ССЫЛКИ НА РОДСТВЕННЫЕ ЗАЯВКИ
Настоящая заявка заявляет приоритет предварительной заявки на патент США No. 60/673 443, зарегистрированной 21 апреля 2005 года, которая включена в настоящее описание полностью в виде ссылки, включая все приведенные в ней чертежи, таблицы, последовательности нуклеиновых кислот, последовательности аминокислот и рисунки.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
"Кашель в условиях конуры", или инфекционный трахеобронхит (ITB), представляет собой острую контагиозную респираторную инфекцию у собак, характеризующуюся, в основном, кашлем (Ford et al, 1998). ITB собак рассматривается как наиболее распространенное в мире инфекционное респираторное заболевание собак, вспышки которого могут достичь масштабов эпидемии, если собаки содержатся в условия высокой плотности животных, какие имеются, например, в конурах. Большая часть таких вспышек заболевания связана с непосредственным контактом собак или определяется аэрозольным распространением респираторных выделений (Ford et al, 1998). Клинические признаки вызываются инфицированием одним инфекционным агентом или сочетанием бактериальных и вирусных агентов, которые населяют эпителий верхнего и нижнего отделов дыхательных путей. Вирус парагриппа собак (CPiV) и бактерии Bordetella bronchiseptica представляют организмы, чаще всего выделяемые из организма больных животных, однако на клиническое течение и исход заболевания могут оказать влияние и некоторые другие вирусы, такие как вирус чумы собак (CDV), аденовирус-1 и -2 собак (CAV-1, CAV-2), вместе с бактериями, такими как Streptococcus sp., Pasteurella multicoda и Escherichia coli (Ford et al, 1998). Несмотря на то, что вспышки заболевания протекают выраженно и быстро, с высоким уровнем заболеваемости, в популяциях с высокой плотностью проживания животных, осложненные респираторные инфекции и летальные исходы встречаются не часто. Хотя могут развиваться опасные для жизни вторичные бактериальные пневмонии, в большинстве случаев ITB являются самокупирующимися и разрешаются без какого-либо лечения (Ford et al, 1998).
В июле 1992 респираторная инфекция, которую, предположительно, отнесли к "кашлю в конуре", достигла эпидемических масштабов на нескольких участках выгула борзых собак в Новой Англии, Флориде, Западной Вирджинии, Висконсине, Канзасе, Колорадо, Оклахоме и Аризоне. Согласно заключению ветеринаров, большая часть пораженных собак имела умеренный кашель, который проходил, но более чем у десятка борзых развилась острая геморрагическая пневмония, которая привела к быстрой смерти животных (Greyhound Daily News, 1999).
С конца 1998 до начала 1999 несколько вспышек заболевания по типу "кашель в условиях конуры", возникшие по всей стране на скачках борзых, содержавшихся в конурах, привели к принудительному закрытию треков и введению карантина на всех соревнованиях борзых в США на несколько недель (Greyhound Daily News, 1999). На одном треке во Флориде (Palm Beach Kennel Club) в один день кашель был зарегистрирован примерно у 40% собак (личное сообщение от д-ра William Duggar). Как и в случае вспышки заболевания в 1992 году, кашель проходил у большинства борзых, но 10 собак во Флориде умерли от синдрома геморрагической пневмонии, нехарактерной для инфекции по типу "кашля в конуре" (Putnam, 1999).
В марте-апреле 2003 другая вспышка "кашля в конуре" возникла на участках выгула борзых на востоке США. Эта вспышка, которая, предположительно, возникла в конурах на четырех треках во Флориде, вызвала отмену скачек и карантин собак почти на три недели. Были поражены около 25% собак на треке в Западном Пальм-Бич, и почти у 50% из 1400 собак на озере Дерби в Санкт-Петербурге развился кашель. И, как и прежде, большая часть собак выздоровела, но несколько собак умерло от респираторной инфекции. Экономический ущерб вспышки респираторной инфекции только на треке в районе озера Дерби составил, по оценкам, 2 миллиона долларов.
В настоящее время отсутствуют документальные отчеты с описанием этиологии или клинической картины эпидемии "кашля в конуре", возникшей в собачьих конурах во время скачек борзых в 1992, 1998-1999 или 2003 гг. Было высказано предположение о связи данной инфекции с CPiV и/или B. bronchiseptica, которые являются наиболее частой причиной кашля в конуре. Не подкрепленные соответствующими обоснованиями сообщения, типа информации, появляющейся на веб-сайтах, связывают фатальные случаи геморрагической пневмонии, развившейся у некоторых кашляющих собак, с инфекцией β-гемолитическим стрептококком Streptococcus equi, подвид zooepidemicus, и относят эти случаи к синдрому так называемого "стрептококкового токсического шока собак".
Передача вируса от одного организма-хозяина к другому является ключевым моментом экологии и эпидемиологии вируса гриппа (Webster, 1998). Возможны два основных механизма межвидовой передачи вируса гриппа (Webster et al., 1992; Lipatov et al., 2004). Один из них относится к непосредственной передаче по существу не измененного вируса от одного вида к другому. Примеры такого механизма включают идентифицированные недавно инфекции человека вирусом птичьего гриппа подтипа H5N1 (Subbarao et al., 1998; Peiris et al., 2004; Guan et al., 2004) и, возможно, также пандемию 1918 года, известную как «испанка» (Reid et al., 2004). Второй механизм имеет отношение к сегментированной природе генома вируса гриппа. Совместная инфекция организма-хозяина вирусами из разных видов может привести к перегруппировке сегментированных вирусных генов и образованию рекомбинанта, который способен инфицировать другой вид. Так, например, новые вирусы, образующиеся при генной перегруппировке между вирусами птичьего гриппа и гриппа человека, привели к пандемии гриппа у людей в 1957 и 1968 гг. (Webster et al., 1992; Lipatov et al., 2004; Kawaoka et al., 1989).
В основном, непосредственная передача не измененных вирусов гриппа от природного организма-хозяина к другому виду представляет собой завершенный процесс, поскольку длительная передача между индивидуумами нового вида не происходит. Множественные взаимодействия по типу вирус-организм-хозяин необходимы для репликации и горизонтальной передачи и обеспечивают непреодолимый барьер для длительного пребывания вирусов гриппа в новом организме-хозяине (Webby et al., 2004). В этой связи установление новых линий вируса гриппа, специфичных для организма-хозяина, является необычным процессом и происходит только на птицеводческих фермах, в свинарниках, конюшнях и в человеческих сообществах (Webster et al., 1992; Lipatov et al., 2004).
В связи с серьезностью инфекции вирусом гриппа остается потребность в способах диагностики, профилактики и лечения инфекции вирусом гриппа.
КРАТКОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к выделенному вирусу гриппа, который способен инфицировать представителей группы псовых и вызывать респираторное заболевание у представителей группы псовых. Настоящее изобретение также относится к композициям и способам, подходящим для индукции иммунного ответа против вируса гриппа согласно настоящему изобретению. Настоящее изобретение также относится к композициям и способам, подходящим для идентификации вируса согласно настоящему изобретению, и диагностики наличия у животного инфекции вирусом согласно настоящему изобретению.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Патент или зарегистрированная заявка содержит по меньшей мере один рисунок, выполненный в цвете. Копии данного патента или публикация по патентной заявке с цветными рисунками будут предоставлены Патентным Бюро при направлении соответствующего запроса и оплате необходимой суммы.
Фиг.1A-1B иллюстрируют филогенетическое родство, имеющееся между генами гемагглютинина. На фиг.1A показано дерево генов HA из репрезентативных изолятов из организма собаки, человека, птицы, свиньи и лошади, включая A/Budgerigar/Hokkaido/1/77 (H4), как варианта за пределами рассматриваемой группы. На фиг.1B показано дерево генов HA из вируса гриппа собаки, в сочетании с современными и более древними генами НА лошади A/Duck/Ukraine/63 (H3), как варианта за пределами рассматриваемой группы. Филогенетические деревья были построены на основе нуклеотидных последовательностей, определенных по методу «ближайших соседей», и показаны результаты анализа по методу «расшнурованной выборки» ≥90%. Стрелка указывает число нуклеотидных изменений на единицу длины горизонтальных ветвей дерева.
На фиг.2A-2B проиллюстрировано выявление иммуногистохимическим методом вирусного антигена H3 в легких. Срезы из ткани легкого зондируют мышиным моноклональным антителом против H3 гемагглютинина и выявляют наличие связывания по результатам иммунопероксидазной реакции (коричневый осадок). На фиг.2A показан бронхиальный эпителий, взятый от борзой со спонтанным заболеванием. Вирусный антиген H3 выявляется в цитоплазме эпителиальных клеток бронхов и в макрофагах в просвете дыхательных путей и в альвеолярном пространстве. На фиг.2B показан бронхиальный эпителий, взятый у собаки через 5 дней после инокуляции A/canine/Florida/43/04 (H3N8). Вирусный антиген H3 выявляется в цитоплазме эпителиальных клеток бронхов. Масштаб: 66 мкм.
На фиг.3 показаны характерные гистологические изменения в бронхах борзых, умерших от геморрагической пневмонии, ассоциированной с инфекцией вирусом гриппа. Ткани окрашивают красителем H&E. На верхней панели показаны: нормальный бронх с реснитчатыми клетками, слизистыми клетками и базальными клетками. На нижней панели показаны: бронх от борзой со спонтанным заболеванием. Отмечается некроз и эрозия реснитчатых клеток бронхиального эпителия. Масштаб: 100 мкм.
На фиг.4A-4B проиллюстрировано филогенетическое родство между генами H3 гемагглютинина. На фиг.4A показано филогенетическое дерево генов НА вируса гриппа собаки вместе с современными и более древними генами НА лошади. На фиг.4B показано филогенетическое дерево белка НА вируса гриппа собаки, вместе с современными и более древними НА лошади. Филогенетические деревья построены на основе нуклеотидных последовательностей, определенных по методу «ближайших соседей», и показаны результаты анализа по методу «расшнурованной выборки» ≥80%. Стрелка указывает число нуклеотидных изменений на единицу длины горизонтальных ветвей дерева.
На фиг.5 показан белок H3 из вируса гриппа в эпителиальных клетках бронхов и бронхиальных желез в легких собак, которые умерли от пневмонии, ассоциированной с инфекцией вирусом гриппа. На верхней панели показаны: эрозия реснитчатых клеток бронхиального эпителия в бронхах. Ткани окрашивают красителем H&E. На нижней панели показаны: белок H3 из вируса гриппа в цитоплазме эпителиальных клеток бронхов (слева) и бронхиальной железы (справа). Ткани окрашивают с использованием моноклонального антитела против H3 вируса гриппа, при выявлении результатов иммунопероксидазной реакцией (коричневый осадок) и при проведении контрастного окрашивания гематоксилином.
На фиг.6A-6D приведены графики, описывающие амплификацию H3 и Matrix генов (фиг.6A и фиг.6B), полученных при амплификации 10-кратных серийных разведений транскрибированных in vitro стандартных препаратов РНК. Стандартные кривые для H3 и Matrix генов (фиг.6C и фиг.6D) получают при построении графика зависимости log исходных концентраций РНК от порогового значения цикла (Ct), полученного для каждого разведения.
На фиг.7 проиллюстрирована чувствительность Directigen Flu A, при анализе 10-кратных серийных разведений стандартных вирусных препаратов, включая A/Wyoming/3/2003 и A/canine/FL/242/2003. Фиолетовый треугольник указывает на положительный результат.
КРАТКОЕ ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
SEQ ID NO: 1 представляет собой нуклеотидную последовательность из вируса гриппа собаки (Florida/43/04), кодирующую белок PB2, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 2 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 1.
SEQ ID NO: 3 представляет собой нуклеотидную последовательность из вируса гриппа собаки (Florida/43/04), кодирующую белок PB1, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 4 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 3.
SEQ ID NO: 5 представляет собой нуклеотидную последовательность из вируса гриппа собаки (Florida/43/04), кодирующую белок PA, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 6 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 5.
SEQ ID NO: 7 представляет собой нуклеотидную последовательность из вируса гриппа собаки (Florida/43/04), кодирующую белок NS, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 8 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 7.
SEQ ID NO: 9 представляет собой нуклеотидную последовательность из вируса гриппа собаки (Florida/43/04), кодирующую белок NP, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 10 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 9.
SEQ ID NO: 11 представляет собой нуклеотидную последовательность из вируса гриппа собаки (Florida/43/04), кодирующую белок NA, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 12 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 11.
SEQ ID NO: 13 представляет собой нуклеотидную последовательность из вируса гриппа собаки (Florida/43/04), кодирующую белок МА, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 14 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 13.
SEQ ID NO: 15 представляет собой нуклеотидную последовательность из вируса гриппа собаки (Florida/43/04), кодирующую белок НА, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 16 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 15.
SEQ ID NO: 17 представляет собой нуклеотидную последовательность из вируса гриппа собаки (FL/242/03), кодирующую белок РВ2, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 18 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 17.
SEQ ID NO: 19 представляет собой нуклеотидную последовательность из вируса гриппа собаки (FL/242/03), кодирующую белок РВ1, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 20 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 19.
SEQ ID NO: 21 представляет собой нуклеотидную последовательность из вируса гриппа собаки (FL/242/03), кодирующую белок РА, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 22 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 21.
SEQ ID NO: 23 представляет собой нуклеотидную последовательность из вируса гриппа собаки (FL/242/03), кодирующую белок NS, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 24 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 23.
SEQ ID NO: 25 представляет собой нуклеотидную последовательность из вируса гриппа собаки (FL/242/03), кодирующую белок NP, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 26 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 25.
SEQ ID NO: 27 представляет собой нуклеотидную последовательность из вируса гриппа собаки (FL/242/03), кодирующую белок NA, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 28 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 27.
SEQ ID NO: 29 представляет собой нуклеотидную последовательность из вируса гриппа собаки (FL/242/03), кодирующую белок МА, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 30 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 29.
SEQ ID NO: 31 представляет собой нуклеотидную последовательность из вируса гриппа собаки (FL/242/03), кодирующую белок МА, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 32 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 31.
SEQ ID NO: 33 представляет собой зрелую форму белка HA, показанного в виде SEQ ID NO: 16, в котором удалена N-концевая сигнальная последовательность из 16 аминокислот.
SEQ ID NO: 34 представляет собой зрелую форму белка HA, показанного в виде SEQ ID NO: 32, в котором удалена N-концевая сигнальная последовательность из 16 аминокислот.
SEQ ID NO: 35 представляет собой олигонуклеотид, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 36 представляет собой олигонуклеотид, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 37 представляет собой олигонуклеотид, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 38 представляет собой олигонуклеотид, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 39 представляет собой олигонуклеотид, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 41 представляет собой олигонуклеотид, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 42 представляет собой олигонуклеотид, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 43 представляет собой олигонуклеотид, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 44 представляет собой олигонуклеотид, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 45 представляет собой олигонуклеотид, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 46 представляет собой олигонуклеотид, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 47 представляет собой нуклеотидную последовательность из вируса гриппа собаки (Miami/2005), кодирующую белок PB2, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 48 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 47.
SEQ ID NO: 49 представляет собой нуклеотидную последовательность из вируса гриппа собаки (Miami/2005), кодирующую белок PB1, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 50 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 49.
SEQ ID NO: 51 представляет собой нуклеотидную последовательность из вируса гриппа собаки (Miami/2005), кодирующую белок PA, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 52 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 51.
SEQ ID NO: 53 представляет собой нуклеотидную последовательность из вируса гриппа собаки (Miami/2005), кодирующую белок NS, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 54 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 53.
SEQ ID NO: 55 представляет собой нуклеотидную последовательность из вируса гриппа собаки (Miami/2005), кодирующую белок NP, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 56 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 55.
SEQ ID NO: 57 представляет собой нуклеотидную последовательность из вируса гриппа собаки (Miami/2005), кодирующую белок NA, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 58 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 57.
SEQ ID NO: 59 представляет собой нуклеотидную последовательность из вируса гриппа собаки (Miami/2005), кодирующую белок МА, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 60 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 59.
SEQ ID NO: 61 представляет собой нуклеотидную последовательность из вируса гриппа собаки (Miami/2005), кодирующую белок НА, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 62 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 61.
SEQ ID NO: 63 представляет собой нуклеотидную последовательность из вируса гриппа собаки (Jacksonville/2005), кодирующую белок РВ2, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 64 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 63.
SEQ ID NO: 65 представляет собой нуклеотидную последовательность из вируса гриппа собаки (Jacksonville/2005), кодирующую белок РВ1, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 66 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 65.
SEQ ID NO: 67 представляет собой нуклеотидную последовательность из вируса гриппа собаки (Jacksonville/2005), кодирующую белок РА, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 68 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 67.
SEQ ID NO: 69 представляет собой нуклеотидную последовательность из вируса гриппа собаки (Jacksonville/2005), кодирующую белок NS, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 70 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 69.
SEQ ID NO: 71 представляет собой нуклеотидную последовательность из вируса гриппа собаки (Jacksonville/2005), кодирующую белок NP, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 72 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 71.
SEQ ID NO: 73 представляет собой нуклеотидную последовательность из вируса гриппа собаки (Jacksonville/2005), кодирующую белок NA, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 74 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 73.
SEQ ID NO: 75 представляет собой нуклеотидную последовательность из вируса гриппа собаки (Jacksonville/2005), кодирующую белок МА, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 76 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 75.
SEQ ID NO: 77 представляет собой нуклеотидную последовательность из вируса гриппа собаки (Jacksonville/2005), кодирующую белок НА, который может использоваться в соответствии с настоящим изобретением.
SEQ ID NO: 78 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 77.
SEQ ID NO: 79 представляет собой олигонуклеотид, который может использоваться согласно настоящему изобретению.
SEQ ID NO: 80 представляет собой олигонуклеотид, который может использоваться согласно настоящему изобретению.
SEQ ID NO: 81 представляет собой олигонуклеотид, который может использоваться согласно настоящему изобретению.
SEQ ID NO: 82 представляет собой олигонуклеотид, который может использоваться согласно настоящему изобретению.
SEQ ID NO: 83 представляет собой олигонуклеотид, который может использоваться согласно настоящему изобретению.
SEQ ID NO: 84 представляет собой олигонуклеотид, который может использоваться согласно настоящему изобретению.
SEQ ID NO: 85 представляет собой олигонуклеотид, который может использоваться согласно настоящему изобретению.
SEQ ID NO: 86 представляет собой олигонуклеотид, который может использоваться согласно настоящему изобретению.
SEQ ID NO: 87 представляет собой олигонуклеотид, который может использоваться согласно настоящему изобретению.
SEQ ID NO: 88 представляет собой олигонуклеотид, который может использоваться согласно настоящему изобретению.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к выделенному вирусу гриппа, который способен инфицировать псовых и вызывать респираторное заболевание. В одном варианте вирус гриппа согласно настоящему изобретению включает полинуклеотид, который кодирует белок, имеющий аминокислотную последовательность, показанную в виде любой из последовательностей SEQ ID No: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 33, 34, 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70, 72, 74, 76 или 78, или их функциональный и/или иммуногенный фрагмент или вариант. В конкретных вариантах осуществления настоящего изобретения указанный полинуклеотид включает нуклеотидную последовательность, показанную в виде любой из последовательностей SEQ ID No: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 47, 49, 51, 53, 55, 57, 59, 61, 63, 65, 67, 69, 71, 73, 75 или 77, или их фрагмент или вариант. В конкретном варианте осуществления настоящего изобретения вирус гриппа согласно настоящему изобретению относится к подтипу H3. Вирус может быть выделен от инфицированных собак и из культуральных клеток или яиц, в соответствии с приведенными в настоящем описании методами. В репрезентативном варианте осуществления настоящего изобретения вирус гриппа представляет собой вирус гриппа A.
Настоящее изобретение также относится к полинуклеотидам, которые включают полностью или часть одного или нескольких генов или геномного сегмента из вируса гриппа согласно настоящему изобретению. В одном варианте полинуклеотид согласно настоящему изобретению включает ген гемагглютинина вируса (HA), ген нейраминидазы (NA), ген нуклеопротеина (NP), ген матричного белка (MA или M), ген белка щелочной полимеразы (PB), ген белка кислой полимеразы (PA), ген неструктурного белка (NS) или функциональный фрагмент или вариант любого из указанных генов. В конкретном варианте осуществления настоящего изобретения полинуклеотид согласно настоящему изобретению включает ген гемагглютинина (HA) или его функциональный фрагмент или вариант. В другом варианте осуществления настоящего изобретения ген HA кодирует белок гемагглютинин, включающий одну или несколько следующих характеристик: серин в положении 83; лейцин в положении 222; треонин в положении 328 и/или треонин в положении 483, в сравнении с аминокислотной последовательностью консенсусной последовательности лошади H3. В одном варианте осуществления настоящего изобретения ген HA кодирует полипептид, имеющий аминокислотную последовательность, показанную в виде SEQ ID No: 16, 32, 62 или 78, или их функциональный и/или иммуногенный фрагмент или вариант. В конкретном варианте осуществления настоящего изобретения ген HA включает нуклеотидную последовательность, показанную в виде SEQ ID No: 15, 31, 61 или 77.
В одном варианте осуществления настоящего изобретения полинуклеотид согласно настоящему изобретению кодирует полипептид, имеющий аминокислотную последовательность, показанную в виде SEQ ID No: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 33, 34, 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70, 72, 74, 76 или 78, или их функциональный и/или иммуногенный фрагмент или вариант. В конкретном варианте осуществления настоящего изобретения полинуклеотид, кодирующий аминокислотную последовательность, показанную в виде SEQ ID No: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 33, 34, 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70, 72, 74, 76 или 78, включает нуклеотидную последовательность, показанную в виде SEQ ID No: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 47, 49, 51, 53, 55, 57, 59, 61, 63, 65, 67, 69, 71, 73, 75 или 77, соответственно, или последовательность, кодирующую функциональный и/или иммуногенный фрагмент или вариант любой из SEQ ID No: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 33, 34, 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70, 72, 74, 76 или 78. Таким образом, настоящее изобретение относится к полинуклеотидным последовательностям, включающим нуклеотидную последовательность, показанную в виде любой из SEQ ID No: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 47, 49, 51, 53, 55, 57, 59, 61, 63, 65, 67, 69, 71, 73, 75 или 77, или фрагмент или вариант, включая вырожденный вариант, любой из SEQ ID No: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 47, 49, 51, 53, 55, 57, 59, 61, 63, 65, 67, 69, 71, 73, 75 или 77. В другом конкретном варианте осуществления настоящего изобретения полинуклеотид согласно настоящему изобретению может включать: нуклеотиды 1-2271 из SEQ ID NO: 3; нуклеотиды 1-2148 из SEQ ID NO: 5; нуклеотиды 1-657 из SEQ ID NO: 7; нуклеотиды 1-1494 из SEQ ID NO: 9; нуклеотиды 1-1410 из SEQ ID NO: 11; нуклеотиды 1-756 из SEQ ID NO: 13; нуклеотиды 1-1695 из SEQ ID NO: 15; нуклеотиды 1-2271 из SEQ ID NO: 19; нуклеотиды 1-2148 из SEQ ID NO: 21; нуклеотиды 1-657 из SEQ ID NO: 23; нуклеотиды 1-1494 из SEQ ID NO: 25; нуклеотиды 1-756 из SEQ ID NO: 29; нуклеотиды 1-1695 из SEQ ID NO: 31; нуклеотиды 1-2277 из SEQ ID NO: 47; нуклеотиды 1-2271 из SEQ ID NO: 49; нуклеотиды 1-2148 из SEQ ID NO: 51; нуклеотиды 1-690 из SEQ ID NO: 53; нуклеотиды 1-1494 из SEQ ID NO: 55; нуклеотиды 1-1410 из SEQ ID NO: 57; нуклеотиды 1-756 из SEQ ID NO: 59; нуклеотиды 1-1695 из SEQ ID NO: 61; нуклеотиды 1-2277 из SEQ ID NO: 63; нуклеотиды 1-2271 из SEQ ID NO: 65; нуклеотиды 1-2148 из SEQ ID NO: 67; нуклеотиды 1-690 из SEQ ID NO: 69; нуклеотиды 1-1494 из SEQ ID NO: 71; нуклеотиды 1-1410 из SEQ ID NO: 73; нуклеотиды 1-756 из SEQ ID NO: 75; и нуклеотиды 1-1695 из SEQ ID NO: 77. Нуклеотидные и аминокислотные последовательности вирусных полинуклеотидных и полипептидных последовательностей, входящие в область настоящего изобретения, депонированы в GenBank с номерами доступа No. DQ124147 - DQ124161 и DQ124190, полное раскрытие которых приведено в настоящем описании в качестве ссылки.
Настоящее изобретение также относится к полипептидам, кодируемым полинуклеотидами вируса гриппа согласно настоящему изобретению. Настоящее изобретение также относится к функциональным и/или иммуногенным фрагментам и вариантам рассматриваемых полипептидов. Рассматриваемые полипептиды включают белок HA, белок NA, белок NS, нуклеопротеин, белок щелочной полимеразы, белок кислой полимеразы и матричный белок вируса гриппа согласно настоящему изобретению. В репрезентативном варианте, полипептид согласно настоящему изобретению имеет аминокислотную последовательность, показанную в виде любой из SEQ ID No: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 33, 34, 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70, 72, 74, 76 или 78, или их функциональный и/или иммуногенный фрагмент или вариант.
Настоящее изобретение также относится к полинуклеотидным экспрессирующим конструкциям, включающим полинуклеотидную последовательность согласно настоящему изобретению. В одном варианте осуществления настоящего изобретения рассматриваемая в нем экспрессирующая конструкция включает полинуклеотидную последовательность, кодирующую полипептид, включающий аминокислотную последовательность, показанную в виде любой из SEQ ID No: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 33, 34, 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70, 72, 74, 76 или 78, или их функциональный и/или иммуногенный фрагмент или вариант. В конкретном варианте осуществления настоящего изобретения полинуклеотид, кодирующий аминокислотную последовательность, показанную в виде SEQ ID No: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 33, 34, 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70, 72, 74, 76 или 78, включает нуклеотидную последовательность, показанную в виде SEQ ID No: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 47, 49, 51, 53, 55, 57, 59, 61, 63, 65, 67, 69, 71, 73, 75 или 77, соответственно, или последовательность, кодирующую функциональный и/или иммуногенный фрагмент или вариант любой из SEQ ID No: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 33, 34, 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70, 72, 74, 76 или 78. Таким образом, настоящее изобретение относится к экспрессирующим конструкциям, включающим полинуклеотидную последовательность, содержащую нуклеотидную последовательность, показанную в виде любой из SEQ ID No: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 47, 49, 51, 53, 55, 57, 59, 61, 63, 65, 67, 69, 71, 73, 75 или 77, или фрагмент или вариант, включая вырожденный вариант, любой из SEQ ID No: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 47, 49, 51, 53, 55, 57, 59, 61, 63, 65, 67, 69, 71, 73, 75 или 77. В предпочтительном варианте осуществления настоящего изобретения рассматриваемая в нем экспрессирующая конструкция обеспечивает суперэкспрессию оперативно связанного полинуклеотида согласно настоящему изобретению.
