином или трипсиноподобными протеазами; протеазой Acromobactor lyticus или протеазой карбоксипептидазой. Сайт расщепления, обеспечивающий отщепление соединительного пептида от А-цепи, предпочтительно представляет собой единственный остаток основной аминокислоты Lys или Arg, предпочтительно Lys.
Альтернативно отщепление соединительного пептида от В-цепи можно обеспечить посредством расщепления у природного аминокислотного остатка Lys в В-цепи, приводящего к образованию предшественника инсулина desВ30 или аналога предшественника инсулина desВ30. Требуемый аминокислотный остаток В30 затем можно добавить хорошо известными ферментативными способами in vitro.
В одном варианте соединительный пептид не будет содержать двух близлежащих остатков основных аминокислот (Lys, Arg). В указанном варианте отщепление от А-цепи можно осуществить у единственного Lys или Arg, локализованного на N-конце А-цепи, и природного Lys в положении В29 в В-цепи.
Соединительный пептид может содержать более одного остатка ароматической аминокислоты, но предпочтительно не более 5. Остатки ароматических аминокислот могут быть одинаковыми или отличаться друг от друга. Соединительный пептид предпочтительно не будет содержать более 3 остатков ароматических аминокислот и наиболее предпочтительно он будет содержать только единственный остаток ароматической аминокислоты.
В одном варианте данного изобретения один из остатков ароматических аминокислот в соединительном пептиде находится непосредственно у N-конца сайта расщепления, примыкающего к А-цепи. Кроме того, один из остатков ароматических аминокислот предпочтительно будет расположен на расстоянии менее 5 Å, по меньшей мере, от одного из остатков в положениях B11, B12 или В26 в В-цепи. В одном варианте ароматическая аминокислота, расположенная непосредственно у N-конца сайта расщепления, примыкающего к А-цепи, находится на расстоянии менее 5 Å, по меньшей мере, от одного из остатков в положениях В11, В12 или В26 в В-цепи.
Предшественники инсулина или аналоги предшественников инсулина характеризуются наличием высокой стабильности укладки в растворе. Предшественники согласно данному изобретению будут обладать более высокой стабильностью с более высоким значением Cmid по сравнению с инсулином или аналогами инсулина, которые не содержат остатка ароматической аминокислоты в соединительном пептиде. Стабильность, таким образом, будет выше с Cmid, выше чем примерно 5,5 М GuHCl, обычно выше чем около 6,0 М GuHCl и более типично выше чем около 6,5 М GuHCl.
Соответственно, в одном аспекте данное изобретение относится к предшественникам инсулина или аналогам предшественников инсулина, содержащим соединительный пептид (С-пептид), отщепляемый от А- и В-цепей, при этом указанный соединительный пептид содержит, по меньшей мере, один остаток ароматической аминокислоты и сайт расщепления, обеспечивающий расщепление пептидной связи между А-цепью и соединительным пептидом, где один остаток ароматической аминокислоты расположен непосредственно у N-конца указанного сайта расщепления.
В другом аспекте данное изобретение относится к предшественникам инсулина или аналогам предшественников инсулина, содержащим соединительный пептид (С-пептид), отщепляемый от А- и В-цепей, при этом указанный соединительный пептид содержит до 9 аминокислотных остатков, из которых, по меньшей мере, один является остатком ароматической аминокислоты.
В еще одном аспекте данное изобретение относится к предшественнику инсулина или аналогу предшественника инсулина, содержащему соединительный пептид (С-пептид), отщепляемый от А- и В-цепей, при этом указанный соединительный пептид содержит один остаток ароматической аминокислоты, который расположен на расстоянии менее 5 Å, по меньшей мере, от одного из остатков в положениях B11, B12 или В26 в В-цепи.
В следующем аспекте данное изобретение относится к предшественникам инсулина или аналогам предшественников инсулина, содержащим соединительный пептид (С-пептид), включающий в себя, по меньшей мере, один остаток ароматической аминокислоты и отщепляемый от А- и В-цепей. Указанные предшественники инсулина или аналоги предшественников инсулина обладают повышенной стабильностью с более высоким Cmid по сравнению с предшественником инсулина или аналогами предшественника инсулина, которые не содержат остатка ароматической аминокислоты в соединительном пептиде.
Повышенную активность определяют разными способами, известными специалисту в данной области и описанными ниже. В одном варианте повышенную стабильность измеряют определением посредством КД концентрации гидрохлорида гуанидина (GuHCl), необходимой для достижения половины от максимального разворачивания молекулы предшественника инсулина (Cmid).
В следующем аспекте данное изобретение относится к предшественникам инсулина или аналогам предшественников инсулина, содержащим последовательность формулы:
В (1-27)-X2-X3-X1-Y-A (1-21),
где
X1 представляет собой пептидную последовательность из 1-15 аминокислотных остатков, содержащую один остаток ароматической аминокислоты непосредственно у N-конца Y,
Х2 представляет собой одну из аминокислот Pro, Asp, Lys или Ile в положении 28 В-цепи,
Х3 представляет собой одну из аминокислот Pro, Lys, Ala, Arg или Pro-Thr в положении 29 В-цепи, и
Y является Lys или Arg.
В одном варианте общее количество аминокислотных остатков в X1 будет составлять 1-10, 1-9, 1-8, 1-7, 1-6, 1-5 или 1-4 аминокислотных остатка в длину. В другом конкретном варианте X1 означает 1-3 аминокислотных остатка и предпочтительно 1-2 аминокислотных остатка. Аминокислотным остатком в X1 может быть любой кодируемый аминокислотный остаток, и они могут быть одинаковыми или разными, только при условии, что один является остатком ароматической аминокислоты, расположенным непосредственно у N-конца Y.
В следующем аспекте данное изобретение относится к предшественникам инсулина или аналогам предшественников инсулина, содержащим последовательность формулы:
B(1-27)-X2-X3-X1-Y-A(1-21),
где
X1 представляет собой пептидную последовательность из 1-15 аминокислотных остатков, один из которых является остатком ароматической аминокислоты, который расположен на расстоянии менее 5 Å, по меньшей мере, от одного из аминокислотных остатков в положении В11, В12 или В26 в В-цепи,
Х2 представляет собой одну из аминокислот Pro, Asp, Lys или Ile в положении 28 В-цепи,
Х3 представляет собой одну из аминокислот Pro, Lys, Ala, Arg или Pro-Thr в положении 29 В-цепи, и Y является Lys или Arg.
В одном варианте количество аминокислотных остатков в Xi составляет 1-9, 1-5 или 1-4. В другом варианте количество аминокислотных остатков составляет 1-3 или 1-2.
В другом аспекте данное изобретение относится к предшественникам инсулина или аналогам предшественников инсулина, содержащим последовательность формулы:
B(1-27)-X2-X3-X1-Y-A(1-21),
где
X1 представляет собой пептидную последовательность из 1-8 аминокислотных остатков, по меньшей мере, один из которых является остатком ароматической аминокислоты,
Х2 представляет собой одну из аминокислот Pro, Asp, Lys или Ile в положении 28 В-цепи,
Х3 представляет собой одну из аминокислот Pro, Lys, Ala, Arg или Pro-Thr в положении 29 В-цепи, и
Y является Lys или Arg.
Общее количество аминокислотных остатков в X1 будет составлять 1-7, 1-6, 1-5 или 1-4 аминокислотных остатка. В более конкретном варианте X1 означает 1-3 аминокислотных остатка и предпочтительно 1-2 аминокислотных остатка. Аминокислотным остатком в X1 может быть любой кодируемый аминокислотный остаток, и они могут быть одинаковыми или разными, только при условии, что, по меньшей мере, один аминокислотный остаток в X1 является остатком ароматической аминокислоты.
В указанной выше формуле X1 может содержать до 5 остатков ароматических аминокислот, которые могут быть одинаковыми или разными. В конкретном варианте X1 содержит до 3 остатков ароматических аминокислот, которые могут быть одинаковыми или разными, X1 предпочтительно будет содержать только один остаток ароматической аминокислоты. Остатки ароматических аминокислот представляют собой Trp, Phe или Tyr, предпочтительно Phe или Trp.
В одном варианте Х2 означает Asp, а Х3 означает Lys. Указанный вариант охватывает аналоги предшественников инсулина, содержащие Asp в положении В28 В-цепи (называемые в дальнейшем «AspB28IP»). В другом варианте X2 означает Lys, а Х3 означает Pro. В следующем варианте последовательность X1-Y выбрана из группы: (a) Met-Trp-Lys; (b) Ala-Trp-Lys; (с) Val-Trp-Lys; (d) Ile-Trp-Lys; (e) Leu-Trp-Lys; (f) Glu-Glu-Phe-Lys (SEQ ID NO:15); (g) Glu-Phe-Lys; (h) Glu-Trp-Lys; (i) Ser-Trp-Lys; (j) Thr-Trp-Lys; (k) Arg-Trp-Lys; (1) Glu-Met-Trp-Lys (SEQ ID NO:1); (m) GIn-Met-Trp-Lys (SEQ ID NO:2) и (n) Asp-Trp-Lys.
В другом варианте Х2 означает Pro, а Х3 означает Lys, и X1 означает 1-2 аминокислотных остатка, один из которых является Trp или Phe.
В другом варианте X2 означает Lys, Х3 означает Pro-Thr, и X1 имеет в своем составе до 15 аминокислотных остатков, один из которых является Thr, Туг или Phe. В указанном варианте X1 будет содержать сайт расщепления на С-конце, например, сайт расщепления из одной или двух основных аминокислот (Lys, Arg).
Данное изобретение также относится к полинуклеотидным последовательностям, которые кодируют заявленные предшественники инсулина или аналоги предшественников инсулина. В следующем аспекте данное изобретение относится к векторам, содержащим такие полинуклеотидные последовательности, и к клеткам-хозяевам, содержащим такие полинуклеотидные последовательности или векторы.
