ОБЛАСТЬ ТЕХНИКИ
Изобретение относится к дизрупции генов протеин-маннозилтрансфераз (Protein mannosyi transferase, PTM) Pichia pastoris, приводящей к снижению уровня O-гликозилирования молекулы предшественника инсулина, продуцируемой Pichia sp.Настоящее изобретение обеспечивает нокаутированные с помощью генной инженерии штаммы метилотрофных дрожжей, в том числе рода Pichia, в частности, вида Pichia pastoris, способные продуцировать гетерологичные белки со сниженной степенью гликозилирования. Настоящим изобретением предусмотрены векторы, содержащие кодирующие последовательности генов РМТ1, РМТ2, РМТ4, РМТ5 и РМТ6 для трансформации метилотрофных дрожжей.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Рекомбинантные формы инсулина, аналогов инсулина и/или его производных были продуцированы в различных микробных системах экспрессии. В настоящее время организмы, такие как E. coli и S. cerevisiae, применяют для промышленного производства рекомбинантного человеческого инсулина и его производных. Вследствие некоторых недостатков указанных систем, таких как низкий уровень экспрессии, трудности в последующей очистке и т.п., предпочтение отдается применению метилотрофных дрожжей Pichia pastoris в качестве системы для экспрессии белков. Данная экспрессионная система имеет ряд преимуществ, таких как высокий уровень экспрессии, простая переработка, низкая стоимость производства, высокая плотность культуры (US 6800606).
Дрожжевые экспрессионные системы широко распространены, поскольку их легко выращивать, они характеризуются быстрым ростом и являются масштабируемыми, однако некоторые дрожжевые экспрессионные системы дают противоречивые результаты, и иногда бывает трудно добиться высокого выхода. Одну из дрожжевых экспрессионных систем, показавшую перспективность применения, представляют метилотрофные дрожжи Pichia pastoris. По сравнению с другими эукариотическими экспрессионными системами, Pichia обладает многими преимуществами, поскольку не влечет за собой проблем, связанных с эндотоксинами, ассоциированных с бактериями, или проблем загрязнения белков, продуцируемых в культуре клеток животных, вирусами (Cino, Am Biotech Lab, May 1999).
Несмотря на различные преимущества, характерные для экспрессионных систем на основе дрожжей, таких как Pichia pastoris, одним из основных недостатков этой системы является посттрансляционная модификация образующихся белков, что впоследствии приводит к наличию примесей в конечном продукте, от которых его трудно очистить. Хотя известен целый ряд посттрансляционных модификаций белков, наиболее распространенной формой посттрансляционной модификации является гликозилирование. (Hart G.W, Glycosylation, Curr. Opin. Cell. Biol 1992; 4: 1017). Гликозилирования может быть либо N-связанным, либо O-связанным, в зависимости от экспрессионной системы. (Gemmill TR et al., Overview of N- and O-linked oligosaccharide structures found in various yeast species, Biochemica et Biophysica Acta, 1999; 1426:227). Гликозилирование влияет на стабильность конформации белка, иммуногенность, скорость выведения из организма, защиту от протеолиза и повышает растворимость белка (Walsh G, Biopharmaceutical benchmarks 2006, Nature Biotechnology, 2006; 24:769).
У дрожжей модификация сахарных ветвей в аппарате Гольджи включает в себя ряд присоединений остатков маннозы различными маннозилтрансферазами («внешнецепочечное» гликозилирование). Структура внешнецепочечного гликозилирования является специфичной для конкретного организма. Такое гликозилирование часто является нежелательным, поскольку оно приводит к неоднородности рекомбинантного белкового продукта по составу углеводов и молекулярной массе, что может затруднять очистку белков. Это также может обуславливать более высокую иммуногенность белка, и может провоцировать аллергические реакции, которые являются нежелательными.
Несмотря на значительный прогресс в улучшении биотехнологического производства, не существует общего решения проблемы для каждого белка. Процесс производства каждого конкретного терапевтического белка требует новых и инновационных путей решения проблем, которые могут быть специфичными для данного белка или семейства белков.
Таким образом, желательно создание с помощью генно-инженерных способов штаммов метилотрофных дрожжей, таких как Pichia pastoris, в которых степень гликозилирования белков можно регулировать, существенно снизить, и в которых можно получать рекомбинантные гликопротеины с посттрансляционным паттерном, схожим с таковым для млекопитающих, при этом не затрагивая продуктивность желаемого конечного продукта.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Таким образом, настоящее изобретение относится к способу снижения гликозилирования белка, продуцируемого штаммом метилотрофных дрожжей, посредством инактивации по меньшей мере одного или более гена, выбранного из группы, включающей гены РМТ1, РМТ2, РМТ4, РМТ5 и РМТ6, имеющие нуклеотидную последовательность, которая по меньшей мере на 80% гомологична нуклеотидной последовательности, представленной в SEQ ID NOs 1, 2, 3, 4 и 5 соответственно, при этом указанные последовательности кодируют протеин-маннозилтрансферазу или ее функциональную часть; вектор, содержащий ген протеин-маннозилтрансферазы или его функциональную часть, выбранный из группы, включающей гены РМТ1, РМТ2, РМТ4, РМТ5 и РМТ6, имеющие нуклеотидную последовательность, которая по меньшей мере на 80% гомологична нуклеотидной последовательности, представленной в SEQ ID NOs 1, 2, 3, 4 и 5 соответственно, при этом встраивание вектора в гомологичный локус ингибирует экспрессию функциональной протеин-маннозилтрансферазы в организме-хозяине, предпочтительно, в клетке метилотрофных дрожжей; способу получения нокаутированного штамма метилотрофных дрожжей, в котором (а) вектор, включающий последовательность нуклеиновой кислоты, способную к гомологичной рекомбинации, содержащую целевую последовательность нуклеиновой кислоты, кодирующую по меньшей мере один из генов, выбранных из группы, включающей РМТ1, РМТ2, РМТ4, РМТ5 и РМТ6, имеющие нуклеотидную последовательность, которая по меньшей мере на 80% гомологична нуклеотидной последовательности, представленной в SEQ ID NOs 1, 2, 3, 4 и 5, соответственно, и последовательность нуклеиновой кислоты, кодирующую селективный маркер, (б) культивирование клеток при условиях, делающих возможной гомологичную рекомбинацию между ДНК, кодирующей целевой ген в векторе, и ДНК клетки-хозяина, что таким образом приводит к дизрупции целевого гена в клетке-хозяине, (в) селекцию клеток-хозяев с инактивированным целевым геном; белку, продуцируемому с помощью способа, описанного выше; «нокаутированному» штамму метилотрофных дрожжей, где указанный штамм содержит по меньшей мере один инактивированный ген, выбранный из группы, включающей РМТ1, РМТ2, РМТ4, РМТ5 и РМТ6, имеющий нуклеотидную последовательность, которая по меньшей мере на 80% гомологична нуклеотидной последовательности, представленной в SEQ ID NOs 1, 2, 3, 4 и 5 соответственно; штамм с инактивированными геном РМТ1, как описано выше; штамм с инактивированными геном РМТ4, как описано выше; штамм с инактивированными геном РМТ5, как описано выше; штамм с инактивированными геном РМТ6, как описано выше; белок, продуцируемый «нокаутированным» штаммом, как описано выше; белок, продуцируемый «нокаутированным» штаммом по п.23 формулы изобретения, где «нокаутированный штамм» представляет собой один из штаммов МТСС5515, МТСС5516, МТСС5517, 5518 МТСС, или модифицированный штамм на его основе; и белок, соответствующий любому из вышеуказанных описаний, при этом белок демонстрирует измененный характер гликозилирования.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к способам получения белка со сниженным уровнем гликозилирования, при этом указанный способ предусматривает применение векторов, с помощью которых генетически модифицируют штаммы метилотрофных дрожжей, что позволяет им продуцировать белки с пониженным уровнем гликозилирования.
В предпочтительном варианте реализации векторы для инактивации согласно настоящему изобретению включают нуклеотидную последовательность, кодирующую протеин-маннозилтрансферазу (РМТ, долихил-фосфат-манноза-протеин маннозилтрансферазу, ЕС 2.4.1.109) или ее функциональную часть, и способны нарушать или инактивировать протеин-маннозилтрансферазу или ее функциональную часть в штаммах метилотрофных дрожжей. Предпочтительными нуклеотидными последовательностями являются нуклеотидные последовательности, кодирующие гены РМТ1, РМТ2, РМТ4, РМТ5, РМТ6, как представлено в SEQ ID NO:1, SEQ ID NO:2, SEQ ID NO:3, SEQ ID NO:4, SEQ ID NO:5, и их функциональные части, выбранные для дизрупции генов, как показано в SEQ ID NO:6, SEQ ID NO:7, SEQ ID NO:8, SEQ ID NO:9, SEQ ID NO:10, соответственно.
В соответствии со способами, изложенными в настоящем описании, нуклеотидная последовательность, способная экспрессировать гетерологичный белок, может быть введена в штамм метилотрофных дрожжей, который предварительно был трансформирован с помощью одного или более из векторов настоящего изобретения поэтапным способом. Такой штамм дрожжей может быть трансформирован, последовательно или одновременно, одним или более вектором настоящего изобретения. Кроме того, штамм метилотрофных дрожжей может быть трансформирован одним или более вектором для инактивации согласно настоящему изобретению, включающим нуклеотидную последовательность, кодирующую протеин-маннозилтрансферазу, как указано в списке последовательностей.
Также настоящее изобретение предусматривает штаммы метилотрофных дрожжей, созданные с помощью способов и векторов настоящего изобретения, а также белки, полученные в таких генетически модифицированных штаммах.
Гетерологичные белковые продукты, полученные с помощью способов настоящего изобретения, т.е. белки со сниженной степенью O-гликозилирования, также являются частью настоящего изобретения.
Согласно наиболее значимым аспектам настоящего изобретения, эффективность продукции желаемого конечного продукта остается неизменной.
Дополнительные цели и преимущества настоящего изобретения частично изложены в описании, которое следует ниже, и частично могут быть очевидны из описания или могут быть понятны из практического осуществления настоящего изобретения. Цели и преимущества могут быть реализованы и достигнуты с помощью технических средств и комбинаций, которые особым образом указаны в прилагаемой формуле настоящего изобретения.
Прилагаемые графические материалы, которые включены в настоящую заявку и которые составляют часть настоящей заявки, иллюстрируют различные характерные признаки, применяемые в настоящем описании, и вместе с описанием служат для объяснения различных значимых характерных признаков, формирующих суть настоящего изобретения.
