Область техники
Настоящее изобретение касается парамиксовирусов птиц (avian paramyxoviruses, APMV) и последовательностей APMV. Настоящее изобретение касается вирусных векторов для встраивания и экспрессии чужеродных генов для применения в качестве безопасных носителей при иммунизации для защиты от множества патогенов. Оно касается векторной вакцины в системе «обратной генетики» для производства живых ослабленных вакцин. Оно также касается полинуклеотидов, которые могут применяться для продукции субъединиц в экспрессионном векторе in vitro или последовательностей для интегрирования в вирусный или плазмидный тип экспрессионного вектора в системе in vivo.
Настоящее изобретение касается немодифицированного и модифицированного вируса APMV, способов его получения и применения и определенных последовательностей ДНК и белков. Более конкретно, настоящее изобретение касается вируса APMV, в котором исходный природный геном вируса был изменен («мутанты APMV» или «рекомбинантный APMV») и способов получения и применения таких мутантов APMV или рекомбинантного APMV.
Уровень техники
Вакцины на основе вирусных векторов представляют собой одну из наиболее быстро развивающихся областей в разработке вакцин. Многие вакцины, находящиеся на стадии клинических разработок, для основных глобальных инфекционных заболеваний, ВИЧ, туберкулеза и малярии, созданы на основе вирусных векторов. Вакцины на основе вирусных векторов для животных уже представлены на рынке, например, вакцины на основе авипоксвирусных векторов для домашних животных и домашней птицы, векторные вакцины на основе герпесвирусов птиц для домашней птицы и вакцины на основе вируса коровьей оспы для диких животных. Другие векторные вакцины для домашних животных находятся в стадии разработки. Преимущество вакцин на основе вирусных векторов заключается в том, что их можно безопасно применять, поскольку в них используется векторная основа, которая является сильно ослабленной и не может сама по себе вызвать заболевание у животного. Недостатком применяемых в настоящее время вирусных векторов является существование полученного от матери иммунитета или антител, появившихся в результате перенесенной инфекции. Эти антитела будут нейтрализовать векторный вирус и, следовательно, уменьшать успешность применения векторной вакцины. Одним из главных стимулов для разработки векторных вакцин было появление высокопатогенного вируса гриппа H5N1, появившегося сначала в Азии, а позднее в Европе и в Африке. Было разработано несколько векторных вакцин-кандидатов, включая векторные вакцины на основе поксвируса птиц (Taylor et al, 1988), вируса коровьей оспы (Chambers et al., 1988), вируса саркомы Рауса (Hunt et al, 1988), аденовирусов (Tang et al., 2002, Gao et al, 2006), вируса венесуэльского энцефалита лошадей (Schultz-Cherry et al, 2000), вируса болезни Ньюкасла (US6,719,979, Veits et al., 2006, Swayne et al, 2002, Park et al, 2006), герпесвируса инфекционного ларинготрахеита (Veits et al. 2003), герпесвируса индеек (Darteil et al., 1995) и аденовируса (Hoelscher et al, 2008, Того et al, 2007). Эффективность этих векторных вакцин была протестирована на незараженных птицах, но до настоящего времени не было опубликовано сообщений об эффективности этих векторных вакцин у птиц с уже имеющимся иммунитетом к вирусному вектору и/или к белку, кодируемому вставкой.
Семейство вирусов Paramyxoviridae включает как патогены человека (вирусы кори, свинки, парагриппа, респираторно-синцитиальный вирус), так и животных (вирус болезни Ньюкасла и вирус чумы рогатого скота), которые оказывают значительное влияние на здоровье населения и на глобальную экономику (Lamb et al., 2007). Установлено, что члены этого семейства имеют однокомпонентный отрицательно направленный (с отрицательной полярностью) одноцепочечный РНК-геном. Семейство Paramyxoviridae состоит из двух подсемейств, а именно Paramyxovirinae и Pneumovirinae. Согласно недавней ревизии классификации подсемейство Paramyxovirinae включает пять родов, а именно Morbillivirus, Henipavirus, Rubulavirus, Respirovirus и Avulavirus, тогда как Pneumovirinae включает Pneumovirus и Metapneumovirus (Mayo, 2002). Парамиксовирусы птиц (APMV) относятся к роду Avulavirus и включают девять антигенно различающихся серотипов, которые были определены с помощью тестов ингибирования гемагглютинации (HI) (Alexander, 1988). Среди этих девяти серотипов изоляты, принадлежащие к подтипу APMV-1, могут вызывать опустошительное заболевание в промышленном птицеводстве и классифицируются как велогенный (с высокой вирулентностью) вирус болезни Ньюкасла (Newcastle disease virus, NDV). Более умеренные формы NDV обозначаются как мезогенные (средней вирулентности) и лентогенные (со сниженной вирулентностью) изоляты, где последняя форма у домашней птицы является обычно бессимптомной. Изоляты, относящиеся к APMV-2, 3, 6 и 7, также являются связанными с заболеванием домашней птицы. В частности, инфекции изолятами APMV-2 и 3 могут вызывать умеренное респираторное заболевание и проблемы с качеством и количеством яиц (Bankowski et al., 1981; Redmann et al., 1991; Tumova et al., 1979; Zhang et al., 2007). Известно, что изоляты APMV-6 и 7 инфицируют индеек, уток и перелетных птиц и могут вызывать респираторное заболевание, которое может осложняться вторичной инфекцией (Saif et al., 1997; Shortridge et al., 1980). С другой стороны, изоляты APMV-4, 5, 8 и 9 были выделены из уток, водоплавающей птицы и других диких птиц, но эти птицы редко демонстрируют клинические признаки, характерные для вирусной инфекции (Alexander et al., 1983; Capua et al., 2004; Gough et al., 1984; Maldonado et al., 1995; Shortridge et al., 1980).
Были получены полные геномные последовательности нескольких изолятов NDV, которые использовали для выявления различных детерминант вирулентности NDV (de Leeuw et al., 1999; Krishnamurthy et al., 1998; Zou et al., 2005). В последние два года было опубликовано несколько последовательностей APMV, которые отличаются от APMV1, такие как GenBank accession number EU338414 для APMV-2, EU403085 для APMV-3, FJ177514 для APMV-4, EU622637 для APMV-6, FJ231524 для APMV-7, FJ215863, FJ215864 и FJ619036 для APMV-8, EU910942 для APMV-9. Кроме информации о последовательностях, о факторах вирулентности известно не очень много. Изоляты APMV 2-9 главным образом были выделены из перелетных птиц. Интересно отметить, что имеется всего несколько сообщений об экспериментальном заражении цыплят такими изолятами (Saif et al., 1997). Поскольку эти APMV широко циркулируют среди диких птиц и в некоторых случаях были выделены из промышленных популяций домашней птицы (Zhang et al., 2007), что иногда вызывает заболевание в этих популяциях (Saif et al., 1997; Shihmanter et al., 1998; Shihmanter et al., 1998), знание их вирулентности в отношении домашней птицы является необходимым.
Большинство из изолятов APMV вызывает относительно слабое заболевание, которое может усиливаться в присутствии сопутствующих бактериальных или вирусных инфекций, что может приводить к экономическим потерям. В частности, APMV-2 был впервые изолирован как вторичный патоген в 1956 году в Южной Калифорнии из цыплят, пораженных острым ларинготрахеитом (Bankowski et al., 1960). Впоследствии были выделены многочисленные штаммы этого серотипа из нескольких видов птиц, что означает, что APMV-2 является широко распространенным по всему миру (Andral et al., 1984; Bradshaw et al., 1979; Fleury et al., 1979; Goodman et al., 1988; Lang et al., 1975; Lipkind et al., 1982; Lipkind et al., 1979; Zhang et al., 2006). Bankowski et. al. сообщили, что естественное или искусственное экспонирование несущихся индеек с APMV-2 вызывает резкое снижение вылупляемости птенцов и выхода птицы (Bankowski et al., 1981).
Первыми примерами изоляции APMV-4 были таковые из убитых охотниками диких уток на путях перелета на Миссисипи и Соединенных Штатах (Webster et al., 1976) и из кур, уток и гусей в Гонконге в ходе программ мониторинга гриппа у домашней птицы (Alexander et al., 1979). Не считая изолята из кольчатого чирка, пораженного геморрагическим энтеритом (Gough et al., 1984), все другие изоляты из домашней птицы были по внешним признакам непатогенными и, как было установлено, имеют широкое распространение среди водоплавающей птицы по всему миру (Stanislawek et al., 2002; Tumova et al., 1989; Yamane et al., 1982). Gough et al. сообщили, что после внутриназальной инокуляции однонедельным утятам и двухнедельным цыплятам изолята из кольчатого чирка не наблюдалось клинических проявлений и были получены очень низкие титры HI (1:8 или ниже) (Gough et al., 1984). Подобно этому, первые изоляты APMV-6 были также получены из домашней птицы в Гонконге в результате проведения программы мониторинга гриппа и сообщалось, что у кур они являются непатогенными на основании низких титров HI из экспериментально зараженных кур (Shortridge et al., 1980). Однако были сообщения о том, что инфекция APMV-6 у индеек приводит к умеренному респираторному заболеванию и проблемам с производством яиц (Alexander, 2003).
APMV-8 (Goose/Delaware/1053/76) был впервые изолирован в США из убитой охотниками Канадской казарки {Branta canadensis) (Rosenberger et al., 1974). Серологические исследования (с 1990 по 1992 годы) пернатой дичи в южной Испании показали значительное преобладание антител против APMV-8 в до 43% протестированных сывороток (Maldonado et al., 1995). Другое серологическое исследование, направленное на выяснение статуса живых здоровых крякв в Новой Зеландии в отношении инфицирования APMV, выявило присутствие антител против APMV-8 в 56% протестированных сывороток (Stanislawek et al., 2002). Warke et al (2008) сообщили, что от 16% до 31% из исследованных сывороток кур могут содержать антитела против APMV-8. Но вследствие наличия высоких титров против APMV1 существует вероятность ложноположительного теста HI, поскольку сыворотки не реагируют высокоспецифично при анализе HI. За исключением нескольких изолятов из водоплавающей птицы, изоляты APMV-8 были получены из популяций при их исследовании на вирус птичьего гриппа (Stallknecht et al., 1991), и сохраняется недостаток информации о широкой распространенности и патогенности этого вируса.
Развитие систем «обратной генетики» (условий обратной транскрипции) для РНК-генома с отрицательной полярностью NDV сделало возможным встраивание последовательностей чужеродных генов в такой геном, таким образом, обеспечивая возможность создания рекомбинантных NDV векторов для вакцинации и генной терапии (Krishnamurthy et al., 2000; Peeters et al., 1999; Roemer-Oberdoerfer et al., 1999). Сообщалось о рекомбинантных NDV векторах, экспрессирующих чужеродные вирусные белки, такие как белок НА подтипа HI вируса гриппа A (Nakaya et al., 2001), VP2 белок вируса инфекционного бурсита (IBDV) (Huang et al., 2004), гемагглютинин подтипа Н5 (Veits et al., 2006; Ge et al., 2007) и подтипа H7 (Park et al., 2006) вируса гриппа птиц. Однако эффективность большинства из таких вакцин была продемонстрирована только на свободных от специфических патогенов (SPF) птицах. NDV вызывает опустошительное заболевание у домашней птицы, приводящее к серьезным экономическим потерям в промышленном птицеводстве. Таким образом, куры в промышленном птицеводстве обычно вакцинируются против NDV в большинстве стран мира. Вследствие этого, цыплята от иммунизированных родителей в промышленных популяциях кур имеют высокий уровень полученных от матери антител. Обычные живые NDV вакцины обеспечивают защиту даже в присутствии этих антител. Однако рекомбинантные NDV вакцины (с вставками чужеродных генов) обычно являются более ослабленными по сравнению с живыми NDV вакцинами, и их эффективность может быть ослаблена в присутствии материнских антител против NDV. Таким образом, существует необходимость в платформе векторной вакцины, которая может обеспечить основу для безопасных вакцин для экспрессии гетерологических антигенов. В идеале рекомбинантная вакцина может индуцировать сильный гуморальный иммунный ответ, может применяться для массового введения и является недорогой.
Раскрытие изобретения
Настоящее изобретение касается вакцины или композиции, включающей (i) рекомбинантный APMV и (ii) фармацевтически или ветеринарно приемлемый носитель. Настоящее изобретение включает способы модификации генома APMV для получения рекомбинантного вируса APMV или вирусного вектора на основе APMV; модифицированный APMV, полученный такими способами; последовательности ДНК и белков; и способы инфицирования клеток и животных-хозяев таким рекомбинантным APMV для индукции амплификации экзогенной ДНК и белков, кодируемых экзогенной ДНК, включая антигенные белки, указанными клетками и животными-хозяевами.
Один аспект настоящего изобретения касается вируса APMV, последовательностей ДНК и белков, применяемых при получении модифицированного или рекомбинантного вируса. Одно воплощение настоящего изобретения касается геномной и белковой последовательности APMV-2, 4, 6 или 8.
Другой аспект настоящего изобретения касается модифицированного рекомбинантного вируса APMV, где указанные вирусы имеют повышенную безопасность, сильный гуморальный иммунный ответ, и способа получения таких рекомбинантных вирусов.
Другой аспект настоящего изобретения касается рекомбинантной APMV вирусной вакцины или композиции, имеющей повышенный уровень безопасности по сравнению с известными APMV или другими рекомбинантными вакцинами.
В другом аспекте настоящее изобретение предоставляет немодифицированный и модифицированный APMV вирусный вектор для экспрессии продукта гена в хозяине.
Другой аспект настоящего изобретения направлен на рекомбинантный APMV вирус, модифицированный путем вставки в него ДНК из любого источника в межгенный участок или в несущественный участок генома APMV. Синтетически модифицированные рекомбинанты APMV вируса, несущие гетерологические гены, кодирующие и экспрессирующие антиген, применяются согласно настоящему изобретению для создания новых композиций или вакцин.
Другой аспект настоящего изобретения касается APMV вирусного вектора, который предоставляет обратную генетическую систему (систему обратной транскрипции), где указанный вектор может быть применен в качестве каркаса для рекомбинантных вакцин или композиций в различных животных-хозяевах.
В одном аспекте настоящее изобретение касается фармацевтической композиции или вакцины для индукции иммунологического ответа в животном-хозяине, инокулированном данной композицией или вакциной, композиции или вакцины, включающей фармацевтически приемлемый носитель и модифицированный APMV рекомбинантный вирус или вирусный вектор. В еще одном аспекте настоящего изобретения рекомбинантный APMV вирус или вирусный вектор включает, в пределах несущественного участка вирусного генома, гетерологическую ДНК, которая кодирует антигенный белок, происходящий из патогена, где композиция или вакцина при введении хозяину является способной индуцировать иммунологический ответ, специфичный в отношении белка, кодируемого данным патогенном.
Другой аспект настоящего изобретения касается способа индукции иммунологического ответа у животного на антиген, где способ включает инокуляцию животного вакциной или фармацевтической композицией, содержащей модифицированный рекомбинантный APMV вирус или вирусный вектор, который включает и экспрессирует антигенную детерминанту патогена для указанного животного. Еще один аспект настоящего изобретения касается способа индукции иммунологического ответа у животного на антиген в режиме «прайм-буст».
Другой аспект настоящего изобретения касается способа экспрессии продукта гена в клеточной культуре in vitro путем введения в клетку модифицированного рекомбинантного APMV вируса, где ген может представлять собой ген антигенного белка, происходящий из патогена.
Краткое описание фигур
Следующее далее «Осуществление изобретения», выполненное в виде примеров, которое не направлено на ограничение настоящего изобретения описанными конкретными воплощениями, может быть понятным с привлечением сопровождающих фигур, среди которых:
Фигура 1 представляет собой таблицу, показывающую изоляцию вируса из нескольких органов кур после экспериментального инфицирования APMV-2, 4, 6 зародышей куриных яиц.
Фигура 2 представляет собой таблицу, показывающую результаты гистологического исследования некоторых органов кур после экспериментального инфицирования APMV-2, 4, 6.
Фигура 3 показывает титры HI антител у SPF кур (свободных от специфических патогенов), экспериментально инокулированных APMV-2, 4 или 6. Образцы сыворотки крови кур, собранные в дни 2, 4, 7, 14 и 28 после инфицирования, подвергали тесту HI для анализа присутствия HI антител.
Фигура 4 показывает титры HI антител у SPF кур и уток, экспериментально инфицированных APMV-8. Куры и утки были инфицированы ороназально в дозе 106 EID50 APMV-8. Образцы сыворотки крови были взяты в дни 2, 4, 7, 14 и 28 после инфицирования и проанализированы в тесте HI с антигеном APMV-8. Титры HI сывороток крови (в логарифмической шкале log2) показаны на левой оси.
Фигура 5 показывает развитие титров антител HI у SPF кур в ходе схемы вакцинирования «прайм-буст» с APMV-8. Однодневные SPF цыплята были инфицированы в день 1 (первично) и в день 14 (буст) в дозе 106 EID50 APMV-8. Образцы сыворотки крови были взяты в дни 7 и 14 после первого инфицирования и в дни 7 и 14 после вторичного инфицирования. Сыворотку исследовали в тесте HI. Титры HI сывороток (в логарифмической шкале log2) показаны на левой оси.
