ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к области заболеваний растений. Более конкретно, изобретение относится к новому вирусу растений, выделенному из томата, к способам детекции указанного вируса, к способам детекции устойчивых растений и к способам получения устойчивых растений.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Томат Solanum lycopersicum (ранее Lycopersicon esculentum) является чувствительным к большому числу видов вирусов. Некоторые из наиболее известных вирусов томата включают в себя вирус бронзовости томата (TSWV; род Tospovirus), вирус мозаики пепино (PepMV; род Potexvirus) и вирус желтой курчавости листьев томата (TYLCV; род Begomovirus). Повреждения, которые эти заболевания наносят растению, находятся в диапазоне от обесцвечивания листьев и некротических очагов до серьезной потери урожайности и гибели растения.
Возможность получать устойчивые растения является крайне важной для коммерческих селекционеров, и для некоторых из вирусов, наносящих наибольший экономический ущерб, получены устойчивые варианты растений. Однако время от времени появляются новые вирусы, которые могут наносить значительный ущерб урожаю.
В 1996 г. опубликован новый вирус томата, инфицировавший растения томата в США и Италии с 1993 г, и названный вирусом инфекционного хлороза томатов (TICV; род Crinivirus; Duffus et al., 1996). Другой новый вирус томата из того же рода опубликован в 1998. Показано, что этот вирус инфицировал растения томата в США с 1989, и его назвали вирусом хлороза томатов (ToCV; Wisler et al, 1998). Доказано, что оба этих новых вируса распространяет белокрылка, насекомое, являющееся очень эффективным переносчиком заболевания.
Обычно считают, что географическое распределение известных вирусов будет увеличиваться и что будут продолжать появляться новые вирусы, частично в результате рекомбинации различных вирусов с образованием новых штаммов или новых вирусов. Разработка устойчивых культурных сортов может играть важную роль в успешном контроле этих заболеваний.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Недавно на растениях томатов из Испании обнаружили новый вирус, вызывающий симптомы, которые нельзя приписать какому-либо из известных вирусов. У растений обнаруживали некротические повреждения на листьях и коричневые кольца на плодах, а также пониженный рост. Серологические тесты (ELISA) показали присутствие вируса мозаики пепино (PepMV). Исследования электронной микроскопией действительно выявили палочковидные частицы, типичные для потекс-вирусов (Potexvirus). Однако в инфицированной ткани листа обнаружены также вирусные частицы сферической формы. Авторам настоящего изобретения удалось выделить новый вирус из комплекса с PepMV. Новый вирус предварительно назвали вирус томата торрадо (ToTV).
Очень важной является возможность распознать заболевание, чтобы проследить его источник, осуществить мониторинг его эпидемиологии и предупредить его возможное распространение. Только тогда можно принимать адекватные меры для изоляции растений и начинать фитосанитарную профилактику. На настоящий момент не существует доступных диагностических средств. Следовательно, существует необходимость разработки диагностических средств для данного заболевания. Более того, в настоящее время неизвестны растения, обладающие специфической устойчивостью к этому новому вирусу, в то время как существует необходимость разработки таких устойчивых растений.
Изобретение в первом аспекте относится к вирусу растений, предварительно названному вирус томата торрадо (ToTV), депонированному в немецкой коллекции микроорганизмов и клеточных культур (DSMZ) в Брауншвейге 24 ноября 2004 г. под номером для ссылки депозитов ToTV-E01 (DSM 16999).
Вирус вызывает симптомы заболевания в растениях томата, так же как в других растениях, и может вызывать симптомы самостоятельно или в комплексе с другими вирусами или заболеваниями.
Первые системные симптомы состоят в некротических пятнах на верхушке растения, начиная с основания листочков сложного листа. Некротические пятна набухают и окружены светло-зеленой или желтой областью (см. Фиг.1). Не все системно инфицированные листья проявляют симптомы, однако в этих листьях можно обнаружить вирус, например, посредством электронной микроскопии. На плодах, инфицированных ToTV, наблюдаются некротические кольца. Рост инфицированных растений может быть сниженным по сравнению с неинфицированными растениями.
Приведенное выше описание относится к растениям, заново инфицированным выделенным вирусом, и не обязательно будет точно отражать симптомы, встречающиеся в полевых условиях. Такие факторы, как раса или вариетет растения, стадия развития, воздействие дополнительных заболеваний и абиотические факторы (например, температура и относительная влажность), будут, в конечном счете, определять проявление и характеристики симптомов.
Вирусные частицы являются сферическими (икосаэдрическими) по форме с диаметром приблизительно 28 нм (см. Фиг.2). Вирусные частицы состоят по меньшей мере из трех белков капсида приблизительно по 23, 26 и 35 кДа (см. Фиг.3). После очистки вирус отображает на дисплее по меньшей мере две видимые полосы в градиенте сульфата цезия. Верхняя видимая полоса (верхняя фракция вируса) содержит молекулу РНК приблизительно 5,5 т.н. (более точно, 5,2 т.н.), а нижняя видимая полоса (нижняя фракция) содержит молекулу РНК приблизительно 8 т.н. (более точно, 7,7 т.н.) (см. Фиг.4). Инокуляция растений табака обеими объединенными полосами приводит к инфекции.
ToTV способен механически передаваться нескольким видам Nicotiana. Общепринятый буфер для инокуляции (например, 0,03 M фосфатный буфер с pH 7,7) является пригодным. Для размножения ToTV предпочтительными являются N.glutinosa, N.tabacum и N.benthamina. Виды табака N.hesperis '67A' и N.occidentalis 'Р1' являются очень чувствительными к ToTV и проявляют системные симптомы после 3-4 суток. У этих видов томата развивается сильный некроз за короткое время, и, таким образом, они являются более подходящими для использования в качестве индикаторного растения, чем в качестве хозяина для размножения. N.glutinosa реагирует местными очагами хлороза, системным хлорозом и умеренной деформацией листьев. N.benthamiana не проявляет местных симптомов и реагирует системным хлорозом и деформацией листьев.
Кроме того, изобретение относится к вирусу, содержащему по меньшей мере одну последовательность нуклеиновой кислоты, выбранную из группы, состоящей из SEQ ID NO:1 и SEQ ID NO:2 и последовательностей, обладающих по меньшей мере 30%, предпочтительно по меньшей мере 40%, предпочтительно по меньшей мере 50%, предпочтительно по меньшей мере 60%, более предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 80%, еще более предпочтительно по меньшей мере 90%, еще более предпочтительно по меньшей мере 95%, еще более предпочтительно по меньшей мере 98% и наиболее предпочтительно по меньшей мере 99% гомологией с ними. Такие вирусы также охвачены термином ToTV, как используется в данном описании.
В предпочтительном варианте осуществления вируса по изобретению, обладающего вышеуказанной гомологией последовательности, указанный вирус связан с заболеваниями томата, известными под названиями «Торрадо», «Марчитец» и/или «Заболевание шоколадной пятнистости», и/или указанный вирус, на основании численного таксономического анализа таксономических признаков, в основном, как определено в таблице 1, обладает более близким родством с вирусом, как определено по п.1 формулы изобретения, чем с любым другим изолятом вируса, доступным в публичных коллекциях, и указанный вирус обладает необходимыми характеристиками, связанными с заболеванием, вызывающим некротические повреждения у томата.
В другом аспекте изобретение относится к выделенной или рекомбинантной нуклеиновой кислоте, содержащей последовательность нуклеиновой кислоты, выбранную из группы, состоящей из SEQ ID NO:1, SEQ ID NO:2, последовательностей, обладающих по меньшей мере 50%, предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 80%, еще более предпочтительно по меньшей мере 90%, еще более предпочтительно по меньшей мере 95%, еще более предпочтительно по меньшей мере 98% и наиболее предпочтительно по меньшей мере 99% гомологией нуклеотидной последовательности c SEQ ID NO:1 или SEQ ID NO:2, комплементарных им цепей и их ToTV-специфических фрагментов. Такую нуклеиновую кислоту можно получить из вируса по изобретению.
В другом аспекте изобретение относится к полинуклеотиду, способному гибридизоваться в строгих условиях с выделенной или рекомбинантной нуклеиновой кислотой по изобретению, как описано выше.
В другом аспекте изобретение относится к выделенному или рекомбинантному полипептиду, полученному из вируса по изобретению, или его ToTV-специфическому фрагменту. В предпочтительном варианте осуществления указанный полипептид выбран из группы, состоящей из белков капсида 23, 26 и 35 кДа и их ToTV-специфических фрагментов.
В другом аспекте изобретение относится к антигену, содержащему полипептид по изобретению или его ToTV-специфический фрагмент.
В другом аспекте изобретение относится к антителу, специфически направленному против антигена по изобретению.
В другом аспекте изобретение относится к способу получения антитела против ToTV, включающему стадии: a) получения вируса ToTV или его (рекомбинантного) белка, или фрагмента его пептида; b) иммунизации подходящего позвоночного-хозяина указанным вирусом, белком или фрагментом пептида и c) сбора из крови (включая сыворотку) или спленоцитов указанного позвоночного-хозяина антител против указанного вируса, белка или фрагмента пептида. В предпочтительном варианте осуществления указанный способ дополнительно включает в себя стадии: d) отбора одного продуцирующего антитело спленоцита, e) слияния указанного спленоцита с иммортализованной линией клеток гибридомы и f) обеспечения возможности указанной слитой гибридоме продуцировать моноклональные антитела.
В другом аспекте изобретение относится к антителу, которое можно получить способом получения антитела против ToTV по изобретению.
В другом аспекте изобретение относится к способу идентификации изолята вируса как вируса ToTV, включающему реакцию указанного изолята вируса или его компонента с антителом по изобретению.
В другом аспекте изобретение относится к способу идентификации изолята вируса как вируса ToTV, включающему реакцию указанного изолята вируса или его компонента с полинуклеотидом по изобретению.
В другом аспекте изобретение относится к способу детекции присутствия ToTV в образце, включающему определение в указанном образце присутствия вируса ToTV или его компонента путем реакции указанного образца с полинуклеотидом или с антителом по изобретению.
В другом аспекте изобретение относится к способу идентификации устойчивого к ToTV растения, включающему стадии: a) воздействия на растение или часть растения инфекционной дозы ToTV, и b) идентификации указанного растения как устойчивого к ToTV, когда, после указанного воздействия, или i) симптомы заболевания в указанном растении или части растения остаются отсутствующими, или их проявление задержано, или, по меньшей мере, тяжесть уменьшена, или они являются локализованными по сравнению с чувствительным контрольным растением, и/или ii) вирус ToTV или геномные последовательности ToTV не присутствуют в указанном растении или части растения или присутствие вируса ToTV является по меньшей мере уменьшенным количественно по сравнению с чувствительным контрольным растением. Стадия a) включает период инкубации достаточно долгой продолжительности, чтобы позволить установление поддающихся выявлению симптомов заболевания в чувствительных контрольных растениях, подвергавшихся воздействию сравнимой инфекционной дозы вируса. Посредством осуществления данного способа можно идентифицировать у растения все формы устойчивости, включая полную устойчивость, частичную устойчивость, гиперчувствительность и толерантность. Чтобы подтвердить толерантность, необходимо подтвердить (системное) присутствие вируса в растении (клетках). Стадия b) может включать осуществление способа детекции присутствия ToTV в образце из указанного растения или части растения по настоящему изобретению, где применяют антитело или полинуклеотид по изобретению в общепринятых способах для анализов гибридизации нуклеотидов или иммуноанализов, хорошо известных специалисту в данной области. Альтернативно, стадия b) может включать приведение в контакт части указанного подвергшегося воздействию растения с чувствительным индикаторным растением. Таким образом, можно выявлять наличие системной или местной инфекции в указанном подвергшемся воздействию растении через наблюдение возникновения заболевания в индикаторном растении или даже в дополнительном контактировавшем индикаторном растении, контактировавшем с указанным первым контактировавшим индикаторным растением.
В другом аспекте изобретение относится к способу получения устойчивого к ToTV растения, включающему стадии идентификации устойчивого к ToTV донорного растения одним из вышеуказанных способов для идентификации устойчивого к ToTV растения по изобретению, скрещивания указанного устойчивого к ToTV донорного растения с реципиентным растением (где реципиентное растение может быть либо чувствительным к ToTV, либо устойчивым к ToTV, но является, соответственно, устойчивым к ToTV растением в случае, если устойчивый фенотип обусловлен рецессивным геном), и отбора из растений-потомков (например, F1, F2 и самоопыленного растения) устойчивого растения способом идентификации устойчивости к ToTV у растения, как описано выше. В случае, если признак устойчивости является рецессивным признаком, устойчивые растения можно обнаружить среди растений-потомков самоопыления F1 или F2 или более дальних поколений. В предпочтительных вариантах осуществления по данному аспекту указанное устойчивое к ToTV донорное растение и реципиентное растение представляют собой растения семейства Solanaceae или семейства Cucurbitaceae. В других предпочтительных вариантах осуществления по данному аспекту указанное реципиентное растение представляет собой томат, баклажан, перец, дыню, арбуз или огурец, более предпочтительно растение из вида Solanum lycopersicum, наиболее предпочтительно линии S.lycopersicum, обладающей коммерчески желательными характеристиками.
В другом аспекте изобретение относится к устойчивому к ToTV растению, предпочтительно томату, баклажану, перцу, дыне, арбузу или огурцу, или его части, такой как семя, которые можно получить способом получения устойчивого к ToTV растения по изобретению.
В другом аспекте изобретение относится к диагностическому набору для детекции присутствия ToTV в образце или для идентификации устойчивости к ToTV в растении, содержащем вирус, полинуклеотид, полипептид, антиген и/или антитело по изобретению.
В другом аспекте изобретение относится к применению вируса, полинуклеотида, полипептида, антигена или антитела по изобретению для получения диагностической композиции.
В другом аспекте изобретение относится к диагностической композиции, содержащей вирус, полинуклеотид, полипептид, антиген или антитело по изобретению.
В другом аспекте изобретение относится к применению ToTV или частей вирусного генома ToTV в качестве экспрессирующего вектора.
В другом аспекте изобретение относится к применению ToTV или частей вирусного генома ToTV для получения обусловленной патогеном устойчивости у растений.
В другом аспекте изобретение относится к применению аттенуированной формы вируса ToTV, или его генома, или его частей для премуниции растения.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Определения
Как используется в данном описании, термин «часть растения» обозначает часть растения, включая отдельные клетки и клеточные ткани, такие как клетки растения, являющиеся интактными в растениях, скопления клеток и культуры тканей, из которых можно регенерировать растения. Примеры частей растения включают, без ограничения, отдельные клетки и ткани из пыльцы, семяпочек, листьев, зародышей, корней, кончиков корней, пыльников, цветов, плодов, отростков стебля и семян; так же как пыльцу, семяпочки, листья, зародыши, корни, кончики корней, пыльники, цветы, плоды, стебли, побеги, черенки, корневище, семена, протопласты, каллюсы и т.п.
Термин «образец» включает в себя образец из растения, из части растения или из переносчика, или образец почвы, воды или воздуха.
Термин «переносчик», как используется в данном описании, относится к распространяющему заболевание агенту или веществу. Переносчики ToTV в полевых условиях могут включать, без ограничения, животных, таких как Arthropoda (в частности, из классов Insecta и Arachnida), Nematoda (в частности, из класса Adenophorea), а также более крупных животных, таких как, например, птицы, кролики и мыши, грибы (т.е. тип Eumycota, в частности грибы класса Phycomycota), (паразитические) растения (включая члены семейства Cuscutaceae), пыльцу, семена, воду, твердые частицы и даже руки человека, оборудование и обувь.
Термин растение-«потомок» относится к любому растению, полученному в качестве потомства благодаря вегетативному или половому размножению от одного или нескольких родительских растений или их потомков. Например, растение-потомок можно получить клонированием или самоопылением родительского растения, или скрещиванием двух родительских растений, и оно включает в себя растения, полученные самоопылением, так же как F1 или F2, или более дальние поколения. F1 представляет собой потомство первого поколения, полученное от родителей, по меньшей мере одно из которых применяли первый раз в качестве донора признака, тогда как потомство второго поколения (F2) или последующих поколений (F3, F4 и т.д.) представляет собой экземпляры, полученные от самоопылений F1, F2 и т.д. F1 может, таким образом, быть (и обычно является) гибридом, полученным от скрещивания между двумя родителями из разведения гомозигот (гомозиготы из разведения являются гомозиготными по признаку), тогда как F2 может быть (и обычно является) потомством от самоопыления указанных гибридов F1.
Термин «устойчивый», как используется в данном описании, относится к растению, способному сопротивляться размножению вируса в его клетках и/или (системному) передвижению вируса к другим клеткам и/или развитию симптомов заболевания после инфекции указанным вирусом, где вирус способен к инфицированию и размножению в соответствующих неустойчивых или чувствительных вариететов указанного растения. Термин используют для включения таких отдельно идентифицируемых форм устойчивости, как «полная устойчивость», «иммунитет», «частичная устойчивость», «гиперчувствительность» и «толерантность».
«Полная устойчивость» обозначает полную неспособность вируса развиваться после инфекции и может быть либо результатом неспособности вируса входить в клетку (отсутствие начальной инфекции), либо может быть результатом неспособности вируса размножаться в клетке и инфицировать последующие клетки (отсутствие подпороговой инфекции, отсутствие распространения). Наличие полной устойчивости можно определить установлением отсутствия вирусных частиц или вирусной РНК в клетках растения, так же как отсутствия каких-либо симптомов заболевания в указанном растении после воздействия на указанное растение инфекционной дозы вируса (т.е. после «инфекции»). Среди селекционеров данный фенотип часто обозначают как «иммунный». «Иммунитет», как используется в данном описании, таким образом, относится к форме устойчивости, характеризуемой отсутствием вирусной репликации, даже когда вирус активно переносят в клетки, например, посредством электропорации.
«Инфекционную дозу» определяют как дозу вирусных частиц или нуклеиновой кислоты вируса, способную инфицировать растение, где доза может различаться между растениями и между тестируемыми изолятами ToTV. Теоретически, количество от приблизительно 1-10 до приблизительно 500-5000 вирусных частиц указанного вируса или его нуклеиновых кислот будет являться достаточным. Инфекции, таким образом, можно достигать механической инокуляцией очищенных частиц вируса или нуклеиновой кислоты вируса в растения.
«Частичная устойчивость» обозначает уменьшенное размножение вируса в клетке, уменьшенное (системное) передвижение вируса и/или уменьшенное развитие симптомов после инфекции. Наличие частичной устойчивости можно определить установлением системного присутствия низких титров вирусных частиц или вирусной РНК в растении и присутствия уменьшенных или отсроченных симптомов заболевания в указанном растении после воздействия на указанное растение инфекционной дозы вируса. Титры вируса можно определить с использованием способа количественной детекции (например, способа ELISA или количественной полимеразной цепной реакции с обратной транскрипцией [RT-PCR]). Среди селекционеров данный фенотип часто обозначают как «промежуточно устойчивый».
Термин «гиперчувствительный» относится к форме устойчивости, при которой инфекция остается местной и не распространяется системно, например, из-за местного некроза инфицированной ткани или отсутствия системного передвижения за пределы инокулированной ткани. В гиперчувствительных растениях обнаруживают местные, но тяжелые симптомы заболевания, и в таких растениях можно установить местное присутствие вируса.
