Изобретение относится к области фотоники и вычислительной техники, конкретно к созданию оптических устройств, действие которых основано на фотохромных свойствах молекул ретинальсодержащих белков, и может быть использовано для создания устройств оптической обработки данных высокого быстродействия.
Ретинальсодержащие белки (РСБ) составляют группу светочувствительных мембранных белков, способных преобразовывать световую энергию для осуществления биологических функций живых организмов. В эту группу входят сенсорные родопсины, обеспечивающие восприятие света как носителя информации (светочувствительные зрительные пигменты животных, сенсорные родопсины микроорганизмов), и родопсины, осуществляющие фотоэнергетическую функцию - направленный транспорт ионов через клеточную мембрану (например, галородопсин, бактериородопсин), а также мутантные формы светочувствительных белков и другие искусственно модифицированные РСБ. Эти белки имеют сходную пространственную организацию и общую для всех хромофорную группу - остаток ретиналя (альдегида витамина A), ковалентно связанный с белком путем образования Шиффова основания с остатком лизина полипептидной цепи белка.
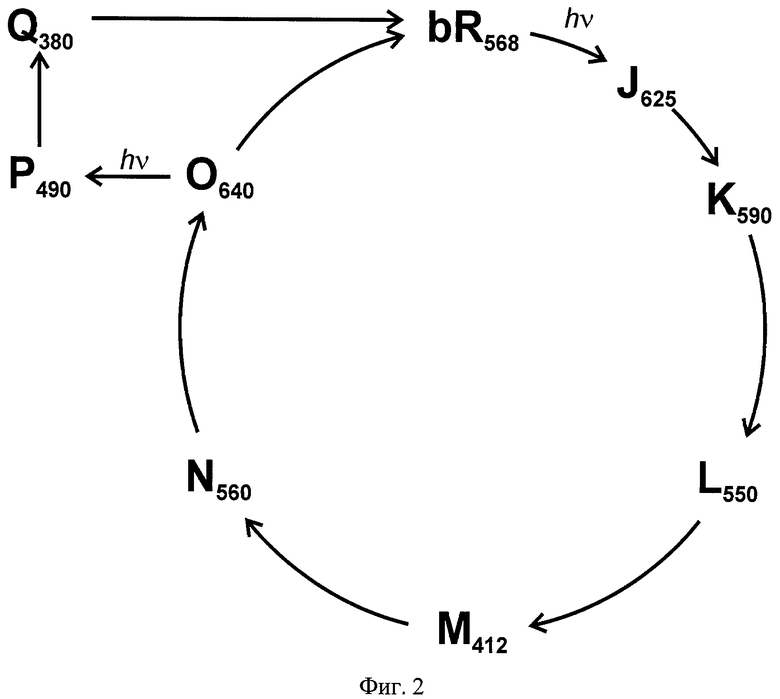
При поглощении света ретинальсодержащим белком происходит изомеризация остатка ретиналя, инициирующая многостадийный процесс превращения исходной молекулы, в результате которого образуются несколько последовательных, различающихся по стабильности и оптическим характеристикам промежуточных состояний. В случае зрительного пигмента родопсина позвоночных животных конформационные превращения приводят к высвобождению ретиналя (фотолизу), а в случае зрительных пигментов беспозвоночных животных и бактериородопсина - к возвращению в исходное состояние (замыканию фотоцикла). На Фиг.1 и 2 в качестве иллюстрации показаны схемы фотопревращений зрительного пигмента родопсина быка и бактериородопсина галобактерий.
Известно, что, воздействуя световыми импульсами с определенными параметрами на долгоживущие состояния (например, состояние M фотоцикла бактериородопсина и др.), можно прервать этот процесс и запустить обратную реакцию, в результате которой молекула белка вернется в начальное состояние или перейдет в другой интермедиат. В дальнейшем по тексту под термином «фотопереключение» будем понимать перевод молекулы светочувствительного белка под действием световых импульсов из одного состояния, характеризующегося определенными оптическими и другими физико-химическими характеристиками, в другое состояние, характеризующееся отличными от первого оптическими и другими физико-химическими характеристиками.
На способности РСБ к фотопереключению под действием световых импульсов, сопровождаемому изменением оптических и других физико-химических свойств, основано их применение в качестве светочувствительной среды в оптических устройствах для записи, передачи и хранения информации. Быстродействие устройств, включающих в качестве рабочей среды РСБ, определяется тем, какие промежуточные состояния РСБ вовлечены в процесс фотопереключения. Общим требованием является возможность реализации выбранных состояний РСБ в интервале температур, в котором предполагается использование оптических устройств, как правило, при комнатной температуре.
В работе [M.A.Ostrovsky and H.H.Weetall "Octopus rhodopsin photoreversibility of a crude extract from whole retina over several weeks' duration", Biosensor & Bioelectronics, 1998, vol.13, No 1, p.61-65] описано фотопереключение зрительного пигмента родопсина осьминога при комнатной температуре в минутном диапазоне из исходного состояния в метародопсин (кислая форма) под действием светового облучения в спектральной области более 570 нм с последующим возвращением его в исходное состояние под действием синего света с длиной волны менее 440 нм. В работе [C.Paternolli et. al. "Photoreversibility and photostability in films of octopus rhodopsin isolated from octopus photoreceptor membranes", Journal of Biomedical Materials Research Part A, 2008, Vol.88A Issue 4, p.947-951] фотопереключение родопсина осьминога осуществляли в секундном диапазоне при использовании спектрально узких вспышек света, центрированных на длинах волн 546 и 431 нм.
В случае бактериородопсина обычно используют промежуточные, относительно долго живущие M-, Q-состояния (Фиг.2). Например, функционирование оптической объемной памяти согласно изобретению, защищенному патентом [US 5559732, опубл. 24.09.1996], основано на фотопереключении бактериородопсина при комнатной температуре из исходного bR-состояния в метастабильное O-состояние под действием первого светового импульса с длиной волны 570-600 нм и последующем переводе молекулы белка из метастабильного O-состояния в более стабильные Р- и Q-состояния, соответствующие разветвленному фотоциклу (см. Фиг.2), под действием второго импульса с длиной волны 680 нм, при этом длительность светового воздействия находится в миллисекундном диапазоне. Использованный в данном изобретении способ фотопереключения бактериородопсина приводит к долгоживущим промежуточным продуктам, пригодным для длительного хранения информации.
