Изобретение относится к инженерной экологии и природоохранному обустройству территорий, в том числе в лесном и сельском хозяйствах, и может быть использовано при сертификации ландшафтов с произрастающими древесными растениями, а также при сертификации лесных массивов по качеству, например молодняков естественного и искусственного происхождения, побегов молодых растений, а также при экологической оценке лесов различного возраста и лесных территорий.
Известен способ испытания побегов древесных растений по геометрическому расположению почек на побегах (см. книгу: Бюсген М. Строение и жизнь наших лесных деревьев / Пер. с нем. - М. - Л.: Гослесбумиздат, 1961. - 424 с. - С.12, 21-23), включающий измерение длины побега, распределение почек и расстояний между почками на побеге (длину междуузлия), хода роста побега в вегетационный период с изменением времени, переменная длина побега по узлам почек.
Недостатком является невозможность учета геометрического расположения почек на побеге, как наиболее постоянного признака растений, при экологической оценке места произрастания, то есть той территории, на которой произрастают испытуемые растения с годичными побегами. Нарушение геометрического расположения почек указывает на плохую экологическую обстановку на месте произрастания.
Известен также способ испытания древесины растущих деревьев с использованием кернов по патенту №2144185, МКИ G01N 33/46, А01G 23/00, включающий взятие кернов, измерение геометрических параметров каждого керна.
Недостатком является то, что этот способ применим преимущественно для средневозрастных, спелых и перестойных деревьев, у которых сформировалась спелая древесина, и она вдоль радиуса сечения ствола четко разделяется на заболонную, рабочую и присердцевинную зоны. Вместе с тем экологическая оценка территорий наиболее целесообразна на молодых деревьях, которые не имеют хозяйственной ценности, но могут быть использованы в инженерной экологии.
Главным недостатком является высокая трудоемкость метода кернов, требующего сложную аппаратуру и трудоемкие действия при камеральных измерениях. При этом с побегов деревьев невозможно брать керны древесины.
Технический результат - повышение простоты и снижение трудоемкости измерений на побегах древесных растений.
Этот технический результат достигается тем, что способ испытания побегов древесного растения, включающий взятие опытных образцов, измерение геометрических параметров каждого образца, отличающийся тем, что взятие образцов в виде годичных побегов проводят в конце вегетационного периода, при этом выбирают учетное древесное растение, срезают опытные образцы побегов около их основания, отмечают на комлевом конце каждого опытного побега номер дерева и номер опытного образца, после этого срезанные и маркированные опытные побеги отправляют для лабораторных измерений, при этом измерение геометрических параметров каждого опытного побега проводят путем измерения геометрического расположения почек на срезанных побегах, по измеренным значениям геометрических параметров расположения почек на каждом опытном побеге статистическим моделированием выявляют закономерность хода роста побега за вегетационное время по следующей статистической формуле:
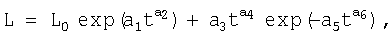
где L - переменная расчетная длина побега от его основания, мм;
L0 - начальная (стартовая) длина побега до момента измерения, мм;
t - время с начала вегетационного периода, дни;
а1...a6 - параметры статистической модели,
или закономерность распределения почек и образования узлов почек по следующей формуле:
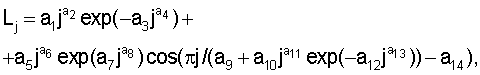
где Lj - переменная расчетная длина побега по узлам почек, мм;
j - номер узла почек, начиная от основания, причем j=1, 2, 3, ...;
a1...a14 - параметры статистической модели,
и по максимальной относительной погрешности найденных статистических моделей судят об экологическом качестве древесного растения и места его произрастания, причем, чем меньше относительная погрешность, тем выше экологическое качество.
Взятие образцов в виде годичных побегов проводят в конце вегетационного периода, например в конце сентября или в октябре месяце, когда годичные побеги выросли полностью в длину и образовались в течение вегетационного периода все почки на следующий вегетационный период.
В лесу или на местности с древесной растительностью выбирают учетное дерево, то есть такое дерево, которое позволяет многократно проводить экологические обследования и эксперименты с годичными побегами и в другие вегетационные периоды, а затем осматривают крону для возможного взятия побегов как опытных образцов.