КРАТКОЕ ОПИСАНИЕ ПРИЛАГАЕМЫХ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
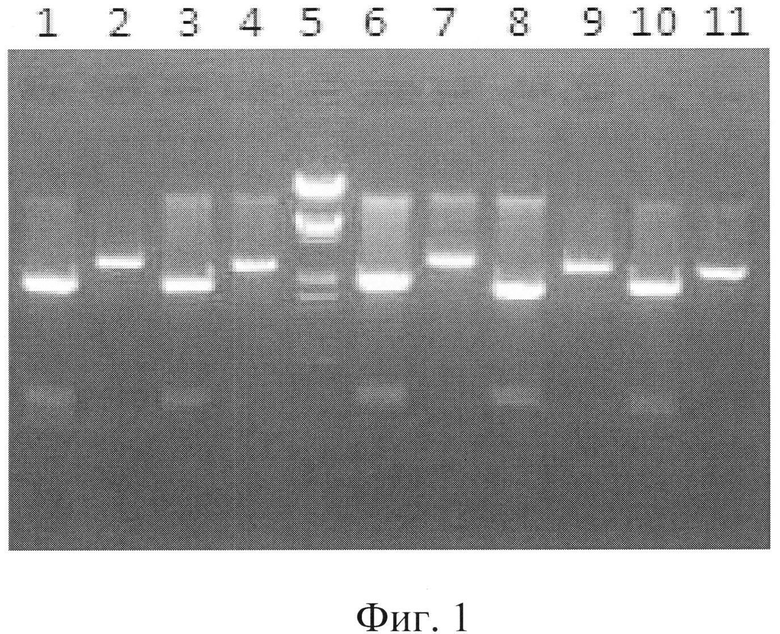
Фигура 1: Клоны, полученные при субклонировании генов РМТ в pPICZ-альфа.
Дорожка 1 = плазмида PMBL184, расщепленная с помощью рестриктаз BamHI и SmaI (фрагменты 1775+576 п.о.),
Дорожка 2 = плазмида PMBL184, расщепленная с помощью рестриктазы BstEII (линеаризованный фрагмент 2351 п.о.),
Дорожка 3 = плазмида PMBL185, расщепленная с помощью рестриктаз BamHI и SmaI (фрагменты 1930+431 п.о.).
Дорожка 4 = плазмида PMBL185, расщепленная с помощью рестриктазы KpnI (линеаризованный фрагмент 2361 п.о.),
Дорожка 5 = Маркер молекулярного веса, ДНК А, рестрицированная с помощью EcoRI и HindIII.
Дорожка 6 = плазмида PMBL186, расщепленная с помощью рестриктаз BamHI и SmaI (фрагменты 1923+428 п.о.),
Дорожка 7 = плазмида PMBL186, расщепленная с помощью рестриктазы XmnI (линеаризованный фрагмент 2351 п.о.),
Дорожка 8 = плазмида PMBL187, расщепленная с помощью рестриктаз BamHI и SmaI (фрагменты 1929+437 п.о.),
Дорожка 9 = плазмида PMBL187, расщепленная с помощью рестриктазы BstEII (линеаризованный фрагмент 2366 п.о.),
Дорожка 10 = плазмида PMBL188, расщепленная с помощью рестриктаз BamHI и SmaI (фрагменты 1929+497 п.о.) и
Дорожка 11 = плазмида PMBL188, расщепленная с помощью рестриктазы NdeI (линеаризованный фрагмент 2426 п.о.).
Фигура 2А: pMBL184 - дизрупция гена РМТ1, клонированного в векторе pPICZ-альфа. Для линеаризации плазмиды использовали сайт рестрикции BstEII.
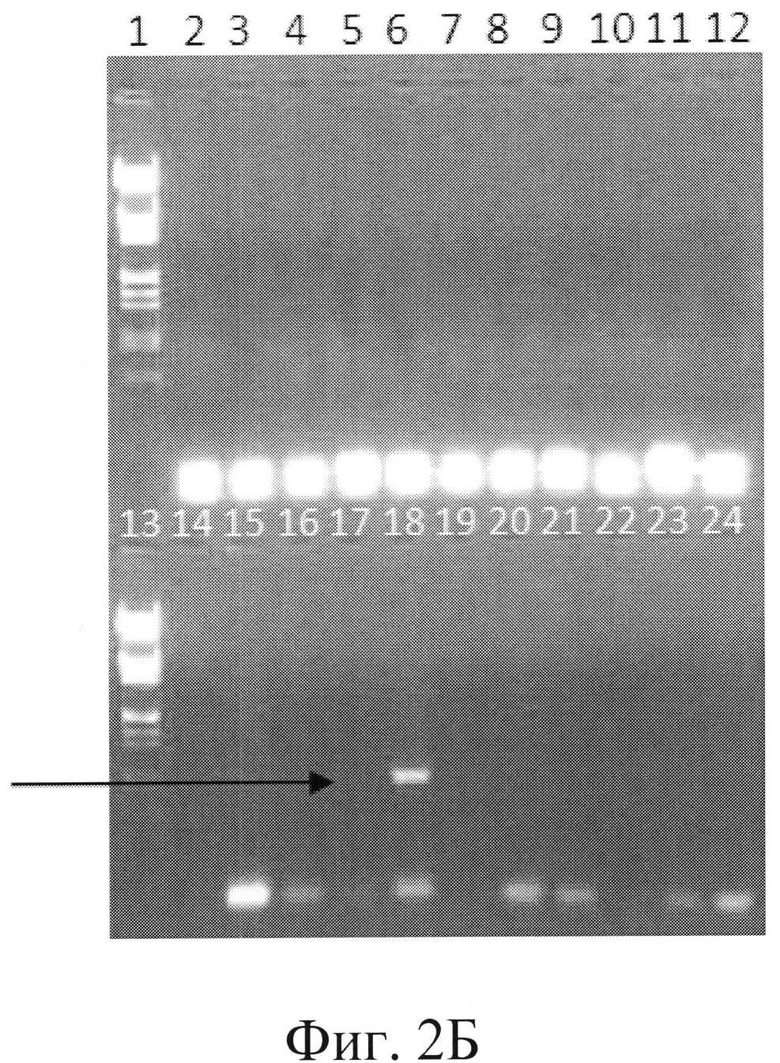
Фигура 2Б: нокаут В1СС#9104 с дизрупцией гена РМТ1, дорожка 1 и 13=маркер молекулярного веса, А, дорожка 18=BICC#9104.
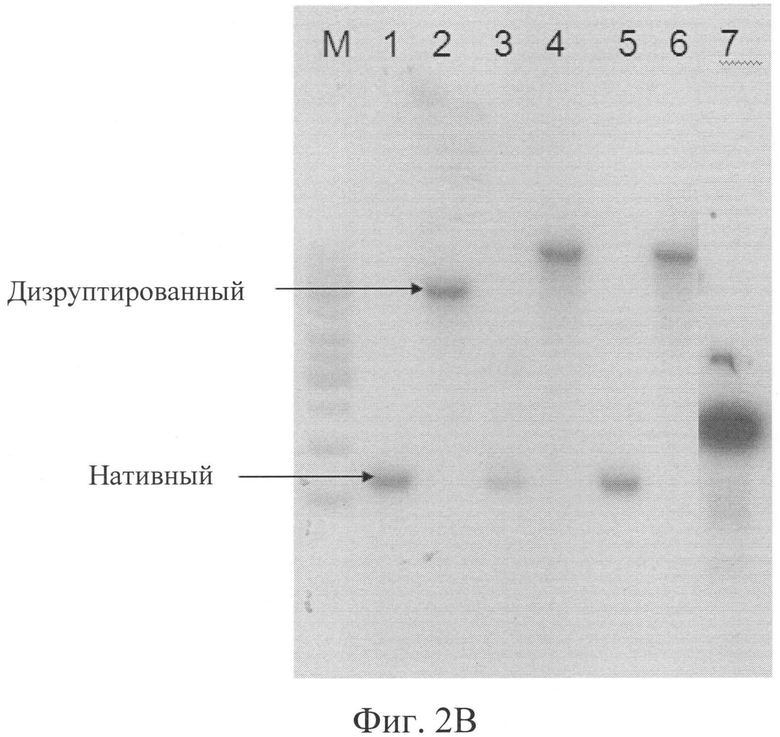
Фигура 2(В): Саузерн-блот-анализ геномной ДНК, рестрицированной с помощью рестриктазы Hind III, с использованием в качестве зонда фрагмента для дизрупции РМТ1.
Дорожка М = Маркер молекулярного веса ДНК, 1 тыс.п.о.,
Дорожка 1 = Продуцирующий предшественник инсулина родительский клон №11,
Дорожка 2 = Нокаутированный по гену РМТ1 продуцирующий предшественник инсулина клон №11,
Дорожка 3 = Продуцирующий предшественник инсулина родительский клон №8,
Дорожка 4 = Нокаутированный по гену РМТ1 продуцирующий предшественник инсулина клон №8,
Дорожка 5 = Продуцирующий предшественник аналога инсулина родительский клон,
Дорожка 6 = Нокаутированный по гену РМТ1 продуцирующий предшественник аналога инсулина клон и контрольная плазмида.
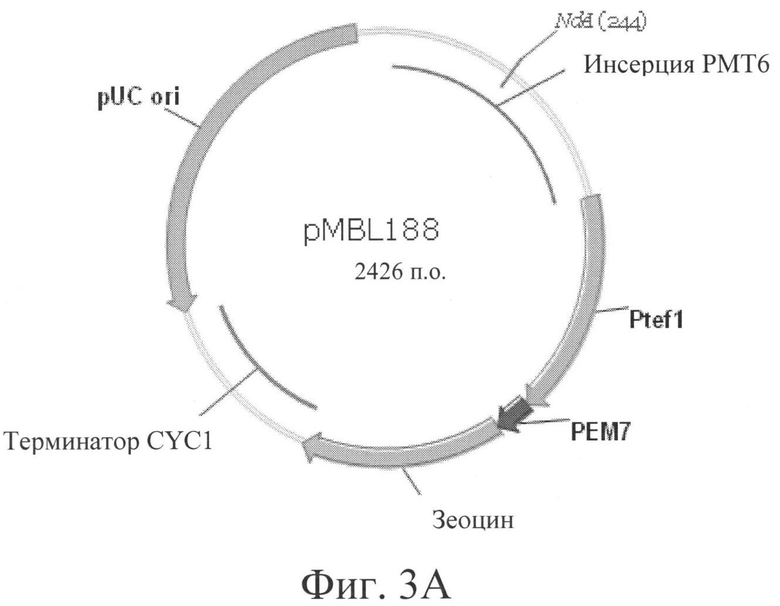
Фигура 3А: pMBL188 - дизрупция гена РМТ6 гена, клонированного в векторе pPICZ-альфа. Для линеаризации плазмиды использовали сайт рестрикции NdeI.