В другом аспекте изобретение относится к способу получения предшественников инсулина или аналогов предшественников инсулина в клетке-хозяине, при этом указанный способ включает в себя (i) культивирование клетки-хозяина, содержащей полинуклеотидную последовательность, кодирующую предшественники инсулина или аналоги предшественников инсулина согласно изобретению, в условиях, подходящих для экспрессии указанного предшественника или аналога предшественника; и (ii) выделение предшественника или аналога предшественника из культуральной среды.
В еще одном аспекте изобретение относится к способу получения инсулина или аналогов инсулина в клетке-хозяине, при этом указанный способ включает в себя (i) культивирование клетки-хозяина, содержащей полинуклеотидную последовательность, кодирующую предшественник инсулина или аналоги предшественника инсулина согласно изобретению; (ii) выделение предшественника или аналога предшественника из культуральной среды и (iii) превращение предшественника или аналога предшественника в инсулин или аналог инсулина путем ферментативного превращения in vitro.
В одном варианте данного изобретения клетка-хозяин представляет собой дрожжевую клетку-хозяина, а в следующем варианте дрожжевая клетка-хозяин выбирается из клеток рода Saccharomyces. В следующем варианте дрожжевая клетка-хозяин выбирается из клеток вида Saccharomyces cerevisiae.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
На фиг.1 представлена экспрессирующая плазмида рАК721 S. cerevisiae, которая экспрессирует слитый белок лидер LA19-ЕЕАЕАЕАЕРК (SEQ ID NO:3)-IP(AlaAlaLys).
На фиг.2 представлена последовательность ДНК и рассчитанная аминокислотная последовательность кодируемого слитого белка (лидер α-фактора-ЕЕАЕАЕАРК (SEQ ID NO:4)-AspB28IP части плазмиды рАК1150, используемой в качестве матрицы в ПЦР (SEQ ID NO:5 и 6).
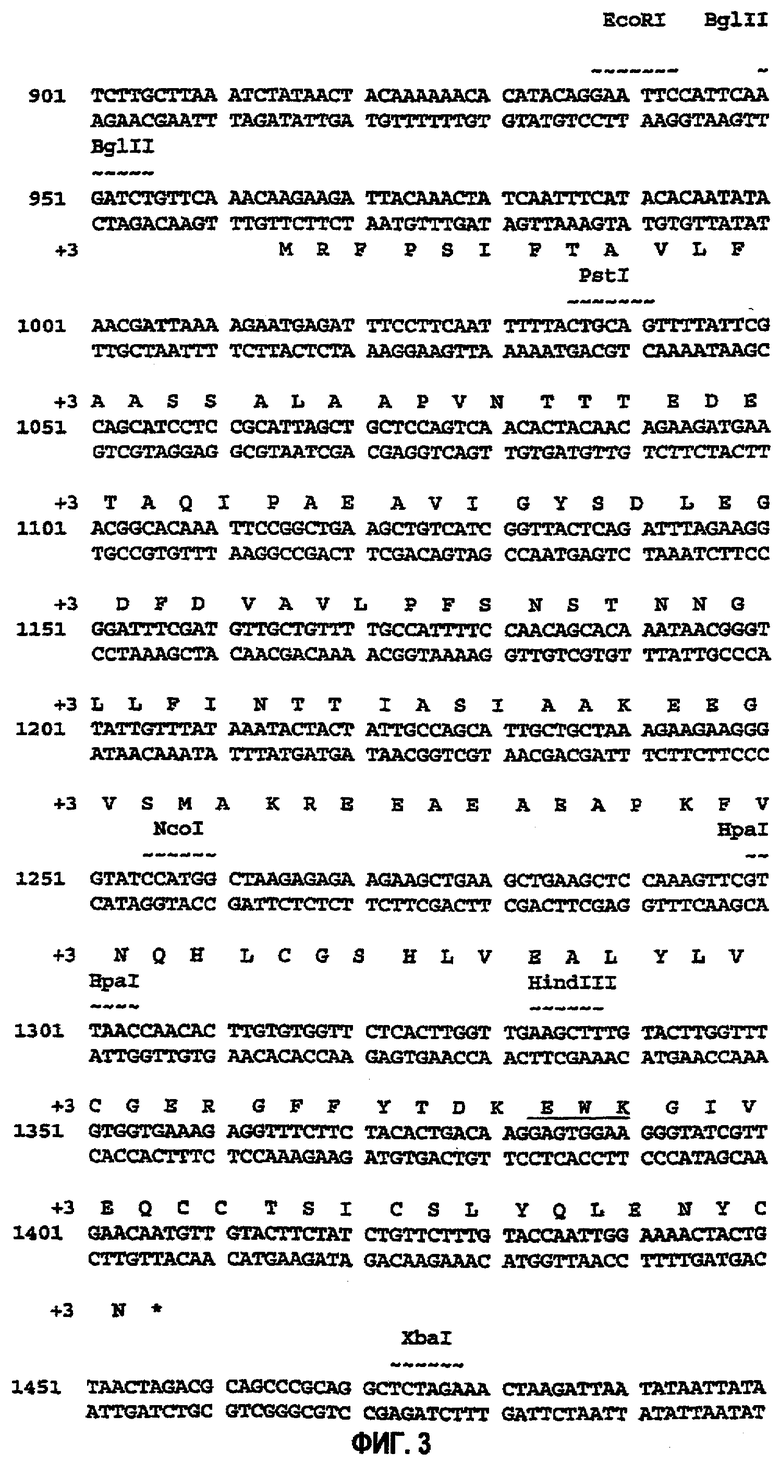
На фиг.3 представлена последовательность ДНК, кодирующая слитый белок лидер α-фактора-AspB28IP(GluTrpLys) с синтетическим мини-С-пептидом GluTrpLys, полученным путем рандомизированной оптимизации (SEQ ID NO:7 и 8). Мини-С-пептид (EWK) указан подчеркиванием.
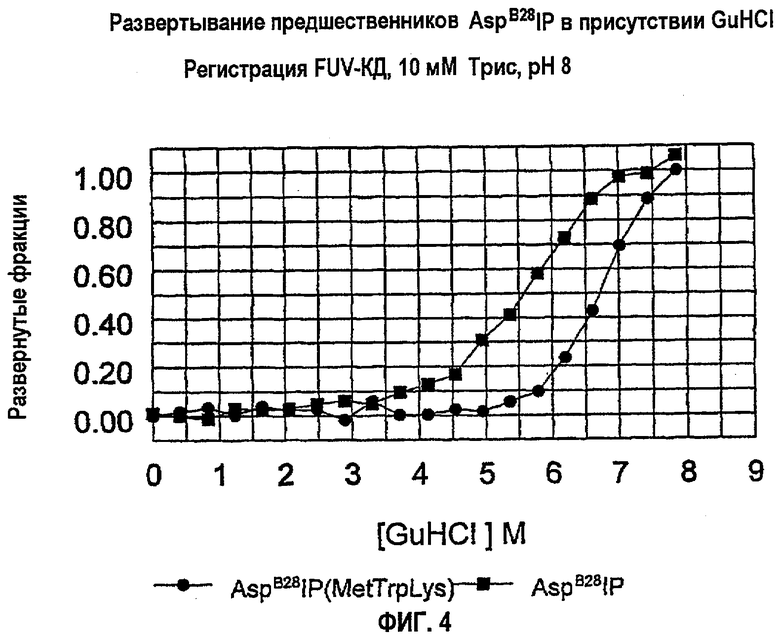
На фиг.4 показана стабильность укладки аналога инсулина AspB28IP(MetTrpLys) по сравнению с AspB28IP.
На фиг.5 показаны структуры AspB28IP(MetTrpLys) в растворе в виде линий основных цепей набора из 20 конвергентных структур.
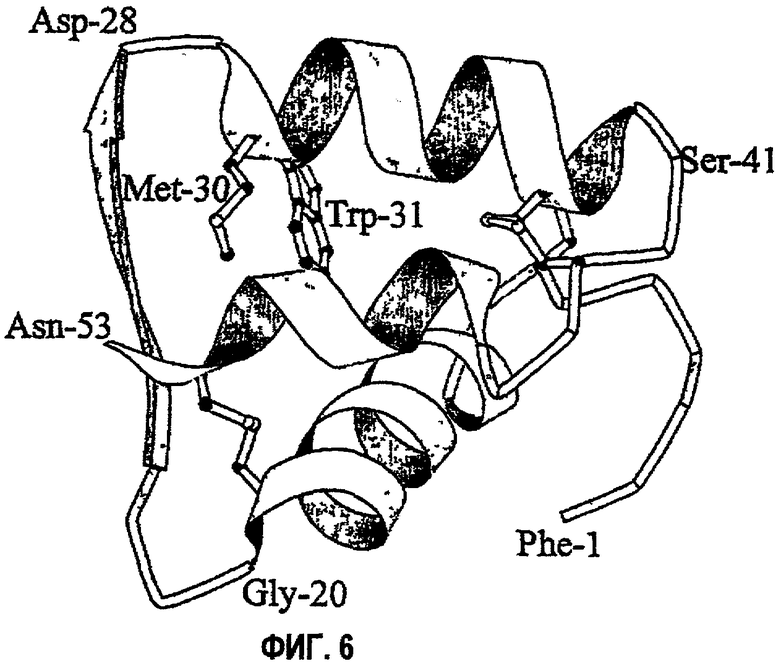
На фиг.6 показано представление AspB28IP(MetTrpLys) в виде ленты. Фигура получена с использованием MOLSCRIPT (Kraulis (1991) J. Appl. Crystallog. 24: 946-950). Аннотация к остаткам аминокислот дана следующим образом: В1-В29 (В-цепь) пронумерованы 1-29, остатки С1-С3 (соединительный пептид) пронумерованы 30-32 и остатки А1-А21 (А-цепь) пронумерованы 33-53.