Сущность изобретения заключается в том, что геометрическое расположение почек на побегах по спирали принадлежит к наиболее постоянным признакам. Изучение геометрии распределения почек по побегу позволяет понять многое, а полученные знания и закономерности становится возможным применять в экологической индикации и биологическом тестировании территории. Особенно, в этом плане, интересны «Ивановы побеги», образующиеся в результате ухудшения внешних условий ростовой деятельности древесных растений.
Такие аномальные побеги (по Бюсгену) наиболее выражены у молодых деревьев. Прежде всего, они появляются с ухудшением условий роста. Предки наших деревьев были вечнозелеными, подобно многим тропическим древесным растениям, имели независимую периодичность в росте побегов. Затем появились сезоны года. Поэтому способность побегов вступать в состояние покоя в середине лета может быть объяснено тем, что предки произрастали в теплых районах и они выработали механизм адаптации к сухости лета.
Сущность заключается также в том, что, если же второй рост происходит поздно, то среди поздних узких элементов древесины снова, как весной, появляются широкие водопроводящие сосуды. Если же почки распускаются раньше, то в древесине возникают и широкие сосуды, но они не образуют ясно выделяющейся зоны. Поэтому разделение годичного слоя на четыре зоны, вместо двух, встречает определенные трудности из-за нечеткости границ между отдельными зонами.
Сущность заключается также в том, что ростовую верхушечную почку имеют хвойные породы, а также ясень, клен, бук и дуб. Причиной отмирания верхушки побега чаще всего является конкуренция верхушечных листьев с вершиной побега за проходящий по старым частым побега поток воды (питательного раствора). Поэтому удаление верхушечных листьев может удлинить жизнь верхушки побега. Уход за побегом позволит увеличить длину самого побега, а значит и всего дерева при уходе за терминальным побегом.
Сущность заключается также в том, что значительные летние осадки и даже травмы вызывают процесс возникновения «Ивановых побегов». Это значит, что излишек влаги также является экологическим загрязнением места произрастания. Аналогично действуют излишнее удобрение, заморозки, засуха, повреждение насекомыми и пр.
Сущность заключается также в том, что растущее дерево и древесные растения всегда готовы откликнуться на благоприятные изменения значений лимитирующих факторов: освещения, тепла, влаги, минеральных веществ, свойств воздушной среды. Но, увы, все это зависит ныне от действий людей. Поэтому необходим действенный и в то же время очень простой способ, позволяющий широко применять в инженерной экологии и природоохранном обустройстве территорий.
Положительный эффект заключается в том, что за последние десятилетия много аномальных побегов появляется у молодняка в загущенных посадках, в которых своевременно не был проведен уход за лесом осветлением или прочисткой. Поэтому необходим способ сертификации молодняков по состоянию побегов отдельных молодых деревьев.
Годичные побеги позволяют изучать влияние загазованной окружающей дерево среды, если поместить молодые деревья в специальные герметичные и прозрачные помещения (например, в США проводили опыты под полиэтиленовыми пленками). Внутри этого помещения регистрируются температура, влажность и концентрации газов, в том числе и экологически вредных для человека.
Положительный эколого-экономический эффект заключается в том, что сертификации подвергаются не только лучшие (с позиций технических целей, которые обычно и экологически лучше), но и худшие (которые технически и экологически худшие) низкокачественные деревья. Поэтому лесная территория получает комплексную оценку, а не только однобоко с позиций промышленника (для которого побеги растения не представляют коммерческого интереса). При этом учитываются свойства не только стволов деревьев, но и условий их роста и развития на геометрических свойствах побегов. Таким образом, предлагаемый способ действительно реализует эколого-экономический подход, а не только утилитарно-потребительский подход в сертификации древесного сырья для изделий из древесины.
В связи с этим техническое решение обладает существенными признаками новизны, положительным эффектом и перспективой расширения практического применения в инженерной экологии, то есть в экологической оценке лесных территорий, а также территорий с произрастающими древесными растениями в лесном и сельском (побеги садовых деревьев) хозяйствах.
