Область техники, к которой относится изобретение
Настоящее изобретение относится к микровезикуле и способу ее получения.
Уровень техники
Известно, что различные клетки секретируют и высвобождают микровезикулы (небольшие мембранные везикулы), например экзосомы, in vivo. Считалось, что одной из функций микровезикул является внеклеточное высвобождение ненужных внутриклеточных компонентов. Однако в последние годы была показана возможность того, что микровезикулы служат средством распространения сигнала при передаче веществ, таких как белки или липиды, между секретирующими клетками и их клетками-мишенями, а также участвуют в межклеточном взаимодействии.
В настоящее время предлагаются различные клинические применения микровезикул, в частности, экзосом. Например, в Патентном документе 1 раскрыто применение выделенных из ретикулоцитов экзосом, содержащих антиген Plasmodium sp., для защиты от малярии. В Патентном документе 2 раскрыто лечение рака с применением экзосом из B-клеток. В Патентном документе 3 раскрыто применение полученных из стволовых клеток микровезикул в регенерации эндотелия или эпителия.
Лентивирус, например, вирус иммунодефицита человека (ВИЧ), инфицирует клетки и обладает способностью интегрироваться в геном как делящихся, так и неделящихся клеток. Поэтому, лентивирусные векторы, основанные на лентивирусной геномной последовательности, широко используются в качестве инструмента для генной трансдукции.
Список литературы
Патентная литература
Патентный документ 1 Международная публикация WO2011/080271.
Патентный документ 2 Международная публикация WO2011/000551.
Патентный документ 3 Международная публикация WO2009/057165.
Сущность изобретения
Техническая проблема
Целью настоящего изобретения является получение микровезикулы и способ ее получения.
Решение проблемы
Авторы настоящего изобретения провели тщательные исследования для достижения цели данного изобретения, в ходе которых обнаружили, что введение трансгена в клетки при помощи лентивирусного вектора, содержащего трансген под контролем промотора гена теломеразной обратной транскриптазы (TERT), может активировать интеграцию трансгена в геном хозяина и его экспрессию, а также может увеличивать внеклеточное высвобождение микровезикул, содержащих трансгенные продукты и т.п., продуцируемые этими клетками. На основании этих результатов было создано настоящее изобретение.
В частности, настоящее изобретение включает в себя следующее:
[1] Способ получения микровезикул, содержащих трансгенный продукт и/или лентивирусную РНК, содержащую трансген, включающий этапы:
культивирования клетки, в которую при помощи лентивирусного вектора in vitro вводится трансген, для внеклеточного высвобождения микровезикул, содержащих трансгенный продукт и/или лентивирусную РНК, содержащую трансген, где указанный лентивирусный вектор является дефицитным, по меньшей мере, по одному гену структурного белка и содержит трансген под контролем промотора гена теломеразной обратной транскриптазы (TERT) в лентивирусной геномной последовательности, и
сбора высвобожденных микровезикул.
[2] Способ по [1], где у указанной клетки отсутствует указанный, по меньшей мере, один ген структурного белка.
[3] Способ по [1] или [2], где указанный лентивирусный вектор является дефицитным по гену env.
[4] Способ по любому из [1]-[3], где указанный промотор гена теломеразной обратной транскриптазы (TERT) является промотором гена TERT человека.
[5] Способ по [4], где указанный промотор гена TERT человека содержит нуклеотидную последовательность SEQ ID NO: 1 или нуклеотидную последовательность, имеющую 90% или более идентичности последовательности с нуклеотидной последовательностью SEQ ID NO: 1.
[6] Способ по [5], где указанный промотор гена TERT человека содержит нуклеотидную последовательность, имеющую 95% или более идентичности последовательности с нуклеотидной последовательностью SEQ ID NO: 1.
[7] Способ по любому из [1]-[6], где указанная лентивирусная РНК содержит последовательность промотора гена TERT перед трансгеном.
[8] Способ по любому из [1]-[7], где указанный лентивирусный вектор представляет собой:
(i) РНК вектор, содержащий лентивирусную геномную последовательность,
(ii) ДНК вектор, кодирующий РНК, содержащую лентивирусную геномную последовательность, или
(iii) вирусную частицу, несущую РНК, содержащую лентивирусную геномную последовательность.
[9] Способ по любому из [1]-[8], где указанная лентивирусная геномная последовательность содержит, по меньшей мере, часть транскрибируемой области гена TERT между промотором гена TERT и трансгеном.
[10] Способ по [9], где указанная, по меньшей мере, часть транскрибируемой области гена TERT содержит нуклеотидную последовательность SEQ ID NO: 2 или нуклеотидную последовательность, имеющую 90% или более идентичности последовательности с нуклеотидной последовательностью SEQ ID NO: 2.
[11] Способ по любому из [1]-[10], где указанная лентивирусная геномная последовательность представляет собой геномную последовательность ВИЧ.
[12] Способ по [11], где указанная геномная последовательность ВИЧ представляет собой геномную последовательность ВИЧ-1.
[13] Способ по [12], где указанная геномная последовательность ВИЧ-1 содержит 5’-LTR; сигнал упаковки ψ; ген gag; ген pol; ген vif; ген vpr; ген tat; ген rev; ген vpu; и 3’-LTR.
[14] Способ по любому из [1]-[13], где указанный трансген кодирует белок или РНК.
[15] Способ по любому из [1]-[14], где указанный лентивирусный вектор содержит указанный трансген, представляющий собой ген-супрессор опухолевого роста.
[16] Способ по [15], где указанный ген-супрессор опухолевого роста представляет собой ген PTEN или p16.
[17] Способ по любому из [1]-[14], где указанный лентивирусный вектор содержит указанный трансген, который кодирует кшРНК.
[18] Способ по [17], где указанная кшРНК специфична по отношению к гену, кодирующему регулятор клеточной пролиферации.
[19] Способ по [18], где указанный регулятор клеточной пролиферации представляет собой CDC6.
[20] Способ по любому из [1]-[19], где указанная клетка является клеткой человека.
[21] Способ по любому из [1]-[20], где указанная клетка является клеткой, выделенной из почки.
[22] Способ по любому из [1]-[21], где указанная клетка является клеткой клеточной линии 293Т эмбриональной почки человека.
[23] Микровезикула, содержащая трансгенный продукт и/или лентивирусную РНК, содержащую трансген, где указанная микровезикула получена способом по любому из [1]-[22].
[24] Способ генной трансдукции, включающий приведение клетки-мишени в контакт с микровезикулой, содержащей трансгенный продукт и/или лентивирусную РНК, содержащую трансген, по [23] для их слияния, чем обеспечивается введение трансгена в клетку.
[25] Способ по [24], где указанная клетка-мишень приводится в контакт с микровезикулой in vitro.
[26] Композиция, содержащая микровезикулу по [23].
[27] Фармацевтическая композиция, содержащая микровезикулу по [23].
[28] Фармацевтическая композиция по [27], которая предназначена для применения в лечении рака.
[29] Фармацевтическая композиция по [28], где указанный рак выбран из группы, состоящей из рака толстой кишки, рака поджелудочной железы, рака почки, рака легкого, нейробластомы, рака молочной железы, рака яичников, рака желудка, рака предстательной железы, рак щитовидной железы и злокачественной лимфомы.
[30] Фармацевтическая композиция по [28] или [29], где указанный рак включает повышенную экспрессию CDC6.
[31] Фармацевтическая композиция по любому из [27]-[30], дополнительно содержащая фармацевтически приемлемый носитель.
[32] Способ лечения пациента, включающий введение микровезикулы по [23] указанному пациенту, нуждающемуся в введении указанного трансгена или указанного трансгенного продукта.
[33] Способ по [32], где указанный пациент страдает от рака.
[34] Способ по [33], где указанный рак выбран из группы, состоящей из рака толстой кишки, рака поджелудочной железы, рака почки, рака легкого, нейробластомы, рака молочной железы, рака яичников, рака желудка, рака предстательной железы, рак щитовидной железы и злокачественной лимфомы.
[35] Способ по [33] или [34], где указанный рак включает повышенную экспрессию CDC6.
Настоящее описание включает в себя содержание Предварительных заявок на патент США Nos. US 61/779556 и US 61/894563, приоритет по которым испрашивается в настоящей заявке.
Эффекты изобретения
Данное изобретение предоставляет микровезикулу и способ ее получения.
Краткое описание чертежей
Фигура 1 представляет собой схематическое изображение геномной области ВИЧ-1 плазмид, имеющих ВИЧ-1 основу. Фигура 1A представляет собой схематичное изображение геномной области ВИЧ-1 плазмиды pNL4-3. Фигура 1B представляет собой схематичное изображение геномной области ВИЧ-1 плазмиды pTHTK.
На Фигуре 2 показаны рекомбинантные лентивирусные плазмидные векторы pRBL0213T и pTHTN. Фигура 2A представляет собой электрофореграмму агарозного геля, показывающую картирование плазмидной ДНК pRBL0213T и pTHTN. Дорожка 1: pRBL0213T, EcoR I; Дорожка 2: pRBL0213T, Mlu I + Xho I; Дорожка 3: pRBL0213T, Nhe I; Дорожка 4: pTHTN, EcoR I; Дорожка 5: pTHTN, Mlu I + Xho I; Дорожка M: 1 kb маркер молекулярной массы. Фигура 2B представляет собой схематическое изображение геномной области ВИЧ-1 плазмиды pRBL0213T. Фигура 2C представляет собой схематическое изображение геномной области ВИЧ-1 плазмиды pTHTN.
На Фигуре 3 показан рекомбинантный лентивирусный плазмидный вектор pRBL001. Фигура 3A представляет собой электрофореграмму агарозного геля, показывающую картирование плазмидной ДНК pRBL001. Дорожка 1: pRBL001, EcoR I; Дорожка 2: pRBL001, EcoR I + Nhe I; Дорожка 3: pRBL001, Mlu I + Xho I; Дорожка 4: pRBL001, Sal I; Дорожка 5: pRBL0213T, Nhe I; Дорожка M: 1 kb маркер молекулярной массы. Фигура 3B представляет собой схематическое изображение геномной области ВИЧ-1 плазмиды pRBL001. Сайты ферментов рестрикции обозначены следующим образом: RI: EcoR I, Sal: Sal I, Mlu: Mlu I, Nhe: Nhe I и Xho: Xho I.
Фигура 4 представляет собой схематическое изображение геномной области ВИЧ-1 рекомбинантных лентивирусных плазмидных векторов. Фигура 4A представляет собой схематическое изображение геномной области ВИЧ-1 плазмиды pNL4-3 (дикий тип ВИЧ-1). Фигура 4B представляет собой схематическое изображение геномной области ВИЧ-1 плазмиды pD64V, которая имеет мутацию интегразы ВИЧ-1 и является дефицитной по интеграции геномной ДНК вируса ВИЧ-1 в геном хозяина. Фигура 4C представляет собой схематическое изображение геномной области ВИЧ-1 плазмиды pTHTK. Фигура 4D представляет собой схематическое изображение геномной области ВИЧ-1 плазмиды pTHTN. Фигура 4E представляет собой схематическое изображение геномной области ВИЧ-1 плазмиды pTHTH, у которой отсутствует 5’-часть промотора hTERT. Фигура 4F представляет собой схематическое изображение геномной области ВИЧ-1 плазмиды pTHTC, имеющей промоторную последовательность цитомегаловируса человека (ЦМВ).
Фигура 5 представляет собой схему, показывающую получение сайта Bpm I для обнаружения LTR-метки. Фигура 5A представляет собой схему, показывающую новый сайт Bpm I, полученный в 5’-LTR геномной области ВИЧ-1 плазмиды pTHTK. Фигура 5B представляет собой схему, показывающую, что фермент рестрикции Bpm I расщепляет хромосомную ДНК хозяина в сайте интеграции (14 нуклеотидов), примыкающем к сайту Bpm I, скопированному в 3’-LTR ВИЧ-1. Фигура 5C представляет собой схематическое изображение концевой структуры лентивирусного генома.
Фигура 6 представляет собой схему, показывающую получение LTR-метки при помощи опосредованной лигированием ПЦР (ОЛ-ПЦР).
Фигура 7 представляет собой электрофореграмму геля результатов ОЛ-ПЦР для обнаружения LTR-метки.
На Фигуре 8 показаны результаты изучения сплайсинга вирусной РНК. Фигуры 8A и 8B представляют собой фотографии, показывающие результаты электрофореза продуктов ОТ-ПЦР в 2,2% агарозном геле в 1x TAE буфере. Фигура 8A представляет собой электрофореграмму, показывающую ПЦР продукты, полученные с использованием праймеров 5’LTRU5 и 3’NL8960. Фигура 8B представляет собой электрофореграмму, показывающую ПЦР продукты, полученные с использованием праймеров 5’LTRU5 и 3’NL5850. Положения зон сплайсированных фрагментов РНК a (E1+A1/3’NL5850), b (E1+A2/3’NL5850), c1 (E1+E2+E3+A3/3’NL5850), c2 (E1+E2+A3/3’NL5850) и c3 (E1+E3+A3/3’NL5850) показаны с правой стороны Фигуры 8B. Фигура 8C представляет собой схематическое изображение несплайсированного транскрипта. Фигура 8D представляет собой схематическое изображение сплайсированной РНК. На Фигуре 8D A1 - A3 обозначают 3’акцепторные сайты сплайсинга, и E1 - E3 обозначают экзоны.
Фигура 9 представляет собой диаграмму, показывающую результаты анализа p24.
Фигура 10 представляет собой схему, показывающую строение плазмидных векторов, использованных в анализе. Фигура 10A показывает pGL3-1375 (Empt.; размер плазмиды: приблизительно 7 т.п.н.) в качестве отрицательного контроля при трансдукции гена PTEN. Фигура 10B показывает pcDNA3.1/CMV-hPTEN (Regul.; размер плазмиды: приблизительно 7,5 т.п.н.) в качестве положительного контроля при трансдукции гена PTEN. Фигура 10C показывает ретровирусный вектор pRBL016Bn (Retro.; размер плазмиды: приблизительно 7,9 т.п.н.) для трансдукции гена PTEN. Фигура 10D показывает рекомбинантный лентивирусный вектор pRBL0213T (Lenti.; размер плазмиды: приблизительно 15 т.п.н.) для трансдукции гена PTEN. Сайты ферментов рестрикции в векторах обозначены следующим образом: BamH: BamH I; Bgl: Bgl II; RI: EcoR I; RV: EcoR V; Hd3: Hind III; Mlu: Mlu I; Nco: Nco I; Nde: Nde I; Sal: Sal I; Stu: Stu I; и Xho: Xho I.
Фигура 11 представляет собой диаграмму, показывающую результаты анализа PTEN при транзиентной трансфекции плазмидным вектором.
Фигура 12 представляет собой диаграмму, показывающую отношение активности PTEN в лизатах МВ к активности PTEN в клеточных лизатах, показанных на Фигуре 11.
Фигура 13 представляет собой диаграмму, показывающую результаты анализа PTEN при инфекции рекомбинантной лентивирусной частицей RBL0213T.
Фигура 14 представляет собой диаграмму, показывающую отношение активности PTEN в МВ лизатах к активности PTEN в клеточных лизатах клеток, трансфицированных рекомбинантным лентивирусным вектором pRBL0213T, или клеток, инфицированных рекомбинантной лентивирусной частицей RBL0213T.
Фигура 15 представляет собой фотографию, показывающую результаты Вестерн-блоттинга вирусных белков или эндогенных или чужеродных белков в МВ или в клетках, инкубированных с МВ. На Фигуре 15A показаны результаты, полученные с использованием анти-Vpu антител. На Фигуре 15B показаны результаты, полученные с использованием анти-RT антител. На Фигуре 15C показаны результаты, полученные с использованием анти-FEN-1 антител.
Фигура 16 представляет собой набор микрофотографий, показывающих доставку содержимого в клетки при помощи МВ, в которые был инкапсулирован c-myc-FEN-1. Фигура 16A представляет собой изображение, после окрашивания анти-c-myc антителами. Фигура 16B представляет собой изображение, после окрашивания ДАПИ. Фигура 16C представляет собой наложенное изображение Фигур 16A и 16B. Фигура 16D представляет собой изображение, полученное из наложенного изображения при помощи программы кривой изменения цвета в методе «настройки» изображения программы Photoshop(R) (Adobe Systems Inc.).
Фигура 17 представляет собой график, показывающий подавляющее действие Lenti-mv2010/CDC6 кшРНК на пролиферацию раковых клеток.
Фигура 18 представляет собой фотографию результатов Вестерн-блоттинга экстрактов клеточных белков и МВ лизатов.
Фигура 19 представляет собой фотографии, показывающие типичные морфологии, наблюдаемые для опухолей из соответствующих экспериментальных групп по результатам патологического исследования. A, группа отрицательного контроля; B, группа, получавшая инъекцию интерферона (ИФН) α-2b; C, группа, получавшая инъекцию Цитомокс PTEN; и D, группа, получавшая инъекцию Цитомокс p53.
Фигура 20 представляет собой фотографии, показывающие типичные морфологии, наблюдаемые для опухолей из соответствующих экспериментальных групп по результатам патологического исследования. A, группа, получавшая инъекцию низкой дозы Цитомокс EX; B, группа, получавшая инъекцию высокой дозы Цитомокс EX; C, группа, получавшая инъекцию низкой дозы Цитомокс HD; и D, группа, получавшая инъекцию высокой дозы Цитомокс HD.
Описание вариантов осуществления изобретения
Далее настоящее изобретение будет описано более подробно.
1. Способ получения микровезикул
Настоящее изобретение относится к способу получения микровезикул. Способ настоящего изобретения включает в себя этапы:
культивирования клетки, в которую при помощи лентивирусного вектора in vitro вводится трансген, для внеклеточного высвобождения микровезикул, содержащих трансгенный продукт и/или лентивирусную РНК, содержащую трансген, где указанный лентивирусный вектор является дефицитным, по меньшей мере, по одному гену структурного белка и содержит трансген под контролем промотора гена теломеразной обратной транскриптазы (TERT) в лентивирусной геномной последовательности, и
сбора высвобожденных микровезикул.
Микровезикулы, полученные способом настоящего изобретения, содержат трансгенный продукт и/или лентивирусную РНК, содержащую трансген. Способом настоящего изобретения можно эффективно получать микровезикулы, содержащие трансгенный продукт и/или лентивирусную РНК, содержащую трансген.
Термин «лентивирусный вектор» в контексте настоящего изобретения относится к вектору для генной трансдукции, имеющему в качестве основы лентивирусную геномную последовательность. Лентивирус представляет собой РНК вирус, имеющий обратную транскриптазу. Лентивирус может интегрировать вирусную геномную ДНК (провирусную ДНК) в хромосомы хозяина не только делящихся клеток, но и неделящихся клеток, таким образом, что вирусные гены могут экспрессироваться этими клетками-хозяевами. Лентивирусный вектор основан на этих свойствах лентивирусов.
Лентивирусный вектор настоящего изобретения может представлять собой:
(i) РНК вектор, содержащий лентивирусную геномную последовательность,
(ii) ДНК вектор, кодирующий РНК, содержащую лентивирусную геномную последовательность, или
(iii) вирусную частицу, несущую РНК, содержащую лентивирусную геномную последовательность.
РНК вектор по пункту (i) может быть получен, например, транскрипции in vitro с экспрессионного вектора или экспрессионной кассеты, содержащих лентивирусную геномную последовательность. ДНК вектор по пункту (ii) может включать в себя плазмидный вектор. Плазмидный вектор, как правило, содержит последовательность ДНК, кодирующую РНК, содержащую лентивирусную геномную последовательность, а также промотор и терминатор для осуществления транскрипции последовательности ДНК, сайт инициации репликации и маркерный ген для скрининга рекомбинантов и т.п. Такой ДНК вектор может быть получен известным в данной области техники способом с применением рекомбинантной технологии или тому подобного. Вирусная частица по пункту (iii) может представлять собой вирусную частицу, псевдотипированную белком оболочки другого вируса, например, гликопротеином оболочки G (ВВС-G) вируса везикулярного стоматита (ВВС). Псевдотипированная вирусная частица может быть получена, например, путем котрансфекции культивируемой клетки ДНК вектором, кодирующим РНК, содержащую лентивирусную геномную последовательность, и плазмидой, кодирующей белок оболочки (например, ВВС-G) другого вируса; сбора высвобождаемых в культуральную среду вирусных частиц; и очистки этих частиц.
Используемый в настоящем изобретении лентивирусный вектор является дефицитным, по меньшей мере, по одному гену вирусного структурного белка в своей лентивирусной геномной последовательности. Такая лентивирусная геномная последовательность может представлять собой последовательность, в которой, по меньшей мере, один ген вирусного структурного белка был нарушен (например, путем делеции части или всей области, или путем вставки молекулы нуклеиновой кислоты) в полноразмерной лентивирусной геномной последовательности. В настоящем изобретении термин «дефицитный», употребляемый по отношению к гену, означает, что целый ген удален, или ген нарушен или мутирован таким образом, что функциональный белок не может экспрессироваться. Ген вирусного структурного белка, по которому лентивирусный вектор является дефицитным, может представлять собой, по меньшей мере, ген, выбранный из группы, состоящей из генов gag, pol и env. Ген gag кодирует белок, принимающий участие в формировании вирусной частицы. Ген pol кодирует фермент, такой как обратная транскриптаза (ОТ). Ген env кодирует белок оболочки, принимающий участие в адсорбции на и проникновении в клетки-хозяева. Например, лентивирусный вектор может быть дефицитным по гену env. Например, область, расположенная между положением 6344 и положением 7611 SEQ ID NO: 4 может быть удалена из лентивирусного вектора, что приведет к дефицитности по гену env ВИЧ-1. В одном варианте осуществления изобретения клетка не содержит в геноме или за пределами хромосомы, по меньшей мере, один ген вирусного структурного белка, по которому лентивирусный вектор является дефицитным. Такая клетка, в которую при помощи лентивирусного вектора был введен трансген, не продуцирует инфекционные лентивирусные частицы. Таким образом, продуцируемые микровезикулы являются в высокой степени безопасными.
Примеры лентивирусной геномной последовательности, используемой в качестве основы в лентивирусном векторе настоящего изобретения, включают в себя без ограничений последовательности из геномов вируса иммунодефицита человека (ВИЧ), вируса иммунодефицита обезьян (ВИО), вируса иммунодефицита кошек (ВИК) и вируса иммунодефицита собак (ВИС). Лентивирусная геномная последовательность в соответствии с настоящим изобретением предпочтительно представляет собой геномную последовательность ВИЧ. В частности, геномная последовательность ВИЧ может представлять собой геномную последовательность ВИЧ-1 или ВИЧ-2. Предпочтительно, геномная последовательность ВИЧ может представлять собой геномную последовательность ВИЧ-1. ВИЧ может представлять собой штамм, принадлежащий к ВИЧ-1 группы M, N, O или P. В частности, ВИЧ может представлять собой любой из штаммов ВИЧ, включая ВИЧ-1 IIIb, ВИЧ-1 SF2, ВИЧ-1 SF162, ВИЧ-1 BRU, ВИЧ-1 NY5, ВИЧ-1L AI, ВИЧ-1 NL4-3 и т.п. Примером геномной последовательности ВИЧ могут являться последовательности, зарегистрированные в базе данных Genbank под номерами EU541617, K03455 и K02013 и т.п. Лентивирусная геномная последовательность может представлять собой РНК или может представлять собой ДНК.
Лентивирусная геномная последовательность в лентивирусном векторе может содержать 5’-LTR и 3’-LTR и при необходимости, по меньшей мере, один элемент, выбранный из группы, состоящей из сигнала упаковки ψ, гена gag, гена pol, гена vif, гена vpr, гена tat, гена rev, гена vpu и гена vpx. В предпочтительном варианте осуществления изобретения лентивирусная геномная последовательность ВИЧ-1 в лентивирусном векторе может содержать 5’-LTR, сигнал упаковки ψ, ген gag, ген pol, ген vif, ген vpr, ген tat, ген rev, ген vpu и 3’-LTR. В другом предпочтительном варианте осуществления изобретения лентивирусная геномная последовательность ВИЧ-2 в лентивирусном векторе может содержать 5’-LTR, сигнал упаковки ψ, ген gag, ген pol, ген vif, ген vpr, ген tat, ген rev, ген vpx и 3’-LTR. Такая лентивирусная геномная последовательность, как правило, содержит, по меньшей мере, один донорный сайт сплайсинга (ДСС) и акцепторный сайт сплайсинга (АСС). В одном варианте осуществления изобретения лентивирусная геномная последовательность в лентивирусном векторе может быть дефицитной по гену nef.
Лентивирусный вектор настоящего изобретения содержит трансген под контролем промотора гена теломеразной обратной транскриптазы (TERT) в лентивирусной геномной последовательности. TERT является ферментом, который синтезирует теломерный повтор во время репликации ДНК. Промотор гена теломеразной обратной транскриптазы (TERT), используемый в настоящем изобретении, может представлять собой промотор гена TERT человека, но не ограничивается им. Предпочтительно, промотор гена TERT человека может содержать нуклеотидную последовательность SEQ ID NO: 1, или нуклеотидную последовательность, имеющую 90% или более идентичности последовательности с нуклеотидной последовательностью SEQ ID NO: 1. Более предпочтительно, промотор гена TERT человека может содержать нуклеотидную последовательность, имеющую 95%, 97%, 99%, 99,5% или 99,9% или более идентичности последовательности с нуклеотидной последовательностью SEQ ID NO: 1.
В одном также предпочтительном варианте осуществления изобретения лентивирусный вектор настоящего изобретения содержит нуклеотидную последовательность SEQ ID NO: 5, или последовательность, имеющую 90% или более, предпочтительно 95% или более, более предпочтительно 99% или более, например, 99,8% или более идентичности с ней по последовательности, в качестве лентивирусной геномной последовательности; и последовательность, содержащую промотор гена TERT, и трансген, находящийся под его контролем, который был дополнительно встроен в эту лентивирусную геномную последовательность (предпочтительно в ген nef).
В настоящем изобретении термин «трансген под контролем промотора гена TERT» означает, что транскрипция трансгена инициируется активностью промотора гена TERT. Предпочтительно, трансген расположен после промотора гена TERT.
