ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Область изобретения
Настоящее изобретение относится к областям молекулярной биологии, вирусологии, иммунологии и медицины. Изобретение относится к композиции, содержащей массив упорядоченных и повторяющихся антигенов или антигенных детерминант, а в частности массив грелина или полученных из грелина пептидов. Более конкретно изобретение относится к композиции, содержащей вирусоподобную частицу и связанный с ней, по меньшей мере, один грелин или полученный из грелина пептид.
Изобретение также относится к способу получения конъюгатов и упорядоченных и повторяющихся массивов соответственно. Композиции по изобретению пригодны для получения вакцин для лечения ожирения и других заболеваний, ассоциированных с увеличенным потреблением пищи или увеличенной массой тела, и для эффективной индукции иммунных ответов, в частности ответов антителами. Кроме того, композиции по изобретению особенно пригодны для эффективной индукции аутоспецифичного иммунного ответа в рамках указанного контекста.
Связанная область
Ожирение представляет собой заболевание, поражающее миллионы людей по всему миру. Хотя лежащие в основе ожирения причины могут являться различными, общей причиной почти для всех форм ожирения является повышенное потребление пищи. Чувство голода и пищевое поведение регулируют многие факторы, включающие в себя лептин, гормон роста, нейропептид Y (NPY), родственный агути-белок (AGRP) и другие. Недавно идентифицированным ключевым регулятором пищевого поведения является грелин, ацилированный пептид, продуцируемый в желудке, а также в некоторых отделах головного мозга (гипоталамус) (Kojima et al. Nature 402: 656-660 (1999)). Грелин образуется при ферментативном расщеплении из формы предшественника пробелка, состоящей из 117 аминокислот, с образованием пептида из 28 аминокислот с н-октаноилированием по серину 3. н-Октаноилирование по данному положению необходимо для биологической активности грелина. Идентифицирована вторая изоформа грелина из 27 а.к. (грелин-desQ14) с отсутствующим в положении 14 глутамином (Q), однако данная изоформа представляет собой только незначительную составляющую циркулирующего грелина. Подобно полноразмерному грелину биологическая активность грелина-desQ14 зависит от н-октаноильной группы на серине 3. Однако наибольшая функциональная активность достигается за счет изоформы грелина из 28 а.к. (Hosoda et al. Biochem. Biophys. Res. Commun. 279 (3): 909-913 (2000)). Грелин высококонсервативен, так как человеческий и крысиный грелин отличаются только на 2 аминокислоты (GSSFLSPEHQRVQQRKESKKPPAKLQPR (SEQ ID № 31) в сравнении с GSSFLSPEHQKAQQRKESKKPPAKLQPR (SEQ ID № 32)) (Kojima et al. Nature 402: 656-660 (1999)).
Рецепторы для грелина (GHS-R) экспрессированы в различных областях головного мозга, включающих в себя дугообразное ядро (Arc) и вентромедиальное ядро гипоталамуса и гипофиза (Howard etal. Science 273: 974-977 (1996)); McKee et al. Mol Endokrin. 11: 415-423 (1997); Guan et al., Mol Brain Research 48: 23-29 (1997)), указывая на то, что грелин преимущественно действует в головном мозге. В дополнение к стимуляции грелином высвобождения GH из гипофиза (Kojima et al. Nature 402: 656-660 (1999)) недавно его идентифицировали как ключевой центральный регулятор питания (Nakazato et al., Nature 409: 194-198 (2001)). Конкретно показано, что после интрацеребровентрикулярного введения грелин стимулирует потребление пищи. Кроме того, интрацеребровентрикулярное введение антител против грелина подавляет потребление пищи. Инъекция грелина индуцировала повышенное высвобождение NPY, а антитела против NPY совместно с антагонистами AGRP блокировали индуцированное грелином потребление пищи, указывая на то, что грелин модулирует потребление пищи посредством усиления экспрессии NPY и AGRP (Nakazato et al., Nature 409: 194-198 (2001)). Кроме того, периферийное ежедневное введение грелина индуцировало увеличение массы тела мышей и крыс, а сывороточные концентрации грелина увеличивались у голодающих крыс и уменьшались при кормлении, дополнительно указывая на то, что грелин играет ключевую роль в регуляции потребления пищи (Tschop et al, Nature 407: 908-912). Для трансгенных крыс, экспрессирующих в Arc антисмысловую к GHS-R РНК, показана меньшая масса тела и меньшее количество адипозной ткани, что поддерживает точку зрения о том, что грелин регулирует массу тела (Shuto et al., JCI 109: 14291436 (2002)). Также существуют доказательства того, что грелин играет ключевую роль в пищевом поведении людей. Периферическое введение грелина людям усиливало аппетит и увеличивало потребление пищи у людей (Wren et al., J Clin Endocrinol Metab 86: 5992-5998 (2001)). У людей с синдромом Прадера-Вилли с наиболее общей формой человеческого синдромного ожирения выявляют чрезвычайно высокие уровни грелина (Cummings et al. Nat Med 8: 643-644 (2002)). Кроме того, у людей уровни грелина в плазме сильно увеличены после вызванной диетой потери массы, коррелируя с быстрым восстановлением массы после прекращения людьми диеты. Напротив, у пациентов с операцией обходного желудочного анастомоза уровни грелина оставались низкими и после диеты, и у пациентов в данных условиях их масса обычно не восстанавливалась (Cummings et al., N Engl J Med 21: 1623-1630 (2002)). Следовательно, по-видимому, грелин представляет собой ключевой регулятор потребления пищи и массы тела у людей.
Так как периферическое введение грелина могло увеличивать потребление пищи, ведя к увеличению массы тела (Tschop et al, Nature 407: 908-912), вероятно, что грелин, продуцируемый в желудке, достигает головного мозга через кровоток и переключает пищевое поведение. Таким образом, возможно, что блокирование поступления грелина из крови в головной мозг остановит потребление пищи у животных и людей. Так как показано, что специфические антитела могут блокировать действие грелина в головном мозге (Nakazato et al., Nature 409: 194-198 (2001)), вероятно, что периферические антитела также будут способны блокировать действие периферического грелина. Кроме того, так как антитела неэффективно проходят через гематоэнцефалический барьер, специфичные к грелину антитела, вероятно, смогут изолировать грелин от головного мозга, но не будут действовать на грелин в головном мозге. Это могло бы являться особенно притягательной возможностью, так как грелин также продуцируется в головном мозге, где он, вероятно, выполняет отличные от регулирования потребления пищи функции (Nakazato et al., Nature 409: 194-198 (2001)). Следовательно, потенциальной терапией ожирения могла бы являться индукция специфичных к грелину антител у хозяина, ведущая к долговременной блокаде грелина, приводящей к уменьшению потребления пищи, подобно той, которую наблюдают у пациентов с обходным желудочным анастомозом.
В WО98/42840 описано влияние грелина и полученных из грелина фрагментов на желудочно-кишечный тракт и, таким образом, в частности, их эффекты на перистальтику желудка и опорожнение желудка. Кроме того, в US 6420521 описано применение коротких пептидов грелина для воздействия на функцию желудка, включая опорожнение желудка, сократительную способность желудка и всасывание глюкозы.
Однако, как правило, трудно индуцировать антитело, отвечающее против пептидов, в особенности против собственных пептидов. Один из способов увеличить эффективность вакцинации представляет собой увеличение степени повторяемости применяемого антигена. В отличие от отдельных белков вирусы индуцируют быстрые и эффективные иммунные ответы в отсутствие каких-либо адъювантов и при помощи и без помощи T-клеток (Bachmann and Zinkernagel, Ann. Rev. Immunol: 15: 235-270 (1991)). Хотя вирусы часто состоят из немногих белков, они способны инициировать значительно более сильные иммунные ответы, чем их отдельные компоненты. Для B-клеточных ответов известно, что одним из ключевых факторов для иммуногенности вирусов является повторяемость и порядок поверхностных эпитопов. Многие вирусы обладают квазикристаллической поверхностью, обнаруживающей регулярный набор эпитопов, эффективно связывающих специфичные к эпитопам иммуноглобулины на B-клетках (Bachmann and Zinkernagel, Immunol. Today 17: 553-558 (1996)). Данное связывание поверхностных иммуноглобулинов на B-клетках представляет собой сильный активационный сигнал, прямо индуцирующий продвижение по клеточному циклу и продукцию антител IgM. Далее, такие инициированные B-клетки способны активировать хелперные T-клетки, в свою очередь индуцирующие в B-клетках переключение продукции антител с IgM на IgG и образование долгоживущих B-клеток памяти - цель любой вакцинации (Bachmann and Zinkernagel, Ann. Rev. Immunol. 15: 235-270 (1997)). Структура вируса даже связана с образованием анти-антител при аутоиммунном заболевании и как часть естественного ответа на патогены (см. Fehr, T., et al., J Exp. Med. 185: 1785-1792 (1997)). Таким образом, антитела, представляемые высокоорганизованной вирусной поверхностью, способны индуцировать сильные ответы против антител.
Однако, как указано, иммунная система, как правило, не может образовывать антитела против собственных структур. Для растворимых антигенов, представленных в низких концентрациях, это происходит вследствие толерантности на уровни Th-клеток. В данных условиях связывание собственного антигена с носителем, который способен обеспечивать помощь T-клеток, может нарушить толерантность. Для растворимых белков, присутствующих в высоких концентрациях, или мембранных белков в низких концентрациях B- и Th-клетки могут являться толерантными. Однако толерантность B-клеток может являться обратимой (анэргия), и ее можно разрушить введением антигена в высокоорганизованной форме, связанным с чужеродным носителем (Bachmann and Zinkernagel, Ann. Rev. Immunol. 15: 235-270 (1997)).
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Авторы обнаружили, что грелин или пептиды грелина, связанные с коровой частицей, обладающей структурой с собственной повторяющейся организацией и, в частности, для данного случая с вирусоподобными частицами (VLP) и субъединицами VLP соответственно приводят к высокоорганизованным и повторяющимся конъюгатам, представляющим собой сильные иммуногены для индукции специфических антител. Настоящее изобретение относится к профилактическому и терапевтическому средству для лечения ожирения и связанных заболеваний, основанному на упорядоченном и повторяющемся массиве коровых частиц с грелином или полученными из грелина пептидами и, в частности, на конъюгате и массиве VLP-грелин/пептид грелина соответственно. Данная профилактическая и терапевтическая композиция способна индуцировать высокие титры антител против грелина у вакцинированного животного или человека. Следовательно, настоящее изобретение относится к грелину и его имеющим отношение к головному мозгу свойствам. Кроме того, настоящее изобретение относится к центральным эффектам грелина на головной мозг, что более важно, к регуляции аппетита, секреции гормона роста и энергетическому гомеостазу. Следовательно, предпочтительно, чтобы антитела, индуцированные посредством стратегии вакцинации авторов настоящего изобретения, являлись способными связываться с н-октаноилированной формой(ами) грелина. Как указано, грелин или более короткие пептидные фрагменты грелина можно применять, когда они связаны с коровой частицей, и альтернативно или предпочтительно вводить в адъюванте для индукции специфичных к грелину антител у людей или животных.
Следовательно, короткие пептидные фрагменты грелина, особенно более короткие пептиды из остатков 24-33, 42-51, 31-41 и 28-37, связанные или C- или N-концом с коровой частицей или вирусоподобной частицей соответственно, способны индуцировать высокоспецифичные антитела против грелина, способные нейтрализовать периферически циркулирующий грелин до его проникновения в ЦНС и воздействия на гормон роста и, следовательно, на потребление пищи.
Таким образом, в предпочтительном осуществлении настоящего изобретения антиген или антигенную детерминанту выбирают из группы пептидов грелина, соответствующих остаткам 24-33, 42-51, 31-41 и 28-37 любой из указанных последовательностей с SEQ ID № 144 по 146, где указанные предпочтительные пептидные фрагменты грелина выбирают из группы, состоящей из (a) человеческого грелина; (b) коровьего грелина; (c) овечьего грелина; (d) собачьего грелина; (e) кошачьего грелина; (f) мышиного грелина; (g) свиного грелина; и (h) лошадиного грелина.
Более конкретно в настоящем изобретении авторы смогли индуцировать высокие уровни антител, неожиданно распознающих н-октаноилированную форму грелина, как показано здесь, а в частности, в примере 17. Кроме того, полученные антитела также распознавали альтернативную изоформу грелин-desQ14. Как результат, антитела, полученные при вакцинации грелином и пептидом грелина соответственно, связанными C- или N-концом с коровой частицей или предпочтительно с VLP, являлись способными блокировать проникновение н-октаноилированного грелина в головной мозг и модулировали потребление пищи у мышей. Следовательно, настоящее изобретение относится к стратегиям вакцинации против активного грелина, как лечению от ожирения и других связанных заболеваний.
Как показано здесь, а в частности в примере 18, вакцинация грелином и пептидом грелина соответственно, связанными C- или N-концом с коровой частицей или предпочтительно с VLP ведет к уменьшению потребления пиши у мышей, что позволяет предположить, что грелин и пептид грелина соответственно представляют собой ключевой регулятор потребления пищи, и полагают, что антитела, направленные соответственно к грелину и пептиду грелина, представляют собой возможное терапевтическое средство против ожирения и других связанных заболеваний.
Таким образом, настоящее изобретение относится к композиции содержащей: (a) коровую частицу, по меньшей мере, с одним первым участком прикрепления и (b) по меньшей мере, один антиген или антигенную детерминанту, по меньшей мере, с одним вторым участком прикрепления, где указанный антиген или антигенная детерминанта представляет собой грелин или полученный из грелина пептид и где указанный второй участок прикрепления выбран из группы, состоящей из (i) участка прикрепления, не встречающегося в природе с указанным антигеном или антигенной детерминантой, и (ii) участка прикрепления, встречающегося в природе с указанным антигеном или антигенной детерминантой, где указанный второй участок прикрепления способен к ассоциации с указанным первым участком прикрепления и где указанный антиген или антигенная детерминанта и указанная коровая частица взаимодействуют посредством указанной ассоциации для формирования упорядоченного и повторяющегося массива антигенов. Предпочтительные осуществления коровых частиц, пригодных для применения по настоящему изобретению, представляют собой вирусы, вирусоподобные частицы, бактериофаг, вирусоподобную частицу РНК-фага, бактериальный пиль или жгутик или любую другую коровую частицу с собственной повторяющейся структурой, способной к формированию упорядоченного и повторяющегося массива антигенов по настоящему изобретению.
Более конкретно изобретение относится к композиции, содержащей упорядоченный и повторяющийся массив антигенов или антигенных детерминант и, в частности для данного случая, конъюгаты грелина или полученных из грелина пептидов с VLP. Более конкретно изобретение относится к композиции, содержащей вирусоподобную частицу и, по меньшей мере, один связанный с ней грелин или полученный из грелина пептид. Также изобретение относится к способу получения конъюгатов и упорядоченных и повторяющихся массивов соответственно. Композиции по изобретению пригодны для получения вакцин для лечения ожирения и связанных заболеваний и в качестве фармакцины для профилактики или лечения ожирения и связанных заболеваний и для эффективной индукции иммунных ответов, в особенности ответов антителами. Кроме того, композиции по изобретению особенно пригодны для эффективной индукции аутоспецифичных иммунных ответов в указанном контексте.
В настоящем изобретении грелин или полученный из грелина пептид связан с коровой частицей и VLP соответственно, как правило, ориентированным образом с получением упорядоченного и повторяющегося массива антигенов грелина или полученного из грелина пептида. Кроме того, высокоповторяющаяся и организованная структура коровых частиц и VLP соответственно опосредует представление грелина или полученного из грелина пептида в высокоупорядоченном и повторяющемся виде, приводя к высокоорганизованному и повторяющемуся массиву антигенов. Кроме того, связывание грелина или полученного из грелина пептида с коровой частицей и VLP соответственно обеспечивает эпитопы хелперных T-клеток, так как коровая частица и VLP являются чужеродными для хозяина, иммунизированного массивом коровая частица-грелин или полученный из грелина пептид и массивом VLP-грелин или полученный из грелина пептид соответственно. Данные массивы отличаются от ранее известных в данной области конъюгатов, в частности их высокоорганизованной структурой, размерами и повторяемостью антигена на поверхности массива.
В одном аспекте изобретения грелин или полученный из грелина пептид экспрессируют в подходящем экспрессирующем хозяине или синтезируют, тогда как коровую частицу и VLP соответственно экспрессируют и очищают из экспрессирующего хозяина, подходящего для укладки и сборки коровой частицы и VLP соответственно. Грелин или полученные из грелина пептиды можно синтезировать химически. Так как биологически активный грелин включает в себя н-октаноилированный серин в третьем положении, химический синтез представляет собой предпочтительный путь получения грелина для содержащего биологически активные формы грелина вакцинного препарата. Затем массив грелина или полученного из грелина пептида собирают посредством связывания грелина или полученного из грелина пептида с коровой частицей или VLP соответственно.
В другом аспекте настоящее изобретение относится к композиции, содержащей (a) вирусоподобную частицу и (b) по меньшей мере, один антиген или антигенную детерминанту, где указанный антиген или указанная антигенная детерминанта представляет собой грелин или полученный из грелина пептид и где, по меньшей мере, один указанный антиген или антигенная детерминанта связаны с указанной вирусоподобной частицей.
В дополнительном аспекте изобретение относится к фармацевтической композиции, содержащей (a) композицию по настоящему изобретению и (b) приемлемый фармацевтический носитель.
В еще одном дополнительном аспекте настоящее изобретение относится к вакцинной композиции, содержащей композицию, содержащую (a) коровую частицу, по меньшей мере, с одним первым участком прикрепления и (b) по меньшей мере, один антиген или антигенную детерминанту, по меньшей мере, с одним вторым участком прикрепления, где указанный антиген или антигенная детерминанта представляет собой грелин или полученный из грелина пептид и где указанный второй участок прикрепления выбран из группы, состоящей из (i) участка прикрепления, не встречающегося в природе с указанным антигеном или антигенной детерминантой, и (ii) участка прикрепления, встречающегося в природе с указанным антигеном или антигенной детерминантой, где указанный второй участок прикрепления способен к ассоциации с указанным первым участком прикрепления и где указанный антиген или антигенная детерминанта и указанная коровая частица взаимодействуют посредством указанной ассоциации для формирования упорядоченного и повторяющегося массива антигенов.
В дополнительном аспекте настоящее изобретение относится к вакцинной композиции, содержащей композицию, где указанная композиция содержит (a) вирусоподобную частицу и (b) по меньшей мере, один антиген или антигенную детерминанту, где указанный антиген или указанная антигенная детерминанта представляет собой грелин или полученный из грелина пептид и где, по меньшей мере, один указанный антиген или антигенная детерминанта связаны с указанной вирусоподобной частицей.
В еще одном дополнительном аспекте настоящее изобретение относится к способу получения композиции по п. 1 формулы изобретения, включающему в себя: (a) получение вирусоподобной частицы и (b) получение, по меньшей мере, одного антигена и антигенной детерминанты, где указанный антиген или указанная антигенная детерминанта представляет собой грелин или полученный из грелина пептид; (c) объединение указанной вирусоподобной частицы и, по меньшей мере, одного указанного антигена или антигенной детерминанты так, чтобы, по меньшей мере, один указанный антиген или антигенная детерминанта являлись связанными с указанной вирусоподобной частицей.
Аналогично настоящее изобретение относится к процессу для получения композиции по изобретению, включающему в себя: (a) получение коровой частицы, по меньшей мере, с одним первым участком прикрепления; (b) получение, по меньшей мере, одного антигена или антигенной детерминанты, по меньшей мере, с одним вторым участком прикрепления, где указанный антиген или антигенная детерминанта представляет собой грелин или полученный из грелина пептид, и где указанный второй участок прикрепления выбран из группы, состоящей из (i) участка прикрепления, не встречающегося в природе с указанным антигеном или антигенной детерминантой, и (ii) участка прикрепления, встречающегося в природе с указанным антигеном или антигенной детерминантой, и где указанный второй участок прикрепления способен к ассоциации с указанным первым участком прикрепления; и (c) объединение указанной коровой частицы и, по меньшей мере, одного указанного антигена или антигенной детерминанты, где указанный антиген или антигенная детерминанта и указанная коровая частица взаимодействуют посредством указанной ассоциации для формирования упорядоченного и повторяющегося массива антигенов.
В другом аспекте настоящее изобретение относится к способу иммунизации, включающему в себя введение композиции по п. 1 формулы изобретения животному или человеку.
В дополнительном аспекте настоящее изобретение относится к применению композиции по п. 1 формулы изобретения для получения лекарственного средства для лечения ожирения или связанного заболевания.
В еще одном дополнительном аспекте настоящее изобретение относится к применению композиции по п. 1 формулы изобретения для получения лекарственного средства для терапевтического или профилактического лечения ожирения или связанного заболевания. Кроме того, в еще одном дополнительном аспекте настоящее изобретение относится к применению композиции по п. 1 формулы изобретения или отдельно, или в сочетании с другими средствами для получения композиции, вакцины или лекарственного средства для лечения или профилактики ожирения или связанного заболевания и/или для стимуляции иммунной системы млекопитающего.
Следовательно, изобретение, в частности, относится к вакцинным композициям, подходящим для профилактики и/или уменьшения или лечения ожирения или связанных с ним состояний. Изобретение дополнительно относится к способам иммунизации и вакцинации соответственно для предотвращения и/или уменьшения или лечения ожирения или связанных с ним состояний у животных и особенно у домашних животных, таких как кошки или собаки, а также у людей. Композиции по изобретению можно применять профилактически или терапевтически.
В конкретных осуществлениях изобретение относится к способам профилактики, лечения и/или уменьшения ожирения или связанных с ним состояний, вызываемых или усиливаемых "собственными" генными продуктами, т.е. "аутоантигенами", как применяют здесь. В связанных осуществлениях изобретение относится к способам индукции иммунологических ответов у животных и индивидуумов соответственно, ведущих к образованию антител, что предотвращает, лечит и/или уменьшает ожирение или связанные с ним состояния, вызываемые или усиливаемые "собственными" генными продуктами.
Как может понять специалист в данной области, когда композиции по изобретению вводят животному или человеку, они могут находиться в композиции, содержащей соли, буферы, адъюванты или другие вещества, желательные для увеличения эффективности композиции. Примеры пригодных для применения в получении фармацевтических композиций веществ предоставлены в многочисленных источниках, включающих в себя Remington's Pharmaceutical Sciences (Osol, A, ed., Mack Publishing Co. (1990)).
Указано, что композиции по изобретению являются "фармакологически приемлемыми" если получающий их индивид может переносить их введение. Кроме того, композиции по изобретению вводят в "терапевтически эффективном количестве" (т.е. в количестве, вызывающем желательный физиологический эффект).
Композиции по настоящему изобретению можно вводить различными известными в данной области способами, но обычно их вводят посредством инъекции, инфузии, ингаляции, орального введения или других физических способов. Композиции альтернативно можно вводить внутримышечно, внутривенно или подкожно. Компоненты композиций для введения включают в себя стерильные водные (например, физиологический раствор) или неводные растворы и суспензии. Примеры неводных растворителей представляют собой пропиленгликоль, полиэтиленгликоль, растительные масла, такие как оливковое масло, и инъецируемые органические сложные эфиры, такие как этилолеат. Для увеличения проницаемости через кожу и увеличения всасывания антигена можно применять носители или непроницаемое покрытие.
Специалисту в данной области в свете того, что известно в данной области, следующего ниже описания изобретения и формулы изобретения станут очевидны другие осуществления настоящего изобретения.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ/ФИГУР
На фигуре 1 представлены продукты связывания реакции мышиного C-грелина (SEQ ID: № 77) или мышиного грелина-GC (SEQ ID: № 105) с капсидным белком Qβ. Полоса M представляет собой маркер, полоса 1 представляет собой дериватизированный VLP Qβ, полоса 2 представляет собой Qβ-C-грелин в растворимой фракции, полоса 3 представляет собой Qβ-C-грелин в нерастворимой фракции, полоса 4 представляет собой грелин-GC-Qβ в растворимой фракции, полоса 5 представляет собой грелин-GC-Qβ в нерастворимой фракции. В нерастворимой фракции находится очень мало продукта.
На фигуре 2 представлены средние титры специфичных к грелину антител IgG, обнаруженные в общей сыворотке мышей, иммунизированных мышиным Qβ-C-грелином (Qb-cGhr), мышиным грелином-GC-Qβ (Qb-GhrC) или PBS на 0, 14, 21 и 42 сутки. Титры ELISA представлены как разведения сыворотки, приводящие к половине максимальной OD при анализе ELISA. Планшеты для ELISA покрывали мышиным грелином с октаноилированным серином (Bachem, Product № H-4862) в концентрации 20 мкг/мл. Планшеты блокировали, а затем инкубировали с серийно разведенными мышиными сыворотками от 14, 21 и 42 суток. Связавшиеся антитела выявляли с применением ферментативно меченного антитела против мышиных IgG. В качестве контроля также тестировали сыворотку от тех же мышей перед иммунизацией.
На фигуре 3 представлено общее потребление пищи мышами, иммунизированными мышиным Qβ-C-грелином (Qb-cGhr), мышиным грелином-GC-Qβ (Qb-GhrC) или PBS. В качестве контроля мышей иммунизировали PBS. Всех мышей от 0 до 14 суток содержали на нормальной диете. Впоследствии всех мышей содержали на диете с высоким содержанием жира (45%) для облегчения развития вызванного диетой ожирения. Пищу и воду давали без ограничений. Через регулярные интервалы после иммунизации (на 5, 11, 14, 21, 28, 35, 40, 49 и 55 сутки после иммунизации) у каждой мыши контролировали изменения массы тела, а также контролировали расход пищи и воды на клетку (т.е. группу).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Если не определено иначе, все применяемые здесь технические и научные термины обладают теми же значениями, как и обычно понимаемые специалистом в данной области, к которой принадлежит данное изобретение. Хотя в практическом осуществлении или для тестирования настоящего изобретения можно применять любые способы и вещества, подобные или эквивалентные тем, которые описаны здесь, ниже описаны предпочтительные способы и вещества.
1. Определения:
Адъювант: Как применяют здесь, термин "адъювант" относится к неспецифическим стимуляторам иммунного ответа или веществам, позволяющим образование у хозяина депо, которое в сочетании с вакциной или фармацевтической композицией по настоящему изобретению соответственно может обеспечить даже более сильный иммунный ответ. Можно применять множество адъювантов. Примеры включают в себя полный и неполный адъювант Фрейнда, гидроксид алюминия и модифицированный мурамилдипептид. Дополнительными адъювантами являются минеральные гели, такие как гидроксид алюминия, поверхностно-активные вещества, такие как лизолецитин, плюрониловые полиолы, полианионы, пептиды, масляные эмульсии, гемоцианины морского блюдца, динитрофенол и потенциально пригодные человеческие адъюванты, такие как BCG (бацилла Кальмета-Герена) и Corynebacterium parvum. Такие адъюванты также хорошо известны в данной области. Дополнительные адъюванты, которые можно вводить с композициями по настоящему изобретению, включают в себя в качестве неограничивающих примеров монофосфорильный липидный иммуномодулятор, AdjuVax 100a, QS-21, QS-18, CRL1005, соли алюминия (квасцы), MF-59, OM-174, OM-197, OM-294 и виросомальную адъювантную технологию. Адъюванты также могут содержать смесь данных веществ.
В данной области известны иммунологически активные сапониновые фракции, полученные из обладающей активностью адъюванта, коры южноамериканского дерева Quillaja Saponaria Molina. Например, QS21, также известный как QA21, представляет собой очищенную при помощи ВЭЖХ фракцию из дерева Quillaja Saponaria Molina, а способ его получения (как QA21) описан в патенте США № 5057540. Также как адъюванты у Scott et al, Int. Archs. Allergy Appl. Immun., 1985, 77, 409 описаны сапонины килайи. В данной области известен монофосфориллипид A и его производные. Предпочтительное производное представляет собой 3-де-O-ацилированный монофосфориллипид A, и оно известно из патента Великобритании № 2220211. Дополнительные предпочтительные адъюванты описаны в WO00/00462, описание которого включено сюда в качестве ссылки.
Однако выгодной особенностью настоящего изобретения является высокая иммуногенность композиций по изобретению. Как уже указывали здесь и станет очевидным по ходу данной спецификации, в дополнительных альтернативных или предпочтительных осуществлениях предоставлены вакцины и фармацевтические композиции без адъювантов, что приводит к вакцинам и фармацевтическим композициям для лечения AD без адъювантов и, таким образом, обладающих более высоким профилем безопасности, так как адъюванты могут вызывать побочные эффекты. Как применяют здесь в контексте вакцин и фармацевтических композиций для лечения AD, термин "без" относится к вакцинам и фармацевтическим композициям, применяемым без адъювантов.
Аминокислотный линкер: Как применяют здесь, "аминокислотный линкер", или также просто обозначаемый в пределах данной спецификации как "линкер", или ассоциирует антиген или антигенную детерминанту со вторым участком прикрепления или более предпочтительно уже включает в себя или содержит второй участок прикрепления, как правило, но не обязательно, в виде одного аминокислотного остатка, предпочтительно в виде цистеинового остатка. Однако, как применяют здесь, термин "аминокислотный линкер" не подразумевает, что такой аминокислотный линкер состоит исключительно из аминокислотных остатков, даже если аминокислотный линкер, состоящий из аминокислотных остатков, представляет собой предпочтительное осуществление настоящего изобретения. Аминокислотные остатки аминокислотного линкера предпочтительно состоят из встречающихся в природе аминокислот или неприродных аминокислот, известных в данной области, всех L или всех D или их смесей. Однако изобретение также включает в себя аминокислотный линкер, содержащий молекулу с сульфгидрильной группой или цистеиновый остаток. Такая молекула предпочтительно содержит C1-C6-алкильную, циклоалкильную (C5, C6), арильную или гетероарильную функциональную группу. Однако кроме аминокислотного линкера в область изобретения также необходимо включить линкер, предпочтительно содержащий C1-C6-алкильную, циклоалкильную (C5, C6), арильную или гетероарильную функциональную группу и не содержащий аминокислот(ы). Ассоциация между антигеном или антигенной детерминантой или, необязательно, вторым участком прикрепления и аминокислотным линкером предпочтительно осуществляется, по меньшей мере, посредством одной ковалентной связи, более предпочтительно - по меньшей мере, посредством одной пептидной связи.
Животное: Как применяют здесь, термин "животное" предназначен для включения в себя, например, людей, овец, лосей, оленя, чернохвостого оленя, норок, млекопитающих, обезьян, лошадей, крупного рогатого скота, свиней, коз, собак, кошек, крыс, мышей, птиц, цыплят, рептилий, рыб, насекомых и паукообразных.
Антитело: Как применяют здесь, термин "антитело" относится к молекулам, способным к связыванию эпитопов или антигенных детерминант. Термин предназначен для включения в себя полных антител и их антигенсвязывающих фрагментов, включая одноцепочечные антитела. Наиболее предпочтительные антитела представляют собой антигенсвязывающие фрагменты человеческих антител и включают в себя в качестве неограничивающих примеров Fab, Fab' и F(ab')2, Fd, одноцепочечные Fvs (scFv),одноцепочечные антитела, связанные дисульфидными связями Fvs (sdFv) и фрагменты, содержащие или VL- или VH-домен. Антитела могут происходить от любого животного, включая птиц и млекопитающих. Предпочтительно антитела являются человеческими, мышиными, кроличьими, козьими, антителами морских свинок, верблюжьими, лошадиными или куриными. Как применяют здесь, термин "человеческие" антитела включает в себя антитела с аминокислотной последовательностью человеческого иммуноглобулина и включает в себя антитела, выделенные из библиотек человеческих иммуноглобулинов или из животных, трансгенных по одному или нескольким человеческим иммуноглобулинами и не экспрессирующих эндогенные иммуноглобулины, как описано, например, в патенте США № 5939598, выданному Kucherlapati et al.
