Изобретение относится к мутантным канальным родопсинам, обладающим улучшенными свойствами, к конструкциям нуклеиновых кислот, кодирующим мутантные канальные родопсины, к экспрессирующим векторам, несущим конструкцию нуклеиновой кислоты, к клеткам, содержащим указанную конструкцию нуклеиновой кислоты или экспрессирующий вектор, и к их соответствующему применению.
Уровень техники
Светозависимый катионный канал внутреннего выпрямления канальный родопсин-2 (ChR2) стал предпочтительным инструментом направленной световой активации нейронов in vitro и in vivo1-4. Хотя для индуцируемой светом деполяризации можно использовать ChR2 дикого типа (WT), продолжаются исследования мутантов ChR2 с повышенной светочувствительностью для возможных будущих клинических применений (WO 03/084994 и 5-7). Большая эффективность позволила бы осуществить деполяризацию клеточных слоев, находящихся на расстоянии от применяемого источника света, несмотря на низкий оптический коэффициент пропускания, например, в тканях головного мозга. Повышение светочувствительности также позволило бы решить проблему возможных повреждений клеток при длительном облучении светом высокой интенсивности синего света, необходимых для полной активации ChR2 WT (1018-1019 фот с-1см-2 при 480 нм). Варианты с более высокой светочувствительностью также являются ключевыми для исследований, относящихся к восстановлению зрения8,9. На уровне белков более высокую эффективность света можно получать только путем повышения времени жизни в открытом состоянии и/или повышением единичной проводимости канала, т.к. светочувствительность как таковую можно улучшить лишь незначительно за счет природы ретиналя хромофоров ChR2. Ранее проведенные исследования показали, что мутации в положениях C128 и D156 спирали 3 и 4, соответственно, приводят к значительно замедленной кинетики каналов со временем жизни в открытом состоянии до 30 минут и более, обеспечивая в 500 раз выше или даже более высокую светочувствительность5 ' 6. Такие мутанты C128 и D156 можно выключать при различных периодах времени жизни в открытом состоянии красным светом. Несмотря на очень высокую светочувствительность, их медленно затухающая кинетика остается ограничивающим фактором их применимости.
Таким образом, существует необходимость в активируемых светом катионных каналах, обладающих более высокой светочувствительностью и более быстрой кинетикой ответной реакции.
Сущность изобретения
Как известно, потенциал внутренней поверхности мембраны клетки зависит от Ca++, и изменения субмембранных уровней внутриклеточного Ca++ приводят к деполяризации мембраны и к активации потенциалзависимых Na+-каналов в нейронах. Таким образом, авторы настоящего изобретения предположили, что светочувствительность нейрона можно опосредовано повышать, увеличивая потенциал внутренней поверхности их мембраны за счет притока Ca++. Неожиданно авторы изобретения обнаружили мутант ChR2 с повышенной проницаемостью для Ca++, далее обозначен как CatCh, т.е. переносящий кальций канальный родопсин. CatCh обладает в четыре раза более высокой проницаемостью для Ca++, в 70 раз более высокой светочувствительностью и более быстрой кинетикой ответной реакции при экспрессии в нейронах гиппокампа по сравнению с ChR2 WT. Показано, что повышенная светочувствительность и быстрая кинетика обусловлены относительно высоким светозависимым притоком Ca++, который повышает потенциал внутренней поверхности мембраны и активирует Ca++-активируемые калиевые каналы высокой проводимости (BK). Увеличение [Ca++]i повышает потенциал внутренней поверхности, способствуя активации потенциалзависимых Na+-каналов и опосредованно увеличивая светочувствительность. Следующая за стимуляцией светом реполяризация заметно ускорена активацией Ca++-зависимого BK канала. CatCh представляет собой пример нового принципа, на основании которого можно сконструировать светозависимые каналы для повышения светочувствительности нейрональной стимуляции. Его характеристики, такие как стимуляция с высокой точностью и быстрые потенциалы действия наряду с необходимостью низких интенсивностей света для активации, открывают возможность использования светозависимых каналов в клинической практике.
Таким образом, в первом аспекте изобретение относится к активируемому светом ионному каналу, где индуцируемый светом ионный канал содержит аминокислотную последовательность, которая обладает по меньшей мере на 70% гомологична аминокислотной последовательности, в положениях 1-309 SEQ ID NO:1 (CHOP-2), и которая содержит мутацию в положении, соответствующем L132 SEQ ID NO: 1.
В сходном втором аспекте изобретение также относится к канальному родопсину, содержащему активируемый светом ионный канал по первому аспекту и ретиналь или производное ретиналя. Кроме того, в третьем аспекте изобретение относится к конструкции нуклеиновой кислоты, содержащей нуклеотидную последовательность, кодирующую индуцируемый светом ионный канал по первому аспекту. В другом аспекте изобретение относится к экспрессирующему вектору, содержащему нуклеотидную последовательность, кодирующую активируемый светом ионный канал по первому аспекту или конструкцию нуклеиновой кислоты по третьему аспекту.
Кроме того, изобретение относится к клетке, содержащей канальный родопсин по второму аспекту, конструкцию нуклеиновой кислоты по третьему аспекту или экспрессирующий вектор по четвертому аспекту.
Кроме того, изобретение относится к применению активируемого светом ионного канала по первому аспекту, канального родопсина второго аспекта, конструкции нуклеиновой кислоты или экспрессирующего вектора по изобретению и клетки по изобретению в качестве лекарственного средства. В частности, изобретение относится к применению экспрессирующего вектора по изобретению в генотерапии.
Более конкретно, изобретение относится к применению активируемого светом ионного канала, канального родопсина, конструкции нуклеиновой кислоты, экспрессирующего вектора или клетки по изобретению при лечении слепоты или вызванного ослабления зрения.
В другом аспекте изобретение относится к применению активируемого светом ионного канала по первому аспекту, дополнительно содержащего треонин, серин или аланин в положении, соответствующем положению 128 SEQ ID NO: 1, и/или аланин в положении, соответствующем положению 156 SEQ ID NO: 1, при абляции злокачественных клеток.
В последнем аспекте изобретение относится к применению активируемого светом ионного канала по первому аспекту или канального родопсина по второму аспекту, или клетки по изобретению в высокопроизводительном скрининге.
Подробное описание предпочтительных вариантов осуществления
В первом аспекте изобретение относится к активируемому светом ионному каналу, где активируемый светом ионный канал содержит аминокислотную последовательность, которая по меньшей мере на 70% идентична аминокислотной последовательности положений 1-309 SEQ ID NO: 1 (CHOP-2), более предпочтительно, к аминокислотной последовательности положений 1-315 SEQ ID NO: 1, или к аминокислотной последовательности положений 1-737 SEQ ID NO: 1, и которая содержит мутацию в положении, соответствующем L132 в SEQ ID NO: 1.
CHOP2 дикого типа имеет следующую аминокислотную последовательность:
MDYGGALSAV GRELLFVTNP VVVNGSVLVP EDQCYCAGWI ESRGTNGAQT ASNVLQWLAA GFSILLLMFY AYQTWKSTCG WEEIYVCAIE MVKVILEFFF EFKNPSMLYL ATGHRVQWLR YAEWLLTCPV ILIHLSNLTG LSNDYSRRTM GLLVSDIGTI VWGATSAMAT GYVKVIFFCL GLCYGANTFF HAAKAYIEGY HTVPKGRCRQ VVTGMAWLFF VSWGMFPILF ILGPEGFGVL SVYGSTVGHT IIDLMSKNCW GLLGHYLRVL IHEHILIHGD IRKTTKLNIG GTEIEVETLV EDEAEAGAVN KGTGKYASRE SFLVMRDKMK EKGIDVRASL DNSKEVEQEQ AARAAMMMMN GNGMGMGMGM NGMNGMGGMN GMAGGAKPGL ELTPQLQPGR VILAVPDISM VDFFREQFAQ LSVTYELVPA LGADNTLALV TQAQNLGGVD FVLIHPEFLR DRSSTSILSR LRGAGQRVAA FGWAQLGPMR DLIESANLDG WLEGPSFGQG ILPAHIVALV AKMQQMRKMQ QMQQIGMMTG GMNGMGGGMG GGMNGMGGGN GMNNMGNGMG GGMGNGMGGN GMNGMGGGNG MNNMGGNGMA GNGMGGGMGG NGMGGSMNGM SSGVVANVTP SAAGGMGGMM NGGMAAPQSP GMNGGRLGTN PLFNAAPSPL SSQLGAEAGM GSMGGMGGMS GMGGMGGMGG MGGAGAATTQ AAGGNAEAEM LQNLMNEINR LKRELGE (SEQ ID NO: 1)
Активируемый светом ионный канал по изобретению представляет собой мембранный белок по меньшей мере с 5 трансмембранными спиралями, который способен связывать чувствительный к свету полиен. Трансмембранные белки с 6 или 7 трансмембранными спиралями являются предпочтительными. Однако трансмембранные белки с более чем 7 спиралями, например, 8, 9 или 10 трансмембранными спиралями также включены в изобретение. Кроме того, изобретение относится к трансмембранным белкам, которые в дополнение к трансмембранной части содержат C- и/или N-концевые последовательности, где C-концевые последовательности могут простираться внутрь ограниченного мембранной просвета, например, в цитоплазму клетки или внутрь липосомы, или могут также располагаться на внешней поверхности мембраны. То же самое необязательно касается содержащейся N-концевой последовательности, которая может аналогичным образом располагаться в просвете, а также на внешней поверхности мембраны. Для длины C- и/или N-концевых последовательностей по существу не установлены ограничения, однако предпочтительными являются активируемые светом ионные каналы с не погруженными в мембрану C-концевыми последовательностями от 1 до 1000 аминокислот, предпочтительно, от 1 до 500, особенно предпочтительно, от 5 до 50 аминокислот. Независимо от длины C-концевые последовательности, N-концевые локализованные последовательности, не погруженные в мембрану, предпочтительно, содержат от 1 до 500 аминокислот, особенно предпочтительно, от 5 до 50 аминокислот. Специалистам с данной области хорошо известна концепция трансмембранной спирали. В основном они представляют собой α-спиральные белковые структуры, которые, как правило, содержат от 20 до 25 аминокислот. Однако в зависимости от природы мембраны, которая может представлять собой природную мембрану, например, клеточную или плазматическую мембрану, или также синтетическую мембрану, трансмембранные сегменты также могут быть короче или длиннее. Например, трансмембранные сегменты в искусственных мембранах могут содержать до 30 аминокислот, но, с другой стороны, также могут содержать только несколько аминокислот, например, от 12 до 16.
В предпочтительном варианте осуществления активируемый светом ионный канал содержит аминокислотную последовательность, которая по меньшей мере на 70% идентична, предпочтительно, по меньшей мере на 75% идентична, более предпочтительно, по меньшей мере на 80% идентична, еще более предпочтительно, по меньшей мере на 85% идентична, например, по меньшей мере на 90% идентична, и наиболее предпочтительно, по меньшей мере на 95% идентична аминокислотной последовательности положений 1-309 SEQ ID NO: 1.
В другом предпочтительном варианте осуществления активируемый светом ионный канал содержит аминокислотную последовательность, которая по меньшей мере на 70% идентична, предпочтительно, по меньшей мере на 75% идентична, более предпочтительно, по меньшей мере на 80% идентична, еще более предпочтительно, по меньшей мере на 85% идентична, например, по меньшей мере на 90% идентична, и наиболее предпочтительно, по меньшей мере на 95% идентична аминокислотной последовательности положений 1-315 SEQ ID NO: 1.
Как правило, аминокислотная последовательность "по меньшей мере на x% идентична" другой аминокислотной последовательности или указанной выше SEQ ID NO: 1, если идентичность последовательности указанной последовательностью и последовательностью сравнения составляет по меньшей мере x%. Такое выравнивание можно проводить с использованием, например, общедоступных компьютерных программ поиска гомологии, таких как программа "BLAST", предоставляемая на главной странице NCBI по ссылке http://www.ncbi.nlm.nih.gov/blast/blast.cgi, с использованием установок по умолчанию, предоставленных по указанной выше ссылке. Дополнительные способы определения процента идентичности последовательности набора последовательностей нуклеиновой кислоты известны в данной области.
Примерами таких активируемых светом ионных каналов, содержащих аминокислотную последовательность, которая по меньшей мере на 70% идентична аминокислотной последовательности положений 1-309 или 1-315 SEQ ID NO: 1, являются CHOP1 из C. reinhardtii (gi: 15811379), CHOP2 (gi: 167650748) и CHOP1 (gi: 167650744) из Volvox carteri, или любой другой ортолог или аллельный вариант CHOP2 или CHOP 1.
Даже в более предпочтительном варианте осуществления активируемый светом ионный канал содержит, предпочтительно состоит из аминокислотной последовательности, показанной в положениях 1-309 SEQ ID NO: 1 (CHOP-2), за исключением мутации в положении L132.
В другом даже более предпочтительном варианте осуществления активируемый светом ионный канал содержит, предпочтительно состоит из аминокислотной последовательности, показанной в положениях 1-315 SEQ ID NO: 1 (CHOP-2), за исключением мутации в положении L132.
Мутация в положении L132, или в положении, соответствующем L132 в SEQ ID NO: 1, может представлять собой замену, добавление и/или делецию. Однако предпочтительно мутация представляет собой замену, более предпочтительно выбранную из L132C, L132S, L132E, L132D и L132T, наиболее предпочтительно, где замена представляет собой L132C. Даже несмотря на то, что экспериментальные данные ограничены L132C, предполагают, что замены L132S, L132E, L132D и L132T будут проявлять аналогичные свойства, т.к. все эти замены будут увеличивать полярность канала.
Кроме того, активируемый светом канал содержит дополнительные (полу)консервативные замены. Консервативные замены представляют собой такие, которые происходят в семействе аминокислот, которые связаны с их боковыми цепями и химическими свойствами. Примерами таких семейств являются аминокислоты с основными боковыми цепями, с кислотными боковыми цепями, с неполярными алифатическими боковыми цепями, с неполярными ароматическими боковыми цепями, с незаряженными полярными боковыми цепями, с маленькими боковыми цепями, с большими боковыми цепями и т.д. Конкретные полуконсервативные и консервативные замены представляют собой:
Кроме того, специалисту в данной области будет понятно, что не следует заменять глицины в необходимых стерических положениях, и что не следует вводить пролин в участки белка, которые содержат альфа-спиральную или бета-складчатую структуру.
В другом предпочтительном варианте осуществления активируемый светом канал содержит консенсусный мотив L(I)DxxxKxxW(F,Y). Приведенные в скобках аминокислоты в каждом случае могут заменять предшествующую аминокислоту. Такой консенсусный мотив представляет собой мотив, окружающий ретиналь-связывающую аминокислоту лизин.
Возможность активировать CatCh интенсивностью света природного происхождения при поддержании высокой временной точности, делает его уникальным кандидатом, в частности в попытках поиска способов генной терапии для восстановления зрения, а также для других биомедицинских применений. Вследствие его пониженных потребностей в свете, спайки CatCh могут быть вызваны даже возбуждением, далеким от его спектрального максимума 474 нм, например, зеленым светом (532 нм - см. фиг.4d). Возможным является функционирование на внешних границах спектра активации вследствие его пониженных потребностей в свете, и она способствует проникновению в ткани.
Таким образом, светочувствительность мутантного активируемого светом ионного канала по изобретению предпочтительно повышена более чем в 5 раз, предпочтительно, более чем в 10 раз, более предпочтительно, более чем в 20 раз, например, в 30 раз, еще более предпочтительно, более чем в 40 раз, например, в 50 раз, и наиболее предпочтительно, более чем в 60 раз, или еще более чем в 70 раз, по сравнению с CHOP-2 WT в нейронах гиппокампа. Кроме того, для мутантного активируемого светом ионного канала по изобретению показана частота стимуляции, которая повышена по меньшей мере в 1,5 раза, более предпочтительно, в 2 раза, или еще более предпочтительно, в 2,5 раза по сравнению с CHOP-2 WT, как определено электрофизиологической регистрацией целой клетки в нейронах гиппокампа. Как показано в примерах, для Chop2 WT выявлена частота стимуляции в нейронах гиппокампа приблизительно от 10 Гц приблизительно до 20 Гц, где при 20 Гц сигналы уже являются неточными. Кроме того, специалисту будет понятно, что характерная частота повторения спайков также зависит от типа клеток. Например, для слуховых клеток характерная частота повторения спайков составляет до 500 Гц. Кроме того, эксперименты проводили in vitro, т.е. при температуре окружающей среды. Однако специалист может допустить, что частота стимуляции даже будет выше у теплокровных животных, таких как млекопитающие, поскольку кинетика также зависит от температуры. Таким образом, в зависимости от типа клеток и температуры можно ожидать, что мутантный активируемый светом ионный канал по изобретению может также характеризоваться частотой стимуляции, которая повышена по меньшей мере в 5 раз, предпочтительно, по меньшей мере в 10 раз, такой как по меньшей мере в 20 раз, или по меньшей мере в 30 раз, или более предпочтительно, по меньшей мере в 40 раз, по меньшей мере в 50 раз, такой как по меньшей мере в 60 раз, или по меньшей мере в 70 раз, еще более предпочтительно, по меньшей мере в 80 раз, по меньшей мере в 90 раз, или по меньшей мере в 100 раз, наиболее предпочтительно, по меньшей мере в 125 раз, например, по меньшей мере в 150 раз, или по меньшей мере в 175 раз и, еще более предпочтительно, по меньшей мере в 200 раз по сравнению с CHOP-2 WT, что определено с помощью электрофизиологической регистрации целой клетки. Культура нейронов гиппокампа и электрофизиологическая регистрация сигналов от нейронов гиппокампа проиллюстрированы в приведенных ниже примерах.