По меньшей мере, часть транскрибируемой области гена TERT может находиться между промотором гена TERT и трансгеном, встроенным в лентивирусную геномную последовательность. По меньшей мере, часть транскрибируемой области гена TERT может содержать, по меньшей мере, первый экзон гена TERT. По меньшей мере, часть транскрибируемой области гена TERT может содержать нуклеотидную последовательность SEQ ID NO: 2, или нуклеотидную последовательность, имеющую 90%, 95%, 97%, 99% или 99,5% или более идентичности последовательности с нуклеотидной последовательностью SEQ ID NO: 2.
Используемый в настоящем изобретении трансген может кодировать любой белок или РНК, такую как микроРНК, малую интерферирующую РНК (миРНК) или короткую шпилечную РНК (кшРНК). В одном варианте осуществления изобретения трансген может представлять собой ген-супрессор опухолевого роста. Примеры гена-супрессора опухолевого роста включают в себя без ограничений гены-супрессоры опухолевого роста, известные специалистам в данной области техники, такие как гены p53, BRCA1, Rb, PTEN и p16. Предпочтительно, ген-супрессор опухолевого роста может представлять собой ген PTEN или p16. Белок PTEN представляет собой фосфатазу, которая катализирует дефосфорилирование фосфатидилинозитол 3,4,5-трифосфата (PtdIns(3,4,5)P3). Ген PTEN может содержать нуклеотидную последовательность SEQ ID NO: 6, или нуклеотидную последовательность, имеющую 90% или более, предпочтительно 95% или более, более предпочтительно 99% или более, например, 99,5% или 99,9% или более идентичности с ней по последовательности. Ген PTEN также может представлять собой нуклеиновую кислоту, кодирующую белок PTEN, который состоит из аминокислотной последовательности SEQ ID NO: 22 (регистрационные номера в GenBank AAD13528 и NP_000305), или аминокислотной последовательности, имеющей 90% или более, предпочтительно 95% или более, более предпочтительно 99% или более, например, 99,5% или 99,7% или более идентичности с ней по последовательности. Белок p16 (также именуемый p16INK4a) известен как ингибитор циклинзависимой киназы (ЦЗК). Например, ген p16 может содержать нуклеотидную последовательность SEQ ID NO: 23 (номер регистрации в GenBank L27211), или нуклеотидную последовательность, имеющую 80% или более, предпочтительно 90% или более, более предпочтительно 95% или более, еще более предпочтительно 99% или более, например, 99,5% или 99,9% или более идентичности с ней по последовательности. Ген p16 также может представлять собой нуклеиновую кислоту, кодирующую белок состоящий из аминокислотной последовательности SEQ ID NO: 24, или аминокислотной последовательности, имеющей 90% или более, предпочтительно 95% или более, более предпочтительно 99% или более идентичности с ней по последовательности. Ген p16 предпочтительно кодирует белок, обладающий ЦЗК-ингибирующей активностью. Примеры гена p16 включают в себя нуклеиновую кислоту, содержащую нуклеотидную последовательность от положения 434 до положения 480 в SEQ ID NO: 25. Кроме того, ген p53 может содержать нуклеотидную последовательность SEQ ID NO: 26 (номер регистрации в GenBank BC003596), или нуклеотидную последовательность, имеющую 80% или более, предпочтительно 90% или более, более предпочтительно 95% или более, еще более предпочтительно 99% или более, например, 99,5% или 99,9% или более идентичности с ней по последовательности. Ген p53 также может представлять собой нуклеиновую кислоту, кодирующую белок, состоящий из аминокислотной последовательности SEQ ID NO: 26, или аминокислотной последовательности, имеющей 90% или более, предпочтительно 95% или более, более предпочтительно 99% или более идентичности с ней по последовательности. Ген p53 предпочтительно кодирует белок, обладающий активностью фактора транскрипции. В контексте настоящего изобретения нуклеиновая кислота может представлять собой ДНК или РНК и может содержать модифицированное основание.
В другом варианте осуществления изобретения трансген может кодировать кшРНК. кшРНК представляет собой одноцепочечную РНК, у которой антисмысловая последовательность, комплементарная последовательности-мишени, и смысловая последовательность (как правило, имеющая выступающую поли-U последовательность на 3’-конце), комплементараная антисмысловой последовательности, соединены через линкер и образуют шпилечную структуру за счет образования внутримолекулярной двухцепочечной структуры. Такая кшРНК внутри клетки расщепляется в своей двухцепочечной части на миРНК, которые, в свою очередь, вызывают РНК-интерференцию, подавляющую экспрессию целевого гена, содержащего последовательность-мишень. Предшественник кшРНК может транскрибироваться с трансгена и затем подвергаться редактированию и процессингу с образованием кшРНК. Мишень для кшРНК может включать в себя без ограничений ген, кодирующий регулятор клеточной пролиферации. Примеры регулятора клеточной пролиферации включают в себя белки, участвующие в репликации ДНК или регуляции клеточного цикла, такие как CDC6, циклин E, CDK2, CDT1, ORC2 и MCM7. Предпочтительно, регулятор клеточной пролиферации может представлять собой CDC6. CDC6 представляет собой белок, который играет центральную роль в инициации репликации ДНК. Было обнаружено, что нокдаун CDC6 приводит к апоптозу раковых клеток человека (Feng, et al., Cancer Res., 2003, Vol. 63, p. 7356-7364; Lau et al., EMBO Rep., 2006, Vol. 7, p. 425-430; and Feng et al., Mol. Cell. Biochem., 2008, Vol. 311, p. 189-197). Ген CDC6 может содержать нуклеотидную последовательность SEQ ID NO: 7, или нуклеотидную последовательность, имеющую 90% или более, предпочтительно 95% или более, более предпочтительно 99% или более, например, 99,5% или 99,9% или более идентичности с ней по последовательности. Трансген, кодирующий кшРНК CDC6, может содержать последовательность ДНК, состоящую из нуклеотидной последовательности SEQ ID NO: 10, или нуклеотидной последовательности, имеющей 90% или более, предпочтительно 95% или более, более предпочтительно 98% или более идентичности с ней по последовательности. Как правило, кшРНК CDC6 содержит: антисмысловую последовательность кшРНК CDC6, состоящую из нуклеотидной последовательности SEQ ID NO: 19, или нуклеотидной последовательности, имеющей 90% или более идентичности с ней по последовательности; линкер, состоящий из нуклеотидной последовательности SEQ ID NO: 20, или нуклеотидной последовательности, имеющей 80% или более идентичности с ней по последовательности; и смысловой последовательности, состоящей из последовательности, комплементарной антисмысловой последовательности, и выступающей поли-U последовательности на 3’-конце из двух или более оснований. Трансгенный продукт может содержать РНК из трансгена, ка описано выше, или белок, транслированный из РНК из трансгена.
Каждый лентивирусный вектор может содержать один или несколько трансгенов. Два или более трансгена могут быть введены в клетку при помощи одного лентивирусного вектора или при помощи двух или нескольких лентивирусных векторов. Например, лентивирусный вектор может содержать трансген, который кодирует кшРНК, и трансген, который представляет собой ген-супрессор опухолевого роста. В предпочтительном варианте осуществления изобретения лентивирусный вектор содержит трансген, кодирующий кшРНК CDC6, и/или трансген, кодирующий белок p16.
В соответствии с настоящим изобретением трансген вводится в клетку при помощи лентивирусного вектора in vitro. Вид организма, от которого получена клетка, используемая в настоящем изобретении, предпочтительно является тем же видом, к которому относится реципиент получаемых микровезикул. Клетка может представлять собой, но без ограничений, клетку млекопитающего, например, клетку собаки, кошки, коровы, овцы, мыши, крысы или примата, такого как обезьяна или человек. Клетка человека является предпочтительной. Также предпочтительно, чтобы клетка была клеткой, выделенной из почки, клеткой, выделенной из матки, клеткой, имеющей лимфоцитарное происхождение, или фибробластом. Клетка может включать в себя клетки клеточной линии 293Т эмбриональной почки человека, клетки клеточной линии HeLa рака шейки матки человека, клетки лимфоцитарной клеточной линии CEM человека, фибробласты линии N144 и другие клеточные линии человека. Предпочтительно, клетка может представлять собой клетку клеточной линии 293Т эмбриональной почки человека или клетку клеточной линии HeLa рака шейки матки человека.
Введение трансгена в клетку при помощи лентивирусного вектора может быть выполнено способом, известным в данной области техники. Введение трансгена в клетку с использованием в качестве лентивирусного вектора РНК вектора или ДНК вектора может быть выполнено кальциево-фосфатным методом, методом липофекции, диэтиламиноэтилдекстрановым методом или методом электропорации или тому подобным методом, известным в данной области техники, и может быть выполнено с использованием коммерчески доступных реагентов для трансфекции, таких как Lipofectamine® 2000 (Invitrogen) и FuGene® 6 (Roche). Введение трансгена в клетку с использованием в качестве лентивирусного вектора вирусной частицы может быть выполнено путем добавления вирусной частицы в культуральную среду клетки для инфицирования этой клетки вирусом.
Кроме лентивирусного вектора в клетку также может быть введен дополнительный экспрессионный вектор. Такой экспрессионный вектор может содержать трансген, кодирующий белок или РНК, которые в дальнейшем инкапсулируются в микровезикулы, под контролем промотора. Клетку, в которую таким образом был введен трансген, затем культивируют. Культивирование может быть осуществлено при помощи любого подходящего для клетки способа. Например, культивирование может проводиться в течение 1-5 суток, например, 2-4 суток, в частности, от 36 часов до 96 часов, или от 36 часов до 72 часов в полной среде DMEM с высоким содержанием глюкозы с добавлением 10% фетальной бычьей сыворотки и соответствующего антибиотика. В клетке, в которую трансген был введен при помощи лентивирусного вектора, лентивирусная ДНК (например, получаемая из лентивирусной РНК под действием обратной транскриптазы) интегрируется во внутриклеточный геном по действием интегразы или тому подобного фермента для образования провирусной ДНК. С этой провирусной ДНК лентивирусная РНК продуцируется путем транскрипции с участием внутриклеточной РНК-полимеразы II. Из полученной таким образом лентивирусной РНК трансгенный продукт, как правило, образуется путем сплайсинга РНК и/или трансляции белка и т.п. Трансгенный продукт и/или лентивирусная РНК, продуцируемые таким образом в клетке, инкорпорируются в микровезикулы. Клетка высвобождает такие микровезикулы наружу во время культивирования, и таким образом микровезикулы накапливаются в среде. Предпочтительно, лентивирусный вектор, используемый в настоящем изобретении, является дефицитным по гену структурного белка, и поэтому он не образует вирус-подобных частиц и не обеспечивает накопление вирус-подобных частиц в среде.
Способ получения микровезикул настоящего изобретения включает в себя этап сбора высвобожденных микровезикул. Сбор микровезикул может быть выполнен путем сбора клеточной культуральной среды. Микровезикулы, собранные таким образом, затем могут быть очищены. Например, крупные везикулы могут быть осаждены и удалены путем центрифугирования при 1000-10000×g (в предпочтительном варианте осуществления изобретения при 9000×g), благодаря чему происходит очистка собранных микровезикул. Такое центрифугирование может быть выполнено один или несколько раз (предпочтительно с различной центробежной силой). Альтернативно или дополнительно, например, микровезикулы могут быть очищены путем ультрафильтрации с использованием мембраны с отсечением по молекулярной массе 1000 кДа. Кроме того, микровезикулы также могут быть осаждены путем смешивания с раствором ПЭГ/NaCl и центрифугирования, чем достигается очистка микровезикул. После такой очистки микровезикулы могут быть пегилированы. Пегилирование микровезикул может быть выполнено с использованием различных типов пегилирующих реагентов, и может быть выполнено с использованием, например, метокси ПЭГ сукцинимидилкарбоната NHS (mPEG-NHS) (Croyle et al., J. Virol., 2004, Vol. 78, p. 912-921). Например, метокси ПЭГ сукцинимидилкарбонат NHS (mPEG-NHS, м.м. 10К (NANOCS, USA)) добавляют в раствор, содержащий микровезикулы, и смесь может быть инкубирована при комнатной температуре в течение 60 минут на качающейся платформе, для обеспечения пегелирования микровезикул. Пегилированные или непегилированные микровезикулы затем могут быть подвергнуты замене буфера путем диализа (например, с использованием 1×ФСБ), концентрированию путем ультрафильтрации и фильтрации через шприцевой фильтр (например, имеющий размер пор 0,45 мкм) и т.п.
Микровезикулы, полученные способом настоящего изобретения содержат трансгенный продукт и/или лентивирусную РНК, содержащую трансген. Лентивирусная РНК может содержать последовательность промотора гена TERT перед трансгеном. Трансген вводится в клетку при помощи лентивирусного вектора, содержащего промотор гена TERT, благодаря чему обеспечивается интеграция трансгена в геном хозяина и его экспрессия, а также увеличивается высвобождение микровезикул из клетки. Таким образом, микровезикулы могут быть эффективно получены.
В одном варианте осуществления настоящее изобретение также относится к:
способу получения микровезикул, содержащих трансгенный продукт и/или лентивирусную РНК, содержащую трансген, включающему в себя этапы:
введения трансгена в клетку при помощи лентивирусного вектора in vitro, где указанный лентивирусный вектор является дефицитным, по меньшей мере, по одному гену структурного белка и содержит трансген под контролем промотора гена теломеразной обратной транскриптазы (TERT) в лентивирусной геномной последовательности,
культивирования клетки для внеклеточного высвобождения микровезикул, содержащих трансгенный продукт и/или лентивирусную РНК, содержащую трансген, и
сбора высвобожденных микровезикул.
2. Микровезикула, полученная способом настоящего изобретения
Настоящее изобретение также относится к микровезикуле, содержащей трансгенный продукт и/или лентивирусную РНК, содержащую трансген, где эта микровезикула получена способом настоящего изобретения.
Термин «микровезикула (МВ)» в контексте настоящего изобретения относится к мембранной везикуле размером от 5 нм до 5 мкм, предпочтительно от 10 нм до 1 мкм, более предпочтительно от 20 нм до 500 нм, которая продуцируется клетками и высвобождается или отпочковывается за пределы клетки. Размер микровезикулы может быть определен методом электронной микроскопии. Примеры микровезикулы, как правило, включают в себя без ограничений экзосомы, отпочковывающиеся микровезикулы и апоптозные тельца. Как правило, микровезикула настоящего изобретения представляет собой экзосому. Экзосома представляет собой мембранную везикулу, состоящую из липидного бислоя. Экзосома имеет размер 150 нм или меньше, как правило, от 20 до 120 нм или от 40 до 100 нм. Экзосома внеклеточно секретируется и продуцируется посредством экзоцитоза, происходящего в результате слияния клеточной мембраны мультивезикулярного тельца (МВТ), образованного путем внутреннего выпячивания эндосомальной мембраны.
Микровезикула настоящего изобретения может содержать белок и/или РНК, такие как трансгенный продукт и/или лентивирусная РНК, содержащая трансген, которые были транспортированы в нее из клетки-хозяина и инкапсулированы в ней. Примеры РНК, которая может содержаться в микровезикуле, включают в себя без ограничений мРНК, микроРНК, кшРНК, миРНК и лентивирусную РНК (включая различные варианты сплайсинга лентивирусной РНК). Примеры белка, который может содержаться в микровезикуле, включают в себя без ограничений вирусные белки (например, белок Vpu и белок обратной транскриптазы (ОТ) ВИЧ-1), эндогенные клеточные белки (например, белки цитоскелета, сигнальные белки и ферменты) и чужеродные трансгенные продукты. Например, экзосома, как правило, может содержать белок цитоскелета (например, тубулин, актин и актинсвязывающий белок), белок, связанный с мембранным транспортом (например, аннексин и белок Rab), сигнальный белок (например, протеинкиназу и 14-3-3), метаболический фермент (например, ГАФДГ, АТФазу и элонгазу), белок из семейства тетраспанинов (например, CD9, CD63, CD81 и CD82), белок теплового шока (например, HSP90 и HSP70), белок биосинтеза МВТ (например, Alix и TSG101) и иммуномодулирующую молекулу (например, MHCI и MHCII) и т.п.
Лентивирусная ДНК (провирусная ДНК), содержащая лентивирусную геномную последовательность, содержащую вышеописанный трансген, которая была введена в клетку способом настоящего изобретения, интегрируется в геном организма-хозяина. Лентивирусная РНК, содержащаяся в микровезикуле настоящего изобретения, транскрибируется с этой провирусной ДНК. Лентивирусная РНК может содержать последовательность РНК промотора гена TERT (например, промотора гена TERT человека) на 5’-конце трансгена.
В особенно предпочтительном варианте осуществления микровезикула настоящего изобретения может содержать белок-супрессор опухолевого роста, такой как PTEN или p16, или кшРНК, такую как кшРНК CDC6, или ее РНК-предшественник, в качестве трасгенного продукта.
Микровезикула настоящего изобретения может быть поглощена другими клетками для доставки в эти клетки содержащегося в микровезикуле трансгенного продукта и/или лентивирусной РНК, содержащей трансген.
3. Способ генной трансдукции при помощи микровезикулы настоящего изобретения
Настоящее изобретение также относится к способу генной трансдукции, включающему в себя приведение клетки-мишени в контакт с микровезикулой, содержащей трансгенный продукт и/или лентивирусную РНК, содержащую трансген, для их слияния, чем обеспечивается введение трансгена в клетку, где микровезикула получена способом настоящего изобретения. В одном варианте осуществления изобретения клетка-мишень может быть приведена в контакт с микровезикулой in vitro или in vivo.
Микровезикула, в частности экзосома, может проникать в находящиеся по соседству с ней другие клетки для участия в межклеточном взаимодействии. Считается, что микровезикула способна проникать внутрь клетки-мишени, благодаря слиянию мембран, или эндоцитозподобным путем, хотя настоящее изобретение не ограничивается этой теорией. В способе генной трансдукции настоящего изобретения приведение клетки-мишени в контакт с микровезикулой может быть осуществлено любым способом, известным специалистам в данной области техники. Например, приведение клетки-мишени в контакт с микровезикулой in vitro может быть осуществлено путем добавления микровезикулы в культуруальную клеточную среду. Приведение клетки-мишени в контакт с микровезикулой in vivo может быть осуществлено, например, путем перорального введения микровезикулы или путем парентерального введения, такого как непосредственное нанесение или инъекция микровезикулы в целевую область (например, внутрипеченочное, внутрисуставное, внутрижелудочковое и интраназальные введение). Другие способы и области введения in vivo, которые могут быть использованы, будут описаны ниже в отношении способов введения и областей введения для фармацевтической композиции. При помощи способа генной трансдукции настоящего изобретения можно эффективно доставлять в клетку-мишень содержащиеся в микровезикуле трансгенный продукт и/или лентивирусную РНК, содержащую трансген.
4. Композиция, содержащая микровезикулу настоящего изобретения
Настоящее изобретение также относится к композиции, содержащей микровезикулу, полученную способом настоящего изобретения. Кроме микровезикулы, композиция может содержать любой ингредиент, необходимый для ее предполагаемого использования. Например, композиция может быть предназначена для использования в генной трансдукции. В этом случае, композиция может содержать лекарственное средство, стимулирующее генную трансдукцию, и/или лекарственное средство, стабилизирующее нуклеиновую кислоту, и т.п.
5. Фармацевтическая композиция, содержащая микровезикулу настоящего изобретения, и способ лечения
Настоящее изобретение также относится к фармацевтической композиции, содержащей, микровезикулу, полученную способом настоящего изобретения.
В одном варианте осуществления изобретения фармацевтическая композиция может быть предназначена для применения в лечении таких заболеваний как рак, диабет, нейродегенеративное заболевание, иммунная дисфункция, воспаление, цирроз печени, атеросклероз, тромбоз и инфекция. Предпочтительно, фармацевтическая композиция может быть предназначена для применения в лечении рака. В частности, рак может быть выбран из группы, состоящей, например, из рака толстой кишки, рака поджелудочной железы, рака почки, рака легкого, нейробластомы, рака молочной железы, рака яичников, рака желудка, рака предстательной железы, рака щитовидной железы и злокачественной лимфомы. В одном варианте осуществления изобретения трансгенный продукт, содержащийся в микровезикуле, может вызывать уменьшение экспрессии CDC6. Например, трансгенный продукт может представлять собой кшРНК, специфичную по отношению к CDC6. В этом случае, заболевание, подлежащее лечению, например, рак, может быть связано с повышенным уровнем экспрессии CDC6.
В случае использования микровезикулы в фармацевтической композиции трансген или трансгенный продукт, содержащийся в микровезикуле настоящего изобретения, оказывает действие, направленное на предотвращение и/или лечение заболевания. Например, такой трансген может представлять собой ген-супрессор опухолевого роста, такой как ген PTEN или p16, и/или может представлять собой ген, кодирующий кшРНК, специфичную по отношению к гену, кодирующему регулятор клеточной пролиферации или его предшественник. В одном варианте осуществления изобретения трансген представляет собой ген PTEN и/или ген, кодирующий кшРНК CDC6. В этом варианте осуществления изобретения фармацевтическая композиция может содержать, например, в комбинации микровезикулу, полученную способом настоящего изобретения, в которой трансген представляет собой ген PTEN, и микровезикулу, полученную способом настоящего изобретения, в которой трансген представляет собой ген, кодирующий кшРНК CDC6. В другом варианте осуществления изобретения трансген представляет собой ген p16 и/или ген, кодирующий кшРНК CDC6. В этом варианте осуществления изобретения фармацевтическая композиция может содержать, например, в комбинации микровезикулу, полученную способом настоящего изобретения, в которой трансген представляет собой ген p16, и микровезикулу, полученную способом настоящего изобретения, в которой трансген представляет собой ген, кодирующий кшРНК CDC6.
Кроме микровезикулы настоящего изобретения фармацевтическая композиция настоящего изобретения содержит жидкую среду. Примеры жидкой среды включают в себя воду, физиологически приемлемые буферные растворы (фосфатно-солевой буферный раствор и т.п.) и биосовместимые водные среды, такие как пропиленгликоль и полиоксиэтиленовый сложный эфир сорбита и жирной кислоты. Такую среду желательно стерилизовать и предпочтительно доводить значение осмотического давления до изотоничного с плазмой крови, если это необходимо.
Фармацевтическая композиция может содержать фармацевтически приемлемый носитель. Термин «фармацевтически приемлемый носитель» относится к добавке, как правило, используемой в области фармацевтических технологий. Примеры фармацевтически приемлемого носителя включают в себя суспендирующие агенты, регулирующие тоничность агенты, буферы и консерванты. Такой носитель в основном используется для облегчения изготовления и поддержания лекарственной формы и эффекта лекарственного средства, и может быть соответствующим образом использован при необходимости.
Например, глицерилмоностеарат, моностеарат алюминия, метилцеллюлоза, карбоксиметилцеллюлоза, гидроксиметилцеллюлоза и лаурилсульфат натрия могут быть использованы в качестве суспендирующих агентов. Примеры регулирующих тоничность агентов включают в себя хлорид натрия, глицерин и D-маннит. Примеры буферов включают в себя фосфатный, ацетатный, карбонатный и цитратный буферные растворы. Примеры консервантов включают в себя хлорид бензалкония, парагидроксибензойную кислоту и хлорбутанол.
Фармацевтическая композиция при необходимости также может содержать корригирующий агент, загуститель, солюбилизирующий агент, регулятор pH, разбавитель, поверхностно-активное вещество, экспандер, стабилизатор, стимулятор абсорбции, смачивающий агент, увлажнитель, адсорбент, покрывающий агент, краситель, антиоксидант, ароматизатор, подсластитель, вспомогательное вещество, связующее вещество, разрыхлитель, ингибитор распадения, наполнитель, эмульгатор, добавка, регулирующая расход, смазывающее вещество или тому подобное, в дополнение к тому, что описано выше.
Фармацевтическая композиция настоящего изобретения также может содержать дополнительный лекарственный препарат без потери фармакологического эффекта, которым обладает микровезикула настоящего изобретения. Например, фармацевтическая композиция может содержать заранее определенное количество антибиотика.
Лекарственная форма фармацевтической композиции не ограничивается и может быть любой формой, которая не инактивирует микровезикулу и не инактивирует трансгенный продукт и/или лентивирусную РНК, содержащиеся в микровезикуле. Лекарственная форма фармацевтической композиции может представлять собой, например, жидкую, твердую или полутвердую форму. Конкретные примеры лекарственной формы включают в себя: парентеральные лекарственные формы, такие как инъекции, суспензии, эмульсии, кремы, глазные капли, капли в нос, мази, гипсовые повязки, пластыри и суппозитории; и пероральные лекарственные формы, такие как жидкие препараты, капсулы, подъязычные препараты, пастилки, порошки, таблетки и гранулы. Лекарственная форма фармацевтической композиции предпочтительно представляет собой жидкий препарат, такой как инъекция.
Фармацевтическая композиция может быть введена в организм в фармацевтически эффективном количестве для лечения целевого заболевания. Организм-реципиент может быть позвоночным, например, млекопитающим, птицей, земноводным или пресмыкающимся, и предпочтительно является млекопитающим. Примеры млекопитающего включают в себя неприматов, таких как собака, кошка, лошадь, свинья, корова, коза, овца, мышь и крыса; и приматов, таких как человек, шимпанзе и горилла. Млекопитающее предпочтительно является человеком.