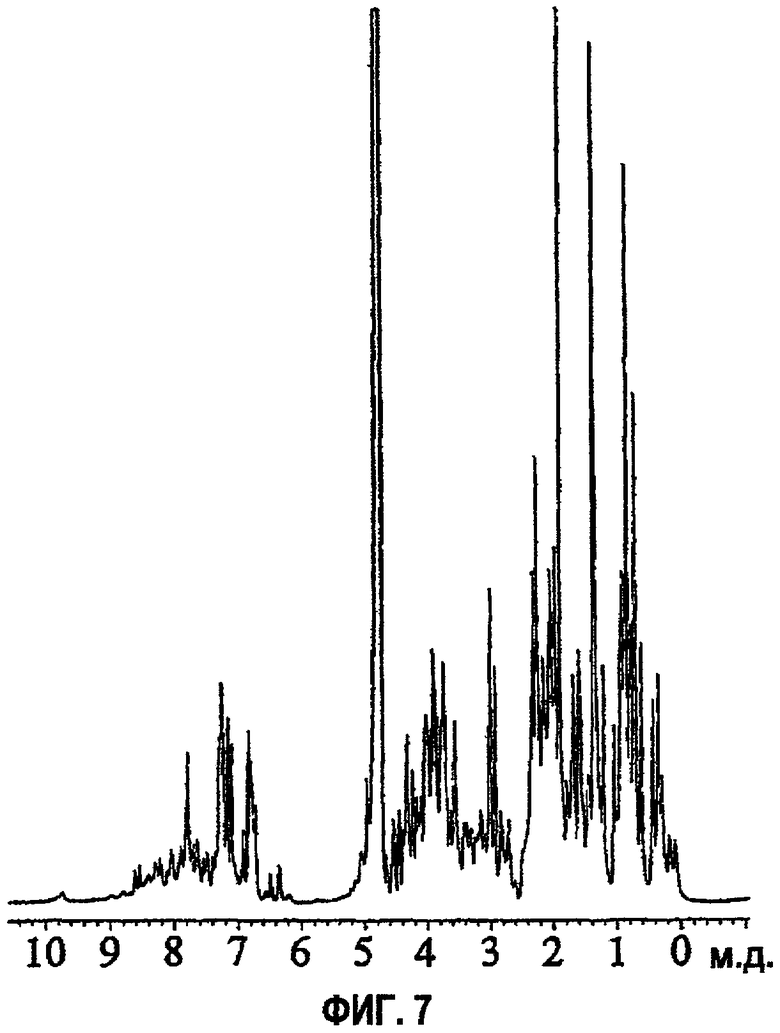
Фиг.7 представляет собой идентификационный спектр ЯМР протонов для AspB28IP(MetTrpLys), записанный при 27°С при концентрации 1,0 мМ в 10%/90% D2O/H2O с 10 мМ фосфатным буфером при рН 8,0.
На фиг.8 представлена ДНК и рассчитанная последовательность аминокислот экспрессирующей кассеты, которая экспрессирует слитый белок YAP3-TA39-EEGEPK (SEQ ID NO:11)-AspB28IP с синтетическим мини-С-пептидом GluTrpLys (SEQ ID NO:9 и 10) и
На фиг.9 представлена ДНК и рассчитанная аминокислотная последовательность экспрессирующей кассеты, которая экспрессирует слитый белок YAP3-TA57-EEGEPK (SEQ ID NO:17)-AspB28IP с синтетическим мини-С-пептидом GluTrpLys (SEQ ID NO:11 и 12).
На фиг.10 представлена кривая расщепления предшественника инсулина Aspart - протеазой Achromobacter Cyticus.
ПОДРОБНОЕ ОПИСАНИЕ
Сокращения и номенклатура
Под «соединительным пептидом» или «С-пептидом» подразумевается соединительный фрагмент «С» полипептидной последовательности В-С-А одноцепочечной молекулы, подобной препроинсулину. В частности, в природной цепи инсулина С-пептид связывает положение 30 В-цепи и положение 1 А-цепи. «Мини-С-пептид» или «соединительный пептид», такой как описанные здесь пептиды, связывает В29 или В30 с А1 и отличается по последовательности и длине от природного С-пептида.
Под «IP» подразумевается одноцепочечный предшественник инсулина, в котором dеsВ30-цепь связана с А-цепью инсулина посредством соединительного пептида. Одноцепочечный предшественник инсулина будет содержать правильно расположенные дисульфидные мостики (три), как в инсулине человека.
Термины «dеsВ30» или «В(1-29)» означают В-цепь природного инсулина, в которой отсутствует остаток аминокислоты В30, «А(1-21)» означает А-цепь природного инсулина, «В(1-27)» означает природную В-цепь, в которой отсутствуют аминокислотные остатки В28, В29 и В30; «AspB28IP» означает одноцепочечный предшественник инсулина с аспарагиновой кислотой в положении 28 В-цепи и без С-пептида) (В29 связан с A1). Мини-С-пептид и его аминокислотная последовательность указаны трехбуквенным кодом аминокислот в круглых скобках после IP; таким образом, «AspB28IP(MetTrpLys)» означает одноцепочечный предшественник инсулина с аспарагиновой кислотой в положении 28 В-цепи и мини-С-пептидом с последовательностью Met-Trp-Lys, связывающим В29 с A1.
Термин «предшественник инсулина» означает одноцепочечный полипептид, который в результате одного или нескольких последовательных химических и/или ферментативных процессов можно превратить в инсулин человека.
Термин «аналог предшественника инсулина» означает молекулу предшественника инсулина, имеющую одну или несколько мутаций, замен, делеций и/или присоединений в А- и/или В-аминокислотных цепях, по сравнению с молекулой инсулина человека. Предпочтительны такие аналоги инсулина, в которых один или несколько аминокислотных остатков природного происхождения, предпочтительно один, два или три остатка, были заменены другим кодируемым аминокислотным остатком. В одном варианте данное изобретение включает в себя молекулы аналогов, в которых положение 28 В-цепи изменено по сравнению с природной молекулой инсулина человека. В указанном варианте положение 28 изменено с природного остатка Pro на один из остатков Asp, Lys или Ile. В предпочтительном варианте природный остаток Pro в положении В28 изменен на остаток Asp. В другом варианте Lys в положении В29 заменен на Pro; Asn в положении А21 также может быть изменен на Ala, Gin, Glu, Gly, His, Ile, Leu, Met, Ser, Thr, Trp, Tyr или Val, в частности на Gly, Ala, Ser или Thr и предпочтительно на Gly. Кроме того, Asn в положении В3 может быть изменен на Lys. Следующими примерами аналогов предшественников инсулина являются инсулин человека des(B30), аналоги инсулина, в которых был делегирован PheB1; аналоги инсулина, в которых А-цепь и/или В-цепь имеют N-концевое удлинение, и аналоги инсулина, в которых А-цепь и/или В-цепь имеют удлинение С-конца. Таким образом, к положению В1 могут быть добавлены один или два Arg.
Термин «непосредственно у N-конца» предназначен для иллюстрации ситуации, когда аминокислотный остаток или пептидная последовательность непосредственно связана своим С-концом с N-концом другого аминокислотного остатка или аминокислотной последовательности посредством пептидной связи.
В данном контексте термин «функциональный аналог инсулина» и тому подобное, предназначен для обозначения полипептида со сходным биологическим действием, как и у нативного белка инсулина человека.
Выражение расстояние «короче 5 Å» между двумя аминокислотными остатками означает самое короткое межатомное расстояние менее 5 Å между любым атомом в первой аминокислоте и любым атомом во второй аминокислоте. Атомные расстояния измеряют из трехмерной структуры молекулы, определенной либо посредством ЯМР (Wüthrich, К., 1986, NMR of Proteins and Nucleic Acids, Wiley, New York), либо способом рентгеновской кристаллографии (Drenth, J., 1994, Principles of Protein X-ray crystallography. Springer Verlag Berlin). Расстояние от одной аминокислоты до другой измеряют в виде самого короткого межатомного расстояния между любым атомом в первой аминокислоте и любым атомом во второй аминокислоте, если не оговорено особо.
Особенностью данного изобретения являются новые мини-С-пептиды, связывающие положение 29 В-цепи инсулина и положение 1 А-цепи инсулина, которые существенно увеличивали выход продукции в дрожжевой клетке-хозяине. Под термином «существенно увеличенная продукция», «увеличенный выход ферментации» и тому подобным понимают увеличение секретируемого количества молекулы предшественника инсулина или молекулы аналога предшественника инсулина в надосадке культуры по сравнению с выходом предшественника инсулина или аналога предшественника инсулина, не содержащего остатка ароматической аминокислоты в мини-С-пептиде. «Повышенный» выход ферментации представляет собой абсолютное количество, большее чем в контроле; предпочтительно увеличение представляет собой повышение на 50% или больше, чем уровень (AspB28 IP) в контроле; еще более предпочтительно увеличение представляет собой превышение на 100% или более по сравнению с контрольными уровнями.
Под термином «повышение стабильности» понимают, например, увеличенное значение Cmid в растворе по сравнению со значением, полученным для предшественника аналога инсулина (например. AspB28IP), не содержащего остатка ароматической аминокислоты в мини-С-пептиде. Под термином «Cmid» понимают концентрацию GuHCl, необходимую для развертывания половины популяции белков при анализе, в котором измеряется круговой дихроизм молекулы инсулина в дальнем УФ как функция возрастающих концентраций денатурирующего агента.
«РОТ» означает ген триозофосфатизомеразы Schizosaccharomyces pombe, а «TPI1» означает ген триозофосфатизомеразы S. cerevisiae.
Под «лидером» понимают аминокислотную последовательность, состоящую из препептида (сигнального пептида) и пропептида.
