Настоящее изобретение касается прямой корреляции между сверхэкспрессией или функциональными молекулярными модификациями человеческих гомологов семейства sgk и гипертензией. В частности, изобретение относится к обнаружению прямой связи между двумя различными полиморфизмами отдельного нуклеотида (single nucleotide polymorphisms = SNP) в гене hsgk1 и генетической предрасположенностью к гипертензии.
Многочисленные внеклеточные сигналы индуцируют внутриклеточные каскады фосфорилирования/дефосфорилирования, обеспечивающие быструю передачу этих сигналов от плазматической мембраны и ее рецепторов в цитоплазму и клеточное ядро. Специфичность этих обратимых каскадов трансдукции сигналов обеспечивается многочисленными отдельными протеинами, прежде всего киназами, которые переносят фосфатную группу на специфические субстраты.
Зависящую от сыворотки и глюкокортикоида киназу (sgk), серин/треонинкиназу, экспрессия которой повышается в присутствии сыворотки и глюкокортикоидов, сначала клонировали из клеток карциномы молочной железы крысы (Webster и др., 1993). Человеческую версию sgk, обозначенную как hsgk1, клонировали из клеток печени (Waldegger и др., 1997). Было установлено, что на экспрессию hsgk1 можно воздействовать путем регуляции объема клетки. До настоящего времени не было обнаружено такой зависимости экспрессии крысиного sgk от объема клетки. Кроме того, было установлено, что крысиная киназа стимулирует эпителиальный Na+-каналы (ENaC) (Chen и др., 1999; Naray-Pejes-Toth и др., 1999). В свою очередь, ENaC играет решающую роль в выделении Na+ почками. Повышенная активность ENaC приводит к увеличению задержания ионов натрия в почках и вследствие этого к развитию гипертензии.
Наконец, были клонированы два других представителя семейства человеческого гена sgk: hsgk2 и hsgk3 (Kobayashi и др., 1999), которые оба, так же, как и hsgk1, активируются инсулином и IGF1 посредством РI3-киназного пути. Результаты электрофизиологических экспериментов свидетельствуют о том, что совместная экспрессия hsgk2 и hsgk3 также приводит к повышению активности ENaC.
В DE 19708173 A1 описано, что hsgk1 представляет собой эффективное средство для диагностики многих заболеваний, при которых изменения объема клетки играют решающую патофизиологическую роль, таких, например, как гиперантремия, гипоантремия, сахарный диабет, почечная недостаточность, гиперкатаболизм, печеночная энцефалопатия и бактериальные или вирусные инфекции.
В WO 00/62781 уже было описано, что активация эндотелиального Na+-канала с помощью hsgk1 приводит к повышению почечной ресорбции Na+. Поскольку такое повышение ресорбции Na+ в почках связано с гипертензией, то можно предположить, что повышенная экспрессия hsgk1 может приводить к гипертензии, а пониженная экспрессия hsgk1 в конце концов может приводить к гипотензии.
Аналогичная связь между сверхэкспрессией или гиперактивностью человеческих гомологов hsgk2 и hsgk3 и сверхактивацией ENaC, приводящей к повышенной почечной ресорбции Na+ и развивающейся на основе этого гипертензии, описана также в неопубликованной имеющей более ранний приоритет заявке на немецкий патент, озаглавленной "sgk2 и sgk3 в качестве мишеней для диагностических и терапевтических целей" (внутреннее обозначение А 35 048) от 28.08.00. Кроме того, уже обсуждалась возможность использования киназ hsgk2 и hsgk3 для диагностики артериальной гипертензии.
В основу настоящего изобретения была положена задача выявить экспериментальным путем прямую корреляцию, т.е. прямую связь между сверхэкспрессией или функциональной молекулярной модификацией человеческих гомологов семейства sgk и гипертензией.
В контексте настоящего описания гомолог семейства sgk, который имеет вышеуказанную функциональную молекулярную модификацию, представляет собой гомолог семейства sgk, который подвергнут такой мутации, в результате которой изменяются свойства, прежде всего каталитические свойства или даже субстратную специфичность соответствующего протеина.
Следующей задачей изобретения является использование этой прямой корреляции или связи между между сверхэкспрессией или функциональной молекулярной модификацией человеческих гомологов семейства sgk и гипертензией в способе диагностики предрасположенности к обусловленной генетическими факторами форме гипертензии.
Согласно настоящему изобретению можно проводить обнаружение прямой корреляции между сверхэкспрессией или функциональной молекулярной модификацией человеческих генов sgk и гипертензией, и, в частности, она экспериментально доказана на примере гена hsgk1.