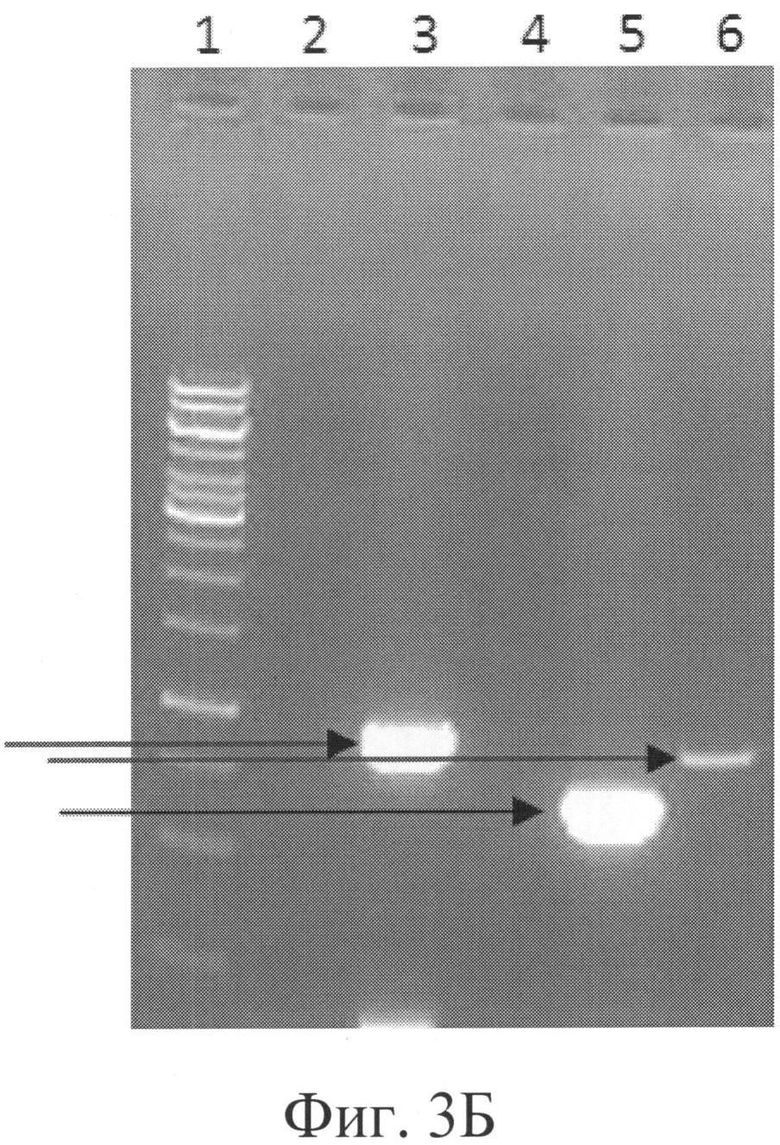
Фигура 3Б: Результаты ПЦР, подтверждающие нокаут гена РМТ6.
Дорожка 1 = Маркер молекулярного веса ДНК, лестница 1 тыс.п.о.,
Дорожка 2 = ПЦР родительского клона с InsteZRP и PMT6DSCHK (нет продукта),
Дорожка 3 = ПЦР клона В1СС#9107 с InsteZRP и PMT6DSCHK (продукт 895 п.о.),
Дорожка 4 = ПЦР родительского клона с TEFDSRP и SPMT6DCFP (нет продукта),
Дорожка 5 = ПЦР клона В1СС#9107 с TEFDSRP и SPMT6DCFP (продукт 639 п.о.),
Дорожка 6 = ПЦР клона В1СС#9107 с ISCHKFP и PMT6DSCHK (продукт 835 п.о.).
Фигура 4А: pMBL186 - дизрупция гена РМТ4 нарушения гена, клонированного в плазмиде pPICZ-альфа. Для линеаризации плазмиды использовали сайт рестрикции XcmI.
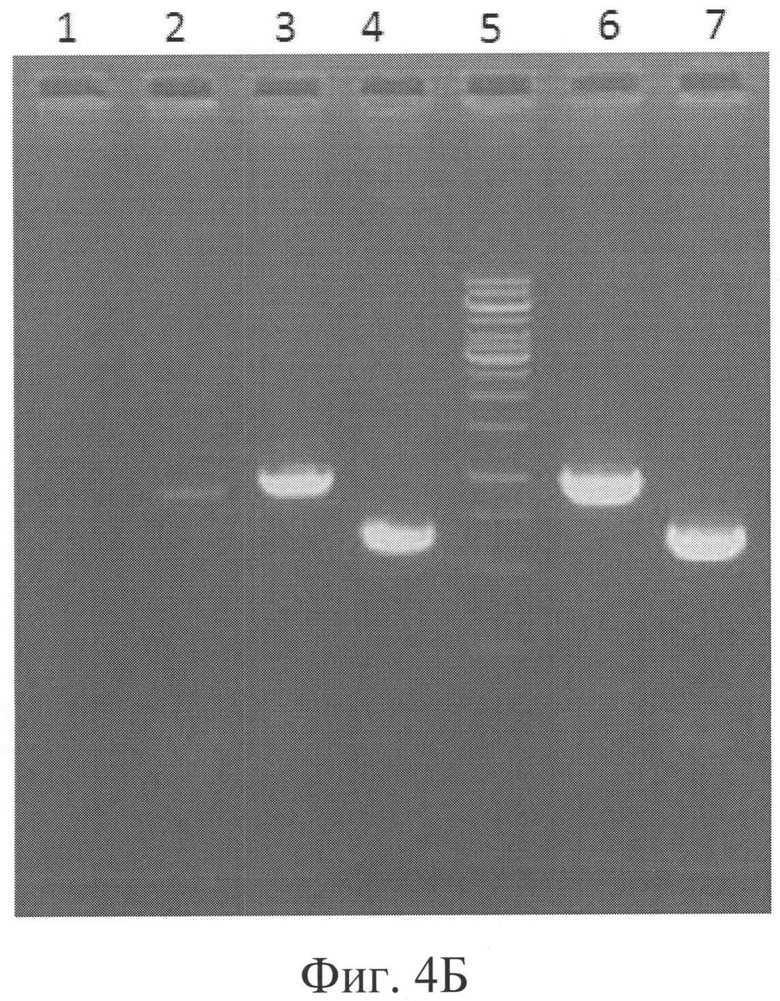
Фигура 4Б: Результаты ПЦР, подтверждающие дизрупцию гена РМТ4.
Дорожка 1 = ПЦР родительского клона с InsteZRP и PMT4DSCHK (нет продукта, контроль),
Дорожка 2 = Положительный контроль РМТ6#79 с InsteZRP и PMT6DSCHK (продукт 895 п.о.),
Дорожка 3 = ПЦР клона В1СС#9105 с InsteZRP и PMT4DSCHK (продукт 981 п.о.) (4-я субкультура),
Дорожка 4=ПЦР клона В1СС#9105 с TEFDSRP и SPMT4DCFP (продукт 649 п.о.) (4-я субкультура),
Дорожка 5=ДНК-маркер, лестница 1 тыс.п.о.,
Дорожка 6 = ПЦР клона В1СС#9105 с InsteZRP и PMT4DSCHK (продукт 981 п.о.) (1-я субкультура), Дорожка 7=ПЦР В1СС#9105 с TEFDSRP и SPMT4DRP (продукт 649 п.о.) (1-я субкультура).
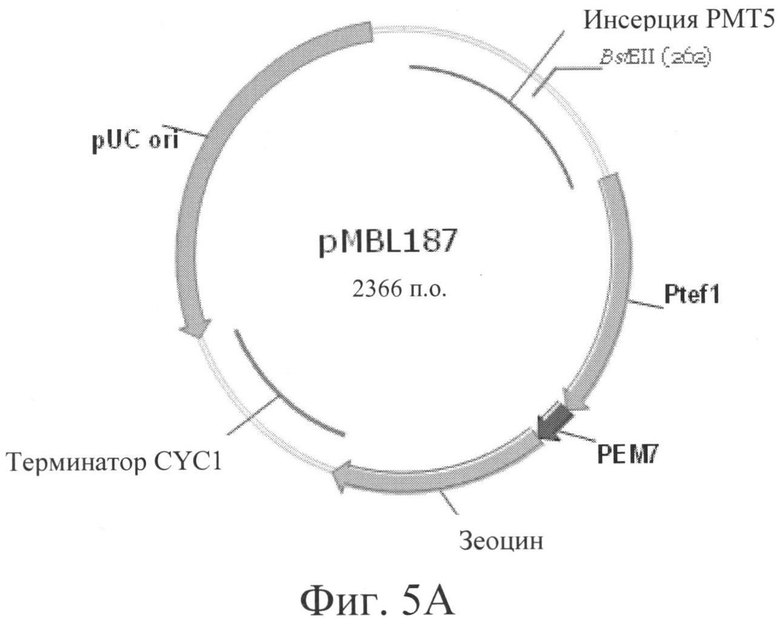
Фигура 5А: pMBL-187 - дизрупция гена РМТ5, клонированного в плазмиде pPICZ-альфа. Для линеаризации плазмиды использовали сайт рестрикции BstEII.