Подразумевается, что термин «сигнальный пептид» означает препептид, который присутствует в виде N-концевой последовательности в форме предшественника белка. Функция сигнального пептида состоит в том, чтобы обеспечить возможность для облегчения транслокации гетерологичного белка в эндоплазматический ретикулум. Сигнальный пептид обычно отщепляется в ходе этого процесса. Сигнальный пептид может быть гетерологичным или гомологичным по отношению к организму дрожжей, продуцирующему белок. Ряд сигнальных пептидов, которые можно использовать с конструкцией ДНК согласно изобретению, включает сигнальный пептид аспарагиновой протеазы 3 дрожжей (YАР3) или любой функциональный аналог (Egel-Mitani et al. (1990) YEAST 6: 127-137 и патент США 5726038) и сигнал α-фактора гена MFα1 (Thorner (1981) in The Molecular Biology of the Yeast Saccharomyces cerevisiae, Strathern et al., eds., p.143-180, Cold Spring Harbor Laboratory, NY и патент США 487000.
Термин «пропептид» означает полипептидную последовательность, функция которой состоит в обеспечении того, чтобы экспрессированный полипептид был направлен из эндоплазматического ретикулума в аппарат Гольджи и далее в секретируемую везикулу для секреции в культуральную среду (т.е. экспорта полипептида через клеточную стенку или, по меньшей мере, через клеточную мембрану в периплазматическое пространство дрожжевой клетки). Пропептидом может быть пропептид α-фактора дрожжей, смотри патенты США 4546082 и 4870008. Альтернативно, пропептидом может быть синтетический пропептид, что говорит о том, что пропептид не встречается в природе. Подходящими синтетическими пропептидами являются пропептиды, заявленные в патентах США 5395922; 5795746; 5162498 и заявке WO 98/32867. Пропептид предпочтительно будет содержать сайт процессинга эндопептидазой на С-конце, такой как последовательность Lys-Arg или любой его функциональный аналог.
Полинуклеотидную последовательность согласно изобретению можно получить синтетически стандартными принятыми способами, например, фосфоамидитным способом, описанным Beaucage et al. (1981) Tetrahedron Letters 22: 1859-1869, или способом, описанным Matthes et al. (1984) EMBO Journal 3: 801-805.
Согласно фосфоамидитному способу олигонуклеотиды синтезируют, например, в автоматическом синтезаторе ДНК, очищают, образуют дуплексы и лигируют, чтобы образовать синтетическую конструкцию ДНК. Предпочтительным в настоящее время способом получения ДНК-конструкции является полимеразная цепная реакция (ПЦР).
Полинуклеотидная последовательность согласно изобретению также может происходить из смешанной геномной, кДНК или синтетической ДНК. Например, последовательность геномной или кДНК, кодирующую лидерный пептид, можно соединить с последовательностью геномной ДНК или кДНК, кодирующей А- и В-цепи, после чего последовательность ДНК можно модифицировать в сайте встраиванием синтетических олигонуклеотидов, кодирующих желаемую аминокислотную последовательность для гомологичной рекомбинации, в соответствии с хорошо известными способами, или предпочтительно созданием желаемой последовательности с помощью ПЦР, используя подходящие олигонуклеотиды.
Изобретение включает в себя вектор, который способен к репликации в выбранном микроорганизме или клетке-хозяине и который несет полинуклеотидную последовательность, кодирующую предшественники инсулина или аналоги предшественников инсулина согласно изобретению. Рекомбинантный вектор может быть автономно реплицируюшимся вектором, т.е. вектором, который существует как внехромосомная единица, репликация которой не зависит от репликации хромосом, например, плазмидой, в не хромосомным элементом, минихромосомой или искусственной хромосомой. Вектор может содержать любые средства для обеспечения автономной репликации. В альтернативном случае вектором может быть вектор, который при введении в клетку-хозяина интегрируется в геном и реплицируется вместе с хромосомой(ами), в которую он был интегрирован. Кроме того, можно использовать отдельный вектор или плазмиду, или два или несколько векторов или плазмид, которые вместе содержат полную ДНК, которую необходимо ввести в геном клетки-хозяина, или транспозон. Векторы могут быть линейными или замкнутыми кольцевыми плазмидами, и предпочтительно будут содержать элемент(ты), который делает возможной стабильную интеграцию вектора в геном клетки-хозяина или автономную репликацию вектора в клетке независимо от генома.
В предпочтительном варианте рекомбинантный экспрессирующий вектор способен реплицироваться в организме дрожжей. Примерами последовательностей, которые дают возможность вектору реплицироваться в дрожжах, являются гены репликации 2 мкм-плазмиды дрожжей REP 1-3 и начало репликации.
Векторы согласно данному изобретению предпочтительно содержат один или несколько селектируемых маркеров, которые обеспечивают простую селекцию трансформированных клеток. Селектируемый маркер представляет собой ген, продукт которого обеспечивает биоцидную или вирусную резистентность, резистентность к тяжелым металлам, прототрофность ауксотрофам и тому подобное. Примерами бактериальных селектируемых маркеров являются гены dal Bacillus subtilis или Bacillus licheniformis, или маркеры, которые придают устойчивость к антибиотикам, такую как устойчивость к ампициллину, канамицину, хлорамфениколу или тетрациклину. Селектируемые маркеры для применения в клетке-хозяине нитчатых грибов включают amdS (ацетамидаза), argB (орнитинкарбамоилтрансфераза), pyrG (оротидин-5'-фосфатдекарбоксилаза) и trpC (антранилатсинтаза). Подходящими маркерами для дрожжевых клеток-хозяев являются ADE2, HIS3, LEU2, LYS2, МЕТ3, TRP1 и URA3. Предпочтительным селектируемым маркером для дрожжей является ген TPI Schizosaccharomyces pombe (Russell (1985) Gene 40: 125-130).
В векторе полинуклеотидная последовательность оперативно связана с подходящей последовательностью промотора. Промотор может быть любой последовательностью нуклеиновой кислоты, которая проявляет транскрипционную активность в клетке-хозяине, выбранной из мутантных, укороченных или гибридных промоторов, и может быть получен из генов, кодирующих внеклеточные или внутриклеточные полипептиды либо гомологичные, либо гетерологичные по отношению к клетке-хозяину.
Примерами подходящих промоторов для управления транскрипцией в бактериальной клетке-хозяине являются промоторы, полученные из lac-оперона Е coli, гена агаразы Streptomyces coelicolor (dagA), гена левансахаразы Bacillus subtilis (sacB), гена альфа-амилазы Bacillus licheniformis (amyL), гена мальтогенной амилазы Bacillus stearothermophilus (amyM), гена альфа-амилазы Bacillus amyloliquefaciens (amyQ) и гена пенициллиназы Bacillus licheniformis (penP). Примерами подходящих промоторов для управления транскрипцией в клетке-хозяине нитчатых грибов являются промоторы, полученные из генов амилазы Aspergillus oryzae ТАКА, аспарагиновой протеиназы Rhizomucor miehei, нейтральной альфа-амилазы Aspergillus niger и устойчивой к кислотам альфа-амилазы Aspergillus niger. В дрожжевой клетке-хозяине пригодными промоторами являются промоторы Mal, TPI, ADH или PGK Saccharomyces cerevisiae.
Полинуклеотидная конструкция согласно изобретению также, как правило, будет оперативно связана с подходящим терминатором. У дрожжей подходящим терминатором является терминатор TPI (Alber et al. (1982) J. Mol. Appl. Genet. 1: 419-434).
Способы, используемые для того, чтобы лигировать соответственно полинуклеотидную последовательность согласно изобретению, промотор и терминатор, и чтобы встроить их в подходящие дрожжевые векторы, содержащие информацию, необходимую для репликации у дрожжей, хорошо известны специалистам в данной области. Будет понятно, что вектор можно сконструировать либо путем получения сначала конструкции ДНК, содержащей полную последовательность ДНК, кодирующую предшественники инсулина или аналоги предшественников инсулина согласно изобретению, и затем встраиванием этого фрагмента в подходящий экспрессирующий вектор, или посредством последовательного встраивания фрагментов ДНК, содержащих генетическую информацию для отдельных элементов (таких как сигнал, пропептид, мини-С-пептид, А- и В-цепи) с последующим лигированием.
Данное изобретение также относится к рекомбинантным клеткам-хозяевам, содержащим полинуклеотидную последовательность, кодирующую предшественники инсулина или аналоги предшественников инсулина согласно изобретению. Вектор, содержащий такую полинуклеотидную последовательность, вводят в клетку-хозяина, так что вектор сохраняется в виде хромосомного компонента или в виде автономно реплицирующегося внехромосомного вектора, как описано ранее. Термин «клетка-хозяин» охватывает любое потомство родительской клетки, которое не идентично родительской клетке вследствие мутаций, которые происходят во время репликации. Выбор клетки-хозяина будет в большой степени зависеть от гена, кодирующего полипептид, и его источника. Клеткой-хозяином может быть одноклеточный микроорганизм, например, прокариот, или не одноклеточный микроорганизм, например, эукариот. Пригодными клетками одноклеточных являются бактериальные клетки, такие как грамположительные бактерии, включая, но не ограничивая указанным, клетку Bacillus, клетку Streptomyces, или грамотрицательные бактерии, такие как Е. coli и Pseudomonas sp. Эукариотические клетки могут быть клетками млекопитающих, насекомых, растений или грибов. В предпочтительном варианте клеткой-хозяином является дрожжевая клетка. Дрожжевым организмом, используемым в способе согласно изобретению, может быть любой подходящий организм дрожжей, который при культивировании продуцирует большие количества предшественника инсулина и аналогов предшественников инсулина согласно изобретению. Примерами подходящих организмов дрожжей являются штаммы, выбранные из видов дрожжей Saccharomyces cerevisiae, Saccharomyces kluyveri, Schizosaccharomyces pombe, Saccharomyces uvarum, Kluyveromyces lactis, Hansenula polymorpha, Pichia pastoris, Pichia methanolica, Pichia kluyveri, Yarrowia lipolytica, Candida sp., Candida utilis, Candida cacaoi, Geotrichum sp. и Geotrichum fermentans.
