ПРЕДПОСЫЛКА ИЗОБРЕТЕНИЯ
Область изобретения
Данное изобретение относится к областям молекулярной биологии, вирусологии, иммунологии и медицины. Изобретение относится к композиции, содержащей упорядоченную и повторяющуюся матрицу антигенов или антигенных детерминант и, в частности, матрицу белка RANKL, фрагмента RANKL или пептида RANKL. Более конкретно, изобретение относится к композиции, содержащей вирусоподобную частицу и по меньшей мере один связанный с ней белок RANKL, фрагмент RANKL или пептид RANKL. Изобретение также относится к способу получения конъюгатов и, соответственно, упорядоченных и повторяющихся матриц. Композиции согласно изобретению применимы для получения вакцин для лечения заболеваний кости и в качестве фармацевтической вакцины для профилактики или лечения заболеваний кости и для эффективной индукции иммунных ответов, в частности гуморальных ответов. Кроме того, композиции согласно изобретению особенно применимы для эффективной индукции специфичных для аутоантигенов иммунных ответов в указанном контексте.
Связанная область
Живая кость постоянно обновляется в результате сбалансированных и согласованных процессов ремоделирования. В основном два типа клеток вносят вклад в указанное ремоделирование: остеобласты необходимы для образования кости, тогда как остеокласты стимулируют распад костного матрикса и солюбилизацию гидроксиапатита. У молодых людей с растущими костями скорость образования кости превышает скорость резорбции кости, тогда как у более старших людей скорость резорбции может превышать образование и приводить к чистой потере минеральной плотности кости и/или костной массы. В последнем случае прочность костей ослабевает, и это приводит к повышенному риску переломов, а также медленному и неполному восстановлению сломанных костей. Известно, что множество состояний у человека связано с дисбалансом в ремоделировании костей.
В последнее время описаны три белка, которые являются ключевыми белками, вовлеченными в образование остеокластов из гематопоэтических клеток-предшественников и в регуляцию ремоделирования кости. RANKL (лиганд активатора рецептора NFkB), который также называют TNFSF11 (представитель 11 суперсемейства фактора некроза опухоли), TRANCE (TNF-родственный индуцируемый при активации цитокин), ODF (фактор дифференцировки остеокластов) или OPGL (лиганд остеопротегерина), является трансмембранным белком из 245 аминокислот, который образует гомотримеры. Часть внеклеточной области RANKL может быть удалена TACE-подобной протеазой. Кроме того, описаны варианты сплайсинга, в которых отсутствует трансмембранная область. Удаляемая часть RANKL содержит домен, который в высокой степени гомологичен TNF-α (Lum, L., et al., J. Biol. Chem. 274: 13613-13618 (2000)).
Способы получения белка RANKL и фрагментов RANKL описаны в WO 9846751, US 5843678, WO 98259958, US 6242586, WO 9828426, US 6242213, WO 9929865, JP 2000102390 и WO 0015807.
RANKL взаимодействует с трансмембранной молекулой на остеокластах, называемой RANK (активатор рецептора NFkB). Это взаимодействие приводит к активации предшественника остеокласта и заканчивается образованием активных резорбирующих кость остеокластов. In vivo растворимый рецептор-ловушка, называемый остеопротегерином, вовлечен в регуляцию остеокластогенеза благодаря его способности связываться с RANKL и ингибировать взаимодействие RANKL с его рецептором RANK. Указанное ингибирование приводит к подавлению остеокластогенеза и, таким образом, обеспечивает способ, позволяющий остановить чрезмерную резорбцию кости. Взаимодействие RANKL с его рецептором RANK можно подавить рекомбинантным остеопротегерином и растворимым слитым белком RANK-Fc. В соответствии с данными наблюдениями, у RANKL- и RANK-дефицитных мышей развивается остеопетроз, тогда как у сверхэкспрессирующих RANKL трансгенных мышей, а также у мышей с дефицитом остеопротегерина развивается остеопороз (Kong YY., et al., Nature 397: 315-322 (1999), Kim, N., et al., Proc. Natl. Acad. Sci USA 97: 10905-10910 (2000), Dougall, B., et al., Proc. Natl. Acad. Sci USA 97: 1566-1571 (1999), Bucay, N., et al., Genes Dev. 12: 1260-1268 (1998)).
Важное значение системы RANKL-RANK-остеопротегерин, кроме того, подтверждено в животной модели остеопороза на грызунах, индуцированного дефицитом эстрогенов. Рекомбинантный остеопротегерин полностью отменял индуцированную овариэктомией потерю костной ткани (Simonet, W.S., et al. Cell 89: 309-319 (1997).
В модели индуцированного адъювантом артрита инъекцией остеопротегерина можно было предотвратить потерю кости и деструкцию хряща, но не воспаление (опухание лапы). Помимо экспрессии на клетках стромы, RANKL также экспрессируется на T-клетках, и RANK обнаружен на антигенпрезентирующих клетках. Предполагается, что во время артритической реакции активированные T-клетки с повышенной экспрессией RANKL опосредуют усиление остеокластогенеза и последующую потерю кости. Взаимодействие RANKL с RANK также увеличивает долговечность и адъювантные свойства дендритных клеток (Kong Y.Y., et al., Nature 402: 304-309 (1999)).
Разрушение альвеолярной кости и последующая утрата зубов наблюдается при периодонтальных инфекциях. In vivo ингибирование функции остеопротегерином уменьшает деструкцию альвеолярной кости и уменьшает количество периодонтальных остеокластов после заражения микробами (Teng, Y.T.A., et al., J. Clin. Invest. 106: R59-R67 (2000).
Опухоли костей и некоторые опухолевые метастазы характеризуются повышенной резорбцией кости вследствие повышенного остеокластогенеза (Hofbauer, L.C. and Heufelder A.E., J. Clin. Endocrin. Met. 85: 2355-2363 (2000). Показано, что остеопротегерин ингибирует индуцированный раком простаты остеокластогенез и предотвращает рост опухоли простаты в костях мышей (Zhang Y., et al., J. Clin. Invest. 107:1219-1220 (2001). Он также уменьшает боль на поздних стадиях рака кости у мышей (Luger N.M., et al., Cancer Res. 61: 4038-4047 (2001)). Множественная миелома является злокачественным B-клеточным заболеванием, характеризуемым накоплением плазматических клеток в костном мозге и развитием остеолитического заболевания кости. В мышиных моделях множественной миеломы инъекция остеопротегерина или слитого белка RANK-Fc предотвращала развитие литических повреждений кости и препятствовала прогрессированию миеломы (Pearse RN., et al., Proc. Natl. Acad. Sci USA 98: 11581-11586 (2001).
Центральным в этиологии асептического ослабления имплантатов простаты является перипростатический остеолизис на границе кость-имплантат, который вызван воспалением, индуцированным частицами износа. Фибробластоподобные синовиоциты, трансфицированные остеопротегерином, были способны предотвращать индуцированный частицами износа остеокластогенез в мышиной модели (Gouter J.J., et al., J. Orthop. Res. 202:169-173 (2002)).
С высокой клинической частотой наблюдается кальцификация сосудов в популяции пациентов с остеопорозом. Участие системы RANKL-RANK-остеопротегерин показано посредством обнаружения того, что у мышей с дефицитом остеопротегерина наблюдалась кальцификация артерий, которая могла быть обратима с помощью рекомбинантного остеопротегерина (Min, H., et al., J. Exp. Med. 192: 463-474 (2000)).
Все указанные данные указывают на ключевое значение системы RANKL-RANK-остеопротегерин в регуляции резорбции кости при различных патологических состояниях. До настоящего времени ингибирование потери кости главным образом показано при инъекции рекомбинантного остеопротегерина или слитого белка RANK-Fc. Теоретически иммунизация животного RANKL должна обеспечивать продукцию RANKL-специфичных антител, которые посредством связывания с сайтом связывания RANK или стерического ингибирования должны препятствовать остеокластогенезу.
Однако до настоящего времени ничего не сообщалось о вакцинации белком или пептидом RANKL. Более того, не получено свидетельств того, что вакцины могут быть эффективными для защиты от заболеваний кости, так как, в частности, обычно трудно индуцировать гуморальные ответы на собственные молекулы в результате обычной вакцинации.
Одним из способов повышения эффективности вакцинации является увеличение степени повторяемости применяемого антигена. В отличие от изолированных белков вирусы индуцируют немедленные и эффективные иммунные ответы в отсутствие каких-либо адъювантов, как с помощью T-клеток, так и без нее (Bachmann and Zinkernagel, Ann. Rev. Immunol. 15: 235-270 (1991)). Хотя вирусы часто состоят из небольшого количества белков, они способны запускать намного более сильные иммунные ответы, чем их изолированные компоненты. В случае ответов B-клеток известно, что одним из ключевых факторов для иммуногенности вирусов является повторяемость и порядок поверхностных эпитопов. У многих вирусов обнаружена квазикристаллическая поверхность, на которой экспонирована регулярная матрица эпитопов, которая эффективно перекрестно связывает специфичные для эпитопов иммуноглобулины на B-клетках (Bachmann and Zinkernagel, Immunol. Today 17: 553-558 (1996)). Указанное перекрестное связывание поверхностных иммуноглобулинов на B-клетках является мощным сигналом активации, который непосредственно индуцирует прохождение клеточного цикла и продукцию IgM-антител. Кроме того, такие стимулированные B-клетки способны активировать хелперные T-клетки, которые, в свою очередь, индуцируют переключение с продукции IgM- на продукцию IgG-антител в B-клетках и образование долгоживущих B-клеток памяти - цель любой вакцинации (Bachmann and Zinkernagel, Ann. Rev. Immunol. 15:235-270 (1997)). Вирусная структура связана даже с образованием анти-антител, происходящим при аутоиммунном заболевании и являющимся частью естественного ответа на патогены (см. Fehr, T., et al., J Exp. Med. 185: 1785-1792 (1997)). Таким образом, антитела, презентированные на высокоорганизованной вирусной поверхности, способны индуцировать мощные ответы в виде анти-антител.
Однако, как указано, обычно иммунная система не может продуцировать антитела против структур собственного организма. В случае растворимых антигенов, присутствующих в низкой концентрации, это является следствием толерантности на уровне Th-клеток. При таких условиях связывание аутоантигена с носителем, который может обеспечивать T-помощь, может нарушить толерантность. Для растворимых белков, присутствующих в высоких концентрациях, или мембранных белков в низкой концентрации толерантными могут быть B- и Th-клетки. Однако B-клеточная толерантность может быть обратимой (анергия) и может быть нарушена введением антигена в высокоорганизованной форме, связанного с чужеродным носителем (Bachmann and Zinkernagel, Ann. Rev. Immunol. 15: 235-270 (1997)).
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Авторы обнаружили, что белки RANKL, фрагменты RANKL или пептиды RANKL, которые связаны с центральной частицей, имеющей структуру с присущей ей повторяющейся организацией и, таким образом, в частности с вирусоподобными частицами (VLP) и субъединицами VLP, соответственно, приводящими к образованию высокоупорядоченных и повторяющихся конъюгатов, представляют собой эффективные иммуногены для индукции антител, специфичных в отношении RANKL. Антитела способны, соответственно, блокировать и нейтрализовать взаимодействие RANKL с его рецептором RANK. Таким образом, данное изобретение относится к терапевтическому способу лечения заболеваний кости, который основан на упорядоченной и повторяющейся матрице RANKL-центральная частица, и, в частности, к VLP-RANKL-конъюгату и -матрице, соответственно. Указанное терапевтическое средство способно индуцировать высокие титры анти-RANKL-антител у вакцинированного животного.
Таким образом, данное изобретение относится к композиции, содержащей (a) центральную частицу по меньшей мере с одним первым сайтом связывания; и (b) по меньшей мере один антиген или антигенную детерминанту по меньшей мере с одним вторым сайтом связывания, где указанный антиген или антигенная детерминанта является белком RANKL, фрагментом RANKL или пептидом RANKL и где указанный второй сайт связывания выбран из группы, состоящей из (i) сайта связывания неприродного происхождения с указанным антигеном или антигенной детерминантой; и (ii) сайта связывания природного происхождения с указанным антигеном или антигенной детерминантой, где указанный второй сайт связывания способен к ассоциации с указанным первым сайтом связывания; и где указанный антиген или антигенная детерминанта и указанная центральная частица взаимодействуют посредством указанной ассоциации, образуя упорядоченную и повторяющуюся антигенную матрицу. Предпочтительными вариантами центральных частиц, подходящих для применения в данном изобретении, являются вирус, вирусоподобная частица, бактериофаг, бактериальная фимбрия или жгутик или любая другая центральная частица, имеющая присущую ей повторяющуюся структуру, способную образовывать упорядоченную и повторяющуюся антигенную матрицу согласно данному изобретению.
Более конкретно, изобретение относится к композиции, содержащей упорядоченную и повторяющуюся матрицу антигенов или антигенных детерминант, и, таким образом, в частности конъюгаты белок RANKL-, фрагмент RANKL- или пептид RANKL-VLP. Более конкретно, изобретение относится к композиции, содержащей вирусоподобную частицу и по меньшей мере один связанный с ней белок RANKL, фрагмент RANKL или пептид RANKL. Изобретение также относится к способу получения конъюгатов и упорядоченных и повторяющихся матриц, соответственно. Композиции согласно изобретению применимы для получения вакцин для лечения заболеваний кости и в качестве фармацевтических вакцин для профилактики или лечения заболеваний кости и для эффективной индукции иммунных ответов, в частности гуморальных ответов. Кроме того, композиции согласно изобретению особенно применимы для эффективной индукции специфичных для аутоантигенов иммунных ответов в указанном контексте.
В данном изобретении белок RANKL, фрагмент RANKL или пептид RANKL обычно связан с центральной частицей и, соответственно, с VLP ориентированным образом, образуя упорядоченную и повторяющуюся матрицу антигенов белка RANKL, фрагмента RANKL или пептида RANKL. Кроме того, высокоповторяющаяся и организованная структура центральных частиц и, соответственно, VLP опосредует экспонирование белка RANKL, фрагмента RANKL или пептида RANKL высокоупорядоченным и повторяющимся образом, приводя к образованию высокоорганизованной и повторяющейся антигенной матрицы. Кроме того, связывание белка RANKL, фрагмента RANKL или пептида RANKL с центральной частицей и, соответственно, с VLP обеспечивает эпитопы хелперных T-клеток, так как центральная частица и VLP являются чужеродными по отношению к хозяину, иммунизируемому матрицей центральная частица-белок RANKL, -фрагмент RANKL или -пептид RANKL и, соответственно, VLP-белок RANKL, -фрагмент RANKL или -пептид RANKL. Указанные матрицы отличаются от конъюгатов предшествующего уровня техники своей высокоорганизованной структурой, размерами и повторяемостью антигена на поверхности матрицы.
В одном аспекте изобретения белок RANKL, фрагмент RANKL или пептид RANKL экспрессируют в подходящем хозяине экспрессии, совестимом с правильной укладкой белка RANKL или фрагмента RANKL, или синтезируют, тогда как центральную частицу и, соответственно, VLP, экспрессируют и очищают из экспрессирующего хозяина, подходящего для укладки и сборки центральной частицы и, соответственно, VLP. Белок RANKL, фрагмент RANKL или пептид RANKL также можно синтезировать химическим способом. Затем собирают матрицу белка RANKL, фрагмента RANKL или пептида RANKL посредством связывания белка RANKL, фрагмента RANKL или пептида RANKL с центральной частицей и, соответственно, с VLP.
В другом аспекте данное изобретение относится к композиции, содержащей (a) вирусоподобную частицу и (b) по меньшей мере один антиген или антигенную детерминанту, где указанный антиген или указанная антигенная детерминанта является белком RANKL, фрагментом RANKL или пептидом RANKL и где указанный по меньшей мере один антиген или антигенная детерминанта связаны с вирусоподобной частицей.
В следующем аспекте данное изобретение относится к фармацевтической композиции, содержащей (a) композицию по п.1 или п.22 и (b) приемлемый фармацевтический носитель.
В еще одном аспекте данное изобретение относится к композиции вакцины, содержащей композицию, включающую в себя (a) центральную частицу по меньшей мере с одним первым сайтом связывания; и (b) по меньшей мере один антиген или антигенную детерминанту по меньшей мере с одним вторым сайтом связывания, где указанный антиген или антигенная детерминанта является белком RANKL, фрагментом RANKL или пептидом RANKL и где указанный второй сайт связывания выбран из группы, состоящей из (i) сайта связывания неприродного происхождения с указанным антигеном или антигенной детерминантой; и (ii) сайта связывания природного происхождения с указанным антигеном или антигенной детерминантой, где указанный второй сайт связывания способен к ассоциации с указанным первым сайтом связывания; и где указанный антиген или антигенная детерминанта и указанная центральная частица взаимодействуют посредством указанной ассоциации, образуя упорядоченную и повторяющуюся антигенную матрицу.
В следующем аспекте данное изобретение относится к композиции вакцины, содержащей композицию, где указанная композиция содержит (a) вирусоподобную частицу; и (b) по меньшей мере один антиген или антигенную детерминанту, где указанный антиген или указанная антигенная детерминанта является белком RANKL, фрагментом RANKL или пептидом RANKL; и где указанный по меньшей мере один антиген или антигенная детерминанта связаны с указанной вирусоподобной частицей.
Еще в одном аспекте данное изобретение относится к способу получения композиции по п.1, включающему в себя (a) получение вирусоподобной частицы; и (b) получение по меньшей мере одного антигена или антигенной детерминанты, где указанный антиген или указанная антигенная детерминанта является белком RANKL, фрагментом RANKL или пептидом RANKL; (c) объединение указанной вирусоподобной частицы и указанного по меньшей мере одного антигена или антигенной детерминанты, так чтобы указанный по меньшей мере один антиген или антигенная детерминанта были связаны с указанной вирусоподобной частицей.
Еще в одном аспекте данное изобретение относится к способу получения композиции по п.22, включающему в себя (a) получение центральной частицы по меньшей мере с одним первым сайтом связывания; (b) получение по меньшей мере одного антигена или антигенной детерминанты по меньшей мере с одним вторым сайтом связывания, где указанный антиген или антигенная детерминанта является белком RANKL, фрагментом RANKL или пептидом RANKL и где указанный второй сайт связывания выбран из группы, состоящей из (i) сайта связывания неприродного происхождения с указанным антигеном или антигенной детерминантой; и (ii) сайта связывания природного происхождения с указанным антигеном или антигенной детерминантой; и где указанный второй сайт связывания способен к ассоциации с указанным первым сайтом связывания; и (c) объединение указанной центральной частицы и указанного по меньшей мере одного антигена или антигенной детерминанты, где указанный антиген или антигенная детерминанта и указанная центральная частица взаимодействуют путем указанной ассоциации, образуя упорядоченную и повторяющуюся антигенную матрицу.
В другом аспекте данное изобретение относится к способу иммунизации, включающему в себя введение композиции по п.1 или п.22 животному или человеку.
В следующем аспекте данное изобретение относится к применению композиции по п.1 или п.22 для производства лекарственного средства для лечения заболеваний кости.
Еще в одном аспекте данное изобретение относится к применению композиции по п.1 или п.22 для приготовления лекарственного средства для терапевтического или профилактического лечения заболеваний кости, предпочтительно энцефалопатий млекопитающих. Кроме того, еще в одном аспекте данное изобретение относится к применению композиции по п.1 или п.22 либо отдельно, либо в комбинации с другими агентами, для производства композиции, вакцины, лекарственного средства или медицинского препарата для терапии или профилактики заболеваний кости, в частности энцефалопатий млекопитающих, и/или для стимуляции иммунной системы млекопитающего.
Таким образом, изобретение, в частности, относится к композициям вакцин, которые подходят для профилактики и/или ослабления заболеваний кости или связанных с ними состояний. Изобретение, кроме того, относится к способам иммунизации и вакцинации, соответственно, для профилактики и/или ослабления заболеваний кости и связанных с ними состояний у животных, в частности у коров, овец и крупного рогатого скота, а также у человека. Композиции согласно изобретению можно использовать профилактически или терапевтически.
В конкретных вариантах изобретение относится к способам профилактики и/или ослабления заболеваний кости или связанных с ними состояний, которые вызваны или обострены продуктами «собственных» генов, т.е. в используемом в данном описании смысле «аутоантигенами». В родственных вариантах изобретение относится к способам индуцирования иммунологических ответов у животных и человека, соответственно, которые приводят к продукции антител, которые предотвращают и/или ослабляют заболевания кости или связанные с ними состояния, которые вызваны или обострены продуктами «собственных» генов.
Как будет понятно специалисту в данной области, когда композиции согласно изобретению вводят животному или человеку, они могут быть в композиции, которая содержит соли, буферные вещества, адъюванты или другие вещества, которые требуются для повышения эффективности композиции. Примеры веществ, подходящих для применения при получении фармацевтических композиций, приведены во многих источниках, включая Remington's Pharmaceutical Sciences (Osol, A, ed., Mack Publishing Co. (1990)).
Говорят, что композиции согласно изобретению являются «фармакологически приемлемыми», если может быть допустимо их введение человеку-реципиенту. Кроме того, композиции согласно изобретению будут вводиться в «терапевтически эффективном количестве» (т.е. в количестве, которое дает требуемый физиологический эффект).
Композиции согласно изобретению можно вводить различными способами, известными в данной области, но обычно они будут вводиться путем инъекции, инфузии, ингаляции, перорального введения или другими подходящими физическими способами. Альтернативно композиции можно вводить внутримышечно, внутривенно или подкожно. К компонентам композиций для введения относятся стерильные водные растворы (например, физиологический раствор соли) или неводные растворы и суспензии. Примерами неводных растворителей являются пропиленгликоль, полиэтиленгликоль, растительные масла, такие как оливковое масло, и инъекционные органические сложные эфиры, такие как этилолеат. Можно использовать носители или окклюзионные повязки, чтобы увеличить проницаемость кожи и усилить абсорбцию антигена.
Другие варианты данного изобретения будут очевидны для специалиста в свете того, что известно в данной области, следующих чертежей и описания изобретения и формулы изобретения.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг.1 показана экспрессия и очистка C-RANKL. Очистку C-RANKL анализировали в SDS-геле в восстанавливающих условиях. Гель красили Кумасси бриллиантовым голубым. Молекулярные массы маркерных белков приведены на левом поле. Дорожка 1: маркер низкой молекулярной массы. Дорожка 2 и 3: надосадок клеточных лизатов клеток BL21/DE3, трансформированных пустым вектором pGEX6p1 и pGEX-RANKL, соответственно, после шестнадцати часов индукции с помощью IPTG 0,4 мМ. Дорожка 4: очищенный белок GST-PS-C-RANKL после колонки FF, улавливающей GST. Дорожка 5: Фракция, не связанная с колонкой FF, улавливающей GST. Дорожка 6: очищенный белок GST-PS-C-RANKL после расщепления протеазой PreScission. Дорожка 7: несвязанная фракция с колонки FF, улавливающей GST, на которую наносили продукт расщепления GST-RANKL, которая содержит очищенный C-RANKL. Дорожка 8: связанная фракция с колонки FF, улавливающей GST, на которую наносили продукт расщепления GST-PS-C-RANKL и элюировали с помощью GSH.
На фиг.2 показана экспрессия и очистка RANKL-C.
На фиг.2A показана очистка GST-EK-RANKL-C. Образцы белков анализировали в SDS-ПААГ в восстанавливающих условиях. Гель красили Кумасси бриллиантовым голубым. Молекулярные массы маркерных белков приведены на левом поле. Дорожка 1: предварительно окрашенный белковый маркер для широкого диапазона (New England Biolabs). Дорожка 2: осветленный клеточный лизат клеток BL21/DE3, трансформированных плазмидой pMod-GST-EK-mRANKL-C1 после индукции в течение ночи с использованием 0,1 мМ IPTG. Дорожка 3: поток, проходящий через колонку FF, улавливающую GST, нагруженную осветленным лизатом с дорожки 2. Дорожка 4: первая промывка колонки FF, улавливающей GST. Дорожка 5: вторая промывка колонки FF, улавливающей GST. Дорожка 6: третья промывка колонки FF, улавливающей GST. Дорожки 7-15: элюированные фракции 1-9 с колонки FF, улавливающей GST, содержащие очищенный слитый белок GST-EK-RANKL-C и небольшое количество белка GST-EK.
На фиг.2B показано расщепление GST-EK-RANKL-C энтерокиназой MaxTM.
Расщепление GST-EK-RANKL-C анализировали в SDS-ПААГ в восстанавливающих условиях. Гель красили Кумасси бриллиантовым голубым. Молекулярные массы маркерных белков приведены на левом поле. Дорожка 1: предварительно окрашенный белковый маркер для широкого диапазона (New England Biolabs). Дорожка 2: Очищенный слитый белок GST-EK-RANKL-C. Дорожка 3: продукты расщепления после 16 час инкубации при 4°C с энтерокиназой MaxTM.
На фиг.2C показана очистка RANKL-C.
Очистку RANKL-C после удаления GST-EK методом аффинной хроматографии на глутатион-сефарозе анализировали в SDS-ПААГ в восстанавливающих условиях. Гель красили Кумасси бриллиантовым голубым. Молекулярные массы маркерных белков приведены на левом поле. Дорожка 1: предварительно окрашенный белковый маркер для широкого диапазона (New England Biolabs). Дорожка 2 и 3: продукты расщепления GST-EK и RANKL-C после 16 час инкубации GST-EK-RANKL-C при 4°C с энтерокиназой MaxTM. Дорожка 4 и 5: различные количества несвязанной фракции с колонки FF, улавливающей GST, которая содержит белок RANKL-C высокой чистоты.
На фиг.3 показано связывание C-RANKL с капсидным белком Qβ.
На фиг.3A показан SDS-ПААГ-анализ продуктов связывания: белки анализировали в 16% SDS-гелях в восстанавливающих условиях. Гель красили Кумасси бриллиантовым голубым. Молекулярные массы маркерных белков приведены на левом поле. Идентичность полос белка показана на правом поле. Дорожка 1: предварительно окрашенный белковый маркер для широкого диапазона (New England Biolabs). Дорожка 2: дериватизованный капсидный белок Qβ. Дорожка 3: очищенный белок C-RANKL. Дорожка 4: реакция связывания C-RANKL/Qβ.
Фиг.3B и фиг.3C: Вестерн-блот-анализ продуктов связывания. Белки разгоняли в 16% SDS-гелях в восстанавливающих условиях, подвергали блоттингу на нитроцеллюлозные мембраны и регистрировали с помощью анти-Qβ-антисыворотки (фиг.3B) или анти-RANKL-антитела (фиг.3C). Молекулярные массы маркерных белков приведены на левом поле. Идентичность белковых полос указана на правом поле. Дорожка 1: предварительно окрашенный белковый маркер для широкого диапазона (New England Biolabs). Дорожка 2: дериватизованный капсидный белок Qβ. Дорожка 3: очищенный белок C-RANKL. Дорожка 4: реакция связывания C-RANKL/Qβ.
На фиг.4 показан ELISA для RANKL-специфичного IgG у мышей, иммунизированных C-RANKL, связанным с Qβ.
Самок мышей Balb/c вакцинировали подкожно 25 мкг C-RANKL, связанного с Qβ, в PBS в 0 день, 16 день и 64 день с добавлением или без добавления квасцов. Сыворотку, полученную в 0, 16, 23, 64 и 78 дни, анализировали в отношении антител, специфичных для RANKL. Титры ELISA выражали в виде среднего для тех разведений сывороток, которые давали половину максимальной OD420 в анализе ELISA.
На фиг.5 показана нейтрализующая активность антител, индуцированных у мышей, иммунизированных C-RANKL, связанным с Qβ.
На фиг.5A показан анализ связывания C-RANKL и его родственного лиганда RANK. Планшеты для ELISA покрывали 10 мкг/мл C-RANKL и инкубировали с серийными разведениями слитого белка RANK-Fc или неродственного слитого с Fc белка. Регистрацию связанного RANK осуществляли с помощью конъюгированных с HRP анти-Fc-антител.
На фиг.5B показано ингибирование связывания C-RANKL/RANK-Fc антителами сыворотки мышей, вакцинированных C-RANKL, связанным с Qβ. Планшеты для Elisa покрывали 10 мкг/мл C-RANKL и подвергали совместной инкубации с серийными разведениями сывороток мышей, полученных на 78 день, и 1 нМ слитого белка RANK-Fc. Связывание слитого белка с C-RANKL регистрировали с использованием конъюгированного с пероксидазой хрена анти-Fc-антитела.
На фиг.6A-C изображена очистка белков AP205 для применения в VLP, которую анализировали с помощью SDS-ПААГ и Вестерн-блоттинга.
На фиг.7A-B изображены электронные микрофотографии сравнения фаговых частиц AP205 с вирусоподобными частицами AP205, спонтанно собираемыми из рекомбинантного белка, экспрессированного в E. coli и очищенного.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Если не оговорено особо, все технические и научные термины, используемые в данном описании, имеют такие же значения, которые обычно подразумеваются специалистами в области, к которой относится данное изобретение. Хотя на практике или при проверке данного изобретения можно использовать любые способы и вещества, сходные или эквивалентные способам и веществам, описанным в данной заявке, предпочтительные способы и вещества описаны ниже.
1. Определения:
Аминокислотный линкер: «аминокислотный линкер» или называемый также в данном описании «линкер» в используемом смысле либо связывает антиген или антигенную детерминанту со вторым сайтом связывания, либо - более предпочтительно - уже содержит или включает в себя второй сайт связывания, обычно - но не обязательно - в виде аминокислотного остатка, предпочтительно в виде остатка цистеина. Однако термин «аминокислотный линкер» в используемом в данном описании смысле не предназначен для обозначения того, что такой аминокислотный линкер состоит исключительно из аминокислотных остатков, хотя аминокислотный линкер, состоящий из аминокислотных остатков, является предпочтительным вариантом согласно данному изобретению. Остатки аминокислот аминокислотного линкера предпочтительно состоят из аминокислот природного происхождения или неприродных аминокислот, известных в данной области, всех L или всех D или их смесей. Однако аминокислотный линкер, содержащий молекулу с сульфгидрильной группой или остаток цистеина, также входит в данное изобретение. Такая молекула предпочтительно содержит остаток C1-C6-алкила, циклоалкила (C5,C6), арила или гетероарила. Однако? кроме аминокислотного линкера, в объем данного изобретения также следует включить линкер, предпочтительно содержащий остаток C1-C6-алкила, циклоалкила (C5,C6), арила или гетероарила и не имеющий никакой аминокислоты (аминокислот). Связь между антигеном или антигенной детерминантой или необязательно вторым сайтом связывания и аминокислотным линкером предпочтительно осуществляется с помощью по меньшей мере одной ковалентной связи, более предпочтительно с помощью по меньшей мере одной пептидной связи.
Животные: В используемом в данном описании смысле подразумевается, что термин «животное» включает в себя, например, человека, овец, лосей, оленей, оленей-мулов, норок, млекопитающих, обезьян, лошадей, крупный рогатый скот, свиней, коз, собак, кошек, крыс, мышей, птиц, кур, рептилий, рыб, насекомых и паукообразных.
Антитело: В используемом в данном описании смысле термин «антитело» относится к молекулам, которые способны связывать эпитоп или антигенную детерминанту. Подразумевается, что термин включает в себя целые антитела и их антигенсвязывающие фрагменты, включая одноцепочечные антитела. Наиболее предпочтительно антитела являются антигенсвязывающими фрагментами антител человека и включают в себя, не ограничиваясь указанным, Fab, Fab' и F(ab')2, Fd, одноцепочечные Fv (scFv), одноцепочечные антитела, связанные дисульфидными связями Fv (sdFv) и фрагменты, содержащие либо VL-, либо VH-домен. Источником антител может быть любое животное, включая птиц и млекопитающих. Предпочтительно антитела получены от человека, мыши, кролика, козы, морской свинки, верблюда, лошади или цыпленка. В используемом в данном описании смысле «человеческие» антитела включают в себя антитела, имеющие аминокислотную последовательность иммуноглобулина человека, и включают антитела, выделенные из библиотек иммуноглобулинов человека или из животных, трансгенных по одному или нескольким иммуноглобулинам человека и которые не экспрессируют эндогенных иммуноглобулинов, как описано, например, в патенте США № 5939598 Kucherlapati et al.
Антиген: В используемом в данном описании смысле «антиген» относится к молекуле, способной подвергаться связыванию антителом или рецептором T-клеток (TCR), если она презентирована молекулами MHC. Термин «антиген» в используемом в данном описании смысле также охватывает T-клеточные эпитопы. Кроме того, антиген может узнаваться иммунной системой и/или способен индуцировать гуморальный иммунный ответ и/или клеточный иммунный ответ, приводящий к активации B- и/или T-лимфоцитов. Однако это может требовать, по меньшей мере в некоторых случаях, чтобы антиген содержал или был связан с Th-клеточным эпитопом и был введен в адъюванте. Антиген может иметь один или несколько эпитопов (B- и T-эпитопов). Подразумевается, что указанная выше специфичная реакция указывает на то, что антиген будет предпочтительно реагировать, обычно высокоизбирательным образом, с соответствующим ему антителом или TCR и не будет реагировать со множеством других антител или TCR, которые могут быть вызваны другими антигенами. В используемом в данном описании смысле антигены также могут представлять собой смеси нескольких отдельных антигенов.
Антигенная детерминанта: В используемом в данном описании смысле подразумевается, что термин «антигенная детерминанта» относится к такой части антигена, которая специфично распознается либо B-, либо T-лимфоцитами. B-лимфоциты, отвечающие на антигенные детерминанты, продуцируют антитела, тогда как T-лимфоциты отвечают на антигенные детерминанты пролиферацией и установлением эффекторных функций, необходимых для опосредования клеточного и/или гуморального иммунитета.
Ассоциация: В используемом в данном описании смысле термин «ассоциация» в применении к первому и второму сайтам связывания относится к связыванию первого и второго сайтов связывания, которое предпочтительно осуществляется посредством по меньшей мере одной непептидной связи. Природа ассоциации может быть ковалентной, ионной, гидрофобной, полярной или любой их комбинацией, предпочтительно природа ассоциации является ковалентной.
Сайт связывания, первый: В используемом в данном описании смысле фраза «первый сайт связывания» относится к элементу неприродного или природного происхождения, с которым может вступать в ассоциацию второй сайт связывания, расположенный на антигене или антигенной детерминанте. Первый сайт связывания может быть белком, полипептидом, аминокислотой, пептидом, сахаром, полинуклеотидом, природным или синтетическим полимером, вторичным метаболитом или соединением (биотин, флуоресцеин, ретинол, дигоксигенин, ионы металлов, фенилметилсульфонилфторид) или их комбинацией или их химически реакционно-способной группой. Первый сайт связывания обычно и предпочтительно расположен на поверхности центральной частицы, такой как предпочтительно вирусоподобная частица. Многочисленные первые сайты связывания обычно присутствуют на поверхности центральной частицы и, соответственно, вирусоподобной частицы в повторяющейся конфигурации.
Сайт связывания, второй: В используемом в данном описании смысле фраза «второй сайт связывания» относится к элементу, находящемуся в ассоциации с антигеном или антигенной детерминантой, с которым может вступать в ассоциацию первый сайт связывания, расположенный на поверхности центральной частицы и, соответственно, вирусоподобной частицы. Второй сайт связывания антигена или антигенной детерминанты может быть белком, полипептидом, пептидом, сахаром, полинуклеотидом, природным или синтетическим полимером, вторичным метаболитом или соединением (биотин, флуоресцеин, ретинол, дигоксигенин, ионы металлов, фенилметилсульфонилфторид) или их комбинацией или их химически реакционно-способной группой. На антигене или антигенной детерминанте присутствует по меньшей мере один второй сайт связывания. Термин «антиген или антигенная детерминанта по меньшей мере с одним вторым сайтом связывания», таким образом, относится к антигену или антигенной конструкции, содержащей по меньшей мере антиген или антигенную детерминанту и второй сайт связывания. Однако, в частности, в случае второго сайта связывания, который имеет неприродное происхождение, т.е. не естественным образом возник в антигене или антигенной детерминанте, указанные антиген или антигенные конструкции содержат «аминокислотный линкер».
Заболевания кости: Термин «заболевания кости» в используемом в данном описании смысле охватывает, в частности, состояния, характеризующиеся повышенной резорбцией кости. Термин «заболевания кости» включает в себя, но не ограничен указанным, остеопороз в его различных формах, такой как первичный остеопороз (такой как идиопатический, постменопаузный, инволюционный или старческий остеопороз) и вторичный остеопороз. Последний включает в себя остеопороз, вызванный избытком глюкокортикоидов, гиперпаратиреоидизмом, гипертиреоидизмом, гипергонадизмом, гиперпролактинемией, сахарным диабетом, остеопороз, индуцированный лекарственными средствами, такой как остеопороз, вызванный глюкокортикостероидами, этанолом, дилантином, табаком, барбитуратами или гепарином), остеопороз, индуцированный прекращением приема лекарственных средств, и разнообразные состояния, связанные с повышенной резорбцией кости, такие как хроническая почечная недостаточность, болезнь печени, синдромы мальабсорбции, хроническое обструктивное легочное заболевание, саркоидоз. Следующие состояния с повышенной резорбцией кости включают в себя болезнь Пэджета, семейный распространяющийся остеолизис, спонтанный остеолизис, потерю кости, связанную с ревматоидным артритом, резорбцию кости и потерю зубов при периодонтальном заболевании, остеомиелит, гиперкальцемию при злокачественных опухолях, опухоли кости, злокачественные опухоли, связанные с метастазами в костях, и потерю кости, вызванную потерей массы, которая обнаружена при космическом полете. Специалисты в данной области могут идентифицировать другие состояния, характеризующиеся повышенной резорбцией кости.
Связь: В используемом в данном описании смысле термин «связь» относится к связыванию или присоединению, которое может быть ковалентным, например посредством химического связывания, или нековалентным, например ионные взаимодействия, гидрофобные взаимодействия, водородные связи и т.д. Ковалентные связи, например, могут быть сложноэфирными, эфирными, фосфоэфирными, амидными, пептидными, имидными связями, связями углерод-сера, связями углерод-фосфор и тому подобными. Термин «связанный» является более широким и включает в себя такие термины, как «соединенный», «слитый» и «прикрепленный».
Покрывающий белок (белки): В используемом в данном описании смысле термин «покрывающий белок (белки)» относится к белку (белкам) бактериофага или РНК-фага, которые могут быть включены в сборку капсида бактериофага или РНК-фага. Однако по отношению к специфичному генному продукту гена покрывающего белка РНК-фага используют термин «CP». Например, специфичный генный продукт гена покрывающего белка РНК-фага Qβ называют «CP Qβ», тогда как «покрывающие белки» бактериофага Qβ содержат «CP Qβ», а также белок A1. Капсид бактериофага Qβ главным образом состоит из CP Qβ с небольшим содержанием белка A1. Подобным образом, покрывающий белок VLP Qβ главным образом содержит CP Qβ с небольшим содержанием белка A1.
Центральная частица: В используемом в данном описании смысле термин «центральная частица» относится к жесткой структуре с присущей ей повторяющейся организацией. Центральная частица в используемом в данном описании смысле может быть продуктом процесса синтеза или продуктом биологического процесса.
Связанный: Термин «связанный» в используемом в данном описании смысле относится к связыванию посредством ковалентных связей или сильных нековалентных взаимодействий, обычно и предпочтительно к связыванию посредством ковалентных связей. В данном изобретении можно использовать любой способ, обычно используемый специалистами в данной области для связывания биологически активных веществ.
Эффективное количество: В используемом в данном описании смысле термин «эффективное количество» относится к количеству, необходимому или достаточному, чтобы осуществить требуемое биологическое действие. Эффективным количеством композиции может быть количество, которое дает возможность достичь данного выбранного результата, и определение такого количества может быть обычной практикой для специалиста в данной области. Например, эффективным количеством для лечения недостаточности иммунной системы может быть количество, необходимое для того, чтобы вызвать активацию иммунной системы, приводящую к развитию специфичного для антигена иммунного ответа при воздействии антигена. Этот термин также является синонимом «достаточного количества».
Эффективное количество для любого конкретного применения может варьировать в зависимости от таких факторов, как заболевание или состояние, подвергаемое лечению, конкретная вводимая композиция, масса субъекта и/или тяжесть заболевания или состояния. Специалист в данной области может эмпирически определить эффективное количество конкретной композиции согласно данному изобретению без необходимости в чрезмерном экспериментировании.
Эпитоп: В используемом в данном описании смысле «эпитоп» относится к непрерывным или прерывающимся частям полипептида, обладающим антигенной или иммуногенной активностью у животного, предпочтительно млекопитающего и наиболее предпочтительно человека. Эпитоп распознается антителом или T-клеткой посредством его T-клеточного рецептора в контексте молекулы MHC. «Иммуногенный эпитоп» в используемом в данном описании смысле определяют как часть полипептида, которая вызывает гуморальный ответ или индуцирует T-клеточный ответ у животного, который определяется любым способом, известным в данной области (см., например, Geysen et al., Proc. Natl. Acad. Sci. USA 81: 3998-4002 (1983)). Термин «антигенный эпитоп» в используемом в данном описании смысле определяют как часть белка, посредством которой антитело может иммуноспецифически связывать свой антиген, что определяют любым способом, хорошо известным в данной области. Иммуноспецифическое связывание исключает неспецифическое связывание, но не обязательно исключает перекрестную реакционную способность по отношению к другим антигенам. Антигенные эпитопы не обязательно должны быть иммуногенными. Антигенные эпитопы также могут быть T-клеточными эпитопами, и в этом случае они могут иммуноспецифически связываться T-клеточным рецептором в контексте молекулы MHC.
Эпитоп может содержать 3 аминокислоты в пространственной конформации, которая уникальна для эпитопа. Как правило, эпитоп состоит по меньшей мере примерно из 5 таких аминокислот и более обычно состоит по меньшей мере примерно из 8-10 таких аминокислот. Если эпитоп является органической молекулой, он может быть небольшим, таким как нитрофенил.
Слияние: В используемом в данном описании смысле термин «слияние» относится к комбинации аминокислотных последовательностей разного происхождения в одной полипептидной цепи посредством комбинирования в рамке считывания кодирующих их нуклеотидных последовательностей. Термин «слияние» особо включает в себя внутренние слияния, т.е. инсерцию последовательностей разного происхождения в полипептидную цепь, кроме слияния с одним из ее концов.
Иммунный ответ: В используемом в данном описании смысле термин «иммунный ответ» относится к гуморальному иммунному ответу и/или клеточному иммунному ответу, приводящему к активации или пролиферации B- и/или T-лимфоцитов и/или антигенпрезентирующих клеток. Однако в некоторых случаях иммунные ответы могут быть низкой интенсивности, и они становятся регистрируемыми только при использовании по меньшей мере одного вещества согласно изобретению. «Иммуногенный» относится к агенту, используемому для стимуляции иммунной системы живого организма с тем, чтобы усилить и направить на иммуногенный агент одну или несколько функций иммунной системы. «Иммуногенный полипептид» является полипептидом, который вызывает клеточный и/или гуморальный иммунный ответ либо отдельно, либо в связанном с носителем виде в присутствии или в отсутствие адъюванта. Предпочтительно может быть активирована антигенпрезентирующая клетка.
Вещество, которое «усиливает» иммунный ответ, относится к веществу в том случае, когда наблюдается иммунный ответ, который становится больше или усиливается или каким-либо образом отклоняется при добавлении вещества, по сравнению с таким же иммунным ответом, измеренным без добавления вещества. Например, может быть измерена литическая активность цитотоксических T-клеток, например, с использованием анализа высвобождения 51Cr в образцах, полученных с применением и без применения вещества в ходе иммунизации. Говорят, что количество вещества, при котором литическая активность CTL усилена по сравнению с литической активностью CTL без вещества, является количеством, достаточным для усиления иммунного ответа животного на антиген. В предпочтительном варианте иммунный ответ усилен по меньшей мере примерно в 2 раза, более предпочтительно примерно в 3 раза или больше. Также может быть изменено количество или тип секретируемых цитокинов. Альтернативно может быть изменено количество индуцируемых антител или их подклассов.
Иммунизация: В используемом в данном описании смысле термины «иммунизировать» или «иммунизация» или связанные термины относятся к приданию способности вырабатывать достаточно сильный иммунный ответ (включая гуморальный и/или клеточный иммунитет, такой как эффекторные CTL) против антигена или эпитопа, являющегося мишенью. Указанные термины не требуют, чтобы был сформирован полный иммунитет, а скорее, чтобы был получен иммунный ответ, который значительно выше исходного уровня. Например, млекопитающее может считаться иммунизированным против антигена-мишени, если после применения способов согласно изобретению возникает клеточный и/или гуморальный иммунный ответ на антиген-мишень.
Природное происхождение: В используемом в данном описании смысле термин «природное происхождение» означает, что целое или часть целого не синтезируют, а оно существует или продуцируется в природе.
Неприродный: В используемом в данном описании смысле термин, в общем, означает полученный не из природы, более конкретно, термин означает полученный искусственно человеком.
Неприродное происхождение: В используемом в данном описании смысле термин «неприродное происхождение» в общем означает «синтезирован или получен не из природы»; более конкретно, термин означает «получен искусственно человеком».
Упорядоченная и повторяющаяся матрица антигенов или антигенных детерминант: В используемом в данном описании смысле термин «упорядоченная и повторяющаяся матрица антигенов или антигенных детерминант» в общем относится к повторяющемуся образцу антигенов или антигенных детерминант, обычно и предпочтительно характеризующемуся однородным пространственным расположением антигенов или антигенных детерминант по отношению к центральной частице и, соответственно, вирусоподобной частице. В одном варианте изобретения повторяющийся образец может иметь геометрический рисунок. Типичными и предпочтительными примерами подходящих упорядоченных и повторяющихся матриц антигенов или антигенных детерминант являются матрицы, которые имеют строго повторяющиеся паракристаллические порядки расположения антигенов или антигенных детерминант, предпочтительно с расстояниями от 1 до 30 нанометров, предпочтительно от 5 до 15 нанометров.
Фимбрии: В используемом в данном описании смысле термин «фимбрии» (в единственном числе «фимбрия») относится к внеклеточным структурам бактериальных клеток, состоящим из белковых мономеров (например, мономеров пилина), которые организованы в упорядоченные и повторяющиеся структуры. Кроме того, фимбрии являются структурами, которые вовлечены в такие процессы, как связывание бактериальных клеток с поверхностными рецепторами клетки-хозяина, межклеточные генетические обмены и распознавание клетками друг друга. Примеры фимбрий включают фимбрии типа 1, P-фимбрии, фимбрии F1C, S-фимбрии и фимбрии 987P. Дополнительные примеры фимбрий указаны ниже.
Структуры, подобные фимбриям: В используемом в данном описании смысле фраза «структура, подобная фимбриям» относится к структурам, имеющим характеристики, сходные с характеристиками фимбрий, и состоящим из белковых мономеров. Одним примером «структуры, подобной фимбриям» является структура, образованная бактериальной клеткой, которая экспрессирует модифицированные белки пилина, которые не образуют упорядоченные и повторяющиеся матрицы, которые идентичны матрицам природных фимбрий.
Полипептид: В используемом в данном описании смысле термин «полипептид» относится к молекуле, состоящей из мономеров (аминокислот), линейно связанных амидными связями (также называемыми пептидными связями). Термин указывает на молекулярную цепь аминокислот и не относится к конкретной длине продукта. Таким образом, пептиды, дипептиды, трипептиды, олигопептиды и белки включены в определение полипептида. Подразумевается, что данный термин также относится к модификациям полипептида после экспрессии, например гликозилированию, ацетилированию, фосфорилированию и тому подобному. Рекомбинантный или производный полипептид необязательно транслируют с определенной последовательности нуклеиновой кислоты. Его также можно создать любым способом, включая химический синтез.
Белок RANKL: Термин «белок RANKL» в используемом в данном описании смысле относится к белку, кодируемому геном RANKL. Различные варианты белка RANKL могут быть обусловлены точечными мутациями нуклеотидов и полиморфизмом, соответственно, а также инсерциями, делециями и/или заменами одного или нескольких нуклеотидов и должны быть включены в объем данного изобретения. Дополнительная вариабельность может быть обусловлена посттрансляционными модификациями, такими как дифференциально гликозилированные формы RANKL, а также протеолитически расщепленные формы RANKL (Lum, L., et al., J. Biol. Chem. 274: 13613-13618 (2000). Имеется ряд известных в настоящее время вариантов сплайсинга гена RANKL человека и гена RANKL других видов, которые вместе с возможными вариантами, указанными выше, также входят в объем изобретения. Таким образом, термин «белок RANKL» в используемом в данном описании смысле также должен охватывать варианты белка RANKL, включая, но не ограничиваясь этим, указанные выше предпочтительные примеры.
В используемом в данном описании смысле термин «фрагмент RANKL» имеет широкое определение как любой полипептид длиной по меньшей мере 50 аминокислот, который представляет собой часть белка RANKL, наиболее предпочтительно подвергнутую укладке часть RANKL и наиболее предпочтительно внеклеточную часть RANKL, еще более предпочтительно область, гомологичную TNF-α. Термин фрагмент RANKL также охватывает рекомбинантно полученные белки и, соответственно, полипептиды, соответствующие изоформам сплайсинга и протеолитически расщепленным формам RANKL и все его варианты, описанные выше.
В используемом в данном описании смысле термин «пептид RANKL» имеет широкое определение как любой пептид, который представляет собой часть белка RANKL или фрагмента RANKL и содержит по меньшей мере два, предпочтительно по меньшей мере три, более предпочтительно по меньшей мере четыре, более предпочтительно по меньшей мере пять, более предпочтительно по меньшей мере шесть, следующих друг за другом аминокислот исходного белка RANKL или фрагмента RANKL, наиболее предпочтительно внеклеточной части RANKL. Кроме того, термин «пептид RANKL» предпочтительно должен охватывать любую часть указанного пептида RANKL, где указанная часть предпочтительно может быть получена путем делеции одной или нескольких аминокислот на N- и/или C-конце. Пептид RANKL может быть получен рекомбинантной экспрессией в эукариотических или прокариотических системах экспрессии в виде отдельного пептида RANKL или в виде слияния с другими аминокислотами или белками, например, чтобы облегчить укладку, экспрессию или растворимость пептида RANKL или облегчить очистку пептида RANKL. Чтобы сделать возможным связывание пептидов RANKL и белков субъединиц VLP или капсидов, к пептиду RANKL может быть добавлен по меньшей мере один второй сайт связывания. Альтернативно пептиды RANKL можно синтезировать с использованием способов, известных в данной области. Термин пептид RANKL в используемом в данном описании смысле также предпочтительно должен включать в себя пептид, который имитирует трехмерную структуру поверхности RANKL. Такой пептид RANKL не обязательно получен из непрерывной аминокислотной последовательности RANKL, а может быть образован несмежными аминокислотными остатками из RANKL. Такие пептиды даже могут содержать аминокислоты, которые не присутствуют в соответствующем белке RANKL.
Остаток: В используемом в данном описании смысле подразумевается, что термин «остаток» означает специфичную аминокислоту в полипептидном остове или боковой цепи.
Аутоантиген (собственный антиген): В используемом в данном описании смысле термин «аутоантиген» относится к белкам, кодируемым ДНК хозяина, и продукты, образованные белками или РНК, кодируемыми ДНК хозяина, определяют как собственные. Кроме того, белки, которые являются результатом комбинации двух или нескольких собственных молекул или которые представляют собой часть собственной молекулы, и белки, которые обладают высокой гомологией с двумя собственными молекулами, которые определены выше (>95%, предпочтительно >97%, более предпочтительно >99%), также могут считаться собственными.
Лечение: В используемом в данном описании смысле термины «лечение», «лечить», «подвергнутый лечению» или «процесс лечения» относится к профилактике и/или терапии. Например, в случае использования по отношению к инфекционному заболеванию термин относится к профилактическому лечению, которое повышает устойчивость субъекта к инфекции патогеном или, другими словами, уменьшает вероятность того, что субъект станет инфицированным патогеном или будет проявлять симптомы болезни, которые можно отнести к инфекции, а также к лечению, после того, как субъект стал инфицированным, чтобы бороться с инфекцией, например, чтобы уменьшить или уничтожить инфекцию или не допустить того, чтобы субъекту стало хуже. В случае использования по отношению к заболеванию кости термин «лечение» относится к профилактическому или терапевтическому лечению, которое ингибирует или снижает повышенную резорбцию кости, которая связана с заболеваниями кости.
Вакцина: В используемом в данном описании смысле термин «вакцина» относится к препарату, который содержит композицию согласно данному изобретению и который находится в форме, которую можно вводить животному. Обычно вакцина содержит среду в виде обычного физиологического раствора или забуференного водного раствора, в которой суспендирована или растворена композиция согласно данному изобретению. В указанной форме композицию согласно данному изобретению легко можно использовать для того, чтобы предотвратить, улучшить или другим образом воздействовать на состояние. При введении в хозяина вакцина способна возбуждать иммунный ответ, включая, но не ограничиваясь указанным, продукцию антител и/или цитокинов и/или активацию цитотоксических T-клеток, антигенпрезентирующих клеток, хелперных T-клеток, дендритных клеток и/или другие клеточные ответы.
Необязательно вакцина согласно данному изобретению дополнительно содержит адъювант, который может присутствовать либо в небольшой, либо в большой пропорции относительно соединения согласно данному изобретению. Термин «адъювант» в используемом в данном описании смысле относится к неспецифичным стимуляторам иммунного ответа или веществам, которые делают возможным создание депо в организме хозяина, которые при комбинировании с вакциной согласно данному изобретению обеспечивают еще более усиленный иммунный ответ. Можно использовать множество адъювантов. Примеры включают полный и неполный адъювант Фрейнда, гидроксид алюминия и модифицированный мурамилдипептид.
Вирусоподобная частица (VLP): В используемом в данном описании смысле термин «вирусоподобная частица» относится к структуре, подобной вирусной частице. Кроме того, вирусоподобная частица согласно изобретению является нерепликативной и неинфекционной, так как не содержит всего или части вирусного генома, в частности репликативных и инфекционных компонентов вирусного генома. Вирусоподобная частица согласно изобретению может содержать нуклеиновую кислоту, отличную от ее генома. Типичным и предпочтительным вариантом вирусоподобной частицы согласно данному изобретению является капсид вируса, такой как вирусный капсид соответствующего вируса, бактериофага или РНК-фага. Термины «вирусный капсид» или «капсид», которые используются в данном описании, взаимозаменяемо относятся к макромолекулярной конструкции, состоящей из белковых субъединиц вируса. Обычно и предпочтительно белковые субъединицы вируса собираются в вирусный капсид и, соответственно, «капсид», имеющий структуру с присущей ей повторяющейся организацией, где указанная структура обычно является сферической или трубчатой. Например, капсиды РНК-фагов или HBcAg имеют сферическую форму с икосаэдрической симметрией. Термин «капсидоподобная структура» в используемом в данном описании смысле относится к собранной из макромолекул конструкции, состоящей из белковых субъединиц вируса, имеющей аналогичную капсиду морфологию в определенном выше смысле, но имеющей отклонения от типичной симметричной сборки при сохранении достаточной степени порядка и повторяемости.
Вирусоподобная частица бактериофага: В используемом в данном описании смысле термин «вирусоподобная частица бактериофага» относится к вирусоподобной частице, аналогичной по структуре бактериофагу, но являющейся нерепликативной и неинфекционной, и не содержащей по меньшей мере гена или генов, кодирующих аппарат репликации бактериофага, и обычно также не содержащей гена или генов, кодирующих белок или белки, отвечающие за прикрепление вируса или проникновение вируса в хозяина. Однако данное определение также должно охватывать вирусоподобные частицы бактериофагов, в которых вышеуказанный ген или гены еще присутствуют, но неактивны и, следовательно, также приводят к нереплицирующимся и неинфекционным вирусоподобным частицам бактериофагов.
VLP покрывающего белка РНК-фага: Капсидную структуру, образованную в результате самосборки 180 субъединиц покрывающего белка РНК-фага и необязательно содержащую РНК хозяина, называют «VLP покрывающего белка РНК-фага». Конкретным примером является VLP покрывающего белка Qβ. В данном конкретном случае VLP покрывающего белка Qβ может быть собрана либо исключительно из субъединиц CP Qβ (образованных при экспрессии гена CP Qβ, содержащего, например, стоп-кодон TAA, препятствующий любой экспрессии более длинного белка A1 посредством супрессии, см. Kozlovska, T.M., et al., Intervirology 39: 9-15 (1996)), либо дополнительно содержит субъединицы белка A1 в собранной конструкции капсида.
Вирусная частица: Термин «вирусная частица» в используемом в данном описании смысле относится к морфологической форме вируса. У некоторых типов вирусов она содержит геном, окруженный белковым капсидом; другие имеют дополнительные структуры (например, оболочки, хвосты и т.д.).
Один или форма единственного числа: При использовании в данном описании терминов «один» или форм единственного числа они означают «по меньшей мере» один или «один или несколько», если не оговорено особо.
Как будет понятно специалистам в данной области, некоторые варианты изобретения касаются применения методик на основе рекомбинантной нуклеиновой кислоты, таких как клонирование, полимеразная цепная реакция, очистка ДНК и РНК, экспрессия рекомбинантных белков в прокариотических и эукариотических клетках и т.д. Такие методики хорошо известны специалистам в данной области, и их легко можно найти в опубликованных руководствах по лабораторным способам (например, Sambrook, J. et al., eds., Molecular Cloning, A Laboratory Manual, 2nd. edition, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1989); Ausubel, F. et al., eds., Current Protocols in Molecular Biology, John H. Wiley and Sons, Inc. (1997)). Фундаментальные лабораторные методики для работы с линиями клеток культур тканей (Celis, J., ed., Cell Biology, Academic Press, 2nd edition, (1998)) и методики на основе антител (Harlow, E. and Lane, D., Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y. (1988); Deutscher, M.P., «Guide to Protein Purification», Meth. Enzymol. 128, Academic Press San Diego (1990); Scopes, R.K., Protein Purification Principles and Practice, 3rd ed., Springer-Verlag, New York (1994)) также в достаточной мере описаны в литературе, все указанные публикации включены в данное описание в виде ссылки.
2. Композиции и способы усиления иммунного ответа
Заявленное изобретение относится к композициям и способам усиления иммунного ответа против белка RANKL, фрагмента RANKL или пептида RANKL у животного. Композиции согласно изобретению содержат или альтернативно состоят из (a) центральной частицы по меньшей мере с одним первым сайтом связывания; и (b) по меньшей мере одного антигена или антигенной детерминанты по меньшей мере с одним вторым сайтом связывания, где указанный антиген или антигенная детерминанта является белком RANKL, фрагментом RANKL или пептидом RANKL и где указанный второй сайт связывания выбран из группы, состоящей из (i) сайта связывания неприродного происхождения с указанным антигеном или антигенной детерминантой; и (ii) сайта связывания природного происхождения с указанным антигеном или антигенной детерминантой, где указанный второй сайт связывания способен вступать в ассоциацию с указанным первым сайтом связывания; и где указанный антиген или антигенная детерминанта и указанная центральная частица взаимодействуют благодаря указанной ассоциации, образуя упорядоченную и повторяющуюся антигенную матрицу. Более конкретно, композиции согласно изобретению содержат - или альтернативно - состоят из вирусоподобной частицы и по меньшей мере одного антигена или антигенной детерминанты, где антиген или антигенная детерминанта является белком RANKL, фрагментом RANKL или пептидом RANKL и где по меньшей мере один антиген или антигенная детерминанта связаны с вирусоподобной частицей, так что образуется упорядоченная и повторяющаяся антиген-VLP-матрица. Кроме того, изобретение дает возможность специалисту-практику легко конструировать такую композицию, наряду с прочим, для лечения и/или профилактического предотвращения костных заболеваний, характеризующихся повышенной резорбцией кости.
В одном варианте центральная частица содержит вирус, фимбрию бактерии, структуру, образованную из бактериального пилина, бактериофаг, вирусоподобную частицу, частицу вирусного капсида или их рекомбинантную форму. Любой вирус, известный в данной области, имеющий упорядоченную и повторяющуюся структуру покрывающего и/или корового белка, может быть выбран в качестве центральной частицы согласно изобретению; примеры подходящих вирусов включают в себя вирус Синдбис и другие альфавирусы, рабдовирусы (например, вирус везикулярного стоматита), пикорнавирусы (например, риновирус человека, вирус Аиши), тогавирусы (например, вирус краснухи), ортомиксовирусы (например, вирус Тогото, вирус Баткен, вирус чумы птиц), вирусы полиомы (например, полиомавирус BK, полиомавирус JC, вирус полиомы птиц BFDV), парвовирусы, ротавирусы, вирус Норуолк, вирус ящура, ретровирус, вирус гепатита B, вирус табачной мозаики, вирус овечьих кошар и вирус папилломы человека, и предпочтительно РНК-фаг, бактериофаг Qβ, бактериофаг R17, бактериофаг M11, бактериофаг MX1, бактериофаг NL95, бактериофаг fr, бактериофаг GA, бактериофаг SP, бактериофаг MS2, бактериофаг f2, бактериофаг PP7 (например, см. таблицу 1 в Bachmann, M.F. and Zinkernagel, R.M., Immunol. Today 17: 553-558 (1996)).
В следующем варианте в изобретении используют генетическую инженерию вируса, чтобы создать слияние между упорядоченным и повторяющимся белком вирусной оболочки и первым сайтом связывания, содержащим гетерологичный белок, пептид, антигенную детерминанту или предпочтительно реакционно-способный аминокислотный остаток. Другие генетические манипуляции, известные специалистам в данной области, могут быть включены в конструирование композиций согласно изобретению; например, может требоваться ограничение способности рекомбинантного вируса к репликации посредством генетической мутации. Кроме того, вирус, используемый для данного изобретения, является некомпетентным по репликации вследствие химической или физической инактивации или, как указано, вследствие отсутствия компетентного по репликации генома. Вирусный белок, выбранный для слияния с первым сайтом связывания, должен иметь организованную и повторяющуюся структуру. Такая организованная и повторяющаяся структура включает в себя паракристаллические организации с расстояниями 5-30 нм, предпочтительно 5-15 нм, на поверхности вируса. Результатом создания слитого белка данного типа будут множественные, упорядоченные и повторяющиеся первые сайты связывания на поверхности вируса при отражении нормальной организации нативного вирусного белка. Как будет понятно специалистам в данной области, первый сайт связывания может быть частью или может быть любым подходящим белком, полипептидом, сахаром, полинуклеотидом, пептидом (аминокислотой), природным или синтетическим полимером, вторичным метаболитом или их комбинацией, которые могут служить для специфичного связывания антигена или антигенной детерминанты, приводящего к образованию упорядоченной и повторяющейся матрицы антигена или антигенной детерминанты.
В другом варианте изобретения центральная частица является рекомбинантным альфавирусом и более конкретно рекомбинантным вирусом Синдбис. Альфавирусы являются вирусами с позитивной нитью РНК, которые реплицируют свою геномную РНК полностью в цитоплазме инфицированной клетки и без промежуточных ДНК-продуктов (Strauss, J. and Strauss, E., Microbiol. Rev. 58: 491-562 (1994)). Большое внимание было обращено на несколько представителей семейства альфавирусов, Синдбис (Xiong, C. et al., Science 243: 1188-1191 (1989); Schlesinger, S., Trends Biotechnol. 11: 18-22 (1993)), вирус леса Семлики (SFV) (Liljeström, P. and Garoff, H., Bio/Technology 9: 1356-1361 (1991)) и другие (Davis, N.L. et al., Virology 171: 189-204 (1989)) в качестве основанных на вирусах экспрессирующих векторов для множества различных белков (Lundstrom, K., Curr. Opin. Biotechnol. 8: 578-582 (1997); Liljeström, P., Curr. Opin. Biotechnol. 5: 495-500 (1994)) и в качестве кандидатов для разработки вакцин. Недавно выпущен ряд патентов, направленных на применение альфавирусов для экспрессии гетерологичных белков и разработки вакцин (см. патенты США № 5766602, 5792462, 5739026, 5789245 и 5814482). Конструирование альфавирусных центральных частиц согласно изобретению можно осуществить способами, широко известными в области технологии рекомбинантной ДНК, которые описаны в вышеуказанных статьях, которые включены в данное описание в виде ссылки.
Множество различных рекомбинантных клеток-хозяев можно использовать для получения основанной на вирусе центральной частицы для связывания антигена или антигенной детерминанты. Например, известно, что альфавирусы имеют широкий круг хозяев; вирус Синдбис инфицирует культивируемые клетки млекопитающих, рептилий и амфибий, а также некоторые клетки насекомых (Clark, H., J. Natl. Cancer Inst. 51:645 (1973); Leake, C., J. Gen. Virol. 35:335 (1977); Stollar, V. in The Togaviruses, R.W. Schlesinger, Ed., Academic Press, (1980), pp.583-621). Таким образом, в практике данного изобретения можно использовать множество рекомбинантных клеток-хозяев. Клетки BHK, COS, Vero, HeLa и CHO особенно подходят для продукции гетерологичных белков, так как они имеют возможность гликозилировать гетерологичные белки сходным с клетками человека образом (Watson, E. et al., Glycobiology 4: 227, (1994)) и могут быть отобраны в ходе селекции (Zang, M. et al., Bio/Technology 13:389 (1995)) или сконструированы генетическим способом (Renner W. et al., Biotech. Bioeng. 4: 476 (1995); Lee K. et al. Biotech. Bioeng. 50:336 (1996)), так, чтобы они росли в среде, не содержащей сыворотки, а также в суспензии.
Введение полинуклеотидных векторов в клетки-хозяева можно осуществить способами, описанными в стандартных лабораторных руководствах (см., например, Sambrook, J. et al., eds., Molecular Cloning, A Laboratory Manual, 2nd. edition, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1989), Chapter 9; Ausubel, F. et al., eds., Current Protocols in Molecular Biology, John H. Wiley and Sons, Inc. (1997), Chapter 16), включая такие способы как электропорация, опосредованная DEAE-декстраном трансфекция, трансфекция, микроинъекция, опосредованная катионными липидами трансфекция, трансдукция, введение с помощью царапин, баллистическое введение и инфекция. Способы введения экзогенных последовательностей ДНК в клетки-хозяева обсуждается в Felgner, P. et al., патент США № 5580859.
Упакованные последовательности РНК также можно использовать для инфекции клеток-хозяев. Указанные упакованные последовательности РНК можно ввести в клетки-хозяева путем добавления их в культуральную среду. Например, получение неинфекционных альфавирусных частиц описано в ряде источников, включая «Sindbis Expression System», Version C (Invitrogen, номер в каталоге K750-1).
Когда в качестве рекомбинантных клеток-хозяев для получения основанных на вирусах центральных частиц используют клетки млекопитающих, указанные клетки обычно выращивают в культуре ткани. Способы выращивания клеток в культуре хорошо известны в данной области (см., например, Celis, J., ed., Cell Biology, Academic Press, 2nd edition, (1998); Sambrook, J. et al., eds., Molecular Cloning, A Laboratory Manual, 2nd. edition, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. (1989); Ausubel, F. et al., eds., Current Protocols in Molecular Biology, John H. Wiley and Sons, Inc. (1997); Freshney, R., Culture of Animal Cells, Alan R. Liss, Inc. (1983)).
Следующие примеры РНК-вирусов, подходящих для применения в качестве центральных частиц в данном изобретении, включают, но не ограничены указанным, следующие вирусы: представителей семейства Reoviridae, включая род Orthoreovirus (множественные серотипы ретровирусов млекопитающих и птиц), род Orbivirus (вирус синего языка, вирус Эугенанги, вирус Кемерово, вирус африканской болезни лошадей и вирус колорадской клещевой лихорадки) и род Rotavirus (ротавирус человека, вирус диареи телят Небраски, ротавирус мышей, ротавирус обезьян, бычий или овечий ротавирус, ротавирус птиц); семейство Picomaviridae, включая род Enterovirus (полиовирус, вирус Коксаки A и B, кишечные цитопатические «сиротские» вирусы человека (ECHO), вирусы гепатита A, C, D, E и G, энтеровирусы обезьян, вирусы энцефаломиелита мышей (ME), Poliovirus muris, бычьи энтеровирусы, энтеровирусы свиней, род Cardiovirus (вирус энцефаломиокардита (EMC), вирус Менго), род Rhinovirus (риновирусы человека, включая по меньшей мере 113 подтипов; другие риновирусы), род Apthovirus (вирус ящура (FMDV)); семейство Calciviridae, включая вирус везикулярной экзантемы свиней, вирус морских львов Сан-Мигель, кошачий пикорнавирус и вирус Норуолк; семейство Togaviridae, включая род Alphavirus (вирус восточного энцефалита лошадей, вирус леса Семлики, вирус Синдбис, вирус Чикунгунья, вирус О'Ньонг-Ньонг, вирус реки Росс, вирус венесуэльского энцефалита лошадей, вирус западного энцефалита лошадей), род Flavirius (вирус москитной желтой лихорадки, вирус Денге, вирус японского энцефалита, вирус энцефалита Сан-Луи, вирус энцефалита долины Мюррей, вирус Западного Нила, вирус Кунжин, вирус центральноевропейского клещевого энцефалита, вирус дальневосточного клещевого энцефалита, вирус леса Киасанур, вирус Louping III, вирус Повассан, вирус Омской геморрагической лихорадки), род Rubivirus (вирус краснухи), род Pestivirus (вирус болезни слизистых оболочек, вирус холеры свиней, вирус пограничной болезни); семейство Bunyaviridae, включая род Bunyvirus (вирус Буньямвера и родственные вирусы, группа вирусов Калифорнийского энцефалита), род Phlebovirus (вирус сицилийской москитной лихорадки, вирус лихорадки долины Рифт), род Nairovirus (вирус геморрагической лихорадки Крым-Конго, вирус болезни овец Найроби) и род Uukuvirus (вирус Уукуниеми и родственные вирусы); семейство Orthomyxoviridae, включая род вируса гриппа (вирус гриппа типа A, множество серотипов у человека); вирус гриппа свиней и вирусы гриппа птиц и лошадей; вирус гриппа типа B (множество серотипов у человека) и вирус гриппа типа C (возможно отдельный род); семейство Paramyxoviridae, включая род Paramyxovirus (вирус парагриппа типа 1, вирус Сендай, вирус гемадсорбции, вирусы парагриппа типов 2-5, вирус ньюкаслской болезни, вирус свинки), род Morbillivirus (вирус кори, вирус подострого склерозирующего панэнцефалита, вирус чумки, вирус чумы рогатого скота), род Pneumovirus (респираторно-синцитиальный вирус (RSV), бычий респираторно-синцитиальный вирус и вирус пневмонии мышей); вирус леса, семейство Rhabdoviridae, включая род Vesiculovirus (VSV), вирус Чандипура, вирус Фландерс-Харт Парк), род Lyssavirus (вирус бешенства), рабдовирусы рыб и филовирусы (вирус Марбург и вирус Эбола); семейство Arenaviridae, включая вирус лимфоцитарного хориоменингита (LCM), вирус комплекса Такарибе и вирус Ласса; семейство Coronoaviridae, включая вирус инфекционного бронхита (IBV), вирус гепатиты мышей, кишечный коронавирус человека и вирус инфекционного кошачьего перитонита (кошачий коронавирус).
Иллюстративные ДНК-вирусы, которые можно использовать в качестве центральных частиц, включают, но не ограничены указанным: семейство Poxviridae, включая род Orthopoxvirus (вирус натуральной оспы, вирус белой оспы, вирус оспы обезьян, вирус оспы коров, вирус оспы буйволов, вирус оспы кроликов, вирус эктромелии), род Leporipoxvirus (миксома, фиброма), род Avipoxvirus (вирус оспы домашних птиц и вирус оспы других птиц), род Capripoxvirus (оспа овец, оспа коз), род Suipoxvirus (оспа свиней), род Parapoxvirus (вирус инфекционного пустулезного дерматита, вирус псевдооспы коров («узелков доярок»), вирус папулезного стоматита крупного рогатого скота); семейство Iridoviridae (вирус африканской лихорадки свиней, вирусы лягушки 2 и 3, вирус лимфоцистоза рыб); семейство Herpesviridae, включая альфа-герпесвирусы (вирусы простого герпеса типа 1 и 2, вирус Варицелла-Зостер, вирус аборта лошадей, вирус герпеса лошадей типа 2 и 3, вирус псевдобешенства, вирус инфекционного бычьего кератоконъюнктивита, вирус инфекционного ринотрахеита крупного рогатого скота, вирус кошачьего ринотрахеита, вирус инфекционного ларинготрахеита), бета-герпесвирусы (цитомегаловирус человека и цитомегаловирусы свиней, обезьян и грызунов); гамма-герпесвирусы (вирус Эпштейна-Барр (EBV), вирус болезни Марека, Herpes saimiri (герпес беличьих обезьян), Herpesvirus ateles (вирус герпеса паукообразных обезьян), Herpesvirus sylvilagus (вирус герпеса лесных кроликов), вирус герпеса морских свинок, вирус опухоли Люкке); и семейство Adenoviridae, включая род Mastadenovirus (подгруппы A, B, C, D и E человека и не относящиеся к подгруппам; аденовирусы обезьян (по меньшей мере 23 серотипа), инфекционный гепатит собак и аденовирусы крупного рогатого скота, свиней, овец, лягушек и многих других видов, род Aviadenovirus (аденовирусы птиц); и некультивируемые аденовирусы; семейство Papoviridae, включая род Papillomavirus (вирусы папилломы человека, вирусы папилломы крупного рогатого скота, вирусы папилломы кроликов Шоупа и различные патогенные вирусы папилломы других видов), род Polyomavirus (полиомавирус, вакуолизирующий вирус обезьян (SV-40), вакуолизирующий вирус кроликов (RKV), K-вирус, BK-вирус, JC-вирус и другие вирусы полиомы приматов, такие как лимфотропный вирус папилломы); семейство Parvoviridae, включая род аденоассоциированных вирусов, род Parvovirus (вирус кошачьей панлейкопении, парвовирус крупного рогатого скота, парвовирус собак, вирус алеутской болезни норок и т.д.). Наконец, ДНК-вирусы могут включать такие вирусы, как агенты хронической инфекционной невропатии (вирус CHINA).
В других вариантах бактериальный пилин, субфрагмент бактериального пилина или слитый белок, который содержит либо бактериальный пилин, либо его субфрагмент, используют для приготовления композиций и, соответственно, вакцинных композиций согласно изобретению. Примеры белков пилина включают пилины, продуцируемые Escherichia coli, Haemophilus influenzae, Neisseria meningitidis, Neisseria gonorrhoeae, Caulobacter crescentus, Pseudomonas stutzeri и Pseudomonas aeruginosa. Аминокислотные последовательности белков пилина, подходящие для применения в случае данного изобретения, включают в себя последовательности, указанные в записях GenBank AJ000636 (SEQ ID NO:1), AJ132364 (SEQ ID NO:2), AF229646 (SEQ ID NO:3), AF051814 (SEQ ID NO:4), AF051815 (SEQ ID NO:5) и X00981 (SEQ ID NO:6), полное описание которых включено в данную заявку в виде ссылки.
Белки бактериального пилина обычно процессируются с удалением N-концевых лидерных последовательностей перед экспортом белков в бактериальную периплазму. Кроме того, как будет понятно специалисту в данной области, белки бактериального пилина, используемые для получения композиций и, соответственно, композиций вакцин согласно изобретению, как правило, не будут иметь лидерной последовательности, присутствующей в естественных условиях.
Одним конкретным примером белка пилина, подходящего для применения в данном изобретении, является P-пилин E. coli (запись в GenBank AF237482 (SEQ ID NO:7)). Примером пилина E. coli типа 1, подходящего для применения в изобретении, является пилин, имеющий аминокислотную последовательность, указанную в GenBank, запись P04128 (SEQ ID NO:8), которая кодируется нуклеиновой кислотой, имеющей нуклеотидную последовательность, указанную в GenBank, запись M27603 (SEQ ID NO:9). Полное описание указанных данных GenBank включено в данную заявку в виде ссылки. И снова в большинстве случаев зрелая форма указанного выше белка может быть использована для приготовления композиции и, следовательно, композиции вакцины согласно изобретению.
Бактериальные пилины или субфрагменты пилинов, подходящие для применения в практике данного изобретения, как правило, будут способны вступать в ассоциацию с образованием упорядоченных и повторяющихся антигенных матриц.
Способы получения фимбрий и подобных фимбриям структур in vitro хорошо известны в данной области. Например, Bullitt et al., Proc. Natl. Acad. Sci. USA 93: 12890-12895 (1996) описали реконструирование in vitro субъединиц P-фимбрий E. coli. Кроме того, Eshdat et al., J. Bacteriol. 148: 308-314 (1981) описали способы, подходящие для диссоциации фимбрий типа-1 E. coli и реконструирование фимбрий. Коротко, указанные способы заключаются в следующем: фимбрии подвергают диссоциации путем инкубации при 37°C в насыщенном гидрохлориде гуанидина. Затем белки пилина очищают путем хроматографирования, после чего димеры пилина образуют при диализе против 5 мМ гидрохлорида трис(гидроксиметил)аминометана (pH 8,0). Eshdat et al. также обнаружили, что димеры пилина снова подвергаются сборке, образуя фимбрии, при диализе против 5 мМ трис(гидроксиметил)аминометана (pH 8,0), содержащего 5 мМ MgCl2.
Кроме того, используя, например, обычные способы генетической инженерии и модификации белка, белки пилина можно модифицировать так, чтобы они содержали первый сайт связывания, с которым связывают антиген или антигенную детерминанту посредством второго сайта связывания. Альтернативно антигены или антигенные детерминанты можно непосредственно связывать посредством второго сайта связывания с аминокислотными остатками, которые присутствуют в указанных белках в природе. Затем указанные модифицированные белки пилина можно использовать в композициях вакцин согласно изобретению.
Бактериальные белки пилина, используемые для получения композиций и, соответственно, композиций вакцин согласно изобретению, можно модифицировать способом, подобным способу, приведенному в данном описании для HBcAg. Например, остатки цистеина и лизина могут быть либо делетированы, либо заменены другими аминокислотными остатками, и к полученным белкам может быть добавлен первый сайт связывания. Кроме того, белки пилина могут быть либо экспрессированы в модифицированной форме, либо химически модифицированы после экспрессии. Подобным образом можно собрать интактные фимбрии из бактерий и затем модифицировать химическим способом.
В другом варианте фимбрии или подобные фимбриям структуры собирают из бактерий (например, E. coli) и используют для образования композиций и композиций вакцин согласно изобретению. Одним примером фимбрий, подходящих для получения композиций и вакцинных композиций, является фимбрия типа-1 E. coli, которая образуется из мономеров пилина, имеющих аминокислотную последовательность, указанную в SEQ ID NO:8.
В данной области известен ряд способов сбора бактериальных фимбрий. Например, Bullitt and Makowski (Biophys. J. 74: 623-632 (1998)), описали способ очистки фимбрий в случае сбора P-фимбрий E. coli. Согласно данному способу фимбрии срезают с E. coli, имеющих гиперколичество фимбрий и содержащих P-фимбрия-плазмиду, и очищают, используя циклы растворения и осаждения с MgCl2 (1,0 М).
После сбора фимбрии или подобные фимбриям структуры можно модифицировать множеством способов. Например, к фимбриям можно добавить первый сайт связывания, с которым могут быть связаны антигены или антигенные детерминанты посредством второго сайта связывания. Другими словами, бактериальные фимбрии или подобные фимбриям структуры можно собирать и модифицировать, получая в результате упорядоченные и повторяющиеся антигенные матрицы.
Антигены или антигенные детерминанты могут быть связаны с остатками цистеина или остатками лизина природного происхождения, присутствующими в фимбиях или подобных фимбриям структурах. В таких случаях высокая упорядоченность и повторяемость аминокислотного остатка природного происхождения могут регулировать связывание антигенов или антигенных детерминант с фимбриями или подобными фимбриям структурами. Например, фимбрии или подобные фимбриям структуры могут быть связаны со вторыми сайтами связывания антигенов или антигенных детерминант с использованием гетеробифункционального перекрестно сшивающего агента.
Когда для получения композиций и вакцинных композиций согласно изобретению используют структуры, которые синтезируются организмами в природе (например, фимбрии), часто преимущество будет давать генетическое конструирование указанных организмов так, чтобы они продуцировали структуры, обладающие требуемыми характеристиками. Например, когда используют фимбрии типа-1 E. coli, E. coli, из которых собирают указанные фимбрии, можно модифицировать так, чтобы они продуцировали структуры с конкретными характеристиками. Примеры возможных модификаций белков пилина включают инсерцию одного или нескольких остатков лизина, делецию или замену одного или нескольких естественно присущих остатков лизина и делецию или замену одного или нескольких естественно присущих остатков цистеина (например, остатков цистеина в положениях 44 и 84 в SEQ ID NO:8).
Кроме того, могут быть осуществлены дополнительные модификации в генах пилина, которые приводят к продуктам экспрессии, содержащим другой первый сайт связывания, отличный от остатка лизина (например, домен FOS или JUN). Конечно, подходящий первый сайт связывания в общем будет ограничен сайтами, которые не препятствуют образованию из белков пилина фимбрий или подобных фимбриям структур, подходящих для применения в композициях вакцин согласно изобретению.
Гены пилина, которые в природе имеются в бактериальных клетках, могут быть модифицированы in vivo (например, посредством гомологичной рекомбинации), или в указанные клетки могут быть встроены гены пилина с конкретными характеристиками. Например, гены пилина могут быть введены в бактериальные клетки в виде компонента либо реплицирующегося клонирующего вектора, либо вектора, который встраивается в бактериальную хромосому. Встроенные гены пилина также могут быть связаны с последовательностями, контролирующими регуляцию экспрессии (например, lac-оператором).
В большинстве случаев фимбрии или подобные фимбриям структуры, используемые в композициях и, соответственно, в композициях вакцин согласно изобретению, будут построены из субъединицы пилина одного типа. Как правило, будут использоваться фимбрии или подобные фимбриям структуры, состоящие из идентичных субъединиц, так как предполагается, что они образуют структуры, которые представляют собой высокоупорядоченные и повторяющиеся антигенные матрицы.
Однако композиции согласно изобретению также включают в себя композиции и вакцины, содержащие фимбрии и подобные фимбриям структуры, образованные из гетерогенных субъединиц пилина. Субъединицы пилина, которые образуют указанные фимбрии или подобные фимбриям структуры, могут быть экспрессированы с генов, присутствующих в бактериальных клетках в природе, или могут быть введены в клетки. Когда оба гена - и присутствующий в природе ген пилина, и введенный ген экспрессируются в клетке, которая образует фимбрии или подобные фимбриям структуры, результатом, как правило, будут структуры, образованные из смеси указанных белков пилина. Кроме того, когда в бактериальной клетке экспрессируются два или более генов пилина, относительная экспрессия каждого гена пилина обычно будет фактором, который определяет соотношение различных субъединиц пилина в фимбриях или подобных фимбриям структурах.
В том случае, когда требуются фимбрии или подобные фимбриям структуры, имеющие конкретный состав из смешанных субъединиц пилина, экспрессию по меньшей мере одного из генов пилина можно регулировать гетерологичным индуцируемым промотором. Такие промоторы, а также другие генетические элементы можно использовать для того, чтобы регулировать относительные количества различных субъединиц пилина, продуцируемых в бактериальной клетке, а следовательно, состав фимбрий или подобных фимбриям структур.
Кроме того, антиген или антигенная детерминанта может быть связана с бактериальными фимбриями или подобными фимбриям структурами связью, которая не является пептидной связью, бактериальные клетки, которые продуцируют фимбрии или подобные фимбриям структуры, используемые в композициях согласно изобретению, можно генетически сконструировать так, чтобы создавать белки пилина, которые слиты с антигеном или антигенной детерминантой. Такие слитые белки, которые образуют фимбрии или подобные фимбриям структуры, подходят для применения в композициях вакцин согласно изобретению.
Вирусоподобные частицы в контексте данной заявки относятся к структурам, аналогичным вирусной частице, но которые являются непатогенными. В общем, вирусоподобные частицы не содержат вирусного генома и, следовательно, являются неинфекционными. Также вирусоподобные частицы могут продуцироваться в больших количествах в результате гетерологичной экспрессии и могут быть легко очищены.
В предпочтительном варианте вирусоподобной частицей является рекомбинантная вирусоподобная частица. Специалист в данной области может получить VLP, используя методику рекомбинантной ДНК и вирусные кодирующие последовательности, которые легко- и общедоступны. Например, кодирующую последовательность белка оболочки или кора вируса можно сконструировать для экспрессии в бакуловирусном экспрессирующем векторе, используя коммерчески доступный бакуловирусный вектор, под регуляторным контролем вирусного промотора с соответствующими модификациями последовательности, чтобы обеспечить функциональную связь кодирующей последовательности с регуляторной последовательностью. Кодирующую последовательность белка оболочки или кора вируса также можно сконструировать для экспрессии, например, в бактериальном экспрессирующем векторе.
Примеры VLP включают, но не ограничены указанным, белки капсида вируса гепатита B (Ulrich, et al., Virus Res. 50: 141-182 (1998)), вируса кори (Warnes, et al., Gene 160: 173-178 (1995)), вируса Синдбис, ротавируса (патент США 5071651 и патент США 5374426), вируса везикулярного стоматита, болезни «ноги и рта» (Twomey, et al., Vaccine 13: 1603-1610, (1995)), вируса Норуолк (Jiang, X., et al., Science 250: 1580-1583 (1990); Matsui, S.M., et al., J. Clin. Invest. 87: 1456-1461 (1991)), ретровирусный белок GAG (WO 96/30523), белок p1 ретротранспозона Ty, поверхностный белок вируса гепатита B (WO 92/11291), вируса папилломы человека (WO 98/15631), РНК-фагов, Ty, fr-фага, GA-фага и Qβ-фага.
Как легко будет понятно специалистам в данной области, VLP согласно изобретению не ограничена какой-либо конкретной формой. Частицу можно синтезировать химическим способом или посредством биологического процесса, который может быть природным или неприродным. В качестве примера такой тип варианта включает вирусоподобную частицу или ее рекомбинантную форму.
В более конкретном варианте, VLP может содержать или, альтернативно, по существу состоять или, альтернативно, состоять из рекомбинантных полипептидов или их фрагментов, выбранных из рекомбинантных полипептидов ротавируса, рекомбинантных полипептидов вируса Норуолк, рекомбинантных полипептидов альфавируса, рекомбинантных полипептидов вируса ящура, рекомбинантных полипептидов вируса кори, рекомбинантных полипептидов вируса Синдбис, рекомбинантных полипептидов вируса полиомы, рекомбинантных полипептидов ретровируса, рекомбинантных полипептидов вируса гепатита B (например, HBcAg), рекомбинантных полипептидов вируса табачной мозаики, рекомбинантных полипептидов вируса овечьих кошар, рекомбинантных полипептидов вируса папилломы человека, рекомбинантных полипептидов бактериофагов, рекомбинантных полипептидов РНК-фагов, рекомбинантных полипептидов Ty, рекомбинантных полипептидов fr-фага, рекомбинантных полипептидов GA-фага и рекомбинантных полипептидов Qβ-фага. Вирусоподобная частица, кроме того, может содержать, или, альтернативно, по существу состоять, или, альтернативно, состоять из одного или нескольких фрагментов таких полипептидов, а также вариантов таких полипептидов. Варианты полипептидов могут иметь, например, по меньшей мере 80%, 85%, 90%, 95%, 97% или 99% идентичности на аминокислотном уровне со своими аналогами дикого типа.
В предпочтительном варианте вирусоподобная частица содержит, по существу состоит или, альтернативно, состоит из рекомбинантных белков или их фрагментов РНК-фага. Предпочтительно РНК-фаг выбран из группы, состоящей из a) бактериофага Qβ; b) бактериофага R17; c) бактериофага fr; d) бактериофага GA; e) бактериофага SP; f) бактериофага MS2; g) бактериофага M11; h) бактериофага MX1; i) бактериофага NL95; k) бактериофага f2 и l) бактериофага PP7.
В другом предпочтительном варианте данного изобретения вирусоподобная частица содержит или, альтернативно, по существу состоит или, альтернативно, состоит из рекомбинантных белков или их фрагментов, РНК-бактериофага Qβ или РНК-бактериофага fr.
В другом предпочтительном варианте данного изобретения рекомбинантные белки содержат или, альтернативно, по существу состоят или, альтернативно, состоят из покрывающих белков РНК-фага.
Покрывающие белки РНК-фага, образующие капсиды или VLP, или фрагменты покрывающих белков бактериофага, совместимые с самосборкой в капсид или VLP, таким образом, являются следующими предпочтительными вариантами данного изобретения. Покрывающие белки бактериофага Qβ, например, могут быть рекомбинантно экспрессированы в E. coli. Кроме того, при такой экспрессии указанные белки спонтанно образуют капсиды. Кроме того, указанные капсиды образуют структуру с присущей ей повторяющейся организацией.