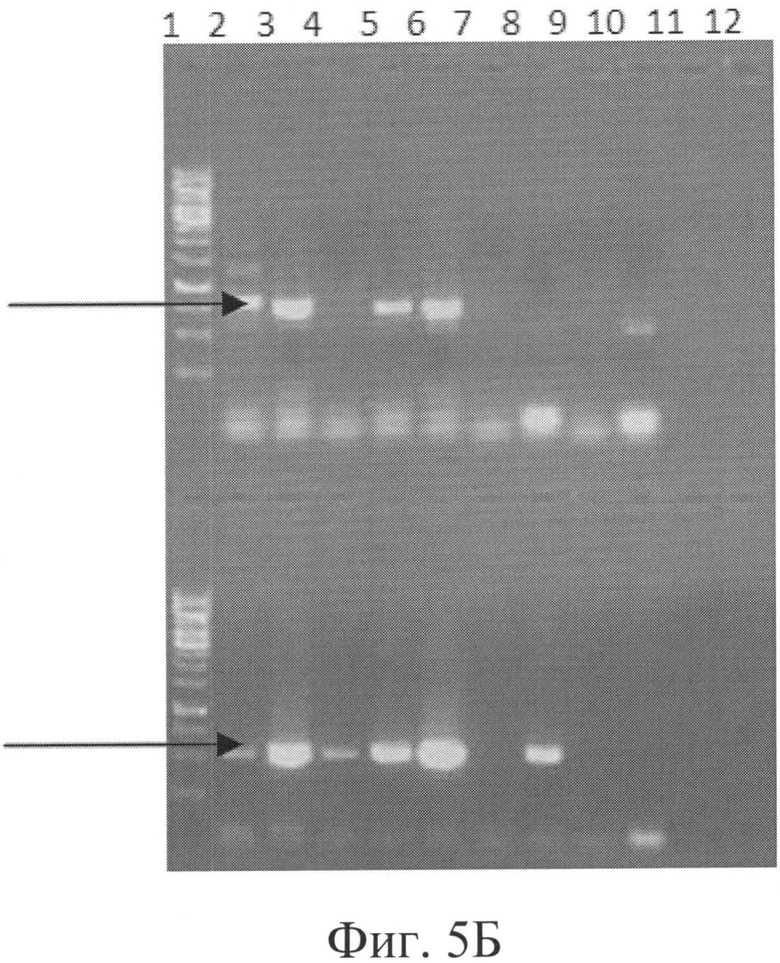
Фигура 5Б: ПЦР-скрининг клонов с дизрупцией РМТ5.
Дорожка 1 и 13 = ДНК-маркер молекулярного веса,
Дорожка 2-12 и 14-21 = скрининг разных клонов с дизрупцией РМТ5.
Фигура 6А: pMBL185 - дизрупция гена РМТ2, клонированного в плазмиде pPICZ-альфа. Для линеаризации плазмиды использовали сайт рестрикции KpnI.
Фигура 6Б: Положительный скрининг клонов с дизрупцией гена РМТ2.
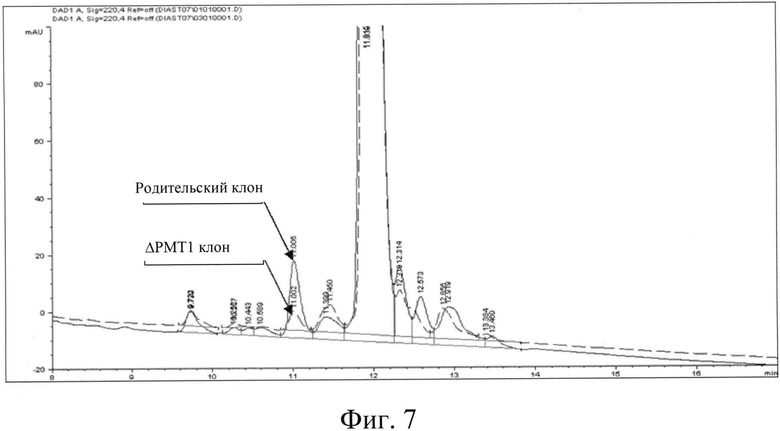
Фигура 7: Наложение хроматограмм указывает на сниженную степень гликозилирования инсулина/клон РМТ1.
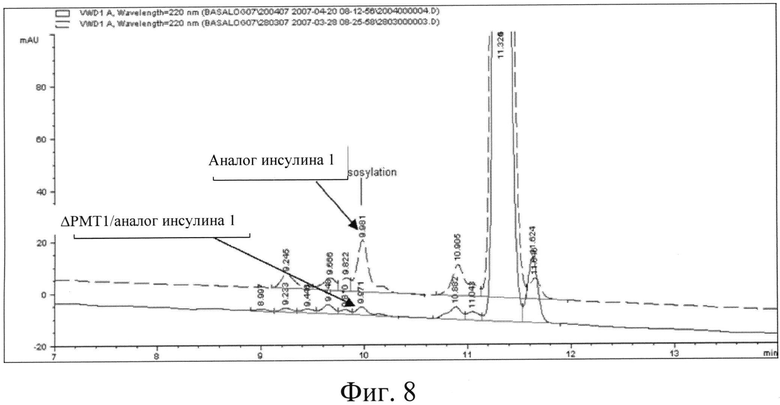
Фигура 8: Профиль ОФ-ВЭЖХ, указывающий на профиль гликозилирования аналога инсулина 1 до и после инактивации гена РМТ1.
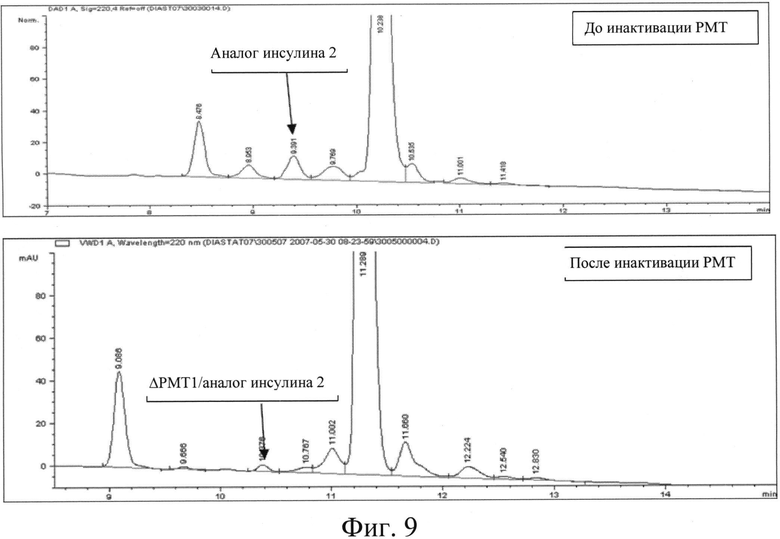
Фигура 9: Профиль ОФ-ВЭЖХ, указывающий на профиль гликозилирования аналога инсулина 2 до и после инактивации гена РМТ1.
ОПИСАНИЕ ПРИЛАГАЕМОГО ПЕРЕЧНЯ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
SEQ ID NO:1: нуклеотидная кодирующая последовательность РМТ1
SEQ ID NO:2: нуклеотидная кодирующая последовательность РМТ2
SEQ ID NO:3: нуклеотидная кодирующая последовательность РМТ4
SEQ ID NO:4: нуклеотидная кодирующая последовательность РМТ5
SEQ ID NO:5: нуклеотидная кодирующая последовательность РМТ6
SEQ ID NO:6: Последовательность с дизрупцией РМТ1
SEQ ID NO:7: Последовательность с дизрупцией РМТ2
SEQ ID NO:8: Последовательность с дизрупцией РМТ4
SEQ ID NO:9: Последовательность с дизрупцией РМТ5
SEQ ID NO:10: Последовательность с дизрупцией РМТ6'
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к способу снижения гликозилирования белка, продуцируемого в клетках метилотрофных дрожжей, что обеспечивается инактивацией по меньшей мере одного или более гена, выбранного из группы, включающей гены РМТ1, РМТ2, РМТ4, РМТ5 и РМТ6, имеющего нуклеотидную последовательность, которая по меньшей мере на 80% гомологична нуклеотидной последовательности, представленной в SEQ ID NOs 1, 2, 3, 4 и 5, соответственно, при этом указанная последовательность кодирует протеин-маннозилтрансферазу или ее функциональную часть.
В одном из вариантов реализации настоящего изобретения метилотрофные дрожжи принадлежат к роду Pichia sp.В другом варианте реализации настоящего изобретения метилотрофные дрожжи относятся к виду Pichia pastoris.
В еще одном варианте реализации настоящего изобретения, белок представлен формулой
X-B-Y-A,
где
Х представляет собой последовательность лидерного пептида, содержащую по меньшей мере одну аминокислоту.
В представляет собой аминокислотной последовательностью В-цепи молекулы инсулина, его производного или аналога
Y представляет собой линкерный пептид, содержащий по меньшей мере одну аминокислоту.
А представляет собой аминокислотную последовательность А-цепи молекулы инсулина, его производного или аналога,
и А- и В-цепи могут быть модифицированы в результате замены, делеции и/или добавления аминокислоты.
В другом варианте реализации настоящего изобретения способ гликозилирования представляет собой 0-гликозилирование.
В еще одном варианте реализации настоящего изобретения, степень гликозилирования снижается по меньшей мере на 10% до примерно 99%.
В еще одном варианте реализации настоящего изобретения степень гликозилирования снижается на 25%.
В еще одном варианте реализации настоящего изобретения степень гликозилирования снижается на 65%.
Настоящее изобретение относится к вектору, содержащему ген протеин-маннозилтрансферазы или его функциональную часть, выбранный из группы, включающей гены РМТ1, РМТ2, РМТ4, РМТ5 и РМТ6, имеющий нуклеотидную последовательность, которая по меньшей мере на 80% гомологична нуклеотидной последовательности, представленной в SEQ ID NOs 1, 2, 3, 4 и 5, соответственно, при этом встраивание вектора в гомологичный локус ингибирует экспрессию функциональной протеин-маннозилтрансферазы в клетке-хозяине, предпочтительно клетке метилотрофных дрожжей.
В одном из вариантов реализации настоящего изобретения предусмотрен способ снижения или изменения степени гликозилирования белков, продуцированных метилотрофными дрожжами, включающий трансформацию указанных дрожжей вектором, как описано выше.
В другом варианте реализации настоящего изобретения метилотрофные дрожжи относятся к роду Pichia sp.
В еще одном варианте реализации настоящего изобретения метилотрофные дрожжи относятся к виду Pichia pastoris.
Настоящее изобретение относится к способу производства нокаутированного штамма метилотрофных дрожжей, в котором (а) вектор, включающий последовательность нуклеиновой кислоты, способную к гомологичной рекомбинации, включающую целевую последовательность нуклеиновой кислоты, кодирующую по меньшей мере один из генов, выбранных из группы, включающей гены РМТ1, РМТ2, РМТ4, РМТ5 и РМТ6, имеющие нуклеотидную последовательность, которая по меньшей мере на 80% гомологична нуклеотидной последовательности, представленной в SEQ ID NOs 1, 2, 3, 4 и 5 соответственно, и последовательность нуклеиновой кислоты, кодирующую селективный маркер, (б) культивирование клеток при условиях, при которых возможно осуществление гомологичной рекомбинации между ДНК, кодирующих целевой ген в векторе и в клетке-хозяине, что, таким образом, приводит к нарушению целевого гена в клетке-хозяине, (в) селекцию клеток-хозяев с инактивированным целевым геном. Настоящее изобретение также относится к белку, продуцируемому с помощью способа по любому из указанных предыдущих пунктов.
