Настоящее изобретение относится к области сельского хозяйства и огородничества, в особенности к защите растений от вирусов, бактерий, грибов или других паразитов. В частности, изобретение описывает бактериальные белки, вызывающие общую устойчивость растений к микробным патогенам и вредителям.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Патогенные микроорганизмы и вредители ответственны за существенные экономические потери в сельскохозяйственном производстве во всем мире. Современные методы контроля фитопатогенов и вредителей имеют каждый свои серьезные недостатки. В принципе, селекция новых сельскохозяйственных сортов, которые генетически более устойчивы к патогенам, может предотвратить потери урожая. На практике, однако, каждый новый сорт в конечном счете теряют свои преимущества, так как патогенны постепенно преодолевают эту устойчивость. Применение синтетических неприродных химических средств защиты представляет значительный риск для экологии. Только сравнительно недавно начали использовать похожие вещества природного происхождения. Однако такие природные вещества дороги, их распыление требует применения специальных устройств и является трудоемким процессом.
В течение последних двадцати лет для придания растениям устойчивости к вирусным патогенам были разработаны новые подходы, использующие трансгенные растения с чужеродными генами. Как правило, в таких растениях экспрессировали определенные вирусные гены (например, ген белка оболочки). К сожалению, приобретенная устойчивость эффективна только против определенного вирусного штамма, которым растение было «вакцинировано». Например, устойчивость к Y-вирусу картофеля была не эффективна против вирусных штаммов, которые отличались от использованного штамма всего на 22% на нуклеотидном уровне. Поэтому этот вид устойчивости имел ограниченное практическое применение, так как при изменении климатических условий и других факторов доминируют различные патогены. Фермеры ожидают, что их инвестиции в дорогостоящий семенной материал должны быть выгодными каждый год, а не только в определенные годы. Тем не менее, из-за отсутствия более универсальных решений такая ограниченная устойчивость была получена к различным вирусам в широком кругу сельскохозяйственных культур. Также ситуация должна учитывать растущую значимость общественного мнения и опасения относительно использования трансгенных растений: выгоды должны быть достаточно значимыми, чтобы перевесить обеспокоенность общества.
Системная приобретенная устойчивость (SAR) впервые описана Честер (1933). SAR - обычная защитная реакция, когда растение системно производит различные защитные молекулы, такие как лигнин, фитоалексины и PR-белки, чтобы предотвратить распространение патогенов (см. Обзор Sticher et al. 1997, Ann.Rev.Phytopathol. 35, 235-270). SAR может быть вызвана в течение нескольких часов многими патогенами, а затем устойчивость длится в течение нескольких недель. SAR - реакция устойчивости, зависящая от салициловой кислоты, но первичная роль салициловой кислоты до сих пор не ясна (Ryal et al. 1996, Plant Cell 8, pp.1809-1819). Распространение патогена ограничено небольшим количеством клеток растения, в которых индуцируется SAR; поэтому болезнетворные микроорганизмы, практически, не могут повредить растение.
Тогда как существует огромная потребность в быстром получении новых сортов растений с высокой устойчивостью к различным болезням и вредителям и с высокой экологической безопасностью, до сих пор не было предложено ни одного ясного решения этой проблемы.
Настоящее изобретение предлагает совершенно новую концепцию увеличения устойчивости растения, основанную на удивительной находке, что определенный бактериальный белок, названный как MF3, может вызывать в растениях системную устойчивость широкого спектра, включающую устойчивость к вирусам, бактериям, грибам и таким вредителям, как Колорадский картофельный жук и нематоды. Широкий защитный потенциал белка MF3 показан на экспериментальных результатах, подтверждающих формулу изобретения, что белок может вызывать полный спектр устойчивости растения к патогенам и вредителям. Хотя действие открытого нами белка в растениях напоминают эффекты SAR, никакой механистической связи между ними не вырисовывается.
Ранее мы обнаружили другой белок с совершенно другой структурой, чем MF3; этот белок выделен из штамма Bacillus thuringiensis (MF2, Djavakhia V., et al. US 6,528,480). Трансгенные растения табака, экспрессирующие MF2, обладали усиленной устойчивостью к вирусам и грибам (вирус мозаики табака и Alternaria longipes соответственно). В настоящем изобретении мы предлагаем полностью различный бактериальный белок с улучшенной активностью и значительно более широкими приложениями. Поэтому настоящее изобретение обеспечивает существенное усовершенствование нашего предыдущего изобретения, показывая, что новая молекула может вызывать множественную устойчивость растений к фитопатогенам, включая микроорганизмы, а также насекомых и нематод, в особенности, которые известны как опасные паразиты растений. Мы также показываем, что MF3 может быть использован как индуктор устойчивости в различных трансгенных растениях без любой потери урожайности.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ.
Чертеж. Номограмма для определения потерь урожая картофеля от фитофтороза в результате преждевременного отмирания листьев.
ДЕТАЛЬНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Ключевой аспект активной защиты организма против вторжения микробов или других патогенов - способность различать свои компоненты от чужих. В растениях устойчивость, базирующаяся на узнавании патогенов, наиболее полно и успешно изучена в случае наличия специфических генов устойчивости, которые придают растениям устойчивость к специфическим расам патогенов. Было показано, что некоторые из этих генов устойчивости участвуют в хемовосприятии факторов, специфически присущих конкретным штаммам патогенов, кроме того, у растений есть система более широкого контроля, базирующая на чувствительном восприятии факторов, характерных для целых групп или классов микроорганизмов, и растения отвечают на эти общие элиситоры активацией сигнальных путей для того, чтобы запустить защитные механизмы.
Возможные примеры элиситоров белковой природы описаны в настоящем изобретении. Известны примеры белков, активирующие устойчивость. Однако эти белки были выделены из зараженных тканей растений, и они специфически действуют только в определенной паре патоген-хозяин, что является существенным отличием от настоящего изобретения. Кроме того, в отличие от известных белков и механизмов устойчивости в настоящем изобретении мы представляем бактериальный белок, названный MF3, с новой энзиматической активностью; этот белок не участвует ни в одном известном процессе фитопатогенеза. Обработка растительных тканей раствором этого белка приводит к активации механизма устойчивости.
Очевидна полезность MF3 при создании трансгенных растений с низким уровнем экспрессии этого белка. Главное преимущество использования - это производство MF3 в больших количествах и его введение в растительные клетки в подобранных составах с помощью соответствующих методов.
Методы получения трансгенных растений хорошо известны по учебникам, а первые трансгенные растения известны еще с 80-х годов прошлого столетия. Генетическая инженерия помогла исследователям понять глубины фундаментальных процессов, которые управляют развитием растений, а первые такие генетически модифицированные растения уже коммерциализованы и сейчас широко используются.
Одно из самых многообещающих направлений, которые открывает техника переноса генов, - устойчивость к болезням. Захватывающие результаты были достигнуты в создании растений, устойчивых к вирусам; это важный вопрос, потому что в настоящее время не существует прямых методов обработки растений, зараженных вирусом. Эксперименты с генетически модифицированными растениями показали, что экспрессия вирусных генов, как правило, придает устойчивость только к тем же самым вирусным штаммам (см. обзор Baulcombe, 1994). В каждом из этих случаев проблема ограниченной устойчивости вызывает беспокойство при практическом использовании этих растений. Проблема может быть преодолена путем использования генов факторов, вызывающих множественную устойчивость растений, факторов, описанных в настоящем изобретении.
Мы выделили белок MF2 из штамма Bacillus thuringiensis, который индуцировал устойчивость растений к вирусным и грибковым инфекциям (Djavakhia V., и др., патент США 6528480). В настоящем изобретении мы с удивлением обнаружили, что не только Bacillus thuringiensis может содержать такой индуктор. Используя определенные новые методы анализа к очень большому количеству почвенных микробов, мы сумели выделить из штамма Pseudomonas fluorescence новый индуктор MF3. Оказалось, что этот индуктор при чрезвычайно низкой концентрации вызывает общую устойчивость у различных растений не только к вирусной, но также и к бактериальной, и к грибковой инфекциям.
Позже было показано, что MF3 вызывает устойчивость даже к нематоде и насекомым как при прямом введении MF3 в растения через кутикулу листа, так и в эндогенном виде при его синтезе в трансгенных растениях. Структуры белков MF2 и MF3 не имеют никаких очевидных гомологичных последовательностей. Единственная общая черта - оба белка термостабильны. Их молекулярные массы также отличаются: MF2 имеет молекулярный вес 7239 Дальтон; молекулярная масса MF3 - порядка 17600 Дальтон. MF3 - это новая белковая последовательность, как оказалось, наиболее близко напоминает фермент пептидил-пролил-цис-транс-изомеразу SlyD из Pseudomonas aeruginosa.