Известен оптический логический элемент для хранения и обработки данных, также содержащий в качестве светочувствительного оптического материала бактериородопсин [RU 2186418, C2, опубл. 27.07.2002], причем в качестве исходного состояния используют метастабильное состояние М, созданное заранее под воздействием желтого света высокой интенсивности с длиной волны около 570 нм и длительностью менее 1 мс, или Q-интермедиат, получаемый воздействием на исходный бактериородопсин света высокой интенсивности с диапазоном длин волн 570-640 нм длительностью более 2 мс. Оба описанных способа фотопереключения бактериородопсина приводят к стабильным долгоживущим состояниям, которые не могут обеспечить быстродействия оптических устройств в субпикосекундной шкале времени.
Известно оптическое устройство, предназначенное для использования в качестве логического элемента, оптического переключателя или фотосенсора [US 2003152358, A1, опубл. 14.08.2003], представляющее собой волновод, покрытый материалом с нелинейными оптическими свойствами, преимущественно бактериородопсином, изменяющим показатель преломления при переходе из исходного bR-состояния в метастабильное M-состояние под действием наносекундного импульса. В описании упомянуто о том, что увеличить быстродействие устройства можно, варьируя использование промежуточных состояний фотоцикла бактериородопсина или его аналогов, однако примеров осуществления этой возможности авторы не приводят.
Наиболее привлекательным, с точки зрения возможности достижения максимального быстродействия, является использование ранних стадий фотоинициированных превращений, например фото- и батородопсина для зрительного пигмента родопсина, K-состояния для бактериородопсина.
В работе [M.Yan, L.Rothberg, R.Callender "Femtosecond dynamics of rhodopsin photochemistry probed by a double pump spectroscopic approach", J. Phys. Chem. В 2001, 105, p.856-859] описана попытка осуществить обратимое сверхбыстрое фотопереключение интермедиатов зрительного пигмента родопсина быка, в частности батородопсина, под действием фемтосекундных (300 фс) лазерных импульсов: первого импульса с длиной волны 500 нм, энергией 1 мкДж и второго с длиной волны 605 нм, энергией 10 мкДж. Однако в работе не содержится убедительных подтверждений того, что фотопереключение осуществлено в субпикосекундной шкале времени.
В работе [V.Bazhenov, P.Schmidt and G.Н.Atkinson "Nanosecond photolytic interruption of bacteriorhodopsin photocycle K-590→BR-570 reaction", Biophys. J., Vol.61, June 1992, p.1630-1637] описан способ фотопереключения бактериородопсина, позволяющий прервать фотоцикл в наносекундной шкале времени. Способ включает воздействие на белок двумя последовательными световыми импульсами: первым с длиной волны 532 нм и вторым с длиной волны в интервале 620-700 нм. Длительность импульсов - 10 нс, задержка второго импульса относительно первого - 20 нс. Плотность энергии первого импульса - 0,3 мДж/мм2, второго - до 20 мДж/мм2. Способ позволяет переводить бактериородопсин в К-состояние и обратно в наносекундном диапазоне.
В качестве прототипа настоящего изобретения принят способ фотопереключения бактериородопсина, описанный в работе [J.K.Delaney, P.K.Schmidt, T.L.Brack and G.Н.Atkinson "Photochemistry of K-590 in the room-temperature bacteriorhodopsin photocycle" J. Phys. Chem. В 2000, 104, p.10827-10834]. Способ включает воздействие на белок при температуре 10°C двумя последовательными световыми импульсами с длиной волны соответственно 578 нм и 650-660 нм. Длительность импульсов 4-5 пс, задержка второго импульса относительно первого 50 пс и более, энергия около 40 нДж на импульс. Получено обратимое фотопереключение молекул бактериородопсина из исходного bR-состояния в интермедиат K и обратно, при этом быстродействие фотопереключения измеряется в пикосекундном диапазоне.
Для современных оптических компьютеров необходимы элементы с быстродействием переключения в субпикосекундной шкале времени, достигаемым при комнатной температуре.
Задачей настоящего изобретения является разработка способа сверхбыстрого фотопереключения молекул РСБ при комнатной температуре в субпикосекундной шкале времени, а также разработка оптического логического элемента, действие которого основано на применении этого способа и предназначенного для использования в сверхбыстрых оптических устройствах, работающих в субпикосекундной шкале времени.
Поставленная задача решается предлагаемым способом фотопереключения РСБ, включающим перевод белка при комнатной температуре из исходного состояния в промежуточное состояние путем воздействия на него первым световым импульсом с длиной волны, находящейся в диапазоне, соответствующем области поглощения исходного состояния, и последующее его возвращение в исходное состояние путем воздействия вторым световым импульсом с длиной волны, находящейся в диапазоне, соответствующем области, в которой происходит поглощение промежуточного состояния, но не происходит поглощение исходного состояния, при этом длительность указанных световых импульсов не превышает 50 фс, а промежуток времени между указанными световыми импульсами не превышает 6 пс.
Поставленная задача решается также предлагаемым оптическим логическим элементом, содержащим ретинальсодержащий белок, который подвергают фотопереключению из исходного состояния в промежуточное состояние и обратно, а указанное фотопереключение сопровождается соответствующими изменениями значения оптического логического элемента, при этом фотопереключение осуществляют при комнатной температуре под действием двух последовательных световых импульсов, причем длина волны первого светового импульса находится в диапазоне, соответствующем области поглощения исходного состояния, длина волны второго светового импульса находится в диапазоне, соответствующем области, в которой происходит поглощение промежуточного состояния, но не происходит поглощение исходного состояния, при этом длительность указанных световых импульсов не превышает 50 фс, а промежуток времени между указанными световыми импульсами не превышает 6 пс.