Термин «толерантный» применяют в данном описании для обозначения фенотипа растения, при котором симптомы заболевания остаются отсутствующими после воздействия на указанное растение инфекционной дозы вируса, при котором можно установить присутствие системной или местной вирусной инфекции, размножение вируса, по меньшей мере присутствие вирусных геномных последовательностей в клетках указанного растения и/или их интеграцию в геном. Таким образом, толерантные растения являются устойчивыми для проявления симптомов, однако бессимптомные растения являются носителями вируса. Иногда вирусные последовательности могут присутствовать или даже размножаться в растениях, не вызывая симптомов заболевания. Этот феномен известен также как «латентная инфекция». Некоторые ДНК- и РНК-вирусы могут становиться не подлежащими выявлению после первичной инфекции, но появляться вновь позже и приводить к острому заболеванию. При латентной инфекции вирус может существовать в истинно латентной неинфекционной скрытой форме, возможно, в виде интегрированного генома или эписомального агента (так что вирусные частицы нельзя обнаружить в цитоплазме, в то время как способами PCR можно показать присутствие последовательностей вирусной нуклеиновой кислоты) или в виде инфекционного и постоянно реплицирующегося агента. Реактивированный вирус может распространяться и вызывать эпидемию среди контактирующих чувствительных растений. Наличие «латентной инфекции» нельзя отличить от наличия «толерантного» фенотипа у растения.
Термин «чувствительный» используют в данном описании для обозначения растения, не обладающего устойчивостью к вирусу, что приводит к проникновению вируса в клетки растения, размножению и системному распространению вируса, вызывая симптомы заболевания. Термин «чувствительный», таким образом, является эквивалентным «неустойчивому». В чувствительном растении обнаруживают нормальные титры вируса в клетках после инфекции. Таким образом, чувствительность можно определить посредством установления нормальных (т.е. относительно других вирусных инфекций у растений) титров вирусных частиц или вирусной РНК в клетках растения и присутствия нормальных симптомов заболевания (т.е. относительно симптомов заболевания, как описано здесь для растения, из которого первоначально выделен ToTV) в указанном растении после воздействия на указанное растение инфекционной дозы вируса.
Термин «чувствительный» отражает симптоматическую реакцию чувствительного растения после вирусной инфекции. Реакция или симптомы могут быть более или менее тяжелыми в зависимости от уровня чувствительности растения. Если вирус повреждает или даже вызывает гибель растения, указанное растение определяют как «чувствительное».
Растения, искусственно инокулированные аттенуированными штаммами вируса, впоследствии являются защищенными от близкородственных вирулентных вирусов. В качестве защищающих вирусов можно использовать либо существующие в природе умеренные штаммы, либо аттенуированный штамм (искусственно полученный умеренный мутант). Предпочтительно, чтобы достигнуть премуниции растения против ToTV, можно использовать аттенуированный штамм ToTV, который не вызывает симптомов или для которого показано по меньшей мере уменьшенное проявление симптомов в инфицированном растении относительно вирулентного штамма ToTV. Способы получения аттенуированного вируса могут включать, например, случайный мутагенез генома ToTV и скрининг штаммов с аттенуацией симптомов. Приведена непосредственная ссылка на способы получения аттенуированных вирусов растений, как описано в статьях Takeshita et al., 2001; Lu et al., 2001; Hagiwara, et al., 2002; Hirata et al., 2003.
Как используется в данном описании, термин «томат» означает любое растение, линию или популяцию Lycopersicon или Solanum, включающие, без ограничения, представленные в списке ниже. Недавно номенклатура Lycopersicon была изменена.
Новая номенклатура Lycopersicon представлена в следующем списке (из: Peralta, Knapp & Spooner, неопубликованная монография (см.: http://www.sgn.cornell.edu "Guide to revised Solanum nomenclature")).
«Экспрессирующий вектор» определяют как молекулу нуклеиновой кислоты, содержащую ген, обычно гетерологичный ген, экспрессирующийся в клетке-хозяине. Как правило, этот ген содержит кодирующую белок последовательность. Экспрессию гена всегда помещают под контроль промотора, и такой ген, как говорят, является «функционально связанным с» промотором. Термин «гетерологичный» относится к молекуле ДНК или к популяции молекул ДНК, которая в природных условиях не существует в данной клетке-хозяине.
Термин «полинуклеотид», как используется в данном описании, является взаимозаменяемым с термином «нуклеиновая кислота» и относится к мультимеру нуклеотидов или полимерной форме нуклеотидов, обладающим любым числом нуклеотидов, например дезоксирибонуклеотидов или рибонуклеотидов, или к полученным синтетически соединениям (например, PNA, как описано в патенте США № 5948902 и процитированных там ссылках), и может быть либо двух-, либо одноцепочечным. Полинуклеотид может гибридизоваться с существующими в природе полинуклеотидами специфическим к последовательности образом, аналогичным гибридизации двух существующих в природе полинуклеотидов, например может участвовать во взаимодействиях Уотсон-Криковского спаривания оснований. Термин также включает в себя модифицированные, например, метилированием и/или кэппированием и немодифицированные формы полинуклеотида.
Термины «рибонуклеиновая кислота» и «РНК», как используется в данном описании, обозначают полимер, состоящий из рибонуклеотидов.
Термины «дезоксирибонуклеиновая кислота» и «ДНК», как используется в данном описании, обозначают полимер, состоящий из дезоксирибонуклеотидов.
Термин «олигонуклеотид» обозначает короткую последовательность мономеров нуклеотидов (обычно от 6 до 100 нуклеотидов), соединенных фосфорными связями (например, фосфодиэфирной, алкил- и арилфосфатной, фосфоротиоатной) или нефосфорными связями (например, пептидной, сульфаматной и другими). Олигонуклеотид может содержать модифицированные нуклеотиды, имеющие модифицированные основания (например, 5-метилцитозин) и модифицированные группы сахаров (например, 2'-O-метилрибозил, 2'-O-метоксиэтилрибозил, 2'-фторорибозил, 2'-аминорибозил и т.п.). Олигонуклеотиды могут представлять собой природные или синтетические молекулы двух- и одноцепочечной ДНК и двух- и одноцепочечной РНК кольцевой, разветвленной или линейной формы и, не обязательно, содержат домены, способные формировать вторичные структуры (например, стебель-петля, псевдоузлы и структуры соприкасающихся петель).
Термин «гомология нуклеотидной последовательности», как используется в данном описании, обозначает наличие гомологии между двумя полинуклеотидами. Полинуклеотиды обладают «гомологичными» последовательностями, если последовательность нуклеотидов в двух последовательностях является одинаковой при сравнении по максимальному соответствию. Сравнение последовательности между двумя или более полинуклеотидами, как правило, проводят сравнением частей двух последовательностей в окне сравнения для идентификации и сравнения локальных областей сходства последовательности. Окно сравнения обычно составляет приблизительно от 20 до 200 непрерывных нуклеотидов. «Процент гомологии последовательности» полинуклеотидов, например 50, 60, 70, 80, 90, 95, 98, 99 или 100 процентов гомологии последовательности, можно определить сравнением двух оптимально выровненных последовательностей в окне сравнения, где часть последовательности полинуклеотида в окне сравнения может содержать добавления или делеции (т.е. пропуски) по сравнению с контрольной последовательностью (которая не содержит добавлений или делеций) для оптимального выравнивания двух последовательностей. Процент вычисляют посредством (a) определения числа положений, в которых идентичные основания нуклеиновой кислоты присутствуют в обеих последовательностях, для получения числа совпадающих положений; (b) деления числа совпадающих положений на общее число положений в окне сравнения; и (c) умножения результата на 100 для получения процента гомологии последовательности. Оптимальное выравнивание последовательностей для сравнения можно проводить компьютеризированным исполнением известных алгоритмов или визуальным контролем. Легко доступными алгоритмами сравнения последовательностей и множественного выравнивания последовательностей являются, соответственно, основная программа поиска локального выравнивания (BLAST) (Altschul et al., 1990; Altschul et al., 1997) и программа ClustalW, обе доступны через интернет. Другие подходящие программы включают, без ограничения, GAP, BestFit, PlotSimilarity и FASTA в пакете программного обеспечения Wisconsin Genetics (Genetics Computer Group (GCG), Madison, WI, USA) (Devereux et al., 1984).
Как используется в данном описании, «в основном комплементарные» означает, что две последовательности нуклеиновой кислоты обладают по меньшей мере приблизительно 65%, предпочтительно приблизительно 70%, более предпочтительно приблизительно 80%, даже более предпочтительно 90% и наиболее предпочтительно приблизительно 98% комплементарностью последовательностей друг с другом. Это означает, что праймеры и зонды должны обладать достаточной комплементарностью с их матрицей и нуклеиновой кислотой-мишенью, соответственно, чтобы гибридизоваться в строгих условиях. Таким образом, последовательности праймера и зонда не должны отражать точную комплементарную последовательность связывающей области на матрице, и можно применять вырожденные праймеры. Например, некомплементарный нуклеотидный фрагмент можно присоединять к 5'-концу праймера, причем остальная последовательность праймера является комплементарной цепью. Альтернативно, в праймер можно вставлять некомплементарные основания или более длинные последовательности при условии, что праймер обладает достаточной комплементарностью с последовательностью одной из подлежащих амплификации цепей для гибридизации с ней и, таким образом, формирования структуры дуплекса, которую можно достроить способами полимеризации. Некомплементарные нуклеотидные последовательности праймеров могут включать участки для рестрикционных ферментов. Добавление участка для рестрикционного фермента на конец(концы) последовательности-мишени может быть особенно полезным для клонирования последовательности-мишени. В основном комплементарная последовательность праймера представляет собой последовательность, обладающую достаточной комплементарностью последовательности с матрицей для амплификации, чтобы приводить к связыванию праймера и синтезу второй цепи. Специалистам в данной области известно, что праймеры должны обладать достаточной комплементарностью последовательности с матрицей для амплификации.
Термин «гибрид» в контексте нуклеиновых кислот относится к двухцепочечной молекуле нуклеиновой кислоты, или дуплексу, образованной с помощью водородных связей между комплементарными нуклеотидными основаниями. Термины «гибридизоваться» или «отжигаться» относятся к процессу, при помощи которого отдельные цепи последовательности нуклеиновой кислоты образуют двуспиральные отрезки путем водородного связывания между комплементарными основаниями.
Термин «гибрид» в контексте селекции растений относится к растению, являющемуся потомком генетически разнородных родителей, полученным путем скрещивания растений из различных линий, пород или видов.
Термин «зонд» относится к одноцепочечной олигонуклеотидной последовательности, которая будет распознавать комплементарную последовательность в последовательности-мишени аналита нуклеиновой кислоты или производной от нее кДНК и формировать с ней дуплекс с водородными связями.
Термин «праймер», как используется в данном описании, относится к олигонуклеотиду, способному отжигаться с мишенью для амплификации, позволяя присоединение ДНК-полимеразы, таким образом, выполняя функцию точки инициации синтеза ДНК при помещении в условия, в которых индуцируется синтез продукта удлинения праймеров, т.е. в присутствии нуклеотидов и средства для полимеризации, такого как ДНК-полимераза, и при подходящих температуре и pH. Праймер (для амплификации) предпочтительно является одноцепочечным для максимальной эффективности амплификации. Предпочтительно, праймер представляет собой олигодезоксирибонуклеотид. Праймер должен быть достаточно длинным для праймирования синтеза продуктов удлинения в присутствии средства для полимеризации. Точные длины праймеров будут зависеть от многих факторов, включая температуру и состав (содержание A/T и G/C) праймера. Пара двусторонних праймеров состоит из одного прямого и одного обратного праймера, как общепринято в области амплификации ДНК, такой как амплификация PCR.
Понятно, что «праймер», как используется в данном описании, может относиться к более чем одному праймеру, в частности, в случае, где существует некоторая неопределенность информации относительно концевой последовательности(последовательностей) области-мишени, подлежащей амплификации. Таким образом, «праймер» включает набор олигонуклеотидов-праймеров, содержащий последовательности, представляющие возможные варианты последовательности, или включает нуклеотиды, позволяющие типичное спаривание оснований.
Олигонуклеотидные праймеры можно получать любым подходящим способом. Способы получения олигонуклеотидов конкретной последовательности известны в данной области и включают в себя, например, клонирование и рестрикцию подходящих последовательностей, и прямой химический синтез. Способы химического синтеза могут включать, например, фосфо ди- или триэфирный способ, диэтилфосфорамидатный способ и твердофазный способ, описанные, например, в патенте США № 4458066. Праймеры, если желательно, можно метить путем введения средств, способных к выявлению, например, спектроскопическими, флуоресцентными, фотохимическими, биохимическими, иммунохимическими или химическими способами.
Зависимое от матрицы удлинение олигонуклеотидного праймера(праймеров) катализирует полимеризующее средство в присутствии подходящих количеств четырех дезоксирибонуклеотидтрифосфатов (dATP, dGTP, dCTP и dTTP, т.е. dNTP) или аналогов, в реакционной среде, состоящей из подходящих солей, катионов металлов, и системы для забуферивания pH. Подходящими полимеризующими средствами являются ферменты, известные как катализирующие зависимый от праймера и матрицы синтез ДНК. Известные ДНК-полимеразы включают, например, ДНК-полимеразу I E.coli или ее фрагмент Кленова, ДНК-полимеразу T4 и ДНК-полимеразу Taq. Условия реакции для катализа синтеза ДНК этими ДНК-полимеразами известны в данной области.
Продукты синтеза представляют собой молекулы дуплекса, состоящего из цепей матрицы и цепей после удлинения праймера, содержащих последовательность-мишень. Эти продукты, в свою очередь, служат матрицей для другого цикла репликации. Во втором цикле репликации цепь после удлинения праймера из первого цикла отжигается с комплементарным ей праймером; в результате синтеза получают «короткий» продукт, который как на 5'-, так и на 3'-конце ограничен последовательностями праймеров или комплементарными им. Повторяющиеся циклы денатурации, отжига праймеров и достройки приводят к экспоненциальному накоплению области-мишени, определяемой праймерами. Проводят достаточно циклов для достижения желаемого количества полинуклеотида, содержащего область-мишень нуклеиновой кислоты. Желаемое количество может меняться и определяется назначением, для которого должен служить полинуклеотидный продукт.
Способ PCR хорошо описан в руководствах и известен специалистам в данной области.
После амплификации PCR полинуклеотиды можно выявлять гибридизацией с полинуклеотидом-зондом, образующим стабильный гибрид с последовательностью-мишенью в условиях гибридизации и отмывки от строгих до умеренно строгих. Если ожидают, что зонды будут по существу полностью комплементарными (т.е. приблизительно на 99% или более) последовательности-мишени, то применяют строгие условия. Если ожидают некоторые несоответствия, например ожидают, что варианты штаммов будут приводить к неполной комплементарности зондов, строгость гибридизации можно уменьшать. Однако выбирают условия, исключающие неспецифическое/случайное связывание. Условия, влияющие на гибридизацию, которые выбирают против неспецифического связывания, известны в данной области и описаны, например, в Sambrook et al., (2001). Как правило, более низкая концентрация соли и более высокая температура увеличивают строгость связывания. Например, обычно считают, что строгие условия представляют собой инкубации в растворах, содержащих приблизительно 0,1×SSC, 0,1% SDS, при температуре инкубации/отмывки приблизительно 65°C, а умеренно строгие условия представляют собой инкубации в растворах, содержащих приблизительно 1-2×SSC, 0,1% SDS, при температуре инкубации/отмывки приблизительно 50°-65°C. Условия низкой строгости представляют собой 2×SSC и приблизительно 30-50°C.
Термины «строгость» или «строгие условия гибридизации» относятся к условиям гибридизации, влияющим на стабильность гибридов, например температуре, концентрации соли, pH, концентрации формамида и т.п. Эти условия эмпирически оптимизируют для максимизации специфического связывания и минимизации неспецифического связывания праймера или зонда с последовательностью-мишенью нуклеиновой кислоты. Термины, как используется, включают в себя обозначение условий, при которых зонд или праймер будет гибридизоваться с его последовательностью-мишенью в большей степени, чем с другими последовательностями, что можно выявить (например, по меньшей мере в 2 раза выше фона). Строгость условий зависит от последовательности и будет различаться в различных обстоятельствах. Более длинные последовательности специфически гибридизуются при более высоких температурах. Как правило, выбирают условия строгости приблизительно на 5°C ниже, чем температурная точка плавления (Tm) для конкретной последовательности при определенной ионной силе и pH. Tm представляет собой температуру (при определенных ионной силе и pH), при которой 50% комплементарной последовательности-мишени гибридизуется с точно совпадающим зондом или праймером. Как правило, строгими условиями являются такие, при которых концентрация соли составляет менее чем приблизительно 1,0 M иона Na+, обычно, концентрация иона Na+ (или других солей) составляет приблизительно 0,01-1,0 M при pH 7,0-8,3, а температура составляет по меньшей мере приблизительно 30°C для коротких зондов или праймеров (например, 10-50 нуклеотидов) и по меньшей мере приблизительно 60°C для длинных зондов или праймеров (например, более чем 50 нуклеотидов). Строгих условий можно достигать также добавлением дестабилизирующих средств, таких как формамид. Характерные условия с низкой строгостью или «условия пониженной строгости» включают в себя гибридизацию в буферном растворе 30% формамида, 1M NaCl, 1% SDS при 37°C и отмывку в 2×SSC при 40°C. Характерные условия с высокой строгостью включают в себя гибридизацию в 50% формамиде, 1M NaCl, 1% SDS при 37°C и отмывку в 0,1×SSC при 60°C. Способы гибридизации хорошо известны в данной области и описаны, например, в Ausubel et al., 1998 и Sambrook et al., 2001.
Термин «антиген» относится к веществу, способному запускать иммунный ответ у позвоночного, приводящий к продукции антитела в качестве части защиты против указанного вещества. Антигены могут представлять собой белки вируса, которые могут провоцировать продукцию антитела, например, в клетках крови, клетках лимфатических узлов и селезенке позвоночных.
Термин «антитело» включает антигенсвязывающие пептиды и относится к антителам, моноклональным антителам, полноразмерному иммуноглобулину или антителу, или любому функциональному фрагменту молекулы иммуноглобулина. Примеры таких пептидов включают полные молекулы антитела, фрагменты антител, такие как Fab, F(ab')2, определяющие комплементарность области (CDR), VL (вариабельную область легкой цепи), VH (вариабельную область тяжелой цепи) и любое их сочетание или любую другую функциональную часть пептида антитела. Термин «антитело» относится к полипептиду, в основном кодируемому геном иммуноглобулина или генами иммуноглобулинов, или его фрагментам, которые специфически связывают и узнают аналит (антиген). Однако в то время как различные фрагменты антител можно определить исходя из расщепления интактного антитела, специалист в данной области будет принимать во внимание, что такие фрагменты можно синтезировать de novo, либо химически, либо с использованием способов рекомбинантной ДНК. Таким образом, термин антитело, как используется в данном описании, включает в себя также фрагменты антител, такие как одноцепочечный Fv, химерные антитела (т.е. содержащие константные и вариабельные области из различных видов), гуманизированные антитела (т.е. содержащие определяющую комплементарность область (CDR) из не относящегося к человеку источника) и гетероконъюгированные антитела (например, биспецифические антитела).