В кратком изложении, гиппокамп выделяют у крыс Sprague-Dawley в первые сутки постнатального периода (Jackson Laboratory) и обрабатывают папаином (20 ед. мл-1) в течение 20 мин при 37ºC. Гиппокамп отмывают DMEM (Invitrogen/Gibco, с высоким содержанием глюкозы) с добавлением 10% эмбриональной телячьей сыворотки и измельчают в небольшом объеме этого раствора. ≈75000 клеток высевают на покрытые поли-D-лизином/ламинином стеклянные покровные стекла в 24-луночные планшеты. Через 3 часа среду для посева заменяют культуральной средой (Neurobasal A, содержащей 2% добавки B-27, 2 мМ Glutamax-I и 100 ед./мл пенициллина и 100 мкг/мл стрептомицина). Мутантный ChR2(L132C)-YFP и ChR2 (WT)-YFP трансфицируют через 5-10 суток после посева с использованием реагента липофектамин 2000 (Invitrogen). Альтернативно, можно добавлять 2-5×109 геномных копий/мл вируса (AAV2/7-CAG-ChR2(L132C)-2A-EGFP-WPRE-bGH) в каждую лунку через 4-9 суток после посева. Характерная конструкция конструкции аденоассоциированного вирусного вектора подробно описана в следующих ниже примерах. Экспрессия становится видимой на 5 сутки после трансдукции. Неполный транс-ретиналь добавляли в культуральную среду или регистрирующую среду для любого из экспериментов.
Для регистраций целой клетки в культивируемых нейронах гиппокампа пэтч-пипетки с сопротивлением 5-10 МОм наполняют 129 мМ глюконатом калия, 10 мМ HEPES, 10 мМ KCl, 4 мМ MgATP и 0,3 мМ Na3GTP, оттитрованными до pH 7,2. В качестве внеклеточного раствора применяют раствор Тироде (125 мМ NaCl, 2 мМ KCl, 2 мМ CaCl2, 1 мМ MgCl2, 30 мМ глюкозы и 25 мМ HEPES, оттитрованными до pH 7,4). Условно не содержащий Ca++ внеклеточный раствор содержит аналогичный раствор за исключением того, что он содержит 0 мМ Ca++ и 3 мМ Mg++. Регистрации проводят в присутствии ингибиторов возбуждающей синаптической передачи сигналов 1,2,3,4-тетрагидро-6-нитро-2,3-диоксо-бензо[f]хиноксалин-7-сульфонамида (NBQX, 10 мкМ, Sigma) и D(-)-2-амино-5-фосфонопентановой кислоты (AP-5, 50 мкМ, Sigma). Для регистраций способом фиксации потенциалов к внеклеточному раствору добавляют 1 мкМ тетродотоксина. Для ингибирования активности BK каналов добавляют 1 мМ TEA. Регистрации проводят на инвертированном микроскопе Zeiss Axiovert 25, оборудованном флуоресцентной лампой. Успешную экспрессию белка подтверждают EGFP- или YFP-опосредованной флуоресценцией. Входное сопротивление нейронов составляет 15-40 МОм, и наблюдают за его стабильностью на всем протяжении эксперимента. Электрофизиологические сигналы амплифицируют с использованием амплификатора Axopatch 200A (Axon Instruments, Union City, CA), фильтруют при 10 кГц, преобразовывают в цифровую форму устройством Axon Digidata 1600 (50 Гц) и регистрируют, и анализируют с использованием программного обеспечения pClamp9 (Axon Instruments). Фототоки вызывают с использованием световых импульсов различных длин твердотельного лазера с диодной накачкой (Pusch Opto Tech GmbH, λ1=473 нм, P1=100 мВт, λ2=532 нм, P2=50 мВт) или 10 нс вспышками лазера на красителях с эксимерной накачкой (кумарин 2, λ=450 нм). Конкретные интенсивности света представляют собой интенсивности на конце кварцевого оптического волокна диаметром 400 мкм (STE-F100/400-Y-VIS/NIR, Laser 2000, Wessling, Germany) на расстоянии ≈500 мкм от клетки. Токи, измеряемые от нейронов, экспрессирующих ChR2(L132C)-YFP и ChR2(L132C)-2A-EGFP, являются идентичными.
Кроме того, в другом предпочтительном варианте осуществления проводимость кальция мутантного активируемого светом ионного канала по изобретению повышена по меньшей мере в два раза, предпочтительно, по меньшей мере в три раза, более предпочтительно, по меньшей мере в четыре раза по сравнению с CHOP-2 WT, как определено визуализацией Fura-2 на клетках HEK293. Для определения проводимости кальция проводят нагрузку Fura-2 AM (5 мМ, Invitrogen) при комнатной температуре в течение от 30 мин до 1 часа. После нагрузки клетки восстанавливают в 140 мМ растворе NaCl без Ca++ (140 мМ NaCl, 7 мМ EGTA, 2 мМ MgCl2 и 10 мМ HEPES). Желтый флуоресцентный белок возбуждают 500 мсек воздействием света с использованием фильтра 460/40 нм (Visitron Systems, Puchheim, Germany) для оценки уровня экспрессии каждой клетки от его флуоресценции YFP. Затем раствор заменяют внеклеточным раствором с Ca++, который состоит из 90 мМ CaCl2, 7 мМ EGTA, 2 мМ MgCl2 и 10 мМ HEPES. После 15 мин в темноте светозависимые каналы стимулируют в течение 10 сек синим светом (460/40 нм). Fura-2 возбуждают при 340 нм (340/20) и 380 нм (380/20), и излучаемый свет (540/80 нм) детектируют камерой CCD (все фильтры от Visitron Systems, Puchheim, Germany).
Как указано выше, мутантный активируемый светом ионный канал может дополнительно содержать дополнительные мутации, предпочтительно, замены. В одном предпочтительном варианте осуществления активируемый светом ионный канал может дополнительно содержать по меньшей мере один из следующих аминокислотных остатков: аспарагиновую кислоту в положении, соответствующем положению 253 SEQ ID NO: 1, лизин в положении, соответствующем положению 257 SEQ ID NO: 1, триптофан в положении, соответствующем положению 260 SEQ ID NO: 1, глутаминовую кислоту в положении, соответствующем положению 123 SEQ ID NO: 1, гистидин или аргинин, предпочтительно аргинин, в положении, соответствующем положению 134 SEQ ID NO: 1, треонин, серин или аланин в положении, соответствующем положению 128 SEQ ID NO: 1, и/или аланин в положении, соответствующем положению 156 SEQ ID NO: 1. Таким образом, мутантный активируемый светом ионный канал может содержать одно из следующих сочетаний аминокислотных остатков в указанных положениях, соответствующих SEQ ID NO: 1:
Cys 132 + Asp 253; Cys 132 + Lys 257; Cys 132 + Trp 260; Cys 132 + Glu 123; Cys 132 + His 134; Cys 132 + Arg 134; Cys 132 + Thr 128; Cys 132 + Ser 128; Cys 132 + Ala 128; Cys 132 + Ala 156;
Cys 132 + Asp 253 + Lys 257; Cys 132 + Asp 253 + Trp 260; Cys 132 + Asp 253 + Glu 123; Cys 132 + Asp 253 + His 134; Cys 132 + Asp 253 + Arg 134; Cys 132 + Asp 253 + Thr 128; Cys 132 + Asp 253 + Ser 128; Cys 132 + Asp 253 + Ala 128; Cys 132 + Asp 253 + Ala 156;
Cys 132 + Lys 257 + Trp 260; Cys 132 + Lys 257 + Glu 123; Cys 132 + Lys 257 + His 134; Cys 132 + Lys 257 + Arg 134; Cys 132 + Lys 257 + Thr 128; Cys 132 + Lys 257 + Ser 128; Cys 132 + Lys 257 + Ala 128; Cys 132 + Lys 257 + Ala 156;
Cys 132 + Trp 260 + Glu 123; Cys 132 + Trp 260 + His 134; Cys 132 + Trp 260 + Arg 134; Cys 132 + Trp 260 + Thr 128; Cys 132 + Trp 260 + Ser 128; Cys 132 + Trp 260 + Ala 128; Cys 132 + Trp 260 + Ala 156;
Cys 132 + Glu 123 + His 134; Cys 132 + Glu 123 + His 134; Cys 132 + Glu 123 + Arg 134; Cys 132 + Glu 123 + Thr 128; Cys 132 + Glu 123 + Ser 128; Cys 132 + Glu 123 + Ala 128; Cys 132 + Glu 123 + Ala 156;
Cys 132 + His 134 + Thr 128; Cys 132 + His 134 + Ser 128; Cys 132 + His 134 + Ala 128; Cys 132 + His 134 + Ala 156;
Cys 132 + Arg 134 + Thr 128; Cys 132 + Arg 134 + Ser 128; Cys 132 + Arg 134 + Ala 128; Cys 132 + Arg 134 + Ala 156;
Cys 132 + Thr 128 + Ala 156; Cys 132 + Ser 128 + Ala 156; Cys 132 + Ala 128 + Ala 156;
Cys 132 + Asp 253 + Lys 257 + Trp 260; Cys 132 + Asp 253 + Lys 257 + Glu 123; Cys 132 + Asp 253 + Lys 257 + His 134; Cys 132 + Asp 253 + Lys 257 + Arg 134; Cys 132 + Asp 253 + Lys 257 + Thr 128; Cys 132 + Asp 253 + Lys 257 + Ser 128; Cys 132 + Asp 253 + Lys 257 + Ala 128; Cys 132 + Asp 253 + Lys 257 + Ala 156;
Cys 132 + Lys 157 + Trp 260 + Glu 123; Cys 132 + Lys 157 + Trp 260 + His 134; Cys 132 + Lys 157 + Trp 260 + Arg 134; Cys 132 + Lys 157 + Trp 260 + Thr 128; Cys 132 + Lys 157 + Trp 260 + Ser 128; Cys 132 + Lys 157 + Trp 260 + Ala 128; Cys 132 + Lys 157 + Trp 260 + Ala 156;
Cys 132 + Trp 260 + Glu 123 + His 134; Cys 132 + Trp 260 + Glu 123 +Arg 134; Cys 132 + Trp 260 + Glu 123 + Thr 128; Cys 132 + Trp 260 + Glu 123 + Ser 128; Cys 132 + Trp 260 + Glu 123 + Ala 128; Cys 132 + Trp 260 + Glu 123 + Ala 156;
Cys 132 + Glu 123 + His 134 + Thr 128; Cys 132 + Glu 123 + His 134 + Ser 128; Cys 132 + Glu 123 + His 134 + Ala 128; Cys 132 + Glu 123 + His 134 + Ala 156;
Cys 132 + Glu 123 + Arg 134 + Thr 128; Cys 132 + Glu 123 + Arg 134 + Ser 128; Cys 132 + Glu 123 + Arg 134 + Ala 128; Cys 132 + Glu 123 + Arg 134 + Ala 156;
Cys 132 + His 134 + Thr 128 + Ala 156; Cys 132 + His 134 + Ser 128 + Ala 156; Cys 132 + His 134 + Ala 128 + Ala 156;
Cys 132 + Arg 134 + Thr 128 + Ala 156; Cys 132 + Arg 134 + Ser 128 + Ala 156; Cys 132 + Arg 134 + Ala 128 + Ala 156;
Cys 132 + Asp 253 + Lys 257 + Trp 260 + Glu 123; Cys 132 + Asp 253 + Lys 257 + Trp 260 + His 134; Cys 132 + Asp 253 + Lys 257 + Trp 260 + Arg 134; Cys 132 + Asp 253 + Lys 257 + Trp 260 + Thr 128; Cys 132 + Asp 253 + Lys 257 + Trp 260 + Ser 128; Cys 132 + Asp 253 + Lys 257 + Trp 260 + Ala 128; Cys 132 + Asp 253 + Lys 257 + Trp 260 + Ala 156;
Cys 132 + Lys 257 + Trp 260 + Glu 123 + His 134; Cys 132 + Lys 257 + Trp 260 + Glu 123 + Arg 134; Cys 132 + Lys 257 + Trp 260 + Glu 123 + Thr 128; Cys 132 + Lys 257 + Trp 260 + Glu 123 + Ser 128; Cys 132 + Lys 257 + Trp 260 + Glu 123 + Ala 128; Cys 132 + Lys 257 + Trp 260 + Glu 123 + Ala 156;
Cys 132 + Trp 260 + Glu 123 + His 134 + Thr 128; Cys 132 + Trp 260 + Glu 123 + His 134 + Ser 128; Cys 132 + Trp 260 + Glu 123 + His 134 + Ala 128; Cys 132 + Trp 260 + Glu 123 + His 134 + Ala 156;
Cys 132 + Trp 260 + Glu 123 + Arg 134 + Thr 128; Cys 132 + Trp 260 + Glu 123 + Arg 134 + Ser 128; Cys 132 + Trp 260 + Glu 123 + Arg 134 + Ala 128; Cys 132 + Trp 260 + Glu 123 + Arg 134 + Ala 156;
Cys 132 + Glu 123 + Arg 134 + Thr 128 + Ala 156; Cys 132 + Glu 123 + Arg 134 + Ser 128 + Ala 156; Cys 132 + Glu 123 + Arg 134 + Ala 128 + Ala 156;
Cys 132 + Glu 123 + His 134 + Thr 128 + Ala 156; Cys 132 + Glu 123 + His 134 + Ser 128 + Ala 156; Cys 132 + Glu 123 + His 134 + Ala 128 + Ala 156;
Cys 132 + Asp 253 + Lys 257 + Trp 260 + Glu 123 + His 134; Cys 132 + Asp 253 + Lys 257 + Trp 260 + Glu 123 + Arg 134; Cys 132 + Asp 253 + Lys 257 + Trp 260 + Glu 123 + Thr 128; Cys 132 + Asp 253 + Lys 257 + Trp 260 + Glu 123 + Ser 128; Cys 132 + Asp 253 + Lys 257 + Trp 260 + Glu 123 + Ala 128; Cys 132 + Asp 253 + Lys 257 + Trp 260 + Glu 123 + Ala 156;
Cys 132 + Lys 257 + Trp 260 + Glu 123 + His 134 + Thr 128; Cys 132 + Lys 257 + Trp 260 + Glu 123 + His 134 + Ser 128; Cys 132 + Lys 257 + Trp 260 + Glu 123 + His 134 + Ala 128; Cys 132 + Lys 257 + Trp 260 + Glu 123 + His 134 + Ala 156;
Cys 132 + Lys 257 + Trp 260 + Glu 123 + Arg 134 + Thr 128; Cys 132 + Lys 257 + Trp 260 + Glu 123 + Arg 134 + Ser 128; Cys 132 + Lys 257 + Trp 260 + Glu 123 + Arg 134 + Ala 128; Cys 132 + Lys 257 + Trp 260 + Glu 123 + Arg 134 + Ala 156;
Cys 132 + Trp 260 + Glu 123 + Arg 134 + Thr 128 + Ala 156; Cys 132 + Trp 260 + Glu 123 + Arg 134 + Ser 128 + Ala 156; Cys 132 + Trp 260 + Glu 123 + Arg 134 + Ala 128 + Ala 156;
Cys 132 + Trp 260 + Glu 123 + His 134 + Thr 128 + Ala 156; Cys 132 + Trp 260 + Glu 123 + His 134 + Ser 128 + Ala 156; Cys 132 + Trp 260 + Glu 123 + His 134 + Ala 128 + Ala 156;
Cys 132 + Asp 253 + Lys 257 + Trp 260 + Glu 123 + His 134 + Thr 128; Cys 132 + Asp 253 + Lys 257 + Trp 260 + Glu 123 + His 134 + Ser 128; Cys 132 + Asp 253 + Lys 257 + Trp 260 + Glu 123 + His 134 + Ala 128; Cys 132 + Asp 253 + Lys 257 + Trp 260 + Glu 123 + His 134 + Ala 156;
Cys 132 + Asp 253 + Lys 257 + Trp 260 + Glu 123 + Arg 134 + Thr 128; Cys 132 + Asp 253 + Lys 257 + Trp 260 + Glu 123 + Arg 134 + Ser 128; Cys 132 + Asp 253 + Lys 257 + Trp 260 + Glu 123 + Arg 134 + Ala 128; Cys 132 + Asp 253 + Lys 257 + Trp 260 + Glu 123 + Arg 134 + Ala 156;
Cys 132 + Lys 257 + Trp 260 + Glu 123 + His 134 + Thr 128 + Ala 156; Cys 132 + Lys 257 + Trp 260 + Glu 123 + His 134 + Ser 128 + Ala 156; Cys 132 + Lys 257 + Trp 260 + Glu 123 + His 134 + Ala 128 + Ala 156;
Cys 132 + Lys 257 + Trp 260 + Glu 123 + Arg 134 + Thr 128 + Ala 156; Cys 132 + Lys 257 + Trp 260 + Glu 123 + Arg 134 + Ser 128 + Ala 156; Cys 132 + Lys 257 + Trp 260 + Glu 123 + Arg 134 + Ala 128 + Ala 156;
Cys 132 + Asp 253 + Lys 257 + Trp 260 + Glu 123 + His 134 + Thr 128 + Ala 156; Cys 132 + Asp 253 + Lys 257 + Trp 260 + Glu 123 + His 134 + Ser 128 + Ala 156; Cys 132 + Asp 253 + Lys 257 + Trp 260 + Glu 123 + His 134 + Ala 128 + Ala 156;
Cys 132 + Asp 253 + Lys 257 + Trp 260 + Glu 123 + Arg 134 + Thr 128 + Ala 156; Cys 132 + Asp 253 + Lys 257 + Trp 260 + Glu 123 + Arg 134 + Ser 128 + Ala 156; Cys 132 + Asp 253 + Lys 257 + Trp 260 + Glu 123 + Arg 134 + Ala 128 + Ala 156.