Фигура 6 показывает исследование продуктов ОТ-ПЦР (обратная транскрипция с последующей ПЦР) с помощью электрофореза в агарозном геле. Ткани трахеи были взяты в день 2 после инфицирования у неинфицированных уток (С1-С5) и инфицированных APMV-8 уток (11-15). Ткани гомогенизировали и готовили РНК для ОТ-ПЦР. Параллельно ставили контроль с водой (W). Продукты реакции разделяли на 1,5% агарозном геле. Размер фрагмента контролировали с использованием лестницы маркеров в 100 п.о. (New England Biolabs). Размеры фрагментов ДНК показаны справа.
Фигура 7 представляет собой таблицу, показывающую выделение вируса из кур и уток, экспериментально инфицированных APMV-8. Выделение вируса из тканей кур было проведено на зародышах куриных яиц, и определение вирусной РНК в тканях уток проводили с помощью ОТ-ПЦР.
Фигура 8 представляет собой таблицу, показывающую результаты гистологического исследования некоторых органов после инфицирования кур и Пекинских уток APMV-8.
Фигура 9 показывает развитие титров HI антител у SPF кур после инфицирования разными дозами APMV-8. Однодневные SPF цыплята были инфицированы в день 1 разными инфицирующими дозами (ID) APMV-8 или ложно инфицированы средой для переноса вируса (virus transport medium, VTM). Кровь брали в дни 7 и 14 после инфицирования, и полученные образцы сывороток крови анализировали в тесте HI. Титры HI сывороток (в логарифмической шкале log2) показаны на левой оси. Средний геометрический титр (geometric mean titer, GMT) образцов сывороток показан в самом нижнем ряду.
Фигура 10 показывает развитие титров антител HI у Пекинских уток после инфицирования разными дозами APMV-8. Однодневные SPF Пекинские утки были инфицированы в день 1 разными инфекционными дозами (ID) APMV-8 или были ложно инфицированы средой для переноса вируса (VTM). Кровь брали в дни 7 и 14 после инфицирования, и полученные образцы сывороток анализировали в тесте HI. Титры сывороток HI (в логарифмической шкале log2) показаны на левой оси. Средний геометрический титр (GMT) образцов сывороток показан в самом нижнем ряду.
Фигура 11 представляет собой таблицу, показывающую SEQ ID Nos соответствующих последовательностей ДНК и белков.
Фигура 12 показывает полную геномную последовательность штамма APMV-8 (APMV-8: SCWDS ID: МА-7, изолирован из кряквы) и генетическую карту полноразмерного генома APMV-8.
Фигура 12А показывает последовательность ДНК (SEQ ID NO:2), кодирующую Нуклеопротеин (NP) APMV-8 и последовательность белка NP (SEQ ID NO:3).
Фигура 13 показывает последовательность ДНК (SEQ ID NO:4), кодирующую Фосфопротеин (Р) APMV-8 и последовательность белка Р (SEQ ID NO:5).
Фигура 14 показывает последовательность ДНК (SEQ ID NO:6), кодирующую Матрикспротеин (М) APMV-8 и последовательность белка М (SEQ ID NO:7).
Фигура 15 показывает последовательность ДНК (SEQ ID NO:8), кодирующую Фьюженпротеин (F) APMV-8 и последовательность белка F (SEQ ID NO:9).
Фигура 16 показывает последовательность ДНК (SEQ ID NO:10), кодирующую Гемагглютинин/нейраминидазу (HN) APMV-8 и последовательность белка HN (SEQ ID NO:11).
Фигура 17 показывает последовательность ДНК (SEQ ID NO:12), кодирующую Полимеразу (L) APMV-8 и последовательность белка L (SEQ ID NO:13). Этот белок L(1) APMV-8 транслируется, начиная с кодона ATG, расположенного в положениях 8273-8275 bSEQ ID NO:1.
Фигура 18 показывает последовательность белка (2) APMV-8 Полимеразы (L) (SEQ ID NO:14). Этот белок L(2) APMV-8 транслируется, начиная с кодона ATG, расположенного в положениях 8297-8299 в SEQ ID NO:1. SEQ ID NO:14 не содержит первых 8 аминокислот SEQ ID NO:13.
Фигура 19А представляет собой схему системы обратной генетики для вируса APMV-8. Фигура 19В показывает результат репликации вируса APMV-8 в клетках MDCK.
Фигура 20 показывает результат теста HI у коммерчески приобретенных кур-бройлеров через 2 недели после вакцинации APMV-8.
Фигура 21 показывает результат теста HI у коммерчески приобретенных кур-бройлеров через 4 недели после вакцинации APMV-8.
Фигура 22 показывает результаты теста HI после вакцинации in ovo в день 18 (исследование 1).
Фигура 23 показывает результаты теста HI после вакцинации in ovo в день 19 (исследование 2).
Фигура 24 показывает результаты теста HI после вакцинации in ovo в день 18 (исследование 3).
Фигура 25 показывает последовательности 5'-полноразмерного генома (5'-FLG) и 3'-полноразмерного генома (З'-FLG), включая фланкирующие последовательности.
Фигура 26 показывает карты плазмид pcNDA-NP, pcNDA-P, pcDNA-L и pcDNA3-Т7.
Фигура 27 показывает карты плазмид pUC 18-MG-APMV-8 и pCITE4A-EGFP.
Фигура 28 показывает карту плазмиды pUC57-FL-APMV-8.
Фигура 29 показывает последовательность минигенома APMV-8.
Фигура 30 показывает выравнивание последовательностей белка NP и идентичность последовательностей на уровнях ДНК и белка.
Фигура 31 показывает выравнивание последовательностей белка Р и идентичность последовательностей на уровнях ДНК и белка.
Фигура 32 показывает выравнивание последовательностей белка М и идентичность последовательностей на уровнях ДНК и белка.
Фигура 33 показывает выравнивание последовательностей белка F и идентичность последовательностей на уровнях ДНК и белка.
Фигура 34 показывает выравнивание последовательностей белка HN и идентичность последовательностей на уровнях ДНК и белка.
Фигура 35 показывает выравнивание последовательностей белка L и идентичность последовательностей на уровнях ДНК и белка.
Осуществление изобретения
Следует указать, что в настоящем «Осуществлении изобретения», и в особенности в «Формуле изобретения», такие термины как «включает», «включено», «включая» и тому подобное должны иметь значение, приписываемое им Законом о Патентах США; например, они могут означать «содержит в себе», «включает в себя», «включая в себя» и тому подобное; и что термины, такие как «состоя по существу из» и «состоит по существу из» имеют значение, приписываемое им Законом о Патентах США, например они позволяют элементам, которые не были в точности перечислены, входить в них, за исключением элементов, которые известны из предшествующего уровня техники, или которые влияют на основные или новые характеристики настоящего изобретения.
Если не указано по-другому, технические термины используются в соответствии с их обычным значением. Определения обычных терминов в молекулярной биологии можно найти в Benjamin Lewin, Genes V., опубликованной Oxford University Press, 1994 (ISBN 0-19-854287-9); Kendrew et al. (eds.), The Encyclopedia of Molecular Biology, опубликованной Blackwell Science Ltd., 1994 (ISBN 0-632-02182-9); и Robert A. Meyers (ed.), Molecular Biology and Biotechnology: a Comprehensive Desk Reference, опубликованной VCH Publishers, Inc., 1995 (ISBN 1-56081-569-8).
Термины в единственном числе включают и множественное число, если из контекста четко не следует другое. Подобно этому, слово «или» включает в себя «и», если контекст четко не указывает на другое. Слово «или» обозначает любого одного члена из определенного списка и также включает любую комбинацию из членов такого списка.
Следует указать, что в настоящем «Осуществлении изобретения» и в прилагаемой «Формуле изобретения» и/или в пунктах, термины «Парамиксовирусы птиц» или «APMV» используются взаимозаменяемо и обозначают и включают в себя APMV-1, APMV-2, APMV-3, APMV-4, APMV-5, APMV-6, APMV-7, APMV-8 и APMV-9.
Термин «животное» в используемом здесь значении включает в себя всех млекопитающих, птиц и рыб. Животное в используемом здесь значении может быть выбрано из группы, состоящей из лошадиных (например, лошадь), псовых (например, собаки, волки, лисы, койоты, шакалы), кошачьих (например, львы, тигры, домашние кошки, дикие кошки, другие крупные кошки, и другие кошачьи, включая гепарда и рысь), бычьих (например, крупный рогатый скот), свиней (например, домашняя свинья), овечьих (например, овцы, козы, ламы, бизоны?), птичьих (например, курица, утка, гусь, индейка, перепелка, фазан, попугай, мелкие птицы (вьюрки), сокол, ворона, африканский страус, эму и казуар), приматов (например, полуобезьяны, долгопят, мартышка, гиббон, человекообразная обезьяна), людей и рыб. Термин «животное» также включает индивидуальное животное на всех стадиях развития, включая эмбриональные и зародышевые стадии.
Термины «полипептид» и «белок» используются здесь взаимозаменяемо для обозначения полимера из соединенных последовательно аминокислотных остатков.
Термины «нуклеиновая кислота», «нуклеотид» и «полинуклеотид» используются взаимозаменяемо и относятся к РНК, ДНК, кДНК (комплементарной ДНК) или кРНК (комплементарной РНК) и их производным, таким как таковые, содержащие модифицированные каркасы. Необходимо понимать, что настоящее изобретение предоставляет полинуклеотиды, включая последовательности, комплементарные к таковым, которые здесь описаны. Описываемые в настоящем изобретении «полинуклеотиды» включают как прямо направленную цепь (от 5' к 3'-концу), так и обратно направленную комплементарную цепь (от 3' к 5'-концу). Полинуклеотиды согласно настоящему изобретению могут быть получены разными способами (например, химическим синтезом, клонированием генов, и т.д.) и могут принимать различные формы (например, линейные или ветвящиеся, одноцепочечные или двухцепочечные, или их гибриды, праймеры, зонды и т.д.).
Термин «геномная ДНК» или «геном» используется взаимозаменяемо и относится к передаваемой по наследству генетической информации организма-хозяина. Геномная ДНК включает ДНК ядра (также обозначается как хромосомная ДНК), а также ДНК пластид (например, хлоропластов) и других клеточных органелл (например, митохондрий). Термины геномная ДНК или геном, применяемые в настоящем изобретении, также относятся к РНК вируса. РНК может представлять собой положительную (положительной полярности) цепь или отрицательную (отрицательной полярности) цепь РНК. Термин «геномная ДНК», применяемый в настоящем изобретении, включает геномную ДНК, содержащую последовательности, которые комплементарны к описываемым здесь последовательностям. Термин «геномная ДНК» также относится к информационной (матричной) РНК (мРНК), комплементарной ДНК (кДНК) и комплементарной РНК (кРНК). Термин «Геномная НК (нуклеиновая кислота)» в используемом здесь значении включает РНК, мРНК, кРНК, ДНК и кДНК.
Термин «ген» широко используется для обозначения любого сегмента полинуклеотида, связанного с биологической функцией. Таким образом, гены или полинуклеотиды включают интроны и экзоны, как в геномной последовательности, или только кодирующие последовательности, как в кДНК, такие как открытая рамка считывания (open reading frame, ORF), начиная со стартового кодона (метионинового кодона) и заканчивая сигналом терминации (стоп-кодоном). Гены и полинуклеотиды также могут включать участки, которые регулируют их экспрессию, например, на уровне инициации транскрипции, трансляции и терминации транскрипции. Таким образом, в данный термин также включаются промоторы и участки связывания с рибосомой (обычно эти регуляторные элементы лежат примерно между 60 и 250 нуклеотидами, расположенными выше стартового кодона кодирующей последовательности или гена; Doree S M et al; Pandher К et al; Chung J Y et al), терминаторы транскрипции (обычно терминатор располагается в пределах примерно 50 нуклеотидов ниже стоп-кодона в кодирующей последовательности или гене; Ward CKef al). Геном или полинуклеотидом также называется фрагмент нуклеиновой кислоты, который экспрессирует мРНК или функциональную РНК, или кодирует определенный белок, и который включает регуляторные последовательности.
Термин «гетерологическая ДНК» в используемом здесь значении относится к ДНК, которая происходит из другого организма, такого как другой тип клеток или другой вид по отношению к реципиенту. Данный термин также относится к ДНК или ее фрагменту из того же самого генома ДНК хозяина, когда гетерологическая ДНК вставлена в участок генома, который отличается от места ее исходной локализации.
В используемом здесь значении термин «антиген» или «иммуноген» обозначает субстанцию, которая индуцирует специфический иммунный ответ у животного-хозяина. Антиген может включать целый организм, убитый, ослабленный или живой; субъединицу или часть организма; рекомбинантный вектор, содержащий вставку с иммуногенными свойствами; часть или фрагмент ДНК, способный индуцировать иммунный ответ при презентации в животном-хозяине; полипептид, эпитоп, гаптен или любую их комбинацию. Альтернативно этому, иммуноген или антиген могут включать в себя токсин или антитоксин.
Термин «иммуногенный белок или пептид» в используемом здесь значении включает в себя полипептиды, которые являются иммунологически активными в том смысле, что при введении такого белка хозяину он способен вызывать иммунный ответ гуморального и/или клеточного типа, направленный против этого белка. Предпочтительно белковый фрагмент является таким, что он обладает практически такой же иммунологической активностью, как и целый белок. Таким образом, белковый фрагмент согласно настоящему изобретению включает в себя или состоит, по меньшей мере, из одного эпитопа или антигенной детерминанты. «Иммуногенный белок или пептид» в используемом здесь значении включает в себя полноразмерную последовательность белка, ее аналог или ее иммуногенные фрагменты. Под «иммуногенным фрагментом» понимается фрагмент белка, который включает в себя один или более эпитопов и, таким образом, вызывает иммунологический ответ, описанный выше. Такие фрагменты могут быть идентифицированы с использованием любого набора методов эпитопного картирования, хорошо известных в данной области техники. Смотри, например, Epitope Mapping Protocols в Methods in Molecular Biology, Vol.66 (Glenn E. Morris, Ed., 1996). Например, линейные эпитопы могут быть определены, например, путем конкурентного синтеза большого числа пептидов на твердых подложках, пептидов, соответствующих участкам белковой молекулы, и анализа реагирования этих пептидов с антителами в условиях, когда пептиды еще остаются связанными с подложками. Такие методы известны в данной области техники и описаны, например, в Патенте США No. 4708871; Geysen et al., 1984; Geysen et al., 1986. Подобно этому, конформационные эпитопы легко идентифицируются путем определения пространственной конформации аминокислот с помощью, например, рентгеновской кристаллографии и двухмерного ядерного магнитного резонанса. Смотри, например, Epitope Mapping Protocols, supra.
Термин «иммуногенный белок или пептид» дополнительно включает в себя делеции, вставки и замены в последовательности, до тех пор, пока полипептид функционирует в плане индукции иммунологического ответа, как здесь определено. Термин «консервативная вариация (замена)» означает замену аминокислотного остатка на другой биологически сходный остаток, или замену нуклеотида в последовательности нуклеиновой кислоты, такую, что кодируемый аминокислотный остаток не изменится или будет заменен на другой биологически сходный остаток. В этом отношении особенно предпочтительные замены будут, как правило, консервативными в природе, то есть, это такие замены, которые имеют место в пределах одного семейства аминокислот. Например, аминокислоты обычно подразделяются на четыре семейства: (1) кислые - аспартат и глутамат; (2) щелочные - лизин, аргинин, гистидин; (3) неполярные - аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан; и (4) незаряженные полярные - глицин, аспарагин, глутамин, цист(е)ин, серии, треонин, тирозин. Фенилаланин, триптофан и тирозин иногда классифицируются как ароматические аминокислоты. Примеры консервативных вариаций включают замену одного гидрофобного остатка, такого как изолейцин, валин, лейцин или метионин на другой гидрофобный остаток, или замену одного полярного остатка на другой полярный остаток, например, замену аргинина на лизин, глутаминовой кислоты на аспарагиновую кислоту, или глутамина на аспарагин, и тому подобное; или подобную консервативную замену одной аминокислоты на структурно сходную аминокислоту, что не будет оказывать значительного влияния на биологическую активность. Белки, имеющие в значительной степени одинаковую аминокислотную последовательность в качестве референтных (эталонных) молекул, но имеющие минорные аминокислотные замены, которые не влияют значительно на иммуногенность белка, попадают, таким образом, под определение референтного полипептида. Все полипептиды, полученные путем таких модификаций, являются включенными в это определение. Термин «консервативная вариация» также включает использование замененной аминокислоты на месте незамененной исходной аминокислоты, при условии, что антитела, полученные против полипептида с заменой, также являются иммунореактивными в отношении незамещенного полипептида.