Антиген: Как применяют здесь, термин "антиген" относится к молекуле, которую способно связывать антитело или T-клеточный рецептор (TCR), если она представлена молекулами MHC. Как применяют здесь, термин "антиген" также охватывает T-клеточные эпитопы. Антиген дополнительно способен распознаваться иммунной системой и/или способен индуцировать гуморальный иммунный ответ и/или клеточный иммунный ответ, приводя к активации B- и/или T-лимфоцитов. Это, однако, может потребовать, чтобы, по меньшей мере, в некоторых случаях антиген содержал эпитоп Th-клеток или являлся связанным с ним и чтобы его вводили в адъюванте. Антиген может обладать одним или несколькими эпитопами (B- и T-эпитопы). Как предполагают, указанная выше конкретная реакция указывает на то, что антиген предпочтительно реагирует, как правило, высокоизбирательно с соответствующим антителом или TCR, а не с множеством антител или TCR индуцируемых другими антигенами. Как применяют здесь, антигены могут представлять собой смеси нескольких отдельных антигенов.
Антигенная детерминанта: Как применяют здесь, термин "антигенная детерминанта" предназначен для обозначения той части антигена, которая специфически распознается B- или T-лимфоцитами. B-лимфоциты, реагирующие на антигенные детерминанты, образуют антитела, тогда как T-лимфоциты отвечают на антигенные детерминанты пролиферацией и развитием эффекторных функций, важных для опосредования клеточного и/или гуморального иммунитета.
Ассоциация: Как применяют здесь, термин "ассоциация", как его применяют к первому и второму участкам прикрепления, относится к связыванию первого и второго участка прикрепления, предпочтительно посредством, по меньшей мере, одной непептидной связи. Связь может являться ковалентной, ионной, гидрофобной, полярной или любым их сочетанием, предпочтительно тип связи является ковалентным.
Участок прикрепления, первый: Как применяют здесь, фраза "первый участок прикрепления" относится к элементу неприродного или природного происхождения, с которым может ассоциироваться второй участок прикрепления, расположенный на антигене или антигенной детерминанте. Первый участок прикрепления может представлять собой белок, полипептид, аминокислоту, пептид сахар, полинуклеотид, природный или синтетический полимер, вторичный метаболит или соединение (биотин, флуоресцеин, ретинол, дигоксигенин, ионы металлов, фенилметилсульфонилфторид), или их сочетание, или их химически активные группы. Первый участок прикрепления, как правило и предпочтительно расположен на поверхности коровой частицы, предпочтительно такой как вирусоподобная частица. На поверхности коровой или вирусоподобной частиц соответственно находятся множество первых участков прикрепления, как правило, в повторяющейся конфигурации.
Участок прикрепления, второй: Как применяют здесь, фраза "второй участок прикрепления" относится к элементу, ассоциированному с антигеном или антигенной детерминантой, с которым может ассоциироваться первый участок прикрепления, расположенный на поверхности коровой частицы или вирусоподобной частицы соответственно. Второй участок прикрепления антигена или антигенной детерминанты может представлять собой белок, полипептид, пептид, сахар, полинуклеотид, природный или синтетический полимер, вторичный метаболит или соединение (биотин, флуоресцеин, ретинол, дигоксигенин, ионы металлов, фенилметилсульфонилфторид), или их сочетание, или их химически активные группы. На антигене или антигенной детерминанте находится, по меньшей мере, один второй участок прикрепления. Следовательно, термин "антиген или антигенная детерминанта, по меньшей мере, с одним вторым участком прикрепления" относится к антигену или антигенной конструкции, содержащей, по меньшей мере, антиген или антигенную детерминанту и второй участок прикрепления. Однако в антигене или антигенной детерминанте, в частности для второго участка прикрепления неприродного происхождения, т.е. не встречающегося в природе, данный антиген или антигенные конструкции содержат "аминокислотный линкер".
Связь: Как применяют здесь, термин "связь" относится к связыванию или прикреплению, которое может являться ковалентным, например, посредством химического связывания, или нековалентным, например, ионные взаимодействия, гидрофобные взаимодействия водородные связи и т.п. Ковалентные связи могут, например, представлять собой сложные эфиры, простые эфиры, фосфоэфиры, амиды, пептиды, имиды, сероуглеродные связи, фосфоуглеродные связи и т.п. Термин "связан" является более широким, чем такие термины, как "соединен", "слит" и "прикреплен", и включает их в себя.
Белок(ки) оболочки: Как применяют здесь, термин "белок(ки) оболочки" относится к белку(ам) бактериофага или РНК-фага, способного встраиваться в капсидную конструкцию бактериофага или РНК-фага. Однако при ссылке на конкретный генный продукт гена белка оболочки РНК-фагов применяют термин "CP". Например, конкретный генный продукт гена белка оболочки РНК-фага Qβ обозначен "CP Qβ", тогда как "белки оболочки" бактериофага Qβ включают в себя "CP Qβ", а также белок A1. Капсид бактериофага Qβ преимущественно состоит из CP Qβ с небольшим содержанием белка A1. Подобным образом белок оболочки VLP Qβ преимущественно состоит из CP Qβ с небольшим содержанием белка A1.
Коровая частица: Как применяют здесь, термин "коровая частица" относится к жесткой структуре с собственной повторяющейся структурой. Как применяют здесь, коровая частица может представлять собой продукт синтетического процесса или продукт биологического процесса.
Связан: Как применяют здесь, термин "связан" относится к прикреплению посредством ковалентных связей или посредством сильных нековалентных взаимодействий, как правило и предпочтительно к прикреплению посредством ковалентных связей. По данному изобретению можно применять любой способ, обычно применяемый специалистами в данной области для связывания биологически активных веществ.
Эффективное количество: Как применяют здесь, термин "эффективное количество" относится к количеству, необходимому или достаточному для реализации желаемого биологического эффекта. Эффективное количество композиции представляет собой количество, с применением которого достигают данного выбранного результата, и такое количество специалист в данной области может определить, по сути, обычными способами. Например, эффективное количество для лечения недостаточности иммунной системы может являться количеством, необходимым для того, чтобы послужить причиной активации иммунной системы, приводящего к развитию специфичного к антигену иммунного ответа при экспозиции антигена. Термин также является синонимом с "достаточным количеством".
Эффективное количество для любого конкретного применения может варьировать, в зависимости от таких факторов как заболевание или состояние, которое необходимо лечить, конкретной композиции для введения, размера субъекта и/или тяжести заболевания или состояния. Специалист в данной области, без необходимости чрезмерного экспериментирования, может эмпирически определить эффективное количество конкретной композиции по настоящему изобретению.
Эпитоп: Как применяют здесь, термин "эпитоп" относится к непрерывным или прерывающимся частям полипептида с антигенной или иммуногенной активностью у животного, предпочтительно млекопитающего, а наиболее предпочтительно - у человека. Эпитоп распознается антителом или T-клеткой посредством ее T-клеточного рецептора в окружении молекулы MHC. Как применяют здесь, "иммуногенный эпитоп" определен как часть полипептида, вызывающая ответ антителами или индуцирующая T-клеточный ответ у животного, как определено любым известным в данной области способом. (См., например, Geysen et al., Proc. Natl. Acad. Sci. USA 81: 3998-4002 (1983)). Как применяют здесь, термин "антигенный эпитоп" определен как часть белка, с которым антитело может иммуноспецифически связывать свой антиген, как определено любым хорошо известным в данной области способом. Иммуноспецифическое связывание исключает неспецифическое связывание, но не обязательно исключает перекрестную реактивность с другими антигенами. Антигенным эпитопам не обязательно необходимо являться иммуногенными. Антигенные эпитопы также могут представлять собой T-клеточные эпитопы, и в данном случае они могут иммуноспецифически связываться T-клеточным рецептором в окружении молекулы MHC.
Эпитоп может включать в себя 3 аминокислоты в пространственной конформации, уникальной для эпитопа. Как правило, эпитоп состоит, по меньшей мере, приблизительно из 5 таких аминокислот, а более обычно состоит, по меньшей мере, приблизительно из 8-10 таких аминокислот. Если эпитоп представляет собой органическую молекулу, он может являться таким же малым, как нитрофенил.
Слияние: Как применяют здесь, термин "слияние" относится к объединению аминокислотных последовательностей разного происхождения в одной полипептидной цепи посредством объединения внутри одной рамки считывания их кодирующих нуклеотидных последовательностей. Термин "слияние" в дополнение к слиянию с одним из его концов явно включает в себя внутренние слияния, т.е. вставки последовательностей другого происхождения в полипептидную цепь.
Грелин: Как применяют здесь, термин "грелин" относится к пептиду, полученному из белка, кодируемого геном грелина. Как применяют здесь, грелин включает в себя все формы грелинов, известные у людей, кошек, собак и всех домашних животных, а также других животных. Как применяют здесь, грелин включает в себя грелин с н-октаноиловой модификацией или без нее. Кроме того, грелин также включает в себя все варианты сплайсинга, существующие у грелина. Кроме того, ввиду высокой гомологии последовательностей у грелинов разных видов (у крысы и человека отличаются только 2 а.к.) и человеческого грелина (Kojima et al. Nature 402: 656-660 (1999)), все природные варианты грелина с более чем 80% идентичности, предпочтительно - более чем с 90%, более предпочтительно - более чем с 95% и даже более предпочтительно - более чем с 99% с человеческим грелином обозначают здесь как "грелин".
Как применяют здесь, термин "полученный из грелина пептид" или "пептиды грелина" в широком контексте определен как любой пептид, представляющий собой часть грелина или содержащий, по меньшей мере, два, предпочтительно - по меньшей мере, три, более предпочтительно - по меньшей мере, четыре, более предпочтительно - по меньшей мере, пять или, по меньшей мере, шесть и даже более предпочтительно - по меньшей мере, восемь или девять или даже более предпочтительно - по меньшей мере, десять последовательных аминокислот исходного пептида грелина. Как правило, "пептид грелина" и "фрагмент грелина" применяют попеременно. Кроме того, как применяют здесь, термины "пептид грелина и его фрагменты" должны включать кроме пептида грелина любую часть указанного пептида грелина, где указанную часть предпочтительно можно получить посредством удаления одной или нескольких аминокислот от N- и/или C-конца. Пептид грелина можно получить посредством рекомбинантной экспрессии в эукариотических или прокариотических экспрессионных системах, как пептид грелина отдельно или как гибрид с другими аминокислотами или белками, например, для облегчения укладки, экспрессии или растворимости пептида грелина или для облегчения очистки пептида грелина. Для облегчения или возможности правильной укладки гибридных белков пептидов грелина и белков субъединиц VLP или капсидов к пептидам грелина можно добавлять одну или несколько аминокислот на N- или C-конце. Для обеспечения возможности связывания пептидов грелина с белками субъединиц VLP или капсидов или коровых частица к пептиду грелина можно добавить, по меньшей мере, один второй участок прикрепления. Альтернативно пептиды грелина можно синтезировать с применением известных в данной области способов. Такие пептиды даже могут содержать аминокислоты, не содержащиеся в соответствующем белке грелина. Пептиды можно модифицировать н-октаноилированием.
Остаток: Как применяют здесь, термин "остаток" означает конкретную аминокислоту в полипептидном остове или боковой цепи.
Иммунный ответ: Как применяют здесь, термин "иммунный ответ" относится к гуморальному иммунному ответу и/или клеточному иммунному ответу, приводящим к активации или пролиферации B- и/или T-лимфоцитов и/или антигенпредставляющих клеток. Однако в некоторых случаях иммунные ответы могут обладать малой интенсивностью и их можно обнаружить только при применении, по меньшей мере, одного вещества по настоящему изобретению. "Иммуногенный" относится к средству, применяемому для стимуляции иммунной системы живого организма так, что усиливаются и направляются против иммуногенного средства одна или несколько функций иммунной системы. "Иммуногенный полипептид" представляет собой полипептид, вызывающий клеточный и/или гуморальный иммунный ответ, или сам или когда связан с носителем в присутствии или в отсутствие адъюванта. Предпочтительно антигенпредставляющая клетка может являться активированной.
Вещество, "усиливающее" иммунный ответ представляет собой такое вещество, когда при добавлении вещества наблюдают иммунный ответ, больший или более интенсивный или, во всяком случае, отличающийся при сравнении с таким же иммунным ответом, измеренным без добавления вещества. Например, литическую активность цитотоксических T-клеток можно измерить, например, применяя анализ высвобождения 51Cr в образцах, полученных с применением во время иммунизации вещества или без него. Полагают, что количество вещества, при котором увеличивается литическая активность CTL при сравнении с литической активностью CTL без вещества, является количеством, достаточным для усиления иммунного ответа животного на антиген. В предпочтительном осуществлении иммунный ответ увеличен, по меньшей мере, приблизительно в 2 раза, более предпочтительно - приблизительно в 3 раза и более. Также можно изменять количество или тип секретируемых цитокинов. Альтернативно также можно изменять количество индуцируемых антител или их подклассов.
Иммунизация: Как применяют здесь, термины "иммунизировать" или "иммунизация" или родственные термины относятся к приданию способности проявлять значительный иммунный ответ (включающий в себя антитела и/или клеточный иммунитет, такой как эффекторные CTL) против целевого антигена или эпитопа. Данные термины не требуют того, чтобы создавался полный иммунитет, а скорее чтобы образовывался иммунный ответ, существенно больший, чем на исходном уровне. Например, млекопитающее можно рассматривать как иммунизированное против целевого антигена, если после применения способов по изобретению развивается клеточный и/или гуморальный иммунный ответ на целевой антиген.
Природное происхождение: Как применяют здесь, термин "природное происхождение" означает, что целое или его часть не синтезированы и существуют или получены в природных условиях.
Неприродный: Как применяют здесь, термин, как правило, означает не из природы, более конкретно термин означает сделанный человеком.
Неприродного происхождения: Как применяют здесь, термин "неприродного происхождения", как правило, означает синтетический или не из природы; более конкретно, термин означает сделанный человеком.
Упорядоченный и повторяющийся массив антигенов или антигенных детерминант: Как применяют здесь, термин "упорядоченный и повторяющийся массив антигенов или антигенных детерминант", как правило, относится к повторяющейся структуре антигенов или антигенных детерминант, характеризующейся типичным и предпочтительно однообразным пространственным расположением антигенов или антигенных детерминант по отношению к коровой частице или вирусоподобной частице соответственно. В одном осуществлении данного изобретения повторяющаяся структура может являться геометрической структурой. Типичные и предпочтительные примеры пригодных упорядоченных и повторяющихся массивов антигенов или антигенных детерминант представляют собой массивы, обладающие точно повторяющимися паракристаллическими последовательностями антигенов или антигенных детерминант, предпочтительно с шагом от 1 до 30 нанометров, предпочтительно от 5 до 15 нанометров.
Пили: Как применяют здесь, термин "пили" (единственное число представляет собой "пиль") относится к внеклеточным структурам бактериальных клеток, состоящих из белковых мономеров (например, мономеров пилина), организованных в упорядоченные и повторяющиеся структуры. Кроме того, пили представляют собой структуры, вовлеченные в такие процессы, как прикрепление бактериальной стенки к поверхностным рецепторам клеточной поверхности хозяина, межклеточный генетический обмен и узнавание клетка-клетка. Примеры пилей включают в себя пили типа 1, P-пили, F1C-пили, S-пили и 987P-пили. Дополнительные примеры пилей рассмотрены ниже.
Пилеобразная структура: Как применяют здесь, фраза "пилеобразная структура" относится к структурам со сходными с пилями характеристиками и состоящим из белковых мономеров. Одним примером "пилеобразной структуры" является структура, формируемая бактериальной стенкой, экспрессирующей модифицированные пилиновые белки, не формирующие упорядоченные и повторяющиеся массивы, идентичные таковым природных пилей.
Полипептид: Как применяют здесь, термин "полипептид" относится к молекуле, состоящей из мономеров (аминокислот), линейно связанных амидными связями (также известными как пептидные связи). Он означает молекулярную цепь аминокислот и не относится к конкретной длине продукта. Таким образом, пептиды, дипептиды, трипептиды, олигопептиды и белки включены в определение полипептида. Данный термин также предназначен для обозначения постэкспрессионных модификаций полипептида, например гликозилирований, ацетилирований, фосфорилирований и т.п. Рекомбинантный или производный полипептид не обязательно транслирован с указанной последовательности нуклеиновой кислоты. Его также можно получить любым способом, включая химический синтез.
Аутоантиген: Как применяют здесь, термин "аутоантиген" относится к белкам, кодируемым ДНК хозяина, а продукты, образуемые белками или РНК, кодируемыми ДНК хозяина, определены как собственные. Кроме того, белки, образующиеся при объединении двух или нескольких собственных молекул или представляющие собой часть собственной молекулы и белков, обладающих высокой гомологией со второй собственной молекулой, как определено выше (>95%, предпочтительно - >97%, более предпочтительно - >99%), также можно рассматривать как собственные.
Лечение: Как применяют здесь, термины "лечение", "лечить", "леченый" или "лечащий" относятся к профилактике и/или лечению. Когда его применяют, например, по отношению к инфекционному заболеванию, термин относится к профилактическому лечению, увеличивающему устойчивость субъекта к инфекции патогеном или, другими словами, уменьшает вероятность того, что субъект будет инфицирован данным патогеном или продемонстрирует признаки заболевания, свойственные инфекции, а также лечению после инфицирования субъекта для борьбы с инфекцией, например уменьшению или устранению инфекции или профилактике ухудшения. Когда его применяют по отношению к ожирению или связанным заболеваниям, термин "лечение" относится к профилактическому или терапевтическому лечению, увеличивающему устойчивость субъекта к ожирению и/или обращающему его.
Вакцина: Как применяют здесь, термин "вакцина" относится к препарату, содержащему композицию по настоящему изобретению и находящемуся в форме, которую можно ввести животному. Как правило, вакцина содержит традиционную солевую или забуференную среду водного раствора, в которой суспендирована или растворена композиция по настоящему изобретению. В данной форме композицию по настоящему изобретению без труда можно применять для профилактики, ослабления или другого лечения состояния. После введения хозяину вакцина способна провоцировать иммунный ответ, включающий в себя в качестве неограничивающих примеров образование антител и/или цитокинов и/или активацию цитотоксических T-клеток, антигенпредставляющих клеток, хелперных T-клеток, дендритных клеток и/или другие виды клеточного ответа.
Необязательно вакцина по настоящему изобретению дополнительно включает в себя адъювант, который относительно соединения по настоящему изобретению может присутствовать в меньшей или в большей пропорции.
Вирусоподобная частица (VLP): Как применяют здесь, термин "вирусоподобная частица" относится к структуре, подобной вирусной частице. Кроме того, вирусоподобная частица по настоящему изобретению является нереплицируемой и неинфекционной, так как у нее отсутствуют весь или часть вирусного генома, в частности, репликативный и инфекционный компоненты вирусного генома. Вирусоподобная частица по настоящему изобретению может содержать нуклеиновую кислоту, отличную от ее генома. Типичным и предпочтительным осуществлением вирусоподобной частицы по настоящему изобретению является вирусный капсид, такой как вирусный капсид соответствующего вируса, бактериофага или РНК-фага. Как поочередно применяют здесь, термины "вирусный капсид" или "капсид" относятся к макромолекулярной структуре, состоящей из субъединиц вирусного белка. Типично и предпочтительно субъединицы вирусного белка собираются в вирусный капсид и капсид соответственно, обладающий структурой с собственной повторяющейся организацией, где указанная структура, как правило, является сферической или трубчатой. Например, капсиды РНК-фагов или HBcAg обладают сферической формой икосаэдрической симметрии. Как применяют здесь, термин "структура подобная капсиду" относится к макромолекулярной структуре, состоящей из субъединиц вирусного белка, подобной морфологии капсида в определенном выше смысле, но отклоняющейся от обычной симметрической структуры, хотя и при поддержании достаточной степени упорядоченности и повторяемости.
Вирусоподобная частица бактериофага: Как применяют здесь, термин "вирусоподобная частица бактериофага" относится к вирусоподобной частице подобной структуре бактериофага, не являющейся реплицирующейся и инфекционной и в которой отсутствуют, по меньшей мере, ген или гены, кодирующие аппарат репликации бактериофага, и, как правило, также отсутствует ген или гены, кодирующие белок или белки, ответственные за прикрепление вируса или проникновение вируса в хозяина. Однако данное определение должно также охватывать вирусоподобные частицы бактериофагов, в которых указанный выше ген или гены все еще присутствуют, но не активны и, следовательно, также приводят к нереплицирующейся и неинфекционной вирусоподобной частице бактериофага.
Белок оболочки VLP или РНК-фага: Капсидную структуру, сформированную посредством самосборки 180 субъединиц белка оболочки РНК-фага и необязательно содержащую РНК хозяина, обозначают как "белок оболочки VLP или РНК-фага". Конкретный пример представляет собой белок оболочки VLP Qβ. В данном конкретном случае белок оболочки VLP Qβ может собираться или исключительно из субъединиц CP Qβ (образуемых посредством экспрессии гена CP Qβ, содержащего, например, стоп-кодон TAA, предотвращающий любую экспрессию более длинного белка A1 вследствие супрессии, см. Kozlovska, T. M., et al., Intervirology 39: 9-15 (1996)), или дополнительно содержать в структуре капсида субъединицы белка A1.
Вирусная частица: Как применяют здесь, термин "вирусная частица" относится к морфологической форме вируса. Некоторые типов вирусов содержит геном, окруженный белковым капсидом; другие обладают дополнительными структурами (например, оболочки, хвосты и т.п.).
Один или какой-либо: Когда в данном описании применяют термины "один" или "какой-либо", они означают "по меньшей мере, один" или "один или несколько", если не указано иначе.
Как станет ясно специалистам в данной области, определенные осуществления данного изобретения включают в себя применение технологий рекомбинантных нуклеиновых кислот, таких как клонирование, полимеразная цепная реакция, очистка ДНК и РНК, экспрессия рекомбинантных белков в прокариотических и эукариотических клетках и т.п. Такие способы хорошо известны специалистам в данной области, и их без труда можно найти в опубликованных руководствах по лабораторным методам (например, Sambrook, J. et al., eds., Molecular Cloning, A Laboratory Manual, 2nd. edition, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N. Y. (1989); Ausubel, F. et al., eds., Current Protocols in Molecular Biology, John H. Wiley & Sons, Inc. (1997)). Фундаментальные лабораторные технологии для работы с клеточными линиями тканевых культур (Celis, J., ed., Cell Biology, Academic Press, 2nd edition, (1998)) и технологии, основанные на антителах (Harlow, E. and Lane, D., Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, N. Y. (1988); Deutscher, M. P., "Guide to Protein Purification", Meth. Enzymol. 128, Academic Press San Diego (1990); Scopes, R. K., Protein Purification Principles and Practice, 3rd ed., Springer-Verlag, New York (1994)) также в достаточной мере описаны в литературе, и все включены сюда в качестве ссылки.
2. Композиции и способы для усиления иммунного ответа.
Раскрываемое изобретение относится к композициям и способам для усиления иммунного ответа против грелина или пептида грелина у животного. Композиции по изобретению содержат или, альтернативно, состоят из (a) коровой частицы, по меньшей мере, с одним первым участком прикрепления и (b) по меньшей мере, одного антигена или антигенной детерминанты, по меньшей мере, с одним вторым участком прикрепления, где указанный антиген или антигенная детерминанта представляет собой грелин или полученный из грелина пептид и где указанный второй участок прикрепления выбран из группы, состоящей из (i) участка прикрепления, не встречающегося в природе с указанным антигеном или антигенной детерминантой, и (ii) участка прикрепления, встречающегося в природе с указанным антигеном или антигенной детерминантой, где указанный второй участок прикрепления способен к ассоциации с указанным первым участком прикрепления и где указанный антиген или антигенная детерминанта и указанная коровая частица взаимодействуют посредством указанной ассоциации для формирования упорядоченного и повторяющегося массива антигенов. Более конкретно, композиции по изобретению содержат или, альтернативно, состоят из вирусоподобной частицы и, по меньшей мере, одного антигена или антигенной детерминанты, где антиген или антигенная детерминанта представляет собой грелин или пептид грелина и где, по меньшей мере, один антиген или антигенная детерминанта связаны с вирусоподобной частицей, так чтобы формировать упорядоченный и повторяющийся массив антиген-VLP. Кроме того, изобретение позволяет практикам легко сконструировать такую композицию, в частности, для лечения и/или профилактического предотвращения ожирения.
В одном осуществлении коровая частица содержит или выбрана из группы, состоящей из вируса, бактериального пиля, структуры, сформированной бактериальным пилином, бактериофага, вирусоподобной частицы, вирусоподобной частицы РНК-фага, частицы вирусного капсида или их рекомбинанта. В качестве коровой частицы по изобретению можно выбрать любой известный в данной области вирус с упорядоченной и повторяющейся оболочкой и/или структурой белков оболочки; примеры пригодных вирусов включают в себя вирус синдбис и другие альфавирусы, рабдовирусы (например, вирус везикулярного стоматита), пикорнавирусы (например, человеческий риновирус, вирус Айчи), тогавирусы (например, вирус краснухи), ортомиксовирусы (вирус Тогото, вирус Баткен, вирус птичьей чумы), вирусы полиомы (например, вирус полиомы BK, вирус полиомы JC, птичий вирус полиомы BFDV), парвовирусы, ротавирусы, вирус Норфолка, вирус ящура, ретровирус, вирус гепатита B, вирус табачной мозаики, вирус Овечьих кошар и вирус папилломы человека и предпочтительно РНК-фаг, бактериофаг Qβ, бактериофаг R17, бактериофаг M11, бактериофаг MX1, бактериофаг NL95, бактериофаг fr, бактериофаг GA, бактериофаг SP, бактериофаг MS2, бактериофаг f2, бактериофаг PP7 (например, см. таблицу 1 у Bachmann, M. F. and Zinkernagel, R. M., Immunol. Today 17: 553-558 (1996)).
В дополнительном осуществлении в изобретении применяют генетическую инженерию вируса для создания гибрида между упорядоченным и повторяющимся вирусным белком оболочки и первым участком прикрепления, содержащим гетерологичный белок, пептид, антигенную детерминанту или активный аминокислотный остаток выбора, или, альтернативно или предпочтительно, являющимся им. При конструировании композиций по изобретению можно включать другие известные специалистам в данной области генетические манипуляции; например может являться желательным ограничить репликационную способность рекомбинантного вируса посредством генетической мутации. Кроме того, применяемый для настоящего изобретения вирус является репликационно неполноценным вследствие химической или физической инактивации или, как указано, вследствие отсутствия способного к репликации генома. Вирусный белок, выбранный для слияния с первым участком прикрепления, должен обладать упорядоченной и повторяющейся структурой. Такая организованная и повторяющаяся структура включает в себя паракристаллические структуры с шагом 5-30 нм, предпочтительно - 5-15 нм, на поверхности вируса. Создание гибридного белка данного типа дает в результате множественные, упорядоченные и повторяющиеся первые участки прикрепления на поверхности вируса и отражает нормальную структуру природного вирусного белка. Как будет понятно специалистам в данной области, первый участок прикрепления может являться или не являться частью любого пригодного белка, полипептида, сахара, полинуклеотида, пептида (аминокислоты), природного или синтетического полимера, вторичного метаболита или их сочетаний, которые могут служить для специфического прикрепления антигена или антигенной детерминанты, ведущего к упорядоченному и повторяющемуся массиву антигенов.
В другом осуществлении изобретения коровая частица представляет собой рекомбинантный альфавирус, а более конкретно, рекомбинантный вирус синдбис. Альфавирусы представляют собой РНК-содержащие вирусы с положительными цепями, которые полностью и без промежуточной ДНК реплицируют свою геномную РНК в цитоплазме инфицированной клетки (Strauss, J. and Strauss, E., Microbiol. Rev. 58: 491-562 (1994)). На некоторых представителей семейства альфавирусов, синдбис (Xiong, C. et al., Science 243: 1188-1191 (1989); Schlesinger, S., Trends Biotechnol. 11: 18-22 (1993)), вирус леса Семлики (SFV) (Liljestrom, P. & Garoff, H., Bio/Technology 9: 1356-1361 (1991)) и другие (Davis, N. L. et al., Virology 171: 189-204 (1989)), обратили значительное внимание в направлении применения в качестве основанных на вирусах экспрессирующих векторов из-за множества различных белков (Lundstrom, K., Curr. Opin. Biotechnol. 8: 578-582 (1997); Liljestrom, P., Curr. Opin. Biotechnol. 5: 495-500 (1994)) и в качестве кандидатов для разработки вакцин. Недавно был выдан ряд патентов, направленных на применение альфавирусов для экспрессии гетерологичных белков и для разработки вакцин (см. патенты США №№ 5766602; 5792462; 5739026; 5789245 и 5814482). Конструирование альфавирусных коровых частиц по изобретению можно произвести средствами, в основном известными в данной области технологии рекомбинантных ДНК, как описано в указанных выше статьях, включенных сюда в качестве ссылки.
Для получения основанных на вирусах коровых частиц для прикрепления антигенов или антигенных детерминант можно применять множество различных рекомбинантных клеток-хозяев. Например, известно, что альфавирусы обладают широким кругом хозяев; вирус синдбис инфицирует культивируемые клетки млекопитающих, рептилий и амфибий, а также некоторые клетки насекомых (Clark, H., J. Natl. Cancer Inst. 51: 645 (1973); Leake, C., J. Gen. Virol. 35: 335 (1977); Stollar, V. в THE TOGAVIRUSES, R. W. Schlesinger, Ed., Academic Press, (1980), pp. 583-621). Таким образом, в практическом применении изобретения можно использовать множество рекомбинантных клеток-хозяев. Клетки BHK, COS, Vero, HeLa и CHO особенно пригодны для получения гетерологичных белков, так как они обладают потенциалом для гликозилирования гетерологичных белков сходным с человеческим клетками образом (Watson, E. et al., Glycobiology 4: 227, (1994)) и их можно подвергать селекции (Zang, M. et al., Bio/Technology 13: 389 (1995)) или генетически модифицировать (Renner W. et al., Biotech. Bioeng 4: 476 (1995); Lee K. et al. Biotech. Bioeng 50: 336 (1996)) для роста в среде без сыворотки, а также в суспензии.
Введение полинуклеотидных векторов в клетки-хозяева можно проводить способами, описанными в стандартных лабораторных руководствах (см., например, Sambrook, J. et al., eds., MOLECULAR CLONING, A LABORATORY MANUAL, 2nd. edition, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N. Y. (1989), глава 9; Ausubel, F. et al., eds., CURRENT PROTOCOLS IN MOLECULAR BIOLOGY, John H. Wiley & Sons, Inc. (1997), глава 16), включающих в себя такие способы, как электропорация, опосредованная DEAE-декстраном трансфекция, трансфекция, микроинъекция, катионная опосредованная липидами трансфекция, трансдукция, scrape loading, баллистическое введение и инфицирование. Способы введения экзогенных последовательностей ДНК в клетки-хозяева обсуждаются у Felgner, P. et al., патент США № 5580859.