Однако в указанном выше списке Cys 132 также может быть замещен на Ser 132, Glu 132, Asp 132 или Thr 132.
В основном ретиналь или производное ретиналя, необходимое для функционирования активируемого светом ионного канала, продуцируется клеткой для трансфекции с указанным выше ионным каналом. В зависимости от своей конформации ретиналь может быть полностью транс-ретиналем, 11-цис-ретиналем, 13-цис-ретиналем или 9-цис-ретиналем. Однако также предполагают, что мутантный активируемый светом ионный канал по изобретению можно встраивать в везикулы, липосомы или другие искусственные клеточные мембраны. Таким образом, во втором аспекте изобретение относится к канальному родопсину, содержащему активируемый светом ионный канал по первую аспекту и ретиналь или производное ретиналя. Предпочтительно производное ретиналя выбрано из группу, состоящей из 3,4-дегидроретиналя, 13-этилретиналя, 9-диметил-ретиналя, 3-гидроксиретиналя, 4-гидроксиретиналя, нафтилретиналя, 3,7,11-триметил-додека-2,4,6,8,10-пентаеналя, 3,7-диметил-дека-2,4,6,8-тетраеналя, 3,7-диметил-окта-2,4,6-триеналя и ретиналей с заблокированным вращением в положении 6-7, ретиналей с заблокированным вращением в положении 8-9 и ретиналей с заблокированным вращением в положении 10-11. Кроме того, предпочтительные варианты осуществления по первому аспекту соответствуют предпочтительным вариантам осуществления по второму аспекту.
В третьем аспекте изобретение также относится к конструкции нуклеиновой кислоты, содержащей нуклеотидную последовательность, кодирующую активируемый светом ионный канал по первому аспекту. Для обеспечения оптимальной экспрессии кодирующую ДНК также можно соответствующим образом модифицировать, например, добавляя подходящие регуляторные последовательности и/или целевые последовательности и/или подбирая кодирующие ДНК последовательности к предпочтительной частоте использования кодона выбранного хозяина. Мишеневая последовательность может кодировать C-концевое удлинение, направляющее активируемый светом ионный канал к конкретному сайту или компартменту в клетке, такому как синапс или постсинаптический рецептор, к аксональному холмику или эндоплазматическому ретикулуму. Нуклеиновую кислоту можно комбинировать с дополнительными элементами, например, промотором и сигналом начала и остановки транскрипции, и сигналом начала и остановки трансляции, и сигналом полиаденилирования для обеспечения экспрессии последовательности белка по изобретению. Промотор может представлять собой индуцируемый или конститутивный, общий или клеткоспецифический промотор. Примером клеткоспецифического промотора является специфический для биполярных клеток промотор mGlu6. Выбор промоторов, векторов и других элементов является предметом общепринятого дизайна среднего уровня компетентности в данной области. Многие такие элементы описаны в литературе и являются доступными от коммерческих поставщиков.
Таким образом, в четвертом аспекте изобретение относится к экспрессирующему вектору, содержащему нуклеотидную последовательность, кодирующую активируемый светом ионный канал по первому аспекту или конструкцию нуклеиновой кислоты по третьему аспекту. В предпочтительном варианте осуществления вектор является подходящим для генотерапии, в частности, где вектор является подходящим для опосредованного вирусом переноса генов. Термин "подходящий для опосредованного вирусом переноса генов" в настоящем описании означает, что указанный вектор можно упаковывать в вирус и таким образом доставлять в представляющий интерес участок или клетки. Примеры подходящих для генотерапии вирусов представляют собой ретровирусы, аденовирусы, аденоассоциированные вирусы, лентивирусы, поксвирусы, альфавирусы, вирус бешенства, вирус леса Семлики и вирусы герпеса. Эти вирусы различаются по способности переноса генов в клетки, которые они распознают и которые способны инфицировать, и возможности изменения ДНК клетки, постоянно или временно. Однако генотерапия также предусматривает не основанные на применении вирусов способы, такие как применение "голой" ДНК, липоплексов и полиплексов и дендримеров.
Как описано выше, получаемую последовательность нуклеиновой кислоты можно вводить в клетки, например, с использованием вируса в качестве носителя или посредством трансфекции, включая, например, химические трансфектанты (такие как липофектамин, Fugene и т.д.), электропорацию, кальций-фосфатную копреципитацию и прямую диффузию ДНК. Способ трансфекции клетки подробно описан в примерах, и его можно адаптировать к соответствующей клетке-реципиенту. Трансфекцией ДНК получают стабильные клетки или клеточные линии, если трансфицированная ДНК интегрирована в геном, или нестабильные (транзиентные) клетки или клеточные линии, где трансфицированная ДНК существует в экстрахромосомной форме. Кроме того, стабильные клеточные линии можно получать с использованием эписомальной реплицирующейся плазмиды, что означает, что наследование экстрахромосомной плазмиды находится под контролем контролирующих элементов, которые интегрированы в клеточный геном. В основном, выбор подходящего вектора или плазмиды зависит от клетки-хозяина.
Таким образом, в пятом аспекте изобретение относится к клетке, содержащей канальный родопсин по второму аспекту, конструкцию нуклеиновой кислоты по третьему аспекту или экспрессирующий вектор по четвертому аспекту.
Как описано ниже, одно из применений мутантного активируемого светом ионного канала по настоящему изобретению представляет собой лечение лишенных зрения индивидуумов, таких как люди или животные. Существует ряд заболеваний, при которых природные зрительные клетки больше не функционируют, но все нервные соединения способны продолжать функционировать. В настоящее время в различных исследовательских центрах предпринимают попытки имплантировать тонкие слои с искусственными керамическими фотоэлементами на сетчатку. Такие фотоэлементы предназначены для деполяризации вторичных все еще интактных клеток сетчатки и, таким образом, для инициирования нервного импульса (бионический глаз). Целенаправленная экспрессия активируемых светом ионных каналов по изобретению в таких ганглионарных клетках, амакриновых клетках или биполярных клетках была бы более чем элегантным решением и обеспечивала бы лучшую трехмерную разрешающую способность зрения.
Встраивание мутантного активируемого светом ионного канала в мембрану клеток, которые в природе не экспрессируют соответствующие каналы, например, можно просто проводить с использованием известных способов технологии рекомбинантной ДНК, где кодирующую ДНК для такого ионного канала сначала встраивают в подходящий экспрессирующий вектор, например, плазмиду, космиду или вирус, а затем трансформируют клетки-мишени такими элементами, и белок экспрессируется в хозяине. Затем клетки обрабатывают подходящим образом, например, ретиналем, для обеспечения связывания основаниями Шиффа белка и ретиналя.
В предпочтительном варианте осуществления это происходит в различных дрожжах, таких как Saccharomyces cerevisiae, Schizosaccharomyces pombe или Pichia pastoris, как уже успешно проведено для родопсинов, таких как бактериородопсин и/или бычий родопсин.
Экспрессию также можно проводить в определенных клеточных системах млекопитающих или клеточных системах насекомых. Таким образом, в предпочтительном варианте осуществления клетка представляет собой клетку млекопитающего или клетку насекомого. Экспрессию также проводят эписомальными векторами в виде транзиторной экспрессии предпочтительно в меланомных клетках (например, клеточной линии BLM), клетках COS (полученных инфицированием клеток "почек CV1 африканской зеленой мартышки") или клетках HEK ("эмбриональных клетках почек человека", например, клетках HEK293), или клетках BHK ("клетках почек детенышей хомячка"), или в виде стабильно экспрессии (интеграцией в геном) в клетках CHO ("клетках яичника китайского хомяка"), миеломных клетках или клетках MDCK ("клетках почки собаки Madine-Darby") или в клетках Sf9 насекомого, инфицированных бакуловирусами. Таким образом, в более предпочтительном варианте осуществления клетка млекопитающего представляет собой клетку COS, клетку BHK, клетку HEK293, клетку CHO, миеломную клетку или клетку MDCK.
В отношении восстановления зрения в наиболее предпочтительном варианте осуществления клетка млекопитающего представляет собой фоторецепторную клетку, палочковидную клетку сетчатки, колбочковидную клетку сетчатки, ганглиозную клетку сетчатки, биполярный нейрон, ганглиозную клетку, псевдоуниполярный нейрон, мультиполярный нейрон, пирамидальный нейрон, клетку Пуркинье или гранулярную клетку.
Нейрон представляет собой электрически возбудимую клетку, которая обрабатывает и передает информацию посредством электрической и химической передачи сигналов, где химическая передача сигналов происходит посредством синапсов, специализированных соединений с другими клетками. Существует ряд специализированных типов нейронов, таких как сенсорные нейроны, отвечающие на прикосновение, звук, свет и многочисленные другие стимулы, воздействующие на клетки сенсорных органов, моторные нейроны, получающие сигналы от головного мозга и спинного мозга и вызывающие мышечные сокращения и воздействующие на железы, и вставочные нейроны, соединяющие нейроны с другими нейронами в одной и той же области головного мозга или спинного мозга. Как правило, нейрон обладает телом, дендритами и аксоном. Дендриты представляют собой филаменты, которые возникают из тела клетки, часто протягивающиеся на сотни микрон и разветвляющиеся множество раз. Аксон представляет собой специальный клеточный филамент, который возникает из тела клетки на участке, называемом аксональным холмиком. Часто из тела клетки нейрона возникают множественные дендриты, но никогда более одного аксона, хотя аксон может разветвляться сотни раз до того, как он закончится. В большинстве синапсов сигналы посылаются от аксона одного нейрона к дендриту другого. Однако из этих правил существует много исключений: нейроны, у которых отсутствуют дендриты, нейроны, не имеющие аксона, синапсы, соединяющие аксон с другим аксоном или дендрит с другим дендритом и т.д. Большинство нейронов можно дополнительно анатомически охарактеризовать как униполярные или псевдополярные (дендрит и аксон выходят из одного и того же отростка), биполярные (аксон и единственные дендрит на противоположных концах тела), мультиполярные (содержащие более двух дендритов, и их можно дополнительно классифицировать как (i) нейроны Гольджи I с длинными выступающими отростками аксона, такие как пирамидальные клетки, клетки Пуркинье и клетки переднего рога спинного мозга, и (ii) Гольджи II: нейроны, чей отросток аксона проецирован локально, например, гранулярные клетки).
Фоторецепторная клетка представляет собой специализированный нейрон, обнаруженный в сетчатке, который способен к фотопреобразованию. Два классических фоторецептора представляют собой палочки и колбочки, где каждый вклад информации используется зрительной системой. Ганглиозная клетка сетчатки представляет собой тип нейрона, локализованного вблизи внутренней поверхности сетчатки глаза. Эти клетки имеют дендриты и длинные аксоны, проецирующиеся в протектум (средний мозг), супрахиазмальное ядро в гипоталамусе и боковое коленчатое тело (таламус). Небольшой процент способствует незначительно или вообще не способствует зрению, но само по себе они являются светочувствительными. Их аксоны образуют ретино-гипоталамический тракт и вносят вклад в циркадные ритмы и зрачковый световой рефлекс, изменение размера зрачка. Они получают зрительную информацию от фоторецепторов посредством двух типов вставочных нейронов: биполярных клеток и амакриновых клеток. Амакриновые клетки представляют собой вставочные нейроны в сетчатке и ответственны за 70% поступающей информации к ганглиозным клеткам сетчатки. Биполярные клетки, которые ответственны за другие 30% поступающей информации к ганглиям сетчатки, регулируются амакриновыми клетками. Как часть сетчатки биполярная клетка находится между фоторецепторами (клетками палочек и клетками колбочек) и ганглиозными клетки. Они действуют прямо или опосредованно для передачи сигналов от фоторецепторов к ганглиозным клеткам.
Клетку можно выделять (и генетически модифицировать), поддерживать и культивировать при соответствующей температуре и смеси газов (как правило, 37ºC, 5% CO2), необязательно в клеточном инкубаторе, как известно специалисту и как проиллюстрировано для определенных клеточных линий или типов клеток в примерах. Условия культивирования могут отличаться для каждого типа клеток, и изменение условий для конкретного типа клеток может привести к различным фенотипам. Независимо от температуры и смеси газов чаще всего изменяющимся фактором в системах клеточных культур является среда для выращивания. Рецепт среды для выращивания может изменяться по значению pH, концентрации глюкозы, фактора роста и наличию других питательных компонентов наряду с другими. Среды для выращивания являются коммерчески доступными, или их можно составлять согласно композициям, которые доступны от American Tissue Culture Collection (ATCC). Факторы роста, используемые для дополнения сред, часто получают из крови животных, такой как телячья сыворотка. Дополнительно, к средам для выращивания можно добавлять антибиотики. К числу общепринятых обработок, проводимых на культивируемых клетках, относятся замены среды и пассирование клеток.
Существуют дополнительные потенциальные области применения CatCh. Поскольку Ca++ является важным внутриклеточным регулятором, CatCh открывает путь к оптическому воздействию на точно регулируемый гомеостаз Ca++ клетки, модулируя его состояние и активность. В фундаментальных исследованиях CatCh можно использовать для оптической регуляции Ca++-зависимого экзоцитоза в качестве альтернативы содержащемуся в клетке Ca++26 (например, выделение трансмиттера в синапсе), для оптической активации последующих внутриклеточных процессов посредством кальций-активируемых киназ и фосфатаз или индукции апоптоза посредством направления CatCh во внутриклеточные компартменты, такие как комплекс Гольджи или эндоплазматический ретикулум.
Таким образом, в дополнительном аспекте изобретение относится к применению активируемого светом ионного канала по первому аспекту или канального родопсина по второму аспекту, или конструкции нуклеиновой кислоты по третьему аспекту, или клетки по изобретению в качестве лекарственного средства. В частности, экспрессирующий вектор по изобретению можно использовать в генотерапии. Более конкретно, активируемый светом ионный канал по первому аспекту, канальный родопсин по второму аспекту, конструкцию нуклеиновой кислоты по третьему аспекту или клетку по изобретению можно применять при лечении слепоты или вызванного ослабления зрения. Однако вследствие его быстрого спайка активности до 300 Гц и ускоренной реполяризации также предусматривают использование активируемого светом ионного канала в восстановлении слуха или при лечении глухоты.
Дополнительно мутантный активируемый светом ионный канал по первому аспекту может содержать дополнительные замены (так называемые SFO или медленные мутанты, см. таблицу 1), которые приводят к постоянному активируемому светом притоку кальция, который в свою очередь приводит к гибели клеток. Таким образом, предусмотрено использование активируемого светом ионного канала по изобретению, дополнительно содержащего треонин, серин или аланин в положении, соответствующем положению 128 SEQ ID NO: 1, и/или аланин в положении, соответствующем положению 156 SEQ ID NO: 1, в абляции злокачественных клеток. Например, экспрессирующий вектор по изобретению можно направлять опосредованной вирусом трансфекцией генов посредством маркера клеточной поверхности злокачественной клетки к злокачественным клеткам. Кроме того, следует отметить, что, в частности, ретровирусы предпочтительно интегрировать в быстро делящиеся клетки, такие как злокачественные клетки. Как следствие, активируемый светом ионный канал по настоящему изобретению преимущественно экспрессируется и встроен в клеточную мембрану злокачественных клеток. После стимуляции светом эти ионные каналы откроются и индуцируют постоянный приток кальция, таким образом, приводя к гибели злокачественной клетки. Такое применение является преимуществом, в частности при абляции злокачественных клеток, которых естественным образом подвергают действию света, таких как злокачественные клетки меланомы. Таким образом, в предпочтительном варианте осуществления злокачественная опухоль представляет собой меланомный рак.
В последнем аспекте изобретение относится к использованию индуцируемого светом ионного канала по первому аспекту, или канального родопсина по второму аспекту, или клетки по изобретению в высокопроизводительном скрининге. Высокопроизводительный скрининг (HTS) представляет собой способ научного исследования, в частности, применяемый при поиске новых лекарственных средств и относящийся к области биологии и химии. HTS позволяет исследователю проводить миллионы биохимических, генетических или фармакологических тестов за короткий период времени, часто посредством сочетания современной робототехники, обработки данных и управляющего программного обеспечения, погрузочно-разгрузочного устройства для жидкостей и чувствительных детекторов. Этим способом можно быстро определять активные соединения, которые модулируют конкретный бимолекулярный путь, в частности вещество, модифицирующее ионный канал, такой как индуцируемый светом ионный канал по изобретению, Ca++-активируемый калиевый канал или BK канал. Например, в клетке-хозяине может совместно экспрессироваться Ca++-активируемый калиевый канал и активируемый светом ионный канал. После стимуляции светом активируемый светом ионный канал откроется, и внутриклеточная концентрация Ca++ увеличится, таким образом, активируя калиевый канал. Таким образом, произойдет изменение мембранного потенциала, которое можно наблюдать посредством чувствительных к потенциалу красителей, таких как RH 421 (N-(4-сульфобутил)-4-(4-(4-(дипентиламино)фенил)бутадиенил)пиридиний, внутренняя соль). Таким образом, такой HTS может включать следующие этапы: (i) контактирование клетки, экспрессирующей Ca++-активируемый (калиевый) канал и активируемый светом ионный канал по изобретению с соединением-кандидатом, направленным против Ca++-активируемого канала, (ii) применение светового стимула для активации активируемого светом канала, (iii) определение изменения мембранного потенциала (смешанный сигнал) и (iv) сравнение сигнала, определяемого на этапе (iii), с сигналом, определяемом в клетке, экспрессирующей только активируемый светом ионный канал по изобретению, подвергнутой этапу (ii) (единичный сигнал). Уменьшение изменения мембранного потенциала указывало бы на потенциальный модулятор Ca++-активируемого (калиевого) канала. Предполагают, что таким подходом можно получать соотношение сигнала к шуму приблизительно 5:1, которое является достаточно улучшенным по сравнению с прямыми измерениями, проводимыми на клетке, экспрессирующей только Ca++-активируемый канал. Вследствие улучшенного соотношения сигнала к шуму указанный способ, в частности с применением активируемого светом ионного канала, может являться, в частности, подходящим для HTS.