Термин «фармацевтически эффективное количество» в данном описании относится к дозе, необходимой для того, чтобы микровезикула, содержащаяся в фармацевтической композиции настоящего изобретения, предотвращала или лечила целевое заболевание или облегчала симптомы, имея при этом небольшое количество или совсем не имея вредных побочных эффектов в отношении организма-реципиента. Конкретная доза изменяется в зависимости от типа заболевания, которое необходимо предотвратить и/или лечить, механизма действия, лежащего в основе возникновения заболевания, используемой лекарственной формы, информации о субъекте и способа введения и т.п. Диапазон фармацевтически эффективного количества и предпочтительный путь введения фарамацевтической композиции, которая вводится человеку, как правило, устанавливаются на основе данных полученных в результате анализа культуры клеток и экспериментов с животными. Конечная доза может быть определена и скорректирована в соответствии с мнением, например, врача для конкретного субъекта. Информация о субъекте, принимаемая во внимание в этом случае, включает в себя степень прогрессирования или тяжести заболевания, общее состояние здоровья, возраст, массу тела, пол, рацион питания, чувствительность к лекарственному средству и устойчивость к лечению и т.п. В одном варианте осуществления изобретения, когда трансген кодирует белок, фармацевтическая композиция настоящего изобретения может быть введена в одной или нескольких дозах в количестве от 1×104 до 1×108 трансфекционных единиц (ТЕ) на кг массы тела, например, от 1×105 до 1×107 ТЕ на кг массы тела или от 2×105 до 5×106 ТЕ на кг массы тела на одну дозу путем прямой инъекции в пораженные области или внутривенной инъекции. Здесь трансфекционная единица может быть определена путем введения трансгена в клетку (например, клетку клеточной линии 293Т эмбриональной почки человека), используя 10 мкг лентивирусного вектора настоящего изобретения (например, ДНК вектора, такого как плазмидный вектор, например, pRBL0213T); определения количества трансгенного продукта (например, белка, такого как белок PTEN) во фракции микровезикул (МВ), высвобожденных из клетки, например, при помощи ИФА; и нормализации определенного количества (эффективность трансфекции) как одной трансфекционной единицы, эквивалентной 20 пкг трансгенного продукта (например, белка) на 1000 клеток. Фармацевтическая композиция настоящего изобретения может вводиться два или более раз через определенные интервалы времени, например, каждые 1 час, 3 часа, 6 часов или 12 часов, каждый день, каждые 2 дня, 3 дня или 7 дней, или каждые 1 месяц, 2 месяца, 3 месяца, 6 месяцев или 12 месяцев. Любое другое парентеральное введение или пероральное введение может быть осуществлено в количестве, которое соответствует количествам, описанным выше. В случае особенно тяжелых симптомов доза может быть увеличена в соответствии с симптомами.
Введение фармацевтической композиции может быть системным введением или местным введением, и может быть надлежащим образом выбрано в соответствии с типом заболевания, местом, где возникло заболевание, или степенью его прогрессирования и т.п. Если заболевание возникает в локальной области, то фармацевтическую композицию предпочтительно вводить местно путем непосредственного введения в локальную область (например, опухоль) и ее окружение при помощи инъекции или постоянного катетера или тому подобного. Это обусловлено тем, что микровезикулы настоящего изобретения могут быть введены в достаточном количестве в область (ткань или орган), подлежащую лечению, и при этом не оказывать влияния на другие ткани. В то же время, при метастатическом раке область, подлежащая лечению, не может быть идентифицирована, или заболевание может возникать системно. В этом случае предпочтительным является системное введение посредством внутривенной инъекции или тому подобного. Это обусловлено тем, что микровезикулы настоящего изобретения могут распространяться системно с помощью кровотока, обеспечивая тем самым введение даже в пораженные участки, которые не могут быть определены при диагностике.
Фармацевтическая композиция может быть введена любым подходящим способом, который не инактивирует содержащийся в ней активный ингредиент. Например, введение может быть парентеральным (например, инъекция, аэрозоль, наружное нанесение, глазные капли, капли в нос или постоянный катетер) или пероральным. Предпочтительным способом введения является инъекция.
В случае введения путем инъекции область инъекции может быть не ограниченной областью, где микровезикула настоящего изобретения может осуществить свои функции и обеспечить достижение цели фармацевтической композиции. Примеры областей инъекций включают в себя внутривенное, внутриартериальное, внутрипеченочное, внутримышечное, внутрисуставное, интрамедуллярное, интраспинальное, внутрижелудочковое, чрескожное, подкожное, внутрикожное, внутрибрюшинное, интраназальное, кишечное и подъязычное введение. В одном варианте осуществления изобретения непосредственное введение в опухоль также является предпочтительным.
Фармацевтическая композиция настоящего изобретения может применяться для обеспечения эффективного предотвращения и/или лечения заболевания при помощи трансгенного продукта или тому подобного, содержащегося в микровезикуле.
Таким образом, настоящее изобретение также относится к способу лечения пациента, включающему в себя введение микровезикулы, полученной способом настоящего изобретения, пациенту, нуждающемуся в ведении трансгена или трансгенного продукта. Пациент может страдать такими заболеваниями, как рак, диабет, нейродегенеративное заболевание, иммунная дисфункция, воспаление, цирроз печени, атеросклероз, тромбоз и инфекция. Предпочтительно, пациент страдает от рака. В частности, рак может быть выбран из группы, состоящей, например, из рака толстой кишки, рака поджелудочной железы, рака почки, рака легкого, нейробластомы, рака молочной железы, рака яичников, рака желудка, рака предстательной железы, рака щитовидной железы и злокачественной лимфомы. В одном варианте осуществления изобретения трансгенный продукт, содержащийся в микровезикуле, может вызывать уменьшение экспрессии CDC6. Например, трансгенный продукт может представлять собой кшРНК, специфичную по отношению к CDC6. В этом случае, заболевание, подлежащее лечению, например, рак, может быть связано с повышенным уровнем экспрессии CDC6. Для введения микровезикул пациенту могут быть использованы способы введения и области введения, описанные выше в отношении способов введения и областей введения для фармацевтической композиции.
Способом лечения настоящего изобретения можно эффективно лечить у пациента такое заболевание как рак. В одном предпочтительном варианте осуществления изобретения способ лечения рака настоящего изобретения может ингибировать (уменьшать) рост опухолей.
6. Описание последовательностей
SEQ ID NO: 1 представляет собой нуклеотидную последовательность промотора гена теломеразной обратной транскриптазы (TERT) Homo sapiens, которая был использован для получения плазмид pRBL0213T, pTHTN и pRBL001 (см., Фигуры 2A и 2B и Фигуру 3B). Последовательность геномной ДНК, содержащая ген TERT человека, доступна в базе данных Genbank под регистрационным номером AF128893.
SEQ ID NO: 2 представляет собой нуклеотидную последовательность 5’-части транскрибируемой области гена TERT Homo sapiens, которая была использована для получения плазмид pRBL0213T, pTHTN и pRBL001.
SEQ ID NO: 3 представляет собой нуклеотидную последовательность, содержащую промотор гена TERT (SEQ ID NO: 1) и 5’-часть (SEQ ID NO: 2) транскрибируемой области гена TERT Homo sapiens, которая была использована для получения плазмид pRBL0213T, pTHTN и pRBL001. SEQ ID NO: 3 содержит апстрим последовательность, весь первый экзон и часть второго экзона гена TERT. Первый экзон начинается в положении 1390 и заканчивается в положении 1670 SEQ ID NO: 3. Второй экзон начинается в положении 1775 SEQ ID NO: 3.
SEQ ID NO: 4 представляет собой последовательность клона рекомбинантной плазмиды pNL4-3, содержащую нуклеотидную последовательность полноразмерной геномной ДНК штамма NL4-3 ВИЧ-1 (регистрационный номер в Genbank M19921). Нуклеотидная последовательность с положения 1 до положения 9709 SEQ ID NO: 4 соответствует геному ВИЧ-1 (с 5’-LTR до 3’-LTR). Нуклеотидная последовательность с положения 6221 до положения 8785 SEQ ID NO: 4 кодирует белок env. Структура генома ВИЧ-1 приведена на Фигуре 1A.
SEQ ID NO: 5 представляет собой нуклеотидную последовательность геномной области ВИЧ-1 в плазмиде pTHTK, т.е. нуклеотидную последовательность от 5’-LTR до 3’-LTR в pTHTK (см. Фигуру 1B). Нуклеотидная последовательность с положения 6344 до положения 7611 SEQ ID NO: 4 была удалена из pNL4-3. Кроме того, pNL4-3 расщепляли между положениями 8650 и 8651 SEQ ID NO: 4 ферментом рестрикции Hpa I, и встраивали в нее нуклеотидную последовательность от положения 7383 до положения 7674 SEQ ID NO: 5 для получения pTHTK.
SEQ ID NO: 6 представляет собой нуклеотидную последовательность кодирующей последовательности PTEN (от старт-кодона до стоп-кодона) Homo sapiens, которая была использована для получения плазмиды pRBL0213T (см. Фигуру 2B). Последовательность мРНК PTEN человека доступна в базе данных Genbank под регистрационным номером NM_000314.
SEQ ID NO: 7 представляет собой нуклеотидную последовательность кодирующей последовательности CDC6 (от старт-кодона до стоп-кодона) Homo sapiens. Последовательность мРНК CDC6 человека доступна в базе данных Genbank под регистрационным номером NM_001254.
SEQ ID NOs: 8 и 9 представляют собой олигонуклеотидные последовательности Cdc6-5-A и Cdc6-3-A, соответственно, которые были использованы для получения двухцепочечного олигонуклеотида, встроенного в плазмиду pRBL001 (см., Фигуру 3B).
SEQ ID NO: 10 представляет собой последовательность ДНК, кодирующую кшРНК CDC6.
SEQ ID NO: 11 представляет собой нуклеотидную последовательность праймера B-NLR8950.
SEQ ID NO: 12 представляет собой нуклеотидную последовательность олигонуклеотида HD-A.
SEQ ID NO: 13 представляет собой нуклеотидную последовательность олигонуклеотида HD-S.
SEQ ID NO: 14 представляет собой нуклеотидную последовательность прямого праймера BHU5-S2.
SEQ ID NO: 15 представляет собой нуклеотидную последовательность обратного праймера HDA/SBOT.
SEQ ID NO: 16 представляет собой нуклеотидную последовательность праймера 5'LTRU5.
SEQ ID NO: 17 представляет собой нуклеотидную последовательность праймера 3'NL5850.
SEQ ID NO: 18 представляет собой нуклеотидную последовательность праймера 3'NL8960.
SEQ ID NO: 19 представляет собой нуклеотидную последовательность антисмысловой последовательности кшРНК CDC6.
SEQ ID NO: 20 представляет собой нуклеотидную последовательность линкера в кшРНК CDC6.
SEQ ID NO: 21 представляет собой нуклеотидную последовательность частичного фрагмента гена большого Т-антигена вируса SV40.
SEQ ID NO: 22 представляет собой аминокислотную последовательность белка PTEN человека, кодируемого геном PTEN, имеющим последовательность SEQ ID NO: 6.
SEQ ID NO: 23 представляет собой нуклеотидную последовательность (кодирующую последовательность) гена p16INK4a человека (регистрационный номер в Genbank L27211).
SEQ ID NO: 24 представляет собой аминокислотную последовательность белка p16INK4a человека, кодируемого геном p16INK4a, имеющим последовательность SEQ ID NO: 23.
SEQ ID NO: 25 представляет собой нуклеотидную последовательность плазмиды pCMV p16INK4a.
SEQ ID NO: 26 представляет собой нуклеотидную последовательность (кодирующую последовательность) гена p53 человека (регистрационный номер в Genbank BC003596).
SEQ ID NO: 27 представляет собой аминокислотную последовательность белка p53 человека, кодируемого геном p53, имеющим последовательность SEQ ID NO: 26.
Примеры
Далее настоящее изобретение будет описано более подробно путем приведения Примеров. Однако технический объем настоящего изобретения не ограничивается этими Примерами.
Пример 1 Рекомбинантный лентивирусный плазмидный вектор
Рекомбинантные лентивирусные плазмидные векторы, используемые в описанных ниже Примерах, получали из вектора pNL4-3 на основе геномной ДНК ВИЧ-1 (Фигуры 1A и 4A) (Adachi A et al., J. Virol., 1986, p. 284-291). Последовательность клона рекомбинантной плазмиды pNL4-3, содержащая нуклеотидную последовательность полноразмерной геномной ДНК штамма NL4-3 ВИЧ-1 доступна в базе данных Genbank под регистрационным номером в M19921 (SEQ ID NO: 4). Рекомбинантный лентивирусный плазмидный вектор pD64V (Фигура 4B) имеет мутацию, которая приводит к замене аспарагиновой кислоты в положении 64 интегразы ВИЧ-1 на валин в pNL4-3. Плазмида pD64V была любезно предоставлена доктором Сэмом Чоу, Факультет молекулярной и медицинской фармакологии Калифорнийского университета Лос-Анджелеса (UCLA).
Другие использованные рекомбинантные лентивирусные плазмидные векторы получали из конструкции pTHTK (Фигура 1B). pTHTK получали из плазмиды pNL4-3, описанной выше, (Фигуры 1A и 1B). У pTHTK отсутствуют нуклеотиды с 6344 по 7611 (по последовательности SEQ ID NO: 4; это относится и к дальнейшему описанию, если не указано иное) в геномной ДНК ВИЧ-1 плазмиды pNL4-3 в результате обработки ферментами рестрикции Kpn I и Bgl II. В частности, у фрагмента, находящегося между двумя сайтами Kpn I (положения с 6343 по 6348) и Bgl II (положения с 7611 по 7616) в основе ВИЧ-1, «тупили» концы и повторно лигировали для получения плазмиды pTHTK, дефицитной по гену env ВИЧ-1, кодирующему гликопротеин оболочки p120. Затем pTHTK расщепляли в только одном сайте Hpa I (положения с 8648 по 8653) и модифицировали плазмиду путем добавления Mlu I линкера (нуклеоотидная последовательность с положения 7383 до 7674 последовательности SEQ ID NO: 5). Нуклеотидная последовательность геномной области ВИЧ-1 (т.е. c 5’-LTR до 3’-LTR) pTHTK показана в SEQ ID NO: 5.
Для лигирования фрагмента ДНК (экспрессирующей трансген кассеты) для клонирования, вектор pTHTK расщепляли со стороны 5’-конца в сайте Mlu I и со стороны 3’-конца в сайте Xho I (положения с 8887 по 8892). Вставка экспрессирующей трансген кассеты в сайт Mlu I-Xho I этого вектора разрывала открытую рамку считывания гена nef ВИЧ-1. Белок nef, кодируемый геном nef, играет важную роль в вирусной инфекции и распространении, а также во внеклеточном высвобождении вириона. Поэтому лентивирусный плазмидный вектор, содержащий вставку (т.е. nef-дефектный), будет продуцировать вирусные частицы менее инфекционные для CD4+ клеток, в отличие от штамма дикого типа NL4-3 ВИЧ-1. В то же время, в случае ВВС-G-псевдотипированных RBL0213T, RBL001, THTN и т.п., как дефицитных, так и недефицитных по Nef, продуцировалось больше вирусных частиц, чем в случае штамма дикого типа NL4-3, как описано ниже (см., например, Фиг.9).
Полученную таким образом основу pTHTK использовали для конструирования рекомбинантных лентивирусных плазмидных векторов, как описано ниже.
(a) pRBL0213T
pRBL0213T представляет собой плазмиду, в которой ДНК, кодирующая ген PTEN человека, была встроена под контролем промотора гена TERT человека (промотор hTERT) (Фигура 2B).
Плазмиду pGL-1375 использовали в качестве источника промотора hTERT (Takakura et al., Cancer Res., 1999, p. 551-557). Плазмида pGL-1375 была любезно предоставлена доктором Сатору Кио (Университет Канадзавы, Япония). Плазмида pGL-1375 содержала фрагмент ДНК промотора hTERT, встроенный между сайтами Mlu I и Bgl II. Последовательность 5’-части hTERT доступна в базе данных Genbank под регистрационным номером AF128893. Фрагмент ДНК промотора hTERT вырезали из плазмиды pGL-1375 и встраивали в основу pTHTK.
Последовательность мРНК PTEN доступна в базе данных Genbank под регистрационным номером NM_000314 (SEQ ID NO: 6). Кодирующую ген PTEN ДНК, используемую для конструирования pRBL0213T, вырезали из вектора pcDNA3.1/CMV-hPTEN (любезно предоставленного доктором Хонг Ву, Факультет молекулярной медицины Калифорнийского университета Лос-Анджелеса (UCLA)). Кодирующую ген PTEN ДНК затем встраивали в основу pTHTK для получения плазмиды pRBL0213T.
Плазмида pRBL0213T содержит нуклеотидную последовательность SEQ ID NO: 3, состоящую из последовательности промотора hTERT (SEQ ID NO: 1) и 5’-части транскрибируемой области hTERT (SEQ ID NO: 2), содержащей первый экзон и часть второго экзона гена hTERT; и кодирующую последовательность гена PTEN.
(b) pTHTN
pTHTN представляет собой плазмиду, в которой частичный фрагмент гена большого Т-антигена вируса SV40 был встроен под контролем промотора hTERT (Фигура 2C). Частичный фрагмент гена большого Т-антигена вируса SV40 (SEQ ID NO: 21) встраивали после промотора hTERT, встроенного в основу pTHTK, для получения плазмиды pTHTN. Геномная последовательность вируса обезьян 40, содержащая ген большого Т-антигена вируса SV40, доступна в базе данных Genbank под регистрационным номером NC_001669. Плазмида pTHTN содержит нуклеотидную последовательность SEQ ID NO: 3, состоящую из последовательности промотора hTERT (SEQ ID NO: 1) и 5’-части транскрибируемой области hTERT (SEQ ID NO: 2), содержащей первый экзон и часть второго экзона гена hTERT; и частичный фрагмент (SEQ ID NO: 21; с нуклеотида 5175 (Hind III) до нуклеотида 4863 (Hae III) последовательности ДНК, зарегистрированной в Genbank под номером NC_001669), гена большого Т-антигена вируса SV40.
(c) pRBL001
pRBL001 представляет собой плазмиду (плазмиду для получения рекомбинантной лентивирусной частицы THTD), в которой ДНК, кодирующая кшРНК CDC6, была встроена под контролем промотора hTERT (Фигура 3B).
Последовательности синтетических олигонуклеотидов, используемых для конструирования ДНК, кодирующей кшРНК CDC6, показаны ниже.
Cdc6-5-A: 5’-GATCCCCAGGCACTTGCTACCAGCAATTCAAGAGATTGCTGGTAGCAAGTGCCTTTTTTGGAAA-3’ (SEQ ID NO: 8)
Cdc6-3-A: 5’-AGCTTTTCCAAAAAAGGCACTTGCTACCAGCAATCTCTTGAATTGCTGGTAGCAAGTGCCTGGG-3’ (SEQ ID NO: 9)
Для конструирования pRBL001 синтетические олигонуклеотиды Cdc6-5-A и Cdc6-3-A смешивали в эквимолярных количествах, денатурировали и повторно отжигали для получения двухцепочечного олигонуклеотида. У этого двухцепочечного олигонуклеотида «тупили» концы и затем встраивали в сайт EcoR V субклонирующего вектора pBlueScript. Проводили секвенирование ДНК для подтверждения того, что двухцепочечный олигонуклеотид CDC6 был встроен в правильной ориентации, чтобы иметь правильную последовательность. Полученный субклон линеаризировали в сайте Xba I, «тупили» концы и модифицировали путем лигирования Mlu I линкера, описанного выше. Фрагмент ДНК, содержащий последовательность промотора hTERT, получали путем расщепления описанной выше плазмиды pGL3-1375 ферментами рестрикции Mlu I и Bgl II, и клонировали его по сайтам Mlu I и BamH I субклонирующего вектора, содержащего двухцепочечный олигонуклеотид CDC6. Фрагмент ДНК, содержащий промотор hTERT и двухцепочечный олигонуклеотид CDC6, вырезали из полученного субклона путем двойной рестрикции Mlu I и Xho I. Полученный фрагмент очищали и встраивали в основу pTHTK, которую расщепляли ферментами рестрикции Mlu I и Xho I, и проводили очистку в геле для получения pRBL001.
Плазмида pRBL001 содержит нуклеотидную последовательность SEQ ID NO: 3, состоящую из последовательности промотора hTERT (SEQ ID NO: 1) и 5’-части транскрибируемой области hTERT (SEQ ID NO: 2), содержащей первый экзон и часть второго экзона гена hTERT; и последовательность, кодирующую кшРНК CDC6 (SEQ ID NO: 10). кшРНК CDC6 содержит антисмысловую последовательность кшРНК CDC6 (SEQ ID NO: 19); линкер (SEQ ID NO: 20); и смысловую последовательность, состоящую из последовательности, комплементарной антисмысловой последовательности и поли-U последовательности, выступающей на 5 оснований на 3’-конце.
кшРНК, специфичная по отношению к мРНК CDC6 человека (кшРНК CDC6), получаемая в результате транскрипции pRBL001, может специфично вызывать деградацию мРНК CDC6 путем РНК-интерференции, что приводит к нокдауну белка инициации репликации ДНК CDC6. Последовательность мРНК CDC6 человека доступна в базе данных Genbank под регистрационным номером NM_001254. Нуклеотидная последовательность кодирующей последовательности CDC6 человека приведена в списке последовательностей под номером SEQ ID NO: 7. Гидовая цепь антисмысловой последовательности кшРНК CDC6 SEQ ID NO: 19 связывается с мРНК CDC6, чтобы вызывать РНК-интерференцию.
(d) pTHTH
pTHTH представляет собой плазмиду, в которой частичный фрагмент гена большого Т-антигена вируса SV40 был встроен под контролем промотора hTERT, у которого отсутствует 5’-часть (1 т.п.н.) (Фигура 4E).
Для конструирования pTHTH плазмиду pGL-378 (любезно предоставлена доктором Сатору Кио, Университет Канадзавы, Япония), содержащую укороченную последовательность промотора hTERT с делецией 5’-части (1 т.п.н.) и ген люциферазы, расщепляли ферментами рестрикции Mlu I и Hind III для получения фрагмента ДНК, содержащего укороченную последовательность промотора hTERT, у которого отсутствует 5’-часть (1 т.п.н.). Этот фрагмент ДНК очищали и встраивали в плазмиду pBluescript по сайтам Mlu I и Hind III для получения плазмиды pEND-HTPs. Частичный фрагмент гена большого Т-антигена вируса SV40 (SEQ ID NO: 21) вырезали из плазмиды, содержащей геномную ДНК вируса SV40, ферментами рестрикции Hind III и Hae III и встраивали в pEND-HTPs по сайтам Hind III и Hinc II. Полученный плазмидный субклон расщепляли ферментами рестрикции Mlu I и Xho I, и вырезанный фрагмент ДНК, содержащий укороченную последовательность промотора hTERT, у которого отсутствует 5’-часть (1 т.п.н.), и частичный фрагмент гена большого Т-антигена вируса SV40, очищали и встраивали в pTHTK по сайтам Mlu I и Xho I для получения pTHTH.
Плазмида pTHTH содержит нуклеотидную последовательность, состоящую из последовательности промотора hTERT, у которого отсутствует 5’-часть (1 т.п.н.), и 5’-части транскрибируемой области hTERT (SEQ ID NO: 2), содержащей первый экзон и часть второго экзона гена hTERT; и последовательность частичного фрагмента гена большого Т-антигена вируса SV40 (SEQ ID NO: 21).
(e) pTHTC
pTHTC представляет собой плазмиду, в которой частичный фрагмент гена большого Т-антигена вируса SV40 был встроен под контролем промотора цитомегаловируса человека (ЦМВ) (Фигура 4F).
Промотор ЦМВ встраивали в основу pTHTK. Частичный фрагмент гена большого Т-антигена вируса SV40 (SEQ ID NO: 21) встраивали после промотора ЦМВ в pTHTK для получения плазмиды pTHTC.
Пример 2 Получение псевдотипированной рекомбинантной лентивирусной частицы
Рекомбинантные лентивирусные плазмидные векторы, дефицитные по гену env ВИЧ-1, описанные в Примере 1, сами по себе не могут продуцировать инфекционные вирусные частицы. Таким образом, в этом Примере рекомбинантные лентивирусные частицы (псевдотипированные) получали путем котрансфекции клеток клеточной линии 293Т эмбриональной почки человека каждым рекомбинантным лентивирусным плазмидным вектором, описанным в Примере 1, и плазмидой pCMV-VSV/G, экспресирующей гликопротеин оболочки G вируса везикулярного стоматита (ВВС). Плазмида pCMV-VSV/G была любезно предоставлена доктором Сэмом Чоу, Факультет молекулярной и медицинской фармакологии Калифорнийского университета Лос-Анджелеса (UCLA). Конкретные экспериментальные процедуры описаны ниже.
1. Трансфекция плазмидной ДНК
15 мл полной среды DMEM с высоким содержанием глюкозы (Hyclone, Utah, USA) (с добавлением 10% фетальной бычьей сыворотки, 100 единиц/мл пеницилина и 100 мкг/мл стрептомицина (Hyclone, Utah, USA)) добавляли в колбу T75, в которую затем инокулировали клетки клеточной линии 293Т эмбриональной почки человека. Клетки 293Т размножали в инкубаторе в атмосфере с 5% CO2 при температуре 37°С и наращивали количество культуры до десяти колб T75 путем пересевания.
2×HEPBS (100 мл) получали следующим образом: 1 г HEPES (кислая соль), 1,6 г NaCl, 0,75 мл Na2HPO4 (0,25 M) и 1 мл KCl (1 M) растворяли в соответствующем количестве ddH2O. Значение pH раствора доводили до 6,9 при помощи NaOH (5 M) и затем точно доводили до 7,12-7,14 при помощи NaOH (1 M). В этот раствор добавляли ddH2O до общего объема 100 мл, и полученный раствор пропускали через шприцевой фильтр с размером пор 0,22 мкм для получения 2×HEPBS.
Следующие компоненты добавляли в 50-мл пробирку:
(a) 500 мкл раствора плазмидной ДНК, содержащего 170 мкг каждого рекомбинантного лентивирусного плазмидного вектора, дефицитного по гену env ВИЧ-1, описанного в Примере 1, и 30 мкг pCMV-VSV/G;
(b) 1650 мкл ddH2O;
(c) 350 мкл 2 M CaCl2.
Раствор осторожно перемешивали, и затем в него по каплям добавляли 2500 мкл 2×HEPBS при аккуратном перемешивании, чтобы избежать образования большого осадка, получая, таким образом, трансфекционную смесь.
Пробирку, содержащую трансфекционную смесь, оставляли при комнатной температуре в течение 20 минут. Из десяти колб T75, в которых выращивали клетки 293T, удаляли среду. Затем в каждую колбу добавляли 12 мл свежей полной среды DMEM с высоким содержанием глюкозы. В каждую колбу постепенно добавляли 500 мкл трансфекционной смеси. Клетки инкубировали в инкубаторе в атмосфере с 5% CO2 при температуре 37°С в течение 8 часов.
2. Клеточная культура и сбор среды
Среду удаляли, и в каждую колбу добавляли 15 мл свежей полной среды DMEM с высоким содержанием глюкозы. Клетки инкубировали в инкубаторе в атмосфере с 5% CO2 при температуре 37°С в течение 36 часов и собирали среду (среда после 36 часов). В каждую колбу добавляли 15 мл свежей полной среды DMEM с высоким содержанием глюкозы. Клетки инкубировали в инкубаторе в атмосфере с 5% CO2 при температуре 37°С в течение дополнительных 36 часов и собирали среду (среда после 72 часов). В каждую колбу добавляли 15 мл свежей полной среды DMEM с высоким содержанием глюкозы. Клетки также инкубировали в инкубаторе в атмосфере с 5% CO2 при температуре 37°С в течение 24 часов и собирали среду (среда после 96 часов). Каждую 50-мл пробирку, содержащую собранную среду, центрифугировали при 3000 об./мин и температуре 4°С в течение 5 минут.