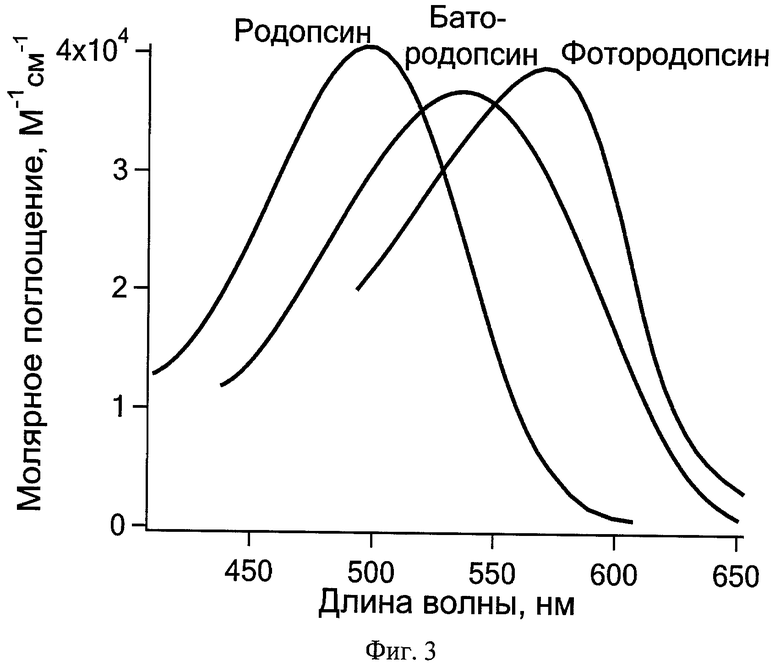
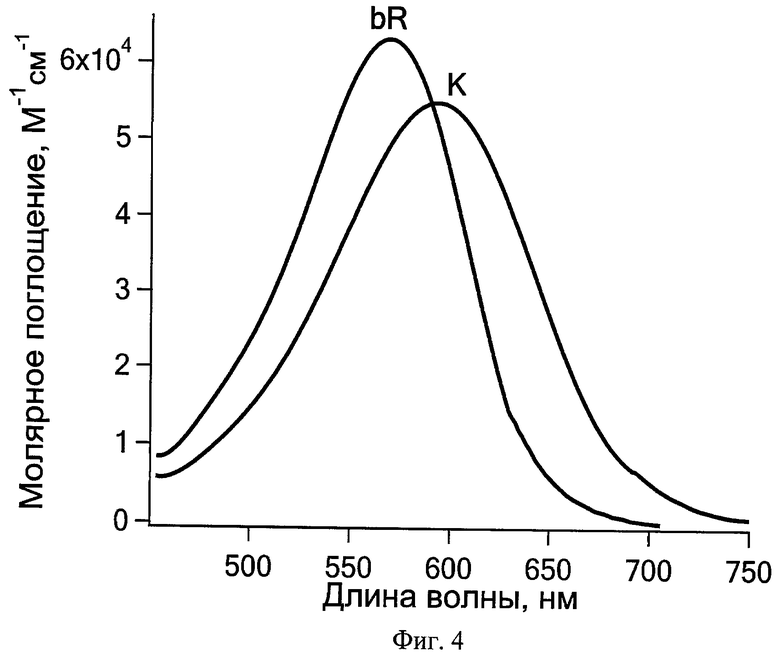
В основе решения поставленной задачи лежат результаты проведенных авторами исследований, которые показали, что с помощью следующих друг за другом первого hv1 и второго hv2 импульсов фемтосекундной длительности при комнатной температуре возможно осуществить возвращение в исходное состояние интермедиатов фотопревращений РСБ, содержащих изомеризованный ретиналь: для зрительного пигмента родопсина животных - фотородопсина (время образования - 200 фс, квантовый выход - 0,65), для бактериородопсина - K-состояния (время образования - примерно 3 пс, квантовый выход - 0,64). Спектры интермедиатов и исходных состояний зрительного пигмента родопсина быка и бактериородопсина галобактерий представлены на Фиг.3 и 4.
В качестве фотохромных материалов можно использовать различные ретинальсодержащие белки: зрительные пигменты родопсина беспозвоночных и позвоночных животных, сенсорные родопсины и светозависимые ионные насосы микроорганизмов. Возможно использование рекомбинантных белков, различных мутантных и модифицированных форм белков, причем модификации могут касаться как пептидной части молекулы, так и хромофора. Сходство строения молекул различных РСБ и сходство механизмов превращений, связанных с изомеризацией остатка ретиналя, дают основание считать, что предлагаемым способом может быть осуществлено сверхбыстрое фотопереключение любого РСБ, независимо от его происхождения. Перспективным является использование и других материалов, содержащих ретиналь в качестве хромофора. Например, в работе [H.Kandori, T.Ichioka, M.Sasaki "Photoisomerization of the rhodopsin chromophore in clay interlayers at 77K", Chemical Physics Letters, 2002, 354, p.251-255] показана возможность фотоизомеризации ретиналя в модифицированных детергентом слоях бентонитовой глины.
Диапазон длин волн первого светового импульса, используемого при реализации заявляемого способа, определяется спектральным положением полосы поглощения ретиналя в белковом окружении (α-полосы). Для бычьего зрительного пигмента родопсина значительное поглощение ретиналя в белковом окружении соответствует интервалу λ=470-530 нм, для бактериородопсина - интервалу λ=540-600 нм. При возбуждении в более коротковолновом диапазоне β- или γ-полос происходит передача энергии с аминокислот на ретиналь, что может привести к замедлению фотопереключения.
Диапазон длин волн второго светового импульса выбирают так, чтобы его селективно поглощал интермедиат, а не исходное состояние. Диапазон длин волн второго светового импульса для бычьего зрительного пигмента родопсина составляет λ=600-650 нм, для бактериородопсина λ=670-720 нм.
Длительность первого и второго импульсов не превышает 50 фс. Использование фемтосекундных импульсов позволяет достигать фотопереключения при низкой энергии импульсов (E) и предотвратить деструкцию фотохромного материала. Меняя различные параметры фемтосекундных импульсов, например частотно-фазовые характеристики, поляризацию, можно повысить эффективность фотопереключения РСБ из того или иного состояния.
Временной промежуток, разделяющий первый и второй импульсы, зависит от времени фотоизомеризации ретиналя в конкретном белковом окружении и составляет от 200 фс до 6 пс.
Фотопереключение регистрируют с помощью третьего (зондирующего) импульса в широком спектральном диапазоне λ=400-750 нм с полной энергией менее 10 нДж и длительностью менее 800 фс.
Концентрацию РСБ выбирают в соответствии с техническими возможностями детекции фотопереключения. В общем случае можно использовать сколь угодно малые концентрации, поскольку возможно фотопереключение одной молекулы, однако для получения достоверного сигнала предпочтительно использовать концентрации РСБ, составляющие несколько мг/мл.
Существенные отличия заявляемого способа от известных аналогов и прототипа состоят в том, что длительность первого и второго импульсов не превышает 50 фс, а разделяющий их временной промежуток составляет от 200 фс до 6 пс, что позволяет осуществить сверхбыстрое субпикосекундное переключение РСБ.