Термины «в основном чистый» и «выделенный» используются взаимозаменяемо и описывают белок, пептид или нуклеиновую кислоту, которые являются в основном отделенными от других (суб)клеточных компонентов, сопровождающих их в природе. Термин охватывает нуклеиновую кислоту или белок, удаленные из их природного окружения, и включает в себя рекомбинантные или клонированные изоляты ДНК и химически синтезированные аналоги, или аналоги, биологически синтезированные в гетерологичных системах. Как правило, термин относится к очищенным белку и нуклеиновой кислоте, обладающим чистотой по меньшей мере приблизительно 75%, например, 85%, 95% или 98% по массе. Незначительные изменения или химически модификации обычно разделяют одну и ту же полипептидную или нуклеотидную последовательность. В основном чистый белок или нуклеиновая кислота обычно будут содержать приблизительно 85-100% (масс./масс.) образца белка или нуклеиновой кислоты, более обычно, приблизительно 95%, и предпочтительно будут обладать более чем приблизительно 99% чистотой. Чистоту или гомогенность белка или нуклеиновой кислоты можно показать рядом способов, хорошо известных в данной области, таких как электрофорез образца белка в полиакриламидном геле с последующей визуализацией отдельной полосы полипептида после окрашивания или электрофорез образца нуклеиновой кислоты в агарозном геле с последующей визуализацией отдельной полосы полинуклеотида в агарозном геле после окрашивания. «Окрашивание» либо может обозначать применение неспецифических для пептида или нуклеиновой кислоты красителей, таких как красители с серебром и Кумасси или красители бромид этидия и SYBR®, либо может обозначать применение специфического для пептида или нуклеиновой кислоты окрашивания, такого как контактирование пептида с антителом и визуализация антитела с использованием меченого вторичного антитела (например, конъюгированного со щелочной фосфатазой) в случае белков или пептидов, или контактирование нуклеиновой кислоты с комплементарным зондом, меченым для визуализации наличия гибридизации между нуклеиновой кислотой и зондом. Для конкретных целей можно обеспечить более высокое разрешение с использованием для очистки высокоэффективной жидкостной хроматографии (ВЭЖХ) или подобных способов. Такие способы находятся в области общеизвестных общих знаний (см., например, Katz, et al., 1998).
Идентификация и таксономия
Изобретение относится к выделенному вирусу, содержащему по меньшей мере одну последовательность нуклеиновой кислоты согласно SEQ ID NO:1 и/или SEQ ID NO:2 и последовательности, обладающие по меньшей мере 30%, 35%, 40%, 45%, 50%, 55%, 60% или предпочтительно 65% гомологией нуклеотидной последовательности с ними. Как указано выше, полинуклеотиды обладают «гомологичными» последовательностями, если последовательность нуклеотидов в двух последовательностях является одинаковой при выравнивании для максимального соответствия. Поисками BLAST с использованием нуклеотидных последовательностей, полученных из изолята вируса ToTV, не выявили значительной гомологии с какой-либо из известных вирусных или невирусных последовательностей в базах данных GenBank и EMBL. Наибольший процент гомологии, обнаруженный таким образом, составлял 46,5% между мотивами геликазы в ORF1 SEQ ID NO:2, соответствующими RNA1 ToTV, с мотивами геликазы других вирусов растений (см. примеры ниже).
Следует понимать, что гомология может быть большой, когда две последовательности сравнивают в небольшом окне сравнения, поскольку часто можно найти локальные области сходства последовательности при сравнении двух длинных нуклеотидных последовательностей. Однако специалист в данной области сознает, что гомология последовательности требует установления общих мотивов между последовательностями, среди которых идентичность последовательности локально может быть настолько высокой, как от 35 до 100%, однако может быть настолько низкой, как 10-20% в других частях геномной последовательности из той же самой ORF. Таким образом, когда в данном описании указано, что последовательность обладает гомологией нуклеотидной последовательности по меньшей мере 50% с SEQ ID NO:1 или SEQ ID NO:2, это может относиться к гомологии последовательности между областями среди общих мотивов, где гомология является наибольшей, а также между кодирующей последовательностью двух гомологичных белков. Необходимо отметить, что гомология последовательности может различаться между различными генами в геноме (см. примеры).
В качестве выявления родства между вновь идентифицированным изолятом вируса ToTV и другими вирусами (включая вирусы, подлежащие сравнению с ним) можно обычно провести филогенетический анализ на основе (части) информации о геномной последовательности вирусов.
Несколько таких анализов представлены в примерах ниже. Информация, поученная таким образом, далее указывает на то, что ToTV обладает наивысшим уровнем гомологии с вирусами из родов Sequivirus и Waikavirus (Sequiviridae) и родов Cheravirus и Sadwavirus. Вирусы из этих родов различают на основании числа вирусных РНК - Sequiviridae обладает 1 РНК, в то время как Cheravirus и Sadwavirus обладают двумя РНК; и числом белков капсида - Sadwavirus обладает двумя CP, Cheravirus - тремя CP. Эти критерии позволяют предполагать, что вирус томата торрадо наиболее вероятно входит в группу рода Cheravirus. Однако филогенетические анализы с использованием нескольких различных мотивов из предположительных белков RdRp и геликазы располагают ToTV четко отдельно от родов Cheravirus и Sadwavirus и фактически позволяют предполагать, что он является более близко родственным с Sequiviridae. К сожалению, в настоящее время доступны полные последовательности только одного Cheravirus (CRLV) и двух Sadwavirus (SDV и SMoV). Предварительные данные о переносчике позволяют предполагать, что ToTV могут переносить белокрылки, в то время как природные переносчики CRLV и Sadwavirus неизвестны. На основании доступной в настоящее время информации, включая информацию о последовательности, так же как дополнительную таксономическую информацию, как представлено в таблице 1 ниже, вновь обнаруженный вирус, наиболее вероятно, представляет собой новый род.
Таблица 1: Таксономические описания для недавно обнаруженного вируса ToTV (как перечислено в международно принятых способах из руководства "Matthew's Plant Virology" (“Matthew's Plant Virology”, Fourth Edition, Roger Hull (ed.) Academic Press, San Diego, p 15, таблица 2.1) (Нумерацию из таблицы в Matthews соблюдают в таблице 1 ниже).
Авторы настоящего изобретения недавно обнаружили, что геномные последовательности вируса, связанного с заболеванием томата в Центральной Америке (например, в Мексике и Гватемале) под названием «Заболевание шоколадной пятнистости», «Шоколад» или «Марчитец», являются идентичными последовательностям описанного здесь ToTV (см. пример 2). Таким образом, возбудитель заболевания, связанный с данным заболеванием, является аспектом настоящего изобретения.
По мере того как больше вирусных последовательностей станут доступными как из не относящихся к ToTV вирусов, которые могут быть или могут не быть близкородственными с ним, так и из вирусов, близко родственных с вирусом ToTV или существенно напоминающих его, филогенетический анализ будет служить полезным способом определения объема таксона или клада ToTV на основе филогенетического родства между изолятами.
Филогенетическое родство можно определить, например, на основании любой или всех нуклеотидных последовательностей RNA 1 и/или РНК 2 из вирусного генома или по данным последовательности (гена) белка капсида. Филогенетические анализы хорошо известны специалисту в данной области и могут, например, включать в себя анализ путем реконструкции деревьев на основе расстояния (например, от соседних узлов), способами максимального правдоподобия или экономии с использованием таких программ, как ClustalX (Thompson et al., 1997), PAUP (Swofford, 2000) или PHYLIP (Felsenstein, 1989).
Чтобы провести анализ филогенетического родства между новым изолятом, последовательностью ToTV, как представлено в данном описании, и контрольными последовательностями из вирусных штаммов, например, из баз данных GenBank, EMBL или DDBJ, геномную РНК из указанного нового изолята можно выделить непосредственно из инфицированных растений, и геномные последовательности можно амплифицировать с нее. Способы амплификации PCR с обратной транскрипцией (RT-PCR) можно выполнять, например, с использованием вырожденных олигонуклеотидных праймеров, где такие праймеры, например, способны действовать как праймер для амплификации последовательностей нуклеиновой кислоты из геномов дивергентных изолятов ToTV, так же как из геномов близкородственных видов вирусов. Предпочтительно, но не обязательно, таким образом можно получить продукты амплификации полноразмерного генома из контрольных штаммов (например, дивергентных изолятов), тестируемых цепей (предположительных изолятов ToTV) и близкородственных видов. Предпочтительно, для сравнения амплифицируют конкретные интересующие генетические области. Продукты амплификации (ДНК) затем можно секвенировать, например, прямым секвенированием двухцепочечных нуклеотидов с использованием флуоресцентно меченных останавливающих дидезоксинуклеотидов (Smith et al., 1986) с помощью вырожденных олигонуклеотидных праймеров, используемых для RT-PCR. Редактирование, анализ, необязательное предсказание аминокислотной последовательности и выравнивание нуклеотидной последовательности можно проводить с помощью доступных пакетов программ, таких как пакет для анализа последовательности LaserGene версии 5 (DNASTAR, Inc., Madison, Wis.) и программное обеспечение IntelliGenetics Gene Works версии 2.5.1 (IntelliGenetics, Mountain View, Calif.). Затем филогенетические анализы можно дополнить филогенетическим анализом с использованием, например, программного обеспечения для поиска максимальной экономии (PAUP) с помощью алгоритма соседних узлов с использованием абсолютных расстояний с последующим эвристическим поиском и 1000 независимыми повторами, и можно получить филогенетическое дерево анализом максимальной экономии выравниваемых непрерывных нуклеотидных или аминокислотных последовательностей, при котором в таких деревьях номера, как правило, представляют собой уровни достоверности после повторов. После 1000 повторов уровень достоверности выше 60%, предпочтительно выше 70%, более предпочтительно выше 80%, 90%, 95% или 98% в филогенетическом дереве считают достаточным доказательством правильного филогенетического заключения (помещения изолятов в конкретный клад), при условии, что дерево является достаточно разветвленным, в силу чего, не обязательно, разветвление можно улучшить образованием корня с подходящими видами вне групп. Таким образом, можно определить, какие изоляты являются наиболее близкородственными с последовательностями ToTV, как представлено в данном описании. Родство, как правило, выражают в переводе на процент сходства последовательностей, обозначенный здесь гомологией последовательности.
Хотя филогенетические анализы предоставляют подходящий способ идентификации вируса в случае достаточной гомологии нуклеиновых кислот, для известных вирусов в данном описании также представлено несколько других, возможно, более прямых, хотя несколько более грубых способов для идентификации указанного вируса или вирусных белков, или нуклеиновых кислот из указанного вируса. Как эмпирическое правило, вирус ToTV можно идентифицировать по процентам гомологии подлежащих идентификации вирусных белков или нуклеиновых кислот по сравнению с вирусными белками или нуклеиновыми кислотами, идентифицированными в соответствии с изобретением по последовательности. Общеизвестно, что виды вирусов, особенно виды РНК-вирусов, часто состоят из псевдовидов, где для кластеров указанных вирусов показана гетерогенность среди их членов. Таким образом, ожидают, что каждый изолят может обладать до некоторой степени различным процентом гомологии с последовательностями изолята, представленного в данном описании. Таким образом, другие изоляты вируса, обладающие достаточной гомологией последовательности с ToTV (например, более чем 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95%, 98% или 99% гомологией последовательности), считают принадлежащими к тому же самому вирусу. Таким образом, вирус ToTV по настоящему изобретению представляет собой вирус, обладающий по меньшей мере 50 или 60% гомологией, предпочтительно по меньшей мере 70%, более предпочтительно по меньшей мере 80%, еще более предпочтительно по меньшей мере 90%, еще более предпочтительно по меньшей мере 95%, еще более предпочтительно по меньшей мере 98% и наиболее предпочтительно по меньшей мере 99% гомологией с последовательностями белка или нуклеиновой кислоты, представленными в данном описании, и вызывающий симптомы индуцированного ToTV заболевания у Solanaceae, более конкретно, у Solanum и Nicotiana, где симптомы заболевания могут быть или могут не быть подобными симптомам, описанным в данном документе. У Cucurbitaceae вирус, по-видимому, не вызывает видимых симптомов, хотя вирус способен к размножению в растениях.
Если желательно сравнить выделенный изолят вируса с последовательностями белка или нуклеиновой кислоты, описанными в данном документе, изобретение предоставляет выделенный вирус (ToTV), который можно идентифицировать как филогенетически соответствующий ToTV путем определения последовательности белка или нуклеиновой кислоты указанного выделенного изолята вируса и определения того, что последовательность указанного белка или нуклеиновой кислоты обладает процентной гомологией последовательности по меньшей мере 60%, предпочтительно по меньшей мере, 70%, более предпочтительно по меньшей мере 80%, еще более предпочтительно по меньшей мере 90%, еще более предпочтительно по меньшей мере 95%, еще более предпочтительно по меньшей мере 98% и наиболее предпочтительно по меньшей мере 99% с последовательностями, перечисленными в данном описании.
Изоляты вируса, имеющие отдельные белки или нуклеиновые кислоты с более высокой гомологией, чем эти указанные максимальные значения, считают филогенетически соответствующими и, таким образом, таксономически соответствующими вирусу ToTV, и как правило, белки будут кодировать последовательность нуклеиновой кислоты, структурно соответствующую последовательности, указанной в данном описании. При этом изобретение относится к вирусу, филогенетически соответствующему выделенному вирусу, последовательности которого перечислены в данном описании.
Следует заметить, что, подобно другим вирусам, можно ожидать, что будет обнаружена определенная степень изменчивости между вирусами ToTV, выделенными из различных источников.
Кроме того, нуклеотидные или аминокислотные последовательности вируса ToTV или его фрагментов, как приведено в данном описании, проявляют, например, менее чем 95%, предпочтительно менее чем 90%, более предпочтительно менее чем 80%, более предпочтительно менее чем 70% и наиболее предпочтительно менее чем 60% гомологию нуклеотидной последовательности, или менее чем 95%, предпочтительно менее чем 90%, более предпочтительно менее чем 80%, более предпочтительно менее чем 70% и наиболее предпочтительно менее чем 60% гомологию аминокислотной последовательности с соответствующей нуклеотидной или аминокислотной последовательностью любого не относящегося к вирусу ToTV или ближайшего родственного ему.
Отклонения последовательности штаммов ToTV в мире могут быть до некоторой степени выше по аналогии с другими вирусами растений.
Изобретение относится к выделенному вирусу (ToTV), который можно идентифицировать как филогенетически соответствующий этому путем определения последовательности нуклеиновой кислоты подходящего фрагмента генома указанного вируса и тестирования в филогенетических анализах, где получают деревья с максимальным правдоподобием с использованием, например, 1000 повторов, как описано выше, и обнаружения того, что он является филогенетически более близко родственным с изолятом вируса, содержащим последовательности из SEQ ID NO:1 или SEQ ID NO:2, как перечислено в данном описании для ToTV, чем родственным с изолятом вируса из не относящегося к ToTV контрольного штамма или ближайшего родственного ему.
Подходящими фрагментами нуклеиновокислотного генома, каждый из которых можно применять для таких филогенетических анализов, является, например, любая часть последовательности нуклеиновой кислоты из фрагментов РНК 5,5 т.н. или 8 т.н. (соответственно обозначенных здесь РНК 2 и РНК 1), как описано в примере.
С обеспечением информации о последовательности данного вируса ToTV изобретение предоставляет диагностические средства и способы для использования при детекции вируса ToTV в образце. Предпочтительно, детекцию вируса ToTV проводят с помощью реагентов, которые являются наиболее специфическими для вируса ToTV. Это ни в коем случае, однако, не исключает возможности использовать вместо этого менее специфичные, но в меру перекрестно реактивные реагенты, например, потому что они являются более легко доступными и подходят для решаемой задачи.
Изобретение относится, например, к способу детекции присутствия ToTV в растениях, предпочтительно в растениях томата, более предпочтительно в растениях S.lycopersicum. Указанный способ может, например, включать определение в указанном образце присутствия вируса ToTV или его компонента с помощью реакции указанного образца со специфической для ToTV нуклеиновой кислотой или антителом по изобретению. Хотя контактная инфекция индикаторного растения также является подходящим способом для детекции присутствия вируса в тестируемом растении.
Изобретение относится к частичной нуклеотидной последовательности нового выделенного вируса (называемого здесь также вирус ToTV) и к специфическим для вируса ToTV компонентам или их синтетическим аналогам. Дополнительные геномные последовательности вируса ToTV, кроме представленных в данном описании, можно определить способами секвенирования, известными специалистам в данной области. Теперь, когда по настоящему изобретению предоставлен вирус ToTV, так же как его частичные геномные последовательности, определение геномной последовательности легко доступно специалисту в данной области. Эти способы включают в себя, например, способы, описанные в примере ниже. Как правило, такие способы секвенирования включают в себя выделение нуклеиновых кислот из вирусного генома способами выделения нуклеиновой кислоты и определения нуклеотидной последовательности выделенной нуклеиновой кислоты, например способами дидезокси-остановки цепи (Sanger et al., 1977), не обязательно предваряемыми обратной транскрипцией РНК в ДНК.
Изобретение, среди прочего, относится к выделенной или рекомбинантной нуклеиновой кислоте или ее специфическому для вируса функциональному фрагменту, которые можно получить из вируса по изобретению. Выделенные или рекомбинантные нуклеиновые кислоты включают в себя последовательности, перечисленные в данном описании, или последовательности гомологов, способные гибридизоваться с ними в строгих условиях. В частности, изобретение относится к праймерам и/или зондам, пригодным для идентификации нуклеиновой кислоты вируса ToTV. Дополнительные зонды и праймеры, способные гибридизоваться с последовательностью нуклеиновой кислоты вируса ToTV, можно разрабатывать способами, известными специалисту в данной области.
Экспрессирующие векторы и экспрессия кодирующих генов вируса
Кроме того, изобретение относится к экспрессирующему вектору, содержащему нуклеиновую кислоту по изобретению. Во-первых, экспрессирующие векторы, такие как плазмидные векторы, содержащие (части) двухцепочечной последовательности вирусного генома ToTV, вирусные векторы, содержащие (части) генома ToTV (например, без ограничения, вирус осповакцины, ретровирусы, бакуловирус), или вирус ToTV, содержащий (части) генома других вирусов или других патогенов, являются частью настоящего изобретения.
Экспрессирующий вектор может содержать геномную последовательность ToTV или ее часть, которая находится под контролем регуляторного элемента, такого как промотор, или является функционально связанной с ним. Фрагмент ДНК, обозначенный как промотор, является ответственным за регуляцию транскрипции ДНК в мРНК. Экспрессирующий вектор может содержать один или несколько промоторов, подходящих для экспрессии гена, предпочтительно для экспрессии гена, кодирующего вирусный белок, в клетках растений, клетках грибов, бактериальных клетках, клетках дрожжей, клетках насекомых или других эукариотических клетках. Экспрессирующие векторы по изобретению очень полезны для получения антигенов вируса в системах экспрессии гена.
Изобретение относится также к клетке-хозяину, содержащей нуклеиновую кислоту или экспрессирующий вектор по изобретению. Плазмидные или вирусные векторы, содержащие нуклеиновые кислоты, кодирующие белковые компоненты вируса ToTV, можно получить в прокариотических клетках для экспрессии компонентов в подходящих типах клеток (клетках растений, клетках грибов, бактериях, клетках насекомых, клетках растений или других эукариотических клетках). Плазмидные или вирусные векторы, содержащие полноразмерные или частичные копии генома вируса ToTV, можно получить в прокариотических клетках для экспрессии вирусных нуклеиновых кислот in vitro или in vivo.
Способы выделения и очистки ToTV
Вирус ToTV можно выделить из инфицированных растений или других источников любым доступным способом. Выделение может включать в себя очистку или частичную очистку вирусных частиц ToTV из подходящего источника. Для выделения и очистки вируса доступен широкий ряд способов (например, см. Dijkstra and De Jager, 1998). Очистку ToTV можно, например, проводить с использованием общепринятых способов, например, для неповирусов или лютеовирусов (с помощью органических растворителей) (см., например, Walker, 2004). Хотя такие способы могут приводить к потере инфекционности вируса, эти способы еще могут быть применимыми для получения вирусного материала для других целей.