Конкретные предпочтительные примеры покрывающих белков бактериофага, которые можно использовать для получения композиций согласно изобретению, включают покрывающие белки таких РНК-бактериофагов, как бактериофаг Qβ (SEQ ID NO:10; база данных PIR, инвентарный No. VCBPQβ, относящаяся к CP Qβ и SEQ ID NO:11; инвентарный No. AAA16663, относящаяся к белку A1 Qβ), бактериофаг R17 (SEQ ID NO:12; PIR, инвентарный No. VCBPR7), бактериофаг fr (SEQ ID NO:13; PIR, инвентарный No. VCBPFR), бактериофаг GA (SEQ ID NO:14; GenBank, инвентарный No. NP-040754), бактериофаг SP (SEQ ID NO:15; GenBank, инвентарный No. CAA30374, относящаяся к CP SP, и SEQ ID NO:16; инвентарный No. NP 695026, относящаяся к белку A1 SP), бактериофаг MS2 (SEQ ID NO:17; PIR, инвентарный No. VCBPM2), бактериофаг M11 (SEQ ID NO:18; GenBank, инвентарный No. AAC06250), бактериофаг MX1 (SEQ ID NO:19; GenBank, инвентарный No. AAC14699), бактериофаг NL95 (SEQ ID NO:20; GenBank, инвентарный No. AAC14704), бактериофаг f2 (SEQ ID NO:21; GenBank, инвентарный No. P03611), бактериофаг PP7 (SEQ ID NO:22). Кроме того, белок A1 бактериофага Qβ или C-концевые укороченные формы, в которых отсутствуют до 100, 150 или 180 аминокислот из его C-конца, могут быть включены в сборку капсида из покрывающих белков Qβ. Как правило, процентное содержание белка A1 Qβ относительно CP Qβ в собранной конструкции капсида будет ограничено, чтобы обеспечить образование капсида.
Также обнаружено, что покрывающий белок Qβ подвергается самосборке в капсиды при экспрессии в E. coli (Kozlovska TM. et al., GENE 137: 133-137 (1993)). Полученные капсиды или вирусоподобные частицы имели икосаэдрическую структуру капсида, подобную структуре фага, с диаметром 25 нм и квазисимметрию T=3. Кроме того, определена кристаллическая структура фага Qβ. Капсид содержит 180 копий покрывающего белка, которые связаны в ковалентные пентамеры и гексамеры дисульфидными мостиками (Golmohammadi, R. et al., Structure 4: 543-5554 (1996)), обеспечивая поразительную стабильность капсида из покрывающего белка Qβ. Однако капсиды или VLP, полученные из рекомбинантного покрывающего белка Qβ, могут содержать субъединицы, не связанные дисульфидными связями с другими субъединицами в капсиде или не полностью связанные. Таким образом, при загрузке рекомбинантного капсида Qβ на невосстанавливающий SDS-ПААГ видны полосы, соответствующие мономерному покрывающему белку Qβ, а также полосы, соответствующие гексамеру или пентамеру покрывающего белка Qβ. Не полностью связанные дисульфидными связями субъединицы могут быть видны в виде полосы димера, тримера или даже тетрамера в невосстанавливающем SDS-ПААГ. Белок капсида Qβ также проявляет необычную резистентность к органическим растворителям и денатурирующим агентам. Неожиданно авторы обнаружили, что такие высокие концентрации ДМСО и ацетонитрила как 30%, и такие высокие концентрации гуанидиния как 1 М, не влияют на стабильность капсида. Высокая стабильность капсида из покрывающего белка Qβ является признаком, дающим преимущество, в частности, для его применения при иммунизации и вакцинации млекопитающих и человека согласно данному изобретению.
При экспрессии в E. coli N-концевой метионин покрывающего белка Qβ обычно удаляется, как обнаружили авторы в результате N-концевого секвенирования по Эдману, как описано в Stoll, E. et al. J. Biol. Chem. 252: 990-993 (1977). VLP, состоящая из покрывающих белков Qβ, где удален N-концевой метионин, или VLP, содержащие смесь покрывающих белков Qβ, где N-концевой метионин либо отщеплен, либо присутствует, также входят в объем данного изобретения.
Кроме того, также показано, что покрывающие белки РНК-фага подвергаются самосборке при экспрессии в бактериальном хозяине (Kastelein, RA. et al., Gene 23: 245-254 (1983), Kozlovskaya, TM. et al., Dokl. Akad. Nauk SSSR 287: 452-455 (1986), Adhin, MR. et al., Virology 170: 238-242 (1989), Ni, CZ., et al., Protein Sci. 5: 2485-2493 (1996), Priano, C. et al., J. Mol. Biol. 249: 283-297 (1995)). Капсид фага Qβ, кроме покрывающего белка, содержит так называемый полностью считываемый белок A1 и белок созревания A2. A1 образуется в результате супрессии в стоп-кодоне UGA и имеет длину 329 а/к. Капсид рекомбинантного покрывающего белка фага Qβ, используемый в данном изобретении, лишен белка лизиса A2 и содержит РНК из хозяина. Покрывающий белок РНК-фагов является РНК-связывающим белком и взаимодействует со стволовой петлей сайта связывания рибосомы гена репликазы, действуя в качестве репрессора трансляции в ходе жизненного цикла вируса. Последовательность и структурные элементы взаимодействия известны (Witherell, GW. and Uhlenbeck, OC. Biochemistry 28: 71-76 (1989); Lim F. et al., J. Biol. Chem. 271: 31839-31845 (1996)). Известно, что стволовая петля и РНК в общем вовлечены в сборку вируса (Golmohammadi, R. et al., Structure 4: 543-5554 (1996)).
В следующем предпочтительном варианте данного изобретения вирусоподобная частица содержит или, альтернативно, по существу состоит или, альтернативно, состоит из рекомбинантных белков или их фрагментов РНК-фага, где рекомбинантные белки содержат, по существу состоят или, альтернативно, состоят из мутантных покрывающих белков РНК-фага, предпочтительно мутантных покрывающих белков РНК-фагов, указанных выше. В другом предпочтительном варианте мутантные покрывающие белки РНК-фага модифицированы удалением по меньшей мере одного остатка лизина путем замены или добавлением по меньшей мере одного остатка лизина путем замены; альтернативно мутантные покрывающие белки РНК-фага модифицированы делецией по меньшей мере одного остатка лизина или добавлением по меньшей мере одного остатка лизина посредством инсерции.
В другом предпочтительном варианте вирусоподобная частица содержит или, альтернативно, по существу состоит или, альтернативно, состоит из рекомбинантных белков или их фрагментов РНК-бактериофага Qβ, где рекомбинантные белки содержат или, альтернативно, по существу состоят, или, альтернативно, состоят из покрывающих белков, имеющих аминокислотную последовательность SEQ ID NO:10, или смеси покрывающих белков, имеющих аминокислотные последовательности SEQ ID NO:10 и SEQ ID NO:11, или мутантов SEQ ID NO:11, и где N-концевой метионин предпочтительно отщеплен.
В следующем предпочтительном варианте данного изобретения вирусоподобная частица содержит, по существу состоит или, альтернативно, состоит из рекомбинантных белков Qβ или их фрагментов, где рекомбинантные белки содержат или альтернативно, по существу состоят или, альтернативно, состоят из мутантных покрывающих белков Qβ. В другом предпочтительном варианте указанные мутантные покрывающие белки модифицированы удалением по меньшей мере одного остатка лизина посредством замены или добавлением по меньшей мере одного остатка лизина посредством замены. Альтернативно указанные мутантные покрывающие белки модифицированы делецией по меньшей мере одного остатка лизина или добавлением по меньшей мере одного остатка лизина посредством инсерции.
На поверхности капсида из покрывающего белка Qβ экспонированы четыре остатка лизина. Мутанты Qβ, для получения которых экспонированные остатки лизина заменяют аргининами, также могут быть использованы для данного изобретения. Таким образом, в практике изобретения могут быть использованы следующие мутанты покрывающего белка Qβ и мутантные VLP Qβ: «Qβ-240» (Lys13-Arg; SEQ ID NO:23), «Qβ-243» (Asn 10-Lys; SEQ ID NO:24), «Qβ-250» (Lys 2-Arg, Lys13-Arg; SEQ ID NO:25), «Qβ-251» (SEQ ID NO:26) и «Qβ-259» (Lys 2-Arg, Lys16-Arg; SEQ ID NO:27). Таким образом, в следующем предпочтительном варианте данного изобретения вирусоподобная частица содержит, по существу состоит или, альтернативно, состоит из рекомбинантных белков мутантных покрывающих белков Qβ, которые содержат белки, имеющие аминокислотную последовательность, выбранную из следующей группы: a) аминокислотной последовательности SEQ ID NO:23; b) аминокислотной последовательности SEQ ID NO:24; c) аминокислотной последовательности SEQ ID NO:25; d) аминокислотной последовательности SEQ ID NO:26; и e) аминокислотной последовательности SEQ ID NO:27. Конструирование, экспрессия и очистка указанных выше покрывающих белков Qβ, VLP и капсидов из мутантных покрывающих белков Qβ, соответственно, описаны в находящейся на рассмотрении заявке на выдачу патента США № 10/050902, поданной настоящим правопреемником 18 января 2002. В частности, ссылка относится к примеру 18 указанной выше заявки.
В следующем предпочтительном варианте данного изобретения вирусоподобная частица содержит или, альтернативно, по существу состоит или, альтернативно, состоит из рекомбинантных белков Qβ или их фрагментов, где рекомбинантные белки содержат, по существу состоят или, альтернативно, состоят из смеси любого одного из вышеупомянутых мутантов Qβ и соответствующего белка A1.
В следующем предпочтительном варианте данного изобретения вирусоподобная частица содержит или, альтернативно, по существу состоит или, альтернативно, состоит из рекомбинантных белков РНК-фага AP205 или их фпагментов.
Геном AP205 состоит из белка созревания, покрывающего белка, репликазы и двух открытых рамок считывания, не присутствующих в родственных фагах; гена лизиса и открытой рамки считывания, играющей роль в трансляции гена созревания (Klovins,J., et al., J. Gen. Virol. 83: 1523-33 (2002)). Покрывающий белок AP205 можно экспрессировать с плазмиды pAP283-58 (SEQ ID NO:111), которая является производной pQb10 (Kozlovska, T.M., et al., Gene 137: 133-37 (1993)) и которая содержит сайт связывания рибосомы AP205. Альтернативно покрывающий белок AP205 может быть клонирован в pQb185 ниже сайта связывания рибосомы, присутствующего в векторе. Оба способа приводят к экспрессии белка и образованию капсидов, как описано в одновременно находящейся на рассмотрении предварительной заявке на выдачу патента США № 60/396126, имеющей название «Молекулярные антигенные матрицы» и поданной настоящим правопреемником 17 июля 2002, которая включена в виде ссылки в полном объеме. Векторы pQb10 и pQb185 являются векторами, полученными из вектора pGEM, и экспрессия клонированных генов в указанных векторах контролируется промотором trp (Kozlovska, T.M., et al., Gene 137:133-37 (1993)). Плазмида pAP283-58 (SEQ ID NO:111) содержит предполагаемый сайт связывания рибосомы AP205 в следующей последовательности, которая расположена ниже сайта XbaI и непосредственно выше стартового кодона ATG покрывающего белка AP205: tctagaATTTTCTGCGCACCCATCCCGGGTGGCGCCCAAAGTGAGGAAAATCACatg (SEQ ID NO:115). Вектор pQb185 содержит последовательность Шайна-Далгарно ниже сайта XbaI и выше стартового кодона (tctagaTTAACCCAACGCGTAGGAGTCAGGCCatg (SEQ ID NO:116), последовательность Шайна-Далгарно подчеркнута).
В следующем предпочтительном варианте данного изобретения вирусоподобная частица содержит или, альтернативно, по существу состоит или, альтернативно состоит из рекомбинантных покрывающих белков РНК-фага AP205 или их фрагментов.
Таким образом, указанный предпочтительный вариант данного изобретения включает в себя покрывающие белки AP205, которые образуют капсиды. Такие белки экспрессируют рекомбинантно или получают из природных источников. Покрывающие белки AP205, продуцированные в бактериях, спонтанно образуют капсиды, что подтверждается электронной микроскопией (ЭМ) и иммунодиффузией. Структурные особенности капсида, образованного покрывающим белком AP205 (SEQ ID NO:112), и особенности капсида, образованного покрывающим белком РНК-фага AP205, почти не отличаются при наблюдении в ЭМ. VLP AP205 являются высокоиммуногенными и могут быть связаны с антигенами и/или антигенными детерминантами с образованием конструкций вакцин, экспонирующих антигены и/или антигенные детерминанты, ориентированные повторяющимся образом. Против экспонированных таким образом антигенов вырабатываются высокие титры антител, что свидетельствует о том, что связанные антигены и/или антигенные детерминанты доступны для взаимодействия с молекулами антител и являются иммуногенными.
В следующем предпочтительном варианте данного изобретения вирусоподобная частица содержит или, альтернативно, по существу состоит, или, альтернативно, состоит из рекомбинантных мутантных покрывающих белков РНК-фага AP205 или их фрагментов.
Компетентные в отношении сборки мутантные формы VLP AP205, включая покрывающий белок AP205 с заменой пролина в положении аминокислоты 5 на треонин (SEQ ID NO:113), также можно использовать в практике изобретения, и они составляют следующий предпочтительный вариант изобретения. Указанные VLP, VLP AP205, полученные из природных источников, или вирусные частицы AP205 могут быть связаны с антигенами с образованием упорядоченных повторяющихся матриц антигенов согласно данному изобретению.
Мутантный покрывающий белок P5-T AP205 может быть экспрессирован из плазмиды pAP281-32 (SEQ ID No. 114), которая получена непосредственно из pQb185 и которая содержит ген мутантного покрывающего белка AP205 вместо гена покрывающего белка Qβ. Векторы для экспрессии покрывающего белка AP205 трансфицировали в E. coli для экспрессии покрывающего белка AP205.
Способы экспрессии покрывающего белка и, соответственно, мутантного покрывающего белка, приводящей к самосборке в VLP, описаны в совместно рассматриваемой предварительной заявке на выдачу патента США, имеющей название «Молекулярные антигенные матрицы» и поданной настоящим правопреемником 17 июля 2002, которая включена в виде ссылки в полном объеме. Подходящие штаммы E. coli включают, но не ограничены указанным, E. coli K802, JM 109, RR1. Подходящие векторы и штаммы и их комбинации можно идентифицировать в результате тестирования экспрессии покрывающего белка и, соответственно, мутантного покрывающего белка с помощью SDS-ПААГ и образования и сборки капсида сначала с помощью необязательной очистки капсидов гель-фильтрацией и затем их тестированием в анализе иммунодиффузии (тест Оухтерлони) или с помощью электронной микроскопии (Kozlovska, T.M., et al., Gene 137:133-37 (1993)).
Покрывающие белки AP205, экспрессированные с векторов pAP283-58 и pAP281-32, могут быть лишены начальной аминокислоты метионина вследствие процессинга в цитоплазме E. coli. Расщепленные, нерасщепленные формы VLP AP205 или их смеси являются следующими предпочтительными вариантами изобретения.
В следующем предпочтительном варианте данного изобретения вирусоподобная частица содержит или, альтернативно, по существу состоит или, альтернативно, состоит из смеси рекомбинантных покрывающих белков РНК-фага AP205 или их фрагментов и рекомбинантных мутантных покрывающих белков РНК-фага AP205 или их фрагментов.
В следующем предпочтительном варианте данного изобретения вирусоподобная частица содержит или, альтернативно, по существу состоит или, альтернативно, состоит из фрагментов рекомбинантных покрывающих белков или рекомбинантных мутантных покрывающих белков РНК-фага AP205.
Фрагменты рекомбинантного покрывающего белка AP205, способные собираться в VLP и, соответственно, капсид, также применимы в практике изобретения. Указанные фрагменты можно образовать путем делеции либо внутри, либо на концах покрывающего белка и мутантного покрывающего белка, соответственно. Инсерции в последовательность покрывающего белка и мутантного покрывающего белка или слияния антигенных последовательностей с последовательностью покрывающего белка и мутантного покрывающего белка, которые совместимы со сборкой в VLP, являются следующими вариантами изобретения, которые приводят к химерным покрывающим белкам AP205 и частицам, соответственно. Последствия инсерций, делеций и слияний с последовательностью покрывающего белка, и то, совместимо ли это со сборкой в VLP, можно определить с помощью электронной микроскопии.
Частицы, образованные покрывающим белком, фрагментами покрывающего белка или химерными покрывающими белками AP205, описанными выше, можно выделить в чистой форме в результате комбинирования стадий фракционирования путем осаждения и стадий очистки гель-фильтрацией с использованием колонок, например, с сефарозой CL-4B, сефарозой CL-2B, сефарозой CL-6B и их комбинаций, как описано в совместно рассматриваемой предварительной заявке на выдачу патента США, имеющей название «Молекулярные антигенные матрицы» и поданной настоящим правопреемником 17 июля 2002, которая включена в виде ссылки в полном объеме. Другие способы выделения вирусоподобных частиц известны в данной области и могут быть использованы для выделения вирусоподобных частиц (VLP) бактериофага AP205. Например, применение ультрацентрифугирования для выделения VLP дрожжевого ретротранспозона Ty описано в патенте США № 4918166, который включен в данное описание в виде ссылки в полном объеме.
Определена кристаллическая структура нескольких РНК-бактериофагов (Golmohammadi, R. et al., Structure 4: 543-554 (1996)). Используя указанную информацию, можно идентифицировать экспонированные на поверхности остатки и, таким образом, покрывающие белки РНК-фага можно модифицировать так, чтобы один или несколько реакционно-способных аминокислотных остатков можно было встроить с помощью инсерции или замены. В итоге указанные модифицированные формы покрывающих белков бактериофага также можно использовать для данного изобретения. Таким образом, варианты белков, которые образуют капсиды или капсидоподобные структуры (например, покрывающие белки бактериофага Qβ, бактериофага R17, бактериофага fr, бактериофага GA, бактериофага SP и бактериофага MS2), также можно использовать для получения композиций согласно данному изобретению.
Хотя последовательность вариантов белков, обсуждаемых выше, будет отличаться от их аналогов дикого типа, указанные варианты белков, как правило, будут сохранять способность образовывать капсиды или капсидоподобные структуры. Таким образом, изобретение, кроме того, включает в себя композиции и, соответственно, композиции вакцин, которые, кроме того, содержат варианты белков, которые образуют капсиды или капсидоподобные структуры, а также способы получения таких композиций и, соответственно, композиций вакцин, отдельные субъединицы белка, используемые для получения таких композиций, и молекулы нуклеиновых кислот, которые кодируют указанные субъединицы белка. Таким образом, в объем изобретения включены вариантные формы белков дикого типа, которые образуют капсиды или капсидоподобные структуры и сохраняют способность вступать в ассоциацию и образовать капсиды или капсидоподобные структуры.
В результате изобретение, кроме того, включает в себя композиции и, соответственно, композиции вакцин, содержащие белки, которые содержат или, альтернативно, по существу состоят или, альтернативно, состоят из аминокислотных последовательностей, которые по меньшей мере на 80%, 85%, 90%, 95%, 97% или 99% идентичны белкам дикого типа, которые образуют упорядоченные матрицы и, соответственно, матрицы, имеющие присущую им повторяющуюся структуру.
Кроме того, в объем изобретения включены молекулы нуклеиновых кислот, которые кодируют белки, используемые для получения композиций согласно данному изобретению.
В других вариантах изобретение, кроме того, относится к композициям, содержащим белки, которые содержат или, альтернативно, по существу состоят или, альтернативно, состоят из аминокислотных последовательностей, которые по меньшей мере на 80%, 85%, 90%, 95%, 97% или 99% идентичны любой из аминокислотных последовательностей, показанных в SEQ ID NO:10-27.
Белки, подходящие для применения в данном изобретении, также включают укороченные на C-конце мутанты белков, которые образуют капсиды или капсидоподобные структуры или VLP. Конкретные примеры таких укороченных мутантов включают белки, имеющие аминокислотную последовательность, показанную в любой из SEQ ID NO:10-27, где 1, 2, 5, 7, 9, 10, 12, 14, 15 или 17 аминокислот были удалены из C-конца. Как правило, указанные укороченные на C-конце мутанты будут сохранять способность образовывать капсиды или капсидоподобные структуры.
Следующие белки, подходящие для применения в данном изобретении, также включают укороченные на N-конце мутанты белков, которые образуют капсиды или капсидоподобные структуры. Конкретные примеры таких укороченных мутантов включают белки, имеющие аминокислотную последовательность, показанную в любой из SEQ ID NO:10-27, где 1, 2, 5, 7, 9, 10, 12, 14, 15 или 17 аминокислот были удалены из N-конца. Как правило, указанные укороченные на N-конце мутанты будут сохранять способность образовывать капсиды или капсидоподобные структуры.
Дополнительные белки, подходящие для применения в данном изобретении, включают укороченные на N- и C-конце мутанты, которые образуют капсиды или капсидоподобные структуры. Подходящие укороченные мутанты включают белки, имеющие аминокислотную последовательность, показанную в любой из SEQ ID NO:10-27, где 1, 2, 5, 7, 9, 10, 12, 14, 15 или 17 аминокислот были удалены из N-конца и 1, 2, 5, 7, 9, 10, 12, 14, 15 или 17 аминокислот были удалены с C-конца. Как правило, указанные укороченные на N-конце и на C-конце мутанты будут сохранять способность образовывать капсиды или капсидоподобные структуры.
Изобретение, кроме того, относится к композициям, содержащим белки, которые содержат или, альтернативно, по существу состоят или, альтернативно, состоят из аминокислотных последовательностей, которые по меньшей мере на 80%, 85%, 90%, 95%, 97% или 99% идентичны описанным выше укороченным мутантам.
Таким образом, изобретение относится к композициям и композициям вакцин, полученным из белков, которые образуют капсиды или VLP, способам получения указанных композиций из отдельных белковых субъединиц и VLP или капсидов, способам получения указанных отдельных белковых субъединиц, молекул нуклеиновой кислоты, которые кодируют указанные субъединицы, и способам вакцинации и/или способам, вызывающим иммунологические ответы у индивидуумов, с использованием указанных композиций согласно данному изобретению.
Как указано ранее, изобретение включает в себя вирусоподобные частицы или их рекомбинантные формы. В одном предпочтительном варианте частицы, используемые в композициях согласно изобретению, состоят из корового белка гепатита B (HBcAg) или фрагмента HBcAg. В следующем варианте частицы, используемые в композициях согласно изобретению, состоят из корового белка гепатита B (HBcAg) или фрагмента белка HBcAg, который был модифицирован, с тем, чтобы либо удалить, либо уменьшить количество свободных остатков цистеина. Zhou et al. (J. Virol. 66: 5393-5398 (1992)) показали, что HBcAg, которые были модифицированы, с тем, чтобы удалить имеющиеся в природных условиях остатки цистеина, сохраняют способность вступать в ассоциацию и образовывать капсиды. Таким образом, VLP, подходящие для применения в композициях согласно изобретению, включают в себя VLP, содержащие модифицированные HBcAg или их фрагменты, в которых один или несколько имеющихся в природных условиях остатков цистеина были либо делетированы, либо заменены другим аминокислотным остатком (например, остатком серина).
HBcAg является белком, образуемым в результате процессинга белка-предшественника корового антигена гепатита B. Идентифицирован ряд изотипов HBcAg, и их аминокислотные последовательности легко доступны специалистам в данной области. В большинстве случаев композиции и, соответственно, композиции вакцин согласно изобретению могут быть получены с использованием процессированной формы HBcAg (т.е. HBcAg, из которого была удалена N-концевая лидерная последовательность белка-предшественника корового антигена гепатита B).
Кроме того, когда HBcAg будут получать в условиях, при которых процессинг не будет происходить, HBcAg, как правило, будет экспрессироваться в «процессированной» форме. Например, когда для получения HBcAg согласно изобретению будет использоваться система экспрессии E. coli, направляющая экспрессию белка в цитоплазме, указанные белки, как правило, будут экспрессироваться так, чтобы в них отсутствовала N-концевая лидерная последовательность белка-предшественника корового антигена гепатита B.
Получение вирусоподобных частиц гепатита B, которые можно использовать для данного изобретения, описано, например, в WO 00/32227 и в указанной заявке, в частности, в примерах с 17 по 19 и с 21 по 24, а также в WO 01/85208 и в ней, в частности, в примерах с 17 по 19, с 21 по 24, с 31 по 41, и в находящейся на рассмотрении заявке на выдачу патента США № 10/050902, поданной настоящим правопреемником 18 января 2002. В случае последней заявки это, в частности, относится к примеру 23, 24, 31 и 51. Все три документа включены в данное описание в виде ссылки в полном объеме.
Данное изобретение также включает в себя варианты HBcAg, которые были модифицированы, чтобы делетировать или заменить один или несколько дополнительных остатков цистеина. В данной области известно, что свободные остатки цистеина могут быть вовлечены в ряд побочных химических реакций. Указанные побочные реакции включают в себя дисульфидные обмены, реакцию с химическими веществами или метаболитами, которые, например, инъецированы или образованы при комбинированной терапии с другими веществами, или прямое окисление и реакцию с нуклеотидами при воздействии УФ-света. Таким образом, могут быть образованы токсичные аддукты, особенно если учесть тот факт, что HBcAg имеют устойчивую тенденцию к связыванию нуклеиновых кислот. Таким образом, токсичные аддукты могут быть представлены множеством видов, каждый из которых отдельно может присутствовать в низкой концентрации, но вместе они достигают токсичных уровней.
С точки зрения указанного выше, одним их преимуществ применения в композициях вакцин HBcAg, которые были модифицированы, чтобы удалить имеющиеся в природных условиях остатки цистеина, является то, что количество сайтов, с которыми могут связываться токсичные элементы в том случае, когда связаны антигены или антигенные детерминанты, может быть уменьшено, или они могут быть вообще удалены.
Идентифицирован ряд вариантов HBcAg природного происхождения, подходящих для применения в практике данного изобретения. Yuan et al., (J. Virol. 73: 10122-10128 (1999)), например, описывают варианты, в которых остаток изолейцина в положении, соответствующем положению 97 в SEQ ID NO:28, заменен либо остатком лейцина, либо остатков фенилаланина. Аминокислотные последовательности ряда вариантов HBcAg, а также нескольких вариантов предшественника корового антигена гепатита B описаны в GenBank, запись AAF121240 (SEQ ID NO:29), AF121239 (SEQ ID NO:30), X85297 (SEQ ID NO:31), X02496 (SEQ ID NO:32), X85305 (SEQ ID NO:33), X85303 (SEQ ID NO:34), AF151735 (SEQ ID NO:35), X85259 (SEQ ID NO:36), X85286 (SEQ ID NO:37), X85260 (SEQ ID NO:38), X85317 (SEQ ID NO:39), X85298 (SEQ ID NO:40), AF043593 (SEQ ID NO:41), M20706 (SEQ ID NO:42), X85295 (SEQ ID NO:43), X80925 (SEQ ID NO:44), X85284 (SEQ ID NO:45), X85275 (SEQ ID NO:46), X72702 (SEQ ID NO:47), X85291 (SEQ ID NO:48), X65258 (SEQ ID NO:49), X85302 (SEQ ID NO:50), M32138 (SEQ ID NO:51), X85293 (SEQ ID NO:52), X85315 (SEQ ID NO:53), U95551 (SEQ ID NO:54), X85256 (SEQ ID NO:55), X85316 (SEQ ID NO:56), X85296 (SEQ ID NO:57), AB033559 (SEQ ID NO:58), X59795 (SEQ ID NO:59), X85299 (SEQ ID NO:60), X85307 (SEQ ID NO:61), X65257 (SEQ ID NO:62), X85311 (SEQ ID NO:63), X85301 (SEQ ID NO:64), X85314 (SEQ ID NO:65), X85287 (SEQ ID NO:66), X85272 (SEQ ID NO:67), X85319 (SEQ ID NO:68), AB010289 (SEQ ID NO:69), X85285 (SEQ ID NO:70), AB010289 (SEQ ID NO:71), AF121242 (SEQ ID NO:72), M90520 (SEQ ID NO:73), P03153 (SEQ ID NO:74), AF110999 (SEQ ID NO:75) и M95589 (SEQ ID NO:76), описания каждой из которых включены в данную заявку в виде ссылки. Указанные варианты HBcAg отличаются по аминокислотной последовательности в ряде положений, включая аминокислотные остатки, которые соответствуют аминокислотным остаткам, распложенным в положениях 12, 13, 21, 22, 24, 29, 32, 33, 35, 38, 40, 42, 44, 45, 49, 51, 57, 58, 59, 64, 66, 67, 69, 74, 77, 80, 81, 87, 92, 93, 97, 98, 100, 103, 105, 106, 109, 113, 116, 121, 126, 130, 133, 135, 141, 147, 149, 157, 176, 178, 182 и 183 в SEQ ID NO:77. Следующие варианты HBcAg, подходящие для применения в композициях согласно изобретению и которые могут быть дополнительно модифицированы согласно изложенному в данном описании, описаны в WO 01/98333, WO 01/77158 и WO 02/14478.
Как указано выше, как правило, в композициях и, соответственно, композициях вакцин согласно изобретению будут использованы процессированные HBcAg (т.е. те, в которых отсутствуют лидерные последовательности). Данное изобретение включает в себя композиции вакцин, а также способы применения таких композиций, в которых используются описанные выше варианты HBcAg.
Имеет ли аминокислотная последовательность полипептида аминокислотную последовательность, которая по меньшей мере на 80%, 85%, 90%, 95%, 97% или 99% идентична одной из указанных выше аминокислотных последовательностей дикого типа или ее части, можно определить стандартным способом с использованием известных компьютерных программ, таких как программа Bestfit. При использовании Bestfit или любой другой программы выравнивания последовательностей для того, чтобы определить, идентична ли конкретная последовательность, например, на 95% эталонной аминокислотной последовательности, параметры устанавливают так, чтобы процент идентичности рассчитывался по всей полной длине эталонной аминокислотной последовательности и чтобы были разрешены пробелы в гомологии до 5% от общего количества аминокислотных остатков в эталонной последовательности.
Варианты и предшественники HBcAg, имеющие аминокислотные последовательности, указанные в SEQ ID NO:29-72 и 73-76, относительно сходны друг с другом. Таким образом, ссылка на аминокислотный остаток варианта HBcAg, расположенный в положении, которое соответствует конкретному положению в SEQ ID NO:77, относится к аминокислотному остатку, который присутствует в данном положении в аминокислотной последовательности, показанной в SEQ ID NO:77. Гомология между указанными вариантами HBcAg среди вирусов гепатита B, которые инфицируют млекопитающих, в большинстве случаев достаточно высока, так что специалист в данной области не будет иметь больших трудностей при просмотре обеих последовательностей, аминокислотной последовательности, показанной в SEQ ID NO:77, и аминокислотной последовательности конкретного варианта HBcAg, и при идентификации «соответствующих» аминокислотных остатков. Кроме того, аминокислотная последовательность HBcAg, показанная в SEQ ID NO:73, в которой представлена аминокислотная последовательность HBcAg, полученного из вируса, который инфицирует лесных сурков, имеет достаточно высокую гомологию с HBcAg, имеющим аминокислотную последовательность, показанную в SEQ ID NO:77, так что легко увидеть, что инсерция из трех аминокислотных остатков присутствует в SEQ ID NO:64 между остатками аминокислот 155 и 156 SEQ ID NO:77.
Изобретение также включает в себя композиции вакцин, которые содержат варианты HBcAg вирусов гепатита B, которые инфицируют птиц, а также композиции вакцин, которые содержат фрагменты указанных вариантов HBcAg. В случае указанных вариантов HBcAg один, два, три или более остатков цистеина, присутствующих в указанных полипептидах в природных условиях, могут быть либо заменены другим аминокислотным остатком, либо делетированы перед их включением в композиции вакцин согласно изобретению.
Как обсуждается выше, удаление свободных остатков цистеина уменьшает количество сайтов, в которых токсичные компоненты могут связываться с HBcAg, а также удаляет сайты, в которых может происходить перекрестное связывание остатков лизина и цистеина одной и той же или соседних молекул HBcAg. Таким образом, в другом варианте данного изобретения один или несколько остатков цистеина белка капсида вируса гепатита B либо делетированы, либо заменены другим аминокислотным остатком.
В других вариантах композиции и, соответственно, композиции вакцин согласно изобретению будут содержать HBcAg, из которых была удалена C-концевая область (например, аминокислотные остатки 145-185 или 150-185 SEQ ID NO:77). Таким образом, дополнительные модифицированные HBcAg, подходящие для применения в практике данного изобретения, включают укороченные на C-конце мутанты. Подходящие укороченные мутанты включают HBcAg, в которых 1, 5, 10, 15, 20, 25, 30, 34, 35 аминокислот были удалены из C-конца.
HBcAg, подходящие для применения в практике данного изобретения, также включают укороченные на N-конце мутанты. Подходящие укороченные мутанты включают модифицированные HBcAg, в которых 1, 2, 5, 7, 9, 10, 12, 14, 15 или 17 аминокислот были удалены из N-конца.
Следующие HBcAg, подходящие для применения в практике данного изобретения, включают укороченные на N- и C-концах мутанты. Подходящие укороченные мутанты включают HBcAg, в которых 1, 2, 5, 7, 9, 10, 12, 14, 15 и 17 аминокислот были удалены из N-конца и 1, 5, 10, 15, 20, 25, 30, 34, 35 аминокислот были удалены из C-конца.
Изобретение, кроме того, включает в себя композиции и, соответственно, композиции вакцин, содержащие полипептиды HBcAg, содержащие или, альтернативно, по существу состоящие или, альтернативно, состоящие из аминокислотных последовательностей, которые по меньшей мере на 80%, 85%, 90%, 95%, 97% или 99% идентичны описанным выше укороченным мутантам.
В некоторых вариантах согласно изобретению в полипептид HBcAg вводят остаток лизина, чтобы опосредовать связывание белка RANKL, фрагмента RANKL или пептида RANKL с VLP HBcAg. В предпочтительных вариантах композиции согласно изобретению получают с использованием HBcAg, содержащего или, альтернативно, состоящего из аминокислот 1-144 или 1-149, 1-185 SEQ ID NO:77, который модифицирован так, чтобы аминокислоты, соответствующие положениям 79 и 80 были заменены пептидом, имеющим аминокислотную последовательность Gly-Gly-Lys-Gly-Gly (SEQ ID NO:117; SEQ ID NO:78). В следующих предпочтительных вариантах остатки цистеина в положениях 48 и 107 SEQ ID NO:77 мутированы в серин. Изобретение, кроме того, включает в себя композиции, содержащие соответствующие полипептиды, имеющие аминокислотные последовательности, показанные в любой из SEQ ID NO:29-74, которые также имеют указанные выше изменения аминокислот. Кроме того, в объем изобретения включены дополнительные варианты HBcAg, которые способны ассоциировать с образованием капсида или VLP и имеют указанные выше изменения аминокислот. Таким образом, изобретение, кроме того, включает в себя композиции и, соответственно, композиции вакцин, содержащие полипептиды HBcAg, которые содержат или, альтернативно, состоят из аминокислотных последовательностей, которые по меньшей мере на 80%, 85%, 90%, 95%, 97% или 99% идентичны любой из аминокислотных последовательностей дикого типа, и формы указанных белков, которые были процессированы в подходящих для этого случаях, чтобы удалить N-концевую лидерную последовательность, и модифицированы указанными выше изменениями.
Композиции или композиции вакцин согласно изобретению могут содержать смеси различных HBcAg. Таким образом, указанные композиции вакцин могут состоять из HBcAg, которые отличаются по аминокислотной последовательности. Например, могут быть получены композиции вакцин, содержащие HBcAg «дикого типа» и модифицированный HBcAg, в котором один или несколько аминокислотных остатков были изменены (например, делетированы, встроены или заменены). Кроме того, предпочтительными композициями вакцин согласно изобретению являются композиции, которые представляют собой высокоупорядоченную и повторяющуюся антигенную матрицу, в которой антигеном является белок RANKL, фрагмент RANKL или пептид RANKL.
В следующем предпочтительном варианте данного изобретения по меньшей мере один белок RANKL, фрагмент RANKL или пептид RANKL связан с указанной центральной частицей и, соответственно, вирусоподобной частицей по меньшей мере одной ковалентной связью. Предпочтительно по меньшей мере один белок RANKL, фрагмент RANKL или пептид RANKL связан с центральной частицей и, соответственно, с вирусоподобной частицей по меньшей мере одной ковалентной связью, при этом указанная ковалентная связь является непептидной связью, что приводит к образованию упорядоченной и повторяющейся матрицы центральная частица-RANKL и, соответственно, RANKL-VLP-матрицы или -конъюгата. Данная матрица или, соответственно, конъюгат RANKL-VLP обычно и предпочтительно имеет повторяющуюся и упорядоченную структуру, так как по меньшей мере один, но обычно несколько белков RANKL, фрагментов RANKL или пептидов RANKL связаны с VLP ориентированным образом. Предпочтительно более 10, 20, 40, 80, 120 белков RANKL, фрагментов RANKL или пептидов RANKL связаны с VLP. Образование повторяющейся и упорядоченной RANKL-VLP-матрицы и, соответственно, конъюгата, обеспечивается ориентированным и направленным, а также определенным связыванием и, соответственно, присоединением по меньшей мере одного белка RANKL, фрагмента RANKL или пептида RANKL к VLP, как будет понятно из нижеследующего. Кроме того, обычная присущая высокоповторяющаяся и организованная структура VLP преимущественно вносит вклад в экспонирование белка RANKL, фрагмента RANKL или пептида RANKL высокоупорядоченным и повторяющимся образом, приводя к образованию высокоорганизованной и повторяющейся RANKL-VLP-матрицы и, соответственно, RANKL-VLP-конъюгата.
Таким образом, предпочтительные конъюгаты и, соответственно, матрицы согласно изобретению отличаются от конъюгатов предшествующего уровня техники своей высокоорганизованной структурой, размерами и повторяемостью антигена на поверхности матрицы. Предпочтительный вариант данного изобретения, кроме того, позволяет осуществлять экспрессию как частицы, так и антигена в экспрессирующем хозяине, обеспечивая правильную упаковку антигена, т.е. по меньшей мере одного белка RANKL, фрагмента RANKL или пептида RANKL и правильную упаковку и сборку VLP.
В данном изобретении заявлены способы связывания белка RANKL, фрагмента RANKL или пептида RANKL с центральными частицами и, соответственно, с VLP. Как указано в одном аспекте данного изобретения белок RANKL, фрагмент RANKL или пептид RANKL связан с центральной частицей и, соответственно, с VLP с помощью химического перекрестного связывания обычно и предпочтительно с использованием гетеробифункционального перекрестно сшивающего агента. В данной области известно несколько гетеробифункциональных перекрестно сшивающих агентов. В предпочтительных вариантах гетеробифункциональный перекрестно сшивающий агент содержит функциональную группу, которая может взаимодействовать с предпочтительными первыми сайтами связывания, т.е. с аминогруппой боковой цепи остатков лизина центральной частицы и VLP или по меньшей мере одной субъединицы VLP, соответственно, и дополнительную функциональную группу, которая может взаимодействовать с предпочтительным вторым сайтом связывания, т.е. остатком цистеина, имеющимся в природных условиях, который сделан доступным для реакции посредством восстановления, или сконструированным на белке RANKL, фрагменте RANKL или пептиде RANKL и необязательно также сделанным доступным для реакции посредством восстановления. Первая стадия способа, обычно называемая дериватизацией, представляет собой взаимодействие центральной частицы или VLP с перекрестно сшивающим агентом. Продуктом данной реакции является активированная центральная частица или активированная VLP, также называемая активированным носителем. На второй стадии непрореагировавший перекрестно сшивающий агент удаляют, используя обычные способы, такие как гель-фильтрация или диализ. На третьей стадии осуществляют взаимодействие белка RANKL, фрагмента RANKL или пептида RANKL с активированным носителем и данную стадию обычно называют стадией связывания. Непрореагировавший белок RANKL, фрагмент RANKL или пептид RANKL необязательно можно удалить на четвертой стадии, например, диализом. В данной области известно несколько гетеробифункциональных перекрестно сшивающих агентов. Указанные агенты включают предпочтительные перекрестно сшивающие агенты SMPH (Pierce), сульфо-MBS, сульфо-EMCS, сульфо-GMBS, сульфо-SIAB, сульфо-SMPB, сульфо-SMCC, SVSB, SIA и другие перекрестно сшивающие агенты, доступные, например, из Pierce Chemical Company (Rockford, IL, USA) и имеющие одну функциональную группу, реакционно-способную по отношению к аминогруппам, и одну функциональную группу, реакционно-способную по отношению к остаткам цистеина. Все указанные выше перекрестно сшивающие агенты приводят к образованию тиоэфирной связи. Другой класс перекрестно сшивающих агентов, подходящих для практики изобретения, характеризуется введением дисульфидной связи между белком RANKL, фрагментом RANKL или пептидом RANKL и центральной частицей или VLP при связывании. Предпочтительные перекрестно сшивающие агенты, относящиеся к данному классу, включают, например, SPDP и сульфо-LC-SPDP (Pierce). На степень дериватизации центральной частицы и, соответственно, VLP перекрестно сшивающим агентом можно влиять различными экспериментальными условиями, такими как концентрация каждого из партнеров реакции, избыток одного реагента по сравнению с другим, pH, температура и ионная сила. Степень связывания, т.е. количество белка RANKL, фрагмента RANKL или пептидов RANKL на субъединицу центральной частицы и, соответственно VLP, можно корректировать различными экспериментальными условиями, описанными выше, чтобы обеспечить соответствие требованиям вакцины. Растворимость белка RANKL, фрагмента RANKL или пептида RANKL может накладывать ограничение на количество белка RANKL, фрагмента RANKL или пептида RANKL, которое можно связать с каждой субъединицей, и в таких случаях, где полученная вакцина может быть нерастворимой, полезным может быть уменьшение количества белка RANKL, фрагмента RANKL или пептидов RANKL на субъединицу.
Особенно предпочтительным способом связывания белка RANKL, фрагмента RANKL или пептидов RANKL с центральной частицей и, соответственно, с VLP является связывание остатка лизина на поверхности центральной частицы и, соответственно, VLP с остатком цистеина на белке RANKL, фрагменте RANKL или пептиде RANKL. Таким образом, в предпочтительном варианте данного изобретения первым сайтом связывания является остаток лизина, а вторым сайтом связывания является остаток цистеина. В некоторых вариантах может требоваться конструирование аминокислотного линкера, содержащего остаток цистеина, в качестве второго сайта связывания или в качестве его части, к белку RANKL, фрагменту RANKL или пептиду RANKL для связывания с центральной частицей и, соответственно, с VLP. Альтернативно цистеин может быть введен в белок RANKL, фрагмент RANKL или пептид RANKL либо посредством инсерции, либо мутации. Альтернативно остаток цистеина или тиоловую группу можно ввести с помощью химического связывания.
Выбор аминокислотного линкера будет зависеть от природы антигена и, соответственно, аутоантигена, т.е. от природы белка RANKL, фрагмента RANKL или пептида RANKL, от его биохимических свойств, таких как pI, распределение заряда и гликозилирование. В общем, предпочтительны гибкие аминокислотные линкеры. Предпочтительные варианты аминокислотного линкера выбраны из группы, состоящей из: (a) CGG; (b) N-концевого линкера гамма 1; (c) N-концевого линкера гамма 3; (d) шарнирных участков Ig; (e) N-концевых глициновых линкеров; (f) (G)kC(G)n при n=0-12 и k=0-5; (g) N-концевых глицин-сериновых линкеров; (h) (G)kC(G)m(S)l(GGGGS)n (SEQ ID NO:118) при n=0-3, k=0-5, m=0-10, l=0-2; (i) GGC; (k) GGC-NH2; (l) C-концевого линкера гамма 1; (m) C-концевого линкера гамма 3; (n) C-концевых глициновых линкеров; (o) (G)nC(G)k при n=0-12 и k=0-5; (p) C-концевых глицин-сериновых линкеров; (q) (G)m(S)l(GGGGS)n(G)oC(G)k (SEQ ID NO:119) при n=0-3, k=0-5, m=0-10, l=0-2 и o=0-8.
Следующими предпочтительными примерами аминокислотных линкеров являются шарнирный участок иммуноглобулинов, глицин-сериновые линкеры (GGGGS)n (SEQ ID NO:120) и глициновые линкеры (G)n, которые все, кроме того, содержат остаток цистеина в качестве второго сайта связывания и необязательно дополнительные остатки глицина. Обычно предпочтительными примерами указанных аминокислотных линкеров являются N-концевой гамма 1: CGDKTHTSPP (SEQ ID NO:121); C-концевой гамма 1: DKTHTSPPCG (SEQ ID NO:122); N-концевой гамма 3: CGGPKPSTPPGSSGGAP (SEQ ID NO:123); C-концевой гамма 3: PKPSTPPGSSGGAPGGCG (SEQ ID NO:124); N-концевой глициновый линкер: GCGGGG (SEQ ID NO:125); C-концевой глициновый линкер: GGGGCG (SEQ ID NO:126); C-концевой глицин-лизиновый линкер: GGKKGC (SEQ ID NO:127); N-концевой глицин-лизиновый линкер: CGKKGG (SEQ ID NO:128).
В следующем предпочтительном варианте данного изобретения и, в частности, в том случае, если антигеном является пептид RANKL, предпочтительными в качестве аминокислотных линкеров являются линкеры GGCG (SEQ ID NO:162), GGC или GGC-NH2 («NH2» означает амидирование) на C-конце пептида или CGG на его N-конце. В общем, остатки глицина будут встроены между основной массой аминокислот и цистеином, используемым в качестве второго сайта связывания, чтобы избежать возможных стерических помех со стороны аминокислоты основной массы в реакции связывания.
Остаток цистеина, присутствующий на белке RANKL, фрагменте RANKL или пептиде RANKL должен быть в своем восстановленном состоянии, чтобы реагировать с гетеробифункциональным перекрестно сшивающим агентом на активированной VLP, то есть должен быть доступным свободный цистеин или остаток цистеина со свободной сульфгидрильной группой. В том случае, когда остаток цистеина для функционирования в качестве сайта связывания находится в окисленной форме, например, если он образует дисульфидный мостик, требуется восстановление указанного дисульфидного мостика, например, с помощью DTT, TCEP или β-меркаптоэтанола.
Связывание белка RANKL, фрагмента RANKL или пептида RANKL с центральной частицей или, соответственно, с VLP с использованием гетеробифункционального перекрестно сшивающего агента согласно описанным выше предпочтительным способам позволяет осуществлять связывание белка RANKL, фрагмента RANKL или пептида RANKL с центральной частицей и, соответственно, с VLP ориентированным образом. Другие способы связывания белка RANKL, фрагмента RANKL или пептида RANKL с центральной частицей и, соответственно, с VLP включают способы, при которых белок RANKL, фрагмент RANKL или пептид RANKL перекрестно сшивают с центральной частицей и, соответственно, с VLP, используя карбодиимид, EDC и NHS. Белок RANKL, фрагмент RANKL или пептид RANKL также сначала может быть тиолирован в ходе реакции, например, с SATA, SATP или иминотиоланом. Затем белок RANKL, фрагмент RANKL или пептид RANKL при необходимости после снятия защиты можно связывать с центральной частицей и, соответственно, VLP следующим образом. После отделения избытка реагента для тиолирования осуществляют взаимодействие белка RANKL, фрагмента RANKL или пептида RANKL с центральной частицей и, соответственно, с VLP, предварительно активированной гетеробифункциональным перекрестно сшивающим агентом, содержащим реакционно-способный по отношению к цистеину остаток и, следовательно, экспонирующим по меньшей мере одну или несколько функциональных групп, реагирующих с остатками цистеина, с которыми может реагировать тиолированный белок RANKL, фрагмент RANKL или пептид RANKL, такой как описано выше. Необязательно в реакционную смесь могут быть включены небольшие количества восстанавливающего агента. В следующих способах белок RANKL, фрагмент RANKL или пептид RANKL связывают с центральной частицей и, соответственно, с VLP, используя гомобифункциональный перекрестно сшивающий агент, такой как глутаральдегид, DSG, BM[PEO]4, BS3, (Pierce Chemical Company, Rockford, IL, USA) или другие известные гомобифункциональные перекрестно сшивающие агенты с функциональными группами, реагирующими с аминогруппами или карбоксильными группами центральной частицы и, соответственно, VLP.
В следующем варианте белок RANKL, фрагмент RANKL или пептид RANKL связывают с центральной частицей и, соответственно, с VLP посредством модификации углеводных остатков, присутствующих на гликозилированном белке RANKL, фрагменте RANKL или пептиде RANKL, и последующего взаимодействия с центральной частицей и, соответственно, с VLP. В одном варианте осуществляют реакцию гликозилированного белка RANKL, фрагмента RANKL или пептида RANKL с периодатом натрия в условиях реакции мягкого окисления углеводного остатка, получая активированный белок RANKL, фрагмент RANKL или пептид RANKL с одной или несколькими функциональными альдегидными группами. Активированный таким образом белок RANKL, фрагмент RANKL или пептид RANKL отделяют от избытка периодата натрия и затем подвергают взаимодействию с центральной частицей и, соответственно, с VLP, при котором остатки лизина центральной частицы и, соответственно, VLP или по меньшей мере одной субъединицы VLP взаимодействуют с ранее образованной альдегидной функциональной группой на белке RANKL, фрагменте RANKL или пептиде RANKL, например, как описано Hermanson, G.T. в Bioconjugate Techniques, Academic Press Inc., San Diego, CA, USA. Самополимеризацию активированного белка RANKL, фрагмента RANKL или пептида RANKL можно контролировать, корректируя pH, как описано в вышеуказанной публикации. Образованное основание Шиффа предпочтительно далее восстанавливают цианоборогидридом натрия, который затем удаляют гель-фильтрацией или диализом. Альтернативно можно осуществлять взаимодействие центральной частицы и, соответственно, VLP с EDC по карбоксильным группам центральной частицы и, соответственно, VLP или по меньшей мере одной субъединицы VLP и дигидразидом, таким как дигидразид адипиновой кислоты, получая остаток гидразида, доступный для реакции с одной или несколькими альдегидными функциональными группами, присутствующими на активированном белке RANKL, фрагменте RANKL или пептиде RANKL. Образованный таким образом гидразон затем можно восстанавливать цианоборогидридом натрия. Альтернативно активированный белок RANKL, фрагмент RANKL или пептид RANKL с одной или несколькими альдегидными функциональными группами подвергают реакции с цистеамином, получая в результате введение группы цистеина в белок RANKL, фрагмент RANKL или пептид RANKL. Дополнительные способы перекрестного сшивания и перекрестно сшивающие агенты, подходящие для связывания белка RANKL, фрагмента RANKL или пептида RANKL с центральной частицей и, соответственно, с VLP, а также руководство по осуществлению реакций связывания и применению химических перекрестно сшивающих агентов и способам химического перекрестного сшивания можно найти в Hermanson, G.T. in Bioconjugate Techniques, Academic Press Inc., San Diego, CA, USA.
Другие способы связывания VLP с белком RANKL, фрагментом RANKL или пептидом RANKL включают способы, при которых центральную частицу и, соответственно, VLP биотинилируют и белок RANKL, фрагмент RANKL или пептид RANKL экспрессируют в виде белка, слитого со стрептавидином, или способы, при которых биотинилируют как белок RANKL, фрагмент RANKL или пептид RANKL, так и центральную частицу и, соответственно, VLP, например, как описано в WO 00/23955. В данном случае белок RANKL, фрагмент RANKL или пептид RANKL сначала можно связать со стрептавидином или авидином, корректируя отношение белка RANKL, фрагмента RANKL или пептида RANKL к стрептавидину так, чтобы свободные сайты связывания еще были доступными для связывания центральной частицы и, соответственно VLP, которую добавляют на следующей стадии. Альтернативно все компоненты можно смешать в реакции «в одном сосуде». Другие пары лиганд-рецептор, где имеется растворимая форма рецептора и лиганда и которые способны к перекрестному сшиванию с центральной частицей и, соответственно, с VLP или белком RANKL, фрагментом RANKL или пептидом RANKL, можно использовать в качестве связывающих агентов для связывания белка RANKL, фрагмента RANKL или пептида RANKL с центральной частицей и, соответственно, с VLP. Альтернативно либо лиганд, либо рецептор могут быть слиты с белком RANKL, фрагментом RANKL или пептидом RANKL и таким образом опосредовать связывание с центральной частицей и, соответственно, с VLP, химически связанной или слитой либо с рецептором, либо с лигандом, соответственно. Слияние также можно осуществлять посредством инсерции или замены.
Как уже говорилось, в предпочтительном варианте данного изобретения VLP является VLP РНК-фага и в более предпочтительном варианте VLP является VLP покрывающего белка РНК-фага Qβ.
Одна или несколько молекул антигена, т.е. белка RANKL, фрагмента RANKL или пептида RANKL, могут быть связаны с одной субъединицей капсида или VLP покрывающих белков РНК-фага, предпочтительно через экспонированные остатки лизина VLP РНК-фагов, если это стерически допустимо. Таким образом, особым признаком VLP покрывающих белков РНК-фагов и, в частности, VLP покрывающего белка Qβ является возможность связывать несколько антигенов на субъединицу. Это обеспечивает возможность образования плотной антигенной матрицы.
В предпочтительном варианте изобретения связывание и, соответственно, присоединение по меньшей мере белка RANKL, фрагмента RANKL или пептида RANKL с центральной частицей и, соответственно, вирусоподобной частицей осуществляют посредством взаимодействия и, соответственно, ассоциации между по меньшей мере одним первым сайтом связывания вирусоподобной частицы и по меньшей мере одним вторым сайтом связывания антигена или антигенной детерминанты.
VLP или капсиды из покрывающего белка Qβ экспонируют определенное количество остатков лизина на своей поверхности с определенной топологией, при этом три остатка лизина направлены внутрь капсида и взаимодействуют с РНК, а четыре других остатка лизина экспонированы на наружную сторону капсида. Указанные определенные свойства предпочтительны для связывания антигенов с наружной стороной частицы, а не с внутренней стороны частицы, где остатки лизина взаимодействуют с РНК. VLP из других покрывающих белков РНК-фагов также имеют определенное количество остатков лизина на своей поверхности и определенную топологию указанных остатков лизина.
В следующих предпочтительных вариантах данного изобретения первый сайт связывания является остатком лизина и/или второй сайт связывания содержит сульфгидрильную группу или остаток цистеина. В очень предпочтительном варианте данного изобретения первый сайт связывания является остатком лизина, а второй сайт связывания является остатком цистеина.
В очень предпочтительных вариантах изобретения белок RANKL, фрагмент RANKL или пептид RANKL связан через остаток цистеина, либо природно присутствующий на белке RANKL, фрагменте RANKL или пептиде RANKL, либо сконструированный, с остатками лизина VLP покрывающего белка РНК-фага и, в частности, VLP покрывающего белка Qβ.
Другим преимуществом VLP, полученных из РНК-фагов, является высокий выход их экспрессии в бактериях, который позволяет получать большие количества материала при допустимой стоимости.
Как указано, конъюгаты и матрицы, соответственно, согласно изобретению отличаются от конъюгатов предшествующего уровня техники своей высокоорганизованной структурой, размерами и повторяемостью антигена на поверхности матрицы. Кроме того, применение VLP в качестве носителей позволяет образовывать прочные антигенные матрицы и конъюгаты, соответственно, с изменяемой плотностью антигенов. В частности, применение VLP РНК-фагов и, таким образом, в частности, применение VLP покрывающего белка РНК-фага Qβ позволяет достигать очень высокой плотности эпитопа. Получение композиций VLP покрывающих белков РНК-фагов с высокой плотностью эпитопов можно осуществлять с использованием инструкций, приведенных в данной заявке.
На антигене или антигенной детерминанте может присутствовать либо природный, либо неприродный второй сайт связывания, который определен в данном описании. В случае отсутствия подходящего второго сайта связывания природного происхождения на антигене или антигенной детерминанте необходимо сконструировать неприродный второй сайт связывания на антигене.
Как описано выше, четыре остатка лизина экспонированы на поверхности VLP покрывающего белка Qβ. Обычно такие остатки дериватизируют в ходе реакции с перекрестно сшивающей молекулой. В том случае, когда не все экспонированные остатки лизина могут быть связаны с антигеном, остатки лизина, которые прореагировали с перекрестно сшивающим агентом, остаются с перекрестно сшивающей молекулой, связанной с ε-аминогруппой после стадии дериватизации. Это приводит к исчезновению одного или нескольких положительных зарядов, которое может быть нежелательным для растворимости и стабильности VLP. Заменяя некоторые остатки лизина аргининами, как в заявленных мутантах покрывающего белка Qβ, описанных ниже, авторы предотвратили чрезмерное исчезновение положительных зарядов, так как остатки аргинина не взаимодействуют с перекрестно сшивающим агентом. Кроме того, замена остатков лизина аргининами может приводить к более определенной антигенной матрице, так как меньше сайтов доступно для реакции с антигеном.
Таким образом, экспонированные остатки лизина заменяли аргининами в следующих мутантах покрывающего белка Qβ и мутантных VLP Qβ, описанных в данной заявке: Qβ-240 (Lys13-Arg; SEQ ID NO:23), Qβ-250 (Lys 2-Arg, Lys13-Arg; SEQ ID NO:25) и Qβ-259 (Lys 2-Arg, Lys16-Arg; SEQ ID NO:27). Конструкции клонировали, белки экспрессировали, VLP очищали и использовали для связывания с пептидными и белковыми антигенами. Также конструировали Qβ-251 (SEQ ID NO:26), и руководство по поводу того, как экспрессировать, очищать и связывать VLP покрывающего белка Qβ-251, можно найти на протяжении данной заявки.
В следующем варианте авторы предложили мутантный покрывающий белок Qβ с одним дополнительным остатком лизина, подходящий для получения матрицы антигенов с еще более высокой плотностью. Данный мутантный покрывающий белок Qβ, Qβ-243 (Asn 10-Lys; SEQ ID NO:24) клонировали, белок экспрессировали и капсид или VLP выделяли и очищали, показав, что введение дополнительного остатка лизина совместимо с самосборкой субъединиц в капсид или VLP. Таким образом, матрицы и, соответственно, конъюгаты белка RANKL, фрагмента RANKL или пептида RANKL можно получить с использованием VLP мутантов покрывающего белка Qβ. Особенно предпочтительным способом связывания антигенов с VLP и, в частности, с VLP покрывающих белков РНК-фага является связывание остатка лизина, присутствующего на поверхности VLP покрывающих белков РНК-фага, с остатком цистеина, природно присутствующим или сконструированным на антигене, т.е. белке RANKL, фрагменте RANKL или пептиде RANKL. Для того чтобы остаток цистеина был эффективным в качестве второго сайта связывания, сульфгидрильная группа должна быть доступной для связывания. Таким образом, остаток цистеина должен быть в своем восстановленном состоянии, то есть должен быть доступным свободный цистеин или остаток цистеина со свободной сульфгидрильной группой. В том случае, когда остаток цистеина для функционирования в качестве второго сайта связывания находится в окисленной форме, например, если он образует дисульфидный мостик, требуется восстановление указанного дисульфидного мостика, например, с помощью DTT, TCEP или β-меркаптоэтанола. Концентрация восстановителя и молярный избыток восстановителя по сравнению с антигеном должен уточняться для каждого антигена. При необходимости тестируют пределы титрования, начиная с таких низких концентраций восстановителя, как 10 мкМ или ниже до 10-20 мМ или выше, и оценивают связывание антигена с носителем. Несмотря на то что низкие концентрации восстановителя совместимы с реакцией связывания, как описано в находящейся на рассмотрении заявке на выдачу патента США № 10/050902, поданной настоящим правопреемником 18 января 2002, более высокие концентрации ингибируют реакцию связывания, как будет понятно специалисту в данной области, и в этом случае восстановитель необходимо удалить диализом или гель-фильтрацией. Преимущественно pH буфера для диализа или уравновешивающего буфера ниже 7, предпочтительно 6. Совместимость буфера с низким значением pH с активностью и стабильностью антигена необходимо проверять.
Плотность эпитопа на VLP покрывающих белков РНК-фага можно модулировать, выбирая перекрестно сшивающий агент и другие условия реакции. Например, перекрестно сшивающие агенты сульфо-GMBS и SMPH обычно обеспечивают достижение высокой плотности эпитопа. На дериватизацию положительно влияет высокая концентрация реагирующих веществ, и манипулирование условиями реакции можно использовать для того, чтобы контролировать количество антигенов, связанных с VLP покрывающих белков РНК-фага и, в частности, с VLP покрывающего белка Qβ.
Перед конструированием неприродного второго сайта связывания необходимо выбрать положение, в котором он должен быть слит, встроен или, в общем, сконструирован. Выбор положения второго сайта связывания в качестве примера может быть основан на кристаллической структуре антигена. Такая кристаллическая структура антигена может давать информацию о доступности C- или N-концов молекулы (определенной, например, исходя из их доступности для растворителя) или об экспонировании по отношению к растворителю остатков, подходящих для использования в качестве вторых сайтов связывания, таких как остатки цистеина. Экспонированные дисульфидные мостики, как в случае Fab-фрагментов, также могут быть источником второго сайта связывания, так как их, как правило, можно превратить в отдельные остатки цистеина посредством умеренного восстановления. Будут выбраны умеренные условия восстановления, не влияющие на иммуногенность белка RANKL, фрагмента RANKL или пептида RANKL. В общем, в том случае, когда целью иммунизации аутоантигеном является ингибирование взаимодействия данного аутоантигена с его природными лигандами, второй сайт связывания будет добавлен таким образом, чтобы он давал возможность образовывать антитела против сайта взаимодействия с природными лигандами. Таким образом, положение второго сайта связывания будет выбрано так, чтобы избежать стерических помех со стороны второго сайта связывания или любого содержащего его аминокислотного линкера. В следующих вариантах требуется гуморальный ответ, направленный на сайт, отличный от сайта взаимодействия аутоантигена с его природным лигандом. В таких вариантах второй сайт связывания может быть выбран так, чтобы он предотвращал образование антител против сайта взаимодействия аутоантигена с его природными лигандами.
Другие критерии для выбора положения второго сайта связывания включают состояние олигомеризации антигена, сайт олигомеризации, присутствие кофактора и наличие экспериментального доказательства обнаружения сайтов в структуре антигена и последовательности, в которой модификация антигена совместима с функцией аутоантигена или с образованием антител, распознающих аутоантиген.
В наиболее предпочтительных вариантах белок RANKL, фрагмент RANKL или пептид RANKL содержит единственный второй сайт связывания или единственный реакционно-способный сайт связывания, способный к ассоциации с первыми сайтами связывания на центральной частице и VLP или субъединицах VLP, соответственно. Это обеспечивает определенное и равномерное связывание и, соответственно, ассоциацию по меньшей мере одного, но обычно нескольких, предпочтительно более 10, 20, 40, 80, 120, антигенов с центральной частицей и VLP, соответственно. Таким образом, обеспечение одного второго сайта связывания или одного реакционно-способного сайта связывания на антигене обеспечивает один тип равномерного связывания и ассоциации, соответственно, приводящий к очень высокоупорядоченной и повторяющейся матрице. Например, если связывание и ассоциацию, соответственно, осуществляют посредством взаимодействия лизина (в качестве первого сайта связывания) и цистеина (в качестве второго сайта связывания), то согласно данному предпочтительному варианту изобретения гарантировано, что только один остаток цистеина на антиген, независимо от того, присутствует ли на антигене данный остаток цистеина природного или неприродного происхождения, способен связываться и ассоциировать, соответственно, с VLP и первым сайтом связывания центральной частицы, соответственно.
В некоторых вариантах конструирование второго сайта связывания на антигене требует слияния аминокислотного линкера, содержащего аминокислоту, подходящую в качестве второго сайта связывания согласно описаниям данного изобретения. Таким образом, в предпочтительном варианте данного изобретения аминокислотный линкер связан с антигеном или антигенной детерминантой посредством по меньшей мере одной ковалентной связи. Предпочтительно аминокислотный линкер содержит или, альтернативно, состоит из второго сайта связывания. В следующем предпочтительном варианте аминокислотный линкер содержит сульфгидрильную группу или остаток цистеина. В другом предпочтительном варианте аминокислотным линкером является цистеин. Некоторые критерии выбора аминокислотного линкера, а также дополнительные предпочтительные варианты аминокислотного линкера согласно изобретению уже указаны выше.
В следующем предпочтительном варианте изобретения по меньшей мере один антиген или антигенная детерминанта, т.е. белок RANKL, фрагмент RANKL или пептид RANKL, слит с центральной частицей и вирусоподобной частицей соответственно. Как указано выше, VLP обычно состоит по меньшей мере из одной субъединицы, собирающейся в VLP. Таким образом, снова в следующем предпочтительном варианте изобретения антиген или антигенная детерминанта, предпочтительно по меньшей мере один белок RANKL, фрагмент RANKL или пептид RANKL слит по меньшей мере с одной субъединицей вирусоподобной частицы или белка, который может быть включен в VLP, с образованием химерного слияния VLP-субъединица-белок RANKL, фрагмент RANKL или пептид RANKL.
Слияние белка RANKL, фрагмента RANKL или пептида RANKL можно осуществить путем инсерции в последовательность субъединицы VLP или путем слияния либо с N-, либо с C-концом VLP-субъединицы или белка, который может быть включен в VLP. В дальнейшем при упоминании слитых белков пептида с субъединицей VLP подразумевают слияние либо с концами последовательности субъединицы либо внутреннюю инсерцию пептида в последовательность субъединицы.
Слияние также можно осуществить путем инсерции последовательностей белка RANKL, фрагмента RANKL или пептида RANKL в вариант субъединицы VLP, где часть последовательности субъединицы была делетирована, полученные продукты далее называют укороченными мутантами. Укороченные мутанты могут иметь N- или C-концевые или внутренние делеции части последовательности субъединицы VLP. Например, конкретный HBcAg VLP, например, с делецией аминокислотных остатков с 79 по 81 является укороченным мутантом с внутренней делецией. Слияние белка RANKL, фрагмента RANKL или пептида RANKL либо с N-, либо с C-концом укороченных мутантов VLP-субъединиц также приводит к образованию вариантов согласно данному изобретению. Подобным образом, слияние эпитопа с последовательностью субъединицы VLP также можно осуществить путем замены, при которой, например, для конкретного HBcAg VLP аминокислоты 79-81 заменяют чужеродным эпитопом. Таким образом, слияние, которое упоминается в дальнейшем, можно осуществить путем инсерции последовательности белка RANKL, фрагмента RANKL или пептида RANKL в последовательность субъединицы VLP путем замены части последовательности субъединицы VLP последовательностью белка RANKL, фрагмента RANKL или пептида RANKL или посредством комбинирования делеции, замены или инсерции.
Химерные конструкции белок RANKL, фрагмент RANKL или пептид RANKL-субъединица VLP, как правило, будут способны к самосборке в VLP. VLP, экспонирующие эпитопы, слитые с их субъединицами, также называют в данном описании химерными VLP. Как указано, вирусоподобная частица содержит или, альтернативно, состоит по меньшей мере из одной субъединицы VLP. В следующем варианте изобретения вирусоподобная частица содержит или, альтернативно, состоит из смеси химерных субъединиц VLP и нехимерных субъединиц VLP, т.е. субъединиц VLP, не имеющих слитого с ними антигена, образуя в результате так называемые мозаичные частицы. Это может иметь преимущества для обеспечения образования и сборки в VLP. В указанных вариантах доля химерных VLP-субъединиц может составлять 1, 2, 5, 10, 20, 30, 40, 50, 60, 70, 80, 90, 95% или больше.
Могут быть добавлены фланкирующие аминокислотные остатки к любому концу последовательности пептида или эпитопа, который необходимо слить с любым из концов последовательности субъединицы VLP, или для внутренней инсерции такой пептидной последовательности в последовательность субъединицы VLP. Остатки глицина и серина являются особенно предпочтительными аминокислотами для использования во фланкирующих последовательностях, добавляемых к белку RANKL, фрагменту RANKL или пептиду RANKL, который необходимо слить. Остатки глицина придают дополнительную гибкость, которая может снижать возможное дестабилизирующее действие слияния чужеродной последовательности в последовательность субъединицы VLP.
В конкретном варианте изобретения VLP является VLP корового антигена гепатита B. Описаны слитые белки либо с N-концом HBcAg (Neyrinck, S. et al., Nature Med. 5: 1157-1163 (1999)), либо инсерции в так называемую основную иммунодоминантную область (MIR) (Pumpens, P. and Grens, E., Intervirology 44:98-114 (2001)), WO 01/98333), и указанные слияния являются предпочтительными вариантами изобретения. Также описаны варианты HBcAg природного происхождения с делециями в MIR (Pumpens, P. and Grens, E., Intervirology 44: 98-114 (2001), публикация специально включена в виде ссылки в полном объеме), и слияния с N- или C-концом, а также инсерции в положение MIR, соответствующее сайту делеции по сравнению с HBcAg дикого типа, являются следующими вариантами изобретения. Также описаны слияния с C-концом (Pumpens, P. and Grens, E., Intervirology 44: 98-114 (2001)). Специалист в данной области легко найдет руководство по поводу того, как конструировать слитые белки, используя классические способы молекулярной биологии (Sambrook, J. et al., eds., Molecular Cloning, A Laboratory Manual, 2nd. edition, Cold Spring Habor Laboratory Press, Cold Spring Harbor, N.Y. (1989), Ho et al., Gene 77:51 (1989)). Описаны векторы и плазмиды, кодирующие HBcAg и слитые белки HBcAg и применимые для экспрессии HBcAg и слитых белков HBcAg (Pumpens, P. and Grens, E. Intervirology 44: 98-114 (2001), Neyrinck, S. et al., Nature Med. 5: 1157-1163 (1999)), которые можно использовать в практике изобретения. Авторы также описали в виде примера (пример 6) инсерцию эпитопа в MIR HBcAg с получением в результате химерного подвергаемого самосборке HBcAg. Важным фактором для оптимизации эффективности самосборки и экспонирования эпитопа, встраиваемого в MIR HBcAg, является выбор места инсерции, а также количество аминокислот, которые необходимо делетировать из последовательности HBcAg в MIR (Pumpens, P. and Grens, E., Intervirology 44: 98-114 (2001); EP 421635; патент США 6231864) при инсерции, или другими словами, какие аминокислоты из HBcAg необходимо заменить новым эпитопом. Например, описана замена аминокислот HBcAg 76-80, 79-81, 79-80, 75-85 или 80-81 чужеродными эпитопами (Pumpens, P. and Grens, E., Intervirology 44: 98-114 (2001); EP 0421635; патент США 6231864). HBcAg содержит длинный аргининовый хвост (Pumpens, P. and Grens, E., Intervirology 44: 98-114 (2001)), который не имеет существенного значения для сборки капсида и способен к связыванию нуклеиновых кислот (Pumpens, P. and Grens, E., Intervirology 44: 98-114 (2001)). HBcAg, либо содержащие аргининовый хвост, либо не имеющие такового, являются вариантами изобретения.
В следующем предпочтительном варианте изобретения VLP является VLP РНК-фага. Основные покрывающие белки РНК-фагов спонтанно собираются в VLP при экспрессии в бактериях и, а частности, в E. coli. Конкретные примеры покрывающих белков бактериофагов, которые можно использовать для получения композиций согласно изобретению, включают покрывающие белки таких РНК-бактериофагов, как бактериофаг Qβ (SEQ ID NO:10; база данных PIR, инвентарный No. VCBPQβ, относящаяся к CP Qβ, и SEQ ID NO:11; инвентарный No. AAA16663, относящаяся к белку A1 Qβ) и бактериофаг fr (SEQ ID NO:4; PIR, инвентарный No. VCBPFR).
В более предпочтительном варианте по меньшей мере один белок RANKL, фрагмент RANKL или пептид RANKL сливают с покрывающим белком Qβ. Описаны конструкции слитых белков, в которых эпитопы были слиты с C-концом укороченной формы белка A1 Qβ или встроены в белок A1 (Kozlovska, T.M., et al., Intervirology, 39: 9-15 (1996)). Белок A1 образуется при супрессии в стоп-кодоне UGA и имеет длину 329 а/к или 328 а/к, если принять во внимание отщепление N-концевого метионина. Отщепление N-концевого метионина перед аланином (вторая аминокислота, кодируемая геном CP Qβ) обычно имеет место в E. coli, и так обстоит дело для N-концов покрывающих белков CP Qβ. Часть гена A1 с 3'-стороны от амбер-кодона UGA кодирует удлинение CP, которое имеет длину 195 аминокислот. Инсерция по меньшей мере одного белка RANKL, фрагмента RANKL или пептида RANKL между положениями 72 и 73 удлинения CP приводит к образованию следующих вариантов согласно изобретению (Kozlovska, T.M., et al., Intervirology 39: 9-15 (1996)). Слияние белка RANKL, фрагмента RANKL или пептида RANKL с C-концом укороченного на C-конце белка A1 Qβ приводит к образованию следующих предпочтительных вариантов согласно изобретению. Например, Kozlovska et al., (Intervirology, 39: 9-15 (1996)) описали слияния белка A1 Qβ, в которых эпитоп слит с C-концом удлинения CP Qβ, укороченным в положении 19.
Как описано Kozlovska et al. (Intervirology, 39: 9-15 (1996)), сборка частиц, экспонирующих слитые эпитопы, обычно требует присутствия как слияния белок A1-белок RANKL, фрагмент RANKL или пептид RANKL, так и CP дикого типа, чтобы образовать мозаичную частицу. Однако варианты, содержащие вирусоподобные частицы, и, таким образом, в частности, VLP покрывающего белка РНК-фага Qβ, которые состоят исключительно из субъединиц VLP, имеющих по меньшей мере один слитый с ними белок RANKL, фрагмент RANKL или пептид RANKL, также входят в объем данного изобретения.
Получение мозаичных частиц можно осуществить рядом способов. Kozlovska et al., Intervirolog., 39: 9-15 (1996) описывают два способа, которые оба можно использовать в практике изобретения. В первом способе эффективное экспонирование слитого эпитопа на VLP опосредовано экспрессией плазмиды, кодирующей слияние белка A1 Qβ, имеющего стоп-кодон UGA между CP и удлинением CP, в штамме E. coli, несущем плазмиду, кодирующую клонированную тРНК-супрессор UGA, который приводит к трансляции кодона UGA в Trp (плазмида pISM3001 (Smiley B.K., et al., Gene 134: 33-40 (1993))). В другом способе стоп-кодон гена CP модифицируют в UAA и проводят котрансфекцию со второй плазмидой, экспрессирующей слияние белка A1 с белком RANKL, фрагментом RANKL или пептидом RANKL. Вторая плазмида кодирует резистентность к различным антибиотикам, и начало репликации совместимо с первой плазмидой (Kozlovska, T.M., et al., Intervirology 39: 9-15 (1996)). В третьем способе CP и слияние белка A1 с белком RANKL, фрагментом RANKL или пептидом RANKL кодируются бицистронным образом при оперативном связывании с таким промотором, как промотор Trp, как описано на фиг.1 в Kozlovska et al., Intervirology, 39: 9-15 (1996).
В следующем варианте белок RANKL, фрагмент RANKL или пептид RANKL встраивают между аминокислотами 2 и 3 (нумерация CP после расщепления, то есть в том случае, когда N-концевой метионин отщеплен) CP fr, таким образом получая белковое слияние белок RANKL, фрагмент RANKL или пептид RANKL-CP fr. Описаны векторы и экспрессирующие системы для конструирования и экспрессии слитых белков CP fr, самособирающихся в VLP и применимых в практике изобретения (Pushko P. et al., Prot. Eng. 6:883-891 (1993)). В конкретном варианте последовательность белка RANKL, фрагмента RANKL или пептида RANKL встраивают в делеционный вариант CP fr после аминокислоты 2, когда остатки 3 и 4 CP fr удалены (Pushko P. et al., Prot. Eng. 6:883-891 (1993)).
Слияние эпитопов с N-концевой выступающей β-шпилькой покрывающего белка РНК-фага MS-2 и последующая презентация слитого эпитопа на образованной при самосборке VLP РНК-фага MS-2 также описаны (WO 92/13081), и слияние белка RANKL, фрагмента RANKL или пептида RANKL путем инсерции или замены в покрывающем белке РНК-фага MS-2 также попадает в объем изобретения.
В другом варианте изобретения белок RANKL, фрагмент RANKL или пептид RANKL сливают с белком капсида вируса папилломы. В более конкретном варианте белок RANKL, фрагмент RANKL или пептид RANKL сливают с основным белком капсида L1 вируса папилломы крупного рогатого скота типа 1 (BPV-1). Описаны векторы и экспрессирующие системы для конструирования и экспрессии слитых белков BPV-1 в системах бакуловирус/клетки насекомых (Chackerian, B. et al., Proc. Natl. Acad. Sci. USA 96: 2373-2378 (1999), WO 00/23955). Замена аминокислот 130-136 L1 BPV-1 белком RANKL, фрагментом RANKL или пептидом RANKL приводит к слитому белку L1 BPV-1-белок RANKL, фрагмент RANKL или пептид RANKL, который является предпочтительным вариантом изобретения. Описано клонирование в бакуловирусном векторе и экспрессия в инфицированных бакуловирусом клетках Sf9, которые могут быть использованы в практике изобретения (Chackerian, B. et al., Proc. Natl. Acad. Sci. USA 96: 2373-2378 (1999), WO 00/23955). Очистку собранных частиц, экспонирующих слитый белок RANKL, фрагмент RANKL или пептид RANKL, можно осуществить рядом способов, таких, например, как гель-фильтрация или ультрацентрифугирование в градиенте сахарозы (Chackerian, B. et al., Proc. Natl. Acad. Sci. USA 96: 2373-2378 (1999), WO 00/23955).
В следующем варианте изобретения белок RANKL, фрагмент RANKL или пептид RANKL сливают с белком Ty, способным включаться в VLP Ty. В более конкретном варианте белок RANKL, фрагмент RANKL или пептид RANKL сливают с p1 или белком капсида, кодируемым геном TYA gene (Roth, J.F., Yeast 16: 785-795 (2000)). Дрожжевые ретротранспозоны Ty1, 2, 3 и 4 выделены из Saccharomyces cerevisiae, тогда как ретротранспозон Tf1 выделен из Schizosaccharomyces pombae (Boeke, J.D. and Sandmeyer, S.B., «Yeast Transposable elements,» in The molecular and Cellular Biology of the Yeast Saccharomyces: Genome dynamics, Protein Synthesis, and Energetics., p. 193, Cold Spring Harbor Laboratory Press (1991)). Ретротранспозоны Ty1 и 2 родственны классу copia элементов растений и животных, тогда как Ty3 относится к семейству ретротранспозонов gypsy, которые родственны ретровирусам растений и животных. В ретротранспозоне Ty1 белок p1, также называемый Gag или капсидным белком, имеет длину 440 аминокислот. P1 расщепляется во время созревания VLP в положении 408, давая белок p2, важный компонент VLP.
Описаны слитые с p1 белки и векторы для экспрессии указанных слитых белков в дрожжах (Adams, S.E., et al., Nature 329: 68-70 (1987)). Таким образом, например, белок RANKL, фрагмент RANKL или пептид RANKL может быть слит с p1 путем инсерции последовательности, кодирующей белок RANKL, фрагмент RANKL или пептид RANKL, в сайт BamH1 плазмиды pMA5620 (Adams, S.E., et al., Nature 329: 68-70 (1987)). Клонирование последовательностей, кодирующих чужеродные эпитопы, в векторе pMA5620 приводит к экспрессии слитых белков, содержащих аминокислоты 1-381 p1 Ty1-15, слитые C-терминально с N-концом чужеродного эпитопа. Подобным образом подразумевается, что N-концевое слияние белка RANKL, фрагмента RANKL или пептида RANKL или внутренняя инсерция в последовательность p1 или замена части последовательности p1 также входит в объем изобретения. В частности, инсерция белка RANKL, фрагмента RANKL или пептида RANKL в последовательность Ty между аминокислотами 30-31, 67-68, 113-114 и 132-133 p1-белка Ty (EP0677111) дает предпочтительные варианты согласно изобретению.
Следующими VLP, подходящими для слияния белка RANKL, фрагмента RANKL или пептида RANKL, являются, например, частицы, подобные ретровирусам (WO9630523), Gag HIV2 (Kang, Y.C., et al, Biol. Chem. 380: 353-364 (1999)), вирус мозаики вигны (Taylor, K.M. et al., Biol. Chem. 380: 387-392 (1999)), VLP парвовируса VP2 (Rueda, P. et al., Virology 263: 89-99 (1999)), HBsAg (патент США 4722840, EP0020416B1).
Примерами химерных VLP, подходящими для практики изобретения, также являются VLP, описанные в Intervirology 39: 1 (1996). Следующими примерами VLP, предполагаемыми для применения в изобретении, являются: HPV-1, HPV-6, HPV-11, HPV-16, HPV-18, HPV-33, HPV-45, CRPV, COPV, GAG HIV, вирус табачной мозаики. Также получены вирусоподобные частицы SV-40, вируса полиомы, аденовируса, вируса простого герпеса, ротавируса и вируса Норуолк, и химерные VLP на основе указанных VLP также входят в объем данного изобретения.
В следующем предпочтительном варианте данного изобретения антигеном или антигенной детерминантой является белок RANKL, фрагмент RANKL или пептид RANKL.
В следующем очень предпочтительном варианте изобретения антигеном или антигенной детерминантой является белок RANKL, фрагмент RANKL или пептид RANKL человека.
В следующем очень предпочтительном варианте изобретения антиген или антигенная детерминанта содержит, альтернативно, по существу состоит или, альтернативно, состоит из аминокислотной последовательности, выбранной из группы, состоящей из a) аминокислотной последовательности SEQ ID NO:79; b) аминокислотной последовательности любого фрагмента SEQ ID NO:79.
В следующем предпочтительном варианте изобретения антигеном или антигенной детерминантой является вариант белка RANKL, фрагмента RANKL или пептида RANKL, например, в частности, содержащий аминокислотные замены или инсерции пептидов или полиморфизмы. Как уже указано, композиции и, соответственно, композиции вакцин, содержащие варианты белка RANKL, фрагмента RANKL или пептида RANKL, включены в объем данного изобретения.
В следующем предпочтительном варианте изобретения антигеном или антигенной детерминантой является фрагмент RANKL. Предпочтительно антигеном или антигенной детерминантой является фрагмент RANKL человека, выбранный из группы, состоящей из a) внеклеточной области RANKL человека, b) изоформы 1 сплайсинга RANKL человека, c) изоформы 2 сплайсинга, результатом которого является секретируемый RANKL человека, d) протеолитически получаемого растворимого участка RANKL человека и e) области гомологии с TNF-α.
В следующем очень предпочтительном варианте изобретения антиген или антигенная детерминанта содержит, альтернативно, по существу состоит или, альтернативно, состоит из аминокислотной последовательности, выбранной из группы, состоящей из a) аминокислотной последовательности SEQ ID NO:79; b) аминокислотной последовательности SEQ ID NO:80; c) аминокислотной последовательности SEQ ID NO:81; d) аминокислотной последовательности SEQ ID NO:82; e) аминокислотной последовательности SEQ ID NO:83; f) аминокислотной последовательности SEQ ID NO:84; g) аминокислотной последовательности SEQ ID NO:100; h) аминокислотной последовательности SEQ ID NO:101; i) аминокислотной последовательности любого фрагмента любой из SEQ ID NO:79-84, 100, 101.
Белок RANKL или фрагменты RANKL можно получить путем экспрессии кДНК RANKL в прокариотических или эукариотических системах экспрессии. Различные их примеры описаны в литературе и могут быть использованы, возможно, после модификаций, чтобы экспрессировать любой белок RANKL, фрагмент RANKL или пептид RANKL любого требуемого вида. Белок RANKL и фрагменты RANKL были экспрессированы в клетках млекопитающих (Anderson, D. M., et al., Nature 390: 175-179 (1997), Lacey, D.L., et al., Cell 93: 165-176 (1998), Wong B.R., et al., J. Biol. Chem. 272: 25190-25194 (1997), Lum, L., et al., J. Biol. Chem. 274: 13613-13618 (2000)), в клетках насекомых (Willard, D., et al., Prot. Express. Purif. 20: 48-57 (2000)) и в прокариотических клетках (Xu, J., et al., J. Bone Mineral Res. 15: 2178-86 (2000), Yasuda et al., Proc. Natl. Acad. Sci USA 95: 3597-3602 (1998)). Описания того, как получить белки и фрагменты RANKL, также приведены в WO 9846751, патенте США 5843678, WO 98259958, патенте США 6242586, WO 9828426, патенте США 6242213, WO 9929865, патенте Японии 2000102390 и WO 0015807.
В следующем предпочтительном варианте изобретения антигеном или антигенной детерминантой является пептид RANKL. Такие пептиды RANKL или их фрагменты можно получить с использованием стандартных методов молекулярной биологии, при которых нуклеотидную последовательность, кодирующую представляющий интерес фрагмент, амплифицируют с помощью ПЦР и клонируют в виде слияния с полипептидной меткой, такой как гистидиновая метка, Flag-метка, myc-метка или константная область антитела (Fc-область). При введении сайта расщепления протеазой между фрагментом RANKL и меткой фрагмент RANKL можно отделить от метки после очистки, расщепляя соответствующей протеазой. В другом способе фрагмент RANKL можно синтезировать in vitro, используя стандартные реакции пептидного синтеза, известные специалисту в данной области. В следующем способе пептиды RANKL или фрагменты RANKL можно получить расщеплением протеазой или химическим расщеплением полноразмерного белка RANKL или фрагментов RANKL, оба способа хорошо известны специалистам в данной области.
В еще одном предпочтительном варианте данного изобретения антиген или антигенная детерминанта, кроме того, содержит по меньшей мере один второй сайт связывания, выбранный из группы, состоящей из (i) сайта связывания неприродного происхождения с указанным антигеном или антигенной детерминантой; и (ii) сайта связывания природного происхождения с указанным антигеном или антигенной детерминантой. Руководство по поводу того, как модифицировать белок RANKL, фрагмент RANKL или пептид RANKL для связывания с вирусоподобной частицей, дано на протяжении данной заявки. Предпочтительные вторые сайты связывания содержат остаток цистеина для связывания с дериватизованной VLP, и примеры приведены в описании выше и в примерах 12 и 13.
Авторы построили модель 3-мерной структуры области RANKL человека, которая гомологична TNF-α. Авторы обнаружили, что цистеин природного происхождения может быть недоступным в подвергнутой укладке структуре для взаимодействия с первый сайтом связывания на VLP согласно данному изобретению. Модель авторов подтверждена рентгеновской структурой RANKL мыши, которая была недавно получена (Lam, J., et al., J. Clin. Invest., 108: 971-979 (2002)). N-конец является предпочтительным для присоединения второго сайта связывания, содержащего аминокислотный линкер с дополнительным остатком цистеина, как показано в примере 12. Однако аминокислотный линкер, содержащий остаток цистеина в качестве второго сайта связывания и слитый с C-концом конструкции RANKL, дает в результате следующий предпочтительный вариант согласно изобретению, который показан в примере 13. Конструкция RANKL человека с N-концевым аминокислотным линкером, содержащим остаток цистеина, слитым с внеклеточной областью RANKL, является очень предпочтительным вариантом изобретения.
Описаны конструкции фрагментов RANKL мыши (SEQ ID NO:96 и SEQ ID NO:99), и также могут быть созданы предпочтительные конструкции фрагментов RANKL человека и иметь, например, последовательность SEQ ID NO:100-104. Следующие предпочтительные конструкции содержат полный белок RANKL человека, фрагмент RANKL человека, выбранный из группы, состоящей из a) внеклеточной области RANKL (SEQ ID NO:82), b) изоформы 1 сплайсинга RANKL (SEQ ID NO:80), c) изоформы 2 сплайсинга, результатом которого является секретируемый RANKL (SEQ ID NO:81), d) протеолитически получаемого растворимого участка RANKL (SEQ ID NO:83) и e) области гомологии с TNF-α (SEQ ID NO:84) или последовательностей пептидов RANKL человека. Иммунизация против белка RANKL, фрагмента RANKL или пептида RANKL с использованием композиций согласно изобретению, содержащих предпочтительно белок RANKL, фрагмент RANKL или пептид RANKL человека, связанный с VLP, может дать способ лечения или профилактики заболеваний кости.
В следующем предпочтительном варианте данного изобретения белок RANKL, фрагмент RANKL или пептид RANKL содержит по меньшей мере один антигенный сайт белка RANKL. Специалисту в данной области известно, как идентифицировать соответствующие пептиды и аминокислотные последовательности, соответственно.
В следующем предпочтительном варианте данного изобретения антигеном или антигенной детерминантой является пептид RANKL, который является ключевым для взаимодействия с рецептором RANK. Моделирование авторов структуры RANKL человека и опубликованная кристаллическая структура RANKL мыши показали, что мономер RANKL состоит из β-сандвича, состоящего из двух плоских антипараллельных β-слоев. Первый слой образован β-нитями A", A, H, C и F, тогда как второй слой образован β-нитями B', B, G, D и E. Внутренний β-слой A"AHCF вовлечен в ассоциацию между субъединицами, тогда как β-слой B'BGDE в значительной степени вносит вклад в наружную поверхность. β-нити связаны через петлю AA", петлю CD, петлю DE, петлю EF. Сборка гомотримера осуществляется так, что один край β-сэндвича в каждом мономере RANKL укладывается против внутренней гидрофобной поверхности β-слоя AHCF соседнего мономера. Предполагается, что сайт связывания RANK включает в себя карман, образованный соседними мономерами гомотримера. На основании гомологии между последовательностью мыши и человека выбраны пептиды, которые включают в себя сайт связывания RANK. В предпочтительном варианте пептиды из сайта взаимодействия RANKL с RANK выбраны из группы, состоящей из a) петли AA", включающей в себя аминокислоты 171-194 (SEQ ID NO:87), b) петли DE, включающей в себя аминокислоты 246-253 (SEQ ID NO:88), c) β-нити D, включающей в себя аминокислоты 235-245 (SEQ ID NO:89), d) петли CD, включающей в себя аминокислоты 223-234 (SEQ ID NO:90), e) петли EF, включающей в себя 262-272 (SEQ ID NO:91), f) β-нити B'-петли B'B, включающих в себя аминокислоты 200-207 (SEQ ID NO:92), g) петли GH, включающей в себя аминокислоты 300-305 (SEQ ID NO:93), h) любого фрагмента указанных пептидов a-g, i) любых N- и/или C-концевых удлинений указанных пептидов a-g по меньшей мере на одну аминокислоту и до 25 аминокислот, j) любого слияния пептидов a-i). Следующие пептиды RANKL, подходящие для применения в данном изобретении, можно определить экспериментально по свойственной им особенности индуцировать T-клеточный или гуморальный ответ. В основном это осуществляется иммунизацией экспериментального животного отдельно выбранными пептидами в иммунологически подходящей композиции и измерением T-клеточного и B-клеточного, т.е. гуморального ответа с использованием способов, известных специалисту в данной области. В том случае, когда антиген является белком, полипептидом или пептидом, указанная область может быть образована непрерывной аминокислотной последовательностью. Альтернативно эпитоп антитела может быть образован прерывистой аминокислотной последовательностью, в которой после трехмерной укладки белка, полипептида или пептида аминокислоты располагаются таким образом, что они пространственно становятся ближе друг к другу и образуют эпитоп. Представляющие интерес непрерывные пептидные фрагменты можно идентифицировать с помощью экспериментов по иммунизации, которые описаны выше.
Следующие предпочтительные пептиды RANKL, подходящие для применения в данном изобретении, можно идентифицировать с использованием существующих или будущих моноклональных или поликлональных антител, способы для осуществления этого известны специалистам в данной области.
Следующие пептиды RANKL, подходящие для применения в данном изобретении, можно идентифицировать посредством скрининга пептидных библиотек в фаговом дисплее с помощью антител, специфичных по отношению к RANKL, способом, хорошо известным специалисту в данной области.
В следующем предпочтительном варианте изобретения антигеном или антигенной детерминантой является изолированный RANKL любого животного, а также любые антигенные фрагменты RANKL любого животного. Специалистам в данной области известно, как получить фрагменты и пептиды из указанного выделенного белка или фрагментов RANKL.
Специалисту в областях, имеющих отношение к данной проблеме, будет понятно, что легко предположить другие подходящие модификации и адаптации способов и применений, описанных в данной заявке, и их можно осуществить, не выходя за пределы объема изобретения и его любого варианта. Теперь, после подробного описания данного изобретения, то же самое будет более четко понятным при обращении к следующим примерам, которые, таким образом, включены только в целях иллюстрации и не предназначены для ограничения изобретения.
ПРИМЕР 1
Конструирование и экспрессия мутантных покрывающих белков Qβ и очистка VLP или капсидов из мутантных покрывающих белков Qβ
Конструирование плазмиды и клонирование мутантных покрывающих белков
Конструирование pQβ-240:
Плазмиду pQβ10 (Kozlovska, TM, et al., Gene 137: 133-137) использовали в качестве исходной плазмиды для конструирования pQβ-240. Мутацию Lys13→Arg создавали обратной ПЦР. Обратные праймеры конструировали в обратных направлениях «хвост к хвосту»:
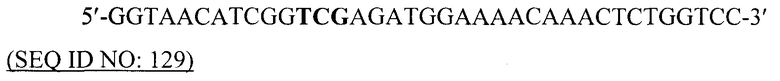
и