Из анализа научно-технической и обзора патентной литературы материалов, порочащих новизну предлагаемого способа, не обнаружено.
Способ испытания побегов древесного растения включает в себя следующие действия.
На участке с древесной растительностью по внешним признакам выбирают учетные деревья. При этом взятие опытных образцов побегов проводят в конце вегетационного периода, чтобы были заметны все почки за прошлый вегетационный период (текущее слежение за ходом роста побега древесного растения значительно повышает трудоемкость измерений).
Лучше всего брать годичные побеги, а не многолетние, так как в этом случае ультразвуковыми волнами можно изучать точнее изменение скорости ультразвука вдоль побега, а также сравнивать между собой побеги с разных мест произрастания древесных растений (например, ивы по берегам водных объектов), имеющих различный уровень загрязнения воды и почвы.
Взятие образцов в виде годичных побегов проводят в конце вегетационного периода, например в конце сентября или в октябре месяце, когда годичные побеги выросли полностью в длину и образовались в течение вегетационного периода все почки на следующий вегетационный период.
У выбранного учетного дерева срезают опытные образцы в виде годичных или многолетних побегов около их основания, отмечают на комлевом конце каждого опытного побега номер пробной площадки, номер дерева и номер опытного образца, после этого срезанные и маркированные опытные побеги отправляют для лабораторных измерений.
При этом измерение геометрических параметров каждого опытного побега проводят путем измерения геометрического расположения почек и размерные параметры срезанных побегов, а после таких измерений разделяют опытный побег, начиная с комлевой части, на отрезки и эти отрезки озвучивают ультразвуком, по измеренным значениям геометрических параметров расположения почек на каждом опытном побеге статистическим моделированием выявляют закономерности распределения почек вдоль каждого побега и по этим закономерностям судят об экологическом качестве древесного растения и места его произрастания.
В лесу или на местности с древесной растительностью выбирают такое учетное дерево или группу кустов, которое позволяет многократно проводить экологические обследования и эксперименты с годичными побегами и в другие вегетационные периоды, а затем осматривают крону для возможного взятия побегов как опытных образцов.
Статистическим моделированием выявляют закономерность хода роста побега за вегетационное время по следующей статистической формуле:

где L - переменная расчетная длина побега от его основания, мм;
L0 - начальная (стартовая) длина побега до момента измерения, мм;
t - время с начала вегетационного периода, дни;
а1...а6 - параметры статистической модели.
Статистическим моделированием выявляют закономерность распределения почек и образования узлов почек по следующей формуле:
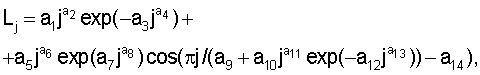
где Lj - переменная расчетная длина побега по узлам почек, мм;
j - номер узла почек, начиная от основания, причем j=1, 2, 3, ...;
а1...а14 - параметры статистической модели.
По найденным статистическим моделированием закономерностям судят об экологическом качестве древесного растения и места его произрастания по максимальной относительной погрешности найденных статистических моделей, причем чем меньше относительная погрешность, тем выше экологическое качество.
Способ испытания побегов древесного растения реализуется, например, на годичных побегах ивы, растущей по берегам водоемов и водотоков, следующим образом.
Опыты проводят в конце вегетационного периода, например в конце сентября или в октябре месяце, когда годичные побеги выросли полностью. Вначале на местности с ивняком закладывают пробную площадку, например, размерами 5×5 или 10×10 м. Пробные площадки закладывают в местах произрастания с различным уровнем загрязнения воды и почвы по берегам водных объектов.
На пробной площадке выбирают учетные деревца или кусты ивы, то есть такие деревья на пробной площадке, которые позволяют многократно проводить экологические обследования и эксперименты с годичными побегами и в другие вегетационные периоды.
Затем срезают опытные образцы годичных побегов около их основания, отмечают на комлевом конце (нижней стороне) побега номер пробной площадки, номер дерева и номер опытного образца. После этого срезанные и маркированные побеги отправляют для лабораторных измерений.
В камеральных условиях измеряют геометрическое расположение почек и размерные параметры срезанных побегов.