Предпочтительно, чтобы поддерживать целостность вируса, для очистки ToTV используют способ мягкой очистки. Такой способ мягкой очистки может включать в себя, например, гомогенизацию инфицированного материала растения (такого как листья) в буфере для гомогенизации (содержащем, например, 0,1 M буфер Трис-HCl при pH приблизительно 8, приблизительно 20 мМ сульфит натрия [Na2SO3], приблизительно 10 мМ диэтилдитиокарбамат натрия [Na-DIECA] и приблизительно 5 мМ этилендиаминтетраацетат натрия (Na-EDTA)), отделение дебриса центрифугированием (например, в течение 30 мин при 49000 g), помещение супернатанта на подходящую подушку сахарозы (например, 20%) и осаждение вирусных частиц (например, центрифугированием в течение 1,5 часов при 70000 g). Затем осадок, содержащий вирусные частицы, можно ресуспендировать в подходящем буфере (например, в Трис-HCl, pH 8), и содержащую вирус фракцию можно отделить от остальной суспензии с использованием центрифугирования в градиенте плотности сахарозы (например, 10-40% сахароза в буфере для гомогенизации, центрифугирование в течение 2 час при 110000 g). Содержащую вирус фракцию можно определить с использованием экспериментов по инфекции. Дополнительную очистку можно проводить с использованием центрифугирования в градиенте плотности сульфата цезия (например, 10-40% градиент сульфата цезия в Трис-HCl, pH 8, центрифугирование в течение 16 час при 125000 g). Обогащенные вирусом полосы можно затем собирать из градиента и дополнительно концентрировать центрифугированием или диализовать (например, против 0,1 M Трис-HCl, pH 8).
Инфекционность ToTV после очистки можно проверить инокуляцией в чувствительное растение (например, N.hesperis '67A').
Способы очистки связанных с ToTV белков и аминокислотного секвенирования
После получения очищенного или частично очищенного вируса ToTV можно получить в основном чистый препарат связанных с вирусом белков (например, кодируемых вирусом белков). Хотя в данной области известны многочисленные способы и стратегии очистки белка, самой подходящей является очистка вирусных белков ToTV, таких как белки вирусной оболочки, либо электрофорезом с использованием, например, полиакриламидного геля с додецилсульфатом натрия (SDS-PAGE), либо аффинной хроматографией. Каждый из этих способов будет описан ниже.
Представляющие интерес белки ToTV можно разделять электрофорезом с использованием, например, трицин-SDS-PAGE (Schagger и Von Jagow, 1987) или глицин-SDS-PAGE (Laemmli, 1970). Можно, конечно, применять другие системы электрофореза, способные разделять различные белки, содержащиеся в изоляте вируса, или транскрибируемые с его генома и экспрессированные в подходящей экспрессирующей системе, такие как не денатурирующий гель-электрофорез. Область геля для PAGE, содержащую заданный белок, можно вырезать, и заданные полипептиды можно элюировать оттуда, например, с использованием устройства Elutrap® (Schleicher & Schuell, Dassel, Germany). Заданный белок можно идентифицировать по его подвижности относительно контрольных полипептидов в геле. Для увеличения степени очистки можно проводить электрофорез элюированного белка во втором геле для SDS-PAGE и элюировать его второй раз. Белок или пептид, содержащийся в вырезанном фрагменте геля, можно затем снова элюировать, и он является пригодным для использования при иммунизации или секвенировании белка.
Представляющие интерес белки ToTV можно очистить также аффинной хроматографией с использованием антитела (такого как моноклональное антитело), специфически связывающегося с белком ToTV. Антитело можно ковалентно присоединить к твердым подложкам, таким как целлюлозы, полистирол, полиакриламид, поперечно-сшитый декстран, гранулированная агароза или стекло с контролируемым размером пор, с использованием бифункциональных сшивающих средств, которые реагируют с функциональными группами на подложке и функциональными группами (т.е. реакционноспособными боковыми цепями аминокислот) в молекуле антитела. Такие способы легко доступны специалисту в данной области. Полученную несущую антитело твердую фазу приводят в контакт с очищенным или частично очищенным вирусом в восстанавливающих условиях с использованием pH, ионной силы, температуры и продолжительности обработки, позволяющих представляющему интерес белку связываться с иммобилизованным антителом. Вирус или белок элюируют с колонки пропусканием элюента, который диссоциирует водородные связи с подложкой. Обычно применяемыми элюентами являются буферы с конкретным pH или более чем приблизительно 2 M растворы NaCl.
Способы осуществления аффинной хроматографии с использованием антител, так же как другие способы иммуноаффинной очистки белков (таких как белки вирусного капсида), хорошо известны в данной области (см., например, Harlow and Lane, 1988).
По предоставленным здесь объяснениям специалист в данной области может выделить специфический для вируса ToTV белок, определить аминокислотную последовательность, например, N-концевой части указанного белка, сконструировать набор вырожденных зондов (ввиду вырожденности генетического кода) для гибридизации с ДНК, кодирующей область указанного белка, с использованием этих зондов для массива генов в геномной библиотеке, полученной из вируса, получить положительную гибридизацию и локализовать соответствующие гены. Затем специалист в данной области может идентифицировать структурную область гена и, не обязательно, области последовательности против хода транскрипции и по ходу транскрипции от нее. После этого специалист в данной области может установить правильную последовательность аминокислотных остатков, формирующих белок.
Получение антитела
Антитела, либо моноклональные, либо поликлональные, можно получить к очищенному или частично очищенному белку или фрагменту пептида вируса ToTV множеством способов, известных специалистам в данной области, включая инъекцию белка в качестве антигена животным, слияние гибридомы и рекомбинантные способы, включающие бактериальные или фаговые системы (см. Marks et al, 1992a; Marks et al., 1992b; Lowman et al., 1991; Lerner et al., 1992, где в каждой из этих ссылок описаны подходящие способы).
Антитела против вирусных частиц, белков или пептидов вируса можно получить иммунизацией подходящего хозяина-позвоночного, предпочтительно млекопитающего, например кроликов, коз, крыс, кур и мышей, частицами, белками или пептидами самостоятельно или в объединении с адъювантом. Обычно предусматривают две или более иммунизации, и кровь или селезенку собирают через несколько дней после последней инъекции. В случае поликлональной антисыворотки, иммуноглобулины можно осадить, выделить и (аффинно) очистить. В случае моноклональных антител, спленоциты обычно сливают с иммортализованным лимфоцитом, например миелоидной линией, в условиях для селекции гибридом. Затем гибридомы можно клонировать в условиях лимитирующих разведений и проводить скрининг их супернатантов по антителам, обладающим желаемой специфичностью. Способы продукции (моноклональных) антител и способы их получения и применения в различных способах хорошо известны в литературе (см., например патенты США №№ 4381292, 4451570 и 4618577; Harlow and Lane, 1988; Ausubel et al., 1998; Rose et al., 1997; Coligan et al., 1997). Обычно антитело, направленное против связанного с вирусом белка, будет обладать аффинностью связывания по меньшей мере 1×105-1×107 M-1.
Рекомбинантный белок, полученный из вируса ToTV, такой как может быть получен путем экспрессии кодирующей белок геномной последовательности вируса в подходящей экспрессирующей системе, является предпочтительным в качестве антигена. Однако можно использовать также очищенные белки. Антигены, подходящие для детекции антителами, включают в себя любой белок ToTV, который соединяется с любым специфическим к ToTV антителом млекопитающего, подвергавшегося воздействию вируса ToTV или инфицированного им. Предпочтительные антигены по изобретению включают в себя антигены, вызывающие иммунный ответ у млекопитающих, подвергавшихся воздействию ToTV, которые, таким образом, как правило, наиболее легко узнают антитела млекопитающих. В частности, предпочтительные антигены включают в себя белки капсида ToTV. Наиболее предпочтительными являются структурные белки из очищенного вируса.
Способы клонирования геномных последовательностей в экспрессирующие векторы и из них для манипуляции с геномными последовательностями и для экспрессии белка, кодируемого геномной последовательностью в гетерологичном хозяине, хорошо известны, и эти способы можно использовать для получения экспрессирующих векторов, клеток-хозяев и клонированных геномных последовательностей, кодирующих антигены, где последовательности экспрессированы в хозяине для получения антител для применения в диагностических анализах (см., например, Sambrook et al., 2001 и Ausubel, et al., 1998).
Для получения антигенов ToTV можно использовать множество экспрессирующих систем. Например, описано множество экспрессирующих векторов, пригодных для получения белков в E.coli, B.subtilis, дрожжах, клетках насекомых, клетках растений и клетках млекопитающих, каждый из которых можно использовать для получения антигена ToTV, подходящего для получения анти-ToTV антитела или его фрагмента. Разумеется, собственно ToTV также можно использовать в качестве экспрессирующего вектора для данной цели.
Одним из применений антител по изобретению является скрининг экспрессирующих библиотек кДНК для идентификации клонов, содержащих вставки кДНК, кодирующие представляющие интерес белки или структурно родственные, перекрестно иммунореактивные белки. Такие скрининги экспрессирующих библиотек кДНК хорошо известны в данной области (см., например, Young and Davis, 1983), ссылка на который приведена в данном контексте, так же как другие опубликованные источники. Другим применением этих антител является использование в аффинной хроматографии для очистки белков ToTV. Эти антитела также применимы для анализа инфекции ToTV.
Настоящее изобретение, таким образом, относится к ToTV-специфическому вирусному белку или его фрагменту, в дальнейшем в этом документе обозначенному белковой молекулой. Применимыми белковыми молекулами являются, например, производные от любой из геномных последовательностей или их фрагменты, которые можно получить из вируса по изобретению. Такие белковые молекулы или их антигенные фрагменты, как приведено в данном описании, применимы, например, в диагностических способах или наборах и в диагностических композициях. В частности, применимыми являются такие белковые молекулы, кодируемые фрагментами рекомбинантной нуклеиновой кислоты, которые идентифицированы как вызывающие образование специфических для вируса ToTV антител, либо in vivo (например, для получения диагностических антител), либо in vitro (например, с использованием технологии фагового дисплея или другого способа, применимого для получения синтетических антител или их частей).
Также в настоящем документе предусмотрены антитела, являются ли они природными поликлональными или моноклональными, или синтетическими антителами (например, полученными из (фаговой) библиотеки связывающимися молекулами), которые специфически реагируют с антигеном, содержащим белковую молекулу или ее специфический для вируса ToTV функциональный фрагмент, такой как белок капсида по изобретению.
Способы идентификации изолята вируса как вируса ToTV
Помимо детекции вируса ToTV, которая включает в себя диагностические способы, настоящее изобретение также относится к способам идентификации, т.е. подтверждения, что данный изолят является ToTV. Такие способы могут быть основаны на филогенетическом заключении, как описано выше, и определении уровня гомологии нуклеотидной или аминокислотной последовательности между неидентифицированным изолятом вируса и одним или несколькими контрольными штаммами подтвержденных вирусов ToTV и вирусов, не относящихся к ToTV. Такие способы могут, например, включать в себя секвенирование (части) генома изолята вируса или белка капсида и сравнение уровня гомологии данной последовательности с последовательностями, предоставленными здесь для ToTV. Изолят, обладающий более чем 50% гомологией последовательности с SEQ ID NO:1 или SEQ ID NO:2, как представлено в данном описании, считают таксономически соответствующим ToTV или принадлежащим к таксону вируса ToTV. Такой вирус является частью настоящего изобретения.
Чтобы идентифицировать вирус как вирус томата торрадо (ToTV), можно также использовать таксономические признаки, как представлено в таблице 1 выше, и путем сравнения определить, что новый изолят принадлежит к предположительному новому роду, типичным видом которого можно принять штамм ToTV, идентифицируемый по предоставленным здесь последовательностям нуклеиновой кислоты SEQ ID NO:1 и 2, и депонированный в немецкой коллекции микроорганизмов и клеточных культур DSM 24 ноября 2004 под номером для ссылки депозитов ToTV-E01 (DSM 16999). Таким образом, не является необходимым проводить сравнение последовательности, чтобы определить то, что вирус является ToTV. Предпочтительнее, способ идентификации вируса как вируса томата торрадо (ToTV) может включать в себя стадии определения наличия сочетания таксономических признаков, выбранных из группы, состоящей из:
a) морфологических признаков, таких как сферические частицы вириона без внешней оболочки диаметром приблизительно 28 нм;
b) признаков свойств генома, таких как свойство вируса обладать одноцепочечной линейной положительной смысловой РНК на основе двух фрагментов РНК, содержащих поли(A)-хвосты, которые кодируют полибелки 5,5 и 8 кДа, соответственно, и содержат кодирующие области или мотивы для 3 белков капсида, геликазы, протеазы, RdRP и предположительного двигательного белка, где фрагменты РНК, и/или полибелки, и/или мотивы обладают гомологией, на основании сравнения последовательности, в основном, как описано в данном документе, и
c) признаков биологических свойств, таких как продуцирование некротических повреждений и ожогоподобных симптомов у томата, обладание кругом хозяев, связью с переносчиком и/или географическим распределением в основном, как описано в данном документе, и наличие связи с заболеваниями растений томата, известными в пределах определенной местности под такими названиями, как Торрадо, Марчитец и/или шоколадная пятнистость.
Сочетанием таксономических признаков, приводящим к положительной идентификации изолята вируса как являющегося вирусом томата торрадо (ToTV), является такое сочетание, которое указывает на то, что изолят является более близко родственным (на основании многочисленных таксономических способов, хорошо известных специалисту в данной области) с ToTV, как описано в данном документе, чем с другими вирусами, и где указанный изолят предпочтительно вызывает у томата симптомы заболевания, типичные для торрадо, как описано в данном документе.
Таким образом, вирус, который, на основе многочисленных таксономических анализов таксономических признаков, в основном, определенных в таблице 1, является более близко родственным с вирусом, как определено в пункте 1 формулы изобретения в настоящем документе, чем с любым другим вирусом, известным ко времени подачи настоящей заявки, и который является вирусом, связанным с заболеванием, вызывающим некротические повреждения у томата, считают здесь являющимся ToTV и входящим в объем настоящего изобретения.
Таким образом, изобретение относится к изоляту вируса, который можно идентифицировать способом по изобретению как вирус растений, таксономически соответствующий вирусу, идентифицируемому как возможно принадлежащий к таксону вируса ToTV. В зависимости от филогенетического родства или своеобразия по отношению к другим вирусным таксонам таксон вируса ToTV может быть изолятом, видом, родом или даже семейством вируса.
Альтернативно, способы идентификации изолята вируса как вируса ToTV могут быть основаны на симптоматологии, т.е. идентификации вируса по связанным с ним симптомам заболевания.
Однако в предпочтительном варианте осуществления в качестве способа идентификации изолята вируса как вируса ToTV применяют антитела по изобретению при условии, что эффективно исключают перекрестную реактивность таких антител для родственных не относящихся к ToTV штаммам. Такие способы включают в себя стадию реакции указанного вирусного изолята или его компонента с антителом, как предусмотрено в данном описании. Реакцией в данном документе называют обеспечение возможности связывания антитело-антиген. Это можно осуществить, например, с использованием очищенного или неочищенного вируса ToTV или его частей (белков, пептидов). Предпочтительно, инфицированные клетки или культуры клеток используют для идентификации вирусных антигенов с применением любого подходящего иммунологического способа. Особенно полезными в этом отношении являются антитела, индуцированные против белков капсида вируса ToTV.
Другие предпочтительные способы идентификации изолята вируса как вируса ToTV включают в себя реакцию указанного изолята вируса или его компонента со специфическим для вируса полинуклеотидом по изобретению, где полинуклеотид способен гибридизоваться в строгих условиях с последовательностью нуклеиновой кислоты, выбранной из группы, состоящей из SEQ ID NO:1, SEQ ID NO:2, последовательностей, обладающих по меньшей мере 60% гомологией нуклеотидной последовательности с SEQ ID NO:1 или SEQ ID NO:2, комплементарных им цепей и их ToTV-специфических фрагментов. Такую реакцию гибридизации можно проводить в любом формате, доступном специалисту в данной области, и она, как правило, включает в себя получение отпечатков тканей, способы дот-блоттинга, Саузерн/Нозерн-блоттинг или гибридизацию, гибридизацию in situ, PCR, RT-PCR и т.п.
Иммунологические способы детекции
Способы по изобретению, которыми выявляют антигены, можно, в принципе, осуществлять с использованием любого иммунологического способа, такого как, например, классическая иммунофлуоресценция (IF), иммуногистохимические способы или сравнимые форматы анализа для детекции антигена. Предпочтительные способы детекции ToTV, основанные на детекции белка вирусной оболочки, могут, например, включать в себя такие способы, как тесты преципитации и агглютинации, радиоиммуноанализ (RIA), иммунологический метод мечения золотом, иммуносорбционную электронную микроскопию (ISEM), твердофазный иммуноферментный анализ (ELISA), вестерн-блоттинг и иммуноблоттинг. Примерами типов иммунологических анализов, в которых можно использовать антитела по изобретению, являются конкурентный и не конкурентный иммунологические анализы либо в прямом, либо в непрямом формате. Антитела можно использовать в иммунологических анализах в жидкой фазе или связанными с твердофазным носителем. Кроме того, антитела в данных иммунологических анализах можно различными способами метить детектируемой меткой. Специалистам в данной области известны или легко понятны без излишних экспериментов подходящие форматы иммунологических анализов. Форматы анализов хорошо известны в литературе и описаны, например, у Harlow and Lane (1988).
Множество форматов иммунологического анализа можно использовать для отбора антител, специфически реагирующих с конкретным полипептидом по изобретению, таким как белки оболочки вируса ToTV. Например, для данной цели обычно используют твердофазные иммунологические анализы ELISA. См. описание форматов иммунологического анализа и условия, которые можно использовать для определения избирательного связывания, у Harlow and Lane (1988).
Антитела можно связывать со многими различными носителями и использовать для детекции присутствия молекул-мишеней. Альтернативно, антигены можно связывать со многими различными носителями и использовать для детекции присутствия антитела. Примеры хорошо известных носителей включают в себя стекло, полистирол, полипропилен, полиэтилен, декстран, нейлон, амилазы, природные и модифицированные целлюлозы, полиакриламиды, агарозы и магнетит. Для целей настоящего изобретения носитель по своей природе может быть либо растворимым, либо нерастворимым. Специалистам в данной области известны другие подходящие носители для связывания антител, или они способны определить такие носители с использованием общепринятых экспериментов.
Анализы вестерн-блоттингом в общих чертах описаны у Harlow and Lane (1988). По данному способу вирусные белки (и другие белки в препарате вируса) разделяют гель-электрофорезом и переносят на твердую фазу (т.е. мембрану, такую как нитроцеллюлоза). Последовательно проводят реакцию иммобилизованного антигена с антителом и системой детекции (например, конъюгированным со щелочной фосфатазой вторичным антителом). Как будет очевидно специалисту в данной области, предпочтительным будет включение в анализ соответствующих материалов для отрицательного и положительного контроля (таких как в основном очищенный антиген или вирус ToTV).
Твердофазные иммуноферментные анализы (ELISA) в общих чертах описаны у Harlow and Lane (1988). Анализ включает в себя реакцию вирусного компонента (например, белка капсида) с антителом. В одном из вариантов осуществления образец может содержать ткань растения, которую перемалывают и проводят ее реакцию с антителом, которым покрыта твердая фаза, такая как планшет для тестирования. Если в образце присутствует вирус, меченное ферментом специфическое антитело будет связываться с комплексом антитело-вирус, и это можно выявлять реакцией фермента с субстратом с получением цветной реакции. Предпочтительными способами анализов ELISA являются ELISA с прямым сэндвичем с двойным антителом (DAS) (Clark and Adams, 1977), непрямой ELISA с DAS (Vela et al 1986) или TAS-ELISA. Специалистам в данной области будет очевидно, что иногда в ELISA или любом другом типе анализа будет желательным определение присутствия более одного белка вирусного капсида в одной реакционной смеси, например, путем смешивания двух или более антител с различными специфичностями и анализа любого или всех вместе связанных с защитным капсидом белков по изобретению.