Настоящее изобретение также относится к нокаутированному штамму метилотрофных дрожжей, при этом такой штамм содержит по меньшей мере один инактивированный ген, выбранный из группы, включающей РМТ1, РМТ2, РМТ4, РМТ5 и РМТ6, имеющий нуклеотидную последовательность, которая по меньшей мере на 80% гомологична нуклеотидной последовательности, представленной в SEQ ID NOs 1, 2, 3, 4 и 5 соответственно.
Настоящее изобретение относится к штамму с инактивированным геном РМТ1, как описано выше, имеющим номер доступа МТСС5515.
Настоящее изобретение относится к штамму с инактивированным геном РМТ4, как описано выше, имеющим номер доступа МТСС5516.
Настоящее изобретение относится к штамму с инактивированным геном РМТ5, как описано выше, имеющим номер доступа МТСС5517.
Настоящее изобретение также относится к штамму с инактивированным геном РМТ6, как описано выше, имеющим номер доступа МТСС5518.
В одном из вариантов реализации настоящего изобретения штамм-хозяин продуцирует белок с пониженным уровнем гликозилирования, по сравнению с белковым продуктом, экспрессируемым в немодифицированном штамме-хозяине.
В еще одном варианте реализации настоящего изобретения степень гликозилирования снижается по меньшей мере на 25%.
В еще одном варианте реализации настоящего изобретения степень гликозилирования снижается по меньшей мере на 65%.
Настоящее изобретение относится к белку, продуцируемому нокаутированным штаммом, как описано выше.
Настоящее изобретение относится к белку, продуцируемому нокаутированным штаммом, в соответствии с п.23, при этом указанный нокаутированный штамм является одним из штаммов с номерами доступа МТСС5515, МТСС5516, МТСС5517, МТСС5518 или модифицированным штаммом на его основе.
В одном из вариантов настоящего изобретения селективный маркер представляет собой маркер устойчивости к зеоцину.
В другом варианте реализации настоящего изобретения белок представляет собой молекулу инсулина/аналога инсулина/молекулу предшественника инсулина.
В еще одном варианте реализации настоящего изобретения белок представляет собой гетерологичный белковый продукт.
В еще одном варианте реализации настоящего изобретения белок характеризуется изменением степени гликозилирования. В еще одном варианте реализации настоящего изобретения производительность желаемого конечного белкового продукта остается неизменной.
Далее будут подробно описаны предпочтительные варианты реализации настоящего изобретения, которые вместе со следующими ниже примерами служат для объяснения принципов настоящего изобретения.
Примеры, которые следуют ниже, приведены для облегчения понимания настоящего изобретения, но они ни в коей мере не ограничивают объем настоящего изобретения. Примеры не включают подробные описания стандартных методов, применяемых при конструировании векторов, встраивании генов, кодирующих полипептиды, в такие векторы или введении образующихся в результате плазмид в клетку-хозяина. Примеры также не включают в себя подробное описание стандартных методов, применяемых для анализа полипептидов, полученных с помощью таких векторных систем хозяина. Такие методы хорошо известны специалистам в данной области и описаны в многочисленных публикациях, некоторые из которых приведены в качестве примеров.
Стандартные методы применяются для различных технологий рекомбинантной ДНК, трансформации (например, электропорация, липофекция) и систем анализа. Методы и процедуры рекомбинации, как правило, производят в соответствии со стандартными методами, хорошо известными в данной области, и описанными в различных общих и более частных источниках, которые процитированы и обсуждены в настоящем описании. См., например, справочник Sambrook et al. Molecular Cloning: A Laboratory Manual (3rd ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (2001)), включенный в настоящую заявку посредством ссылки.
В описании и в формуле настоящего изобретения используется следующая терминология в соответствии с определениями, изложенными в настоящей заявке.
Если не определено иначе в настоящем документе, научные и технические термины, используемые в связи с настоящим изобретением, имеют значения, которые обычно понимают специалисты в данной области. Кроме того, если контекстом не предусмотрено иное, термины, употребляемые в единственном числе, включают значения во множественном числе, и термины, используемые во множественном числе, включают значения в единственном числе. Способы и методики настоящего изобретения, как правило, осуществляют в соответствии со стандартными способами, хорошо известными в данной области. В целом, используемая терминология, а также методы молекулярной и клеточной биологии, биохимии, химии белков и нуклеиновых кислот и методы гибридизации, описанные в настоящей заявке, являются хорошо известными и широко применимыми в данной области. Способы и технологии настоящего изобретения, как правило, осуществляют в соответствии со стандартными методами, хорошо известными в данной области.
Используемый в настоящей заявке термин «аминокислота» относится к пептидной или белковой последовательности или ее части. Термины «белок», «пептид» и «полипептид» используются как синонимы.
Гетерологичный белок в соответствии с предпочтительными вариантами реализации настоящего изобретения представляет собой молекулу предшественника инсулина или аналога инсулина.
ДНК кодирует гетерологичный белок, представленный формулой I
X-B-Y-A,
где
Х представляет собой последовательность лидерного пептида, содержащую по меньшей мере одну аминокислоту.
В представляет собой аминокислотную последовательность В-цепи молекулы инсулина, его производного или аналога,
Y представляет собой линкерный пептид, содержащий по меньшей мере одну аминокислоту.
А представляет собой аминокислотную последовательность А-цепи молекулы инсулина, его производного или аналога, и А- и В-цепи могут быть модифицированы в результате замены, делеции и/или добавления аминокислоты.
Термин «С-пептид» или «линкерный пептид», используемый в настоящей заявке, включает в себя все формы инсулинового С-пептида, в том числе нативного или синтетического пептида. Такой инсулиновый С-пептид может представлять собой человеческий пептид, или может происходить от других видов и родов животных, предпочтительно от млекопитающего.
Таким образом, также охвачены варианты и модификации нативного инсулинового С-пептида при условии, что они сохраняют активность инсулинового С-пептида. Как известно в данной области, модификация последовательности белков или пептидов, при сохранении их необходимой активности, может быть осуществлена с помощью методов, которые являются стандартными в данной области и широко описаны в литературе, например, случайного или сайт-направленного мутагенеза, расщепления и лигирования нуклеиновых кислот и т.д. Таким образом, функционально эквивалентные варианты или производные последовательности нативного инсулинового С-пептида можно легко получить в соответствии со способами, хорошо известными в данной области, и они включают пептидные последовательности, имеющие функциональную, например, биологическую, активность нативного инсулинового С-пептида. Все такие аналоги, варианты, производные или фрагменты инсулинового С-пептида, в частности, включены в объем настоящего изобретения и входят в понятие термина «инсулиновый С-пептид».
Линкерная последовательность может представлять собой любую последовательность, имеющую по меньшей мере две аминокислоты. Линкерный участок может включать от 2 до 25, от 2 до 15, от 2 до 12 или от 2 до 10 аминокислотных остатков, хотя его длина не является критичной и может быть выбрана для удобства или по желанию, или же может отсутствовать.
Линкерный пептид может представлять собой любую последовательность, включающую по меньшей мере две аминокислоты, при условии, что первые две аминокислоты представляют собой «RR».
В соответствии с другими вариантами реализации настоящего изобретения полипептиды, упоминаемые в настоящей заявке, могут обладать активностью инсулина гларгина, например, представляют собой инсулин гларгин, который имеет аминокислотную последовательность с двумя изменениями в структуре человеческого инсулина: заменой на аминокислоту глицин нативного аспарагина в положении А21 А-цепи человеческого инсулина и добавлением двух молекул аргинина с СООН-конца В-цепи человеческого инсулина, и такие изменения произведены с помощью технологии рекомбинантной ДНК.
Первичным действием любого инсулина, в том числе инсулина гларгина, является регуляция метаболизма глюкозы. Инсулин и его аналоги снижают уровень глюкозы в крови путем стимуляции периферического поглощения глюкозы, в частности, в скелетных мышцах и жировой ткани, а также путем ингибирования образования глюкозы в печени.
Термин «инсерционная инактивация» обозначает прерывание кодирующей области гена путем введения экзогенной ДНК, что приводит к потере функции гена. Такой подход широко используется в генной технологии и позволяет легко осуществить селекцию рекомбинантов после трансформации.
Термин «нокаут» относится к дизрупции гена, и при этом такая дизрупция приводит к функциональной инактивации нативного гена; делеции нативного гена или его части, или мутации в нативном гене. В частности, в отношении настоящего изобретения «нокаут» относится к дизрупции генов РМТ1, РМТ2, РМТ4, РМТ5, РМТ6.
Нокаутированный штамм может быть получен в соответствии с любым из различных способов, известных в данной области. Например, клетка-хозяин может быть трансформирована вектором, обуславливающим гомологичную рекомбинацию, содержащим гомологичные целевому гену последовательности гена в направлении 5' и 3' от последовательности, которую необходимо делегировать. В идеале, при гомологичной рекомбинации может быть осуществлен нокаут целевого гена, кодирующего фермент.
«Гомологичная рекомбинация» обозначает обмен фрагментами ДНК между двумя молекулами ДНК пары хромосом в области идентичных или почти идентичных нуклеотидных последовательностей. При гомологичной рекомбинации входящая ДНК взаимодействует с и встраивается в сайт в геноме, который содержит по существу гомологичную последовательность ДНК. При негомологичном («случайном» или «незаконном») встраивании входящая ДНК встраивается не в гомологичную последовательность в геноме, а в другом месте, в одном из множества потенциальных сайтов.
Например, мутантные, функциональные или нефункциональные гены, фланкированные ДНК, гомологичной эндогенному гену-мишени (например, кодирующие области, фланкирующие ген-мишень) можно применять совместно или без селективного маркера и/или отрицательного селективного маркера, для трансформации клеток, содержащих нежелательную форму гена-мишени in vivo. Встраивание конструкции ДНК посредством целевой гомологичной рекомбинации приводит к инактивации эндогенного гена.