Таким образом, решение указанной выше задачи заключается в использовании этой прямой корреляции между сверхэкспрессией или функциональной молекулярной модификацией человеческих гомологов семейства sgk, прежде всего гена hsgk1 gene, и гипертензией для диагностики обусловленной генетическими факторами формы гипертензии.
Решение указанной выше задачи заключается, в частности, в том, что при создании настоящего изобретения в гене hsgk1 были идентифицированы два различных SNP, которые, если они присутствуют в конкретной версии гена hsgk1, вызывают у пациента определенную предрасположенность к гипертензии. Присутствие таких SNP в гене hsgk1 или также в других человеческих гомологах семейства генов sgk, обнаруженное в образцах, взятых из организма пациентов, может служить диагностическим показателем генетической предрасположенности к развитию гипертензии.
Кроме того, указанная выше задача решается с помощью предложенного в изобретении способа диагностики, позволяющего осуществлять количественный диагноз определенной формы генетической гипертензии, заключающийся в том, что сверхэкспрессию человеческого гомолога семейства sgk или функциональную молекулярную модификацию этих гомологов выявляют путем количественного обнаружения гомологов в образце, взятом из организма пациента, с помощью антитела к протеинам гомологов или полинуклеотидов, которые могут гибридизоваться с ДНК или мРНК гомологов в строгих условиях, и набора для диагностики, пригодного для осуществления этого способа.
Набор по изобретению предпочтительно содержит антитела к протеину hsgk1 или полипептиды, которые могут гибридизоваться с геном hsgk1 в строгих условиях.
Такой набор для диагностики содержит, в частности, антитела, обладающие специфической активностью против областей протеина hsgk1, которые включают фрагмент протеина hsgk1, несущий мутацию в гене hsgk1, соответствующую определенному SNP. Однако набор может содержать также антитела против наиболее часто встречающихся аллелей гена hsgk1 или других гомологов семейства sgk, с помощью которых можно количественное определить модифицированный уровень экспрессии этих гомологов или hsgk1.
Кроме того, набор для диагностики по изобретению предпочтительно включает полинуклеотиды, имеющие специфические области, содержащие одну или несколько версий связанных с гипертензией SNP в гене hsgk1, и таким образом пригодных для обнаружения специфических SNP в гене hsgk1 путем гибридизации в строгих условиях с геномной ДНК, кДНК или мРНК, полученных из образцов, взятых из организма пациента.
Согласно настоящему описанию наличие прямой корреляции между гипертензией и человеческими гомологами семейства sgk означает, что у некоторых пациентов могут происходить индивидуальные мутации генов hsgk1, hsgk2 или hsgk3, приводящие к изменению уровня экспрессии или функциональных свойств киназ hsgk1, hsgk2 или hsgk3, обусловливая тем самым вызванную генетическими факторами предрасположенность к гипертензии. Такие мутации могут возникать, например, в регуляторных областях гена или также в интронных последовательностях в локусе гена sgk и вследствие этого вызывать сверхэкспрессию соответствующей киназы и сверхактивацию ENaC. С другой стороны, индивидуальные различия в генетической структуре локуса sgk могут влиять также на кодирующую область гена. Мутации в кодирующей области могут, кроме того, приводить к функциональному изменению соответствующей киназы, например к изменению каталитических свойств киназы. Поэтому оба типа описанных выше мутаций могут вызывать повышенную активацию ENaC и, следовательно, в конце концов приводить к возникновению обусловленной генетическими факторами формы гипертензии у пациента.
Такие мутации в человеческих гомологах семейства sgk, которые вызывают развитие обусловленной генетическими факторами формы гипертензии у пациента, как правило, представляют собой так называемые полиморфизмы отдельного нуклеотида (SNP) либо в области экзона, либо в области интрона этих гомологов. SNP в области экзона генов hsgk могут, в их сравнительно редко встречающейся версии, которая в настоящем описании названа мутантной версией, приводить к аминокислотным заменам в соответствующем протеине hsgk и следовательно к функциональной модификации киназы. SNP в области интрона или в регуляторных последовательностях генов hsgk в их мутантных версиях могут приводить к изменению уровня экспрессии соответствующей киназы.
При создании настоящего изобретения проводили исследование корреляции между наличием генотипа гена hsgk1 у различных пациентов (близнецов) и величинами измеренного у них систолического и диастолического кровяного давления, которое в каждом случае измеряли при различных положениях тела (сидя, стоя, лежа), и производили статистический анализ.