Техническим результатом заявляемого способа фотопереключения РСБ является то, что он позволяет при комнатной температуре осуществить фотопереключение РСБ в субпикосекундном диапазоне.
Использование предлагаемого способа фотопереключения РСБ позволит многократно повысить быстродействие оптических устройств, действие которых основано на применении РСБ в качестве светочувствительной рабочей среды.
Краткое описание чертежей
На Фиг.1 показана схема фотолиза зрительного пигмента родопсина быка.
На Фиг.2 показана схема фотоцикла светоадаптированного бактериородопсина галобактерий.
На Фиг.3 показаны спектры поглощения бычьего зрительного пигмента родопсина в исходном состоянии, фотородопсина и батородопсина.
На Фиг.4 показаны спектры поглощения бактериородопсина и K-интермедиата.
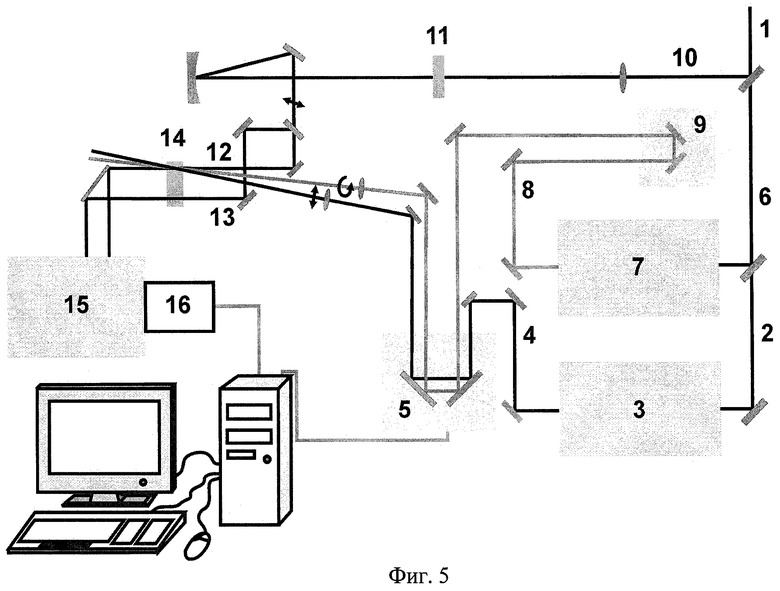
На Фиг.5 показана схема фемтосекундной оптической установки, использованной для реализации заявляемого способа.
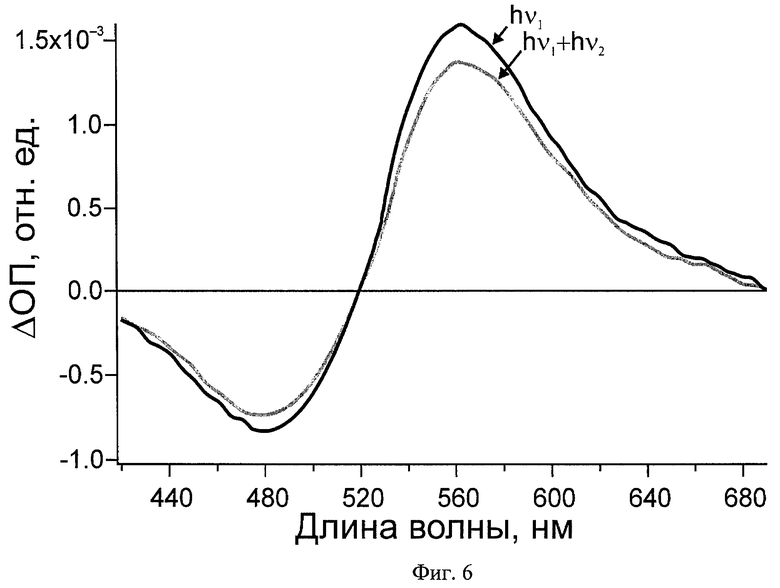
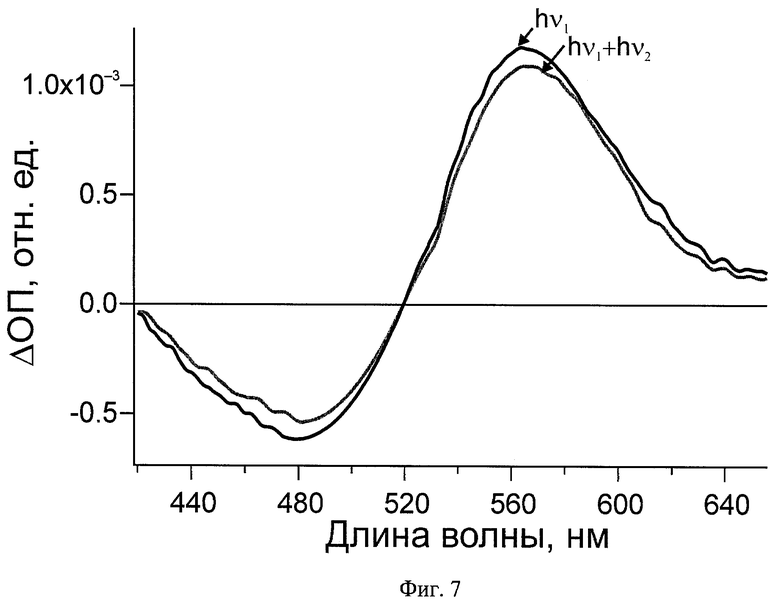
На Фиг.6 и 7 показаны дифференциальные спектры поглощения зрительного пигмента родопсина быка при реализации заявляемого способа: hν1 - дифференциальный спектр после действия первого импульса, hν1+hν2 - дифференциальный спектр после действия первого и второго импульсов.