Упакованные последовательности РНК также можно применять для инфекции клеток-хозяев. Данные упакованные последовательности РНК можно вводить клеткам-хозяевам посредством добавления их к среде культивирования. Например, получение неинфекционных альфавирусных частиц описано в ряде источников, включающих в себя "Sindbis Expression System", Version C (Invitrogen Catalog № K750-1).
Когда в качестве рекомбинантных клеток-хозяев для получения основанных на вирусах коровых частиц применяют клетки млекопитающих, данные клетки, как правило, растут в тканевой культуре. Способы выращивания клеток в культуре хорошо известны в данной области (см., например, Celis, J., ed., CELL BIOLOGY, Academic Press, 2nd edition, (1998); Sambrook, J. et al., eds., MOLECULAR CLONING, A LABORATORY MANUAL, 2nd. edition, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N. Y. (1989); Ausubel, F. et al., eds., CURRENT PROTOCOLS IN MOLECULAR BIOLOGY, John H. Wiley & Sons, Inc. (1997); Freshney, R., CULTURE OF ANIMAL CELLS, Alan R. Liss, Inc. (1983)).
Дополнительные примеры РНК-вирусов, пригодных для применения в качестве коровой частицы по настоящему изобретению, включают в себя в качестве неограничивающих примеров следующее: представители семейства Reoviridae, включающие в себя род Orthoreovirus (множество серотипов ретровирусов млекопитающих и птиц), род Orbivirus (вирус катаральной лихорадки овец, вирус Eugenangee, вирус Кемерово, вирус африканской болезни лошадей и вирус колорадской клещевой лихорадки), род Rotavirus (ротавирус человека, вирус диареи телят Небраски, мышиный ротавирус, ротавирус обезьян, ротавирус коров или овец, птичий ротавирус); семейство Picomaviridae, включающее в себя род Enterovirus (полиовирус, вирус коксаки A и B, кишечные цитопатические человеческие "сиротские" (ECHO) вирусы, вирусы гепатитов A, C, D, E и G, обезьяньи энтеровирусы, вирусы мышиного энцефаломиелита (ME), вирус полиомиелита мышей, коровьи энтеровирусы, энтеровирусы свиней), род Cardiovirus (вирус энцефаломиокардита (EMC), менговирус), род Rhinovirus (человеческий риновирус, включающий в себя, по меньшей мере, 113 подтипов; другие риновирусы), род Apthovirus (ящур (FMDV)); семейство Calciviridae, включающее в себя вирус везикулярной экзантемы свиней, вирус морских львов Сан-Мигеля, Кошачий пикорнавирус и вирус Норфолк; семейство Togaviridae, включающее в себя род Alphavirus (восточный вирус лошадиного энцефалита, вирус леса Семлики, вирус синдбис, вирус чикунгунья, вирус O'Nyong-Nyong, вирус Росс-Ривер, венесуэльский вирус лошадиного энцефалита, западный вирус лошадиного энцефалита), род Flavirius (передаваемый комарами вирус желтой лихорадки, вирус тропической лихорадки, японский вирус энцефалита, вирус энцефалита Сент-Льюиса, вирус энцефалита долины Муррей, вирус Западного Нила, вирус Кунжин, центральноевропейский передаваемый клещами вирус, дальневосточный передаваемый клещами вирус, киасанурский лесной вирус, вирус шотландского энцефалита, вирус Powassan, омский вирус геморрагической лихорадки), род Rubivirus (вирус краснухи), род Pestivirus (вирус заболевания слизистой, вирус холеры свиней, вирус пограничной болезни овец); семейство Bunyaviridae, включающее в себя род Bunyvirus (Буньямвера и родственные вирусы, вирусы группы калифорнийского энцефалита), род Phlebovirus (сицилийский вирус флеботомной лихорадки, вирус лихорадки долины Рифт), род Nairovirus (вирус конго-крымской геморрагической лихорадки, вирус болезни найробианских овец), и род Uukuvirus (Uukuniemi и родственные вирусы); семейство Orthomyxoviridae, включающее в себя род Influenza virus (вирус гриппа типа A, множество человеческих подтипов); вирус гриппа свиней и обезьян и вирусы гриппа лошадей; грипп типа B (множество человеческих подтипов) и грипп типа C (возможно отдельный род); семейство Paramyxoviridae, включающее в себя род Paramyxovirus (вирус парагриппа типа 1, вирус сендай, вирус гемадсорбции, вирусы парагриппа типов со 2 по 5, вирус болезни Ньюкастла, вирус свинки), род Morbillivirus (вирус кори, вирус подострого склерозирующего панэнцефалита, вирус чумки, вирус чумы рогатого скота), род Pneumovirus (респираторно-синцитиальный вирус (RSV), бычий респираторно-синцитиальный вирус и вирус пневмонии мышей); семейство Rhabdoviridae, включающее в себя род Vesiculovirus (VSV), вирус Шандипура, вирус фландрийского оленьего парка), род Lyssavirus (вирус бешенства), рабдовирусы рыб и филовирусы (марбургский вирус и вирус Эбола); семейство Arenaviridae, включающее в себя вирус лимфоцитарного хориоменингита (LCM), вирусный комплекс Tacaribe и вирус Ласса; семейство Coronoaviridae, включающее в себя вирус инфекционного бронхита (IBV), вирус мышиного гепатита, человеческий кишечный коронавирус и вирус кошачьего перитонита (кошачий коронавирус).
Иллюстративные ДНК-содержащие вирусы, которые можно применять в качестве коровых частиц, включают в себя в качестве неограничивающих примеров семейство Poxviridae, включающее в себя род Orthopoxvirus (натуральная оспа, аластрим, обезьянья поксвакцина, коровья оспа, оспа буйволов, оспа кроликов, эктромелия), род Leporipoxvirus (миксома, фиброма), род Avipoxvirus (оспа птиц, другие птичьи поксвирусы), род Capripoxvirus (оспа овец, оспа коз), род Suipoxvirus (оспа свиней), род Parapoxvirus (вирус контагиозного пустулезного дерматита, ложная коровья оспа, вирус бычьего папулезного стоматита); семейство Iridoviridae (вирус африканской лихорадки свиней, вирусы лягушки 2 и 3, вирус лимфоцистоза рыб); семейство Herpesviridae, включающее в себя альфа-герпесвирусы (простой герпес типов 1 и 2, ветряная оспа, вирус абортов лошадей, лошадиный вирус герпеса 2 и 3, вирус псевдобешенства, вирус инфекционного бычьего кератоконъюнктивита, вирус инфекционного бычьего ринотрахеита, вирус кошачьего ринотрахеита, вирус инфекционного ларинготрахеита) бета-герпесвирусы (человеческий цитомегаловирус и цитомегаловирусы свиней, обезьян и грызунов); гамма-герпесвирусы (вирус Эпштейна-Бар (EBV), вирус болезни Марека, герпес Saimiri, герпесвирус Ateles, герпесвирус Sylvilagus, вирус герпеса морских свинок, вирус опухоли Lucke); семейство Adenoviridae, включающее в себя род Mastadenovirus (человеческие подгруппы A, B, C, D и E и несгруппированные; обезьяньи аденовирусы (по меньшей мере, 23 серотипа), инфекционный собачий гепатит и аденовирусы крупного рогатого скота, свиней, овец, лягушек и многих других видов, род Aviadenovirus (птичьи аденовирусы) и некультивируемые аденовирусы; семейство Papoviridae, включающее в себя род Papillomavirus (вирусы папилломы человека, вирусы папилломы коров, вирус папилломы кроликов Шоупа и различные патогенные вирусы папилломы других видов), род Polyomavirus (вирус полиомы, вакуолизирующий обезьяний вирус (SV-40), вакуолизирующий кроличий вирус (RKV), вирус K, вирус BK, вирус JC и другие вирусы полиомы приматов, такие как лимфотропный вирус папилломы); семейство Parvoviridae включающее в себя род аденоассоциированных вирусов, род Parvovirus (кошачий вирус панлейкопении, коровий парвовирус, собачий парвовирус, вирус алеутской болезни норок и т.п.). Наконец, ДНК-содержащие вирусы могут включать в себя такие вирусы, как хронические инфекционные нейропатические агенты (вирус CHINA).
В других осуществлениях для получения композиций и вакцинных композиций по изобретению соответственно применяют бактериальный пилин, субфрагмент бактериального пилина или гибридный белок, содержащий или бактериальный пилин или его субфрагмент. Примеры пилиновых белков включают в себя пилины, продуцируемые Escherichia coli, Haemophilus influenzae, Neisseria meningitidis, Neisseria gonorrhoeae, Caulobacter crescentus, Pseudomonas stutzeri и Pseudomonas aeruginosa. Аминокислотные последовательности пилиновых белков, пригодные для применения по настоящему изобретению, включают в себя последовательности, приведенные в протоколах GenBank AJ000636, AJ132364, AF229646, AF051814, AF051815 и X00981, полное описание которых включено сюда в качестве ссылки.
Бактериальные пилиновые белки до экспорта белков в бактериальную периплазму, как правило, процессируются с удалением N-концевых лидирующих последовательностей. Далее, как может понять специалист в данной области, бактериальные пилиновые белки, применяемые для композиций и вакцинных композиций по изобретению соответственно, как правило, не несут присутствующих от природы лидирующих последовательностей.
Один конкретный пример пилинового белка, пригодного для применения по настоящему изобретению, представляет собой P-пилин E. coli (протокол GenBank AF237482 (SEQ ID № 1)). Пример пилина E. coli типа 1, пригодного для применения по настоящему изобретению, представляет собой пилин с аминокислотной последовательностью, приведенной в протоколе GenBank P04128 (SEQ ID № 2), кодируемого нуклеиновой кислотой с нуклеотидной последовательностью, приведенной в протоколе GenBank M27603 (SEQ ID № 3). Полные описания данных протоколов GenBank включены сюда в качестве ссылки. Кроме того, зрелую форму указанного выше белка, как правило, можно применять для приготовления композиций и вакцинных композиций по изобретению соответственно.
Бактериальные пилины или субфрагменты пилинов, пригодные для применения в практическом применении настоящего изобретения, как правило, способны ассоциировать с формированием упорядоченных и повторяющихся массивов антигенов.
Способы получения пилей и пилеобразных структур in vitro известны в данной области. Например, Bullitt et al., Proc. Natl. Acad. Sci. USA 93: 12890-12895 (1996) описывают восстановление in vitro субъединиц P-пилей E. coli. Кроме того, Eshdat et al., J. Bacteriol. 148: 308-314 (1981) описывают способы, пригодные для диссоциирующих пилей E. coli типа 1 и восстановления пилей. Коротко данные способы состоят в следующем: пили диссоциируют посредством инкубации при 37°C в насыщенном гуанидингидрохлориде. Пилиновые белки затем очищают хроматографией, после чего формируют пилиновые димеры посредством диализа против 5 мМ трис(гидроксиметил)аминометангидрохлорида (pH 8,0). Eshdat et al. также обнаружили, что пилиновые димеры вновь собираются в форму пилей при диализе против 5 мМ трис(гидроксиметил)аминометана (pH 8,0), содержащего 5 мМ MgCl2.
Кроме того, с применением, например, традиционной генетической инженерии и способов модификации белков пилиновые белки можно модифицировать так, чтобы они содержали первый участок прикрепления, с которым через второй участок прикрепления связывается антиген или антигенная детерминанта. Альтернативно, антигены и антигенные детерминанты посредством второго участка прикрепления можно прямо связать с аминокислотными остатками, присутствующими в данных белках от природы. Данные модифицированные пилиновые белки затем можно применять в вакцинных композициях по изобретению.
Бактериальные пилиновые белки, применяемые для получения композиций и вакцинных композиций по изобретению соответственно, можно модифицировать способом, подобным способу, описанному здесь для HBcAg. Например, остатки цистеина и лизина можно или удалить или заменить другими аминокислотными остатками, а к данным белкам можно добавить первые участки прикрепления. Кроме того, пилиновые белки можно или экспрессировать в модифицированной форме или модифицировать химически после экспрессии. Подобным образом интактные пили можно отделить от бактерий и затем химически модифицировать.
В другом осуществлении пили или пилеобразные структуры выделяют из бактерий (например, E. coli) и применяют для получения композиций и вакцинных композиций по изобретению. Один пример пилей, пригодных для получения композиций и вакцинных композиций, представляет собой пиль E. coli типа 1, который сформирован из пилиновых мономеров с аминокислотной последовательностью, приведенной в SEQ ID № 2.
В данной области известен ряд способов выделения бактериальных пилей. Например, Bullitt and Makowski (Biophys. J. 74: 623-632 (1998)) описывают способ очистки пилей для выделения P-пилей из E. coli. По данному способу пили срезают с гиперпилированной E. coli, содержащей плазмиду с P-пилем, и очищают посредством циклов солюбилизации и осаждения в MgCl2 (1,0 M).
После выделения пили или пилеобразные структуры можно модифицировать множеством способов. Например, к пилям можно добавлять первый участок прикрепления, к которому посредством второго участка прикрепления могут прикрепляться антигены или антигенные детерминанты. Другими словами, бактериальные пили или пилеобразные структуры можно выделить и модифицировать для получения упорядоченных и повторяющихся массивов антигенов.
Антигены и антигенные детерминанты можно связать с природными цистеиновыми или лизиновыми остатками, находящимися в пилях или пилеобразных структурах. В таких случаях высокий порядок и повторяемость природного аминокислотного остатка могла бы направлять связывание антигенов или антигенных детерминант с пилями или пилеобразными структурами. Например, пили или пилеобразные структуры можно связать со вторым участком прикрепления антигенов или антигенных детерминант с применением гетеробифункционального сшивающего средства.
Когда структуры, которые синтезируются организмами в природе (например, пили), применяют для получения композиций и вакцинных композиций по изобретению, часто выгодно генетически модифицировать данные организмы так, чтобы они продуцировали структуры, обладающие желательными характеристиками. Например, когда применяют пили E. coli типа 1, E. coli, из которых выделяют данные пили, можно модифицировать так, чтобы продуцировать структуры со специфическими характеристиками. Примеры возможных модификаций пилиновых белков включают в себя вставку одного или нескольких лизиновых остатков, удаление или замену одного или нескольких природных лизиновых остатков и уделение или замену одного или нескольких природных цистеиновых остатков (например, цистеиновых остатков в позициях 44 и 84 в SEQ ID № 2).
Кроме того, пилиновые гены можно подвергать дополнительным модификациям, которые приводят к экспрессии продуктов, содержащих первый участок прикрепления, отличный от лизинового остатка (например, домен FOS или JUN). Конечно, подходящие первые участки прикрепления, как правило, ограничены теми, которые не мешают пилину формировать пили или пилеобразные структуры, пригодные для применения в вакцинных композициях по изобретению.
Пилиновые гены, находящиеся в природе в бактериальных клетках, можно модифицировать in vivo (например, посредством гомологической рекомбинации), или в данные клетки можно внести пилиновые гены с конкретными характеристиками. Например, пилиновые гены можно ввести в бактериальные клетки как компонент или реплицирующегося клонирующего вектора, или вектора, который интегрирует в бактериальную хромосому. Внесенные пилиновые гены можно также связать с регулирующими экспрессию контрольными последовательностями (например, lac-оператор).
В большинстве случаев пили или пилеобразные структуры, применяемые в композициях и вакцинных композициях по изобретению соответственно, состоят из пилиновых субъединиц одного типа. Пили или пилеобразные структуры, состоящие из идентичных субъединиц, как правило, применяют, так как ожидают, что они сформируют структуры, представляющие собой высокоупорядоченные и повторяющиеся массивы антигенов.
Однако композиции по изобретению также включают в себя композиции или вакцины, содержащие пили или пилеобразные структуры, сформированные гетерогенными пилиновыми субъединицами. Пилиновые субъединицы, формирующие данные пили или пилеобразные структуры, можно экспрессировать с генов, находящихся в бактериальной клетке в природе, или можно внести в клетки. Когда природный пилиновый ген и внесенный ген оба экспрессированы в клетке, формирующей пили или пилеобразные структуры в результате, как правило, образуются структуры, сформированные из смеси данных пилиновых белков. Кроме того, когда в бактериальной клетке экспрессированы два или более пилиновых гена, относительная экспрессия каждого пилинового гена, как правило, является фактором, определяющим соотношение различных пилиновых субъединиц в пилях или пилеобразных структурах.
Когда желательны пили или пилеобразные структуры, обладающие конкретным составом смешанных пилиновых субъединиц, экспрессию, по меньшей мере, одного из пилиновых генов можно регулировать гетерологичным индуцибельным промотором. Такие промоторы, а также другие генетические элементы можно применять для регуляции относительных количеств различных пилиновых субъединиц, продуцируемых бактериальной клеткой и, следовательно, состава пилей или пилеобразных структур.
Кроме того, антиген или антигенную детерминанту можно связать с бактериальными пилями или пилеобразными структурами посредством связи, которая не является пептидной связью, бактериальные клетки, продуцирующие пили или пилеобразные структуры, применяемые в композициях по изобретению, можно генетически сконструировать для получения пилиновых белков, слитых с антигеном или антигенной детерминантой. Такие гибридные белки, формирующие пили или пилеобразные структуры, пригодны для применения в вакцинных композициях по изобретению.
Вирусоподобные частицы в контексте настоящей заявки относятся к структурам, подобным вирусной частице, но не являющимся патогенными. В общих чертах в вирусоподобных частицах отсутствует вирусный геном и, следовательно, они являются неинфекционными. Также вирусоподобные частицы можно получить в больших количествах посредством гетерологичной экспрессии, и их можно легко очистить.
В предпочтительном осуществлении коровая частица представляет собой вирусоподобную частицу, где вирусоподобная частица представляет собой рекомбинантную вирусоподобную частицу. Специалист в данной области может получить VLP с применением технологии рекомбинантных ДНК и кодирующих вирусы последовательностей, которые без труда доступны публично. Например, кодирующую последовательность оболочки вируса или корового белка можно сконструировать для экспрессии в бакуловирусном экспрессирующем векторе с применением коммерчески доступного бакуловирусного вектора под регуляторным контролем вирусного промотора с соответствующими модификациями последовательности, чтобы позволить функциональное связывание кодирующей последовательности с регуляторной последовательностью. Кодирующую последовательность оболочки вируса или корового белка также можно сконструировать для экспрессии, например, в бактериальном экспрессирующем векторе.
Примеры VLP включают в себя в качестве неограничивающих примеров капсидные белки вируса гепатита B (Ulrich, et al., Virus Res. 50: 141-182 (1998)), вирус кори (Warnes, et al., Gene 160: 173-178 (1995)), вирус синдбис, ротавирус (US 5071651 и US 5374426), вирус ящура (Twomey, et al., Vaccine 13: 1603-1610, (1995)), вирус Норфолка (Jiang, X., et al., Science 250: 1580-1583 (1990); Matsui, S. M., et al., J. Clin. Invest. 87: 1456-1461 (1991)), ретровирусный белок GAG (WO 96/30523), белок ретротранспозона Ty p1, поверхностный белок вируса гепатита B (WO 92/11291), вирус папилломы человека (WO 98/15631), РНК-фаги, Ty, fr-фаг, GA-фаг, AP205-фаг и Qβ-фаг.
Как легко увидят специалисты в данной области, VLP по изобретению не ограничена какой-либо конкретной формой. Частицу можно синтезировать химически или посредством биологических процессов, которые могут являться природными или неприродными. В качестве примера данный тип осуществления включает в себя вирусоподобную частицу или ее рекомбинантные формы.
В более конкретном осуществлении VLP может включать в себя или, альтернативно, по существу состоять из или, альтернативно, состоять из рекомбинантных полипептидов или их фрагментов, выбранных из рекомбинантных полипептидов ротавируса, рекомбинантных полипептидов вируса Норфолка, рекомбинантных полипептидов альфавируса, рекомбинантных полипептидов вируса ящура, рекомбинантных полипептидов вируса кори, рекомбинантных полипептидов вируса синдбис, рекомбинантных полипептидов вируса полиомы, рекомбинантных полипептидов ретровируса, рекомбинантных полипептидов вируса гепатита B (например, HBcAg), рекомбинантных полипептидов вируса табачной мозаики, рекомбинантных полипептидов вируса овечьих кошар, рекомбинантных полипептидов человеческого вируса папилломы, рекомбинантных полипептидов бактериофагов, рекомбинантных полипептидов РНК-фагов, рекомбинантных полипептидов Ty, рекомбинантных полипептидов фага fr, рекомбинантных полипептидов фага GA и рекомбинантных полипептидов фага Qβ. Вирусоподобная частица дополнительно может содержать или, альтернативно, по существу состоять из или, альтернативно, состоять из одного или нескольких фрагментов таких полипептидов, а также вариантов таких полипептидов. Варианты полипептидов могут обладать, например, по меньшей мере, 80%, 85%, 90%, 95%, 97% или 99% идентичности на уровне аминокислот с их копиями дикого типа.
В предпочтительном осуществлении вирусоподобная частица содержит, по существу состоит из или, альтернативно, состоит из рекомбинантных белков РНК-фага или их фрагментов. Предпочтительно РНК-фаг выбран из группы, состоящей из a) бактериофага Qβ; b) бактериофага R17; c) бактериофага fr; d) бактериофага GA; e) бактериофага SP; f) бактериофага MS2; g) бактериофага M11; h) бактериофага MX1; i) бактериофага NL95; k) бактериофага f2; 1) бактериофага PP7 и m) бактериофага AP205.
В другом предпочтительном осуществлении настоящего изобретения вирусоподобная частица содержит или, альтернативно, по существу состоит из или, альтернативно, состоит из рекомбинантных белков РНК-бактериофага Qβ или РНК-бактериофага fr, или РНК-бактериофага AP205 или их частей.
В дополнительном предпочтительном осуществлении настоящего изобретения рекомбинантные белки включают в себя, или, альтернативно, по существу состоят из, или, альтернативно, состоят из белков оболочки РНК-фагов.
Белки оболочки РНК-фагов, формирующие капсиды или VLP или фрагменты белков оболочки бактериофагов, согласующиеся с самостоятельной сборкой в капсид или VLP, являются, следовательно, дополнительными предпочтительными осуществлениями настоящего изобретения. Белки оболочки бактериофага Qβ, например, можно рекомбинантно экспрессировать в E. coli. Далее, после такой экспрессии данные белки спонтанно формируют капсиды. Дополнительно, данные капсиды формируют структуру с собственной повторяющейся организацией.
Конкретные предпочтительные примеры белков оболочки бактериофагов, которые можно применять для получения композиций по изобретению, включают в себя белки оболочки РНК-бактериофагов, таких как бактериофаг Qβ (SEQ ID № 4; PIR Database, инвентарный номер № VCBPQβ, указывающий на CP Qβ и SEQ ID № 5; инвентарный номер № AAA16663, указывающий на белок A1 Qβ), бактериофаг R17 (SEQ ID № 6; инвентарный номер PIR № VCBPR7), бактериофаг fr (SEQ ID № 7; инвентарный номер PIR № VCBPFR), бактериофаг GA (SEQ ID № 8; инвентарный номер GenBank № NP-040754), бактериофаг SP (SEQ ID № 9; инвентарный номер GenBank № CAA30374, указывающий на SP CP и SEQ ID № 10; Accession № NP 695026, указывающий на SP белка A1), бактериофаг MS2 (SEQ ID № 11; инвентарный номер PIR № VCBPM2), бактериофаг M11 (SEQ ID № 12; инвентарный номер GenBank № AAC06250), бактериофаг MX1 (SEQ ID № 13; инвентарный номер GenBank № AAC14699), бактериофаг NL95 (SEQ ID № 14; инвентарный номер GenBank № AAC14704), бактериофаг f2 (SEQ ID № 15; инвентарный номер GenBank № P03611), бактериофаг PP7 (SEQ ID № 16) и бактериофаг AP205 (SEQ ID № 28). Кроме того, в конструкцию капсида белков оболочки Qβ можно вносить белок A1 бактериофага Qβ (SEQ ID № 5) или укороченные на C-конце формы без 100, 150 или 180 аминокислот на его C-конце. Как правило, доля белка A1 Qβ относительно CP Qβ в конструкции капсида ограничена, для гарантии формирования капсида.
Также обнаружили, что белок оболочки Qβ самостоятельно собирается в капсиды при экспрессии в E. coli (Kozlovska TM. et al., GENE 137: 133-137 (1993)). Полученные капсиды или вирусоподобные частицы демонстрировали икосаэдрическую фагоподобную капсидную структуру с диаметром 25 нм и квазисимметрией T=3. Кроме того, разрешили кристаллическую структуру фага Qβ. Капсид содержит 180 копий белка оболочки, связанных в ковалентные пентамеры и гексамеры дисульфидными мостиками (Golmohammadi, R. et al., Structure 4: 543-5554 (1996)), что приводит к поразительной стабильности капсида из белка оболочки Qβ. Однако капсиды или VLP, полученные из рекомбинантного белка оболочки Qβ, могут содержать в капсиде субъединицы, не связанные дисульфидными связями с другими субъединицами или связанными частично. Однако в VLP, как правило, приблизительно более чем 80% субъединиц связаны друг с другом дисульфидными мостиками. Таким образом, после помещения рекомбинантного капсида Qβ на невосстанавливающий SDS-PAGE видны полосы, соответствующие мономерному белку оболочки Qβ, а также полосы, соответствующие гексамерному или пентамерному белку оболочки Qβ. Частично связанные дисульфидными связями субъединицы на невосстанавливающем SDS-PAGE могут выявляться как димерная, тримерная или даже тетрамерная полоса. Капсидный белок Qβ также демонстрирует необычную устойчивость к органическим растворителям и денатурирующим средствам. Неожиданно авторы изобретения обнаружили, что ДМСО и ацетонитрил в концентрациях до 30% и гуанидин в концентрациях до 1 М не влияют на стабильность капсида. Высокая стабильность капсидного белка оболочки Qβ представляет собой выгодную черту, в особенности для его применения при иммунизации и вакцинации млекопитающих и людей по настоящему изобретению.
При экспрессии в E. coli N-концевой метионин белка оболочки Qβ, как правило, удаляется, как наблюдали авторы при N-концевом секвенировании по Эдману, как описано у Stoll, E. et al. J. Biol. Chem. 252: 990-993 (1977). VLP, составленный из белков оболочки Qβ, где N-терминальный метионин не удален, или VLP, включающие в себя смесь белков оболочки Qβ, где N-терминальный метионин или отщеплен или присутствует - также входят в объем настоящего изобретения.
Дополнительные предпочтительные вирусоподобные частицы РНК-фагов, в особенности Qβ, по настоящему изобретению описаны в WO 02/056905, описание которого полностью включено сюда в качестве ссылки.
Показано, что дополнительные белки оболочки РНК-фагов также самостоятельно собираются после экспрессии в бактериальном хозяине (Kastelein, RA. et al., Gene 23: 245-254 (1983), Kozlovskaya, TM. et al., Dokl. Akad. Nauk SSSR 287: 452-455 (1986), Adhin, MR. et al., Virology 170: 238-242 (1989), Ni, CZ., et al., Protein Sci. 5: 2485-2493 (1996), Priano, C. et al., J. Mol. Biol. 249: 283-297 (1995)). Дополнительно к белку оболочки капсид фага Qβ содержит так называемый прочитанный до конца белок A1 и зрелый белок A2. A1 образуется посредством супрессии в стоп-кодоне UGA и обладает длиной 329 а.к. Капсид из рекомбинантного белка оболочки фага Qβ, применяемый по изобретению, лишен белка лизиса A2 и содержит РНК хозяина. Белок оболочки РНК-фагов представляет собой связывающий РНК белок и взаимодействует с петлей стебля рибосомсвязывающего участка гена репликазы, действуя как трансляционный репрессор в течение жизненного цикла вируса. Последовательность и структурные элементы взаимодействия известны (Witherell, GW. & Uhlenbeck, OC. Biochemistry 28: 71- 76 (1989); Lim F. et al., J Biol. Chem. 271: 31839-31845 (1996)). В общих чертах известно, что петля стебля и РНК вовлечены в сборку вируса (Golmohammadi, R. et al., Structure 4: 543-5554 (1996)).
В дополнительном предпочтительном осуществлении настоящего изобретения вирусоподобная частица содержит, или, альтернативно, по существу состоит из, или, альтернативно, состоит из рекомбинантных белков РНК-фага или их фрагментов, где рекомбинантные белки включают в себя, или, альтернативно, по существу состоят из, или, альтернативно, состоят из мутантных белков оболочки РНК-фага, предпочтительно - из мутантных белков оболочки указанных выше РНК-фагов. В другом предпочтительном осуществлении мутантные белки оболочки РНК-фагов модифицированы посредством удаления, по меньшей мере, одного или, альтернативно, по меньшей мере, двух лизиновых остатков посредством замещения, или посредством добавления, по меньшей мере, одного лизинового остатка посредством замещения; альтернативно, мутантные белки оболочки РНК-фагов модифицированы посредством удаления, по меньшей мере, одного или, альтернативно, по меньшей мере, двух лизиновых остатков, или посредством добавления, по меньшей мере, одного лизинового остатка посредством вставки. Удаление, замещение или добавление, по меньшей мере, одного лизинового остатка позволяет варьировать степень связывания, т.е. количество Aβ1-6 пептидов на субъединицу VLP РНК-фагов, в частности, для соответствия и приспосабливания к требованиям вакцины.
В другом предпочтительном осуществлении вирусоподобная частица содержит, или, альтернативно, по существу состоит из, или, альтернативно, состоит из рекомбинантных белков РНК-бактериофага Qβ или их фрагментов, где рекомбинантные белки включают в себя, или, альтернативно, по существу состоят из, или, альтернативно, состоят из белков оболочки с аминокислотной последовательностью SEQ ID № 4 или смеси белков оболочки с аминокислотными последовательностями SEQ ID № 4 и SEQ ID № 5 или мутантных SEQ ID № 5 и где N-концевой метионин предпочтительно отщеплен.
В дополнительном предпочтительном осуществлении настоящего изобретения вирусоподобная частица содержит, или, альтернативно, по существу состоит из, или, альтернативно, состоит из рекомбинантных белков Qβ или их фрагментов, где рекомбинантные белки включают в себя, или, альтернативно, по существу состоят из, или, альтернативно, состоят из мутантных белков оболочки Qβ. В другом предпочтительном осуществлении данные мутантные белки оболочки модифицированы посредством удаления, по меньшей мере, одного лизинового остатка посредством замещения или посредством добавления, по меньшей мере, одного лизинового остатка посредством замещения. Альтернативно данные мутантные белки оболочки модифицированы посредством удаления, по меньшей мере, одного лизинового остатка или посредством добавления, по меньшей мере, одного лизинового остатка посредством вставки.