Продукты первой ПЦР использовали в качестве матриц для второй реакции ПЦР, в которой использовали верхний праймер
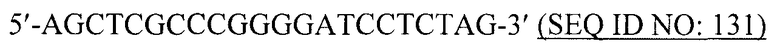
и нижний праймер
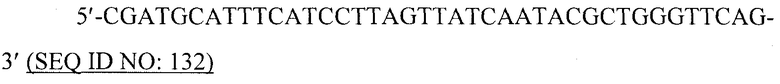
Продукт второй ПЦР расщепляли XbaI и Mph1103I и клонировали в экспрессирующем векторе pQβ10, который расщепляли такими же ферментами рестрикции. Реакции ПЦР осуществляли с использованием реагентов из набора для ПЦР и согласно протоколу производителя (MIX Fermentas, Vilnius, Lithuania).
Секвенирование с использованием способа прямого включения метки подтвердило требуемые мутации. Клетки E. coli, несущие pQβ-240, поддерживали эффективный синтез белка 14 кД, мигрирующего в SDS-ПААГ совместно с контрольным покрывающим белком Qβ, выделенным из частиц фага Qβ.
Полученная в результате аминокислотная последовательность: (SEQ ID NO:23)

Конструирование pQβ-243:
Плазмиду pQβ10 использовали в качестве исходной плазмиды для конструирования pQβ-243. Мутацию Asn10→Lys создавали обратной ПЦР. Обратные праймеры конструировали в обратных направлениях «хвост к хвосту»:

и
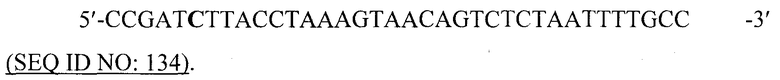
Продукты первой ПЦР использовали в качестве матриц для второй реакции ПЦР, в которой использовали верхний праймер
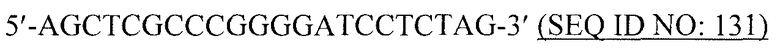
и нижний праймер
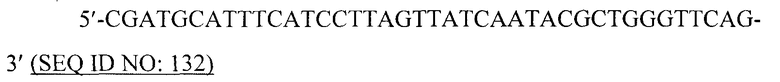
Продукт второй ПЦР расщепляли XbaI и Mph1103I и клонировали в экспрессирующем векторе pQβ10, который расщепляли такими же ферментами рестрикции. Реакции ПЦР осуществляли с использованием реагентов из набора для ПЦР и согласно протоколу производителя (MBI Fermentas, Vilnius, Lithuania).
Секвенирование с использованием способа прямого включения метки подтвердило требуемые мутации. Клетки E. coli, несущие pQβ-243, поддерживали эффективный синтез белка 14 кД, мигрирующего в SDS-ПААГ совместно с контрольным покрывающим белком Qβ, выделенным из частиц фага Qβ.
Полученная в результате аминокислотная последовательность: (SEQ ID NO:24)