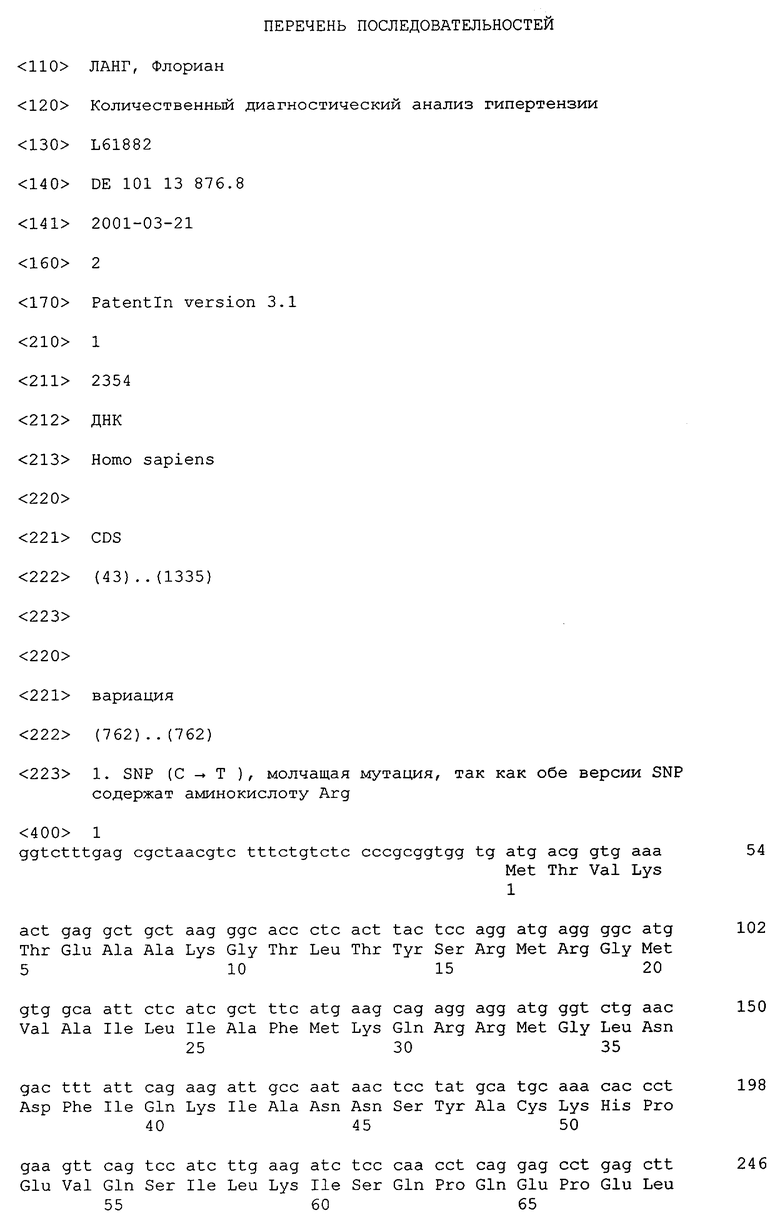
При создании изобретения было установлено, что наличие замены (С→Т) в экзоне 8 (1-ый SNP, см. SEQ ID NO.1) в обоих аллелях (гомозиготные ТТ-носители SNP в экзоне 8), которая не приводит к аминокислотной замене на уровне протеина (см. SEQ ID NO.2), обусловливает существенно более высокие уровни кровяного давления и тем самым генетическую предрасположенность к гипертензии (таблица 3).
Кроме того, было установлено, что замена (Т→С) (2-ой SNP), локализованная на расстоянии 551 пар оснований от 1-го SNP в донорном сайте сплайсинга интрона 6 и экзона 7, в ее гомозиготной форме приводит к более низким величинам кровяного давления и тем самым к более слабой генетической предрасположенности к гипертензии (таблица 3).
Поскольку оба SNP в гене hsgk1 по изобретению не приводят к аминокислотным заменам на уровне протеина, то вызываемая ими более или менее выраженная генетическая предрасположенность к гипертензии, по-видимому, обусловлена измененным уровнем экспрессии гена hsgk1.
Первый SNP в экзоне 8 (С→Т) более подробно проиллюстрирован на чертеже. На чертеже представлены индивидуальные экзоны гена hsgk1 и в каждом случае указаны номер экзона, идентификационный номер (ID) экзона, связанная с ним "непрерывная последовательность (sequence-contig)" и номер цепи, а также начало, конец и длина экзона. Точное положение замены (С→Т) в рамке SNP в экзоне 8 обозначено с помощью черного маркера на букве С в экзоне 8. Светлая маркировка в экзоне 8 на чертеже обозначает SNP-фланкирующую последовательность в гене hsgk1, которая однозначно определяет положение в геноме.