Экспрессирующая конструкции согласно настоящему изобретению в основном включают регуляторные элементы, функционирующие в целевой клетке-хозяине, в которой должна экспрессироваться указанная экспрессирующая конструкция. При этом любой специалист со средним уровнем знаний в данной области может выбрать регуляторные элементы для целей использования, например, в человеческих клетках-хозяевах, клетках-хозяевах других млекопитающих, клетках-хозяевах насекомых, дрожжевых клетках-хозяевах, бактериальных клетках-хозяевах и растительных клетках-хозяевах. В одном варианте осуществления настоящего изобретения рассматриваемые в нем регуляторные элементы включают такие регуляторные элементы, которые функционируют в клетках собаки. Указанные регуляторные элементы включают промоторы, последовательности терминации транскрипции, последовательности терминации трансляции, энхансеры и сигналы полиаденилирования. В контексте настоящего описания термин "экспрессирующая конструкция" относится к сочетанию последовательностей нуклеиновых кислот, которая обеспечивает транскрипцию оперативно связанной последовательности нуклеиновой кислоты. В контексте настоящего описания термин "оперативно связанный" относится к такому взаимному расположению описанных компонентов, при котором указанные компоненты находятся в определенной взаимосвязи, позволяющей им функционировать заданным образом. В основном оперативно связанные компоненты расположены непосредственно вблизи друг от друга.
Экспрессирующая конструкция согласно настоящему изобретению может включать промоторную последовательность, оперативно связанную с полинуклеотидной последовательностью, кодирующей полипептид согласно настоящему изобретению. Промоторы могут быть введены в состав полинуклеотида с использованием стандартных методик, известных в данной области. В экспрессирующей конструкции согласно настоящему изобретению могут использоваться множественные копии промоторов или множественные промоторы. В предпочтительном варианте осуществления настоящего изобретения промотор может быть расположен примерно на том же примерно расстоянии от сайта старта транскрипции в экспрессирующей конструкции, как это имеет место применительно к сайту старта транскрипции в его природном генетической окружении. Допускаются некоторые вариации в этом расстоянии, которые не снижают в заметной мере промоторную активность. В экспрессирующую конструкцию в типичном случае включается сайт старта транскрипции. Предпочтительно, промотор, ассоциированный с экспрессирующей конструкцией согласно настоящему изобретению, обеспечивает суперэкспрессию оперативно связанного полинуклеотида согласно настоящему изобретению.
Промоторы, используемые в экспрессирующих конструкциях в эукариотических клетках, согласно настоящему изобретению, могут быть вирусного или клеточного происхождения. Вирусные промоторы включают, без ограничения, генные промоторы цитомегаловируса (CMV), ранние или поздние промоторы SV40 или генные промоторы вируса саркомы Рауса (RSV). Промоторы клеточного происхождения включают, без ограничения, промотор гена десмина и промотор гена актина. Промоторы, подходящие для использования в сочетании с конструкцией, экспрессируемой в дрожжевых клетках, включают, без ограничения, промотор 3-фосфоглицерилкиназы, промотор глицеральдегид-3-фосфатдегидрогеназы, промотор металлотионеина, промотор алкогольдегидрогеназы-2 и промотор гексокиназы.
Если экспрессирующая конструкция находится в растительной клетке или встраивается в нее, то могут также использоваться промоторы вирусов растений, такие как, например, вирус табачной мозаики (CaMV) 35S (включая усиленный промотор CaMV 35S (см., например, патент США No. 5 106 739 и An, 1997)) или промотор CaMV 19S. Другие промоторы, которые также могут использоваться в конструкциях, экспрессируемых в растениях, включают, например, промотор prolifera, Ap3 промотор, промоторы белков теплового шока, T-ДНК 1'- или 2'- промотор из A. tumefaciens, промотор полигалактуроназы, промотор халконсинтазы A (CHS-A) из петуньи, промотор PR-1a из табака, убихитиновый промотор, промотор актина, промотор гена alcA, промотор pin2 (Xu et al., 1993), промотор WipI из кукурузы, промотор гена trpA из кукурузы (патент США No. 5 625 136), промотор гена CDPK из кукурузы и промотор RUBISCO SSU (патент США No. 5 034 322). В конструкциях согласно настоящему изобретению могут использоваться корне-специфические промоторы, такие как промоторные последовательности, описанные в патенте No. 6 455 760, или в патенте США No. 6 696 623, или в публикациях заявок на патент США No. 20040078841; 20040067506; 20040019934; 20030177536; 20030084486 или 20040123349. Конститутивные промоторы (такие как промотор CaMV, убихина, актина или NOS), промоторы, регулируемые в процессе развития, и индуцибельные промоторы (такие как промоторы, которые могут быть индуцированы теплом, светом, гормонами или химическими веществами) также рассматриваются как подходящие для использования с полинуклеотидными экспрессирующими конструкциями согласно настоящему изобретению. Могут также использоваться тканеспецифичные промоторы, например специфичные для плодов промоторы, такие как промотор E8 из томатов (номер хранения депонированного материала: AF515784; Good et al. (1994)). Могут также использоваться специфичные для семян промоторы, такие как промотор из гена β-фазеолина (например, в фасоли обыкновенной) или гена глицина (например, в сое) и другие промоторы.
Для экспрессии в прокариотических системах экспрессирующая конструкция согласно настоящему изобретению может включать промоторы, такие как, например, промотор щелочной фосфатазы, промотор триптофана (trp), промотор ламбда PL, промотор β-лактамазы, промотор лактозы, промотор phoA, промотор T3, промотор T7 или промотор tac (de Boer et al., 1983).
Экспрессирующие конструкции согласно настоящему изобретению могут необязательно содержать последовательность терминации транскрипции, последовательность терминации трансляции, последовательность, кодирующую сигнальный пептид, и/или энхансерные элементы. Участки терминации транскрипции в типичном случае могут быть получены из 3'-нетранслируемой области эукариотической или вирусной генной последовательности. Последовательности терминации транскрипции могут быть локализованы по направлению считывания кодирующей последовательности, обеспечивая эффективную терминацию. Последовательность сигнального пептида представляет собой короткую аминокислотную последовательность, присутствующую обычно на амино-конце белка, отвечающего за перемещение оперативно связанного зрелого полипептида как в пределах клетки, по самым разным направлениям, варьирующим от специфических клеточных отсеков до сайта действия белка, так и во внеклеточную среду. Применительно к полипептидам согласно настоящему изобретению рассматриваются целевые генные продукты, действующие в направлении заданной мишени внутри клетки и/или во внеклеточной среде, ассоциированные с использованием оперативно связанной сигнальной пептидной последовательности. Классические энхансеры представляют собой цис-энхансеры, которые повышают уровень транскрипции гена и которые также могут включаться в экспрессирующую конструкцию. Такие классические энхансерные элементы известны в данной области и включают, без ограничения, энхансерный элемент CaMV 35S, энхансерный элемент раннего промотора цитомегаловируса (CMV) и энхансерный элемент SV40. Интрон-опосредованные энхансерные элементы, которые повышают генную экспрессию, известны в данной области. Указанные элементы должны присутствовать в транскрибируемой области и ориентированы определенным образом.
Последовательности ДНК, которые направляют полиаденилирование мРНК, транскрибированной на основе экспрессирующей конструкции и которые также могут вводиться в экспрессирующую конструкцию, включают, без ограничения, сигнальную последовательность октопинсинтазы или нопалинсинтазы.
Экспрессирующие конструкции могут также включать один или несколько доминантных селектируемых маркерных генов, включающих, например, гены, кодирующие антибиотикорезистентность и/или гербицидную резистентность выбранных трансформированных клеток. Гены антибиотикорезистентности могут обеспечивать резистентность к одному или нескольким антибиотикам, таким как гигромицин, канамицин, блеомицин, G418, стрептомицин, паромомицин, неомицин и спектиномицин. Резистентность к канамицину может быть достигнута за счет неомицинфосфотрансферазы (NPT II). Гены резистентности к гербицидам могут обеспечивать резистентность к ацетилтрансферазе фосфинотрицина или глифозата. Другие маркеры, используемые для скрининга клеточных трансформантов, включают, без ограничения, гены, кодирующие β-глюкуронидазу (GUS), β-галактозидазу, люциферазу, нопалинсинтазу, хлорамфениколацетилтрансферазу (CAT), зеленый флуоресцентный белок (GFP) или усиленный GFP (Yang et al., 1996).
Настоящее изобретение также относится к полинуклеотидным векторам, включающим полинуклеотидную последовательность согласно настоящему изобретению, которая кодирует полипептид согласно настоящему изобретению. Уникальные сайты для ферментов рестрикции могут быть включены на 5' и 3' концах экспрессирующей конструкции или полинуклеотида согласно настоящему изобретению, с тем чтобы можно быть ввести инсерцию в полинуклеотидный вектор. В контексте настоящего описания термин "вектор" относится к любому генетическому элементу, включающему, например, плазмиды, космиды, хромосомы, фаг, вирус и т.п., которые способны к репликации в сочетании с соответствующими контрольными элементами и которые могут переносить полинуклеотидные последовательности между клетками. Векторы содержат нуклеотидную последовательность, которая позволяет вектору реплицироваться в выбранной клетке-хозяине. Доступно множество векторов для экспрессии и/или клонирования, которые включают, без ограничения, вектор pBR322, векторы серии pUC, векторы серии M13, векторы серии pGEM и вектор pBLUESCRIPT (Stratagene, La Jolla, CA и Promega, Madison, WI).
Настоящее изобретение также относится к олигонуклеотидным зондам и праймерам, таким как праймеры для полимеразно-цепной реакции (ПЦР), которые могут гибридизоваться с кодирующей или некодирующей последовательностью полинуклеотида согласно настоящему изобретению. Олигонуклеотидные зонды согласно настоящему изобретению могут использоваться в рамках методик выявления последовательностей нуклеотидных кислот вируса гриппа. Олигонуклеотидные праймеры согласно настоящему изобретению могут использоваться в методиках ПЦР, а также в других методиках, включающих амплификацию нуклеиновой кислоты. В предпочтительном варианте зонд или праймер согласно настоящему изобретению может гибридизоваться с полинуклеотидом согласно настоящему изобретению в жестких условиях. Праймеры и зонды согласно настоящему изобретению могут необязательно включать выявляемую метку или репортерную молекулу, такие как флуоресцентные молекулы, ферменты, радиоактивный фрагмент и т.п. Зонды и праймеры согласно настоящему изобретению могут иметь любую длину, приемлемую для метода или теста, в котором они используются. В типичном случае, зонды и праймеры согласно настоящему изобретению включают в длину от 10 до 500 или более нуклеотидов. В область настоящего изобретения включаются зонды и праймеры, которые включают в длину 10-20, 21-30, 31-40, 41-50, 51-60, 61-70, 71-80, 81-90, 91-100 или 101 или более нуклеотидов. В одном варианте осуществления настоящего изобретения рассматриваемые в нем зонды и праймеры включают в длину 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29 или 30 нуклеотидов. Зонды и праймеры согласно настоящему изобретению могут характеризоваться полной (100%) идентичностью по нуклеотидной последовательности к полинуклеотидной последовательности, или указанная идентичность по последовательности может быть ниже, чем 100%. Например, идентичность по последовательности между зондом или праймером и рассматриваемой последовательностью может составлять 99%, 98%, 97%, 96%, 95%, 90%, 85%, 80%, 75%, 70% или характеризоваться любым процентным показателем идентичности по последовательности, при условии, что зонд или праймер могут гибридизоваться в жестких условиях с нуклеотидной последовательностью полинуклеотида согласно настоящему изобретению. Репрезентативные зонды и праймеры согласно настоящему изобретению включают такие зонды и праймеры, которые имеют любую из последовательностей, показанную в виде SEQ ID NO: 35, SEQ ID NO: 36, SEQ ID NO: 37, SEQ ID NO: 38, SEQ ID NO: 39, SEQ ID NO: 40, SEQ ID NO: 41, SEQ ID NO: 42, SEQ ID NO: 43, SEQ ID NO: 44, SEQ ID NO: 45 и SEQ ID NO: 46, или функциональный фрагмент или вариант любой из SEQ ID NO: 35-46.
В контексте настоящего описания термины "нуклеиновая кислота", "полинуклеотид" и "олигонуклеотид" относятся к дезоксирибонуклеотиду, рибонуклеотиду или смеси дезоксирибонуклеотидного и рибонуклеотидного полимера в одноцепочечной или двухцепочечной форме и, в отсутствие иных ограничений, включают известные аналоги природных нуклеотидов, которые могут функционировать аналогично природным нуклеотидам. Полинуклеотидные последовательности включают последовательность цепи ДНК, которая может транскрибироваться в РНК, и цепи РНК, которая может транслироваться в белок. В область настоящего изобретения также включается комплементарная последовательность любой рассматриваемой в нем нуклеиновой кислоты, полинуклеотида или олигонуклеотида. Полинуклеотидные последовательности также включают полноразмерные последовательности, равно как и более короткие последовательности, полученные из полноразмерных последовательностей. Настоящее изобретение также включает те полинуклеотиды, которые комплементарны на уровне последовательности рассматриваемым в нем полинуклеотидам. Полинуклеотиды и полипептиды согласно настоящему изобретению могут быть представлены в очищенной форме или в виде выделенного продукта.
В связи с вырожденностью генетического кода, полипептид согласно настоящему изобретению может кодироваться множеством разных полинуклеотидных последовательностей. В работе Левина (1985) приводится таблица, описывающая возможные триплетные кодоны (где U также означает T) и аминокислоты, кодируемые каждым из этих кодонов. Кроме того, специалисты в данной области могут получить альтернативные полинуклеотидные последовательности, кодирующие одни и те же, или по существу одни и те же, полипептиды согласно настоящему изобретению. Указанные вырожденные вариантные и альтернативные полинуклеотидные последовательности охватываются рамками настоящего изобретения. В контексте настоящего описания, использование применительно к последовательностям фразы "по существу одни и те же" относится к таким последовательностям, которые кодируют аминокислотные замещения, делеции, добавления или инсерции и которые физически не меняют функциональную и/или иммуногенную активность полипептида, кодируемого полинуклеотидами согласно настоящему изобретению.
Настоящее изобретение также относится к вариантам полинуклеотидов согласно настоящему изобретению, которые кодируют полипептиды согласно настоящему изобретению. Вариантные последовательности включают такие последовательности, в которых один или несколько нуклеотидов были замещены, делетированы и/или встроены. Нуклеотиды, которые могут замещать природные нуклеотиды в ДНК, содержат фрагмент основания, который включает, без ограничения, инозин, 5-фторурацил, 5-бромурацил, гипоксантин, 1-метилгуанин, 5-метилцитозин и тритилированные основания. Сахарный фрагмент нуклеотида в такой последовательности может быть также модифицирован и включает, без ограничения, арабинозу, ксилулозу и гексозу. Кроме того, адениновые, цитозиновые, гуаниновые, тиминовые и урациловые основания нуклеотидов могут быть модифицированы ацетильной, метильной и/или тиогруппами. Последовательности, содержащие нуклеотидные замещения, делеции и/или инсерции, могут быть получены и проанализированы с использованием стандартных методик, известных в данной области.
Настоящее изобретение также охватывает своими рамками замещение аминокислот, отличных от конкретно указанных примеров или от присутствующих в полипептиде согласно настоящему изобретению, в его природной форме. Так, например, неприродные аминокислоты могут замещать аминокислоты в полипептиде, при условии, что такой полипептид с замещенными аминокислотами сохраняет по существу ту же самую функциональную активность, что и полипептид, в котором аминокислоты не были замещены. Примеры неприродных аминокислот включают, без ограничения, орнитин, цитруллин, гидроксипролин, гомосерин, фенилглицин, таурин, йодтирозин, 2,4-диаминомасляную кислоту, α-аминомасляную кислоту, 4-аминомасляную кислоту, 2-аминомасляную кислоту, γ-аминомасляную кислоту, ε-аминогексаноевую кислоту, 6-аминогексаноевую кислоту, 2-аминоизомасляную кислоту, 3-аминопропионовую кислоту, норлейцин, норвалин, саркозин, гомоцитруллин, цистеиновую кислоту, τ-бутилглицин, τ-бутилаланин, фенилглицин, циклогексилаланин, β-аланин, фтораминокислоты, сконструированные аминокислоты, такие как β-метиламинокислоты, C-метиламинокислоты, N-метиламинокислоты и, в целом, аналоги аминокислот. Неприродные аминокислоты также включают аминокислоты, содержащие дериватизированные боковые группы. Кроме того, любая из аминокислот в белке может быть в D (правовращающей) форме или в L (левовращающей) форме. В область настоящего изобретения также включаются аллельные варианты белковой последовательности полипептида согласно настоящему изобретению.
Аминокислоты могут быть классифицированы, в основном, на несколько классов: неполярные, незаряженные полярные, основные и кислотные аминокислоты. Настоящее изобретение охватывает своими рамками консервативные замещения, в ходе которых полипептид согласно настоящему изобретению содержит аминокислоту одного определенного класса, замещенную другой аминокислотой того же класса, если полипептид, содержащий такое замещение, все еще сохраняет по существу ту же самую функциональную активность, что и полипептид, который не содержит замещения. Полинуклеотиды, кодирующие полипептид, содержащий в своей последовательности одно или несколько аминокислотных замещений, также входят в область настоящего изобретения. В приведенной ниже таблице 11 дан перечень примеров аминокислот, принадлежащих к каждому из указанных классов. Однобуквенные сокращения названий аминокислот определены в таблице 12.
Фрагменты и варианты полипептидов вируса гриппа согласно настоящему изобретению могут быть получены с использованием стандартных методик, известных в данной области, и далее проанализированы на наличие в них функции или иммуногенности с использованием стандартных методик, известных в данной области. Так, например, при анализе фрагментов и/или вариантов полипептида нейраминидазы согласно настоящему изобретению может быть определена его ферментативная активность. Таким образом, любой специалист со средним уровнем знаний в данной области может без труда получить и протестировать фрагменты и варианты полипептида согласно настоящему изобретению и определить, сохраняет ли указанный фрагмент или вариант активность, в сравнении с полноразмерным полипептидом или полипептидом, который не является вариантом.
Полинуклеотиды и полипептиды, включенные в область настоящего изобретения, могут быть определены в терминах большей идентичности и/или близких диапазонов, в сравнении с конкретно приведенными в настоящем описании последовательностями согласно настоящему изобретению. Идентичность по последовательности в типичном случае выше чем 60%, предпочтительно, выше чем 75%, более предпочтительно, выше чем 80%, и еще более предпочтительно, выше чем 90%, и может быть даже выше чем 95%. Идентичность и/или сходство такой последовательности может составлять 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98 или 99%, в сравнении с последовательностью, конкретно приведенной в настоящем описании. Если особо не указано иное, приводимый в описании процент идентичности и/или сходства на уровне последовательности двух последовательностей может быть определен с использованием алгоритма Карлина и Альтшуля (Karlin and Altschul (1990)), в модификации Карлина и Альтшуля (Karlin and Altschul (1993)). Указанный алгоритм включен в программы NBLAST и XBLAST, разработанные Альтшулем с соавт. (Altschul et al. (1990)). Поиски в режиме BLAST могут быть осуществлены на основе программы NBLAST, включающей следующие параметры: очки = 100, длина слов = 12, для получения последовательностей с желательным значением процента идентичности по последовательности. Для выравнивания последовательностей по наличию двухцепочечных разрывов, с целью сравнения, может использоваться программа Gapped BLAST, описанная в работе Альтшуля с соавт. (Altschul et al. (1997)). При использовании программ BLAST и Gapped BLAST, могут использоваться заданные параметры соответствующих программ (NBLAST и XBLAST). См. веб-сайт NCBI/NIH.
Настоящее изобретение также относится к таким полинуклеотидным молекулам, которые имеют последовательности, обладающие достаточной степенью гомологии с полинуклеотидными последовательностями, приведенными в качестве примеров в настоящем изобретении, что позволяет им гибридизоваться с каждой из таких последовательностей в условиях стандартной жесткости и в рамках стандартных методик (Maniatis et al., 1982). В контексте настоящего описания термин "жесткие" условия применительно к гибридизации относится к условиям, в случае которых гибридизация проводится в течение ночи при температуре, которая на 20-25ºC ниже температуры плавления (Tm) гибридной ДНК в смеси, включающей 6x SSPE, 5x растворе Денхардта, 0,1% SDS, 0,1 мг/мл денатурированной ДНК. Температура плавления, Tm, описывается приведенной ниже формулой (Beltz et al., 1983):
Tm=81,5ºC+16,6 Log[Na+]+0,41(%G+C)-0,61(% формамид)-600/длина дуплекса пар оснований.
Промывку проводят в типичном случае следующим образом:
(1) Два раза при комнатной температуре в течение 15 минут в 1x SSPE, 0,1% ДСН (промывка в условиях низкой жесткости).
(2) Один раз при Tm-20ºC в течение 15 минут в 0,2x SSPE, 0,1% ДСН (промывка в условиях умеренной жесткости).
Настоящее изобретение также относится к вирусным белкам и пептидам, кодируемым генами вируса гриппа согласно настоящему изобретению. В одном варианте осуществления настоящего изобретения указанный вирусный белок представляет собой зрелый белок HA. В конкретном варианте осуществления настоящего изобретения зрелый белок HA включает одну или несколько следующих особенностей: содержит серин в положении 82; лейцин в положении 221; треонин в положении 327; и/или треонин в положении 482. В репрезентативном варианте осуществления настоящего изобретения зрелый белок HA содержит аминокислотную последовательность, показанную в виде SEQ ID NO: 33 или SEQ ID NO: 34, или функциональный и/или иммуногенный фрагмент или вариант SEQ ID NO: 33 или SEQ ID NO: 34. В другом варианте осуществления настоящего изобретения указанный вирусный белок представляет собой белок NA, белок NS, белок PB, белок PA или белок MA. Вирусные белки и пептиды согласно настоящему изобретению могут использоваться для получения антител, которые специфически связываются с указанным белком или пептидом. Вирусные белки и пептиды согласно настоящему изобретению могут также использоваться в качестве иммуногенов и в составе вакцинных композиций.
Настоящее изобретение также относится к композициям и способам, используемым для индукции иммунного ответа против вируса гриппа, обладающего способностью инфицировать организм чувствительного животного-хозяина и вызвать респираторное заболевание. Настоящее изобретение может также использоваться для индукции иммунного ответа против вируса гриппа любого подтипа у чувствительного животного-хозяина. Вирус гриппа может представлять собой, например, вирус HA, подтипа H1, H2, H3, H4, H5, H6, H7, H8, H9, H10, H11, H12, H13, H14, H15 или H16, а также вирус NA, подтипа N1, N2, N3, N4, N5, N6, N7, N8 или N9. В одном варианте осуществления настоящего изобретения соответствующим подтипом вируса HA является H3 или H5. В другом варианте осуществления настоящего изобретения соответствующим подтипом вируса NA является N7 или N8. В конкретном варианте осуществления настоящего изобретения иммунный ответ индуцируется против вируса гриппа подтипа H3N8. В одном варианте осуществления настоящего изобретения соответствующим животным-хозяином является представитель псовых. Псовые в контексте настоящего изобретения включают диких, содержащихся в зоопарке, и домашних собак, таких как волки, койоты и лисы. Псовые в контексте настоящего изобретения также включают собак, в частности домашних собак, таких как, например, чистокровные и/или помесные собаки, декоративные собаки, служебные собаки, пастушьи собаки, охотничьи собаки, сторожевые собаки, полицейские собаки, гончие собаки и/или лабораторные собаки. В конкретном варианте осуществления настоящего изобретения животное-хозяин представляет собой домашнюю собаку, такую как борзая. В одном варианте осуществления настоящего изобретения животному вводят эффективное количество иммуногенной композиции согласно настоящему изобретению, достаточное для индукции иммунного ответа против вируса гриппа согласно настоящему изобретению. Указанный иммунный ответ может представлять собой гуморальный и/или клеточный иммунный ответ. В конкретном варианте осуществления настоящего изобретения указанный иммунный ответ представляет собой защитную иммунную реакцию, способную предотвратить или минимизировать проявление вирусной инфекции в иммунизированном организме животного-хозяина в течение некоторого периода времени после иммунизации. Таким образом, настоящее изобретение также относится к вакцинным композициям и методам, которые обеспечивают генерирование у вакцинированного животного защитной иммунной реакции против вируса согласно настоящему изобретению.
Как приведено в настоящем описании, вакцинные или иммуногенные композиции согласно настоящему изобретению могут включать бесклеточный полный вирус, включающий аттенуированный или инактивированный вирус, или части такого вируса, в том числе субвирионные частицы (включая субъединичную вакцину, в составе которых вирион подвергают обработке с целью удаления некоторой части или всех вирусных липидов), вирусные белки (включая индивидуальные белки и макромолекулярные комплексы множества белков), полипептиды и пептиды, а также инфицированные вирусом клеточные линии или любое сочетание указанных форм. Вакцинные или иммуногенные композиции, содержащие инфицированные вирусом клеточные линии, могут включать множество клеточных линий, каждая из которых инфицирована другим штаммом вируса.
Вакцинные или иммуногенные композиции согласно настоящему изобретению относятся также к рекомбинантным конструкциям, созданным с использованием вирусного вектора, которые могут включать, например, гены, кодирующие белок HA, белок NA, нуклеопротеин, основной белок полимеразы, кислый белок полимеразы и/или матричный белок вируса гриппа согласно настоящему изобретению. Для использования в рамках настоящего изобретения может рассматриваться любой подходящий вирусный белок, который может быть использован для получения рекомбинантной вектор/вирусной конструкции. Так, например, в композициях и способах настоящего изобретения могут использоваться вирусные векторы, полученные из аденовируса, вируса птичьей оспы, герпесвируса, вируса осповакцины, вируса оспы канареек, вируса оспы насекомых, вируса свиной оспы, вируса лихорадки Западного Нила и других известных в данной области векторов. Рекомбинантные полинуклеотидные векторы, которые кодируют и экспрессируют соответствующие компоненты, могут быть сконструированы с использованием методик, известных в данной области. Кроме того, приведенные в настоящем описании различные вакцинные композиции могут использоваться как по отдельности, так и в сочетании друг с другом. Так, например, при первичных иммунизациях животных могут использоваться рекомбинантные конструкции, построенные на основе векторов, содержащих компоненты одного или многих штаммов, с последующим проведением вторичного бустинга вакцинными композициями, содержащими инактивированный вирус или клеточные линии, инфицированные инактивированным вирусом. Другие протоколы иммунизации с использованием вакцинных композиций согласно настоящему изобретению известны специалистам в данной области и охватываются объемом настоящего изобретения.