Термин «клетка-хозяин» обозначает прокариотическую или эукариотическую клетку, которая была генетически изменена или способна быть генетически измененной путем введения экзогенного полинуклеотида, такого как рекомбинантная плазмида или вектор. При обозначении генетически измененных клеток данный термин относится как к исходно измененной клетке, так и к ее потомству. Полинуклеотиды, включающие желаемую последовательность, могут быть вставлены в подходящий вектор для клонирования или экспрессионный вектор, и данный вектор в свою очередь может быть введен в подходящую клетку-хозяина для репликации и амплификации. Полинуклеотиды могут быть введены в клетку-хозяина любым способом, известным в данной области техники. Векторы, содержащие представляющие интерес полинуклеотиды, могут быть введены в клетку-хозяина с помощью любого набора подходящих методов, включая прямой перенос (поглощение), эндоцитоз, трансфекцию, спаривание (f-mating), электропорацию, трансфекцию с помощью хлорида кальция, хлорида рубидия, фосфата кальция, ДЭАЭ-декстрана или других соединений; бомбардировку микропулями; липофекцию; и инфицирование (когда вектор является инфекционным, например, как в случае ретровирусного вектора). Выбор вводимых в клетку векторов или полинуклеотидов часто будет зависеть от свойств клетки-хозяина.
«Иммунологический ответ» на композицию или вакцину представляет собой развитие в хозяине клеточного и/или опосредованного антителами иммунного ответа на представляющую интерес композицию или вакцину. Как правило, «иммунологический ответ» включает, но этим не ограничивается, один или более из следующих эффектов: продукцию антител, В-клеток, клеток Т-хелперов и/или цитотоксичных Т-клеток, нацеленных специфически на антиген или антигены, включенные в представляющую интерес композицию или вакцину. Предпочтительно, хозяин будет демонстрировать либо терапевтический, либо защитный иммунологический ответ, так что устойчивость к новой инфекции будет повышена и/или клиническая тяжесть заболевания будет снижена. Такая защита будет проявляться либо в ослаблении, либо в отсутствии симптомов, обычно наблюдаемых у инфицированного хозяина, более быстром времени восстановления и/или сниженном титре вируса у инфицированного хозяина.
Одно воплощение настоящего изобретения предоставляет последовательность геномной ДНК и кодирующие белковые последовательности APMV-8. Последовательность комплементарной геномной ДНК (кДНК) штамма APMV-8 настоящего изобретения имеет полинуклеотидную последовательность, как показано в SEQ ID NO:1. Последовательность геномной кДНК APMV-8 (SEQ ID NO:1) имеет 48% идентичности последовательности с геномной ДНК APMV-1 (SEQ ID NO:15), 61% идентичности последовательности с геномной ДНК APMV-2 (SEQ ID NO:16), 47,2% идентичности последовательности с геномной ДНК APMV-3 (SEQ ID NO:17), 47,6% идентичности последовательности с геномной ДНК APMV-4 (SEQ ID NO:18), 52% идентичности последовательности с геномной ДНК APMV-6 (SEQ ID NO:19), 53% идентичности последовательности с геномной ДНК APMV-7 (SEQ ID NO:20), 99,1% идентичности последовательности с геномной ДНК APMV-8 (SEQ ID NO:37), 96,5% идентичности последовательности с геномной ДНК APMV-8 (SEQ ID NO:38), 96,4% идентичности последовательности с геномной ДНК APMV-8 (SEQ ID NO:39), 48% идентичности последовательности с геномной ДНК APMV-9 (SEQ ID NO:40). В другом воплощении, настоящее изобретение предоставляет полинуклеотид с последовательностью, показанной в SEQ ID NO: 1, 2, 4, 6, 8, 10 или 12, и его вариант или фрагмент. Настоящее изобретение дополнительно включает комплементарную цепь к описанному здесь полинуклеотиду. В еще одном воплощении настоящее изобретение предоставляет полипептид с последовательностью, показанной в SEQ ID NO: 3, 5, 7, 9, 11, 13 или 14, и его вариант или фрагмент.
Более того, гомологи полинуклеотидов или полипептидов из APMV, например, из штаммов APMV-8, APMV-2, APMV-4, APMV-6, являются входящими в рамки настоящего изобретения. В используемом здесь значении термин «гомологи» включает ортологи, аналоги и паралоги. Термин «аналоги» относится к двум полинуклеотидам или полипептидам, которые имеют одинаковую или сходную функцию, но которые эволюционировали независимо в неродственных организмах. Термин «ортологи» относится к двум полинуклеотидам или полипептидам из разных организмов, которые эволюционировали из общего предкового гена в процессе видообразования. Как правило, ортологи кодируют полипептиды, имеющие одинаковые или сходные функции. Термин «паралоги» относится к двум полинуклеотидам или полипептидам, которые связаны родством за счет дупликации в пределах генома. Паралоги обычно имеют разные функции, но эти функции могут быть родственными. Аналоги, ортологи и паралоги полипептида дикого типа APMV могут отличаться от полипептида дикого типа APMV своими посттрансляционными модификациями, различиями в аминокислотной последовательности, или и тем, и другим. В частности, гомологи настоящего изобретения будут, как правило, демонстрировать, по меньшей мере, 80-85%, 85-90%, 90-95%, или 95%, 96%, 97%, 98%, 99% идентичности последовательности со всей или с частью последовательностей полинуклеотида или полипептида APMV-8, и будут выполнять сходную функцию.
В другом аспекте настоящее изобретение предоставляет геномную кДНК APMV-8 с последовательностью, показанной в SEQ ID NO:1. В еще одном воплощении полинуклеотид представляет собой обратную комплементарную цепь полинуклеотида с последовательностью, показанной в SEQ ID NO:1. В еще одном воплощении полинуклеотид или обратная комплементарная цепь полинуклеотида настоящего изобретения имеет, по меньшей мере, 70%, по меньшей мере, 75%, по меньшей мере, 80%, по меньшей мере, 85%, по меньшей мере, 90%, по меньшей мере, 95%, 96%, 97%, 98% или 99% идентичности последовательности с полинуклеотидом, имеющим последовательность, показанную в SEQ ID NO:1.
В одном воплощении, настоящее изобретение предоставляет фрагмент полинуклеотида, колирующего полипептид AMPV-8, такой как полинуклеотид, кодирующий полипептид, имеющий последовательность, показанную в SEQ ID NO: 3, 5, 7, 9, 11, 13 или 14. В еще одном аспекте настоящее изобретение предоставляет полинуклеотид, кодирующий полипептид, имеющий, по меньшей мере, 70%, по меньшей мере, 75%, по меньшей мере, 80%, по меньшей мере, 85%, по меньшей мере, 90%, по меньшей мере, 95%, 96%, 97%, 98% или 99% идентичности последовательности с полипептидом, имеющим последовательность, показанную в SEQ ID NO: 3, 5, 7, 9, 11, 13 или 14, или консервативный вариант, аллельный вариант, гомолог или иммуногенный фрагмент, включающий, по меньшей мере, восемь, или, по меньшей мере, десять последовательных аминокислот одного из этих полипептидов, или комбинацию этих полипептидов.
В другом аспекте настоящее изобретение предоставляет полинуклеотид, имеющий нуклеотидную последовательность, показанную в SEQ ID NO:1, 2, 4, 6, 8, 10 или 12, или его вариант. В еще одном воплощении полинуклеотид представляет собой обратную комплементарную цепь полинуклеотида с последовательностью, показанной в SEQ ID NO:1. В еще одном аспекте настоящее изобретение предоставляет полинуклеотид или обратную комплементарную цепь полинуклеотида, имеющего, по меньшей мере, 70%, по меньшей мере, 75%, по меньшей мере, 80%, по меньшей мере, 85%, по меньшей мере, 90%, по меньшей мере, 95%, 96%, 97%, 98% или 99% идентичности последовательности с одним из полинуклеотидов, имеющим последовательность, показанную в SEQ ID NO:1, 2, 4, 6, 8, 10 или 12, или его вариант.
В другом аспекте настоящее изобретение предоставляет полипептид, имеющий, по меньшей мере, 70%, по меньшей мере, 75%, по меньшей мере, 80%, по меньшей мере, 85%, по меньшей мере, 90%, по меньшей мере, 95%, 96%, 97%, 98% или 99% идентичности последовательности с полипептидом, имеющим последовательность, показанную в SEQ ID NO:3, 5, 7, 9, 11, 13 или 14. В еще одном аспекте настоящее изобретение предоставляет фрагменты и варианты полипептидов APMV, идентифицированных выше (SEQ ID NO: 3, 5, 7, 9, 11, 13 или 14), которые могут быть легко получены специалистом в данной области техники с использованием хорошо известных методов молекулярной биологии.
Варианты представляют собой гомологичные полипептиды, имеющие аминокислотную последовательность, по меньшей мере, с 75%, 80%, 85%, 90%, 95%>, 96%, 97%, 98% или 99% идентичностью с аминокислотной последовательностью, показанной в SEQ ID NO: 3, 5, 7, 9,11, 13 или 14.
Варианты включают в себя аллельные варианты. Термин «аллельный вариант» относится к полинуклеотиду или к полипептиду, содержащему полиморфизмы, которые приводят к изменениям в аминокислотных последовательностях белка, и которые присутствуют в природной популяции (например, вида или варианта вируса). Такие природные аллельные вариации обычно приводят к 1-5% различиям в полинуклеотиде или в полипептиде. Аллельные варианты могут быть идентифицированы путем секвенирования представляющей интерес последовательности нуклеиновой кислоты у ряда разных видов, что может быть легко проведено с использованием гибридизационных зондов для идентификации одного и того же генетического локуса у этих видов. Любой и все такие варианты нуклеиновых кислот и появляющиеся в результате этого полиморфизмы или вариации аминокислот, которые являются результатом природной аллельной вариации и которые не изменяют функциональную активность представляющего интерес гена, являются включенными в рамки настоящего изобретения.
Термин «идентичность» в отношении последовательностей может относиться, например, к числу положений с идентичными нуклеотидами или аминокислотами, разделенными некоторым количеством нуклеотидов или аминокислот в более короткой из этих двух последовательностей, где выравнивание этих двух последовательностей может быть проведено в соответствии с алгоритмом Вилбура и Липмана (Wilbur and Lipman). Идентичность последовательностей или сходство последовательностей двух аминокислотных последовательностей, или идентичность последовательностей между двумя нуклеотидными последовательностями может быть определена с помощью пакета программ Vector NTI software package (Invitrogen, 1600 Faraday Ave., Carlsbad, С А). Когда говорится, что последовательности РНК являются сходными или имеют определенную степень идентичности последовательности или гомологии с последовательностями ДНК, тимидин (Т) в последовательности ДНК рассматривается как соответствующий (равный) урацилу (У) в последовательности РНК. Таким образом, последовательности РНК находятся в рамках настоящего изобретения и могут являться производными от последовательностей ДНК с учетом того, что тимидин (Т) в последовательности ДНК рассматривается как соответствующий (равный) урацилу (U) в последовательностях РНК.
В одном аспекте настоящее изобретение касается фармацевтической композиции или вакцины для индукции иммунологического ответа в животном-хозяине, которому введена вакцина или композиция, где вакцина или композиция включает фармацевтически приемлемый носитель и модифицированный рекомбинантный вирус APMV или вирусный вектор. В еще одном аспекте настоящего изобретения рекомбинантный вирус APMV или вирусный вектор включает в себя, в пределах несущественного участка вирусного генома, гетерологическую последовательность ДНК, которая кодирует антигенный белок, происходящий из патогена, где композиция или вакцина при введении хозяину является способной индуцировать иммунологический ответ, специфичный в отношении белка, кодируемого патогеном.
Термин «вектор» относится к рекомбинантной ДНК- или РНК-плазмиде, бактериофагу или вирусу, которые включают гетерологический полинуклеотид, который необходимо доставить в клетку-мишень, либо in vitro, либо in vivo. Гетерологический полинуклеотид может включать представляющую интерес последовательность для целей профилактики или лечения, и может не обязательно находиться в форме экспрессионной кассеты. В используемом здесь значении вектор должен не быть способен к репликации в конечной клетке-мишени или в субъекте. Данный термин включает в себя векторы для клонирования и вирусные векторы.
Термин «генноинженерный» или «рекомбинантный» обозначает полинуклеотид полусинтетического или синтетического происхождения, который либо не встречается в природе, либо соединен с другим полинуклеотидом в таком порядке, который не найден в природе.
Термин «несущественный участок» относится к участку вирусного генома, который не является существенным для репликации и размножения вируса в культуре ткани, и удаление или инактивация которого могут снижать вирулентность в разнообразных животных системах. Любой несущественный участок или его часть могут быть удалены из генома APMV или в них может быть вставлена чужеродная последовательность, и жизнеспособность и стабильность рекомбинантного APMV, полученного в результате такой делеции или вставки, может использоваться для определения того, действительно ли удаленный участок или его часть являются несущественными. В одном воплощении, несущественный участок генома APMV представляет собой любой участок генома APMV-2, 4, 6 или 8, который не кодирует Полимеразу (L). В еще одном воплощении несущественный участок включает в себя открытую рамку считывания, кодирующую несущественный белок. В этом аспекте открытая рака считывания выбирается из группы, состоящей из нуклеопротеина (NP), фосфопротеина (Р), матрикспротеина (М), фьюженпротеина (F), и гемагглютинина/нейраминидазы (HN). В одном воплощении, несущественный участок локализован выше гена NP. В другом воплощении, несущественный участок локализован ниже гена L. В еще одном воплощении, несущественный участок представляет собой некодирующий или межгенный участок. В этом аспекте некодирующий или межгенный участок может быть участком между генами NP и Р, между генами Р и М, между генами М и F, или между генами F и HN в геноме APMV-2, 4, 6 или 8. В еще одном воплощении, несущественный участок может находится в участке положений нуклеотидов 1-140, 1526-1692, 2910-3085, 4195-4498, 6130-6382, 8116-8272, 8116-8289 или 15013-15342 в последовательности SEQ ID NO: 1.
В другом аспекте настоящее изобретение включает химеры APMV, в которых одна часть или целый ген или несколько частей или целых генов вектора APMV заменены подобными генами из других вирусов, в частности вирусов, относящихся к семейству Paramyxoviridae.
В одном воплощении настоящего изобретения вакцина или фармацевтическая композиция включает в себя антиген, выбираемый из группы патогенов птиц, включая, но ими не ограничиваясь, Salmonella typhimurium, Salmonella enteritidis, вирус инфекционного бронхита (Infectious Bronchitis virus, IBV), вирус болезни Ньюкасла (NDV), вирус синдрома снижения яйценоскости (несучести) (egg drop syndrome virus, EDS), или вирус инфекционного бурсита (Infectious Bursal Disease virus, IBDV), вирус инфекционного ларинготрахеита (Infectious Laryngotracheitis virus, ILTV), аденовирусы птиц, вирус болезни Марека (Marek's disease virus, MDV), вирус оспы кур, вирус энтерита уток (duck enteritis virus, DEV), парвовирусы уток, вирус птичьего гриппа, APMV, такой как APMV-1 и тому подобное, и их комбинации.
В другом воплощении, вакцина или фармацевтическая композиция включают в себя антиген, выбираемый из патогена кошачьих, такого как, но ими не ограничиваясь, герпесвирус кошачьих (FHV), калицивирус кошачьих (FCV), вирус лейкемии кошачьих (FeLV), вирус иммунодефицита кошачьих (FIV), парвовирус кошачьих (FPV), вирус инфекционного перитонита кошачьих (FIPV), вирус бешенства и тому подобное, и их комбинации.
В еще одном воплощении, вакцина или фармацевтическая композиция настоящего изобретения включает в себя антиген, выбираемый из патогена псовых, включая, но ими не ограничиваясь, вирус бешенства, герпесвирус псовых (CHV), парвовирус псовых (CPV), вирус собачьей чумки (CDV), вирус парагриппа 2 псовых (CPI2), коронавирус псовых, Leptospira canicola, Leptospira icterohaemorragiae, Leptospira grippotyphosa, Borrelia burgdorferi, Bordetella bronchiseptica и тому подобное, и их комбинации.
В еще одном воплощении, вакцина или фармацевтическая композиция включает в себя антиген, выбираемый из патогена лошадиных, такого как гепресвирус лошадей (тип 1 или тип 4), вирус гриппа лошадей, столбняка, вирус лихорадки Западного Нила, артеривирус лошадей и тому подобное, и их комбинации.
В еще одном воплощении, вакцина или фармацевтическая композиция включает в себя антиген, выбираемый из патогена бычьих, овец или коз, такого как вирус бешенства, ротавирус крупного рогатого скота, вирус парагриппа типа 3 крупного рогатого скота (bPIV-З), коронавирус крупного рогатого скота, вирус вирусной диареи крупного рогатого скота (BVDV), вирус ящура (FMDV), вирус чумы крупного рогатого скота (Rinderpest virus, RPV), вирус чумы мелких жвачных животных (Peste des Petits Ruminants virus, PPRV), вирусы злокачественной катаральной лихорадки крупного рогатого скота, респираторно-синцитиальный вирус крупного рогатого скота (BRSV), вирус инфекционного ринотрахеита крупного рогатого скота (IBR), Escherichia coli, Pasteurella multocida, Pasteurella haemolytica и тому подобное, и их комбинации.
В другом воплощении вакцина или фармацевтическая композиция включает в себя антиген, выбираемый из патогена свиней, такого как, но ими не ограничиваясь, вирус свиного гриппа (SIV), цирковирус свиней типа 2 (PCV-2), вирус репродуктивно-респираторного синдрома свиней (PRRS), вирус ложного бешенства (PRV), парвовирус свиней (PPV), FMDV, Mycoplasma hyopneumoniae, Erysipelothrix rhusiopathiae, Pasteurella multocida, Bordetella bronchiseptica, Escherichia coli и тому подобное, и их комбинации.