По существу в HTS используют подход сбора большого количества экспериментальных данных о действии большого количества веществ на конкретную мишень за относительно короткий период времени. Скрининг в этом контексте представляет собой более крупный эксперимент с одной целью (как правило, проверки научной гипотезы), к которой затем можно применить все эти данные. Для HTS клетки по изобретению можно рассевать в культуральный планшет, такой как многолуночный планшет, например, 96-луночный планшет. Затем клетку в планшете подвергают контактированию с тестируемым веществом в течение периода времени, достаточного для взаимодействия с целевым ионным каналом. Тестируемое вещество может быть различным от лунки к лунке во всем планшете. После окончания времени инкубацию измерения проводят во всех лунках планшета, вручную или автоматическим устройством и необязательно в сравнении с измерениями клетки, которую не подвергали контактированию с тестируемым веществом. Проводимые вручную измерения могут быть необходимыми, когда исследователь использует пэтч-кламп, определяя эффекты, которые еще определяют автоматизированным образом. В других случаях специализированным устройством автоматического анализа можно анализировать ряд экспериментов в лунках (таких как анализ света определенной частоты или высокопроизводительное измерение пэтч-кламп). В этом случае устройство выводит результат каждого эксперимента, например, таблицу числовых значений, где каждое число соответствует значению, полученному от одной лунки. В зависимости от результатов такого первого анализа исследователь может проводить последующие анализы в том же скрининге с использованием веществ, аналогичных тем, которые определили как активные (например, изменяющие уровень внутриклеточных циклических нуклеотидов), в новых аналитических планшетах, а затем повторно проводить эксперимент для сбора дополнительных данных, оптимизации структуры химического соединения для улучшения действия соединения на клетку. Автоматизация является важным элементом применимости HTS. Специализированный робот часто ответственен за большую часть процессов на протяжении всего анализа планшета, от получения до конечного анализа. Робот HTS, как правило, составляет и анализирует одновременно много планшетов, дополнительно ускоряя процесс сбора данных. Примеры устройств, подходящих для HTS в соответствии с настоящим изобретением, включают флуоресцентный спектрофотометр для считывания планшетов (FLIPRTM, Molecular Devices), FLEXstationTM (Molecular Devices), анализатор напряжения/ионов в пробе (VIPR, Aurora Biosciences), Attofluor® Ratio Vision® (ATTO).
Ниже настоящее изобретение проиллюстрировано фигурами и примерами, которые не предназначены для ограничения объема настоящего изобретения.
Описание фигур
Фигура 1. Гомологичная модель ChR2, основанного на структуре сенсорного родопсина 2 (код PDB 1H2S). Целевая область сканирования цистеина (от R115 до T139) находится в трансмембранной спирали 3 (TM3) и выделена красным. Вставка показывает предположительное расположение мутантного L132C, связанного с водородом C128 и D156, соединяющим TM3 и TM4, как показано пунктирной линией, и гомологичные остатки для донора протона (H134) и акцептора протона (E123) соответственно. Хромофор образован из полного транс-ретиналя (ATR) и K257, ковалентно связанных основанием Шиффа. Полость, образованная удалением метильных групп лейцина, изображена в виде сфер и наложена на изменившуюся сульфгидрильную группу цистеинового остатка (желтый шар). Фигуру получали с помощью VMD30.
Фигура 2. Биофизическая характеристика CatCh в клетках HEK293 и ооцитах Xenopus. (a) Слева, сводка амплитуд стационарного тока в ответ на 500 мсек импульсы синего света, измеренных при -60 мВ в клетках HEK293, экспрессирующих CatCh (черный) и ChR2 WT (красный), показано в виде среднего значения ± стандартное отклонение (n=6). Справа, сравнение кинетики спада фототоков, нормализованных на стационарные токи. (b) Слева, существующие фототоки в ответ на 1 с 473 нм импульсов синего света. Кривые нормализованы на пик амплитуды фототока для иллюстрации увеличения соотношения стационарного состояния к пиковому току CatCh (черный) по сравнению с WT (красный). Справа, сравнение кинетики нарастания фототоков, нормализованных на пиковые токи. (c) Реакции на свет 473 нм экспрессирующих CatCh и ChR2 WT ооцитов Xenopus в 80 мМ внеклеточного Ca++ (pH 9) при -120 мВ (постоянно более низкие кривые). Введение хелатора Ca++ BAPTA в конечной концентрации в цитозоле 1 мМ устранило наложенные токи конкретных Ca++-активируемых хлорных каналов, в то время как остаточные Ca++-токи канального родопсина сохранялись (пунктирные верхние кривые). Токи нормализовали на пиковый ток ChR2 WT, и они являются характерными для шести других экспериментов. До и после введения BAPTA отмечена большая разница амплитуды фототоков CatCh, свидетельствующая о его увеличенной проницаемости для Ca++ по сравнению с ChR2 WT. (d) Характеристика потока ионов CatCh в клетках HEK293 при -80 мВ (n=6, см. способы). (e) Вольт-амперные характеристики ChR2 WT (-■-) и CatCh (-▲-) в 90 мМ CaCl2 в сравнении с 140 мМ NaCl (-•-, ChR2 WT и наложенного CatCh). Токи нормализованы на ток ChR2 WT при -100 мВ. Потенциал реверсии CatCh в CaCl2 смещен к положительным потенциалам, свидетельствуя об увеличенной проницаемости Ca++ (среднее значение ± стандартное отклонение, n=5). (f) Измерения с Fura-2 потока Ca++ в клетках HEK293, экспрессирующих ChR2 WT (•) и CatCh (■), в ответ на 10 сек 460 нм света (синяя составляющая) в присутствии 90 мМ внеклеточного Ca++ (n=10), демонстрирующие четырехкратно увеличенное повышение внутриклеточного Ca++ в CatCh (контроль нетрансфицированные клетки HEK293, ▲).
Фигура 3. Экспрессия CatCh в культивируемых нейронах гиппокампа. (a) Полученное на конфокальном микроскопе изображение культивируемого нейрона гиппокампа, экспрессирующего ChR2(L132C)-2A-EGFP под промотором CAG. Масштаб 20 мкм. (b) Сравнение характерных фототоков CatCh (черный) и WT (красный), вызванных 600 мсек импульсами 473 нм синего света (J473нм 1×1019 фотонов с-1см-2). (c) Сводка амплитуд стационарных токов (-60 мВ, n=6).
Фигура 4. Быстрая с высокочувствительная фотостимуляция нейрона. (a-d) Характерные регистрации измерений токов целой клетки от экспрессирующего CatCh нейрона гиппокампа в ответ на 2 с световые импульсы. (a) Необходимая для WT 473 нм интенсивность света блокирует деполяризацию (J473нм 2,5×1017 фотонов с-1см-2). (b) Уменьшение светочувствительности восстанавливает активность (J473нм 2,5×1016 фотонов с-1см-2). (c) Характерная кривая частотной настройки света для пиковой активности (Jmax 9,7×1016 фотонов с-1см-2, среднее значение ± стандартное отклонение, 2 прочтения). (d) Умеренное зеленое освещение 532 нм также вызывает последовательность потенциалов действия (J532нм 2,5×1017 фотонов с-1см-2). (e) Время запаздывания пика светового импульса к спайку на всем протяжении последовательностей световых импульсов, состоящих из 25 1 мсек 473 нм световых импульсов (J473нм 3×1018 фотонов с-1см-2, среднее значение ± стандартное отклонение [колебание]), в 2 мМ внеклеточного Ca++ (■) и в качестве контроля при 5 Гц в 3 мМ внеклеточного Mg++ (■), который увеличивает время запаздывания до аналогичных значений ChR2 WT. (f) Спайк активности в ответ на 1 мсек 473 нм импульсы при частоте 50 Гц (J473нм 2,8×1019 фотонов с-1см-2) и (g) в ответ на 10 нс 473 мн импульсов света при 10 Гц (J473нм 1,1×1025 фотонов с-1см-2). (h) Неполная реполяризация мембраны (двунаправленная стрелка) в результате ингибирования BK каналов 1 мМ TEA. Перекрытие 3-го спайка последовательностью импульсов (черный), 1-го спайка (красный) и 3-го спайка (синий) после применения TEA (J473нм 1,8×1018 фотонов с-1см-2). (i) Замена Ca на Mg во внеклеточном растворе замедляет спайки реполяризации и вызывает пролонгированную деполяризацию (5 Гц, слева) и формирование многих спайков при более высоких частотах (20 Гц, справа) (J473нм 8,3×1018 фотонов с-1см-2).
Фигура 5. Спектроскопическая характеристика CatCh. После возбуждения светом мутант CatCh (черные линии) претерпевает фотоцикл, сравнимый с WT (красные линии) по кинетике и в присутствии фотоинтермедиатов. На фигуре показаны изменения спектров после 450 нм возбуждения длинами волн, характерными для депротонированного основания Шиффа, P390 (381 нм, верхняя панель), для P520, преобладающего в открытом состоянии (541 нм, вторая панель), и в состоянии покоя (440 нм, третья панель). Первый смещенный в красную область интермедиат, предположительно P500, невозможно регистрировать, и его детектируют только как смещение. Депротонированные основания Шиффа во временной шкале микросекунд (τ=50 мкс), событие, которое трудно наблюдать вследствие низкой амплитуды при 381 нм, сопровождающееся увеличением при 541 нм. Увеличение интермедиата P520 происходит при следующем процессе (t=1,5 мс) до его распада (t=9 мс), таким образом, давая второе более долгоживущее соединение (P480). Состояние покоя (D470) возвращается в следующем процессе (t=10 сек). Переходы в фотоцикле сходны с наблюдаемыми для WT. Что касается кинетики в открытом и закрытом состояниях при измерениях тока, мутации не вызывают грубых изменений функциональных состояний. Открытое состояние определяют в основном посредством интермедиата P520. Основное отличие обнаружено в величине амплитуды P390 по сравнению с P520, которая ниже, чем в WT. Таким образом, мутация L132C не влияет на реакцию на свет в участке хромофора. Отмечают, что спектрально-кинетические данные фотоцикла не изменялись в присутствии 50 мМ Ca++.
Фигура 6. Спектры действия CatCh, определяемые двухэлектродным способом фиксации потенциала в ооцитах Xenopus. Амплитуды токов измеряли при различных длинах волн (λ) в отсутствие Ca++ (как указано в примерах), нормализовали на поток фотонов (n=6). Сравнение состояния покоя (-) и спектра действия (■).
Фигура 7. Вызванные Ca++ изменения поверхностного потенциала. Известно, что падение потенциала на мембране зависит от применяемой разницы потенциалов (Ψ') и изменяется поверхностным потенциалом (Φ0). В основном Φ0 зависит от плотности отрицательного заряда поверхности, которую можно изменять экранированием противоионами. Таким образом, на активацию потенциалзависимых натриевых каналов (и других потенциалзависимых каналов) можно влиять изменением поверхностного заряда на внешней или внутренней стороне мембраны. В данном случае, проводимый через CatCh Ca++ нейтрализует отрицательные поверхностные заряды на внутренней стороне мембраны нейрона. Таким образом индуцируют деполяризующий эффект на мембранный потенциал, приводящий к вызыванию потенциалов действия при более низких интенсивностях света. Схематический чертеж этого механизма предоставлен на a-c (после Hille 2001). (a) В темноте канал CatCh закрыт, разница потенциалов на мембране, EM (применяемого внешнего потенциала), равна мембранному потенциалу покоя (в данном случае установленного как -60мВ). Для простоты, Φ0 устанавливали как Φ0'. (b) После активации светом CatCh возникает обычный деполяризующий мембрану приток Na+. Однако дополнительный Ca++, который поступает в нейрон, увеличивает поверхностный потенциал на внутренней стороне мембраны (Φ0"). Чем больше приток Ca++, тем более положительный Φ0" (указано двунаправленной стрелкой), и тем меньше падение потенциала на мембране. Это облегчает активацию потенциалзависимых натриевых каналов. (c) При замене внеклеточного Ca++ на Mg++, который не проникает через CatCh и всегда содержится в цитозоле ~4 мМ, возникает только незначительный деполяризующий эффект. Это происходит в результате более слабого связывания Mg++ с внутриклеточной стороной мембраны по сравнению с Ca++, который слегка понижает внутриклеточный поверхностный потенциал Φ0'. Отмечают, что деполяризующий эффект на поверхностный потенциал увеличивается при уменьшении градиента падения потенциала на мембране.
Примеры
Конструкция и биофизическая характеристика CatCh
В отличие от предшествующих подходов, целью авторов изобретения являлось определение остатков в ChR2 WT, мутации которых изменяют проницаемость катионов. Авторы изобретения уделили особое внимание третьему трансмембранному домену, т.к. было показано, что различные мутантные остатки в этом домене изменяют фотоцикл и открытие канала (фиг.1)5-7,12. Каждый остаток от Arg115 до Thr139 по отдельности замещали цистеином и проводили скрининг на обнаружение функциональных изменений на ооцитах Xenopus laevis.
Спектроскопия. CatCh экспрессировали и очищали из Pichia pastoris, как описано ранее 5'13. Проводили исследование импульсного фотолиза и измеряли изменения оптической плотности после возбуждения 10 нс лазерной вспышкой от лазера на красителях с эксимерной накачкой (450 нм, 2-3 мДж)13.
Показано, что мутация L132C (CatCh) вызывает значительные изменения амплитуды и формы кривых токов.
Клеточная культура HEK293 и молекулярная биология. Укороченный с C-конца ChR2(L132C)-YFP (вектор: pcDNA3(-)-chop2-309-(L132C)-EYFP) трансфицировали в клетки HEK293 и отбирали по G418 на протяжении всего периода времени (0,6 мг/мл, PAA Germany, Cölbe, Germany). Для ChR2 WT дикого типа укороченный с C-конца ChR2-YFP (вектор: pcDNA4TO-chop2-309-EYFP) стабильно трансфицировали в клетки HEK293-Trex (Invitrogen), которые культивировали и индуцировали, как описано. Отношения пика к стационарному состоянию определяли в клетках HEK293 транзиторно трансфицированных (Effectene, QIAGEN) конструкциями pcDNA3,1(-)-ChR2-YFP (WT, H134R или L132C) c оптимизированными для экспрессии в клетках человека кодонами за 24 часа до измерений.
Электрофизиологическая регистрация на клетках HEK293. Пэтч-пипетки с сопротивлениями 2-4 МОм получали из тонкостенного боросиликатного стекла (GB150-8P, Science Products, Hofheim, Germany) на устройстве для горизонтального вытягивания пипеток DMZ-Universal (Serial No. 5318904120B, Zeitz-Instruments, Augsburg, Germany). Фототоки регистрировали способом фиксации потенциала целой клетки и активировали световыми импульсами от твердотельного лазера с диодной накачкой (Pusch Opto Tech GmbH, Baden Baden, Germany; λ=473 нм), сфокусированными в 400 мкм оптическом волокне. Световые импульсы получали посредством быстродействующего управляемого компьютером оптического модулятора (Uniblitz LS6ZM2, Vincent Associates). Все получаемые интенсивности света измеряют в конце световода. Для оценки проницаемости для различных катионов, авторы измеряли отношения фототока к напряжению и определяли потенциал реверсии. Внутриклеточный раствор содержал 140 мМ NaCl, 7 мМ EGTA, 2 мМ MgCl2 и 10 мМ Tris (pH=9) и внеклеточный раствор содержал 140 мМ NaCl, 2 мМ MgCl2 и 10 мМ Tris (pH=9). Для проницаемостей катионов внешний 140 мМ NaCl заменяли на 140 мМ KCl, 90 мМ CaCl2 или 90 мМ MgCl2, соответственно. Проницаемость фотонов определяли из сдвига потенциала реверсии отношения тока к напряжению при понижении pH от 9 до 7,4 (или 6). Коэффициент проницаемости рассчитывали по уравнению Голдмана-Ходжкина-Катца (GHK), включая условия для Na+, K+, H+ и Ca++.
В клетках HEK293 амплитуда стационарных токов CatCh, индуцированных синим светом, являлась в ~2,5 раза выше по сравнению с ChR2 WT через 24 часа после трансфекции (CatCh: 25,0±8,8 пА/пФ, WT: 10,1±4,1 пА/пФ, среднее значение ± стандартное отклонение, n=6, -60 мВ, фиг.2a). Отношение стационарного тока к пиковому току также увеличивалось от 0,37±0,18 в WT до 0,71±0,16 в CatCh (фиг.2b). Во время повторяющейся стимуляции синим светом пиковый ток CatCh пропадал. Он восстанавливался через минуту в темноте, когда желтый свет не индуцировал преждевременного восстановления. Наоборот, полное восстановление пикового тока ChR2 WT при одинаковых условиях занимает 20 секунд13. Константы времени активации и деактивации CatCh (τon=590±3 мкс, τoff=15±2 мс, n=9, pH 7,4, -60 мВ, среднее значение ± стандартное отклонение) были немного больше по сравнению с WT ChR2 (τon=214±2 мкс, τoff=10±1 мс, n=9, pH 7,4, -60 мВ, среднее значение ± стандартное отклонение, фиг.2a,b, фиг.5 нижняя панель, таблица 1).