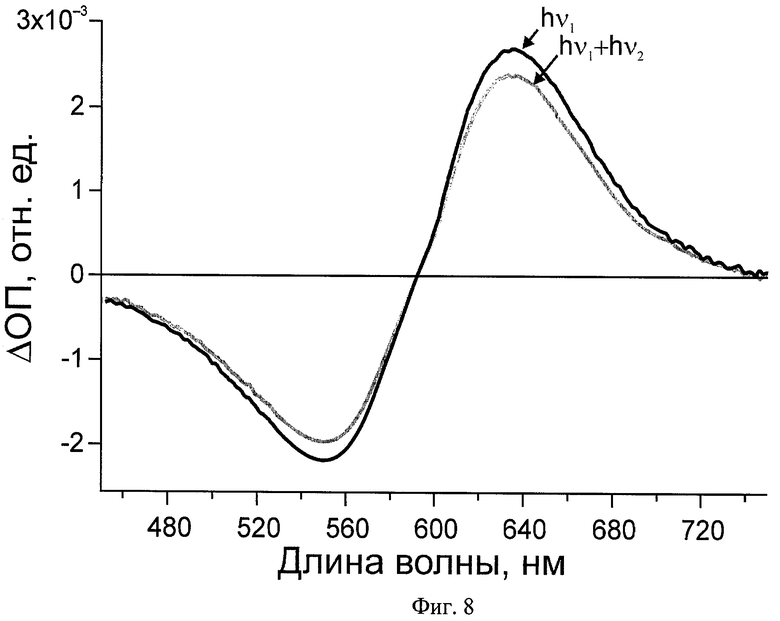
На Фиг.8 показаны дифференциальные спектры поглощения бактериородопсина при реализации заявляемого способа: hν1 - дифференциальный спектр после действия первого импульса, hν1+hν2 - дифференциальный спектр после действия первого и второго импульсов.
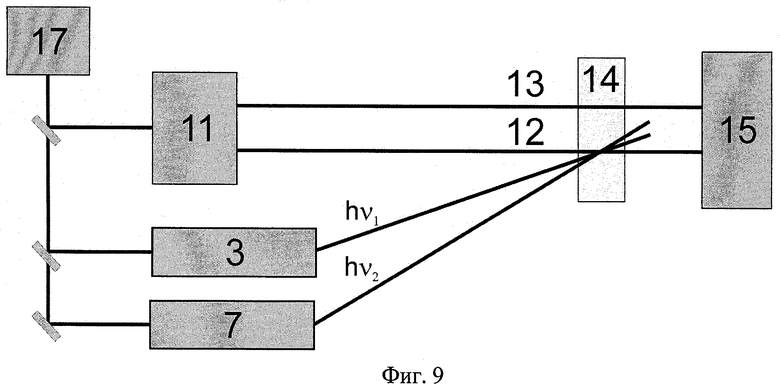
На Фиг.9 показана схема оптического логического элемента, действующего на основе заявляемого способа.
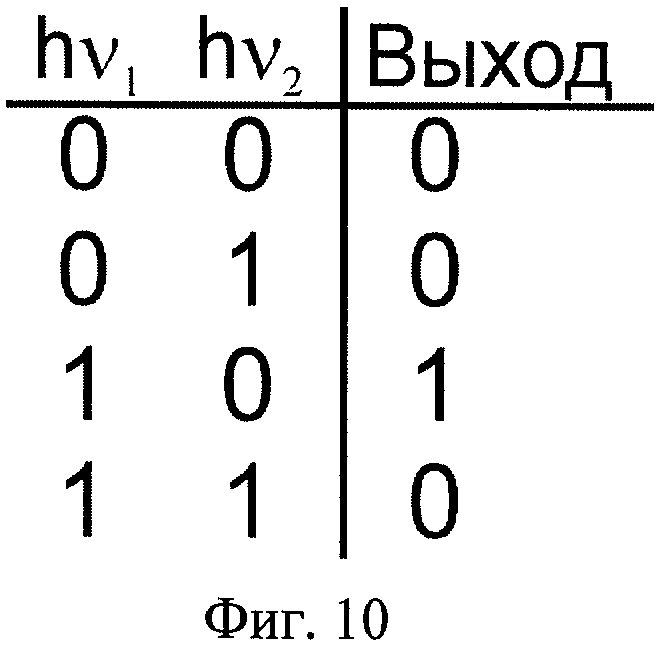
На Фиг.10 показана таблица истинности оптического логического элемента, действующего на основе заявляемого способа.
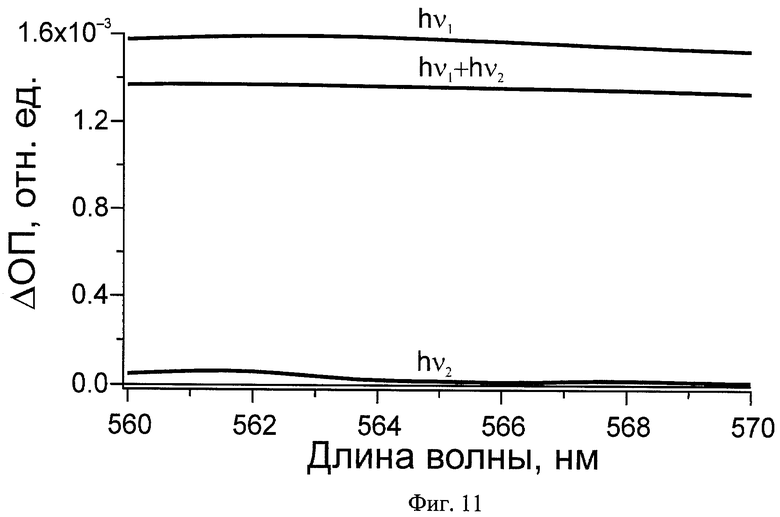
На Фиг.11 показано изменение выходного сигнала при разных режимах работы оптического логического элемента, действующего на основе заявляемого способа: hν1 - дифференциальный спектр после действия первого импульса, hν1+hν2 - дифференциальный спектр после действия первого и второго импульсов, hν2 - дифференциальный спектр после действия второго импульса.
Для осуществления заявляемого способа фотопереключения ретинальсодержащих белков используют фемтосекундную оптическую установку, схема которой показана на Фиг.5.
Фемтосекундные импульсы генерируют в титан-сапфировом лазере Tsunami «Spectra Physics» с накачкой непрерывным излучением твердотельного лазера MilleniaVs «Spectra Physics» с диодной накачкой и усиливают в регенеративном усилителе Spitfire «Spectra Physics» с накачкой излучением лазера Evolution X «Spectra Physics».