Второй SNP (Т→С) в интроне 6 идентифицировали путем прямого секвенирования, и он однозначно характеризуется тем, что локализован в гене hsgk1 (содержащем экзоны и интроны) точно на расстоянии 551 пары оснований от первого SNP в экзоне 8 против хода транскрипции в донорном сайте сплайсинга интрона 6 и экзона 7 гена 1 и связан с заменой Т на С.
Кроме того, было установлено, что все величины систолического и диастолического кровяного давления, измеренные у пациентов при различных положениях тела, в равной мере зависят от генотипа гена hsgk1 (таблица 4). Данные, приведенные в таблице 4, свидетельствуют о том, что корреляция между измеренным у пациентов кровяным давлением и встречаемостью указанных выше полиморфизмов (SNP) в их генах hsgk1 действительно является статистически достоверной.
Кроме того, при анализе двух SNP в гене hsgk1 был выявлен большой дисбаланс в частоте их взаимной встречаемости (таблица 5). В то время как большинство СС-носителей SNP в экзоне 8 являются также ТТ-носителями SNP в интроне 6 (64%), обратное соотношение является другим (только 2% ТТ-носителей в экзоне 8 являются также СС-носителями в экзоне 6).
При создании изобретения впервые была установлена корреляция между кровяным давлением у пациентов и их индивидуальной генетической версией локуса гена hsgk1, которая свидетельствует о том, что специфические антитела или полинуклеотиды к hsgk1 можно применять для диагноза специфической генетической предрасположенности к гипертензии. Эта специфическая обусловленная генетическими факторами гипертензия может отличаться повышенной экспрессией hsgk1, т.е сверхэкспрессией, или возможно также модифицированными функциональными свойствами hsgk1.
Поскольку две гомологичные киназы семейства sgk, т.е. hsgk2 и hsgk3, активируют также ENaC, то специфические антитела и полинуклеотиды по изобретению к hsgk2 или hsgk3 можно применять также для диагностического анализа определенных обусловленных генетическими факторами форм гипертензии.
Сделанное при создании изобретения открытие, что встречаемость двух SNPs в гене hsgk1 коррелирует с предрасположенностью к гипертензии, свидетельствует, частности, о том, что полинуклеотиды, несущие ту или иную версию двух SNP в гене hsgk1, особенно предпочтительно применять для диагностики обусловленной генетическими факторами формы гипертензии для гибридизации с эндогенной ДНК (кДНК или геномной ДНК) или мРНК, полученной из образца, выделенного из организма пациента. Аналогично этому представленные в настоящем описании данные свидетельствуют о том, что для диагностики генетической предрасположенности к гипертензии можно применять антитела к специфическим связанным с гипертензией полиморфизмам (SNP) в протеине hsgk1 или в одном из его человеческих гомологов. Такие SNP, которые приводят также к связанному с гипертензией полиморфизму на уровне протеина, могут быть связаны, в частности, с функциональной модификацией протеина hsgk1 и тем самым обусловливать предрасположенность к гипертензии.
Настоящее изобретение относится также к использованию прямой корреляции, т.е. корреляции между сверхэкспрессией или функциональной молекулярной модификацией человеческих гомологов семейства sgk, прежде всего hsgk1, и гипертензией, для количественного анализа определенной формы генетической гипертензии.
В частности, для количественного анализа определенной формы генетической гипертензии применяют два SNP в гене hsgk1, которые коррелируют с предрасположенностью к гипертензии.
Кроме того, изобретение относится к способу количественной диагностики обусловленной генетическими факторами формы гипертензии, заключающемуся в том, что сверхэкспрессию человеческого гомолога семейства sgk или функциональную молекулярную модификацию этих гомологов выявляют путем количественного обнаружения гомологов в образце, выделенном из организма пациента, с помощью антител к протеинам гомологов или полинуклеотидов, которые могут гибридизоваться с геномной ДНК, кДНК или мРНК гомологов в строгих условиях.