3. Титрование
200 мкл надосадочной среды переносили в другую пробирку и разбавляли от 50 до 200 раз путем добавления 1×ФСБ. Эту разбавленную надосадочную среду анализировали на количество белка p24 вирусного происхождения, высвободившегося в среду из клеток, инфицированных рекомбинантными лентивирусными частицами, с использованием набора реагентов для ИФА антигена p24 ВИЧ-1 (Coulter Inc., Miami, FL, USA), и таким образом определяли титры вирусных частиц.
4. Очистка
Надосадочную среду, собранную при помощи центрифугирования на описанном выше этапе «2. Клеточная культура и сбор среды» переносили в 250-мл высокоскоростную центрифужную пробирку и центрифугировали при 9000×g и температуре 4°С в течение 60 минут. Полученный супернатант подвергали 5-кратному концентрированию путем ультрафильтрации при помощи концентратора Vivaspin® 20 (1000 кДа отсечение по молекулярной массе (м.м. отсеч.)) (Sartorius, NY, USA). В полученный раствор добавляли раствор мМP, и вирусные частицы осаждали в течение ночи при температуре 4°С. Раствор мМP (300 мл) получали следующим образом: 90 г ПЭГ 8000 (молекулярно-биологическая степень очистки) и 30 мл 5M раствора NaCl (высокая степень очистки) растворяли в соответствующем количестве ddH2O и в этот раствор добавляли ddH2O до конечного объема 300 мл.
Раствор, в котором осаждались вирусные частицы, центрифугировали при 9000×g и температуре 4°С в течение 30 минут для получения осадка, содержащего вирусные частицы. Супернатант удаляли, и осадок ресуспендировали в 10 мл 1×ФСБ для получения раствора, содержащего вирусные частицы.
Вирусные частицы в аликвоте раствора подвергали пегилированию (Croyle et al., J. Virol., 2004, Vol. 78, p. 912-921). Пегилирование проводили путем добавления 0,4 мл пегилирующего раствора (33 мг/мл метокси ПЭГ сукцинимидилкарбоната NHS (mPEG-NHS, м.м. 10К (NANOCS, USA)), 30 мМ HEPES-KOH, pH 7,5, 500 мМ NaCl) в приблизительно 10 мл раствора, содержащего вирусные частицы, и инкубирования этой смеси при комнатной температуре в течение 60 минут на качающейся платформе. Раствор, содержащий пегилированные или непегилированные вирусные частицы диализировали против 1×ФСБ при температуре 4°С при помощи кассеты Slide-A-Lyzer (20 кДа м.м. отсеч.) (Thermo Scientific, IL, USA), и 1×ФСБ заменяли свежим буфером каждые 24 часа в течение 2 суток. Диализированный таким образом раствор вирусных частиц концентрировали в 30 раз с использованием фильтра AmiconUltra 15 (100 кДа м.м. отсеч.) (Millipore, MA, USA). Концентрированный раствор вирусных частиц пропускали через шприцевой фильтр с размером пор 0,45 мкм, и полученный препарат хранили при температуре -80°С.
Таким образом, успешно получали псевдотипированные рекомбинантные лентивирусные частицы. Псевдотипированные рекомбинантные лентивирусные частицы, полученные из рекомбинантных лентивирусных плазмидных векторов pTHTK, pRBL0213T, pTHTN, pRBL001, pTHTH и pTHTC, описанных в Примере 1, в последующих Примерах обозначали как THTK, RBL0213T, THTN, THTD, THTH и THTC, соответственно.
Полученные таким образом рекомбинантные лентивирусные частицы могли инфицировать различные человеческие пролиферирующие клетки и неделящиеся и делящиеся клетки, как описано ниже. Клетки-мишени, инфицированные рекомбинантными лентивирусными частицами, синтезируют лентивирусную ДНК, которая в свою очередь встраивается в хромосомную ДНК хозяина. Таким образом, полученные рекомбинантные лентивирусные частицы могут быть использованы в качестве лентивирусных векторов.
Пример 3 Изучение способности рекомбинантной лентивирусной частицы продуцировать инфекционную вирусную частицу
Способность полученной из pTHTK псевдотипированной рекомбинантной вирусной частицы THTK продуцировать инфекционные вирусные частицы после инфекции клеток изучали путем обнаружения LTR-метки, полученной при помощи опосредованной лигированием ПЦР (ОЛ-ПЦР).
1. Получение сайта Bpm I в pTHTK
Для проведения ОЛ-ПЦР pTHTK сначала модифицировали, чтобы создать новый сайт Bpm I в геноме ВИЧ-1 (Фигура 5A). Область 5’-LTR ВИЧ-1 имеет три элемента (U3, R и U5). Сайт Bpm I получали при помощи перекрывающейся ПЦР 3’-конца элемента 5’-LTR в pTHTK. Bpm I представляет собой фермент рестрикции класса II S. Bpm I распознает последовательность 5’-CTGGAG-3’. Bpm I расщепляет ДНК через 16 нуклеотидов от 3’-конца своего сайта узнавания в цепи, содержащей этот сайт 5’-CTGGAG-3’, и через 14 нуклеотидов от 5’-конца своего сайта узнавания в другой цепи, с образованием на3’-конце 2 выступающих нуклеотидов (Фигура 5B).
На Фигуре 5C показана концевая структура лентивирусного генома. Лентивирусная РНК образуется путем транскрипции от 5’-LTR R элемента до 3’-LTR R элемента провирусной ДНК. Соответственно, лентивирусная РНК не имеет на концах ни 5’-LTR U3 элемента ни 3’-LTR U5 элемента. Однако, при обратной транскрипции лентивирусной РНК в лентивирусную ДНК, 3’-LTR U3 элемент и 5’-LTR U5 элемент копируются в 5’-LTR U3 элемент и 3’-LTR U5 элемент, соответственно. Затем эта лентивирусная ДНК интегрируется в геном хозяина, образуя провирусную ДНК.
Таким образом, новый сайт Bpm I в 5’-LTR U5 элементе, созданный в геноме ВИЧ-1 pTHTK, копируется в 3’-LTR U5 элемент, когда лентивирусная ДНК синтезируется в инфицированных клетках. Этот новый сайт Bpm I не оказывает влияния на сборку вириона THTK, инфекцию вирусной частицы и интеграцию провирусной ДНК в инфицированных клетках. Клетки-хозяева инфицируются вирусной частицей THTK, полученной из pTHTK, имеющей этот новый сайт Bpm I, таким образом, что провирусная ДНК, имеющая сайт Bpm I, скопированный в положение, удаленное на 2 нуклеотида (5’-CA-3’) от конца, в 3’-LTR U5 элементе, интегрируется в геном хозяина. Таким образом, геномная ДНК из инфицированных клеток в результате обработки Bpm I расщепляется в положении, удаленном на 14 нуклеотидов от сайта интеграции провирусной ДНК (Фигура 5B).
2. Получение LTR-метки
Клетки клеточной линии HeLa рака шейки матки человека, экспрессирующие белок Tat ВИЧ-1 (клетки HeLa/tat), инфицировали путем совместной инкубации с вирусными частицами THTK, полученными как описано в Примере 2 из pTHTK, имеющей новый сайт Bpm I, описанный выше. Клетки HeLa/tat были любезно предоставлены доктором Сэмом Чоу, Факультет молекулярной и медицинской фармакологии Калифорнийского университета Лос-Анджелеса (UCLA). Через 3, 5, 10 и 15 суток после инфицирования из клеток HeLa/tat выделяли геномную ДНК хозяина. Геномную ДНк подвергали двойной рестрикции Kas I и Xho I. Сайты Kas I (положения с 637 по 642; по последовательности SEQ ID NO: 4; это относится и к дальнейшему описанию) и Xho I (положения с 8887 по 8892) являются уникальными сайтами в геноме ВИЧ-1. Сайт Kas I расположен через 4 нуклеотида после 5’-LTR, а сайт Xho I расположен приблизительно за 190 нуклеотидов перед 3’-LTR. Двойная рестрикция Kas I и Xho I отделяет провирусную ДНК от геномной ДНК хозяина, при этом последовательности 5’-LTR и 3’-LTR провирусной ДНК остаются в геномной ДНК хозяина. Кроме того, сайт узнавания Kas I расположен между сайтом узнавания Bpm I (положения с 627 по 632), созданным в 5’-LTR, и сайтом узнавания Bpm I, расположенным через 16 нуклеотидов после него. Поэтому даже рестрикция Kas I с последующей рестрикцией Bpm I не приводит к расщеплению Bpm I по сайту Bpm I в 5’-LTR.
Затем проводили специальную реакцию достраивания ДНК, как описано ниже. В этой реакции использовали праймеры, полученные из последовательности приблизительно 100 нуклеотидов перед 3’-LTR. Таким образом, в значительной степени амплифицируются 3’-LTR провирусной ДНК и геномная ДНК хозяина после него, а относительное количество контаминирующей ДНК, в частности ДНК, расположенной перед провирусной ДНК, резко уменьшается. Нуклеотидная последовательность праймера B-NLR8950 для достраивания ДНК приведена ниже.
B-NLR8950: 5’-B-GTGCCTGGCTAGAAGCACAAG-3’ (SEQ ID NO: 11)
Эта последовательность имеет меченные биотином нуклеотиды, и она соответствует положениям 8950-8970 в геномной ДНК штамма NL4-3 ВИЧ-1. Этот праймер был использован для проведения реакции достраивания ДНК, как показано ниже.
Смесь для реакции достраивания ДНК:
B-NLR8950 50 мкМ;
Геномная ДНК хозяина, обработанная Kas I + Xho I 10 мкг;
dNTPs 200 мкМ;
Taq ДНК полимераза 5 единиц.
Пробирку, содержащую смесь для реакции достраивания ДНК, описанную выше, инкубировали при температуре 94°С в течение 5 минут, при температуре 55°С в течение 5 минут и при температуре 72°С в течение 30 минут для осуществления реакции достраивания ДНК, чтобы получить биотинилированную ДНК.
Далее эту биотинилированную ДНК очищали путем связывания с покрытыми стрептавидином магнитными частицами (Dynabeads M-280): приблизительно 5 пмоль биотинилированной ДНК смешивали с 40 пмоль Dynabeads M-280 в пробирке, и инкубировали эту смесь при комнатной температуре в течение 30 минут. Пробирку помещали на магнитную подставку (Dynal MPC stand) и оставляли на 2 минуты. Затем частицы промывали TE буфером (pH 8,0). Для промывания частиц использовали 1 мл TE буфера, и затем повторяли процедуру два раза. Частицы центрифугировали в течение 2 секунд и удаляли супернатант для очистки биотинилированной ДНК.
Затем биотинилированную ДНК подвергали рестрикции Bmp I. Рестрикцию биотинилированной ДНК ферментом Bmp I осуществляли в течение ночи при температуре 37°С. Расщепленную биотинилированную ДНК очищали при помощи Dynabeads M-280 и затем лигировали с двухцепочечным олигонуклеотидным линкером. Один конец этого двухцепочечного олигонуклеотидного линкера имел выступающие с 3’-конца 2 нуклеотида и был комплементарен концу, образующемуся при рестрикции Bmp I, Другой конец этого двухцепочечного олигонуклеотидного линкера имел выступающие с 5’-конца 3 нуклеотида и эффективно предотвращал самолигирование линкера. Нуклеотидные последовательности олигонуклеотидов HD-A и HD-S (любезно предоставлены доктором Сэмом Чоу, Калифорнийский университет, UCLA), образующих двухцепочечный линкер, приведены ниже.
HD-A: 5’-CACGCGTCGCATCATATCTCCAGGTGTGACAG-3’ (SEQ ID NO: 12)
HD-S: 5’-CCTCTGTCACACCTGGAGATATGATGCGACGCGTGNN-3’ (SEQ ID NO: 13)
3’-концевые нуклеотиды «NN» олигонуклеотида HD-S обозначают вырожденную последовательность и представляют собой любую комбинацию «A», «T», «G» и «C».
Лигирование биотинилированной ДНК с двухцепочечным олигонуклеотидным линкером проводили при комнатной температуре в течение 16 часов. Биотинилированную ДНК, связанную с двухцепочечным олигонуклеотидным линкером, очищали при помощи Dynabeads M-280.
3. Обнаружение LTR-метки
Для обнаружения LTR-метки проводили ПЦР-амплификацию. В ПЦР реакции использовали следующие праймеры:
Прямой праймер BHU5-S2: 5’-GAGTGCTCAAAGTAGTGGT-3’ (SEQ ID NO: 14)
Обратный праймер HDA/SBOT: 5’-CTGTCACACCTGGAGATATGAT-3’ (SEQ ID NO: 15)
Нуклеотидные последовательности прямого праймера и обратного праймера получены из последовательности геномной ДНК ВИЧ-1 (положения 9617-9636) и одной цепи HD-S двухцепочечного олигонуклеотидного линкера, соответственно.
Проводили следующую ПЦР реакцию: 1 цикл, включающий в себя 94°С в течение 4 минут, и 30 циклов, каждый из которых включал в себя 94°С в течение 60 секунд, 60°С в течение 30 секунд и 72°С в течение 90 секунд, а затем 72°С в течение 10 минут и 4°С в течение 1-12 часов. Биотинилированную ДНК, связанную с двухцепочечным олигонуклеотидным линкером, описанным выше, использовали в качестве матрицы ДНК в ПЦР реакции.
После ПЦР, 5 мкл каждого продукта реакции наносили на гель (2,0% агароза) и проводили электрофорез в 1×TAE. Наблюдали ПЦР продукт (LTR-метка) размером примерно 138 п.н. в том случае, если провирусная ДНК была интегрирована в геном клетки-хозяина (Фигура 6).
4. Результаты
Результаты показаны на Фигуре 7. Обнаруженная LTR-метка (положительный результат по LTR-метке) указывает на то, что провирусная ДНК была интегрирована в геном клетки-хозяина. Это означает, что клетки-хозяева были инфицированы вирусом. Псевдотипированная рекомбинантная лентивирусная частица THTK оказалась положительной по LTR-метке, и было показано, что она успешно инфицирует клетки (Фигура 7, после инфицирования).
Затем культуральную среду инфицированных клеток собирали, фильтровали через фильтр с размером пор 0,45 мкм и добавляли к свежим культивируемым клеткам («повторное инфицирование»). Из этих клеток выделяли геномную ДНК и анализировали ее на наличие LTR-метки. В результате вирусная частица, отрицательная по LTR-метке, указывает на то, что клетки не были инфицированы этой вирусной частицей (Фигура 7, после повторного инфицирования). Эти результаты показали, что клетки, инфицированные рекомбинантной лентивирусной частицей THTK, не продуцируют инфекционные вирусные частицы. ОЛ-ПЦР для проверки того, является ли рекомбинантная лентивирусная частица неспособной реплицироваться в инфицированных клетках, оказалась точной и привела к отличной воспроизводимости без ложноположительных результатов, т.е. контаминации прямой ПЦР последовательности ВИЧ-1.
Описанные выше результаты показали, что псевдотипированная рекомбинантная лентивирусная частица THTK может инфицировать клетки, но инфицированные клетки не продуцируют инфекционные вирусные частицы. Это указывает на то, что способ получения микровезикул настоящего изобретения, в котором в качестве лентивирусного вектора используются псевдотипированные рекомбинантные лентивирусные частицы, не приводит к образованию инфекционных вирусных частиц.
Пример 4 Прямая инъекция рекомбинантной лентивирусной частицы THTD в опухоль человека, трансплантированную голой мыши
Проведенный эксперимент с животными предусматривал прямую инъекцию рекомбинантной лентивирусной частицы THTD, полученной как описано в Примере 2, в опухоль рака молочной железы человека Bcap-37, трансплантированную голой мыши. THTD (содержание белка 275 мкг) вводили путем инъекции в несколько мест опухолевого очага животного два раза в неделю в течение 3 недель. Через 48 часов после последнего введения животное умерщвляли путем смещения шейных позвонков. Каждому животному в контрольной группе в опухолевый очаг вводили 1×ФСБ. Длину и ширину каждой опухоли измеряли дважды в неделю после инъекции, при помощи стандартного штангенциркуля. После того, как животных умерщвляли путем смещения шейных позвонков, у них извлекали опухоли, которые измеряли и обрабатывали для патологоанатомического исследования.
Пролиферация обработанных THTD опухолей подавлялась на 34,70% по сравнению с контрольной группой, и средняя масса опухоли была значимо меньше, чем масса опухоли в контрольной группе (p<0,02). Инъекция THTD вызывала сильный фиброз в опухолевой ткани, подавляя тем самым пролиферацию опухоли.
Описанные выше результаты показали, что псевдотипированная рекомбинантная лентивирусная частица THTD инфицирует мышиные клетки, и затем кшРНК CDC6, кодируемая вирусным геномом, продуцировалась в этих клетках, подавляя экспрессию CDC6, что приводило к подавлению пролиферации опухоли. Это указывает на возможность того, что микровезикула настоящего изобретения, содержащая кшРНК CDC6, также обладает способностью подавлять пролиферацию опухоли.
Пример 5 Усиление сплайсинга вирусной геномной РНК промотором hTERT
Активность в отношении генной транскрипции и генной трансдукции рекомбинантной вирусной частицы THTN, полученной как описано в Примере 2, сравнивали с этой активностью у других рекомбинантных лентивирусных частиц, и изучали путем выявления активности сплайсинга РНК при помощи ОТ-ПЦР (Фигура 8).
1. Вирусная инфекция и сбор клеток
Клетки клеточной линии HeLa рака шейки матки человека, экспрессирующие белок Tat ВИЧ-1 (клетки HeLa/tat), инокулировали в концентрации приблизительно 2×105 клеток на лунку в 6-луночный планшет, и добавляли в него полную среду DMEM с высоким содержанием глюкозы в количестве 3 мл на лунку. Клетки размножали в инкубаторе при температуре 37°С в атмосфере с 5% CO2 в течение 24 часов до тех пор, пока они не достигали 80% конфлюентности при наблюдении в инвертированный микроскоп. Псевдотипированную рекомбинантную лентивирусную частицу THTN, THTH или THTC, полученную, как описано в Примере 2, или вирусную частицу NL4-3 или D64V, полученную путем трансфекции клеток клеточной линии 293Т эмбриональной почки человека плазмидой pNL4-3 или pD64V, добавляли в лунки (две лунки для каждой лентивирусной частицы), в которых пролиферировали клетки HeLa/tat, для того, чтобы инфицировать эти клетки каждым вирусом. Раствор вируса, содержащий приблизительно 4×105 инфекционных единиц, эквивалентных 400 нг вирусного белка p24, использовали для инфицирования 2×105 клеток-мишений при каждом заражении (множественность заражения (МЗ): 1,3).
В то же время проводили имитацию заражения путем инкубации клеток с «мертвыми» вирионами THTN. Эти вирионы THTN получали путем кипячения при температуре 100°С в течение 5 минут, чтобы гарантировать инактивацию вируса. Это ложное заражение «мертвым» вирусом полностью гарантирует, что ни одна вирусная частица не проникает в клетку, и поэтому вирусная РНК не транскрибируется в анализе при помощи ОТ-ПЦР.
Инфицированные таким образом клетки размножали в инкубаторе в атмосфере с 5% CO2 при температуре 37°С в течение 8 часов. Клетки наблюдали под микроскопом, для подтверждения того, что клетки являются здоровыми и не контаминированы. Среду осторожно удаляли и добавляли к клеткам свежую полную среду DMEM с высоким содержанием глюкозы в количестве 3 мл на лунку. Клетки размножали в инкубаторе при температуре 37°С в атмосфере с 5% CO2 в течение 48 часов. Затем клетки три раза промывали 1×ФСБ и собирали клетки путем соскабливания их с поверхности лунок при помощи резинового скребка. Клетки собирали в 15-мл низкоскоростную центрифужную пробирку, в которую затем добавляли 1×ФСБ до конечного объема 12 мл. Пробирку центрифугировали при 3000 об./мин для получения клеточного осадка. Супернатант удаляли, и клеточный осадок замораживали в сухом льду в течение 30 минут.
2. Экстракция РНК
Буфер GTC готовили следующим образом: 212 г гуанидин тиоцианата (м.м. 118,16), 2.2 г цитрата натрия (м.м. 294,1) и 15 мл саркозила (10% масс./об.) растворяли в соответствующем количестве ddH2O и в этот раствор добавляли ddH2O до конечного объема 300 мл (конечные концентрации: 6 M, 5 мМ и 0,5%, соответственно). Для приготовления буфера GTC перед использованием на каждые 20 мл добавляли 1,4 мл β-меркаптоэтанола.
1000 мкл буфера GTC добавляли к клеточному осадку в каждой пробирке. Клетки пять раз пропускали через иглу 18G 1/2 при помощи шприца для гомогенизации клеток.
Затем в каждую пробирку добавляли по 1000 мкл насыщенного водой фенола и 1000 мкл хлороформа и перемешивали пробирку на вортексе. Затем в каждую пробирку добавляли по 3 мл 1 M ацетата натрия pH 5,3, перемешивали пробирку на вортексе и центрифугировали при 9000×g и температуре 4°С в течение 10 минут. Образовавшийся верхний слой раствора переносили в новую 50-мл пробирку, в которую добавляли от 0,6 до 1,0 объема изопропанола. Раствор перемешивали путем переворачивания пробирки пять раз, и оставляли на ночь при комнатной температуре. Пробирку перемешивали на вортексе в течение 30 секунд. От 200 до 300 мкл раствора переносили в 1,5-мл микропробирку и центрифугировали при 13000 об./мин и температуре 4°С в течение 15 минут для получения осадка РНК. Супернатант удаляли и осадок РНК промывали 1 мл 75% этанола и центрифугировали при 13000 об./мин и температуре 4°С в течение 5 минут. Супернатант удаляли и осадок РНК сушили на воздухе.
3. ОТ-ПЦР
Осадок РНК повторно растворяли в 10 мкл ddH2O. Реакцию обратной транскрипции и ПЦР (ОТ-ПЦР) проводили для амплификации кДНК. В этой ОТ-ПЦР для каждой обратной транскрипции использовали 200 мкг РНК.
Реакцию обратной транскрипции (ОТ) проводили следующим образом: 10 мкл обработанной ДНКазой I РНК (приблизительно 200 мкг), 1 мкл исходного раствора 10 мМ dATP/dCTP/dGTP/dTTP, 1 мкл ингибитора РНКазы Rnasin® (30 единиц/мкл, Promega), 2 мкл 10x ПЦР-буфера, 1 мкл раствора случайного праймера 0,1 мкг/мл (случайный гексамер) и 1 мкл 50 мМ MgCl2 осторожно перемешивали и затем добавляли 1 мкл обратной траскриптазы M-MuLV (200 единиц/мкл) и перемешивали в пробирке путем центрифугирования в течение 2 секунд. Пробирку инкубировали сначала при комнатной температуре в течение 10 минут, а затем при температуре 42°С в течение 60 минут. Затем пробирку кипятили на водяной бане при температуре 100°С в течение 15 минут и охлаждали на льду в течение 5 минут. Раствор, полученный в пробирке в результате реакции ОТ, распределяли по четырем новым пробиркам (в количестве, соответствующем 50 мкг РНК в каждой пробирке) и хранили при температуре -80 для дальнейшего использования.
Затем ПЦР реакцию проводили следующим образом: 2,5 мкл праймера 5’LTRU5 (20 мкM), 2,5 мкл праймера 3’NL5850 (20 мкМ) или праймера 3’NL8960, 5 мкл раствора после реакции ОТ и 40 мкл смеси ПЦР смешивали и при помощи термоциклера для ПЦР реакции (Applied Biosystems) подвергали воздействию следующих температурных условий: 1 цикл, включающий в себя 94°С в течение 4 минут, и 16 циклов, каждый из которых включал в себя 94°С в течение 60 секунд, 60°С в течение 30 секунд и 72°С в течение 90 секунд, а затем 72°С в течение 10 минут.
Нуклеотидные последовательности использованных праймеров показаны ниже.
Праймер 5’LTRU5: 5’-TCTGGCTAACTAGGGAACCCACTG-3’ (SEQ ID NO: 16).
Праймер 3’NL5850: 5’-GCTATGTCGACACCCAATTCTGAA-3’ (SEQ ID NO: 17).
Праймер 3’NL8960: 5’-TGTGCTTCTAGCCAGGCACAAGC-3’ (SEQ ID NO: 18).
ПЦР смесь (для шести образцов) готовили в конечном объеме 240 мкл путем смешивания следующих компонентов:
Исходный раствор 10 мМ dATP/dCTP/dGTP/dTTP 6 мкл;
32P-α-dCTP (3000 Ки/ммоль, 10 мкрКи/мкл, GE, USA) 1 мкл;
10×ПЦР буфер 30 мкл;
Taq ДНК полимераза (5 единиц/мкл) 3 мкл;
ddH2O 201 мкл.
После ПЦР, в 20 мкл каждого продукта ПЦР реакции добавляли 3 мкл буфера для нанесения ДНК на гель. Смесь наносили на 2,2% агарозный гель в 1x TAE буфере. Гель заворачивали, сушили и экспонировали с рентгеновской пленкой в течение ночи при температуре -80°С. 1x TAE буфер готовили путем растворения 242 г трис-основания (молекулярно-биологическая степень очистки), 57,1 мл ледяной уксусной кислоты (молекулярно-биологическая степень очистки) и 100 мл 0,5М ЭДТА pH 8,0 (молекулярно-биологическая степень очистки) в соответствующем количестве ddH2O, добавления в этот раствор ddH2O до конечного объема 1000 мл, и разбавления полученного таким образом 50x TAE буфера ddH2O.
В клетках HeLa/tat, инфицированных различными рекомбинантными лентивирусными частицами, проводили сравнение сплайсинга молекул РНК, полученных из этих частиц. Вирусную частицу D64V, дефицитную по интегразе ВИЧ-1, и таким образом дефицитную по интеграции провирусной ДНК ВИЧ-1, использовали в качестве отрицательного контроля.