Для приготовления первого, второго и зондирующего импульсов усиленное излучение 1 делят на три пучка. Луч 2, предназначенный для получения первого импульса, направляют в неколлинеарный параметрический усилитель света (NOPA) «Clark MXR» 3, который преобразовывает несущую длину волны 805 нм для фотопереключения зрительного пигмента родопсина быка в 500 нм, а для фотопереключения бактериородопсина - в 560 нм, после чего первый импульс 4 направляют в линию задержки 5. Луч 6, предназначенный для получения второго импульса, направляют в неколлинеарный параметрический усилитель света «Light Conversion» Topas-white 7, где импульс приобретает несущую длину волны 620 нм для фотопереключения зрительного пигмента родопсина быка и 680 нм - для бактериородопсина. Затем луч 8 проходит линию задержки 9, с помощью которой устанавливают задержку между первым и вторым импульсами. После этого его направляют на общую для обоих пучков линию задержки 5, которая позволяет задерживать первый и второй импульсы относительно зондирующего импульса 12 с минимальным шагом 3,3 фс. Третью часть усиленного лазерного излучения 10 направляют на приготовление зондирующего импульса. Пучок ослабляют до энергии менее 2 мкДж и фокусируют в кювету с водой 11, где генерируют импульс суперконтинуума со спектром 400-1000 нм и полной энергией менее 10 нДж. Полученный импульс суперконтинуума делят на два луча: зондирующий 12 и опорный 13 импульсы. Лучи 4, 8 и 12 фокусируют внутри кварцевой кюветы 14 толщиной 0,5 мм, через которую прокачивают образец белка объемом 3 мл. Диаметры перетяжек первого и второго импульсов в образце - 150 мкм, зондирующего импульса - 100 мкм. Зондирующий импульс линейно поляризован и направлен под магическим углом 54,7° относительно первого, а второй импульс имеет циркулярную поляризацию. Опорный луч 13 проходит через кювету 14 в области, не подверженной действию первого и второго импульсов. Зондирующий и опорные импульсы после кюветы попадают в полихроматор «Acton» SP-300 15. Их спектры регистрирует CCD-камера 16 системы «Roper Scientific» SPEC-10, после чего данные поступают в компьютер для первичной обработки.
Для получения спектров дифференциального поглощения лучи 4 и 8 при каждом времени задержки перекрывают механическим затвором, управляемым компьютером. Спектры дифференциального поглощения вычисляют по формуле:
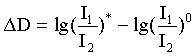 ,
,
где I1 и I2 - интенсивности импульсов 12 и 13, измеряемые при открытом (*) и закрытом (0) затворе накачки.
Для получения достоверного сигнала производят усреднение 10000 спектров.
Спектры первого и второго импульсов близки к Гауссову распределению, и их ширина на полувысоте не превышает 20 нм. Используют низкие плотности энергии первого и второго импульсов (менее 5 Дж/м2), которые позволяют осуществлять однофотонное поглощение. Фотопереключения и измерения проводят при комнатной температуре.
Приведенные ниже примеры демонстрируют возможность осуществления заявляемого способа с достижением заявляемого технического результата.
Пример 1. Фотопереключение зрительного пигмента родопсина Bos taurus за 200 фс
Наружные сегменты палочек сетчатки глаза быка Bos taurus получают при слабом красном освещении из свежих (не позднее 3-х часов после забоя) глаз в соответствии с методом препаративного центрифугирования в градиенте плотности сахарозы, описанным в работе [Smith H.G., Stubb G.W., Litman B.J. "The isolation and purification of osmotically intact discs from retinal rod outer segments" Exp. Eye Res., 1975, vol.20: p.211-217].
Экстракты зрительного пигмента родопсина из наружных сегментов палочек получают модифицированным методом Окады, используя 1,6% раствор детергента н-нонил-β-D-гептилтиоглюкозида (HTG), приготовленный на буфере (0,1 M CH3COONa, 0,1 М CH3COOZn, 0,3 М сульфат аммония, 0,01% NaN3, pH 6,0). Экстракт инкубируют 3 ч при 20° и 12 ч при 4°. После 30-минутного центрифугирования при 41400 g экстракт родопсина фильтруют с помощью фильтров Millipore (Millex GS PVDF 0,22 µm) и концентрируют с помощью фильтров Millipore (Amicon Ultra-4 Ultracell 30k). Получают препарат родопсина в HTG с концентрацией 4 мг/мл.
Полученный фотохромный материал, содержащий зрительный пигмент родопсин, используют сначала в режиме с первым и зондирующим импульсами. В результате воздействия первого импульса 4 (λ=500 нм, t=25 фс, E=70 нДж) происходит фотопереключение родопсина в фотородопсин. Соответствующее изменение дифференциального спектра поглощения через 100 пс после первого импульса hν1, регистрируемое с помощью зондирующего импульса 12 во всем видимом диапазоне, показано на Фиг.6.
Фотопереключение из стадии фотородопсина в родопсин осуществляют через 200 фс при помощи второго импульса 8 (λ=620 нм, t=40 фс, E=450 нДж), который переводит часть фотородопсина в родопсин, что подтверждает изменение дифференциального спектра поглощения при 100 пс (Фиг.6, hν1+hν2). В области обесцвечивания родопсина (отрицательное дифференциальное поглощение) сигнал увеличивается за счет частичного восстановления исходного состояния. В более длинноволновой области поглощения фотопродуктов наблюдается спад сигнала, поскольку второй импульс индуцирует обратную реакцию.
Детектирование в широком спектральном диапазоне убедительно подтверждает обратимое фотопереключение зрительного пигмента родопсина за 200 фс и дает возможность выбора подходящей области регистрации для реализации способа в различных оптических устройствах.
Использование низких значений плотности энергии и длительности зондирующего импульса позволяет сохранять большую часть молекул белка в детектируемом состоянии, что дает возможность длительно использовать образцы РСБ в оптических устройствах.
Пример 2. Фотопереключение зрительного пигмента родопсина Bos taurus за 6 пс
Фотопереключение зрительного пигмента родопсина быка осуществляют, как описано в примере 1, за исключением того, что концентрация родопсина составляет 2,8 мг/мл, а промежуток времени между первым импульсом (λ=500 нм, t=30 фс, E=70 нДж) и вторым импульсом (λ=620 нм, t=50 фс, E=470 нДж) составляет 6 пс. Полученые дифференциальные спектры, представленные на Фиг.7, подтверждают фотопереключение родопсина в батородопсин и обратно.
Пример 3. Фотопереключение бактериородопсина Halobacterium salinarum за 3 пс
Препарат мембран Halobacterium salinarum готовят по модифицированной методике, описанной в [Xu ZJ, Moffett DB и др. J Biol Chem. 1995, v.270, p.24858-24863].