При использовании в настоящей заявке термин «гомологичный» обозначает либо I) белок или пептид, который имеет последовательность аминокислот, которая является по существу схожей (то есть, по меньшей мере на 70, 75, 80, 85, 90, 95 или 98%) с последовательностью данного исходного белка или пептида и который сохраняет нужную функцию исходного белка или пептида, либо II) нуклеиновую кислоту, имеющую последовательность, которая является по существу схожей (то есть, по меньшей мере на 70, 75, 80, 75, 90, 95 или 98%) с последовательностью данной нуклеиновой кислоты и которая сохраняет нужную функцию исходной последовательности нуклеиновой кислоты. Во всех вариантах реализации настоящего изобретения любой описанный белок, пептид или любая нуклеиновая кислота может быть замещена гомологичным или по существу гомологичным белком, пептидом или нуклеиновой кислотой, которая сохраняет необходимую функцию. Во всех вариантах реализации настоящего изобретения при описании (раскрытии) любой нуклеиновой кислоты следует предположить, что описание также включает в себя все нуклеиновые кислоты, которые гибридизуются с описанной нуклеиновой кислотой.
Под «функциональной частью» понимают фрагмент гена РМТ, соответствующий белок которого по существу сохраняет ферментативную активность полноразмерного белка. «По существу» означает, что сохраняется по меньшей мере около 40% или предпочтительно по меньшей мере 50% или более ферментативной активности полноразмерного РМТ.
Нуклеиновые кислоты «функционально связаны», когда они находятся в функциональной связи с другой последовательностью нуклеиновой кислоты. Таким образом, когда говорят, что кодирующая последовательность «функционально связана» с контролирующими последовательностями, это относится к конфигурации, в которой кодирующая последовательность может экспрессироваться под контролем этих последовательностей, и при этом последовательности ДНК прилегают друг к другу.
При использовании в настоящей заявке термин «экспрессия» относится к процессу, при котором на основе нуклеиновой кислоты гена образуется полипептид. Этот процесс включает в себя как транскрипцию, так и трансляцию. В одном аспекте, изолированная молекула нуклеиновой кислоты содержит нуклеотидную последовательность, имеющую по меньшей мере около 80% идентичности последовательности нуклеиновой кислоты, как альтернатива, по меньшей мере около 81% идентичности последовательности нуклеиновой кислоты, как альтернатива, по меньшей мере около 82% идентичности последовательности нуклеиновой кислоты, как альтернатива, по меньшей мере около 83% идентичности последовательности нуклеиновой кислоты, как альтернатива, по меньшей мере около 84% идентичностью последовательности нуклеиновой кислоты, альтернативно по меньшей мере около 85% идентичностью последовательности нуклеиновой кислоты, альтернативно по меньшей мере около 86% идентичностью последовательности нуклеиновой кислоты, как альтернатива, по меньшей мере около 87% нуклеиновых кислот идентичности последовательностей, как альтернатива, по меньшей мере около 88% идентичности последовательности нуклеиновой кислоты, как альтернатива, по меньшей мере около 89% идентичности последовательности нуклеиновой кислоты, как альтернатива, по меньшей мере около 90% идентичности последовательности нуклеиновой кислоты, как альтернатива, по меньшей мере около 91% идентичности последовательности нуклеиновой кислоты, как альтернатива, по меньшей мере около 92% идентичности последовательности нуклеиновой кислоты, как альтернатива, по меньшей мере около 93% идентичности последовательности нуклеиновой кислоты, как альтернатива, по меньшей мере около 94% идентичности последовательности нуклеиновой кислоты, как альтернатива, по меньшей мере около 95% идентичности последовательности нуклеиновой кислоты, как альтернатива, по меньшей мере около 96% идентичности последовательности нуклеиновой кислоты, как альтернатива, по меньшей мере около 97% идентичности последовательности нуклеиновой кислоты, как альтернатива, по меньшей мере около 98% идентичности последовательности нуклеиновой кислоты и как альтернатива, по меньшей мере около 99% идентичности последовательности нуклеиновой кислоты к молекуле ДНК, кодирующей гены РМТ1, РМТ2, РМТ4, РМТ5, РМТ6.
Кроме того, оптимальное выравнивание последовательностей для сравнения может быть осуществлено с помощью алгоритма локальной идентичности Смита и Уотермана, (Smith and Waterman (1981) Add. APL. Math 2:482), с помощью алгоритма выравнивания Нидлмана и Вунша (Needleman and Wunsch (1970) J. Mol. Biol. 48:443), и при помощи методов поиска сходства Пирсона и Липмана (Pearson and Lipman (1988) Proc. Natl. Acad. Sci. USA 85: 2444,), с помощью автоматизированной реализации указанных алгоритмов (BLAST, FASTA, GAP, BESTFIT и TFASTA в пакете программ Wisconsin Genetics Software Package, Genetics Computer Group (GCG), 575 Science Dr., Madison, Wis.), или путем наблюдения.
Одним из примеров алгоритмов, которые можно применять для определения процента идентичности последовательностей и сходства последовательностей являются алгоритмы BLAST и BLAST 2,0, которые описаны в работах Altschul et al. (1977) Nucl. Acids Res. 25:3389 3402 и Altschul et al. (1990) J. Mol. Biol. 215:403 410, соответственно.
В частности, настоящее изобретение относится к последовательности нуклеиновой кислоты, в частности изолированной последовательности нуклеиновой кислоты, которая включает или содержит по меньшей мере SEQ ID NO:1, SEQ ID NO:2, SEQ ID NO:3, SEQ ID NO:4, SEQ ID NO:5, их варианты или их части, или по меньшей мере один из генов РМТ1, РМТ2, РМТ4, РМТ5, РМТ6, включая его варианты или части. Настоящее изобретение также относится к изолированной нуклеиновой кислоте, способной к гибридизации в жестких условиях с такими последовательностями нуклеиновых кислот.
Настоящее изобретение обеспечивает векторы, содержащие ДНК, кодирующую любой из описанных в настоящей заявке генов. Клетки-хозяева, содержащие такие векторы, также предусмотрены настоящим изобретением. Например, клетки-хозяева могут представлять собой клетки бактерий, грибов или млекопитающих.
Настоящее изобретение также относится к рекомбинантной клетке-хозяину, в которой по меньшей мере часть последовательности нуклеиновой кислоты, как определено выше, нарушается, в результате чего такая рекомбинантная клетка-хозяин характеризуется снижением уровня гликозилирования предшественника инсулина, по сравнению с соответствующей родительской рекомбинантной клеткой-хозяином. Рекомбинантные экспрессионные системы выбраны из прокариотических и эукариотических клеток-хозяев. Эукариотические хозяева включают дрожжевые клетки (например, Saccharomyces cerevisiae или Pichia pastoris), клетки млекопитающих или клетки растений. Бактериальные и эукариотические клетки можно получить из ряда различных источников, включая коммерческие источники, известные специалистам в данной области, например, из Американской коллекции типовых культур (American Type Culture Collection, ATCC, Rockville, Md.). Коммерческие источники клеток, применяемых для экспрессии рекомбинантного белка, также предоставляют инструкции по применению таких клеток. Выбор экспрессионной системы зависит от желаемых характеристик экспрессируемого полипептида.
Наиболее предпочтительными в отношении аспектов настоящего изобретения клетками-хозяевами являются метилотрофные дрожжи. Штаммы метилотрофных дрожжей, которые могут быть модифицированы с помощью настоящего изобретения, включают, но не ограничиваются перечисленными, штаммы дрожжей, способные расти на метаноле, такие как дрожжи родов Pichia, Candida, Hansenula или Torulopsis. Предпочтительные метилотрофные дрожжи относятся к роду Pichia. Штаммы метилотрофных дрожжей, которые могут быть модифицированы с помощью способов настоящего изобретения, также включают штаммы метилотрофных дрожжей, модифицированные с помощью генно-инженерного подхода для экспрессии одного или более целевого гетерологичного белка. Степень гликозилирования гетерологичных белков, экспрессирующихся в таких модифицированных генно-инженерными методами штаммах, может быть снижена путем трансформации таких штаммов с помощью одного или более вектора настоящего изобретения.
Клетки-хозяева или организм может быть модифицирован генно-инженерным способом для экспрессии рекомбинантного белка или пептида с применением стандартных способов. Например, рекомбинантный белок может экспрессироваться с вектора или экзогенного гена, встроенного в геном хозяина.
Векторы, которые можно применять для экспрессии экзогенных белков, хорошо известны в данной области и описаны ниже. Гены для экспрессии рекомбинантных белков или пептидов также могут быть встроены в геном с применением таких подходов, как гомологичная и гетерологичная рекомбинация, как описано в настоящей заявке. Предпочтительные векторы настоящего изобретения, содержащие гены протеин-маннозилтрансферазы, включают, но неограничены pPICZ альфа и pTZ57R.
В другом аспекте настоящее изобретение обеспечивает векторы для инактивации, которые при встраивании в дрожжевой метилотрофный штамм, инактивируют или повреждают (дизруптируют) ген, что, таким образом, способствует снижению степени гликозилирования целевого конечного белкового продукта, синтезируемого в дрожжевом метилотрофном штамме, при этом не оказывая влияния на производительность конечного продукта.
Рекомбинантный белок или пептид может экспрессироваться после индукции химическим соединением или под действием экспрессии эндогенного гена или продукта гена. Рекомбинантный белок может также экспрессироваться в том случае, когда хозяйская клетка находится на определенной среде. Специфические промоторные элементы описаны ниже.
При использовании в настоящей заявке термины «трансформированный» и «стабильно трансформированный» относятся к клеткам, которые были созданы путем введения ненативных (гетерологичных) полинуклеотидных последовательностей, интегрированных в эписомальные плазмиды, которые поддерживаются на протяжении не менее двух поколений.
При использовании в настоящей заявке термин «рекомбинантный» относится к клетке или вектору, которые были модифицированы с помощью введения гетерологичной нуклеотидной последовательности, или клетку, которая произошла от клетки, модифицированной таким образом.
Векторы могут быть введены в клетки-хозяева с помощью способов, включающих, но не ограниченных перечисленными, электропорацию, вирусную инфекцию, кальций-фосфатную преципитацию, с помощью DEAE-декстрана, направленную микроинъекцию, с помощью липосом, загруженных ДНК, и комплексов липофектамин-ДНК, ультразвуковую трансформацию, бомбардировку с применением высокоскоростных микрочастиц, или любых других способов, описанных в настоящей заявке или известных в данной области.
Настоящее изобретение также относится к способу получения желаемого белка, включающему ферментирование при условиях и на среде, подходящей для получения таких белковых соединений или их аналагов, в организме, таком как Pichia sp., в который перенесены гены, кодирующие полипептиды, достаточные для направленного синтеза желаемого конечного продукта.