Настоящее изобретение также относится к перегруппированному вирусу, включающему по меньшей мере один ген или генный сегмент из вируса гриппа согласно настоящему изобретению, тогда как оставшаяся часть вирусных генов или сегментов генома происходит из другого вируса гриппа согласно настоящему изобретению или из вируса гриппа, отличного от вируса согласно настоящему изобретению. Перегруппированный вирус может быть получен путем генетической реаранжировки нуклеиновой кислоты из донорного вируса гриппа согласно настоящему изобретению с нуклеиновой кислотой реципиентного вируса гриппа, с последующим отбором перегруппированного вируса, который включает нуклеиновую кислоту из вируса-донора. Способы продуцирования и выделения перегруппированного вируса известны в данной области (Fields et al., 1996). В одном варианте осуществления настоящего изобретения перегруппированный вирус согласно настоящему изобретению включает гены или сегменты генома из вируса гриппа человека, птицы, свиньи или лошади. Перегруппированный вирус согласно настоящему изобретению может включать любо сочетание нуклеиновых кислот из донорного и реципиентного вируса гриппа при условии, что перегруппированный вирус включает по меньшей мере один ген или генный сегмент из донорного вируса согласно настоящему изобретению. В одном варианте осуществления настоящего изобретения реципиентный вирус гриппа может представлять собой вирус гриппа лошади.
Природные, рекомбинантные или синтетические полипептиды вирусных белков и их пептидные фрагменты также могут использоваться в качестве вакцинных композиций, в соответствии со способами, предлагаемыми в настоящем изобретении. В одном варианте осуществления настоящего изобретения вирусные полипептиды, получаемые из множества штаммов, могут быть объединены в составе вакцинной композиции и далее использоваться для вакцинации животного-хозяина. Так, например, в составе одной вакцины могут быть объединены полипептиды, основанные на HA белках, взятых по меньшей мере из двух штаммов вируса гриппа согласно настоящему изобретению. Указанные полипептиды могут быть гомологичны по отношению к одному штамму или могут включать «гибридные», или «химерные» полипептиды, аминокислотная последовательность которых получена из присоединенных или связанных полипептидов, взятых по меньшей мере из двух разных штаммов. Процедуры получения вирусных полипептидов известны в данной области. Так, например, вирусные полипептиды и пептиды могут быть синтезированы с использованием методик твердофазного синтеза (Merrifield, 1963). Вирусные полипептиды и пептиды могут быть также получены с использованием методик рекомбинантных ДНК, в рамках которых полинуклеотидную молекулу, кодирующую вирусный белок или пептид, подвергают экспрессии в клетке-хозяине, такой как бактериальная клетка, дрожжевая клетка или клеточные линии млекопитающих, после чего экспрессированный белок подвергают очистке с использованием стандартных методик, широко применяемых в данной области.
Вакцинные композиции согласно настоящему изобретению включают композиции на основе оголенных нуклеиновых кислот. В одном варианте осуществления настоящего изобретения нуклеиновая кислота может включать нуклеотидную последовательность, кодирующую белок HA и/или NA из вируса гриппа согласно настоящему изобретению. Способы вакцинации нуклеиновыми кислотами известны в данной области и описаны, например, в патентах США No. 6 063 385 и 6 472 375. Указанная нуклеиновая кислота может иметь форму плазмиды или кассеты генной экспрессии. В одном варианте осуществления настоящего изобретения рассматриваемая в нем нуклеиновая кислота инкапсулирована в состав липосомы, вводимой в организм животного.
Вакцинные композиции и иммуногены, такие как полипептиды и нуклеиновые кислоты, которые могут использоваться в соответствии с настоящим изобретением, могут поставляться вместе с фармацевтически приемлемым носителем или разбавителем. Соединения и композиции, используемые согласно настоящему изобретению, могут быть изготовлены с использованием известных методов получения фармацевтически полезных композиций. Процессы изготовления композиций подробно описаны во многих источниках, которые хорошо известны специалистам в данной области и доступны для использования. См., например, руководство Ремингтона (Remington's Pharmaceutical Science by E.W. Martin, Easton Pennsylvania, Mack Publishing Company, 19th ed., 1995), в котором описываются композиции, которые могут использоваться в контексте настоящего изобретения. В основном, композиции согласно настоящему изобретению изготавливают путем объединения эффективного количества иммуногена с приемлемым носителем, с тем чтобы достичь эффективности при введении такой композиции. Композиции, используемые в соответствии с настоящим изобретением, могут быть представлены в виде разнообразных форм. Указанные формы включают, например, твердые, полутвердые и жидкие дозированные формы, такие как таблетки, пилюли, порошки, жидкие растворы или суспензии, суппозитории, растворимые, вводимые инъекцией и инфузией формы, а также распыляемые формы. Предпочтительная форма определяется предполагаемым способом введения и характером терапевтического применения. Предпочтительно, композиции также включают традиционные фармацевтически приемлемые носители и разбавители, известные специалистам в данной области. Примеры носителей или разбавителей, которые могут использоваться вместе с пептидомиметиками, включают, без ограничения, воду солевой раствор, масла, включая минеральное масло, этанол, диметилсульфоксид, желатин, циклодекстраны, стеарат магния, декстрозу, целлюлозу, сахара, карбонат кальция, глицерин, глинозем, крахмал и эквивалентные указанным, носители и разбавители, или их смеси. Композиции на основе иммуногена согласно настоящему изобретению могут также включать суспензию агентов, защитных веществ, замасливателей, буферов, консервантов и стабилизаторов. Для целей введения таких дозировок в направлении проведения желательного терапевтического воздействия, фармацевтические композиции согласно настоящему изобретению могут с успехом включать примерно от 0,1 вес.% до 45 вес.%, и в особенности, от 1 до 15 вес.% одного или нескольких иммуногенов от веса всей композиции, включая носитель или разбавитель.
Вакцинные и иммуногенные композиции согласно настоящему изобретению могут быть изготовлены в соответствии с процедурами, хорошо известными в данной области. Например, в типичном случае вакцину и иммуногены изготавливают в виде инъецируемых форм, например в виде жидких растворов или суспензий. Вакцину или иммуногены вводят с использованием такого способа, который будет совместим с данной дозированной формой, и в таком количестве, которая будет терапевтически эффективной и иммуногенной для реципиента. Оптимальные дозировки и характер введения, подходящие для конкретной композиции вакцины или иммуногенов, могут быть без труда определены специалистом в данной области.
Пептиды и/или полипептиды согласно настоящему изобретению также могут быть представлены в виде конструкции из множества антигенных пептидов (MAP). Получение конструкций MAP описано в работе Tam (1988). Конструкции MAP включают наличие ядерной матрицы из лизиновых остатков, на основе которой синтезируют множество копий иммуногена (Posnett et al., 1988). Может быть получено множество конструкций MAP, которые содержат одинаковые или разные иммуногены, и введено в составе вакцинной композиции согласно способам настоящего изобретения. В одном варианте осуществления настоящего изобретения конструкция MAP представлена в объединенной форме вместе с одним или несколькими адъювантами и/или вводится вместе с одним или несколькими адъювантами. Полипептиды из вируса гриппа согласно настоящему изобретению также могут быть получены и далее введены в виде макромолекулярных белковых структур, включающих один или несколько полипептидов. См. публикацию заявки на патент США US2005/0009008, в которой описываются способы получения вирусоподобных частиц в качестве вакцины против вируса гриппа.
В соответствии со способами настоящего изобретения описываемые в нем вакцинные и иммуногенные композиции вводят в чувствительный организм-хозяин, в типичном случае псовых, и в более типичном случае, домашним собакам, в эффективном количестве и по способу, позволяющему индуцировать защитный иммунитет против последующих провокации или инфекции данного организма-хозяина вирусом. В одном варианте осуществления настоящего изобретения указанный организм животного-хозяина включает представителя псовых. Псовые включают диких, содержащихся в зоопарке и домашних собак, таких как волки, койоты и лисы. Псовые также включают собак, в частности, домашних собак, таких как, например, чистокровные и/или помесные собаки, декоративные собаки, служебные собаки, пастушьи собаки, охотничьи собаки, сторожевые собаки, полицейские собаки, гончие собаки и/или лабораторные собаки. В конкретном варианте осуществления настоящего изобретения животное-хозяин представляет собой домашнюю собаку, такую как борзая. Вакцины или иммуногены в типичном случае вводят парентерально, путем инъекции, например, подкожно, внутрибрюшинно или внутримышечно. Другие подходящие способы введения включают пероральное или назальное введение. Обычно вакцины и иммуногены вводят животному по меньшей мере два раза, с интервалом между введениями в одну или несколько недель. Однако в изобретении рассматриваются также другие режимы начального и бустерного введения вакцины или иммуногенов, выбор которых может определяться решением соответствующего специалиста, с учетом конкретного организма-хозяина, подлежащего лечению.
Вирус и инфицированные вирусом клетки в составе вакцинной композиции могут быть инактивированы или ослаблены с использованием известных в данной области способов. Например, целый вирус и инфицированные вирусом клетки могут быть инактивированы или ослаблены за счет воздействия на них параформальдегида, формалина, фенола, УФ-облучения, повышенной температуры и т.п. Количество бесклеточного целого вируса в дозе вакцины обычно варьирует в диапазоне от примерно 0,1 мг до примерно 5 мг, и чаще, составляет от примерно 0,2 мг до примерно 2 мг. Дозировки вакцинных композиций, включающих клеточные линии, инфицированные вирусом, обычно содержат от примерно 106 до примерно 108 клеток на дозу, и чаще, от примерно 5 × 106 до примерно 7,5 × 107 клеток на дозу. Количество белкового или пептидного иммуногена, содержащегося в дозе для одного животного, может варьировать примерно от 0,1 мкг до 10000 мкг или примерно от 1 мкг до 5000 мкг, или примерно от 10 мкг до 1000 мкг, или примерно от 25 мкг до 750 мкг, или примерно от 50 мкг до 500 мкг, или от 100 мкг до 250 мкг, в зависимости от размера, возраста и других характеристик животного, получающего дозу.
Иммуногенная или вакцинная композиция согласно настоящему изобретению, такая как композиция на основе вируса или клеток, инфицированных вирусом, или вирусных белков или пептидов, может быть объединена с адъювантом, что обычно делают перед введением. Адъюванты, которые могут рассматриваться как подходящие для использования в вакцинных композициях, включают треонилмурамильный дипептид (MDP) (Byars et al., 1987), сапонин, агент на основе Corinebacterium parvum, полный и неполный адъюванты Фрейнда, глинозем или смесь любых из указанных агентов. В настоящем изобретении рассматривается также множество других адъювантов, подходящих для использования в сочетании с методами и вакцинами согласно настоящему изобретению, хорошо известные в данной области, такие как глинозем.
Настоящее изобретение также относится к антителам, которые способны специфически связываться с белком или пептидом согласно настоящему изобретению. Антитела согласно настоящему изобретению включают композиции на основе моноклональных и поликлональных антител. Предпочтительно, антитела согласно настоящему изобретению представляют собой моноклональные антитела. В настоящем изобретении рассматриваются как полные антитела, так и их антиген-связывающие фрагменты. В этой связи, например, подходящие антиген-связывающие фрагменты включают Fab2, Fab и Fv антительные фрагменты. Антитела согласно настоящему изобретению могут быть мечены детектируемым фрагментом, таким как флуоресцентная молекула (например, флуоресцеин или фермент).
Настоящее изобретение также относится к способам и композициям, используемым для детекции и идентификации вируса гриппа согласно настоящему изобретению и для диагностики наличия у животного инфекции вирусом гриппа согласно настоящему изобретению. Способы согласно настоящему изобретению включают детекцию наличия гриппа собак в биологическом образце, взятом у животного. Детекция гриппа собак в образце используется для диагностики собачьего гриппа у животного. В свою очередь, эта информация может позволить сделать прогноз развития заболевания у животного на основе оценки характерных уровней прогрессирования собачьего гриппа с течением времени и стать подспорьем в выборе терапевтических агентов и методов лечения животного, а также использоваться при мониторинге проводимой терапии. Данный способ также позволяет установить отсутствие собачьего гриппа у исследуемого животного.
Возможность выявления собачьего гриппа у животного позволит проводить оценку вспышек заболеваемости собачьим гриппом в разных географических областях. Такого рода информация также поможет в достижении ранней детекции, так что в этом случае инфицированные животные могут быть вовремя изолированы, что будет способствовать ограничению распространения заболевания, а также позволит провести более ранние соответствующие терапевтические процедуры. Кроме того, такая информация, в случае ее доступности, может стать основой для организации и отправки медицинского персонала, готового лечить большое число больных животных, включая подготовку продукции и устройств медицинского назначения, и, при необходимости, вакцин.
В одном варианте осуществления настоящего изобретения рассматриваемый в нем способ включает отбор биологического образца из организма исследуемого животного, такого как собака. Биологический образец может представлять собой любой биологический материал, включающий клетки, ткань, волосы, цельную кровь, сыворотку, плазму, аспират из соска, лаваж легких, цереброспинальную жидкость, слюну, пот и слезы.
Исследуемый образец животного может быть взят от животного, в отношении которого подозревается наличие инфекции вирусом собачьего гриппа, независимо от того, проявляются ли у животного симптомы этого заболевания. Могут быть также получены или собраны контрольные образцы от тех животных, в отношении которых известно, что они не имеют собачьего гриппа. Могут быть также использованы дополнительные контроли, например, с целью снижения риска ложных и отрицательных результатов, а также для подтверждения того, что используемые в тесте реагенты активны по выявлению вируса собачьего гриппа типа А.
В дополнение к выявлению наличия или отсутствия собачьего гриппа в биологическом образце методы детекции, используемые в настоящем изобретении, могут выявлять мутации в вирусе собачьего гриппа, такие как изменения в последовательности нуклеиновой кислоты, которые могут быть следствием воздействия условий окружающей среды, лечения лекарственными препаратами, генетических манипуляций или мутаций, поражения, изменений в режиме питания, возраста или любых других одной или нескольких характеристик животного. Мутации могут также приводить к тому, что вирус собачьего гриппа A становится резистентным к тому лекарственному веществу, которое раньше было эффективным, или которые будут позволять вирусу инфицировать и размножаться в другом виде животного или в организме человека. Например, было показано, что вирус птичьего гриппа A становится способным инфицировать других животных и людей.
В одном варианте детекции вируса гриппа в организме животного согласно настоящему изобретению диагностика облегчается за счет отбора образцов высокого качества, их быстрой транспортировки к месту анализа, а также за счет создания соответствующих условий хранения, до лабораторного исследования. Вирус выявляется лучше всего в образцах, содержащих инфицированные клетки и их секреты. В одном варианте осуществления настоящего изобретения образцы для непосредственного выявления вирусных антигенов и/или нуклеиновых кислот и/или вирусных изолятов в клеточных культурах отбирают в течение первых 3 дней после появления первых клинических симптомов. Приемлемо множество разных типов образцов для целей диагностики вирусных инфекций верхних дыхательных путей, включающих, без ограничения, назальный мазок, носоглоточный мазок, носоглоточный аспират, назальную промывку и мазок из горла. Кроме отбора мазков могут быть отобраны образцы ткани или сыворотки, а также могут проводиться инвазивные процедуры.
В одном варианте осуществления настоящего изобретения респираторные образцы отбирают и переносят в 1-5 мл среды для переноса вируса. Коммерчески доступно большое число сред, которые подходят для восстановления множества вирусов. К среде для переноса добавляют клинические образцы. Назальные или носоглоточные мазки также могут транспортироваться в среде для переноса вирусов. Один из примеров среды для переноса включает среду, содержащую 10 г телячьего инфузионного бульона и 2 г бычьего альбумина, фракцию V, доведенную стерильной дистиллированной водой до объема 400 мл. К среде могут быть добавлены антибиотики, такие как 0,8 мл раствора сульфата гентамицина (50 мг/мл) и 3,2 мл амфотерицина B (250 мкг/мл). Среду предпочтительно стерилизуют фильтрованием. Назальные промывные жидкости, такие как полученные в стерильном солевом растворе (0,85% NaCl), также могут использоваться для целей отбора респираторных образцов с целью оценки наличия в них вирусов.
В одном варианте осуществления настоящего изобретения сыворотки собирают в количестве, соответствующем 1-5 мл цельной крови от животного в острой фазе заболевания, вскоре после появления клинических симптомов и предпочтительно не позже, чем через 7 дней. Может быть также отобран образец сыворотки в фазе выздоровления, например по прошествии приблизительно 14 дней после появления симптомов заболевания. Образцы сыворотки могут использоваться для выявления антител против респираторных вирусов в тесте на нейтрализацию.
В некоторых случаях образцы могут быть отобраны от индивидуальных животных в течение некоторого периода времени (например, с периодичностью один раз в день, один раз в неделю, один раз в месяц, один раз в два года или ежегодно). Различные образцы, полученные от индивидуального животного в течение определенного периода времени, могут использоваться для подтверждения результатов более ранних детектирующих тестов и/или для идентификации реакции или резистентности в ответ на конкретное лечение, например, в ответ на выбранный терапевтический препарат.
Способы согласно настоящему изобретению могут использоваться для детекции наличия одного или нескольких патологических агентов в исследуемом образце от животного и для определения уровня каждого такого патологического агента. При этом может использоваться любой способ детекции патологического агента, включающий, без ограничения, антительные тесты, в том числе тесты на основе иммуноферментного твердофазного анализа (ELISA), тесты, основанные на непрямой флуоресценции антитела (IFA), тесты с использованием реакции гемагглютинации и тесты, основанные на ингибировании реакции гемагглютинации (HI), а также вестерн-блот анализ. Могут также использоваться известные методы культивирования клеток. Положительные по нужному признаку культуры могут быть далее идентифицированы с использованием феномена флуоресценции клеточных культур или на основе HI анализа среды для клеточной культуры (супернатанта).
Кроме того, могут использоваться методы детекции нуклеиновой кислоты (ДНК или РНК) или белка. Такие методы включают, без ограничения, полимеразно-цепную реакцию (ПЦР) и тесты на основе ПЦР с обратной транскриптазой (ОТ), а также тесты в масштабе реального времени и количественные тесты по нуклеазной защите. Имеются коммерчески доступные наборы, подходящие для проведения указанных видов тестирования. Так, например, компания QIAGEN (Valencia, CA) продает набор для проведения одностадийного тестирования в рамках реакции ОТ-ПЦР, а также набор для экстрагирования вирусной РНК.
В одном варианте осуществления настоящего изобретения данный метод включает использование антитела, специфичного для вируса или вирусного белка согласно настоящему изобретению. В конкретном варианте осуществления настоящего изобретения используют антитело, специфичное для белка HA из вируса согласно настоящему изобретению. В другом варианте осуществления настоящего изобретения используют антитело, специфичное для белка NP из вируса согласно настоящему изобретению. При этом из организма животного отбирают подходящий образец, например, из назальной или носоглоточной области и выделяют из него вирус или вирусный белок. Далее вирусные компоненты подвергают скринингу для оценки их способности связываться с антителом, специфичным для белка, таким как HA или NP, из вируса согласно настоящему изобретению. В другом варианте осуществления настоящего изобретения из организма животного отбирают сыворотку (или другой антитело-содержащий образец) и далее сыворотку подвергают скринингу для выявления наличия антитела, которое связывается с белком из вируса согласно настоящему изобретению. Так, например, может быть проведен тест ELISA, в рамках которого ячейки планшета содержат белок HA и/или NP, или их пептид, связанные с ячейкой. Затем ячейки в планшете подвергают контакту с сывороткой или антителом из исследуемого организма. Наличие антитела в организме животного, которое специфически связывается с белком HA и/или NP, является показателем того, что исследуемое животное инфицировано вирусом гриппа согласно настоящему изобретению.
В одном варианте осуществления настоящего изобретения наличие патологического агента выявляется посредством определения наличия или отсутствия в биологическом образце антител против данного агента. Этот процесс может потребовать некоторого времени (например, месяцы) после инфицирования животного и перед выявлением антител при анализе крови. Однако, после своего появления, антитела обычно персистируют в течение многих лет, даже если заболевание было успешно излечено. Обнаруженные антитела против вируса собачьего гриппа A необязательно следует рассматривать как указание на то, что инфекция была недавно, поскольку они могут относиться к инфекции, имевшей место в прошлом.
Анализ на наличие антител может быть также выполнен в одной или нескольких жидкостях в качестве образца. Тесты для выявления антител включают тесты на основе иммуноферментного твердофазного анализа (ELISA), тесты, основанные на непрямой флуоресценции антител (IFA), а также вестерн-блот анализ. Предпочтительно, анализ на наличие антител проводят с использованием для этого множества тестов, например ELISA или IFA, с последующим проведением вестерн-блоттинга. Тесты на наличие антител могут быть выполнены в рамках двустадийного процесса, с проведением вначале ELISA или IFA анализов и затем вестерн-блоттинга. ELISA считается более достоверным и точным методом, чем IFA, однако IFA может использоваться в тех случаях, кода выполнение анализа ELISA по каким-либо причинам недоступно. Вестерн-блот анализ (представляющий собой более специфичный тест) может проводиться на всех животных, в особенности на тех из них, у которых по результатам ELISA или IFA анализов были получены позитивные или погранично-позитивные (неоднозначные) результаты.
Другие основанные на антителах тесты, которые могут использоваться для выявления вируса гриппа, включают тесты на ингибирование реакции гемагглютинации. Активность в реакции гемагглютинации может быть выявлена в биологическом образце, отобранном из организма животного, с использованием эритроцитов кур или индейки, как описано в литературе (Burleson et al., 1992; и Kendal et al., 1982). В одном варианте осуществления настоящего изобретения вирус гриппа или белок HA или пептид согласно настоящему изобретению подвергают контакту с исследуемым образцом, содержащим сыворотку или антитело. Добавляют эритроциты (RBC), взятые от животного, такого как птица. Если имеется антитело против HA, то RBC не агглютинируют. Если антитело к HA не присутствует, то RBC будут агглютинировать в присутствии HA. В данной области известны вариации и модификации стандартного теста на ингибирование реакции агглютинации, которые охватываются областью настоящего изобретения.
Инфекция у животного может быть также установлена путем выделения вируса из образца, такого как назальный или носоглоточный мазок. Выделение вируса может быть осуществлено с использованием стандартных методов, включающих культивирование клеток и инокуляцию яйца.
В другом варианте осуществления настоящего изобретения может быть использован тест на основе нуклеиновой кислоты для детекции вируса согласно настоящему изобретению. В одном варианте осуществления настоящего изобретения получают образец нуклеиновой кислоты из организма животного и нуклеиновую кислоту подвергают реакции ПЦР с использованием праймеров, способных генерировать продукт амплификации, если нуклеиновая кислота содержит последовательность, специфичную для вируса гриппа согласно настоящему изобретению. В конкретном варианте осуществления настоящего изобретения в тесте для оценки наличия целевого вируса используют ОТ-ПЦР. В репрезентативном варианте осуществления настоящего изобретения используют ОТ-ПЦР в масштабе реального времени для оценки наличия вируса гриппа согласно настоящему изобретению. Методы, основанные на проведении ПЦР, ОТ-ПЦР и ПЦР в масштабе реального времени, известны в данной области и описаны в патентах США No. 4 683 202; 4 683 195; 4 800 159; 4 965 188; 5 994 056; 6 814 934; и в работах Saiki et al. (1985); Sambrook et al. (1989); Lee et al. (1993); и Livak et al. (1995). В одном варианте осуществления настоящего изобретения в тесте на основе ПЦР используют олигонуклеотиды, специфичные для матричного гена (MA) и/или гена HA из вируса гриппа. Продукт амплификации может быть подвергнут секвенированию для определения, содержит ли данный продукт последовательность из вируса гриппа согласно настоящему изобретению. Другие основанные на нуклеиновых кислотах тесты могут использоваться для детекции и диагностики вирусной инфекции, вызванной вирусом согласно настоящему изобретению, и такие тесты охватываются областью настоящего изобретения. В одном варианте осуществления настоящего изобретения образец, содержащий нуклеиновую кислоту, подвергают реакции амплификации по методу ПЦР с использованием прямых и обратных праймеров, где указанные праймеры специфичны для полинуклеотида или генной последовательности вируса. Если нуклеиновая кислота в данном образце представлена РНК, то проводят реакцию ОТ-ПЦР. Для проведения ПЦР в масштабе реального времени с праймерами используют выявляемый зонд.
Наборы праймеров, специфичных для гена гемагглютинина (HA) из многих циркулирующих вирусов гриппа, известны и постоянно разрабатываются. Геном вируса гриппа представлен одноцепочечной РНК, и должна быть получена копия ДНК (кДНК) с использованием полимеразы обратной транскриптазы (ОТ). Реакция амплификации геномной РНК, например, с использованием ОТ-ПЦР, требует наличия пары олигонуклеотидных праймеров, обычно разрабатываемых на основе известной последовательности HA из вируса гриппа, подтипа A, и нейраминидазы (NM)-1. Соответствующие праймеры могут быть отобраны таким образом, что они будут специфически амплифицировать РНК только одного подтипа вируса. ДНК, генерируемые при использовании праймеров, специфичных для определенного подтипа, могут быть далее проанализированы в рамках молекулярно-генетических методик, таких как секвенирование. Данный тест проводят, предпочтительно, с включением в него позитивного контроля, или идентичность продуктов подтверждают путем их секвенирования и сравнения с известными последовательностями. Отсутствие целевых продуктов ПЦР (т.е. "отрицательный" результат) необязательно исключает наличие вируса. Результаты могут быть получены позже, через несколько часов, когда станут доступны данные анализа клинических мазков или инфицированных клеточных культур. Тесты на основе ПЦР и ОТ-ПЦР для вируса гриппа А описаны в работах Fouchier et al., 2000 и Maertzdorf et al., 2004.
Настоящее изобретении также относится к методам скрининга соединений или лекарственных агентов, которые обладают антивирусной активностью против вируса согласно настоящему изобретению. В одном варианте осуществления настоящего изобретения клетки, инфицированные вирусом согласно настоящему изобретению, подвергают контакту с исследуемым соединением или лекарственным агентом. Затем определяют количество вируса или вирусную активность после такого контакта. Те соединения или лекарственные агенты, которые демонстрируют противовирусную активность, могут быть отобраны для дальнейшей оценки.
Настоящее изобретение также относится к выделенным клеткам, инфицированным вирусом гриппа согласно настоящему изобретению. В одном варианте осуществления настоящего изобретения указанная клетка представляет собой клетку собаки, например, эпителиальные клетки почки собаки.