Конструирование рекомбинантного вируса хорошо известно в данной области техники, как описано, например, в Патентах США Nos. 4,769,330, 4,722,848, 4,603, 112, 5,174, 993, и 5,756,103, 6,719,979. В частности, рекомбинантный APMV вирус может быть сконструирован в две стадии. Во-первых, ген, представляющий интерес для вставки в вирус, такой как открытая рамка считывания антигена из APMV-1 (NDV) или вируса птичьего гриппа или другого организма, помещается в плазмидную конструкцию E.coli, в которую вставлена кДНК, гомологичная участку кДНК APMV. Отдельно от этого, кДНК последовательности гена, который должен быть вставлен, предваряется промоторным участком (стартовым участком гена), а за ней следует участок конца гена, который специфичен для вектора APMV. Фрагмент ДНК, состоящий из стартового участка гена/чужеродного антигена/конца гена, фланкируется фрагментом кДНК, гомологичным APMV-8 кДНК, содержащим уникальные сайты (участки) для расщепления ферментами рестрикции. Затем полученная плазмидная конструкция амплифицируется путем выращивания в бактериях E.coli и выделяется. Далее рекомбинантная плазмида используется для расщеплении ферментом рестрикции для вырезания фрагмента ДНК, включающего стартовый участок гена/чужеродный антиген/конец гена, который фланкирован кДНК, гомологичной кДНК APMV-8, и последующего лигирования этого фрагмента в подходящим образом расщепленную полноразмерную конструкцию APMV-8.
Полноразмерная конструкция, содержащая представляющий интерес ген, трансфецируется в клетки вместе с плазмидами, содержащими полинуклеотиды для экспрессии нуклеопротеина (NP) APMV, фосфопротеина (Р) APMV и РНК-полимеразы (L) APMV, а также Т7 РНК-полимеразы. Все кДНК-конструкции APMV находятся под контролем промотора Т7-полимеразы. Сохранение инфекционности вируса осуществляется, как описано в Romer-Oberdorfer et al., 1999, и как показано на фигуре 19А. Экспрессия Т7 РНК-полимеразы в трансфецированных клетках может быть достигнута разными способами, включая трансфекцию плазмидной ДНК, содержащей экспрессионную кассету для Т7 РНК-полимеразы, рекомбинантный вирус (такой как вирус куриной оспы или оспы канареек), экспрессирующий Т7 РНК-полимеразу, или в клетках, которые экспрессируют Т7 РНК-полимеразу. В другом аспекте полинуклеотиды для экспрессии нуклеопротеина (NP) APMV, фосфопротеина (Р) APMV и РНК-полимеразы (L) APMV и полноразмерная вирусная кРНК находятся под контролем истинно раннего промотора цитомегаловируса человека. Сохранение вируса проводится, как описано в Inoue К, et al., 2003.
Успешная экспрессия вставленной представляющей интерес кДНК (чужеродной кДНК или гетерологической кДНК) модифицированным инфекционным вирусом требует двух условий. Во-первых, вставка должна быть введена в участок генома вируса таким образом, чтобы модифицированный вирус оставался жизнеспособным. Вторым условием для экспрессии вставленной кДНК является присутствие регуляторных последовательностей, обеспечивающих экспрессию данного гена в составе вирусного генома (например, начало гена, конец гена, промотор, энхансер, сигналы полиаденилирования, межгенные и нетранслируемые участки).
В целом, желательно применять сильный промотор, функциональный в эукариотических клетках. В одном воплощении, промотор, используемый для транскрипции вирусной мРНК вирусной РНК-полимеразой, представляет собой «последовательность старта гена». «Последовательность старта гена» представляет собой сайт связывания для L белка, необходимый для связывания и транскрибирования расположенной ниже вирусной РНК в вирусную мРНК.
В одном воплощении, настоящее изобретение предоставляет введение терапевтически эффективного количества вакцины APMV для доставки и экспрессии антигена, эпитопа или иммуногена в клетку-мишень. Определение терапевтически эффективного количества представляет собой рутинную задачу для специалиста средней квалификации в данной области техники. В одном воплощении, композиция вакцины APMV включает экспрессионный вектор, содержащий полинуклеотид, который кодирует антиген, эпитоп или иммуноген, и фармацевтически или ветеринарно приемлемый носитель, связующий материал или наполнитель. В другом воплощении, фармацевтически или ветеринарно приемлемый носитель, связующий материал или наполнитель облегчают трансфекцию и/или улучшают сохранение вектора или белка.
Фармацевтически или ветеринарно приемлемые носители, или связующие материалы, или наполнители хорошо известны специалисту в данной области техники. Например, фармацевтически или ветеринарно приемлемый носитель, или связующий материал, или наполнитель могут представлять собой 0,9% раствор NaCl (то есть, физиологический раствор) или фосфатный буфер. Другие фармацевтически или ветеринарно приемлемые носители, или связующие материалы, или наполнители, которые могут применяться в способах настоящего изобретения, включают, но ими не ограничиваются, поли-(L-глутамат) или поливинилпирролидон. Фармацевтически или ветеринарно приемлемый носитель, или связующий материал, или наполнитель могут представлять собой любое соединение или комбинацию соединений, облегчающих введение вектора (или белка, экспрессируемого вектором настоящего изобретения in vitro), или облегчающих трансфекцию и/или улучшающих сохранность вектора (или белка). Дозы и объемы дозировок здесь обсуждаются в общем описании и также могут быть определены специалистом в данной области техники, исходя из прочтения данного «Раскрытия изобретения» с учетом знаний в данной области техники, без какого-либо специального экспериментирования.
В другом воплощении, фармацевтически или ветеринарно приемлемый носитель, наполнитель или связующий материал могут представлять собой эмульсию воды в масле. Примеры пригодных эмульсий воды в масле включают водно-масляные эмульсии для вакцинации на масляной основе, которые стабильны и остаются жидкими при 4°С, содержащие: от 6 до 50% (по объему) антиген-содержащей водной фазы, от 12 до 25% (по объему), от 50 до 94% (по объему) масляной фазы, состоящей полностью или частично из неметаболизируемого масла (например, минерального масла, такого как жидкий парафин), и/или метаболизируемого масла (например, растительного масла, или эфиров жирных кислот, полиолов или спиртов), от 0,2 до 20 (p/v) % сурфактантов, от 3 до 8 (p/v) %, последние представлены полностью или частично, или в смеси либо с эфирами полиглицерола, где указанные эфиры полиглицерола являются полиглицерол(поли)рицинолеатами, либо полиоксиэтиленрициновыми маслами, или же гидрированными полиоксиэтиленрициновыми маслами. Примеры сурфактантов, которые могут применяться в эмульсии вода в масле, включают этоксилированные эфиры сорбитана (например, полиоксиэтилен(20)сорбитан моноолеат (TWEEN 80®), доступный от AppliChem, Inc., Cheshire, СТ) и эфиры сорбитана (например, сорбитан моноолеат (SPAN 80®), доступный от Sigma Aldrich, St. Louis, МО). Кроме того, в отношении эмульсии вода в масле, смотри также Патент США No. 6919084. В некоторых воплощениях содержащая антиген водная фаза включает солевой раствор, содержащий один или более буферных агентов. Примером подходящего буферного раствора является фосфатный буферный раствор. В одном воплощении, эмульсия воды в масле может представлять собой тройную эмульсию вода/масло/вода (W/O/W) (смотри, например, Патент США No. 6358500). Примеры других пригодных эмульсий описаны в Патенте США No. 7371395.
Фармацевтические композиции и вакцины согласно настоящему изобретению могут включать в себя или состоять в основном из одного или более адъювантов. Подходящими адъювантами для применения при осуществлении настоящего изобретения являются (1) полимеры акриловой или метакриловой кислоты, малеиновый ангидрид и полимеры производных алкенилов, (2) иммуностимулирующие последовательности (ISS), такие как олигодезоксирибонуклеотидные последовательности, имеющие одну или более неметилированных CpG единиц (Klinman et al., 1996; W098/16247), (3) эмульсии масла в воде, такие как эмульсия SPT, описанная на стр.147 в "Vaccine Design, The Subunit and Adjuvant Approach" опубликованной M. Powell, M. Newman, Plenum Press 1995, и эмульсия MF59, описанная на стр.183 в той же работе, (4) катионные липиды, содержащие четвертичную соль аммония, например, DDA, (5) цитокины, (6) гидроокись алюминия или фосфат алюминия, (7) сапонин или (8) другие адъюванты, обсуждаемые в любом процитированном здесь документе, которые все включаются в настоящее изобретение путем отсылки, или (9) любые их комбинации или смеси.
Эмульсия масла в воде (3), которая является особенно пригодной для вирусных векторов, может иметь своей основой: легкое жидкое парафиновое масло (Тип Европейской фармакопеи), изопреноидное масло, такое как сквалан, сквален, масло, получаемое при олигомеризации алкенов, например, изобутена или децена, эфиры кислот или спиртов, имеющих алкильную группу с неразветвленной цепью, такие как растительные масла, этилолеат, пропиленгликоль, ди(каприлат/капрат), глицеролтри(каприлат/капрат) и пропиленгликольдиолеат, или эфиры разветвленных жирных спиртов или кислот, особенно эфиры изостеариновой кислоты. Масло используется в комбинации с эмульгирующими агентами для образования эмульсии. Эмульгирующими агентами могут быть неионные сурфактанты, такие как: эфиры, с одной стороны, сорбитана, маннида (например, ангидроманнитололеат), глицерола, полиглицерола или пропиленгликоля и, с другой стороны, олеиновой, изостеариновой, рицинолеиновой или гидроксистеариновой кислот, где указанные эфиры могут не обязательно быть этоксилированы, или представлять собой полиоксипропилен-полиоксиэтилен блок-сополимеры, такие как Плюроник, например, L121. Среди адъювантных полимеров типа (1) предпочтение отдается полимерам поперечно сшитой акриловой или метакриловой кислоты, особенно сшитым полиалкениловыми эфирами Сахаров или полиспиртов. Эти соединения известны под названием карбомер (Pharmeuropa, vol.8, no. 2, June 1996). Специалист в данной области техники может также обратиться к Патенту США No. 2909462, который предоставляет такие акриловые полимеры, поперечно сшитые полигидроксильным соединением, имеющим, по меньшей мере, три гидроксильные группы, предпочтительно не более, чем восемь таких групп, где атомы водорода, по меньшей мере, трех гидроксильных групп заменены на ненасыщенные алифатические радикалы, имеющие, по меньшей мере, два атома углерода. Предпочтительными радикалами являются таковые, которые содержат от 2 до 4 атомов углерода, например, винилы, аллилы и другие этиленированные ненасыщенные группы. Ненасыщенные радикалы могут также содержать другие заместители, такие как метил. Продукты, продаваемые под названием Carbopol (BF Goodrich, Ohio, USA), являются особенно предпочтительными. Они являются поперечно сшитыми аллилсахарозой или аллилпентаэритритолом. Среди них можно указать на Carbopol 974Р, 934Р и 971Р.
В отношении сополимеров, производных малеинового ангидрида/алкенилов, предпочтение отдается EMA (Monsanto), которые представляют собой сополимеры этилен/малеиновый ангидрид с прямой цепью или поперечно сшитые, и которые, например, сшиты дивиниловым эфиром. Можно также сослаться на работу J. Fields et al., 1960.
В отношении структуры, полимеры акриловой или метакриловой кислоты и ЕМА являются предпочтительно образованными из базовых единиц, имеющих следующую формулу:
в которой:
-R1 и R2, которые могут быть одинаковыми или разными, представляют собой Н или СН3
-х=0 или 1, предпочтительно х=1
-у=1 или 2, где х+у=2.
Для ЕМА, х=0иу=2и для карбомеров х=у=1.
Эти полимеры являются растворимыми в воде или физиологическом солевом растворе (20 г/л NaCl) и pH может быть доведен до 7,3-7,4, например, щелочью (NaOH), для получения раствора адъюванта, в который может (могут) быть включен (включены) вектор (векторы) экспрессии. Концентрация полимера в конечной иммунологической композиции или вакцине может находиться в диапазоне между 0,01 и 1,5% (вес/объем) между 0,05 и 1% (вес/объем) и между 0,1 и 0,4% (вес/объем).
Другой аспект настоящего изобретения касается способа индукции иммунологического ответа у животного к антигену, включающего введение животному вакцины или фармацевтической композиции, содержащей модифицированный рекомбинантный вирус APMV, который включает в себя и кодирует антиген патогена для указанного животного. Еще другой аспект настоящего изобретения касается способа индукции иммунологического ответа у животного на антиген в режиме введения «прайм-буст», который включает, по меньшей мере, одно первичное введение и, по меньшей мере, одно бустерное введение с применением, по меньшей мере, одного общего полипептида, антигена, эпитопа или иммуногена. Иммунологическая композиция или вакцина, применяемая при первичном введении, может быть той же самой или может отличаться по своей природе от таковой, используемой при бустерном введении. В одном аспекте протокола «прайм-буст» настоящего изобретения композиция или вакцина, включающая рекомбинантный вирус APMV (вирусный вектор) настоящего изобретения вводится перед последующим введением инактивированной вирусной вакцины или композиции, включающей антиген, или вакцины или композиции, включающей субъединицу (белок, антиген), или ДНК-плазмидной вакцины или композиции, которая содержит или экспрессирует антиген. Подобно этому, протокол «прайм-буст» может включать введение инактивированной вирусной вакцины или композиции, включающей антиген, или вакцины или композиции, включающей субъединицу (белок, антиген), или ДНК-плазмидной вакцины или композиции, которая содержит или экспрессирует антиген, перед последующим введением композиции или вакцины, включающей рекомбинантный вирус APMV (вирусный вектор) настоящего изобретения Следуют дополнительно указать, что оба введения, и первичное, и вторичное, могут включать композицию или вакцину, включающую рекомбинантный вирус APMV (вирусный вектор) настоящего изобретения.
Первичное введение может включать одно или более введений одних и тех же иммунологических композиций вакцины на основе вирусного вектора. Подобно этому, бустерное введение может включать одно или более введений той же иммунологической композиции вакцины на основе вирусного вектора. Пути введения при первичном и бустерном введении могут быть одинаковыми или разными. Подобно этому, происхождение защитного гена, присутствующего в первичном и в бустерном введениях, может быть одним и тем же или разным (например, разные штаммы).
Разные введения проводятся предпочтительно с интервалом от 1 до 6 недель одно от другого, более предпочтительно примерно через 3 недели. Согласно предпочтительной схеме также предусматриваются ежегодные бустерные введения, предпочтительно с применением иммунологической композиции вакцины на основе вирусного вектора. Животные предпочтительно имеют возраст, по меньшей мере, в один день при первом введении.
Может применяться множество путей введения, таких как подкожно или внутримышечно, внутрикожно или трансдермально, в виде спрея, питьевой воды, глазных капель, внутри назально, перорально, с пищей, in ovo, или их комбинация (например, в глаза и нос, в рот и нос).
Настоящее изобретение далее будет детально описано с помощью следующих нелимитирующих примеров.
ПРИМЕРЫ
Пример 1 APMV-2, APMV-4 и APMV-6
A. Вирусы и Птицы
Однодневные SPF (свободные от специфического антигена) цыплята (Merial, Gainesville, GA) содержались в изолированных клетках Horsfall-Baur под положительным давлением. Корм и вода предоставлялись ad libitum, и птицы осматривались дважды в день. Используемые в экспериментальных исследованиях вирусы (APMV-2, 4 и 6) были выделены из диких птиц и классифицированы в National Veterinary Service Laboratory (NVSL, Ames Iowa, USA). Вирусы размножали в 9-дневных зародышах SPF куриных яиц (SunRise Farms, Catskill, NY, USA) путем инокуляции через аллантоис. Аллантоисную жидкость собирали в день 3 после инокуляции, разделяли на аликвоты и хранили при -80°С. Подтип APMV подтверждали в тесте HI с использованием стандартных сывороток (NVSL, Ames, Iowa, USA). Инфекционная доза для 50% яиц (EID50) для каждого изолята была определена с помощью инокуляции 10-кратных серийных разведений аллантоисной жидкости в зародыши SPF яиц. Титр рассчитывали по методу, описанному Reed and Muench (Reed, LJ et al., 1938, Am. J. Epidemiol. 27:493-497).