4. Результаты
Результаты ПЦР с использованием праймеров 5’LTRU5 и 3’NL5850 показаны на Фигуре 8B. ВИЧ-1 имеет семь основных сплайсированных фрагмента РНК, связанных с образованием различных вирусных мРНК. В результате использования праймеров 5’LTRU5 и 3’NL5850 основные ПЦР продукты пяти из этих фрагментов были обнаружены во всех проанализированных клетках, инфицированных лентивирусными частицами. В частности, клетки, инфицированные вирусной частицей THTN, содержащей промотор hTERT в вирусной геномной РНК, обладали очень высокой активностью сплайсинга РНК, которая была, по меньшей мере, в 10 раз выше, чем уровень сплайсинга РНК в клетках, инфицированных вирусной частицей NL4-3 дикого типа. В отличие от этого, уровень сплайсинга РНК не повышался в клетках, инфицированных рекомбинантной лентивирусной частицей THTH, у которой отсутствует 5’-часть (1 т.п.н.) промотора hTERT, или THTC, имеющей промотор ЦМВ, по сравнению с клетками, инфицированными вирусной частицей NL4-3 дикого типа.
Результаты ПЦР с использованием праймеров 5’LTRU5 и 3’NL8960 показаны на Фигуре 8A. Полученный уровень ПЦР амплификации с использованием праймеров 5’LTRU5 и 3’NL8960 отражает количество вирусной геномной РНК, полученной из каждой лентивирусной частицы. Однако при помощи ПЦР с использованием праймеров 5’LTRU5 и 3’NL8960 не удалось эффективно получить ПЦР продукты, и в этом случае были получены слабые сигналы. Клетки, инфицированные вирусной частицей THTN, содержащей промотор hTERT в вирусной геномной РНК, имели уровень вирусной геномной РНК, эквивалентный этому уровню у клеток, инфицированных другими вирусными частицами (THTH, THTC или NL4-3).
Описанные выше результаты показали, что сплайсинг лентивирусной РНК усиливается в клетках, инфицированных псевдотипированной рекомбинантной лентивирусной частицей, несущей лентивирусную РНК, содержащую промотор гена теломеразной обратной транскриптазы человека (hTERT), тем самым повышая экспрессию трансгена.
Пример 6 Усиление генной трансдукции и экспрессии промотором hTERT
Влияние промотора hTERT на активность в отношении генной трансдукции оценивали при помощи ИФА анализа p24, используя в качестве показателя экспрессию вирусного белка.
Клетки 293T и клетки HeLa/tat инокулированные в концентрации 2×106 клеток инфицировали (множественность заражения (МЗ): 1,0) псевдотипированной рекомбинантной лентивирусной частицей THTN, THTH или THTC, полученной в Примере 2, или вирусной частицей NL4-3 или D64V. После инфицирования вирусная геномная ДНК синтезируется из вирусной геномной РНК в лентивирусной частице и интегрируются в виде провирусной геномной ДНК в геном клетки-хозяина. Белки, например вирусный антиген p24, экспрессирующиеся в результате интеграции гена (генной трансдукции) в геном хозяина, могут секретироваться в культуральную среду. Таким образом, среду собирали, и количество вирусного белка p24 в среде измеряли при помощи набора реагентов для ИФА антигена p24 ВИЧ-1 (Coulter Inc., Miami, FL, USA), чтобы оценить активность генной трансдукции каждой вирусной частицы.
Сначала культуральную клеточную среду собирали в различные моменты времени после инфицирования. Каждую собранную среду центрифугировали для удаления клеточного дебриса и затем разбавляли 1×ФСБ и подвергали анализу на p24. Степень разбавления, как правило, составляла от 1:50 до 1:200. В результате, уровень p24 повышался приблизительно через 36 часов после инфицирования, достигал пика через 72 часа после инфицирования и быстро снижался через 96 часов после инфицирования. Таким образом, уровень p24 указывали в виде среднего значения в среде, собранной через 72 часа после инфицирования.
Результаты показаны на Фигуре 9. Клетки 293T, инфицированные вирусной частицей THTC, содержащей промотор ЦМВ в вирусной геномной РНК, имели уровень p24 выше, чем уровень этого белка в клетках 293T, инфицированных вирусной частицей NL4-3 дикого типа. Это свидетельствует о том, что промотор ЦМВ активирует генную трансдукцию и экспрессию в клетках 293T. Такая активация генной трансдукции и экспрессии была более выражена в случае вирусной частицы THTN, которая содержала промотор hTERT в вирусной геномной РНК и обладала активностью в отношении сплайсинга РНК, как показано в Примере 5. Клетки 293T, инфицированные THTN имели уровень p24 выше, чем уровень этого белка в клетках 293T, инфицированных THTC. В отличие от этого, клетки 293T, инфицированные вирусной частицей THTH, у которой отсутствует 5’-часть (1 т.п.н.) промоторной последовательности hTERT, содержащей имели уровень p24 эквивалентный уровню этого белка в клетках, инфицированных вирусной частицей NL4-3 дикого типа. Кроме того, p24 присутствовал в незначительных количествах в случае D64V, которая не вызывает генной трансдукции, показывая, что уровень p24 является показателем уровней генной трансдукции и экспрессии.
В случае клеток HeLa/tat, клетки, инфицированные THTC или THTH, продуцировали p24 на уровне, эквивалентном этому уровню в случае инфицирования вирусной частицей NL4-3 дикого типа. Только клетки HeLa/tat, инфицированные вирусной частицей THTN, содержащей промотор hTERT в вирусной геномной РНК, имели уровень экспрессии p24 2500 нг/мл, которые был явно выше, чем в случае инфицирования другими вирусными частицами.
Описанные выше результаты показали, что, генная трансдукция и экспрессия усиливаются промотором hTERT, как видно из того, что экспрессия вирусного белка усиливалась в клетках, инфицированных псевдотипированной рекомбинантной лентивирусной частицей, несущей лентивирусную РНК, содержащую промотор hTERT.
Пример 7 Повышение высвобождения микровезикул, опосредованное промотором hTERT
Клетки, в которые был введен лентивирусный плазмидный вектор pRBL0213T, или клетки, инфицированные вирусной частицей RBL0213T, оценивали по их уровню высвобождения микровезикул (МВ), используя в качестве показателя значение активности трансгена PTEN в клетках или в МВ, определенное при помощи анализа PTEN.
1. Трансфекция плазмидной ДНК
200 мкл раствора pRBL0213T с концентрацией 1мкг/мл и 50 мкл 3M ацетата натрия pH 5,3 добавляли в 1,5-мл микропробирку, в которую затем добавляли 1 мл 100% этанола. Раствор перемешивали и охлаждали при температуре -80°С в течение 60 минут. Затем пробирку центрифугировали при 13000 об./мин в течение 15 минут для получения осадка ДНК. Супернатант удаляли. В пробирку добавляли 1 мл 70% этанола и затем центрифугировали при 13000 об./мин в течение 5 минут. Осторожно удаляли супернатант и осадок ДНК сушили на воздухе. Осадок ДНК растворяли путем добавления 500 мкл ddH2O для получения раствора плазмидной ДНК pRBL0213T. pRBL0213T представляет собой рекомбинантный лентивирусный плазмидный вектор, содержащий ген PTEN под контролем промотора hTERT (Фигура 10D, Lenti.).
Аналогичным образом готовили растворы ДНК плазмид GL3-1375, pcDNA3.1/CMV-hPTEN и pRBL016Bn. Плазмида pGL3-1375 (Takakura et al., Cancer Res., 1999, p. 551-557) имела промотор hTERT, но не имела гена PTEN, и была использована в качестве отрицательного контроля при анализе PTEN (Фигура 10A, Empt.). Плазмида pcDNA3.1/CMV-hPTEN представляла собой плазмиду, в которой ген PTEN был встроен под контролем промотора ЦМВ основной плазмиды pcDNA3.1, и она была использована в качестве положительного контроля при анализе PTEN (Фигура 10B, Regul.). Плазмида pRBL016Bn представляет собой ретровирусную плазмиду Mo-MLV, содержащую ген PTEN под контролем промотора крысиного рецептора фактора роста нервной ткани (rNGFR) (Фигура 10C, Retro.).
500 мкл раствора любого из этих четырех видов плазмидной ДНК (20 мкг плазмидной ДНК), 400 мкл ddH2O и 75 мкл 2M CaCl2 добавляли в новую 50-мл пробирку и осторожно перемешивали. Затем в нее по каплям добавляли 525 мкл 2×HEPBS при аккуратном перемешивании, чтобы избежать образования большого осадка. Пробирку выдерживали при комнатной температуре в течение 20 минут для получения трансфекционной смеси.
Среду удаляли из колбы T75, в которой росли клеткой клеточной линии 293Т эмбриональной почки человека, как описано в Примере 2. Затем в каждую колбу добавляли 12 мл свежей полной среды DMEM с высоким содержанием глюкозы. В каждую колбу постепенно добавляли 1000 мкл трансфекционной смеси. Клетки инкубировали в инкубаторе в атмосфере с 5% CO2 при температуре 37°С в течение 8 часов, чтобы трансфицировать клетки каждой плазмидной ДНК. Затем удаляли среду и в каждую колбу добавляли по 15 мл свежей полной среды DMEM с высоким содержанием глюкозы. Клетки инкубировали в инкубаторе в атмосфере с 5% CO2 при температуре 37°С в течение 60 часов. Также ложную трансфекцию проводили при помощи аналогичных процедур, используя вместо растворов плазмидной ДНК воду.
2. Инфицирование вирусной частицей RBL0213T
Как описано в Примере 2, рекомбинантную лентивирусную частицу RBL0213T получали, используя плазмиду pRBL0213T, затем пегилировали и очищали. Полученной рекомбинантной лентивирусной частицей RBL0213T инфицировали (МЗ: 1,3) клетки 293T, клетки CEM или клетки HeLa. Клетки инфицировали вирусной частицей RBL0213T путем инкубации в течение 12 часов. К клеткам добавляли свежую среду, и выращивали клетки в течение 48 часов.
В то же время химически инактивированную вирусную частицу получали путем приготовления 100 мМ раствора AT-2 (2,2’-дипиридилдисульфид) (Aldrithiol-2) в ДМСО (Fluka), добавления AT-2 в концентрации 1 мМ в раствор, содержащий вирусную частицу, и выдерживания этой смеси в течение ночи при температуре 4°С (Rossio JL, J. Virol., 1998, Vol. 72, p. 7992-8001). AT-2 представляет собой реагент, который окисляет цистеин в белках вирионов, ингибируя функции обратной транскриптазы, и таким образом инактивирует вирус. Обработка AT-2 устраняет инфекционность ВИЧ, сохраняя при этом целостность и конформацию поверхностных вирусных белков.
3. Получение клеточного лизата
Раствор, содержащий свободный фосфат не является предпочтительным для анализа PTEN, так как он создает высокий фон. Рекомендуется, чтобы буферы и пробирки были бесфосфатными.
После описанных выше трансфекции или инфицирования клетки собирали в новую 50-мл пробирку и центрифугировали при 3000 об./мин в течение 5 минут для осаждения клеток. Для использования на описанном ниже этапе «4. Получение лизата МВ», среду переносили в новую 5-мл пробирку. Осажденные клетки промывали 30 мл холодного 1×ФСБ и снова центрифугировали для осаждения клеток. Супернатант удаляли. К клеткам (2×106 клеток) добавляли 250 мкл лизирующего буфера (25 мМ Tris-HCl, pH 8,0, 150 мМ NaCl, 1% NP-40, 1 мМ ЭДТА, 5% глицерин). Клетки десять раз пропускали через иглу 24G при помощи шприца для гомогенизации клеток, получая тем самым клеточные экстракты.
Клеточные экстракты инкубировали при температуре 4°С в течение 60 минут на качающейся платформе и центрифугировали при 13000 об./мин при температуре 4°С в течение 20 минут. Супернатант (клеточные лизаты) переносили в новую пробирку. 5 мкл клеточных лизатов добавляли в другую пробирку. Затем в нее добавляли 1 мл раствора для анализа белков Bio-Rad (Bio-Rad, USA) и измеряли OD600. При определении концентрации белка в качестве стандарта использовали бычий сывороточный альбумин. Оставшиеся клеточные лизаты хранили при температуре -80°С для последующего использования.
4. Получение лизата МВ
Культуральную среду после трансфекции или инфицирования переносили в 50-мл пробирку на описанном выше этапе «3. Получение клеточного лизата» и центрифугировали при 9000×g и температуре 4°С в течение 60 минут. Супернатант переносили в новую пробирку и подвергали ультрафильтрации с использованием Vivaspin® 20 (м.м. отсеч. 1000 кДа) (Sartorius, NY, USA, заменяя тем самым буфер TBS-D. TBS-D готовили путем смешивания 6,5 мл 1 M Hepes-KOH, pH 7,6, 7 мл 5 M NaCl и 0,25 мл 1 M KCl с соответствующим количеством ddH2O, добавления к этой смеси ddH2O до конечного объема 250 мл, и добавления в полученный таким образом раствор TBS 500 мкл 1M дитиотреитола (DTT) на каждые 50 мл раствора TBS перед использованием.
К 1-2 мл раствора, полученного из среды таким образом, добавляли равный объем раствора ПЭГ 8000/NaCl, и инкубировали эту смесь в течение ночи при температуре 4°С на качающейся платформе. Раствор ПЭГ 8000/NaCl (300 мл) получали путем смешивания 90 г ПЭГ 8000 и 180 мл 5 M NaCl с соответствующим количеством ddH2O, и добавления к этой смеси ddH2O до 300 мл (конечный объем).
Раствор, который инкубировали таким образом, центрифугировали при 13000 об./мин в течение 20 минут для получения осадка, содержащего микровезикулы (МВ). Супернатант удаляли и осадок ресуспендировали в 65 мкл лизирующего буфера и инкубировали при температуре 4°С в течение 2 часов на качающейся платформе для лизиса осадка. Этот раствор центрифугировали при 13000 об./мин в течение 20 минут и супернатант (лизаты МВ) переносили в новую пробирку. 5 мкл лизатов МВ добавляли в другую пробирку и измеряли OD600 для определения концентрации белка. Оставшиеся лизаты МВ хранили при температуре -80°С для последующего использования.
5. Иммунопреципитация (ИП) PTEN
70 мкл клеточных лизатов или лизатов МВ (содержащих 500 мкг белка) и 5 мкл мышиных моноклональных анти-PTEN антител (клон 6H2.1, 1 мг/мл, Upstate, USA) добавляли в новую пробирку, в которую затем добавляли TBS-D до 100 мкл (конечный объем) для получения каждого образца для ИП. Образец для ИП инкубировали в течение ночи при температуре 4°С на качающейся платформе. Затем в каждый образец для ИП добавляли 50 мкл раствора предварительно промытой белок-A-агарозы (Thermo Scientific, Illinois, USA), и эту смесь инкубировали в течение ночи при температуре 4°С на качающейся платформе. После инкубации микрогранулы агарозы промывали 1 мл TBS-D и центрифугировали при 6500 об./мин в течение 5 минут. Промывку затем повторяли три раза. После центрифугирования супернатант удаляли и микрогранулы агарозы ресуспендировали в 75 мкл TBS-D для получения раствора микрогранул агарозы.
6. Анализ PTEN
Анализ PTEN проводили по частично модифицированному протоколу для набора реагентов для анализа фосфатазы PTEN с малахитовым зеленым (Echelon Biosciences, Utah, USA). Для каждого анализа первоначальный объем 25 мкл доводили до конечного объема 100 мкл и измеряли поглощение при длине волны 620 нм с помощью считывающей системы Promega GloMax-Multi Jr. (Promega, USA). Анализ проводили, используя жидкий субстрат PtdIns(3,4,5)P3 (Echelon, Utah, USA), по инструкции фирмы-производителя. Для каждого анализа смешивали 23 мкл буфера TBS-D, 70 мкл раствора микрогранул агарозы, полученного на описанном выше этапе «5. Иммунопреципитация (ИП) PTEN», и 7 мкл 1мМ исходного раствора PtdIns(3,4,5)P3. Эту смесь инкубировали при температуре 37°С в течение 2 часов. В эту смесь добавляли 450 мкл раствора малахитового зеленого (Echelon, Utah, USA) комнатной температуры. Пробирку, содержащую смесь, оборачивали алюминиевой фольгой для защиты от света и инкубировали при комнатной температуре в течение 30-60 минут. Поглощение при длине волны 620 нм измеряли, используя раствор малахитового зеленого в качестве «пустого» образца и субстрат в качестве фосфатазного фона. Значения активности PTEN получали от трех различных трансфекций или инфицирований. Значения активности субстрата (фон) вычитали из полученных значений и получали среднее полученных значений.
7. Результаты
Результаты анализа PTEN клеток 293T, трансфицированных каждой плазмидой, показаны на Фигуре 11. Самая высокая активность PTEN в клеточных лизатах была подтверждена для клеток, трансфицированных вектором Regul., содержащим сильный промотор ЦМВ, который управлял транскрипцией гена PTEN. Активность PTEN также была высокой в случае трансфекции вектром Retro., содержащим промотор rNGFR, который управлял транскрипцией гена PTEN. Трансфекция вектром Retro., содержащим промотор hTERT, который управлял транскрипцией гена PTEN, обеспечила умеренное значение активности, по сравнению с векторами Regul. и Retro.. Для лизатов МВ самая высокая активность PTEN была подтверждена в случае трансфекции вектором Lenti., по сравнению с векторами Regul. и Retro..
На Фигуре 12 показано отношение активности PTEN в лизатах МВ к активности PTEN в клеточных лизатах, показанных на Фигуре 11. Трансфекция вектром Lenti. обеспечила самое высокое отношение активности PTEN в лизатах МВ (27,37%). Кроме того, следует отметить, что более высокое отношение активности PTEN в лизатах МВ наблюдалось в результате трансфекции вектором Empt., который, как и вектор Lenti., содержал промотор hTERT для генной транскрипции, но не содержал ген PTEN, по сравнению с векторами Regul. и Retro. Эти результаты показали, что трансфекция вектором Empt. стимулирует инкапсуляцию в МВ эндогенного PTEN клеток-хозяев, в то время как трансфекция вектором Lenti. стимулирует инкапсуляцию в МВ как эндогенного, так и трансгенного PTEN-продукта, тем самым, увеличивая высвобождение МВ во внеклеточную среду.
Кроме того, результаты анализа PTEN для клеток 293T, клеток HeLa и клеток CEM, инфицированных вирусной частицей RBL0213T, показаны на Фигуре 13. Клетки, инкубированные с рекомбинантной лентивирусной частицей RBL0213T, высвобождали большее количество МВ, чем клетки в случае трансфекции, независимо от того, является ли частица инфекционной или неактивной (ложное инфицирование) (Фигура 13, лизаты МВ). В отличие от этого, клеточные лизаты клеток, инфицированных рекомбинантной лентивирусной частицей RBL0213T или ложноинфицированных, имели активность PTEN практически равную активности PTEN в случае трансфекции (Фигура 13, клеточные лизаты). Однако повышенная активность PTEN наблюдалась в лизатах клеток HeLa, инфицированных вирусной частицей RBL0213T. Эти результаты показали, что инфицирование рекомбинантной лентивирусной частицей RBL0213T приводит к увеличению активности в лизатах МВ, но не в клеточных лизатах, по сравнению с трансфекцией вектором Lenti. и pRBL0213T. Кроме того, было показано, что в случае инфицирования лентивирусной частицей клетки клеточной линии HeLa рака шейки матки человека образуют и высвобождают большее количество трансгенного продукта PTEN, инкапсулированного в МВ, среди всех проанализированных клеточных линий (Фигура 13, лизаты МВ). Анализ PTEN раствора вирусной частицы RBL0213T показал, что значение активности PTEN было очень небольшим (OD620 = 0,065), что свидетельствует о том, что PTEN плохо инкорпорируется в вирусную частицу.
На Фигуре 14 показано отношение активности PTEN в лизатах МВ к активности PTEN в клеточных лизатах клеток 293T, трансфицированных рекомбинантным лентивирусным плазмидным вектором pRBL0213T, или клеток 293T, CEM или HeLa, инфицированных рекомбинантной лентивирусной частицей RBL0213T или ложноинфицированных. Инфицирование (Инф.) и ложное инфицирование (Контроль) показаны значениями, которые компенсируют неспецифичную стимуляцию высвобождения МВ в случае ложного инфицирования. Отношение активности PTEN в лизатах МВ к активности PTEN в клеточных лизатах было больше в случае инфицирования, чем в случае ложного инфицирования, что свидетельствует о том, что инфицирование рекомбинантной лентивирусной частицей RBL0213T усиливает высвобождение МВ из клеток. В отличие от отношения высвобождения МВ, равного 27,37%, в клетках, трансфицированных вектором Lenti. (Фигура 12), отношение высвобождения МВ достигало 38,75% в клетках, инфицированных рекомбинантной лентивирусной частицей RBL0213T (Фигура 14), что свидетельствует о том инфицирование рекомбинантной лентивирусной частицей RBL0213T также значительно стимулирует высвобождение МВ.
Описанные выше результаты показали, что клетки, трансфицированные рекомбинантным лентивирусным плазмидным вектором, содержащим промотор hTERT в лентивирусной РНК, внутриклеточно продуцируют большое количество микровезикул и обладают увеличенным высвобождением микровезикул, содержащих трансгенный продукт. Было показано, что клетки, инфицированные лентивирусной частицей, полученной с использованием такой плазмиды, обладают более сильным увеличенным высвобождением микровезикул.
Пример 8 Получение микровезикулы
Микровезикулу (МВ) настоящего изобретения получали способом, описанным ниже.
1. Трансфекция плазмидной ДНК или вирусное инфицирование
15 мл полной среды DMEM с высоким содержанием глюкозы (Hyclone, Utah, USA) (с добавлением 10% фетальной бычьей сыворотки, 100 единиц/мл пеницилина и 100 мкг/мл стрептомицина (Hyclone, Utah, USA)) добавляли в колбу T75, в которую затем инокулировали клетки клеточной линии 293Т эмбриональной почки человека. Клетки 293Т размножали в инкубаторе в атмосфере с 5% CO2 при температуре 37°С и наращивали количество культуры до десяти колб T75 путем пересевания. Клетки 293Т подвергали трансфекции плазмидной ДНК или вирусному инфицированию, как описано ниже.
(A) Трансфекция плазмидной ДНК
Следующие компоненты добавляли в 50-мл пробирку:
(a) 500 мкл раствора плазмидной ДНК, содержащего 170 мкг рекомбинантного лентивирусного плазмидного вектора pTHTN или pRBL001, описанного в Примере 1,
(b) 1650 мкл ddH2O, и
(c) 350 мкл 2 M CaCl2
Раствор осторожно перемешивали, и затем в него по каплям добавляли 2500 мкл 2×HEPBS при аккуратном перемешивании, чтобы избежать образования большого осадка, получая, таким образом, трансфекционную смесь. Пробирку, содержащую трансфекционную смесь, оставляли при комнатной температуре в течение 20 минут. Из десяти колб T75, в которых выращивали клетки 293T, удаляли среду. Затем в каждую колбу добавляли 12 мл свежей полной среды DMEM с высоким содержанием глюкозы. В каждую колбу постепенно добавляли 500 мкл трансфекционной смеси. Клетки инкубировали в инкубаторе в атмосфере с 5% CO2 при температуре 37°С в течение 8 часов.
(B) Вирусное инфицирование
Клетки 293T инфицировали путем совместной инкубации в инкубаторе в атмосфере с 5% CO2 при температуре 37°С в течение 16 часов с псевдотипированными рекомбинантным лентивирусными частицами THTN или THTD, полученными, как описано в Примере 2, (множественность заражения: 0,3-0,4).
2. Клеточная культура и сбор среды
Среду удаляли, и в каждую колбу добавляли 15 мл свежей полной среды DMEM с высоким содержанием глюкозы. Клетки инкубировали в инкубаторе в атмосфере с 5% CO2 при температуре 37°С в течение 60 часов и собирали среду.
3. Очистка
50-мл пробирку, содержащую собранную среду, центрифугировали при 3000 об./мин и температуре 4°С в течение 5 минут. Полученную таким центрифугированием надосадочную среду переносили в 250-мл высокоскоростную центрифужную пробирку и центрифугировали при 9000×g и температуре 4°С в течение 60 минут. Полученный супернатант подвергали 5-кратному концентрированию путем ультрафильтрации при помощи концентратора Vivaspin® 20 (1000 кДа отсечение по молекулярной массе (м.м. отсеч.)) (Sartorius, NY, USA). В полученный раствор добавляли раствор мМP, и МВ осаждали в течение ночи при температуре 4°С.
Раствор, в котором осаждались МВ, центрифугировали при 9000×g и температуре 4°С в течение 30 минут для получения осадка, содержащего МВ. Супернатант удаляли и осадок ресуспендировали в 10 мл 1×ФСБ для получения раствора, содержащего МВ.
МВ в аликвоте раствора подвергали пегилированию (Croyle et al., J. Virol., 2004, Vol. 78, p. 912-921). Пегилирование проводили путем добавления 0,4 мл пегилирующего раствора (33 мг/мл метокси ПЭГ сукцинимидилкарбоната NHS (mPEG-NHS, м.м. 10К (NANOCS, USA)), 30 мМ HEPES-KOH, pH 7,5, 500 мМ NaCl) в приблизительно 10 мл раствора, содержащего МВ, и инкубирования этой смеси при комнатной температуре в течение 60 минут на качающейся платформе.
Раствор, содержащий пегилированные или непегилированные МВ диализировали против 1×ФСБ при температуре 4°С при помощи кассеты Slide-A-Lyzer (20 кДа м.м. отсеч.) (Thermo Scientific, IL, USA), и 1×ФСБ заменяли свежим буфером каждые 24 часа в течение 2 суток. Диализированный таким образом раствор МВ концентрировали в 30 раз с использованием фильтра AmiconUltra 15 (100 кДа м.м. отсеч.) (Millipore, MA, USA). Концентрированный раствор вирусных частиц пропускали через шприцевой фильтр с размером пор 0,45 мкм, и полученный препарат хранили при температуре -80°С.
Таким образом, микровезикулу, содержащую в качестве трансгенного продукта частичный фрагмент большого Т-антигена вируса SV40, получали с использованием рекомбинантного лентивирусного плазмидного вектора pTHTN или псевдотипированной рекомбинантной лентивирусной частицы THTN. Кроме того, микровезикулу Lenti-mv2010/CDC6 кшРНК, содержащую трансгенный продукт кшРНК CDC6, получали с использованием рекомбинантного лентивирусного плазмидного вектора pRBL001 или псевдотипированной рекомбинантной лентивирусной частицы THTD.