На поверхности капсида из белка оболочки Qβ представлены четыре лизиновых остатка. По настоящему изобретению можно также применять мутантные Qβ, для которых представленные лизиновые остатки замещены на аргининовые остатки. Таким образом, в практическом применении данного изобретения можно применять следующие мутантные по белку оболочки Qβ или мутантные VLP Qβ: "Qβ-240" (Lys13-Arg; SEQ ID № 17), "Qβ-243" (Asn10-Lys; SEQ ID № 18), "Qβ-250" (Lys2-Arg, Lys13-Arg; SEQ ID № 19), "Qβ-251" (SEQ ID № 20) и "Qβ-259" (Lys2-Arg, Lys16-Arg; SEQ ID № 21). Таким образом, в дополнительном предпочтительном осуществлении настоящего изобретения вирусоподобная частица содержит, или, альтернативно, по существу состоит из, или, альтернативно, состоит из рекомбинантных белков мутантных белков оболочки Qβ, включающих в себя белки с аминокислотной последовательностью, выбранной из группы a) аминокислотной последовательности SEQ ID № 17; b) аминокислотной последовательности SEQ ID № 18; c) аминокислотной последовательности SEQ ID № 19; d) аминокислотной последовательности SEQ ID № 20 и e) аминокислотной последовательности SEQ ID № 21. Структура, экспрессия и очистка указанных выше белков оболочки Qβ, VLP и капсиды из мутантных белков оболочки Qβ соответственно описаны в WO 02/056905. В особенности это относится к примеру 18 указанной выше заявки.
В дополнительном предпочтительном осуществлении настоящего изобретения вирусоподобная частица содержит, или, альтернативно, по существу состоит из, или, альтернативно, состоит из рекомбинантных белков Qβ или их фрагментов, где рекомбинантные белки включают в себя, по существу состоят из или, альтернативно, состоят из смеси одного из указанных ранее мутантных Qβ и соответствующего белка A1.
В дополнительном предпочтительном осуществлении настоящего изобретения вирусоподобная частица содержит, или, альтернативно, по существу состоит из, или, альтернативно, состоит из рекомбинантных белков РНК-фага AP205 или их фрагментов.
Геном AP205 состоит из белка созревания, белка оболочки, репликазы и двух открытых рамок считывания, не существующих в родственных фагах - гена лизиза и открытой рамки считывания играющей роль в трансляции гена созревания (Klovins, J., et al., J. Gen. Virol. 83: 1523-33 (2002)). Белок оболочки AP205 можно экспрессировать с плазмиды pAP283-58 (SEQ ID № 27), являющейся производной pQb10 (Kozlovska, T. M. et al., Gene 137: 133-37 (1993)) и содержащей рибосомсвязывающий участок AP205. Альтернативно белок оболочки AP205 можно клонировать в pQb185, ниже рибосомсвязывающего участка, присутствующего в векторе. Оба подхода приводят к экспрессии белка и формированию капсидов, как описано в совместно рассматриваемой предварительной патентной заявке США с заголовком "Молекулярные антигенные массивы", поданной правопреемником настоящего изобретения 17 июля 2002, полностью включенной сюда в качестве ссылки. Векторы pQb10 и pQb185 представляют собой векторы, полученные из вектора pGEM, а экспрессия клонированных генов в данных векторах находится под контролем промотора trp (Kozlovska, T. M. et al., Gene 137: 133-37 (1993)). Плазмида pAP283-58 (SEQ ID № 27) содержит предполагаемый рибосомсвязывающий участок AP205 в следующей последовательности, находящейся ниже участка XbaI, и непосредственно выше стартового кодона ATG белка оболочки AP205: tctagaATTTTCTGCGCACCCATCCCGGGTGGCGCCCAAA-GTGAGGAAAATCACatg (SEQ ID № 57). Вектор pQb185 содержит последовательность Шайна-Дальгарно ниже участка XbaI и выше стартового кодога (tctagaTTAACCCAACGCGTAGGAGTCAGGCCatg (SEQ ID № 58), последовательность Шайна-Дальгарно подчеркнута).
В дополнительном предпочтительном осуществлении настоящего изобретения вирусоподобная частица содержит, или, альтернативно, по существу состоит из, или, альтернативно, состоит из рекомбинантных белков оболочки РНК-фага AP205 или их фрагментов.
Таким образом, данное предпочтительное осуществление настоящего изобретения включает в себя формирующие капсиды белки оболочки AP205. Такие белки экспрессируют рекомбинантно или получают из натуральных источников. Белки оболочки AP205, продуцированные в бактерии, спонтанно формируют капсиды, как доказано электронной микроскопией (EM) и иммунодиффузией. Структурные свойства капсида, сформированного белком оболочки AP205 (SEQ ID № 28) и структурные свойства капсида, сформированного белком оболочки РНК-фага AP205, при просмотре с применением EM почти не отличаются. VLP AP205 являются высокоиммуногенными, и их можно связать с антигенами и/или антигенными детерминантами для получения вакцинных конструкций представляющих антигены и/или антигенные детерминанты расположенными повторяющимся образом. Высокие титры, образующиеся против представленных таким образом антигенов, демонстрируют, что связанные антигены и/или антигенные детерминанты доступны для взаимодействия с молекулами антител и являются иммуногенными.
В дополнительном предпочтительном осуществлении настоящего изобретения вирусоподобная частица содержит, или, альтернативно, по существу состоит из, или, альтернативно, состоит из рекомбинантных мутантных белков оболочки РНК-фага AP205 или их фрагментов.
Пригодные для сборки мутантные формы VLP AP205, включающие в себя белки оболочки AP205 с заменой пролина в положении 5 на треонин (SEQ ID № 29), также можно применять для практического применения настоящего изобретения, и они приводят к дополнительному предпочтительному осуществлению изобретения. Данные VLP, VLP AP205, полученные из природных источников, или вирусные частицы AP205 можно связывать с антигенами для получения упорядоченных повторяющихся массивов антигенов по настоящему изобретению.
Мутантный белок оболочки AP205 P5-T можно экспрессировать с полученной непосредственно из pQb185 плазмиды pAP281-32 (SEQ ID № 30), содержащей ген мутантного белка оболочки AP205 вместо гена мутантного белка оболочки Qβ. Векторы для экспрессии белка оболочки AP205 трансфицируют в E. coli для экспрессии белка оболочки AP205.
Способы для экспрессии белка оболочки и мутантного белка оболочки соответственно, приводящие к самостоятельной сборке в VLP, описаны в примерах 20 и 21. Пригодные штаммы E. coli включают в себя в качестве неограничивающих примеров K802, JM 109, RR1 E. coli. Пригодные векторы и штаммы и их сочетания можно определить тестированием экспрессии белка оболочки и мутантного белка оболочки соответственно посредством SDS-PAGE и формированием капсида и сборки посредством необязательной первой очистки капсидов гель-фильтрацией и последующего тестирования их в анализе иммунодиффузии (Ouchterlony test) или электронной микроскопией (Kozlovska, T. M. et al., Gene 137: 133-37 (1993)).
У белков оболочки AP205, экспрессированных с векторов pAP283-58 и pAP281-32, может отсутствовать начальная аминокислота - метионин, вследствие процессинга в цитоплазме E. coli. Расщепленные, нерасщепленные формы VLP AP205 или их смеси представляют собой дополнительные предпочтительные осуществления изобретения.
В дополнительном предпочтительном осуществлении настоящего изобретения вирусоподобная частица содержит, или, альтернативно, по существу состоит из, или, альтернативно, состоит из смеси рекомбинантных белков оболочки РНК-фага AP205 или их фрагментов и рекомбинантных мутантных белков оболочки РНК-фага AP205 или их фрагментов.
В дополнительном предпочтительном осуществлении настоящего изобретения вирусоподобная частица содержит, или, альтернативно, по существу состоит из, или, альтернативно, состоит из фрагментов рекомбинантных белков оболочки или рекомбинантных мутантных белков оболочки РНК-фага AP205.
Фрагменты рекомбинантных белков оболочки AP205, способные к сборке в VLP или капсид соответственно, также пригодны в практическом применении данного изобретения. Данные фрагменты можно получить посредством удаления или внутренней части, или концов белка оболочки и мутантного белка оболочки соответственно. Вставки в последовательности белка оболочки и мутантного белка оболочки или слияния антигенных последовательностей с последовательностями белка оболочки и мутантного белка оболочки, совместимые со сборкой в VLP, представляют собой дополнительные осуществления изобретения и дают в результате гибридные белки оболочки AP205 и частицы соответственно. Результат вставок, удалений и слияний с последовательностью белка оболочки и совместим ли он со сборкой в VLP, можно определить посредством электронной микроскопии.
Сформированные описанными выше белком оболочки AP205, фрагментами белка оболочки или гибридными белками оболочки частицы, можно выделить в чистом виде посредством объединения стадий фракционирования посредством преципитации и стадий очистки посредством гель-фильтрации с применением, например, колонок с сефарозой CL-4B, сефарозой CL-2B, сефарозой CL-6B и их сочетаний. В данной области известны другие способы выделения вирусоподобных частиц, и их можно применять для выделения вирусоподобных частиц (VLP) бактериофага AP205. Например, в патенте США № 4918166 полностью включенном сюда в качестве ссылки, описано применение ультрацентрифугирования для выделения VLP дрожжевого ретротранспозона Ty.
Определена кристаллическая структура некоторых РНК-бактериофагов (Golmohammadi, R. et al., Structure 4: 543-554 (1996)). С применением такой информации можно идентифицировать представленные на поверхности остатки и, таким образом, можно модифицировать белки оболочки РНК-фагов так, чтобы посредством вставки или замещения являлось возможным вставить один или несколько химически активных аминокислотных остатков. Как следствие, такие модифицированные формы белков оболочки бактериофагов также можно применять для настоящего изобретения. Таким образом, варианты белков, формирующих капсиды или подобные капсидам структуры (например, белки оболочки бактериофага Qβ, бактериофага R17, бактериофага fr, бактериофага GA, бактериофага SP, бактериофага AP205 и бактериофага MS2) также можно применять для получения композиций по настоящему изобретению.
Хотя последовательность вариантов белков, обсуждаемых выше, отличается от их копий дикого типа, данные варианты белков, как правило, сохраняют способность формировать капсиды или подобные капсидам структуры. Таким образом, изобретение дополнительно включает в себя композиции и вакцинные композиции соответственно, дополнительно включающие в себя варианты белков, формирующие капсиды или подобные капсидам структуры, а также способы получения таких композиций и вакцинных композиций соответственно отдельных белковых субъединиц, применяемых для получения таких композиций и молекул нуклеиновых кислот, кодирующих данные белковые субъединицы. Таким образом, в объем изобретения включены вариантные формы белков дикого типа, формирующие капсиды или подобные капсидам структуры и сохраняющие способность ассоциировать и формировать капсиды или подобные капсидам структуры.
Как результат, изобретение дополнительно включает в себя композиции и вакцинные композиции соответственно, включающие в себя белки, содержащие, или, альтернативно, по существу состоящие из, или, альтернативно, состоящие из аминокислотных последовательностей, которые, по меньшей мере, на 80%, 85%, 90%, 95%, 97% или 99% идентичны белкам дикого типа, формирующим упорядоченные массивы и обладающие собственной повторяющейся структурой соответственно.
Дополнительно в объем изобретения включены молекулы нуклеиновых кислот, кодирующие белки, применяемые для получения композиций по настоящему изобретению.
В других осуществлениях изобретение дополнительно включает в себя композиции, содержащие белки, включающие в себя, или, альтернативно, по существу состоящие из, или, альтернативно, состоящие из аминокислотных последовательностей, по меньшей мере, на 80%, 85%, 90%, 95%, 97% или 99% идентичных любой из аминокислотных последовательностей, представленных в SEQ ID №№: 4-21.
Белки, пригодные для применения по настоящему изобретению, также включают в себя укороченные на C-конце мутанты белков, формирующие капсид или подобные капсиду структуры или VLP. Конкретные примеры таких укороченных мутантов включают в себя белки с аминокислотной последовательностью, представленной в любой из SEQ ID №№: 4-21, где с C-конца удалены 1, 2, 5, 7, 9, 10, 12, 14, 15 или 17 аминокислот. Как правило, данные укороченные на C-конце мутанты сохраняют способность формировать капсиды или подобные капсидам структуры.
Дополнительно белки, пригодные для применения по настоящему изобретению, также включают в себя укороченные на N-конце мутанты белков, формирующих капсиды или подобные капсидам структуры. Конкретные примеры таких укороченных мутантов включают в себя белки с аминокислотной последовательностью, представленной в любой из SEQ ID №№: 4-21, где с N-конца удалены 1, 2, 5, 7, 9, 10, 12, 14, 15 или 17 аминокислот. Как правило, данные укороченные на N-конце мутанты сохраняют способность формировать капсиды или подобные капсидам структуры.
Дополнительные белки, пригодные для применения по настоящему изобретению, также включают в себя укороченные на N- и C-конце мутанты белков, формирующих капсиды или подобные капсидам структуры. Пригодные укороченные мутанты включают в себя белки с аминокислотной последовательностью, представленной в любой из SEQ ID №№: 4-21, где 1, 2, 5, 7, 9, 10, 12, 14, 15 или 17 аминокислот удалены с N-конца и 1, 2, 5, 7, 9, 10, 12, 14, 15 или 17 аминокислот удалены с C-конца. Как правило, данные укороченные на N-конце и на C-конце мутанты сохраняют способность формировать капсиды или подобные капсидам структуры.
Изобретение дополнительно включает в себя белки, включающие в себя, или, альтернативно, по существу состоящие из, или, альтернативно, состоящие из аминокислотных последовательностей, по меньшей мере, на 80%, 85%, 90%, 95%, 97% или 99% идентичных описанным выше укороченным мутантам.
Таким образом, изобретение включает в себя композиции и вакцинные композиции, полученные из белков, формирующих капсиды или VLP, способы получения данных композиций из отдельных белковых субъединиц и VLP или капсидов, способы получения данных отдельных белковых субъединиц, молекул нуклеиновых кислот, кодирующих данные субъединицы, и способы вакцинирования и/или вызова иммунологических ответов у индивидов с применением данных композиций по настоящему изобретению.
В одном осуществлении изобретение относится к вакцинной композиции по изобретению, дополнительно содержащей адъювант. В другом осуществлении в вакцинной композиции адъювант отсутствует. В дополнительном осуществлении вакцинная композиция содержит коровую частицу по изобретению, где коровая частица содержит, а предпочтительно является вирусоподобной частицей, где предпочтительно указанная вирусоподобная частица представляет собой рекомбинантную вирусоподобную частицу. Предпочтительно вирусоподобная частица содержит, или, альтернативно, по существу состоит из, или, альтернативно, состоит из рекомбинантных белков РНК-фага или их частей, предпочтительно из белков оболочки РНК-фагов. В предпочтительном осуществлении белок оболочки РНК-фагов обладает аминокислотной последовательностью выбранной из группы, состоящей из:(a) SEQ ID № 4; (b) смеси SEQ ID № 4 и SEQ ID № 5; (c) SEQ ID № 6; (d) SEQ ID № 7; (e) SEQ ID № 8; (f) SEQ ID № 9; (g) смеси SEQ ID № 9 и SEQ ID № 10; (h) SEQ ID № 11; (i) SEQ ID № 12; (k) SEQ ID № 13; (1) SEQ ID № 14; (m) SEQ ID № 15; (n) SEQ ID № 16 и (o) SEQ ID № 28. Альтернативно рекомбинантные белки вирусоподобной частицы вакцинной композиции по изобретению содержат, или, альтернативно, по существу состоят из, или, альтернативно, состоят из мутантных белков оболочки РНК-фагов, где указанный РНК-фаг выбран из группы, состоящей из (a) бактериофага Qβ; (b) бактериофага R17; (c) бактериофага fr; (d) бактериофага GA; (e) бактериофага SP; (f) бактериофага MS2; (g) бактериофага M11; (h) бактериофага MX1; (i) бактериофага NL95; (k) бактериофага f2; (1) бактериофага PP7; и (m) бактериофага AP205.
В предпочтительном осуществлении мутантные белки оболочки указанного РНК-фага модифицируют посредством удаления или путем добавления, по меньшей мере, одного лизинового остатка посредством замещения. В другом предпочтительном осуществлении мутантные белки оболочки указанного РНК-фага модифицируют посредством удаления, по меньшей мере, одного лизинового остатка или путем добавления, по меньшей мере, одного лизинового остатка посредством вставки. В предпочтительном осуществлении вирусоподобная частица содержит рекомбинантные белки РНК-фага Qβ, или, альтернативно, РНК-фага fr, или РНК-фага AP205 или их фрагменты.
Как указано ранее, изобретение включает в себя вирусоподобные частицы или их рекомбинантные формы. В одном предпочтительном осуществлении применяемые в композициях по изобретению частицы состоят из корового белка вируса гепатита B (HBcAg) или фрагмента HBcAg. В дополнительном осуществлении применяемые в композициях по изобретению частицы состоят из корового белка вируса гепатита B (HBcAg) или фрагмента HBcAg, модифицированных или удалением или восстановлением ряда свободных цистеиновых остатков. Zhou et al. (J. Virol. 66: 5393-5398 (1992)) продемонстрировали, что HBcAg, модифицированные удалением природных цистеиновых остатков, сохраняют способность к ассоциации и формированию капсидов. Таким образом, VLP, пригодные для применения в композициях по изобретению, включают в себя VLP, содержащие модифицированные HBcAg или их фрагменты, в которых один или несколько природных цистеиновых остатков или удалены, или замещены другим аминокислотным остатком (например, сериновым остатком).
HBcAg представляет собой белок, образуемый при процессинге белка-предшественника корового антигена гепатита B. Ряд изотипов HBcAg идентифицированы, и их аминокислотные последовательности легко доступны специалистам в данной области. В большинстве случаев композиции и вакцинные композиции по изобретению соответственно получают с применением процессированной формы HBcAg (т.е., HBcAg, из которого удалена N-концевая лидирующая последовательность белка-предшественника корового антигена гепатита B).
Кроме того, когда HBcAg получают в условиях, где не происходит процессинг, HBcAg, как правило, экспрессируют в "процессированной" форме. Например, когда для получения HBcAg по изобретению применяют экспрессирующую систему E. coli, направляющую экспрессию белка в цитоплазму, данные белки, как правило, экспрессируют так, чтобы N-концевая лидирующая последовательность белка-предшественника корового антигена гепатита B не была представлена.
Получение подобных вирусу гепатита B частиц, которые можно применять для настоящего изобретения, описано, например, в WO 00/32227, а там, в частности, в примерах с 17 по 19 и с 21 по 24, а также в WO 01/85208, а там, в частности, в примерах с 17 по 19, с 21 по 24, с 31 по 41 и в WO 02/056905. Для последней заявки на него, в частности, ссылаются в примерах 23, 24, 31 и 51. Все три документа полностью включены сюда в качестве ссылки.
Настоящее изобретение также включает в себя варианты HBcAg, модифицированные удалением или замещением одного или нескольких дополнительных цистеиновых остатков. В данной области известно, что свободные цистеиновые остатки могут вовлекаться в ряд побочных химических реакций. Данные побочные реакции включают в себя дисульфидные обмены, реакции с химическими веществами или метаболитами, которые, например, вводят или формируют при сочетанной с другими веществами терапии, или прямое окисление и реакцию с нуклеотидами при экспозиции в UV-свете. Таким образом, могут образоваться токсические продукты, особенно принимая во внимание факт того, что HBcAg обладает сильной тенденцией связывать нуклеиновые кислоты. Таким образом, токсические продукты будут распределены между многообразием молекул, которые индивидуально могут присутствовать в малых концентрациях, но достигать токсических уровней вместе.
Ввиду указанного выше одним из преимуществ применения в вакцинных композициях HBcAg, модифицированного удалением природных цистеиновых остатков, является то, что уменьшено количество участков, с которыми при ассоциации антигенов или антигенных детерминант могут связываться токсические молекулы, или они все удалены.
Определен ряд встречающихся в природе вариантов HBcAg, пригодных для практического применения настоящего изобретения. Например, Yuan et al., (J. Virol. 73: 10122-10128 (1999)) описывают варианты, в которых изолейциновый остаток в позиции, соответствующей позиции 97 в SEQ ID № 22, замещен или лейциновым остатком или фенилаланиновым остатком. Аминокислотные последовательности ряда вариантов HBcAg, а также некоторые варианты предшественников корового антигена гепатита B описаны в протоколах GenBank AAF121240, AF121239, X85297, X02496, X85305, X85303, AF151735, X85259, X85286, X85260, X85317, X85298, AF043593, M20706, X85295, X80925, X85284, X85275, X72702, X85291, X65258, X85302, M32138, X85293, X85315, U95551, X85256, X85316, X85296, AB033559, X59795, X85299, X85307, X65257, X85311, X85301 (SEQ ID № 23), X85314, X85287, X85272, X85319, AB010289, X85285, AB010289, AF121242, M90520 (SEQ ID № 24), P03153, AF110999 и M95589, описания каждого из которых включены сюда в качестве ссылки. Последовательности указанных выше вариантов предшественника корового антигена гепатита B дополнительно описаны в WO 01/85208 в SEQ ID №№: 89-138. Данные варианты HBcAg в аминокислотной последовательности отличаются по ряду позиций, включающих в себя аминокислотные остатки, соответствующие аминокислотным остаткам, расположенным в позициях 12, 13, 21, 22, 24, 29, 32, 33, 35, 38, 40, 42, 44, 45, 49, 51, 57, 58, 59, 64, 66, 67, 69, 74, 77, 80, 81, 87, 92, 93, 97, 98, 100, 103, 105, 106, 109, 113, 116, 121, 126, 130, 133, 135, 141, 147, 149, 157, 176, 178, 182 и 183 в SEQ ID № 25. Дополнительно варианты HBcAg, которые пригодны для применения в композициях по настоящему изобретению и которые можно дополнительно модифицировать согласно описанию данной спецификации, описаны в WO 00/198333, WO 00/177158 и WO 00/214478.
Как отмечено выше, в композициях или вакцинных композициях по настоящему изобретению соответственно применяют, как правило, процессированные HBcAg (т.е. те, в которых отсутствуют лидирующие последовательности). Настоящее изобретение включает в себя вакцинные композиции, а также способы применения данных композиций, в которых применяют описанные выше варианты HBcAg.
Обладает ли аминокислотная последовательность полипептида аминокислотной последовательностью, которая, по меньшей мере, на 80%, 85%, 90%, 95%, 97% или 99% идентична одной из указанных выше аминокислотных последовательностей дикого типа или ее части, можно условно определить с применением известных компьютерных программ, таких как программа Bestfit. При применении Bestfit или другой программы выравнивания последовательностей для определения того, обладает ли конкретная последовательность, например, 95% идентичности с референсной аминокислотной последовательностью, параметры устанавливают так, чтобы процент идентичности рассчитывался по полной длине референсной аминокислотной последовательности и чтобы в референсной последовательности допускались пропуски в гомологии до 5% общего числа аминокислотных остатков.
Аминокислотные последовательности указанных выше вариантов HBcAg и предшественники относительно сходны друг с другом. Таким образом, ссылка на аминокислотный остаток варианта HBcAg, расположенный в позиции, соответствующей конкретной позиции в SEQ ID № 25, относится к аминокислотному остатку, находящемуся в данной позиции в аминокислотной последовательности, приведенной в SEQ ID № 25. Среди вирусов гепатита B, инфицирующих млекопитающих, гомология между данными вариантами HBcAg в целом является достаточно высокой, так что специалист в данной области не испытает больших затруднений, рассматривая и аминокислотную последовательность, приведенную в SEQ ID № 25, и аминокислотную последовательность конкретного HBcAg, и идентифицируя "соответствующие" аминокислотные остатки. Кроме того, приведенная в SEQ ID № 24 аминокислотная последовательность HBcAg, демонстрирующая аминокислотную последовательность HBcAg, полученного из вируса, инфицирующего лесного североамериканского сурка, обладает достаточной гомологией с аминокислотной последовательностью HBcAg, приведенной в SEQ ID № 25, так что легко понять, что три вставленных аминокислотных остатка присутствуют в SEQ ID № 23 между аминокислотными остатками 155 и 156 SEQ ID № 25.
Изобретение также включает в себя вакцинные композиции, содержащие варианты HBcAg вирусов гепатита B, инфицирующие птиц, а также вакцинные композиции, содержащие фрагменты данных вариантов HBcAg. Для данных вариантов HBcAg один, два, три или более цистеиновых остатков, присутствующих в природе в данных полипептидах, могут являться или замещенными другим аминокислотным остатком, или удаленными до их включения в вакцинные композиции по изобретению.
Как обсуждалось выше, удаление свободных цистеиновых остатков уменьшает количество участков, где с HBcAg могут связываться токсические компоненты, а также удаляет участки, где может происходить сшивание лизиновых и цистеиновых остатков той же или соседних молекул HBcAg. Следовательно, в другом осуществлении настоящего изобретения один или несколько цистеиновых остатков капсидного белка вируса гепатита B или удалены или замещены другим аминокислотным остатком.
В других осуществлениях композиции и вакцинные композиции по изобретению соответственно содержат HBcAg, из которого удалена C-концевая область (например, аминокислотные остатки 145-185 или 150-185 из SEQ ID № 25). Таким образом, дополнительно модифицированные HBcAg, пригодные для применения в практическом осуществлении настоящего изобретения, включают в себя укороченные на C-конце мутанты. Пригодные укороченные мутанты включают в себя HBcAg, где с C-конца удалены 1, 5, 10, 15, 20, 25, 30, 34, 35 аминокислот.
Пригодные для применения в практическом осуществлении настоящего изобретения HBcAg также включают в себя укороченные на N-конце мутанты. Пригодные укороченные мутанты включают в себя модифицированные HBcAg, где с N-конца удалены 1, 2, 5, 7, 9, 10, 12, 14, 15 или 17 аминокислот.
Дополнительно пригодные для применения в практическом осуществлении настоящего изобретения HBcAg также включают в себя мутанты, укороченные на N- и C-конце. Пригодные укороченные мутанты включают в себя модифицированные HBcAg, где с N-конца удалены 1, 2, 5, 7, 9, 10, 12, 14, 15 или 17 аминокислот и с C-конца удалены 1, 5, 10, 15, 20, 25, 30, 34, 35 аминокислот.
Дополнительно изобретение включает в себя композиции и вакцинные композиции соответственно, содержащие полипептиды HBcAg, содержащие, или, альтернативно, по существу состоящие из, или, альтернативно, состоящие из аминокислотных последовательностей, по меньшей мере, на 80%, 85%, 90%, 95%, 97% или 99% идентичных описанным выше укороченным мутантам.
В определенных осуществлениях изобретения для опосредования связывания грелина или пептида грелина с VLP из HBcAg в полипептид HBcAg вводят лизиновый остаток. В предпочтительных осуществлениях композиции по изобретению получают с применением HBcAg содержащих, или, альтернативно, по существу состоящих из, или, альтернативно, состоящих из аминокислот 1-144 или 1-149, 1-185 из SEQ ID № 25, модифицированных так, что соответствующие положениям 79 и 80 аминокислоты замещены пептидом с аминокислотной последовательностью Gly-Gly-Lys-Gly-Gly (SEQ ID № 33), что приводит к полипептиду HBcAg с последовательностью, приведенной в SEQ ID № 26. В дополнительных предпочтительных осуществлениях цистеиновые остатки в положениях 48 и 107 в SEQ ID № 25 заменены на серин. Изобретение дополнительно включает в себя композиции, содержащие соответствующие полипептиды, с аминокислотными последовательностями, представленными в любом из указанных выше вариантов предшественника корового антигена гепатита B, которые также обладают указанными выше изменениями аминокислот. Кроме того, в объем изобретения входят дополнительные варианты HBcAg, способные к ассоциации с формированием капсида или VLP и обладающие указанными выше изменениями аминокислот. Таким образом, изобретение дополнительно включает в себя композиции и вакцинные композиции соответственно, содержащие полипептиды HBcAg, содержащие, или, альтернативно, по существу состоящие из, или, альтернативно, состоящие из аминокислотных последовательностей, по меньшей мере, на 80%, 85%, 90%, 95%, 97% или 99% идентичных любой из аминокислотных последовательностей дикого типа и формы данных белков, которые, где подходит, преобразованы удалением N-концевой лидирующей последовательности, и где проведены указанные выше изменения.
Композиции или вакцинные композиции по изобретению могут содержать смеси различных HBcAg. Таким образом, данные вакцинные композиции могут состоять из HBcAg, отличающихся по аминокислотной последовательности. Например, можно получить вакцинные композиции, содержащие HBcAg "дикого типа" и модифицированный HBcAg, в котором изменены один или более аминокислотных остатков (например, удалены, вставлены или замещены). Кроме того, предпочтительные вакцинные композиции по изобретению представляют собой вакцинные композиции, представляющие высокоупорядоченный и повторяющийся массив антигенов, где антиген представляет собой грелин или пептид грелина.
В дополнительном предпочтительном осуществлении настоящего изобретения, по меньшей мере, один грелин или пептид грелина связан с указанной коровой частицей и вирусоподобной частицей соответственно, по меньшей мере, одной ковалентной связью. Предпочтительно, по меньшей мере, один грелин или пептид грелина связан с коровой частицей или вирусоподобной частицей, по меньшей мере, одной ковалентной связью где указанная ковалентная связь, является непептидной связью, приводящей к упорядоченному и повторяющемуся массиву коровая частица - грелин и массиву или конъюгату грелин - VLP соответственно. Данный массив и конъюгат грелин - VLP соответственно, как правило и предпочтительно обладает повторяющейся и упорядоченной структурой, так как, по меньшей мере, один, но, как правило, более чем один грелин или пептид грелина связан с VLP и коровой частицей соответственно ориентированным образом. Предпочтительно с VLP связаны более чем 120, предпочтительно - более чем 180, предпочтительно - более чем 270 и предпочтитльно - более чем 360 грелинов или пептидов грелина. Формирование повторяющегося и ориентированного массива и конъюгата соответственно грелин - VLP и коровая частица соответственно обеспечивается ориентированным и направленным, а также определенным связыванием и присоединением соответственно по меньшей мере, одного грелина или пептида грелина с VLP и коровой частицей соответственно, как станет ясно в дальнейшем. Кроме того, типичная собственная высокоповторяющаяся и организованная структура VLP и коровых частиц соответственно преимущественно содействует представлению грелина или пептида грелина в высокоориентированной и повторяющейся форме, приводя к высокоорганизованному и повторяющемуся массиву или конъюгату грелин-VLP/коровая частица соответственно.
Следовательно, предпочтительные конъюгаты и массивы по изобретению соответственно отличаются от предыдущих конъюгатов в данной области их высокоорганизованной структурой, размерами и повторяемостью антигена на поверхности массива. Кроме того, предпочтительное осуществление данного изобретения допускает экспрессию и частицы, и антигена в экспрессирующем хозяине, гарантируя правильную укладку антигена, т.е. по меньшей мере, одного грелина или пептида грелина, и правильную укладку и сборку VLP.