Способы детекции на основе нуклеиновой кислоты
ToTV состоит по меньшей мере из двух рибонуклеиновых кислот (РНК). Существуют серьезные указания на присутствие трех белков защитного капсида. Способы, описанные выше, направлены на иммунологическую детекцию вирусных белков для детекции вируса. Способом рекомбинантной ДНК можно получать зонды, напрямую или опосредованно гибридизующиеся с вирусными РНК (или комплементарными им) или с кДНК, полученными с них обратной транскрипцией, которые можно использовать в анализах для детекции вируса. Способы амплификации нуклеиновой кислоты позволяют амплификацию фрагментов вирусных нуклеиновых кислот, которые могут присутствовать в очень малых количествах.
Чтобы разработать способы детекции на основе нуклеиновой кислоты, необходимо определить специфические для вируса последовательности, для которых можно затем разработать праймеры или зонды. Для детекции ToTV посредством амплификации и/или гибридизации с зондом нуклеиновой кислоты белок капсида ToTV можно секвенировать или, альтернативно, можно выделить вирусную геномную РНК из очищенного вируса, провести обратную транскрипцию в кДНК и напрямую клонировать и/или секвенировать. Либо с использованием клонированной нуклеиновой кислоты в качестве зонда для гибридизации, либо с использованием информации о последовательности, полученной из клона, или путем конструирования вырожденных праймеров на основании аминокислотной последовательности белка ToTV можно сконструировать зонды для гибридизации нуклеиновой кислоты и/или праймеры для амплификации нуклеиновой кислоты и использовать в анализе детекции для детекции присутствия вируса в образце, как определено в данном описании.
Способы по изобретению, которыми выявляют нуклеиновые кислоты, можно, в принципе, осуществлять с использованием любого способа амплификации нуклеиновой кислоты, такого как полимеразная цепная реакция (PCR; Mullis and Faloona, 1987; патенты США №№ 4683195; 4683202 и 4800159), или с использованием таких реакций амплификации, как лигазная цепная реакция (LCR; Barany, 1991; EP 0320308), самоподдерживающаяся репликация последовательности (3SR; Guatelli et al., 1990), амплификация с замещением цепей (SDA; Walker et al., 1992; патенты США №№ 5270184 и 5455166), система транскрипционной амплификации (TAS; Kwoh et al., 1989), с использованием Q-бета репликазы (Lizardi et al., 1988), амплификации по типу катящегося кольца (RCA; патент США № 5871921), амплификации на основе последовательности нуклеиновой кислоты (NASBA; Compton, 1991), полиморфизма длины фрагментов расщепления клевазой (патент США № 5719028), изотермической и инициируемой химерным праймером амплификации нуклеиновой кислоты (ICAN), способа амплификации с разветвленным удлинением (RAM; патенты США №№ 5719028 и 5942391) или других подходящих способов амплификации нуклеиновых кислот.
Поскольку вирус является РНК-вирусом (т.е. последовательности на Фиг.5 и 6 являются ДНК-эквивалентами вирусного РНК-генома), подходящий способ детекции может включать в себя выделение вирусных нуклеиновых кислот из образца, например из инфицированного растения, с использованием способов, известных по существу специалисту в данной области (например, Chomczynski and Sacchi, 1987; Boom et al., 1990) или коммерчески доступных систем (например, набор для выделения общей РНК RNeasy или набор для выделения РНК растений RNeasy plant от QIAGEN GmbH, Hilden, Germany, или High-Pure-RNA-Isolation-Kit® (Roche Diagnostics, отделение F. Hoffmann-La Roche Ltd, Basel, Switzerland).
Тотальную РНК можно, например, выделить из материала листа или протопластов клеток растений, и тотальную РНК, или конкретно вирусную геномную РНК, или ее часть, можно затем подвергать обратной транскрипции в кДНК с использованием, например, обратной транскриптазы вируса миелобластоза птиц (AMV) или обратной транскриптазы вируса лейкемии мышей Молони (M-MuLV). Подходящий способ может, например, включать в себя смешивание в подходящей водной буферной системе (например, в коммерчески доступном буфере для RT) подходящего количества тотальных РНК (например, 1-5 мкг), подходящего количества (например, 10 пмоль) праймера для обратной транскрипции, подходящего количества dNTP и обратной транскриптазы, денатурацию нуклеиновых кислот кипячением в течение 1 минуты и охлаждением их на льду с последующей обратной транскрипцией, например, при 45°C в течение 1 часа, как рекомендовано для конкретной используемой обратной транскриптазы, для получения кДНК-копий вирусных последовательностей.
В качестве праймера для обратной транскрипции можно использовать полинуклеотид по настоящему изобретению, например 18-26-мерный олигонуклеотид, содержащий нуклеотидную последовательность, комплементарную геномной последовательности ToTV, или предпочтительно, по меньшей мере способную гибридизоваться в строгих условиях с последовательностью нуклеиновой кислоты SEQ ID NO:1 или SEQ ID NO:2, или ее ToTV-специфический фрагмент. Альтернативно, можно использовать неспецифический полиT-праймер (олигоdT-праймер), чтобы начинать обратную транскрипцию с мотивов поли-A РНК.
После RT-стадии полученную кДНК можно амплифицировать с помощью PCR, используя, например, ДНК-полимеразы Pfu и Taq и праймеры для амплификации, специфические для последовательностей вирусной геномной кДНК. Для RT-PCR можно использовать также полные коммерчески доступные системы (например, системы для RT-PCR Access и AccessQuick™ от Promega [Madison WI, USA], или система для RT-PCR в одной пробирке Titan™, или системы для двухступенчатой RT-PCR, поставляемые Roche Diagnostics [отделение F. Hoffmann-La Roche Ltd, Basel, Switzerland]).
Чтобы амплифицировать нуклеиновую кислоту с небольшим количеством несовпадений с одним или несколькими праймерами для амплификации, реакцию амплификации можно проводить в условиях пониженной строгости (например, амплификация PCR с использованием температуры отжига 38°C или в присутствии 3,5 мМ MgCl2). Специалист в данной области будет способен выбрать условия подходящей строгости.
Праймеры в данном описании выбирают «в основном» комплементарными (т.е. по меньшей мере на 65%, более предпочтительно по меньшей мере на 80% точно комплементарными) их областям-мишеням, присутствующим на различных цепях каждой конкретной последовательности, подлежащей амплификации. Можно использовать последовательности праймеров, содержащие, например, остатки инозита или неопределенные основания, или даже праймеры, содержащие одно или несколько несовпадений по сравнению с последовательностью-мишенью. Как правило, последовательности, обладающие по меньшей мере 65%, более предпочтительно по меньшей мере 80% гомологией с олигонуклеотидными последовательностями-мишенями ДНК или РНК, считают подходящими для использования в способе по настоящему изобретению. Несовпадения последовательности также не являются критическими при использовании условий гибридизации низкой строгости.
Детекцию продуктов амплификации можно, в принципе, осуществлять любым подходящим способом, известным в данной области. Амплифицированные фрагменты можно окрашивать напрямую или метить радиоактивными метками, антителами, люминесцентными красителями, флуоресцентными красителями или ферментативными реагентами. Прямые красители для ДНК включают в себя, например, интеркалирующие красители, такие как красители акридин оранжевый, бромид этидия, моноазид этидия или краситель Хехста.
Альтернативно, фрагменты ДНК или РНК можно выявлять посредством включения меченых оснований dNTP в синтезированные фрагменты. Метки для детекции, которые могут быть связанными с нуклеотидными основаниями, включают в себя, например, флуоресцеин, краситель цианин, дигоксигенин (DIG) или бромдезоксиуридин (BrdUrd).
При использовании систем детекции, основанных на зондах, подходящий способ детекции для применения по настоящему изобретению может, например, включать в себя формат ферментного иммунологического анализа (EIA) (Jacobs et al., 1997). Для осуществления детекции способом процедуры EIA либо прямой, либо обратный праймер, используемый в реакции амплификации, может содержать группу для захвата, такую как группа биотина для иммобилизации ампликонов PCR ДНК-мишени, например, на покрытых стрептавидином лунках планшета для микротитрования или на покрытых стрептавидином Dynabeads® (Dynal Biotech, Oslo, Norway) для последующей детекции EIA ампликонов ДНК-мишени. Специалисту в данной области понятно, что для иммобилизации ампликонов ДНК-мишени в формате EIA можно использовать другие группы.
Зонды, применимые для детекции последовательностей нуклеиновой кислоты-мишени, как описано в данном документе, предпочтительно связывают только по меньшей мере часть области последовательности нуклеиновой кислоты, амплифицированной способом амплификации нуклеиновой кислоты. Специалисты в данной области без излишних экспериментов могут получить подходящие зонды для детекции на основе нуклеотидной последовательности нуклеиновой кислоты-мишени, как описано в данном документе. Также в качестве типоспецифических зондов для детекции в способе по изобретению, соответственно, можно использовать комплементарные нуклеиновой кислоте-мишени нуклеотидные последовательности, либо ДНК, либо РНК, либо химически синтезированные аналоги, при условии, что такую комплементарную цепь амплифицируют в используемой реакции амплификации.
Подходящие способы детекции для применения в данном изобретении могут, например, включать в себя иммобилизацию ампликонов и гибридизацию с зондами последовательностей их нуклеиновой кислоты, например, посредством нозерн- и Саузерн-блоттинга. Другие форматы могут включать в себя формат EIA, как описано выше. Для облегчения детекции связывания специфические зонды для детекции ампликона могут содержать группу метки, такую как флуорофор, хромофор, фермент или радиоактивная метка, так чтобы облегчать мониторинг связывания зондов с продуктом реакции амплификации. Такие метки хорошо известны специалистам в данной области и включают в себя, например, изотиоцианат флуоресцеина (FITC), β-галактозидазу, пероксидазу хрена, стрептавидин, биотин, дигоксигенин, 35S, 14C, 32P или 125I. Другие примеры будут очевидны специалистам в данной области.
Детекцию можно проводить также путем так называемого анализа блота с перевернутыми дорожками (RLB), такого как, например, описанный Van den Brule et al. (2002). Для данной цели зонды для RLB предпочтительно синтезируют с 5'-аминогруппой для последующей иммобилизации, например, на покрытых карбоксилом нейлоновых мембранах. Преимуществами формата RLB являются простота системы и ее скорость, что, таким образом, позволяет более высокую производительность обработки образцов.
Использование нуклеиновокислотных зондов для детекции фрагментов РНК или ДНК хорошо известно в данной области. Большинство данных способов включают в себя гибридизацию нуклеиновой кислоты-мишени с зондом с последующими отмывками после гибридизации. Специфичность, как правило, является функцией отмывок после гибридизации, где критическими факторами являются ионная сила и температура конечного раствора для отмывки. Для гибридов нуклеиновой кислоты Tm можно приближенно вывести из уравнения Meinkoth-Wahl (1984): Tm=81,5°C+16,6 (log M)+0,41(%GC)-0,61 (%form)-500/L, где M представляет собой молярность моновалентных катионов, %GC представляет собой процент нуклеотидов гуанозина и цитозина в нуклеиновой кислоте, %form представляет собой процентное содержание формамида в растворе для гибридизации, и L представляет собой длину гибрида в парах оснований. Tm представляет собой температуру (при определенной ионной силе и pH), при которой 50% комплементарной последовательности-мишени гибридизуется с полностью совпадающим зондом. Tm уменьшают приблизительно на 1°C для каждого 1% несоответствия; таким образом, условия гибридизации и/или отмывки можно регулировать для гибридизации с последовательностями желаемой идентичности. Например, при поиске последовательностей с идентичностью >90% Tm можно увеличить на 10°C. Как правило, строгие условия выбирают приблизительно на 5°C ниже температурной точки плавления (Tm) для конкретной последовательности и комплементарной ей при определенной ионной силе и pH. Однако для чрезвычайно строгих условий можно использовать гибридизацию и/или отмывку на 1, 2, 3 или 4°C ниже температурной точки плавления (Tm); для умеренно строгих условий можно использовать гибридизацию и/или отмывку на 6, 7, 8, 9 или 10°C ниже температурной точки плавления (Tm); для условий с низкой строгостью можно использовать гибридизацию и/или отмывку на 11, 12, 13, 14, 15 или 20°C ниже температурной точки плавления (Tm). Обычному специалисту в данной области будет понятно, что с использованием уравнения, составов для гибридизации и отмывки и желаемой Tm по существу описывают варианты строгости гибридизации и/или растворов для отмывки. Если желаемая степень несоответствия приводит к Tm менее чем 45°C (раствор в воде) или 32°C (раствор в формамиде), предпочтительным является увеличение концентрации SSC, чтобы можно было использовать более высокую температуру. Обширное руководство по гибридизации нуклеиновых кислот представлено в Tijssen, 1993; Ausubel et al., 1998.
В другом аспекте изобретение относится к олигонуклеотидным зондам для детекции РНК или кДНК ToTV. Зонды для детекции здесь выбирают как «в основном» комплементарные одноцепочечной молекуле РНК или одной из цепей двухцепочечных нуклеиновых кислот, полученных посредством реакции амплификации по изобретению. Предпочтительно, зонды являются в основном комплементарными, не обязательно, иммобилизованным (например, меченным биотином) антисмысловым цепям ампликонов, полученных с мишени РНК или ДНК.
Для зондов для детекции по настоящему изобретению допустимо содержать одно или несколько несоответствий их последовательности-мишени. Как правило, последовательности, обладающие по меньшей мере 65%, более предпочтительно по меньшей мере 80% гомологией с олигонуклеотидными последовательностями-мишенями, считают подходящими для использования в способе по настоящему изобретению.
Устойчивые к ToTV растения
Кроме того, изобретение относится к способу идентификации устойчивого к ToTV растения или его части. Существуют различные возможности идентифицировать устойчивые к ToTV растения. В первом наборе вариантов осуществления такого способа можно применять активный/инфекционный вирус или полноразмерные инфекционные клоны, тогда как в альтернативном варианте осуществления используют только средства для детекции вируса.
Первая стадия способа идентификации устойчивого к ToTV растения с использованием активного/инфекционного вируса включает в себя воздействие на растение или часть растения, такую как фрагмент листа или стебля, инфекционной дозы ToTV, целью чего является достичь инфекции. Воздействие во многих случаях включает в себя установление физического контакта. Инфекционная доза может различаться между растениями и между тестируемыми изолятами ToTV. Теоретически, количество от приблизительно 1-10 до приблизительно 500-5000 вирусных частиц указанного вируса или его нуклеиновых кислот будет являться достаточным. Инфекции, таким образом, можно достигать механической инокуляцией очищенных частиц вируса или нуклеиновой кислоты вируса в здоровые растения.
Альтернативно, инфекции можно достигать, например:
- выращиванием здорового черенка на зараженном ToTV корневище или наоборот;
- воздействием на здоровое растение переносчиков, содержащих вирус (включая инфицированные растения, например растения-паразиты, подобные видам Cuscuta);
- введением в здоровое растение экспрессирующего вектора, несущего кодирующую область генома вируса ToTV;
- применением агроинфекционных клонов, таких как штаммы Agrobacterium tumefaciens, содержащих экспрессирующий вектор, несущий кодирующую область генома вируса ToTV.
В контексте настоящего изобретения способы воздействия на растение или часть растения инфекционной дозы ToTV не являются ограниченными каким-либо конкретным способом.
Как указано, инфекция может включать в себя механическую инокуляцию вируса в здоровые растения. Например, часть пораженного заболеванием листа можно втереть непосредственно в лист подлежащего инфицированию растения. По альтернативному способу инокулят можно получить, например, перемалыванием содержащей вирус ткани растения, предпочтительно молодых листьев с выраженными симптомами, с помощью ступки и пестика, или любого другого типа гомогенизатора, например, в буфере, пригодном для инокуляции (например 0,03 M фосфатный буфер, pH 7,7). После перемалывания полученный гомогенат (сок) предпочтительно фильтруют, например, через суровую марлю. Затем сок можно инокулировать, например, путем осторожного приведения листьев в контакт с каким-либо количеством сока. Листья предпочтительно предварительно обрабатывают, чтобы разрушить нижний эпидермис и усилить проникновение вируса. Этого можно достигать, например, предварительным припудриванием листьев порошком карбида кремния. Предпочтительно избегают избыточного повреждения. Предпочтительно используют порошок карбида кремния, обладающий микроскопически малыми частицами карбида кремния угловатой формы (400-500 меш). Порошок карбида кремния можно также добавлять непосредственно в сок, в этом случае предварительную обработку исключают. Сок можно наносить, например, указательным пальцем, тампоном из пропитанного соком пенопласта или ткани или даже пестиком, используемым для перемалывания, стеклянным шпателем, жесткой кистью или пульверизатором. После инокуляции листья предпочтительно немедленно промывают водой.
Вторая стадия способа идентификации устойчивого к ToTV растения включает в себя идентификацию указанного растения как устойчивого к ToTV растения, когда, после указанного воздействия, или i) симптомы заболевания в указанном растении или части растения остаются отсутствующими, или их проявление задержано, или по меньшей мере тяжесть уменьшена, или они являются локализованными по сравнению с восприимчивым и/или чувствительным контрольным растением, и/или ii) вирус ToTV или геномные последовательности ToTV не присутствуют в указанном растении или части растения, или присутствие вируса ToTV является по меньшей мере уменьшенным количественно по сравнению с чувствительным контрольным растением. Как используется в данном описании, термин локализованный означает ограниченный инокулированным листом.
Определение развития симптомов индуцированного ToTV заболевания в инфицированных растениях можно осуществлять количественными способами, например, где отмечают период, необходимый для развития явных (например, видимых) симптомов заболевания, или количественными способами, где после прекращения определенного периода растение проверяют на проявление симптомов и отмечают присутствие или тяжесть симптомов.
В дополнение к определению развития индуцированных ToTV-симптомов заболевания или в качестве альтернативы ему, в зависимости от подлежащего детекции типа устойчивости к ToTV, выявляют присутствие вируса в растении или части растения. Для детекции отсутствия вируса в тестируемых растениях, в принципе, можно использовать любой способ. Например, можно применять способ, в котором используют специфическое для ToTV антитело, набор праймеров или зонд по настоящему изобретению. Альтернативно, часть тестируемого растения можно приводить в контакт с чувствительным индикаторным растением (например, N.hesperis '67A') для определения, присутствует или отсутствует вирус в тестируемом растении. Специалисту в данной области понятно, что для таких способов является важным дезинфицировать поверхность тестируемого растения, чтобы провести различие между переносчиком, толерантным тестируемым растением и устойчивым тестируемым растением, поскольку необходимо показать только присутствие вируса в клетках растения.
При проведении второй стадии способа идентификации устойчивого к ToTV растения можно получить следующие результаты. Если после успешной инокуляции (например, после осуществления контакта растения с вирусом в условиях, приводящих к инфекции в восприимчивом и чувствительном контрольном растении):
i) симптомы заболевания остаются отсутствующими; или вирусные частицы, или вирусная РНК не поддаются выявлению: растение является устойчивым;
ii) проявление симптомов заболевания задержано, или их тяжесть уменьшена; или можно выявить низкие системные титры вирусных частиц или вирусной РНК: растение является частично устойчивым;
iii) симптомы заболевания являются тяжелыми, но остаются локализованными, ограниченными инокулированным листом и не распространяются системно за пределы инокулированной ткани; или вирусные частицы, или вирусную РНК можно детектировать только локализованно: растение является гиперчувствительным;
iv) если симптомы заболевания остаются отсутствующими и можно детектировать вирусные частицы, или вирусную РНК: растение является толерантным.
v) если у растения развиваются симптомы заболевания и оно обладает высокими системными титрами вируса, тогда растение является восприимчивым и чувствительным. Примерами таких растений являются растения, из которых выделен вирус по настоящему изобретению. Эти растения могут служить подходящими контрольными растениями в способах по настоящему изобретению.