Пептидил-пролил-цис-транс-изомеразы играют ключевую роль в фосфорилировании белков животных (см. обзор Zavyalov, V. et al. APMIS Vol.103, pp.401-415, 1995). Поскольку могут существовать общие молекулярные связи или корреляции между механизмами устойчивости у животных и в растениях, возможно, что MF3 является ключевым фактором механизма устойчивости растений и имеет, таким образом, общую ценность как молекулярный инструмент. Следовательно, MF3 может быть использован в решении различных проблем фитопатологии.
Препараты MF3 могут эффективно использоваться в технологии культуры меристемы для получения чистого вещества, без микробов ростков, опрыскивая культуру меристемы надлежащим соединением MF3, или при использовании MF3, или его активных пептидов, возможно в комбинации с термообработкой клеточных культур. Использование этого экологически чистого, нефитотоксичного вещества могло бы быть эффективным для клонального размножения безмикробных побегов различных сельскохозяйственных растений. Знание структуры MF3 обеспечивает возможность для получения генетических конструкций для того, чтобы создать трансгенные растения, устойчивые к вирусным и другим болезням.
Очевидны преимущества трансгенных растений, содержащих MF3. Известно, что трансгенные растения, содержащие часть вирусного генома, как правило, специфически устойчивы только к близкородственным вирусам. Микроорганизм, используемый согласно настоящему изобретению, не имеет никакой очевидной структурной связи. Мы показываем здесь, что MF3 индуцирует неспецифическую устойчивость растений табака к вирусу табачной мозаики (TMV), к вирусу Х картофеля (PVX) и к вирусу Y картофеля (PVY). Более того, обработка растений картофеля MF3 приводит к индуцированию устойчивости к фитофторозу, вызываемому Phytophthora infestans. Было также показано, что обработка клубней картофеля MF3 индуцирует устойчивость к бактериальной мягкой мокрой гнили (Erwinia carotovora). Кроме того, MF3 индуцирует устойчивость растений риса к пирикуляриозу риса - болезни, вызываемой Pyricularia oryzae. MF3 также индуцирует устойчивость растений пшеницы к Fusarium culmorwn and Septoria nodorum. Как показано в специальных примерах, индуцируется также устойчивость к нематодам и насекомым. С высокой степенью вероятности устойчивость проявляется по отношению к насекомым на разных возрастных стадиях, таких как яйца, личинки и взрослые особи. Эти различные возрастные формы атакуют разные части растения (н-р, корни, листья). Аналогично, как показано в специальных примерах, у широкого круга растений формируется устойчивость после in vitro или in vivo введения MF3 в их клетки или в межклеточное пространство. Следует специально отметить, что MF3 может также иметь специфические рецепторы внутри растительной ткани. Поэтому настоящее изобретение нужно рассматривать как создание универсальной защиты растений от различных растительных паразитов и болезней.
Последовательность ДНК, кодирующая белок MF3, может быть клонирована в любой вектор для клонирования и/или экспрессии в любом организме от бактерий до высших эукариот, включая растения, с помощью обычно используемых методов генетической инженерии, как описано, например, в J.Sambrook, E.F.Fritsch and T.Maniatis, Molecular Cloning: A Laboratory Manual, 2nd ed. (Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press, 1989). Мы клонировали ген, кодирующий белок MF3, и определили его нуклеотидную последовательность. Также мы определили аминокислотную последовательность белка MF3. На основании этих результатов был сконструирован штамм Escherichia coli - супер-продуцент белка MF3.
Следующая цель настоящего изобретения - метод выделения и очистки белков, обладающих антивирусной, антибактериальной, фунгицидной, общей антимикробной, антинематодной и инсектицидной активностями, из предпочтительно генетически модифицированного микроба, экспрессирующего вышеназванный(ые) белок(ки). Такой метод состоит из:
а) культивирование микробного штамма-продуцента и экстракция из клеток в соответствующем буферном растворе при повышенной температуре, предпочтительно на кипящей водяной бане, для того чтобы исключить основную часть температурочувствительных веществ из экстракционной среды;
в) осаждение неочищенного MF3 полипептида при низкой температуре с соответствующим осадителем, чтобы исключить органические соединения низкой молекулярной массы из белковой фракции;
г) фракционирование растворенного осадка согласно п.в) с помощью анион-обменной колоночной хроматографии и сбор фракций с антимикробной, антинематодной и/или инсектицидной активностями:
д) проведение электрофоретического анализа в ПААГ белковых фракций с антивирусной, антибактериальной, фунгицидной, антинематодной и/или инсектицидной активностями согласно п.в);
е) рефолдинг белка, элюированного из геля.
Принимая во внимание, что вышеописанный предпочтительный метод очистки антимикробного белка может быть модифицирован, существенные особенности метода должны включать термообработку при температуре между 60-110°С. Также одинаково существенны методы измерения вышеназванных биологических активностей в различных фракциях. Различные применения антимикробных белков не требуют чистого белка в виде, как описано в предпочтительном методе очистке. На стадии термообработки микробные клетки такие, как Pseudomonas fluorescence, экстрагируют предпочтительно с калийфосфатным буфером рН7,4, содержащим EDTA, PMSF (phenyl methyl sulphonyl fluoride), b-меркаптоэтанол и Triton Х-100 (polyoxyethylene ether). Осаждение предпочтительно выполнять при температуре между 2°С и 6°С с охлажденным во льду хлороформом, и/или пропанолом и/или раствором сульфата аммония.
Следующий объект изобретения - использование MF3 в качестве защитного средства растения от различных микробов. MF3 предпочтительно используется в составе смесей веществ, включающих стабилизаторы, носители и/или адъюванты. Поскольку MF3 относительно устойчив, такие добавки главным образом предназначены для помощи активным ингредиентам переносить активный белок или его активные фрагменты в клетки растения и/или делать их (белки) доступными для рецепторов. Техника составления составов для различных растительных протектантов хорошо известна и описана (N.M.Golishin, 1982).
Мы показали защитные свойства MF3 на растениях табака против вируса табачной мозаики (VTM), вируса Х картофеля (PVX) и вируса Y картофеля (PVY). Мы также показали защитную (протекторную) активность на растениях картофеля против Phytophthora infestans и Erwinia carotovora, на растениях риса против Pyricularia oiyzae и на растениях пшеницы против Fusarium culmorum и Septoria nodorum, a также против типа Nematoda. В тестах использовали штамм 197 Pseudomonas fluorescence из Всероссийской Микробиологической Коллекции. Это один из бактериальных штаммов, выделенных на корневых волосках растений пшеницы в поле одной из ферм в Одинцовском районе в Московской области.
Хотя MF3 выделен из определенного микробного штамма, следует полагать, что родственные активные белки могут производиться также другими организмами, и эти белки также покрываются (включаются) в соответствии с настоящим изобретением. Принимая во внимание, что последовательность MF3 напоминает структуру известного фермента, ферментативная активность не обязательно требуется для заявленной биологической активности MF3. Напротив, потому что MF3 может быть подвергнут специфическому протеолизу или кипячению с сохранением заявленной активности, более вероятно, что другие биологические свойства MF3 обеспечивают возникновение защиты растения.
Изобретение далее иллюстрировано, но не ограничено, следующими примерами определенных воплощений изобретения.
ПРИМЕР 1. Культуральные и морфологические признаки Pseudomonas fluorescence штамм 197
Изолят культивируется на синтетической среде, содержащей 20 г/л пептона, 2,5 г/л K2HPO4, 6 г/л MgSO4, 20 г/л глюкозы. При инкубации в течение 18 часов при 28°С на агаризованной культуральной среде бактерии образуют маленькие (1,5-2 мм) сферические колонии с гладкими краями и матовой поверхностью. При инкубации в течение 18 часов при 28°С в жидкой среде при качании бактерии достигают плотности 1010 клеток/мл. Бактерии производят зеленовато-желтый прозрачный и способный флюоресцировать пигмент. Клетки - грамотрицательные, представляют собой короткие, маленькие малоподвижные палочки со жгутиками. Оптимальная температура роста для штамма 197 - это 28°С, минимальная - +4°С, а максимальная - +43°С. Оптимум рН - около 7,0. Бактерии не фиксируют атмосферный азот и не используют соединения с одним атомом углерода в качестве источника углерода. Источником углерода для бактерий служат сахароза, глюкоза, глицерин и/или другие соединения, содержащие более одного атома углерода. Бактерия является хемо-органотрофом, аэробом, содержит оксидазы и каталазы. Метаболизм - дыхательный, но не ферментативный. Результаты морфологического, культурального, физиологического и биохимического анализов позволили нам заключить, что выделенный штамм принадлежит к виду Pseudomonas fluorescence (Cion, 1948; The shorter Bergey's manual of determinative bacteriology, 1980).