Настоящее изобретение описывает способы связывания грелина или пептида грелина с коровыми частицами и VLP соответственно. Как указано в одном аспекте изобретения, грелин или пептид грелина связан с коровой частицей и VLP соответственно посредством образования поперечных химических связей, как правило и предпочтительно с применением гетеробифункционального сшивающего средства. В данной области известно несколько гетеробифункциональных сшивающих средств. В предпочтительных осуществлениях гетеробифункциональные сшивающие средства содержат функциональную группу, которая может реагировать с предпочтительными первыми участками прикрепления, т.е. с аминогруппой боковой цепи лизиновых остатков коровой частицы и VLP или, по меньшей мере, одной субъединицы VLP соответственно, и дополнительную функциональную группу, которая может реагировать с предпочтительным вторым участком прикрепления, т.е. цистеиновым остатком, присутствующим в природе, сделанным доступным для реакции посредством восстановления, или встроенным в грелин или пептид грелина и, необязательно, также сделанным доступным для реакции посредством восстановления. Первая стадия процедуры, как правило, называемой дериватизацией, представляет собой реакцию коровой частицы или VLP со сшивающим средством. Продукт данной реакции представляет собой активированную коровую частицу или активированную VLP, также называемую активированный носитель. На второй стадии непрореагировавшее сшивающее средство удаляют с применением обычных способов, таких как гель-фильтрация или диализ. На третьей стадии грелин или пептид грелина реагирует с активированным носителем и данную стадию, как правило, называют стадией связывания. Непрореагировавший грелин или пептид грелина необязательно можно удалить на четвертой стадии, например, посредством диализа. В данной области известно несколько гетеробифункциональных сшивающих средств. Они включают в себя предпочтительные сшивающие средства SMPH (Pierce), Sulfo-MBS, Sulfo-EMCS, Sulfo-GMBS, Sulfo-SIAB, Sulfo-SMPB, Sulfo-SMCC, SVSB, SIA и другие сшивающие средства, доступные, например, из Pierce Chemical Company (Rockford, IL, USA), с одной функциональной группой, реагирующей с аминогруппами и одной функциональной группой, реагирующей с цистеиновыми остатками. Все указанные выше сшивающие средства после реакции с аминогруппой приводят к формированию амидной связи, а после реакции с цистеином - тиоэфирной связи. Другой класс сшивающих средств, пригодных в практическом осуществлении изобретения, характеризуется введением дисульфидой связи между грелином или пептидом грелина и коровой частицы или VLP при связывании. Предпочтительные сшивающие средства, принадлежащие к данному классу, включают в себя, например, SPDP и Sulfo-LC-SPDP (Pierce). На степень дериватизации коровой частицы и VLP соответственно сшивающим средством можно влиять изменением экспериментальных условий, таких как концентрация каждого из реагирующих партнеров, избыток одного реагента по сравнению с другим, pH, температура и ионная сила. Для соответствия требованиям вакцины степень связывания, т.е. количество грелина или пептида грелина на субъединицу коровой частицы и VLP соответственно, можно регулировать изменением описанных выше экспериментальных условий. Растворимость грелина или пептида грелина может накладывать ограничение на количество грелина или пептида грелина, которое может связываться с каждой субъединицей и в тех случаях, когда полученная вакцина может являться нерастворимой, уменьшение количества грелина или пептида грелина на субъединицу является выгодным.
Особенно предпочтительный способ связывания грелина или пептида грелина с коровой частицей и VLP соответственно представляет собой связывание лизинового остатка на поверхности коровой частицы и VLP соответственно с цистеиновым остатком на грелине или пептиде грелина. Таким образом, в предпочтительном осуществлении настоящего изобретения первый участок прикрепления представляет собой лизизовый остаток, а второй участок прикрепления представляет собой цистеиновый остаток. В некоторых осуществлениях для связывания с коровой частицей или VLP соответственно может потребоваться конструирование содержащего в качестве второго участка прикрепления или его части цистеиновый остаток аминокислотного линкера с грелином или пептидом грелина. Альтернативно цистеин можно ввести или вставкой или мутацией в пределах грелина или пептида грелина. Альтернативно цистеиновый остаток можно ввести посредством химического связывания.
Выбор аминокислотного линкера зависит от природы антигена и аутоантигена соответственно, т.е. от природы грелина или пептида грелина, от его биохимических свойств, таких как pI, распределение заряда и гликозилирование. Как правило, предпочтительными являются гибкие аминокислотные линкеры. Предпочтительные осуществления аминокислотных линкеров выбирают из группы, состоящей из (a) CGG; (b) N-концевой гамма-1-линкер; (c) N-концевой гамма-3-линкер; (d) области шарнира Ig; (e) N-концевые глициновые линкеры; (f) (G)kC(G)n с n=0-12 и k=0-5 (SEQ ID № 34); (g) N-концевые глицин-сериновые линкеры; (h) (G)kC(G)m(S)i(GGGGS)n с n=0-3, k=0-5, m=0-10, l=0-2 (SEQ ID № 35); (i) GGC; (k) GGC-NH2; (1) C-концевой гамма-1-линкер; (m) C-концевой гамма-3-линкер; (n) C-концевые глициновые линкеры; (o) (G)nC(G)k с n=0-12 и k=0-5 (SEQ ID № 36); (p) C-концевые глицин-сериновые линкеры; (q) (G)m(S)i(GGGGS)n(G)oC(G)k с n=0-3, k=0-5, m=0-10, l=0-2 и o=0-8 (SEQ ID № 37).
Дополнительными предпочтительными примерами аминокислотных линкеров являются шарнирная область иммуноглобулинов, глицин-сериновые линкеры (GGGGS)n (SEQ ID № 38) и глициновые линкеры (G)n, все дополнительно содержащие цистеиновый остаток в качестве второго участка прикрепления и, необязательно, дополнительные глициновые остатки. Как правило, предпочтительные примеры указанных аминокислотных линкеров представляют собой N-концевые гамма-1: CGDKTHTSPP (SEQ ID № 39); C-концевые гамма-1: DKTHTSPPCG (SEQ ID № 40); N-концевые гамма-3: CGGPKPSTPPGSSGGAP (SEQ ID № 41); Сконцевые гамма-3: PKPSTPPGSSGGAPGGCG (SEQ ID № 42); N-концевой глициновый линкер: GCGGGG (SEQ ID № 43); C-концевой глициновый линкер: GGGGCG (SEQ ID № 44); C-концевой глицин-лизиновый линкер: GGKKGC (SEQ ID № 45); N-концевой глицин-лизиновый линкер: CGKKGG (SEQ ID № 46).
В дополнительном предпочтительном осуществлении настоящего изобретения, и особенно если антиген представляет собой пептид грелина, предпочтительными в качестве аминокислотных линкеров являются линкеры GGCG (SEQ ID № 47), GGC или GGC-NH2 ("NH2" стоит для амидирования) на C-конце пептида или линкеры CGG на его N-конце. Как правило, глициновые остатки вставляют между объемными аминокислотами и цистеином для применения в качестве второго участка прикрепления во избежание потенциальной помехи более объемной аминокислоты в реакции связывания.
Цистеиновый остаток, присутствующий на грелине или пептиде грелина, для реакции с гетеробифункциональным сшивающим средством на активированной VLP должен находиться в восстановленном состоянии, то есть свободный цистеин или цистеиновый остаток со свободной сульфгидрильной группой должен являться доступным. Например, когда цистеиновый остаток для функционирования в качестве участка прикрепления находится в окисленной форме, например, если он формирует дисульфидный мостик, необходимо восстановление данного дисульфидного мостика, например, DTT, TCEP или β-меркаптоэтанолом.
Связывание грелина или пептида грелина с коровой частицей и VLP соответственно с применением гетеробифункционального сшивающего средства предпочтительными описанными выше способами обеспечивает ориентированное связывание грелина или пептида грелина с коровой частицей и VLP соответственно. Другие способы связывания грелина или пептида грелина с коровой частицей и VLP соответственно включают в себя способы, где грелин или пептид грелина сшит с коровой частицей и VLP соответственно с применением карбодиимид-EDC и NHS. Грелин или пептид грелина также сначала можно модифицировать тиольной группой в реакции, например, с SATA, SATP или иминотиоланом. Грелин или пептид грелина после снятия защиты, если необходимо, затем можно связывать с коровой частицей и VLP соответственно, как изложено ниже. После отделения избытка тиоилирующего реагента, грелин и пептид грелина вступают в реакцию с коровой частицей и VLP соответственно, ранее активированными гетеробифункциональными сшивающими средствами, содержащими реагирующую с цистеином группу и, следовательно, представляющими, по меньшей мере, одну или несколько функциональных групп, вступающих в реакцию с цистеиновыми остатками, с которыми может реагировать тиоилированный грелин или пептид грелина так, как описано выше. Необязательно, в реакционную смесь добавляют небольшие количества восстанавливающего средства. В дополнительных способах грелин или пептид грелина связывают с коровой частицей и VLP соответственно с применением гомобифункционального сшивающего средства, такого как глутаровый альдегид, DSG, BM[PEO]4, BS3 (Pierce Chemical Company, Rockford, IL, USA), или другие известные гомобифункциональные сшивающие средства с функциональными группами, вступающими в реакцию с аминогруппами или карбоксильными группами на коровой частице и VLP соответственно.
Другие способы связывания VLP с грелином или пептидом грелина включают в себя способы, где коровая частица и VLP соответственно биотинилированы, а грелин или пептид грелина экспрессированы как гибридный белок со стрептавидином, или способы, где и грелин или пептиды грелина и коровая частица и VLP соответственно биотинилированы, например, как описано в WO 00/23955. В данном случае грелин или пептид грелина можно сначала связать со стрептавидином или авидином, регулируя соотношение грелина или пептида грелина к стрептавидину так, чтобы свободные участки прикрепления все еще являлись доступными для связывания коровой частицы и VLP соответственно, которые добавляют на следующей стадии. Альтернативно все компоненты можно смешать в реакции "в одном сосуде". В качестве связывающих средств для связывания грелина или пептида грелина с коровой частицей и VLP соответственно можно применять другие пары лиганд-рецептор, где возможна растворимая форма рецептора и лиганда, способные сшиваться с коровой частицей и VLP соответственно и с грелином или пептидом грелина. Альтернативно, с грелином или пептидом грелина можно слить или лиганд, или рецептор и, таким образом, опосредовать связывание с коровой частицей и VLP соответственно, химически связанной или слитой или с рецептором или с лигандом соответственно. Образование гибрида можно проводить посредством вставки или замены.
Как уже указывалось, в предпочтительном осуществлении настоящего изобретения VLP представляет собой VLP РНК-фага, а в еще более предпочтительном осуществлении VLP представляет собой VLP из белка оболочки РНК-фага Qβ.
С одной субъединицей капсида или VLP белков оболочки РНК-фагов можно связать одну или несколько молекул антигена, т.е. грелина или пептида грелина, предпочтительно посредством незащищенных лизиновых остатков VLP РНК-фагов, если они стерически доступны. Специфической особенностью VLP из белков оболочки РНК-фагов, а, в частности, VLP из белка оболочки Qβ, таким образом, является возможность связывать несколько антигенов с одной субъединицей. Это позволяет образование плотных массивов антигенов.
В предпочтительном осуществлении изобретения связывание и прикрепление соответственно, по меньшей мере, одного грелина или пептида грелина к коровой частице и вирусоподобной частице соответственно происходит посредством взаимодействия и ассоциации соответственно между, по меньшей мере, одним первым участком прикрепления вирусоподобной частицы и, по меньшей мере, одним вторым участком прикрепления антигена или антигенной детерминанты.
VLP или капсиды из белков оболочки Qβ представляют на своей поверхности определенное количество лизиновых остатков с определенной топологией с тремя лизиновыми остатками, направленными внутрь капсида и взаимодействующими с РНК, и четырьмя другими лизиновыми остатками, выставленными наружу капсида. Данные определенные свойства помогают связыванию антигенов с внешней поверхности частицы, а не с внутренней поверхности частицы, где лизиновые остатки взаимодействуют с РНК. VLP из белков оболочки других РНК-фагов также обладают определенным количеством лизиновых остатков на своей поверхности и определенной топологией данных лизиновых остатков.
В дополнительных предпочтительных осуществлениях настоящего изобретения первый участок прикрепления представляет собой лизиновый остаток и/или второй участок прикрепления содержит сульфгидрильную группу или цистеиновый остаток. В очень предпочтительном осуществлении настоящего изобретения первый участок прикрепления представляет собой лизиновый остаток, а второй участок прикрепления представляет собой цистеиновый остаток.
В очень предпочтительном осуществлении настоящего изобретения грелин или пептид грелина связан посредством присутствующего на грелине или пептиде грелина в природе или сконструированного цистеинового остатка с лизиновыми остатками VLP белка оболочки РНК-фага, а в частности с VLP из белка оболочки Qβ.
Другим преимуществом VLP, полученных из РНК-фагов, является высокий уровень их экспрессии у бактерий, обеспечивающий образование больших количеств материала при допустимой стоимости.
Как указано, конъюгаты и массивы по изобретению соответственно отличаются от предыдущих конъюгатов в данной области их высокоорганизованной структурой, размерами и повторяемостью антигена на поверхности массива. Кроме того, применение VLP в качестве носителей обеспечивает формирование устойчивых массивов и конъюгатов антигенов соответственно с различной антигенной плотностью. В частности, применение VLP РНК-фагов и, в частности, для данного случая применение VLP из белков оболочки РНК-фага Qβ обеспечивает достижение очень высокой плотности эпитопов. Получение композиций VLP из белков оболочки РНК-фагов с высокой плотностью эпитопов можно осуществлять применением указания данной заявки. В предпочтительном осуществлении изобретения, когда пептид грелина предпочтительно связан с VLP из белков оболочки Qβ, предпочтительным является среднее количество пептидов грелина на субъединицу в количестве 0,5, 0,6, 0,7, 0,8, 0,9, 1,0, 1,1, 1,2, 1,3, 1,4, 1,5, 1,6, 1,7, 1,8, 1,9, 2,0, 2,1, 2,2, 2,3, 2,4 2,5, 2,6, 2,7, 2,8, 2,9 или более.
Как определено здесь, второй участок прикрепления может являться представленным на антигене или антигенной детерминанте в природе или являться неприродным.
В случае отсутствия на антигене или антигенной детерминанте пригодного встречающегося в природе второго участка прикрепления на антигене необходимо сконструировать неприродный второй участок прикрепления.
Как описано выше, четыре лизиновых остатка выставлены на поверхность VLP из белка оболочки Qβ. Как правило, данные остатки при реакции дериватизированы сшивающей молекулой. В случае когда не все экспонированные лизиновые остатки могут связываться с антигеном, лизиновые остатки, прореагировавшие с сшивающим средством, остаются с сшивающей молекулой связанной с ε-аминогруппой после стадии дериватизации. Это приводит к исчезновению одного или нескольких положительных зарядов, что может являться вредным для растворимости и стабильности VLP. Замещением некоторых из лизиновых остатков на остатки аргинина, как в описанных ниже предоставляемых мутантах по белкам оболочки Qβ, авторы предотвращают избыточное исчезновение положительных зарядов, так как аргининовые остатки не реагируют со сшивающим средством. Кроме того, замещение лизиновых остатков аргининовыми может приводить к образованию более определенных массивов антигенов, так как для реакции с антигеном доступно меньше участков.
Таким образом, выставленные лизиновые остатки замещены аргининовыми в следующих мутантах по белку оболочки Qβ и мутантных VLP Qβ, представляемых в данной заявке: Qβ-240 (Lys13-Arg; SEQ ID № 17), Qβ-250 (Lys2-Arg, Lys13-Arg; SEQ ID № 19) и Qβ-259 (Lys2-Arg, Lys16-Arg; SEQ ID № 21). Конструкции клонированы, белки экспрессированы, VLP очищены и применены для связывания пептидных или белковых антигенов. Также сконструированы Qβ-251; (SEQ ID № 20), руководство о том как экспрессировать, очищать и связывать VLP из белка оболочки Qβ-251, можно найти по ходу данной заявки.
В дополнительном осуществлении авторы представляют мутантный белок оболочки Qβ с одним дополнительным лизиновым остатком, пригодный для получения даже более плотных массивов антигенов. Данный мутантный белок Qβ, Qβ-243 (Asn 10-Lys; SEQ ID № 18) клонирован, белок экспрессирован и выделен, и очищен капсид или VLP, демонстрируя, что введение дополнительного остатка совместимо с самостоятельной сборкой субъединиц в капсид или VLP. Таким образом, массивы и конъюгаты грелина или пептида грелина соответственно можно получить с применением VLP из мутантных белков оболочки Qβ. Предпочтительным на практике способом связывания антигенов к VLP и, в частности, к VLP из белков оболочки РНК-фагов является связывание лизинового остатка, представленного на поверхности VLP из белков оболочки РНК-фагов с цистеиновым остатком, присутствующем в природе или сконструированном на антигене, т.е. грелине или пептиде грелина. Для того чтобы цистеиновый остаток являлся эффективным в качестве второго участка прикрепления, сульфгидрильная группа должна являться доступной для связывания. Таким образом, необходимо, чтобы цистеиновый остаток находится в восстановленном состоянии, то есть свободный цистеин или цистеиновый остаток со свободной сульфгидрильной группой должен являться доступным. В случае, где цистеиновый остаток для функционирования в качестве второго участка прикрепления находится в окисленной форме, например, если он формирует дисульфидный мостик, необходимо восстановление данного дисульфидного мостика, например DTT, TCEP или β-меркаптоэтанолом. Для каждого антигена необходимо отрегулировать концентрацию восстановителя и молярный избыток восстановителя над антигеном. Если необходимо, начиная с таких малых концентраций как 10 мкМ или менее до концентраций от 10 до 20 мМ или более, тестируют диапазон титрования восстановителя и определяют связывание антигена. Хотя низкие концентрации восстановителя совместимы с реакцией связывания, как описано в WO 02/056905, более высокие концентрации ингибируют реакцию связывания, как известно специалисту в данной области, и в данном случае восстановитель необходимо удалять диализом или гель-фильтрацией. Преимущественно pH буфера для диализа или равновесного буфера ниже чем 7, предпочтительно - 6. Совместимость буфера с низким pH с активностью или стабильностью антигена на тестировали.
Плотность эпитопов на VLP из поверхностных белков РНК-фагов можно изменять выбором сшивающего средства и других условий реакции. Например, сшивающие средства Sulfo-GMBS и SMPH, как правило, позволяют достигать высокой плотности эпитопов. На дериватизацию позитивно влияет высокая концентрация реагентов, а для контроля количества связанных с VLP из белков оболочки РНК-фагов, а в особенности с VLP белка оболочки Qβ, антигенов можно применять управление условиями реакции.
До конструирования неприродного второго участка прикрепления необходимо выбрать положение, в котором его необходимо слить, вставить или подвергнуть конструированию в общем смысле. Таким образом, положение второго участка прикрепления необходимо выбрать так, чтобы избежать стерических препятствий от второго участка прикрепления или содержащего его аминокислотного линкера. В дополнительных осуществлениях желателен ответ антителами, направленный на участок, отличный от участка взаимодействия аутоантигена с его природным лигандом. В таких осуществлениях второй участок прикрепления можно выбрать так, чтобы он предотвращал образование антител против участка взаимодействия аутоантигена с его природными лигандами.
В наиболее предпочтительных осуществлениях грелин или пептид грелина содержат второй участок прикрепления или единственный реакционноспособный участок прикрепления, способный к ассоциации с первыми участками прикрепления на коровой частице и VLP или субъединицах VLP соответственно. Это обеспечивает определенное и однородное связывание и ассоциацию соответственно, по меньшей мере, одного, но, как правило, более чем одного, предпочтительно - более чем 10, 20, 40, 80, 120, 150, 180, 210, 240, 270, 300, 360, 400, 450 антигенов с коровой частицей и VLP соответственно. Таким образом, обеспечение единственного второго участка прикрепления или единственного реакционноспособного участка прикрепления на антигене гарантирует единый и однообразный тип связывания и ассоциации соответственно, приводящий к очень высокоупорядоченным и повторяющимся массивам. Например, если связывание и ассоциацию соответственно осуществляют посредством взаимодействия лизина (в качестве первого участка прикрепления) и цистеина (в качестве второго участка прикрепления) по предпочтительному осуществлению изобретения, гарантировано, что только один цистеиновый остаток на антиген независимо от того, является ли присутствующий на антигене цистеиновый остаток природным или неприродным, способен к связыванию и ассоциации соответственно с VLP и первым участком прикрепления коровой частицы соответственно.
В некоторых осуществлениях конструирование второго участка прикрепления на антигене требует слияния аминокислотного линкера, содержащего аминокислоту, пригодную в качестве второго участка прикрепления по описаниям данного изобретения. Следовательно, в предпочтительном осуществлении настоящего изобретения аминокислотный линкер связан с антигеном или антигенной детерминантой, по меньшей мере, одной ковалентной связью. Предпочтительно аминокислотный линкер содержит второй участок прикрепления или, альтернативно, состоит из него. В дополнительном предпочтительном осуществлении аминокислотный линкер содержит сульфгидрильную группу или цистеиновый остаток. В другом предпочтительном осуществлении аминокислотный линкер представляет собой цистеин. Некоторые критерии выбора аминокислотного линкера, а также дополнительные предпочтительные осуществления аминокислотного линкера по изобретению уже указаны выше.
В дополнительном предпочтительном осуществлении изобретения с коровой частицей и вирусоподобной частицей соответственно слиты, по меньшей мере, один антиген или антигенная детерминанта, т.е. грелин или пептид грелина. Как указано выше, VLP, как правило, состоит, по меньшей мере, из одной субъединицы, собирающейся в VLP. Таким образом, в еще одном дополнительном осуществлении изобретения антиген или антигенная детерминанта предпочтительно, по меньшей мере, один грелин или пептид грелина слиты, по меньшей мере, мере с одной субъединицей вирусоподобной частицы или белка, способного внедряться в VLP, образуя гибридный белок субъединицы VLP-грелин или пептид грелина.
Слияние грелина или пептидов грелина можно проводить вставкой в последовательность субъединицы VLP или слиянием с N- или с C-концом субъединицы VLP или белка, способного к включению в VLP. Ниже при указании на гибридные белки пептида с субъединицей VLP имеют в виду слияние с каким-либо из концов последовательности субъединицы или вставку пептида внутрь последовательности субъединицы.
Слияние также можно проводить посредством вставки последовательностей грелина или пептида грелина в вариант субъединицы VLP, где удалена часть последовательности субъединицы, то, что далее обозначают как укороченные мутанты. У укороченных мутантов могут являться удаленными N- или C-конец или внутренние части последовательности субъединицы VLP. Например, конкретная VLP HBcAg, например, с удалением аминокислотных остатков с 79 по 81 представляет собой укороченный мутант с внутренней делецией. Слияние грелина или пептида грелина или N- или C-концом укороченного мутанта субъединиц VLP также приводит к осуществлениям изобретения. Подобным образом слияние эпитопа с последовательностью субъединицы VLP также можно проводить посредством замены, где, например, для конкретной VLP HBcAg аминокислоты 79-81 затененены на чужеродный эпитоп. Таким образом, слияние, как указано далее, можно проводить вставкой последовательности грелина или пептида грелина в последовательность субъединицы VLP посредством замены части последовательности субъединицы VLP последовательностью грелина или пептида грелина или посредством сочетания удаления, замены или вставок.
Гибридная с грелином или пептидом грелина субъединица VLP, как правило, способна к самостоятельной сборке в VLP. VLP, представляющую эпитопы, слитые с ее субъединицами, здесь также обозначают как на гибридные VLP. Как указано, вирусоподобная частица содержит или, альтернативно, состоит, по меньшей мере, из одной субъединицы VLP. В дополнительном осуществлении изобретения вирусоподобная частица содержит или, альтернативно, состоит из смеси гибридных субъединиц и негибридных субъединиц, т.е. субъединиц VLP, не несущих слитого с ними антигена, что приводит к так называемым мозаичным частицам. Это может являться выгодным для обеспечения формирования и сборки VLP. В данных осуществлениях соотношение гибридных субъединиц VLP может составлять 1, 2, 5, 10, 20, 30, 40, 50, 60, 70, 80, 90, 95% или более.
К любому концу последовательности пептида или эпитопа для слияния или с концом последовательности субъединицы VLP, или для внутренней вставки такой пептидной последовательности в последовательность субъединицы VLP можно добавлять фланкирующие аминокислотные остатки. Особенно предпочтительными аминокислотами для применения во фланкирующих последовательностях, добавляемых к грелину или пептиду грелина для слияния, являются остатки глицина и серина. Глициновые остатки дают дополнительную гибкость, что может уменьшить потенциально дестабилизирующее действие слияния чужеродной последовательности с последовательностью субъединицы VLP.
В конкретном осуществлении изобретения VLP представляет собой VLP корового антигена гепатита B. Описаны и представляют собой предпочтительные осуществления изобретения гибридные белки или с N-концом HBcAg (Neyrinck, S. et al., Nature Med. 5: 1157-1163 (1999)) или вставки в так называемую главную иммунодоминантную область (MIR) (Punpens, P. and Grens, E., Intervirology 44: 98-114 (2001)), WO 01/98333). Также описаны встречающиеся в природе варианты HBcAg с делециями в MIR (Pumpens, P. and Grens, E., Intervirology 44: 98-114 (2001), специально включенные в качестве ссылки полностью), а слияния с N- или C-концом, а также вставки в положение MIR, соответствующие участку делеции, в сравнении с wt HBcAg, являются дополнительными осуществлениями изобретения. Также описаны слияния с C-концом (Pumpens, P. and Grens, E., Intervirology 44: 98-114 (2001)). Специалист в данной области легко найдет руководство о том, как конструировать гибридные белки с применением классических способов молекулярной биологии (Sambrook, J. et al., eds., Molecular Cloning, A Laboratory Manual, 2nd. edition, Cold Spring Habor Laboratory Press, Cold Spring Harbor, N. Y. (1989), Ho et al., Gene 77: 51 (1989)). Описаны векторы и плазмиды, кодирующие HBcAg и гибридные белки HBcAg и пригодные для экспрессии HBcAg и гибридных белков HBcAg (Pumpens, P. & Grens, E. Intervirology 44: 98-114 (2001), Neyrinck, S. et al., Nature Med. 5: 1157-1163 (1999)), и их можно применять в практическом осуществлении изобретения. Также посредством примера (пример 6) авторы описывают вставку эпитопа в MIR HBcAg, приводящую к гибридному самостоятельно собирающемуся HBcAg. Важным фактором для оптимизации эффективности самостоятельной сборки и представления эпитопа, который будет вставлен в MIR HBcAg, является выбор участка вставки, а также количество аминокислот, которое будет удалено из последовательности HBcAg в пределах MIR (Pumpens, P. and Grens, E., Intervirology 44: 98-114 (2001); EP 421635; US 6231864) при вставке, или, иными словами, какие аминокислоты, формирующие HBcAg, подлежат замене новым эпитопом. Например, описана замена аминокислот 76-80,79-81, 79-80, 75-85 или 80-81 HBcAg чужеродными эпитопами (Pumpens, P. and Grens, E., Intervirology 44: 98-114 (2001); EP0421635; US 6231864). HBcAg содержит длинный конец из аргининовых остатков (Pumpens, P. and Grens, E., Intervirology 44: 98-114 (2001)), который несущественен для сборки капсида и способен к связыванию нуклеиновых кислот (Pumpens, P. and Grens, E., Intervirology 44: 98-114 (2001)). Содержащие данный конец из аргининовых остатков или не содержащие его HBcAg представляют собой осуществления данного изобретения.
В дополнительном предпочтительном осуществлении изобретения VLP представляет собой VLP РНК-фага. Основные белки оболочки РНК-фагов при экспрессии в бактериях и, в частности, в E. coli спонтанно собираются в VLP. Конкретные примеры белков оболочки бактериофагов, которые можно применять для получения композиций по изобретению, включают в себя белки оболочки таких РНК-бактериофагов, как бактериофаг Qβ (SEQ ID № 4; PIR Database, инвентарный № VCBPQβ, указывающий на CP Qβ и SEQ ID № 5; инвентарный № AAA16663, указывающий на белок A1 Qβ) and бактериофаг fr (SEQ ID № 7; инвентарный № в PIR VCBPFR).
В более предпочтительном осуществлении, по меньшей мере, один грелин или пептид грелина слит с белком оболочки Qβ. Описаны конструкции гибридных белков, где эпитопы слиты c C-концом укороченной формы белка A1 Qβ или вставлены в белок A1 (Kozlovska, T. M., et al., Intervirology, 39: 9-15 (1996)). Белок A1 получают супрессией стоп-кодона UGA, и он обладает длиной 329 а.к. или 328 а.к., если принять в расчет отщепление N-концевого метионина. Отщепление N-концевого метионина перед аланином (вторая аминокислота, кодируемая геном CP Qβ), как правило, происходит в E. coli, и это имеет место для N-концов белков оболочки CP Qβ. Часть гена A1 в направлении 3' от амбер-кодона UGA кодирует продолжение CP с длиной 195 аминокислот. Вставка, по меньшей мере, одного грелина или пептида грелина между положениями 72 и 73 в продолжении CP приводит к дополнительным осуществлениям изобретения (Kozlovska, T. M., et al., Intervirology 39: 9-15 (1996)). Слияние грелина или пептида грелина с C-концом укороченного на C-конце белка A1 Qβ ведет к дополнительным осуществлениям изобретения. Например, Kozlovska et al., (Intervirology, 39: 9-15 (1996)) описывают гибридные белки A1 Qβ, где эпитоп слит с C-концом удлинения CP Qβ, укороченного по положению 19.
Как описано у Kozlovska et al. (Intervirology, 39: 9-15 (1996)), сборка частиц представляющих слитые эпитопы для формирования мозаичной частицы, как правило, требует присутствия и гибрида белок A1-грелин или пептид грелина, и wt CP. Однако осуществления, включающие в себя вирусоподобные частицы и, в частности, для данного случая, VLP из белка оболочки РНК-фага Qβ, исключительно состоящие из субъединиц VLP, по меньшей мере, с одним слитым с ними грелином или пептидом грелина, также находятся в объеме настоящего изобретения.
Образование мозаичных частиц можно проводить рядом способов. Kozlovska et al., Intervirolog, 39: 9-15 (1996), описывают два способа, оба из которых можно применять при практическом осуществлении изобретения. В первом подходе эффективное представление слитого эпитопа на VLP опосредуют экспрессией плазмиды, кодирующей белок A1 Qβ со стоп-кодоном UGA между CP и удлинением CP в штамме E. coli, несущем плазмиду, кодирующую клонированную супрессорную для UGA тРНК, что ведет к трансляции кодона UGA в Trp (плазмида pISM3001 (Smiley B. K., et al., Gene 134: 33-40 (1993))). В другом подходе стоп-кодон гена CP модифицируют в UAA, совместно трансформируют вторую плазмиду, экспрессирующую гибрид белок A1-грелин или пептид грелина. Вторая плазмида кодирует различную устойчивость к антибиотикам, а начало репликации сочетается с первой плазмидой (Kozlovska, T. M., et al., Intervirology 39: 9-15 (1996)). В третьем подходе гибрид белок CP и белок A1-грелин или пептид грелина кодируют в виде двух цистронов, функционально связанных с таким промотором, как Trp-промотор, как описано на фигуре 1 в Kozlovska et al., Intervirology, 39: 9-15 (1996).
В дополнительном осуществлении грелин или пептид грелина вставлен между аминокислотами 2 и 3 (нумерация расщепленного CP, то есть там, где отщеплен N-концевой метионин) CP fr, таким образом приводя к гибридному белку грелин или пептид грелина-CP fr. Описаны векторы и экспрессирующие системы для конструирования и экспрессии гибридных белков CP fr, самостоятельно собирающихся в VLP и пригодных в практическом осуществлении изобретения (Pushko P. et al., Prot. Eng. 6: 883-891 (1993)). В конкретном осуществлении последовательность грелина или пептида грелина вставлена в делеционный вариант CP fr после аминокислоты 2, где остатки CP fr 3 и 4 удалены (Pushko P. et al., Prot. Eng. 6: 883-891 (1993)).
Также описано слияние эпитопов с N-концевой выступающей β-шпилькой белка оболочки РНК-фага MS-2 и последующее представление слитого эпитопа на самостоятельно собирающейся VLP РНК-фага MS-2 (WO 92/13081), а слияние грелина или пептида грелина посредством вставки или замены в белке оболочки РНК-фага MS-2 также попадают в область изобретения.