Пример 9 Обнаружение белка, содержащегося в микровезикуле
В этом Примере изучали белки, содержащиеся в микровезикулах. Также изучали опосредованную микровезикулами доставку веществ в клетки.
Клетку клеточной линии 293Т эмбриональной почки человека трансфицировали плазмидами (основа pcDNA3.1), экспрессирующими N-терминально c-myc-меченную ДНК флэп-эндонуклеазу 1 человека (c-myc-FEN-1) для эктопической экспрессии c-myc-меченной FEN-1. Было показано, что FEN-1 функционирует в качестве важного клеточного хелперного фактора для созревания вирусной геномной ДНК ВИЧ-1 и ее интеграции в геном хозяина. Микровезикулу Lenti-mv2010, полученную из клеток, инфицированных вирусной частицей THTN, полученную как описано в Примере 8, совместно инкубировали с некоторыми из клеток 293T, трансфицированных экспрессирующей c-myc-FEN-1 плазмидой, или с нетрансфицированными клетками 293T. После инкубации в течение 60 часов из клеток получали ядерные и цитоплазматические экстракты. Кроме того, собирали клеточную культуральную среду и получали МВ, способом (за исключением пегилирования), описанным на этапе «3. Очистка» Примера 8. Полученные ядерные и цитоплазматические экстракты и МВ анализировали при помощи Вестерн-блоттинга.
Результаты обнаружения белка Vpu с использованием анти-Vpu ВИЧ-1 антител показаны на Фигуре 15A. Белок Vpu был обнаружен в цитоплазматических экстрактах клеток 293T, которые трансфицировали экспрессирующей c-myc-FEN-1 плазмидой и затем совместно инкубировали с Lenti-mv2010, и в МВ, высвобожденных из этих клеток (Фигура 15A, THTN), в то время как этот белок не был обнаружен в клетках 293T, которые трансфицировали экспрессирующей c-myc-FEN-1 плазмидой, но не инкубировали совместно с Lenti-mv2010 (Фигура 15A, pcDNA3.1). Vpu представляет собой белок, который подавляет тетерин- или CD317-опосредованное прикрепление вирионов к клеточным мембранам, усиливая тем самым высвобождение вирионов ВИЧ-1 (Sauter et al., Cell, 2010, Vol. 141, p. 392-398). Это показало, что: микровезикула Lenti-mv2010, полученная из клеток 293T, инфицированных рекомбинантной лентивирусной частицей THTN, содержит вирусный белок Vpu; содержимое было доставлено в другие клетки посредством микровезикулы; и микровезикулы, высвобожденные из клеток, которые получили доставленное содержимое, также содержали белок Vpu.
Результаты обнаружения белка RT с использованием анти-обратная транскриптаза (RT) ВИЧ-1 антител показаны на Фигуре 15B. Белок RT был обнаружен в микровезикуле Lenti-mv2010, полученной из клеток 293T, инфицированных рекомбинантной лентивирусной частицей THTN (Фигура 15B, Lenti-mv2010). В отличие от этого, белок RT не был обнаружен в МВ, полученных из клеток 293T, ложноинфицированных вирусной частицей THTN, химически инактивированной AT-2 (Фигура 15B, Контроль). Белок RT был также обнаружен в цитоплазматических экстрактах (Фигура 15B, THTN-цитоплазм) клеток 293T, которые трансфицировали экспрессирующей c-myc-FEN-1 плазмидой и затем инкубировали с Lenti-mv2010, и в МВ (Фигура 15B, THTN-МВ), полученных из этих клеток. В отличие от этого, белок RT не был обнаружен ни в МВ (Фигура 15B, МВ/pcDNA3.1), полученных из клеток 293T, которые трансфицировали экспрессирующей c-myc-FEN-1 плазмидой, но не инкубировали совместно с Lenti-mv2010, ни в необработанных клетках 293T ((Фигура 15B, 293T). Это показало, что: микровезикула Lenti-mv2010, полученная из клеток 293T, инфицированных рекомбинантной лентивирусной частицей THTN, содержит вирусный белок RT; содержимое было доставлено в другие клетки посредством микровезикулы; и микровезикулы, высвобожденные из клеток, которые получили доставленное содержимое, также содержали белок RT.
Результаты обнаружения эндогенного FEN-1 и чужеродного FEN-1 с использованием анти-FEN-1 антител показаны на Фигуре 15C. эндогенный FEN-1 и чужеродный FEN-1 (т.е. c-myc-FEN-1) обнаружили в ядрах и цитоплазме, полученных из клеток 293T, которые трансфицировали экспрессирующей c-myc-FEN-1 плазмидой и затем совместно инкубировали с Lenti-mv2010, и в МВ, высвобожденных из этих клеток (Фигура 15C, THTN/FEN-1). В случае клеток 293T, которые совместно инкубировали с Lenti-mv2010, но не трансфицировали экспрессирующей c-myc-FEN-1 плазмидой, в ядрах и цитоплазме и МВ, полученных из этих клеток, обнаружили только эндогенный FEN-1 (Фигура 15C, THTN). Это показало, что как эндогенный клеточный белок (FEN-1), так и чужеродный белок (c-myc-FEN-1) инкапсулируются в микровезикулы.
Описанные выше результаты показали, что вирусные белки и другие эндогенные клеточные и чужеродные белки инкапсулируются в микровезикулы, особенно в экзосомы, вероятно с помощью вирусного белка Vpu и клеточной системы транспорта белков, называемой клеточный эндосомальный сортинговый комплекс, необходимый для транспортного пути (ESCRT) в клетках. Эти результаты также показали, что инкапсулированное в микровезикулах содержимое доставляется в другие клетки.
Пример 10 Опосредованная микровезикулами доставка веществ
Доставку (генную трансдукцию) веществ при помощи очищенных пегилированных МВ изучали с использованием культивируемых клеток.
Содержащую c-myc-FEN-1 пегилированную МВ Lenti-mv2010/c-myc-FEN-1 получали способом для получения микровезикул посредством инфицирования THTN, описанным в Примере 8, за исключением того, что используемые клетки заменяли на клетки 293T, трансфицированные экспрессирующей c-myc-FEN-1 плазмидой. Lenti-mv2010/c-myc-FEN-1 вводили в клетки клеточной линии HeLa рака шейки матки человека путем совместной инкубации. Клетки HeLa, инкубированные совместно с МВ, подвергали непрямому иммунофлуоресцентному окрашиванию с использованием анти-c-myc антител.
Результаты показаны на Фигуре 16. Фигура 16A представляет собой изображение, после окрашивания анти-c-myc антителами. Фигура 16B представляет собой изображение, после окрашивания ДАПИ. Фигура 16C представляет собой наложенное изображение Фигур 16A и 16B. Фигура 16D представляет собой изображение, полученное из наложенного изображения при помощи программы кривой изменения цвета в методе «настройки» изображения программы Photoshop(R) (Adobe Systems Inc.). На Фигуре 16D более интенсивное красное окрашивание соответствует увеличению уровня c-myc-FEN-1, а зеленовато-желтое окрашивание соответствует отсутствию c-myc-FEN-1. Много красных клеток, экспрессирующих c-myc-FEN-1, наблюдалось на Фигуре 16D, указывая на то, что c-myc-FEN-1 был доставлен в большее количество клеток в результате совместной инкубации с Lenti-mv2010/c-myc-FEN-1. Было показано, что пегилированные микровезикулы захватываются большим количеством клеток и способны доставлять инкапсулированное в микровезикулах содержимое в другие клетки, так как они являются очень стабильными и активными.
Пример 11 Подавление пролиферации раковых клеток при помощи микровезикулы, содержащей кшРНК CDC6
Изучали подавляющее пролиферацию раковых клеток действие микровезикул, содержащих кшРНК CDC6 (Lenti-mv2010/CDC6 кшРНК).
Клетки клеточной линии MCF7 рака молочной железы человека, клетки клеточной линии LA-N-2 нейробластомы человека и клетки клеточной линии KANR нейробластомы по отдельности инкубировали с микровезикулой Lenti-mv2010/CDC6 кшРНК, содержащей кшРНК CDC6, полученной, как описано в Примере 8. В качестве контроля использовали микровезикулу Lenti-mv2010, содержащую частичный фрагмент большого Т-антигена вируса SV40, полученную, как описано в Примере 8. Затем пролиферацию клеток изучали с использованием набора реагентов для MTT-анализа выживаемости и пролиферации клеток (ScienCell, USA). В этом анализе пролиферацию культивируемых клеток оценивали количественно, используя в качестве показателя поглощение субстрата, восстанавливаемого живыми клетками.
Результаты показаны на Фигуре 17. По горизонтальной оси показано количество инкубированных МВ на клетку, а по вертикальной оси показано отношение (индекс роста) полученного значения оптической плотности к значению оптической плотности, полученному без инкубации с МВ. Пролиферация клеток MCF7 и клеток LA-N-2 значительно подавлялась инкубацией с Lenti-mv2010/CDC6 кшРНК, но не подавлялась инкубацией с Lenti-mv2010 (контроль) (Фигура 17). С другой стороны, пролиферация клеток KANR не подавлялась даже инкубацией с Lenti-mv2010/CDC6 кшРНК. Клетки MCF7 и клетки LA-N-2 имели повышенный уровень экспрессии CDC6, в то время как в клетках KANR экспрессия CDC6 практически отсутствовала. Это показало, что кшРНК CDC6, содержащаяся в микровезикулах, вызывала нокдаун CDC6, что приводило к подавлению пролиферации раковых клеток.
Описанные выше результаты показали, что микровезикула, содержащая кшРНК CDC6, способна доставлять кшРНК CDC6 в раковые клетки, экспрессирующие CDC6, подавляя тем самым их пролиферацию.
Пример 12 Молекулярный механизм подавления пролиферации раковых клеток при помощи микровезикулы, содержащей кшРНК CDC6
Изучали молекулярный механизм, лежащий в основе подавления пролиферации раковых клеток при помощи Lenti-mv2010/CDC6 кшРНК. Предыдущее исследование выявило, что: высокий уровень белка CDC6 связан с онкогенной активностью при раке человека; и уровни белка CDC6 и белка-супрессора опухолевого роста p16INK4a имеют обратную корреляцию между собой (Gonzalez, S. et al., Nature, 2006, p. 702-706). Прогрессирование клеточного цикла у клеток MCF7 не подавляется даже в присутствии p16INK4a. Это свидетельствует об инактивации p16INK4a-Rb пути в раковых клетках. Однако p16INK4a-Rb путь может быть снова активирован в раковых клетках MCF7 через Lenti-mv2010/CDC6 кшРНК опосредованный нокдаун CDC6. Таким образом, был проведен нерадиоактивный иммунопреципитационный анализ киназной активности для изучения восстановления p16INK4a-Rb пути. CDC6 был обнаружен в киназном комплексе ЦЗК4, и он необходим для фосфорилирования Rb-c.
CDC6 в клетках MCF7 удаляли (нокдаун) путем инкубации с Lenti-mv2010/CDC6 кшРНК. В результате было фосфорилирование Rb-c ингибировано. Нокдаун CDC6 увеличил ЦЗК-ингибирующую активность p16INK4a, по меньшей мере, в 25 раз. Это показало восстановление p16INK4a-Rb пути.
Описанные выше результаты показали, что микровезикула, содержащая кшРНК CDC6, вызывала нокдаун CDC6 в раковых клетках и снова активировала p16INK4a-Rb путь, и таким образом осуществляла свою функцию по подавлению пролиферации раковых клеток.
Пример 13 Вестерн-блоттинг анализ
В этом примере белок PTEN обнаруживали при помощи Вестерн-блоттинга в белковых экстрактах клеток, трансфицированных кодирующими PTEN векторами, и в экстрактах микровезикул, полученных при культивировании этих клеток. В качестве вектора были использованы плазмидные векторы pGL3-1375, pcDNA3.1/CMV-hPTEN, pRBL016Bn и pRBL0213T, описанные в Примере 7 и Фиг.10, и вектор на основе лентивирусной частицы RBL0213T, описанный в Примере 2 и 7.
1. Получение белковых экстрактов
Примерно 1×106 клеток 293T трансфицировали вышеуказанными различными плазмидными векторами. После 60 часов трансфекции клетки собирали, осаждали, гомогенизировали, и отбирали супернатант для приготовления экстрактов клеточных белков, как описано на этапе «3. Получение клеточного лизата» Примера 7.
Также собирали среду после 60 часов трансфекции, и получали из нее лизаты микровезикул, как описано на этапе «4. Получение лизата МВ» Примера 7.
Кроме того, примерно 1×106 клеток 293T инфицировали вектором на основе лентивирусной частицы RBL0213T с МЗ 0,3 в 25 мл полной среды DMEM, как описано на этапе «2. Инфицирование вирусной частицей RBL0213T3» Примера 7. После инфицирования в течение 60 часов экстракты клеточных белков получали, как описано на этапе «3. Получение клеточного лизата» Примера 7. Также собирали среду после инфицирования в течение 60 часов, и получали из нее лизаты микровезикул, как описано на этапе «4. Получение лизата МВ» Примера 7.
Полученные экстракты клеточных белков (примерно 20 микрограмм белка) или лизаты микровезикул (примерно 50 микрограмм белка) смешивали 2x ДСН буфером для нанесения (4% ДСН, 250 мМ Трис-HCl, рН 6,8, 3% β-меркаптоэтанола, 15% глицерина, 0,05% бромфенолового синего) и инкубировали в кипящей водяной бане в течение 15 минут. Затем образцы белка разделяли при помощи электрофореза в 12% ДСН-ПААГ (белковый гель PreciseTM), используя систему для электрофореза с минигелем для белков OwlTM P82 (Thermo Scientific). После электрофореза белки переносили из геля на ПВДФ мембрану и инкубировали с зондом, в качестве которого использовали мышиное моноклональное анти-PTEN антитела (первичное антитело; клон 6H2.1, MilliPore), а затем с козьим антимышиным вторичным антителом (разбавление 1:5000, MilliPore), для обнаружения белка PTEN.
Результаты обнаружения в экстрактах клеточных белков (дорожки 1-5) и лизатах микровезикул (дорожки 6-10) показаны на Фигуре 18. Дорожки 1 и 6 соответствуют клеткам, трансфицированным pGL3-1375 (Empt.). Дорожки 2 и 7 соответствуют клеткам, трансфицированным pcDNA3.1/CMV-hPTEN (Regul.). Дорожки 3 и 8 соответствуют клеткам, трансфицированным pRBL016Bn (Retro.). Дорожки 4 и 9 соответствуют клеткам, трансфицированным pRBL0213T (Lenti.). Дорожки 5 и 10 соответствуют клеткам, инфицированным RBL0213T.
Эндогенный PTEN был обнаружен (дорожки 1 и 6) и трансгенный продукт PTEN был обнаружен в экстрактах клеточных белков (дорожки 2-5) и в лизатах микровезикул (дорожки 7-10). Этот результат показывает, что трансгенный продукт PTEN синтезировался и инкапсулировался в микровезикулы, и эти микровезикулы (генно-инженерные микровезикулы) высвобождались из клеток.
Уровни трансгенного продукта PTEN, полученного из клеток, трансфицированных Regul., Retro. или Lenti. (дорожки 2-4), были повышены в 2-3 раза по сравнению уровнем PTEN, полученного из клеток, трансфицированных Empt. (дорожка 1). Кроме того, уровень трансгенного продукта PTEN, полученного из клеток, инфицированных лентивирусным вектором RBL0213T (дорожка 5), был значительно выше уровня эндогенного PTEN (дорожка 1).
Кроме того, уровень трансгенного продукта PTEN в лизатах микровезикул, полученных с использованием клеток, инфицированных лентивирусным вектором RBL0213T (дорожка 10), был явно повышен по сравнению с уровнем трансгенного продукта PTEN в лизатах микровезикул, полученных с использованием клеток, трансфицированных Empt., Regul., Retro. или Lenti. (дорожки 6-9). Это указывает на то, что инфицирование подобным вирусной частице лентивирусным вектором настоящего изобретения является более подходящим для усиления продуцирования и высвобождения микровезикул, содержащих трансгенные продукты.
Пример 14 Ингибирующее действие генно-инженерных микровезикул на рост опухоли
В этом Примере действие генно-инженерных микровезикул, содержащих продукт гена-супрессора опухолевого роста (в настоящем описании также именуются Цитомокс), направленное на ингибирование роста опухоли, изучали с применением прямой инъекции микровезикул в опухоли.
1. Получение исследуемого образца
Следующие исследуемые образцы использовали для внутриопухолевой инъекции.
i) Цитомокс HD (emvp 130001)
На основе раздела «4. Получение лизата МВ» Примера 7 и Примера 8, клетки клеточной линии 293Т эмбриональной почки человека трансфицировали обоими лентивирусными векторами pRBL001 и pRBL0203 и собирали среду путем осаждения культивируемых клеток и получали микровезикулы. Эти микровезикулы назвали Цитомокс HD. Лентивирусный вектор pRBL001 описан в Примере 1, и он использовался здесь в качестве вектора для получения кшРНК CDC6. Лентивирусный вектор pRBL0203 представляет собой плазмиду, содержащую ген p16INK4a человека, который был встроен в основу pTHTK под контролем промотора hTERT, и он был получен таким же способом, как в Примере 1. В частности, pCMV p16INK4a (Плазмида 10916; SEQ ID NO: 25; Medema et al., Proc. Natl. Acad. Sci. USA., (1995) 92(14): 6289-6293), которая представляет собой плазмидную конструкцию, содержащую кДНК p16INK4a, расщепляли EcoR I и Xho I и полученный фрагмент (кДНК p16 примерно 0,45 т.п.н.) субклонировали в сайт Hind III-Xho I плазмиды pEND-HTPl, в которую 1,5 т.п.н. промотора hTERT из pGL3-1375 было клонировано в сайт Mlu I-BamH I. Фрагмент ДНК, содержащий промотор hTERT и кДНК p16INK4a, расположенную после него, расщепляли Mlu I и Xho I и фрагмент размером 2,0 т.п.н. встраивали в основу pTHTK для получения вектора pRBL0203.
ii) Цитомокс p53 (emvp 130003)
На основе Примера 8, клетки клеточной линии 293Т эмбриональной почки человека трансфицировали вектором pGLQ-p53EX и собирали среду путем осаждения культивируемых клеток и получали микровезикулы. Вектор pGLQ-p53EX представляет собой плазмидный экспрессионный вектор, содержащий ген p53 человека (SEQ ID NO: 26; кодирующая последовательность в GenBank под регистрационным номером BC003596). В частности, субклонирующий вектор pBS-TP53PU, содержащий фрагмент кДНК гена-супрессора опухолевого роста p53 человека размером примерно 2,0 т.п.н., полученный из плазмиды pBSH19, содержащей кДНК p53, линеаризировали в сайте Hind III на его 3’-конце. У полученного сайта Hind III «тупили» концы, после чего проводили рестрикцию вектора эндонуклеазой Sal I. Основу очищали и лигировали в фрагментом ДНК размером примерно 0,3 т.п.н. (который имел «затупленный» конец сайта Xba I и конец сайта Sal I), выделенным из pGL3-378, для получения pBS-TP53PA. Фрагмент кДНК p53 вырезали из pBS-TP53PA рестриктазами Stu I и Sal I и встраивали в вектор pGL3-378 в сайт, который имел один «затупленный» конец сайта Xba I и другой конец сайта Sal,I для получения pGLQ-3UTR. Фрагмент кДНК p53 размером примерно 1,4 т.п.н. (содержащий полноразмерную кодирующую область гена p53 человека) вырезали из другого субклонирующего вектора pUTK-p53TT рестриктазами Hind III и Xho I и встраивали в pGLQ-3UTR для получения pGLQ-p53EX.
iii) Цитомокс PTEN (emvp 130006)
На основе раздела «4. Получение лизата МВ» Примера 7 и Примера 8, клетки клеточной линии 293Т эмбриональной почки человека трансфицировали лентивирусным вектором pRBL0213T и собирали среду путем осаждения культивируемых клеток и получали микровезикулы. Эти микровезикулы назвали Цитомокс PTEN. Лентивирусный вектор pRBL0213T описан в Примере 1, и он использовался здесь в качестве вектора для получения белка PTEN человека.
iv) Цитомокс EX (T 130075)
Как описано в Примере 2, рекомбинантные лентивирусные частицы RBL001 и RBL0203 получали, используя плазмиды pRBL001 и pRBL0203. На основе раздела «4. Получение лизата МВ» Примера 7 и Примера 8, клетки клеточной линии 293Т эмбриональной почки человека инфицировали лентивирусной частицей и собирали среду путем осаждения культивируемых клеток и получали микровезикулы. В частности, клетки трансфицировали обоими лентивирусными векторами pRBL001 и pRBL0203, как описано в Примере 8, и после 60 часов инфицирования собирали среду и клеточный дебрис удаляли путем центрифугирования. Среду подвергали высокоскоростному центрифугированию при 9000×g и температуре 4°С в течение 60 минут. Супернатанты собирали для очистки микровезикул, как описано в Примере 8, и следовые количества осадка ресуспендировали в 1×ФСБ и подвергали диализу с последующим концентрированием с использованием фильтра Amicon Ultra 15 (100 кДа м.м. отсеч.) (Millipore). Полученный препарат (микровезикулы) назвали Цитомокс EX.
v) Рекомбинантный интерферон α-2b (положительный контроль)
Рекомбинантный интерферон α-2b (Schering-Plough (Brinny) Co., Ireland) использовали в качестве положительного контроля.
2. Животные
Мышей линии BALB/c-nu (самки, всего восемьдесят животных, свободных от патогенной флоры, 14-16 грамм каждое животное) приобретали у Центра лабораторных животных Китайской Академии Медицинских Наук для использования в этом Примере.
Всех мышей содержали для разведения в виварии с вытяжной системой вентиляции. Температуру в виварии поддерживали на уровне 20-25°С (±<3°С) с относительной влажностью примерно 40-60%. Продолжительность освещения вивария составляла 12 часов в сутки с притоком чистого воздуха примерно 100%, освещенность рабочей зоны составляла примерно 150-300 лк, для зоны с клетками для животных - 100-200 лк. Концентрация аммиака составляла менее 14 мг/м3, уровень шума составлял менее 60 дБ. Ежедневно мышам давали свежие корм и питьевую воду (деионизированную и ультрафильтрованную).
3. Имплантация опухоли
Клетки клеточной линии Bcap-37 рака молочной железы человека имплантировали мыши BALB/c-nu, чтобы получить животное-хозяин, имеющее опухоль Bcap-37. В боксе биологической безопасности в стерильных условиях опухоль Bcap-37 извлекали из имеющего опухоль животного-хозяина, и ткани опухоли промывали 1×ФСБ. Области опухоли с хорошим ростом без распада опухоли собирали и разрезали скальпелем на маленькие кусочки в виде блоков размером 2 мм3. Блоки опухоли промывали 1×ФСБ и по одному имплантировали под кожу в правую подмышечную область каждой мыши при помощи игольной трубки. В общей сложности получили 8 групп (десять животных в группе). Через одиннадцать дней после имплантации опухоли вырастали до размера 110-120 мм3.
4. Инъекция микровезикулы (Цитомокс)
Микровезикулы вводили вышеуказанным мышам, имеющим рост опухоли. Внутриопухолевую инъекцию микровезикул проводили два раза в неделю в течение трех недель, где каждому животному в опухоли вводили инъекционный раствор, содержащий выделенные микровезикулы. То есть, всего было проведено шесть инъекций.
В случае Цитомокс HD и Цитомокс EX первые 3 инъекции были сделаны с более низкой дозой, находящейся в диапазоне от 10,0 до 30,0 мкг белка на кг массы тела, и последующие 3 инъекции были сделаны с более высокой дозой от 1,0 до 3,0 мг белка на кг массы тела. Группу, которая получала Цитомокс HD в дозе 10,0 мкг белка на кг массы тела и затем 1,0 мг белка на кг массы тела, в данном описании обозначали как группа, получавшая низкую дозу Цитомокс HD. Группу, которая получала Цитомокс HD в дозе 30,0 мкг белка на кг массы тела и затем 3,0 мг белка на кг массы тела, в данном описании обозначали как группа, получавшая высокую дозу Цитомокс HD. Кроме того, две другие группы, которые по аналогичной схеме получали Цитомокс EX, в данном описании обозначали как группа, получавшая низкую дозу Цитомокс EX, и группа, получавшая высокую дозу Цитомокс EX, соответственно. В случае Цитомокс p53 и Цитомокс PTEN первые 3 инъекции были сделаны с дозой 10,0 мкг белка на кг массы тела, и последующие 3 инъекции были сделаны с более высокой дозой 1,0 мг белка на кг массы тела (группа, получавшая Цитомокс p53, и группа, получавшая Цитомокс PTEN). В качестве положительного контроля вводили рекомбинантный интерферон α-2b в количестве 250×104 МЕ/кг массы тела два раза в неделю в течение трех недель (группа положительного контроля). В качестве отрицательного контроля в опухоли имеющих опухоль мышей вводили 1×ФСБ два раза в неделю в течение трех недель (группа отрицательного контроля). Животных в каждой группе умерщвляли путем смещения шейных позвонков через 48 часов после введения последней дозы. У мертвых мышей извлекали опухоли, фиксировали их и обрабатывали для патологоанатомического исследования.
В результате в этом эксперименте у подопытных животных не было отмечено острых токсических эффектов, даже при получении ими высокой дозы Цитомокс, также ни одно животное не погибло в результате инъекции генно-инженерных микровезикул (Цитомокс).
5. Определение массы тела, массы опухоли и размера опухоли
Массу тела мышей измеряли перед инъекцией исследуемого образца (масса тела с опухолью) и перед умерщвлением. Кроме того, массу тела мертвых мышей измеряли после извлечения из их тел опухолей (чистая масса тела). Массы удаленных опухолей также измеряли.
На основе измеренных значений масс опухолей эффективность ингибирования роста опухоли вычисляли следующим образом.
Эффективность ингибирования роста опухоли(I)(%)=[1-T/C]x100
T: Средняя масса опухоли (г) в каждой группе, получавшей каждый исследуемый образец (Цитомокс или положительный контроль)
C: Средняя масса опухоли (г) в группе отрицательного контроля.