После культивирования при оптимальных условиях (см. выше) бактериальные клетки собирали центрифугированием при 6000 об/мин в течение 15 минут при комнатной температуре (Sorvall-RC28S centrifuge, rotor GS-3) и дважды промывали дистиллированной водой. Клетки ресуспендировали в 50 мМ калийфосфатном буфере, рН 7,0, содержащем 1 мМ EDTA, чтобы конечная концентрация была 4-5×1010 клеток/мл. Суспензию прогревали на бане с кипящей водой в течение 20 мин и затем центрифугировали при 6000g в течение 15 минут, чтобы убрать бактериальные клетки и клеточный дебрис. Супернатант последовательно обрабатывали 1 объемом (вес/объем) холодного хлороформа и 2 объемами (вес/объем) холодного пропанола. Осадок удаляли центрифугированием в течение 15 минут при 6000g. Супернатант смешивали с холодным пропанолом так, чтобы конечная концентрация пропанола была 5 объемов. Осадок собирали центрифугированием при 10000g в течение 20 минут, а затем растворяли в 0,1М буфере Tris-HCl (рН 7,0), содержащем 0,1М NaCl, 1 мМ EDTA и 1% Triton X-100. Получающийся материал после этого прогревали в течение 2 минут в бане с кипящей водой и после охлаждения наносили на колонку с Сефадексом G-50 (1,2×90 см), уравновешенным тем же буфером. Активные фракции объединяли, упаривали и затем для осаждали, добавляя 5 объемов холодного этанола. На следующей стадии очистки проводили вертикальный гель-электрофорез в полиакриламидном геле, описанный Лэммли (1970), за исключением того, что разделяющий гель и буфер содержали Triton Х-100 вместо додецил-сульфата натрия. Использовали 20% полиакриламидный гель, содержащий 0,1% Triton X-100. Вымывание активных полос было сделано возобновлением гель-электрофореза. Для визуализации белковых полос гель окрашивался раствором Coomassie Brilliant Blue R-250. Количество белка определялось по методу Брэдфордта (1976) с бычим сывороточным альбумином в качестве стандарта.
Следующие примеры служат иллюстрацией определенных аспектов настоящей заявки. Очищенный кипяченный экстракт штамма Р. fluorescence 197 использовался в этих экспериментах. Следует отметить, что препарат может быть также получен из генетически модифицированных организмов-продуцентов таких, например, как Е. coli, описанный далее.
ПРИМЕР 2. Защитные свойства белка MF3 против вируса табачной мозаики (TMV) на растениях табака
Все штаммы и микробов, и нематод, а также сорта растений были из коллекции культур Всероссийского научно-исследовательского института фитопатологии, Голицино, Московская область, Российская Федерация. Полученный экспериментальный материал подвергался математической обработке методом статистического анализа при 95% уровне достоверности.
Растения табака (Nicotiana tabacum var. Virginia и Nicotiana glutinosa) выращивали до стадии шести листьев (около 3-х недель) в горшках с почвой в камере искусственного климата с относительной влажностью (RH) 60% и температурой 24°С при 12-часовой длине дня и ночи. Листья растений табака инокулировали кисточкой для рисования с карборундом в качестве абразива. Каждая половинка листа 3-недельных растений табака протиралась карборундом с 50 мкл экстракта Р.fluorescence. В качестве контроля брались листья растений, обработанные с таким же объемом буфера. Через два дня эти же листья были протерты с суспензией вируса табачной мозаики (0,3 мкг/мл в 10 мМ калийфосфатном буфере (рН 7,0); брали 0,3 мл/половина листа). Количество инфекционных повреждений (некрозов) на каждой половинке листа подсчитывали через 3 дня. Развитие болени измерялось как отношение числа повреждений в опыте к контролю. Результаты эксперимента показаны в Таблице 1. Очищенный экстракт из Р.fluorescence показал на растениях табака защитный эффект против TMV инфекции.
ПРИМЕР 3. Системный характер активности MF3 против вируса табачной мозаики на растениях табака
Растения табака (Nicotiana tabacum var. Virginia и Nicotiana glutinosa) выращивали до стадии шести листьев (около 3-х недель) в горшках с почвой в камере искусственного климата с относительной влажностью (RH) 60% и температурой 24°С при 12-часовой длине дня и ночи. Два нижних листа на растении были протерты с карборундом кисточкой для рисования с 50 мкл раствора MF3. Два дня спустя те же и верхние листья были инокулированы суспензией вируса табачной мозаики. В контроле листья растений обрабатывали буфером. Результаты эксперимента представлены в Таблице 2.
ПРИМЕР 4. Активность MF3 против Pyricularia oryzae на растениях риса
Был использован природный изолят Н-5-3 of Pyricularia oryzae Cav. Грибок культивировался при 28°С на агаризованной минимальной среде, содержащей 3 мг/мл гидролизата казеина (Sigma). Споры (конидии) 10-дневной культуры промывались с дистиллированной водой при 4°С. Примесь мицелия удаляли фильтрованием через нетканный фильтр (Miracloth, Calbiochem-Boehring Corp.) и через два слоя стальных сит (размер пор 50 мкм). Суспензия спор промывали двойным центрифугированием в течение 15 минут при 7000g и ресуспендировали в дистиллированной воде. Концентрацию спор определяли подсчетом в гемоцитометре под микроскопом.
Для анализа использовали растения риса (Oryza saliva L. of cv. Sha-tiao-tsao), чувствительные к вынеупомянотому штамму грибка. Растения выращивали до стадии четырех листьев (около 13-15-ти дней) в горшках с почвой в камере искусственного климата с относительной влажностью (RH) 95% и температурой 30°С днем и 23°С ночью при 12-часовой длине дня и ночи. Для освещения (20 клюкс) использовали 10 кВт ксеноновые лампы (DKsT-10000) с водяным фильтром. Растения риса опрыскивали суспензией спор (100000 спор/мл, 5 мл/горшок). Обработанные растения инкубировали в течение 18-24 часов во влажной камере в темноте при 23°С, а затем переносили на свет в камеру искусственного климата для наблюдения за развитием симптомов заболевания в течение 10 дней. Чтобы оценить жизнеспособность инокулята, каплю суспензии спор инкубировали в течение 15 часов в иммунологической плашке в темноте при 23°С. Затем подсчитывали проростание спор. Раствор препаратов MF3 в буфере добавляли в инокулят со спорами. Контрольные образцы содержали равный объем буфера.
Все препараты MF3 в использованных концентрациях не подавляли прорастание в воде спор Р.oiyzae. В то же время добавление препаратов в инокуляты защищало растения риса от болезни в значительной степени.
Развитие болезни вычисляли по формуле:
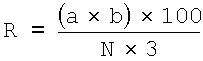
R - развитие болезни (%);
а - количество инфицированных
растений;
b - тип инфекции;
N - общее количество растений;
3 - высший тип инфекции.
ПРИМЕР 5. Активность MF3 против Septoria nodorum на растениях пшеницы
Использовали природный изолят Septoria nodorum. Растения пшеницы (Мироновская 808) выращивали до стадии двух листов (около 13-15-дней) в горшках с почвой в камере искусственного климата.
Срезанные листья пшеницы помещали в чашки Петри, содержащие 2% агар и 40 мг/л бензимидазола. На верхнюю часть каждого листа капали 5 мкл MF3 (в концентрации 1,25 мг/мл и 2,5 мг/мл), а на нижнюю часть каждого листа - 5 мкл буфера (для контроля). Через два дня все капли с листьев удаляли с помощью стерильной марли и наносили по 5 мкл суспензии спор (106 /мл). Чашки Петри помещали в темную комнату на один день, а затем переносили на свет (8-10 klux) под 10 кВт ксеноновую лампу (DKsT-10000) при температуре 20-22°С и длине светового дня 16 часов. Симптомы заболевания анализировали через 7 дней.
Как видно по результатам в Таблице 4, MF3 защищает листья растений пшеницы от поражения Septoria nodorum в значительной степени.
ПРИМЕР 6. Заражение зерновки пшеницы бактериями Pseudomonas fluorescence штамм 197 вместе с Fusarium culmorum (W.G.Sm) Sacc. и стимуляция роста растений
Инокулят патогена был получен из 10-дневной культуры двух изолятов F.culmorum, которые выращивали на картофельно-декстрозном агаре. Концентрация конидий в суспензии была примерно 2×106 /мл. Тест проводили на растениях пшеницы Мироновская 808. Обработка зерновки пшеницы суспензией клеток Pseudomonas fluorescence штамм 197 или очищенным экстрактом из бактериального штамма защищала семена пшеницы от F. culmorum, вызывающей корневые гнили пшеницы, при смешенном заражении на рулонах бумаги для прорастания в лабораторных экспериментах (см. детали ниже).
Бактерии культивировали на следующей среде:
2 г/л гидролизата казеина, 10 г/л сахарозы, 3 г/л дрожжевого экстракта, 2,5 г/л NaNO3,
0,5 г/л MgSO4*7H2O, 1 г/л КН3PO4, 20 г/л агар в дистиллированной воде.