Трансформацию дрожжевых клеток можно, например, осуществить путем образования протопластов с последующей трансформацией по существу известным способом. Средой, используемой для культивирования клеток, может быть любая обычная среда, подходящая для роста организмов дрожжей. Секретированный предшественник инсулина или аналоги предшественника инсулина согласно изобретению, значительная часть которых будет присутствовать в среде в правильно процессированной форме, можно извлечь из среды традиционными способами, включая отделение дрожжевых клеток от среды центрифугированием, фильтрацией или улавливанием предшественника инсулина или аналога предшественника инсулина с помощью ионообменного матрикса или адсорбирующего матрикса обращенной фазы, осаждением белковых компонентов надосадка или фильтрата с помощью соли, например сульфата аммония, с последующей очисткой разными хроматографическими способами, например ионообменной хроматографией, аффинной хроматографией и тому подобного.
Предшественники инсулина или аналоги предшественников инсулина согласно изобретению могут быть экспрессированы с удлиненным аминокислотными остатками N-концом, как описано в патенте США 5395922 и Европейском патенте 765395А, оба этих патента специально включены здесь в виде ссылки. Обнаружено, что во время ферментации удлинение стабильно связано с предшественником инсулина или аналогами предшественника инсулина согласно изобретению, защищая N-конец предшественника инсулина или аналога предшественника инсулина от протеолитической активности дрожжевых протеаз, таких как DPAP. Присутствие удлинения N-конца в предшественнике инсулина или аналоге предшественника инсулина также может служить в качестве защиты N-концевой аминогруппы в ходе химической обработки белка, т.е. может служить как заместитель ВОС (трет-бутилоксикарбонила) или сходной защитной группы. N-концевое удлинение можно удалить из выделенного предшественника инсулина или аналога предшественника инсулина с помощью протеолитического фермента, который специфичен в отношении основной аминокислоты (например, Lys), так что концевое удлинение отщепляется у остатка Lys. Примерами таких протеолитических ферментов являются трипсин или протеаза Achromobacter lyticus.
После секреции в культуральную среду и выделения предшественник инсулина или аналоги предшественника инсулина согласно изобретению будут подвергнуты различным процедурам in vitro, чтобы удалить возможную последовательность удлинения N-конца и мини-С-пептид, чтобы получить инсулин или желаемый аналог инсулина. Такие способы включают ферментативное превращение с помощью трипсина или протеазы Achromobacter lyticus в присутствии сложного эфира L-треонина с последующим превращением сложного треонинового эфира инсулина или аналога инсулина в инсулин или аналог инсулина посредством основного или кислотного гидролиза, как описано в спецификации патента США 4343898 или 4916212 или Research Disclosure, September 1994/487, сообщения которых включены здесь в виде ссылки.
Как описано ниже, конструировали предшественники инсулина или аналоги предшественников инсулина с синтетическими С-пептидами, содержащими, по меньшей мере, одну ароматическую аминокислоту (пример 1). Экспрессирующие плазмиды Saccharomyces cerevisiae, содержащие полинуклеотидную последовательность, кодирующую заявленные предшественники инсулина или аналоги предшественников инсулина конструировали с помощью ПЦР и использовали для трансформации клетки-хозяина S. cerevisiae. Количество экспрессированного продукта, например аналога инсулина, измеряли в виде процента от соответствующего контрольного уровня, например AspB28IP, в котором отсутствует мини-С-пептид (таблица 1) и AspB28IP(AlaAlaLys) с мини-С-пептидом без остатка ароматической аминокислоты (таблица 2). Новые С-пептиды согласно изобретению давали увеличенные выходы до уровней в 7 раз выше контрольного.
Данное изобретение далее подробно описано на следующих примерах, которые никоим образом не предназначены для того, чтобы ограничить рамки заявленного изобретения. Предполагается, что прилагаемые фигуры должны рассматриваться как неотъемлемые части спецификации и описания изобретения. Все цитированные ссылки специально включены здесь в виде ссылки в отношении всего, что в них описано.
ПРИМЕРЫ Общие процедуры
Все экспрессирующие плазмиды являются плазмидами С-РОТ-типа, сходными с плазмидами, описанными в ЕР 171142, характерной особенностью которых является содержание гена триозофосфатизомеразы (РОТ) Schizosaccharomyces pombe с целью селекции и стабилизации плазмид в S. cerevisiae. Плазмиды также содержат промотор и терминатор триозофосфатизомеразы S. cerevisiae. Указанные последовательности сходны с соответствующими последовательностями в плазмиде pKFN1003 (описанной в WO 90/100075), так же как и все последовательности, за исключением последовательности EcoRI-XbaI фрагмента, кодирующего слитый белок лидера и продукта предшественника инсулина. Чтобы экспрессировать разные слитые белки EcoRI-XbaI фрагмент плазмиды pKFN1003 просто заменяют EcoRI-XbaI фрагментом, кодирующим представляющее интерес слияние лидер-предшественник инсулина. Такие EcoRI-XbaI фрагменты можно синтезировать, используя синтетические олигонуклеотиды и ПЦР, стандартными способами.
Трансформанты дрожжей получали путем трансформации штамма-хозяина S. cerevisiae, штамма МТ663 (MATalMATα рер4-3/рер4-3 HIS4/his4 tpi::LEU2/tpi::LEU2 Cir+). Штамм дрожжей МТ663 депонирован в Немецкой коллекции микроорганизмов и культур клеток (Deutsche Sammlung von Mikroorganismen und Zellkulturen) в связи с подачей заявки на патент WO 92/11378, и получил номер депозита DSM 6278.
МТ663 выращивали на YPGaL (1% дрожжевой бактоэкстракт, 2% бактопептон, 2% галактоза, 1% лактат) до OD при 600 нм, равной 0,6. 100 мл культуры собирали центрифугированием, промывали 10 мл воды, повторно центрифугировали и ресуспендировали в 10 мл раствора, содержащего 1,2 М сорбит, 25 мМ Na2EDTA, pH 8,0 и 6,7 мг/мл дитиотреитола. Суспензию инкубировали при 30°С в течение 15 минут, центрифугировали и клетки ресуспендировали в 10 мл раствора, содержащего 1,2 М сорбит, 10 мМ Na2EDTA, 0,1 М цитрат натрия, pH 5,8, и 2 мг Novozym®234. Суспензию инкубировали при 30°С в течение 30 минут, клетки собирали центрифугированием, промывали в 10 мл 1,2 М сорбита и 10 мл CAS (1,2 М сорбит, 10 мМ CaCl2, 10 мМ Трис-HCl (Трис = Трис(гидроксиметил)аминометан) pH 7,5) и ресуспендировали в 2 мл CAS. Для трансформации 1 мл клеток, суспендированных в CAS, смешивали примерно с 0,1 мг плазмидной ДНК и оставляли при комнатной температуре в течение 15 мин. Добавляли 1 мл раствора (20% полиэтиленгликоль 4000, 10 мМ CaCl2, 10 мМ Трис-HCl, рН 7,5) и смесь оставляли еще на 30 минут при комнатной температуре. Смесь центрифугировали, осадок ресуспендировали в 0,1 мл SOS (1,2 М сорбит, 33% об./об. YPD, 6,7 мМ CaCl2) и инкубировали при 30°С в течение 2 часов. Затем суспензию центрифугировали и осадок ресуспендировали в 0,5 мл 1,2 М сорбита. Затем добавляли 6 мл верхнего агара (среда SC Sherman et al. (1982) Methods in Yeast Genetics, Cold Spring Harbor Laboratory, содержащая 1,2 М сорбита плюс 2,5% агара) при 52°С и суспензию наливали сверху чашек, содержащих такую же агаризованную среду, содержащую сорбит.
Штамм МТ663 S. cerevisiae, трансформированный экспрессирующими плазмидами, выращивали в YPD в течение 72 час при 30°С. Количественный анализ выхода предшественника инсулина в надосадках культур осуществляли анализом обращенно-фазовой ВЭЖХ, используя в качестве внешнего стандарта инсулин человека (Snel and Damgaard (1988) Proinsulin heterogenity in pigs. Horm. Metabol. Res. 20: 476-488).
Пример 1
Конструирование синтетических С-пептидов с ароматической аминокислотой(ами)
Синтетические гены, кодирующие слитые белки, состоящие из Asp8B8IP, связанного с лидерной последовательностью, состоящей из препептида (сигнального пептида) и пропептида, конструировали, используя ПЦР в стандартных условиях (Sambrook et al. (1989) Molecular Cloning, Cold Spring Harbor Laboratory Press) и полимеразу E.H.F. (Boehringer Mannheim GmbH, Sandhoefer Strasse 116, Mannheim, Germany). Полученные в результате фрагменты ДНК выделяли и расщепляли эндонуклеазами, очищали, используя набор для очистки генов (Bio101 Inc., La Jolla, CA, USA). Для лигирования ДНК использовали стандартные способы, трансформацию клеток Е. coli осуществляли CaCl2-способом (Sambrook et al. (1989), выше). Плазмиды очищали из трансформированных клеток, используя колонки QIAGEN (QIAGEN, Hilden, Germany). Определяли нуклеотидные последовательности, используя систему секвенирования ДНК ALF Pharmacia Biotech с очищенной двунитевой плазмидной ДНК в качестве матрицы. Олигонуклеотидные праймеры для ПЦР получали из DNA technology (Arhus, Denmark).
Экспрессию секретируемого AspB28IP в S. cerevisiae осуществляли, используя штамм МТ663 S. cerevisiae и экспрессирующий дрожжевой вектор СРОТ на основе 2 мкм-плазмиды (смотри фиг.1), как описано в Thim, L. et al. (1986) Proc. Natl. Acad. Sci. USA 83: 6766-6770. Экспрессирующий дрожжевой вектор содержит ген триозофосфатизомеразы (РОТ) Schizosaccharomyces pombe для селекции и стабилизации плазмиды в S. cerevisiae. Кроме того, используют промотор и терминатор гена триозофосфатизомеразы (ТРИ) S. cerevisiae для инициации и терминации транскрипции рекомбинантного гена, кодирующего слитый белок лидер-AspB28IP. Секрецию AspB28IP обеспечивали лидером α-фактора, хотя можно использовать множество известных дрожжевых лидерных последовательностей.