Статистическим моделированием выявляют закономерность хода роста побега по данным измерений расположения почек за вегетационное время по следующей статистической формуле:
где L - переменная расчетная длина побега от его основания, мм;
L0 - начальная (стартовая) длина побега до момента измерения, мм;
t - время с начала вегетационного периода, дни;
a1...a6 - параметры статистической модели.
Статистическим моделированием выявляют закономерность распределения почек и образования узлов почек вдоль годичного побега по следующей формуле:
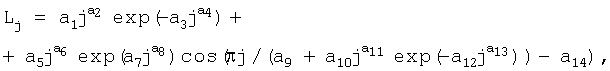
где Lj - переменная расчетная длина побега по узлам почек, мм;
j - номер узла почек, начиная от основания, причем j=1, 2, 3, ...;
a1...а14 - параметры статистической модели.
По найденным статистическим моделированием закономерностям судят об экологическом качестве древесного растения и места его произрастания по максимальной относительной погрешности найденных статистических моделей, причем чем меньше относительная погрешность, тем выше экологическое качество.
Примеры. Здесь приведены несколько примеров обработки данных, полученных измерениями еще в XIX веке М.Бюсгеном. Весь текстовый материал и исходные данные для статистического моделирования закономерностей изменения геометрических параметров побегов были приняты по книге: Бюсген М. Строение и жизнь наших лесных деревьев / Пер. с нем. - М. - Л.: Гослесбумиздат, 1961. - 424 с.
Геометрическое расположение почек на побеге является наиболее постоянным признаком растений. Его нарушение указывает на плохую экологическую обстановку на месте произрастания. Как правило, нижние почки сначала очень часто следуют одна за другой, затем расстояние между ними возрастает, а на верхушке побега снова уменьшается.
Часть стебля у годичного побега между почками называют термином междуузлие. Узел - это место расположения почки или листа. Расстояние между этими узлами измеряется с помощью миллиметровой линейки. Тогда статистические модели можно идентифицировать с получением значений параметров по последовательному изменению расстояния между узлами (прирост побега по длине) в зависимости от номера узлов. В дальнейшем возможно сопоставление с вегетационным периодом (по неделям или дням роста и развития годичного побега), а также с климатическими и метеорологическими данными (влажность, температура и освещенность солнечным светом).
Пример 1. Годичный побег терминальный (верхушечный у дерева). Измеренный М.Бюсгеном верхушечный побег имеет ход роста длины (высоты) по дням, причем за t=0 начала отсчета времени вегетационного периода принята дата 27 апреля. После идентификации измеренных данных годичного побега было получено уравнение (табл.1) вида
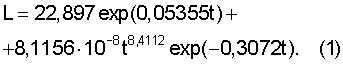
Ход роста верхушечного побега бука европейского (по [Бюсген, с.22]), мм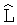
Первая составляющая показывает предысторию изучаемого процесса, в данном случае медленного повышения длины годичного побега еще до 27 апреля. Однако, в дальнейшем, эта составляющая экспоненциально нарастает. Таким образом, основной и естественной закономерностью хода роста становится экспоненциальная динамика длины побега. Стрессовое возбуждение растения на действие комплекса климатических факторов в вегетационный период характеризуется второй составляющей. Она получает максимальное значение роста 222,1 мм на дату 24 мая.
В табл.1 приняты следующие условные обозначения:
 - фактические (измеренные) значения переменной длины побега в ходе его роста за вегетационный период, мм;
- фактические (измеренные) значения переменной длины побега в ходе его роста за вегетационный период, мм;
L - расчетная по формуле (1) длина растущего побега, мм;
ε - остатки, не позволяющие в дальнейшем выявление других факторов (кроме времени), то есть абсолютная погрешность формулы (1) по разнице, вычисляемой как 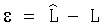 ;
;
Δ - относительная погрешность статистической модели, вычисляемая по далее неделимым остаткам ε с использованием математического выражения  .
.
Адекватность статистической модели (1) оценивается максимальной относительной погрешностью Δmax, значение которой в табл.1 подчеркнуто. В этом случае доверительная вероятность готовой статистической модели (1) вычисляется как разность D=100-|Δmax|. В данном примере доверие к формуле (1) будет равно не ниже 100-7,14=92,86%, что является достаточно высоким доверием к результатам экологических исследований.