Бактерии использовали на 6-й день культивирования. Концентрация клеток в использованной суспензии была 107-108 кл/мл.
Зерновку озимой пшениы стерилизовали в 96% этаноле 1 минуту, затем промывали в стерильной воде, помещали во влажные чашки Петри и инкубировали 24 часа при 23-24°С. Затем проросшие зерна переносили в другие чашки Петри (по 30 зерен на чашку) и смачивали их суспензией конидий F. culmorum и/или клетками штамма 197. Общий объем суспензии - 10 мл/чашка (5 мл суспензии F.culmorum и 5 мл суспензии клеток штамма 197 при смешенном заражении или 5 мл одной из суспензий и 5 мл стерильной воды при раздельном заражении).
Через 24 часа зерна помещали между двумя листами (16×8 5 см и 6×85 см) сухой бумаги для прорастания (30 зерен на пару листов), которые затем покрывали стерильной полиэтиленовой пленкой (6×90 см) и скручивали в рулон. По три рулона/реплика было использовано в каждой обработке. Рулоны помещали в отдельные сосуды, смачивали водой и инкубировали в темноте при 22-23°С в течение 6 дней. Затем рулоны переносили на лабораторный стол и инкубировали при 19-20°С и 12-часовом дне в течение 6 дней. Рулоны смачивали водой по мере их высыхания. Затем определяли количество проросших зерен, измеряли длину побегов и самого длинного из корней и их сухой вес.
Анализ данных в Таблице 5 свидетельствует о том, что штамм 197 существенно уменьшает развитие болезни. Длина побегов была на 22-23%, максимальная длина корней на 18-21%, сухой вес побегов на 22-23%, а сухой вес корней на 19-26% больше при смешанном заражении, чем при заражении только F.culmorum. He было существенной разницы между обработками бактериями в концентрациях 107-108 кл/мл при смешанном заражении.
При оценке влияния клеточного экстракта Pseudomonas fluorescence штамм 197 бактерии культивировали в жидкой среде, содержащей 20 г/л пептона, 2,5 г/л KH2PO4, 6 г/л MgSO4×7H2O, 20 г/л сахарозы, и использовали на второй день культивирования. Концентрация суспензии была около 1010 клеток/мл.
MF3 готовили следующим образом: бактериальные клетки промывали дважды водой и суспендировали в 10 мМ натрийфосфатном буфере (рН 7,5), чтобы концентрация клеток была более чем в 10 раз выше, чем в культуральной среде. Затем суспензию кипятили в течение 30 минут на бане с кипящей водой. Клеточный дебрис убирали центрифугированием, а супернатант использовали в эксперименте, предварительно разведя его в 15 раз.
Нестерильные зерна пшеницы помещали в мелкую посуду (90 зерен на сосуд) и заливали 5 мл клеточной суспензии, или клеточного экстракта Pseudomonas fluorescence, или стерильной воды, и инкубировали при 20°С в течение 5 часов. Затем добавляли в сосуды 5 мл суспензии спор F.culmorum (2×106 спор/мл) или 5 мл стерильной воды в зависимости от эксперимента. После 24-часовой инкубации в сосудах зерна помещали между двумя (16×85 см и 6×85 см) сухой бумаги для прорастания (30 зерен на пару листов), которые затем покрывали стерильной полиэтиленовой пленкой (6×90 см) и скручивали в рулон. По три рулона/реплика было использовано в каждой обработке. Рулоны помещали в отдельные сосуды, смачивали водой и инкубировали в темноте при 22-23°С в течение 6 дней. Затем рулоны переносили на лабораторный стол и инкубировали при 19-20°С и 12-часовом дне в течение 6 дней. Рулоны смачивали водой по мере их высыхания. Количество проросших зерен, длина побегов и самого длинного из корней и их сухой вес представлены в Таблице 6.
Длина побегов была на 17.3-18.6%, максимальная длина корней на 6.6-17.4%, сухой вес побегов на 23.1-23.6%, а сухой вес корней на 12.3-24.3% больше при смешанном заражении, чем при заражении только F.culmorum. Длина побегов и вес проростков, обработанных клеточным экстрактом Pseudomonas fluorescence (str.197), были на 14.7% и 22.0%, соответственно, больше, чем в контроле.
ПРИМЕР 7. Защитная активность MF3 против фитофтороза
В тесте использовали природный изолят Phytophtora infestans и растения картофеля сорт «Лорх». Чтобы исследовать влияние обработки препаратами штамма 197 на инфекцию Ph.infestans, клубни картофеля смачивали бактериальной суспензией (107 кл/мл). Через 7-10 дней эти клубни высаживали в теплицу. Листья с выросших растений картофеля срезали и заражали вирулентной расой Ph.infestans, чтобы определить степень проникновения, скорость развития симптомов, интенсивность споруляции и скорость колонизации клубней. Листья, срезанные с растений, полученных из необработанных перед посадкой клубней, использовали как контроли.
Интенсивность проникновения измеряли, заражая нижнюю сторону срезанных листьев картофеля путем опрыскивания суспензией спор Ph.iInfestans (103 спор/мл) с последующей инкубацией в течение 2-3-дней при 18-20°С во влажной камере. Далее определяли количество повреждений на см2 поверхности листа. Результаты типичного эксперимента представлены в Таблице 7.
Для определения скорости развития повреждений нижнюю сторону срезанных листьев картофеля заражали каплей суспензии спор Ph.infestans (около 103 спор/мл). Скорость развития симптомов определяли, измеряя диаметр повреждений на 4-й или 5-й день после заражения. Результаты анализа представлены в Таблице 8.
Интенсивность споруляции определяли, вычисляя количество спор, образуемых при единичном повреждении. Заражение проводили аналогично описанному выше тесту. На 5-й день споры смывали с листа с определенным количеством повреждений и определяли количество спор на одно повреждение листа. Результаты эксперимента представлены в Таблице 9.
Эффект обработки клубней картофеля MF3 оценивали, определяя скорость колонизации клубней. Клубни картофеля смачивали бактериальной суспензией штамма 197 (107 кл/мл). Через 7-10 дней из клубней вырезали брусочки размером 0,5×0,5×5 см. Один из этих фрагментов заражали спорами Ph.infestans, после чего фрагменты клубней помещали во влажную камеру и инкубировали при 18-20°С. Длину колонизированного патогенном участка каждого брусочка клубня измеряли на 8-9-й день инкубации. Результаты этих экспериментов представлены в Таблице 10.
Определение развития болезни на клубнях картофеля, обработанных MF3, выполняли следующим образом: клубни картофеля смачивали препаратами бактерий штамма 197 (107 кл/мл). Через 7-10 дней эти клубни высаживали по стандартной технологии на делянки размером 2,8×9 м. Эти делянки находились на большом картофельном поле с естественным инфекционным фоном. Расстояние между экспериментальными делянками составляло более 1 м. Клубни, собранные с экспериментальных делянок и ранее обработанные MF3, были поражены Ph.infestans только на 15%, в то время как контрольные клубни были поражены на 27%.
В других экспериментах необработанные клубни картофеля высаживали на делянках размером 2,8×7 м. Эти растения опрыскивали бактериальной суспензией штамма 197 (104 кл/мл) в количестве 400 л/га четыре раза с различным интервалом между обработками в течение вегетативного периода. В этих экспериментах использовали чувствительный к заражению сорт «Лорх». В течение вегетационного периода анализ развития болезни контролировали с 10-дневным интервалом. После окончания вегетационного периода каждую делянку делили на 5-6 частей. Степень повреждений клубней картофеля определяли на каждом из этих участков. Растения на контрольных делянках не обрабатывали бактериальной суспензией. Клубни, собранные на обработанных штаммом 197 делянках, были повреждены Ph.infestans только на 11%, тогда как клубни с контрольных делянок были повреждены на 27%.
ПРИМЕР 8. Влияние MF3 на личинок Колорадского жука
Личинки Колорадского жука выращивали из яиц (собранных на картофельном поле ВНИИФ) на листьях картофеля во влажной стеклянной камере. В экспериментах использовали листья картофеля сорт Сантэ, собранные с растений в возрасте 40-50 дней, растущих в поле.
Эксперимент проводили с личинками 2-й стадии. Высечки из листьев (16 мм диаметром) смачивали в растворе MF3 или в воде (контроль). После высушивания высечки переносили в чашки Петри на влажную бумагу и помещали на них по 3 личинки. Через 24 часа высечки высушивали и определяли размер области, съеденной личинками. Каждый эксперимент проводили в трех повторностях (Таблица 11).
В экспериментах с личинками 4-й стадии увеличили размер высечек листа до 36 мм в диаметре и на каждую высечку высаживали по одной личинке. Результаты эксперимента представлены в Таблице 12.