В другом осуществлении изобретения грелин или пептиды грелина слиты с белком капсида вируса папилломы. В более конкретном осуществлении грелин или пептиды грелина слиты с главным белком капсида L1 коровьего вируса папилломы типа 1 (BPV-1). Описаны векторы и экспрессионные системы для конструирования и экспрессии гибридных белков BPV-1 в клеточных системах бакуловирус/клетки насекомых (Chackerian, B. et al., Proc. Natl. Acad. Sci. USA 96: 2373-2378 (1999), WO 00/23955). Замена аминокислот 130-136 L1 BPV-1 на грелин или пептид грелина приводит к гибридному белку L1 BPV-1-грелин или пептид грелина, который представляет собой предпочтительное осуществление изобретения. Описано клонирование в бакуловирусном векторе и экспрессия в инфицированных бакуловирусом клетках Sf9, и его можно применять в практическом осуществлении изобретения (Chackerian, B. et al., Proc. Natl. Acad. Sci. USA 96: 2373-2378 (1999), WO 00/23955). Очистку собранных частиц, представляющих слитые грелин или пептиды грелина, можно проводить рядом способов, например, таких как гель-фильтрация или ультрацентифугирование на градиенте сахарозы (Chackerian, B. et al., Proc. Natl. Acad. Sci. USA 96: 2373-2378 (1999), WO 00/23955).
В дополнительном осуществлении изобретения грелин или пептиды грелина слиты с белком Ty, способным включаться в VLP Ty. В более конкретном осуществлении грелин или пептиды грелина слиты с p1 или капсидным белком, кодируемым геном TYA gene (Roth, J. F., Yeast 16: 785-795 (2000)). Дрожжевые ретротранспозоны Ty1, 2, 3 и 4 выделены из Saccharomyces Cerevisiae, тогда как ретротранспозон Tf1 выделен из Schizosaccharomyces Pombae (Boeke, J. D. and Sandmeyer, S. B., "Yeast Transposable elements", в The molecular and Cellular Biology of the Yeast Saccharomyces: Genome dynamics, Protein Synthesis, and Energetics., p. 193, Cold Spring Harbor Laboratory Press (1991)). Ретротранспозоны Ty1 и 2 относятся к классу copia элементов растений и животных, тогда как Ty3 принадлежит семейству ретротранспозонов gypsy, относящегося к ретровирусам растений и животных. В ретротранспозоне Ty1 белок p1, также обозначаемый как Gag или капсидный белок, обладает длиной 440 аминокислот. P1 в течение созревания VLP расщепляется по положению 408, что ведет к белку p2, неотъемлемому компоненту VLP.
Описаны гибридные белки с p1 и векторы для экспрессии указанных гибридных белков в дрожжах (Adams, S. E., et al., Nature 329: 68-70 (1987)). Так, например, грелин или пептид грелина можно слить с p1 посредством вставки последовательности, кодирующей грелин или пептид грелина в участок BamH1 плазмиды pMA5620 (Adams, S. E., et al., Nature 329: 68-70 (1987)). Клонирование последовательностей, кодирующих чужеродные эпитопы, в вектор pMA5620 приводит к экспрессии гибридных белков, содержащих аминокислоты 1-381 p1 Ty1-15, C-концом слитых с N-концом чужеродного эпитопа. Подобным образом N-концевое слияние грелина или пептидов грелина, или внутреннюю вставку в последовательность p1, или замену части последовательности p1 также подразумевают попадающими в объем изобретения. В частности, вставка грелина или пептидов грелина в последовательность Ty между аминокислотами белка p1 Ty 30-31, 67-68,113-114 и 132-133 (EP0677111) приводит к предпочтительным осуществлениям изобретения.
Дополнительные VLP, пригодные для слияния с грелином или пептидами грелина, представляют собой, например, подобные ретровирусам частицы (WO 9630523), Gag HIV2 (Kang, Y. C., et al, Biol. Chem. 380: 353-364 (1999)), вирус мозаики коровьего гороха (Taylor, K. M. et al., Biol. Chem. 380: 387-392 (1999)), VLP парвовируса VP2 (Rueda, P. et al., Virology 263: 89-99 (1999)), HBsAg (US 4722840, EP0020416B1).
Примеры гибридных VLP, пригодных для практического осуществления изобретения, также представляют собой VLP, описанные в Intervirology 39: 1 (1996). Дополнительные примеры VLP, рассматриваемых для применения в изобретении, представляют собой: HPV-1, HPV-6, HPV-11, HPV-16, HPV-18, HPV-33, HPV-45, CRPV, COPV, HIV GAG, вирус табачной мозаики. Также сконструированы вирусоподобные частицы SV-40, вируса полиомы, аденовируса, вируса простого герпеса, ротавируса и вируса Норфолка, а гибридные VLP данных VLP также находятся в объеме настоящего изобретения.
В дополнительном предпочтительном осуществлении настоящего изобретения антиген или антигенная детерминанта представляют собой грелин или пептид грелина.
В дополнительном очень предпочтительном осуществлении изобретения антиген или антигенная детерминанта выбраны из группы, состоящей из a) человеческого грелина; b) кошачьего грелина; c) собачьего грелина; d) коровьего грелина; e) овечьего грелина; f) лошадиного грелина; g) свиного грелина; h) пептида или его фрагмента любого грелина из a)-g).
В дополнительном очень предпочтительном осуществлении изобретения антиген или антигенная детерминанта содержит, или, альтернативно, по существу состоит из, или, альтернативно, состоит из аминокислотной последовательности, выбранной из группы, состоящей из:
(a) GSSFLSPEHQRVQRKESKKPPAKLQPR (SEQ ID № 48)
(b) GSSFLSPEHQRVQQRKESKKPPAKLQPR (SEQ ID № 31)
(c) GSSFLSPEHQKLQQRKESKKPPAKLQPR (SEQ ID № 49)
(d) GSSFLSPEHQKLQRKESKKPPAKLQPR (SEQ ID № 50)
(e) GSSFLSPEHQKAQQRKESKKPPAKLQPR (SEQ ID № 32)
(f) GSSFLSPEHQKAQRKESKKPPAKLQPR (SEQ ID № 51)
(g) KKPPAKLQPR (SEQ ID № 52)
(h) PPAKLQPR (SEQ ID № 53)
(i) AKLQPR (SEQ ID № 54)
(j) GSSFLSPEHQ (SEQ ID № 55)
(k) EHQRVQQRKE (SEQ ID № 56)
(l) KLQPR (SEQ ID № 59)
(m) GSSFLSPEHQRVQ (SEQ ID № 60)
(n) QRKESKKPPAKLQPR (SEQ ID № 61)
(o) GSSFLSPEHQKLQ (SEQ ID № 62)
(p) QRKESKKPPAKLQPR (SEQ ID № 63)
(q) EHQRVQQRKES (SEQ ID № 111)
(r) EHQKAQQRKE (SEQ ID № 112)
(s) EHQKAQQRKES (SEQ ID № 113)
(t) EHQKLQQRKE (SEQ ID № 114)
(u) EHQKLQQRKES (SEQ ID № 115)
(v) LSPEHQRVQQ (SEQ ID № 116)
(w) LSPEHQKAQQ (SEQ ID № 117)
(x) LSPEHQKLQQ (SEQ ID № 118).
В дополнительном очень предпочтительном осуществлении изобретения антиген или антигенная детерминанта представляет собой человеческий, собачий, свиной, лошадиный, овечий, бычий, морских свинок, собачий или мышиный грелин и пептид грелина соответственно. Грелины и пептиды грелинов соответственно можно получить посредством экспрессии ДНК, кодирующей грелин и пептид грелина соответственно под контролем сильного промотора. Предпочтительно ДНК кодирует не предшественник прогрелина, а только пептидный каркас активного модифицированного н-октаноилом пептида. Различные примеры этого описаны в литературе, и их можно применять, возможно, после модификаций, для экспрессии грелина любых желаемых видов.
В дополнительном предпочтительном осуществлении изобретения антиген или антигенная детерминанта представляет собой полученный из грелина пептид или его фрагмент. Предпочтительно пептид грелина выбран из группы, состоящей из a) пептида человеческого грелина; b) пептида коровьего грелина; c) пептида кошачьего грелина; d) пептида собачьего грелина; e) пептида свиного грелина; f) пептида грелина куриц; g) пептида мышиного грелина; h) пептида лошадиного грелина; i) пептида овечьего грелина; и j) фрагмента любого пептида грелина из a)-i).
Такие пептиды грелина или их фрагменты можно получить с применением стандартных способов молекулярной биологии, где нуклеотидную последовательность, кодирующую интересующий фрагмент, амплифицируют посредством GWH и клонируют как слияние с полипептидной меткой, такой как гистидиновая метка, метка Flag, метка myc или константная область антитела (область Fc). Посредством внесения участка расщепления энтерокиназой между фрагментом грелина и меткой фрагмент грелина после очистки можно отделить от метки посредством расщепления энтерокиназой. В другом подходе фрагмент грелина можно синтезировать in vitro с модификацией н-октаноилом или без нее с применением стандартных реакций пептидного синтеза, известных специалисту в данной области. В дополнительном подходе фрагменты грелина можно получить посредством расщепления протеазой или химическим расщеплением полноразмерного грелина, где оба способы хорошо известны специалистам в данной области.
В дополнительном предпочтительном осуществлении настоящего изобретения пептид грелина содержит, по меньшей мере, один антигенный участок грелина. Специалист в данной области знает, как идентифицировать соответствующие пептиды и аминокислотные последовательности соответственно. Предпочтительно фрагменты могут включать в себя N- или C-конец грелина. C-конец может являться особенно пригодным, так как модификация н-октаноилом, которая возможно может препятствовать связыванию специфичных к немодифицированной форме антител, находится вблизи N-конца.
Предпочтительные человеческие пептиды грелина включают в себя:
KKPPAKLQPR (SEQ ID № 52)
PPAKLQPR (SEQ ID № 53)
AKLQPR (SEQ ID № 54)
KLQPR (SEQ ID № 59)
GSSFLSPEHQ (SEQ ID № 55)
EHQRVQQRKES (SEQ ID № 111)
LSPEHQRVQQ (SEQ ID № 116)
GSSFLSP (SEQ ID № 119)
Однако пептиды из внутренних участков грелина также могут являться привлекательными кандидатами для вакцин. Так как грелины различных видов являются высокогомологичными, вероятно то, что можно индуцировать ответы перекрестно-реактивными антителами. Таким образом, антитело, отвечающее против собачьего или мышиного грелина, также распознает человеческий грелин и наоборот. Следовательно, в объеме данного изобретения находятся то, что все грелины и пептиды, полученные из них, с аминокислотами более чем на 80%, предпочтительно - более чем на 85%, более предпочтительно - более чем на 90% и даже боле предпочтительно - более чем на 95%, 97% или даже 99% идентичными человеческому грелину, можно применять для вакцинации.
Руководство о том, как модифицировать человеческий грелин или пептид грелина, в частности, для связывания с вирусоподобной частицей, дано в данной заявке. Иммунизация против грелина с применением композиций по изобретению, содержащих предпочтительно человеческий грелин или пептид грелина, связанный с коровой частицей и VLP соответственно, может предоставить способ лечения ожирения.
В дополнительном очень предпочтительном осуществлении настоящего изобретения антиген или антигенная детерминанта представляет собой грелин или пептид грелина, содержащий, или, альтернативно, по существу состоящий из, или, альтернативно, состоящий из аминокислотной последовательности, выбранной из группы, состоящей из:
(a) GSSFLSPEHQRVQRKESKKPPAKLQPR (человек) (SEQ ID № 48)
(b) GSSFLSPEHQRVQQRKESKKPPAKLQPR (человек) (SEQ ID № 31)
(c) GSSFLSPEHQRVQ (человек) (SEQ ID № 60)
(d) QRKESKKPPAKLQPR (человек) (SEQ ID № 61)
(е) PPAKLQPR (человек) (SEQ ID № 53)
(f) GSSFLSPEHQKLQQRKESKKPPAKLQPR (собака) (SEQ ID № 49)
(g) GSSFLSPEHQKLQRKESKKPPAKLQPR (собака) (SEQ ID № 50)
(h) GSSFLSPEHQKLQ (собака) (SEQ ID № 62)
(i) QRKESKKPPAKLQPR (собака) (SEQ ID № 63)
(k) EHQRVQQRKE (человек) (SEQ ID № 56)
(l) GSSFLSPEHQKAQQRKESKKPPAKLQPR (мышь) (SEQ ID № 32)
(n) аминокислотной последовательности любого фрагмента любого из (a)-(l)
В еще одном дополнительном предпочтительном осуществлении настоящего изобретения антиген или антигенная детерминанта дополнительно содержат, по меньшей мере, один второй участок прикрепления, выбираемый из: (i) участка прикрепления, не встречающегося в природе с указанным антигеном или антигенной детерминантой, и (ii) участка прикрепления, встречающегося в природе с указанным антигеном или антигенной детерминантой. В предпочтительном осуществлении указанный участок прикрепления содержит аминокислотный линкер по изобретению, предпочтительно линкерную последовательность из C, CG или CGG. Предпочтительно антиген или антигенная детерминанта с указанным, по меньшей мере, одним вторым участком прикрепления содержит, или, альтернативно, по существу состоит из, или, альтернативно, состоит из аминокислотной последовательности, выбранной из группы, состоящей из:
(a) CGSSFLSPEHQRVQRKESKKPPAKLQPR (человек) (SEQ ID № 64)
(b) CGSSFLSPEHQRVQQRKESKKPPAKLQPR (человек) (SEQ ID № 65)
(c) GSSFLSPEHQRVQRKESKKPPAKLQPRC (человек) (SEQ ID № 66)
(d) GSSFLSPEHQRVQRKESKKPPAKLQPRGC (человек) (SEQ ID № 120)
(e) GSSFLSPEHQRVQQRKESKKPPAKLQPRC (человек) (SEQ ID № 67)
(f) GSSFLSPEHQRVQQRKESKKPPAKLQPRGC (человек) (SEQ ID № 121)
(g) GSSFLSPEHQRVQC (человек) (SEQ ID № 68)
(h) GSSFLSPEHQRVQGC (человек) (SEQ ID № 122)
(i) CQRKESKKPPAKLQPR (человек) (SEQ ID № 69)
(j) CPPAKLQPR (человек) (SEQ ID № 70)
(k) CGSSFLSPEHQKLQQRKESKKPPAKLQPR (собака) (SEQ ID № 71)
(l) CGSSFLSPEHQKLQRKESKKPPAKLQPR (собака) (SEQ ID № 72)
(m) GSSFLSPEHQKLQQRKESKKPPAKLQPRC (собака) (SEQ ID № 73)
(n) GSSFLSPEHQKLQQRKESKKPPAKLQPRGC (собака) (SEQ ID № 123)
(o) GSSFLSPEHQKLQRKESKKPPAKLQPRC (собака) (SEQ ID № 74)
(p) GSSFLSPEHQKLQRKESKKPPAKLQPRGC (собака) (SEQ ID № 124)
(q) GSSFLSPEHQKLQC (SEQ ID № 75)
(r) GSSFLSPEHQKLQGC (SEQ ID № 125)
(s) CEHQRVQQRKE (SEQ ID № 76)
(t) CGSSFLSPEHQKAQQRKESKKPPAKLQPR (мышь) (SEQ ID № 77)
(u) GSSFLSPEHQKAQRKESKKPPAKLQPRC (мышь) (SEQ ID № 126)
(v) GSSFLSPEHQKAQRKESKKPPAKLQPRGC (мышь) (SEQ ID № 127)
(w) GSSFLSPEHQKAQQRKESKKPPAKLQPRC (мышь) (SEQ ID № 128)
(x) GSSFLSPEHQKAQQRKESKKPPAKLQPRGC (мышь) (SEQ ID № 129)
(y) GSSFLSPEHQKAQC (мышь) (SEQ ID № 130)
(z) GSSFLSPEHQKAQGC (мышь) (SEQ ID № 131)
(aa) GGSSFLSPEHQGC (SEQ ID № 132)
(bb) CKKPPAKLQPR (SEQ ID № 133)
(cc) CEHQKAQQRKE (SEQ ID № 134)
(dd) CEHQKAQQRKES (SEQ ID № 135)
(ee) CLSPEHQKAQQ (SEQ ID № 136)
(ff) CEHQRVQQRKES (SEQ ID № 137)
(gg) CLSPEHQRVQQ (SEQ ID № 138)
аминокислотной последовательности любого фрагмента любого из (a)-(gg)
Некоторые из очень предпочтительных пептидов грелина описаны в примере 13, где, в частности, описаны соответствующие мышиные пептиды. Данные пептиды в качестве второго участка прикрепления для связывания с VLP и пилями содержат N- или C-концевой цистеиновый остаток. Данные очень предпочтительные короткие пептидные фрагменты грелина способны обладать очень высокой иммунногенностью при связывании с VLP и коровой частицей соответственно. Кроме того, предпочтительные фрагменты грелина также способны преодолеть проблемы безопасности, возникающие, когда целевые собственные белки как более короткий фрагмент со значительно меньшей вероятностью содержат T-клеточные эпитопы. Как правило, чем короче пептиды - тем безопаснее в отношении активации T-клеток. Однако слишком короткие пептиды могут не вызвать индукции высокоаффинных антител, способных к прочному связыванию грелина в растворе.
Сначала выбрали очень предпочтительные фрагменты пептидов грелина, соответствующие фрагменту мышиного грелина из остатков 1-10 (GSSFLSPEHQ) (SEQ ID № 55), так как N-концевой сегмент грелина идентичен у всех известных видов. Кроме того, вероятно, что он идентичен у видов, где грелин еще не идентифицирован. Кроме того, C-концевой остаток, глутамин, может увеличивать растворимость, облегчая получение растворимого вакцинного продукта при связывании с VLP и с коровой частицей соответственно. Фактически растворимость пептидов часто представляет собой лимитирующий фактор для эффективности связывания и стабильности вакцины. Дальнейшее рассуждение включает в себя предотвращение потенциального T-клеточного эпитопа. Выбор меньшего фрагмента пептида уменьшает вероятность присутствия T-клеточного эпитопа. Связывание остатков 1-10 мышиного грелина через C-конец с VLP, при иммунизации мышей индуцирует специфические к N-концу антитела, способные связывать активный грелин и, следовательно, предотвращают его прохождение через гематоэнцефалический барьер, приводя к уменьшенному потреблению пищи.
Дополнительные очень предпочтительные фрагменты пептидов грелина, соответствующие фрагменту пептида мышиного грелина из остатков 42-51 (KKPPAKLQPR) (SEQ ID № 52), выбрали исходя из оснований, сходных с указанными выше. Единственная разница, состоит в том, что данный фрагмент пептида, связанный через N-конец, способен при иммунизации у мышей индуцировать антитела, специфичные к C-концевой части грелина. Кроме того, можно усилить узнавание антител, поскольку можно избежать н-октаноильной группы на серине 3, следовательно, уменьшая ей возможность служить помехой. Связывание остатков 42-51 мышиного грелина через N-конец с VLP индуцирует антитела, способные нейтрализовать активный грелин и, следовательно, предотвращают его прохождение через гематоэнцефалический барьер, приводя к уменьшенному потреблению пищи.
Кроме того, выбрали еще несколько дополнительных очень предпочтительных фрагментов пептидов грелина, соответствующих фрагментам пептида мышиного грелина из остатков 31-41 (EHQKAQQRKES) (SEQ ID № 113) и остатков 28-37 (LSPEHQKAQQ) (SEQ ID № 117), охватывающих центральный сегмент грелина. Подобным образом данные фрагменты грелина лишены потенциальных T-клеточных эпитопов, а также лишены н-октаноильной группы на серине 3. Связывание данных пептидов через N- или C-конец с VLP индуцирует антитела, способные нейтрализовать активный грелин и, следовательно, предотвращают его прохождение через гематоэнцефалический барьер, приводя к уменьшенному потреблению пищи.
Дополнительные предпочтительные пептиды грелина, пригодные для применения по настоящему изобретению, можно идентифицировать посредством применения существующих или будущих моноклональных или поликлональных антител.
Дополнительные предпочтительные молекулы и пептиды грелина, полученные из молекул, можно выявлять в природе в видах, где еще не доступна информация о последовательности.
Специалист в подходящих областях поймет, что без отклонения от объема изобретения или любого его осуществления легко увидеть и сделать другие пригодные модификации и адаптации к описанным здесь способам и применениям. Теперь, после подробного описания настоящего изобретения, то же самое будет более легко понять при ссылке к следующим примерам, включенным сюда только с целью иллюстрации и не предназначенным являться ограничением изобретения.
ПРИМЕРЫ
ПРИМЕР 1
Конструирование и экспрессия мутантных белков оболочки Qβ и очистка VLP или капсидов из мутантных белков оболочки Qβ
Конструирование плазмиды и клонирование мутантных белков оболочки.
Конструирование pQβ-240:
В качестве исходной плазмиды для конструирования pQβ-240 применяли плазмиду pQβ10 (Kozlovska, TM, et al., Gene 137: 133-137). Посредством обращенной ПЦР создавали мутацию Lys13→Arg. Обращенные праймеры разрабатывали в обращенных направлениях хвост к хвосту:
5'-GGTAACATCGGTCGAGATGGAAAACAAACTCTGGTCC-3' (SEQ ID № 78)
и
5'-GGACCAGAGTTTGTTTTCCATCTCGACCGATGTTACC-3' (SEQ ID № 79).
Продукты первой ПЦР применяли как матрицы для второй реакции ПЦР, в которой применяли прямой праймер
5'-AGCTCGCCCGGGGATCCTCTAG-3' (SEQ ID № 80)
и обратный праймер
5'-CGATGCATTTCATCCTTAGTTATCAATACGC-TGGGTTCAG3' (SEQ ID № 81).
Продукт второй ПЦР расщепляли XbaI и Mph1103I и клонировали в экспрессирующий вектор pQβ10, расщепленный теми же рестрикционными ферментами. Проводили реакции ПЦР с набором реагентов для ПЦР и согласно протоколу производителя (MBI Fermentas, Vilnius, Lithuania).
Секвенированием с применением способа прямого включения метки подтверждали наличие желательных мутаций. Клетки E. coli, несущие pQβ-240, поддерживали эффективный синтез белка массой 14 кДа, совместно мигрирующего на SDS-PAGE с контрольным белком оболочки Qβ, выделенным из частиц фага Qβ.
Полученная аминокислотная последовательность: (SEQ ID № 17)
AKLETVTLGNIGRDGKQTLVLNPRGVNPTNGVASLSQAGAVP
ALEKRVTVSVSQPSRNRKNYKVQVKIQNPTACTANGSCDPSVTRQ
KYADVTFSFTQYSTDEERAFVRTELAALLASPLLIDAIDQLNPAY
Конструирование pQβ-243:
В качестве исходной плазмиды для конструирования pQβ-243 применяли плазмиду pQβ10. Посредством обращенной ПЦР создавали мутацию Asn10→Lys. Обращенные праймеры разрабатывали в обращенных направлениях хвост к хвосту:
5'-GGCAAAATTAGAGACTGTTACTTTAGGTAAGATCGG-3' (SEQ ID № 82)
и
5'-CCGATCTTACCTAAAGTAACAGTCTCTAATTTTGCC-3' (SEQ ID № 83).
Продукты первой ПЦР применяли как матрицы для второй реакции ПЦР, в которой применяли прямой праймер
5'-AGCTCGCCCGGGGATCCTCTAG-3' (SEQ ID № 80)
и обратный праймер
5'-CGATGCATTTCATCCTTAGTTATCAATACGCTGGGTTCAG-3' (SEQ ID № 81).
Продукт второй ПЦР расщепляли XbaI и Mph1103I и клонировали в экспрессирующий вектор pQβ10, расщепленный теми же рестрикционными ферментами. Проводили реакции ПЦР с набором реагентов для ПЦР и согласно протоколу производителя (MBI Fermentas, Vilnius, Lithuania).
Секвенированием с применением способа прямого включения метки подтверждали наличие желательных мутаций. Клетки E. coli, несущие pQβ-243, поддерживали эффективный синтез белка массой 14 кДа, совместно мигрирующего на SDS-PAGE с контрольным белком оболочки Qβ, выделенным из частиц фага Qβ.
Полученная аминокислотная последовательность: (SEQ ID № 18)
AKLETVTLGKIGKDGKQTLVLNPRGVNPTNGVASLSQAGAVP
ALEKRVTVSVSQPSRNRKNYKVQVKIQNPTACTANGSCDPSVTRQ
KYADVTFSFTQYSTDEERAFVRTELAALLASPLLIDAIDQLNPAY
Конструирование pQβ-250:
В качестве исходной плазмиды для конструирования pQβ-250 применяли плазмиду pQβ-240. Посредством сайт-специфического мутагенеза создавали мутацию Lys2→Arg. Для синтеза мутантного фрагмента ПЦР, вставляемого по уникальным участкам рестрикции NcoI and HindIII в экспрессирующий вектор pQβ-185, применяли прямой праймер
5'-GGCCATGGCACGACTCGAGACTGTTACTTTAGG-3' (SEQ ID № 84)
и обратный праймер
5'-GATTTAGGTGACACTATAG-3' (SEQ ID № 85).
Проводили реакции ПЦР с набором реагентов для ПЦР и согласно протоколу производителя (MBI Fermentas, Vilnius, Lithuania).
Секвенированием с применением способа прямого включения метки подтверждали наличие желательных мутаций. Клетки E. coli, несущие pQβ-250, поддерживали эффективный синтез белка массой 14 кДа, совместно мигрирующего на SDS-PAGE с контрольным белком оболочки Qβ, выделенным из частиц фага Qβ.
Полученная аминокислотная последовательность: (SEQ ID № 19)
ARLETVTLGNIGRDGKQTLVLNPRGVNPTNGVASLSQAGAVP
ALEKRVTVSVSQPSRNRKNYKVQVKIQNPTACTANGSCDPSVTRQ
KYADVTFSFTQYSTDEERAFVRTELAALLASPLLIDAIDQLNPAY
Конструирование pQβ-251:
В качестве исходной плазмиды для конструирования pQβ-251 применяли плазмиду pQβ10. Посредством обращенной ПЦР создавали мутацию Lys16→Arg. Обращенные праймеры разрабатывали в обращенных направлениях хвост-к-хвосту:
5'-GATGGACGTCAAACTCTGGTCCTCAATCCGCGTGGGG-3' (SEQ ID № 86)
и
5'-CCCCACGCGGATTGAGGACCAGAGTTTGACGTCCATC-3' (SEQ ID № 87).
Продукты первой ПЦР применяли как матрицы для второй реакции ПЦР, в которой применяли прямой праймер
5'-AGCTCGCCCGGGGATCCTCTAG-3' (SEQ ID № 80)
и обратный праймер
5'-CGATGCATTTCATCCTTAGTTATCAATACGCTGGGTTCAG-3' (SEQ ID № 81).
Продукт второй ПЦР расщепляли XbaI и Mph1103I и клонировали в экспрессирующий вектор pQβ10, расщепленный теми же рестрикционными ферментами. Проводили реакции ПЦР с набором реагентов для ПЦР и согласно протоколу производителя (MBI Fermentas, Vilnius, Lithuania).
Секвенированием с применением способа прямого включения метки подтверждали наличие желательных мутаций. Клетки E. coli, несущие pQβ-251, поддерживали эффективный синтез белка массой 14 кДа, совместно мигрирующего на SDS-PAGE с контрольным белком оболочки Qβ, выделенным из частиц фага Qβ.
Полученная аминокислотная последовательность, кодируемая данной конструкцией, приведена в SEQ ID № 20.
Конструирование pQβ-259:
В качестве исходной плазмиды для конструирования pQβ-259 применяли плазмиду pQβ-251. Посредством сайт-специфического мутагенеза создавали мутацию Lys2→Arg. Для синтеза мутантного фрагмента ПЦР, вставляемого по уникальным участкам рестрикции NcoI and HindIII в экспрессирующий вектор pQβ-185, применяли прямой праймер
5'-GGCCATGGCACGACTCGAGACTGTTACTTTAGG-3' (SEQ ID № 84)
и обратный праймер
5'-GATTTAGGTGACACTATAG-3' (SEQ ID № 85).
Проводили реакции ПЦР с набором реагентов для ПЦР и согласно протоколу производителя (MBI Fermentas, Vilnius, Lithuania).
Секвенированием с применением способа прямого включения метки подтверждали наличие желательных мутаций. Клетки E. coli, несущие pQβ-259, поддерживали эффективный синтез белка массой 14 кДа, совместно мигрирующего на SDS-PAGE с контрольным белком оболочки Qβ, выделенным из частиц фага Qβ.
Полученная аминокислотная последовательность: (SEQ ID № 21)
AKLETVTLGNIGKDGKQTLVLNPRGVNPTNGVASLSQAGAVP
ALEKRVTVSVSQPSRNRKNYKVQVKIQNPTACTANGSCDPSVTRQ
KYADVTFSFTQYSTDEERAFVRTELAALLASPLLIDAIDQLNPAY
Основные способы экспрессии и очистки Qβ и мутантов Qβ.
Очистка
JM109 E. coli трансформировали экспрессирующей белок оболочки Qβ плазмидой. 5 мл жидкой среды LB, содержащей 20 мкг/мл ампициллина, инокулировали клонами, трансформированными экспрессирующими белок оболочки Qβ плазмидами. Инокулированную культуру инкубировали 16-24 часа при 37°C без перемешивания. Полученный инокулят последовательно разводили 1:100 в 100-300 мл свежей среды LB, содержащей 20 мкг/мл ампициллина, и инкубировали в течение ночи при 37°C без перемешивания. Полученный второй инокулят разводили 1:50 в среде M9, содержащей 1% казаминовых кислот и 0,2% глюкозы, в колбах и инкубировали в течение ночи при 37°C при перемешивании.
Очистка
Растворы и буферы для процедуры очистки
1. Буфер для лизиза LB
50 мМ Tris-HCl pH 8,0 с 5 мМ ЭДТА, 0,1% TritonХ100 и свежеприготовленный PMSF в концентрации 5 микрограмм на мл. Без лизоцима и ДНКазы.
2. SAS
Насыщенный сульфат аммония в воде
3. Буфер NET.
20 мМ Tris-HCl, pH 7,8 с 5 мМ ЭДТА и
150 мМ NaCl.
4. PEG
40% (мас./об.) полиэтиленгликоля 6000 в NET
Разрушение и лизис
Замороженные клетки ресуспендировали в LB при 2 мл/г клеток. Смесь подвергали воздействию ультразвука пять раз по 15 секунд при 22 кГц с интервалами в 1 мин для охлаждения раствора на льду. Затем лизат 1 час центрифугировали при 14000 об/мин с применением ротора Janecki K 60. Все стадии центрифугирования, описанные ниже, проводили с применением этого же ротора, если не указано иначе. Супернатант хранили при 4°C, в то время как клеточный дебрис дважды отмывали LB. После центрифугирования супернатанты лизата и отмытые фракции объединяли.
Фракционирование
К объединенному выше лизату при перемешивании капельно добавляли насыщенный раствор сульфата аммония. Объем SAS доводили до одной пятой общего объема для получения 20% насыщения. Раствор оставляли на ночь, а на следующие сутки центрифугировали 20 минут при 14000 об/мин. Осадок отмывали малым количеством 20% сульфата аммония и снова центрифугировали. Полученные супернатанты объединяли и капельно добавляли SAS для получения 40% насыщения. Раствор оставляли на ночь, а на следующие сутки центрифугировали 20 минут при 14000 об/мин. Полученный осадок растворяли в буфере NET.