Культуру Halobacterium salinarum, штамм DSM 671, высевают на агаризованную питательную среду A, содержащую 250 г/л NaCl, 20 г/л MgSO4, 2 г/л KCl, 3 г/л Na-цитрата, 5 г/л пептона, 2 г/л дрожжевого экстракта, и выращивают в течение 10 дней при 37°C. Колонии собирают, пересевают в 50 мл среды А и культуру выращивают 7 дней при 37°C в режиме интенсивного перемешивания и непрерывного освещения лампами ЛБ-40.
50 мл клеточной суспензии центрифугируют (10 мин, 3000 g), клетки лизируют путем суспендирования осадка клеток в 2 мл H2O и 0,1 мг ДНК-азы I в присутствии 5 mM MgCl2 в течение 20 мин при 4°С, затем озвучивают на приборе Branson Sonifier в течение 2 мин. Мембраны осаждают центрифугированием при 80000 g в течение 1,5 час при 10°C, осадок мембран ресуспендируют в 2 мл H2O путем озвучивания и процедуру центрифугирования и ресуспендирования повторяют еще два раза. Для спектрофотометрического анализа осадок мембран суспендируют в 1 мл H2O. Концентрация бактериородопсина в препарате составляет 3 мг/мл.
Светоадаптированный препарат пурпурных мембран подвергают воздействию первого импульса 4 (λ=560 нм, t=20 фс, E=100 нДж), что приводит к запуску фотоцикла (Фиг.2). Регистрацию дифференциального спектра бактериородопсина проводят при 100 пс зондирующим импульсом 12 (Фиг.8, hν1).
Фотоцикл бактериородопсина прерывают на стадии интермедиата K и запускают обратную реакцию путем воздействия на него вторым импульсом 8 (λ=680 нм, t=30 фс, E=560 нДж), задержанным по времени относительно первого на 3 пс. Фотопереключение бактериородопсина регистрируют в широком спектральном диапазоне при 100 пс зондирующим импульсом 12 (Фиг.8, hν1+hν2). Таким образом, фотопереключение бактериородопсина осуществлено за 3 пс.
Такой же результат наблюдается при фотопереключении бактериородопсина Halobacterium salinarum фемтосекундными импульсами, задержка между которыми 5 пс.
Описанный способ фотопереключения ретинальсодержащих белков лежит в основе функционирования заявляемого оптического логического элемента, предназначенного для использования в оптических устройствах передачи и обработки информации, обладающих быстродействием, превышающим быстродействие существующих аналогов.
Примером логического элемента может служить элемент, схематически изображенный на Фиг.9, осуществляющий логическую функцию в двоичной логике, которую обычно реализуют сочетанием простейших логических элементов "И", "НЕ". Заявляемая оптическая логическая ячейка осуществляет логическую функцию декремента, запрет импликации по второму импульсу, инверсию импликации от первого ко второму световому импульсу (в англоязычной литературе INH от inhibit). Выходной сигнал равен единице, только когда первый вход 1, а второй 0 (см. таблицу истинности на Фиг.10).
Оптический логический элемент содержит оптически связанные источник фемтосекундных импульсов 17, устройства формирования параметров световых пучков 3, 7, 11, рабочий элемент 14, а также устройство для анализа результатов 15.
Рабочий элемент 14 представляет собой емкость или пленку, изготовленную из светопропускающего материала произвольной формы, содержащую в качестве светочувствительного материала ретинальсодержащий белок. В качестве емкости может быть использована кварцевая, стеклянная или пластмассовая кювета произвольной формы. Емкость содержит светочувствительный материал, в качестве которого используют РСБ, выбранный из группы: зрительные пигменты животных, сенсорные родопсины и светозависимые ионные насосы микроорганизмов, а также другие аналоги ретинальсодержащих белков. РСБ в емкости может быть в виде экстракта в подходящем детергенте. В качестве детергента может быть использован н-нонил-β-D-гептилтиоглюкозид. РСБ может быть введен в пленку, изготовленную из прозрачного материала (полимер, стекло и т.п.).
Источником фемтосекундных импульсов может служить титан-сапфировый лазер. Параметры световых лучей задаются в неколлинеарных параметрических усилителях света 3, 7. Зондирующий 12 и опорный 13 импульсы, выходящие из источника 11, проходят через рабочий элемент 14. Световые импульсы 12, 13 могут попадать на следующую логическую ячейку или на детектор 15.
Оптический логический элемент работает следующим образом.
Входными сигналами логической ячейки служат лучи 4, 8, которые попадают на рабочий элемент 14. Изменение интенсивности зондирующего импульса 12 относительно опорного импульса 13 служит выходным сигналом.
Когда нет входных сигналов (hν1=0, hν2=0), то детектор регистрирует нулевой выходной сигнал (выход=0).
Когда на вход логической ячейки подается первый световой импульс (hν1=1, hν2=0), то детектор регистрирует высокий выходной сигнал (выход = 1).
Когда подаются первый импульс, а через время задержки второй импульс (hν1=1, hν2=1), то детектор регистрирует низкий сигнал (выход = 0).
Когда на вход логической ячейки подается второй импульс (hν1=0, hν2=1), то детектор регистрирует низкий выходной сигнал (выход = 0).
Заявляемая оптическая логическая ячейка функционирует с наибольшим быстродействием, когда в качестве РСБ используют зрительный пигмент родопсин при использовании условий фотопереключения, описанных в примере 1, при этом время фотопереключения составляет 200 фс.
Как показано на Фиг.11, первый входной сигнал, в качестве которого используют первый световой импульс (λ=500 нм, t=25 фс, E=70 нДж), логическая ячейка преобразует в высокий оптический сигнал, которому присвоено значение 1. Приход через 200 фс второго импульса (λ=620 нм, t=40 фс, E=450 нДж) снижает выходное значение зондирующего импульса ниже порогового. Уменьшение дифференциального сигнала поглощения зрительного пигмента родопсина в области 560-570 нм на 10% выбрано в качестве порогового значения (см. Фиг.11). Если на вход подают только второй световой импульс, то выходной сигнал остается нулевым, как и при отсутствии импульсов.
Если в качестве рабочей среды используют бактериородопсин, то параметры первого светового импульса составляют: λ=560 нм, t=20 фс, E=100 нДж, второго λ=680 нм, t=30 фс, E=560 нДж. Время задержки между импульсами 3 пс. В этом случае также происходит функционирование логической ячейки в различных режимах. Только при подаче на вход первого импульса на выходе получается единичное значение, в иных случаях - нулевое.