ПРИМЕР 9. Устойчивость трансгенных растений картофеля к Erwinia carotovora
Способность Erwinia carotovora мацерировать растительные ткани указывает на ее пектолитическую активность. Однако это не доказывает патогенность бактерий в естественных условиях. Ложные положительные результаты возникают вследствие естественного присутствия эндофитичесих или эпифитических микроорганизмов на заражаемой ткани. Дезинфекцию поверхности растительной ткани проводили, погружая ее на 10 минут в 10% раствор домашнего хлорсодержащего моющего средства (5,25% гипохлорид натрия), после чего высушивали на воздухе. Затем нарезанные на кусочки клубни выкладывали на влажную фильтровальную бумагу в чашки Петри и инокулировали 0,1-1 мл бактериальной суспензии (около 106 кл/мл) суточной культуры. Через 48 часов инкубации при 20-27°С проверяли состояние ткани вокруг места заражения с помощью шпателя или иглы на предмет степени мацерации.
Как видно из Таблицы 13, линии трансгенного картофеля N53, N56 и N71 демонстрируют устойчивость к Erwinia carotovora.
ПРИМЕР 10. Устойчивость трансгенных растений картофеля к фитофторозу, вызываемому Phytophthora infestans
Лабораторный тест проводили на листьях, срезанных с трансгенных растений картофеля, экспрессирующих MF3. Заражение проводили популяцией Ph.infestans Московской области. В качестве контроля брали нетрансгенные растения картофеля тех же сортов, что использовали для трансформации растений, и тот же изолят Ph.infestans. По десять листьев с растений каждого сорта заражали путем распыления суспензии зооспор (5-6 зооспор согласно световой микроскопии при 20-кратном увеличении). После заражения листья инкубировали во влажной камере при 18°С в течение 12 часов. Через три дня подсчитывали количество некрозов на 1 см листа.
Листья также заражали каплей 8 мкл суспензии зооспор (1-2 капли/лист). Концентрация зооспор аналогична вышеописанному тесту. Зараженные листья инкубировали во влажной камере в темноте в течение 18 часов. Затем остатки суспензии удаляли фильтровальной бумагой и переносили листья во влажную камеру при 20°С. Через 5-6 дней измеряли площадь повреждения (мм2) и способность к споруляции (по 4-балльной шкале).
На основе полученных результатов этих измерений мы определяли динамику болезни в течение вегетационного периода, а затем вычисляли соответствующие ей потери урожая картофеля (в %).
При этом используется зависимость:
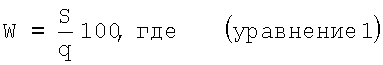
W - потери урожая, %
S - площадь под кривой, описывающей нарастание степени пораженностя растений (AUDP);
q - продолжительность периода от фазы образования соцветий до окончания вегетации растений картофеля, не пораженных фитофторозом, дни.
Величина q определяется скороспелостью сорта картофеля и условиями его выращивания. Средняя величина q для разных групп спелости сортов следующая:
для раннеспелых - 52 дня;
для среднеспелых - 84 дня;
для позднеспелых - 97 дней.
Расчеты проводятся с помощью компьютерной программы или по номограмме (чертеж) для сортообразцов картофеля трех групп спелости (раннеспелых, среднеспелых и позднеспелых). Номограмма включает следующие шкалы:
А - индекс заражения (произведение числа некрозов на их размер), в долях от эталона;
В - инкубационный период, в долях от эталона;
Г - продуктивность спороношения, в долях от эталона;
Б - потенциальные потери урожая картофеля от фитофтороза в результате преждевременного отмирания ботвы, в %.
Шкала Б представлена тремя отрезками для сортов разной скороспелости: для поздних (L), среднеспелых (М), и ранних (Е).
Принятая шкала оценки устойчивости сортов картофеля представлена ниже.
В Таблицах 14, 15 показаны результаты лабораторных тестов на устойчивость трансгенных растений картофеля к Ph.infestans. В Таблице 16 представлены результаты полевых тестов.
ПРИМЕР 11. Устойчивость трансгенных растений картофеля к цистообразующей нематоде Globodera rostochiensis Rol-типа (эксперимент в горшках)
Цисты Globodera rostochiensis Rol-типа были выделены из почвы на Сельскохозяйственной экспериментальной станции ВНИИ картофелеводства, Коренево, Московская область. Анализировали трансгенные растения картофеля, экспрессирующие MF3.
Почву в 300-мл пластиковых горшках заражали цистами Globodera rostochiensis (5000 яиц/100 мл почвы). Одновременно по одному клубню трансгенного картофеля высаживали в каждый горшок. Через 2,5-3 месяца определяли количество цист на каждом растении (Таблица 17).
Количество цист Globodera rostochiensis, собранных с трансгенных растений картофеля
Количество цист, собранных с трансгенных растений линии N53, было существенно меньше, чем на контрольном нетрансгенном растении картофеля.
ПРИМЕР 12. Устойчивость трансгенных растений картофеля к цистообразующей нематоде Globodera rostochiensis Rol-типа (полевой эксперимент)
Цисты Globodera rostochiensis Rol-типа были выделены из почвы на Сельскохозяйственной экспериментальной станции ВНИИ картофелеводства, Коренево, Московская область. Анализировали трансгенные растения картофеля, экспрессирующие MF3. Клубни от каждой линии трансгенного картофеля были высажены в поле на Сельскохозяйственной экспериментальной станции ВНИИ картофелеводства, Коренево, Московская область, где содержались 4816 молодых (неполовозрелых) особей Globodera rostochiensis Rol-типа на 100 мл почвы. Растения картофеля убрали 12 августа 2002 г. и подсчитали количество цист на корнях каждого растения.
Количество цист, собранных с корней трансгенных растений картофеля с геном MF3, выращенных на экспериментальном поле, было меньше, чем на нетрансгенных растениях. Более того, линия N71 показала устойчивость к нематоде. Количество цист, собранных с этой линии, было на 27% меньше, чем на контроле.
ПРИМЕР 13. Тестирование устойчивости к вирусу Х картофеля (PVX) трансгенных растений табака с геном MF3
В качестве источника вируса использовали экстракт из листьев полностью инфицированных PVX растений. Экстракт хранили замороженным при -70°С. По четыре растения табака в возрасте 3-х недель (трансгенные и нетрансгенные) анализировали для каждой линии. Заражали вирусом Х картофеля третий и четвертый сверху лист на каждом растении.
Содержание PVX во всех обработанных (инфицированных) листьях проверяли через 1, 2 и 3 недели после заражения с помощью стандартного иммуноферментного анализа с использованием набора ИФА-PVX (производства ВНИИ картофелеводства, Коренево, Московская область). Измеряли поглощение 0,1% сока из инфицированных листьев согласно прилагающемуся к набору протоколу анализа. Результаты этих экспериментов представлены в Таблице 19. Эти результаты свидетельствуют о высокой степени устойчивости трансгенных растений табака к PVX (более 90%).
ПРИМЕР 14. Испытание трансформантов табака с геном mf3 на устойчивость к Вирусу Мозаики Табака (TMV)
Тестирование и материал - аналогичны Примеру 13. Результаты этих экспериментов представлены в Таблице 20. Как видно, трансгенные растения 177, 152, 171 демонстрируют высокую устойчивость к TMV в течение первых двух недель, а растения 391, 286, 409, 279 - в течение 3 недель после заражения.
ПРИМЕР 15. Тестирование трансгенных растений табака с геном mf3 на устойчивость к вирусу Y картофеля (PVY)
Тестирование и материал - аналогичны Примеру 13. Результаты этих экспериментов представлены в Таблице 21. Согласно этим результатам высокую устойчивость к PVY показали 21 линия из 27 (78%) проанализированных трансгенных линий табака.
ПРИМЕР 16. Тестирование трансгенных растений картофеля с геном MF3 на устойчивость к вирусу Х картофеля (PVX) на естественном инфекционном фоне (Таблица 22)
ПРИМЕР 17. Уровень экспрессии гена MF3 в трансгенных растениях
Мышиные моноклональные антитела к белку MF3 сорбировали на поверхность иммунологической плашки в течение ночи при +4°С, после чего плашку промывали трижды промывочным буфером (PBS-буфер, содержащий 0,05% Твин20). 100 мкл разведенного в 100 и 10 раз, соответственно, сока растений в PBS-буфере раскапывали в лунки иммунологической плашки и инкубировали при 37°С в течение 1 часа, затем промывали ячейки трижды промывочным буфером. По 100 мкл моноклональных мышиных антител к белку MF3, конъюгированных с пероксидазой хрена (HRP) с концентрацией 5 мкг/мл, раскапывали в лунки и инкубировали в течение 1 часа при 37°С. После этого плашку трижды промывали промывочным буфером, а затем в лунки добавляли субстрат (3,3',5,5'-тетраметилбензидин, ТМВ, Sigma). Реакцию окрашивания останавливали через 20 минут, добавляя 2М раствор HCl. Уровень поглощения измеряли согласно инструкции Sigma Chem. Co., USA.