Было обнаружено, что протеин-маннозилтрансферазы (РМТ) катализируют O-гликозилирование сериновых и треониновых остатков в эндоплазматическом ретикулуме. В настоящем изобретении было продемонстрировано, что дизрупция генов РТМ резко снижает уровень O-гликозилирования продуцируемых молекул предшественников инсулина. Дизрупция гена РМТ1 приводит к формированию предшественника инсулина, характеризующегося снижением присоединения маннозных остатков на 65%. Дизрупции генов РМТ5 и РМТ6 в отдельности приводят к снижению уровня гликозилирования предшественника инсулина на 31% и 28%, соответственно. Дизрупции генов РМТ2 и РМТ4 не влияют на присоединение маннозных остатков.
В настоящей заявке термин «снижение уровня экспрессии» используется в широком смысле и включает снижение продукции целевого белка. Пониженная экспрессия представляет собой такую экспрессию, уровень которой ниже уровня нормальной экспрессии в соответствующем хозяйском штамме, который не был модифицирован в соответствии со способами, описанными в настоящей заявке, но был выращен при тех же условиях. В контексте настоящего изобретения целевые ферменты или белки представляют собой маннозилтрансферазы, которые играют важную роль в гликозилировании маннозных остатков молекул предшественников инсулина/аналогов инсулина.
В соответствии с одним из аспектов настоящего изобретения снижение степени гликозилирования может составлять по меньшей мере 20%, предпочтительно по меньшей мере 25%, предпочтительно по меньшей мере 30%, предпочтительно по меньшей мере 35%, наиболее предпочтительно по меньшей мере 40%, наиболее предпочтительно по меньшей мере 45%, наиболее предпочтительно по меньшей мере 50%, наиболее предпочтительно по меньшей мере 55%, наиболее предпочтительно по меньшей мере 60%, наиболее предпочтительно по меньшей мере 65% и наиболее предпочтительно по меньшей мере 70%. В соответствии с другим аспектом настоящего изобретения снижение степени гликозилирования достигает 100%.
При использовании в настоящей заявке, термин «праймер» относится к олигонуклеотиду, либо природному, в очищенном рестрицированном виде, либо синтетическому, который способен инициировать синтез при условиях, при которых индуцируется синтез продукта удлинения праймера, комплементарного цепи нуклеотидной последовательности (например, в присутствии нуклеотидов и агента, такого как ДНК-полимераза и при подходящей температуре и рН). Методика ПЦР описана в многочисленных публикациях, в том числе в руководствах PCR: А Practical Approach, M.J. McPherson et al., IRL Press (1991), PCR Protocols: A Guide to Methods and Applications, by Innis et al., Academic Press (1990), и PCR Technology: Principals and Applications of DNA Amplification, H.A. Eriich, Stockton Press (1989).
Метод ПЦР также описан во многих патентах США, включая патенты с номерами 4, 683,195, 4,683,202; 4,800,159; 4,965,188; 4, 889,818; 5, 075, 216; 5,079, 352; 5,104,792, 5,023,171; 5,091,310; и 5, 066,584, которые включены в качестве ссылок.
В соответствии с наиболее значимыми аспектами настоящего изобретения предусмотрен способ производства «нокаутированных» штаммов метилотрофных дрожжей, включающий (а) вектор, содержащий нуклеотидную последовательность, кодирующую по меньшей мере один ген, выбранный из группы, включающей РМТ1, РМТ2, РМТ4, РМТ5 и РМТ6, как представлено в SEQ ID NOs 1, 2, 3, 4 и 5, соответственно, и нуклеотидную последовательность, кодирующую селективный маркер, (б) культивирование клеток при условиях, при которых происходит гомологичная рекомбинация, в результате которой происходит дизрупция целевой нуклеотидной последовательности в клетках-хозяевах, (в) селекцию клеток, в которых произошла гомологичная рекомбинация и (г) оценку степени снижения уровня гликозилирования в модифицированных штаммах.
Трансформацию Pichia pastoris осуществляли с помощью электропорации.
Трансформанты, содержащие вектор с геном устойчивсти к зеоцину, были отобраны на чашках с YPD, содержащих 100 мкг/мл зеоцина.
Таким образом, способы настоящего изобретения приводят к изменению дрожжевого штамма, который становится способным производить целевой белок, при этом (а) вышеупомянутый штамм содержит по меньшей мере один инактивированный ген из группы, включающей РМТ1, РМТ2, РМТ4, РМТ5 и РМТ6, как представлено в SEQ ID NOs 1, 2, 3, 4 и 5, соответственно, и (б) выращивание вышеупомянутого измененного штамма при условиях, при которых происходит снижение степени гликозилирования конечного продукта, по сравнению с белковым продуктом, продуцированным неизмененным штаммом-хозяином.
Следует понимать, что настоящее изобретение не ограничено описанными конкретными способами, протоколами, клеточными линиями, векторами, видами или родами и компонентами сред, которые могут варьироваться. Также следует понимать, что терминология используется только с целью описания конкретных вариантов реализации и не предназначена для ограничения области настоящего изобретения, которое ограничено лишь формулой настоящего изобретения. Приведенное выше описание предназначено для того, чтобы указать специалисту, как применять настоящее изобретение, и его целью не является детальное описание всех явных модификаций и вариантов настоящего изобретения, которые очевидны специалисту в данной области при прочтении настоящего описания.
Депонирование штаммов с инактивированными генами РМТ
В соответствии с требованиями полного описания изобретения штаммы настоящего изобретения были депонированы в IMTEC Института микробных технологий, сектор 3 9-А, Чандигарх, 160036, Индия (в соответствии с международным депонированием на основании Будапештского договора). Штаммы являются следующими (номера доступа указаны в скобках):
Настоящее изобретение может быть более полно описано и понято с помощью следующих примеров, которые приведены в качестве иллюстрации и ни в коей мере не предназначены для ограничения настоящего изобретения.
ПРИМЕР 1: ДИЗРУПЦИЯ РМТ1:
Около 477 п.о. кодирующей последовательности, включающей часть кодирующей области РМТ1 Pichia pastoris, амплифицировали с помощью следующих праймеров РМТ1.
РМТ1 FP=5' GGA ТСС ТАА TAG CCC ACT CTG АТС TAG CTC ACT 3'
РМТ1 RP=5' GGA ТСС ААА GCC CTC ATG ТСС АТА AGC AGA 3'.
Два стоп-кодона были включены в прямой праймер, чтобы избежать какого-либо прочтения с ранних потенциальных сайтов начала трансляции в векторе. Продукт ПЦР был клонирован в вектор pTZ57R, и для подтверждения он был секвенирован с помощью M13FP и M13RP. Фрагмент РМТ1 был вырезан при помощи BamHI и клонирован в pPICZ-альфа по сайтам BamHI и BgI II. Клон, давший фрагменты размером 1775 п.о. и 576 п.о., был отобран. Кассета для дизрупции РМТ1 была перенесена в Pichia Pastoris GS115. Плазмида была линеаризована по уникальному сайту BstEII, который находится почти в середине фрагмента для дизрупции РМТ1, перенесенного в штамм Pichia с помощью электропорации.
Трансформированные клоны были отобраны с помощью 100 мкг/мл зеоцина. Была получена пара сотен колоний, и отдельные колонии были высажены штрихом на чашки со средой YPD. Геномная ДНК была изолирована из каждого клона, и для проверки клонов с дизрупцией проводили ПЦР. Нокаут РМТ1 подтверждали с помощью ПЦР (см. фиг.2Б), а также с помощью Саузерн-блота (см. фиг 2В). Отобранные клоны были депонированы как МТСС5515. Последовательности праймеров:
InSTEzRP: 5' TAG CAG AGC GAG GTA TGT AGG CGG TGC 3'
TEFDSRP: 5' GAG ТСС GAG ААА АТС TGG AAG AGT 3'
ISCHKFP: 5' GCT АСА СТА GAA GGA CAG TAT TTG GTA 3'
SPMT1 DCFP: 5' GGA CTT ATG GTT CAT CAT TGG TGA 3'
ПРИМЕР 2: ДИЗРУПЦИЯ РМТ6:
Около -515 п.о. кодирующей последовательности, включающей часть кодирующей области РМТ6, амплифицировали с применением следующих праймеров:
PMT6FP=5' GGA ТСС ТАА TAG CTT GCC GTT AAG AGA TAC GAT GA
3' PMT6RP=5'GGA ТСС TGA GAA TGC AAG TTT GCA ССА GTA 3'
Два стоп-кодона были включены в прямой праймер, чтобы избежать какого-либо прочтения с ранних потенциальных сайтов начала трансляции в векторе. Продукт ПЦР был клонирован в вектор pTZ57R, и для подтверждения он был секвенирован с помощью M13FP и M13RP. Кассета для дизрупции РМТ6 была перенесена в Pichia Pastoris GS115. Плазмида была линеаризована по уникальному сайту NdeI, который находится почти в середине фрагмента для дизрупции РМТ6, перенесенного в штамм Pichia с помощью электропорации. Трансформированные клоны отбирали с помощью 100 мкг/мл зеоцина. Была получена пара сотен колоний, и отдельные колонии были высажены штрихом на чашки со средой YPD. Геномная ДНК была изолирована из каждого клона, и для проверки клонов с дизрупцией проводили ПЦР (см. фиг.3Б). Отобранные клоны были депонированы как МТСС5518.
ПРИМЕР 3: ДИЗРУПЦИЯ РМТ4:
Около -516 п.о. кодирующей последовательности, включающей часть кодирующую область РМТ4, амплифицировали с применением следующих праймеров:
PMT4FP=51 GGA ТСС ТАА TAG GTT CAT TTC GCT АТТ СТА AGC А 3'
PMT4RP=5' GGA ТСС TTT CGA CTT CAA AGG ACG GGT Т 3'
Два стоп-кодона были включены в прямой праймер, чтобы избежать какого-либо прочтения с ранних потенциальных сайтов начала трансляции в векторе. Продукт ПЦР был клонирован в вектор pTZ57R, и для подтверждения он был секвенирован с помощью M13FP и M13RP. Кассета для дизрупции РМТ4 была введена в Pichia Pastoris GS115. Плазмида была линеаризована по уникальному сайту XcmI, который находится почти в середине фрагмента для дизрупции РМТ4, перенесенного в штамм Pichia с помощью электропорации. Трансформированные клоны отбирали с помощью 100 мкг/мл зеоцина. Была получена пара сотен колоний, и отдельные колонии были высажены штрихом на чашки со средой YPD. Геномная ДНК была изолирована из каждого клона, и для проверки клонов с дизрупцией проводили ПЦР (см. фиг.4Б). Отобранные клоны были депонированы как МТСС5516.