B. Экспериментальное инфицирование
По двадцать пять однодневных SPF цыплят в каждой группе инфицировали 10б EID50 на одного цыпленка окулярно-назальным путем (через глаза и нос). Цыплят в контрольной группе ложно инокулировали PBS (фосфатный буферный раствор). В дни 2, 4, 7, 14 и 28 после инфицирования у пяти птиц из каждой группы забирали кровь из плечевой вены для сбора образцов сыворотки, птиц умерщвляли под СО2, и вскрывали. Брали образцы тканей трахеи, легких, поджелудочной железы и кишечника. Для каждого органа использовали новую пару стерильных ножниц и пинцетов. Половину образца ткани помещали в пробирку Lysing Matrix D tube (MP Biomedicals, Solon, OH), содержащую среду для переноса вирусов (IX минимальная основная среда, 7,5% бикарбонат натрия, 15 мМ HEPES, 1% фетальная сыворотка теленка, 4000 Ед/мл пенициллин, 400 мкг/мл гентамицин, 8 мкг/мл амфотерицин В, 4000 мкг/мл стрептомицин, 1000 мкг/мл канамицин сульфат). Вторая половина образца ткани фиксировалась в 10-процентном забуференном формалине и заливалась в парафин. Срезы залитых в парафин тканей прокрашивали гематоксилином Майера и эозином (Н&Е).
Результаты показали, что слабая диарея наблюдалась в день четыре и день семь после инфицирования у птиц, инфицированных APMV-2 или APMV 4. При вскрытии у птиц, инфицированных APMV-2, наблюдалось небольшое увеличение поджелудочной железы в дни два и четыре после инфицирования. Никаких других крупных повреждений ни в одной группе обнаружено не было.
C. Серология
Для выявления вируса в аллантоисной жидкости и анализа присутствия HI антител в собранных образцах сыворотки использовались тесты гемагглютинации (НА) и ингибирования гемагглютинации (HI) соответственно. Тесты проводили стандартным способом, используя 0,8% эритроциты цыплят, ресуспендированные в PBS. Тест HI проводили методом разведения сыворотки при постоянном уровне антигена. Для каждого разведения сыворотки использовали восемь НА единиц вирусного антигена.
Титры HI антител исследовали в дни 2, 4, 7, 14 и 28 после инфицирования. Положительные титры HI (≥1:16) были обнаружены в образцах сыворотки инфицированных APMV-2 птиц в день 7 (1/5), день 14 (5/5)и день 28 после инфицирования (5/5). Интересно отметить, что только один цыпленок, инфицированный APMV-4, показал титр HI, который был оценен как положительный во время всего хода эксперимента в день 14 после инфицирования. Подобно этому для группы APMV-6 два цыпленка из пяти показали титр HI 1:16 только в день 28 после инфицирования. Ложно инфицированные птицы оставались отрицательными на наличие HI антител против всех трех APMV, использованных в этом эксперименте. Кроме того, для исключения перекрестного заражения все сыворотки были протестированы на наличие HI антител против двух других антигенов и показали отрицательные результаты.
D. Выделение вируса
Образцы тканей, собранные в Lysing Matrix D tubes (MP Biomedicals, Solon, OH, USA), были дважды прогомогенизированы с использованием Fastprep®-24 (MP Biomedicals, Solon, ОН) в режиме 4,0 M/S в течение 20 секунд. После инкубации в течение 15 мин при комнатной температуре гомогенизированные образцы центрифугировали 20 мин при 2000xg при 4°С. Стерильность проверяли после инокуляции 50 мкл полученного супернатанта в 2 мл триптозо-фосфатного бульона (ТРВ) (DIFCO, Becton Dickenson, Sparks, MD, USA), обогащенного 10% гидролактальбумина, при инкубации при 37°С на вращающемся (орбитальном) шейкере в течение ночи. Нестерильные образцы фильтровали через 0,45 мкм шприцевые фильтры (Whatman Inc., Florham Park, NJ, USA). Образцы хранили при -80°C. Выделение вируса проводили путем инокуляции 0,1 мл в аллантоисную полость 9-дневных зародышей SPF куриных яиц. После инкубации в течение трех дней при 37,5°С аллантоисную жидкость собирали и анализировали на наличие гемагглютинирующей активности в тесте НА.
Для выявления мест размножения вируса несколько органов (трахея, легкие, кишечник, поджелудочная железа) анализировали на инфекционный вирус путем изолирования вируса из зародышей яиц (фигура 1). В целом, размножающийся вирус был обнаружен только в нескольких цыплятах. Вкратце, APMV-4 был выделен в день 2 из трахеи, легких и поджелудочной железы, тогда как APMV-6 был выделен из легких и поджелудочной железы. В день 4 APMV-2 был выделен из трахеи и легких, тогда как APMV-6 был выделен из всех тестируемых органов за исключением кишечника. В день 7 APMV-2 был выделен из единственного образца кишечника, APMV-4 был выделен из поджелудочной железы, тогда как APMV-6 был выделен из образцов легких и поджелудочной железы. Неожиданно в день 14 после инфицирования вирусы не были выделены, а в день 28 после инфицирования APMV-2, 4 и 6 были выделены из поджелудочной железы. Из ложно инокулированных птиц вирусов выделено не было. Идентичность вновь выделенных вирусов была подтверждена в тесте HI с использованием стандартных сывороток, предоставляемых NVSL.
Для оценки патологического потенциала исследуемых вирусов были проанализированы микроскопические поражения в полученных органах (фигура 2). В день 2 после инфицирования у всех инфицированных цыплят наблюдались катаральный трахеит в комбинации с потерей ресничек в респираторном (мерцательном) эпителии и слабый энтерит. В день 4 после инфицирования инфицированные APMV-2 цыплята демонстрировали увеличенное количество гипертрофированных слизистых желез в трахее и фокальное (очаговое) изъязвление респираторного эпителия. Инфицированные APMV-4 птицы демонстрировали изменения, позволяющие отнести их к респираторной инфекции, такие как слабый трахеит, слабый до умеренного многоочаговый лимфоцитарный панкреатит, а также фокальная гиперплазия ассоциированной с бронхами лимфоидной ткани (BALT, Bronchus-Associated Lymphoid Tissue) в день 4 после инфицирования. Исследование органов инфицированных APMV-6 цыплят выявило изменения в трахее, такие как катаральный и язвенный трахеит, и фокальный панкреатит, что согласуется с поражением вирусом. В день 7 после инфицирования инфицированные APMV-2 цыплята демонстрировали ослабление фокального трахеита или восстановление респираторного эпителия, что является признаками заживления. Птицы, инфицированные APMV-4, демонстрировали слабую гиперплазию BALT, тогда как инфицированные APMV-6 птицы демонстрировали кистозную (пузырчатую) энтеропатию, фокальный энтерит и инфильтрат лимфоцитов в поджелудочную железу. Помимо слабого лимфоцитарного энтерита и слабой гиперплазии GALT, инфицированные APMV-2 птицы также демонстрировали признаки заживления, такие как ослабление поражения трахеи в день 14 после инфицирования. Образцы органов цыплят, инфицированных APMV-4 или 6, демонстрировали изменения, позволяющие сделать вывод о вирусной инфекции, такие как слабая интерстициальная пневмония, катаральный трахеит и гиперплазия BALT или GALT в день 14 после инфицирования. Все исследованные образцы, полученные из инфицированных цыплят, продемонстрировали поражения, такие как гиперплазия GALT, лимфоцитарный панкреатит и лимфоцитарный бронхит в день 28 после инфицирования. В день 2 птицы контрольной группы продемонстрировали слабый катаральный трахеит, который мог быть связан с факторами окружающей среды.
Фигура 3 показывает низкие титры HI (до 1:32 в день 14) у SPF цыплят, инфицированных APMV-4 или APMV-6, указывая на то, что только инфекция APMV-2 вызывает HI ответ, который может быть охарактеризован как сероположительный. Тем не менее, все три вируса выделялись из трахеи, легких, кишечника и поджелудочной железы инфицированных птиц вплоть до дня 7 после инфицирования и из поджелудочной железы вплоть до дня 28 после инфицирования. Инфекция APMV-2, 4 или 6 демонстрирует характерные гистопатологические поражения (суммированы в фигуре 2) у всех инфицированных птиц, что указывает на стимуляцию вирусным антигеном. Выделение вирусов и гистологические профили инфицированных птиц четко показывают тропизм вирусов в инфицированных птицах (трахея и легкие в дни от 2 до 7, и кишечник, легкие и поджелудочная железа после дня 7). Все изоляты были выявлены в поджелудочной железе вплоть до дня 28 после инфицирования, но выделение вируса было невозможным в день 14 после инфицирования. Это указывает на то, что исследуемые APMV, по-видимому, могут сохраняться и позднее вновь активироваться. Таким образом, носители вируса могут присутствовать в инфицированных группах/популяциях птицы. Только APMV-2 индуцировал образование HI антител, тогда как HI антитела у цыплят, инфицированных APMV-4 и 6, не определялись.
Пример 2. APMV-8
А. Вирусы и птицы
Однодневные SPF цыплята (Merial, Gainesville, GA, USA) и Пекинские утки (Metzer Farms, Gonzales, CA, USA) содержались в изолированных клетках Horsfall Baur под положительным давлением. Корм и вода предоставлялись ad libitum, и птицы осматривались дважды в день. Используемый в экспериментальных исследованиях вирус APMV-8 (APMV-8: SCWDS ID: МА-7) был изолирован из диких уток (кряква) и классифицирован в National Veterinary Service Laboratory (NVSL, Ames Iowa, USA). Вирус размножали в 9-дневных зародышах SPF куриных яиц путем инокуляции через аллантоис. Аллантоисную жидкость собирали в день 3 после инокуляции, объединяли, разделяли на аликвоты и хранили при -80°С. Подтип APMV-8 подтверждали в тесте HI с использованием стандартных сывороток, предоставляемых National Veterinary Service Laboratory (Ames, IA, USA). Доза EID50 была определена с помощью инокуляции 10-кратных серийных разведений аллантоисной жидкости в зародыши SPF яиц. Титр рассчитывали по методу, описанному Reed andMuench (Reed & Muench, 1938).
B. Экспериментальное инфицирование
По двадцать пять однодневных SPF цыплят или Пекинских уток на группу были инфицированы окулярно-назальным путем 106 EID50 на каждую птицу в растворе PBS. Птиц в контрольных группах цыплят и уток ложно инокулировали PBS. У пяти птиц из каждой группы брали кровь из плечевой вены для сбора образцов сыворотки, птиц гуманно умерщвляли с помощью СО2, и проводили вскрытие в дни 2, 4, 7, 14 и 28 после инфицирования. Брали образцы тканей трахеи, легких, поджелудочной железы и кишечника (двенадцатиперстная кишка). Для каждого органа использовали новую пару стерильных ножниц и пинцетов. Половину образца ткани помещали в пробирку Lysing Matrix D tube (MP Biomedicals, Solon, ОН), содержащую среду для переноса вирусов (IX минимальная основная среда, 7,5% бикарбонат натрия, 15 мМ HEPES, 1% фетальная сыворотка теленка, 4000 Ед/мл пенициллин, 400 мкг/мл гентамицин, 8 мкг/мл амфотерицин В, 4000 мкг/мл стрептомицин, 1000 мкг/мл канамицин сульфат). Вторая половина образца ткани фиксировалась в 10-процентном забуференном формалине и стандартно обрабатывалась, заливалась в воск, были получены срезы, которые прокрашивали гематоксилином и эозином (Н&Е).
C. Выделение вируса
Образцы тканей, собранные в Lysing Matrix D tubes (MP Biomedicals, Solon, OH, USA), были дважды прогомогенизированы с использованием Fastprep®-24 (MP Biomedicals) в режиме 4,0 M/S в течение 20 секунд. Гомогенизированные образцы инкубировали в течение 15 мин при комнатной температуре, после чего центрифугировали 20 мин при 2000xg при 4°С. Стерильность проверяли после инокуляции 50 мкл полученного супернатанта в 2 мл стерильного ТРВ, обогащенного 10% гидролактальбумина, с последующей инкубацией при 37°С на вращающемся (орбитальном) шейкере в течение ночи. Нестерильные образцы фильтровали через 0,45 мкм шприцевые фильтры (Whatman Inc.). Образцы хранили при -80°С. Выделение вируса проводили путем инокуляции 0,1 мл в аллантоисную полость 9-дневных зародышей SPF куриных яиц. После инкубации в течение трех дней при 37,5°С аллантоисную жидкость собирали и анализировали на наличие гемагглютинирующей активности в тесте НА.
D. Серология
Для выявления вируса в аллантоисной жидкости и анализа присутствия HI антител в собранных образцах сыворотки использовались тесты гемагглютинации (НА) и ингибирования гемагглютинации (HI) соответственно. Тесты проводили стандартными способами, используя 0,8% эритроциты цыплят, ресуспендированные в PBS. Тест HI проводили методом разведения сыворотки при постоянном уровне антигена. Для каждого разведения сыворотки использовали восемь НА единиц вирусного антигена. Средний геометрический титр был определен, как описано ранее (Brugh, 1978).
Фигура 4 показывает титры HI антител у SPF цыплят и уток, инфицированных
APMV-8. Цыплята и утки были инфицированы ороназально в дозе 10^ EID50 APMV-8.
Образцы сыворотки брали в дни 2, 4, 7, 14, и 28 после инфицирования и анализировали в тесте HI с антигеном APMV-8. HI титры сыворотки показаны в логарифмической шкале (log2) на левой оси.
E. Показатели патогенности APMV-8 у цыплят
Для оценки вирулентности APMV-8 был определен индекс интрацеребральной патогенности (intracerebral pathogenicity index, ICPI) согласно методу World Organization for Animal Health (OIE, 2008), предложенному для вируса болезни Ньюкасла. Среднее время смерти (mean dead time, MDT) для зародышей цыплят было определено, как описано ранее (Swayne et al., 1998) с использованием серийного разведения APMV-8 от 10-1 до 10-8.
Определение среднего времени смерти (MDT) для зародышей яиц, также как определение индекса интрацеребральной патогенности (ICPI), представляет собой важную характеристику патогенности вируса. Что касается MDT для зародышей цыплят, ни один из зародышей в инокулированных яйцах не умер после 7-дневного периода и, таким образом, данный изолят APMV-8 может быть классифицирован как лентогенный (со сниженной вирулентностью). Присутствие вируса было подтверждено в тесте НА с использованием аллантоисной жидкости из яиц, инокулированных с разведением 10"6. Все цыплята, инокулированные интрацеребрально, не показали клинических признаков за все время наблюдения, что говорит о значении ICPI равном нулю, что свидетельствует о лентогенном фенотипе.
F. Бустерная вакцинация APMV-8
По десять однодневных SPF цыплят на группу были инфицированы окуло-назальным путем в дозе 106 EID50 на птицу. Птицы из контрольной группы цыплят были ложно инокулированы раствором PBS. Птицы были повторно инфицированы той же дозой в день 14 после первого инфицирования. Присутствие инфекционного вируса анализировалось в дни 2, 4, 7 и 14 после первой инфекции и в дни 2 и 4 после второй инфекции путем выделения вируса из мазков из трахеи с использованием 9-дневных зародышей SPF куриных яиц. Ответ антител регистрировали по титру HI в образцах сыворотки, взятых в дни 0, 7 и 14 после каждой вакцинации.
Для исследования того, обеспечивает ли вторая иммунизация более устойчивый титр HI, был проведен эксперимент по схеме «прайм-буст» (фигура 5). В день 7 после первой инфекции, все десять птиц продемонстрировали HI в диапазоне от 64 до 1024 (GMT 207). В день 14 после инфицирования после первой инфекции титр упал (GMT 84), но увеличился после бустерной инфекции в день 14 после первой инфекции. В день 7 после бустерной вакцинации GMT увеличился до 137 и снова снизился до GMT 73 в день 14 после второй инфекции. Инфекционный вирус может быть изолирован из мазков из трахеи в день 2 (5/10 птиц) и в день 4 (4/10 птиц) после первой инфекции. После второй инфекции из мазков, взятых в дни 2 и 4 после инфицирования, вирус изолировать не удалось.
G. Выявление вирусной РНК с помощью ОТ-ПЦР
Определение вирусной РНК в образцах тканей проводили после гомогенизации образцов тканей и последующего выделения РНК с использованием набора High Pure RNA isolation kit (Roche, Mannheim, Germany). При проведении ОТ-ПЦР использовалась пара праймеров (8NPfl, 8NPr, смотри таблицу 1) с использованием набора Supercript III One Step RT-PCR kit с Platinum Taq (Invitrogen, Carlsbad, CA, USA) в соответствии с инструкциями производителя. Полученные продукты реакции анализировали в 1% агарозном геле (фигура 6). Ткани трахеи были взяты в день 2 после инфицирования из неинфицированных уток (С1-С5) и инфицированных APMV-8 уток (11-15). Ткани были прогомогенизированы и РНК приготовлена для ОТ-ПЦР. Параллельно был приготовлен контроль с водой (W). Продукты реакции разделяли в 1,5% агарозном геле. Размер фрагмента контролировали с использованием лестницы маркеров по 100 п.о. (New England Biolabs, Boston, MA, USA). Размеры фрагментов ДНК показаны справа.
H. Определение минимальной инфекционной дозы для цыплят и уток
Для того чтобы определить, какой вирусный титр будет достаточным у цыплят для выявления сероконверсии, однодневные SPF цыплята были инфицированы разными дозами APMV-8. Птицы содержались, как описано выше. По десять цыплят на группу были инфицированы каждый в дозе 101, 102, 103, 104, 105 или 106 EID50. Вирус разводили VTM. Одна группа была инокулирована VTM и использовалась в качестве контроля. У птиц брали кровь в дни 7 и 14 после инфицирования из плечевой вены. На основании результатов, полученных в эксперименте на цыплятах, трехдневные Пекинские утки были инфицированы разными количествами вируса. Были выбраны инфицирующие дозы 103, 104, 105 или 106 EID50 на одну утку. Одна группа была ложно инфицирована VTM. У птиц брали кровь в дни 7 и 14 после инфицирования из бедренной вены. Образцы сыворотки анализировали на наличие вирус-специфических антител в тесте HI как описано выше.