Для целей получения устойчивых растений и с точки зрения фитосанитарии только исходы i), ii) и iii) можно считать представляющими интерес. Для целей получения растений, пригодных для получения бессимптомного урожая и продуктов, исход iv) может представлять также конкретный коммерческий интерес.
В альтернативном варианте осуществления способа для идентификации устойчивого к ToTV растения используют только средства для детекции вируса. Например, устойчивое к ToTV растение можно идентифицировать в полевых условиях посредством обнаружения или идентификации бессимптомного растения среди обладающих симптомами растений и определения отсутствия вируса в указанном растении любым из способов детекции вируса по настоящему изобретению. Фактически, это соответствует способу идентификации устойчивого к ToTV растения, где стадию a) воздействия на растение или часть растения инфекционной дозы ToTV проводят пассивно (например, естественным образом). При осуществлении такого способа предпочтительно использовать способ детекции присутствия ToTV в образце по настоящему изобретению, где присутствие вируса ToTV или его компонента демонстрируют посредством реакции указанного образца с полинуклеотидом или антителом по настоящему изобретению. Предпочтительно, для способа идентификации устойчивого к ToTV растения необходимо использование либо вируса по настоящему изобретению, либо полинуклеотида или антитела по настоящему изобретению.
Кроме того, изобретение относится к способу получения устойчивого к ToTV растения или его части. После идентификации устойчивого к ToTV растения это растение может служить донорным растением для генетического материала, который можно переносить от указанного донорного растения к реципиентному растению, чтобы обеспечить указанное реципиентное растение генетическим материалом. Перенос генетического материала от донорного растения к реципиентному растению можно осуществлять любым подходящим способом, известным в данной области. Генетический материал в большинстве случаев будет геномным материалом. Однако является важным переносить по меньшей мере одну сообщающую устойчивость часть генома донорного растения. В отсутствие способов определения, какие части генома донорного растения сообщают устойчивость к ToTV, перенос можно осуществлять, соответственно, посредством переноса целых хромосом. Предпочтительно, устойчивое к ToTV растение служит мужским или женским родителем в скрещивании для получения устойчивых растений-потомков, таким образом, растение-потомок получает геномный материал от устойчивого донора и действует как реципиентное растение. Хотя чувствительный родитель при скрещивании не обязательно является реципиентным растением в буквальном смысле, такой чувствительный родитель здесь также включен в термин реципиентное растение.
В способе получения устойчивого к ToTV растения можно также использовать слияние протопластов для переноса сообщающего устойчивость геномного материала от донорного растения к реципиентному растению, т.е. в качестве способа скрещивания указанных растений. Слияние протопластов является индуцированным или спонтанным объединением, таким как соматическая гибридизация, между двумя или более протопластами (клетками, клеточные стенки которых удалены ферментативной обработкой) для получения одной би- или многоядерной клетки. Слитая клетка, которую можно получить даже из видов растений, которые не могут скрещиваться в природе, представляет собой ткань, культивируемую до гибридного растения, обладающего желаемым сочетанием признаков. Более конкретно, первый протопласт можно получить из растения томата или другой линии растений, обладающей устойчивостью к инфекции ToTV. Например, можно использовать протопласт из устойчивой к ToTV линии (томата, баклажана, перца, дыни, арбуза или огурца). Второй протопласт можно получить из второй, чувствительной, линии растений, не обязательно, из других видов растений или вариетета, предпочтительно, из тех же самых видов растений или вариетета, обладающих коммерчески желательными характеристиками, такими как, без ограничения, устойчивость к заболеванию, устойчивость к насекомым, различные характеристики плодов и т.д. Затем протопласты сливают с использованием общепринятых способов слияния протопластов, известных в данной области для получения скрещивания.
Альтернативно, можно использовать спасение эмбриона для переноса сообщающего устойчивость геномного материала от донорного растения реципиентному растению, т.е. как способ скрещивания указанных растений. Спасение эмбриона можно использовать как способ выделения эмбрионов из потомков скрещивания, когда растения неспособны производить жизнеспособные семена. По данному способу оплодотворенную завязь или незрелое семя растения культивируют для получения новых растений (этот способ подробно описан в Pierik, 1999).
Таким образом, способ получения устойчивого к ToTV растения в одном из вариантов осуществления включает в себя стадии идентификации устойчивого к ToTV донорного растения, как описано здесь выше, и скрещивания указанного устойчивого к ToTV донорного растения с реципиентным растением, как описано выше, таким образом, получая растения-потомки.
Способ получения устойчивого к ToTV растения дополнительно включает в себя стадию отбора из растений-потомков устойчивого растения способом идентификации устойчивого к ToTV растения, как описано ранее.
Предпочтительно, указанное устойчивое к ToTV донорное растение представляет собой растение семейства Solanaceae или Cucurbitaceae, даже более предпочтительно томат, баклажан, перец, дыню, арбуз или огурец.
Предпочтительно, указанное реципиентное растение представляет собой растение семейства Solanaceae или Cucurbitaceae, даже более предпочтительно томат, баклажан, перец, дыню, арбуз или огурец. Еще более предпочтительно, указанное реципиентное растение представляет собой томат видов Solanum lycopersicum, более предпочтительно растение S.lycopersicum, обладающее коммерчески желательными характеристиками. Реципиентное растение может быть восприимчивым к ToTV растением, чувствительным к ToTV растением или устойчивым к ToTV реципиентным растением. Как объясняется выше, выбор растения будет в первую очередь определяться тем, является ли признак устойчивости доминантным или рецессивным. Специалист в данной области будет сознавать, что различные способы доступны для решения таких задач.
Также аспектом по настоящему изобретению является устойчивое к ToTV растение, или его часть, которые можно получить способом по изобретению.
Как указано, предпочтительный вариант осуществления способа для получения устойчивого к ToTV растения включает в себя перенос посредством интрогрессии указанной сообщающей устойчивость последовательности нуклеиновой кислоты от устойчивого к ToTV донорного растения к реципиентному растению путем скрещивания указанных растений. В устойчивых растениях, полученных по данному предпочтительному варианту осуществления, большинство признаков успешно могут происходить из реципиентного растения, а устойчивость к ToTV - происходить из донорного растения.
В одном из способов, который обозначается как племенное разведение, донорное растение, обладающее устойчивостью к ToTV, скрещивают с реципиентным растением, предпочтительно обладающим коммерчески желательными характеристиками, такими как, без ограничения, устойчивость к заболеванию, устойчивость к насекомым, полезные характеристики плодов, и т.д. Полученную популяцию растений (представляющих собой гибриды F1) затем подвергают самоопылению и обеспечивают возможность образования семян (семена F2). Затем растения F2, выращенные из семян F2, скринируют по устойчивости к ToTV. Проводить скрининг популяции можно рядом различных способов, предпочтительно способом визуального контроля по настоящему изобретению.
Поскольку идентификация устойчивых к ToTV растений первоначально стала возможной только по настоящему изобретению, способ получения устойчивого растения является аспектом изобретения. Также аспектом настоящего изобретения является устойчивое к ToTV растение или его часть, которую можно получить способом по изобретению.
Настоящее изобретение относится к способам предотвращения распространения инфекции ToTV в растениях томата путем получения устойчивых растений томата, так же как путем уничтожения растений, несущих вирус ToTV. Данные меры могут составлять часть общей стратегии улучшения фитосанитарной профилактики по отношению к вирусу ToTV. Таким образом, можно идентифицировать и уничтожать толерантные растения, чтобы уничтожить такие источники вируса ToTV.
В одном из вариантов осуществления способа получения устойчивого к ToTV растения или его части, настоящее изобретение относится к способу получения толерантного к ToTV растения. Из толерантного растения можно получать полноценный урожай, плоды и семена, поскольку, хотя растение может быть носителем вируса, для него не обнаруживают симптомов заболевания. Такой способ будет включать в себя идентификацию толерантных растений и использование таких толерантных растений в качестве источников или доноров желательного генетического материала. Целью является не получение растения, способного сопротивляться проникновению или размножению вируса в клетках, а получение растения, которое не страдает от симптомов.
Таким образом, настоящее изобретение относится к способу идентификации устойчивого к ToTV растения, включающему стадии: a) воздействия на растение или часть растения инфекционной дозы ToTV, и b) идентификации указанного растения как толерантного к ToTV растения, когда после указанного воздействия симптомы заболевания в указанном растении или части растения остаются отсутствующими, а ToTV присутствует в указанном растении или части растения.
Определение развития симптомов индуцированного ToTV заболевания в инфицированных растениях можно осуществлять количественными способами, например, где отмечают период, необходимый для развития явных (например, видимых) симптомов заболевания, или количественными способами, где после прекращения определенного периода растение проверяют на отсутствие проявления симптомов или отмечают уменьшение тяжести симптомов.
В предпочтительном варианте осуществления присутствие ToTV в указанном растении или части растения определяют на стадии b) способом, включающим определение в указанном растении или части растения присутствия вируса ToTV или его компонентов с помощью реакции указанного растения или части растения с полинуклеотидом по изобретению или антителом по изобретению.
Диагностические наборы
Способы и средства, предоставленные в данном описании, являются, в частности, применимыми в диагностическом наборе для диагностики инфекции вируса ToTV посредством вирусологического диагноза. Такие наборы или анализы могут, например, содержать вирус, нуклеиновую кислоту, белковую молекулу или ее фрагмент и/или антитело по изобретению.
Изобретение относится также к диагностическому набору для диагностики инфекции ToTV, содержащему вирус ToTV, специфическую для вируса ToTV нуклеиновую кислоту, белковую молекулу или ее фрагмент и/или антитело по изобретению, и предпочтительно, средства для детекции указанного вируса ToTV, специфической для вируса ToTV нуклеиновой кислоты, белковой молекулы или ее фрагмента и/или антитела, где указанные средства, например, включают в себя возбуждаемую группу, такую как флуорофор, или ферментативную систему детекции, применяемую в данной области (примеры подходящих форматов диагностических наборов включают IF, ELISA, анализ нейтрализации, анализ RT-PCR, анализы гибридизации). Подходящие анализы для детекции включают прямые и непрямые анализы, сэндвич-анализы, твердофазные анализы, такие как, среди прочих, анализы с использованием планшетов или гранул, и анализы в жидкой фазе. Подходящие анализы включают анализы с использованием первичных и вторичных антител и анализы с использованием связывающих антитело реагентов, таких как белок A. Более того, по изобретению можно использовать множество способов детекции, включая колориметрический, флуоресцентный, фосфоресцентный, хемилюминесцентный, люминесцентный и радиоактивный способы.
Чтобы определить, можно ли компонент еще не идентифицированного вируса или его синтетический аналог, такой как нуклеиновая кислота, белковая молекула или ее фрагмент, идентифицировать как специфический для вируса ToTV, достаточно анализировать последовательность нуклеиновой кислоты или аминокислоты указанного компонента, например фрагмент указанной нуклеиновой кислоты или аминокислоты, предпочтительно по меньшей мере 10, более предпочтительно по меньшей мере 25, более предпочтительно по меньшей мере 40 нуклеотидов или аминокислот (соответственно), путем сравнения последовательности с предоставленными последовательностями вируса ToTV и с известными последовательностями, не относящимися к вирусу ToTV (предпочтительно используют ближайшие филогенетические родственники ToTV), с использованием, например, филогенетических анализов, как предусмотрено в данном описании. В зависимости от степени родства с указанными вирусными последовательностями ToTV или не относящимися к ToTV, можно идентифицировать компонент или синтетический аналог.
Набор для детекции вируса ToTV может, в зависимости от формата анализа, содержать одно или несколько антител, специфических для белка, предпочтительно специфических по меньшей мере для одного белка капсида ToTV, и предпочтительно также содержать в основном очищенный белок ToTV или антиидиотипическое антитело для использования в качестве положительного контроля.
Противовирусные средства
Изобретение относится также к способам получения противовирусного средства, применимого для лечения инфекции ToTV у растений, включающим получение культуры клеток или экспериментального растения, содержащего вирус по изобретению, обработку указанной культуры или растения потенциальным противовирусным средством и определение эффекта указанного средства на указанный вирус или его инфекцию указанной культуры или растения. Примеры такого противовирусного средства включают в себя нейтрализующее ToTV антитело или его функциональный компонент, как предусмотрено в данном описании, однако можно получать также противовирусные средства другой природы.
Существуют различные противовирусные средства, применимые для растений, такие как химические продукты, бактерии, грибы, насекомые и вирусы. Большинство из них связано с системной приобретенной устойчивостью (SAR). Настоящее изобретение относится к применению генома ToTV или его части в качестве индуктора системной приобретенной устойчивости у растения. Системная приобретенная устойчивость может быть направленной против ToTV или против других заболеваний. По данному аспекту ToTV, его геном или его сообщающие устойчивость части можно применять в качестве противовирусного агента.
Изобретение относится также к применению противовирусного средства по изобретению для получения композиции для лечения, в частности для лечения инфекции ToTV у растений, и относится к фармацевтической композиции, содержащей противовирусное средство по изобретению, применимой в способе лечения или предупреждения инфекции вирусом ToTV, где указанный способ включает введение такой композиции для лечения в отдельное растение.
Изобретение также относится к растительной модели, применимой для тестирования способов и/или композиций для лечения. По-видимому, некоторые виды Nicotiana можно инфицировать вирусом ToTV, благодаря чему наблюдать симптомы заболевания, отличающиеся от симптомов, обнаруженных у растений томата, страдающих от вируса ToTV. Подвергая растения Nicotiana противовирусному лечению либо до, либо во время инфекции вирусом можно иметь предсказуемое значение применения такого противовирусного средства для растений томата.
Изобретение также относится к применению ToTV или частей генома вируса ToTV в качестве экспрессирующего вектора, например, для применения для индуцированного вирусом молчания генов (VIGS). VTGS представляет собой технологию, использующую РНК-опосредованный механизм противовирусной защиты у растений. У растений, инфицированных немодифицированными вирусами, механизм специфически направлен против вирусного генома. С использованием вирусных экспрессирующих векторов, несущих вставки, полученные из генов хозяев, механизм можно использовать также для атаки против соответствующих РНК растений. VIGS широко использовали в растениях для анализа функции генов и адаптировали для высокопроизводительной функциональной геномики. До настоящего времени большинство применений VIGS относилось к Nicotiana benthamiana. Однако настоящее изобретение относится к применению ToTV в качестве новых экспрессирующих векторных систем, позволяющих анализировать функцию генов в других растениях, таких как томат или другие виды семейства Solanaceae, такие как перец и картофель, и в видах семейства Cucurbitaceae.
Изобретение далее объясняется в примерах без ограничения ими.
ПРИМЕРЫ
Пример 1. Выделение и характеризация ToTV из растений томата
Методы
Введение
Образцы растений томата из Испании получали для диагностических исследований. Симптомы у растений томата проявлялись в виде некротических пятен и хлороза на листьях и коричневых колец на плодах. Серологические тесты (ELISA) указывали на присутствие вируса мозаики пепино (PepMV), однако, принимая во внимание симптомы, по-видимому, присутствовал другой, еще не идентифицированный агент.
Электронно-микроскопическими исследованиями ткани инфицированного листа были обнаружены сферические вирусные частицы.
Затем проводили тест на инфекцию и обнаружили, что восприимчивыми к ToTV являлись многие образцы томата, некоторые из которых реагировали с явными симптомами (некроз листьев, начинающийся в основании отдельных листочков (см. Фиг.1).
Перенос и размножение вируса
ToTV выделяли, как описано ниже, из пораженных болезнью растений, полученных из Испании. ToTV можно было механически перенести к нескольким видам Nicotiana. Показано, что общепринятый буфер для инокуляции является пригодным (например, 0,03 M фосфатный буфер, pH 7,7). Везде в данном примере ToTV механически инокулировали в N.glutinosa или N.benthamiana и размножали в них. Очистку вируса проводили приблизительно через 14 дней после инокуляции.
Очистка вируса
Выполняли несколько попыток очистки ToTV согласно общепринятым способам, например, для неповирусов или лютеовирусов (с помощью органических растворителей). Эти способы всегда приводили к потере инфекционности вируса. Более того, ToTV проявлял тенденцию к агрегации, когда на стадиях центрифугирования использовали низкие температуры (ниже 5°C).
В конечном счете, очень мягким способом очистки в результате получали приемлемо чистые препараты вируса, на которых можно было проводить дальнейшие эксперименты.
Для очистки ToTV использовали следующий способ (все стадии центрифугирования проводили при 6°C). Инфицированные листья N.glutinosa или N.benthamiana гомогенизировали в 5 частях 0,1 M Трис-HCl (pH 8) плюс 20 мМ Na2SO3, 10 мМ Na-DIECA и 5 мМ Na-EDTA (буфер для гомогенизации) и гомогенат центрифугировали в течение 30 минут при 49000×g. Супернатант помещали на подушку 20% сахарозы и центрифугировали в течение 1,5 часов при 70000×g. Осадок ресуспендировали в 2 мл Трис-HCl, pH 8, суспензию наносили на градиент сахарозы (10-40% сахарозы в буфере для гомогенизации) и центрифугировали в течение 2 часов при 110000×g. Поскольку полоса с вирусом не была видимой, градиент аликвотировали на отдельные фракции и присутствие вируса в каждой фракции определяли путем инокуляции части указанной фракции в листья N.hesperis '67A', как описано в данном документе, и наблюдения наличия инфекции. Содержащую вирус фракцию помещали в 10-40% градиент сульфата цезия (в Трис-HCl, pH 8) и центрифугировали в течение 16 час при 125000×g. Полосы с вирусом собирали и концентрировали центрифугированием или диализовали против 0,1 M Трис-HCl, pH 8.
Инфекционность ToTV после каждой стадии очистки проверяли инокуляцией в N.hesperis '67A' (перед инокуляцией фракции градиента сульфата цезия диализовали против Трис-HCl, pH 8).
Электронная микроскопия
Суспензии вируса «монтировали» на сетке, покрытой Formvar®, известным иначе как поливинилформаль, окрашивали 2% уранилацетатом и исследовали на электронном микроскопе Philips CM12.
Анализ PAGE
Вирусные белки разделяли электрофорезом в 12% денатурирующем полиакриламидном геле (SDS-PAGE, Laemmli, 1970) и окрашивали серебром.
Выделение и оценка нуклеиновой кислоты
Очищенный вирус концентрировали центрифугированием (2 часа при 115000 g). Осадки подвергали экстракции РНК по способу очистки Qiagen RNeasy MinElute (Qiagen, Hilden, Germany). Концентрацию РНК определяли на УФ-спектрофотометре (Beckman Coulter, Inc., Fullerton, USA).
Целостность РНК проверяли электрофорезом в агарозном геле. После электрофореза в течение 2 часов при 60 В РНК окрашивали с использованием ортотолуидина синего.