Настоящее изобретение также относится к клеткам, трансформированным полинуклеотидом согласно настоящему изобретению, кодирующим полипептид согласно настоящему изобретению. Предпочтительно, полинуклеотидная последовательность входит в экспрессирующую конструкцию согласно настоящему изобретению. Более предпочтительно, экспрессирующая конструкция обеспечивает суперэкспрессию в клетке оперативно связанного полинуклеотида согласно настоящему изобретению. В одном варианте осуществления настоящего изобретения клетку трансформируют полинуклеотидной последовательностью, включающей последовательность, кодирующую аминокислотную последовательность, показанную в виде любой из последовательностей SEQ ID No: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 33, 34, 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70, 72, 74, 76 или 78, или ее функциональный фрагмент или вариант. В конкретном варианте осуществления настоящего изобретения клетку трансформируют полинуклеотидом, кодирующим аминокислотную последовательность, показанную в виде SEQ ID No: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 33, 34, 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70, 72, 74, 76 или 78, который включает нуклеотидную последовательность, показанную в виде SEQ ID No: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 47, 49, 51, 53, 55, 57, 59, 61, 63, 65, 67, 69, 71, 73, 75 или 77, соответственно, или последовательность, кодирующую функциональный фрагмент или вариант любой из SEQ ID No: 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 33, 34, 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70, 72, 74, 76 или 78. Таким образом, настоящее изобретение относится к клеткам, трансформированным полинуклеотидной последовательностью, включающей нуклеотидную последовательность, показанную в виде любой из SEQ ID No: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 47, 49, 51, 53, 55, 57, 59, 61, 63, 65, 67, 69, 71, 73, 75 или 77, или фрагмент или вариант, включая дегенеративный вариант, любой из SEQ ID No: 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 47, 49, 51, 53, 55, 57, 59, 61, 63, 65, 67, 69, 71, 73, 75 или 77.
Трансформированная клетка может представлять собой эукариотическую клетку, например растительную клетку, включая протопласты, или трансформированная клетка может быть прокариотической клеткой, например бактериальной клеткой, такой как E. coli или B. subtilis. Клетки животных включают человеческие клетки, клетки других млекопитающих, частично клетки собаки, клетки птиц и клетки насекомых. Растительные клетки включают, без ограничения, клетки двудольных, однодольных и хвойных растений.
Настоящее изобретение также относится к растениям, включая трансгенные растения, которые экспрессируют и продуцируют вирусный белок или полипептид согласно настоящему изобретению. Растения, растительные ткани и растительные клетки, трансформированные или выращенные таким образом, что они будут содержать полинуклеотид согласно настоящему изобретению, охватываются областью настоящего изобретения. Предпочтительно, полинуклеотид согласно настоящему изобретению подвергается суперэкспрессии в растении, растительной ткани или растительной клетке. Растения могут использоваться для получения противовирусных вакцинных композиций согласно настоящему изобретению и далее такие вакцины могут вводиться в организм при употреблении в пищу этих растений (см., например, патенты США No. 5 484 719 и 6 136 320).
Настоящее изобретение также относится к наборам для выявления вируса или для диагностики инфекции вирусом согласно настоящему изобретению. В одном варианте осуществления настоящего изобретения набор включает антитело согласно настоящему изобретению, которое специфически связывается с вирусом гриппа согласно настоящему изобретению или с его антигенной частью. В другом варианте осуществления настоящего изобретения набор включает один или несколько полипептидов или пептидов согласно настоящему изобретению. В конкретном варианте осуществления настоящего изобретения указанные полипептиды имеют аминокислотную последовательность, показанную в виде любой из SEQ ID No. 2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 33, 34, 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70, 72, 74, 76 или 78, или их функциональный и/или иммуногенный фрагмент или вариант. В еще одном варианте осуществления настоящего изобретения набор включает один или несколько полинуклеотидов или олигонуклеотидов согласно настоящему изобретению. В конкретном варианте осуществления настоящего изобретения указанные полинуклеотиды имеют нуклеотидную последовательность, показанную в виде любой из SEQ ID No. 1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 47, 49, 51, 53, 55, 57, 59, 61, 63, 65, 67, 69, 71, 73, 75 или 77, или их фрагмент или вариант. Набор может необязательно включать одно или несколько контрольных антител, контрольный полипептид или пептид и/или контрольный полинуклеотид или олигонуклеотид. Антитело, полипептиды, пептиды, полинуклеотиды и/или олигонуклеотиды в наборе могут быть введены в подходящий контейнер или соответствующую упаковку.
Настоящая заявка также относится к использованию помесных собак в качестве модели для оценки инфекции и патогенеза заболевания, вызванного вирусом гриппа. В одном варианте осуществления настоящего изобретения помесной собаке инокулируют вирус гриппа, такой как вирус собачьего гриппа согласно настоящему изобретению. Необязательно, указанной собаке после инокуляции также вводят терапевтические агенты. Дополнительно, данной собаке, перед инокуляцией вирусом, может быть также введена композиция, которая вызывает иммунный ответ против вируса гриппа. Образцы ткани, крови, сыворотки и других биологических материалов могут быть отобраны до и/или после инокуляции, после чего устанавливают наличие вируса и исследуют протекающий в ткани патогенез с использованием известных методик, включающих, без ограничения, ПЦР, ОТ-ПЦР, секвенирование нуклеиновой кислоты и иммуногистохимический анализ.
Любой элемент любого из раскрытых в настоящем описании вариантов осуществления настоящего изобретения может быть объединен с любым другим элементом или вариантом, приведенных в данной заявке, и такие сочетания специфически охватываются областью настоящего изобретения.
Все патенты, патентные заявки, предварительные заявки и публикации, относящиеся к данному описанию или процитированные в указанных материалах, включены в настоящее описании полностью, включая все чертежи и таблицы, в той мере, в какой они соответствуют тематике настоящего изобретения и помогают прояснить его описание.
МАТЕРИАЛЫ И МЕТОДЫ, ОТНОСЯЩИЕСЯ К ПРИМЕРАМ 1-6
Отбор образцов крови и назальных мазков у борзых собак
Образцы крови в острый период заболевания и на стадии выздоровления отбирают путем венопункции яремной вены у борзых с клиническими признаками заболевания или у нормальных борзых, находящихся в конурах в период скачек, где выявлены вспышки респираторного заболевания. Образцы, рассматриваемые как образцы со стадии выздоровления, отбирают через 4-12 недель после отбора образца в острой стадии. Получают сыворотку и хранят ее при температуре -80°C. Собирают назальные мазки и помещают их в среду для транспортировки Amies, содержащую древесный уголь (Becton Dickinson Biosciences), с тем, чтобы эти образцы использовать далее для выделения бактерий.
Посмертное обследование борзых
Полное посмертное обследование было проведено в отделе анатомии и патологии Колледжа ветеринарной медицины при Университете во Флориде (Anatomic Pathology Service at the University of Florida College of Veterinary Medicine (UF CVM)) на 5 из 8 собак, которые умерли в январе 2004 г. при вспышке заболевания на соревнованиях во Флориде. Посмертный осмотр другой собаки был проведен в частной ветеринарной клинике в ходе анализа по программе UF CVM для постановки гистопатологического диагноза. Ткани фиксировали в 10% нейтральном забуференном растворе формалина, погруженном в парафин, и срезы толщиной по 5 мкм окрашивали либо гематоксилином, либо эозином, для постановки гистопатологического диагноза, или подвергали обработке с целью подготовки к проведению иммуногистохимического анализа по процедуре, описанной ниже. Нефиксированные ткани подвергают культивированию для оценки бактериального роста и далее также хранят при температуре -80°C.
Серологические тесты для оценки вирусных патогенов, вызывающих респираторное заболевание у собак
Образцы сыворотки крови, полученные в острый период заболевания и на стадии выздоровления, попарно, подвергают анализу в лаборатории диагностики заболеваний животных (Animal Health Diagnostic Laboratory (AHDL)) в Колледже ветеринарной медицины при Корнельском Университете (Cornell University College of Veterinary Medicine) в рамках проведения тестов на способность сыворотки к нейтрализации вируса собачьей чумы, аденовируса типа 2 и вируса парагриппа. Титры антител выражают в виде последнего разведения сыворотки, которое еще ингибирует вирусную инфекцию в клеточных культурах. Сероконверсия, определяемая как ≥ 4-кратное повышение титра антител между образцами, отобранными в остром периоде заболевания и на стадии выздоровления, указывает на наличие вирусной инфекции. Однако, применительно к указанных вирусным патогенам сероконверсия не была выявлена.
Микробные тесты, используемые для анализа бактериальных патогенов, вызывающих респираторное заболевание
Образцы назальных мазков и тканей для посмертного анализа, попарно, направляют в Центр Диагностической Клинической Микробиологии/Паразитологии/Серологии (Diagnostic Clinical Microbiology/Parasitology/Serology Service) в UF CVM для выделения из них бактерий и последующей идентификации. Полученные образцы культивируют на неселективных средах, а также на средах, специфичных для видов Bordetella (Regan-Lowe; Remel) и видов Mycoplasma (Remel). Все культуры выдерживают в течение 21 дня для того, чтобы сделать вывод об отсутствии роста. Назальные мазки, отобранные от некоторых борзых, также направляют в Отдел Диагностической Медицины/Патобиологии Колледжа ветеринарной медицины при Канзасском Государственном Университете (Department of Diagnostic Medicine/Pathobiology at the Kansas State University College of Veterinary Medicine) для культивирования с целью оценки роста бактерий. Из 70 исследованных собак с клиническими признаками заболевания, Bordetella bronchiseptica была выделена из полости носа у 1 собаки, тогда как виды Mycoplasma spp. были выделены из полости носа у 33 собак. Pasteurella multocida обычно выделяется из носовой полости собак с гнойными выделениями из носа. У двух собак, умерших во время вспышки заболевания в январе 2004 г., отмечался очень слабый рост Escherichia coli в посмертных образцах легких, у одной собаки отмечался очень слабый рост E. coli и Streptococcus canis, и у другой собаки отмечался также очень слабый рост Pseudomonas aeruginosa и дрожжевых форм. Ни Bordetella bronchiseptica, ни Mycoplasma не были выделены из трахей или легких умерших собак.
Выделение вируса из посмертных образцов тканей
Замороженные ткани оттаивают и гомогенизируют в 10 объемах минимальной основной среды (MEM) с добавкой 0,5% бычьего сывороточного альбумина (БСА) и антибиотиков. Твердые клеточные остатки удаляют центрифугированием, и супернатанты инокулируют в культивируемые клетки или в куриные яйца с 10-дневными эмбрионами. Тканевые гомогенаты от умерших борзых инокулируют в различные клеточные культуры, которые поддерживают репликацию большого числа вирусных патогенов. Используемые клеточные культуры включают клетки Vero (эпителиальные клетки из почки Африканской зеленой мартышки, ATCC No. CCL-81), A-72 (фибробласты из опухолевой ткани собаки, CRL-1542), HRT-18 (эпителиальные клетки из прямой кишки человека, CRL-11663), MDCK (эпителиальные клетки из почки собаки, CCL-34), первичные эпителиальные клетки из почки собаки (AHDL, Cornell University), первичные эпителиальные клетки из легкого собаки (AHDL) и первичные тестикулярные клетки быка (AHDL). Клетки MDCK и HRT культивируют в среде MEM с добавкой 2,5 мкг/мл TPCK, обработанного трипсином (Sigma); оставшиеся клеточные линии культивируют в среде MEM с добавкой 10% фетальной сыворотки теленка и антибиотиков. Клетки растят в колбах объемом 25 см2 при температуре 37°C в увлажненной атмосфере с содержанием 5% CO2. В контрольную культуру инокулируют среду MEM с добавками. Рост культур оценивают ежедневно с точки зрения появления морфологических изменений и собирают на 5 день после инокуляции. Собранные жидкости и клетки осветляют путем центрифугирования и инокулируют в свежие клетки по той же процедуре, которая была описана выше применительно к начальной инокуляции; в тест вводят два слепых пассажа. Гемагглютинирующую активность в осветленных супернатантах определяют с использованием эритроцитов курицы или индейки, как описано в литературе (Burleson et al., 1992; Kendal et al., 1982). Для выделения вируса из куриных эмбрионов, 0,1 мл тканевого гомогената инокулируют в аллантоиновый мешок с последующей инкубацией в течение 48 часов при температуре 35°C. После двух слепых пассажей, определяют гемагглютинирующую активность в аллантоиновых жидкостях, по описанной в литературе процедуре (Burleson et al., 1992; Kendal et al., 1982).
ОТ-ПЦР, нуклеотидное секвенирование и филогенетический анализ
Суммарную РНК экстрагируют из супернатанта культуры ткани или из аллантоиновой жидкости с использованием набора RNeasy (Qiagen, Valencia, CA), в соответствии с инструкциями производителя. Суммарную РНК (10 нг) подвергают обратной транскрипции до кДНК с использованием набора для одностадийной реакции ОТ-ПЦР (Qiagen, Valencia, CA), в соответствии с инструкциями производителя. Амплификацию по методу ПЦР кодирующей области 8 генов из вируса гриппа проводят по ранее описанной методике (Klimov et al., 1992a) с использованием универсальных наборов ген-специфических праймеров. Полученные ампликоны ДНК используют в качестве матриц для автоматизированного секвенирования на автоматическом секвенаторе ДНК Applied Biosystems 3100 по процедуре секвенирования, в сочетании с окрашиванием красителем в конце цикла (ABI). Нуклеотидные последовательности анализируют с использованием пакета прикладных программ GCG Package© версия 10.0 (Accelyrs) (Womble, 2000). Используют также пакет программ Phylogeny Inference Package© версия 3.5 для оценки филогенетических взаимосвязей и расчета показателей, полученных по методу «расшнурованной выборки» на основе данных нуклеотидных последовательностей (Felsenstein, 1989). Построенные филогенетические деревья сравнивают с деревьями, полученными при анализе по методу анализа ближайших соседей с использованием Tamura-Nei-гамма модели, встроенной в программу MEGA© (Kumar et al., 2004), и подтверждают полученные результаты с использованием программы PAUP© 4.0 Beta (Sinauer Associates).
Эксперименты, включающие инокуляцию собакам
Используют четырех 6-месячных, конкретно подтвержденных как не содержащие патогенов собак бигль [2 самца и 2 самки (Liberty Research)]. Физический осмотр и анализ крови на базовом уровне, включая результаты полного/дифференциального подсчета клеток крови, анализа панели для оценки химических показателей сыворотки, и анализ мочи показывают, что животные были здоровы. Животных поместили вместе в устройство BSL, одобренное Ассоциацией по контролю за содержанием животных, подвергающихся лабораторным экспериментам (Association for Assessment and Accreditation of Laboratory Animal Care). Базовый уровень ректальных температур регистрируют два раза в день в течение 7 дней. Собак подвергают анестезии путем внутривенного введения пропофола (Diprivan®, Zeneca Pharmaceuticals, 0,4 мг/кг веса тела) для интубации эндотрахеальными трубками. Каждой собаке инокулируют суммарную дозу 106,6 средних цитопатогенных доз для культуры тканей (TCID50) вируса A/Canine/Florida/43/2004 (Canine/FL/04) (H3N8), при этом половину дозы вводят в дистальную трахею через эндотрахеальную трубку, а другую половину вводят глубоко в носовой проход с помощью катетера. Физический осмотр и регистрацию ректальной температуры проводят два раза в день в течение 14 дней после инокуляции (p.i.). Образцы крови (по 4 мл) отбирают венопункцией яремной вены в дни 0, 3, 5, 7, 10 и 14 после инокуляции. Назальные и орофарингеальные образцы отбирают с помощью полиэфирных тампонов (Fisher Scientific) от каждой собаки в дни 0-5, 7, 10 и 14 после инокуляции. Тампоны помещают в среды для транспортировки вируса (Remel) и хранят при температуре -80°C. Две собаки (1 самец и 1 самка) были подвергнуты эвтаназии путем внутривенной инокуляции раствора Beuthanasia-D® solution (1 мл/5 кг веса тела; Schering-Plough Animal Health Corp) в 5 день после инокуляции, а оставшихся 2 собак подвергли эвтаназии на 14 день, для посмертного исследования образцов. Ткани для гистологического анализа были подвергнуты обработке по описанной выше процедуре. Ткани для культивирования вирусов хранят при температуре -80°C. Данное исследование было разрешено Комитетом по контролю за обращением с лабораторными животными при Университете Флориды (University of Florida Institutional Animal Care and Use Committee).
Рассеивание вируса от животных, инокулированных в ходе эксперимента
Серийные разведения гомогенов легких и экстрактов тампонов, полученных путем осветления при проведении центрифугирования сред для транспортировки тампонов, проводят в среде MEM с добавкой 0,5% БСА и антибиотиков. Проводят тест на бляшкообразование, как описано в литературе (Burleson et al., 1992), с использованием монослойных клеток MDCK в 6-ячеечных планшетах для культуры тканей. На инокулированные монослои клеток наслаивают среду MEM с добавками, включающими 0,8% агарозы и 1,5 мкг/мл TPCK-трипсина. Клетки культивируют в течение 72 часов при температуре 37°C в увлажненной атмосфере, содержащей 5% CO2, перед фиксацией и окрашиванием кристаллическим фиолетовым. Концентрацию вируса выражают в виде бляшкообразующих единиц (БОЕ) на грамм ткани или на тампон.
Иммуногистохимия
Депрафинизированные и регидратированные срезы из ткани легкого толщиной по 5 мкм, отобранные от борзых и собак бигль, вносят на предметные стекла Bond-Rite™ (Richard-Allan Scientific, Kalamazoo, MI) и затем обрабатывают протеиназой K (DakoCytomation, Carpenteria, CA), с последующей обработкой реагентом, блокирующим пероксидазу (Dako® EnVision™ Peroxidase Kit, Dako Corp.). Срезы инкубируют со взятыми в разведении 1:500 моноклональных антител против вируса чумы собак (VMRD, Inc.), аденовируса собак, типа 2 (VMRD, Inc.), вируса парагриппа собак (VMRD, Inc.) или вируса гриппа A H3 (Chemicon International, Inc.) в течение 2 часов при комнатной температуре. Контроли включают проведение инкубации тех же самых срезов с мышиным IgG (1 мг/мл, Serotec, Inc.) и инкубацию моноклональных антител со срезами из нормальных легких собак. После обработки первичными антителами, срезы инкубируют с вторичными антителами реагентами-субстратами иммунопероксидазы и пероксидазы (Dako® EnVision™ Peroxidase Kit, Dako Corp.), в соответствии с инструкциями производителя. Срезы подвергают контрастному окрашиванию гематоксилином, обрабатывают Clarifier #2 и Bluing реагентом (Richard-Allan Scientific, Kalamazoo, MI), дегидратируют и закрывают покровными стеклами Permount (ProSciTech).
Тест на ингибирование гемагглютинации (HI)
Образцы сыворотки инкубируют с рецептор-разрушающим ферментом (RDE, Denka) (1 часть сыворотки: 3 части RDE) в течение 16 часов при температуре 37°C до проведения тепловой инактивации в течение 60 минут при температуре 56°C. Вирус гриппа A/Canine/FL/04 (H3N8) растят в клетках MDCK в течение 36-48 часов при температуре 37°C. Супернатанты вирусной культуры собирают, осветляют центрифугированием и хранят при температуре -80°C. Проводят HI тест по описанной ранее методике (Kendal et al., 1982). В общих чертах, тест состоит в том, что 4 гемагглютинирующих единицы вируса в 25 мкл добавляют к равному объему серийно разведенной сыворотки в ячейках микротитрационного планшета и инкубируют при комнатной температуре в течение 30 минут. Добавляют равный объем 0,5% (объем/объем) эритроцитов индейки и оценивают титры гемагглютинации через 30 минут. Конечная точка HI титра определяется как последнее разведение сыворотки, которое полностью ингибирует гемагглютинацию. Сероконверсию определяют как ≥ 4-кратное возрастание HI титра между парой образцов, взятых в острой фазе заболевания и в период выздоровления. Серопозитивность одного образца определяют как титр HI антитела ≥1:32.
Тест на микронейтрализацию (MN)
Реакции нейтрализующих антител сыворотки в ответ на вирус A/Canine/FL/04 (H3N8) выявляют в MN тесте по описанной ранее методике (Rowe et al., 1999), с тем исключением, что сыворотки от собак обрабатывают RDE, как было указано выше, перед описанием тестирования. Конечный титр определяют как наивысшее разведение сыворотки, которое приводит к 50% нейтрализации 100 TCID50 вируса. Сероконверсию определяют как ≥ 4-кратное возрастание HI титра между парой образцов, взятых в острой фазе заболевания и в период выздоровления. Серопозитивность одного образца определяют как титр MN ≥1:80.
Ниже приведены примеры, которые иллюстрируют процедуры, используемые в практике осуществления настоящего изобретения. Однако указанные примеры не следует рассматривать в ограничительном контексте. Все процентные показатели даны применительно к весу, и все пропорции в смеси растворителей даны по объему, если особо не указано иное.
ПРИМЕР 1
В январе 2004 произошла вспышка респираторного заболевания у 22 борзых на скачках, размещенных 2 зданиях с конурами для собак на треке во Флориде и на местной ферме, которая поставляла собак в указанные конуры. Было приблизительно 60 животных в каждом из зданий для собак и 300 собак - на ферме. Вспышка произошла в течение 6-дневного периода, после которого не было идентифицировано новых случаев. У четырнадцати из 22 собак отмечалась лихорадка с температурой от 39,5 до 41,5°C, слабый, рвотный кашель в течение периода времени от 10 до 14 дней, с постепенным выздоровлением. Среди оставшихся 8 собак, 6 явно здоровых неожиданно умерли от кровотечения изо рта и носа. Две другие собаки были подвергнуты эвтаназии в течение 24 часов в связи с тем, что кровотечение изо рта и носа быстро усиливалось. Обе этих собаки имели лихорадку с температурой до 41°C. Четыре из 8 летальных случаев произошли в зданиях с конурами собак, и 4 произошли на ферме. Пятьдесят процентов летальных исходов произошло на 3 день вспышки заболевания. Указанные 22 собаки были в возрасте от 17 месяцев до 4 лет, и 73% были в возрасте от 17 до 33 месяцев.
Были отмечены два клинических синдрома: слабое заболевание, характеризующееся вначале лихорадкой и затем кашлем в течение 10-14 дней (у 14 собак) с последующим выздоровлением, или подострое течение со смертельным исходом, ассоциированным с кровотечением в дыхательных путях (8 собак, при среднем уровне смертности 36%). Посмертные исследования проводились в 6 из 8 летальных случаев. У всех собак отмечалась обширная геморрагия в легких, средостении и плевральной полости. Гистологическое исследование дыхательных путей показало, что, кроме легочной геморрагии, у всех собак отмечался трахеит, бронхит, бронхиолит и гнойная бронхопневмония (фиг.3). Эпителиальная выстилка и просвет дыхательных путей в этих тканях инфильтрованы нейтрофилами и макрофагами. Гомогенаты легких, полученные из указанных собак, инокулируют в большое число клеточных линий обезьян, человека, быка и собаки для культивирования вируса. Гомогенат легкого от одной собаки вызывал цитопатические эффекты в эпителиальных клетках почки собаки Madin-Darby (MDCK), культивируемых в присутствии трипсина, а клеточный супернатант вызывал агглютинацию куриных эритроцитов. Предварительные данные о том, что это был вирус гриппа типа A, были получены при проведении метода ELISA, в его коммерческом варианте, для детекции нуклеопротеина вирусов гриппа A и B, а также по результатам анализа по методу ПЦР с использованием праймеров, специфичных для матричного гена вирусов гриппа А. Кроме того, гемагглютинирующая активность ингибировалась эталонной антисывороткой против лошадиного вируса гриппа А, подтипа H3, но не антисывороткой, специфичной для человеческого вируса гриппа А, подтипов H1-H11 и H13 (Таблица 3). Для характеристики молекулярных свойств вируса авторы определили нуклеотидные последовательности 8 сегментов РНК в вирусном геноме. Сравнение последовательностей с известными генами вируса гриппа и результаты филогенетического анализа указывают на то, что 8 генов изолята от собаки были наиболее близки к современным изолятам вирусов гриппа лошади A (H3N8), с которым они имели идентичность по последовательности ≥96-97% (фиг.1A, Таблица 4). И наоборот, репрезентативные гены из изолятов вируса гриппа А из птиц, свиньи и человека характеризовались ≤94% идентичностью с изолятом из собаки (Таблица 4). Эти данные позволили идентифицировать изолят из собаки A/Canine/Florida/43/04 (Canine/FL/04) как вирус гриппа A H3N8, очень близкий к современной линии вирусов гриппа лошади. Поскольку все гены изолята из собаки происходят из вируса гриппа лошади, авторы сделали вывод о том, что полный геном вируса гриппа лошади был перенесен к собаке.
ПРИМЕР 2
Для исследования роли вируса Canine/FL/04 в проявлении клинических и патологических признаков у борзых авторы провели иммуногистохимическое окрашивание (IHC) тканей легкого с использованием моноклонального антитела против вируса гриппа A H3. Вирусный антиген H3 неизменно выявляется в цитоплазме эпителиальных клеток бронхов и бронхиол, эпителиальных клетках бронхиальной железы, а также в макрофагах в просвете дыхательных путей и в альвеолярном пространстве (фиг.2A). Эти данные подтверждают диагноз инфекции легкого, вызванной вирусом H3 подтипа, у множества собак.
ПРИМЕР 3
Для установления участия вируса, подобного вирусу Canine/FL/04, в этиологии вспышки респираторного заболевания, авторы проанализировали, попарно, сыворотку, взятую в остром периоде заболевания и в фазе выздоровления у 11 больных собак и у 16 животных, бывших в контакте, но не демонстрирующих симптомов заболевания, с использованием тестов на ингибирование гемагглютинации (HI) и на микронейтрализацию (MN). Сероконверсия, определяемая как ≥ 4-кратное повышение титра антител против Canine/FL/04, и которая имела место в период от острой фазы заболевания до стадии выздоровления, отмечалась 8 из 11 (73%) больных собак, по данным обоих тестов (Таблица 1). Сероконверсия была выявлена у 6 из 16 (38%) животных без симптомов заболевания, которые были в контакте с больными, по данным HI теста, тогда как по данным MN теста (Таблица 1), сероконверсия имеет место у 8 из 16 (50%) исследованных животных. Данные по оценке сероконверсии указывают на наличие инфекции собак вирусом, подобным вирусу Canine/FL/04, которая совпала по времени с началом респираторного заболевания у большинства животных.
Через 3 месяца после вспышки заболевания отобрали отдельные образцы сыворотки еще у 46 собак, которые не имели симптомов заболевания, но которые были размещены вместе с больными собаками. Из них, по данным обоих тестов, 43 животных (93%) были серопозитивными. Из общей популяции 73 исследованных собак, как было выявлено по данным обоих тестов, 93% животных были серопозитивными, включая 82% (9/11) больных собак и 95% (59/62) здоровых животных, бывших в контакте с больными. Высокий уровень серопозитивности у собак, у которых ранее не было зарегистрировано респираторное заболевание, указывает на то, что по большей части инфекция вирусом собачьего гриппа является субклинической по своему течению, что предполагает эффективное распространение указанного вируса в популяции собак. Однако, пока неизвестно, вносят ли субклинические инфекции какой-либо вклад в распространение вируса.