Хроматография
Белок капсида или VLP повторно растворенный в буфере NET помещали в колонку с сефарозой CL-4B. В ходе хроматографии элюировали три пика. Первый пик в основном содержал мембраны и фрагменты мембран, и его не собирали. Капсиды содержались во втором пике, тогда как третий пик содержал другие белки E. coli.
Пиковые фракции объединяли и концентрацию NaCl доводили до конечной концентрации 0,65 М. При перемешивании капельно добавляли объем раствора PEG, соответствующий половине объединенной фракции пиков. Раствор оставляли на ночь без перемешивания. Капсидный белок осаждали центрифугированием 20 мин при 14000 об/мин. Затем его растворяли в минимальном объеме NET и снова помещали на колонку с сефарозой CL-4B. Пиковые фракции объединяли и осаждали сульфатом аммония при 60% насыщении (мас./об.). После центрифугирования и повторного растворения в буфере NET капсидный белок помещали на колонку с сефарозой CL-4B для повторной хроматографии.
Диализ и сушка
Полученные выше пиковые фракции объединяли и подвергали интенсивному диализу против стерильной воды и лиофилизировали для хранения.
Экспрессия и очистка Qβ-240
Клетки (JM 109 E. coli, трансформированные плазмидой pQβ-240) ресуспендировали в LB, пять раз по 15 секунд подвергали воздействию ультразвука (в бане из ледяной воды) и 1 час центрифугировали при 13000 об/мин. Супернатанты хранили при 4°C до дальнейшей обработки, в то время как дебрис 2 раза отмывали 9 мл LB, а в заключение 9 мл 0,7 М мочевины в LB. Все супернатанты объединяли и помещали на колонку с сефарозой CL-4B. Собранные пиковые фракции осаждали сульфатом аммония и центрифугировали. Затем повторно растворенный белок дополнительно очищали на колонке с сефарозой 2B, а в заключение на колонке с сефарозой 6B. Капсидный пик в заключение подвергали интенсивному диализу против воды и лиофилизировали, как описано выше. Сборку белка оболочки в капсид подтверждали электронной микроскопией.
Экспрессия и очистка Qβ-243
Клетки (RR1 E. coli) ресуспендировали в LB и обрабатывали, как описано в основной процедуре. Белок очищали двумя последовательными стадиями гель-фильтрации на колонке с сефарозой CL-4B и в заключении на колонке с сефарозой CL-2B. Пиковые фракции объединяли и лиофилизировали, как описано выше. Сборку белка оболочки в капсид подтверждали электронной микроскопией.
Экспрессия и очистка Qβ-250
Клетки (JM109 E. coli, трансформированные Qβ-250) ресуспендировали в LB и обрабатывали, как описано в основной процедуре. Белок очищали гель-фильтрацией на колонке с сефарозой CL-4B и в заключении на колонке с сефарозой CL-2B и лиофилизировали, как описано выше. Сборку белка оболочки в капсид подтверждали электронной микроскопией.
Экспрессия и очистка Qβ-259
Клетки (JM109 E. coli, трансформированные Qβ-259) ресуспендировали в LB и обрабатывали ультразвуком. Дебрис однократно отмывали 10 мл LB и второй раз 10 мл 0,7 М мочевины в LB. Белок очищали двумя последовательными стадиями гель-фильтрации на колонке с сефарозой CL-4B. Белок подвергали диализу и лиофилизировали, как описано выше. Сборку белка оболочки в капсид подтверждали электронной микроскопией.
ПРИМЕР 2
Вставка содержащего лизиновый остаток пептида в эпитоп c/e1 HBcAg (1-149)
Эпитоп c/e1 (остатки с 72 по 88) HBcAg расположен в верхней области на поверхности капсида вируса гепатита B(HBcAg). Часть данной области (пролин 79 и аланин 80) генетически заменили пептидом Gly-Gly-Lys-Gly-Gly (SEQ ID № 33), что привело к конструкции HBcAg-Lys (SEQ ID № 26). Введенный лизиновый остаток содержит на его боковой цепи реакционноспособную аминогруппу, которую можно применять для внутримолекулярного химического сшивания частиц HBcAg с любым антигеном, содержащим свободную цистеиновую группу.
ДНК HBcAg-Lys с аминокислотной последовательностью, приведенной в SEQ ID № 78, получали посредством ПЦР: Два фрагмента, кодирующих фрагменты HBcAg (аминокислотные остатки с 1 по 78 и с 81 по 149), раздельно амплифицировали посредством ПЦР. Праймеры, применяемые для данных ПЦР, также вносили последовательность ДНК, кодирующую пептид Gly-Gly-Lys-Gly-Gly (SEQ ID № 33). Фрагмент HBcAg (с 1 по 78) амлифицировали с pEco63 с применением праймеров EcoRIHBcAg(s) и Lys-HBcAg(as). Фрагмент HBcAg (с 81 по 149) амлифицировали с pEco63 с применением праймеров Lys-HBcAg(s) и HBcAg(1-149)Hind(as). Праймеры Lys-HBcAg(as) и Lys-HBcAg(s) на концах продуктов ПЦР вносили в комплементарные последовательности ДНК, позволяя слияние данных двух продуктов ПЦР при последующей объединяющей ПЦР. Собранные фрагменты амплифицировали посредством ПЦР с применением праймеров EcoRIHBcAg(s) и HbcAg(1-149)Hind(as).
Для ПЦР применяли 100 пмоль каждого олигонуклеотида и 50 нг матричной ДНК в 50 мл реакционных смесей с 2 единицами полимеразы Pwo, 0,1 мМ dNTP и 2 мМ MgSO4. Для обеих реакций цикличное изменение температуры проводили, как указано ниже: 2 минуты - 94°C; 30 циклов, состоящих из 94°C (1 минута), 50°C (1 минута), 72°C (2 минуты).
Последовательность праймеров:
EcoRIHBcAg(s):
(5'-CCGGAATTCATGGACATTGACCCTTATAAAG-3') (SEQ ID № 88);
Lys-HBcAg(as):
(5'-CCTAGAGCCACCTTTGCCACCATCTTCTAAATTAGTACCCACCCAG
GTAGC-3') (SEQ ID № 89);
Lys-HBcAg(s):
(5'-
GAAGATGGTGGCAAAGGTGGCTCTAGGGACCTAGTAGTCAGTTAT
GTC -3') (SEQ ID № 90);
HBcAg(1-149)Hind(as):
(5'-CGCGTCCCAAGCTTCTAAACAACAGTAGTCTCCGGAAG-3') (SEQ ID № 91).
Для слияния двух фрагментов ПЦР применяли 100 пмоль праймеров EcoRIHBcAg(s) и HBcAg(1-149)Hind(as) со 100 нг двух очищенных фрагментов ПЦР в 50 мл реакционной смеси, содержащей 2 единицы полимеразы Pwo, 0,1 мМ dNTP и 2 мМ MgSO4. Параметры циклов ПЦР составляли: 94°C - 2 минуты; 30 циклов, состоящих из 94°C (1 минута), 50°C (1 минута), 72°C (2 минуты). Объединенный продукт ПЦР анализировали электрофорезом в агарозном геле, очищали и расщепляли рестрикционными ферментами EcoRI и HindIII 19 часов в соответствующем буфере. Расщепленные фрагменты ДНК лигировали в расщепленный EcoRI/HindIII вектор pKK для получения экспрессиурющего вектора pKK-HBcAg-Lys. Вставку продукта ПЦР в вектор анализировали рестрикционным анализом EcoRI/HindIII и определением последовательности вставки ДНК.
ПРИМЕР 3
Экспрессия и очистка HBcAg-Lys.
Штаммы K802 или JM109 E. coli трансформировали pKK-HBcAg-Lys. 1 мл ночной культуры бактерий применяли для инокуляции 100 мл среды LB содержащей 100 мкг/мл ампициллина. Данную культуру 4 часа растили при 37°C до тех пор, пока OD при 600 нм не достигала приблизительно 0,8. Индукцию синтеза HBcAg-Lys проводили добавлением IPTG до конечной концентрации 1 мМ. После индукции бактерии дополнительно перемешивали 4 часа при 37°C. Бактерии собирали центрифугированием 15 минут при 5000×g. Осадок замораживали при -80°C. Осадок размораживали и ресуспендировали в буфере для лизиса бактерий (10 мМ Na2HPO4, pH 7,0, 30 мМ NaCl, 0,25% Tween-20, 10 мМ ЭДТА), дополненном 200 мкг/мл лизоцимом и 10 мкл бензоазы (Merck). Клетки 30 минут инкубировали при комнатной температуры и разрушали обработкой ультразвуком. Для индукции экспрессии HBcAg-Lys посредством IPTG применяли клетки E. coli, несущие экспрессирующую плазмидуpKK-HBcAg-Lys или контрольную плазмиду. До добавления IPTG из бактериальной культуры, несущей плазмиду pKK-HBcAg-Lys, и из культуры, несущей контрольную плазмиду, отбирали образцы. Через четыре часа после добавления IPTG из бактериальной культуры, содержащей плазмиду pKK-HBcAg-Lys, и из контрольной культуры снова отбирали образцы. Экспрессию белка контролировали посредством SDS-PAGE с последующим окрашиванием Кумасси.
Затем лизат 30 минут центрифугировали при 12000×g для удаления нерастворимого клеточного дебриса. Супернатант и осадок анализировали посредством вестерн-блоттинга с применением моноклонального антитела против HBcAg (YVS1841, полученного из Accurate Chemical and Scientific Corp., Westbury, NY, USA), указывавшего на то, что значительное количество белка HBcAg-Lys являлось растворимым. Кратко, лизаты из клеток E. coli, экспрессирующих HBcAg-Lys, и из контрольных клеток 30 минут центрифугировали при 14000×g. Супернатант (= растворимая фракция) и осадок (= нерастворимая фракция) разделяли и разбавляли буфером с SDS для образца до равных объемов. Образцы анализировали посредством SDS-PAGE с последующим вестерн-блоттингом с моноклональным антителом против HBcAg YVS 1841.
Очищенный клеточный лизат применяли для центрифугирования в ступенчатом градиенте с применением ступенчатого градиента сахарозы, состоящего из 4 мл 65% раствора сахарозы, с наслоенными 3 мл 15% раствора сахарозы и с последующими 4 мл бактериального лизата. Образец центрифугировали 3 часа при 100000×g при 4°C. После центрифугирования собирали 1 мл фракций сверху градиента и анализировали посредством SDS-PAGE с последующим окрашиванием Кумасси. Белок HBcAg-Lys выявляли посредством окрашивания Кумасси.
Белок HBcAg-Lys являлся обогащенным на границе раздела 15 и 65% сахарозы, означая, что он сформировал капсидную частицу. Большинство бактериальных белков оставались в верхнем слое градиента без сахарозы, следовательно, центрифугирование частиц HBcAg-Lys в ступенчатом градиенте вело к обогащению и к частичной очистке частиц.
Экспрессию и очистку HBcAg-Lys в целом проводили, как указано ниже. Ночную культуру готовили инокулированием одной колонии в 100 мл LB, 100 мкг/мл ампициллина и выращиванием культуры в течение ночи при 37°C. 25 мл предварительной культуры на следующие сутки разбавляли 800 мл среды LB с ампициллином и культуру растили до оптической плотности OD600, равной 0,6-0,8. Затем культуру индуцировали 1 мМ IPTG, и оставляли расти еще 4 часа. Клетки собирали и лизировали, по существу, как описано выше.
Затем HBcAg-Lys очищали посредством первого осаждения белка из очищенного клеточного лизата в сульфате аммония (30% насыщения), загружая затем повторно растворенный осадок на колонку для гель-фильтрации (сефакрил S-400, Pharmacia). Объединенные фракции снова осаждали сульфатом аммония, повторно растворяли и второй раз помещали на ту же колонку для гель-фильтрации. В заключение фракции собирали и концентрировали, а концентрации оценивали с применением способа Брэдфорда (BioRad).
ПРИМЕР 4
Конструирование лишенного цистеиновых остатков и содержащего вставленный лизиновый остаток HBcAg
Коровый антиген гепатита (HBcAg), обозначаемый здесь как HBcAg-lys-2cys-Mut, лишенный цистеиновых остатков в позициях, соответствующих 48 и 107 в SEQ ID № 25 и содержащий вставленный лизиновый остаток, сконструировали с применением следующих способов.
Посредством первой раздельной амплификации трех фрагментов гена HBcAg-Lys, полученных, как описано выше в примере 2, со следующими ниже сочетаниями праймеров для ПЦР внесены две мутации. Для получения гена HBcAg-lys-2cys-Mut применяли способы ПЦР и традиционные способы клонирования.
Кратко, для получения фрагмента 1 применяли следующие ниже праймеры:
Праймер 1: EcoRIHBcAg(s)
CCGGAATTCATGGACATTGACCCTTATAAAG (SEQ ID № 88)
Праймер 2: 48as
GTGCAGTATGGTGAGGTGAGGAATGCTCAGGAGACTC (SEQ ID № 92)
Для получения фрагмента 2 применяли следующие ниже праймеры:
Праймер 3: 48s
GAGTCTCCTGAGCATTCCTCACCTCACCATACTGCAC (SEQ ID № 93)
Праймер 4: 107as
CTTCCAAAAGTGAGGGAAGAAATGTGAAACCAC (SEQ ID № 94)
Для получения фрагмента 3 применяли следующие ниже праймеры:
Праймер 5: HBcAg149hind-as
CGCGTCCCAAGCTTCTAAACAACAGTAGTCTCCGGAAGCGTTGATAG (SEQ ID № 95)
Праймер 6: 107s
GTGGTTTCACATTTCTTCCCTCACTTTTGGAAG (SEQ ID № 96)
Затем фрагменты 1 и 2 объединяли с праймерами для ПЦР EcoRIHBcAg(s) и 107as для получения фрагмента 4. Затем фрагмент 4 и фрагмент 3 объединяли с праймерами EcoRIHBcAg(s) and HBcAg149hind-as для получения полноразмерного гена. Затем полноразмерный ген расщепляли ферментами EcoRI (GAATTC) и HindIII (AAGCTT) и клонировали в вектор pKK (Pharmacia), расщепленный по тем же участкам рестрикции. Экспрессию и очистку HBcAg-lys-2cys-Mut проводили, как указано в примере 3.
ПРИМЕР 5
Конструирование HBcAg1-185-Lys.
Коровый антиген гепатита (HBcAg) 1-185 модифицировали, как описано в примере 2. Часть (пролин 79 и аланин 80) области эпитопа c/e1 (остатки с 72 по 88) генетически заменили пептидом Gly-Gly-Lys-Gly-Gly (SEQ ID № 33), получив в результате конструкцию HBcAg-Lys (SEQ ID № 26). Введенный лизиновый остаток содержит на его боковой цепи реакционноспособную аминогруппу, которую можно применять для внутримолекулярного химического сшивания частиц HBcAg с любым антигеном, содержащим свободную цистеиновую группу. Для получения гена HBcAg1-185-Lys применяли способы ПЦР и традиционные способы клонирования
Последовательность Gly-Gly-Lys-Gly-Gly (SEQ ID № 33) вставляли посредством амплификации двух отдельных фрагментов гена HBcAg с pEco63, как описано выше в примере 2, и последующего слияния двух фрагментов посредством ПЦР для сборки полноразмерного гена. Применяли следующие сочетания праймеров для ПЦР:
Фрагмент 1:
Праймер 1: EcoRIHBcAg(s) ((SEQ ID № 88), см. пример 2)
Праймер 2: Lys-HBcAg(as) ((SEQ ID № 89), см. пример 2)
Фрагмент 2:
Праймер 3: Lys-HBcAg(s) ((SEQ ID № 90), см. пример 2)
Праймер 4: HBcAgwtHindIIII
CGCGTCCCAAGCTTCTAACATTGAGATTCCCGAGATTG (SEQ ID № 97)
Сборка:
Праймер 1: EcoRIHBcAg(s) ((SEQ ID № 88), см. пример 2)
Праймер 2: HBcAgwtHindIIII (SEQ ID № 97)
Собранный полноразмерный ген затем расщепляли ферментами EcoRI (GAATTC) и HindIII (AAGCTT) и клонировали в вектор pKK (Pharmacia), расщепленный по тем же участкам рестрикции.
ПРИМЕР 6
Слияние эпитопа пептида в области MIR HBcAg.
Остатки HBcAg1-185 79 и 80 заменяли эпитопом CεH3 с последовательностью VNLTWSRASG (SEQ ID № 98). Последовательность CεH3 происходит из последовательности третьего константного домена тяжелой цепи человеческого IgE. Эпитоп вставляли в последовательность HBcAg1-185 с применением способа объединяющей ПЦР. На первой стадии ПЦР ген HBcAg1-185, происходящий из клона ATCC pEco63 и амплифицированный с праймерами HBcAg-wt EcoRI fwd и HBcAg-wt Hind III rev, применяли в качестве матрицы в двух отдельных реакциях для амплификации двух фрагментов, содержащих элементы последовательности, кодирующей последовательность CεH3. Затем данные два фрагмента объединяли на второй стадии ПЦР, в объединяющей реакции ПЦР.
Сочетания праймеров на первой стадии ПЦР: СεР3fwd с HBcAg-wt HindIII rev и HBcAg-wt EcoRI fwd с СεР3rev. В объединяющей реакции ПЦР два фрагмента, выделенных на первой стадии ПЦР, сначала объединяли в течение 3 циклов ПЦР без внешних праймеров, которые впоследствии добавляли в реакционную смесь для следующих 25 циклов. Внешние праймеры: HBcAg-wt EcoRI fwd и HBcAg-wt Hind III rev.
Продукт ПЦР клонировали в pKK223.3 с применением участков EcoRI и HindIII для экспрессии в E. coli (см. пример 2). Гибридную VLP экспрессировали в E. coli и очищали, как описано в примере 2. Объем элюции, при котором HBcAg1-185-СεР3 элюировал при гель-фильтрации, показывал сборку гибридных белков в гибридную VLP.
Последовательности праймеров:
CεH3fwd:
5'GTT AAC TTG ACC TGG TCT CGT GCT TCT GGT GCA TCC AGG GAT CTA GTA GTC 3'(SEQ ID № 99)
V N L T W S R A S G A80 S R D L V V86 (SEQ ID № 100)
CεH3rev:
5' ACC AGA AGC ACG AGA CCA GGT CAA GTT AAC ATC TTC CAA ATT ATT ACC CAC 3' (SEQ ID № 101)
D78 E L N N G V72 (SEQ ID № 102)
HBcAg-wt EcoRI fwd:
5' CCGgaattcATGGACATTGACCCTTATAAAG (SEQ ID № 103)
HBcAG-wt Hind III rev:
5' CGCGTCCCaagcttCTAACATTGAGATTCCCGAGATTG (SEQ ID № 104)
ПРИМЕР 7
Слияние эпитопа пептида грелина с областью MIR HBcAg.
Остатки 79 и 80 HBcAg1-185 замещали эпитопом пептида грелина с последовательностью: EHQRVQQRKE (SEQ ID № 56). Сконструировали два перекрывающихся праймера с применением той же стратеги, что и описанная в примере 6, и посредством объединяющей ПЦР конструировали гибридный белок. Продукт ПЦР клонировали в вектор pKK223.3 и экспрессировали в K802 E. coli. Гибридные VLP экспрессировали и очищали, как описано в примере 3.
ПРИМЕР 8
Слияние эпитопа пептида грелина с C-концом укороченного по позиции 19 удлинения CP белка A1 Qβ.
В реакции ПЦР с pQβ10 в качестве матрицы применяли праймер, отжигающийся к 5'-концу гена A1 Qβ, и праймер, отжигающийся к 3'-концу гена A1 Qβ и дополнительно содержащий элемент последовательности, кодирующий фрагмент грелина с последовательностью KKPPAKLQPR (SEQ ID № 52). Продукт ПЦР клонировали в pQβ10 (Kozlovska T. M. et al., Gene 137: 133-37 (1993)), а гибридную VLP экспрессировали и очищали, как описано в примере 1.
ПРИМЕР 9
Вставка эпитопа пептида грелина между положениями 2 и 3 белка оболочки fr.
В участок Bsp119I вектора pFrd8 (Pushko, P. et al., Prot. Eng. 6: 883-91 (1993)) посредством стандартных способов молекулярной биологии вставляли комплементарные праймеры, кодирующие последовательность пептида грелина с последовательностью EHQRVQQRKE (SEQ ID № 56) и содержащие совместимые с Bsp119I концы и дополнительные нуклеотиды nucleotides, дающие возможность вставки с сохранением рамки считывания. Альтернативно "липкие концы" вектора pFrd8 после расщепления Bsp119I дополняли фрагментом Кленова и в pFrd8 лигировали олигонуклеотиды, кодирующие последовательность пептида грелина мышиного пептида грелина и дополнительные нуклеотиды для клонирования с сохранением рамки считывания, после обработки фрагментом Кленова. Клоны со вставкой в правой ориентации анализировали посредством определения последовательности. Экспрессию и очистку химерного гибридного белка в JM109 E. coli или K802 E. coli проводили, как описано у Pushko, P. et al, Prot. Eng. 6: 883-91 (1993), кроме стадий хроматографии, которые проводили с применением сефарозы CL-4B или сефакрила S-400 (Pharmacia). Клеточный лизат осаждали сульфатом аммония и очищали посредством двух последовательных стадий очистки гель-фильтрацией подобно процедуре, описанной для Qβ в примере 1.
ПРИМЕР 10
Вставка эпитопа пептида грелина между положениями 67 и 68 белка p1 Ty1 в вектор pOGS8111.
Синтезировали два комплементарных нуклеотида, кодирующих последовательность EHQRVQQRKE (SEQ ID № 56) пептида человеческого грелина, с концами, совместимыми с участком NheI pOGS8111. Добавляли дополнительные нуклеотиды для обеспечения вставки без сдвига рамки считывания последовательности, кодирующей эпитоп мышиного грелина, по описанию EP06777111. Фланкирующие вставленный эпитоп аминокислоты AS и SS кодируются измененными участками NheI, что происходит в результате вставки олигонуклеотида в ген TyA(d) pOGS8111.
POGS8111 трансформировали в штамм MC2 S. cervisiae для экспрессии гибридной VLP Ty, как описано в EP0677111 и указано здесь. Гибридные VLP Ty очищали посредством ультрацентрифугирования на градиенте сахарозы, как описано в EP0677111.
ПРИМЕР 11
Вставка эпитопа пептида грелина в главный капсидный белок L1 вируса папилломы типа 1 (BPV-1).
Последовательностью, кодирующей эпитоп грелина с последовательностью EHQRVQQRKE (SEQ ID № 56), замещали последовательность, кодирующую аминокислоты 130-136 гена L1 BPV-1, клонированного в вектор pFastBac1 (GIBCO/BRL), как описано (Chackerian, B. et al., Proc. Natl. Acad. USA 96: 2373-2378 (1999)). Последовательность конструкции проверяли посредством анализа последовательности нуклеотидов. С применением бакуловирусной системы GIBCO/BRL получали рекомбинантный бакуловирус, как описано у производителя. Гибридные VLP очищали от зараженных бакуловирусом клеток Sf9, как описано у Kirnbauer, R. et al., Proc. Natl. Acad. Sci. 89: 12180-84 (1992) и Greenstone, H. L., et al., Proc. Natl. Acad. Sci. 95: 1800-05 (1998).
ПРИМЕР 12
Иммунизация мышей слитыми с VLP пептидами грелина.
Гибридные VLP, представляющие эпитоп мышиного грелина с последовательностью EHQRVQQRKE (SEQ ID № 56), полученный в примерах 7, 9, 10 и 11, применяли для иммунизации мышей, как описано в примере 15. Полученные у иммунизированных мышей сыворотки анализировали в специфичном к грелину ELISA, как описано в примере 14.
Эффект вакцины проверяли последующим увеличением массы мышей и измерением потребления пищи.
ПРИМЕР 13
Связывание мышиного грелина и пептидов мышиного грелина с белком капсида Qβ: вакцины из пептидов грелина.
Химически синтезировали следующие пептиды грелина:
cGhrel: CGSSFLSPEHQKAQQRKESKKPPAKLQPR (SEQ ID № 77)
GhreIC: GSSFLSPEHQKAQQRKESKKPPAKLQPRGC (SEQ ID № 105)
cGhrQ14: CGSSFLSPEHQKAQRKESKKPPAKLQPR(SEQ ID № 106)
GhrQ14C: GSSFLSPEHQKAQRKESKKPPAKLQPRGC (SEQ ID № 107)
Ghrel24-33C GGSSFLSPEHQGC (SEQ ID № 132)
cGhrel42-51 CKKPPAKLQPR (SEQ ID № 133)
cGhrel31-40 CEHQKAQQRKE (SEQ ID № 134)
cGhrel31-41 CEHQKAQQRKES (SEQ ID № 135)
cGhrel28-37 CLSPEHQKAQQ (SEQ ID № 136)
содержащие добавленный на N- или C-конце цистеиновый остаток для связывания с VLP или пилями и применяемые для химического связывания с Qβ, как описано далее.
Раствор 5 мл 140 мМ капсидного белка Qβ в 20 мМ Hepes, 150 мМ NaCl pH 7,4 на 30 минут подвергают взаимодействию со 108 мкл 65 мМ раствора SMPH (Pierce) в H2O при 25°C на качающемся шейкере. Затем реакционный раствор дважды подвергают диализу в течение 2 часов против 5 л 20 мМ Hepes, 150 мМ NaCl, pH 7,2 при 4°C. Затем 100 мкл диализированной реакционной смеси подвергали взаимодействию или с 28,6 мкл 10 мМ маточного раствора (в ДМСО) пептида грелина (соотношение пептид/капсидный белок Qβ составляет 1:10). Реакцию связывания проводят 2 часа при 15°C в водяной бане. Затем реакционную смесь подвергают диализу в течение 24 часов против 2×5 л 20 мМ Hepes, 150 мМ NaCl, pH 7,2 при 4°C.
Продукты связывания центрифугируют, а супернатанты и осадки анализируют на 16% гелях SDS-PAGE в восстанавливающих условиях. Гели окрашивают кумасси бриллиантовым голубым.
Связывание пептидов грелина с белком капсида fr
Раствор 120 мМ белка капсида fr в 20 мМ Hepes, 150 мМ NaCl pH 7,2 30 минут подвергают взаимодействию с 10-кратным молярным избытком SMPH (Pierce)), разведенным из маточного раствора в ДМСО, при 25°C на качающемся шейкере. Затем реакционный раствор дважды подвергают диализу в течение 2 часов против 1 л 20 мМ Hepes, 150 мМ NaCl, pH 7,2 при 4°C. Затем диализированную реакционную смесь fr в течение ночи при 16°C на качающемся шейкере подвергают взаимодействию с эквимолярной концентрацией пептида грелина или с соотношением пептид грелина/fr, составляющим 1:2. Продукты связывания анализируют посредством SDS-PAGE.
Связывание пептидов грелина с HBcAg-Lys-2cys-Mut
Раствор 120 мМ HBcAg-Lys-2cys-Mut в 20 мМ Hepes, 150 мМ NaCl pH 7,2 30 минут подвергают взаимодействию с 10-кратным молярным избытком SMPH (Pierce)), разведенным из маточного раствора в ДМСО, при 25°C на качающемся шейкере. Затем реакционный раствор дважды подвергают диализу в течение 2 часов против 1 л 20 мМ Hepes, 150 мМ NaCl, pH 7,2 при 4°C. Затем диализированную реакционную смесь HBcAg-Lys-2cys-Mut в течение ночи при 16°C на качающемся шейкере подвергают взаимодействию с эквимолярной концентрацией пептида грелина или с соотношением пептид грелина/HBcAg-Lys-2cys-Mut, составляющим 1:2. Продукты связывания анализируют посредством SDS-PAGE.
Связывание пептидов грелина с пилями
Раствор 125 мМ пилей типа 1 E. coli в 20 мМ Hepes, pH 7,2 60 минут подвергают взаимодействию с 50-кратным молярным избытком сшивающего средства SMPH (Pierce)), разведенным из маточного раствора в ДМСО, при RT на качающемся шейкере. Реакционную смесь обессоливают на колонке PD-10 (Amersham-Pharmacia Biotech). Собирают содержащие белок фракции, элюирующиеся с колонки, и обессоленный дериватизированный белок пилей в течение ночи при 16°C на качающемся шейкере подвергают взаимодействию с пептидами грелина в эквимолярном соотношении или в соотношении пептида с пилями, составляющем 1:2. Продукты связывания анализируют посредством SDS-PAGE.
ПРИМЕР 14
Иммунизация мышей конъюгатами VLP-пептид грелина.
Иммунизация мышей связанными с Qβ пептидами грелина
Самок мышей C57BL/6 иммунизируют перечисленными в примере 13 пептидами мышиного грелина, связанными с Qβ, смешанными с квасцами, растворенными в IFA, или без адъюванта. 50 мкг инъецируют подкожно на 0, 14 и 28 сутки. У мышей после иммунизации ретроорбитально собирали кровь на 14, 21, 28, 42 сутки и затем через месячные интервалы после этого. Сыворотку анализировали с применением специфичного к пептидам грелина ELISA.
ПРИМЕР 15
Связывание C-грелина и грелина-GC с белком оболочки Qβ
2 мл раствора 2,0 мг/мл капсидного белка Qβ в 20 мМ Hepes, 150 мМ NaCl pH 7,2 30 минут при 25°C подвергали взаимодействию со 114,4 мкл раствора SMPH (Pierce) (из 50 мМ маточного раствора, разведенного ДМСО). Затем реакционный раствор дважды подвергали диализу в течение 2 часов против 2 л 20 мМ Hepes, 150 мМ NaCl, pH 7,2 при 4°C.
Затем диализированную дериватизированную VLP Qβ подвергали реакции с пептидом мышиного C-грелина (SEQ ID: № 77) или мышиного грелина-GC (SEQ ID: № 105) в течение 2 часов при 15°C в 20 мМ Hepes, 150 мМ NaCl, pH 7,2. Реакцию связывания проводили в нескольких аликвотах, где 1 мл дериватизированной VLP Qβ (в концентрации 2 мг/мл) подвергали реакции с 286 мкл 10 мМ раствора пептида. Для реакции связывания добавляли 10% ацетонитрил. Реакционный раствор однократно подвергали диализу в течение 2 часов, а затем - в течение ночи против 2 л 20 мМ Hepes, 150 мМ NaCl, pH 7,2 при 4°C.
Продукты связывания анализировали на 16% гелях SDS-PAGE. Гели окрашивали кумасси бриллиантовым голубым. Результаты представлены на фигуре 1. Продукты связывания можно выявить в окрашенных кумасси гелях и отчетливо демонстрируют ковалентное связывание C-грелина или грелина-GC с капсидным белком Qβ. Полосы, соответствующие связыванию одного, двух, трех или четырех пептидов, связанных с одной субъединицей, обозначены стрелками.
На фигуре 1 представлены продукты связывания реакции мышиного C-грелина или мышиного грелина-GC с капсидным белком Qβ. Полосы 2 и 4 демонстрируют продукты связывания в растворимой фракции Qβ-C-грелин и грелин-GC-Qβ соответственно. В нерастворимой фракции находится очень мало продукта.