Детектирование предпочтительно осуществлять в тех спектральных областях, в которых изменение дифференциального спектра после одного и двух импульсов максимально. Использование низких значений энергии и длительности зондирующего импульса позволяют не влиять на работу элемента.
Изобретение относится к области фотоники и вычислительной техники и направлено на обеспечение возможности сверхбыстрого фотопереключения молекул ретинальсодержащих белков (РСБ) при комнатной температуре в субпикосекундной шкале времени, а также на создание оптического логического элемента, содержащего такой ретинальсодержащий белок, что обеспечивается за счет того, что способ фотопереключения ретинальсодержащего белка включает перевод белка при комнатной температуре из исходного состояния в промежуточное состояние путем воздействия на него первым световым импульсом с длиной волны, находящейся в диапазоне, соответствующем области поглощения исходного состояния, и последующее его возвращение в исходное состояние путем воздействия вторым световым импульсом с длиной волны, находящейся в диапазоне, соответствующем области, в которой происходит поглощение промежуточного состояния, но не происходит поглощение исходного состояния, при этом длительность указанных световых импульсов не превышает 50 фс, а промежуток времени между указанными световыми импульсами не превышает 6 пс. 2 н. и 8 з.п. ф-лы, 11 ил.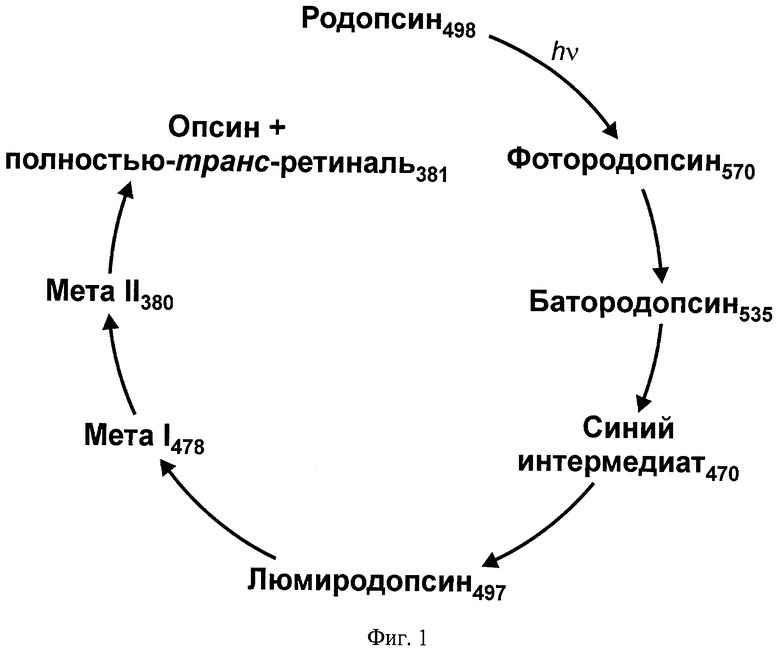
1. Способ фотопереключения ретинальсодержащего белка, включающий перевод белка при комнатной температуре из исходного состояния в промежуточное состояние путем воздействия на него первым световым импульсом с длиной волны, находящейся в диапазоне, соответствующем области поглощения исходного состояния, и последующее его возвращение в исходное состояние путем воздействия вторым световым импульсом с длиной волны, находящейся в диапазоне, соответствующем области, в которой происходит поглощение промежуточного состояния, но не происходит поглощение исходного состояния, при этом длительность указанных световых импульсов не превышает 50 фс, а промежуток времени между указанными световыми импульсами не превышает 6 пс.
2. Способ по п.1, отличающийся тем, что ретинальсодержащий белок представляет собой зрительный пигмент родопсин животных.
3. Способ по п.2, отличающийся тем, что длина волны первого импульса находится в диапазоне 470-530 нм, а длина волны второго импульса находится в диапазоне 600-650 нм.
4. Способ по п.1, отличающийся тем, что ретинальсодержащий белок представляет собой бактериородопсин.
5. Способ по п.4, отличающийся тем, что длина волны первого импульса находится в диапазоне 540-600 нм, а длина волны второго импульса находится в диапазоне 670-720 нм.
6. Оптический логический элемент, содержащий ретинальсодержащий белок, который подвергают фотопереключению из исходного состояния в промежуточное состояние и обратно, а указанное фотопереключение сопровождается соответствующими изменениями логического значения оптического логического элемента, отличающийся тем, что фотопереключение осуществляют при комнатной температуре под действием двух последовательных световых импульсов, причем длина волны первого светового импульса находится в диапазоне, соответствующем области поглощения исходного состояния, длина волны второго светового импульса находится в диапазоне, соответствующем области, в которой происходит поглощение промежуточного состояния, но не происходит поглощение исходного состояния, при этом длительность указанных световых импульсов не превышает 50 фс, а промежуток времени между указанными световыми импульсами не превышает 6 пс.
7. Оптический логический элемент по п.6, отличающийся тем, что ретинальсодержащий белок представляет собой зрительный пигмент родопсин животных.
8. Оптический логический элемент по п.7, отличающийся тем, что длина волны первого импульса находится в диапазоне 470-530 нм, а длина волны второго импульса находится в диапазоне 600-650 нм.
9. Оптический логический элемент по п.6, отличающийся тем, что ретинальсодержащий белок представляет собой бактериородопсин.
10. Оптический логический элемент по п.9, отличающийся тем, что длина волны первого импульса находится в диапазоне 540-600 нм, а длина волны второго импульса находится в диапазоне 670-720 нм.
| DELANEY J.K., SCHMIDT P.K., BRACK T.L., ATKINSON G.H | |||
| Промывально-чесальная машина для выделения волокон из хвои | 1920 |
|
SU590A1 |
| PHYS.CHEM., 2000, 104, P.10827-10834 | |||
| ОПТИЧЕСКИЙ ЛОГИЧЕСКИЙ ЭЛЕМЕНТ И ОПТИЧЕСКОЕ ЛОГИЧЕСКОЕ УСТРОЙСТВО | 1997 |
|
RU2172975C2 |
| WO 9316476 A1, 26.12.1995 | |||
| KR 20000016570 A, 25.03.2000. | |||