В этом способе диагностики по изобретению предназначенные для применения образцы, выделенные из организма пациента, предпочтительно представляют собой образцы крови или образцы слюны, содержащие клеточный материал, и их можно выделять из организма пациента относительно дешевым методом. Можно применять, однако, и другие образцы, выделенные из организма пациента, которые также содержат клетки, например образцы ткани и т.д. Из этого содержащего клетки материала образцов, выделенных из организма, можно получать стандартными методами как геномную ДНК или кДНК, а также мРНК (Sambrook J. и Russel D.W. Cold Spring Harbor, NY, изд-во CSHL Press, 2001), и при необходимости подвергать амплификации и последующей гибридизации в строгих условиях с полинуклеотидами, которые могут специфически гибридизоваться с указанной геномной ДНК, кДНК или также с мРНК. Кроме того, протеиновый экстракт можно выделять также из содержащего клетки материала образцов, выделенных из организма (кровь, слюна, ткань и т.д.) с помощью стандартных методов (Sambrook J. и Russel D.W. Cold Spring Harbor, NY, изд-во CSHL Press, 2001), и затем соответствующий протеин sgk можно количественно обнаруживать путем инкубации с антителом к этому протеину.
В способе по изобретению предпочтительно применяют антитела к протеину hsgk1 или полинуклеотидам, которые могут гибридизоваться с геномной ДНК, кДНК или мРНК гена hsgk1.
В способе по изобретению применяют, в частности, полинуклеотиды, которые могут гибридизоваться в строгих условиях с геномной ДНК, кДНК или мРНК версии SNP в интроне 6 гена hsgk1 или версии SNP в экзоне 8 гена hsgk1. В контексте настоящего описания гибридизация в строгих условиях означает гибридизацию в таких условиях гибридизации с точки зрения температуры гибридизации и содержания формамида в растворе для гибридизации, уоторые описаны с соответствующей технической литературе (Sambrook J. и Russel D.W. Cold Spring Harbor, NY, изд-во CSHL Press, 2001).
Кроме того, объектом изобретения является набор для количественной диагностики определенной обусловленной генетическими факторами формы генетической гипертензии, включающий антитела к человеческим гомологам протеина семейства sgk, или полинуклеотидам, которые могут гибридизоваться в строгих условиях с человеческими гомологами гена семейства sgk, или такие антитела совместно с полинуклеотидами, предназначенный для количественного определения сверхэкспрессии или функциональной молекулярной модификации этих гомологов.
Антитела, содержащиеся в наборе, предпочтительно представляют собой антитела к протеину hsgk1, а содержащиеся в наборе полинуклеотиды, предпочтительно обладают способностью гибридизоваться с геном hsgk1.
Наиболее предпочтительно набор для диагностики может содержать полинуклеотиды, которые могут гибридизоваться с геномной ДНК, кДНК или мРНК версии SNP в интроне 6 (Т→С) или SNP в экзоне 8 (С→Т).
Ниже изобретение более подробно проиллюстрировано на примерах.
Пример 1
Анализ корреляции проводили в группе, состоящей из 75 пар дизиготных близнецов (Busjahn и др., J. Hypertens., 14, 1996, cc.1195-1199; Busjahn и др., Hypertension, 29, 1997, c.165-170). Все участвовавшие в эксперименте индивидуумы принадлежали к германо-кавказской расе и происходили из различных областей Германии. Для проверки того, что они являются дизиготными, и для дальнейшего молекулярно-генетического анализа у пар близнецов, а также у их родителей брали образцы крови. Перед этим проводили медицинское обследование каждого участвующего в эксперименте индивидуума. Ни у одного из индивидуумов, участвующего в эксперименте, не было выявлено хронического медицинского заболевания. У тестируемых индивидуумов после их нахождения в течение 5 мин в сидячем положении квалифицированный специалист измерял кровяное давление с помощью стандартного ртутного сфигмоманометра (2 измерения с интервалом времени 1 мин). Среднее значение, полученное по двум измерениям, использовали в качестве величины кровяного давления.
Преимущество эксперимента на дизиготных близнецах с точки зрения исследования корреляции заключается в том, что они имеют совершенно одинаковый возраст и можно считать, что внешние воздействия на их фенотипы являются минимальными (Martin и др., Nat. Genet., 17, 1997, c.387-392).
На важность исследований на близнецах для объяснения комплексных генетических заболеваний недавно было указано в работе Martin и др., 1997. Тот факт, что пары близнецов являлись дизиготными, подтверждали с помощью амплификации пяти микросателлитных маркеров с использованием полимеразной цепной реакции (ПЦР). В этом анализе микросателлитные маркеры, представляющие собой фрагменты дезоксирибонуклеиновой кислоты (ДНК), амплифицировали с помощью ПЦР с использованием выделенных из организма различных людей специфических олигонуклеотидов, содержащих высоковариабельные области. Высокую вариабельность этих областей генома можно обнаруживать по небольшим различиям размеров амплифицированных фрагментов, и, если существуют различия в соответствующем сайте гена, то после разделения продуктов ПЦР с помощью гель-электрофореза образуются двойные полосы, так называемые микросателлитные полосы (Becker и др., J. Reproductive Med., 42 1997, c.260-266).