ПРИМЕР 16
Иммунизация мышей связанным с капсидным белком Qβ C-грелином или грелином-GC
Самок мышей C57BL/6 (5 на группу) вакцинировали или мышиным C-грелином (SEQ ID: № 77) или мышиным грелином-GC (SEQ ID: № 105), связанным с капсидным белком Qβ. 50 мкг диализированной вакцины из каждого образца разводили в PBS до объема 200 мкл и инъецировали подкожно (по 100 мкл с двух сторон брюшка) на 0, 14 и 42 сутки. Вакцину вводили без дополнительного адъюванта. В качестве контроля группу мышей иммунизировали PBS. У мышей на 0, 14, 21, 42 сутки ретроорбитально собирали кровь и их сыворотку анализировали с применением специфичного к грелину ELISA (описанном в примере 17). Кроме того, у мышей в течение эксперимента через регулярные контролировали массу тела и потребление пищи (описано примере 18).
ПРИМЕР 17
Выявление специфичных к грелину антител в ELISA
Планшеты ELISA покрывали мышиным грелином с октаноилированным серином (Bachem, Product № H-4862) в концентрации 20 мкг/мл. Планшеты блокировали, а затем инкубировали с серийно разведенными мышиными сыворотками от 14, 21 и 42 суток. Связывание антител выявляли с применением ферментативно меченных антител против мышиных IgG. В качестве контроля также тестировали сыворотку до иммунизации (фигура 2).
У мышей, иммунизированных Qβ-C-грелином или грелином-CG-Qβ, средний титр на 21 сутки достигал 13000 и 8000 соответственно (фигура 2). Сыворотки до иммунизации или сыворотки от мышей, иммунизированных PBS, не показали какой-либо активности в отношении грелина. Это четко демонстрирует, что конъюгат грелин-VLP способен индуцировать высокий титр антител против грелина, даже если он является собственным белком.
ПРИМЕР 18
Эффективность экспериментов с C-грелином или грелином-GC, связанным с капсидным белком Qβ
Самок мышей C57BL/6 (5 на группу) вакцинировали мышиным C-грелином или грелином-GC, связанным с капсидным белком Qβ, как описано в примере 16. В качестве контроля группу мышей иммунизировали PBS. Всех мышей между сутками 0 и 14 содержали на нормальной диете. Затем всех мышей содержали на диете с высоким содержанием жира (45%) для облегчения развития индуцированного диетой ожирения. Пищу и воду вводили свободно. У отдельных мышей контролировали изменения массы тела, а также через регулярные интервалы после иммунизации (на 5, 11, 14, 21, 28, 35, 40, 49 и 55 сутки после иммунизации) контролировали потребление пищи и воды на клетку (т.е. группу) (см. фигуру 3).
У мышей, иммунизированных мышиным Qβ-C-грелином или мышиным грелином-CG-Qβ, достигалось среднее уменьшение потребления пищи приблизительно на 30-40% по сравнению с контрольными мышами, иммунизированными PBS (фигура 3). Это отчетливо демонстрирует, что конъюгат грелин-VLP способен подавлять аппетит и уменьшать потребление пищи.
ПРИМЕР 19
Эффективность экспериментов с C-грелином или грелином-GC, связанным с капсидным белком Qβ в животной модели ожирения
Самок мышей C57BL/6 ob/ob (5 на группу) вакцинировали мышиным C-грелином или грелином-GC, связанным с капсидным белком Qβ. Контрольных мышей иммунизировали PBS или только VLP Qβ. 50 мкг диализированной вакцины или контрольного белка растворяют в объеме PBS 200 мкл и подкожно инъецируют (по 100 мкл с двух сторон брюшка) с квасцами или без на 0 сутки. Мышей подвергали вторичной стимуляции соответствующим препаратом на 14, 28 и 42 сутки.
У мышей ретроорбитально собирают кровь на 0, 14, 21, 42, 56, 70 сутки и затем через месячные интервалы. Сыворотки анализируют на наличие антител, специфичных к грелину, в специфичном к грелину ELISA, как описано в примере 17. Если титры специфичных к грелину антител значительно уменьшаются в течение периода эксперимента, мышей соответственно подвергают вторичной стимуляции. Действие вакцины проверяли измерением потребления пищи в условиях отсутствия ограничений и последующего увеличения массы мышей, как описано в примере 18.
ПРИМЕР 20
Клонирование гена белка оболочки AP205
кДНК белка оболочки (CP) AP205 (SEQ ID № 28) собирали из двух фрагментов кДНК, полученных из РНК фага AP205 с применением способа обратной транскрипции-ПЦР и клонирования в коммерческую плазмиду pCR 4-TOPO для определения последовательности. Способы обратной транскрипции хорошо известны специалистам в соответствующей области. Первый фрагмент, содержащийся в плазмиде p205-246, содержал 269 нуклеотидов вверх от последовательности CP и 74 нуклеотида, кодирующих первые 24 N-концевых аминокислот CP. Второй фрагмент, содержащийся в плазмиде p205-262, содержал 364 нуклеотидов, кодирующих аминокислоты l2-131 CP и дополнительные 162 нуклеотида вниз от последовательности CP. И p205-246 и p205-262 представляли собой великодушный подарок J. Klovins.
Для слияния обоих фрагментов CP из плазмид p205-246 и p205-262 в одну полноразмерную последовательность CP посредством двухстадийной ПЦР сконструировали плазмиду 283.-58.
Примеряли прямой праймер p1.44, содержащий участок NcoI для клонирования в плазмиду pQb185, или p1.45,содержащий участок XbaI для клонирования в плазмиду pQb10, и обратный праймер p1.46, содержащий рестрикционный участок HindIII (последовательности, распознаваемые ферментами рестрикции, подчеркнуты):
p1.44 5'-AACCATGGCAAATAAGCCAATGCAACCG-3' (SEQ ID № 139)
p1.45 5'-AATCTAGAATTTTCTGCGCACCCATCCCGG-3' (SEQ ID № 140)
p1.46 5'-AAAAGCTTAAGCAGTAGTATCAGACGATACG-3' (SEQ ID № 141)
Для амплификации фрагментов первой ПЦР применяли два дополнительных праймера: p1.47, отжигающийся к 5'-концу фрагмента, содержащегося в p205-262, и p1.48, отжигающийся к 3'концу фрагмента, содержащегося в плазмиде p205-246. Праймеры p1.47 и p1.48 комплементарны друг другу.
p1.47: 5'-GAGTGATCCAACTCGTTTATCAACTACATTTTCAGCAAGTCTG-3' (SEQ ID № 142)
p1.48: 5'-CAGACTTGCTGAAAATGTAGTTGATAAACGAGTTGGATCACTC-3' (SEQ ID № 143)
В первых двух реакциях ПЦР получали два фрагмента. Первый фрагмент получали с праймерами p1.45 и p1.48 и матрицей p205-246. Второй фрагмент получали с праймерами p1.47 и p1.46, и матрицей p205-262. Оба фрагмента применяли в качестве матриц для второй реакции ПЦР, расширение сращивания-перекрывания, с сочетанием праймеров p1.45 и p1.46 или p1.44 и p1.46. Продукт двух реакций ПЦР второй стадии расщепляли XbaI или NcoI соответственно и HindIII и клонировали в те же участки рестрикции в pQb10 или pQb185 соответственно два полученных из pGEM экспрессирующих вектора под контролем промотора триптофанового оперона E. coli.
Получали две плазмиды, pAP283-58 (SEQ ID № 27), содержащую ген, кодирующий CP wt AP205 (SEQ ID № 28) в pQb10, и pAP281-32 (SEQ ID № 30) с мутацией Pro5→Thr (SEQ ID № 29), в pQbl85. Последовательности белков оболочки проверяли посредством определения последовательности ДНК. PAP283-58 содержала 49 нуклеотидов вверх от кодона ATG CP, вниз от участка XbaI и содержала предполагаемый исходный участок прикрепления рибосомы мРНК белка оболочки.
ПРИМЕР 21:
Экспрессия и очистка рекомбинантной VLP AP205
A. Экспрессия рекомбинантной VLP AP205
JM109 E. coli трансформировали плазмидой pAP283-58. 5 мл жидкой среды LB с 20 мкг/мл ампициллина инокулировали одной колонией и 16-24 часов инкубировали при 37°C без перемешивания.
Полученный инокулят разбавляли 1:100 в 100-300 мл среды LB, содержащей 20 мкг/мл ампициллина, и в течение ночи инкубировали при 37°C без перемешивания. Полученный второй инокулят разбавляли 1:50 в среде 2TY medium, содержащей 0,2 % глюкозы и фосфата для буферизации, и в течение ночи инкубировали при 37°C на шейкере. Клетки собирали центрифугированием и замораживали при -80°C.
B. Очистка рекомбинантной VLP AP205
Растворы и буферы:
Буфер для лизиса
50 мМ Tris-HCl pH 8,0 с 5 мМ ЭДТА, 0,1% TritonХ100 и PMSF при 5 микрограммах на мл.
SAS
Насыщенный сульфат аммония в воде
Буфер NET.
20 мМ Tris-HCl, pH 7,8 с 5 мМ ЭДТА и
150 мМ NaCl.
PEG
40% (мас./об.) полиэтиленгликоля 6000 в NET
Лизис
Замороженные клетки ресуспендировали в буфере для лизиса при 2 мл/г клеток. Смесь подвергали воздействию ультразвука пять раз по 15 секунд при 22 кГц с интервалами в 1 мин для охлаждения раствора на льду. Затем лизат 20 минут центрифугировали при 12000 об/мин с применением ротора F34-6-38 (Eppendorf). Все стадии центрифугирования, описанные ниже, проводили с применением этого же ротора, если не указано иначе. Супернатант хранили при 4°C, в то время как клеточный дебрис дважды отмывали буфером для лизиса. После центрифугирования супернатанты лизата и отмытые фракции объединяли.
Для очистки VLP AP205 дополнительно можно применять осаждение сульфатом аммония. На первой стадии выбирают концентрацию сульфата аммония, при которой VLP AP205 не осаждается. Полученный осадок отбрасывают. На следующей стадии выбирают концентрацию сульфата аммония, при которой VLP AP205 осаждается количественно, а VLP AP205 выделяют из осадка, образованного на данной стадии, посредством центрифугирования (14000 об/мин, 20 мин). Полученный осадок растворяют в буфере NET.
Хроматография:
Капсидный белок из объединенных супернатантов помещали на колонку с сефарозой 4B column (2,8×70 см) и элюировали буфером NET при 4 мл/час/фракцию. Фракции 28-40 собирали и осаждали сульфатом аммония при насыщении 60%. Фракции анализировали посредством SDS-PAGE и вестерн-блоттинга с антисывороткой, специфичной к AP205 до осаждения. Выделенный при центрифугировании осадок повторно растворяли в буфере NET и помещали на колонку с сефарозой 2B (2,3×65 см), проводя элюцию при 3 мл/час/фракцию. Фракции анализировали посредством SDS- PAGE, а фракции 44-50 собирали, объединяли и осаждали сульфатом аммония при насыщении 60%. Выделенный при центрифугировании осадок повторно растворяли в буфере NET и очищали на колонке с сефарозой 6B (2,5×47 см), проводя элюцию при 3 мл/час/фракцию. Фракции анализировали посредством SDS-PAGE. Собирали фракции 23-27, концентрацию соли доводили до 0,5 M и осаждали PEG 6000, добавленного из 40% маточного раствора в воде, до конечной концентрации 13,3%. Выделенный посредством центрифугирования осадок повторно растворяли в буфере NET и помещали на ту же колонку с сефарозой 2B, что и выше, проводя элюцию таким же способом. Собирали фракции 43-53 и осаждали сульфатом аммония при насыщении 60%. Выделенный посредством центрифугирования осадок повторно растворяли в воде и полученный раствор белка интенсивно диализировали против воды. Возможным получить являлось приблизительно 10 мг очищенного белка на грамм клеток.
Проверка вирусоподобных частиц посредством электронной микроскопии показала, что они идентичны фаговым частицам.
Теперь, после полного описания настоящего изобретения в некоторых деталях посредством иллюстрации и примера с целью четкости понимания, специалисту в данной области будет очевидно, что то же самое можно исполнять посредством модификации или изменения изобретения в широком и эквивалентном диапазоне условий, композиций и других параметров без влияния на объем изобретения или любое его конкретное осуществление и что такие модификации или изменения подразумевают включенными в объем прилагаемой формулы изобретения.
Все публикации, патенты и патентные заявки, указанные в данной спецификации, указывают на уровень, достигнутый специалистами в данной области, к которой принадлежит данное изобретение, и включены сюда в качестве ссылки в такой же степени, как если бы каждую отдельную публикацию, патент или патентную заявку конкретно и отдельно указывали бы как включенную в качестве ссылки.
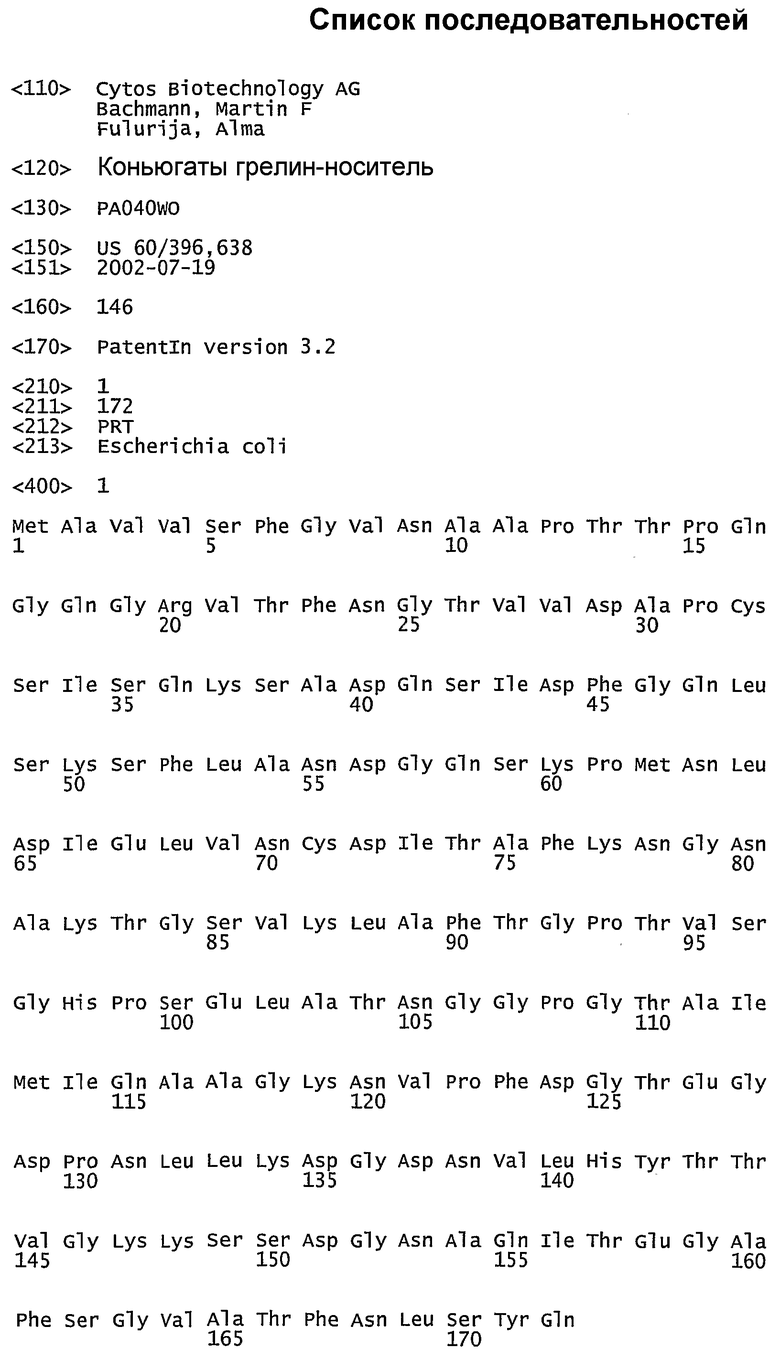


















































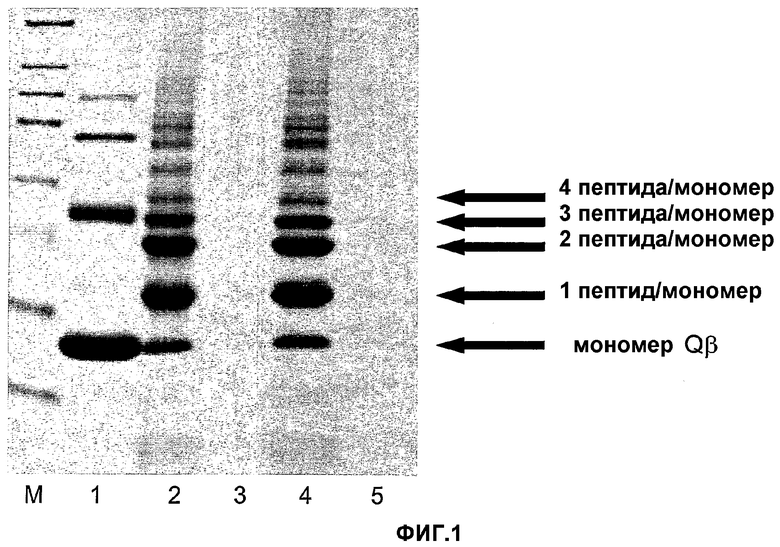
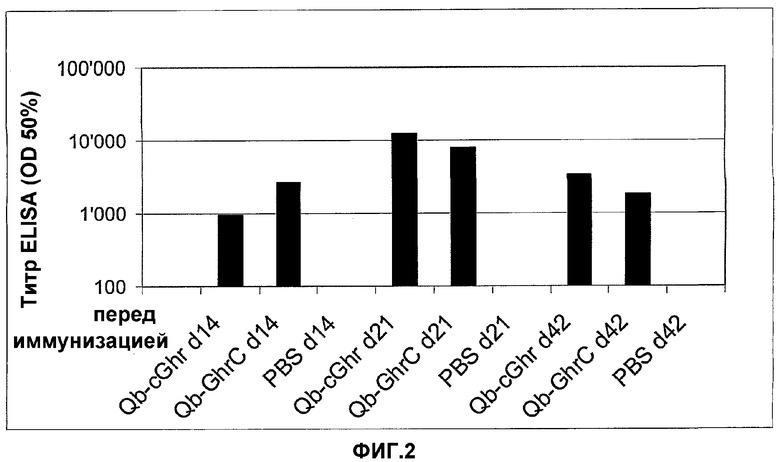
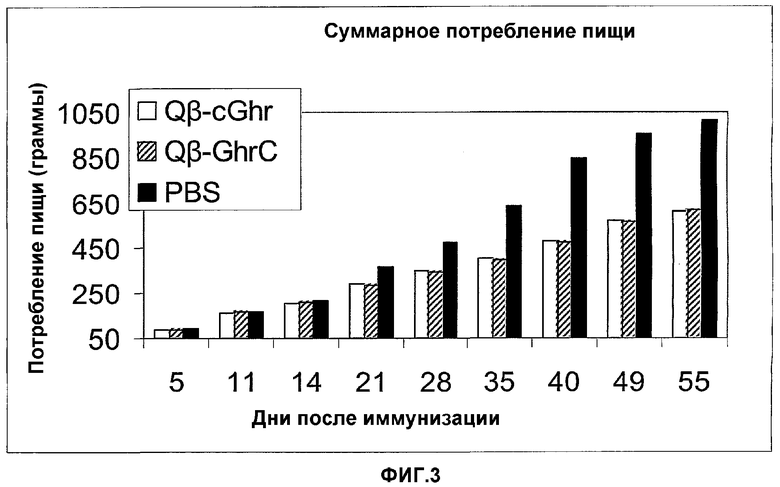
Настоящее изобретение относится к областям молекулярной биологии, вирусологии, иммунологии и медицины. Изобретение относится к композиции, содержащей вирусоподобную частицу и, по меньшей мере, один связанный с ней грелин или полученный из грелина пептид. Изобретение также относится к способу получения конъюгатов и упорядоченных и повторяющихся массивов. Композиции по изобретению пригодны для получения вакцин для лечения ожирения и другого заболевания, ассоциированного с увеличенным потреблением пищи или увеличенной массой тела, и для эффективной индукции иммунных ответов, в частности ответов антителами. Кроме того, композиции по изобретению особенно пригодны для эффективной индукции аутоспецифичных иммунных ответов. 5 н. и 25 з.п. ф-лы, 3 ил.
(a) коровую частицу, по меньшей мере, с одним первым участком прикрепления, где указанная коровая частица представляет собой вирусоподобную частицу, где предпочтительно указанная вирусоподобная частица представляет собой рекомбинантную вирусоподобную частицу и
(b) по меньшей мере, один антиген или антигенную детерминанту, по меньшей мере, с одним вторым участком прикрепления, где указанный антиген или антигенная детерминанта представляет собой грелин или пептид грелина и где указанный второй участок прикрепления выбран из группы, состоящей из:
(i) участка прикрепления, не встречающегося в природе с указанным антигеном или антигенной детерминантой и
(ii) участка прикрепления, встречающегося в природе с указанным антигеном или антигенной детерминантой,
где указанный второй участок прикрепления способен к ассоциации с указанным первым участком прикрепления и где указанный грелин или пептид грелина и указанная коровая частица взаимодействуют посредством указанной ассоциации с образованием упорядоченного и повторяющегося массива антигенов.
(b) рекомбинантных белков вируса кори;
(c) рекомбинантных белков вируса синдбис;
(d) рекомбинантных белков ротавируса;
(e) рекомбинантных белков вируса ящура;
(f) рекомбинантных белков ретровируса;
(g) рекомбинантных белков вируса Норфолка;
(h) рекомбинантных белков альфавируса;
(i) рекомбинантных белков вируса папилломы человека;
(j) рекомбинантных белков вируса полиомы;
(k) рекомбинантных белков бактериофагов;
(l) рекомбинантных белков РНК-фагов;
(m) рекомбинантных белков Ту;
(n) рекомбинантных белков фага Qβ;
(о) рекомбинантных белков фага GA;
(р) рекомбинантных белков фага fr;
(q) рекомбинантных белков фага АР205 и
(r) фрагментов любого из рекомбинантных белков от (а) до (q).
(a) бактериофага Qβ;
(b) бактериофага R17;
(c) бактериофага fr;
(d) бактериофага GA;
(e) бактериофага SP;
(f) бактериофага MS2;
(g) бактериофага M11;
(h) бактериофага МХ1;
(i) бактериофага NL95;
(k) бактериофага f2;
(l) бактериофага РР7; и
(m) бактериофага АР205.
(a) бактериофага Qβ;
(b) бактериофага R17;
(c) бактериофага fr;
(d) бактериофага GA;
(e) бактериофага SP;
(f) бактериофага MS2;
(g) бактериофага M11;
(h) бактериофага МХ1;
(i) бактериофага NL95;
(k) бактериофага f2;
(l) бактериофага РР7; и
(m) бактериофага АР205.
(a) человеческого грелина или пептида человеческого грелина;
(b) коровьего грелина или пептида коровьего грелина;
(c) овечьего грелина или пептида овечьего грелина;
(d) собачьего грелина или пептида собачьего грелина;
(e) кошачьего грелина или пептида кошачьего грелина;
(f) мышиного грелина или пептида мышиного грелина;
(g) свиного грелина или пептида свиного грелина;
(h) лошадиного грелина или пептида лошадиного грелина; и
(k) пептида или его фрагмента любого грелина из (а)-(h).
(a) GSSFLSPEHQRVQRKESKKPPAKLQPR (SEQ ID NO: 48)
(b) GSSFLSPEHQRVQQRKESKKPPAKLQPR (SEQ ID NO: 31)
(c) GSSFLSPEHQKLQQRKESKKPPAKLQPR (SEQ ID NO: 49)
(d) GSSFLSPEHQKLQRKESKKPPAKLQPR (SEQ ID NO: 50)
(e) GSSFLSPEHQKAQQRKESKKPPAKLQPR (SEQ ID NO: 32)
(f) GSSFLSPEHQKAQRKESKKPPAKLQPR (SEQ ID NO: 51)
(q) KKPPAKLQPR (SEQ ID NO: 52)
(h) PPAKLQPR (SEQ ID NO: 53)
(i) AKLQPR (SEQ ID NO: 54)
(j) GSSFLSPEHQ (SEQ ID NO: 55)
(k) EHQRVQQRKE (SEQ ID NO: 56)
(l) KLQPR (SEQ ID NO: 59)
(m) GSSFLSPEHQRVQ (SEQ ID NO: 60)
(n) QRKESKKPPAKLQPR (SEQ ID NO: 61)
(о) GSSFLSPEHQKLQ (SEQ ID NO: 62)
(p) QRKESKKPPAKLQPR (SEQ ID NO: 63)
(q) EHQRVQQRKES (SEQ ID NO: 111)
(r) EHQKAQQRKE (SEQ ID NO: 112)
(s) EHQKAQQRKES (SEQ ID NO: 113)
(t) EHQKLQQRKE (SEQ ID NO: 114)
(u) EHQKLQQRKES (SEQ ID NO: 115)
(v) LSPEHQRVQQ (SEQ ID NO: 116)
(w) LSPEHQKAQQ (SEQ ID NO: 117)
(x) LSPEHQKLQQ (SEQ ID NO: 118) и
(у) GSSFLSP (SEQ ID NO: 119).
(a) GSSFLSPEHQRVQRKESKKPPAKLQPR (SEQ ID NO: 48)
(b) GSSFLSPEHQRVQQRKESKKPPAKLQPR (SEQ ID NO: 31)
(c) GSSFLSPEHQKLQQRKESKKPPAKLQPR (SEQ ID NO: 49)
(d) GSSFLSPEHQKLQRKESKKPPAKLQPR (SEQ ID NO: 50)
(e) GSSFLSPEHQKAQQRKESKKPPAKLQPR (SEQ ID NO: 32)
(f) GSSFLSPEHQKAQRKESKKPPAKLQPR (SEQ ID NO: 51)
(g) KKPPAKLQPR (SEQ ID NO: 52)
(h) PPAKLQPR (SEQ ID NO: 53)
(i) AKLQPR (SEQ ID NO: 54)
(j) GSSFLSPEHQ (SEQ ID NO: 55)
(k) EHQRVQQRKE (SEQ ID NO: 56)
(l) KLQPR (SEQ ID NO: 59)
(m) GSSFLSPEHQRVQ (SEQ ID NO: 60)
(n) QRKESKKPPAKLQPR (SEQ ID NO: 61)
(о) GSSFLSPEHQKLQ (SEQ ID NO: 62)
(p) QRKESKKPPAKLQPR (SEQ ID NO: 63)
(q) EHQRVQQRKES (SEQ ID NO: 111)
(r) EHQKAQQRKE (SEQ ID NO: 112)
(s) EHQKAQQRKES (SEQ ID NO: 113)
(t) EHQKLQQRKE (SEQ ID NO: 114)
(u) EHQKLQQRKES (SEQ ID NO: 115)
(v) LSPEHQRVQQ (SEQ ID NO: 116)
(w) LSPEHQKAQQ (SEQ ID NO: 117)
(x) LSPEHQKLQQ (SEQ ID NO: 118) и
(у) GSSFLSP (SEQ ID NO: 119).
(a) GGC;
(b) GGC-CONH2,
(c) GC;
(d) GC-CONH2,
(e) С; и
(f) C-CONH2.
(a) CGSSFLSPEHQRVQRKESKKPPAKLQPR (SEQ ID NO: 64)
(b) CGSSFLSPEHQRVQQRKESKKPPAKLQPR (SEQ ID NO: 65)
(c) CGSSFLSPEHQKLQQRKESKKPPAKLQPR (SEQ ID NO: 71)
(d) CGSSFLSPEHQKLQRKESKKPPAKLQPR (SEQ ID NO: 72)
(e) CGSSFLSPEHQKAQQRKESKKPPAKLQPR (SEQ ID NO: 77)
(f) CGSSFLSPEHQKAQRKESKKPPAKLQPR (SEQ ID NO: 106)
(g) GSSFLSPEHQRVQRKESKKPPAKLQPRC (SEQ ID NO: 66)
(h) GSSFLSPEHQRVQRKESKKPPAKLQPRGC (SEQ ID NO: 120)
(i) GSSFLSPEHQRVQQRKESKKPPAKLQPRC (SEQ ID N0:67)
(j) GSSFLSPEHQRVQQRKESKKPPAKLQPRGC (SEQ ID NO: 121)
(k) GSSFLSPEHQKLQQRKESKKPPAKLQPRC (SEQ ID NO: 73)
(l) GSSFLSPEHQKLQQRKESKKPPAKLQPRGC (SEQ ID NO: 123)
(m) GSSFLSPEHQKLQRKESKKPPAKLQPRC (SEQ ID NO: 74)
(n) GSSFLSPEHQKLQRKESKKPPAKLQPRGC (SEQ ID NO: 124)
(о) GSSFLSPEHQKAQQRKESKKPPAKLQPRC (SEQ ID NO: 105)
(p) GSSFLSPEHQKAQRKESKKPPAKLQPRC (SEQ ID NO: 107)
(q) CKKPPAKLQPR (SEQ ID NO: 108)
(r) CPPAKLQPR (SEQ ID NO: 70)
(s) CAKLQPR (SEQ ID NO: 109)
(t) GSSFLSPEHQC (SEQ ID NO: 110)
(u) CEHQRVQQRKE (SEQ ID NO: 76)
(v) GSSFLSPEHQRVQC (SEQ ID NO: 68)
(w) GSSFLSPEHQRVQGC (SEQ ID NO: 122)
(x) CQRKESKKPPAKLQPR (SEQ ID NO: 69)
(у) GSSFLSPEHQKLQC (SEQ ID NO: 75)
(z) GSSFLSPEHQKLQGC (SEQ ID NO: 125)
(aa) GSSFLSPEHQKAQRKESKKPPAKLQPRC (SEQ ID NO: 126)
(bb) GSSFLSPEHQKAQRKESKKPPAKLQPRGC (SEQ ID NO: 127)
(cc) GSSFLSPEHQKAQQRKESKKPPAKLQPRC (SEQ ID NO: 128)
(dd) GSSFLSPEHQKAQQRKESKKPPAKLQPRGC (SEQ ID NO: 129)
(ее) GSSFLSPEHQKAQC (SEQ ID NO: 130)
(ff) GSSFLSPEHQKAQGC (SEQ ID NO: 131)
(gg) GGSSFLSPEHQGC (SEQ ID NO: 132)
(hh)CKKPPAKLQPR(SEQ ID NO: 133)
(ii) CEHQKAQQRKE (SEQ ID NO: 134)
(jj) CEHQKAQQRKES (SEQ ID NO: 135)
(kk) CLSPEHQKAQQ (SEQ ID NO: 136)
(ll) CEHQRVQQRKES (SEQ ID NO: 137) и
(mm) CLSPEHQRVQQ (SEQ ID NO: 138).
(a) композицию по п.1 и
(b) приемлемый фармацевтический носитель,
и где предпочтительно указанная фармацевтическая композиция дополнительно содержит адъювант или где предпочтительно указанная фармацевтическая композиция не содержит адъювант.
| 0 |
|
SU185208A1 | |
| EP 1197496, 17.04.2002. | |||