ПРИМЕР 4: ДИЗРУПЦИЯ РМТ5:
Около -455 п.о. кодирующей последовательности, включающей часть кодирующую область РМТ5, амплифицировали с применением следующих праймеров:
PMT5FP=51 АСА ТСТ ТАА TAG АТС СТА ССА GTG АТС АТТ ТАС СТ 3'
PMT5RP=5' AGA ТСТ ТСА СТА АТТ GGA AGG ТСТ AGA АТС 3'
Два стоп-кодона были включены в прямой праймер, чтобы избежать какого-либо прочтения с ранних потенциальных сайтов начала трансляции в векторе. Продукт ПЦР был клонирован в вектор pTZ57R, и для подтверждения он был секвенирован с помощью M13FP и M13RP. Кассета для дизрупции РМТ5 была введена в Pichia Pastoris GS115. Плазмида была линеаризована по уникальному сайту BstEII, который находится почти в середине фрагмента для дизрупции РМТ5, перенесенного в штамм Pichia с помощью электропорации. Трансформированные клоны отбирали с помощью 100 мкг/мл зеоцина. Была получена пара сотен колоний, и отдельные колонии были высажены штрихом на чашки со средой YPD. Геномная ДНК была изолирована из каждого клона, и для проверки клонов с дизрупцией проводили ПЦР (см. фиг.5Б). Отобранные клоны были депонированы как МТСС5517.
ПРИМЕР 5: ДИЗРУПЦИЯ РМТ2:
Около -439 п.о. кодирующей последовательности, включающей часть кодирующую область РМТ2, амплифицировали с применением следующих праймеров:
PMT2FP=5' GGA ТСС ТАА TAG GTG GGT ТТА ТТТ GTC AC A GTA 3'
Pmt2RP=5' GGA ТСС GAA АСА ССС ААТ CAT TGT TGG СА 3'
Два стоп-кодона были включены в прямой праймер, чтобы избежать какого-либо прочтения с ранних потенциальных сайтов начала трансляции в векторе. Продукт ПЦР был клонирован в вектор pTZ57R, и для подтверждения он был секвенирован с помощью M13FP и M13RP.
Кассета для дизрупции РМТ2 была введена в Pichia Pastoris GS115. Плазмида была линеаризована по уникальному сайту KpnI, который находится почти в середине фрагмента для дизрупции РМТ2, перенесенного в штамм Pichia с помощью электропорации. Трансформированные клоны отбирали с помощью 100 мкг/мл зеоцина. Была получена пара сотен колоний, и отдельные колонии были высажены штрихом на чашки со средой YPD. Геномная ДНК была изолирована из каждого клона, и для проверки клонов с дизрупцией проводили ПЦР (см. фиг.6).
ПРИМЕР 6: СНИЖЕНИЕ СТЕПЕНИ ГЛИКОЗИЛИРОВАНИЯ ИНСУЛИНА С ПОМОЩЬЮ НОКАУТА РМТ
Для получения клона В1СС#7743, уровень гликозилирования в котором использовали в качестве контроля, штамм Pichia pastoris GS115 трансформировали конструкцией для экспрессии инсулина; степень гликозилирования секретируемого инсулина в таком клоне составила 1,90-2,0 (принята за 100%).
Были также клонированы штаммы с нокаутом РМТ МТСС5515, МТСС5517 и МТСС5518 с конструкцией для экспрессии инсулина для сравнения уровня гликозилирования секретируемого инсулина с контрольным уровнем. Было отмечено снижение (см. фиг.7) степени гликозилирования на 61%, 31% и 28%, соответственно, по сравнению с инсулином, продуцированным контрольным клоном В1СС#7743.
ПРИМЕР 7: СНИЖЕНИЕ СТЕПЕНИ ГЛИКОЗИЛИРОВАНИЯ АНАЛОГА ИНСУЛИНА 1 С ПОМОЩЬЮ НОКАУТА РМТ
Для получения клона В1СС#7744, уровень гликозилирования в котором использовали в качестве контроля, штамм Pichia pastoris GS115 трансформировали конструкцией для экспрессии инсулина; степень гликозилирования секретируемого инсулина в таком клоне составила 1,66 (принята за 100%). Был также клонирован штамм с нокаутом РМТ МТСС5515 с конструкцией для экспрессии инсулина для сравнения уровня гликозилирования секретируемого инсулина с контрольным уровнем. Было отмечено снижение (см. фиг.8) степени гликозилирования на 46%, по сравнению с аналогом инсулина 1, продуцированным контрольным клоном В1СС#7744.
ПРИМЕР 8: СНИЖЕНИЕ СТЕПЕНИ ГЛИКОЗИЛИРОВАНИЯ АНАЛОГА ИНСУЛИНА 2 С ПОМОЩЬЮ НОКАУТА РМТ
Для получения клона BICC #7996, уровень гликозилирования в котором использовали в качестве контроля, штамм Pichia pastoris GS115 трансформировали конструкцией для экспрессии инсулина; степень гликозилирования секретируемого инсулина в таком клоне составила 1,92 (принята за 100%). Был также клонирован штамм с нокаутом РМТ МТСС5515 с конструкцией для экспрессии инсулина для сравнения уровня гликозилирования секретируемого инсулина с контрольным уровнем. Было отмечено снижение (см. фиг.9) степени гликозилирования на 65%, по сравнению с аналогом инсулина 1, продуцированным контрольным клоном BICC #7744.















Изобретение относится к области биотехнологии, а именно к способу снижения уровня O-гликозилирования молекул-предшественников инсулина или аналогов инсулина, синтезируемых Pichia sp. Изобретение раскрывает нокаутированные штаммы метилотрофных дрожжей, полученные с помощью генной инженерии. Штаммы содержат дизрупцию в генах протеин-маннозилтрансферазы (РМТ) и способны синтезировать гетерологичные белки со сниженной степенью гликозилирования. Также описан вектор, содержащий кодирующие последовательности генов РМТ1, РМТ2, РМТ4, РМТ5 и РМТ6 для трансформации метилотрофных дрожжей. Изобретение может быть использовано в медицине. 5 н. и 5 з.п. ф-лы, 15 ил., 8 пр.
1. Вектор, содержащий ген протеин-маннозилтрансферазы или его функциональную часть, выбранный из группы, включающей гены PMT1, PMT5 и РМТ6, имеющий нуклеотидную последовательность, представленную в SEQ ID NOs 1, 4 и 5 соответственно, при этом указанный вектор предназначен для трансформации метилотрофных дрожжей для снижения степени или изменения степени гликозилирования предшественника инсулина или предшественника аналога инсулина.
2. Вектор по п. 1, в котором указанный штамм метилотрофных дрожжей принадлежит роду Pichia sp.
3. Вектор по п. 2, в котором указанный штамм метилотрофных дрожжей принадлежит виду Pichia pastoris.
4. Способ получения нокаутированного штамма метилотрофных дрожжей для продуцирования предшественника инсулина или предшественника аналога инсулина со сниженной степенью гликозилирования, включающий:
(а) получение вектора, включающего последовательность нуклеиновой кислоты, способную к гомологичной рекомбинации, содержащую целевую последовательность нуклеиновой кислоты, кодирующую по меньшей мере один из генов, выбранных из группы, включающей гены PMT1, PMT5 и PMT6, имеющий нуклеотидную последовательность, представленную в SEQ ID NOs 1, 4 и 5 соответственно, и последовательность нуклеиновой кислоты, кодирующую селективный маркер,
(б) культивирование клеток в условиях, делающих возможной гомологичную рекомбинацию между ДНК, кодирующей целевой ген в векторе, и ДНК, кодирующей целевой ген в клетке-хозяине, что приводит к нарушению целевого гена в клетке-хозяине;
(в) селекцию клеток-хозяев с инактивированным целевым геном для получения нокаутированного штамма метилотрофных дрожжей для продуцирования предшественника инсулина или предшественника аналога инсулина со сниженной степенью гликозилирования.
5. Нокаутированный штамм метилотрофных дрожжей, содержащий инактивированный ген РМТ1, имеющий нуклеотидную последовательность, представленную в SEQ ID NO 1, где штамм обладает номером доступа MTCC5515 и продуцирует предшественник инсулина или предшественник аналога инсулина, уровень гликозилирования которых снижен по сравнению с инсулиновым продуктом, экспрессированным в немодифицированном штамме-хозяине.
6. Нокаутированный штамм метилотрофных дрожжей, содержащий инактивированный ген РМТ5, имеющий нуклеотидную последовательность, представленную в SEQ ID NO 4, где штамм обладает номером доступа МТСС5517 и продуцирует предшественник инсулина или предшественник аналога инсулина, уровень гликозилирования которых снижен по сравнению с инсулиновым продуктом, экспрессированным в немодифицированном штамме-хозяине.
7. Нокаутированный штамм метилотрофных дрожжей, содержащий инактивированный ген РМТ6, имеющий нуклеотидную последовательность, представленную в SEQ ID NO 5, где штамм обладает номером доступа МТСС5518 и продуцирует предшественник инсулина или предшественник аналога инсулина, уровень гликозилирования которых снижен по сравнению с инсулиновым продуктом, экспрессированным в немодифицированном штамме-хозяине.
8. Нокаутированные штаммы по любому из пп. 5-7, где указанный штамм-хозяин экспрессирует предшественник инсулина или предшественник аналога инсулина, в котором степень гликозилирования снижена по меньшей мере на 25%.
9. Нокаутированные штаммы по любому из пп. 5-7, где указанный штамм-хозяин экспрессирует предшественник инсулина или предшественник аналога инсулина, в котором степень гликозилирования снижена по меньшей мере на 65%.
10. Способ по п. 4, где селективный маркер представляет собой маркер устойчивости к зеоцину.
| WO 2009105357 A1, 27.08.2009 | |||
| WO 2007061631 A2, 31.05.2007 | |||
| СПОСОБ ПРОМОТИРОВАНИЯ АКТИВНОЙ КОНФОРМАЦИИ ГЛИКОЗИЛИРОВАННОГО РЕКОМБИНАНТНОГО БЕЛКА, СПОСОБ ПОЛУЧЕНИЯ АКТИВНОГО ГЛИКОЗИЛИРОВАННОГО БЕЛКА И СПОСОБ ПОЛУЧЕНИЯ КОМПОЗИЦИИ УКАЗАННОГО БЕЛКА ДЛЯ ВВЕДЕНИЯ ПОТРЕБИТЕЛЮ И/ИЛИ ПАЦИЕНТУ ИЛИ ПОТРЕБЛЕНИЯ ПОТРЕБИТЕЛЕМ И/ИЛИ ПАЦИЕНТОМ | 2002 |
|
RU2316563C2 |