ПРИМЕР 4
С тем, чтобы наилучшим образом определить способность вируса Canine/FL/04 инфицировать собак, четырем 6-месячным выращенным по специальной программе щенкам бигль инокулировали 106,6 средних цитопатогенных доз для культуры (TCID50) путем интратрахеального и интраназального введения. У всех собак в течение первых 2 дней после инокуляции (p.i.) появляется лихорадка (с ректальной температурой ≥39°C), однако, ни у одной из них не отмечаются респираторные симптомы, такие как кашель или выделения из носа, в течение 14-дневного периода наблюдения. Тест на выделение вируса проводят путем количественной оценки содержания вируса в назальных и орофарингеальных мазках. Только у 2 животных из 4 были выявлены заметные количества вируса. У одного животного вирус выделялся в 1 и 2 дни после инокуляции (1,0-2,5 log10 БОЕ на мазок), тогда как у другой собаки вирус выделялся в течение 4 последовательных дней после инокуляции (1,4-4,5 log10 БОЕ на мазок). Посмертное исследование 2 собак на 5 день после инокуляции выявило некротизирующий и гиперплазирующий трахеит, бронхит, бронхиолит, аналогично тем изменениям, которые наблюдались при спонтанном развитии заболевания у борзых собак, но при этом не отмечалось легочной геморрагии или бронхопневмонии. Вирусный антиген H3 выявлялся в цитоплазме эпителиальных клеток бронхов, бронхиол и бронхиальных желез, по данным иммуногистохимического анализа (фиг.2B). У одной собаки инфицирующий вирус восстанавливался из ткани легкого. Посмертное исследование 2 оставшихся собак на 14 день после инокуляции выявило минимальные гистологические изменения в тканях дыхательных путей, тогда как по данным иммуногистохимического анализа не было выявлено антигена H3, а также из гомогенатов легкого не выделялся вирус. Сероконверсия у этих 2 собак была детектирована по данным MN тестирования, проведенного на 7 после инокуляции, а к 14 дню отмечалось дальнейшее повышение титра антител в 2-3 раза. Полученные результаты указывают на чувствительность собак к инфекции вирусом Canine/FL/04, как следует из результатов продемонстрированной у животных фебрильной реакции, наличия вирусного антигена и инфицирующего вируса в паренхиме легкого, гистопатологических показателей, характерных для гриппа, и сероконверсии. Тот факт, что у собак бигль, которым провели инокуляцию в ходе эксперимента, не развилось тяжелое заболевание с последующим летальным исходом, не стал неожиданностью, поскольку большая часть естественно инфицированных борзых характеризовалась бессимптомным течением заболевания.
ПРИМЕР 5
Для выяснения того, циркулировал ли вирус гриппа, подобный вирусу Canine/FL/04, в популяции борзых во Флориде до вспышки заболевания в январе 2004 г., были исследованы архивные образцы сывороток крови от 65 борзых, с целью установления наличия антител к вирусу Canine/FL/04, с использованием HI и MN тестов. В образцах из 33 собак, отобранных в период 1996-1999 гг., не было продемонстрировано выявляемых количеств антител. Из образцов 32 собак, отобранных в период 2000-2003 гг., 9 образцов были определены как серопозитивные, по данным обоих тестов, при этом 1 из них в 2000 г., 2 - в 2002 г. и 6 образцов - в 2003 г. (Таблица 5). Серопозитивные собаки, размещенные на треках во Флориде, были вовлечены в развитие вспышек респираторного заболевания неясной этиологии, имевшего место в период 1999-2003 гг., что дает основания полагать, что вирус, подобный вирусу Canine/FL/04, мог быть тем агентом, который вызывал эти вспышки. Для дальнейшего исследования этого предположения, авторы исследовали архивные образцы тканей тех борзых, которые умерли от геморрагической бронхопневмонии в марте 2003 г. Для этого проводили экспериментальную инокуляцию в клетки MDCK и куриные эмбрионы гомогенатов легкого, полученных от одной собаки с гриппом, вызванным вирусом H3N8, обозначенным как A/Canine/Florida/242/2003 (Canine/FL/03). Анализ последовательности полного генома вируса Canine/FL/03 показал наличие >99% идентичности к последовательности вируса Canine/FL/04 (Таблица 4), указывая на то, что вирусы, подобные вирусу Canine/FL/04, инфицировали борзых еще до 2004 г.
ПРИМЕР 6
В период с июня по август 2004 г. вспышки респираторного заболевания регистрировались у тысяч борзых на 14 треках во Флориде, Техасе, Алабаме, Арканзасе, Западной Вирджинии и Канзасе.
По официальным данным, на ряде из этих треков по меньшей мере до 80% собак в популяциях имели клиническую форму заболевания. Большая часть собак характеризовалась наличием клинических признаков лихорадки (температура ≥ 39°C) и кашля, аналогично тем признакам, которые были зарегистрированы во время вспышки заболевания в 2004 г., однако у многих собак имелись также гнойно-слизистые выделения из носа. Было зарегистрировано большое число летальных исходов, однако точный уровень смертности не установлен.
Авторы отобрали и проанализировали, попарно, образцы сыворотки крови, отобранные у 94 собак в остром периоде заболевания и на фазе выздоровления, размещенных на 4 треках во Флориде: 56% указанных собак демонстрировали ≥4-кратное повышение титра антител против вируса Canine/FL/04, и 100% были серопозитивными (Таблица 6). В сыворотке, взятой на фазе выздоровления у 29 собак в Западной Вирджинии и Канзасе, также выявлялись антитела против вируса Canine/FL/04. Авторы выделили вирус гриппа A (H3N8) из легких борзых, которые умерли от геморрагической бронхопневмонии на треке в Техасе. Анализ полноразмерной последовательности данного изолята, обозначенного как A/Canine/Texas/1/2004 (Canine/TX/04), выявил наличие ≥99% идентичности к Canine/FL/04 (Таблица 4). Выделение трех близкородственных вирусов гриппа от трех умерших собак, где указанные летальные случаи имели место в течение 13-месячного периода времени и были локализованы в различных географических областях, в сочетании со значимыми серологическими данными о распространении инфекции среди участвовавших в скачках борзых, дали основание говорить о длительной циркуляции вируса, подобного вирусу Canine/FL/04, в популяции собак.
Филогенетический анализ, проведенный на генах HA из вирусов Canine/FL/03, Canine/FL/04 и Canine/TX/04, показал, что они составляют монофилетическую группу, которая соответствует жестким критериям оценки по методу «расшнурованной выборки» и явно отличается от современных генов H3 из вирусов лошади, выделенных в 2002 и 2003 гг. (фиг.1B). Результаты филогенетического анализа и сравнения нуклеотидных последовательностей других 7 сегментов генома, проведенного попарно, подтверждают факт сегрегации генов собаки в качестве четко выделенной сублинии, которая ближе всего стоит к линии лошадиного вируса (данные не проиллюстрированы, а также Таблица 4). Выделение генов собачьего гриппа в виде монофилетической группы, отделенной от лошадиного вируса, также подтверждается наличием 4 характерных изменений в аминокислотном составе HA (Таблица 2). В сочетании с серологическими результатами, полученными в период с 2003 по 2004 гг., эти данные согласуются с предположением о феномене однократной передачи вируса от лошадей к собакам, с последующим горизонтальным распространением вируса в популяции борзых. Однако повторное встраивание уникальной линии вируса гриппа из неидентифицированного вида, в качестве резервуара инфекции, формально не может быть исключено, хотя это маловероятно.
Вирусный антиген HA представляет собой ключевую детерминанту, определяющую видоспецифичность вируса (Suzuki et al., 2000). Для идентификации остатков в составе HA, которые могут быть ассоциированы с адаптацией к организму собаки, как к организму-хозяину, авторы провели сравнение расчетной аминокислотной последовательности HA собачьих вирусов с соответствующей последовательностью, характерной для современных лошадиных вирусов. Наличие четырех изменений на уровне аминокислот определяют различие между зрелыми консенсусными аминокислотными последовательностями НА лошади и собаки: N83S, W222L, I328T и N483T (см. Таблица 2). Вирусы собаки содержат делецию аминокислоты, в сравнении с консенсусными последовательностями лошадиных вирусов. В этой связи аминокислота в положении 7 в последовательности лошадиного HA соответствует аминокислоте в положении 6 в последовательности HA из вирусов собаки, аминокислота в положении 29 в последовательности лошадиного HA соответствует аминокислоте в положении 28 в последовательности HA из вирусов собаки, аминокислота в положении 83 в последовательности лошадиного HA соответствует аминокислоте в положении 82 в последовательности HA из вирусов собаки и т.п. Таким образом, четыре замещенных аминокислоты локализованы в положениях 82, 221, 327 и 482 в аминокислотной последовательности, показанной в виде SEQ ID NO: 33 и SEQ ID NO: 34. Замещение серина на аспарагин в положении 83 в консенсусной последовательности представляет собой изменение неизвестной функциональной значимости, поскольку различные полярные остатки обнаружены в молекулах H3 из других видов. Строго консервативный изолейцин, локализованный в положении 328 в консенсусной последовательности, рядом с сайтом расщепления H3 HA, был замещен треонином. Жизненно важная роль расщепления HA протеазами организма-хозяина в ходе патогенеза дополнительно указывает на то, что это изменение заслуживает дальнейшего исследования. Замещение лейцина на триптофан в положении 222 в консенсусной последовательности является весьма примечательным феноменом, поскольку представляет собой неконсервативное изменение, в непосредственной близости от связывающего кармана сиаловой кислоты, что может привести к модуляции рецепторной функции (Weis et al., 1988). Интересно отметить, что лейцин в положении 222 не является уникальной характеристикой собачьего H3 HA, поскольку, в типичном случае, он обнаруживается в H4, H8, H9 и H12 подтипах HA (Nobusawa et al., 1991; Kovacova et al., 2002). Замещение лейцина может быть более совместимо с сохранением специфичности вируса в отношении млекопитающих, в качестве организмов-хозяев, поскольку имеются сообщения об инфекции свиней подтипом H4 (Karasin et al., 2000), а также и людей, и свиней подтипом H9 (Peiris et al., 1999) вирусов. Замещение аспарагина треонином в положении 483 в консенсусной последовательности приводит к потере сайта гликозилирования в субъединице HA2, которая сохраняется, в качестве консервативного элемента, во всех подтипах HA (Wagner et al., 2002). Несмотря на то что еще предстоит определить значимость указанных аминокислотных замен в составе HA, с точки зрения адаптации лошадиного вируса к организму собак, аналогичные изменения в составе аминокислот ранее были выявлены применительно к межвидовой передаче (Vines et al., 1998; Matrosovich et al., 2000). Различия на уровне аминокислот между другими белками вируса гриппа согласно настоящему изобретению и соответствующей консенсусной последовательностью в лошадином вирусе проиллюстрированы в Таблицах 19-25.
Соображения относительно источника вируса лошадиного гриппа по сей день остаются спекулятивными. Строения с конурами для собак на треках в районе скачек борзых не расположены рядом с лошадьми или с лошадиными рейстреками, и этот факт свидетельствует о том, что контакт между борзыми собаками и распространяющими инфекцию лошадьми является несостоятельным объяснением множества вспышек заболевания, имевших место в разных штатах в 2004 г. Потенциальным источником воздействия лошадиного вируса является поедание борзыми собаками лошадиной пищи, рацион которых включает добавку сырого мяса, поставляемого мясокомбинатами, на которых разделываются туши, включая туши лошадей, которые могли быть носителями вируса гриппа. Прецеденты такого способа инфекции описаны применительно к межвидовой передаче вируса птичьего гриппа H5N1 к свиньям и кошачьим в зоопарке, которых кормили тушками инфицированных кур (Webster, 1998; Keawcharoen et al., 2004; Kuiken et al., 2004). Хотя этот путь внедрения вируса лошадиного гриппа в организм собак выглядит достаточно правдоподобным, он не может объяснить имевшиеся в недавнее время многочисленные вспышки гриппа у тысяч собак в разных штатах. Результаты авторов, полученные в ходе экспериментальной инокуляции, продемонстрировали наличие вируса в носовых проходах и мезофаринксе у собак, хотя и с умеренными значениями титра. Тем не менее, полученные результаты указывают на то, что выделение вируса в среду возможно, и что передача вируса от собаки к собаке в составе крупных аэрозольных капель, через предметы или путем непосредственного контакта через слизистые, могли сыграть определенную роль в эпизоотиологии заболевания.
Межвидовая передача полноразмерного вируса гриппа млекопитающих к неродственным видам млекопитающих представляет собой весьма редкое событие. Предшествующие исследования предоставили ограниченные доказательства, серологического или вирусологического характера, но не одновременно обоих видов, временного инфицирования собак вирусами человеческого гриппа A (H3N2) (Nikitin et al., 1972, Kilbourne, et al., 1975; Chang et al., 1976; Houser et al., 1980). Однако до сих пор отсутствуют доказательства возможностей длительной циркуляции вируса в организме псовых, в качестве хозяев. Несмотря на то что возможность непосредственной передачи вирусов свиного гриппа от свиней к людям хорошо установлена и описана (Dacso et al., 1984; Kimura et al., 1998; Patriarca et al., 1984; Top et al., 1977), отсутствуют достоверные данные, свидетельствующие о возможности их адаптации в организме человека в качестве хозяина. В настоящем описании авторы предоставляют вирусологические, серологические и молекулярные доказательства межвидовой передачи полноразмерного вируса лошадиного гриппа A (H3N8) к другому виду млекопитающих, а именно к собаке. Данные по уникальным замещениям в HA вируса собаки, в сочетании с серологическим подтверждением инфекции у собак во многих штатах США, указывают на возможную адаптацию вируса к организму собаки в качестве хозяина. Поскольку собаки являются основным спутником человека, полученные результаты имеют значимость и для охраны здоровья людей, поскольку собаки могут выполнять роль нового источника в передаче новых вирусов гриппа A к людям.
Образование антител против вируса A/Canine/Florida/43/04 (H3N8)
b Число собак без симптомов заболевания, размещенных в контакте с собаками, имеющими клинические признаки заболевания.
c Тест на ингибирование гемагглютинации (HI), проводимый с использованием вируса A/Canine/Florida/43/04.
d Тест на микронейтрализацию (MN), проводимый с использованием вируса A/Canine/Florida/43/04.
e Процент собак, демонстрирующих по меньшей мере 4-кратное повышение титра антител, при попарном исследовании сывороток, отобранных в остром периоде заболевания и в фазе выздоровления.
f Процент собак, демонстрирующих положительный титр антител (HI титр ≥32: MN титр ≥80) в сыворотке, взятой в фазе выздоровления.
g Среднее геометрическое значение титра в сыворотке, взятой в фазе выздоровления.
Аминокислотные различия между Н3 гемагглютининами
собаки и лошади
† Означает отсутствие изменения относительно консенсуса лошадиных H3 HA.
b Поликлональные антисыворотки получают из африканского хорька, а все другие антисыворотки получают из овцы или коз.
DQ124147
Eq/Kentucky/2/8
M73526
Mall/Alberta/98/85
AY633315
Sw/Ontario/
01911-1/99
AF285892
PR/8/34 (HK/213/03)
AF389115 (AY576381)
DQ124148
Eq/Tennessee/5/86
M25929
Ck/BritishColumbia/04
(Gull/Md/704/77)
AY61675 (M25933)
Sw/Korea/S109/04
(Sw/Saskatch/
18789/02)
AY790287 (AY619955)
WSN/33 (Sing/1/57)
J02178 (M25924)
DQ124149
M26082
Eq/Tennesee/5/86
Ck/Chile/4591/02
(Ostrich/SA/08103/95)
AY303660(AF508662)
Sw/Hong Kong/
126/02
M26081
Taiwan/2/70
(Viet Nam/
1203/04)
AY210199 (AY818132)
DQ124190
Eq/FL/1/93
L39916
Dk/Norway/1/03
AJ841293
Sw/Ontario/42729a/01
AY619977
HK/1/68
AF348176
DQ124150
Eq/Tennesee/5/86
M30758
Ck/Chile/176822/02
AY303658
Sw/Ontario/42729a/01
(Sw/Fujian/1/2003)
AY619974 (AY747611)
HK/1073/99
(Hong Kong /538/97)
AF255742 (AF255751)
DQ124151
Eq/Tennesee/5/86
L06583
Dk/NJ/2000
L06583
DQ124152
Eq/Tennesee/5/86
(Eq/Kentucky/92)
M63529 (AF001683)
Tky/Mn/833/80
AF001683
Sw/Saskatchewan/
18789/02
M63527
WSN/33
(Hong Kong/
1073/99)
J02177 (AJ278646)
DQ124153
Eq/Tn/5/86
(Eq/Kentucky/92)
M80973 (AF001671)
Mal/NY/6750/78
M80945
Sw/China/8/78
(Sw/Korea/s452/04)
M80968 (AY790309)
Brevig Mission/1/18
AF333238
обозначением изолята).
b Не применимо: нет ни одного сообщения о наличии нейраминидазы N8 в человеческих или свиных вирусах.
с 1996 по 2003 гг.
b Титры антител в тесте на микронейтрализацию у серопозитивных собак, включая диапазон, выявленный для шести серопозитивных собак в 2003 г.
b Процент собак с ≥4-кратным повышением титра антител в сыворотках, выявленным в период между острой фазой заболевания и стадией выздоровления.
c Процент собак с позитивным титром антител (титр HI >16) в сыворотках, отобранных в фазе выздоровления.
d Среднее геометрическое значение титра в сыворотках, отобранных в фазе выздоровления.
МАТЕРИАЛЫ И МЕТОДЫ, ОТНОСЯЩИЕСЯ К ПРИМЕРАМ 7-11
Ткани, отобранные у собак
Посмертные исследования животных проводят в отделе анатомии и патологии Колледжа ветеринарной медицины при Университете во Флориде (Anatomic Pathology Service at the University of Florida College of Veterinary Medicine (UF CVM)) на 6 помесных собаках, которые умерли в апреле/мае 2005 г. при вспышке заболевания в приюте на северо-востоке Флориды, и на йоркширском терьере, который умер в мае 2005 г. при вспышке заболевания в ветеринарной клинике на юго-востоке Флориды. Ткани фиксировали в 10% нейтральном забуференном растворе формалина, погруженном в парафин, и срезы толщиной по 5 мкм окрашивали гематоксилином и эозином для постановки гистопатологического диагноза. Нефиксированные ткани хранили при температуре -80°C до проведения вирусологических анализов.
Экстракция РНК из образцов тканей собак
Замороженные ткани легкого, взятые от каждой из 7 собак, оттаивают и гомогенизируют в минимальной основной среде (MEM), содержащей, в качестве добавок, 0,5% бычий сывороточный альбумин (БСА) и антибиотики (гентамицин и ципрофлоксацин), с использованием одноразового измельчителя тканей (Kendall, Lifeline Medical Inc., Danbury, CT). Суммарную РНК экстрагируют с использованием коммерческого набора RNeasy (RNeasy® Mini Kit, QIAGEN Inc., Valencia, CA), в соответствии с инструкциями производителя, и проводят элюцию буфером, взятым в конечном объеме 60 мкл. Суммарную РНК также экстрагируют из тканей легких, собранных от собак, не имевших респираторного заболевания.
ОТ-ПЦР в масштабе реального времени
Одностадийную количественную ОТ-ПЦР в масштабе реального времени проводят на суммарной РНК, экстрагированной из образцов тканей собак, с использованием набора зондов для проведения ОТ-ПЦР (QuantiTect® Probe RT-PCR Kit), содержащего ROX в качестве пассивного эталонного красителя (QIAGEN Inc., Valencia, CA). В общих чертах, процедура состоит в том, что для каждого образца используют 2 набора праймерных зондов для детекции последовательностей вируса A (Таблица 7). Один набор праймерных зондов является селективным для последовательностей гена гемагглютинина собаки (H3). Другой набор праймерных зондов направлен на высококонсервативный участок матричного гена (M) вируса гриппа типа A. При проведении каждой из реакций по процедуре ОТ-ПЦР в масштабе реального времени добавляют 5 мкл экстрагированной суммарной РНК к реакционной смеси, содержащей 12,5 мкл 2Х основной смеси зондов для ОТ-ПЦР (2X QuantiTech® Probe RT-PCR Master Mix), 0,25 мкл смеси ОТ (QuantiTech® RT Mix), обратные и прямые праймеры (в конечной концентрации каждого из них 0,4 мкМ), зонд (в конечной концентрации 0,1 мкМ) и не содержащую РНК-азы воду, в конечном объеме 25 мкл. Используют также рибосомальную РНК, в качестве контрольных реагентов (TaqMan® Ribosomal RNA Control Reagents (Applied Biosystems, Foster City, CA)), в соответствии с инструкциями производителя, для детекции 18S рРНК в качестве эндогенного внутреннего контроля на присутствие РНК, экстрагированной из образцов тканей собак.
Количественную одностадийную ОТ-ПЦР в масштабе реального времени проводят при введении реакционных смесей в систему Mx3000P® QPCR (Stratagene, La Jolla, CA). Условия циклической обработки включают стадию обратной транскрипции, проводимую при температуре 50°C в течение 30 минут, стадию начальной денатурации, проводимую при температуре 95°C в течение 15 минут, для активации ДНК-полимеразы HotStarTaq®, и стадию амплификации, проводимую в течение 40 циклов. Каждый цикл амплификации включает денатурацию при температуре 94°C в течение 15 секунд, с последующим проведением стадии отжига/наращивания при температуре 60°C в течение 1 минуты. Сигналы FAM (длина волны эмиссии при 518 нм) и VIC (длина волны эмиссии при 554 нм) флуоресценции регистрируют в конце каждого цикла. Пороговый показатель цикла (Ct) определяют при установке пороговой флуоресценции (dR) на 1000 в каждом отдельном эксперименте. Для регистрации и анализа данных используют пакет прикладных программ Mx3000P® версия 2.0 (Stratagene, La Jolla, CA). Образцы рассматриваются как позитивные в отношении вируса гриппа A, если пороговый показатель цикла (Ct) для H3 или M гена на 3 единицы меньше, чем значение Ct для тканей легких у собак, не имеющих респираторного заболевания. Позитивный контроль включает проведение амплификации РНК, экстрагированной из вируса A/canine/FL/242/03 (H3N8).
Выделение вируса из клеток MDCK
Замороженные ткани легких от каждой из 7 собак оттаивают и гомогенизируют в 10 объемах среды Игла в модификации Дульбекко (DMEM) с добавкой 0,5% БСА и антибиотиков (гентамицин и ципрофлоксацин). Твердые остатки клеток удаляют центрифугированием, и супернатанты используют для инокуляции в клетки почки собаки Madin-Darby (MDCK), культивируемые в среде DMEM с добавкой 1 мкг/мл TPCK при обработке трипсином (Sigma-Aldrich Corp., St. Louis, MO) и антибиотиков (гентамицин и ципрофлоксацин). Клетки растят в колбах объемом 25 см2 при температуре 37°C в увлажненной атмосфере, содержащей 5% CO2. Культуры ежедневно осматривают для выявления наличия морфологических изменений и собирают через 5 дней после инокуляции. Собранные культуры осветляют центрифугированием, и супернатанты используют для инокуляции в свежие клетки MDCK, как было описано применительно к начальной инокуляции; проводят еще два дополнительных пассажа для образцов, которые не демонстрируют наличия вируса гриппа, по данным оценки гемагглютинации или по результатам реакции ОТ-ПЦР. Гемагглютинирующую активность в осветленных супернатантах определяют с использованием 0,5% эритроцитов индейки, в соответствии с описанной ранее методикой (Burleson, F. et al., 1992; Kendal, P. et al., 1982). ОТ-ПЦР проводят по описанной ниже процедуре.
Выделение вируса из эмбрионов в куриных яйцах
Получают гомогенаты из замороженных тканей, как было описано применительно к инокуляции в клетки MDCK. Гомогенаты (0,2 мл) используют для инокуляции в аллантоиновый мешок 10-дневных эмбрионов в куриных яйцах. После 48 часов инкубации при температуре 35°C яйца охлаждают при температуре 4°C в течение ночи, перед отбором аллантоиновой жидкости. Гемагглютинирующую активность в осветленных супернатантах определяют с использованием 0,5% эритроцитов индейки, в соответствии с описанной ранее методикой (Burleson, F. et al., 1992; Kendal, P. et al., 1982). ОТ-ПЦР проводят по описанной ниже процедуре. Проводят еще два дополнительных пассажа в яйцах с эмбрионами для тех образцов, которые не продемонстрировали наличия вируса после первоначальной инокуляции.
ОТ-ПЦР, секвенирование нуклеотидов и филогенетический анализ
Суммарную РНК экстрагируют из супернатанта культуры клеток MDCK или из аллантоиновой жидкости с использованием мининабора для выделения вирусной РНК QIAamp® Viral RNA Mini Kit (QIAGEN Inc., Valencia, CA), в соответствии с инструкциями производителя. Вирусную РНК подвергают обратной транскрипции до кДНК с использованием набора для проведения одностадийной реакции ОТ-ПЦР QIAGEN® OneStep RT-PCR Kit (QIAGEN, Valencia, CA), в соответствии с инструкциями производителя. Амплификацию по методу ПЦР кодирующей области 8 генов из вируса гриппа проводят по описанной ранее методике (Klimov et al., 1992a) с использованием универсальных наборов ген-специфических праймеров (праймерные последовательности доступны по запросу). Полученные ампликоны ДНК используют в качестве матриц для автоматизированного секвенирования на автоматическом секвенаторе ДНК ABI PRISM® 3100 по процедуре циклического секвенирования, включающего использование красителя в конце цикла (Applied Biosystems, Foster City, CA). Нуклеотидные последовательности анализируют с использованием пакета прикладных программ Lasergene 6 Package® (DNASTAR, Inc., Madison, WI). Используют также пакет программ PHYLIP версия 3.5© для оценки филогенетических взаимосвязей и расчета показателей, полученных при анализе по методу «расшнурованной выборки», на основе данных по нуклеотидным последовательностям (Felsenstein, 1989). Построенные филогенетические деревья сравнивают с деревьями, полученными при анализе по методу анализа ближайших соседей с использованием Tamura-Nei-гамма модели, встроенной в программу MEGA© (Kumar et al., 2004), и подтверждают полученные результаты с использованием программы PAUP© 4.0 Beta (Sinauer Associates, Inc., Sunderland, MA).