ПРИМЕР 18. Защитные свойства MF3 (SEQ ID:1), обработанного эндопептидазой, против заражения вирусом табачной мозаики (TMV) на листьях растений табака
Чтобы локализовать элиситорную активность на специфической аминокислотной последовательности, белок MF3 подвергли пептидазной и химической обработке. Использовали эндопептидазу Arg-C (для секвенирования, из Clostridium histolyticum, Roche Molecular Biochemicals) в количестве 0,5нг/анализ. Обработку проводили при 37°С в течение 24 часов в 50 мкл раствора, содержащего 0,1М Tris-HCl, pH 7.6; 10 мМ CaCl2 (Christophe Breton et al, 2001). Расщепление эндопептидазой показало наличие фрагментов с активностью, присущей интактному MF3.
Проводили расщепление MF3 бромцианом CNBr в 100 мкл 70% растворе муравьиной кислоты, содержащем несколько кристаллов CNBr. Смесь инкубировали в темноте при комнатной температуре в течение 24 часов в атмосфере аргона, и затем реакции останавливали добавлением 500 мкл воды. Объем смеси слегка упаривали под вакуумом, а муравьиную кислоту удаляли заменой растворителя на воду (Christophe Breton et al, 2001). Химическое расщепление бромцианом показало наличие фрагментов с активностью, присущей интактному MF3.
Использовали эндопротеиназу Lys-C (для секвенирования, из Lysobacter enzymogenes, Roche Molecular Biochemicals) в количестве 0, 2нг/анализ. Обработку проводили при 37°С в течение 16-24 часов в 50 мкл раствора, содержащего 25 mM Tris-HCl, рН 8.5; 1 тМ EDTA; 0.1% SDS. Реакции останавливали добавлением Лаэммли буфера (Christophe Breton et al, 2001).
Расщепление MF3 трипсином убирало антивирусную активность белка. Обработку трипсином проводили при 37°С в течение 1 часа в растворе, содержащем 0.5 М мочевину, 50 мМ Tris-HCl, 1 мМ CaCl2 (рН 7.6). Реакцию останавливали добавляя трихлоруксусную кислоту до конечной концентрации 10% согласно протоколу фирмы Promega.
Расщепление белка трипсином удаляло антивирусную активность MF3. Расщепление MF3 трипсином и Lys-C удаляло антивирусную активность, тогда как обработка эндопротеиназой Arg-C, CNBr и Lys-C показывала, что часть MF3 сохраняла защитные свойства. По крайне мере две аминокислотные последовательности: с 1-й по 80-ю аминокислоты и фрагмент 105-149, - содержат последовательности, которые могут обеспечивать новый РАМР (pathogen-associated molecular pattern, связанный с патогеном молекулярный характер), который запускает в растении защитный ответ. Сравнительный анализ гомологичных белков из различных источников (45 доступных последовательностей) показал, что фрагменты 29-85 и 105-149 являются консервативными (выравнивание с пропусками). Вероятно, для проявления антивирусной активности достаточно пептида длиной около 30 аминокислот. Вот эти консервативные участки:
29 - GA PLVYLQGAGN IIPGLEKALE GKAVGDDLEV AVEPEDAYGE
YAAELVSTLS RSMFE - 85, (SEQ ID:3)
105 - MQIVTI ADLDGDDVTV DGNHPLAGQR LNFKVKIVDI
RDASQEEIA - 149. (SEQ ID:4)
ПРИМЕР 19. Клонирование и определение нуклеотидной последовательности гена MF3 (SEQ ID:2)
В соответствии с N-концевой аминокислотной последовательностью MF3 были синтезированы вырожденные олигонуклеотиды. Была выделена из бактериальных клеток, экспрессирующих MF3, высокомолекулярная хромосомная ДНК, которую обработали 6 рестриктазами (BamHl, EcoRI, Psti, HindIII, Sail, Sphi) отдельно и по парам. Продукты рестрикции фракционировали по молекулярному весу с помощью электрофореза в агарозном геле и переносили на мембрану HybondN с помощью блоттинга. Синтетические олигонуклеотиды метили с помощью Т4-полинуклеотид киназы и радиоактивного [γ-32Р] АТР, после чего использовали как радиоактивную пробу в экспериментах по Саузерн-блот гибридизации. Только одна позитивная полоса на рестрикцию появлялась на рентгеновской пленке. На основе молекулярного веса позитивно гибридизующихся фрагментов была реконструирована рестрикционная карта хромосомного гена этого антивирусного белка.
Проводили электрофорез в 0,7% геле низкоплавкой агарозы SalI-рестриктов хромосомной ДНК. Затем из геля выделяли фрагменты ДНК размером около 3,3 тпн. Далее эти фрагменты обработали рестриктазами HindIII и BamHI, после чего провели электрофорез в 1% агарозном геле и выделили из геля фрагменты ДНК размером около 700 п.н. Эти фрагменты были клонированы в вектор pUC18 по сайтам рестрикции HindIII и BamHI. Компетентные клетки Е.coli (штамм XL1-blue) трансформировали этой лигазной смесью, а затем растили на LB-среде, содержащей ампициллин (50 мг/л). Выросшие колонии переносили на мембрану HybondN и проводили гибридизацию колоний, используя радиоактивно меченный олигонуклеотид в качестве гибридизационной пробы. Около 70% колоний гибридизовались с пробой.
Плазмидную ДНК из положительных клонов выделяли и использовали для секвенирования вставок. Последовательность ДНК, кодирующая N-конец антивирусного белка, как оказалось, находится рядом с BamHI-сайтом. Была обнаружена открытая рамка считывания длиной 486 п.н., начинающаяся ATG-кодоном и заканчивающаяся TGA-кодоном. Согласно ДНК кодирующей области, антивирусный белок состоит из 161 аминокислотного остатка. Процедуры, использованные в описанном выше процессе клонирования, могут быть в основном найдены в руководстве J. Sambrook, Е.F.Fritsch and T.Maniatis, Molecular Cloning: A Laboratory Manual 2nd ed. (Cold Spring Harbor, NY: Cold Spring Harbor Laboratory Press) (1989).
ПРИМЕР 20. Экспрессия белка MF3(SEQ ID:1) в Е. coli
Для модификации концов mf3 мы использовали в качестве матрицы плазмидную ДНК В/Н4 и следующие праймеры:
Nde - mf3 5'-GGAATTCCATATGCTGATCGCCGCC-3
Hind - mf3 5'-CCCAAGCTTAGTGGTGATGGCCACC-3'.
После проведения ПЦР образованный фрагмент расщепляли рестриктазами Ndel и Hindlll, после чего клонировали в вектор pGEMEXl вместо гена 10. Реакционная смесь для ПЦР (50 мкл) содержала прмерно 10 нг плазмидной ДНК, 1 мкМ каждого праймера, 0,2 мМ смесь dNTP, 1-кратный Vent-буфер (20 мМ Tris-HCl, рН 8.8, 10 mM KCl, 2 тМ MgSO4, 10 тМ (NH2SO4, 0.1% Triton X-100) и 1U Vent-ДНК полимеразы (New England Biolabs). ПНР-программа состояла из следующих циклов: 5 мин денатурация при 96°С, затем 30 циклов амплификации: денатурация при 96°С в течение 1 мин, синтез при 72°С в течение 1 мин, отжиг праймеров при 45°С в течение 1 мин; окончательная достройка при 72°С в течение 10 минут. Смесь продуктов ПЦР разделяли электрофорезом в 1% агарозном геле, содержащем 1нг/мл этидиум бромид, при 100В в течение 1 часа. ПЦР-продукт выделяли из геля с помощью набора для элюции Prep-A-Gene DNA Purification Kit (Bio-Rad Laboratories) и растворяли в 50 мкл 1 м ТЕ-буфере. Очищенный ПЦР продукт расщепляли Ndel и Hindlll и фрагменты чистили в 1% агарозном геле. Фрагмент ДНК размером около 500 п.н. элюировали из геля с помощью набора для элюции Prep-A-Gene DNA Purification Kit (Bio-Rad Laboratories) и растворяли в 50 мкл 1 м ТЕ-буфере.
NdeI/AatII-фрагмент вектора pGEMEXl, несущий ген устойчивости к ампициллину, фрагмент AatII/HindIII вектора pGEMEXl, содержащий терминатор гена 10 бактериофага Т7, и mf3 модифицированный ПЦР продукт объединяли с помощью лигирования и последующей трансформации клеток E.coli. В результате была получена плазмида, названная pMF, содержащая модифицированный ген mf3 под контролем промотора и терминатора гена 10 бактериофага Т7. Эта плазмидная ДНК была сексенирована по двум цепям с праймеров Т7 и Sp6 на автоматическом ДНК-секвенаторе (BioRad).