Конструирование pQβ-250:
Плазмиду pQβ-240 использовали в качестве исходной плазмиды для конструирования pQβ-250. Мутацию Lys2→Arg создавали сайт-специфичным мутагенезом. Верхний праймер
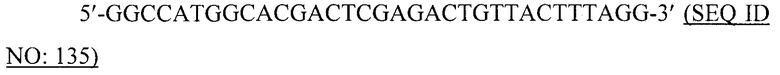
и нижний праймер
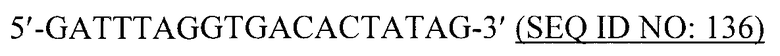
использовали для синтеза мутантного ПЦР-фрагмента, который вводили в экспрессирующий вектор pQβ-185 в уникальные сайты рестрикции NcoI и HindIII. Реакции ПЦР осуществляли с использованием реагентов из набора для ПЦР и согласно протоколу производителя (MBI Fermentas, Vilnius, Lithuania).
Секвенирование с использованием способа прямого включения метки подтвердило требуемые мутации. Клетки E. coli, несущие pQβ-250, поддерживали эффективный синтез белка 14 кД, мигрирующего в ПААГ совместно с контрольным покрывающим белком Qβ, выделенным из частиц фага Qβ.
Полученная в результате аминокислотная последовательность: (SEQ ID NO:25)