Как показано на фиг.1, экспрессирующая плазмида рАК721 S. cerevisiae, которая экспрессирует слитый белок лидер LA19-ЕЕАЕАЕАЕРК (SEQ ID NO:3)-1Р, конструировали на основе челночной плазмиды РОТ S. cerevisiae - E. coli(патент США 5871957). На фиг 1. L-IP обозначает кассету, экспрессирующую слитый белок, кодирующую слитый белок лидер-IP; TPI-ПРОМОТОР означает промотор TPI1 S. cerevisiae, а TPI-ТЕРМИНАТОР означает терминатор TPI1 S. cerevisiae; TPI-POMBE обозначает ген РОТ S. pombe, используемый для селекции в S. cerevisiae; НАЧАЛО РЕПЛИКАЦИИ обозначает начало репликации S. cerevisiae, полученное из 2 мкм-плазмиды; AMP-R обозначает ген (S-лактамазы, придающий устойчивость к ампициллину, облегчающий селекцию в Е coli, и НАЧАЛО РЕПЛИКАЦИИ-PBR322 обозначает начало репликации Е. coli.
ДНК, кодирующую ряд слитых белков лидерных последовательностей и AspB28IP с различными мини-С-пептидами, создавали посредством ПЦР, используя в качестве праймеров соответствующие олигонуклеотиды, которые описаны ниже. Использовали стандартные способы для того, чтобы субклонировать фрагменты ДНК, кодирующие слитые белки лидер-Asp828IP, в экспрессирующем векторе СРОТ в следующем составе: лидер-Lys-Arg-спейсер-AspB28IP, где Lys-Arg является потенциальным диосновным сайтом процессинга эндопротеазой и спейсер является N-концевым удлинением. Чтобы оптимизировать процессинг слитого белка эндопротеазой Кех2 S. cerevisiae, ДНК, кодирующую спейсерный пептид (N-концевое удлинение), например, ЕЕАЕАЕАРК (SEQ ID NO: 4), встраивали между ДНК, кодирующими лидер и AspB28IP (Kjeldsen et al. (1996) Gene 170, 107-112.). Однако наличие спейсерного пептида не обязательно. Зрелый AspB28IP секретировался в виде одноцепочечного предшественника инсулина с удлиненным N-концом и мини-С-пептидом, связывающим LysB29 и GlyA1. После очистки AspB28IP и протеолитического удаления удлинения N-конца и мини-С-пептида к LysB29 можно добавить аминокислоту Thr830 с помощью опосредованной ферментом транспептидации, чтобы получить Asp828-инсулин человека (Markussen, et al. (1987) in «Peptides 1986» (Theodoropoulos, D., Ed.), p.189-194, Walter de Gruyter and Co., Berlin.).
Разработку синтетических мини-С-пептидов осуществляли рандомизацией одного или нескольких кодонов, кодирующих аминокислоты в мини-С-пептиде. Характерной особенностью всех синтетических мини-С-пептидов является сайт ферментативного процессинга (Lys) на С-конце, который позволяет осуществлять ферментативное удаление синтетического мини-С-пептида (патент США 4916212, специально включенный здесь в виде ссылки). Рандомизацию выполняли, используя олигонуклеотиды с изменяемой структурой, которые вводили изменения кодона(нов) в одном или нескольких положениях синтетических мини-С-пептидов. Обычно изменяли структуру одного из двух праймеров (олигонуклеотидов), используемых для ПЦР. Примером пары олигонуклеотидов, используемой для создания методом ПЦР лидер-AspB28IP со случайными синтетическими мини-С-пептидами, применяемой для создаваемых синтетических мини-С-пептидов общей формулы: Xaa-Trp-Lys (XWK), являются следующие олигонуклеотиды:
Праймер А: 5'-ТАААТСТАТААСТАСААААААСАСАТА-3' (SEQ ID NO:13) и
Праймер В: 3'-CCAAAGAAGATGTGACTGTTCNNMACCTTCCCATAGCAACTT GTTACAACATGAAGATAGACAAGAAACATGGTTAACCTTTTGATGACATTGATCAGATCTTT GA-TTC-5' (SEQ ID NO:14), где N означает А, С, G или Т, а М означает С или А.
Полимеразная цепная реакция. ПЦР обычно выполняли, как указано ниже: 5 мкл праймера А (20 пмоль), 5 мкл праймера В (20 пмоль), 10 мкл 10 × буфера для ПЦР, 8 мкл смеси дНТФ, 0,75 мкл фермента E.H.F., 1 мкл плазмиды рАК1150 в качестве матрицы (примерно 0,2 мкг ДНК) и 70,25 мкл дистиллированной воды.
Обычно выполняли от 10 до 15 циклов, один цикл обычно представлял собой 94°С в течение 45 сек.; 55°С в течение 1 мин; 72°С в течение 1,5 мин. Затем смесь ПЦР наносили на 2% агарозный гель и выполняли электрофорез, используя стандартные способы. Полученный в результате фрагмент ДНК вырезали из агарозного геля и выделяли с помощью набора для очистки генов.
На фиг.2 показана последовательность ДНК рАК1150, используемая в качестве матрицы для ПЦР, и рассчитанные аминокислоты кодируемого слитого белка (α-фактор-лидер-ЕЕАЕАЕАРК (SEQ ID NO:4)-AspB28IP плазмиды рАК1150 (SEQ ID NO:5 и 6). Плазмида рАК1150 сходна с рАК721, показанной на фиг.1. С-конец лидера α-фактора модифицировали для того, чтобы ввести сайт рестрикции эндонуклеазой Nco I, который изменяет рассчитанные аминокислотные последовательности с SerLeuAsp на SerMetAla. Кроме того, кодируемый AspB28IP не содержит мини-С-пептида, a Lys непосредственно связан с GlyA1.
Очищенный ПЦР-фрагмент ДНК растворяли в воде и буфере для эндонуклеаз рестрикции и расщепляли подходящими рестрикционными эндонуклеазами (например, Bgl II и Xba I) стандартными способами. BglII-Xbal-фрагменты ДНК подвергали электрофорезу в агарозном геле и очищали, используя набор для очистки генов.
Экспрессирующую плазмиду рАК1150 или сходную плазмиду СРОТ-типа. (смотри фиг.1) расщепляли эндонуклеазами рестрикции Bgl II и Xba I и выделяли фрагмент вектора длиной 10765 пар нуклеотидных оснований, используя набор для очистки генов.
Два расщепленных и выделенных фрагмента ДНК (фрагмент вектора и ПЦР-фрагмент) лигировали вместе, используя ДНК-лигазу Т4 и стандартные условия. Затем лигированной смесью трансформировали компетентный штамм Е. coli (R-, М+), после чего проводили селекцию на устойчивость к ампициллину. Плазмиды из полученных в результате клеток Е. coli выделяли с использованием колонок QIAGEN.
Затем плазмиды использовали для трансформации подходящего штамма-хозяина S. cerevisiae, например, МТ663 (MATalMATα рер4-3/рер4-3 HIS4/his4 tpi::LEU2/tpi::LEU2 Cir+). Отдельные трансформированные клоны S. cerevisiae выращивали в жидкой культуре и определяли количество AspB28IP, секретированного в надосадки культуры, с помощью ОФ-ВЭЖХ. Затем определяли последовательность ДНК, кодирующую синтетический мини-С-пептид экспрессирующих плазмид клонов S. cerevisiae, секретирующих повышенное количество AspB28IP. Затем идентифицированную последовательность синтетического мини-С-пептида можно подвергнуть еще одному раунду рандомизированной оптимизации.
Пример последовательности ДНК, кодирующей слитый белок лидер-Asp828IP(GluTrpLys), содержащий синтетический мини-С-пептид (GluTrpLys), полученный в результате описанного способа рандомизированной оптимизации, показан на фиг.3 (SEQ ID NO:7 и 8).
В таблице 1 и 2 показаны аналоги предшественников инсулина, созданные описанным выше способом, и выход продукции, выраженный в виде процента от контроля. Ферментацию проводили при 30°С в течение 72 час в 5 мл YPD. Выход предшественника инсулина определяли посредством ОФ-ВЭЖХ в надосадке культуры и выражали относительно выхода в контрольном штамме, экспрессирующем либо слитый белок лидер-AspB28IP, в котором остаток В29 связан с остатком А1 пептидной связью; либо слитый белок лидер-AspB28IP, в котором остаток В29 связан с остатком А1 мини-С-пептидом, соответственно. В таблицах «α*» указан лидер α-фактора, в котором С-конец до LysArg был изменен с «SLD (SerLeuAsp)» на «SMA (SerMetAla)», и «ех4» обозначено N-концевое удлинение с аминокислотной последовательностью ЕЕАЕАЕАРК (SEQ ID NO:4). YАР3 означает сигнальную последовательность YAP3, ТА39 означает синтетическую пропоследовательность QPIDDTESNTTSVNLMADDTESRFATNTTLAGGLDVVNLISMAKR (SEQ ID NO:16). Последовательность EEGEPK (SEQ ID NO:17) представляет собой N-концевое удлинение В-цепи аналога инсулина. ТА57 означает синтетическую пропоследовательность QPIDDTESQTTSVNLMADDTESA FATQTNSGGLDVVGLISMAKR (SEQ ID NO:18).
Пример 2
Определение структуры Asp328IP (MetTrpLys) в водном растворе способом ЯМР-спектроскопии ЯМР-спектроскопия.