Для производства MF3 белка плазмидная ДНК pMF была перетрансформирована в штамм E.coli BL21(DE3). Этот штамм синтезирует Т7 РНК-полимеразу, и ее уровень синтеза регулируется добавлением IPTG.
ПРИМЕР 21. Очистка белка MF3 (SEQ ID:1)
100 мл ТВ-среды, содержащей 100 мг/мл ампицилина, в колбе на 1 л инокулировали примерно 100 колониями штамма pMF3/BL21(DE3) и инкубировали при 37°С на орбитальном шейкере "Certomat Н" ("B.Braun Melsungen", Germany) при 260 об/мин. При достижении культурой оптической плотности A55onm, равной 2-2,5, добавляли IPTG до конечной концентрации 0,05 мМ, после чего инкубирование продолжали в течение ночи при тех же условиях. На следующий день клетки собирали центрифугированием при 4000 g в течение 30 минут.
Осадок клеток суспендировали в 50 мл следующего буфера: 50 мМ Tris-НС1 рН 8.0, 0.15 М NaCl, 2 мМ EDTA, 2 мг/мл лизоцима, и инкубировали на льду в течение 30 минут. Просветленный лизат наносили на колонку (25х50 мм) с Chelating Sepharose FF, положительно заряженной Ni2+(Pharmacia, Sweden) и уравновешенной буфером А: 50 мМ Tris-HCl (рН 7.5), 0.25 М NaCl. Сорбент промывали этим же буфером. Белки, связавшиеся с сорбентом, элюировали линейным градиентом в возрастающей концентрацией буферов: буфер А (50 mM Tris-HCl, рН 7.5, 0.25 М NaCl), буфер В (50 мМ Tris-HCl рН (7.5), 0.25 М NaCl, 0.25 М имидазол). Скорость потока была 3 мл/мин, а объем градиента - 300 мл. MF3 элюировался примерно при 35% буфера В.
Присутствие MF3 в собранных фракциях анализировали с помощью электрофореза в ПААГ, содержащем SDS. Фракции, содержащие MF3, объединяли и проводили диализ против 20 мМ Tris-HCl, рН 8.0. Затем раствор белка наносили на Mono Q HR10/10 колонку (Pharmacia, Sweden). Элюцию белков проводили градиентом возрастающей концентрации NaCI: буфер А, 50 мМ Tris-HCl (рН 8.0); буфер В, 50 mM Tris-HCl (pH 8.0), 1 М NaCl. Скорость потока - 1 мл/мин, объем градиента - 60 мл. MF3 элюировался примерно при 40% буфера В. Сульфат аммония добавляли к элюированному белку до 25% насыщения, затем проводили центрифугирование при 4000g в течение 30 минут. Супернатант наносили на колонку Phenyl Sepharose HiLoad 16/10 (Pharmacia, Sweden), уравновешенную 50 мМ натрийфосфатным буфером. Белки элюировали линейным градиентом уменьшающейся концентрации сульфата аммония с 0,5 до 0,2М в течение 30 минут, и с 0,2 до 0М в течение 60 минут. Объем градиента - 200 мл, скорость потока - 2 мл/мин. В качестве буфера А использовали 50 мМ Na2HPO4 (рН 6.5), 1.7 М (NH4)2SO4, буфер В - MilliQ вода. MF3 элюировался примерно при 95% буфера В. Присутствие MF3 в собранных фракциях анализировали с помощью электрофореза в ПААГ, содержащем SDS. Фракции, содержащие MF3, объединяли и проводили диализ против 50 мМ ацетата аммония и лиофилизовали.
Затем белок MF3 растворяли в 1 мл 50 mM Tris-HCl (pH8.0) и наносили на колонку (10×500 мм) с Sephadex-G50 (Pharmacia), уравновешенную 50 мМ ацетатом аммония (рН 8,0). Фракции, содержащие MF3, объединяли и лиофилизировали. Выход MF3 составлял около 200 мг с 1 л культуральной среды.
ТВ-среду готовили следующим образом: растворяли 12 г Bacto Trypton, 24 г дрожжевого экстракта и 4 мл глицерина в 900 мл воды и автоклавировали. После охлаждения до 60°С добавляли 100 мл раствора 0.17 М КН2РО4, 0.72 М К2НРО4.
ПРИМЕР 22. Создание генетической конструкции для получения трансгенных растений
Для получения трансгенных растений использовали Т-ДНК перенос в клетки растений с помощью агробактериальной трансформации. Растительный бинарный вектор (р13К) был создан на основе pBinl9 (Bevan, М. 1984) путем клонирования EcoRI-фрагмента из вектора pGL22/MF3 в EcoRI-сайт pBinl9. Вектор pGL22/MF3 содержит промотор и терминатор 35S РНК вируса мозаики цветной капусты, между которыми был проклонирован модифицированный ген mf3 в BamHI-сайт вместо гена hphll (Pietrzak et al., 1986). Модификацию последовательности mf3 поводили с помощью ПЦР на плазмиде В/Н4 со следующими праймерами:
13К5: 5'-GGCCACCATGCTGATCGCCGCCAATAAGG
13К3: 3'-GGGTCAGTGGTGATGGCCACCTTCG
Плазмида р13К перенесена из клеток Е.coli в клетки Agrobacterium tumefaciens LBA4404 с помощью трехродительской конъюгации согласно Van HauteE. et al., 1983.
ПРИМЕР 23. Агробактериальная трансформация растений картофеля
Растения картофеля (Solarium tuberosum, сорта Невский и Луговской) для трансформации были получены из Центра «Биоинженерия» РАН в виде свободной от вирусов in vitro культуры. Растения размножали стерильно, разрезая на отдельные междоузлия, в 3×11,5 см стеклянных культуральных пробирках на стандартной среде для выращивания (РМ), содержащей смесь солей Мурасига-Скуг с добавлением 20 г/л сахарозы, 0,4 мг/л тиамина, 100 мг/л мио-инозитола и 1,7 г/л фитогеля (Sigma, St Louis, Mo. USA), pH5,7. Растения размножали черенкованием отдельных междоузлий на свежую среду каждый месяц. Для трасформации и регенерации использовали фрагменты стебля без листьев. Эти побеги выращивали в камере искусственного климата при 19°С, смешанном освещении 50:50 лампами Grow-Lux™ и флюороресцентного света при 16-часовом дне и 8-часовой ночи.
Листья удаляли как можно ближе к стеблю, стебли собирали в пучок из 20 штук и нарезали на 5-мм фрагменты. Эти экспланты культивировали в 25 мл чашках Петри в течение 2 дней на жидкой MS-среде без добавления фитогормонов. Для заражения эксплантов среду удаляли с помощью ваккумного отсоса и добавляли разбавленную культуру Agrobacterium, оставляли на 15 минут без качания. Затем раствор Agrobacterium удаляли вакуумным отсосом, а экспланты раскладывали на среде для сокультивирования. Примерно 100 эксплантов сокультивировали с агробактерией на одной чашке Петри. Обычно эта система трасформации дает 40 событий трансгеноза на 100 эксплантов. Чтобы минимизировать самоклональную изменчивость процесс регенерации останавливали прежде сбора всех регенерантов. Следовательно, реальный выход составил около 10 независимых трансгенных растений на 100 эксплантов. Сокультивирование проводили в культуральной комнате в течение 2 дней и 16-часовой длине дня при 19°С.
Для трансформации использовали обезоруженный штамм А. twnefaciens LBA4404, содержащий вектор pBinl3K, производный pBinl9. Этот вектор содержит ген nptII, слитый с промотором и терминатором нопалинсинтазы, и ген mf3 под контролем промотора 35S РНК CaMV и терминатора нопалинсинтазы. Бактерию выращивали при 28°С на минимальной А-среде, содержащей 50 мг/л канамицина, при постоянном перемешивании при 200 об/мин в течение ночи. Для заражения растительной ткани ночную культуру агробактерии разводили в 10 раз средой MSO.
После сокультивирования стеблевые экспланты переносили в чашки Петри со средой для регенерации (RM): MS-соли с добавлением 3% сахарозы, 2.0 мг/л глицина, 0.1 мг/д thiamine-HCL, 5.0 мг/л никотиновой кислоты, 0.5 мг/л pyridoxine-HCL, 0.05 мг/л D-биотина, 0.5 мг/л фолиевой кислоты, 100 мг/л мио-инозитола, 0.3 мн/л GA3, 5 мг/л ZR и 0.1 мг/л IAA. Инкубацию вели в течение 3 дней в темноте. Для селекции трансформантов мы использовали RM-среду, содержащую 100-150 мн/л канамицина и 500 мг/л карбеницилина. Чашки Петри помещали стопкой в пластиковый пакет, заклеивали верх пакета и делали несколько отверстий в пакете для вентиляции, инкубировали в культуральной комнате при 19°С и 16-часовой длине дня. Контроль обрабатывали аналогично, кроме заражения агробактерией. Экспланты переносили каждые 2 недели на свежую RM-среду с антибиотиками.