Конструирование pQβ-251:
Плазмиду pQβ10 использовали в качестве исходной плазмиды для конструирования pQβ-251. Мутацию Lys16→Arg создавали обратной ПЦР. Обратные праймеры конструировали в обратных направлениях «хвост к хвосту»:
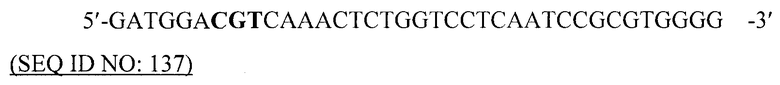
и

Продукты первой ПЦР использовали в качестве матриц для второй реакции ПЦР, в которой использовали верхний праймер
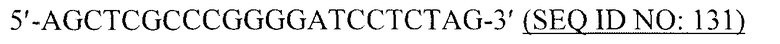
и нижний праймер
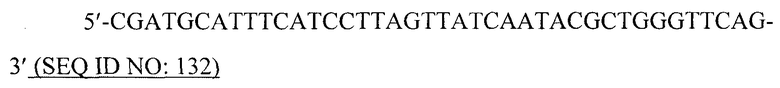
Продукт второй ПЦР расщепляли XbaI и Mph1103I и клонировали в экспрессирующем векторе pQβ10, который расщепляли такими же ферментами рестрикции. Реакции ПЦР осуществляли с использованием реагентов из набора для ПЦР и согласно протоколу производителя (MBI Fermentas, Vilnius, Lithuania).
Секвенирование с использованием способа прямого включения метки подтвердило требуемые мутации. Клетки E. coli, несущие pQβ-251, поддерживали эффективный синтез белка 14 кД, мигрирующего в SDS-ПААГ совместно с контрольным покрывающим белком Qβ, выделенным из частиц фага Qβ. Полученная в результате аминокислотная последовательность, кодируемая данной конструкцией, показана в SEQ ID NO:26.
Конструирование pQβ-259:
Плазмиду pQβ-251 использовали в качестве исходной плазмиды для конструирования pQβ-259. Мутацию Lys2→Arg создавали сайт-специфичным мутагенезом. Верхний праймер
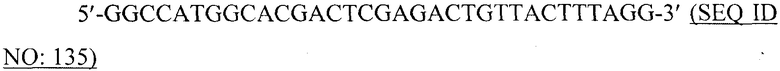
и нижний праймер
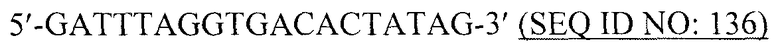
использовали для синтеза мутантного ПЦР-фрагмента, который вводили в экспрессирующий вектор pQβ-185 в уникальные сайты рестрикции NcoI и HindIII. Реакции ПЦР осуществляли с использованием реагентов из набора для ПЦР и согласно протоколу производителя (MBI Fermentas, Vilnius, Lithuania).
Секвенирование с использованием способа прямого включения метки подтвердило требуемые мутации. Клетки E. coli, несущие pQβ-259, поддерживали эффективный синтез белка 14 кД мигрирующего в SDS-ПААГ совместно с контрольным покрывающим белком Qβ, выделенным из частиц фага Qβ.
Полученная в результате аминокислотная последовательность: (SEQ ID NO:27)