Вычисленные значения эффективности ингибирования роста опухоли приведены в Таблице 1.
Как видно из Таблицы 1, прямая инъекция в очаг опухоли рекомбинантного интерферона α-2b вызывала ингибирование роста опухоли на 16,06%. Внутриопухолевая инъекция Цитомокс HD показала ингибирование роста опухоли на 18,37% при высокой дозе (3 мг белка на инъекцию на кг массы тела) и на 14,67% при низкой дозе (1 мг белка на инъекцию на кг массы тела). Таким образом, имело место дозозависимое увеличение ингибирующего рост опухоли действия. Аналогичным образом, внутриопухолевая инъекция Цитомокс EX показала ингибирование роста опухоли на 21,32% при высокой дозе (3 мг белка на инъекцию на кг массы тела) и на 2,83% при низкой дозе (1 мг белка на инъекцию на кг массы тела), и также наблюдалось дозозависимое увеличение ингибирующего рост опухоли действия. Внутриопухолевая инъекция Цитомокс p53 и Цитомокс PTEN также приводила к ингибированию роста опухоли. Эти результаты показывают, что генно-инженерные микровезикулы, содержащие трансгенные продукты, могут проникать в клетки, и трансгенные продукты могут осуществлять свои функции в опухолевых клетках.
Кроме того, измерение длины и ширины каждой опухоли у мыши проводили два раза в неделю после инъекции исследуемых образцов. Для проведения этих измерений использовали стандартный штангенциркуль. На основе измеренных значений объемы опухолей вычисляли следующим образом.
Объем опухоли (мм3) = (Длина опухоли × Ширина опухоли)2/2.
В группе, получавшей высокую дозу Цитомокс HD, группе получавшей высокую дозу Цитомокс EX, и группе, получавшей PTEN, после 4 дней инъекции исследуемых образцов (т.е., через 15 дней после имплантации опухоли) увеличение объема опухоли непрерывно ингибировалось по сравнению с группой отрицательного контроля в последний день измерения (т.е., через 30 дней после имплантации опухоли и через 20 дней после инъекции тестовых образцов). Также группе, получавшей p53, увеличение объема опухоли значительно ингибировалось после 12 дней инъекции исследуемого образца. В частности, в группе, получавшей высокую дозу Цитомокс HD, и группе получавшей высокую дозу Цитомокс EX, различия в объемах опухолей между этими группами и отрицательным контролем увеличивались с течением времени. В 31-й день после имплантации опухоли (последний день измерения) объем опухоли составлял 1250 мм3 для группы отрицательного контроля (ФСБ); примерно 1000 мм3 для группы, получавшей интерферон α-2b; примерно 950 мм3 для группы, получавшей Цитомокс HD (evmp 130001, высокая доза); примерно 1000 мм3 для группы, получавшей Цитомокс p53 (evmp 130003, высокая доза); примерно 1100 мм3 для группы, получавшей Цитомокс PTEN (evmp 130006, высокая доза); и примерно 1000 мм3 для группы, получавшей Цитомокс EX (T130075, высокая доза).
6. Патоморфологический анализ опухоли
Удаленные опухоли подвергали патологоанатомическому исследованию для оценки их морфологии. Критерий оценки был следующим:
«-», имеет место некроз в центральной области очага опухоли, но отсутствует фиброз;
«+», имеет место небольшая степень воспаления в опухолевых тканях с низкой степенью фиброза;
«++», имеет место воспаление в опухолевых тканях и распад опухолевых тканей, и фиброз наблюдается по всей опухоли; и
«+++», имеет место значительное воспаление с распадом опухолевой ткани, и наивысшая степень фиброза наблюдается по всей опухоли.
Результаты показаны в Таблице 2.
1 мг/кг
3 мг/кг
1 мг/кг
1 мг/кг
1 мг/кг
3 мг/кг
Как видно из Таблицы 2, в отличие от отрицательного контроля, введение микровезикул, содержащих продукт гена-супрессора опухолевого роста, в очаг опухоли часто вызывает распад опухолевой ткани и усиливает воспаление и фиброз в опухоли. В частности, инъекция высокой дозы Цитомокс HD или Цитомокс ED в очаг опухоли приводила к воспалению и распаду опухолевых тканей на более высоких уровнях и фиброзу в широком диапазоне, которые отличались от некроза опухоли в результате отсутствия ангиогенеза кровеносных сосудов в центральной области опухолевой массы. Пришли к выводу, что причиной распада опухолевых тканей стал фиброз, который блокировал распространение опухоли.
На Фигурах 19 и 20 показаны типичные морфологии, наблюдаемые в опухолях из соответствующих групп при патологоанатомическом исследовании.
7. Анализ опухоли при помощи Вестерн-блоттинга
Экстракцию белка осуществляли из опухолей, удаленных у животных из группы, получавшей Цитомокс PTEN, и группы отрицательного контроля, при помощи стандартных способов, и обнаружение белка в этих экстрактах проводили при помощи Вестерн-блоттинга, как описано в Примере 13. В результате PTEN обнаружили как в опухоли из группы отрицательного контроля (неинъецированные микровезикулами опухолевые ткани), так и в опухоли из группы, получавшей инъекцию микровезикул, (инъецированные микровезикулами опухолевые ткани). Однако уровень PTEN в инъецированных микровезикулами опухолевых тканях оказался в два раза выше, чем уровень PTEN в неинъецированных микровезикулами опухолевых тканях. Это означает, что эффективность проникновения микровезикул в клетки и доставки трансгенов/транс генных продуктов при помощи микровезикул является достаточно высокой. Кроме того, как видно из этих результатов, при повышенном уровне PTEN в инъецированных Цитомокс PTEN опухолевых клетках, рост опухоли ингибировался (см., 9,13% ингибирования в Таблице 1).
Промышленная применимость
Настоящее изобретение применимо для эффективного получения генно-инженерных микровезикул. Микровезикулы в соответствии с настоящим изобретением могут применяться для доставки биологических веществ в клетки.
Все публикации, патенты и патентные заявки, упомянутые в настоящем описании, включены в него полностью путем ссылки.
| название | год | авторы | номер документа |
|---|---|---|---|
| БИОПРОИЗВОДСТВО ЛЕНТИВИРУСНЫХ ВЕКТОРОВ | 2016 |
|
RU2729385C2 |
| ВЕКТОРНАЯ СИСТЕМА НА ОСНОВЕ ASLV | 2010 |
|
RU2566563C2 |
| УСОВЕРШЕНСТВОВАНИЕ ГЕНЕТИЧЕСКИХ КОНСТРУКЦИЙ ДЛЯ ПОВЫШЕНИЯ ЭФФЕКТИВНОСТИ АНТИВИЧ ТЕРАПИИ | 2013 |
|
RU2533817C1 |
| ГЕНЕТИЧЕСКИЕ КОНСТРУКЦИИ ДЛЯ АНТИВИЧ-ТЕРАПИИ | 2010 |
|
RU2426788C1 |
| РЕКОМБИНАНТНЫЙ ЛЕНТИВИРУСНЫЙ ВЕКТОР, КЛЕТКА-ХОЗЯИН, ТРАНСДУЦИРОВАННАЯ ЛЕНТИВИРУСНЫМ ВЕКТОРОМ, СПОСОБ ЕЕ ТРАНСДУКЦИИ И ПРИМЕНЕНИЕ | 2002 |
|
RU2305708C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ДЛЯ ЛЕЧЕНИЯ ГЕМОГЛОБИНОПАТИЙ | 2015 |
|
RU2707540C2 |
| Генетические конструкции и их смеси для антиВИЧ терапии | 2017 |
|
RU2666991C1 |
| СПОСОБЫ ПОЛУЧЕНИЯ И ПРИМЕНЕНИЯ МУЛЬТИПОТЕНТНЫХ КЛЕТОЧНЫХ ПОПУЛЯЦИЙ | 2008 |
|
RU2535365C2 |
| ВИРУСНЫЕ ВЕКТОРЫ С ЗАВИСИМОЙ ОТ УСЛОВИЙ РЕПЛИКАЦИЕЙ И ИХ ПРИМЕНЕНИЕ | 2001 |
|
RU2301260C2 |
| СПОСОБ ВРЕМЕННОЙ ТРАНСФЕКЦИИ ДЛЯ ПРОДУЦИРОВАНИЯ РЕТРОВИРУСА | 2016 |
|
RU2749717C2 |




















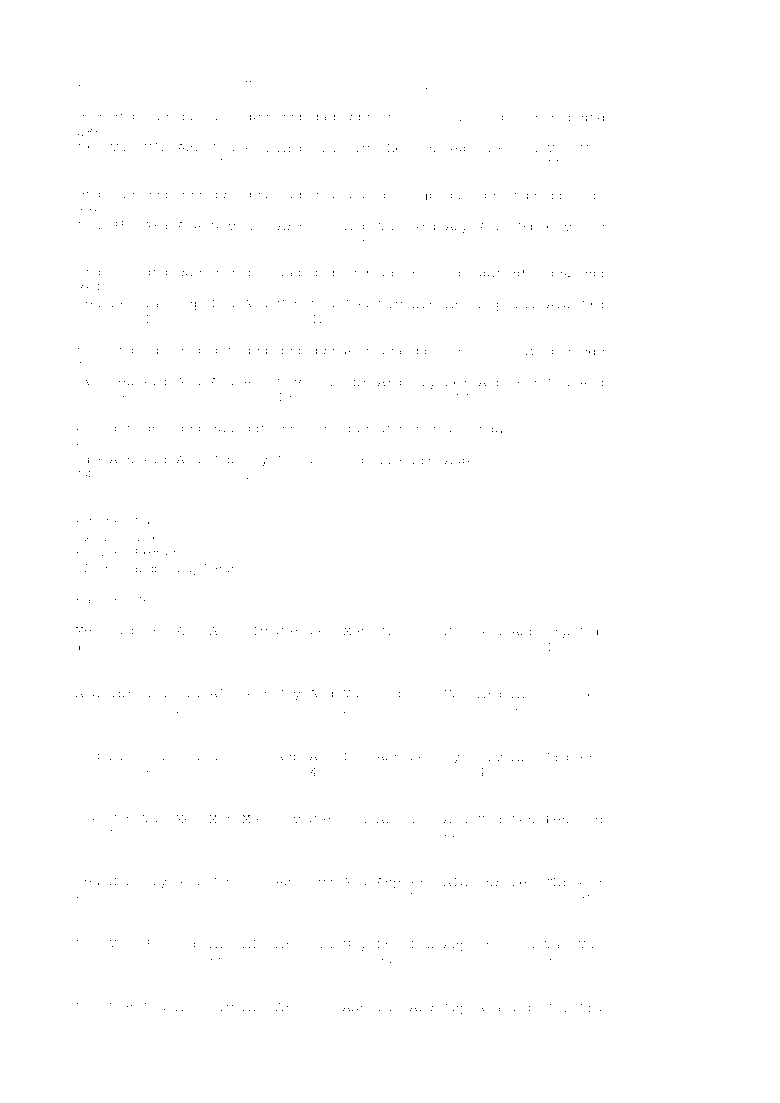




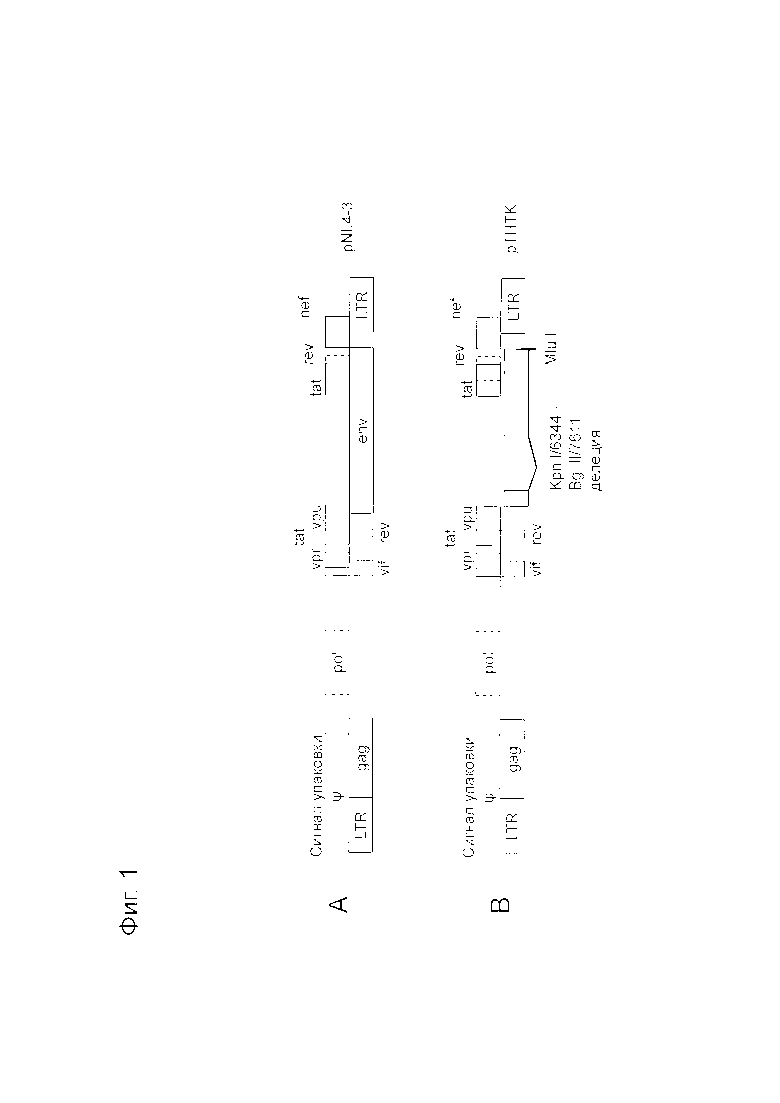
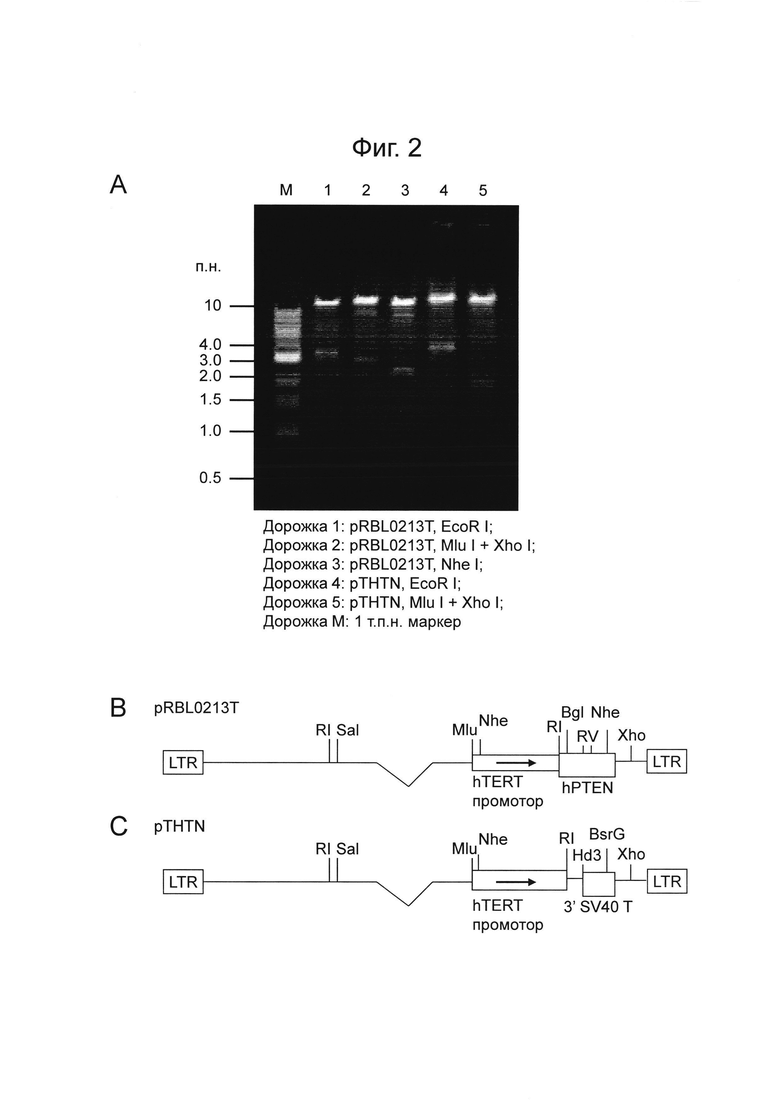