I. Определение патогенности у уток и цыплят
Во время данных экспериментов у цыплят и уток не наблюдалось клинических признаков. При вскрытии у трех инфицированных цыплят было обнаружено незначительное увеличение поджелудочной железы и воспаление двенадцатиперстной кишки в дни 2 и 4 после инфицирования. Во всех группах не было обнаружено никаких других заметных поражений.
Серологический ответ исследовали в дни 2, 4, 7, 14 и 28 после инфицирования путем определения титров HI в сыворотке (фигура 4). Образцы сыворотки инфицированных APMV-8 цыплят показали положительные титры HI (≥16), начиная с дня 7 после инфицирования (5/5, GMT: 111), 14 дня после инфицирования. (5/5, GMT: 48), и 28 дня после инфицирования (5/5, GMT: 48). Образцы сыворотки инфицированных APMV-8 уток также показали положительные титры HI (≥16) в дни 7 (5/5, GMT 21), 14 (5/5, GMT 28) и 28 (4/5, GMT 14) после инфицирования. Диапазон титров HI для цыплят составил от 32 до 256, тогда как для уток диапазон был от 16 до 64. Ложно инокулированные птицы обоих видов оставались отрицательными на наличие HI антител против APMV-8 во всех временных точках исследования.
Для определения участков размножения вируса у цыплят и уток было проанализировано несколько органов (трахея, легкие, двенадцатиперстная кишка и поджелудочная железа) на наличие инфекционного вируса путем выделения вируса из зародышей яиц (фигура 7). У цыплят APMV-8 был получен в день 2 после инфицирования из трахеи, легких и двенадцатиперстной кишки. В день 4 после инфицирования APMV-8 был выделен из всех проанализированных органов, тогда как в день 7 после инфицирования APMV-8 был выделен только из поджелудочной железы. В дни 14 и 28 после инфицирования ни из одного органа вирус выделить не удалось. Из ложно инокулированных птиц вирус выделить не удалось. Идентичность повторно выделенного вируса была подтверждена в тесте HI с использованием стандартной сыворотки, предоставляемой NVSL. Ни из одной собранной ткани уток не было выделено вируса ни в одной из временных точек даже после двух субпассажей в 9-дневные зародыши SPF куриных яиц. Таким образом, были проведен дизайн праймеров на основе доступной информации о последовательности APMV-8 для выявления присутствия вирусной РНК в собранных образцах тканей (фигура 7). В день 2 после инфицирования вирусная РНК была обнаружена в трахее (фигура 6), кишечнике и поджелудочной железе, тогда как в день 4 после инфицирования вирусная РНК была обнаружена во всех проанализированных органах. В дни 7, 14 и 28 после инфицирования вирусная РНК была обнаружена только в трахее и легких. ОТ-ПЦР с использованием РНК, полученной из органов ложно инокулированных птиц, не привела к амплификации ОТ-ПЦР фрагмента, что говорит об отсутствии APMV-8 у этих птиц.
Для оценки патологического потенциала исследуемого вируса органы были проанализированы на наличие микроскопических повреждений (фигура 8). В день 2 после инфицирования у всех инфицированных цыплят был обнаружен слабый многоочаговый пролиферативный трахеит. В остальных органах не было обнаружено отличий от контрольной группы. Инфицированные APMV-8 цыплята продемонстрировали фокальное восстановление или регенерацию респираторного эпителия в трахее в день 4 после инфицирования, что указывает на заживление. Кроме того, эти птицы также демонстрировали слабый многоочаговый лимфоцитарный панкреатит, что указывает на вирусную инфекцию. Инфицированные цыплята демонстрировали изменения в легких в день 7 после инфицирования, такие как от умеренного до тяжелого BALT, изменения в трахее, такие как катаральный трахеит, и многоочаговый лимфоцитарный панкреатит. Эти наблюдения также согласуются с антигенной стимуляцией. В день 14 после инфицирования у инфицированных цыплят наблюдались изменения в трахее, свидетельствующие о заживлении, и изменения в поджелудочной железе, такие как лимфоцитарный панкреатит, позволяющие сделать заключение о вирусной инфекции. В день 28 после инфицирования наблюдались только слабый катаральный трахеит и слабый энтерит у некоторых инфицированных цыплят. У инфицированных уток наблюдались многоочаговый слабый лимфоцитарный трахеит, изменения в легких (интерстициальная пневмония) и изменения в кишечнике (лимфоцитарный энтерит) в день 2 после инфицирования, тогда как изменения в трахее, свидетельствующие о респираторной инфекции, были видны в день 4 после инфицирования. В день 7 после инфицирования у инфицированных уток наблюдались лимфоцитарный трахеит и панкреатит, свидетельствующие о вирусной инфекции, тогда как наблюдаемый катаральный трахеит позволял сделать вывод о заживлении в день 14 после инфицирования. Кроме того, инфицированные утки демонстрировали лимфоцитарный панкреатит в день 14 после инфицирования. Позже, в день 28 после инфицирования у инфицированных уток в легких отмечались гиперплазия BALT и также слабый многоочаговый нейтрофильный трахеит. Оба патологических микроскопических поражения свидетельствовали о вирусной инфекции. У неинфицированных контрольных птиц не наблюдалось никаких изменений в исследуемых органах.
J. Определение минимальной дозы, необходимой для индукции иммунного ответа
Для определения минимальной инфекционной дозы, которая необходима для индукции сероконверсии у цыплят, для инфицирования десяти однодневных SPF цыплят использовались десятикратные разведения APMV-8 (фигура 9). Для теста HI использовали 4 единицы НА, в результате чего порог, равный 16, рассматривался как положительный. Образцы сыворотки, взятые в день 14 после инфицирования, показали, что доза EID50, составляющая 103, была достаточной для индукции иммунного ответа у 4/10 цыплят, что рассматривалось как положительный результат (GMT 11). В день 14 после инфицирования дозой EID50 104 девять из десяти птиц показали титр ≥16 (GMT 34). Инфицирование дозами EID50 105 и 106 индуцировало титр HI≥16 в день 14 после инфицирования у всех птиц с GMT 73 и 137 соответственно.
На основании этих результатов по 8 уток в каждой группе были инфицированы APMV-8, начиная с дозы EID50 103 на птицу до дозы EID50 106 на птицу (фигура 10). Шесть из восьми уток показали значительные титры (≥16) через 14 дней после инфицирования с GMT 14 при инфицировании дозой EID50 104/птицу. В день 14 после инфицирования 6/8 уток, инфицированных дозой EID50 105/птицу, и 7/8 уток, инфицированных дозой EID50 106/птицу, показали значительные титры (≥16) с GMT 17 и 23 соответственно.
Пример 3. Определение полноразмерной последовательности APMV-8
Для определения полноразмерной последовательности APMV-8 сначала необходима информация о последовательности вирусной РНК. Для этого был клонирован 3'-конец вирусного генома с использованием праймера (APMV-polyT, смотри таблицу 1), который содержал вырожденную последовательность, основанную на доступных 3'-последовательностях APMV1 (Genbank accession No. AF077761), APMV-2 (Genbank accession No. EU338414), и APMV-6 (Genbank accession No. EF569970). Вирусная РНК была очищена из аллантоисной жидкости с использованием набора High Pure RNA isolation kit (Roche, Mannheim, Germany). Данная последовательность была амплифицирована с использованием 5' RACE System для быстрой амплификации концов кДНК Version 2.0 (Invitrogen) в соответствии с инструкциями производителя. Было получено несколько фрагментов, которые элюировали из геля и клонировали в вектор для клонирования Торо ТА (Invitrogen), после чего положительные клоны, отобранные в результате селекции, были просеквенированы. Полученные нуклеотидные последовательности были проанализированы с использованием программы nblast search по базе данных NCBI, что не выявило сходства. Анализ tblastx search по базе данных NCBI показал сходство на 83% с нуклеопротеином из парамиксовируса 2 птиц (APMV-2/Chicken/California/Yucaipa/56, Genbank accession No. EU338414) и сходство на 56%>со штаммом парамиксовируса 6 птиц (APMV-6/Goose/FarEast/4440/2003, Genbank accession No. EF569970). С использованием этого праймера был применен метод блуждающего праймера с использованием 5' RACE System для быстрой амплификации концов кДНК Version 2.0 (Invitrogen). При проведении 5'-RACE был получен фрагмент размером примерно 800 п.о. С использованием этого метода была получена новая информация о последовательности на основе информации о последовательности из предыдущих последовательностей, которая была использована для создания новых олигонуклеотидов. 5'-Конец вирусного генома был также определен методом 5'-RACE. 3'-Конец вирусного генома был получен после лигирования РНК с использованием Т4 РНК-лигазы 1 (New England Biolabs). Продукт реакции лигирования был снова очищен с использованием набора High Pure RNA isolation kit (Roche) и была проведена ОТ-ПЦР с использованием набора Superscript III One Step RT-PCR kit с Platinum Taq (Invitrogen). Полученный фрагмент кДНК был клонирован в вектор pCR2.1 (Invitrogen) и секвенирован. Три плазмиды из каждого клонированного фрагмента были просеквенированы в обоих направлениях, в результате чего каждый нуклеотид в последовательности был прочитан 6 раз.
Полноразмерная геномная последовательность анализируемого штамма APMV-8 составила 15342 нуклеотида, что согласуется с «правилом шести» (Calain, Р. & Roux, L., 1993) для Paramyxovirinae. Было выявлено шесть открытых рамок считывания (ORF), кодирующих белки. Порядок белков был определен как 3'-NP-P-M-F-HN-L-5' (геномная последовательность SEQ ID NO:1 является антигеномной (антисмысловой) и имеет направление от 5' к 3') с использованием подобия белковых последовательностей с белками других парамиксовирусов птиц. Предполагаемые старт- и стоп-кодоны в открытых рамках считывания и теоретический молекулярный вес белков (сайт Swiss Institute of Bioinformatics ExPASy) показаны в таблице 2.
Предполагаемые геномные лидерные и трейлерные последовательности были определены путем определения предполагаемого начала гена для гена NP (лидер) и предполагаемой последовательности конца гена белка L (трейлер). Лидерная последовательность располагается от нуклеотида 1 до нуклеотида 55. Предполагаемая последовательность начала гена (нуклеотиды 56-63) гена NP завершает лидерную последовательность. Трейлерная последовательность локализована позади последовательности конца последнего гена в вирусном геноме. Из-за наличия двух возможных последовательностей конца гена для гена РНК Полимеразы (нуклеотиды 15161-15171 или 15288 -15297) были идентифицированы две предполагаемые трейлерные последовательности (нуклеотиды 15172-15342 или 15289-15342). Локализация предполагаемых последовательностей начала генов (последовательности, содержащие поли-G) и последовательностей концов генов (сигнальная последовательность для полиаденилирования) и межгенные последовательности показаны в таблице 3.
Предполагаемые последовательности начальных участков генов для APMV-8 были консервативными и содержали поли-(С)5 последовательность, расположенную перед последовательностью 3'-GCU-5'. Единственным исключением является предполагаемая последовательность начала гена для вирусной РНК-Полимеразы (3'-CUCCCGCU-5'). Предполагаемые последовательности концов генов были также консервативными и содержали поли-(U)6 последовательность на 5'-последовательности генома вируса (таблица 5).
Эти последовательности были предсказаны на основании последовательностей, которые были описаны для других парамиксовирусов из рода Avulavirus (Chang et al., 2001, Nayak et al, 2008, Jeon et al., 2008). Имеется два возможных стартовых кодона для открытой рамки считывания (ORF) РНК Полимеразы. Первый стартовый кодон (нуклеотиды 8273-8275) локализован в участке конец гена-межгенный участок-начало гена между ORF HN и ORF вирусной РНК Полимеразы. Это делает этот стартовый кодон маловероятным, но не невозможным. Второй стартовый кодон (8297 -8299) расположен ниже участка конец гена-межгенный участок-начало гена и может работать как инициирующий кодон для начала трансляции РНК-Полимеразы APMV-8.
Геном APMV-8 имеет длину 15342 нуклеотида. Это больше, чем APMV-1 (SQ ID NO:15, 15186 нуклеотидов, de Leeuw & Peeters, 1999), APMV-2 (SEQ ID NO:16, 14,904 нуклеотида, Subbiah et al., 2008) и APMV-4 (SEQ ID NO:18, 15054 нуклеотида, Nayak et al., 2008), и меньше чем APMV-3 (SEQ ID NO:17, 16,272 нуклеотида, Kumar et al., 2008) и APMV-9 (SEQ ID NO:20, 15,438 нуклеотидов, Samuel et al., 2009). Длина лидерной последовательности, состоящей из 55 нуклеотидов, вероятно, является консервативной для всех APMV (Krishnamurthy & Samal, 1998, de Leeuw & Peeters, 1999, Subbiah et al., 2008, Nayak et al., 2008, Kumar et al., 2008, Samuel et al., 2009), тогда как трейлерная последовательность, по-видимому, является вариабельной по длине. Последовательности начала генов и конца генов вирусных генов также были высококонсервативны для APMV-8 (как показано в таблице 5). Это также описано для последовательностей APMV-2 (Subbiah et al, 2008), APMV-3 (Kumar et al, 2008), APMV-4 (Jeon et al., 2008, Nayak et al, 2008), APMV-6 (Chang et al, 2001), и недавно для APMV-9 (Samuel et al., 2009). Число нуклеотидов в полноразмерной последовательности кратно шести, что согласуется с «правилом шести» для геномов парамиксовирусов (Kolakofsky et al., 1998).
Идентичность последовательностей между последовательностями геномов APMV-1, 2, 3, 4, 6, 8 и 9 показана в таблице 6.
Процент идентичности последовательностей между двумя последовательностями нуклеиновых кислот или полипептидов определяется с использованием программы Vector NTI 11.0 (PC) software package (Invitrogen, 1600 Faraday Ave., Carlsbad, CA). Для определения процента идентичности двух нуклеиновых кислот используются gap opening penalty 15 и gap extension penalty 6,66. Для определения процента идентичности двух полипептидов используются gap opening penalty 10 и gap extension penalty 0,1. Процент идентичности рассчитывается на основе более короткой последовательности.
Пример 4. Вакцинация однодневных цыплят-бройлеров штаммом APMV-8
Двадцать однодневных цыплят были разделены на две группы, как показано ниже в таблице 7.
В день 1 у однодневных цыплят брали кровь для определения статуса антител против вируса болезни Ньюкасла (NDV) и APMV-8 с использованием определения ингибирования гемагтлютинации (тест HI). Тест проводили с использованием аллантоисной жидкости из SPF яиц, инфицированных либо штаммом Lasota NDV, либо APMV-8. Для теста HI использовали четыре НА единицы штамма Lasota NDV или APMV-8 и 1% эритроцитов цыплят. Полученные HI титры показывают, что сыворотка цыплят содержала HI антитела против NDV, но не содержала определяемых количеств антител против вируса APMV-8.
В день 1 группа 1 из 10 цыплят была инфицирована через нос в дозе 106 EID50 штаммом APMV-8, группа 2 из 10 цыплят не была инфицирована и выступала в качестве контроля.
Через 14 дней после инфицирования (день 14) у цыплят брали кровь, и полученные образцы сыворотки снова анализировали на наличие HI антител против NDV и APMV-8 (фигура 20). Результаты показали, что вакцинированные APMV-8 цыплята показывают HI титры с использованием APMV-8 в качестве антигена. Специфические к APMV-8 HI тиры были между 128 и 2048. Титры HI против NDV снижались до HI титра ниже 16, следовательно, они не рассматривались как NDV-положительные. Результаты показали, что цыплята, получившие от матери антитела против NDV, не избегали инфицирования APMV-8, таким образом, интерференция таких антител с вакцинацией APMV-8 является маловероятной.
Через 14 дней после инфицирования (день 14) цыплята в группе 1 и группе 2 были разделены. По пять цыплят из группы 1 (группа 1-1) и из группы 2 (группа 2-1) были снова инфицированы в дозе 106 EID50 штаммом APMV-8 (таблица 7), а оставшиеся пять цыплят в каждой группе (группа 1-2 и группа 2-2) инфицированы не были. Этот эксперимент был проведен для того, чтобы исследовать, будет ли более позднее заражение влиять на инфекцию, и будет ли второе инфицирование (бустер-вакцинация) увеличивать титр антител. Через четырнадцать дней (день 28) у всех цыплят снова брали кровь, и исследовали сыворотку на наличие антител против APMV-8 и NDV. Титры сыворотки (фигура 21) показали, что первая вакцинация в день 14 (группа 2-1) индуцировала титры специфических антител против APMV-8, находящиеся в диапазоне от 32 до 512. У цыплят, вакцинированных только в день 1 (группа 1-2), титры антител упали до уровня между 128 и 512. У цыплят, которые были вакцинированы в день 1 и в день 14 (группа 1-1), титр антител, специфических для APMV-8, не увеличился, что позволяет предполагать, что используемый для второй вакцинации вирус был нейтрализован антителами, специфическими для APMV-8, индуцированными первой инфекцией. Сыворотка невакцинированных контрольных цыплят (группа 2-2) не содержала HI антител, специфических для APMV-8. В день 28 содержание антител против NDV снизилось еще больше, только 11 цыплят из 20 показали какие-либо титры HI с антигеном NDV, тогда как в день 14 четырнадцать цыплят демонстрировали низкие титры антител против NDV.