Общие способы экспрессии и очистки Qβ и мутантов Qβ
Экспрессия
E. coli JM109 трансформировали плазмидами, экспрессирующими покрывающий белок Qβ. В 5 мл жидкой среды LB, содержащей 20 мкг/мл ампициллина, инокулировали клоны, трансформированные плазмидами, экспрессирующими покрывающий белок Qβ. Инокулированную культуру инкубировали при 37°C в течение 16-24 час без встряхивания. Затем полученный инокулят разводили 1:100 в 100-300 мл свежей среды LB, содержащей 20 мкг/мл ампициллина, и инкубировали при 37°C в течение ночи без встряхивания. Полученный в результате второй инокулят разводили 1:50 в среде M9, содержащей 1% казаминовых кислот и 0,2% глюкозы, в колбах и инкубировали при 37°C в течение ночи при встряхивании.
Очистка
Растворы и буферы для способа очистки:
1. Лизирующий буфер LB
50 мМ трис-HCl pH 8,0 с 5 мМ EDTA; 0,1% тритоном X100 и свежеприготовленным PMSF в концентрации 5 микрограмм на мл, без лизоцима и ДНКазы.
2. SAS
Насыщенный раствор сульфата аммония в воде.
3. Буфер NET
20 мМ трис-HCl, pH 7,8 с 5 мМ EDTA и 150 мМ NaCl.
4. ПЭГ
40% (мас./об.) полиэтиленгликоль 6000 в NET.
Разрушение и лизис
Замороженные клетки ресуспендировали в LB из расчета 2 мл/г клеток. Смесь обрабатывали ультразвуком при 22 кГц пять раз по 15 секунд с интервалами 1 мин, чтобы охладить раствор на льду. Затем лизат центрифугировали при 14000 об/мин в течение 1 час, используя ротор Janecki K 60. Все описанные ниже стадии центрифугирования осуществляли с использованием такого же ротора, если не оговорено особо. Надосадок хранили при 4°C, тогда как обломки клеток дважды промывали в LB. После центрифугирования надосадки и промывные фракции объединяли.
Фракционирование
К описанному выше объединенному лизату по каплям при перемешивании добавляли насыщенный раствор сульфата аммония. Объем SAS доводили до одной пятой общего объема, чтобы получить 20% насыщения. Раствор оставляли стоять в течение ночи и центрифугировали на следующий день при 14000 об/мин в течение 20 мин. Осадок промывали небольшим количеством 20% сульфата аммония и снова центрифугировали. Полученные надосадки объединяли и по каплям добавляли SAS, чтобы получить 40% насыщения. Раствор оставляли стоять в течение ночи и на следующий день центрифугировали при 14000 об/мин в течение 20 мин. Полученный осадок растворяли в буфере NET.
Хроматография
Капсид или белок VLP, перерастворенный в буфере NET, наносили на колонку с сефарозой CL-4B. В ходе хроматографии элюировали три пика. Первый пик главным образом содержал мембраны и фрагменты мембран, и его не собирали. Капсиды находились во втором пике, тогда как третий пик содержал другие белки E. coli.
Фракции пика объединяли и концентрацию NaCl доводили до конечной концентрации 0,65 М. По каплям при перемешивании добавляли объем раствора ПЭГ, соответствующий половине объединенной фракции пика. Раствор оставляли стоять в течение ночи без перемешивания. Белок капсида осаждали центрифугированием при 14000 об/мин в течение 20 мин. Затем его растворяли в минимальном объеме NET и снова нагружали на колонку с сефарозой CL-4B. Фракции пика объединяли и осаждали сульфатом аммония при 60% насыщении (мас./об.). После центрифугирования и перерастворения в буфере NET белок капсида нагружали на колонку с сефарозой CL-6B для повторной хроматографии.
Диализ и сушка
Фракции пиков, полученные, как описано выше, объединяли и диализовали против большого объема стерильной воды и лиофилизовали для хранения.
Экспрессия и очистка Qβ-240
Клетки (E. coli JM 109, трансформированные плазмидой pQβ-240) ресуспендировали в LB, обрабатывали ультразвуком пять раз по 15 секунд (рубашка, содержащая воду со льдом) и центрифугировали при 13000 об/мин в течение одного часа. Надосадок хранили при 4°C вплоть до дальнейшей обработки, тогда как обломки клеток 2 раза промывали 9 мл LB и, наконец, 9 мл 0,7 М мочевины в LB. Все надосадки объединяли и нагружали на колонку с сефарозой CL-4B. Объединенные фракции пика осаждали сульфатом аммония и центрифугировали. Затем перерастворенный белок снова очищали на колонке с сефарозой 2B и, наконец, на колонке с сефарозой 6B. Наконец пик капсида диализовали против большого объема воды и лиофилизовали, как описано выше. Сборку покрывающего белка в капсид подтверждали электронной микроскопией.
Экспрессия и очистка Qβ-243
Клетки (E. coli RR1) ресуспендировали в LB и обрабатывали, как описано в общем способе. Белок очищали двумя последовательными стадиями гель-фильтрации на колонке с сефарозой CL-4B и, наконец, на колонке с сефарозой CL-2B. Фракции пика объединяли и лиофилизовали, как описано выше. Сборку покрывающего белка в капсид подтверждали электронной микроскопией.
Экспрессия и очистка Qβ-250
Клетки (E. coli JM 109, трансформированные pOβ-250) ресуспендировали в LB и обрабатывали, как описано выше. Белок очищали гель-фильтрацией на колонке с сефарозой CL-4B и, наконец, на колонке с сефарозой CL-2B и лиофилизовали, как описано выше. Сборку покрывающего белка в капсид подтверждали электронной микроскопией.
Экспрессия и очистка Qβ-259
Клетки (E. coli JM 109, трансформированные pQβ-259) ресуспендировали в LB и обрабатывали ультразвуком. Обломки клеток один раз промывали 10 мл LB и второй раз 10 мл 0,7 М мочевины в LB. Белок очищали двумя стадиями хроматографии на основе гель-фильтрации на колонке с сефарозой CL-4B. Белок диализовали и лиофилизовали, как описано выше. Сборку покрывающего белка в капсид подтверждали электронной микроскопией.
ПРИМЕР 2
Инсерция пептида, содержащего остаток лизина, в c/e1-эпитоп HBcAg (1-149)
Эпитоп c/e1 (остатки с 72 по 88) HBcAg расположен в верхушечной области на поверхности капсида вируса гепатита B (HBcAg). Часть данной области (пролин 79 и аланин 80) генетическим способом заменяли пептидом Gly-Gly-Lys-Gly-Gly (конструкция HBcAg-Lys: (SEQ ID NO:117)). Введенный остаток лизина содержит реакционно-способную аминогруппу в своей боковой цепи, которую можно использовать для межмолекулярного химического перекрестного сшивания частиц HBcAg с любым антигеном, содержащим свободную группу цистеина.
ДНК HBcAg-Lys, кодирующую аминокислотную последовательность, показанную в SEQ ID NO:78, создавали с помощью ПЦР: два фрагмента, кодирующие фрагменты HBcAg (аминокислотные остатки с 1 по 78 и с 81 по 149) амплифицировали отдельно в ПЦР. Праймеры, используемые для указанных ПЦР, также вводили последовательность ДНК, кодирующую пептид Gly-Gly-Lys-Gly-Gly (SEQ ID NO:117). Фрагмент HBcAg (с 1 по 78) амплифицировали с pEco63, используя праймеры EcoRIHBcAg(s) и Lys-HBcAg(as). Фрагмент HBcAg (с 81 по 149) амплифицировали с pEco63, используя праймеры Lys-HBcAg(s) и HBcAg(1-149)Hind(as). Праймеры Lys-HBcAg(as) и Lys-HBcAg(s) вводили комплементарные последовательности ДНК на концах двух продуктов ПЦР, обеспечивая слияние двух продуктов ПЦР в последующей сборке на основе ПЦР. Сблокированные фрагменты амплифицировали в ПЦР, используя праймеры EcoRIHBcAg(s) и HbcAg(1-149)Hind(as).
Для ПЦР использовали 100 пмоль каждого олигонуклеотида и 50 нг ДНК-матриц в 50 мл реакционных смесей, содержащих 2 единицы полимеразы Pwo, 0,1 мМ dNTP и 2 мМ MgSO4. В обеих реакциях температурные циклы осуществляли следующим образом: 94°C в течение 2 минут; 30 циклов 94°C (1 минута), 50°C (1 минута), 72°C (2 минуты).
Последовательности праймеров:
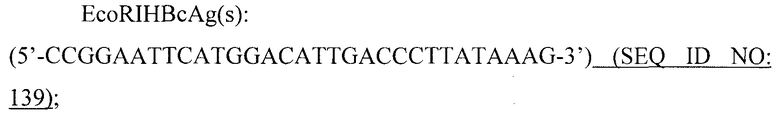

Для слияния двух ПЦР-фрагментов с помощью ПЦР использовали 100 пмоль праймеров EcoRIHBcAg(s) и HBcAg(1-149)Hind(as) со 100 нг двух очищенных ПЦР-фрагментов в 50 мл реакционной смеси, содержащей 2 единицы полимеразы Pwo, 0,1 мМ dNTP и 2 мМ MgSO4. Условия циклов ПЦР представляли собой: 94°C в течение 2 минут; 30 циклов 94°C (1 минута), 50°C (1 минута), 72°C (2 минуты). Продукт, полученный при ПЦР-сборке, анализировали электрофорезом в агарозном геле, очищали и расщепляли в течение 19 часов в соответствующем буфере с ферментами рестрикции EcoRI и HindIII. Полученный при расщеплении фрагмент ДНК лигировали в EcoRI/HindIII-расщепленный вектор pKK, чтобы получить экспрессирующий вектор pKK-HBcAg-Lys. Инсерцию ПЦР-продукта в вектор анализировали с помощью рестрикционного анализа ферментами EcoRI/HindIII и секвенирования ДНК вставки.
ПРИМЕР 3
Экспрессия и очистка HBcAg-Lys
Штаммы E. coli K802 или JM109 трансформировали вектором pKK-HBcAg-Lys. 1 мл ночной культуры бактерий использовали для того, чтобы инокулировать 100 мл среды LB, содержащей 100 мкг/мл ампициллина. Указанную культуру выращивали в течение 4 часов при 37°C до достижения OD при 600 нм, примерно равной 0,8. Индукцию синтеза HBcAg-Lys осуществляли добавлением IPTG до конечной концентрации 1 мМ. После индукции бактерии дополнительно встряхивали при 37°C в течение 4 часов. Бактерии собирали центрифугированием при 5000 x g в течение 15 минут. Осадок замораживали при -80°C. Осадок размораживали и ресуспендировали в буфере для лизиса бактерий (10 мМ Na2HPO4, pH 7,0; 30 мМ NaCl; 0,25% твин-20; 10 мМ EDTA) с добавлением 200 мкг/мл лизоцима и 10 мкг бензоназы (Merck). Клетки инкубировали в течение 30 минут при комнатной температуре и разрушали обработкой ультразвуком. Клетки E. coli, несущие экспрессирующую плазмиду pKK-HBcAg-Lys или контрольную плазмиду, использовали для индукции экспрессии HBcAg-Lys с помощью IPTG. Перед добавлением IPTG из культуры бактерий, несущих плазмиду pKK-HBcAg-Lys и из культуры бактерий, несущих контрольную плазмиду, брали образец. Через четыре часа после добавления IPTG снова брали образцы из культуры, содержащей pKK-HBcAg-Lys, и из контрольной культуры. Экспрессию белка контролировали с использованием SDS-ПААГ с последующим окрашиванием Кумасси.
Затем лизат центрифугировали в течение 30 минут при 12000 x g, чтобы удалить нерастворимые обломки клеток. Надосадок и осадок анализировали Вестерн-блоттингом, используя моноклональное антитело против HBcAg (YVS1841, приобретенное из Accurate Chemical and Scientific Corp., Westbury, NY, USA), показав, что значительное количество белка HBcAg-Lys является растворимым. Коротко, лизаты клеток E. coli, экспрессирующих HBcAg-Lys, и лизаты контрольных клеток центрифугировали при 14000 x g в течение 30 минут. Надосадок (= растворимая фракция) и осадок (= нерастворимая фракция) разделяли и разбавляли в SDS-буфере для образцов до равных объемов. Образцы анализировали в SDS-ПААГ с последующим Вестерн-блоттингом с моноклональным антителом YVS 1841.
Осветленные лизаты клеток использовали для центрифугирования в ступенчатом градиенте, используя ступенчатый градиент сахарозы, состоящий из 4 мл 65% раствора сахарозы, на который наслаивали 3 мл 15% раствора сахарозы, а затем 4 мл бактериального лизата. Образец центрифугировали в течение 3 час при 100000 x g, при 4°C. После центрифугирования собирали фракции объемом 1 мл с верха градиента и анализировали в SDS-ПААГ с последующим окрашиванием Кумасси. Белок HBcAg-Lys выявляли при окрашивании Кумасси.
Обогащенная фракция белка HBcAg-Lys была расположена на границе между 15 и 65% сахарозой, что свидетельствует о том, что белок образовал частицу капсида. Большинство бактериальных белков оставалось в не содержащем сахарозу верхнем слое градиента, таким образом, центрифугирование частиц HBcAg-Lys в ступенчатом градиенте приводило как к обогащению, так и к частичной очистке частиц.
Экспрессию и очистку HBcAg-Lys в крупном масштабе осуществляли следующим образом. Ночную культуру получали, высевая единичную колонию в 100 мл LB, 100 мкг/мл ампициллина и выращивая культуру в течение ночи при 37°C. На следующий день 25 мл предварительной культуры разбавляли в 800 мл среды LB с ампициллином и культуру выращивали до оптической плотности OD600, равной 0,6-0,8. Затем культуру индуцировали 1 мМ IPTG и оставляли расти еще в течение 4 часов. Клетки собирали и лизировали по существу так, как описано выше.
Затем HBcAg-Lys очищали, сначала осаждая белок сульфатом аммония (30% насыщения) из осветленного лизата клеток, затем нанося перерастворенный осадок на колонку для гель-фильтрации (Sephacryl S-400, Pharmacia). Объединенные фракции снова осаждали сульфатом аммония, осадок перерастворяли и наносили второй раз на такую же колонку для гель-фильтрации. Наконец фракции объединяли и концентрировали, и концентрацию оценивали с использованием теста Бредфорда (BioRad).
ПРИМЕР 4
Конструирование HBcAg, лишенного свободных остатков цистеина и содержащего встроенный остаток лизина
Коровый антиген гепатита (HBcAg), называемый в данном описании HBcAg-lys-2cys-Mut, лишенный остатков цистеина в положениях, соответствующих 48 и 107 в SEQ ID NO:77, и содержащий встроенный остаток лизина, конструировали, используя следующие способы.
Две мутации вводили сначала посредством отдельной амплификации трех фрагментов гена HBcAg-Lys, полученных, как описано выше в примере 2, со следующими комбинациями праймеров для ПЦР. Использовали способы ПЦР и обычные способы клонирования для получения гена HBcAg-lys-2cys-Mut.
Коротко, для получения фрагмента 1 использовали следующие праймеры:
Праймер 1: EcoRIHBcAg(s)
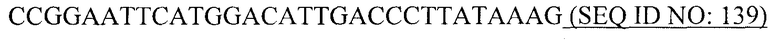
Праймер 2: 48as
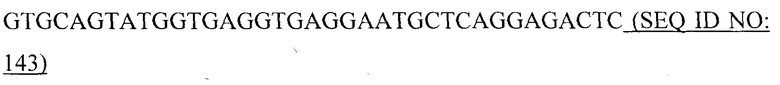
Для получения фрагмента 2 использовали следующие праймеры:
Праймер 3: 48s
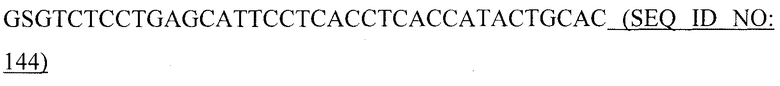
Праймер 4: 107as

Для получения фрагмента 3 использовали следующие праймеры:
Праймер 5: HBcAg149hind-as

Праймер 6: 107s
Затем фрагменты 1 и 2 объединяли с использованием праймеров ПЦР EcoRIHBcAg(s) и 107as, получая фрагмент 4. Затем фрагмент 4 и фрагмент 3 объединяли с использованием праймеров EcoRIHBcAg(s) и HBcAg149hind-as, получая полноразмерный ген. Затем полноразмерный ген расщепляли ферментами EcoRI(GAATTC) и HindIII(AAGCTT) и клонировали в векторе pKK (Pharmacia), разрезанными в таких же сайтах рестрикции. Экспрессию и очистку HBcAg-lys-2cys-Mut осуществляли, как указано в примере 3.
ПРИМЕР 5
Конструирование HBcAg1-185-Lys
Коровый антиген гепатита (HBcAg) 1-185 модифицировали, как описано в примере 2. Часть области эпитопа c/e1 (остатки с 72 по 88) (пролин 79 и аланин 80) генетическим способом заменяли пептидом Gly-Gly-Lys-Gly-Gly (конструкция HBcAg1-185-Lys: SEQ ID NO:117). Введенный остаток лизина содержит реакционно-способную аминогруппу в своей боковой цепи, которую можно использовать для межмолекулярного химического перекрестного сшивания частиц HBcAg с любым антигеном, содержащим свободную группу цистеина. Для получения гена HBcAg1-185-Lys использовали способы ПЦР и обычные способы клонирования.
Последовательность Gly-Gly-Lys-Gly-Gly (SEQ ID NO:117) встраивали путем амплификации двух отдельных фрагментов гена HBcAg из pEco63, как описано выше в примере 2, и затем посредством слияния двух фрагментов с помощью ПЦР, чтобы собрать полноразмерный ген. Использовали следующие комбинации ПЦР-праймеров:
фрагмент 1:
праймер 1: EcoRIHBcAg(s) (см. пример 2)
праймер 2: Lys-HBcAg(as) (см. пример 2)
фрагмент 2:
праймер 3: Lys-HBcAg(s) (см. пример 2)
праймер 4: HBcAgwtHindIIII

Сборка:
праймер 1: EcoRIHBcAg(s) (см. пример 2)
праймер 2: HBcAgwtHindIIII
Затем собранный полноразмерный ген расщепляли ферментами EcoRI(GAATTC) и HindIII(AAGCTT) и клонировали в векторе pKK (Pharmacia), разрезанном в таких же сайтах рестрикции.
ПРИМЕР 6
Слияние пептидного эпитопа в MIR-области HbcAg
Остатки 79 и 80 HBcAg1-185 заменяли эпитопом CεH3 последовательности VNLTWSRASG (SEQ ID NO:149). Последовательность CεH3 происходит из последовательности третьего константного домена тяжелой цепи IgE человека. Эпитоп встраивали в последовательность HBcAg1-185, используя способ ПЦР-сборки. На первой стадии ПЦР ген HBcAg1-185, происходящий из ATCC-клона pEco63 и амплифицированный с праймерами HBcAg-wt EcoRI fwd и HBcAg-wt Hind III rev, использовали в качестве матрицы в двух отдельных реакциях, чтобы амплифицировать два фрагмента, содержащие элементы последовательности, кодирующие последовательность CεH3. Затем указанные два фрагмента подвергали сборке на второй стадии ПЦР в реакции ПЦР-сборки.
Комбинации праймеров на первой стадии ПЦР: CεH3fwd с HBcAg-wt Hind III rev, и HBcAg-wt EcoRI fwd с CεH3rev. В реакции ПЦР-сборки два фрагмента, выделенные на первой стадии ПЦР, сначала подвергали сборке в течение 3 циклов ПЦР без внешних праймеров, которые добавляли к реакционной смеси позже для следующих 25 циклов. Внешние праймеры: HBcAg-wt EcoRI fwd и HBcAg-wt Hind III rev.
Продукт ПЦР клонировали в pKK223.3, используя сайты EcoRI и HindIII, для экспрессии в E. coli (см. пример 2). Химерную VLP экспрессировали в E. coli и очищали, как описано в примере 2. В объеме элюирования, в котором HBcAg1-185-CεH3 элюировался при гель-фильтрации, показана сборка слитых белков в химерную VLP.
Последовательности праймеров:

ПРИМЕР 7
Слияние эпитопа пептида RANKL в MIR-области HbcAg
Остатки 79 и 80 HBcAg1-185 заменяли эпитопом пептида RANKL с последовательностью: SIKIPSSH (SEQ ID NO:88). Конструировали два перекрывающихся праймера с использованием такой же методики, которая описана в примере 6, и слитый белок конструировали с помощью ПЦР-сборки. Продукт ПЦР клонировали в векторе pKK223.3 и экспрессировали в E. coli K802. Химерные VLP экспрессировали и очищали, как описано в примере 3.
ПРИМЕР 8
Слияние эпитопа пептида RANKL с C-концом белка A1 Qβ, укороченного в положении 19 CP-удлинения
Использовали праймер, который отжигается с 5'-концом гена A1 Qβ, и праймер, который отжигается с 3'-концом гена A1 и содержит дополнительный элемент последовательности, кодирующий эпитоп пептида RANKL с последовательностью: SIKIPSSH (SEQ ID NO:88), в реакции ПЦР с pQβ10 в качестве матрицы. Продукт ПЦР клонировали в pQβ10 (Kozlovska T.M. et al., Gene 137: 133-37 (1993)), и химерные VLP экспрессировали и очищали, как описано в примере 1.
ПРИМЕР 9
Инсерция эпитопа пептида RANKL между положениями 2 и 3 покрывающего белка fr
Комплементарные праймеры, кодирующие последовательность эпитопа пептида RANKL с последовательностью: SIKIPSSH (SEQ ID NO:88) и содержащие Bsp119I-совместимые концы и дополнительные нуклеотиды, обеспечивающие инсерцию в рамке считывания, встраивали в Bsp119I-сайт вектора pFrd8 (Pushko, P. et al., Prot. Eng. 6: 883-91 (1993)) стандартными способами молекулярной биологии. Альтернативно выступающие концы вектора pFrd8 заполняли фрагментом Кленова после расщепления Bsp119I и олигонуклеотиды, кодирующие последовательность белка RANKL, фрагмента RANKL или пептида RANKL и дополнительные нуклеотиды для клонирования в рамке, лигировали в pFrd8 после обработки фрагментом Кленова. Клоны со вставкой в правильной ориентации анализировали секвенированием. Экспрессию и очистку химерного слитого белка в E. coli JM109 или E. coli K802 осуществляли, как описано в Pushko, P. et al, Prot. Eng. 6: 883-91 (1993), за исключением стадий хроматографии, которые выполняли с использованием сефарозы CL-4B или сефакрила S-400 (Pharmacia). Лизат клеток осаждали сульфатом аммония и очищали, используя две последовательные стадии гель-фильтрации, подобно способу, описанному для Qβ в примере 1.
ПРИМЕР 10
Инсерция эпитопа пептида RANKL между положениями 67 и 68 p1-белка Ty1 в векторе pOGS8111
Синтезировали два комплементарных олигонуклеотида, кодирующих эпитоп пептида RANKL с последовательностью: SIKIPSSH (SEQ ID NO:88) с концами, совместимыми с NheI-сайтом pOGS8111. Добавляли дополнительные нуклеотиды, чтобы обеспечить инсерцию в рамке считывания последовательности, кодирующей эпитоп RANKL, согласно описанию EP06777111. Аминокислоты AS и SS, фланкирующие встроенный эпитоп, кодируются измененными NheI-сайтами, возникающими в результате инсерции олигонуклеотида в TyA(d)-ген pOGS8111.
POGS8111 трансформировали в штамм MC2 S. cerevisiae для экспрессии химерной VLP Ty, как описано в EP0677111 и ссылках в данном описании. Химерную VLP Ty очищали ультрацентрифугированием в градиенте сахарозы, как описано в EP0677111.
ПРИМЕР 11
Инсерция эпитопа пептида RANKL в основной белок капсида L1 вируса папилломы типа 1 (BPV-1)
Последовательностью, кодирующей эпитоп пептида RANKL с последовательностью SIKIPSSH (SEQ ID NO:88), заменяли последовательность, кодирующую аминокислоты 130-136 гена L1 BPV-1, клонированного в векторе pFastBac1 (GIBCO/BRL), как описано (Chackerian, B. et al., Proc. Natl. Acad. USA 96: 2373-2378 (1999)). Последовательность конструкции проверяли анализом нуклеотидной последовательности. Рекомбинантный бакуловирус создавали с использованием бакуловирусной системы GIBCO/BRL, как описано производителем. Химерные VLP очищали из инфицированных бакуловирусом клеток Sf9, как описано Kirnbauer, R. et al., Proc. Natl. Acad. Sci. 89:12180-84 (1992) и Greenstone, H.L., et al., Proc. Natl. Acad. Sci. 95:1800-05 (1998).
ПРИМЕР 12
Введение N-концевого cys-содержащего линкера, экспрессия и очистка RANKL
Фрагмент RANKL рекомбинантно экспрессировали с N-концевым линкером, содержащим один цистеин для связывания с VLP.
Конструирование экспрессирующей плазмиды
C-концевую кодирующую область гена RANKL амплифицировали в ПЦР с олигонуклеотидами RANKL-UP и RANKL-DOWN (олигонуклеотиды: RANKL-UP: 5'CTGCCAGGGGCCCGGGTGCGGCGGTGGCCATCATCACCACCATCACCAGCGCTTCTCAGGAG-3' (SEQ ID NO:154); RANKL-DOWN: 5'-CCGCTCGAGTTAGTCTATGTCCTGAACTTTGAAAG-3'(SEQ ID NO:155). RANKL-UP имел внутренний сайт ApaI, а RANKL-DOWN имел внутренний сайт XhoI. Продукт ПЦР расщепляли ApaI и XhoI и лигировали в pGEX-6p1 (Amersham Pharmacia). Полученную в результате плазмиду назвали pGEX-RANKL. Все стадии выполняли, используя стандартные протоколы молекулярной биологии, и последовательность проверяли. Плазмида pGEX-RANKL кодирует слитый белок глутатион-S-трансфераза-сайт расщепления PreScission-содержащий цистеин аминокислотный линкер-RANKL (GST-PS-C-RANKL). Содержащий цистеин аминокислотный линкер имел последовательность GPGCGGG (SEQ ID NO:156). Конструкция также содержит гексагистидиновую метку между содержащим цистеин аминокислотным линкером и последовательностью RANKL. Последовательности полученных в результате конструкций кДНК и белка приведены в виде SEQ-ID NO:94 и SEQ-ID NO:95.
Экспрессия и очистка C-RANKL
Компетентные клетки Escherichia coli BL21 (DE3) Gold pLys трансформировали плазмидой pGEX-RANKL. Отдельные колонии из чашек с агаром, содержащим ампициллин, размножали в жидкой культуре (среда LB, 100 мкг/мл ампициллина) и инкубировали при 30°C в течение ночи при встряхивании 220 об/мин. Затем в один литр LB (с 100 мкг/мл ампициллина) высевали ночную культуру 1:100 об./об. и выращивали при 24°C до OD600=1. Экспрессию индуцировали 0,4 мМ IPTG. Клетки собирали через 16 час и центрифугировали при 5000 об/мин. Осадок клеток суспендировали в лизирующем буфере (50 мМ трис-HCl, pH 8,0; 25% сахарозы; 1 мМ EDTA, 1% NaN3; 10 мМ DTT; 5 мМ MgCl2; 1 мг/мл лизоцима; 0,4 U/мл ДНКазы) в течение 30 мин. Затем добавляли 2,5 объема буфера A (50 мМ трис-HCl, pH 8,0; 1% тритон X100; 100 мМ NaCl; 0,1% NaN3; 10 мМ DTT; 1 мМ PMSF) и инкубировали при 37°C в течение 15 мин. Клетки обрабатывали ультразвуком и осаждали при 9000 об/мин в течение 15 мин. Надосадок сразу же использовали для GST-аффинной хроматографии.
Колонку FF, улавливающую GST, объемом 5 мл (Amersham Pharmacia) уравновешивали в PBS, pH 7,3 (140 мМ NaCl; 2,7 мМ KCl; 10 мМ Na2HPO4; 1,8 мМ KH2PO4). Надосадок наносили на колонку FF, улавливающую GST, объемом 5 мл и затем колонку промывали объемом PBS, равным 5 объемам колонки. Белок GST-PS-C-RANKL элюировали, используя 50 мМ трис-HCl, pH 8,0, содержащий 10 мМ восстановленный глутатион.
Очищенный белок GST-PS-C-RANKL расщепляли с использованием протеазы PreScission (Amersham Pharmacia). Расщепление осуществляли при 37°C в течение 1 часа, используя молярное отношение 500/1 GST-PS-C-RANKL к PreScission.
Кроме того, в реакционной смеси для расщепления протеазой осуществляли замену буфера, используя колонку для обессоливания HiPrep 26/10 (Amersham Pharmacia), фракции, содержащие белки, объединяли и сразу же использовали для другой стадии GST-аффинной хроматографии с использованием таких условий, как условия, описанные выше. Очистку C-RANKL анализировали в SDS-ПААГ-геле в восстанавливающих условиях, показанном на фиг.1. Отщепленный C-RANKL присутствует в проходящем потоке (несвязанная фракция), тогда как нерасщепленный GST-PS-C-RANKL, отщепленный GST-PS и PreScission остаются связанными с колонкой. Белок C-RANKL (SEQ ID NO:96) ожидаемого размера 22 кД получали очищенным в высокой степени.
ПРИМЕР 13
Введение C-концевого cys-содержащего линкера, экспрессия и очистка RANKL
Фрагмент RANKL рекомбинантно экспрессировали с C-концевым линкером, содержащим один цистеин для связывания с VLP.
Конструирование экспрессирующей плазмиды
MCS pET22b(+) (Novagen, Inc.) меняли на GTTTAACTTTAAGAAGGAGATATACATATGGATCCGGCTAGCGCTCGAGGGTTTAAACGGCGGCCGCATGCACC (SEQ ID NO:157) заменой исходной последовательности от сайта NdeI до сайта XhoI отжигаемыми олигонуклеотидами primerMCS-1F и primerMCS-1R (отжигание в 15 мМ трис-HCl-буфере, pH 8). Полученную в результате плазмиду назвали pMod00, которая имела сайты рестрикции NdeI, BamHI, NheI, XhoI, PmeI и NotI в своем MCS. Отжигаемую пару олигонуклеотидов Bamhis6-EK-Nhe-F и Bamhis6-EKNhe-R и отжигаемую пару олиго1F-C-глицин-линкер и олиго1R-C-глицин-линкер вместе лигировали в расщепленную BamHI-NotI плазмиду pMod00, чтобы получить pModEC1, которая имела N-концевую гистидиновую метку, сайт расщепления энтерокиназой и C-концевой аминокислотный глициновый линкер, содержащий один остаток цистеина.
Фрагмент ДНК, содержащий ген глутатион-S-трансферазы с C-концевым сайтом расщепления энтерокиназой, амплифицировали с помощью ПЦР, используя олигонуклеотиды GST-UP и GST-EK из плазмиды SP-GST-EK-pCEP-Pu (Wuttke, M., et al., J. Biol. Chem., 276: 36839-36848), расщепленной NheI и BamHI, и клонировали в векторе pModEC1.
Полученная в результате плазмида pMod-GST-EK-C1 содержит ген, кодирующий глутатион-S-трансферазу, слитый с сайтом расщепления энтерокиназой и C-концевым cys-содержащим линкером. Затем C-концевую кодирующую последовательность RANKL амплифицировали с помощью ПЦР, используя олигонуклеотиды mRANKL-1 и mRANKL-2, расщепляли NheI и XhoI и клонировали в плазмиде pMod-GST-EK-C1. Полученная в результате плазмида pMod-GST-EK-mRANKL-C1 кодирует слитый белок, состоящий из глутатион-S-трансферазы, сайта расщепления энтерокиназой, фрагмента RANKL и содержащего цистеин аминокислотного линкера (GST-EK-RANKL-C). Последовательности полученной в результате кДНК и конструкций белка приведены в виде SEQ ID NO:97 и SEQ ID NO:98.
Последовательности олигонуклеотидов:
Экспрессия и очистка RANKL-C
Компетентные клетки Escherichia coli BL21 (DE3) Gold pLys трансформировали плазмидой pMod-GST-EK-mRANKL-C1. Отдельные колонии из чашек с агаром, содержащим ампициллин, размножали в 100 мл жидкой культуры (среда LB, 200 мкг/мл ампициллина) и инкубировали при 30°C в течение ночи и встряхивании 220 об/мин. Затем в один литр среды (SB с 150 мМ MOPS; pH 7,0; 200 мкг/мл Amp) высевали ночную культуру 1:100 об./об. и выращивали при 30°C и встряхивании 125 об/мин до OD600=2,5. Затем культуры перемещали на 18°C и через 30 мин индуцировали экспрессию белка добавлением 0,1 мМ IPTG. Бактерии собирали после ночной культуры при 18°C центрифугированием (SLA-3000, 15 мин, 4°C, 6000 об/мин), ресуспендировали в 40 мл лизирующего буфера (10 мМ Na2HPO4, 30 мМ NaCl, 10 мМ EDTA и 0,25% твин-20) и инкубировали в течение 30 мин на льду с 0,8 мг/мл лизоцима. Затем бактерии лизировали обработкой ультразвуком и инкубировали в течение 30 мин при КТ с 0,2 М MgCl2 и 8 мкл бензоназы. Лизат очищали от нерастворимого материала центрифугированием (SS-34, 30 мин, 4°C, 20000 об/мин) и сразу же использовали для аффинной хроматографии на глутатион-сефарозе.
Колонку FF, улавливающую GST, объемом 5 мл (Amersham Pharmacia) уравновешивали лизирующим буфером (10 мМ Na2HPO4, 30 мМ NaCl, 10 мМ EDTA и 0,25% твин-20) и нагружали осветленный лизат при постоянной скорости потока 0,5 мл/мин. Затем колонку три раза промывали 5 объемами колонки лизирующего буфера и белок GST-EK-RANKL-C элюировали в 9 фракциях по 1 мл элюирующего буфера (50 мМ трис-HCl, pH 8,0; 10 мМ восстановленный глутатион) каждая. Очистка, показанная на фиг.2A, давала слитый белок GST-EK-RANKL-C размером примерно 45 кД с небольшой долей GST-EK.
Элюированные фракции объединяли и очищенный белок GST-EK-RANKL-C расщепляли с использованием энтерокиназы MaxTM (Invitrogen). Расщепление осуществляли при 4°C в течение 16 часов с использованием 10 единиц энтерокиназы Max на мг очищенного GST-EK-RANKL-C. На фиг.2B показано, что реакция расщепления приводила к получению RANKL-C с кажущейся М.м. примерно 16 кД.
После расщепления раствор белка диализовали против PBS, pH 7,2 (140 мМ NaCl; 2,7 мМ KCl; 10 мМ Na2HPO4; 1,8 мМ KH2PO4) и глутатион-S-трансферазу удаляли второй аффинной хроматографией на глутатион-сефарозе. Для этого колонку FF, улавливающую GST, объемом 5 мл уравновешивали PBS pH 7,2 и на колонку нагружали раствор белка с постоянной скоростью потока 0,5 мл/мин. Фракции белка анализировали до и после хроматографии на глутатион-сефарозе в SDS-гелях. Как показано на фиг.2C, отщепленный белок RANKL-C (SEQ ID NO:99) находился в проходящем потоке с высокой степенью чистоты, тогда как белок GST-EK оставался связанным с колонкой.
ПРИМЕР 14
Связывание C-RANKL с белком капсида Qβ
Раствор объемом 1,48 мл капсидного белка Qβ в концентрации 6 мг/мл в 20 мМ Hepes, 150 мМ NaCl, pH 7,2, в течение 30 минут при 25°C подвергали взаимодействию с 14,8 мкл SMPH (Pierce) (из 100 мМ маточного раствора, растворенного в ДМСО). Затем реакционный раствор дважды диализовали в течение 3 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, pH 7,0, при 4°C. 230 мкл раствора белка C-RANKL в концентрации 9,8 мг/мл в 20 мМ Hepes, 150 мМ NaCl, pH 7,2 в течение 30 мин при 25°C подвергали взаимодействию с 1,7 мкл 100 мМ раствора TCEP (Pierce). Для связывания 80 мкл дериватизованного и диализованного Qβ затем смешивали с 24 мкл восстановленного C-RANKL и 96 мкл 20 мМ Hepes, 150 мМ NaCl, pH 7,0, и инкубировали в течение ночи при 25°C.
Связанные продукты анализировали в 16% SDS-ПААГ-гелях в восстанавливающих условиях. Гели либо красили Кумасси бриллиантовым голубым, либо подвергали блоттингу на нитроцеллюлозные мембраны. В последнем случае мембраны блокировали и инкубировали либо с поликлональной кроличьей антисывороткой против Qβ (разведение 1:2000), затем с конъюгированным с пероксидазой хрена антителом козы против IgG кролика (разведения 1:5000) или с мышиным моноклональным анти-RANKL-антителом (разведение 1:2000), затем с конъюгированным с пероксидазой хрена антимышиным антителом козы (разведение 1:5000). Затем блоты проявляли с помощью реагентов для регистрации Вестерн-блоттинга ECLTM (Amersham Pharmacia). Результаты показаны на фиг.3A и фиг.3B. Связанные продукты можно было регистрировать в окрашенных Кумасси гелях (фиг.3A) и с помощью как анти-Qβ-антисыворотки, так и с помощью анти-RANKL-антитела (фиг.3B), что ясно свидетельствует о ковалентном связывании C-RANKL с белком капсида Qβ.
ПРИМЕР 15
Иммунизация мышей C-RANKL, связанным с белком капсида Qβ
Всего 8 самок мышей Balb/c вакцинировали C-RANKL, связанным с белком капсида Qβ. 25 мкг суммарного белка каждого образца разбавляли в PBS до 200 мкл и инъецировали подкожно (100 мкл с двух сторон брюшка) в 0 день, 16 день и 64 день. Четыре мыши получали вакцину без добавления адъювантов, тогда как четыре другие получали вакцину с добавлением квасцов. У мышей брали кровь из ретроорбитальной области в 0, 16, 23, 64 и 78 день и их сыворотку анализировали с использованием RANKL-специфичного ELISA.
ПРИМЕР 16
Выявление RANKL-специфичных антител в ELISA
Планшеты для ELISA покрывали C-RANKL в концентрации 10 мкг/мл. Планшеты блокировали и затем инкубировали с серийными разведениями сыворотки мышей, полученной на 16, 23, 64 и 78 дни. Связанные антитела выявляли с помощью меченного ферментом антитела против IgG мыши. В качестве контроля также тестировали полученные до иммунизации сыворотки тех же самых мышей. На фиг.4 показаны средние титры RANKL-специфичных антител, которые можно выявить в сыворотке мышей, которых иммунизировали Qβ-C-RANKL в присутствии или без квасцов. Титры в ELISA выражали в виде разведений сыворотки, которые дают половину максимальной OD в анализе ELISA. У мышей, иммунизированных без квасцов, средний титр достигал 28000, тогда как средний титр у мышей, иммунизированных с добавлением квасцов, составлял 160000. Сыворотка до иммунизации не проявляла никакой реакционной способности по отношению к C-RANKL. Это ясно свидетельствует о том, что конъюгат RANKL-VLP способен индуцировать высокий титр антител против аутологичного белка.
ПРИМЕР 17
Ингибирование взаимодействия RANKL-RANK сывороткой мышей, иммунизированных Qβ-C-RANKL
Чтобы проверить, обладают ли антитела, образованные у мышей, вакцинированных Qβ-C-RANKL, нейтрализующей активностью, разработали анализ связывания in vitro для RANKL и родственного ему рецептора RANK. Для этого планшеты для ELISA покрывали белком C-RANKL при концентрации 10 мкг/мл и инкубировали с серийными разведениями очищенного слитого белка RANK-Fc или неродственного Fc-слитого белка в качестве негативного контроля. Связывающиеся белки выявляли с использованием конъюгированного с пероксидазой хрена анти-Fc-антитела. На фиг.5A показан результат данного анализа. Обнаружено, что очищенный слитый белок RANK-Fc связывается с высокой аффинностью (половина максимального связывания при 1-3 нМ) с его лигандом RANKL, тогда как при использовании неродственного Fc-слитого белка фактически не наблюдали связывания.
Затем сыворотки мышей, вакцинированных C-RANKL, связанным с Qβ, тестировали в отношении их способности ингибировать связывание RANKL с RANK-Fc. Для этого планшеты ELISA покрывали белком C-RANKL в концентрации 10 мкг/мл и инкубировали вместе с серийными разведениями сывороток мышей, полученных на 78 день, смешанными с 1 нМ слитого белка RANK-Fc. Связывание слитого белка RANK-Fc с C-RANKL регистрировали с помощью конъюгированного с пероксидазой хрена анти-Fc-антитела. На фиг.5B показано, что все 8 мышей, которые получали вакцину, продуцировали антитела, которые специфично ингибировали связывание RANK-Fc с C-RANKL. В среднем, половину максимального ингибирования достигали в случае использования разведений сывороток 1:90. Это ясно свидетельствует о том, что иммунизация конъюгатом RANKL-VLP индуцирует антитела с высокими титрами, которые способны ингибировать взаимодействие RANKL с его рецептором RANK. Таким образом, ингибирование взаимодействия RANK-RANKL посредством инъекции специфичного варианта согласно данному изобретению может устранять или предотвращать заболевания кости, характеризующиеся повышенной резорбцией кости.
ПРИМЕР 18
Экспрессия и очистка рекомбинантных VLP AP205
A. Экспрессия рекомбинантных VLP AP205
E. coli JM109 трансформировали плазмидой pAP283-58. В 5 мл жидкой среды LB с 20 мкг/мл ампициллина инокулировали отдельную колонию и инкубировали при 37°C в течение 16-24 час без встряхивания.
Полученный инокулят разводили 1:100 в 100-300 мл среды LB, содержащей 20 мкг/мл ампициллина, и инкубировали при 37°C в течение ночи без встряхивания. Полученный в результате второй инокулят разводили 1:50 в среде 2TY, содержащей 0,2% глюкозы и фосфат для забуферивания, и инкубировали при 37°C в течение ночи на качалке. Клетки собирали центрифугированием и замораживали при -80°C.
B. Очистка рекомбинантных VLP AP205
Растворы и буферы:
1. Лизирующий буфер
50 мМ трис-HCl, pH 8,0 с 5 мМ EDTA; 0,1% тритоном X100 и PMSF при концентрации 5 микрограмм на мл.
2. SAS
Насыщенный раствор сульфата аммония в воде.
3. Буфер NET.
20 мМ трис-HCl, pH 7,8 с 5 мМ EDTA и 150 мМ NaCl.
4. ПЭГ
40% (мас./об.) полиэтиленгликоль 6000 в NET.
Лизис
Замороженные клетки ресуспендировали в лизирующем буфере из расчета 2 мл/г клеток. Смесь обрабатывали ультразвуком при 22 кГц пять раз по 15 секунд с интервалами 1 мин, чтобы охладить раствор на льду. Затем лизат центрифугировали в течение 20 минут при 12000 об/мин, используя ротор F34-6-38 (Ependorf). Все описанные ниже стадии центрифугирования выполняли с использованием такого же ротора, если не оговорено особо. Надосадок хранили при 4°C, тогда как обломки клеток дважды промывали лизирующим буфером. После центрифугирования надосадки лизата и промывные фракции объединяли.
Фракционирование
Затем можно использовать осаждение сульфатом аммония, чтобы очистить VLP AP205. На первой стадии выбирали концентрацию сульфата аммония, при которой VLP AP205 не осаждаются. Полученный в результате осадок отбрасывали. На следующей стадии выбирали концентрацию сульфата аммония, при которой VLP AP205 осаждается количественно, и VLP AP205 выделяли из осадка, полученного на данной стадии осаждения, центрифугированием (14000 об/мин в течение 20 мин). Полученный осадок растворяли в буфере NET.
Хроматография
Белок капсида из объединенных надосадков наносили на колонку с сефарозой 4B (2,8 X 70 см) и элюировали буфером NET при скорости 4 мл/час/фракцию. Фракции 28-40 собирали и осаждали сульфатом аммония при 60% насыщения. Перед осаждением фракции анализировали с помощью SDS-ПААГ и Вестерн-блоттинга, используя антисыворотку, специфичную по отношению AP205 (фиг.6A и фиг.6B). Осадок, выделенный центрифугированием, перерастворяли в буфере NET и наносили на колонку с сефарозой 2B (2,3 х 65 см), элюировали при скорости 3 мл/час/фракцию. Фракции анализировали в SDS-ПААГ и фракции 44-50 собирали, объединяли и осаждали сульфатом аммония при насыщении 60%. Осадок, выделенный центрифугированием, перерастворяли в буфере NET и очищали на колонке с сефарозой 6B (2,5 X 47 см), элюировали при скорости 3 мл/час/фракцию. Фракции анализировали в SDS-ПААГ. Фракции 23-27 собирали, концентрацию соли доводили до 0,5 М и осаждали ПЭГ 6000, добавляемым из 40% маточного раствора в воде и до конечной концентрации 13,3%. Осадок, выделенный центрифугированием, перерастворяли в буфере NET и наносили на такую же колонку с сефарозой 2B, как описано выше, элюировали таким же образом. Анализ фракций в SDS-ПААГ показан на фиг.6C. Фракции 43-53 собирали и осаждали сульфатом аммония при насыщении 60%. Осадок, выделенный центрифугированием, перерастворяли в воде и полученный раствор белка диализовали против большого объема воды. Можно было выделить примерно 10 мг очищенного белка на грамм клеток.
Определение вирусоподобных частиц с помощью электронной микроскопии показало, что они были идентичны фаговым частицам (фиг.7A и 7B).
На фиг.6A на верхней панели показан окрашенный серебром SDS-ПААГ, разгоняемый в восстанавливающих условиях, фракций, полученных на первой стадии хроматографии на сефарозе 4B. На дорожки 1-13 наносили каждую вторую фракцию из фракций с 20 по 44. Фракцию 50 наносили на дорожку 14. На второй гель наносили такие же фракции и анализировали Вестерн-блоттингом с использованием антисыворотки, специфичной по отношению к AP205, второй гель показан на нижней панели (фиг.6B).
На фиг.6C показан окрашенный серебром SDS-ПААГ, разгоняемый в восстанавливающих условиях, фракций с последней стадии хроматографии на сефарозе 2B. Фракции 38-54 нанесены на дорожки 1-16.
На фиг.7A показана ЭМ-картина частиц фага AP205, тогда как ЭМ-картина частиц рекомбинантной VLP AP205, образованной в ходе самосборки, показана на фиг.7B.
ПРИМЕР 19
Ингибирование RANKL-индуцированного образования остеокластов сывороткой мышей, иммунизированных Qβ-C-RANKL
Для того чтобы проверить, способны ли антитела, образованные у мышей, иммунизированных Qβ-C-RANKL, ингибировать биологическую активность RANKL, осуществляли анализ дифференцировки остеокластов in vitro. Для этого клетки костного мозга выделяли из мышей Balb/c (4-недельного возраста) и инкубировали при плотности 106/мл с рекомбинантным M-CSF мыши (5 нг/мл) в α-MEM/10% FCS в течение 16 часов. Затем неприкрепленные клетки собирали и далее культивировали с M-CSF (30 нг/мл), PGE2 (1 мкМ) и различными концентрациями C-RANKL. На 4 день образование остеокластов оценивали по количеству многоядерных клеток, позитивно окрашиваемых по резистентной к тартрату кислой фосфатазе (TRAP). Обнаружили, что C-RANKL индуцирует значительное количество TRAP-позитивных многоядерных клеток при концентрации 100-1000 нг/мл.
Затем сыворотки мышей, вакцинированных Qβ-C-RANKL, тестировали по их способности ингибировать образование остеокластов из обработанных C-RANKL клеток костного мозга. Для этого клетки-предшественники остеокластов выделяли из мышей Balb/c и инкубировали с M-CSF (30 нг/мл), PGE2 (1 мкМ), C-RANKL (1000 нг/мл) и серийными разведениями сывороток, полученных от иммунизированных мышей, в α-MEM/10% FCS в течение 4 часов. Затем образование остеокластов анализировали путем подсчета количества TRAP-позитивных многоядерных клеток и сравнения с контролями, инкубированными только в C-RANKL и с C-RANKL и сыворотками, полученными до иммунизации.
ПРИМЕР 20
Ингибирование потери кости при вакцинировании Qβ-C-RANKL в мышиной модели овариэктомии
Чтобы проверить, защищает ли вакцинация Qβ-C-RANKL от потери кости, индуцированной дефицитом эстрогена, готовили модель овариэктомии (ovx) у мышей. Для этого всего 40 мышей C57/BL6 в возрасте 12 недель случайным образом распределяли на 4 группы и обрабатывали следующим образом: группу 1 не иммунизировали и подвергали ложной операции, группу 2 не иммунизировали и подвергали ovx, группу 3 иммунизировали 25 мкг Qβ-C-RANKL без квасцов и подвергали ovx и группу 4 иммунизировали 25 мкг Qβ-C-RANKL с квасцами и также подвергали ovx. Животных из групп 3 и 4 иммунизировали в 0, 14, 21 и 42 день и всех животных оперировали (либо ложно, либо ovx) на 28 день и забивали на 63 день. Массу тела измеряли в 0, 28 и 63 дни и образцы крови собирали в 0, 14, 21, 28, 42 и 63 дни. Титры антител, а также маркеры образования кости и деградации кости контролировали непрерывно, а минеральную плотность кости оценивали после забоя животных с помощью DXA-сканирования изолированного позвоночного столба и pQCT-сканирования вырезанных бедренных костей на одном дистальном участке и одном участке средней трети кости.
Теперь на основании полного описания данного изобретения с некоторыми подробностями с использованием иллюстрации и примера в целях четкого понимания специалисту в данной области будет очевидно, что то же самое можно осуществить путем модификации или изменения изобретения в широких и эквивалентных пределах условий, композиций и других параметров, не затрагивая объем изобретения или какого-либо конкретного его варианта, и что предполагается, что такие модификации или изменения входят в объем прилагаемой формулы изобретения.
Все публикации, патенты и заявки на выдачу патентов, упоминаемые в данном описании, являются показателем уровня специалиста в данной области, к которой данное изобретение относится, и включены в виде ссылки в такой же степени, как в случае, когда отдельно указано, что каждая отдельная публикация, патент или заявка на выдачу патента включена в виде ссылки.















































