Синтез и клонирование кДНК
кДНК синтезировали с использованием системы для синтеза кДНК Invitrogen Superscript Choice (Invitrogen, Breda, The Netherlands) согласно инструкциям производителя. Первую цепь кДНК праймировали с использованием либо олиго-d(T), либо случайных гексамерных праймеров. После синтеза второй цепи лигировали адаптеры для EcoRI для облегчения клонирования. После фосфорилирования кДНК с адаптерами для EcoRI нелигированные линкеры удаляли колоночной хроматографией. Полученную кДНК лигировали в предварительно расщепленный EcoRI экспрессирующий вектор pBluescript II (Stratagene, La Jolla, USA) и лигатом трансформировали компетентные клетки Top10 (Invitrogen).
5'-Конец последовательности ToTV определяли с использованием системы быстрой амплификация концов кДНК 5'RACE (LifeTechnologies) с использованием dCTP согласно инструкциям производителя.
Анализ кДНК
После трансформации в рекомбинантных клонах анализировали присутствие вставок посредством PCR с использованием праймеров, специфических как для T3, так и для T7. Размер продуктов PCR анализировали в 1% агарозном геле. Клоны, содержащие вставки приблизительно по 1500 нуклеотидов, использовали для дальнейшего анализа последовательности. Полученные данные о последовательности анализировали с использованием пакета программ DNASTAR.
Определение аминокислотной последовательности трех белков капсида ToTV
Очищенные частицы ToTV наносили на денатурирующий гель для PAGE и разделяли белки капсида. Полосы с разделенными белками капсида вырезали из геля, обрабатывали трипсином и гидролизаты анализировали с использованием тандемного масс-спектрометра (MSMS), по существу с использованием способа, описанного Kinter & Sherman, 2000. В результате этого получали аминокислотные (ак) последовательности малых пептидов, каждая из которых проявляла гомологию с аминокислотной последовательностью, прогнозируемой из нуклеотидной последовательности RNA2 ToTV. Сравнение с другими данными о вирусных последовательностях выполняли с помощью программ из пакета PHYLIP.
Результаты
Перенос и размножение вируса
Для размножения ToTV можно использовать N.glutinosa и N.benthamina. Виды табака N.hesperis '67A' и N.occidentalis 'P1' являются очень восприимчивыми к ToTV и проявляют симптомы после 3-4 суток. В этих видах табака развивается сильный некроз за короткое время, и, таким образом, их считают более подходящими в качестве индикаторного растения, чем хозяина для размножения. N.glutinosa и N.benthamiana реагируют с местными очагами хлороза и системным хлорозом, а также с умеренной деформацией листьев.
Очистка вируса
Очистка вирионов из инфицированной ткани листа оказалась довольно сложной. ToTV является неустойчивым к органическим растворителям и проявляет тенденцию к агрегации при центрифугировании при низких температурах. Используемым протоколом очистки (см. 1.2.) получили две видимые содержащие вирус полосы в градиенте сульфата цезия. Полосы появлялись в нижней части градиента, что указывает на довольно высокую плавучую плотность в Cs2SO4, равную или превышающую 1,4 г/см2.
Сульфат цезия влиял на инфекционность ToTV, однако она не терялась полностью при высокой концентрации вируса в исходном материале. Фракции градиента сульфата цезия, содержащие обе полосы с вирусом, были инфекционными. Инфекционность отдельных полос не определяли.
Электронная микроскопия
Две полосы анализировали электронной микроскопией, и они содержали частицы вируса диаметром приблизительно 28 нм (Фиг.2).
Анализ PAGE
Электрофорезом в полиакриламидном геле (PAGE) очищенных фракций вируса с последующим окрашиванием геля серебром обнаружили три белка капсида (CP) приблизительно 23, 26 и 35 кДа (см. Фиг.3, где показаны очищенные верхняя (TAgV-T) и нижняя фракции (TAgV-B) очищенного вируса).
Выделение нуклеиновой кислоты и анализ кДНК
Выделением РНК из обеих полос с вирусом вместе получили две РНК: приблизительно 5,5 т.н. и 8 т.н. (см. Фиг.4). Выделением РНК из верхней полосы получали только фрагмент РНК 5,5 т.н.
РНК 5,5 т.н. из верхней полосы использовали в качестве матрицы для синтеза и клонирования кДНК. Для различных клонов проводили реакции секвенирования с использованием прямого и обратного праймеров для секвенирования (T3/T7 или M13F/M13R). Для определения точного 5'-конца РНК выполняли быструю амплификацию концов кДНК (RACE). В результате анализа полученных данных о последовательности с использованием пакета программ Lasergene® (DNASTAR, Inc., Madison, WI, USA), получили полную последовательность РНК2 вируса (SEQ ID NO:1; см. фиг. 5). Размер РНК составляет 5389 нуклеотидов, исключая поли A-хвост. В РНК обнаружили две открытые рамки считывания (ORF). ORF1 локализована от нуклеотида 182 до 742, обладает длиной 561 нуклеотидов и кодирует белок из 187 аминокислот. Для данной последовательности не обнаружено гомологии в базах данных NCBI на белковом и нуклеотидном уровне.
ORF2 простирается от нуклеотида 702 до 4298, обладает длиной 3597 нуклеотидов и кодирует белок из 1199 аминокислот. После поиска BLAST в базе данных NCBI обнаружили небольшую гомологию с несколькими вирусными полибелками. Для обнаруженных гомологов инвентарные номера из базы данных NCBI приведены вместе с названием вируса и типом белка. Как последовательности нуклеиновой кислоты, так и производные аминокислотные последовательности во всех трех рамках считывания использовали для анализа BLAST. Для идентификации ORF использовали MAPDRAW из пакета программ Lasergene®.
Карты ORF на обеих РНК
РНК1
РНК1 содержит одну ORF (ORF1) с мотивами, типичными для геликазы, РНК-зависимой РНК-полимеразы (RdRp). Помимо этого, для аминокислот в положениях 106-338 обнаружили низкий уровень гомологии аминокислотной (ак) последовательности с кофактором протеазы (Pro-Co) вируса слабой мозаики пачули (PatMMV) (NP647592.1), с идентичностью 22%.
Типичные мотивы геликазы A (GKS), В (D), С (N) идентифицированы в положениях 398-400, 444 и 495 предположительного полипептида.
Для области геликазы наиболее близкую идентичность обнаружили со сферическим вирусом риса тунгро (RTSV; 42% идентичности в перекрывающихся 140 ак), вирусом карликового хлороза кукурузы (MCDV; 43% идентичности в перекрывающихся 137 ак), вирусом крапчатости земляники (SMoV; 42% идентичности в перекрывающихся 135 ак) и вирусом желтой пятнистости пастернака (PYFV; 42% в перекрывающихся 138 ак). В предположительной области VpG не обнаружено сходства последовательности с другими вирусами.
Наибольшее сходство по протеазе обнаружено для ак 1000-1100, где обнаружено 25% идентичности с вирусом картофеля V (PVV; в перекрывающихся 86 ак протеазы NIa).
Область RdRp между мотивами I (KDE)-VII (FLSR) обнаружили между ак 1303-1554 (Koonin 1991).
Наиболее близкую идентичность последовательности полибелка обнаружили с: сферическим вирусом риса тунгро (RTSV) с 29% идентичности в перекрывающихся 751 ак, вирусом карликового хлороза кукурузы (MCDV) с 28% идентичности в перекрывающихся 742 ак, вирусом желтой пятнистости пастернака (PYFV) с 33% идентичности в 501 ак, сферическим латентным вирусом яблока (ALSV) с 32% идентичности в 472 ак, вирусом крапчатости земляники (SMoV) с 30% идентичности в 680 ак и вирусом рашпилевидности листьев вишни (CRLV) с 33% в 465 ак.
РНК2
РНК2 содержит две потенциальные ORF (Фиг.7). ORF1 кодирует предположительный белок из 187 ак с молекулярной массой 20 кДа. Анализом последовательности не выявили гомологии с каким-либо белком из баз данных EMBL. Неизвестно, кодирует ли данная ORF реально существующий белок.
Вторая ORF, частично перекрывающаяся с ORF1, начинается с трех стартовых кодонов ATG в рамке. Она кодирует предположительный белок из 1198 ак с предсказанной молекулярной массой 134 кДа. Идентификацией белков посредством масс-спектрометрии выделенных белков оболочки четко картировали три цистрона белков оболочки на C-конце РНК2-ORF2.
N-концевая область РНК2-ORF2 для полибелка наиболее вероятно кодирует предположительный двигательный белок (MP), поскольку в положении ак 262-267 обнаружен мотив LRVPML, очень похожий на консенсусную последовательность предположительного двигательного белка LxxPxL (Mushegian, 1994). Других гомологий последовательности для N-конца ORF2 РНК2 не обнаружено.
ORF2 предположительно кодирует четыре белка, которые должны отщепляться из полибелка-предшественника посредством протеолитического расщепления. Однако невозможно идентифицировать очевидную гомологию с участками расщепления известных полибелков. Остается необходимость определить точное положение этих участков расщепления.
Для последовательности полибелка CP1 авторы настоящего изобретения обнаружили только некоторую гомологию с парэховирусом человека (HPeV) с 21% идентичностью в 103 ак. Наиболее близкую идентичность с последовательностью полибелка CP2 обнаружили для вируса обыкновенной черемуховой тли (RhPV) с 25% идентичностью в 168 ак, вируса энцефаломиелита птиц (AEV) с 33% в 74 ак, вируса черного маточника (BQCV) с 43% идентичностью в 37 ак и вируса красного огненного муравья (SINV-1) с 30% идентичностью в 51 ак. Не обнаружено гомологий с последовательностью полибелка CP3.
Нетранслируемые области (UTR)
3'-UTR РНК1 и РНК2 составляют 1210 н. и 1092 н., соответственно. Существует 98% идентичность концевых 988 нуклеотидов обеих РНК. Для подтверждения того, что 3'-области UTR обеих РНК являются идентичными, проводили RT-PCR тотальной вирусной РНК с одним обратным праймером, выведенным из идентичной области 3'-UTR и двух специфических для каждой РНК прямых праймеров. Полученные продукты PCR секвенировали.
По этим результатам заключили следующее: вирус, выделенный из растений томата и предварительно названный вирусом томата торрадо (ToTV), обладает сферическими (икосаэдрическими) частицами диаметром приблизительно 28 нм. При очистке вирус проявляет на дисплее по меньшей мере две видимые полосы в градиенте сульфата цезия. Обе объединенные полосы являются инфекционными при инокуляции растений табака. Частицы вируса, по-видимому, состоят по меньшей мере из трех белков капсида приблизительно 23, 26 и 35 кДа.
Верхняя фракция вируса из градиента сульфата цезия содержит молекулу РНК приблизительно 5,5 т.н. (РНК 2; SEQ ID NO:1), а нижняя фракция - молекулу РНК приблизительно 8 т.н. (РНК 1; SEQ ID NO:2).
В результате синтеза и клонирования кДНК с использованием РНК 5,5 т.н. из верхней вирусной фракции и последующего анализа информации о последовательности несколько контигов собрали в SEQ ID NO:1. Два контига явно содержали поли-A-хвост, что позволяло предполагать, что вирусная РНК обладает поли-A-хвостом. Анализ BLAST нуклеотидной и производной аминокислотной последовательности не выявил какой-либо существенной гомологии с каким-либо известным вирусом из базы данных EMBL.
Приведенная выше информация указывает, что ToTV является новым и до настоящего времени не описанным вирусом. Полученная до настоящего времени информация еще не позволяет группировать вирус с конкретным вирусным семейством или родом.
Для целей детекции и идентификации вируса сконструированы два набора праймеров для RT-PCR (таблица 4) на основе последовательности SEQ ID NO:1.
Праймеры для RT-PCR для детекции ToTV
темп
Подходящий способ RT-PCR включает в себя следующее. Выделяют и очищают общую РНК из приблизительно 100 мкг материала инфицированного листа с использованием набора для очистки РНК, например, такого как Qiagen RNA-Easy. Из этой общей РНК количество приблизительно 1 мкг используют в 50 мкл реакционной смеси для одноступенчатой реакции Superscript RT-PCR (Invitrogen), где реакционная смесь дополнительно содержит 25 мкл 2× реакционной смеси, 1 мкл (100 нг) как верхнего, так и нижнего праймера, 1 мкл смеси RT/Taq и 22 мкл воды MilliQ. Чтобы провести обратную транскрипцию РНК в кДНК и амплифицировать эту кДНК, можно использовать следующую программу RT-PCR: стадия 1: 30 минут при 50°C (реакция обратной транскрипции); стадия 2: 3 минуты при 94°C (активация Taq-полимеразы); стадия 3: 30 секунд при 94°C; стадия 4: 30 секунд при 55°C; стадия 5: 1 минута при 72°C; стадия 6: повтор стадий (3-5) 40×; стадия 7: 10 минут при 72°C; стадия 8: 10°C так долго, как потребуется. Продукты PCR можно анализировать в 1% агарозном геле в буфере TAE или TBE.
2.6. Определение аминокислотных последовательностей трех белков капсида ToTV
Фрагменты наибольшего белка оболочки (CP1: приблизительно 35 кДа) можно выровнять с частями области ORF2 РНК2 (ак 487-729). Фрагменты белка оболочки из средней полосы с белком (CP2: прибл. 26 кДа) можно выровнять с областью ORF2 РНК2 между ак 730 и 983.
Фрагменты наименьшего белка оболочки (CP3: прибл. 23 кДа) можно выровнять с C-концом ORF2 РНК2 (ак 984-1195). Из этих результатов можно заключить, что кодирующие последовательности трех белков капсида локализованы в ORF2 РНК2 (5,5 т.н.) ToTV, и следовательно, что выделенные молекулы вирусной РНК являются частью частиц вируса ToTV.
Пример 2. Выделение и характеризация возбудителя заболевания Марчитец
В 2003 г. обнаружили растения томата с симптомами, сходными с симптомами, вызванными ToTV, выращенные в Центральной Америке (Мексика и Гватемала). Предположили, что возбудитель заболевания является вирусным. Заболевание в пределах определенной местности известно под названиями «Шоколад», «Марчитец (вирус)» или «Заболевание шоколадной пятнистости». Восприимчивые растения, выращенные в полевых условиях, становились сильно инфицированными с 2003 г. и позже.
Целью настоящего исследования являлось сравнение последовательности до настоящего времени неизвестного вируса, вызывающего «Заболевание шоколадной пятнистости», с последовательностью ToTV, выделенного в примере 1.
Методы
РНК выделяли из приблизительно 100 мкг материала листа, инфицированного «Заболеванием шоколадной пятнистости» с использованием системы для выделения тотальной РНК (Promega SV 96). Два мкл тотальной РНК использовали в 50 мкл реакционной смеси для одноступенчатой реакции Superscript RT-PCR (Invitrogen), где реакционная смесь дополнительно содержит 25 мкл 2× реакционной смеси, 1 мкл (100 нг) как прямого праймера, так и обратного праймера, 1 мкл смеси RT/Taq и 22 мкл воды MilliQ. Чтобы провести обратную транскрипцию РНК в кДНК и амплифицировать эту кДНК, использовали следующую программу RT-PCR: стадия 1: 30 минут при 50°C (реакция обратной транскрипции); стадия 2: 3 минуты при 94°C (активация Taq-полимеразы); стадия 3: 30 секунд при 94°C; стадия 4: 30 секунд при 55°C; стадия 5: 1 минута при 72°C; стадия 6: повтор стадий (3-5) 40×; стадия 7: 10 минут при 72°C; стадия 8: 10°C так долго, как потребуется. Продукты PCR анализировали в 1% агарозном геле в буфере TAE или TBE.
Для RT-PCR использовали различные наборы праймеров, известных как отжигающиеся в различных положениях РНК1 или РНК2 из генома ToTV.
Праймеры для RT-PCR на основе последовательности РНК-2 ToTV, используемые для характеризации вируса-возбудителя заболевания шоколадной пятнистости, как применяли в примере 2 и как показано на Фиг.7
Р1048/1049:
P1056/1057:
P1060/1061:
P1064/1065:
Результаты
Как ожидали, использованные наборы праймеров обеспечивали амплификацию части последовательности ToTV, однако при использовании для RT-PCR четырех различных наборов праймеров, основанных на последовательности РНК-2 (P1048/1049, P1056/1057, P1060/1061 и P1064/1065), получили также амплификацию части генома возбудителя «Заболевания шоколадной пятнистости». Участки отжига этих наборов праймеров проиллюстрированы на фиг. 1.
Продукты RT-PCR для четырех наборов праймеров, с которыми амплифицировали фрагменты возбудителя «Заболевания шоколадной пятнистости», секвенировали, и они обладали 100% идентичностью с соответствующими частями последовательности вирусного генома вируса, описанного в примере 1.
ПОЯСНЕНИЯ К ЧЕРТЕЖАМ
На Фиг.1 показана фотография листьев растения томата с симптомами ToTV-заболевания.
На Фиг.2 показана электронная микрофотография очищенных частиц ToTV. Частицы обладают диаметром приблизительно 28 нм.
На Фиг.3 показан результат окраски серебром геля для PAGE с очищенными верхней (TAgV-T) и нижней (TAgV-B) фракциями вируса ToTV, показывающий три белка капсида приблизительно 23, 26 и 35 кДа.
На Фиг.4 показан результат денатурирующего гель-электрофореза в агарозе. Размеры фрагментов указаны в тысячах нуклеотидов (т.н.). Гель окрашивали с использованием ортотолуидина синего. M: стандарт размера молекул. TAgV: 1 мкг выделенной РНК ToTV.
На Фиг.5 показана полная последовательность молекулы РНК 2 ToTV (SEQ ID NO:1).
На Фиг.6 показана полная последовательность молекулы РНК 1 ToTV (SEQ ID NO:2).
На Фиг.7 показана общая структура вируса ToTV с отмеченными участками отжига различных наборов праймеров, как использовали в примере 2 для РНК 2.
Источники информации
Altschul, S.F., Gish, W., Miller, W., Myers, E.W., Lipman, D.J. (1990). Basic local alignment search tool. J. Mol. Biol., 215:403-410.
Altschul, S.F., Madden, T.L., Schaffer, A.A., Zhang, J., Zhang, Z., Miller, W., Lipman, D.J. (1997). Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucl. Acids Res., 25, 3389-3402.
Ausubel, F.M., Brent, R., Kingston, R.E., Moore, D.D., Seidman, J.G., Smith, J.A., Struhl, K. eds. (1998) Current protocols in molecular biology. V.B. Chanda, series ed. New York: John Wiley & Sons.
Barany, F. (1991) Genetic disease detection and DNA amplification using cloned thermostable ligase. Proc. Natl. Acad. Sci. USA 88: 189-193.
Boom, R., Sol, C.J.A., Salimans, M.M.M., Jansen, C.L., Wertheim-van Dillen, P.M.E., Noordaa, van der J. (1990) Rapid and simple method for purification of nucleic acids. J. Clin. Microbiol. 28: 496-503.
Chomczynski, P., Sacchi, N. (1987) Single-step method of RNA isolation by acid guanidinium thiocyanate-phenol-chloroform extraction. Anal. Bio. 162:156-159.
Clark, M.F., Adams A.N. (1977) Characteristics of the microplate method of enzyme-linked immunosorbent assay for the detection plant viruses. J. Gen. Virol. 34:475-483.
Coligan, J.E., Kruisbeek, A.M., Margulies, D.H., Shevach, E.M. Strober, W. (Eds.) (1997) Current Protocols in Immunology. John Wiley & Sons Inc. Baltimore.
Compton, J. (1991) Nucleic acid sequence-based amplification. Nature 1991, 350:91-92.
Devereux, J., Haeberli, P., Smithies, O. (1984) A comprehensive set of sequence analysis programs for the VAX. Nucleic Acids Res. 12: 387-395.
Dijkstra, J., de Jager, С (Eds.) (1998) Practical Plant Virology - Protocols and Exercises, Springer.