Пример 5. In ovo вакцинация зародышей SPF яиц
Это исследование было проведено с целью выяснения, будет ли вакцинация с APMV-8 in ovo приводить к ответу антител у цыплят, и будет ли вакцинация in ovo влиять на их вылупляемость и жизнеспособность.
В исследовании 1, 108 SPF яиц были вакцинированы in ovo вирусом штамма APMV-8 с использованием INOVOJECT (Pfizer Animal Health, NY, USA) в день 18 инкубации. Вирус был разведен стерильным 0,9% раствором NaCl. Обратное титрование разведенного вируса показало титр 105,5 EID50 /100 мкл. В качестве контроля 108 яиц инокулировали стерильным 0,9% раствором NaCl. Объем введения составил 100 мкл на одно яйцо. В контрольной группе вылупилось восемьдесят цыплят, а в вакцинированной APMV-8 группе вылупилось сорок пять цыплят. По десять цыплят из каждой группы пересадили в отдельные клетки Horsefall-Bauer. Кроме того, цыплята из вакцинированной APMV-8 группы и пять цыплят из контрольной группы были посажены вместе в одну клетку Horsefall-Bauer для проверки возможности передачи APMV-8 после вакцинации. Вода и корм предоставлялись ad libitum. Через четырнадцать и через двадцать восемь дней после вылупления были взяты образцы крови и протестированы на присутствие HI антител против APMV-8 с использованием 4 единиц НА и 1% раствора эритроцитов цыплят. Результаты (фигура 22) показали, что вакцинация in ovo в день 18 инкубации яиц вызывает иммунный ответ, на что указывает присутствие титров HI в протестированных образцах сыворотки. Через четырнадцать дней после вылупления наблюдались титры HI от 256 до 4048 у группы, вакцинированной in ovo. В сыворотке контрольных цыплят, контактировавших с вакцинированными цыплятами, через 14 дней после начала контакта наблюдались HI титры от 256 до 2048, что говорит о распространении вируса, применявшегося для вакцинации. В контрольной группе не было обнаружено HI титра, специфического для APMV-8. Еще через четырнадцать дней у цыплят снова брали кровь, в которой были обнаружены титры от 256 до 4096 в вакцинированной APMV-8 группе и от 256 до 1024 в группе, контактировавшей с вакцинированными APMV-8 цыплятами. В контрольной группе HI антител против APMV-8 обнаружено не было.
В исследовании 2, 88 SPF (свободных от специфического антигена) яиц были вакцинированы in ovo с использованием INOVOJECT в день 19 инкубации. Штамм вируса APMV-8 был разведен стерильным 0,9% раствором NaCl. Титр вируса составил 105,75 EID50 /100 мкл, как показало обратное титрование разведенного вируса. В качестве контроля 88 SPF яиц инокулировали стерильным 0,9% раствором NaCl. Объем введения составил 100 мкл на одно яйцо. В контрольной группе (NaCl) вылупилось семьдесят четыре цыпленка, а в группе, вакцинированной APMV-8, вылупилось семьдесят шесть цыплят. По десять цыплят из каждой группы пересадили в отдельные клетки Horsefall-Bauer. Вода и корм предоставлялись ad libitum. Через четырнадцать дней после вылупления были взяты образцы крови и протестированы на присутствие HI антител против APMV-8 с использованием 4 единиц НА и 1% раствора эритроцитов цыплят. Результаты (фигура 23) показали, что вакцинация in ovo в день 19 инкубации яиц приводит к иммунному ответу с HI титрами, специфическими для APMV-8. Через четырнадцать дней после вылупления в группе, вакцинированной APMV-8 in ovo, наблюдался титр HI от 256 до 4048. В контрольной группе не наблюдалось какого-либо титра, специфичного для APMV-8. В день 28 после вылупления у цыплят снова была взята кровь. Титры HI в группе, вакцинированной APMV-8, составили от 512 до 4096 (фигура 23), тогда как сыворотка контрольных цыплят все еще оставалась отрицательной в отношении APMV-8.
В третьем исследовании по 108 SPF яиц в группе 1 и группе 2 были вакцинированы in ovo с использованием INOVOJECT в день 18 инкубации в дозах 103,5 EID50 и 104,5 EID50 соответственно. Штамм вируса APMV-8 был разведен стерильным 0,9% раствором NaCl. В третьей группе 108 SPF яиц были инокулированы стерильным 0,9% раствором NaCl в качестве контроля. В группе 1 вывелось восемьдесят три цыпленка, в группе 2 вывелось семьдесят девять цыплят, и в группе 3 вывелось восемьдесят восемь цыплят (смотри таблицу 8). После вылупления по десять цыплят из каждой группы пересадили в отдельные клетки Horsefall-Bauer. Вода и корм предоставлялись ad libitum. После вылупления десять цыплят из группы 3 были вакцинированы подкожно в шею с использованием 100 мкл 106 EID50 вируса APMV-8. Через четырнадцать дней после вылупления были взяты образцы крови и протестированы на наличие HI антител против APMV-8 с использованием 4 единиц НА и 1% раствора эритроцитов цыплят. Результаты (фигура 24) показали, что вакцинация in ovo в день 18 инкубации приводит к иммунному ответу с титрами HI, специфическими для APMV-8. Через 14 дней после вылупления у группы, вакцинированной in ovo вирусом APMV-8, наблюдались титры HI от 4096 до 16384. В контрольной группе титра HI антител, специфических для APMV-8, обнаружено не было. Группа, вакцинированная APMV-8 подкожно, показала сероконверсию с титром в диапазоне от 64 до 4096. В день 28 после вылупления у цыплят снова брали кровь и тестировали на наличие HI антител, специфических для APMV-8. Титры HI через четыре недели после вылупления снизились и находились в диапазоне от 512 до 4096. В контрольной группе не было обнаружено титра HI, специфического для APMV-8. Титр HI в труппе, которая была вакцинирована подкожно, находился в диапазоне от 32 до 256.
Пример 6. Разработка системы обратной генетики для штамма APMV-8 и создание мутантов APMV-8, экспрессирующих гетерологические гены
Конструирование и экспрессия плазмид, содержащих гены NP, Р, и L вируса APMV-8
Для создания обратной генетической системы для парамиксовирусов необходимо создание плазмид, экспрессирующих белки, участвующие в репликации вирусной РНК. Открытые рамки считывания (ORF) для трех белков APMV-8 (Нуклеопротеин NP, Фосфопротеин Р, белок РНК-зависимая РНК-полимераза или белок L) были клонированы в эукариотический экспрессионный вектор pcDNA3 (Invitrogen, California, USA). Для этого РНК из аллантоисной жидкости, содержащей APMV-8, была очищена с использованием набора High Pure RNA Isolation Kit (Roche, Basel, Switzerland). Очищенная РНК была использована для обратной транскрипции в полимеразной цепной реакции ОТ-ПЦР с использованием набора Titan One Tube RT-PCR Kit (Roche). Открытые рамки считывания белков были амплифицированы с использованием подходящих пар праймеров [NP (NP-FP, NP-RP), Р (P-FP, P-RP), L (L-FP, L-RP), смотри таблицу 9]. Продукты реакции разделяли в 0,7% агарозном геле и элюировали из геля с использованием набора QIAquick Gel Extraction Kit (Qiagen, Hilden, Germany) в соответствии с инструкциями производителя. Фрагменты, полученные с помощью ОТ-ПЦР, инкубировали с подходящими ферментами рестрикции (NP и Р с Eco RI/NotI; L с Kpn I/NotI), снова элюировали из геля и лигировали в эукариотический экспрессионный вектор pcDNA3, расщепленный соответствующими ферментами рестрикции. Клетки Topi OF (Invitrogen) трансформировали продуктами реакции лигирования в, и плазмидную ДНК, выделенную из клеток Topi0F, расщепляли соответствующими ферментами рестрикции (смотри выше). Плазмиды, содержащие фрагменты ДНК (pcDNA-NP, pcDNA-Р, pcDNA-L) нужного размера, секвенировали.
праймера
b Ориентация последовательности праймеров дана в соответствии с вирусной информационной РНК.
с Показано положение вирус-специфических последовательностей в полноразмерном геноме вируса.
Конструирование плазмиды, содержащей полноразмерный геном APMV-8
Для создания плазмиды, содержащей полноразмерный геном APMV-8, была синтезирована кДНК генома вируса (Genscript, New York, USA) на основе созданных консенсусных последовательностей из двух разных частей (5'-FLG, З'-FLG, смотри фигуру 25). 5'-Часть (5'-FLG, SEQ ID NO:47) вирусной последовательности была синтезирована из нуклеотидов 1-5564. Предшествующая последовательность APMV-8 представляет собой последовательность кассеты, состоящей из промотора CMV-IE, расположенной за ним последовательности для расщепления ферментом рестрикции Xmal (для возможных последующих процедур клонирования) и последовательности «хаммерхед» рибозима (в виде головки молотка, hammerhead ribozyme). На 5'-конце и 3'-конце данной последовательности были добавлены сайты для расщепления ферментами
рестрикции Not I и SacII соответственно. Синтезированная 3'-часть (З'-FLG, SEQ ID NO:48) последовательности (нуклеотиды 5503-15342) завершалась последовательностью рибозима гепатита дельта и поли-А сигнальной последовательностью гормона роста быка. Для целей клонирования на 5'-конец З'-FLG была добавлена последовательность для расщепления ферментом рестрикции Not I, и на 3'-конце последовательности была добавлена последовательность для расщепления ферментом рестрикции Sac П.
В пределах перекрывающихся частей 5'-FLG и З'-FLG (нуклеотиды 5503-5564) находилась последовательность для уникального фермента рестрикции (Bmt I), который расщепляет последовательность по нуклеотиду 5541 полноразмерной последовательности. Обе части ДНК (5'-FLG, З'-FLG) были лигированы по отдельности в плазмиду pUC57 (Genscript) с получением плазмид pUC57/5'-FLG и pUC57/3'-FLG. Для клонирования вместе обоих фрагментов pUC57/5'-FLG была расщеплена ферментами Bmtl и Sac II и содержащая 5'-FLG плазмида была элюирована из геля. Параллельно pUC57/3'-FLG была расщеплена теми же ферментами и фрагменты З'-FLG были элюированы из геля. Затем З'-FLG был лигирован в содержащую 5'-FLG плазмиду для получения плазмиды, которая содержит полноразмерную последовательность кДНК генома APMV-8 под контролем промотора CMV-IE (pUC57-FL-APMV-8). Конструирование плазмиды, содержащей минигеном APMV-8
Плазмида, содержащая все функциональные элементы минигенома для APMV-8 (pMG-APMV-8), была сконструирована с использованием метода, описанного Conzelmann, et al. (J Virol. 68:713-719, 1994). Плазмида pMG-APMV-8 содержит трейлерный и лидерный участок генома APMV-8, который фланкирован промотором Т7 и последовательностью рибозима антигенома вируса гепатита дельта (Collins, et al., PNAS USA 88:9663-9667, 1991). После последовательности рибозима антигенома вируса гепатита дельта располагается последовательность терминатора транскрипции Т7. Между трейлерным и лидерным участками располагается кодирующая последовательность усиленного зеленого флуоресцентного белка в антисмысловой ориентации. Перед трейлерной последовательностью и сразу за промотором Т7 располагаются три дополнительных остатка G. Вставка была фланкирована сайтами расщепления ферментов рестрикции Eco RI и Not I и была клонирована по тупым концам в плазмиду pUC57. Эта конструкция была субклонирована в плазмиду pUC18. Для этого плазмида pMG-APMV-8 была расщеплена ферментами Eco RI и Hind III и нужный фрагмент был элюирован из геля и лигирован в расщепленную соответствующим образом плазмиду pUC18 для получения плазмиды puC18-MG-APMV-8. Наличие вставки было подтверждено секвенированием.
Создание экспрессионной плазмиды, обеспечивающей экспрессию Т7-полимеразы
Для создания плазмиды, кодирующей Т7 ДНК-зависимую РНК-полимеразу (Т7 полимеразу), кодирующая последовательность (GenBank accession number AY264778) была синтезирована с использованием Genscript. Последовательность Т7-полимеразы (SEQ ID NO:49) была модифицирована для оптимизации кодонов, используемых для экспрессии в эукариотической системе, и для удаления возможных донорных/акцепторных участков сплайсинга в последовательности. Последовательность, кодирующая Т7-полимеразу, была фланкирована сайтами для EcoRI (5') и NotI (3'). Синтезированный фрагмент был клонирован по тупым концам в вектор pUC57 (pCU57-Т7). Эта плазмида была расщеплена ферментами EcoRI/NotI и кодирующий Т7-полимеразу фрагмент был элюирован из геля. Затем данный фрагмент был клонирован в эукариотический экспрессионный вектор pcDNA3 (Invitrogen) для получения pcDNA3-T7. Наличие фрагмента в векторе pcDNA3-T7 было подтверждено секвенированием. Создание плазмиды для экспрессии усиленного зеленого флуоресцентного белка с использованием внутреннего сайта связывания рибосомы под контролем промотора Т7
Для тестирования функциональной активности Т7-полимеразы методом ПЦР была амплифицирована открытая рамка считывания усиленного зеленого флуоресцентного белка (EGFP) с использованием плазмиды pEGFP-Nl (Clontech, California, USA) и следующей пары праймеров:
EGFP-FP fCCGGATCCATGGTGAGCAAGGGCGAGGAGCTG) SEQ ID NO:50 и EGFP-RP CCCGCGGCCGCTTACTTGTACAGCTCGTCCATGCCG) SEQ ID NO:51
Полученный ПЦР-фрагмент был элюирован из геля и проинкубирован с ферментами рестрикции BamHI и NotI. Продукт реакции был элюирован из геля и лигирован в соответствующим образом расщепленный вектор pCITE 4А (Novagen). Полученная плазмида (pCITE4A-EGFP) была использована в последующих экспериментах. Плазмиды pCITE4A-EGFP и pcDNA3-T7 были трансфецированы по отдельности или вместе в клеточную линию кур DF1, выращиваемую в 24-луночных планшетах, с использованием Lipofectin 2000 (Invitrogen). Через двадцать четыре часа после трансфекции среду удаляли и добавляли стерильный фосфатный буферный раствор (PBS). Клетки исследовали с использованием инвертированного флуоресцентного микроскопа Axiovert 40 CFL (Zeiss, Jena, Germany). Зеленая флуоресценция наблюдалась только в лунках планшета с культурой клеток, которые были одновременно трансфецированы обеими плазмидами. Результаты говорят о том, что обе плазмиды, pCITE4A-EGFP и pcDNA3-T7, были функционально активны.
Подтверждение функциональной активности вирусных белков NP. Р и L. экспрессированных с использованием минигеномной плазмиды
Клетки DF1 были котрансфецированы pcDNA3-T7, pUC18-MG-APMV-8, pcDNA-NP, pcDNA-P и pcDNA-L для подтверждения функциональной активности экспрессированных белков NP, Р и L. Через двадцать четыре часа после трансфекции среду удаляли и добавляли стерильный раствор фосфатного буфера (PBS). Клетки исследовали с использованием инвертированного флуоресцентного микроскопа Axiovert 40 CFL (Zeiss, Jena, Germany). Зеленая флуоресценция наблюдалась только в лунках планшета с культурой клеток, которые были трансфецированы всеми 5 плазмидами. Эти результаты указывают, что экспрессированные вирусные белки NP, Р и L были функционально активны и обеспечивали транскрипцию минигенома APMV-8 в мРНК и экспрессию белка EGFP, кодируемого pUC18-MG-APMV-8.