Пример 2. Годичный побег угнетенный. Они расположены в тени кроны самого или другого дерева. Измерения миллиметровой линейкой выполняются аналогично верхушечному побегу.
Побег с минимальным ростом (угнетенный побег) описывается статистической моделью (табл.2).
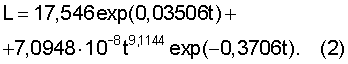
Ход роста отстающего побега бука европейского (по [Бюсген, с.22]), мм
Общая конструкция закономерности сохранилась, изменились только значения параметров модели. Теперь можем сопоставить две модели - терминального побега и отстающего в росте годичного побега. Все остальные побеги будут располагаться в середине этих крайних примеров.
Из первой составляющей видно, что уже из предыстории видна низкая активность хода роста слабого побега, которому, по-видимому, недостает многого (света, тепла и влаги) для быстрого роста. Причем активность стрессового возбуждения отстающего побега почти на порядок меньше. Это видно также и из данных таблиц.
Из расчетных данных видно, что терминальный побег за 42 дня роста по естественной закономерности вырастает от 22,9 до 253,6 мм, то есть в 11,07 раз. А слабый побег за это же время растет по закону экспоненциального роста всего 76,5/17,5=4,37 раз. При этом по второй составляющей максимум у побега с малым ростом достигает 37,0 мм в 25 дней, а у сильного побега максимум наблюдался 222,1 мм на 27-й день. Поэтому биотехническое возбуждение у слабого побега оказалось меньше в 222,1/37,0=5,98 раз.
Таким образом, измеряя дополнительно освещенность у угнетенных годичных побегов, можно оценить влияние условий произрастания каждого годичного побега на растущем дереве, деревце и кустарнике.
Пример 3. Годичный побег дерева ясеня. Приведем еще пример по ходу роста побега №1 у ясеня [Бюсген, с.23] при начале вегетационного периода с 18 апреля. После параметрической идентификации была получена конкретная закономерность (табл.3).
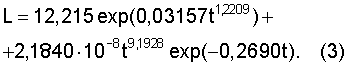
Здесь первая составляющая получила полное конструктивное насыщение до общей формы экспоненциального роста за счет интенсивности роста годичного побега, большей единицы. Поэтому у этого побега естественный рост происходит с ускорением.
Ход роста побега №1 ясеня (по [Бюсген, с.22]), мм
Можно предположить, что вторая составляющая также может достичь биотехнического закона в общей форме.
Поэтому ход роста годичного побега описывается общей моделью
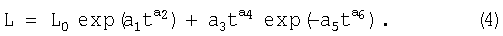
Пример 4. Распределение расстояния между почками на годичном побеге. Начиная с основания побега, измерялись миллиметровой линейкой расстояния между почками вдоль годичного побега, начиная от его основания [Бюсген, с.12]. Часть стебля между почками называется междуузлие. А место, где расположена почка или лист - это узел. У дерева ели узел, в котором по окружности ствола расположены несколько ветвей, получил название мутовка.
Расстояние между почками можно измерить тремя способами: 1) порядковым номером почек, начиная от основания годичного побега (что позволяет отказаться от измерений с помощью линейки); 2) расстоянием (длина) между почками, мм; 3) расстоянием (длиной) от основания до измеряемой почки, мм.
Длина lj между почками у побега граба обыкновенного изменяется, в зависимости от номера j узла, даваемого от основания годичного побега, по двухчленной формуле (табл.4).
lj=9,6183j1,8071ехр(-0,3028j0,9027)+
+acos(πj/p0.5-2,8458),
а=48,4233j-3,2790ехр(0,4711j),
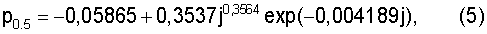
где а - амплитуда колебательного возмущения; p0.5 - половина длины волны (полупериод) волновой второй составляющей.