Тест на ингибирование гемагглютинации (HI)
Образцы сыворотки инкубируют с рецептор-разрушающим ферментом (RDE, DENKA SEIKEN Co., Ltd., Tokyo, Japan) (1 часть сыворотки: 3 части RDE) в течение 16 часов при температуре 37°C до проведения тепловой инактивации в течение 60 минут при температуре 56°C. Вирус гриппа A/Canine/Jacksonville/05 (H3N8) растят в клетках MDCK в течение 72 часов при температуре 37°C в атмосфере 5% CO2. Супернатанты вирусной культуры собирают, осветляют центрифугированием и хранят при температуре -80°C. Все другие вирусы, используемые в HI тесте, растят в куриных яйцах с 10-дневными эмбрионами, из которых отбирают аллантоиновую жидкость и хранят при температуре -80°C. Проводят HI тест по описанной ранее методике (Kendal et al., 1982). В общих чертах, тест состоит в том, что 4 гемагглютинирующих единицы вируса в 25 мкл добавляют к равному объему серийно разведенной сыворотки в ячейках 96-ячеечного пластикового микротитрационного планшета и инкубируют при комнатной температуре в течение 30 минут. Добавляют равный объем 0,5% (объем/объем) эритроцитов индейки, и визуально оценивают титры гемагглютинации через 30 минут. Конечная точка HI титра определяется как последнее разведение сыворотки, которое полностью ингибирует гемагглютинацию.
ПРИМЕР 7 - КЛИНИЧЕСКИЕ СЛУЧАИ
В апреле и мае 2005 г. произошла описанная ранее (Crawford, P.C. et al., 2005) вспышка респираторного заболевания у собак, размещенных в приюте на северо-востоке Флориды. Вспышка охватила 58 собак в возрасте от 3 месяцев до 9 лет и вовлекла как чистопородных собак, так и помесных собак. Самыми распространенными общими для всех клиническими признаками заболевания были гнойные выделения из носа и кашель, продолжавшиеся от 7 до 21 дня. Из 43 собак, демонстрировавших клинические проявления заболевания в течение ≥7 дней, 41 имела титры HI антител против вируса canine/FL/04 (H3N8) в диапазоне от 32 до >1024. По меньшей мере, у 10 собак заболевание прогрессировало до пневмонии, из числа которых 6 животных были подвергнуты эвтаназии. Указанные 6 помесных собак включали 3 самцов и 3 самок в возрасте от 4 месяцев до 3 лет. Длительность проявления клинических признаков варьировала от 2 до 10 дней, как времени эвтаназии. При посмертном осмотре было обнаружено, что эти собаки имели застой и отек легких. Гистологическое исследование дыхательных путей показало наличие ринита, трахеита, бронхита, бронхиолита и гнойной бронхопневмонии. Отмечались явления некроза и эрозии эпителиальных клеток в трахее, бронхах, бронхиолах и бронхиальных железах. Ткани дыхательных путей, как было показано, инфильтрованы нейтрофилами и макрофагами.
В мае 2005 г. вспышка респираторного заболевания охватила 40 домашних собак, находившихся в ветеринарной клинике на юго-востоке Флориды. Самыми распространенными общими для всех клиническими признаками заболевания были гнойные выделения из носа и кашель, продолжавшиеся от 10 до 30 дней. Из 40 указанных собак, 17 были серопозитивными по вирусу canine/FL/04 (H3N8), где титры HI антител варьировали от 32 до >1024. Сероконверсия была выявлена у 10 собак, для которых было возможно провести спаренный тест на сыворотках, отобранных в остром периоде заболевания и на фазе выздоровления. У трех собак заболевание прогрессировало до пневмонии. Одна из этих собак, 9-летний самец йоркширского терьера, умерла через 3 дня после появления клинических признаков заболевания. У этой собаки были выявлены трахеобронхит, отек легких и застойные явления в легких, а также тяжелая бронхопневмония. Аналогично 6 собакам, заболевшим в приюте, в ее дыхательных путях были выявлены некроз и эрозия эпителиальных клеток, а также нейтрофильная инфильтрация в тканях.
ПРИМЕР 8 - ОТ-ПЦР В МАСШТАБЕ РЕАЛЬНОГО ВРЕМЕНИ И ВЫДЕЛЕНИЕ ВИРУСА
Ткани легкого от 7 собак анализируют в рамках количественных тестов ОТ-ПЦР в масштабе реального времени, которые позволяют выявить ген M вируса гриппа типа A и ген H3 вируса собачьего гриппа А подтипа H3N8. Образцы легкого из всех 7 собак дали положительный результат и на ген М вируса гриппа типа А, и на ген H3 вируса собачьего гриппа (Таблица 8). После 3 пассажей в клетках MDCK вирус гриппа А подтипа H3N8 был выделен от собак из приюта, которые умерли через 3 дня после появления пневмонии. Указанному вирусу было дано название A/canine/Jacksonville/05 (H3N8) (canine/Jax/05). После 2 пассажей в куриных яйцах с эмбрионами вирус гриппа A подтипа H3N8 был выделен из легких домашних собак, которые также умерли через 3 дня после появления пневмонии. Указанному вирусу было дано название A/canine/Miami//05 (H3N8) (canine/Miami/05).
ПРИМЕР 9 - ГЕНЕТИЧЕСКИЙ АНАЛИЗ ИЗОЛЯТОВ СОБАЧЬЕГО ВИРУСА ГРИППА А ПОДТИПА H3N8
Анализ на уровне последовательности собачьих вирусов canine/Jax/05 и canine/Miami/05 показал, что их гены гемагглютинина (HA) на 98% идентичны таковым в изолятах собачьих вирусов canine/FL/04, canine/TX/04 и canine/Iowa/05, выделенных из легких борзых, которые умерли от пневмонии во время вспышек заболевания на треках в 2004 и 2005 гг. (Crawford, P.C. et al., 2005; Yoon K-Y. et al., 2005). Кроме того, HA гены собачьих вирусов canine/Jax/05 и canine/Miami/05 были на 98% идентичны таковым в современных вариантах лошадиных вирусов, выделенных после 2000 г. Сравнительный филогенетический анализ HA генов показал, что собачьи вирусы canine/Jax/05 и canine/Miami/05 образуют кластерную группу с изолятами собачьих вирусов canine/FL/04, canine/TX/04 и canine/Iowa/05 из борзых собак и с изолятами современных форм лошадиных вирусов, так что они формируют отдельную группу, отличную от более ранних лошадиных вирусов, выделенных в начале 1990-х гг. (фиг.4). Кроме того, изоляты собачьих вирусов canine/Jax/05, canine/Miami/05 и canine/Iowa/05 ближе стоят к canine/Tx/04, чем к canine/FL/04 или canine/FL/03. Изоляты 2005 г. формируют подгруппу, которая, по всей видимости, является ответвлением более ранних собачьих вирусов 2003 и 2004 гг., при этом различия наблюдаются примерно в 10 сайтах, отобранных по принципу экономии информации. Наличие этих различий поддерживают гипотезу, согласно которой вирус собачьего гриппа был перенесен горизонтально от собаки к собаке, а не встраивался неоднократно с некоторой периодичностью из внешнего источника. Накопление мутаций в период с 2003 по 2005 гг. иллюстрирует протекающий процесс адаптации, которому вирус должен был подвергнуться, с тем, чтобы он мог быть перенесен в новый организм-хозяин, как это, по предположениям, случилось с вирусами собачьего гриппа.
ПРИМЕР 10 - АМИНОКИСЛОТНЫЙ АНАЛИЗ ИЗОЛЯТОВ ВИРУСА СОБАЧЬЕГО ГРИППА A ПОДТИПА H3N8
В 6 изолятах вируса собачьего гриппа было показано наличие консервативных замещений аминокислот, которые отличают их от современных вирусов лошадиного гриппа (Таблица 9). Указанные консервативные замещения включают I15M, N83S, W222L, I328T и N483T. Сравнительный филогенетический анализ зрелого белка HA показал, что вирусы собаки canine/Jax/05, canine/Miami/05 и canine/Iowa/05 формируют одну подгруппу вместе с изолятом вируса собаки canine/TX/04 (фиг.4). Идентифицировано 3 аминокислотных замены (L118V, K261N и G479E), которые отличают данную подгруппу от других вирусов собак (Таблица 9). Идентифицировано также 2 аминокислотных замены (F79L и G218E), которые отличают изоляты 2005 г. от их корневого варианта canine/TX/04. Кроме того, изоляты 2005, взятые от собак, отличных от борзых, а именно canine/Jax/05 и canine/Miami/05, отличаются от изолята, выделенного из борзых, canine/Iowa/05, наличие изменения по одной аминокислоте, R492K. И наконец, собачий вирус canine/Jax/05 отличается от другого вируса, выделенного из собак, canine/Miami/05, всего по одной аминокислоте, S107P. Во всех других вирусах H3N8 лошади и собаки, аминокислота S консервативна в положении 107, за исключением A/Equine/Jilin/1/89, который в этом положении содержит T (Guo Y. et al., 1992).
ПРИМЕР 11 - АНТИГЕННЫЙ АНАЛИЗ ИЗОЛЯТОВ ВИРУСА СОБАЧЬЕГО ГРИППА A ПОДТИПА H3N8
Проводят тесты на ингибирование гемагглютинации (HI) с использованием панели антигенов из более ранних и современных вирусов лошадиного гриппа и вирусов собачьего гриппа, а также на фоне сыворотки, отобранной в 2005 г. из лошадей и собак, которые были инфицированы вирусом гриппа (Таблица 10). В процедуру анализа была также включена сыворотка, взятая у африканских хорьков, иммунизированных против canine/FL/04. Титры HI антител, выявленные в сыворотке лошадей, были в 8-16 раз выше, в случае их тестирования с использованием современных лошадиных вирусов, чем с другими, более ранними изолятами, но снижались по меньшей мере в 4 раза, в случае их тестирования с использованием вирусов собаки. Сыворотка из собак не давала реакции с более ранними лошадиными вирусами, тогда как титр антител повышался в 4 раза, при проведении тестирования с использованием изолятов современных лошадиных вирусов и изолятов вирусов собаки. Указанная картина наблюдалась также для сыворотки, взятой из африканских хорьков, иммунизированных против вируса собачьего гриппа. Такого рода серологическая реакция указывает на антигенное сходство между вирусами гриппа собак и современными лошадиными вирусами, что также согласуется с результатами филогенетического анализа. Титры антител, выявленные в сыворотках лошади, собаки и африканского хорька, против изолята canine/Miami/05, были аналогичны таковым, найденным для изолятов вирусов собаки в 2003 и 2004 гг. Однако в данном случае титры были в 2-4 раза ниже применительно к изоляту canine/Jax/05. Эти результаты позволяют полагать, что canine/Jax/05 отличается по антигенному составу от других изолятов собачьего вируса, что, хотя бы частично, может определяться аминокислотной заменой в положении 107 в зрелом HA.
b Буквы верхнего регистра обозначают остатки зафиксированной нуклеиновой кислоты.
3-й пассаж
2-й пассаж
Сравнение по аминокислотному составу зрелого HA из вирусов собачьего гриппа и современных вирусов лошадиного гриппа
Титры антител в сыворотке лошади, собаки и африканского хорька, выявленные против более ранних и современных вирусов лошадиного гриппа и против вирусов собачьего гриппа
b Сыворотка из африканских хорьков, иммунизированных вирусом canine/FL/04.
МАТЕРИАЛЫ И РЕПРЕЗЕНТАТИВНЫЕ МЕТОДЫ, ОТНОСЯЩИЕСЯ К ПРИМЕРАМ 12-15
Инокулят вируса собачьего гриппа
Вирусный инокулят получают путем инокуляции в эпителиальные клетки почки собаки Madin-Darby (MDCK) основной формы вируса A/canine/FL/43/04 (H3N8), представляющей собой 3 пассаж ранее описанного исходного изолята (Crawford et al., 2005). Инокулированные MDCK клетки в минимальной основной среде в модификации Дульбеккоо (DMEM), содержащей в качестве добавок 1 мкг/мл TPCK в трипсиновой обработке (Sigma-Aldrich Corp., St. Louis, MO) и антибиотики (гентамицин и ципрофлоксацин), растят в колбах на 250 см2 при температуре 37°C в увлажненной атмосфере, содержащей 5% CO2. Культуру осматривают ежедневно на выявление возможных морфологических изменений и собирают через 5 дней после инокуляции. Собранную культуру осветляют центрифугированием, и полученные супернатанты хранят при температуре -80°C до проведения инокуляции собак. Отбирают аликвоту супернатанта для определения титра вируса по методу Рида и Мюнша (Reed and Muench). Указанный титр составляет 107 средней цитопатогенной дозы для культуры (TCID50) вируса A/canine/Florida/43/04 (canine/FL/04) на мл.
Экспериментальная инокуляция
Используют восемь 4-месячных помесных собак из колонии (Marshall BioResources, North Rose, NY) (4 самца и 4 самки) для проведения исследования по экспериментальной инокуляции, разрешенного Комитетом по надзору за использованием животных в эксперименте и уходу за ними при Университете во Флориде (University of Florida Institutional Animal Care and Use Committee). В эксперимент были включены собаки с весом тела от 13 до 17 кг. Указанные собаки были признаны здоровыми, по результатам физического осмотра, данных анализа крови на базовом уровне и температурных показателей, зарегистрированных в течение 2 недель до инокуляции. Ни одна из собак до этого не подвергалась воздействию вируса собачьего гриппа, как следует из результатов серологических анализов, проведенных на спаренных образцах сывороток, отобранных в момент поступления животных в данное учреждение и через 2 недели. Собак повергают анестезии путем внутривенной инъекции пропофола (Diprivan®, Zeneca Pharmaceuticals, 0,4 мг/кг веса тела, для достижения желательного эффекта) с последующей интубацией эндотрахеальными трубками. Каждой из шести собак (3 самца и 3 самки) инокулируют 107 TCID50 вируса canine/FL/04 в 5 мл стерильного солевого раствора, вводимого в дистальную часть трахеи с помощью каучукового катетера небольшого диаметра, встроенного в эндотрахеальную трубку. Двум собакам (1 самец и 1 самка) проводят ложную инокуляцию равного объема стерильного раствора. Ложно инокулированных контрольных собак помещают в комнатах, отдельно от собак, которым был инокулирован вирус, и за ними ухаживают разные люди из числа персонала. Физический осмотр и регистрацию ректальной температуры проводят два раза в день в течение 6 дней после инокуляции (p.i.).
Отбор фарингеальных и ректальных мазков
Для отслеживания выделения вируса во внешнюю среду от каждой собаки дважды в день отбирают орофарингеальные образцы в дни от 0 до 6 после инокуляции, с использованием полиэфирных тампонов (Fisher Scientific International Inc., Pittsburgh, PA). Тампоны помещают в 1 мл стерильного фосфатно-буферного раствора (PBS), содержащего 0,5% бычьего сывороточного альбумина (БСА). Ректальные образцы берут у каждой собаки ежедневно в дни от 0 до 6. Экстракты тампонов получают путем осветления центрифугированием сред, использованных для транспортировки. Аликвоту экстракта тампона анализируют, сразу же отбирают для анализа на наличие нуклеопротеина вируса гриппа A с использованием коммерческого набора для иммуноанализа Directigen™ (BD, Franklin Lakes, NJ), в соответствии с инструкциями производителя. Оставшийся экстракт хранят при температуре -80°C до проведения вирусологических исследований.
Посмертный осмотр
В 1 день после инокуляции одну ложно инокулированную собаку и одну собаку с инокулированным вирусом подвергают эвтаназии путем внутривенной инокуляции раствора Beuthanasia-D® (1 мл/5 кг веса тела; Schering-Plough Animal Health Corp). Аналогично, по одной собаке с инокулированным вирусом подвергают эвтаназии каждый день в период от 2 до 5 дня после эвтаназии. На 6 день после эвтаназии подвергают эвтаназии оставшихся ложно инокулированных собак и собак с инокулированным вирусом. Полный посмертный осмотр проводит один из исследователей (WLC). Ткани фиксируют в 10% нейтральном забуференном формалине, погруженном в парафин, и срезы по 5 мкм либо окрашивают гематоксилином и эозином, либо подвергают обработке для иммуногистохимического анализа по описанной ниже процедуре. Нефиксированные ткани легкого отправляют в Диагностический Центр клинической микробиологии/паразитологии/серологии Колледжа Ветеринарной Медицины при Университете Флориды (Diagnostic Clinical Microbiology/Parasitology/ Serology Service at the University of Florida College of Veterinary Medicine) для выделения и идентификации бактерий. Образцы культивируют на неселективных средах, а также на средах, селективных для видов Bordetella (Regan-Lowe; Remel, Lenexa, KS) и видов Mycoplasma (Remel). Все культуры выдерживают в течение 21 для того, чтобы сделать вывод об отсутствии роста. Нефиксированные ткани также хранят при температуре -80°C до проведения вирусологических исследований.
Иммуногистохимия
Депарафинизированные и регидратированные срезы тканей трахеи и легкого толщиной по 5 мкм помещают на предметные стекла Bond-Rite™ (Richard-Allan Scientific, Kalamazoo, MI), после чего обрабатывают протеиназой K (DAKOCytomation Inc., Carpenteria, CA) и затем реагентом, блокирующим пероксидазу (DAKO® EnVision™ Peroxidase Kit, DAKO Corp., Carpenteria, CA). Срезы инкубируют с моноклональным антителом против вируса гриппа А Н3, в разведении 1:500 (Chemicon International, Inc., Ternecula, CA), в течение 2 часов при комнатной температуре. В качестве контрольных вариантов проводят инкубацию тех же срезов с мышиным IgG (1 мг/мл, Serotec, Inc. Raleigh, NC), а также инкубацию моноклонального антитела со срезами из ткани нормального легкого собаки. После обработки первичным антителом срезы инкубируют со вторичными реагентами субстратов иммунопероксидазы и пероксидазы (Dako® EnVision™ Peroxidase Kit, Dako Corp.), в соответствии с инструкциями производителя. Далее срезы подвергают контрастному окрашиванию гематоксилином, обрабатывают очищающим реагентом #2 и подсинивающим реагентом (Richard-Allan Scientific, Kalamazoo, MI), дегидратируют и закрывают покровными стеклами Permount (ProSciTech, Queensland, Australia).
Экстракция РНК из тампонов и тканей
Ткани трахеи и легкого от каждой из исследуемых собак оттаивают и гомогенизируют в минимальной основной среде (MEM), содержащей, в качестве добавок, 0,5% бычий сывороточный альбумин (БСА) и антибиотики (гентамицин и ципрофлоксацин) с использованием одноразового измельчителя тканей (Kendall, Lifeline Medical Inc., Danbury, CT). Суммарную РНК экстрагируют из гомогенатов тканей, а также из экстрактов, полученных из орофарингеальных и ректальных тампонов, с использованием коммерческого набора (RNeasy® Mini Kit, QIAGEN Inc., Valencia, CA), в соответствии с инструкциями производителя, и элюируют в конечном объеме 60 мкл буфера.
ОТ-ПЦР в масштабе реального времени
На суммарной РНК проводят количественную одностадийную ОТ-ПЦР в масштабе реального времени с использованием набора зондов для ПЦР (QuantiTect® Probe RT-PCR Kit), содержащего ROX, в качестве эталонного пассивного красителя (QIAGEN Inc., Valencia, CA), и набор праймерных зондов, направленных на высококонсервативный участок матричного гена (M) вируса гриппа типа A (Payungporn S. et al., 2006a; Payungporn S. et al., 2006b). Для проведения каждой из реакций ОТ-ПЦР в масштабе реального времени добавляют по 5 мкл экстрагированной суммарной РНК к реакционной смеси, содержащей 12,5 мкл основной смеси зондов для ОТ-ПЦР (2X QuantiTech® Probe RT-PCR Master Mix), 0,25 мкл смеси QuantiTech® RT Mix, прямых и обратных праймеров (в конечной концентрации каждого из них 0,4 мкМ), зонд (в конечной концентрации 0,1 мкМ) и воду, не содержащую РНКазу, до общего конечного объема 25 мкл. Используют контрольные реагенты TaqMan® GAPDH (Applied Biosystems, Foster City, CA), в соответствии с инструкциями производителя, для детекции GAPDH, в качестве эндогенного внутреннего контроля для оценки РНК, экстрагированной из тампона и образцов ткани, и в качестве контрольного варианта для нормализации результатов.
Количественную одностадийную ОТ-ПЦР в масштабе реального времени проводят на реакционных смесях, введенных в систему Mx3000P® QPCR System (Stratagene, La Jolla, CA). Условия циклической обработки включают стадию обратной транскрипции при температуре 50°C в течение 30 минут, стадию начальной денатурации при температуре 95°C в течение 15 минут, для целей активации ДНК-полимеразы HotStarTaq®, и стадию амплификации, осуществляемую в ходе 40 циклов. Каждый цикл амплификации включает стадию денатурации при температуре 94°C в течение 15 секунд, с последующим проведением стадии отжига/наращивания при температуре 60°C в течение 1 минуты. В конце каждого цикла регистрируют флуоресцентные сигналы FAM (длина волны эмиссии при 518 нм) и VIC (длина волны эмиссии при 554 нм). Пороговый показатель цикла (Ct) определяют при установке пороговой флуоресценции (dR) на 1000 в каждом отдельном эксперименте. Для регистрации и анализа данных используют пакет прикладных программ Mx3000P® версия 2.0 (Stratagene, La Jolla, CA). Позитивный контроль включает проведение амплификации РНК, экстрагированной из вируса A/canine/FL/242/03 (H3N8). Полученные результаты нормализуют путем деления значения Ct для М на соответствующее значение Ct для GAPDH, полученное для каждого образца.
Повторное выделение вируса из тканей
Замороженные ткани, взятые от собак с инокулированным вирусом, оттаивают и гомогенизируют в 10 объемах среды DMEM, содержащей, в качестве добавок, 0,5% БСА и антибиотики. Твердые остатки удаляют центрифугированием, а полученные супернатанты используют для инокуляции в клетки MDCK, культивируемые в среде DMEM, содержащей, в качестве добавок, 1 мкг/мл TPCK после трипсиновой обработки (Sigma-Aldrich Corp., St. Louis, MO) и антибиотики, описанные выше. Клетки растят в колбах на 25 см2 при температуре 37°C в увлажненной атмосфере, содержащей 5% CO2. Культуры осматривают ежедневно для выявления возможных морфологических изменений и собирают на 5 день после инокуляции. Собранные культуры осветляют центрифугированием, и полученные супернатанты используют для инокуляции в клетки MDCK, как было описано выше применительно к исходной инокуляции; проводят два дополнительных пассажа для образцов, которые не продемонстрировали наличия вируса гриппа, по данным теста на гемагглютинацию или по данным анализа в рамках ОТ-ПЦР. Гемагглютинирующую активность в осветленных супернатантах определяют с использованием 0,5% эритроцитов индейки, по описанной ранее методике (Crawford et al., 2005). ОТ-ПЦР проводят в соответствии с описанной ниже процедурой.
ОТ-ПЦР, нуклеотидное секвенирование и филогенетический анализ
Вирусную РНК экстрагируют из супернатанта культуры клеток MDCK с использованием мининабора для выделения вирусной РНК QIAamp® Viral RNA Mini Kit (QIAGEN Inc., Valencia, CA), в соответствии с инструкциями производителя. Вирусную РНК подвергают обратной транскрипции до кДНК с использованием набора для проведения одностадийной реакции ОТ-ПЦР QIAGEN® OneStep RT-PCR Kit (QIAGEN, Valencia, CA), в соответствии с инструкциями производителя. Амплификацию по методу ПЦР кодирующей области 8 генов из вируса гриппа проводят по описанной ранее методике (Crawford et al., 2005), с использованием универсальных наборов ген-специфических праймеров (праймерные последовательности доступны по запросу). Полученные ампликоны ДНК используют в качестве матриц для автоматизированного секвенирования на автоматическом секвенаторе ДНК ABI PRISM® 3100 по процедуре циклического секвенирования, включающего использование красителя в конце цикла (Applied Biosystems, Foster City, CA). Нуклеотидные последовательности анализируют с использованием пакета прикладных программ Lasergene 6 Package® (DNASTAR, Inc., Madison, WI). Нуклеотидные последовательности вирусов, выделенных из инфицированных собак, сравнивают с последовательностями вируса в используемом инокуляте, с целью определения возможных изменений, произошедших в ходе репликации в дыхательных путях.
ПРИМЕР 12 - КЛИНИЧЕСКАЯ КАРТИНА ЗАБОЛЕВАНИЯ
У всех 6 собак с инокулированным вирусом развилась временная лихорадка (с ректальной температурой ≥39°C) в течение первых 2 дней после инокуляции, однако, в ходе 6-дневного периода наблюдения не было отмечено респираторных симптомов, таких как кашель или выделения из носа. При этом ложно инокулированные собаки оставались клинически здоровыми.
ПРИМЕР 13 - ВЫДЕЛЕНИЕ ВИРУСА В ОКРУЖАЮЩУЮ СРЕДУ
Нуклеопротеин вируса гриппа A был выявлен в орофарингеальном мазке, взятом от одной собаки с инокулированным вирусом через 24 часа после инокуляции. Тогда как по данным количественной ОТ-ПЦР в масштабе реального времени орофарингеальные мазки, взятые от одной собаки через 72, 84 и 120 часов после инокуляции, и у другой собаки через 108, 120 и 132 часа после инокуляции, дали положительные результаты на наличие вируса (Таблица 11). Абсолютное число копий гена M на мкл экстракта из тампона возрастает со временем, в период от 3 до 6 дней после инокуляции. Тогда как в ректальных мазках вирус выделен не был.
ПРИМЕР 14 - ПОСМЕРТНЫЙ ОСМОТР
В отличие от предшествующих исследований с экспериментальным инфицированием собак бигль, не содержавших патогена (Crawford et al., 2005), у помесных собак с инокулированным вирусом развилась пневмония, как следовало по данным общего осмотра животных и по результатам гистологического анализа легких, проведенного в период от 1 до 6 дня после инокуляции. Кроме пневмонии, указанные собаки имели ринит, трахеит, бронхит и бронхиолит, аналогично тому, что было ранее зарегистрировано для собак, инфицированных естественным образом (Crawford et al., 2005). Отмечаются некроз и эрозия эпителиальной выстилки дыхательных путей и бронхиальных желез, с сопутствующей инфильтрацией нейтрофилов и макрофагов в подслизистые ткани (фиг.5, верхние панели). При проведении иммуногистохимического анализа был выявлен вирусный антиген H3 в эпителиальных клетках бронхов, бронхиол и бронхиальных желез (фиг.5, нижние панели). Не обнаружено бактериальной суперинфекции. Ткани дыхательных путей, взятые от 2 ложно инокулированных собак были нормальными.
ПРИМЕР 15 - РЕПЛИКАЦИЯ ВИРУСА В ТРАХЕЯХ И ЛЕГКИХ
По данным количественной ОТ-ПЦР в масштабе реального времени трахеи и легкие были положительными на наличие вируса у всех собак в период от 1 до 6 дня после инокуляции (Таблица 12). Абсолютное число копий гена М вируса гриппа на мкл гомогената трахей возрастает в период от 1 до 5 дня после инокуляции, и затем снижается к 6 дню. Абсолютное число копий гена М вируса гриппа на мкл гомогената легких снижается в период от 1 до 6 дня после инокуляции. В основном, трахеи содержат ≥ одного log10 больше вируса, чем легкие, на 6 день после инокуляции.