Затем авторы изобретения сравнивали описанные эффекты на свойства каналов со спектральными изменениями в фотоцикле. В экспериментах импульсного фотолиза на очищенном CatCh показаны только несущественные отклонения от спектра ChR2 WT13 (см. фиг.5). 1. Ранний интермедиат P390, который представляет собой депротонированное основание Шиффа, является слабо детектируемым. 2. Для интермедиата P520, который относится к открытому состоянию канала, показана немного увеличенная продолжительность жизни 9 мсек по сравнению со значением τoff, определенным электрофизиологическими методами. Аналогичные значения продолжительности жизни в открытом состоянии получали для мутантного H134R, для которого была показана удвоенная активность при неизмененной единичной проводимости2,14. Таким образом, также в случае CatCh замедленная кинетика открытого состояния может являться ответственной за увеличенные в 2,5 раза стационарные токи, измеренные в клетках HEK293, в то время как единичная проводимость остается неизмененной. Это подтверждали проводимостью одиночного канала CatCh с использованием анализа стационарного шума, как описано ранее14.
Анализ шума. Эксперименты проводили на клетках HEK293, как описано ранее14 и проводили при комнатной температуре (23ºC). Раствор пипетки содержал 1 мМ гуанидин-HCl, 199 мМ NMG-Cl (N-метилглюкамин), 10 мМ EGTA, 2 мМ MgCl2 и 20 мМ Hepes (pH 7,4), омывающий раствор содержал 200 мМ гуанидин-HCl, 2 мМ MgCl2, 2 мМ CaCl2 и 20 мМ Hepes (pH 7,4). Регистрировали отклик тока в ответ на стимуляцию синим светом при применении протокола перепада напряжений в условиях насыщения светом и снова в условиях освещения, где отклик тока при -60 мВ составлял половину максимального тока (I0,5, 2 кГц низкочастотный фильтр Бесселя, частота регистрации: 100 кГц, диаметр клетки: 15 мкм). Проводили регистрации стационарного I0,5 при продолжительном освещении (2 мин) при -60 мВ исходного потенциала для оценки проводимости одного канала (2 кГц низкочастотный фильтр Бесселя, частота регистрации 20 кГц). Проводили альтернативные регистрации без освещения (контроль, 3 регистрации) и с освещением (2 регистрации, 30 сек после начала световой стимуляции), преобразование Фурье и проводимость одиночного канала оценивали для аппроксимации функции Лоренца (для подробного описания см.14). Более низкие интенсивности света выбирали для получения максимальной флуктуации в открытом и закрытом состоянии светозависимого канала.
В экспериментах анализа шума ChR2 WT и H134R в качестве проводящего иона использовали гуанидин. Отмечают, что кинетические свойства канала не зависят от проходящего катиона14. Оценку спектров разницы мощности получали для проводимости одиночного канала γ 140±5 фСм (n=6, -60 мВ) для 200 мМ гуанидина при комнатной температуре (23ºC), которая является аналогичной исследованной при комнатной температуре проводимости одиночного канала ChR2 WT 150 фСм14. Вероятность открытого состояния CatCh, определяемая из анализа шума, остается неизменной по сравнению с H134R (P0~0,6). Таким образом, увеличенная продолжительность жизни канала в открытом состоянии может легко приводить к наблюдаемому увеличению фототоков в 2,5 раза, однако нельзя исключать незначительно увеличенную экспрессию копий CatCh.
Получение ооцитов Xenopus laevis и молекулярная биология. Укороченный с C-конца вариант ChR2 (остатки 1-315) без внеклеточно расположенных цистеиновых остатков (содержащих мутации C34A и C36A) субклонировали в вектор pTLN27. Единичные мутации цистеина вводили с помощью набора для сайт-направленного мутагенеза (QuickChange Site-Directed Mutagenesis) (Stratagene) и подтверждали секвенированием. мРНК получали с использованием набора SP6 mMessage mMachine (Ambion, Austin, TX). 50 нл кРНК, которая содержит 30 нг мРНК ChR2 WT/CatCh, инъецировали в каждый ооцит Xenopus. Ооциты получали обработкой коллагеназы после частичной овариэктомии. После инъекции кРНК ооциты инкубировали в полном транс-ретинале (1 мкМ из 1 мМ исходного раствора в этаноле) и хранили в буфере ORI (90 мМ NaCl, 2 мМ KCl, 2 мМ CaCl2 и 5 мМ Mops, pH 7,4), содержащем 1 мг/мл гентамицина при 18ºC в течение от двух до четырех суток.
Двухэлектродный способ фиксации потенциала в ооцитах Xenoyus laevis. Фототоки индуцировали 75 Вт ксеноновой дуговой лампой и 450±25 нм полосовым фильтром, свет от которых заключали в 1 мм световод с выходом ~1018 фотонов с-1см-2. Спектры действия регистрировали с использованием узкополосных фильтров (398-645 нм, ±10 нм, серия K Balzer) в комбинации с нейтральными фильтрами с получением оптоволоконного выхода ~1,4×1017 фотонов с-1см-2 для каждой длины волны. Для генерации спектров действия Ca++ в растворе ORI заменяли на Ba++ для подавления токов CaCC. Амплитуды токов для каждой длины волны нормализовали на конкретное равное экспонирование фотонами. Спектр покоя, определяемый спектроскопией, затем подбирали к усредненным точкам измерения. Для подавления активации кальций-активируемого хлорного канала (CaCC) в каждый ооцит инъецировали 50 нл 20 мМ раствора быстрого хелатора Ca2+ 1,2-бис(2-аминофенокси)этан-N,N,N',N'-тетраацетат (BAPTA) (~1 мМ конечная концентрация в ооците).
При возбуждении CatCh различными длинами волн в ооцитах Xenopus laevis было показано, что спектр действий является почти идентичным спектру ChR2 WT с максимальной длиной волны возбуждения 474 нм (фиг.6). Показано, что в присутствии внеклеточного Ca++ и при отрицательных исходных потенциалах амплитуды токов CatCh значительно увеличены во время освещения вследствие наложенного выходящего тока, который похож на ток кальций-активируемых хлорных каналов (CaCC)15,16 (фиг.2c). В экспрессирующих ChR2 WT ооцитах также наблюдали токи CaCC, но они являлись заметно более слабыми, чем индуцируемые CatCh (фиг.2c). Для ChR2 WT и CatCh при 80 мМ внеклеточного Ca++ введение быстрого хелатора Ca++ BAPTA 1,2-бис(2-аминофенокси)этан-N,N,N'N'-тетраацетата в клетку устраняло токи CaCC, тогда как остаточный ток Ca++ сохранялся (фиг.2c)1. Более выраженная разница фототоков CatCh до и после введения BAPTA поддерживает гипотезу увеличенного притока Ca++ после активации CatCh.
Увеличенная проницаемость кальция CatCh. Для получения оценки проницаемости ионов CatCh в клетках HEK293 измеряли отношения фототока к напряжению и потенциал реверсии различный катионов. В этих экспериментах было показано, что проницаемости для натрия, калия и магния являются сравнимыми для ChR2 WT (фиг.2d)1. Проницаемость протонов CatCh (pH/pNa=4*106) незначительно увеличена по сравнению с ChR2 WT (pH/pNa=2,5*106). Проницаемость Ca++ (pCa/pNa) определяли посредством сдвига потенциала реверсии, когда 140 мМ Na+ внеклеточного раствора заменяли 90 мМ Ca++. Относительная проницаемость Ca++ CatCh увеличивалась от 0,15 в ChR2 WT до 0,24, как видно из потенциала реверсии (биионный потенциал), сдвигающегося от -30,7±2,7 мВ (ChR2 WT, среднее значение ± стандартное отклонение, n=5) до -21,6±3,8 мВ (CatCh, среднее значение ± стандартное отклонение, n=5, фиг.2e). Для дополнительного количественного определения увеличенной проницаемости Ca++ CatCh авторы проводили визуализацию кальция Fura-2 в экспрессирующих CatCh клетках HEK293 и сравнивали измеренные соотношения 340/380 с соотношениями, измеренными в клетках, экспрессирующих ChR2 WT.
Визуализация Fura-2 на клетках HEK293. Fura-2 AM (5 мМ, Invitrogen) нагружали при комнатной температуре в течение от 30 мин до 1 часа. После нагрузки клетки восстанавливали в 140 мМ растворе NaCl без Ca++ (140 мМ NaCl, 7 мМ EGTA, 2 мМ MgCl2 и 10 мМ HEPES). Для оценки уровня экспрессии каждой клетки по флуоресценции YFP желтый флуоресцентный белок возбуждали, экспонируя 500 мсек светом с использованием 460/40 нм фильтр (Visitron Systems, Puchheim, Germany). Затем раствор заменяли на содержащий внеклеточный Ca++ раствор, который состоял из 90 мМ CaCl2, 7 мМ EGTA, 2 мМ MgCl2 и 10 мМ HEPES. После 15 мин в темноте светозависимые каналы стимулировали в течение 10 сек синим светом (460/40 нм). Fura-2 возбуждался при 340 нм (340/20) и 380 нм (380/20) и излучал свет (540/80 нм), детектируемый камерой на ПЗС (все фильтры от Visitron Systems, Puchheim, Germany).
Чтобы исключить изменения уровня экспрессий белка в зависимости от притока кальция, измеряемые отношения 340/380 нормализовали на значение флуоресценции YFP каждой отдельной клетки. На фигуре 2f показано, что после 10 с фотостимуляции синим светом (470 нм) 90 мМ насыщенного раствора Ca++ увеличение внутриклеточного Ca++ в экспрессирующих CatCh клетках является приблизительно в 4 раза больше, чем в экспрессирующих WT клетках.
Применение в нейронах гиппокампа
Для тестирования пригодности CatCh к применению в нейронах, конструкция экспрессировалась в культивируемых пирамидальных клетках гиппокампа.
Культура нейронов гиппокампа. Гиппокамп выделяли из крыс Sprague-Dawley в первые сутки постнатального периода (Jackson Laboratory) и обрабатывали папаином (20 ед. мл-1) в течение 20 мин при 37°C.
Гиппокамп отмывали DMEM (Invitrogen/Gibco, с высоким содержанием глюкозы) с добавлением 10% эмбриональной телячьей сыворотки и растирали в небольшом объеме этого раствора. ~75000 клеток высевали на покрытые поли-D-лизином/ламинином стеклянные покровные стекла в 24-луночные планшеты. Через 3 часа среду для посадки заменяли культуральной средой (Neurobasal A, содержащей 2% добавки B-27, 2 мМ Glutamax-I и 100 ед/мл пенициллина и 100 мкг/мл стрептомицина). ChR2(L132C)-YFP и ChR2-YFP трансфицировали через 5-10 суток после рассевания с использованием реагента липофектамина 2000 (Invitrogen). Альтернативно, через 4-9 суток после рассевания к каждой лунке добавляли 2-5×109 геномных копий/мл вируса (AAV2/7-CAG-ChR2(L132C)-2A-EGFP-WPRE-bGH). Экспрессия становилась видимой на 5 сутки после трансдукции. В течение всего периода жизни культуры (~5 недель) не наблюдали нейротоксичность. Полностью тарнс-ретиналь не добавляли в культуральную среду или в регистрирующую среду для любого из описанных выше экспериментов.
Конструкция аденоассоциированного вирусного вектора. Энхансер раннего промотора цитомегаловируса/промотор β-актина цыпленка (CAG) амплифицировали ПЦР и вставляли в pAAV2-Rho-EGFP (любезно предоставленный Alberto Auricchio28) для получения pAAV2-CAG-EGFP. Вирусная экспрессирующая плазмида pAAV2-CAG-EGFP дополнительно содержала посттранскрипционный регуляторный элемент вируса гепатита лесного сурка (WPRE) и полиаденилированиую последовательность бычьего гормона роста (BGH). ChR2(L132C)-2A-EGFP (любезно предоставленный Volker Busskampö - 2A саморасщепляющийся пептид/CHYSEL29) конструировали посредством ПЦР с адаптером и субклонировали в pAAV2-CAG-EGFP, заменяя EGFP, с использованием набора Clontech's in fusion. Вирусный вектор (pAAV2-CAG-ChR2(L132C)-2A-EGFP-WPRE-bGH) упаковывали (серотип 7) и аффинно очищали в рамках программы генотерапии Университета Пенсильвании с конечным инфекционным титром вируса 2,26×1011 геномных копий/мл.
Электрофизиологические регистрации от нейронов гиппокампа. Для регистраций целой клетки в культивируемых нейронах гиппокампа пэтч-пипетки с сопротивлением 5-10 МОм наполняли 129 мМ глюконатом калия, 10 мМ HEPES, 10 мМ KCl, 4 мМ MgATP и 0,3 мМ Na3GTP, оттитрованными до pH 7,2. В качестве внеклеточного раствора применяли раствор Тироде (125 мМ NaCl, 2 мМ KCl, 2 мМ CaCl2, 1 мМ MgCl2, 30 мМ глюкозы и 25 мМ HEPES, оттитрованными до pH 7,4). По существу не содержащий Ca++ внеклеточный раствор содержал тот же аналогичный раствор за исключением того, что он содержал 0 мМ Ca++ и 3 мМ Mg++. Регистрации проводили в присутствии ингибиторов возбуждающей синаптической передачи сигналов, 1,2,3,4-тетрагидро-6-нитро-2,3-диоксо-бензо[f]хиноксалин-7-сульфонамида (NBQX, 10 мкМ, Sigma) и D(-)-2-амино-5-фосфонопентановой кислоты (AP-5, 50 мкМ, Sigma). Для регистраций способом фиксации потенциалов к внеклеточному раствору добавляли 1 мкМ тетродотоксина. Для ингибирования активности BK-каналов добавляли 1 мМ TEA. Регистрации проводили на инвертированном микроскопе Zeiss Axiovert 25, оборудованном флуоресцентной лампой. Успешную экспрессию белка подтверждали EGFP- или YFP-опосредованной флуоресценцией. Входное сопротивление нейронов составляло 15-40 МОм, и за его стабильностью наблюдали на всем протяжении эксперимента. Электрофизиологические сигналы амплифицировали с использованием амплификатора Axopatch 200A (Axon Instruments, Union City, CA), фильтровали при 10 кГц, преобразовывали в цифровую форму устройством Axon Digidata 1600 (50 Гц) и регистрировали, и анализировали с использованием программного обеспечения pClamp9 (Axon Instruments). Фототоки вызывали с использованием световых импульсов различных длин твердотельного лазера с диодной накачкой (Pusch Opto Tech GmbH, λ1=473 нм, P1=100 мВт, λ2=532 нм, P2=50 мВт) или 10 нс вспышками лазера на красителях с эксимерной накачкой (кумарин 2, λ=450 нм). Конкретные интенсивности света указаны в легенде на чертеже и в тексте и представляют собой интенсивности на конце кварцевого оптического волокна диаметром 400 мкм (STE-F100/400-Y-VIS/NIR, Laser 2000, Wessling, Germany) на расстоянии ~500 мкм от клетки. Токи, измеряемые от нейронов, экспрессирующих ChR2(L132C)-YFP и ChR2(L132C)-2A-EGFP, являлись идентичными.
Конфокальная микроскопия. Для визуализации покровные стекла с нейронами гиппокампа фиксировали при 4°C в течение 10 мин 4% параформальдегидом в буфере PBS, содержащем 2% сахарозы. Затем клетки инкубировали в течение 1,5 часов с IgG кролика против GFP (Invitrogen, A11122) с последующей 45 минутной инкубацией с конъюгированными с Alexa Fluor 488 антителами осла к IgG кролика (Invitrogen, A21206). Иммунофлуоресценцию покровных стекол, покрытых средой для заливки, фотографировали конфокальным микроскопом Zeiss LSM 510 (Zeiss, Plan-Neofluar 40×/0,75).
Мутант CatCh устойчиво экспрессировался в культурах гиппокампа (фиг.3a) в течение недель без признаков нейротоксичности, и для него в клетках HEK293 было показано более высокое соотношение стационарного состояния к пику при регистрациях целой клетки и приблизительно пониженные в четыре раза амплитуды токов 644±31 пА (-60 мВ, n=6, среднее значение ± стандартное отклонение) по сравнению с WT с 164±39 пА (-60 мВ, n=6, среднее значение ± стандартное отклонение) в ответ на синий свет 473 нм (фиг.3b). В режиме фиксации тока, как правило, используемые завышенные интенсивности света для активации WT (1018-1019 фотонов с-1см-2) приводили к блокированию деполяризации в экспрессирующих CatCh пирамидальных клетках (фиг.4a). Для индукции достоверных последовательностей спайков интенсивность света уменьшали на 2 логарифмические единицы (5×1016-2×1017 фотонов с-1см-2) до интенсивности света природного диапазона, приводящего к фототопическому зрению фоторецепторов колбочек (фиг.4b)11. На фигуре 4c показана характерная кривая частотной настройки для зависящей от интенсивности света частоты возбуждения пирамидальной клетки. Усредненный максимум частоты возбуждения составляет 8,2×1016±2,5×1016 фотонов с-1см-2 (среднее значение ± стандартное отклонение, n=5). Более высокая эффективность активирования светом экспрессирующих CatCh нейронов облегчает активацию длинами волн, далекими от пика чувствительности, как приведено в пример для зеленого света (532 нм) на фигуре 4d. Это может предоставлять преимущества в отношении более эффективного подбора тканей с более глубоким проникновением зеленого света. Авторы предоставляют существенно усиленную интенсивность светочувствительности экспрессирующих CatCh нейронов для увеличенной проницаемости Ca++, таким образом, временно увеличивающую поверхностный потенциал цитозольной поверхности мембраны10,11,18,19 (для объяснения эффекта Ca++ на поверхностный потенциал см. фиг.7). При возбуждении CatCh служит в качестве источника, быстро связывающегося с мембраной Ca++ посредством временного увеличения локальной внутриклеточной поверхностной концентрации Ca++, таким образом, нейтрализуя отрицательные поверхностные заряды (фиг.7). Известно, что это вызывает сдвиг потенциала внутренней поверхности в сторону более положительных значений, таким образом, деполяризуя мембрану (фиг.7)11,18. Следствием является то, что потенциалзависимые Na+-каналы активируются при более отрицательных мембранных потенциалах18. После коротких импульсов света или после выключения стационарного освещения Ca++ быстро приходит в равновесное состояние (микросекунды) в цитоплазме, приводя к быстрому восстановлению и немедленному исчезновению потенциалов действия. Таким образом, для CatCh необходимо меньше фототоков и, как следствие, меньше света для индукции спайков по сравнению с ChR2 WT. Время запаздывания светового импульса к спайку в CatCh было короче (~5-6 мс, фиг.4e) с меньшим колебанием по сравнению с временем запаздывания для WT ChR2 (~10 мс) при аналогичных интенсивностях света (2,8×1018 фотонов с-1см-2)3. Авторы изобретения дополнительно тестировали CatCh на его способность индуцировать единичные потенциалы действия при высокой интенсивности световой стимуляции. Последовательность световых импульсов длительностью 1 мсек 473 нм (2,8×1019 фотонов с-1см-2) приводила к 100% достоверной последовательности спайков до частот 50 Гц (n=8, фиг.4f - большинство пирамидальных клеток не отражают хорошо более 50 Гц даже с инжекциями прямого тока). С другой стороны, для WT необходимо по меньшей мере 2 мсек световые импульсы для достоверной индукции спайков, и происходит это только до частот 20 Гц12. Авторы подбирали еще более короткие времена активации CatCh и индуцировали единичные потенциалы действия до частот 10 Гц при 10 нс импульсов голубого света (1,1×1025 фотонов с-1см-2), где длительность импульсов являлась достаточно короткой для индукции только одного функционального цикла в каждом белке CatCh (фиг.4g). Однако для быстрых частот активации также необходима быстрая реполяризация клетки после каждого спайка. Несмотря на уменьшение τoff CatCh по сравнению с WT, увеличение в 4 раза притока Ca++ при активации CatCh (см. измерения Fura-2 фиг.2f) является достаточным для активации в достаточной мере Ca++-активируемых калиевых каналов высокой проводимости (BK каналы)20 для потенциальной реполяризации клетки до ее исходного потенциала покоя за миллисекунды после каждого потенциала действия. Для доказательства того, что быстрая реполяризация являлась опосредованной BK каналами, авторы добавляли 100 мкМ блокатора калиевых каналов тетраэтиламмония (TEA) к внеклеточному раствору и наблюдали неполную реполяризацию мембраны и образование потенциала плато, как правило, наблюдаемого в протоколах при импульсной стимуляции ChR2 WT3 (фиг.4h).