Duffus, J.E., Liu, H.Y., Wisler, G.C. (1996) Tomato infectious chlorosis virus - a new clostero-like virus transmitted by Trialeurodes vaporariorum. European Journal of Plant Pathology, 102(3), 219-226.
Felsenstein, J. 1989. PHYLIP - Phylogeny Inference Package (Version 3.2). Cladistics 5: 164-166.
Gruatelli, J.C., Whitfield, KM., Kwoh, D.Y., Barringer, K. J., Richman, D.D., Gingeras, T.R. (1990) Isothermal, in vitro amplification of nucleic acids by a mutienzyme reaction modeled after retroviral replication. Proc. Natl. Acad. Sci. USA 87:1874-1878.
Hagiwara, K., Ichiki, T.U., Ogawa, Y., Omura, Т., Tsuda, S. (2002). A single amino acid substitution in 126-kDa protein of Pepper mild mottle virus associates with symptom attenuation in pepper; the complete nucleotide sequence of an attenuated strain, C-1421. Arch Virol 147:833-340.
Harlow, E. and Lane, D. (1988) Antibodies: A Laboratory Manual. Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.
Hirata, H., Lu, X., Yamaji, Y., Kagiwada, S., Ugaki, M. Namba, S. (2003) A single silent substitution in the genome of Apple stem grooving virus causes symptom attenuation. J Gen Virol 84:2579-2583.
Jacobs M.V., Snijders P.J., van der Brule A.J. (1997) A general primer (GP5+/GP6+)-mediated PCR-enzyme immunoassay method for rapid detection of 14 high-risk and 8 low-risk human papillomavirus genotypes in cervical scrapings. J Clin Microbiol 35:791-795.
Katz, E., Eksteen, R., Schoenmakers, P., Miller, N. (eds.) (1998) Handbook of HPLC. Marcel Dekker, New York.
Kinter, M., Sherman, N.E. (2000) Protein Sequencing and Identification Using Tandem Mass Spectrometry. Wiley Interscience.
Koonin, E.V. (1991) The phylogeny of RNA-dependent RNA polymerases of positive-strand RNA viruses. J Gen Virol 72:2197-2206.
Kwoh, D.Y., Davis, G.R., Whitefield, K.M., Chappelle, H.L., DiMichele, L.J., Gingeras, T.R. (1989) Transcription-Based Amplification System and Detection of Amplified Human Immunodeficiency Virus Type 1 with a Bead-Based Sandwich Hybridization Format. Proc. Natl Acad. Sci. USA, 86, 1173-1177.
Laemmli, U.K. (1970) Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227, 680-685.
Lerner, R.A., Kang, A.S., Bain, J.D., Burton, D.R., Barbas, C.F. (1992) Antibodies without immunization. Science 258:1313-1314.
Lizardi, P.M., Guerra, C.E., Lomeli, H., Tussie-Luna, I., Kramer, F.R. (1988) Exponential amplification of recombinant RNA hybridization probes. Biotechnology 6, 1197-1202.
Lowman, H.B., Bass, S.H., Simpson, N., Wells, J.A. (1991) Selecting high-affinity binding proteins by monovalent phage display. Biochem. 30(45): 10832-8.
Lu, X., Hirata, H., Yamaji, Y., Ugaki, M., Namaba, S. (2001). Random mutagenesis in a plant viral genome using a DNA repair-deficient mutator Escherichia coli strain. J Virol Methods 94:37-43.
Marks, J.D., Hoogenboom, H.R., Griffiths, A.D., Winter, G. (1992a) Molecular evolution of proteins on filamentous phage. Journal of Biological Chemistry, 267, 16007-16010.
Marks, J.D., Griffiths, A.D., Malmqvist, M., Clackson, T.P., Bye, J.M. Winter, G. (1992b). By passing immunization: building high affinity human antibodies by chain shuffling. Biotechnology 10: 779:783.
Meinkoth J., Wahl G. (1984) Hybridization of nucleic acids immobilized on solid supports. Anal Biochem, 138(2):267-284.
Mullis, K.B., Faloona, F.A. (1987) Specific synthesis of DNA in vitro via a polymerasecatalyzed chain reaction. Meth. Enzymol. 155:335-350.
Pierik, R.L.M. (1999) In vitro Culture of Higher Plants, 4th edition, 360 pages, ISBN: 0-7923-5267-X.
Rose, N., DeMacrio, E., Fahey, J., Friedman, H., Penn, G. (1997) Manual of Clinical Laboratory Immunology. American Soc. Microbiology Press, Washington, D.C
Sambrook, J., Russell D.W., Sambrook, J. (2001) Molecular Cloning: a Laboratory Manual. Cold Spring Harbor Laboratory Press, Plainview, N.Y.
Sanger, F., Nicklen, S., Coulson, A.R. (1977) DNA sequencing with chain-terminating inhibitors. Proc. Natl. Acad. Sci. U.S.A. 74;5463-5467.
Schagger H., von Jagow, G. (1987) Tricine-sodium dodecyl sulfate-polyacrylamide gel electrophoresis for the separation of proteins in the range from 1 to 100 kDa. Analytical Biochemistry 166, 368-379.
Smith, L.M., Sanders, J.Z., Kaiser, R.J., Hughs, P., Dodd, C, Connell, C.R., Heins, C., Kent, S.B.H., Hood, L.E. (1986) Fluorescent detection in automated DNA sequence analysis. Nature 321:673-681.
Swofford, D.L. (2000). PAUP*: phylogenetic analysis using parsimony (*and other methods). 4th edition. Sinauer Associates, Sunderland, Mass.
Takeshita, M., Suzuki, M., Takanami, Y. (2001). Combination of amino acids in the 3a protein and the coat protein of Cucumber mosaic virus determines symptom expression and viral spread in bottle gourd. Arch Virol 146:697-711.
Thompson, J.D., Gibson, T.J., Plewniak, F., Jeanmougin, F., Higgins, D.G. (1997) The ClustalX windows interface: flexible strategies for multiple sequence alignment aided by quality analysis tools. Nucleic Acids Research, 24:4876-4882.
Thompson, J.R., Perry, K.L., De Jong, W. (2004a) A new potato virus in a new lineage of picorna-like viruses. Arch Virol. 149:2141-2154.
Thompson, J.R., Perry, K.L., De Jong, W. (2004b) The characterization of a new picorna-like virus infecting tomato. Abstract book of the 23rd Annual Meeting of the American Society for Virology, Montreal, Canada, July 2004.
Tijssen, P. 1993. Laboratory Techniques in Biochemistry and Molecular Biology-Hybridization with Nucleic Acid Probes, Part I, Chapter 2 "Overview of principles of hybridization and the strategy of nucleic acid probe assays", Elsevier. New York.
Van den Brule, A.J.C., Pol, R., Fransen-Daalmeijer, N., Schouls, L.M., Meijer, С.J.L.M., Snijders, P.J.F. (2002) GP5+/6+ PCR followed by Reverse Line Blot Analysis Enables Rapid and High-Throughput Identification of Human Papillomavirus Genotypes. J. Clin. Microbiol. 40:779-787.
Vela, C, Cambra, M., Cortes, E., Moreno, P., Miguet, J.G., Perez de San Roman, С., Sanz, A. (1986) Production and characterization of monoclonal antibodies specific for citrus tristeza virus and their use for diagnosis. J. Gen. Virol. 67:91-96.
Walker, G.T., Fraiser, M.S., Schram, J.L., Little, M.C., Nadeau, J.G., Malinowski, D.P. (1992) Strand displacement amplification - an isothermal, in vitro DNA amplification technique Nucleic Acids Res 20:1691-1696.
Walker, J.M. 2004. PCR Protocols on CD. Humana Press.
Wisler, G.C., Li, R.H., Liu, H.Y., Lowry, D.S., Duffus, J.E. (1998) Tomato chlorosis virus: a new whitefly-transmitted, phloem-limited, bipartite closterovirus of tomato. Phytopathology, 88(5), 402-409.
Young R.A., Davis, R.W. (1983) Efficient isolation of genes using antibody probes. Proc. Natl. Acad. Sci. U.S.A. 80:1194-1198.
| название | год | авторы | номер документа |
|---|---|---|---|
| НОВЫЙ ПТИЧИЙ АСТРОВИРУС | 2009 |
|
RU2547589C2 |
| СПОСОБ БЫСТРОГО ОТБОРА ВАРИАНТОВ GP-120 ВИЧ | 2012 |
|
RU2603732C2 |
| СПЕЦИФИЧНЫЕ В ОТНОШЕНИИ ВИРУСА БЕШЕНСТВА НЕЙТРАЛИЗУЮЩИЕ МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА ЧЕЛОВЕКА И НУКЛЕИНОВЫЕ КИСЛОТЫ И СВЯЗАННЫЕ С НИМИ СПОСОБЫ | 2001 |
|
RU2272809C2 |
| ВИРУСЫ PRRS, ИХ ИНФЕКЦИОННЫЕ КЛОНЫ, МУТАНТНЫЕ ФОРМЫ И СПОСОБЫ ПРИМЕНЕНИЯ | 2006 |
|
RU2427646C2 |
| МАТЕРИАЛЫ И МЕТОДЫ, ИСПОЛЬЗУЕМЫЕ ДЛЯ ЛЕЧЕНИЯ РЕСПИРАТОРНЫХ ЗАБОЛЕВАНИЙ У СОБАК | 2011 |
|
RU2711791C2 |
| ШТАММ ВИРУСА ГРИППА А СОБАК (ВАРИАНТЫ), ИММУНОГЕННЫЙ ПОЛИПЕПТИД, ПОЛИНУКЛЕОТИД, КОДИРУЮЩИЙ ЕГО, ВЕКТОР ЭКСПРЕССИИ ПОЛИПЕПТИДА, ИММУНОГЕННАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ ПОЛИПЕПТИД, И СПОСОБ ИНДУКЦИИ ИММУННОГО ОТВЕТА У ЖИВОТНОГО | 2006 |
|
RU2449014C2 |
| ОЛИГОМЕР (ВАРИАНТЫ), СПОСОБ ОБНАРУЖЕНИЯ ПОСЛЕДОВАТЕЛЬНОСТИ HCV (ВАРИАНТЫ), НАБОР ДЛЯ ОБНАРУЖЕНИЯ, СПОСОБ ПОЛУЧЕНИЯ КРОВИ | 1990 |
|
RU2145635C1 |
| СПОСОБ ВЫЯВЛЕНИЯ ПАРВОВИРУСА B19 ЧЕЛОВЕКА В БИОЛОГИЧЕСКОМ ОБРАЗЦЕ (ВАРИАНТЫ) И НАБОР ДЛЯ ЕГО ОСУЩЕСТВЛЕНИЯ | 2002 |
|
RU2301263C2 |
| КОМПОЗИЦИИ, РЕАГЕНТЫ, НАБОРЫ И СПОСОБЫ ДЛЯ ДИАГНОСТИКИ, МОНИТОРИНГА И ЛЕЧЕНИЯ ОЖИРЕНИЯ И/ИЛИ ДИАБЕТА | 2004 |
|
RU2420583C2 |
| ВИРУС ГРИППА, СПОСОБНЫЙ ИНФИЦИРОВАТЬ СОБАЧЬИХ, И ЕГО ПРИМЕНЕНИЕ | 2014 |
|
RU2711807C2 |

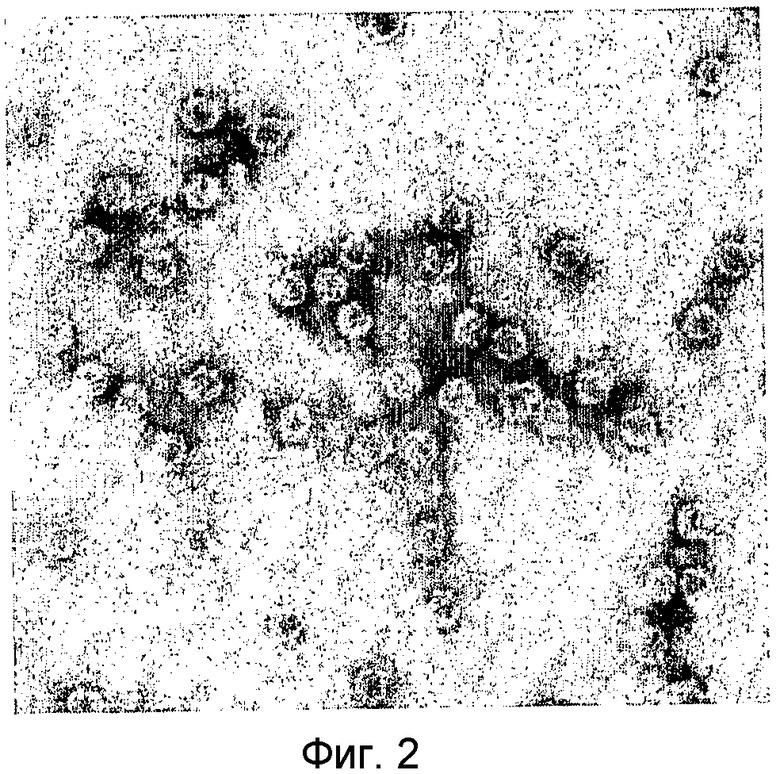
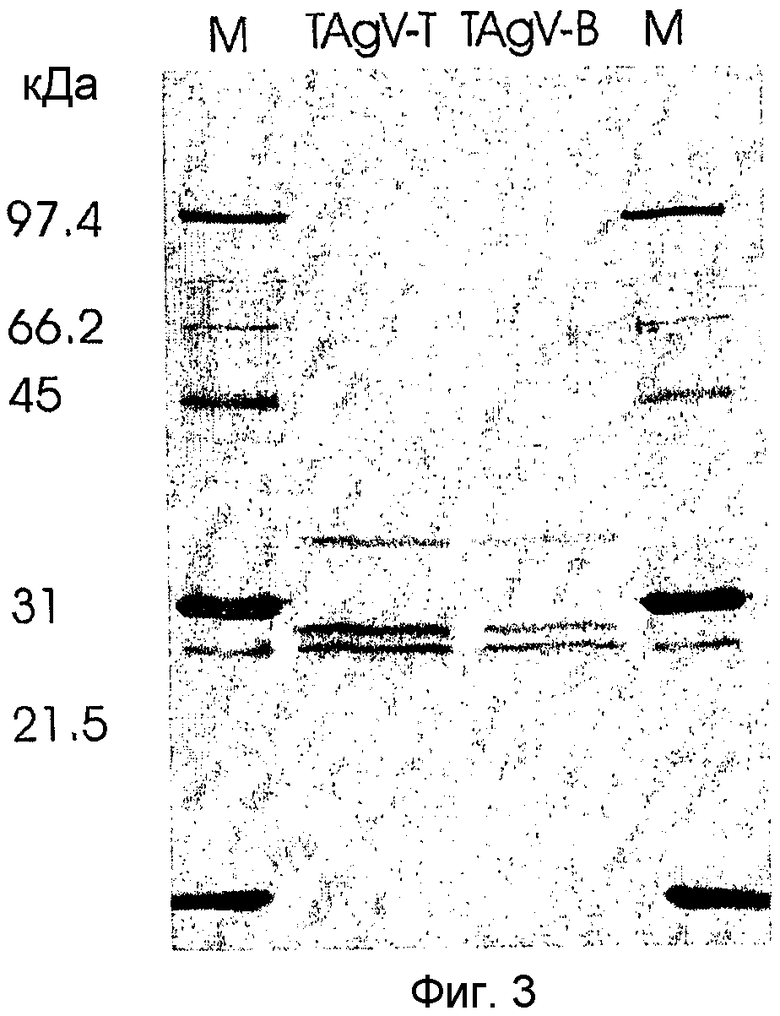






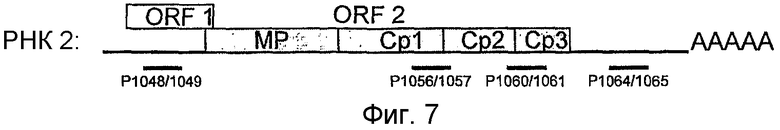
Изобретение относится к выделенному вирусу растений, названному вирус томата торрадо (ToTV), и его компонентам, а также к способам получения устойчивых к ToTV растений. Сначала на растение или его часть воздействуют инфекционной дозой ToTV. Затем идентифицируют растения, у которых отсутствуют или уменьшены симптомы заболевания. Растения, идентифицированные таким образом как устойчивые к вирусу, используют в качестве донорных при скрещивании с реципиентными растениями, и из растений-потомков отбирают устойчивые к ToTV. 6 н. и 2 з.п. ф-лы, 5 табл., 7 ил.
1. Вирус растений, названный вирусом томата торрадо (ToTV), содержащий по меньшей мере одну последовательность нуклеиновой кислоты, выбранную из группы, состоящей из SEQ ID NO:1 и SEQ ID NO:2, и последовательностей, обладающих по меньшей мере 50%-ной гомологией нуклеотидной последовательности с ними, где указанный вирус является агентом, вызывающим заболевание томатов, обозначенное как заболевание торрадо.
2. Вирус растений по п.1, депонированный в немецкой коллекции микроорганизмов и клеточных культур 24 ноября 2004 под номером для ссылки депозитов ToTV-E01 (DSM 16999).
3. Выделенная или рекомбинантная нуклеиновая кислота ToTV или ее ToTV-специфический фрагмент, содержащие последовательность нуклеиновой кислоты, выбранную из группы, состоящей из SEQ ID NO:1, SEQ ID NO:2, последовательностей, обладающих по меньшей мере 50%-ной гомологией нуклеотидной последовательности с SEQ ID NO:1 или SEQ ID NO:2, комплементарных им цепей и их ToTV-специфических фрагментов, где указанная нуклеиновая кислота способна инфицировать растения и/или используется для филогенетического анализа.
4. Полинуклеотид, способный гибридизоваться в строгих условиях с нуклеиновой кислотой по п.3.
5. Антигенный препарат, полученный из белка вируса ToTV по п.1 или 2, выбранного из группы, состоящей из белков капсида 23, 26 и 35 кДа, геликазы, РНК-зависимой РНК-полимеразы, предположительного двигательного белка (MP) ToTV и их ToTV-специфических фрагментов.
6. Способ идентификации устойчивого к ToTV растения, включающий стадии:
a) воздействия на растение или часть растения инфекционной дозой ToTV и
b) идентификации указанного растения как устойчивого к ToTV, когда после указанного воздействия или
симптомы заболевания в указанном растении или части растения остаются отсутствующими, или их проявление задержано, или, по меньшей мере, тяжесть уменьшена, или они являются локализованными по сравнению с чувствительным контрольным растением, и/или
вирус ToTV или геномные последовательности ToTV не присутствуют в указанном растении или части растения, или присутствие вируса ToTV является по меньшей мере уменьшенным количественно по сравнению с чувствительным контрольным растением.
7. Способ по п.6, где на стадии b) присутствие ToTV в указанном растении или части растения определяют путем определения в образце присутствия вируса ToTV или его компонента посредством реакции указанного образца с полинуклеотидом по п.4.
8. Устойчивое к ToTV растение или его часть, полученное способом, включающим стадии:
a) идентификации устойчивого к ToTV донорного растения способом по п.6 или 7,
b) скрещивания указанного устойчивого к ToTV донорного растения с реципиентным растением и
c) отбора из растений-потомков устойчивого растения способом по п.6 или 7.
| DATABASE EMBL, accession no | |||
| Губная помада | 1978 |
|
SU829112A1 |
| DATABASE EMBL, accession no | |||
| LERTUPOLY, 18.06.1992 | |||
| KR 20010086888 A, 15.09.2001 | |||
| CN 1424326 A, 18.06.2003. | |||