Изменение длины между почками на побеге граба обыкновенного, мм
Доверительная вероятность модели (5) составляет не ниже 80,49% (в экологических исследованиях допустимо доверие не ниже 70%). В статистическом моделировании, чтобы получить очень точные закономерности по линейным показателям, необходимо точность инструментальных измерений повысить почти на порядок (то есть не менее 10 раз). Здесь расположение почек измерено с округлением до одного миллиметра, а в новых экспериментах следует добиться точности измерений 0,1 мм.
Из формулы (5) видно, что амплитуда колебания изменяется по аномальной закономерности биотехнического закона, то есть образуется выпуклая кривая с минимальной амплитудой 2,2 мм. Это означает, что на седьмом узле наблюдалось минимальное воздействие колебательного возмущения извне (со стороны места произрастания) на появление почек на побеге. Половина периода колебания также изменяется криволинейно.
Причем дальнейшие вычислительные эксперименты показали, что частота колебания также меняется по аномальному биотехническому закону, а при этом полный период колебания составит 2 узла почек. Однако для точных выводов необходимы новые, более точные, замеры. Поэтому, в дальнейшем, частоту возмущения примем постоянной величиной.
Пример 5. Распределение расстояния между почками на годичном побеге. Один из побегов (закономерности по остальным примерам аналогичны) бузины красной по размещению почек описывается формулой (табл.5).

Изменение длины между почками на побеге бузины красной, мм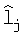
По сравнению с грабом обыкновенным у бузины красной появление почек завершается всего через 8 узлов. Поэтому высока точность модели, которая по максимальной относительной погрешности составляет 3,11%.
Минимум амплитуды 4,0 мм достигается уже на третьем узле, вместо 7-го узла у граба. А затем колебание быстро «идет в разнос» и «биологическая машина» по появлению почек повреждается и ломается. Поэтому значимость второй составляющей с 7-го узла резко нарастает. Однако приспособляемость побега к условиям внешней для побега бузины красной среды не может быть значительной (на 8-м узле коэффициент приспособляемости будет равен 39,0/6,0=6,5), поэтому организм прекращает процесс своего роста.
Пример 6. Распределение почек вдоль побега лещины. Годичный побег лещины обыкновенной резко выбрасывает от основания побега первую почку (или, может оказаться, первых почек просто не видно).
Поэтому второй коэффициент в биотехническом законе превращается в нуль и уравнение приобретает вид (табл.6).
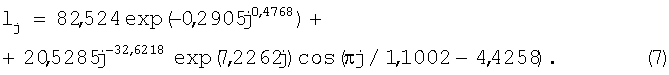
Поэтому для первой составляющей искомой закономерности из биотехнического закона y=ахb=0ехр(-cxd) получаем закон гибели в общей форме y=aexp(-схd).
Изменение длины между почками на побеге лещины обыкновенной, мм
Из данных табл.6 видно, что естественная составляющая показывает снижение расстояний между почками лещины обыкновенной. Волновая составляющая ведет себя оригинально: на узлах 2-9 она пропадает до нуля по амплитуде и полупериоду колебания, а с 10-го узла снова возникает, но уже с отрицательным знаком, вычитая естественные силы побега к росту.
Последовательное сложение длин междуузлий дает увеличение длины по номерам побегов. Поэтому кумулятивные графики хода роста предпочтительнее расстояний между почками, что покажем на примере.
Пример 7. Переменная длина побега по узлам почек. Последовательно просуммируем данные по годичному побегу граба обыкновенного. При этом номер узла превращается в кодовую шкалу переменных расстояний между узлами. Такой прием моделирования можно использовать, если даже не знаем переменного интервала между отдельными кодами (узлами), то есть нет результатов измерений миллиметровой линейкой.
Такой вариант реализации предлагаемого способа удобен для измерений в условиях школьных и студенческих экологических кружков, так как позволяет не следить экспериментально за динамикой роста побега, а измерять расстояния от основания срезанного побега до каждой последующей почки вдоль побега в камеральных условиях за один раз. Например, измерения проводятся миллиметровой линейкой метровой длины в конце вегетационного периода (например, в конце сентября или в октябре месяце, когда годичные побеги выросли полностью в длину и образовались в течение вегетационного периода все почки на следующий вегетационный период).