В совокупности для экспрессирующих CatCh нейронов показаны более быстрое возникновение спайков, более быстрая реполяризация и увеличенная светочувствительность по сравнению с клетками, экспрессирующими WT (для сравнения см. таблицу 1). Контрольные эксперименты при отсутствии внеклеточного Ca++ и в присутствии 3 мМ Mg++, который оказывает менее выраженный эффект на поверхностный потенциал11,18 (фиг.7) и не проходит через ChR2 WT или CatCh, поддерживают указанные выше объяснения: 1) Время запаздывания светового импульса к спайку увеличивалось до значений ChR2 WT (фиг.4e); 2) Вместо быстрого спайка реполяризации, как наблюдали в присутствии Ca++, наблюдали пролонгированную искусственную деполяризацию, аналогичную той, что наблюдают в экспериментах с ChR2 WT (фиг.4i, слева); 3) В отсутствие Ca++ одинаковые интенсивности света приводили к сниженной деполяризации при ~10 мВ при иных равных экспериментальных условиях и 4) Множественные спайки, как и ожидалось вследствие пролонгированного времени в открытом состоянии CatCh, повторяются при отсутствии Ca++ (фиг.4i, справа).
Таким образом, авторы изобретения продемонстрировали, что CatCh, канальный родопсин с повышенной проницаемостью Ca++, объединяет повышенную светочувствительность и быструю кинетику и, таким образом, превосходит по эффективности ChR2 WT и опубликованных быстрых и медленных мутантов (для сравнения свойств различных вариантов ChR2 см. таблицу 1).
Обсуждение
На первый взгляд для CatCh, мутанта L132C ChR2 WT, показаны достаточно неожиданные результаты по сравнению с ChR2 WT: 1) увлечение в два раза времени жизни в открытом состоянии; 2) замедленный спад интермедиата P520 в кинетике фотоцикла; 3) неизмененная проводимость одиночного канала и 4) незначительно смещенный в красную область максимум поглощения (4 нм). Увеличенные в 2,5 раза фототоки можно легко объяснить измеряемыми параметрами без соответствующего увеличения уровня экспрессии. С другой стороны, тем не менее, при более внимательном рассмотрении данных фиксации потенциалов, полученных от экспрессирующих CatCh ооцитов Xenopus laevis, получили первое указание на повышенную проницаемость Ca++, которую затем подтвердили определением потенциала реверсии и экспериментами по визуализации кальция на клетках HEK293.
Рассматривая модель на фигуре 1, увеличению проницаемости Ca++ могло бы способствовать формирование более гибкой структуры и, таким образом, формирование полости, как показано для мутации L94A активируемого светом протонного насоса бактериородопсина (сравнение фиг.1)21. Такая полость могла бы располагаться в гидрофобном участке как часть консервативной трансмембранной спирали три (TM3), только оборота спирали за исключением C128. Манипулирование взаимодействия C128 (TM3) и D156 (TM4) значительно замедляет реакционный цикл ChR25,6, эффект, который также наблюдали в мутанте L93A бактериородопсина22,23, т.е. расположенного рядом остатка L94. В ChR2 взаимодействие TM3 и TM4 вероятно влияет на открытие канала и избирательность, указывая на структурный элемент как на передатчик реакции света в ионную пору24. Вставка меньшего и более гидрофильного цистеина могла бы увеличивать гибкость сегмента спирали, облегчая доступ Ca++.
При доставке в пирамидальные клетки гиппокампа для CatCh выявлено повышение светочувствительности в 70 раз по сравнению с ChR2 WT. Как правило, такая увеличенная эффективность активация светом сопровождается значительно пролонгированным временем жизни канала в открытом состоянии2,6,13. Это не является случаем CatCh. Наблюдаемая светочувствительность заметно отличается от той, которую наблюдали для других канальных родопсинов до настоящего времени. Как объяснено ниже, вторичные эффекты на возбудимость нейронов вызваны притоком Ca++ через CatCh. Несмотря на более медленно затухающую кинетику по сравнению с WT, для CatCh показана увеличенная достоверность спайков и точность при высокочастотной стимуляции светом, уменьшение дополнительных спайков и устранение искусственных потенциалов плато, как правило, наблюдаемых в экспрессирующих WT клетках при стимуляции частотами выше 20 Гц3,7,12. 1 мсек световые импульсы, предоставляемые при комнатной температуре, индуцировали достоверные последовательности спайков до 50 Гц в экспрессирующих CatCh пирамидальных клетках (их естественный предел естественной генерации спайков, фиг.4f). Более высокую частоту опосредованной CatCh индукции спайков на более быстро генерирующих спайки клетках, таких как кортикальные содержащие парвальбумин вставочные нейроны, еще необходимо протестировать12. Поскольку кинетика канального родопсина является зависимой от температуры с Q10 ~2,314, авторы изобретения ожидают увеличенную в 3,2 раза кинетику CatCh в экспериментах in vivo при 37°C без потери светочувствительности. Это позволило бы стимуляцию опосредованных CatCh спайков по меньшей мере до 300 Гц. Повышенная светочувствительность в сочетании с быстрой кинетикой и высокой временной точностью позволила авторам активировать CatCh такими коротким импульсами света как 10 нс, которые активируют один функциональный цикл в каждой молекуле CatCh с последующим одиночным спайком. Наблюдения в возбудимых клетках лучшим образом объясняются увеличенным потоком Ca++ в нейрон при освещении. Отмечают, что вкладом Ca++ в движущую силу вследствие его увеличенной проницаемости можно пренебрегать. Тем не менее, авторы могут рассматривать CatCh как светозависимый связанный с мембраной источник Ca++ ("связанный с мембраной внутриклеточный Ca++"), который временно поставляет Ca++ на цитозольную поверхность клеточной мембраны при условии, что канал CatCh открыт. Это временно нейтрализует отрицательные заряды на внутренней стороне мембраны, таким образом, увеличивая поверхностный потенциал, который равен деполяризации мембраны11 (фиг.7). Как и ожидалось, когда внеклеточный Ca++ заменяли на непроникающий Mg++, все наблюдаемые эффекты Ca++ на потенциал действия исчезали, восстанавливая фенотип ChR2 WT. Это доказывает, что наблюдаемые эффекты Ca++ являлись результатом притока внеклеточного Ca++ и не были вызваны повышением [Ca++]i, опосредованным потенциальной экспрессией CatCh в клеточных органеллах, таких как эндоплазматический ретикулум. Быстрая начальная деполяризация посредством поверхностного потенциала уменьшает вдвое время задержки светового импульса к пику от ~10 мсек в экспрессирующих ChR2 WT клетках3 до ~5 мсек в экспрессирующих CatCh клетках. По сравнению с ChR2 WT отношение пика к стационарному току в CatCh значительно снижено (см. таблицу 1). Таким образом, при длительном освещении CatCh уровень деполяризации клетки остается почти стационарным. Продолжительный приток Ca++ при непрерывном освещении может активировать кальций-активируемые неселективные катионные каналы, которые дополнительно поддерживают сохранение стационарного уровня деполяризации. С другой стороны, предпосылкой успешной высокочастотной импульсной стимуляции является быстрая реполяризация клетки, следующая после каждого потенциала действия. Незначительно увеличенное время жизни CatCh в открытом состоянии по сравнению с WT должно ограничивать его максимальную частоту стимуляции. Однако увеличенная проницаемость Ca++ препятствует этому ограничению посредством эффективной активации кальций-зависимых калиевых каналов высокой проводимости (BK каналов). Это восстанавливает мембранный потенциал покоя нейрона за миллисекунды после каждого потенциала действия. Эту теорию подтвердили ингибированием BK каналов блокатором канала в открытом состоянии TEA, который вызывал непрерывную деполяризацию нейрона при длительности импульса по протоколу импульсной стимуляции.
По сравнению с уже доступными оптогенетическими способами CatCh обладает повышенной светочувствительностью, аналогичной как у медленных мутантов13 или SFO6, но со значительно ускоренной кинетической реакцией вследствие его увеличенной проницаемости для Ca++ и следствие возбудимости нейронов. Это делает CatCh превосходящим доступные варианты ChR2, где получили высокую светочувствительность за счет быстрой кинетики и наоборот в отношении быстрых канальных родопсинов7,12 (для обзора см. таблицу 1).
Принимая во внимание применение в оптогенетике, авторы отмечают, что важным будет являться валидация оптимальных параметров светового импульса в каждой постановке эксперимента, таких как длительность и интенсивность стимуляции, т.к. специфический ответ суммарно будет зависеть от внутренних биофизических свойств нейрона и уровней экспрессии CatCh.
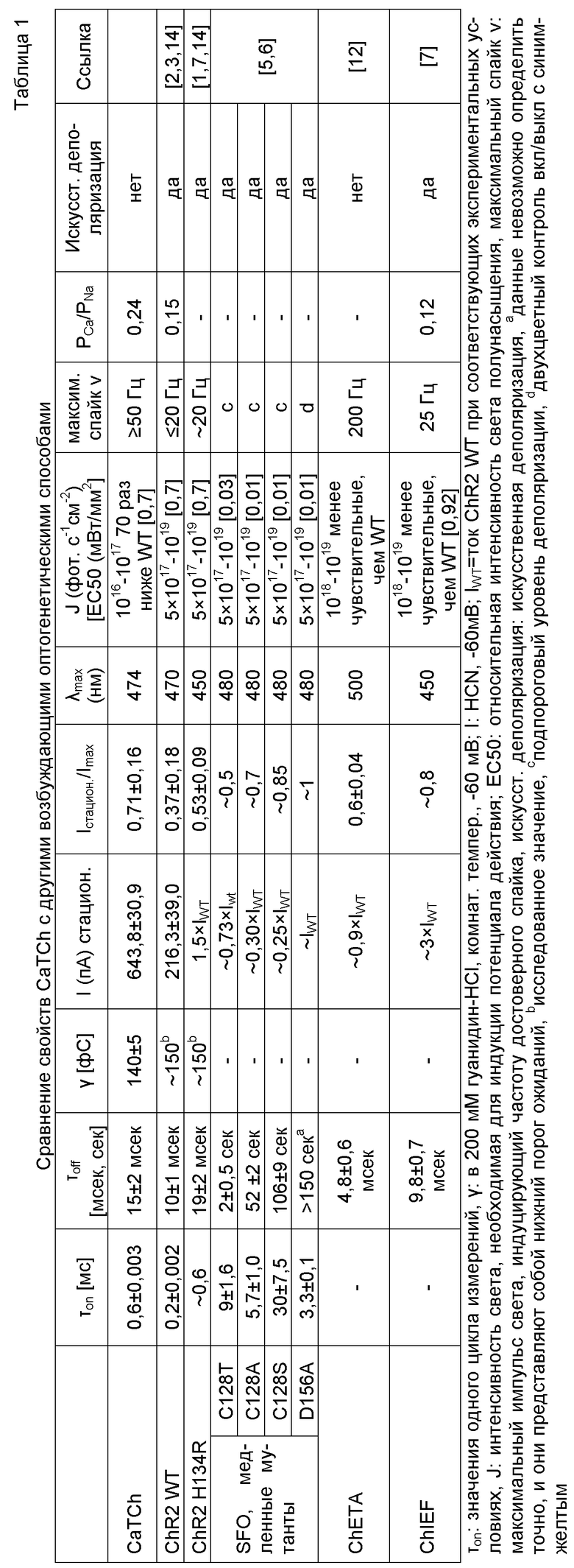
Список ссылок
1. Nagel, G. et al. Channelrhodopsin-2, a directly light-gated cation-selective membrane channel. Proc Natl Acad Sci USA 100, 13940-13945 (2003).
2. Nagel, G. et al. Light activation of channelrhodopsin-2 in excitable cells of Caenorhabditis elegans triggers rapid behavioral responses. Curr Biol 15, 2279-2284 (2005).
3. Boyden, E., Zhang, F., Bamberg, E., Nagel, G. & Deisseroth, K. Millisecond-timescale, genetically targeted optical control of neural activity. Nat Neurosci 8, 1263-1268 (2005).
4. Zhang, F. et al. Multimodal fast optical interrogation of neural circuitry. Nature 446, 633-639 (2007).
5. Bamann, C., Gueta, R., Kleinlogel, S., Nagel, G. & Bamberg, E. Structural guidance of the photocycle of channelrhodopsin-2 by an interhelical hydrogen bond. Biochemistry 49, 267-278 (2010).
6. Berndt, A., Yizhar, O., Gunaydin, L., Hegemann, P. & Deisseroth, K. Bi-stable neural state switches. Nat Neurosci 12, 229-234 (2009).
7. Lin, J., Lin, M., Steinbach, P. & Tsien, R. Characterization of engineered channelrhodopsin variants with improved properties and kinetics. Biophys J 96, 1803-1814 (2009).
8. Lagali, P. et al. Light-activated channels targeted to ON bipolar cells restore visual function in retinal degeneration Nat Neurosci 11, 667-675 (2008).
9. Bi, A. et al. Ectopic expression of a microbial-type rhodopsin restores visual responses in mice with photoreceptor degeneration. Neuron 50, 23-33 (2006).
10. Frankenhäuser, B. & Hodgkin, A. The action of calcium on the electrical properties of squid axons. J Physiol (Lond) 137, 218-244 (1957).
11. Hille, B. 649-662 (Sinauer Associates, Sunderland Massachusetts USA, 2001).
12. Gunaydin, L. et al. Ultrafast optogenetic control. Nat Neurosci 13, 387-392 (2010).
13. Bamann, C., Kirsch, T., Nagel, G. & Bamberg, E. Spectral characteristics of the photocycle of channelrhodopsin-2 and its implication for channel function. J Mol Biol 375, 686-694 (2008).
14. Feldbauer, K. et al. Channelrhodopsin-2 is a leaky proton pump. Proc Natl Acad Sci USA (2009).
15. Caldwell, J. et al. Increases in intracellular calcium triggered by channelrhodopsin-2 potentiate the response of metabotropic glutamate receptor mGluR7. J Biol Chemistry 283, 2430024307 (2008).
16. Weber, W.-M. Ion currents in Xenopus laevis oocytes: state of the art. Biochim Biophys Acta 1421, 213-233 (1999).
17. Thyagarajan, S. et al. Visual function in mice with photoreceptor degeneration and transgenic expression of channelrhodopsin 2 in ganglion cells. J Neurosci 30, 8745-8758 (2010).
18. Hille, B., Woodhull, B. & Shapiro, B. Negative surface charge near sodium channels of nerve: divalent ions, monovalent ions, and pH. Philos Trans R Soc Lond B Biol Sci 270, 301-318 (1975).
19. Muller, R. & Finkelstein, A. The effect of surface charge on the voltage-dependent conductance induced in thin lipid membranes by monazomycin. J General Physiol 60, 285-306 (1972).
20. Faber, E. & Sah, P. Calcium-activateed potassium-channels: multiple contributions to neuronal function. Neuroscientist 9, 181-194 (2003).
21. Joh, N., Oberai, A., Yang, D., Whitelegge, J. & Bowie, J. Similar energetic contributions of packing in the core of membrane and water-soluble proteins. J Am Chem Soc 131, 10846-10847 (2009).
22. Subramaniam, S., Faruqi, A., Oesterhelt, D. & Henderson, R. Electron diffraction studies of light-induced conformational changes in the Leu-93→Ala bacteriorhodopsin mutant. Proc Nat Acad Sci USA 94 1767-1772 (1997).