Через 4 недели экспланты начинают образовывать побеги изконцевых каллусов. Самые большие побеги срезали. Только один побег с одного конца переносили на селекцию, после чего экспланты уничтожали.
По пять побегов помещали в чашки Петри, содержащую 50 мл РМ-среды (с добавлением антибиотиков), срезанным концом в среду. Через две недели здоровые, растущие и дающие корни побеги переносили на свежую чашку Петри с РМ-средой, по одному на чашку. Если побеги не давали корней в течение 4 недель культивирования, их уничтожали.
Черенки из одного междоузлия культивировали для клонального размножения на РМ-среде в 25×150 мм пробирках при 19°С и 16-часовом дне, по одному в пробирке. In vitro культивируемые побеги обычно переносили на свежую среду с интервалом 3-4 недели. Растения, предназначенные для переноса в теплицу, подращивали в течение 7-10 дней.
ПРИМЕР 24. Трансформация листовых дисков табака с помощью А. tumefaciens
Растения табака (Nicotiana tabacum cv. Samsun NN) размножали стерильно в виде черенков из одного междоузлия в 0,8 л стеклянных банках на А 1-среде. Для этого срезали либо верхушку с апикальной меристемой, либо нарезали стебель на фрагменты, состоящие из одного междоузлия с боковой почкой, и помещали в А1-среду. Экспланты формировали корни за 10-14 дней. Из апикальной меристемы или из аксиллярной почки развивался новый побег.
Стерильные побеги растили при 24°С, 16-часовом дне/8-часовой ночи, при рассеянном освещении. Черенкование проводили через 3-4 недели.
Для трансформации срезали листья и переносили их в чашку Петри, содержащую влажную фильтровальную бумагу Whatman 1M. Удаляли среднюю жилку и нарезали диски диаметром около 5 мм стерильным дыроколом. 40-50 дисков помещали верхней стороной вниз в 9-см чашку Петри, содержащую 10 мл инфекционной среды А2. В каждую чашку добавляли по 2,5 мл культуры Agrobacterium AGLO, несущей вектор pBinl3K, производный pBinl9. Этот вектор содержит ген nptll, слитый с промотором и терминатором нопалинсинтазы, и ген mf3 под контролем промотора 35S РНК CaMV и терминатора нопалинсинтазы. Бактерию выращивали при 28°С на минимальной А-среде, содержащей 50 мг/л канамицина, при постоянном перемешивании при 200 об/мин до достижения оптической плотности А600нм 0,6. После 15-20 минут сокультивации листовые экспланты промокали на стерильной фильтровальной бумаге Whatman 1M и помещали в чашку Петри абаксиальной стороной вниз на А2-среду, содержащую 8 г/л агара, и инкубировали в камере искусственного климата при слабом освещении. Через 3 дня листовые диски отмывали в среде А2, содержащей 500 мг/л карбеницилина, просушивали на стерильной фильтровальной бумаге Whatman 1 M и помещали в чашку Петри с А3-средой для каллусообразования, содержащей 100 мг/л канамицина и 500 мг/л карбеницилина. Экспланты инкубировали при 24°С, 16-часовом дне/8-часовой ночи, при рассеянном освещении. Каждую неделю листовые диски перекладывали на свежую A3-среду. Через 3-4 недели морфогенные каллусы отделяли от листовых дисков и помещали на А4-среду с 100 мг/л канамицина. 2-3 недели спустя побеги срезали с каллусов и переносили на среду А5 для укоренения с 100 мг/л канамицина. Укоренившиеся побеги размножали стерильным черенкованием на А 1-среде или переносили в почву для выращивания в теплице.
Для тех, кто имеет соответствующие навыки, будет ясно, что много изменений может быть сделано в вышеописанных деталях предпочтительных воплощений настоящего изобретения, не отступая от основных вышезаявленных принципов. Сфера настоящего изобретения должна поэтому быть определена только в соответствии со следующими целями.
ЛИТЕРАТУРА
1. Pietrzak, M., Shillito, R.D., Hohn, Т., Potrykus, I. Expression in plants of two bacterial antibiotic resistance genes after protoplast transformation with a new plant expression vector. Nucleic Acids Res., 1986, v.l4, pp.5857-5868.
2. Bevan, M. Binary Agrobacterium vectors for plant transformation. Nucleic Acids Res., 1984, v.12, pp.8711-8721.
3. Van Haute E., Joos, H., Maes, S., Warren, G., Van Montagu M., Schell, J.-Intergeneric transferband exchange recombination of restriction fragment cloned in pBR322:a novel strategy for reversed genetics of the Ti plasmids of Agrobacterium tumefaciens. EMBO J., 1983, v.2, pp.411-418.
4. Baulcombe D. 1994. Novel strategies for engineering virus resistance in plants. Current Opinion in Biotechnology., 5, 117-124.
5. Bradford M. 1976. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem., 72, 276-287.
6. Christophe Breton, Hichem Chellil, Majida Kabbaj-Benmansoiir, Eric Carnazzi, Rene Seyer, Sylvie Phalipou, Denis Morin, Thieny Durroux, Hans Zingg, Claude Barberis, and Bernard Mouillac. 2001. Direct Identification of Human Oxytocin Receptor-binding Domains Using a Photoactivatable Cyclic Peptide 20 Antagonist. Comparison with the human Via vasopression receptor. J. Biol. Chem., Vol.276, Issue 29, 26931-26941, July 20, 2001.
7. Cion R.A. The manual of determinative microbiology. OGIZ-SELHOZGIZ. Moscow. 1948. p.484 (In Russian).
8. de Barjac H. and A.Bonnefoi. 1967. A classification of strains of Bacillus 25 thuringiensis Berliner with a key to their differentiation. J Invertebr. Pathol. 11, 335-347.
9. de Barjac H. and A.Bonnefoi. 1967. Classification des souches de Bacillus thuringiensis. С R Acad Sci (Paris) 264, 1811-1813.
10. Golishin N.M. Fungicide in agriculture, Moscow, "Kolos" 1982. pp.20-66.
11. Jackman P.J.H. Microbial systematics based on electrophoretic whole-cell protein patterns. Methods in microbiology. Academic Press, London. Ed. by R.R.Colwell and Grigorova R.. 1987. Vol.19, pp.210-224.
12. Krieg A. 1968. A taxonomic study of Bacillus thuringiensis Berliner. J. Invertebr. Pathol. 12, 366-378.
13. Laemmli U.K. 1970. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature, 227, 680-685.
14. Latterell F.M., Marchetti M.A., Grove B.R. Co-ordination of effort to establish an international system for race identification in Pyricularia oryzae. The Rice Blast Disease. Baltimore, Maryland: The Johns Hopkins Press: 1964. pp.257-268.
15. Sambrook J., Fritsch E.F. and Maniatis Т., 1989. Molecular cloning. A Laboratory Manual 2nd ed. Cold Spring Harbor Laboratory Press.
16. Schroder K, Zuber P., Willimsky G., Wagner B. and Marahiel MA. 1993. Mapping of the Bacillus subtilis csp В gene and cloning of its homologs in thermophilic, mesophilic and psychrotrophic bacilli. Gene, 136, 277-280.
17. Smith N.R., R.E.Gordon and F.E.Clark. 1952. Aerobic sporeforming bacteria. Agr.Monogr. 16 U S Dept. Agr, pp.1-148.
18. The shorter Bergey's manual of determinative bacteriology // The Williams and Wilkins Company, Baltimore. Ed. by Holt. 1980. P.495. 15.



Изобретение относится к области биохимии и биотехнологии и может быть использовано в растениеводстве. Из клеток микроорганизма Pseudomonas fluorescence выделен новый белок (MF3), обладающий способностью повышать устойчивость растений к инфекционным болезням и растительным паразитам. Клонирован ген mf3, определена полная кодирующая белок последовательность и сконструированы экспрессирующие векторы, включающие указанную последовательность или ее часть, кодирующую фрагмент MF3, который сохраняет защитные свойства полноразмерного белка. Путем введения созданных векторных конструкций в клетки организмов-хозяев получены культуры растительных клеток, экспрессирующие MF3, устойчивые к инфекционным болезням и/или растительным паразитам трансгенные растения, а также штаммы микроорганизмов, продуцирующие рекомбинантные белки. Очищенный белок MF3 или его активный фрагмент по изобретению предлагается использовать в составах для защиты растений от патогенов и вредителей. 8 с. и 1 з.п. ф-лы, 1 ил., 24 табл.
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
| GODOY A.V | |||
| ET AL., Plant Science, 2000, v.152, p.123-134. | |||