Кумулятивную кривую получим при условии 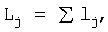 если были замерены расстояния между соседними почками (см. примеры 1-6). Если такие данные не были получены, то выполняют измерения метровой линейкой с миллиметровым делением от основания побега до каждой почки вдоль годичного побега.
если были замерены расстояния между соседними почками (см. примеры 1-6). Если такие данные не были получены, то выполняют измерения метровой линейкой с миллиметровым делением от основания побега до каждой почки вдоль годичного побега.
После идентификации была получена зависимость (табл.7).
Lj=9,7011j1,9132exp(-0,04824j1,1015)+
+acos(πj/p0.5-3,8270),
a=48,2150j-3,7640exp(0,5189j),
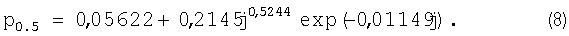
Получена очень точная статистическая модель (8), которая показывает вибрационную адаптацию годичного побега к окружающей древесное растение и отдельно измеренный побег среде по процессу выброса почек в отдельных узлах. Так как вегетационный цикл проходит всего за один сезон календарного года, то поиск влияния загрязнения окружающей среды на процесс выработки побегами почек становится хорошим и практичным методом в дендрометрии для нужд инженерной экологии.
Изменение длины побега по узлам почек граба обыкновенного, мм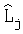
Общая закономерность образования узлов почек имеет вид:
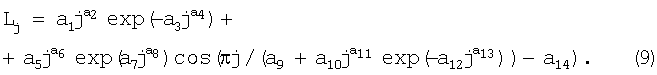
Если принять за нулевой ранг начало побега, то есть его основание на материнском растении, то коэффициент а14 показывает, что волновое изменение начинается гораздо раньше начала роста измеренного годичного побега. Минимальная длина волны равна 2,4 мм, а максимальная длина волны вначале старта в росте побега равна бесконечности. Половина периода 0,5 ранга достигается к четвертому рангу.
В нашем примере для граба обыкновенного получаем этот сдвиг, равным 3,8270×π/0,5=0,609, то есть колебание начинается от старта первой почки на побеге раньше не более чем на один ранг (период образования почки).
Из сравнения всех семи примеров видно, что проще реализовать предлагаемый способ на срезанных в конце вегетационного периода годичных побегах. Вместе с тем, слежение за ходом роста годичного побега в его динамике, причем без срезания нарастающего побега, позволяет на одном и том же древесном растении отбирать множество учетных годичных побегов. А это позволит организовать текущий мониторинг за ходом роста и развития и многолетних растений по отдельным группам годичных побегов.
Предлагаемый способ является универсальным для различных типов ландшафтов с древесной растительностью и лесных массивов вне зависимости от их возраста и позволяет реализовать физико-технологический подход к экологической, экономической и комплексной эколого-экономической оценки растущими деревьями территорий, а также сертифицировать качество ландшафтов и территорий по геометрическому расположению почек на годичных побегах.