23. Subramaniam, S., Greenhalgh, D., Rath, P., Rothschild, K. & Khorana, H. Replacement of leucine-93 by alanine or threonine slows down the decay of the N and O intermediates in the photocycle of bacteriorhodopsin: implications for proton uptake and 13-cis-retinal all-trans-retinal reisomerization. Proc Natl Acad Sci USA 88, 6873-6877 (1991).
24. Nack, M. et al. The DC gate in Channelrhodopsin-2: crucial hydrogen bonding interaction between C128 and D156. Photochemical & Photobiological Sciences 9, 194-198 (2010).
25. Busskamp, V. et al. Genetic reactivation of cone photoreceptors restores visual responses in retinitis pigmentosa. Science 329, 413-417 (2010).
26. Ellis-Davies, G. C. R. Neurobiology with caged calcium. Chem Rev 108, 1603-1613 (2008).
27. Lorenz, C., Pusch, M. & Jentsch, T. Heteromultimeric ClC chloride channels with novel properties. Proc Natl Acad Sci USA 92, 13362-13366 (1996).
28. Allocca, M. et al. Novel adeno-associated virus serotypes efficiently transduce murine photoreceptors. J Virology 81, 11372-11380 (2007).
29. de Felipe, P. et al. E unum pluribus: multiple proteins from a self-processing polyprotein. Trends Biotechnol 24, 68-75 (2006).
30. Humphrey, W., Dalke, A. and Schulten, K. VMD - Visual Molecular Dynamics. J Molec Graphics 14, 33-38 (1996).
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОДАВЛЕНИЕ ЭКСПРЕССИИ ГЕНОВ У НАСЕКОМЫХ-ВРЕДИТЕЛЕЙ | 2018 |
|
RU2725953C2 |
| T-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ, РАСПОЗНАЮЩИЕ МУТАЦИЮ R175H ИЛИ Y220C В P53 | 2020 |
|
RU2830061C2 |
| ВАРИАНТНЫЕ СВЯЗЫВАЮЩИЕ CD3 ДОМЕНЫ И ИХ ПРИМЕНЕНИЕ В КОМБИНИРОВАННОЙ ТЕРАПИИ ПРИ ЛЕЧЕНИИ ЗАБОЛЕВАНИЙ | 2019 |
|
RU2810222C2 |
| МОДИФИЦИРОВАННЫЕ НЕПРИРОДНЫЕ ЛИГАНДЫ NKG2D, КОТОРЫЕ ИЗБИРАТЕЛЬНО ДОСТАВЛЯЮТ ПРИСОЕДИНЕННЫЕ ГЕТЕРОЛОГИЧНЫЕ МОЛЕКУЛЫ К НЕПРИРОДНЫМ РЕЦЕПТОРАМ NKG2D НА CAR-КЛЕТКАХ | 2020 |
|
RU2823728C2 |
| АКТИВИРУЕМЫЕ ПРОТЕАЗОЙ СВЯЗЫВАЮЩИЕ Т-КЛЕТКИ БИСПЕЦИФИЧЕСКИЕ МОЛЕКУЛЫ | 2017 |
|
RU2830288C2 |
| МУЛЬТСПЕЦИФИЧЕСКИЕ АНТИТЕЛА, МУЛЬТСПЕЦИФИЧЕСКИЕ АКТИВИРУЕМЫЕ АНТИТЕЛА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2815683C2 |
| ГИБРИДНЫЕ БЕЛКИ ВАРИАНТА sPD-1—FC | 2019 |
|
RU2785993C2 |
| АКТИВИРУЕМЫЕ ПОЛИПЕПТИДЫ ИНТЕРЛЕЙКИНА-2 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2826454C2 |
| АНТИТЕЛА ПРОТИВ SARS-COV-2 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2021 |
|
RU2832025C1 |
| АНТИТЕЛА К С5 И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2746356C2 |
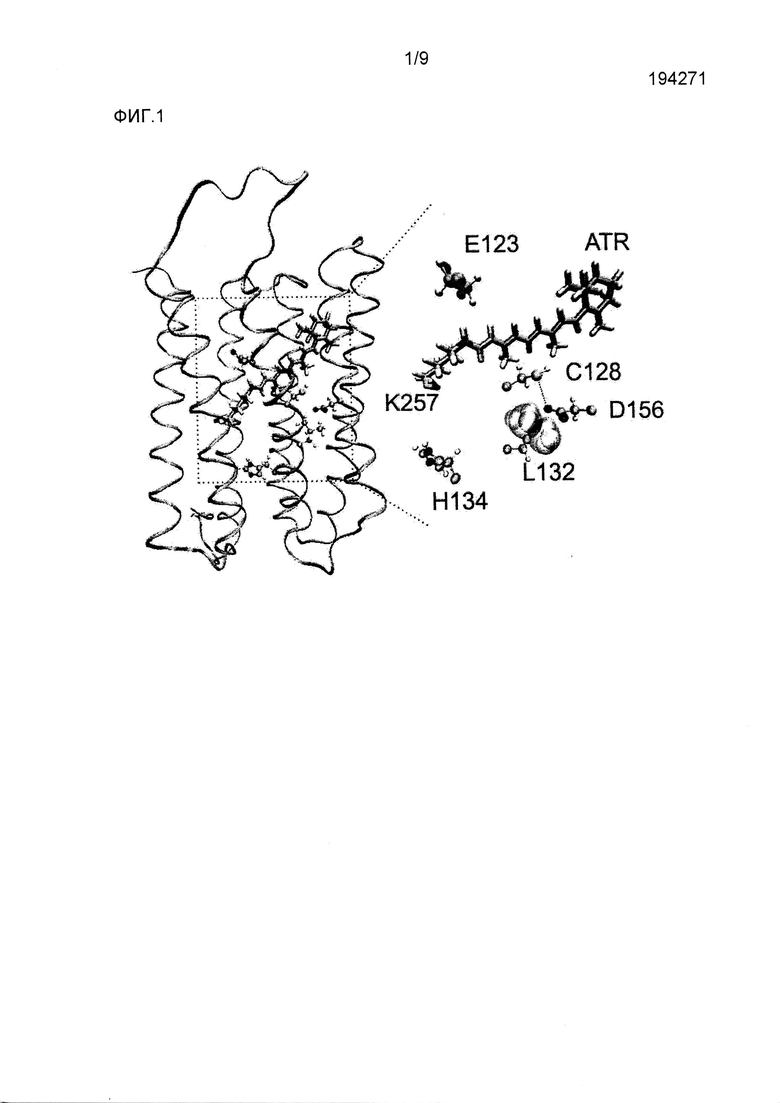
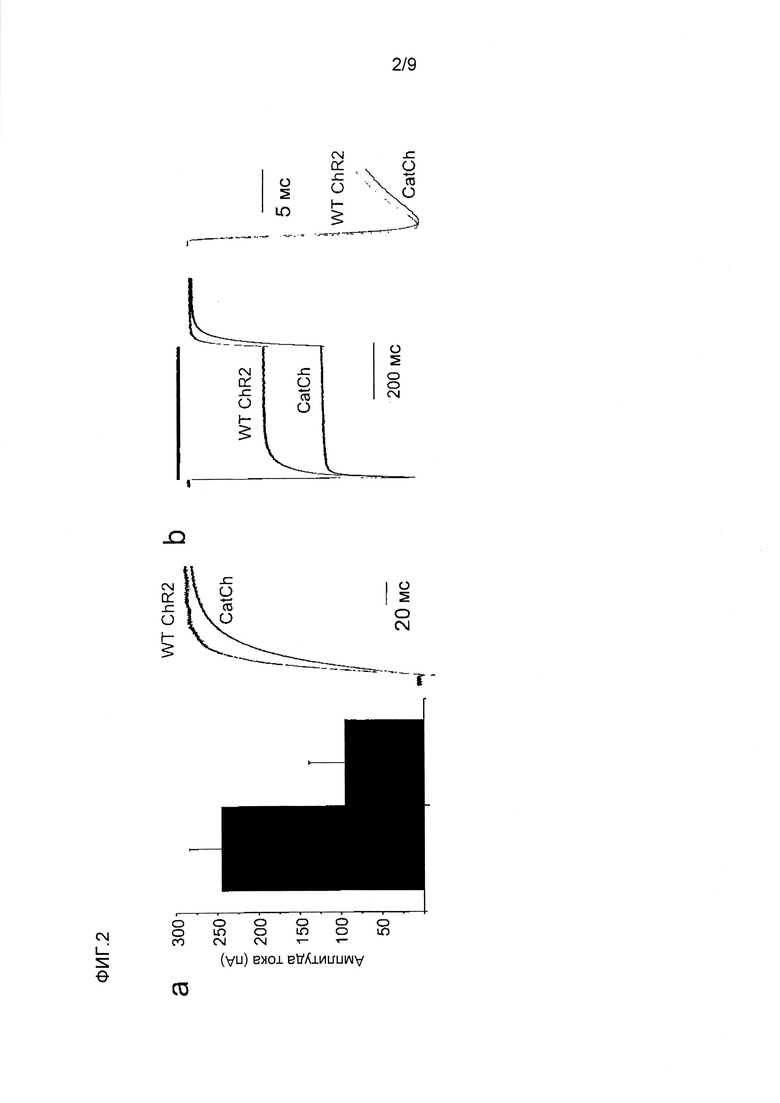
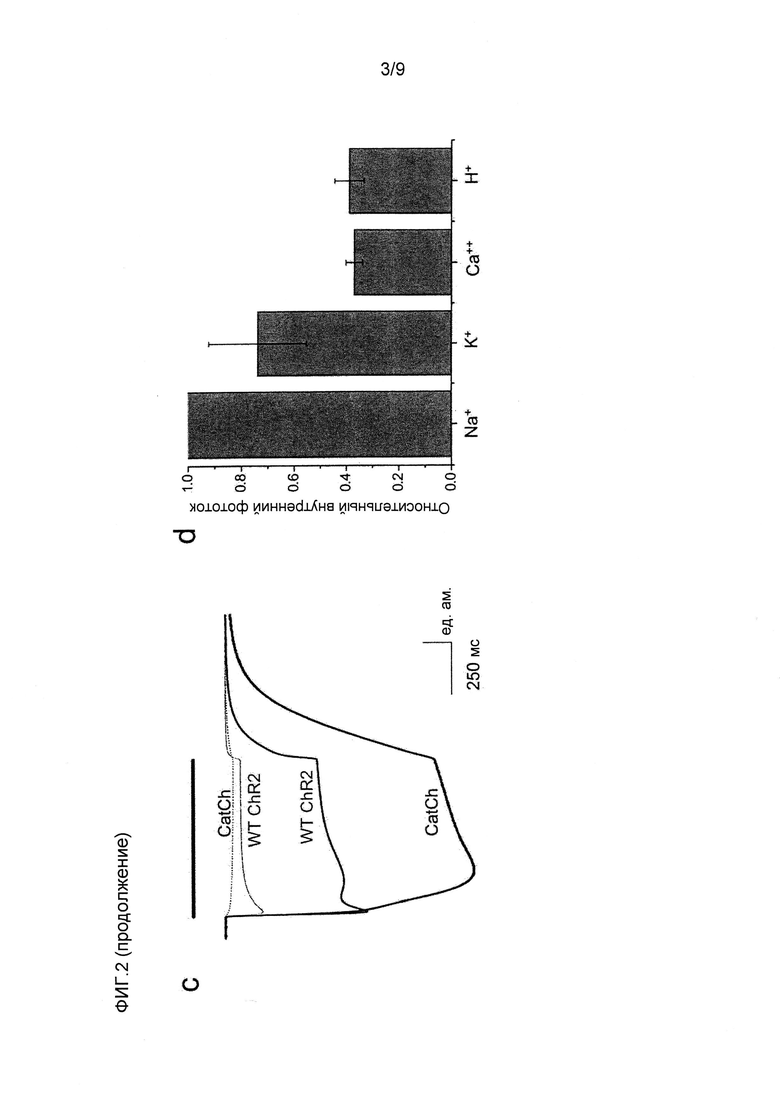
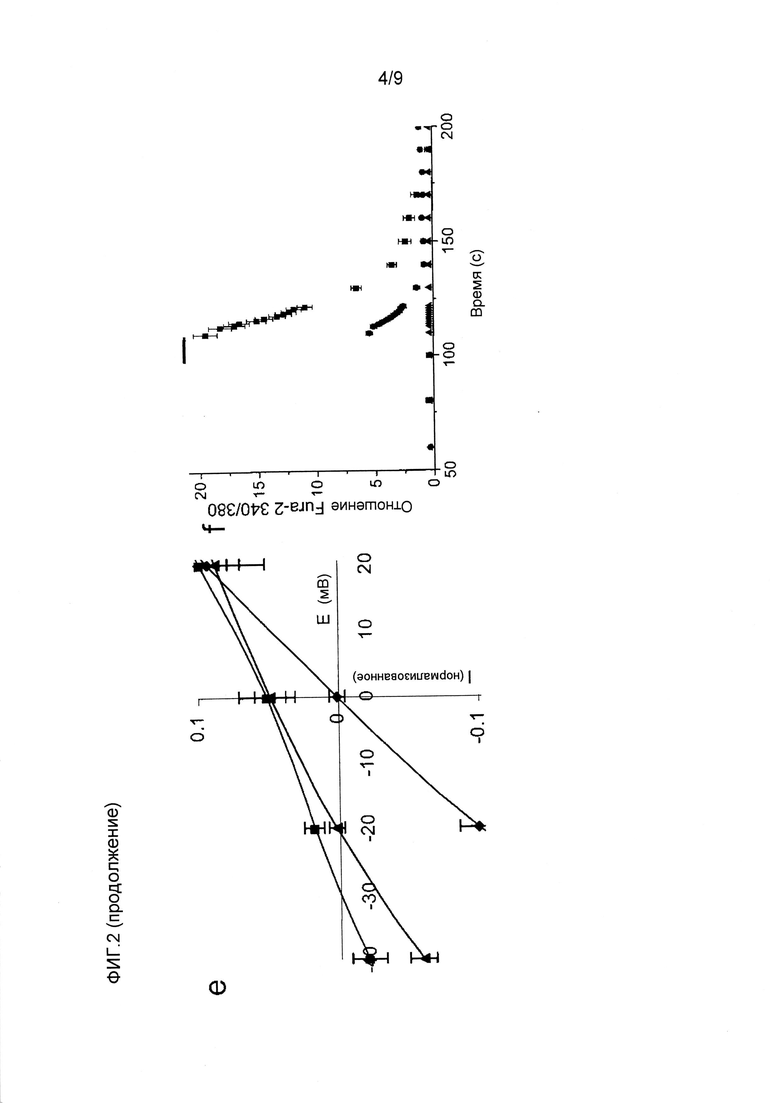
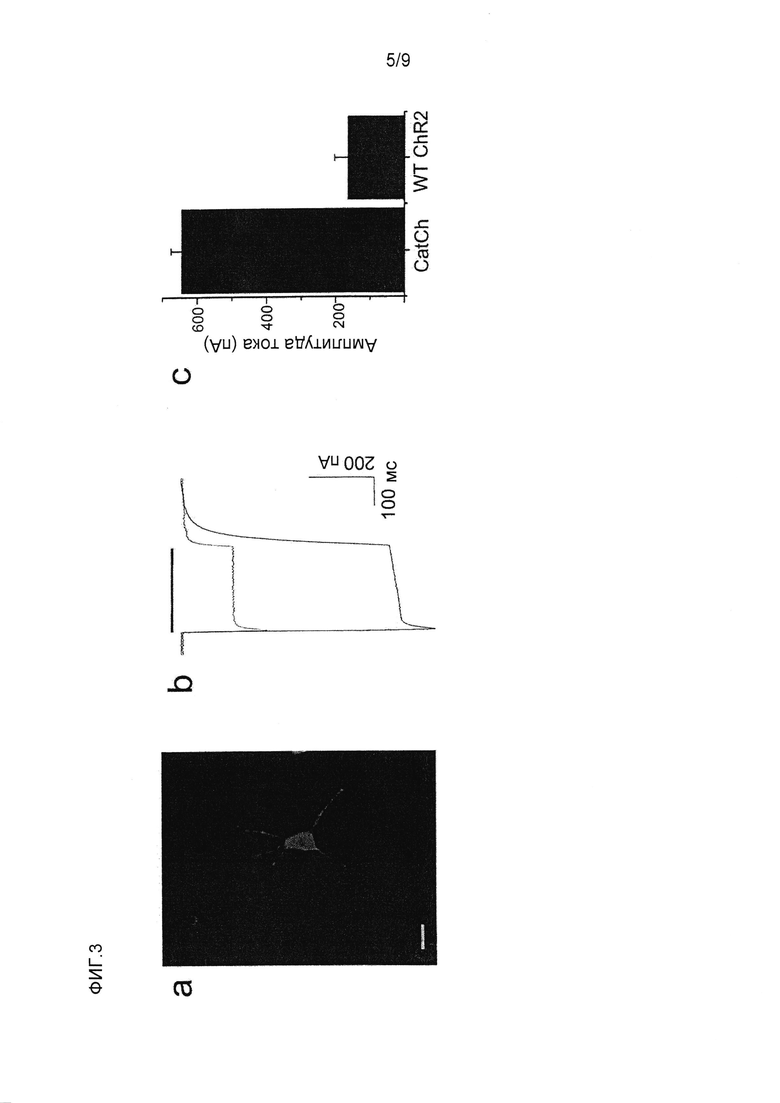
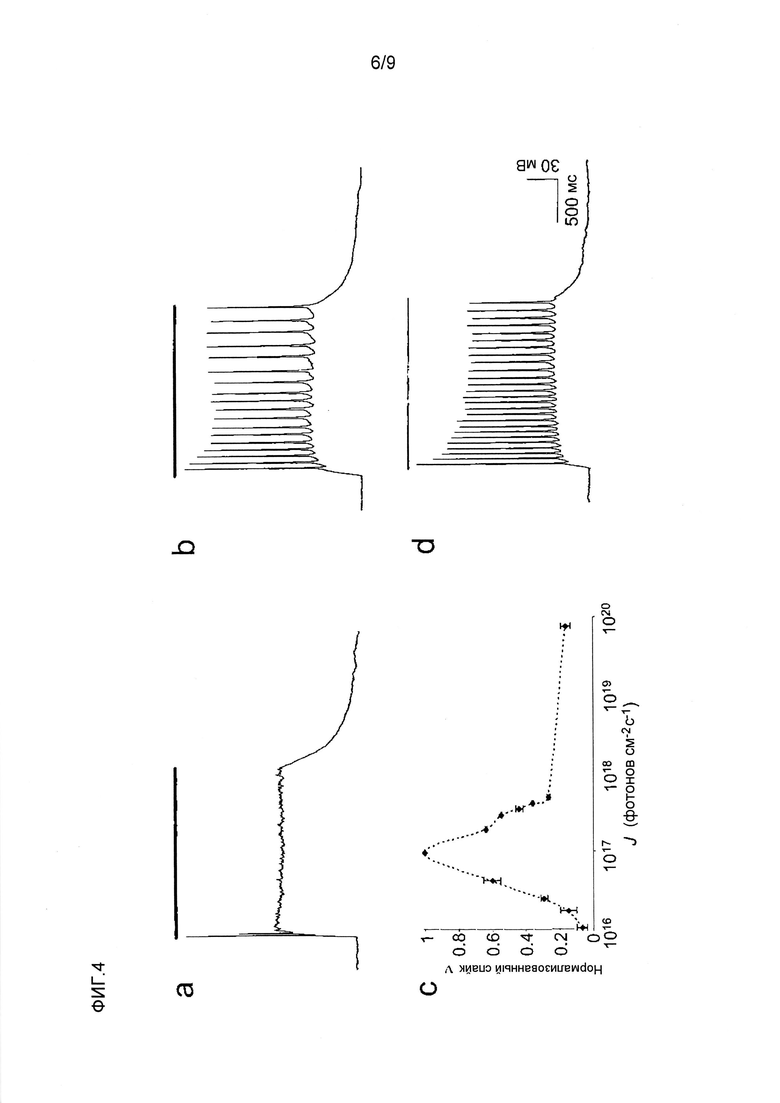
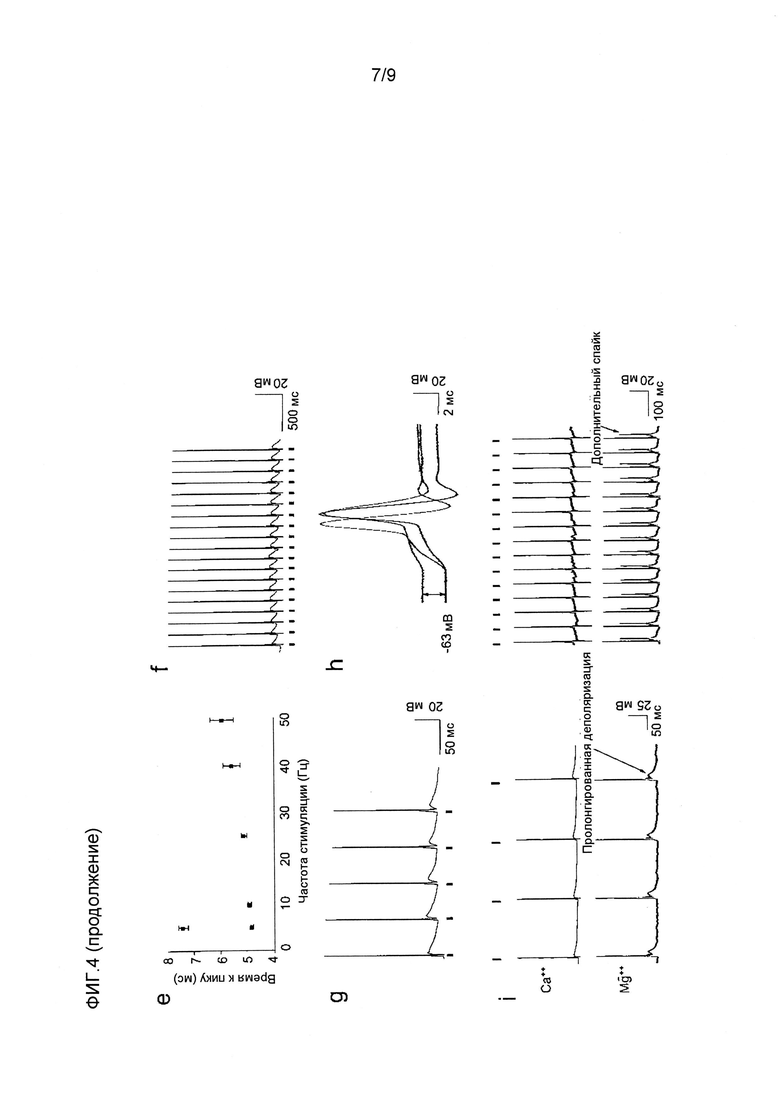
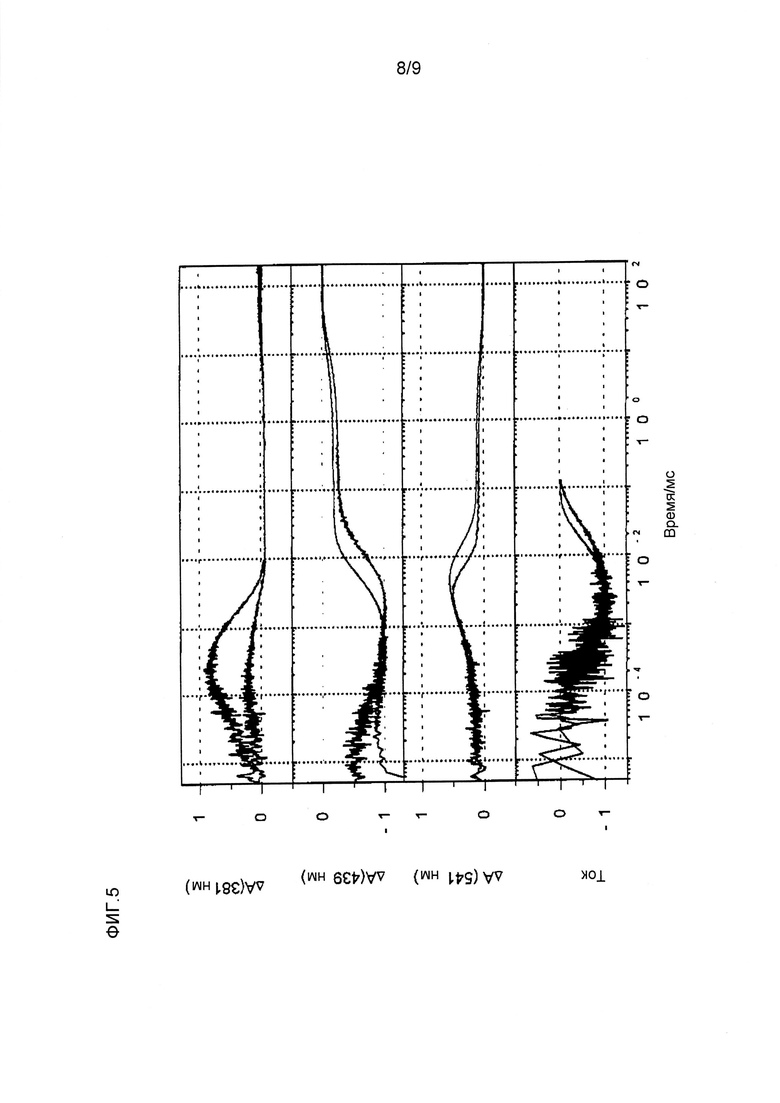
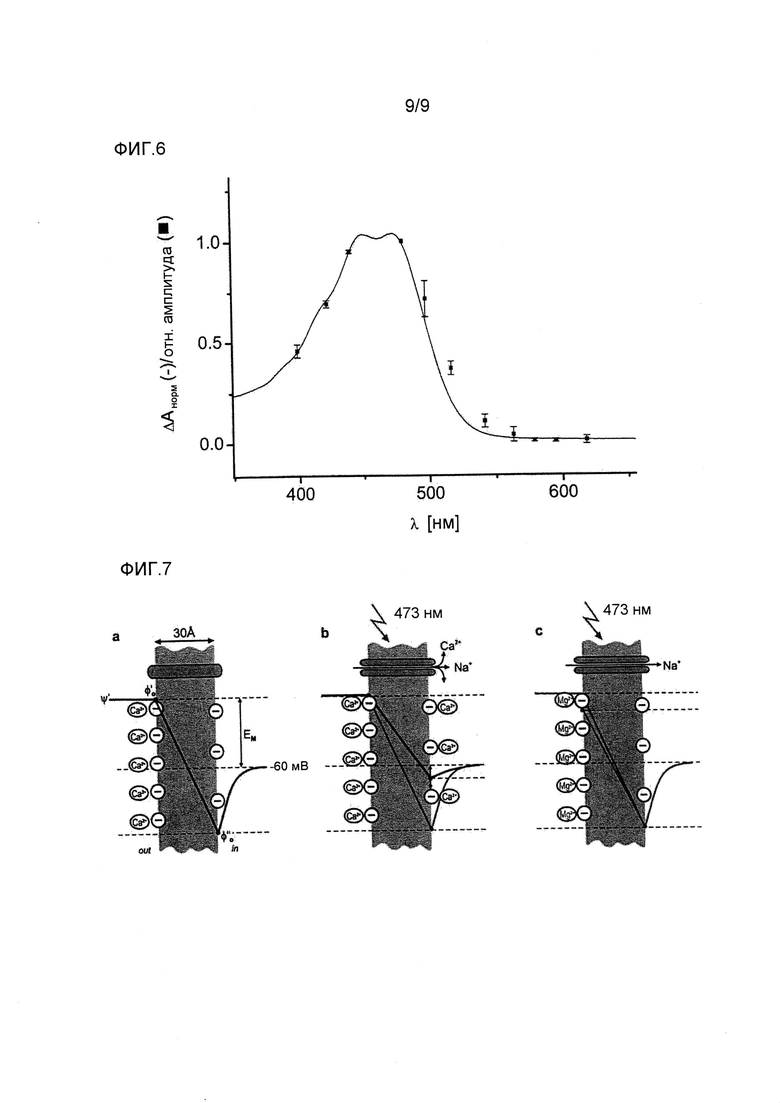






Группа изобретений относится к мутантным канальным родопсинам и к их соответствующим применениям. Активируемый светом ионный канал содержит аминокислотную последовательность, которая по меньшей мере на 70% гомологична аминокислотной последовательности положений 1-309 SEQ ID NO: 1 (СНОР-2) и которая содержит замену в положении, соответствующем L132 в SEQ ID NO: 1. При этом замена повышает полярность канала, и проводимость кальция увеличивается по меньшей мере в два раза по сравнению с диким типом СНОР-2, как определено визуализацией Fura-2 на клетках HEK293. 7 н. и 9 з.п. ф-лы, 7 ил., 1 табл.
1. Активируемый светом ионный канал, где активируемый светом ионный канал содержит аминокислотную последовательность, которая по меньшей мере на 70% гомологична аминокислотной последовательности положений 1-309 SEQ ID NO: 1 (СНОР-2), и которая содержит замену в положении, соответствующем L132 в SEQ ID NO: 1, где замена повышает полярность канала, и
где проводимость кальция увеличивается по меньшей мере в два раза по сравнению с диким типом СНОР-2, как определено визуализацией Fura-2 на клетках HEK293.
2. Активируемый светом ионный канал по п. 1, где активируемый светом ионный канал содержит, предпочтительно, состоит из аминокислотной последовательности положений 1-309 SEQ ID NO: 1 (СНОР-2), за исключением замены в положении L132, где замена повышает полярность канала.
3. Активируемый светом ионный канал по п. 1, где замена выбрана из L132C, L132S, L132E, L132D и L132T, где, предпочтительно, замена представляет собой L132C.
4. Активируемый светом ионный канал по любому из предшествующих пунктов, где
(a) светочувствительность повышена более чем в 5 раз, предпочтительно, более чем в 10 раз, более предпочтительно, более чем в 20 раз, например в 30 раз, еще более предпочтительно, более чем в 40 раз, например в 50 раз, и наиболее предпочтительно, более чем в 60 раз или даже более чем в 70 раз по сравнению с СНОР-2 WT в нейронах гиппокампа, и/или