Для молекулярно-генетического анализа гена-мишени, в данном случае гена hsgk1, с помощью ПЦР амплифицировали еще три микросателлитные маркерные области (d6s472, d6s1038, d6s270), расположенные в непосредственной близости от локуса hsgk1, и затем сравнивали с соответствующими образцами, выделенными из организма второго близнеца и родителей. Таким методом можно определять, имеют ли близнецы наследуемые идентичные или различные аллели (из числа рассматриваемых аллелей) по сравнению с их родителями. Корреляционный анализ проводили с использованием модели на основе так называемого "моделирования структурного уравнения " (SEM) (Eaves и др., Behav. Genet., 26, 1996, c.519-525; Neale, Mx: Statistical modeling. Box 126 MCV, Richmond, VA 23298: Department of Psychiatry, 4-е изд., 1997). Эта модель основана на использовании матриц дисперсий-ковариаций для тестируемых пар, которые отличаются вероятностью того, что они не имеют ни одного, имеют один или два идентичных аллеля. С точки зрения фенотипа вариация подразделяется на вариацию, генетически обусловленную всеми генами (А), вариацию, обусловленную геном-мишенью (Q), в рассматриваемом случае геном hsgk1, и вариацию, обусловленную внешними факторами (Е).
VAR=А2+Q2+Е2
Для трех возможных аллельных комбинаций IBDo, IBDi, IBDz (IBD означает "идентичный с потомком"; 0, 1 или 2 обозначают количество идентичных аллелей), ковариация тестируемой пары определяли следующим образом:
COV(IBD0)=0,5A2 COV(IBD1)=0,5А2+0,5Q2 COV(IBD2)=0,5A2+Q2
Для оценки корреляции между генетической структурой локуса hsgk1 и кровяным давлением тестируемого индивидуума различия между моделями, которые учитывают или не учитывают генетическую вариацию в отношении гена-мишени hsgk1, вычисляли с использованием статистического критерия χ2. Для каждой пары и для каждого локуса гена соотношения аллелей вычисляли с помощью так называемой "многоточечной" модели (MAPMAKER/SIBS; Kruglyak и др., Am. J. Hum. Genet., 57, 1995, c.439-454), основанной на использовании генотипов родителей.
Более высокая информативная ценность метода анализа, основанного на оценке дисперсий-ковариаций, по сравнению с описанным выше статистическим анализом, основанным на использовании χ2-критерия (S.A.G.E. Statistical Analysis for Genetic Epidemiology, Release 2.2. Computer program package, Department of Epidemiology and В io statistics. Case Western Reserve University, Cleveland, OH, USA, 1996), была продемонстрирована недавно в исследовании на моделях (Fulker и др., Behav. Gen., 26, 1996, c.527-532). Для того, чтобы гарантировать значимый уровень корреляции в соответствии с критериями Lander и Kruglyak (Lander и др., Nat. Genet., 11, 1995, c.241-246), был принят уровень значимости (вероятность ошибки) р<0,01.
Результаты этого исследования корреляции приведены в таблице 1.
Как видно из результатов, приведенных в таблице 1, низкие уровни значимости р, которые лишь немного превышают принятый уровень значимости р<0,01 или совсем не превышают его, свидетельствуют о наличии прямой корреляции между генетической дисперсией в сайте гена hsgk1 и фенотипической дисперсией измеренного кровяного давления.
Пример 2
Структура генома гена hsgk1 уже была описана ранее (Waldegger и др., Genomics, 51, 299 [1998]),
http://www.ensembl.org/Homo_sapiens/geneview?gene=ENSG00000118515).
Для идентификации SNP, наличие которых связано с предрасположенностью к развитию гипертензии, сначала проводили изучение известных SNP в гене hsgk1, находящихся в банках данных, с точки зрения того, являются ли они истинными SNP, а не следствием ошибок секвенирования, и обладают ли SNP достаточным полиморфизмом для того, чтобы служить основой для диагностического определения предрасположенности к гипертензии. SNP rs 1057293 в экзоне 8, который связан с заменой С на Т, удовлетворял сформулированным заранее необходимым требованиям
(http://www.ensembl.org/Homo_sapiens/snpview?snp=1057293;
http://www.ncbi.nlm.nih.gov/SNP/snp_ref.cgi?type=rs&rs=1057293). Кроме того, прямым секвенированием был идентифицирован второй SNP, локализованный в гене hsgk1 точно на расстоянии 551 пары оснований от первого SNP в донорном сайте сплайсинга интрона 6 и экзона 7, и который связан с заменой Т на С. Эти два SNP в интроне 6 (Т→С) и в экзоне 8 (С→Т) анализировали согласно описанному ниже методу.