Высокую практическую значимость приобретает применение способа на годичных побегах быстрорастущих древесных растений, произрастающих, например, по берегам водных объектов. Это позволяет наладить надежный, оперативный и в то же время простой в практической реализации, например в школьных лесничествах и экологических кружках, ежегодный экологический мониторинг территорий с различным уровнем загрязнения (например, измерением побегов ивы около свалок), а также за качеством водных и других природных объектов.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ИСПЫТАНИЯ РАСТИТЕЛЬНОГО ПОКРОВА НА УЧАСТКАХ ТРАССЫ ПРОДУКТОПРОВОДА | 2010 |
|
RU2473898C2 |
| СПОСОБ АНАЛИЗА КРОНЫ УЧЕТНОЙ ЕЛИ ПО ИСПЫТАНИЯМ ХВОИНОК ГОДИЧНЫХ ВЕТОЧЕК | 2011 |
|
RU2489717C2 |
| СПОСОБ АНАЛИЗА ВЕТВЕЙ КРОНЫ ДЕРЕВА ЕЛИ | 2011 |
|
RU2495417C2 |
| Способ анализа онтогенеза чистых листьев дерева | 2015 |
|
RU2615363C2 |
| СПОСОБ ИЗМЕРЕНИЯ ФОРМЫ ЛИСТЬЕВ У ДРЕВЕСНЫХ РАСТЕНИЙ | 2011 |
|
RU2477039C1 |
| СПОСОБ АНАЛИЗА СТВОЛА И КРОНЫ ДЕРЕВА ЕЛИ | 2008 |
|
RU2376749C1 |
| СПОСОБ ИЗМЕРЕНИЯ ДИНАМИКИ РОСТА ЛИСТЬЕВ ДЕРЕВА В ЧИСТЫХ ЭКОЛОГИЧЕСКИХ УСЛОВИЯХ | 2015 |
|
RU2597645C2 |
| СПОСОБ УЛЬТРАЗВУКОВОГО ИСПЫТАНИЯ ДРЕВЕСИНЫ РАСТУЩЕГО ДЕРЕВА НА КЕРНАХ | 2006 |
|
RU2327342C2 |
| Способ факторного анализа онтогенеза учетных листьев дерева | 2015 |
|
RU2606189C2 |
| СПОСОБ ИСПЫТАНИЯ ЕЛОВЫХ ХВОИНОК | 2009 |
|
RU2408184C1 |
Изобретение относится к области лесного и сельского хозяйств, в частности к экологической оценке лесных территорий различного возраста. Способ включает взятие опытных образцов в виде годичных побегов в конце вегетационного периода, например в конце сентября или в октябре месяце. Выбирают учетное древесное растение, срезают опытные образцы побегов около их основания, отмечают на комлевом конце каждого опытного побега номер дерева и номер опытного образца, после этого срезанные и маркированные опытные побеги отправляют для лабораторных измерений. При этом измерение геометрических параметров каждого опытного побега проводят путем измерения геометрического расположения почек на срезанных побегах. По измеренным значениям геометрических параметров расположения почек на каждом опытном побеге статистическим моделированием выявляют закономерность хода роста побега за вегетационное время по статистической формуле или закономерность распределения почек и образования узлов почек по соответствующим формулам и по максимальной относительной погрешности найденных статистических моделей судят об экологическом качестве древесного растения и месте его произрастания, причем чем меньше относительная погрешность, тем выше экологическое качество. Способ позволяет снизить трудоемкость измерений на побегах древесных растений. 2 з.п. ф-лы, 7 табл.
L=L0exp(a1ta2)+a3tа4exp(-а5tа6),
где L - переменная расчетная длина побега от его основания, мм;
L0 - начальная (стартовая) длина побега до момента измерения, мм;
t - время с начала вегетационного периода, дни;
a1...a6 - параметры статистической модели,
или закономерность распределения почек и образования узлов почек по следующей формуле:
Lj=a1jа2exp(-a3jа4)+a5ja6exp(a7ja8)cos(πj/(а9+a10jа11exp(-a12jа13))-а14,
где Lj - переменная расчетная длина побега по узлам почек, мм;
j - номер узла почек, начиная от основания, причем j=1, 2, 3, ...;
a1...a14 - параметры статистической модели,
и по максимальной относительной погрешности найденных статистических моделей судят об экологическом качестве древесного растения и месте его произрастания, причем чем меньше относительная погрешность, тем выше экологическое качество.
| СПОСОБ ИСПЫТАНИЯ ДРЕВЕСИНЫ РАСТУЩИХ ДЕРЕВЬЕВ | 1998 |
|
RU2144185C1 |
| ЛЕОНТЬЕВ Н.Л | |||
| Техника испытаний древесины | |||
| - М.: Лесная промышленность, 1970 | |||
| БЮСГЕН М | |||
| Строение и жизнь наших лесных деревьев | |||
| - М.: Гослесбумиздат, 1961 | |||
| РОДИН Л.Е., РЕМЕЗОВ Н.П., БАЗИЛЕВИЧ Н.Н | |||
| Методические указания к изучению динамики и биологического круговорота в фитоценозах | |||
| - Л.: Наука, 1968, с.9-12 | |||
| АНУЧИН Н.П | |||
| Лесная таксация | |||
| - М.: Гослесбумиздат, 1952. | |||