Детекция выделения вируса в орофаринксе помесных собак с инокулированным вирусом собачьего гриппа, по данным количественной ОТ-ПЦР в масштабе реального времени
b Коэффициенты, используемые для нормализации результатов, получаемых при делении показателя Ct для M на Ct для GAPDH для каждого экстракта из тампона.
c Абсолютное число копий матричного гена на мкл экстракта тампона.
Выявление репликации вируса в трахеях и легких помесных собак с инокулированным вирусом собачьего гриппа, по данным количественной ОТ-ПЦР в масштабе реального времени
b Коэффициенты, используемые для нормализации результатов, получаемых при делении показателя Ct для M на Ct для GAPDH для каждого гомогената ткани.
c Абсолютное число копий матричного гена на мкл гомогената ткани.
МАТЕРИАЛЫ И РЕПРЕЗЕНТАТИВНЫЕ МЕТОДЫ ДЛЯ ПРИМЕРА 16
Штаммы вирусов
Штаммы вируса гриппа собаки, а также штаммы соответствующих вирусов из птиц, лошади и человека (перечисленные в Таблице 15), размножают в эмбрионах куриных яиц или в клетках MDCK, и определяют их инфективность путем титрования серийных разведений, с определением конечной точки, в куриных эмбрионах или в рамках теста на бляшкообразование. Быстрым методом количественной оценки вируса является тест на гемагглютинацию, проводимый с использованием эритроцитов индейки.
Образцы, используемые для диагностики
В ходе вспышки 2005 г. отбирают в общей совокупности 60 тканей легкого у собак, в отношении которых имелись основания, подозревать развитие вирусного респираторного заболевания, которые исследуют на наличие у них вируса собачьего гриппа.
Экстракция РНК из образцов тканей собак
Блоки ткани легкого весом от 20 до 30 мг гомогенизируют с использованием одноразового измельчителя тканей (Kendal). Суммарную РНК экстрагируют с использованием коммерческого набора (RNeasy Mini Kit, Qiagen, Valencia, CA), и далее проводят элюцию в конечном объеме 60 мкл, в соответствии с рекомендациями производителя.
Разработанные для анализа праймеры и зонды
Проводят множественные сопоставления последовательностей для H3 и M генов из множества подтипов и различных видов с использованием программы CLUSTAL X (версия 1.8). Выбирают матричные (M) праймеры и зонды из консервативных участков известных последовательностей, соответствующих различных подтипам вируса гриппа A, тогда как ген-специфичные праймеры для гемагглютинина H3 и набор зондов выбирают таким образом, чтобы можно было отслеживать специфическое спаривание генов из вирусов лошадиного и собачьего гриппа A и ошибочное спаривание гомологичных птичьих и человеческих генов (Таблица 13). Был использован соответствующий пакет прикладных программ, используемый для разработки праймеров (OLIGOS версия 9.1), а также информация веб-сайта, построенного на результатах анализа с использованием инструментов, доступных в рамках программы EXIQON (http://lnatools.com), для расчета показателя Tm и прогностических оценок вторичной структуры и возможности самогибридизации. Был использован консервативный участок гена 18S рРНК, в качестве внутреннего эндогенного контроля для оценки наличия РНК, экстрагированной из образца ткани собаки. Были использованы реагенты TaqMan® для проведения теста на наличие эукариотической 18S рРНК (VIC/TAMRA) (Applied Biosystems) для детекции в реальном времени 18S рРНК в образцах ткани.
Условия проведения ОТ-ПЦР в масштабе реального времени
Проводят одностадийную ОТ-ПЦР в масштабе реального времени с использованием набора для ОТ-ПЦР Quantitect Probe RT-PCR, содержащего ROX, в качестве пассивного эталонного красителя (Qiagen, Valencia, CA). Для каждой из ОТ-ПЦР, проводимой в масштабе реального времени, используют 5 мкл образца РНК, в качестве матрицы, в составе реальной смеси, содержащей 10 мкл основной смеси зондов для ОТ-ПЦР (2X QuantiTech Probe RT-PCR Master Mix), 0,2 мкл смеси QuantiTech RT Mix, праймеры (в конечной концентрации 0,4 мкМ, для H3 гена, или в конечной концентрации 0,6 мкМ, для M гена), зонд (в конечной концентрации 0,1 мкМ, для H3 гена, или в конечной концентрации 0,2 мкМ, для M гена) и воду, не содержащую РНКазу, до общего конечного объема 20 мкл. Одностадийную ОТ-ПЦР в масштабе реального времени проводят в системе Mx3005P Real-Time QPCR System (Stratagene). Условия циклической обработки включают стадию обратной транскрипции при температуре 50°C в течение 30 минут. После проведения стадии начальной денатурации при температуре 95°C в течение 15 минут, с целью активации ДНК-полимеразы HotStarTaq, проводят амплификацию в ходе 40 циклов, включающих денатурацию (94°C в течение 15 секунд) и отжиг/наращивание (60°C в течение 30 секунд). Флуоресцентные сигналы FAM (длина волны эмиссии при 516 нм, для детекции H3 и M) и VIC (длина волны эмиссии при 555 нм, для детекции 18S рРНК) получают один раз в конце стадии расширения. Сбор и анализ данных, полученных в ходе ПЦР теста в масштабе реального времени, проводят с использованием программного обеспечения Mx3005P, версия 2.02 (Stratagene).
Специфичность H3 праймеров/набора зондов для вируса собачьего гриппа (H3N8) и универсальность M праймеров/набора зондов для вируса собачьего гриппа типа A
Для оценки специфичности каждого из праймеров/набора зондов используют РНК, экстрагированную из нескольких известных подтипов вирусов гриппа A, в качестве матрицы в тесте на основе ОТ-ПЦР в масштабе реального времени (Таблица 15).
Стандартная РНК, используемая для проведения ОТ-ПЦР в масштабе реального времени
Используют гены вируса собачьего гриппа A (A/canine/Florida/242/2003(H3N8)) с целью создания ПЦР ампликонов для H3 (нт 1-487) и M (нт 1-276) с использованием праймеров, соединенных с T7 промотором (Таблица 13). Затем очищенные ПЦР ампликоны H3 и M генов используют в качестве матриц для проведения транскрипции in vitro с использованием системы транскрипции Riboprobe in vitro Transcription System-T7 (Promega), в соответствии с рекомендациями производителя. Концентрации транскрибированных РНК рассчитывают при измерении поглощения при длине волны 260 нм. Далее делают 10-кратные серийные разведения РНК, в диапазоне от 108 до 10 копий/мкл, для проведения тестов на чувствительность. Кроме того, строят стандартную кривую зависимости log исходных концентраций матричных РНК (копии/мкл) от порогового значения цикла (Ct), полученного для каждого разведения, с тем, чтобы определить общие характеристики ОТ-ПЦР в масштабе реального времени.
Сравнительные тесты на чувствительность между ОТ-ПЦР в масштабе реального времени и набором для анализа Directigen Flu A
Используют основные вирусные препараты двух вирусных штаммов, включающих A/Wyoming/3/2003 (H3N2) с показателем 106,67 EID50/мл (HA=64) и A/canine/Florida/242/2003(H3N8) с показателем 107,17 EID50/мл (HA=16), в методе определения порогового показателя. Используют логарифмические разведения образцов в фосфатно-буферном растворе (PBS) (125 мкл), включенные в набор для быстрой детекции антигена вируса гриппа A, Directigen Flu A (Becton, Dickinson and Company), в соответствии с инструкциями производителя. Устройство для каждого из анализов в рамках теста с использованием набора Directigen Flu A содержит в центре мембраны пятно нанесенного антигена из вируса гриппа H1N1, которое приобретает вид точки фиолетового цвета, указывая на пригодность системы тестирования, основанной на моноклональном антителе против нуклеопротеина (NP). Образование фиолетового треугольника вокруг точки указывает на наличие в исследуемом образце NP вируса гриппа. Интенсивность фиолетового сигнала, регистрируемая от треугольника, ранжируется в следующем порядке: + (очертание треугольника), ++ (слегка окрашенный треугольник), +++ (темно-фиолетовый треугольник) и ++++ (очень темно-фиолетовый треугольник). Вирусную РНК экстрагируют аликвотами по 125 мкл каждого из разведений вируса с использованием мининабора для вирусной РНК QIAamp Viral RNA Mini Kit (Qiagen, Valencia, CA) и далее проводят элюцию в конечном объеме 50 мкл. Экстрагированные вирусные РНК в объеме 5 мкл анализируют в рамках ОТ-ПЦР в масштабе реального времени, для проведения сравнительной оценки относительно метода тестирования, включающего использование набора Directigen Flu A.
ПРИМЕР 16
Тест в рамках ОТ-ПЦР в масштабе реального времени на наличие вируса собачьего гриппа основан на информации, доставляемой тремя молекулярными зондами, которые направляют 18S рРНК из клетки-хозяина, а также M и H3 гены из генома вируса гриппа A (Таблица 14). Амплификация гена-хозяина является показателем качества и целостности образца. Ожидалось, что клинические образцы, равно как и образцы некропсии или лабораторные образцы, содержащие вирус собачьего гриппа (H3N8), продемонстрируют сигнал амплификации с тремя зондами. Образцы, демонстрирующие сигнал амплификации с зондами для M и 18S рРНК, но дающие отрицательную реакцию для H3, могут рассматриваться как индикаторные на наличие вируса гриппа, подтипа H3, взятого от человека, свиньи или птицы, или подтипов, отличных от H3. Такие случаи могут быть далее прояснены в рамках ОТ-ПЦР с использованием универсальных праймеров HA, способных создавать ампликон кДНК, который может быть далее проанализирован секвенированием. Отобранные и обработанные соответствующим образом образцы, которые не содержат вируса гриппа A, демонстрируют только сигнал амплификации для 18S рРНК. Ситуации, в которых только зонд для 18S рРНК и H3 зонды генерируют сигнал амплификации, являются индикаторами ошибочности в проведении методики, если не доказано обратное; или в этом случае следует подтвердить ложно-негативный результат для M зондов или ложно-позитивный результат для H3. И, наконец, образцы, которые не демонстрируют сигналы амплификации с тремя зондами, являются показателями дефектов в процессе отбора образца, его деградации, некорректности в процессе экстракции РНК или наличия ингибиторов полимераз, используемых в ПЦР.
Для оценки специфичности H3 праймеров/набора зондов для вируса собачьего гриппа A (H3N8) и универсальности M праймеров/набора зондов для вируса гриппа типа A, несколько подтипов вирусов гриппа А тестируют по методу ОТ-ПЦР в масштабе реального времени. Полученные результаты показывают, что H3 праймеры/набор зондов дают позитивный сигнал амплификации только с вирусом собачьего гриппа (H3N8). Для других подтипов или для человеческих Н3 штаммов не наблюдалось значимых ложно-позитивных или неспецифических сигналов амплификации. M праймеры/набор зондов генерируют позитивный сигнал амплификации со всеми исследованными штаммами (Таблица 15). Полученные результаты показывают, что H3 праймеры/зонды способны специфически выявлять вирус собачьего гриппа A (H3N8), тогда как M праймеры/зонды выявляют множество подтипов вирусов гриппа типа A.
Результаты тестов в рамках ОТ-ПЦР в масштабе реального времени оценивают по точке конечного разведения транскрибированных in vitro рРНК для M и H3. Как ожидалось, пороговое значение цикла (Ct) возрастает прямо пропорционально разведению стандартов РНК. Флуоресцентные сигналы могут быть выявлены при разведениях стандартов РНК для M и H3 до 103 и 102 копий/мкл соответственно (фиг.6A и 6B). Стандартные кривые для M и H3 генов получают при построении графика зависимости log исходных концентраций РНК от порогового значения цикла (Ct), полученного для каждого разведения (фиг.6C и 6D). Угол наклона стандартной кривой используется в качестве показателя, позволяющего определить эффективность реакции ПЦР, которая теоретически является экспоненциальной; при этом показатель 100% эффективности амплификации будет означать удвоение концентрации ампликона в каждом цикле. Стандартные кривые с углом наклона в диапазоне примерно от -3,1 до -3,6 в типичном случае приемлемы для большинства вариантов применения, требующих точного количественного определения (в плане тщательности выявления) (где эффективность реакции достигает 90-110%). Показатель Rsq представляет собой коэффициент, используемый с целью преставления и анализа всех данных в графической форме. Если все данные укладываются точно на одну линию, то показатель Rsq будет равен 1,00. По мере удаления данных от указанной линии показатель Rsq будет снижаться. Для большинства вариантов тестирования приемлем показатель Rsq ≥ 0,985. Стандартная кривая для M имеет угол наклона -3,576 (эффективность = 90,4%) и, в этом случае, показатель Rsq = 1,00, тогда как стандартная кривая для H3 имеет угол наклона -3,423 (эффективность = 95,9%), и для нее показатель Rsq = 0,999. Приведенные значения представляют собой удовлетворительную эффективность амплификации и, в целом, характера тестирования по методу ОТ-ПЦР в масштабе реального времени. Авторы связывают сниженную эффективность и чувствительность M праймеров/набора зондов, относительно варианта H3 праймеров/набора зондов, с N-кратной вырожденностью праймерных последовательностей для M, необходимых для охвата генных последовательностей М, в связи с более расширенным в этом случае диапазоном, определяемым вариабельностью множества подтипов вирусов, организмов-хозяев и клеточных линий.
Чувствительность процедуры тестирования по методу ОТ-ПЦР в масштабе реального времени сравнивают с данным показателем для коммерческого набора, применяемой с целью быстрой детекции антигена (Directigen Flu A). Анализируют логарифмические разведения вирусов A/Wyoming/3/2003 (H3N2) и A/canine/Florida/242/2003(H3N8) с использованием набора Directigen Flu A и по процедуре ОТ-ПЦР в масштабе реального времени. Результаты оценки с использованием набора Directigen Flu A показывают, что чувствительность против обоих вирусных штаммов характеризуется величиной, равной примерно 100-кратному разведению основных препаратов вирусов, использованных в данных экспериментах (фиг.7). Сигналы (фиолетовое окрашивание), создаваемые образцами с вирусом собаки (A/canine/Florida/242/2003: 106,x БОЕ/мл), были намного слабее, чем соответствующие сигналы, выявленные для человеческого вируса (A/Wyoming/3/2003: 107,x БОЕ/мл), что согласуется с более низкой концентрацией вируса в этих образцах. Альтернативно, более низкий сигнал в случае вируса собачьего гриппа может определяться молекулярной специфичностью моноклональных антител против NP; т.е. может иметь отношение к недостаточной консервации аминокислот в NP эпитопа вирусов собачьего гриппа.
Чувствительность тестирования по методу ОТ-ПЦР в масштабе реального времени применительно к гену M приводит к получению показателей Ct, превышающих порог для реакции с вирусными эквивалентами в 10 и 30 БОЕ применительно к A/canine/Florida/242/2003 и A/Wyoming/3/2003 соответственно (Таблица 16). Различия между показателями чувствительности 2 вирусных штаммов определяются разницей в исходных титрах этих вирусов. Сравнительное исследование вирусов собачьего и человеческого гриппа применительно к возможности детекции в них гена H3 не проводилось, поскольку H3 праймеры/зонды в использованном авторами тесте по методу ОТ-ПЦР в масштабе реального времени приводили к амплификации исключительного вируса собачьего гриппа типа А. При этом тест по процедуре ОТ-ПЦР был в 105 более чувствительным, чем набор для быстрой детекции антигена.
Для оценки возможностей теста по методу ОТ-ПЦР при исследовании образцов некропсии, взятых от собак с острым респираторным заболеванием, провели анализ по методу ОТ-ПЦР в масштабе реального времени шестидесяти образцов тканей легкого, отобранных в 2005 г., с целью выявления вируса собачьего гриппа A. В случае 12 из 60 исследованных образцов (20%) проведенный анализ дал позитивный результат для генов M и H3, тогда как оставшиеся 48 образцов дали отрицательный результат и для гена M, и для гена H3. Были предприняты попытки выделения вируса с использованием метода инокуляции в куриные яйца и клетки MDCK, с тем, чтобы оценить специфичность теста в масштабе реального времени; в случае 2 из 12 образцов, которые были позитивными, по данным ОТ-ПЦР, для вируса собачьего гриппа, было продемонстрировано наличие вируса собачьего гриппа (данные не показаны, рукопись находится в печати). Несмотря на то что все ткани были взяты от собак, у которых было ранее зарегистрировано респираторное заболевание, в большинстве образцов не был выявлен вирус собачьего гриппа, ни по процедуре ОТ-ПЦР в масштабе реального времени, ни при проведении стандартного выделения, что может указывать на высокую вероятность наличия других респираторных патогенов, таких как Bordetella bronchiseptica, возбудитель чумы собак или вирус парагриппа. В этой связи одностадийный тест по методу ОТ-ПЦР в масштабе реального времени обеспечивает быструю, чувствительную и недорогую процедуру для выявления вируса собачьего гриппа A (H3N8). А быстрая лабораторная диагностика инфекций, вызванных вирусом собачьего гриппа A (H3N8), на ранних стадиях заболевания может дать релевантную информацию, важную для оценки клинического состояния животного и соответствующих для данного случая эффективных стратегий.
Праймеры и пробы, использование для детекции по методу ОТ-ПЦР и при транскрипции in vitro
Зонд TaqMan
Интерпретация теста по методу ОТ-ПЦР
в масштабе реального времени
Специфичность теста с H3 праймерами/набором зондов (из собаки) и универсальность теста с M праймерами/набором зондов (из собаки) применительно к нескольким подтипам вирусов гриппа А
Сравнение теста по методу ОТ-ПЦР в масштабе реального времени и теста, основанного на использовании набора Directigen Flu A, по их чувствительности на выявление вируса гриппа A
Различия на уровне аминокислот между белками PB2 из H3N8 вирусов гриппа лошади и собаки
Различия на уровне аминокислот между белками PB1 из H3N8 вирусов гриппа лошади и собаки
Различия на уровне аминокислот между белками PA из H3N8 вирусов гриппа лошади и собаки
Различия на уровне аминокислот между белками NP из H3N8 вирусов гриппа лошади и собаки
Различия на уровне аминокислот между белками NA из H3N8 вирусов гриппа лошади и собаки
Различия на уровне аминокислот между белками М1 из H3N8 вирусов гриппа лошади и собаки
Различия на уровне аминокислот между белками NS1 из H3N8 вирусов гриппа лошади и собаки
Следует понимать, что примеры и варианты осуществления настоящего изобретения, приведенные в данном тексте, даны лишь с целью иллюстрации, и возможны различные модификации или изменения, которые могут быть внесены специалистами в данной области, в свете приведенного описания, и которые охватываются принципами и областью настоящей заявки.
СПИСОК ЦИТИРОВАННОЙ ЛИТЕРАТУРЫ
Патент США No. 5,106,739.
Патент США No. 5,034,322.
Патент США No. 6,455,760.
Патент США No. 6,696,623.
Патент США No. 4,683,202.
Патент США No. 4,683,195.
Патент США No. 4,800,159.
Патент США No. 4,965,188.
Патент США No. 5,994,056.
Патент США No. 6,814,934.
Опубликованная заявка США No. 20040078841.
Опубликованная заявка США No. 20040067506.
Опубликованная заявка США No. 20040019934.
Опубликованная заявка США No. 20030177536.
Опубликованная заявка США No. 20030084486.
Опубликованная заявка США No. 20040123349.
Greyhound Daily News, 1/28/99. National Greyhound Association (NGA), Abilene, Kansas. http://www.NGAgreyhounds.com.
Personal communication, Dr. William Duggar, veterinarian at Palm Beach Kennel Club, West Palm Beach, Florida.
Altschul, S. F. et al. (1990) "Basic Local Alignment Search Tool" J. Mol. Biol. 215:402-410.
Altschul, S. F. et al. (1997) "Gapped BLAST and PSI-BLAST: A New Generation of Protein Database Search Programs" Nucl. Acids Res. 25:3389-3402.
An, G. (1987) "Binary Ti vectors for plant transformation and promoter analysis" Methods Enzymol. 153:292-305.
Beltz, G. A., Jacobs, K. A., Eickbush, T. H., Cherbas, P. T., Kafatos, F. C. (1983) "Isolation of multigene families and determination of homologies by filter hybridization methods" Methods of Enzymology, R. Wu, L. Grossman and K. Moldave [eds.] Academic Press, New York 100:266-285.
Burleson, F. et al. (1992) Virology: A Laboratory Manual (Academic Press).
Byars, N.E., A.C. Allison (1987) "Adjuvant formulation for use in vaccines to elicit both cell-mediated and humoral immunity" Vaccine 5:223-228.
Crawford, P.C. et al. (2005) "Transmission of equine influenza virus to dogs" Science 310:482-485.
Chang, C.P. et al. (1976) "Influenza virus isolations from dogs during a human epidemic in Taiwan" Int J Zoonoses 3:61-64.
Dacso, C.C. et al. (1984) "Sporadic occurrence of zoonotic swine influenza virus infections" J Clin Microbiol 20:833-835.
de Boer, H. A., Comstock, L. J., Vasser, M. (1983) "The tac promoter: a functional hybrid derived from the trp and lac promoters" Proc. Natl. Acad. Sci. USA 80(1):21-25.
Felsenstein, J. (1989) Cladistics 5:164.
Fields et al. (1946) Fields Virology, 3rd ed., Lippincott-Raven publishers.
Ford, R.B., Vaden, S.L. (1998) "Canine infectious tracheobronchitis" In Infectious Diseases of the Dog and Cat, 2nd edition, C.E. Greene, editor, W.B. Saunders Co., Philadelphia, PA, pp. 33-38.
Fouchier et al., (2000) Journal of Clinical Microbiology 38 (11):4096-4101.
Good, X. et al. (1994) "Reduced ethylene synthesis by transgenic tomatoes expressing S-adenosylmethionine hydrolase" Plant Molec. Biol. 26:781-790.
Guan, Y. et al. (2004) "H5N1 influenza: a protean pandemic threat" Proc Natl Acad Sci U S A 101:8156-8161.
Guo, Y. et al. (1992) "Characterization of a new avian-like influenza A virus from horses in China" Virology 188:245-255.
Houser, R.E. et al. (1980) "Evidence of prior infection with influenza A/Texas/77 (H3N2) virus in dogs with clinical parainfluenza" Can J Comp Med 44:396-402.
Karasin, A.I. et al. (2000) "Isolation and characterization of H4N6 avian influenza viruses from pigs with pneumonia in Canada" J Virol 74:9322-9327.
Karlin S. and Altschul, S. F. (1990) "Methods for Assessing the Statistical Significance of Molecular Sequence Features by Using General Scoring Schemes" Proc. Natl. Acad. Sci. USA 87:2264-2268.
Karlin S. and Altschul, S. F. (1993) "Applications and Statistics for Multiple High-Scoring Segments in Molecular Sequences" Proc. Natl. Acad. Sci. USA 90:5873-5877.
Kawaoka, Y. et al., (1989) "Avian-to-human transmission of the PB1 gene of influenza A viruses in the 1957 and 1968 pandemics" J Virol 63:4603-4608.
Keawcharoen, J. et al. (2004) "Avian influenza H5N1 in tigers and leopards" Emerg Infect Dis 10:2189-2191.
Kendal, A. P. et al. (1982) In Concepts and Procedures for Laboratory-based Influenza Surveillance . A. P. Kendal, M. S. Pereira, J. J. Skehel, Eds. (U.S. Department of Health and Human Services, Centers for Disease Control and Prevention and Pan-American Health Organization, Atlanta, GA, United States) pp. B17-B35.
Kilbourne, E.D. et al. (1975) "Demonstration of antibodies to both hemagglutinin and neuraminidase antigens of H3N2 influenza A virus in domestic dogs" Intervirology 6:315-318.
Kimura, K. et al. (1998) "Fatal case of swine influenza virus in an immunocompetent host" Mayo Clin Proc 73:243-245.
Klimov, A. I. et al. (1992a) "Sequence changes in the live attenuated, cold-adapted variants of influenza A/Leningrad/134/57 (H2N2) virus" Virology 186:795-797.
Klimov A. et al. (1992b) "Subtype H7 influenza viruses: comparative antigenic and molecular analysis of the HA-, M-, and NS-genes" Arch Virol. 122:143-161.
Kovacova, A. et al. (2002) "Sequence similarities and evolutionary relationships of influenza virus A hemagglutinins" Virus Genes 24:57-63.
Kuiken, T. et al. (2004) "Avian H5N1 influenza in cats" Science 306:241.
Kumar, S. et al. (2004) "MEGA3: Integrated software for Molecular Evolutionary Genetics Analysis and sequence alignment" Brief Bioinform 5:150-163.
Lee, L.G. et al. (1993) "Allelic discrimination by nick-translation PCR with fluorogenic probes" Nucleic Acids Res. 21(16):3761-3766.
Lewin, B. (1985) Genes II, John Wiley & Sons, Inc., p. 96.
Lipatov, A.S. et al. (2004) "Influenza: emergence and control" J Virol 78:8951-8959.
Livak, K. J. et al. (1995) "Oligonucleotides with fluorescent dyes at opposite ends provide a quenched probe system useful for detecting PCR product and nucleic acid hybridization" PCR Methods Appl. 4(6):357-362.
Maniatis, T., E.F. Fritsch, J. Sambrook (1982) "Nuclease Bal31" Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, NY.
Matrosovich, M. et al. (2000) "Early alterations of the receptor-binding properties of H1, H2, and H3 avian influenza virus hemagglutinins after their introduction into mammals" J Virol 74:8502-8512.
Maertzdorf et al., (2004) Clin Microbiol. 42(3):981-986.
Merrifield, R.B. (1963) "Solid Phase Peptide Synthesis. I. The Synthesis of a Tetrapeptide" J. Amer. Chem. Soc. 85:2149-2154.
Nikitin, A. et al. (1972) "Epidemiological studies of A-Hong Kong-68 virus infection in dogs" Bull World Health Organ 47:471-479.
Nobusawa, E. et al. (1991) "Comparison of complete amino acid sequences and receptor-binding properties among 13 serotypes of hemagglutinins of influenza A viruses" Virology 182:475-485.
Patriarca, P.A. et al. (1984) "Lack of significant person-to person spread of swine influenza-like virus following fatal infection in an immunocompromised child" Am J Epidemiol 119:152-158.
Payungporn S. et al. (2006a) "Detection of canine influenza A virus (H3N8) RNA by real-time RT-PCR" (in preparation for Journal of Clinical Microbiology).
Payungporn S, et al. (2006b) "Isolation and characterization of influenza A subtype H3N8 viruses from dogs with respiratory disease in a shelter and veterinary clinic in Florida" (in preparation for Emerging Infectious Diseases).
Peiris, M. et al. (1999) "Human infection with influenza H9N2" Lancet 354:916-917.
Peiris, J.S. et al. (2004) "Re-emergence of fatal human influenza A subtype H5N1 disease" Lancet 363:617-619.