После ПЦР-амплификации для получения вырожденного ПЦР-праймера и дефосфорилирования дНТФ в каждом случае добавляли 1 единицу щелочной фосфатазы и 1 единицу экзонуклеазы I. ПЦР проводили в следующих условиях: 95°С в течение 10 мин, затем 35 циклов при 95° в течение 15 с, затем 62°С в течение 15 с, затем 72°С в течение 30 с и стадия удлинения при 72°С в течение 10 мин в термоячейке типа 9600 Thermocycler (фирма Applied Biosystems).
Реакции минисеквенирования проводили с использованием праймеров для SNP (Т→С) интрона 6: 5'-СТС СТТ GCA GAG TCC GAA и для SNP (С→Т) экзона 8: 5'-АСС AAG ТСА ТТС TGG GTT GC. 0,15 пмоля очищенного ПЦР-продукта использовали в качестве матрицы при ПЦР-секвенировании. Для ПЦР-секвенирования осуществляли 25 циклов амплификации, состоящих из следующих отдельных стадий: стадия денатурации в течение 10 с при 96°С, стадия отжига в течение 10 с при 50°С и стадия удлинения в течение 30 с при 60°С в термоячейке типа 9600 Thermocycler.
У тех пациентов, у которых был определен SNP-генотип гена hsgk1, измеряли систолическое и диастолическое кровяное давление в положении лежа, стоя и сидя для определения корреляции между SNP-генотипом гена hsgk1 и кровяным давлением.
В таблице 2 представлены некоторые демографические данные, касающиеся близнецов, и результаты анализа корреляции между генетической структурой локуса гена hsgk1 и измеренным кровяным давлением. Результаты свидетельствуют о том, что для подвергавшихся исследованию индивидуумов кровяное давление, измеренное во всех положениях, существенно зависит от генетических факторов.
В таблице 3 приведены дополнительные результаты опытов по оценке корреляции, полученные при создании изобретения. Установлено, что частоты встречаемости в аллелях для SNP в экзоне 8 составляют для С 91% и для Т 9% соответственно, а для SNP в интроне 6 они составляют для Т 79% и для С 21% соответственно (для обоих полиморфизмов поддерживается равновесие Харди-Вайнберга).
Результаты измерения кровяного давления во всех положениях (сидя, лежа, стоя) характеризуются одинаковой тенденцией. Величины кровяного давления у гомозиготных СС-носителей и гетерозиготных СТ-носителей SNP в экзоне 8 не отличаются друг от друга, однако для них характерно существенно более низкое систолическое и диастолическое кровяное давление, чем для гомозиготных ТТ-носителей SNP в экзоне 8.
Соответствующие результаты анализа корреляции для SNP в интроне 6 являются менее статистически достоверными по сравнению с SNP в экзоне 8. Однако было установлено, что гомозиготные СС-носители SNP в интроне 6 в целом характеризуются более низкой величиной кровяного давления, чем гомозиготные ТТ-носители и гетерозиготные ТС-носители SNP в интроне 6.
Результаты, представленные в таблице 4, убедительно свидетельствуют о том, что генетическая организация SNP в интроне 6 практически одинаково достоверно влияет как на величину систолического, так и диастолического давления независимо от положения, в котором производили измерение кровяного давления (сидя, стоя, лежа). Результаты, свидетельствующие о важности генетической организации SNP в экзоне 8, являются сходными, однако достоверность связи между величинами систолического и диастолического кровяного давления, измеренными в различных положениях, несколько меньше, чем для SNP в интроне 6.
Как следует из данных, представленных в таблице 5, существует выраженное корреляционное равновесие между двумя анализируемыми SNP: в то время как большинство СС-носителей SNP в экзоне 8 являются также ТТ-носителями SNP в интроне 6 (64%), обратное соотношение не является таковым (только 2% ТТ-носителей в экзоне 8 являются также СС-носителями в интроне 6).