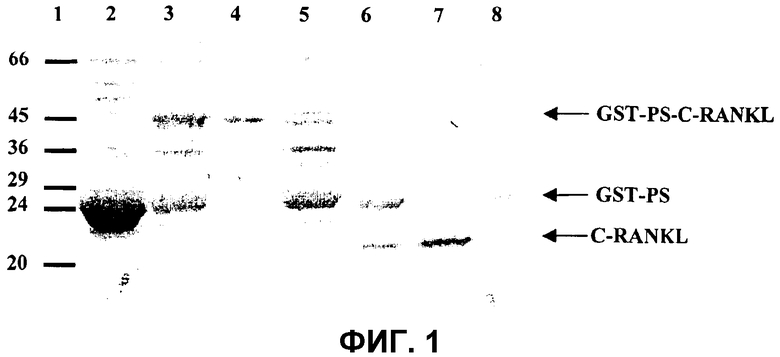
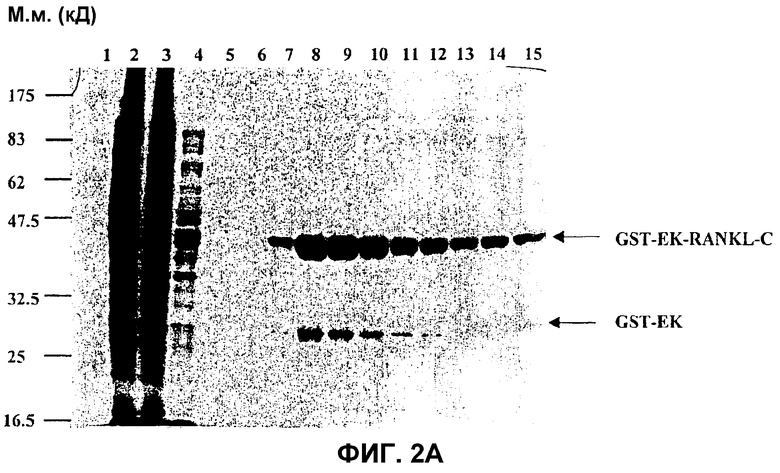
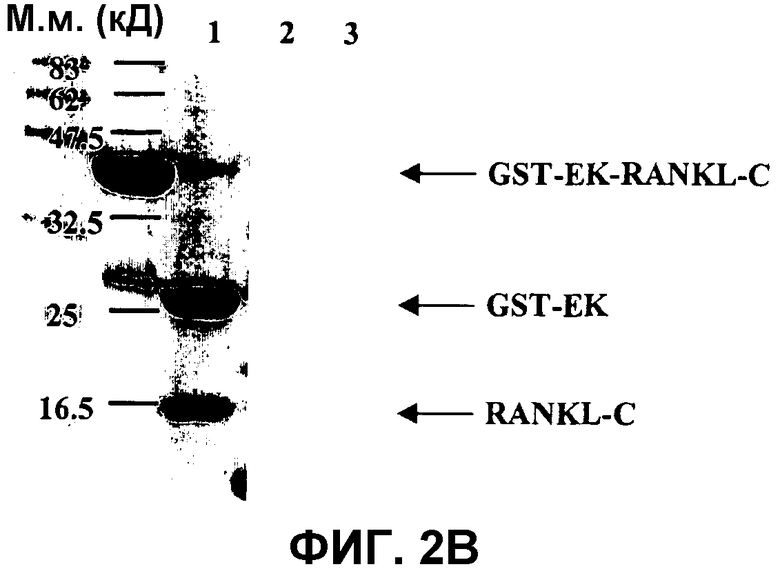
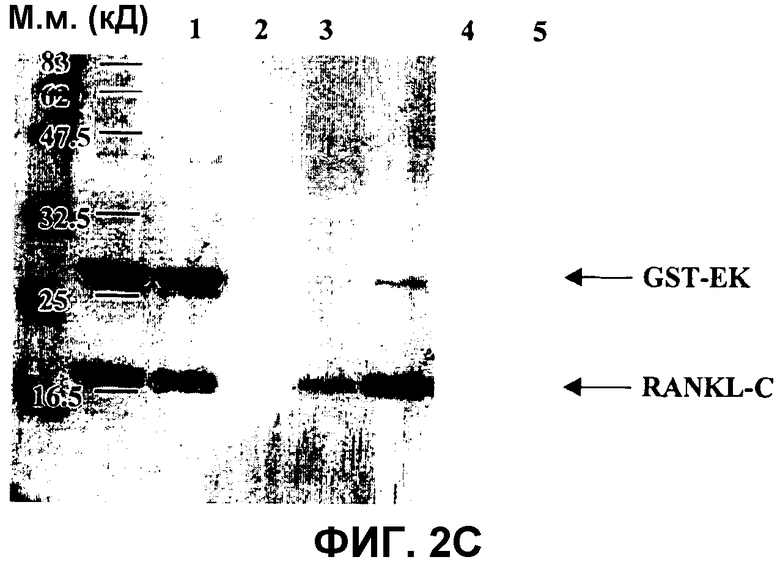

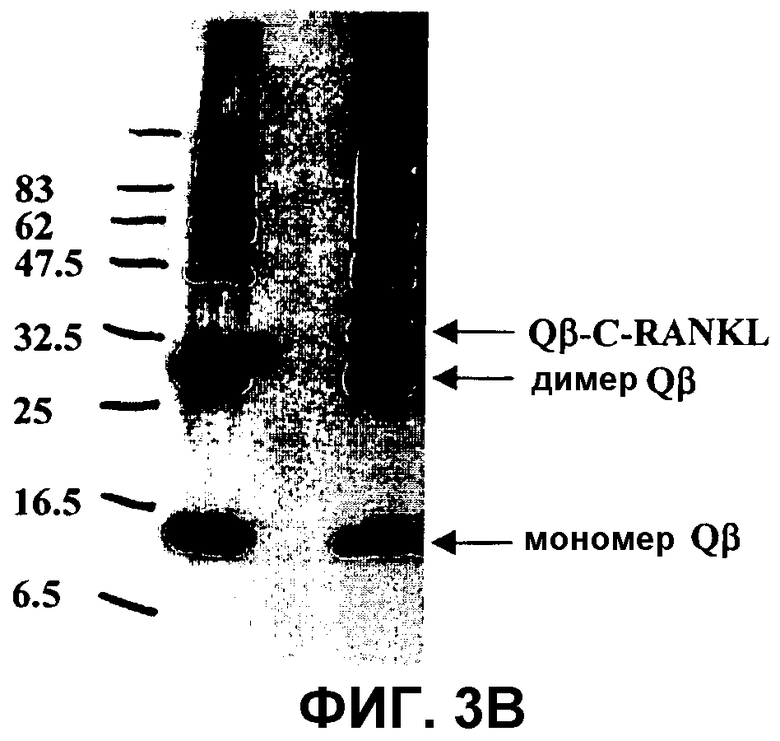

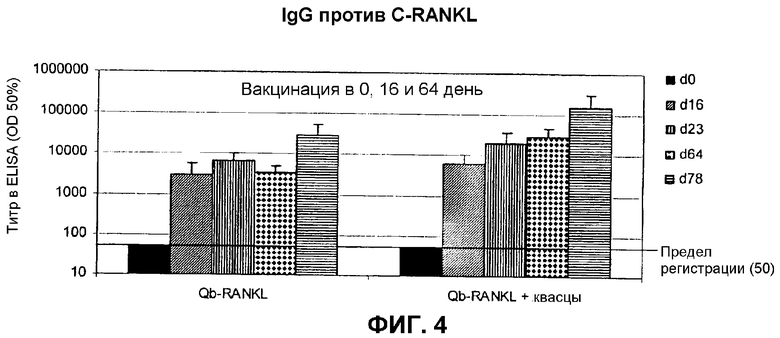
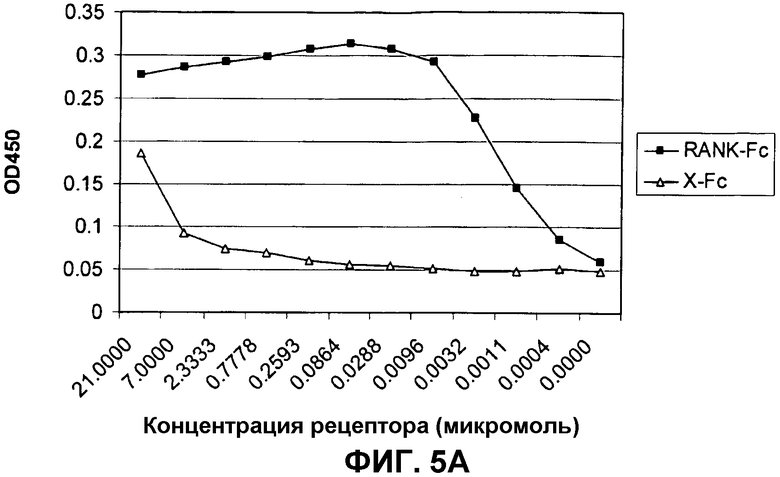



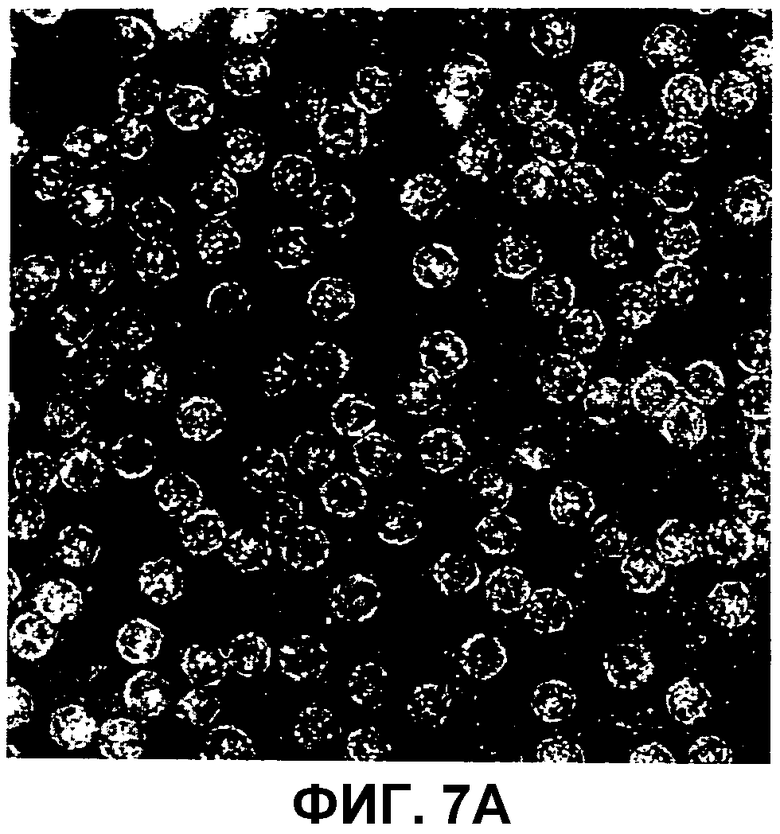
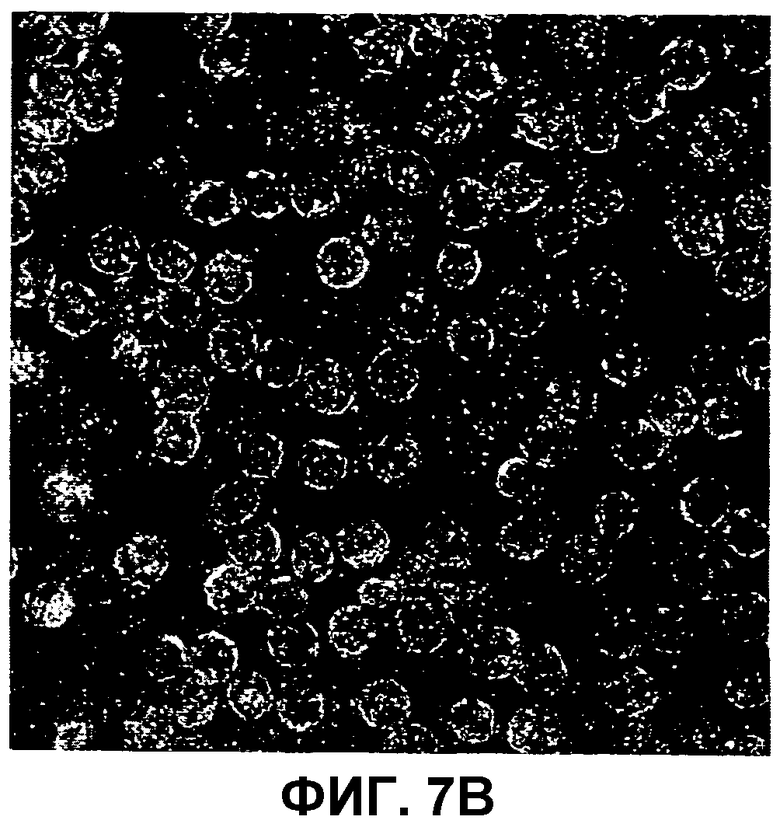
Данное изобретение относится к областям молекулярной биологии, вирусологии, иммунологии и медицины. Изобретение включает композицию, содержащую упорядоченную и повторяющуюся матрицу антигенов или антигенных детерминант и, в частности, матрицу, включающую белок RANKL, фрагмент RANKL или пептид RANKL-VLP. Более конкретно, изобретение относится к композиции, содержащей вирусоподобную частицу и по меньшей мере один связанный с ней белок RANKL, фрагмент RANKL или пептид RANKL, а также к способу получения конъюгатов и упорядоченных и повторяющихся матриц, соответственно. Композиции согласно изобретению применимы для получения вакцин для лечения заболеваний кости и в качестве фармацевтической вакцины для профилактики или лечения заболеваний кости и для эффективной индукции иммунных ответов, в частности гуморальных ответов. Преимущество изобретения заключается в повышении индукции иммунных ответов к белку RANCL. 7 н. и 21 з. п. ф-лы, 7 ил.
(a) вирусоподобную частицу РНК-фага; и
(b) по меньшей мере один антиген или антигенную детерминанту, где указанный антиген или антигенная детерминанта является белком RANKL, фрагментом RANKL или пептидом RANKL, и
где указанный по меньшей мере один антиген или антигенная детерминанта связаны с указанной вирусоподобной частицей по меньшей мере одной непептидной ковалентной связью.
(a) бактериофага Qβ;
(b) бактериофага R17;
(c) бактериофага fr;
(d) бактериофага GA;
(e) бактериофага SP;
(f) бактериофага MS2;
(g) бактериофага M11;
(h) бактериофага МХ1;
(i) бактериофага NL95;
(j) бактериофага f2;
(k) бактериофага РР7; и
(1) бактериофага АР205.
(a) аминокислотной последовательности SEQ ID NO: 79;
(b) аминокислотной последовательности SEQ ID NO: 80;
(c) аминокислотной последовательности SEQ ID NO: 81;
(d) аминокислотной последовательности SEQ ID NO: 82;
(e) аминокислотной последовательности SEQ ID NO: 83;
(f) аминокислотной последовательности SEQ ID NO: 84;
(g) аминокислотной последовательности SEQ ID NO: 100;
(h) аминокислотной последовательности SEQ ID NO: 101;
(i) аминокислотной последовательности фрагмента любой из последовательностей SEQ ID NO: 79-84, 100, 101.
(a) аминокислотной последовательности SEQ ID NO: 87;
(b) аминокислотной последовательности SEQ ID NO: 88;
(c) аминокислотной последовательности SEQ ID NO: 89;
(d) аминокислотной последовательности SEQ ID NO: 90;
(e) аминокислотной последовательности SEQ ID NO: 91;
(f) аминокислотной последовательности SEQ ID NO: 92;
(g) аминокислотной последовательности SEQ ID NO: 93;
(h) аминокислотной последовательности фрагмента любой из последовательностей SEQ ID NO: 87-93.
(i) сайта связывания неприродного происхождения с указанным антигеном или антигенной детерминантой; и
(ii) сайта связывания природного происхождения с указанным антигеном или антигенной детерминантой;
и где указанный второй сайт связывания способен к ассоциации с указанным первым сайтом связывания; и где дополнительно указанный антиген или антигенная детерминанта и указанная вирусоподобная частица взаимодействуют посредством указанной ассоциации, образуя упорядоченную и повторяющуюся антигенную матрицу.
(a) аминокислотной последовательности SEQ ID NO: 104;
(b) аминокислотной последовательности SEQ ID NO: 105;
(c) аминокислотной последовательности SEQ ID NO: 106;
(d) аминокислотной последовательности SEQ ID NO: 107;
(e) аминокислотной последовательности SEQ ID NO: 108;
(f) аминокислотной последовательности SEQ ID NO: 109;
(g) аминокислотной последовательности SEQ ID NO: 110;
(h) аминокислотной последовательности фрагмента любой из последовательностей SEQ ID NO: 104-110.
(a) композицию по п.1; и
(b) приемлемый фармацевтический носитель.
(a) вирусоподобную частицу РНК-фага; и
(b) по меньшей мере один антиген или антигенную детерминанту, где указанный антиген или указанная антигенная детерминанта является белком RANKL, фрагментом RANKL или пептидом RANKL, и
где указанный по меньшей мере один антиген или антигенная детерминанта связаны с указанной вирусоподобной частицей по меньшей мере одной непептидной ковалентной связью.
(a) получение вирусоподобной частицы РНК-фага; и
(b) получение по меньшей мере одного антигена или антигенной детерминанты, где указанный антиген или антигенная детерминанта является белком RANKL, фрагментом RANKL или пептидом RANKL;
(c) объединение указанной вирусоподобной частицы и указанного по меньшей мере одного антигена или антигенной детерминанты, так, чтобы указанный по меньшей мере один антиген или антигенная детерминанта были связаны с указанной вирусоподобной частицей по меньшей мере одной непептидной ковалентной связью.
Приоритеты:
АР205, п.9 в отношении признаков SEQ ID No: 79, 80, 83, 84, 100 и 101, пп.16, 17 и п.20 в отношении признаков РНК фагов АР205 и Fr.
| WO 00/23955 A, 27.04.2000 | |||
| KOSLOVSKA Т | |||
| М | |||
| et al., Recombinant RNA phage Q beta capsid particles synthesized and self-assembled in Esherishia coli, Gene, 1993, Dec | |||
| Прибор с двумя призмами | 1917 |
|
SU27A1 |
| KOSLOVSKA T | |||
| M | |||
| et al., RNA phage Q beta coat protein as a carrier for foreign epitopes, Intervirology, 1996, 39 (1-2), pp.9-15 | |||
| HERTZ M | |||
| et al., A therapeutic | |||