(b) проводимость кальция повышена по меньшей мере в два раза, предпочтительно, по меньшей мере в три раза, более предпочтительно, по меньшей мере в четыре раза по сравнению с СНОР-2 WT, как определено визуализацией Fura-2 на клетках HEK293, и/или
(c) частота стимуляции увеличена по меньшей мере в 1,5 раза, более предпочтительно, в 2 раза или, даже более предпочтительно, в 2,5 раза по сравнению с СНОР-2 WT, как определено электрофизиологическими регистрациями целой клетки в нейронах гиппокампа.
5. Активируемый светом ионный канал по п. 1, где активируемый светом ионный канал дополнительно содержит по меньшей мере один из следующих аминокислотных остатков: аспарагиновую кислоту в положении, соответствующем положению 253 SEQ ID NO: 1; лизин в положении, соответствующем положению 257 SEQ ID NO: 1; триптофан в положении, соответствующем положению 260 SEQ ID NO: 1; глутаминовую кислоту в положении, соответствующем положению 123 SEQ ID NO: 1; гистидин или аргинин, предпочтительно, аргинин, в положении, соответствующем положению 134 SEQ ID NO: 1; треонин, серин или аланин в положении, соответствующем положению 128 SEQ ID NO: 1; и/или аланин в положении, соответствующем положению 156 SEQ ID NO: 1.
6. Активируемый светом ионный канал по п. 1, где активируемый светом ионный канал содержит консенсусный мотив L(I)DxxxKxxW(F,Y).
7. Канальный родопсин, содержащий активируемый светом ионный канал по пп. 1-6 и ретиналь или производное ретиналя, предпочтительно, где производное ретиналя выбрано из группы, состоящей из 3,4-дегидроретиналя, 13-этилретиналя, 9-диметил-ретиналя, 3-гидроксиретиналя, 4-гидроксиретиналя, нафтилретиналя, 3,7,11-триметил-додека-2,4,6,8; 10-пентаеналя, 3,7-диметил-дека-2,4,6,8-тетраеналя, 3,7-диметил-окта-2,4,6-триеналя и ретиналей с заблокированным вращением в положении 6-7, ретиналей с заблокированным вращением в положении 8-9 и ретиналей с заблокированным вращением в положении 10-11.
8. Конструкция нуклеиновой кислоты, содержащая нуклеотидную последовательность, кодирующую активируемый светом ионный канал по любому из пп. 1-6.
9. Вектор экспрессии, содержащий нуклеотидную последовательность, кодирующую активируемый светом ионный канал по любому из пп. 1-6 или конструкцию нуклеиновой кислоты по п. 8, где, предпочтительно, вектор подходит для генотерапии, в частности, где вектор подходит для опосредованного вирусом переноса генов.
10. Клетка для продукции канального родопсина по п. 7, содержащая канальный родопсин по п. 7, конструкцию нуклеиновой кислоты по п. 8 или вектор экспрессии по п. 9.
11. Клетка по п. 10, где клетка представляет собой клетку млекопитающего или клетку насекомого, или где клетка представляет собой клетку дрожжей, предпочтительно, из Saccharomyces cerevisiae, Schizosaccharomyces pombe или Pichia pastoris.
12. Клетка по п. 11, где клетка млекопитающего представляет собой
(a) фоторецепторную клетку, палочковидную клетку сетчатки, колбочковидную клетку сетчатки, ганглиозную клетку сетчатки, биполярный нейрон, ганглиозную клетку, псевдоуниполярный нейрон, мультиполярный нейрон, пирамидальный нейрон, клетку Пуркинье или гранулярную клетку; или
(b) меланомную клетку, клетку COS, клетку BHK, клетку HEK293, клетку СНО, миеломную клетку или клетку MDCK.
13. Конструкция нуклеиновой кислоты по п. 8, вектор экспрессии по п. 9 или клетка по п. 10 для применения в генотерапии.
14. Применение активируемого светом ионного канала по пп. 1-6, или канального родопсина по п. 7, или клетки по п. 10 в высокопроизводительном скрининге для идентификации модулятора Са2+-активируемого канала.
15. Способ идентификации модулятора Са2+-активируемого (калиевого) канала, включающий следующие стадии:
(i) приведение клетки, экспрессирующей Са2+-активируемый (калиевый) канал и активируемый светом ионный канал по пп. 1-6 или канальный родопсин по п. 7, с средством-кандидатом;
(ii) применение светового стимула для активации активируемого светом канала,
(iii) определение изменения мембранного потенциала (смешанный сигнал) и
(iv) сравнение сигнала, определяемого на стадии (iii), с сигналом, определяемом в клетке, экспрессирующей только активируемый светом ионный канал по пп. 1-6, или канальный родопсин по п. 7, подвергнутых стадии (ii) (единичный сигнал);
уменьшение изменения мембранного потенциала идентифицирует
модулятор Са2+-активируемого (калиевого) канала.
16. Способ по п. 15, где средство-кандидат направлено против Са2+-активируемого канала.
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| АНТИАЛКОГОЛЬНОЕ СРЕДСТВО | 1997 |
|
RU2112510C1 |
| BAMANN C | |||
| et al, Structural guidance of the photocycle of channelrhodopsin-2 by an interhelical hydrogen bond, Biochemistery, 2010 Jan., 49 (2), abstract | |||
| PMID:20000562. | |||