Образцы для ЯМР готовили растворением лиофилизированного белкового порошка в 10/90 D2O/H2O с 10 мМ фосфатным буфером, и рН доводили до нужного значения добавлением небольших объемов 1 М DCl или NaOD. Все показания рН-метра даны без корректировки на влияние изотопа. Образцы Asp828IP (MetTrpLys) для ЯМР готовили в концентрациях в пределах от 25 мкМ до 1 мМ при рН 8,0. Двумерные спектры 1H-1Н-ЯМР 1 мМ образцов, DQF-COSY (Piantini et al. (1982) J. Am. Chem. Soc. 104: 6800-6801, Rance et al. (1983) Biochem. Biophys. Res. Commun. 117: 479-485), TOCSY (Braunschweiler et al. (1983) J. Magn. Reson. 53: 521-528, Bax et al. (1985) J. Magn. Reson. 65: 355-360) и NOESY (Jeener et al. (1979) J. Chem. Phys. 71: 4546-4553) записывали при 600 МГц на ЯМР-спектрометре Varian Unity Inova, оборудованном 1H/13С/15N-датчиком тройного резонанса с самоэкранированной градиентной катушкой по трем осям, используя стандартные импульсные последовательности из библиотеки Varian для пользователей. Рабочая температура была установлена на уровне 27°С. Для каждого фазочувствительного двумерного ЯМР-спектра получали 512 t1-инкрементов, каждый с 2048 или 4096 реальными точками данных, способом TPPI-States (Marion et al. (1989) J. Magn. Reson. 85: 393-399). Использовали ширину спектров, равную 6983 Гц, в обоих измерениях, при этом опорная частота была расположена точно на резонансном сигнале воды, который подавляли либо насыщением между сканами в течение 1,5 сек, либо селективным возбуждением импульсной последовательностью возбуждения заданного градиента (WATERGATE, Piotto et al. (1992) J. Biomol. NMR 2: 661-665). Спектры DQFCOSY записывали, используя градиентно-усиленную версию с применением градиентов под магическим углом (Mattiello et al. (1996) J. Am. Ch'em. Soc. 118: 3253-3261). Для спектров TOCSY использовали время смешивания от 30 до 80 мсек, а для NOESY время смешивания составляло от 50 до 200 мсек.
Обработку двумерных ЯМР-спектров осуществляли с использованием пакета программ Xwinnmr (версия 2.5, компьютерная программа для обработки данных ЯМР Bruker Analytische Messtechnik GmbH, D-76275 Ettlingen, Germany). Каждое измерение обрабатывали с аподизацией сдвинутого «колокола» функции синуса и заполнением нулями, выполненном один раз в каждом измерении. При необходимости применяли корректировки базовой линии, используя стандартные способы Xwinnmr.
Спектральное отнесение, интегрирование кросс-пиков, специфичное отнесение последовательностей, стереоспецифичное отнесение и все другие типы обработок выполняли, используя программу PRONTO (PRONTO Software Development and Distribution, Copenhagen Denmark) (Kjaer et al. (1991) NATO ASI Series (Hoch, J.C., Redfield C., and Poulsen, F.M., Eds.) Plenum, New York). Химические сдвиги измеряли в м.д., и резонансный сигнал воды устанавливали при 4,75 м.д.
Расчеты структуры. Ограничения расстояний для последующего расчета структуры получали из интегрированных кросс-пиков NOESY, классифицированных как слабые, средние или сильные, соответствующие верхним ограничениям расстояний 5,5; 3,3 и 2,7 Å, соответственно. Что касается ограничений расстояний, относящихся к метильным группам, к верхнему пределу дополнительно добавляли 0,5 Å (Wagner et al. (1985) J. Mol. Biol. 196: 611-639). Расчеты структур выполняли, используя гибридный способ, объединяющий геометрию расстояний (Crippen et al. (1988) Distance Geometry and Molecular Conformation, Research Studies Press, Taunton, Somerset, England; Kuszewski et al. (1992) J. Biomol NMR 2: 33-56) и модельный отжиг, основанный на идеях Nilges et al. (1988) FEBS Lett. 229: 317-324, используя Х-PLOR 3.0 (Brünger (1992) версия 3.1 X-PLOR: A System for X-ray Crystallography and NMR, Yale University Press, New Haven) в соответствии с примерами, приведенными в руководстве X-PLOR manual (dg_sub_embed.inp.dgsa.inp,refine.inp). Номера остатков взяты из стандартной нумерации остатков инсулина, остатки в В-цепи пронумерованы В1-29, остатки в С-пептиде (например, MetTrpLys) пронумерованы С1-С3, и остатки в А-цепи пронумерованы А1-А21.
При спектральном отнесении ЯМР-спектра для большинства резонансов следовали стандартному способу последовательного отнесения, описанного Wüthrich (1986 NMR of Proteins and Nucleic Acids, Wiley, New York). Стандартный способ отнесения был неудачным в том случае, когда протон амида отдельного остатка аминокислоты быстро обменивался с протонами в воде. При рН 8,0 это имело место для нескольких аминокислотных остатков, однако сравнение с более ранними отнесениями ЯМР-спектров мутантного инсулина и идентификация соседних (в пространстве) аминокислотных остатков с помощью NOE дают возможность для получения почти полного спектрального отнесения. Анализ спектров NOESY показал, что несколько аминокислотных остатков образуют сеть взаимодействия NOE с окружающими остатками, сходную с взаимодействием, которое ранее было установлено для других молекул инсулина, т.е. мутанта HisB16 инсулина человека (Ludvigsen et al. (1994) Biochemistry 33: 7998-8006), и указанные сходные связи были обнаружены для остатков В1-В10, В13-В14, В17-В24 и А4-А21. Кроме того, ограничения двугранных углов для перечисленных выше остатков приняты, исходя из ограничений, использованных ранее (Ludvigsen et al. (1994), выше).
Несколько аминокислот, в частности В27-В29, С1-С3, А1-А3, имеют картины кросс-пиков, которые согласуются с пептидными цепями, которые менее упорядочены, чем хорошо определенные элементы вторичной структуры в большинстве случаев. Таким образом, дополнительные величины NOE превращали в ограничения расстояний без какой-либо дальнейшей классификации, кроме верхних пределов 5,5 Å или 6,0 Å в том случае, если включали метильную группу. Рассчитывали набор 20 конвергентных структур (фиг.5) и соответствующие параметры конвергентных структур перечислены в таблице 3. Каждое значение NOE, которое здесь идентично ограничению расстояния, рассчитано только один раз, даже хотя оно может несколько раз встречаться в спектре NOESY. Качественное отнесение на карте Рамачандрана представляет собой стандартные качественные параметры для оценки особенности локальной геометрии. В общем, описанные качественные параметры сравнимы со структурами белков, основанными на рентгеноструктурном анализе при разрешении 2,5 Å (Laskowski et al. (1996) J. Biomol. NMR 8: 477-486).

Описание рассчитанной структуры
Характерная структура, наиболее близкая к средней из группы, показана на фиг.6. AspB28IP (MetTrpLys) структурно сходен со структурой нативного инсулина в районах, содержащих остатки В1-В10, В14-В23, А4-А21. Различия, главным образом, четко выражены в районах вблизи соединительного пептида в положениях В26-В29, С1-С3, А1-А3 и менее выражены для остатков В11-В13. Структура AspB28IP (MetTrpLys) вблизи С-пептида значительно отличается от структуры, подобной нативной структуре (Ludvigsen (1994) выше). Боковые цепи метионина и триптофана в AspB28IP(MetTrpLys) открывают традиционную структуру кора инсулина, отодвигая в одну сторону боковые цепи отдельных TyrB26 и PheB25 и оставляя интактным другой обычный близлежащий гидрофобный участок, состоящий из боковых цепей LeuB11, ValB12, IleA2 и TyrA19. Указанный карман, полученный передвижением PheB25, TyrB26 и пептидная цепь, образованная остатками с В25 по В29, по-видимому, хорошо подходят для размещения боковых цепей MetC1 и TrpC2 из С-пептида. Несколько NOE от указанных двух боковых цепей к близлежащим в структуре остаткам подтверждают указанное новое расположение боковых цепей, не наблюдаемое ранее ни в одной структуре инсулина. MetC1 размещается в кармане, образованном остатками LeuB15, PheB24, TyrB26, TrpC1 IleA2 и TyrA19, все из которых имеют NOE с MetC1. TrpC2 имеет даже более протяженную сеть NOE, но в результате факта обмена протона амида индола можно отнести только четыре резонанса, относящихся к ароматической кольцевой системе TrpC2. Несмотря на это в спектре NOESY AspB28IP (Met Trp Lys) было обнаружено 21 NOE между остатками между TrpC2 и соседними с ним положениями, занятыми LeuB11, ValB12, LeuB15, TyrB26, MetC1 и IleA2.
Присутствие боковой цепи триптофана в кармане также оказывает большое влияние на химические сдвиги, наблюдаемые в спектре AspB28IP(Met Trp Lys). В условиях, используемых- для ЯМР на спектры AspB28IP(Met Trp Lys) в некоторой степени влияет самоассоциация (фиг.7), но обмен между мономером и димером на временной шкале ЯМР наблюдается только как среднее между двумя состояниями. Между концентрациями 25 мкМ и 0,2 мМ степень самоассоциации не меняется, как видно при ЯМР.
В таблице 4 показаны химические сдвиги AspB28IP (MetTrpLys) при 27° по Цельсию, полученные при 600 мГц, рН 8 в 10%/90% D2O/H2O с 10 мМ фосфатным буфером. Химические сдвиги сравнивали с эталоном при установке остаточного сигнала воды при 4,75 м.д. N/A означает, что нет отнесения. Отнесения AspB28IP (MetTrpLys) - (1-29=В1-В29; 30-32=C1-C3 и 33-53=A1-A21), и таблица 5 дает координаты атомов AspB28IP (MetTrpLys) в форме PDB.
Пример 3
Относительная стабильность укладки аналогов AspB28IP.