Posnett, D. N. et al. (1988) "A Novel Method for Producing Anti-peptide Antibodies" J. Biol. Chem. 263(4):1719-1725.
Putnam, Bob (February 10, 1999) "Two illnesses seen in death of dogs" St. Petersburg Times.
Reid, A.H. et al. (2004) "Evidence of an absence: the genetic origins of the 1918 pandemic influenza virus" Nat Rev Microbiol 2:909-914.
Rowe, T. et al. (1999) "Detection of antibody to avian influenza A (H5N1) virus in human serum by using a combination of serologic assays" J Clin Microbiol 37: 937-943.
Saiki, R. (1985) "Enzymatic amplification of beta-globin genomic sequences and restriction site analysis for diagnosis of sickle cell anemia" Science 230:1350-1354.
Sambrook, J. et al. (1989) "Plasmid Vectors" In: Molecular Cloning: A Laboratory Manual, 2d Edition, pp. 1.82-1.104. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York.
Subbarao, K. et al. (1998) "Characterization of an avian influenza A (H5N1) virus isolated from a child with a fatal respiratory illness" Science 279:393-396.
Suzuki, Y. et al. (2000) "Sialic acid species as a determinant of the host range of influenza A viruses" J Virol 74:11825-11831.
Tam, J. P. (1988) "Synthetic Peptide Vaccine Design: Synthesis and Properties of a High-Density Multiple Antigenic Peptide System" PNAS USA 85(15):5409-5413.
Top, Jr., F.H. et al. (1977) "Swine influenza A at Fort Dix, New Jersey (January-February 1976). IV. Summary and speculation" J Infect Dis 136 Suppl:S376-S380.
Vines, A. et al. (1998) "The role of influenza A virus hemagglutinin residues 226 and 228 in receptor specificity and host range restriction" J Virol 72:7626-7631.
Wagner, R. et al. (2002) "N-Glycans attached to the stem domain of haemagglutinin efficiently regulate influenza A virus replication" J Gen Virol 83:601-609.
Webby, R. et al. (2004) "Molecular constraints to interspecies transmission of viral pathogens" Nat Med 10:S77-S81.
Webster, R.G. (1998) "Influenza: an emerging disease" Emerg Infect Dis. 4:436-441.
Webster, R.G. et al. (1992) "Evolution and ecology of influenza A viruses" Microbiol Rev. 56 :152-179.
Weis, W. et al. (1988) "Structure of the influenza virus haemagglutinin complexed with its receptor, sialic acid" Nature 333:426-431.
Womble, D.D. (2000) "GCG: The Wisconsin Package of sequence analysis programs" Methods Mol Biol 132:3-22.
Xu, D., McElroy, D., Thornburg, R. W., Wu, R. et al. (1993) "Systemic induction of a potato pin2 promoter by wounding, methyl jasmonate, and abscisic acid in transgenic rice plants" Plant Molecular Biology 22:573-588.
Yang, T. T. et al. (1996) "Optimized Codon Usage and Chromophore Mutations Provide Enhanced Sensitivity with the Green Fluorescent Protein" Nucleic Acid Research 24(22):4592-4593.
Yoon K-Y. et al. (2005) "Influenza virus infection in racing greyhounds" Emerg Infect Dis. 11:1974-1975.
| название | год | авторы | номер документа |
|---|---|---|---|
| МАТЕРИАЛЫ И МЕТОДЫ, ИСПОЛЬЗУЕМЫЕ ДЛЯ ЛЕЧЕНИЯ РЕСПИРАТОРНЫХ ЗАБОЛЕВАНИЙ У СОБАК | 2011 |
|
RU2711791C2 |
| ВИРУС ГРИППА, СПОСОБНЫЙ ИНФИЦИРОВАТЬ СОБАЧЬИХ, И ЕГО ПРИМЕНЕНИЕ | 2006 |
|
RU2520081C2 |
| ВИРУС ГРИППА, СПОСОБНЫЙ ИНФИЦИРОВАТЬ СОБАЧЬИХ, И ЕГО ПРИМЕНЕНИЕ | 2014 |
|
RU2711807C2 |
| МАТЕРИАЛЫ И МЕТОДЫ, ИСПОЛЬЗУЕМЫЕ ДЛЯ ЛЕЧЕНИЯ РЕСПИРАТОРНЫХ ЗАБОЛЕВАНИЙ У СОБАК | 2020 |
|
RU2811752C2 |
| ВИРУС СОБАЧЬЕГО ГРИППА, КОМПОЗИЦИИ, СОДЕРЖАЩИЕ ЭТОТ ВИРУС, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2006 |
|
RU2410427C2 |
| ВИРУС ГРИППА, СПОСОБНЫЙ ИНФИЦИРОВАТЬ СОБАЧЬИХ, И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2802222C2 |
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ВИРУСНОГО ВЕКТОРА APMV-8 | 2010 |
|
RU2592544C2 |
| ВАРИАНТЫ ГЕМАГГЛЮТИНИНА И НЕЙРАМИНИДАЗЫ ГРИППА | 2010 |
|
RU2535970C2 |
| ХИМЕРНЫЕ ГЕНЫ OSPA, БЕЛКИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2011 |
|
RU2583289C2 |
| ПОЛИПЕПТИДЫ С НАПРАВЛЕННЫМ ДЕЙСТВИЕМ | 2006 |
|
RU2393874C2 |



























































































































































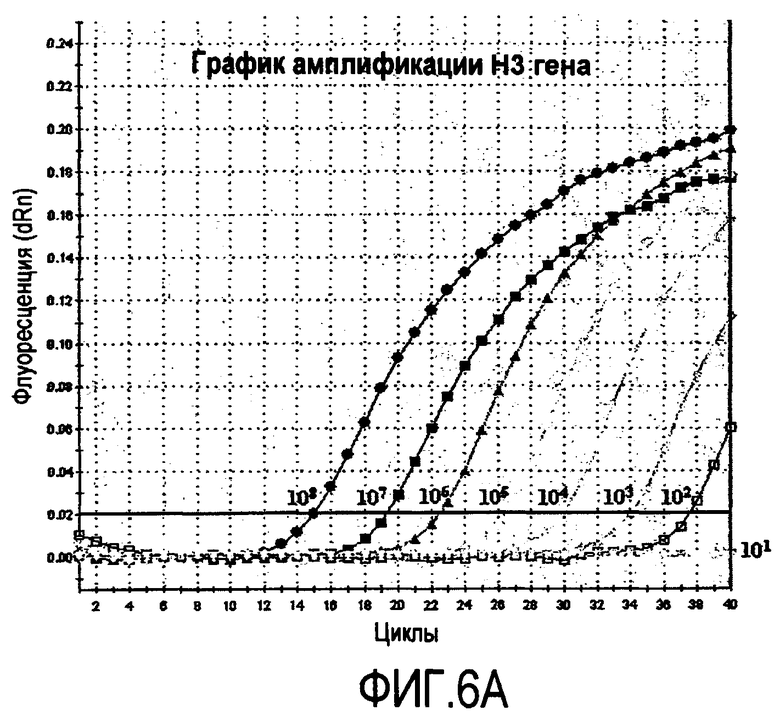
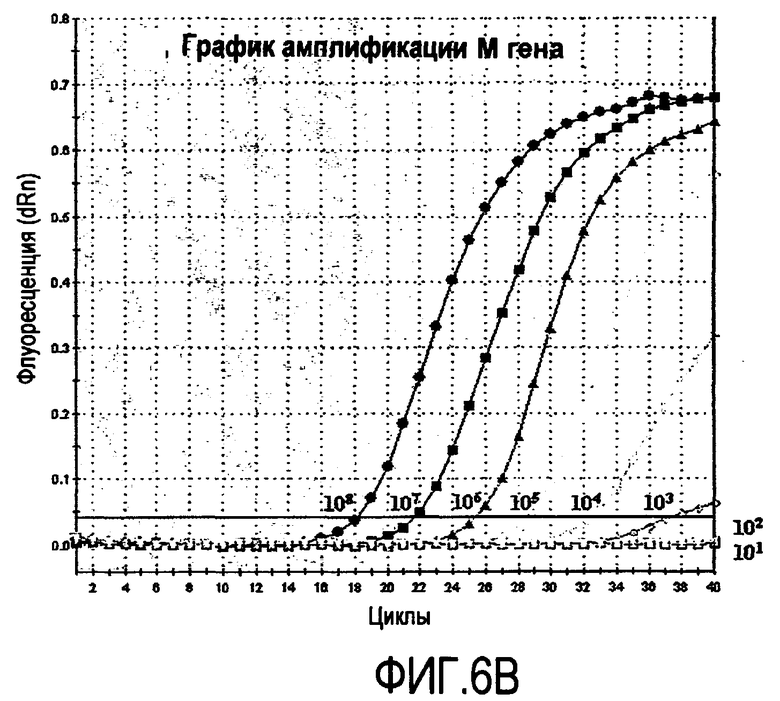
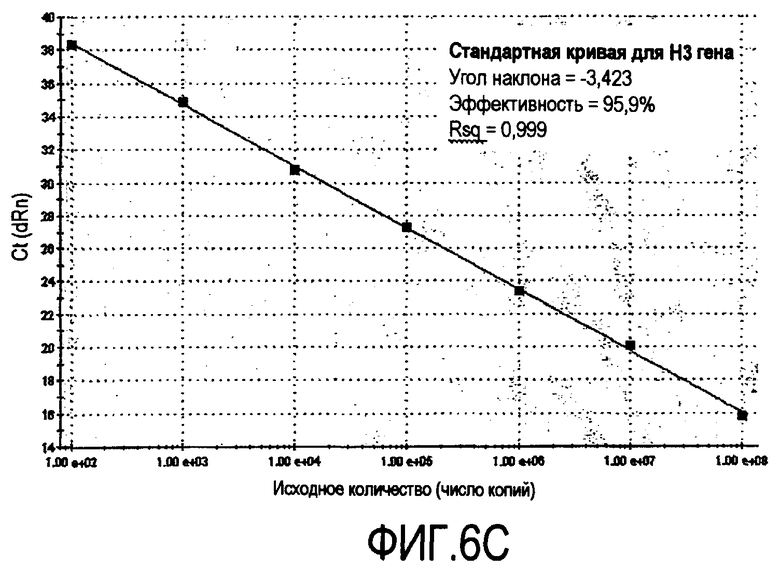
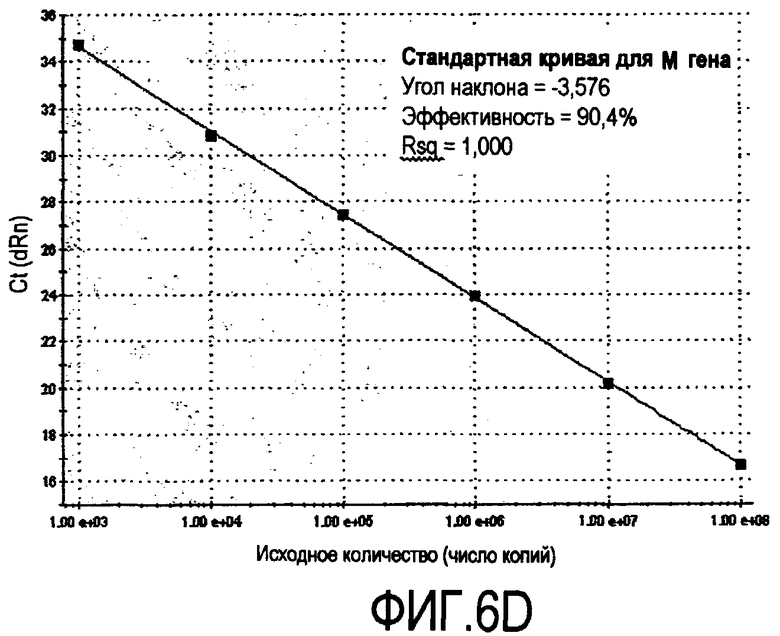
Изобретение относится к области биотехнологии и вирусологии. Раскрыты варианты вируса гриппа, который способен инфицировать животных из группы псовых и вызывать респираторное заболевание у представителей псовых. Также описаны композиции и способы, применяемые для индукции иммунного ответа против вируса гриппа у псовых с использованием новых штаммов, их полинуклеотидов или полипептидов. Изобретение может быть использовано в ветеринарии. 9 н. и 29 з.п. ф-лы, 14 ил., 25 табл., 16 пр.
1. Штамм A/canine/Florida/43/04 вируса гриппа собак А, подтипа H3N8, депонированный в АТТС под номером доступа РТА-7914, являющийся продуцентом полипептидов, индуцирующих иммунный ответ против вируса гриппа, который способен инфицировать животное из группы псовых, при этом геном указанного вируса содержит полинуклеотид, который кодирует полипептид, имеющий аминокислотную последовательность, показанную в виде любой из SEQ ID No:2, 4, 6, 8, 10, 12, 14, 16 или 33, или его функциональный и/или иммуногенный фрагмент, или указанный полинуклеотид кодирует полипептид, характеризующийся 99%-ной или большей идентичностью по последовательности с аминокислотной последовательностью, показанной в виде любой из SEQ ID No:2, 4, 6, 8, 10, 12, 14, 16 или 33.
2. Штамм вируса гриппа по п.1, где указанный полинуклеотид кодирует гемагглютинин (НА), включающий аминокислотную последовательность SEQ ID No:33, или полинуклеотид кодирует полипептид, характеризующийся 99%-ной или большей идентичностью с аминокислотной последовательностью SEQ ID No:33.
3. Штамм вируса гриппа по п.1, где указанный полинуклеотид кодирует гемагглютинин (НА), включающий аминокислотную последовательность SEQ ID No:33, или полинуклеотид кодирует полипептид, характеризующийся 95%-ной или большей идентичностью с аминокислотной последовательностью SEQ ID No:33, где указанный белок НА содержит серин в положении 82, лейцин в положении 221, треонин в положении 327 и треонин в положении 482 указанной последовательности.
4. Штамм вируса гриппа по п.1, где указанный полинуклеотид кодирует гемагглютинин (НА), включающий аминокислотную последовательность SEQ ID No:16, или полинуклеотид кодирует полипептид, характеризующийся 95%-ной или большей идентичностью с аминокислотной последовательностью SEQ ID No:16.
5. Штамм A/canine/Florida/242/2003 вируса гриппа собак А, подтипа H3N8, депонированный в АТТС под номером доступа РТА-7915, являющийся продуцентом полипептидов, индуцирующих иммунный ответ против вируса гриппа, который способен инфицировать животное из группы псовых, при этом геном указанного вируса содержит полинуклеотид, который кодирует полипептид, имеющий аминокислотную последовательность, показанную в виде любой из SEQ ID No: 18, 20, 22, 24, 26, 28, 30, 32 или 34, или его функциональный и/или иммуногенный фрагмент, или указанный полинуклеотид кодирует полипептид, характеризующийся 99%-ной или большей идентичностью по последовательности с аминокислотной последовательностью, показанной в виде любой из SEQ ID No:18, 20, 22, 24, 26, 28, 30, 32 или 34.
6. Штамм canine/Miami/05 вируса гриппа собак А, подтипа H3N8, депонированный в АТТС под номером доступа РТА-7940, являющийся продуцентом полипептидов, индуцирующих иммунный ответ против вируса гриппа, который способен инфицировать животное из группы псовых, при этом геном указанного вируса содержит полинуклеотид, который кодирует полипептид, имеющий аминокислотную последовательность, показанную в виде любой из SEQ ID No:48, 50, 52, 54, 56, 58, 60 или 62, или его функциональный и/или иммуногенный фрагмент, или указанный полинуклеотид кодирует полипептид, характеризующийся 99%-ной или большей идентичностью по последовательности с аминокислотной последовательностью, показанной в виде любой из SEQ ID No:48, 50, 52, 54, 56, 58, 60 или 62.
7. Штамм canine/Jax/05 вируса гриппа собак А, подтипа H3N8, депонированный в АТТС под номером доступа РТА-7941, являющийся продуцентом полипептидов, индуцирующих иммунный ответ против вируса гриппа, который способен инфицировать животное из группы псовых, при этом геном указанного вируса содержит полинуклеотид, который кодирует полипептид, имеющий аминокислотную последовательность, показанную в виде любой из SEQ ID No:64, 66, 68, 70, 72, 74, 76 или 78, или его функциональный и/или иммуногенный фрагмент, или указанный полинуклеотид кодирует полипептид, характеризующийся 99%-ной или большей идентичностью по последовательности с аминокислотной последовательностью, показанной в виде любой из SEQ ID No:64, 66, 68, 70, 72, 74, 76 или 78.
8. Выделенный полинуклеотид из вируса по любому из пп.1, 5-7, который кодирует иммуногенный полипептид, индуцирующий иммунный ответ против вируса гриппа, где полипептид имеет аминокислотную последовательность, показанную в любой из последовательностей SEQ ID No:1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 47, 49, 51, 53, 55, 57, 59, 61, 63, 65, 67, 69, 71, 73, 75 или 77, или указанный полинуклеотид кодирует полипептид, характеризующийся 98%-ной или большей идентичностью по последовательности с аминокислотной последовательностью, показанной в любой последовательностей из SEQ ID No:1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 47, 49, 51, 53, 55, 57, 59, 61, 63, 65, 67, 69, 71, 73, 75 или 77.
9. Полинуклеотид по п.8, отличающийся тем, что указанный полинуклеотид кодирует полипептид, имеющий аминокислотную последовательность, показанную в любой из последовательностей SEQ ID No:2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 33, 34, 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70, 72, 74, 76 или 78, или его функциональный и/или иммуногенный фрагмент, или указанный полинуклеотид кодирует полипептид, характеризующийся 99%-ной или большей идентичностью по последовательности с аминокислотной последовательностью, показанной в любой из последовательностей SEQ ID No:2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 33, 34, 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70, 72, 74, 76 или 78.
10. Полинуклеотид по п.8, отличающийся тем, что указанный полинуклеотид имеет нуклеотидную последовательность, показанную любой из последовательностей SEQ ID No:1, 3, 5, 7, 9, 11, 13, 15, 17, 19,21, 23, 25, 27, 29, 31, 47, 49, 51, 53, 55, 57, 59, 61, 63, 65, 67, 69, 71, 73, 75 или 77, или указанный полинуклеотид обладает 98%-ной или большей идентичностью с нуклеотидной последовательностью, показанной любой из последовательностей SEQ ID No:1, 3, 5, 7, 9, 11, 13, 15, 17, 19, 21, 23, 25, 27, 29, 31, 47, 49, 51, 53, 55, 57, 59, 61, 63, 65, 67, 69, 71, 73, 75 или 77.
11. Полинуклеотид по п.8, отличающийся тем, что указанный полинуклеотид кодирует гемагглютинин (НА), включающий аминокислотную последовательность SEQ ID No:33, или полинуклеотид кодирует полипептид, характеризующийся 99%-ной или большей идентичностью с аминокислотной последовательностью SEQ ID No:33.
12. Полинуклеотид по п.8, отличающийся тем, что указанный полинуклеотид кодирует гемагглютинин (НА), включающий аминокислотную последовательность SEQ ID No:33, или полинуклеотид кодирует полипептид, характеризующийся 95%-ной или большей идентичностью с аминокислотной последовательностью SEQ ID No:33, где указанный белок НА содержит серин в положении 82, лейцин в положении 221, треонин в положении 327 и треонин в положении 482 указанной последовательности.
13. Полинуклеотид по п.8, отличающийся тем, что указанный полинуклеотид кодирует гемагглютинин (НА), включающий аминокислотную последовательность SEQ ID No:16, или полинуклеотид кодирует полипептид, характеризующийся 95%-ной или большей идентичностью с аминокислотной последовательностью SEQ ID No:16.
14. Вектор экспрессии полипептида, который способен индуцировать иммунный ответ против вируса гриппа, где вектор включает полинуклеотид по любому из пп.8-13, и регуляторные последовательности.
15. Вектор по п.14, отличающийся тем, что указанный полинуклеотид кодирует гемагглютинин (НА), включающий аминокислотную последовательность SEQ ID No:33 или полинуклеотид кодирует полипептид, характеризующийся 99%-ной или большей идентичностью с аминокислотной последовательностью SEQ ID No:33.
16. Вектор по п.14, отличающийся тем, что указанный полинуклеотид кодирует гемагглютинин (НА), включающий аминокислотную последовательность SEQ ID No:33, или полинуклеотид кодирует полипептид, характеризующийся 95%-ной или большей идентичностью с аминокислотной последовательностью SEQ ID No:33, где указанный белок НА содержит серин в положении 82, лейцин в положении 221, треонин в положении 327 и треонин в положении 482 указанной последовательности.
17. Вектор по п.14, отличающийся тем, что указанный полинуклеотид кодирует гемагглютинин (НА), включающий аминокислотную последовательность SEQ ID No:16, или полинуклеотид кодирует полипептид, характеризующийся 95%-ной или большей идентичностью с аминокислотной последовательностью SEQ ID No:16.
18. Выделенный полипептид, который способен индуцировать иммунный ответ против вируса гриппа, который способен инфицировать животное из группы псовых, где указанный полипептид имеет аминокислотную последовательность, показанную в любой из последовательностей SEQ ID No:2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 33, 34, 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70, 72, 74, 76 или 78, или указанный полипептид характеризуется 99%-ной или большей идентичностью по последовательности с аминокислотной последовательностью, показанной в любой из последовательностей SEQ ID No:2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 33, 34, 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70, 72, 74, 76 или 78, или его функциональный и/или иммуногенный фрагмент.
19. Полипептид по п.18, отличающийся тем, что указанный полипептид выбирают из группы, состоящей из белка гемагглютинина, белка нейраминидазы, нуклеопротеина, матричного белка, основного белка полимеразы, кислого белка полимеразы и неструктурного белка.
20. Полипептид по п.18, отличающийся тем, что указанный полинуклеотид кодирует гемагглютинин (НА), включающий аминокислотную последовательность SEQ ID No:33, или полинуклеотид кодирует полипептид, характеризующийся 99%-ной или большей идентичностью с аминокислотной последовательностью SEQ ID No:33.
21. Полипептид по п.18, отличающийся тем, что указанный полинуклеотид кодирует гемагглютинин (НА), включающий аминокислотную последовательность SEQ ID No:33, или полинуклеотид кодирует полипептид, характеризующийся 95%-ной или большей идентичностью с аминокислотной последовательностью SEQ ID No:33, где указанный белок НА содержит серин в положении 82, лейцин в положении 221, треонин в положении 327 и треонин в положении 482 указанной последовательности.
22. Полипептид по п.18, отличающийся тем, что указанный полинуклеотид кодирует гемагглютинин (НА), включающий аминокислотную последовательность SEQ ID No:16, или полинуклеотид кодирует полипептид, характеризующийся 95%-ной или большей идентичностью с аминокислотной последовательностью SEQ ID No:16.
23. Иммуногенная композиция, включающая эффективное количество полипептида по любому из пп.18-22 и эксципиенты, где указанная композиция способна индуцировать иммунный ответ против вируса гриппа, который способен инфицировать животное из группы псовых.
24. Композиция по п.23, отличающаяся тем, что указанный полипептид представляет собой полипептид с аминокислотной последовательностью, показанную в любой из последовательностей SEQ ID No:2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 33, 34, 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70, 72, 74, 76 или 78, или его функциональный и/или иммуногенный фрагмент, или указанный полипептид характеризуется 99%-ной или большей идентичностью по последовательности с аминокислотной последовательностью, показанной в виде любой из SEQ ID No:2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 33, 34, 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70, 72, 74, 76 или 78.
25. Способ индукции иммунного ответа у животного против вируса гриппа, который способен инфицировать животное из группы псовых, где указанный способ включает введение указанному животному эффективного количества композиции по п.23 или 24.
26. Способ по п.25, отличающийся тем, что указанная композиция содержит вирусный белок, полипептид или пептид, включающий аминокислотную последовательность, показанную в виде любой из SEQ ID No:2, 4, 6, 8, 10, 12, 14, 16, 18, 20, 22, 24, 26, 28, 30, 32, 33, 34, 48, 50, 52, 54, 56, 58, 60, 62, 64, 66, 68, 70, 72, 74, 76 или 78, или его функциональный и/или иммуногенный фрагмент.
27. Способ по п.25, отличающийся тем, что указанный полинуклеотид кодирует гемагглютинин (НА), включающий аминокислотную последовательность SEQ ID No:33, или полинуклеотид кодирует полипептид, характеризующийся 99%-ной или большей идентичностью с аминокислотной последовательностью SEQ ID No:33.
28. Способ по п.25, отличающийся тем, что указанный полинуклеотид кодирует гемагглютинин (НА), включающий аминокислотную последовательность SEQ ID No:33, или полинуклеотид кодирует полипептид, характеризующийся 95%-ной или большей идентичностью с аминокислотной последовательностью SEQ ID No:33, где указанный белок НА содержит серин в положении 82, лейцин в положении 221, треонин в положении 327 и треонин в положении 482 указанной последовательности.
29. Способ по п.25, отличающийся тем, что указанный полинуклеотид кодирует гемагглютинин (НА), включающий аминокислотную последовательность SEQ ID No:16, или полинуклеотид кодирует полипептид, характеризующийся 95%-ной или большей идентичностью с аминокислотной последовательностью SEQ ID No:16.
30. Способ по п.25, отличающийся тем, что указанный ответ представляет собой защитную иммунную реакцию, которая препятствует инфекции или ингибирует инфицирование указанного животного вирусом гриппа.
31. Способ по п.25, отличающийся тем, что указанный вирус гриппа имеет НА-серотип, выбранный из Н3 и Н5.
32. Способ по п.25, отличающийся тем, что указанный вирус гриппа имеет серотип H3N8.
33. Способ по п.25, отличающийся тем, что указанная композиция включает также адъювант.
34. Способ по п.25, отличающийся тем, что указанное животное является представителем псовых.
35. Способ по п.34, отличающийся тем, что указанное животное из псовых представляет собой домашнюю собаку.
36. Способ по п.25, отличающийся тем, что указанную композицию вводят парентерально.
37. Способ по п.36, отличающийся тем, что указанную композицию вводят подкожно, внутрибрюшинно или внутримышечно.
38. Способ по п.25, отличающийся тем, что указанную композицию вводят назально или перорально.
| MANUGUERRA еt al | |||
| Natural infection of dogs by influenza С virus // RESEARCH IN VIROLOGY, ELSEVIER, PARIS, FR, vol.143, 1 January 1992 (1992-01-01), pages 199-204 | |||
| OXBURGH L | |||
| et al | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| LAI A.C | |||
| et al | |||
| Alternate circulation of recent | |||