| название | год | авторы | номер документа |
|---|---|---|---|
| sgk1 В КАЧЕСТВЕ ДИАГНОСТИЧЕСКОЙ И ТЕРАПЕВТИЧЕСКОЙ МИШЕНИ | 2003 |
|
RU2331072C2 |
| КИНАЗЫ sgk 2 И sgk 3 В КАЧЕСТВЕ МИШЕНЕЙ ДЛЯ ДИАГНОСТИЧЕСКИХ И ТЕРАПЕВТИЧЕСКИХ ЦЕЛЕЙ | 2001 |
|
RU2310471C2 |
| ЛЕКАРСТВЕННЫЕ СРЕДСТВА, СОДЕРЖАЩИЕ ИНГИБИТОРЫ ЧЕЛОВЕЧЕСКОЙ h-sgk КИНАЗЫ, КОНТРОЛИРУЮЩЕЙ КЛЕТОЧНЫЙ ОБЪЕМ | 2000 |
|
RU2288718C9 |
| СПОСОБ ПРОГНОЗИРОВАНИЯ УРОВНЯ АРТЕРИАЛЬНОГО ДАВЛЕНИЯ У БЕРЕМЕННЫХ В ЗАВИСИМОСТИ ОТ ГЕНЕТИЧЕСКОГО ПОЛИМОРФИЗМА ЭНДОТЕЛИНА 1 | 2011 |
|
RU2473912C1 |
| СПОСОБ ВЫЯВЛЕНИЯ ПРЕДРАСПОЛОЖЕННОСТИ К ВЫСОКОГОРНОМУ ОТЕКУ ЛЕГКИХ | 2003 |
|
RU2353656C2 |
| ГЕНЕТИЧЕСКИЙ ТЕСТ НАКОПЛЕНИЯ МЕДИ В ПЕЧЕНИ У СОБАК | 2012 |
|
RU2662660C2 |
| ГЕНЕТИЧЕСКИЙ ТЕСТ НАКОПЛЕНИЯ МЕДИ В ПЕЧЕНИ У СОБАК | 2012 |
|
RU2707814C2 |
| СПОСОБ ЛЕЧЕНИЯ И ПРЕДУПРЕЖДЕНИЯ ПОТЕРИ КОСТНОЙ ТКАНИ | 2007 |
|
RU2358728C2 |
| МУТАЦИИ ГЕНА МТS В ЗАРОДЫШЕВОЙ ЛИНИИ И СПОСОБ ВЫЯВЛЕНИЯ ПРЕДРАСПОЛОЖЕННОСТИ К ЗЛОКАЧЕСТВЕННЫМ ОПУХОЛЯМ В ГЕНЕ МТS | 1995 |
|
RU2161309C2 |
| ГЕН MTS, МУТАЦИИ ДАННОГО ГЕНА И СПОСОБЫ ДИАГНОСТИКИ ЗЛОКАЧЕСТВЕННЫХ ОПУХОЛЕЙ С ИСПОЛЬЗОВАНИЕМ ПОСЛЕДОВАТЕЛЬНОСТИ ГЕНА MTS | 1995 |
|
RU2164419C2 |

Изобретение относится к области медицины и касается способа диагностики in vitro, обусловленной генетическими факторами предрасположенности к гипертонии посредством выявления в образце (а) SNP, который выявляют в экзоне 8(С→Т) гена hsgk1 и представленном в SEQ ID NO:1, а также (б) SNP, который выявляют в интроне 6(Т→С) гена hsgk1 и локализованном во фрагменте размером 551 п.о. против хода транскрипции по отношению к SNP в экзоне 8 согласно подпункту (а), или обоих SNP согласно подпунктам (а) и (б). Изобретение также относится к применению таких SNP для получения диагностикумов и наборов для диагностики обусловленной генетическими факторами предрасположенности к гипертонии. Преимущество изобретения заключается в обнаружении конкретных последовательностей гена hsgk1, обуславливающих предрасположенность к гипертонии. 3 н.п. ф-лы, 5 табл., 1 ил.
(а) коррелирующий с гипертонией SNP, который выявляется в экзоне 8 (С→Т) гена hsgk1 и представлен в SEQ ID NO:1;
(б) коррелирующий с гипертонией SNP, который выявляется в интроне 6 (Т→С) гена hsgk1 и локализован в фрагменте размером 551 п.о. против хода транскрипции по отношению к SNP в экзоне 8 согласно подпункту (а), или
(в) коррелирующий с гипертонией подпункту (а) и коррелирующий с гипертонией SNP подпункту (б).
| WO 00/62781 А, 26.10.2000 | |||
| WAERNTGES S | |||
| et al., Transcriptional upregulation of the human serine-threonine Kinase hSGK1 in glomerulonephritis, Kidney and blood pressure zesearch, 2000, v.23, №3-5, seite 246. |