Для оценки стабильности укладки аналогов предшественников инсулина, образцы денатурации получали, комбинируя разные соотношения аналога предшественника инсулина и исходных растворов GuHCl с 10 мМ Трис/ClO4 -, рН 8,0. Концентрация исходных растворов белков обычно составляла 0,06 мМ в 10 мМ Трис/ClO4 -, рН 8,0. Концентрация исходных растворов GuHCl составляла 8,25 М в 10 мМ Трис/ClO4 -, рН 8,0. Спектры КД записывали на спектрополяриметре Jasco J-715, откалиброванном с помощью (+)-10-камфаросульфоновой кислотой. Все спектры записывали при 20°С. Образцы денатурации сканировали от 250 до 218 нм. Типичная длина оптического пути в кювете и концентрация белка составляли 0,5 см и 3 мкМ соответственно. Все спектры сглаживали перед вычитанием соответствующих пустых контролей с растворителем. Круговой дихроизм выражали в виде Δε на основании молярной концентрации пептидной связи. В целях представления каждую кривую нормализовали по шкале 0-1 делением наблюдаемого изменения в каждой точке на общее изменение, наблюдаемое в эксперименте.
Анализ данных. Кривые денатурации GuHCl анализировали, полагая, что переход укладка/разворачивание представляет собой два состояния, как описано Santoro and Bolen (1988) Biochemistry 27: 8063-8068 and Kaarsholm et al. (1993) Biochemistry 32: 10773-10778, и обе эти публикации специально включены здесь в виде ссылки в качестве пособий в отношении способов расчета стабильности при денатурации GuHCl. Указанный анализ дает ряд параметров, включая концентрацию' GuHCl в средней точке кривой денатурации, Cmid, отражающую концентрацию денатурирующего агента, необходимую для того, чтобы развернуть половину популяции белков. Таким образом, увеличение стабильности укладки проявляется в увеличении значения Cmid. Константы равновесия можно получить при каждой концентрации денатурирующего агента, используя K=(ΔεN-Δε)/(Δε-ΔεU), где Δε означает наблюдаемое значение КД, а ΔεN и ΔεU представляют значения КД для нативной и развернутой форм, соответственно, при данной концентрации GuHCl (Расе, 1975). Значения ΔεN и ΔεU при концентрациях GuHCl в районе перехода получают линейной экстраполяцией базовых линий до и после перехода в район перехода, т.е. ΔεN=Δε°N+mN[GuHCl], и ΔεU=Δε°U+mU[GuHCl], где Δε°N и Δε°U означают пересечения, а mN и mU являются наклонами базовых линий до и после перехода соответственно. Свободная энергия развертывания при данной концентрации денатурирующего агента в зоне перехода имеет вид ΔG=-RTInK. Допуская линейную зависимость ДС от концентрации денатурирующего агента: ΔG=ΔGH2O-m[GuHCl], где ΔGH2O представляет собой значение ΔG в отсутствии денатурирующего агента, a m является мерой зависимости ΔG от концентрации денатурирующего агента. Следовательно, значения ΔG, полученные при К в зоне перехода, можно экстраполировать назад к 0 М денатурирующего агента, чтобы получить ΔGH2O. Взаимосвязь между Δε и [GuHCl] для кривой полного разворачивания показана в уравнении 1 (Santoro and Bolen, 1988):

При Δε в качестве ответного сигнала и [GuHCl] в качестве независимой переменной уравнение (1) подлежит нелинейному анализу методом наименьших квадратов с использованием процедуры NLIN PC SAS (SAS Inc. Cary, North Carolina). В таком случае кривая денатурации описывается шестью параметрами: Δε°N, Δε°U, mN, mU, m и ΔGH2O. Кроме того, концентрация GuHCl в средней точке кривой денатурации, Cmid, определяется ΔGH2O/m.
Оценку относительной стабильности укладки молекул производных AspB28IP с С-пептидом Met Trp Lys (AspB28IP(MetTrpLys)) проводили относительно AspB28IP. Результаты показывают, что молекула AspB28IP(MetTrpLys) гораздо более стабильна, чем AspB28IP (фиг.5), что очевидно на основании изменения Cmid. В то время как Cmid для AspB28IP составляет примерно около 5,5 М GuHCl, для AspB28IP(MetTrpLys) она увеличивается, по меньшей мере, примерно до 6,5 М GuHCl, при этом увеличение составляет примерно 18%.
Пример 4
Предшественник аналога инсулина AspB28IP(EWK) получили, культивируя штамм дрожжей МТ663, трансформированный экспрессирующей плазмидой, которая экспрессирует слитый белок YAP3-TA39-EEGEPK (SEQ ID NO:17)-AspB28IP (EWK) или слитый белок YAP3-TA57-EEGEPK (SEQ ID NO:17)-AspB28IP (EWK).
кДНК, кодирующую лидерные последовательности YAP3-TA39 и YAP3-TA57, и кДНК, кодирующие AspB28IP (EWK) и N-концевое удлинение, клонировали в экспрессирующем векторе С-РОТ-типа, используя стандартные способы (Sambrook J, Fritsch EF and Maniatis Т, Molecular cloning. Cold spring Harbour laboratory press, 1989). Последовательность ДНК и рассчитанные аминокислотные последовательности показаны на фиг.8 и 9.
В таблице 6 показаны выходы. Ферментацию проводили при 30°С в течение 72 час в 5 мл YPD. Выход IP определяли ОФ-ВЭЖХ надосадка культуры и выражали относительно выхода IP контрольного штамма.
В таблице 6 «α*» указан лидер α-фактора, в котором С-конец до LysArg был изменен с «SLD (SerLeuAsp)» на «SMA (SerMetAla)», и «ех4» обозначено N-концевое удлинение с аминокислотной последовательностью ЕЕАЕАЕАРК (SEQ ID NO:4). YAP3 означает сигнальную последовательность YAP3. ТА39 означает синтетическую пропоследовательность QPIDDTESNTTSVNLMADDTESRFATNTTLAGGLDVVNLISMAKR (SEQ ID NO:16). Последовательность EEGEPK (SEQ ID NO:17) представляет собой N-концевое удлинение В-цепи аналога инсулина. ТА57 означает синтетическую пропоследовательность QPIDDTESQTTSVNLMADDTESAFATQTNSGGLDVVGLISMAKR (SEQ ID NO:18).
Пример 5
Расщепление инсулинового предшественника продеазой Achromobacter lyticos (ALP)
В 1 мл 50 мМ глутамата натрия, рН 9,0, растворяли 10 мг одноцепочечного предшественника инсулина B28Asp desB30, содержащего на N-конце последовательность EEGEPK и в качестве соединительного пептида между В29 и А1 последовательность MWK. Послед того как рН доводили до 9,0, добавляли 12 мкл водного раствора, содержащего 100 мкг ALP, и реакционную смесь оставляли при температуре 23 С. Через 0,5 часа, 1 час и 2 часа отбирали образцы для анализа ВЭЖХ с обращенной фазой, и результаты показали 98%-ное преобразование в инсулин des(B30Thr) B28Asp (инсулин Aspart) через 2 часа. Данные показаны на фигуре 10.

















Изобретение относится к области биотехнологии и может быть использовано в медико-биологической промышленности. Предложены новые полипептиды - предшественники инсулина или его аналогов, характеризующиеся повышенной стабильностью структуры и высоким уровнем экспрессии при получении в виде рекомбинантных форм в дрожжевых клетках. Полипептиды по изобретению отличаются от природной формы предшественника инсулина тем, что содержат укороченный С-пептид, включающий остаток ароматической аминокислоты, и имеют общую формулу: В (1-27)-X2-X3-X1-Y-A (1-21), где Х1-пептидная последовательность из 1-3 аминокислотных остатков, в которой рядом с Y расположен остаток ароматической аминокислоты; Х2-Pro или Asp; Х3-Lys; Y-Lys или Arg; A (1-21) - А-цепь инсулина человека, а В (1-27) - первые 27 аминокислотных остатков В-цепи инсулина человека. Раскрыт способ получения новых предшественников инсулина с помощью технологии рекомбинантных ДНК и последующего преобразования их в активный гормон. 5 с. и 4 з.п. ф-лы, 6 табл., 10 ил.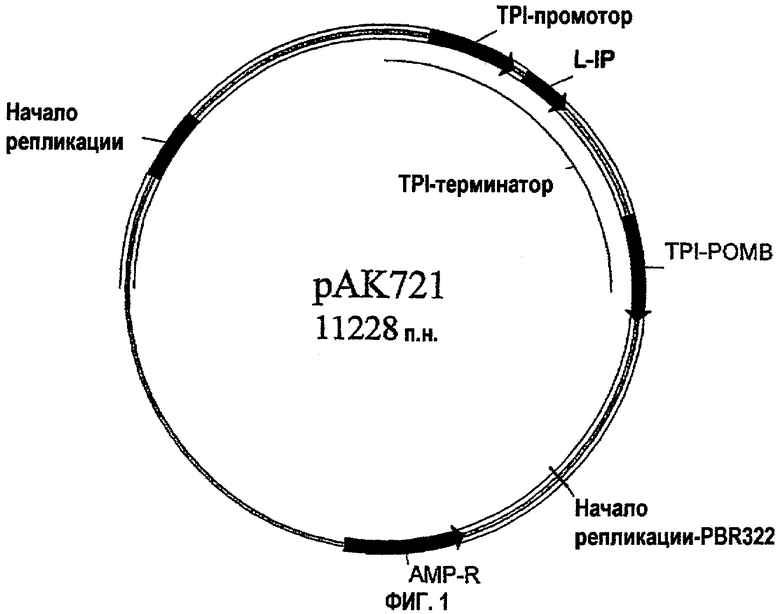
В (1-27)-Х2-Х3-Х1-Y-А (1-21),
где X1 представляет собой пептидную последовательность из 1-3 аминокислотных остатков, содержащую остаток ароматической аминокислоты непосредственно у N-конца Y;
Х2 представляет собой Pro или Asp;
Х3 представляет собой Lys в положении 29 В-цепи; и
Y представляет собой Lys или Arg;
В(1-27) обозначает природную В-цепь, в которой отсутствуют аминокислотные остатки В28, В29 и В30;
А(1-21) обозначает А-цепь инсулина человека.
Приоритет по пунктам:
| WO 9516708 A1, 22.06.1995 | |||
| US 5962267 A, 05.10.1999 | |||
| US 4916212 A, 10.04.1990. |