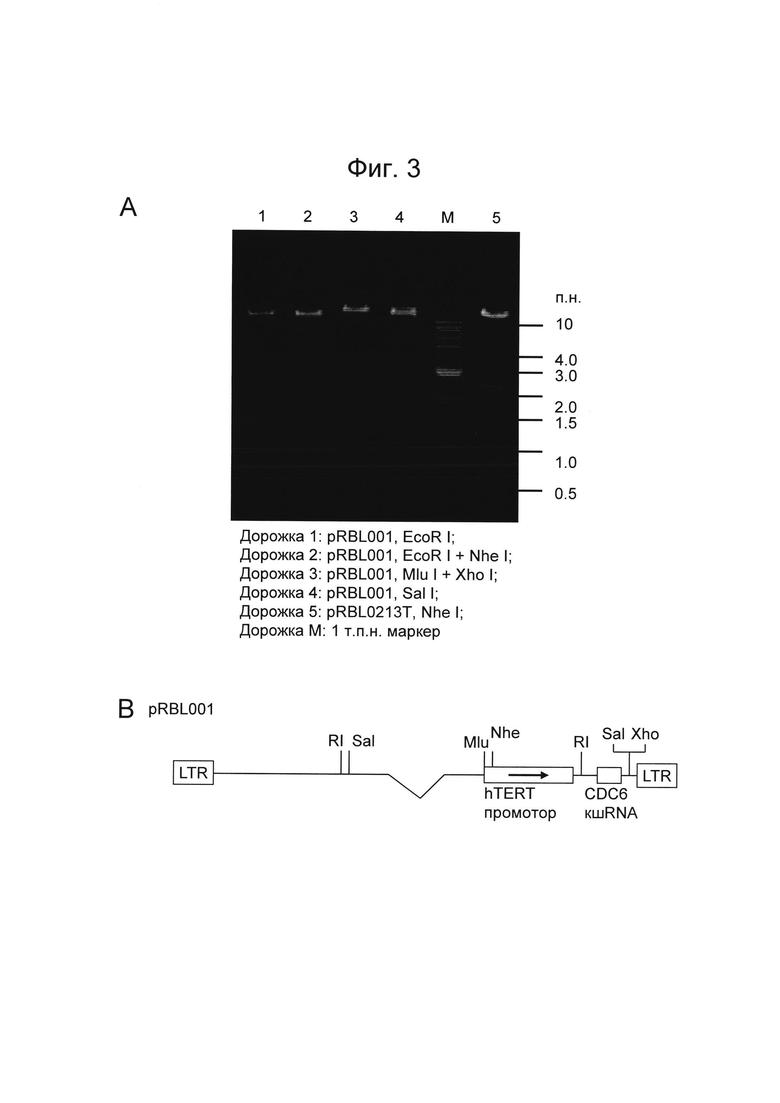

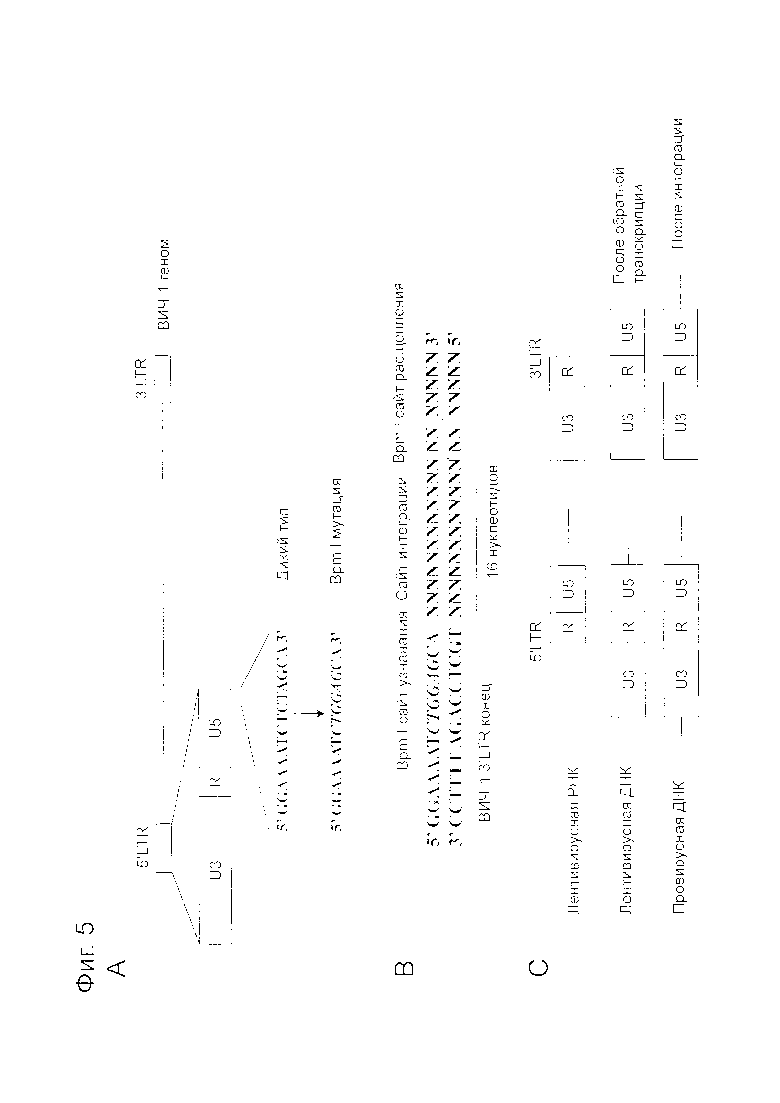


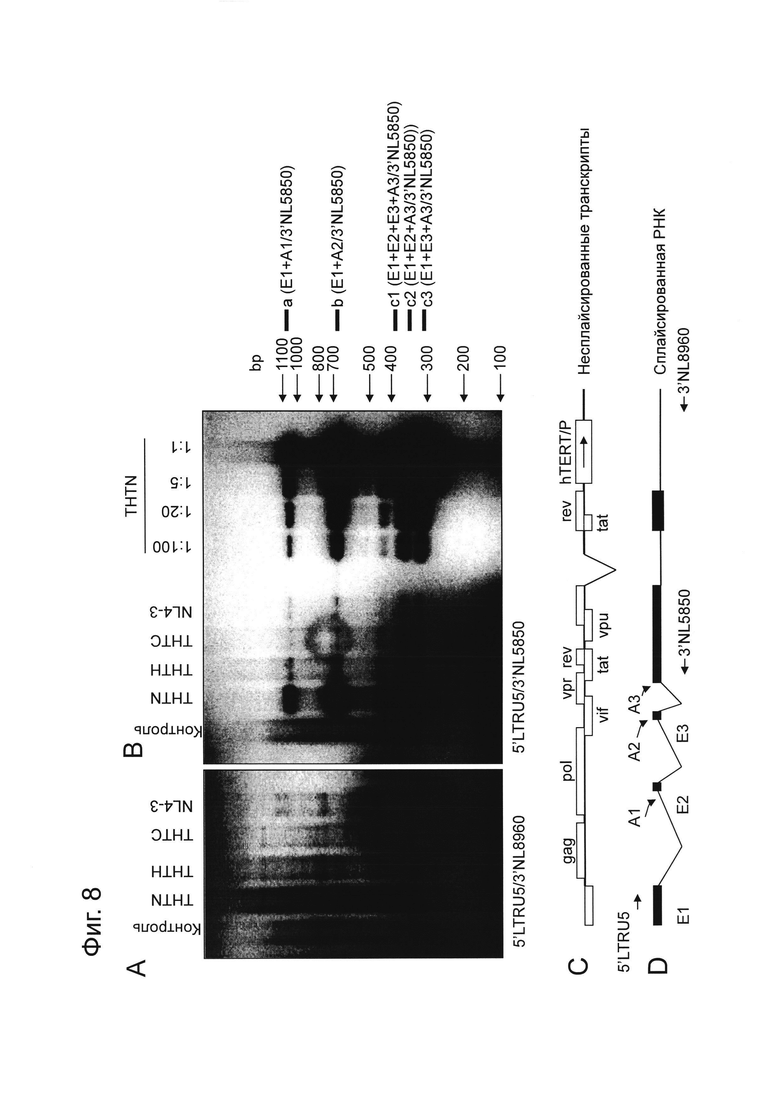
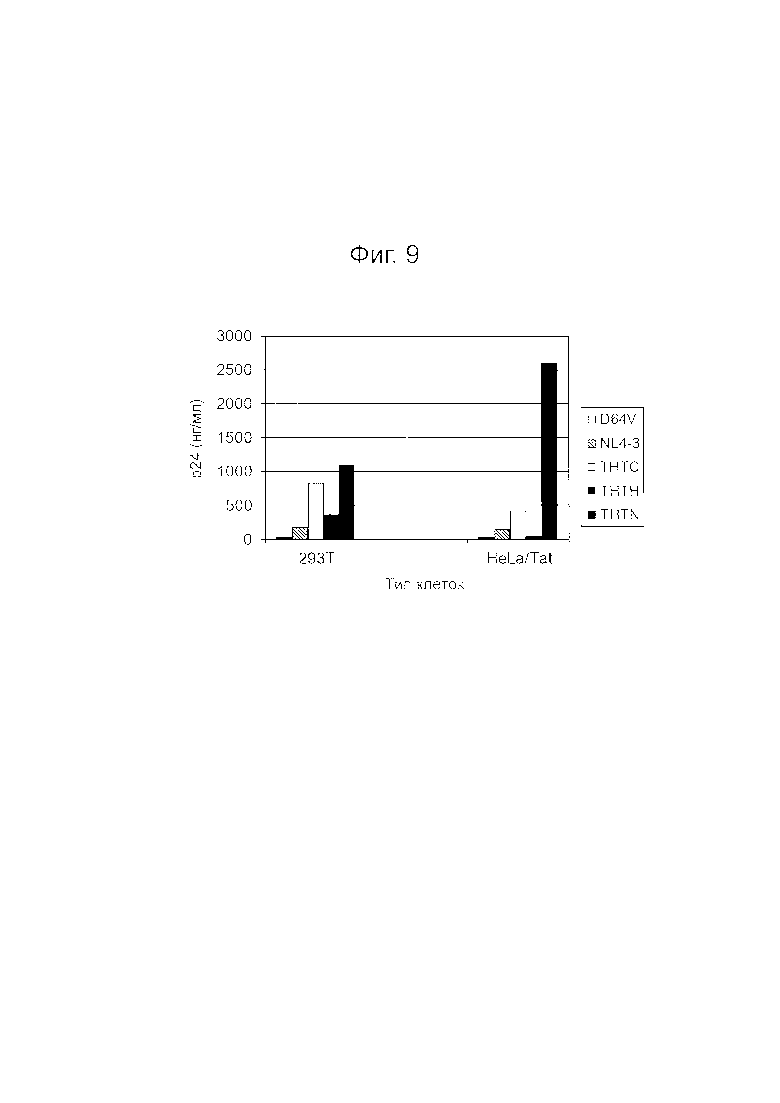
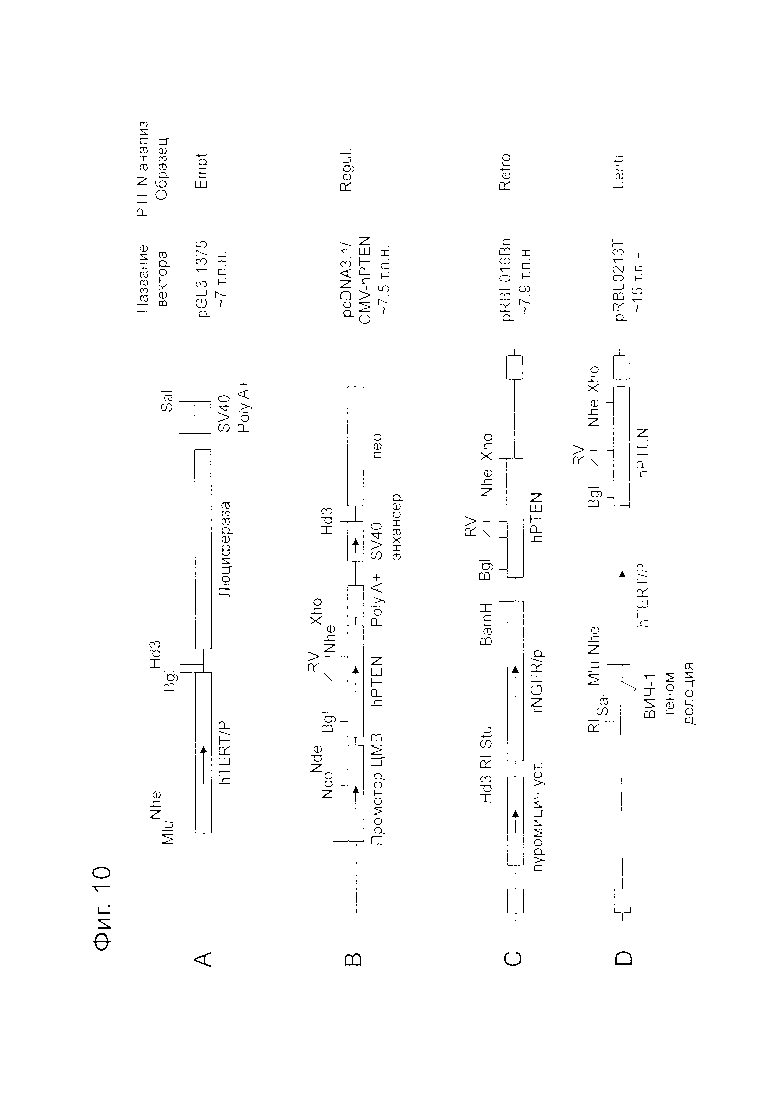
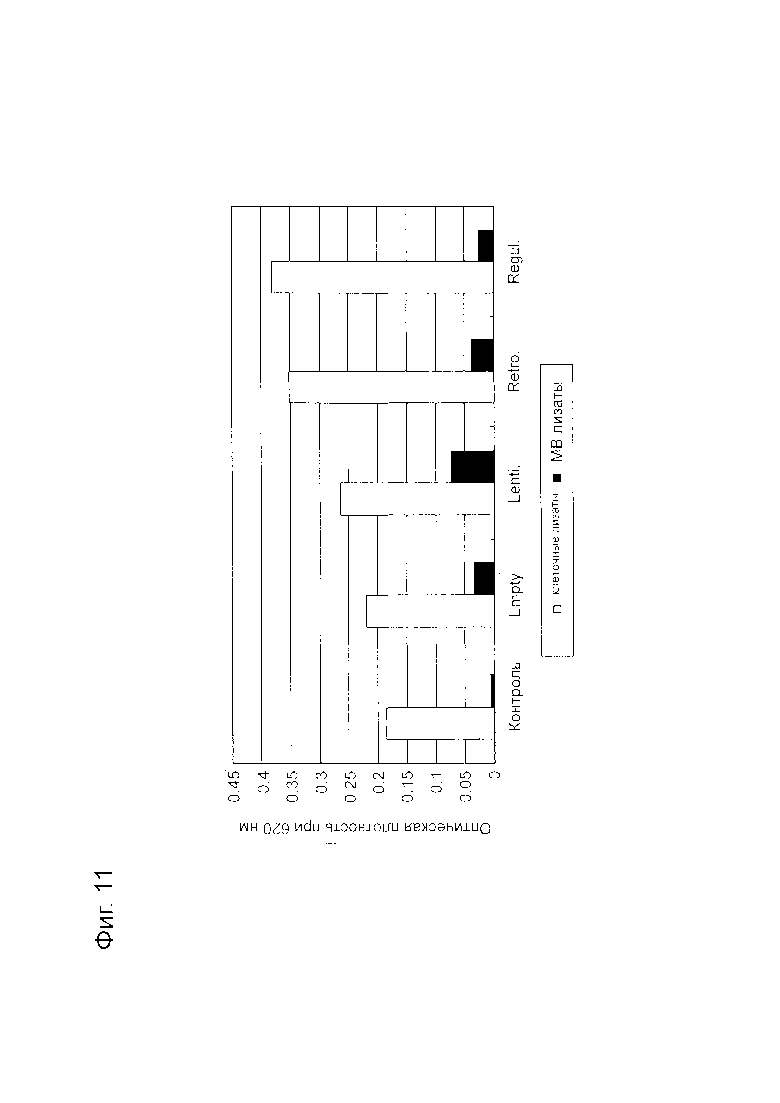

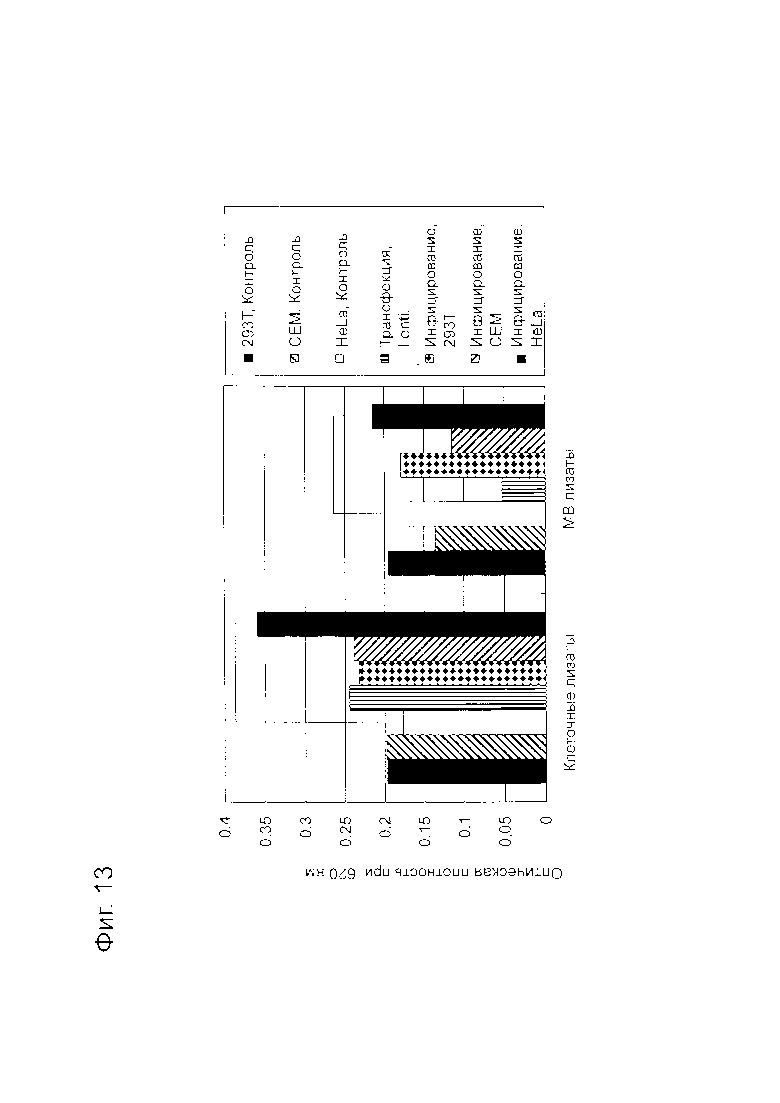

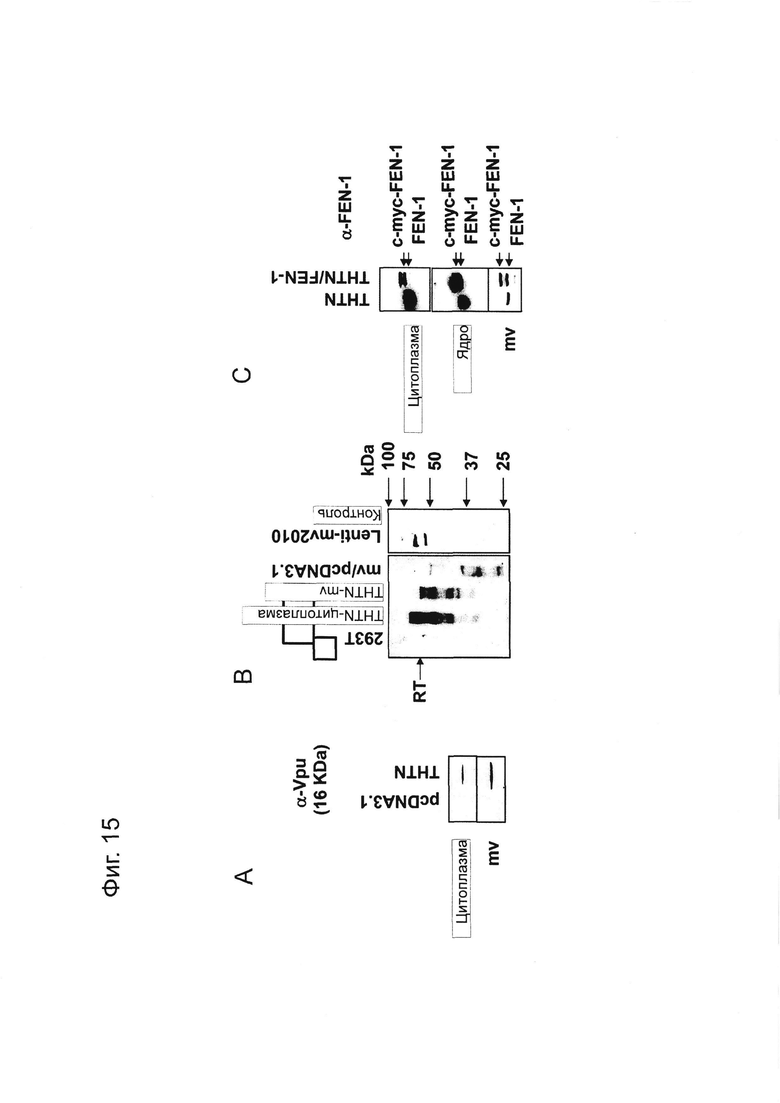

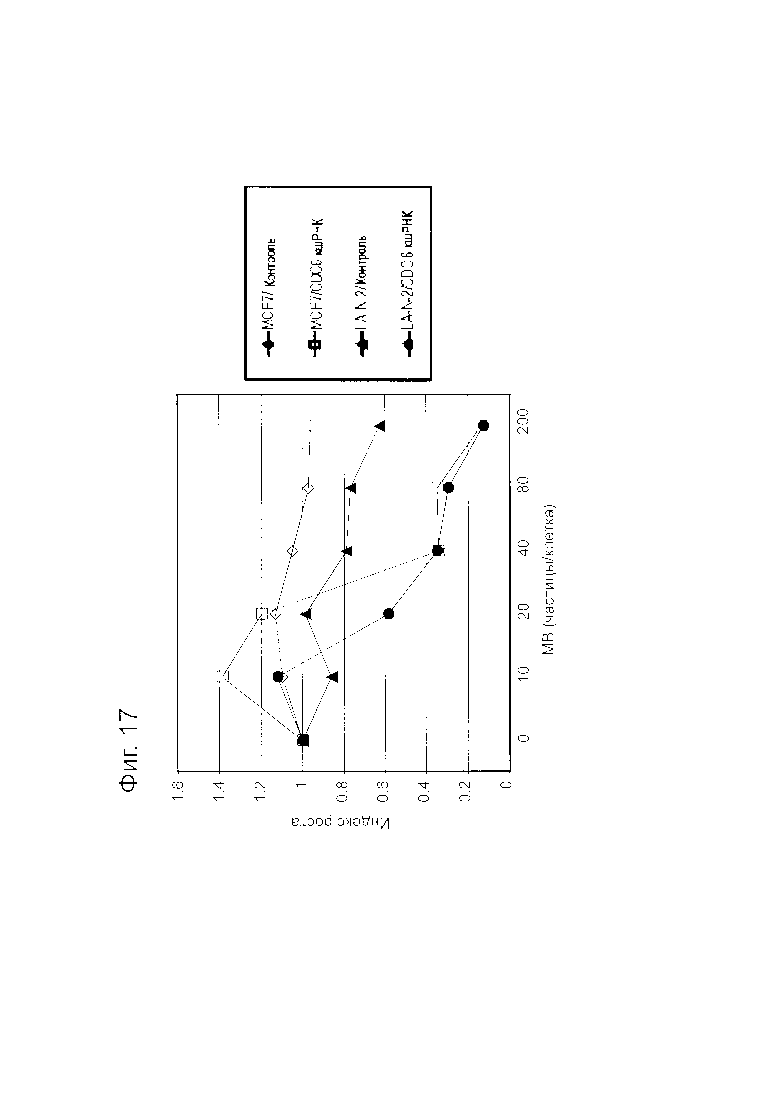
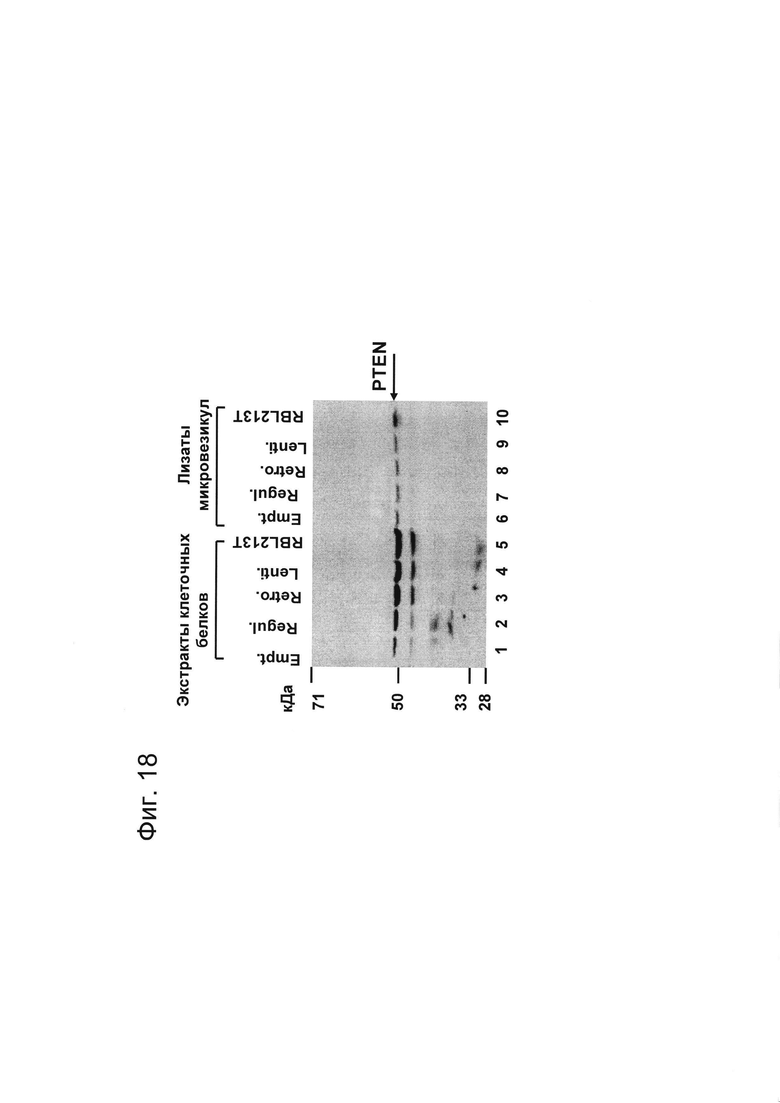
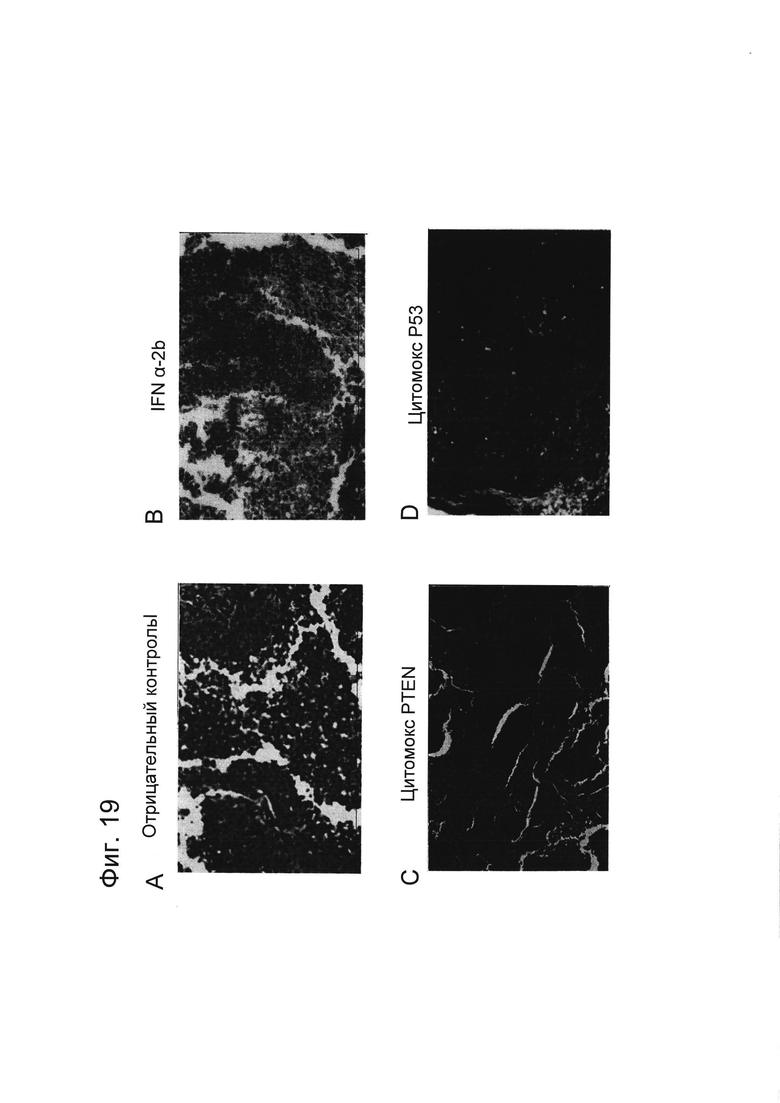
Изобретение относится к биотехнологии. Описан способ получения микровезикул, содержащих трансгенный продукт и/или лентивирусную РНК, содержащую трансген. Культивируют клетки человека, в которых при помощи лентивирусного вектора in vitro вводится трансген, для внеклеточного высвобождения микровезикул, содержащих трансгенный продукт и/или лентивирусную РНК, содержащую трансген. При этом указанный лентивирусный вектор является дефицитным по меньшей мере по одному гену структурного белка, выбранному из генов env, gag и pol, и содержит трансген под контролем промотора гена теломеразной обратной транскриптазы (TERT) в лентивирусной геномной последовательности. Кроме того, указанная лентивирусная РНК, указанный лентивирусный вектор и указанная лентивирусная геномная последовательность представляют собой РНК ВИЧ, вектор ВИЧ и геномную последовательность ВИЧ, соответственно. Собирают высвобожденные микровезикулы. Изобретение также относится к микровезикуле, полученной этим способом, и ее применению. Изобретение расширяет арсенал средств лечения рака. 5 н. и 12 з.п. ф-лы, 20 ил., 2 табл., 14 пр.
1. Способ получения микровезикул, содержащих трансгенный продукт и/или лентивирусную РНК, содержащую трансген, включающий этапы:
культивирования клетки человека, в которую при помощи лентивирусного вектора in vitro вводится трансген, для внеклеточного высвобождения микровезикул, содержащих трансгенный продукт и/или лентивирусную РНК, содержащую трансген, где указанный лентивирусный вектор является дефицитным по меньшей мере по одному гену структурного белка, выбранному из генов env, gag и pol, и содержит трансген под контролем промотора гена теломеразной обратной транскриптазы (TERT) в лентивирусной геномной последовательности, где указанная лентивирусная РНК, указанный лентивирусный вектор и указанная лентивирусная геномная последовательность представляют собой РНК ВИЧ, вектор ВИЧ и геномную последовательность ВИЧ, соответственно, и сбора высвобожденных микровезикул.
2. Способ по п.1, где у указанной клетки отсутствует указанный по меньшей мере один ген структурного белка, выбранный из генов env, gag и pol.
3. Способ по п.1 или 2, где указанный лентивирусный вектор является дефицитным по гену env.
4. Способ по любому из пп.1-3, где указанный промотор гена теломеразной обратной транскриптазы (TERT) является промотором гена TERT человека.
5. Способ по п.4, где указанный промотор гена TERT человека содержит нуклеотидную последовательность SEQ ID NO: 1 или нуклеотидную последовательность, имеющую 90% или более идентичности последовательности с нуклеотидной последовательностью SEQ ID NO: 1.
6. Способ по любому из пп.1-5, где указанный лентивирусный вектор представляет собой:
(i) РНК вектор, содержащий лентивирусную геномную последовательность,
(ii) ДНК вектор, кодирующий РНК, содержащую лентивирусную геномную последовательность, или
(iii) вирусную частицу, несущую РНК, содержащую лентивирусную геномную последовательность.
7. Способ по любому из пп.1-6, где указанный лентивирусный вектор содержит указанный трансген, представляющий собой ген-супрессор опухолевого роста.
8. Способ по п.7, где указанный ген-супрессор опухолевого роста представляет собой ген PTEN или p16.
9. Способ по любому из пп.1-6, где указанный лентивирусный вектор содержит указанный трансген, который кодирует кшРНК.
10. Способ по п.9, где указанная кшРНК специфична по отношению к гену, кодирующему регулятор клеточной пролиферации.
11. Способ по п.10, где указанный регулятор клеточной пролиферации представляет собой CDC6.
12. Способ по любому из пп.1-11, где указанная клетка является клеткой, выделенной из почки.
13. Микровезикула, содержащая трансгенный продукт и лентивирусную РНК, содержащую трансген под контролем промотора гена TERT в лентивирусной геномной последовательности, где указанная лентивирусная РНК является дефицитной по меньшей мере по одному гену структурного белка, выбранному из генов env, gag и pol, и указанная лентивирусная РНК и указанная лентивирусная геномная последовательность представляют собой РНК ВИЧ и геномную последовательность ВИЧ, соответственно; и где указанная микровезикула получена способом по любому из пп.1-12 и указанная микровезикула предназначена для применения в доставке трансгенного продукта и лентивирусной РНК, содержащей трансген, в клетку человека.
14. Способ генной трансдукции, включающий приведение клетки-мишени человека в контакт с микровезикулой, содержащей трансгенный продукт и/или лентивирусную РНК, содержащую трансген, по п.13 для их слияния, чем обеспечивается введение трансгена в клетку человека.
15. Способ по п.14, где указанная клетка человека является клеткой рака человека.
16. Лекарственное средство для лечения рака человека, содержащее микровезикулу, содержащую трансгенный продукт и лентивирусную РНК, содержащую трансген под контролем промотора гена TERT в лентивирусной геномной последовательности, где указанная лентивирусная РНК является дефицитной по меньшей мере по одному гену структурного белка, выбранному из генов env, gag и pol, и указанная лентивирусная РНК и указанная лентивирусная геномная последовательность представляют собой РНК ВИЧ и геномную последовательность ВИЧ, соответственно; где указанная микровезикула получена способом по любому из пп.7, 8, 10 и 11.
17. Способ лечения рака у пациента, включающий введение микровезикулы, содержащей трансгенный продукт и лентивирусную РНК, содержащую трансген под контролем промотора гена TERT в лентивирусной геномной последовательности, указанному пациенту, нуждающемуся в введении указанного трансгена или указанного трансгенного продукта, где указанная лентивирусная РНК является дефицитной по меньшей мере по одному гену структурного белка, выбранному из генов env, gag и pol, и указанная лентивирусная РНК и указанная лентивирусная геномная последовательность представляют собой РНК ВИЧ и геномную последовательность ВИЧ, соответственно; где указанная микровезикула получена способом по любому из пп.7, 8, 10 и 11.
| WO 2011106376 A2, 01.09.2011 | |||
| PAINTER R.G | |||
| ET AL | |||
| Conditional expression of a suicide gene by the telomere reverse transcriptase promoter for potential post-therapeutic deletion of tumorigenesis | |||
| Cancer Sci | |||
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |
| ШПАЛОРЕЗНЫЙ СТАНОК | 1922 |
|
SU607A1 |
| WO 2012135844 A2, 04.10.2012 | |||
| RU 2009118432 A, 27.11.2010. | |||