Восстановление вируса AMPV8 из плазмиды, содержащей полноразмерную последовательность APMV-8
Клетки DF1 были котрансфецированы pUC57-FL-APMV-8, pcDNA-NP, pcDNA-P, и pcDNA-L. Через 48-96 часов супернатанты клеток DF1 инокулировали в 1-дневные зародыши яиц для размножения вируса. Через 3-5 дней собирали аллантоисную жидкость и тестировали на гемагглютинацию (НА) с использованием 1% раствора эритроцитов кур. Аллантоисная жидкость, положительная на НА активность, использовалась в трех исследованиях: 1) клетки DF1 инфицировались аллантоисной жидкостью и тестировались через 36 часов после инфекции с APMV-8-специфической антисывороткой на наличие экспрессии белков APMV-8 в непрямом иммунофлуоресцентном анализе. 2) Аллантоисная жидкость тестировалась на специфичность в отношении APMV-8 с использованием APMV-8-специфичной сыворотки кур (предоставлена National Central Veterinary Laboratory, Ames, Iowa, USA) в исследовании ингибирования гемагглютинации. 3) Восстановление вируса было установлено ОТ-ПЦР с использованием специфических для APMV-8 олигонуклеотидов. Отсутствие вирусной кДНК было подтверждено исключением стадии обратной транскрипции во время реакции. Образцы, положительные по всем трем тестам, далее размножались в зародышах SPF куриных яиц. Размножение APMV-8 в клетках, не являющихся куриными клетками
Клетки из разных видов животных [хомячка (клетки почек хомячка, Baby hamster kidney cells, клетки BHK-21), мартышки (Vero cells, клеточная линия, полученная из почек Африканской зеленой мартышки), собаки (клетки почек собаки, Madin-Darby canine kidney cells, MDCK) и перепела (линия мышечных клеток перепела, Quail muscle cell line QM7)] выращивали в 24-луночных планшетах для культивирования тканей и инфицировали многочисленными инфекциями 0,01. Клетки фиксировали ледяным этанолом через 24 часа после инфекции и анализировали на наличие специфических для APMV-8 белков с помощью непрямой иммунофлуоресценции с использованием специфичной к APMV-8 антисыворотки из инфицированных APMV-8 SPF цыплят. Связывание антител визуализировали с использованием ФИТЦ-конъюгата козы, специфичного против IgY цыплят. Неинфицированные клетки использовались в качестве отрицательного контроля. Зеленая флуоресценция наблюдалась только в инфицированных APMV-8 клетках. Это указывает на то, что APMV-8 был способен инфицировать клетки из других видов животных, а не только клетки кур.
Репликация APMV-8 была увеличена в присутствии трипсина. Клетки MDCK были инфицированы APMV-8, как описано выше. После инфекции клетки промывали бессывороточной средой и переносили либо в бессывороточную среду, содержащую трипсин в концентрации 1 мкг/мл, либо в бессывороточную среду без трипсина. Через двадцать четыре, сорок восемь и девяносто шесть часов после инфекции супернатанты клеток удаляли и в клетках DF1 определяли TCID50 с использованием непрямой иммунофлуоресценции, как описано выше. Полученные данные указывают на то, что в присутствии трипсина APMV-8 реплицируется до более высокого титра, чем в отсутствие этого фермента (фигура 19 В).
Результаты этого эксперимента показали, что подобно AMPV-1, APMV-8 способен проникать в клетки разных видов и инициировать свой репликационный цикл. Таким образом, он представляет собой вектор, подходящий для множества видов. Получение рекомбинантного вируса APMV-8, экспрессирующего чужеродные гены, с использованием системы обратной генетики
Для создания рекомбинантного вируса APMV-8 (вирусного вектора), экспрессирующего ген гемагглютинина (НА) высокопатогенного птичьего гриппа (HPAI), кодирующая последовательность гена НА подтипов Н5 или Н7 вируса HPAI была вставлена в несущественные участки, например, между генами М и F или между генами Р и М генома APMV-8, в плазмиду, содержащую полноразмерный геном APMV-8. Для этой цели кодирующая последовательность открытой рамки считывания гемагглютинина фланкировалась всеми необходимыми регуляторными последовательностями гена F, которые включали последовательность старта гена, 5'-некодирующую последовательность, 3'-некодирующую последовательность и последовательность конца гена. Конструкция была синтезирована таким образом, чтобы сайты расщепления ферментами рестрикции Bsu 361 и Nhe I можно было использовать для лигирования нужного фрагмента в существующую плазмиду, содержащую полноразмерный геном APMV-8, за счет их уникальности в плазмидной конструкции. Полученная плазмида представляла собой желаемую траскрипционную плазмиду, которая содержит ген гемагглютинина в несущественном участке полноразмерного генома APMV-8. С использованием этого подхода можно клонировать кодирующие последовательности множества вирусных и бактериальных антигенов в каркас последовательности APMV-8. Другие возможные антигены, которые могут быть вставлены в геном APMV-8, представляют собой слитый белок вируса болезни Ньюкасла, S-белок вируса птичьего бронхита, другие гены гемагглютинина вирусов птичьего гриппа, отличные от Н5 и Н7, ген структурного белка VP1 вируса куриной анемии, гены гликопротеинов из вируса инфекционного ларинготрахеита.
Пример 7. Вакцинация животных
Животные вакцинируются одним, двумя введениями или по схеме «прайм-буст» композицией или вакциной, содержащей рекомбинантный вирус APMV-8 (вирусный вектор), как описано в примере 6. Для цыплят/птиц проводятся разные режимы введения, например, введение in ovo в день 18 или день 19, подкожное введение однодневным птенцам, или мукозальное введение (спрей, питьевая вода, глазные капли) в разном возрасте. Дозировка составляет между 3 и 7 log10 (предпочтительно 4-6 log10 EID50). Для млекопитающих применяется мукозальный (интраназальный, интраокулярный, пероральный) или парентеральный (внутримышечный, подкожный, трансдермальное или внутрикожное введение без иглы) пути введения. Дозировки составляют от 5 до 9 log (предпочтительно 6-8 log). Обычно проводится два введения с интервалом в 3-4 недели. Гетерологический прайм-буст (например, бустерное введение белков) также является предпочтительным.
Эффективность защиты, индуцируемая композицией или вакциной, оценивается против специфического патогена, вводимого животным. Защитная эффективность оценивается путем клинических наблюдений и/или вирусной нагрузки специфического патогена в тканях, крови или мазках слизистых. Образцы крови вакцинированных животных берутся на разных стадиях и тестируются на серологию. Результаты показывают, что композиция или вакцина настоящего изобретения является иммуногенной и обеспечивает защиту вакцинированных животных.
Таким образом, познакомившись с детальным описанием предпочтительных воплощений настоящего изобретения, следует понимать, что настоящее изобретение, описанное выше, не ограничивается приведенными выше конкретными деталями, и что возможно множество очевидных вариаций настоящего изобретения без выхода за рамки или дух настоящего изобретения.
Все процитированные или упомянутые здесь документы («процитированные здесь документы») и все документы, процитированные или упомянутые в процитированных здесь документах, вместе с любыми инструкциями производителей, описаниями, спецификациями продуктов и технологическими картами для любых продуктов, упомянутых здесь или в любом документе, включенном в настоящее изобретение путем отсылки, являются также включенными в настоящее изобретение путем отсылки и могут применяться при осуществлении настоящего изобретения.
Ссылки:
Alexander, 1988. Newcastle disease, p.x, 378 p.In D. J. Alexander (ed.), Developments in veterinary virology. Kluwer Academic, Boston.
Alexander, Characterization of viruses which represent further distinct serotypes (PMV-8 and PMV-9) of avian paramyxoviruses. Arch Virol 78:29-36.
Alexander, et al., 1979. Properties of a newly isolated, serologically distinct avian paramyxovirus. Arch Virol 60:105-13.
Alexander, 2003. Newcastle Disease, other Paramyxoviruses, and Pneumovirus Avian Paramyxoviruses 2-9, p.63-99. In Y. M. Saif (ed.), Diseases of poultry, 11th ed. Iowa State Press, Ames, Iowa.
Andral, et al., 1984. Isolation of avian paramyxovirus 2 and 3 from turkeys in Brittany. Vet Rec 114:570-1.
Bankowski, et al., 1981, Effect of paramyxovirus yucaipa on fertility, hatchability, and poult yield of turkeys. Avian Dis 25:517-20.
Bankowski, et al., 1960. Isolation of an Unidentified Agent from the Respiratory Tract of Chickens. Science 132:292-293.
Bradshaw, et al., 1979. The Epidemiology of Yucaipa Virus in Relationship to the Acute Respiratory Disease Syndrome in Turkeys. Avian Diseases 23:539-542.
Capua, et al., 2004. Isolation of an avian paramyxovirus type 9 from migratory waterfowl in Italy. Vet Rec 155:156.
Chambers, et al., 1988. Protection of chickens from lethal influenza infection by vaccineexpressed hemagglutinin. Virology 167:414-421.
Collins, P. L., et al., 1991. Rescue of synthetic analogs of respiratory syncytial virus genomic RNA and effect of truncations and mutations on the expression of a foreign reporter gene. Proc. Natl. Acad. Sci. USA 88:9663-9667
Conzelmann KK, and Schnell M., 1994. Rescue of synthetic genomic RNA analogs of rabies virus by plasmid-encoded proteins. J Virol. 68:713-719
Darteil, R., M. Bublot, E. Laplace, J.F. Bouquet, J.C. Audonnet and M. Riviere (1995). Herpesvirus of turkey recombinant viruses expressing infectious bursal disease virus (IBDV) VP2 immunogen induce protection against an IBDV virulent challenge in chickens. Virology 211,481^90.
de Leeuw, et al., 1999. Complete nucleotide sequence of Newcastle disease virus: evidence for the existence of a new genus within the subfamily Paramyxovirinae. J Gen Virol 80:131-136. Fleury, et al., 1979. Isolation of twenty-three Yucaipa-like viruses from 616 wild birds in Senegal, West Africa. Avian Dis 23:742-4.
Gao, et al., (2006). Protection of mice and poultry from lethal H5N1 avian influenza virus through adenovirus-based immunization. J Virol 80:1959-1964.
Ge, et al., (2007) Newcastle disease virus-based live attenuated vaccine completely protects chickens and mice from lethal challenge of homologous and heterologous H5N1 avian influenza viruses. Journal of Virology, 81(1), 150-158.
Goodman, et al., 1988. Isolation of avian paramyxovirus-2 from domestic and wild birds in Costa Rica. Avian Dis 32:713-7.
Gough, et al., 1984. Avian paramyxovirus type 4 isolated from a ringed teal (Calonetta leucophrys). Vet Rec 115:653.
Hoelscher, et al., (2008). A broadly protective vaccine against globally dispersed clade 1 and clade2H5Nl influenza viruses. J Infect Dis. 197:1185-1188.
Huang, et al., (2004) A recombinant Newcastle Disease Virus (NDV) expressing VP2 protein of Infectious Bursal Disease Virus (IBDV) protects against NDV and IBDV. Journal of Virology, 78, 10054-10063.
Hunt, et al., (1988). Retrovirus-expressed hemagglutinin protects against lethal influenza virus infections. J Virol 62:3014-3019.
Inoue K, Shoji Y, Kurane I, Iijima T, Sakai T, Morimoto K. (2003). An improved method for recovering rabies virus from cloned cDNA. J Virol Methods. 107:229-236. Krishnamurthy, et al., 1998. Nucleotide sequences of the trailer, nucleocapsid protein gene and intergenic regions of Newcastle disease virus strain Beaudette С and completion of the entire genome sequence. J Gen Virol 79:2419-2424.
Krishnamurthy, S., Huang, Z. & Samal, S. K. (2000) Recovery of a virulent strain of Newcastle disease virus from cloned cDNA: expression of a foreign gene results in growth retardation and attenuation. Virology, 278, 168-182.
Lamb, et al., 2007. Paramyxoviridae: The viruses and Their Replication, p.1449-1496. In B. N. Fields, D. M. Knipe, and P. M. Howley (ed.), Fields' virology 5th ed. Wolters Kluwer Health/Lippincott Williams & Wilkins, Philadelphia.
Lang, G., A. Gagnon, and J. Howell. 1975. The occurrence of Paramyxovirus yucaipa in Canadian poultry. Can Vet J 16:233-7.
Lipkind, et al., 1982. Isolation of yucaipa-like avian paramyxovirus from a wild mallard duck (Anas platyrhinchos) wintering in Israel. Vet Rec 110:15-6.
Lipkind, et al., 1979. The isolation of yucaipa-like paramyxoviruses from epizootics of a respiratory disease in turkey poultry farms in Israel. Vet Rec 105:577-8.
Maldonado, et al., (1995) Serological survey for avian paramyxoviruses from wildfowl in aquatic habitats in Andalusia. Journal of Wildlife Diseases, 31(1), 66-69.
Mayo, et al., 2002. A summary of taxonomic changes recently approved by ICTV. Arch Virol 147:1655-63.
Nayak B, et al., (2008). Molecular characterization and complete genome sequence of avian
paramyxovirus type 4 prototype strain duck/Hong Kong/D3/75. Virol J. 20;5:124.
Park, et al., (2006) Engineered viral vaccine constructs with dual specificity: Avian influenza and
Newcastle disease. Proceedings of the National Academy of Sciences, 103(21), 8203-8208.
Peeters, et al., (1999) Rescue of Newcastle disease virus from cloned cDNA: evidence that
cleavability of the fusion protein is a major determinant for virulence. Journal of Virology, 73(6),
5001-5009.
Redmann, et al., 1991. [Isolation of a paramyxovirus-3 from turkeys with respiratory tract disease in Germany]. Dtsch Tierarztl Wochenschr 98:138-41.
Reed, et al., 1938. A SIMPLE METHOD OF ESTIMATING FIFTY PER CENT ENDPOINTS. Am. J. Epidemiol. 27:493-497.
Romer-Oberdorfer, et al., (1999) Generation of recombinant lentogenic Newcastle disease virus from cDNA. Journal of General Virology, 80, 2987-2995.
Rosenberger, et al., (1974) Isolation of Newcastle disease and type-A influenza viruses from migratory waterfowl in the Atlantic flyway. Avian Diseases, 18(4), 610-613. Saif, et al., 1997. Natural and Experimental Infection of Turkeys with Avian Paramyxovirus-7. Avian Diseases 41:326-329.
Shihmanter, et al., 1998. Isolation of avian serotype 3 paramyxoviruses from imported caged birds in Israel. Avian Dis 42:829-31.
Shihmanter, et al., 1998. Avian paramyxoviruses serotype 3 isolated from captive birds in Israel: clinical signs, pathology, and antigenic characterization. Avian Dis 42:418-22. Shortridge, et al., 1980. Isolation and properties of viruses from poultry in Hong Kong which represent a new (sixth) distinct group of avian paramyxoviruses. J Gen Virol 49:255-262. Schultz-Cherry, et al., (2000). Influenza virus (A/HK/156/97) hemagglutinin expressed by an alphavirus replicon system protects chickens against lethal infection with Hong Kong-origin H5N1 viruses. Virology 278:55-59.
Stallknecht, et al., (1991) Avian paramyxoviruses from migrating and resident ducks in coastal Louisiana. Journal of Wildlive Diseases. 27:123-128.
Stanislawek, et al., (2002) Avian paramyxoviruses and influenza viruses isolated from mallard ducks (Anas platyrhynchos) in New Zealand. Archives of Virology, V147, 1287-1302. Tang M, et al., 2002. Recombinant adenovirus encoding the HA gene from swine H3N2 influenza virus partially protects mice from challenge with heterologous virus: A/HK/I/68 (H3N2). Arch Virol 147:2125-2141.
Taylor, et al., (1998). Protective immunity against avian influenza induced by a fowlpox virus recombinant. Vaccine 6:504-508.
Того, et al., (2007). Protective avian influenza in ovo vaccination with non-replicating human adenovirus vector. Vaccine 25:2886-2891.
Tumova, et al., 1979. A hitherto unreported paramyxovirus of turkeys. Res Vet Sci 27:135-40. Tumova, et al., 1989. Further evidence of the circulation of PMV-4 and influenza viruses with N2-1957 enzyme in the migratory waterfowls. Acta Virol 33:573-6.
Veits, et al., (2003). Deletion of the non-essential UL0 gene of infectious laryngotracheitis (ILT) virus leads to attenuation in chickens, and UL0 mutants expressing influenza virus haemagglutinin (H7) protect against ILT and fowl plague. J Gen Virol 84:3343-3352. Veits, et al., (2006) Newcastle disease virus expressing H5 hemagglutinin gene protects chickens against Newcastle disease and avian influenza. Proceedings of the National Academy of Sciences, 103(21), 8197-8202.
Webster, et al., 1976. Ortho- and paramyxoviruses from migrating feral ducks: characterization of a new group of influenza A viruses. J Gen Virol 32:217-25.
Yamane, et al., 1982. Characterization of avian paramyxoviruses isolated from feral ducks in northern Japan: the presence of three distinct viruses in nature. Microbiol Immunol 26:557-68. Zhang, et al., 2007. Serological survey on prevalence of antibodies to avian paramyxovirus serotype 2 in China. Avian Dis 51:137-9.
Zhang, et al., 2006. Isolation, identification, and comparison of four isolates of avian paramyxovirus serotype 2 in China. Avian Dis 50:386-90.
Zou, et al., 2005. Complete Genome Sequence and Biological Characterizations of A Novel Goose Paramyxovirus-SF02 Isolated in China. Virus Genes 30:13-21.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНЫЕ ВЕКТОРЫ HVT, ЭКСПРЕССИРУЮЩИЕ АНТИГЕНЫ ПАТОГЕНОВ ПТИЦ И ИХ ПРИМЕНЕНИЕ | 2012 |
|
RU2620936C2 |
| РЕКОМБИНАНТНЫЕ HVT-ВЕКТОРЫ, ЭКСПРЕССИРУЮЩИЕ МНОЖЕСТВЕННЫЕ АНТИГЕНЫ ПАТОГЕНОВ ПТИЦ, И СОДЕРЖАЩИЕ ИХ ВАКЦИНЫ | 2017 |
|
RU2752836C2 |
| РЕКОМБИНАНТНАЯ ВАКЦИНА ОТ ПТИЧЬЕГО ГРИППА И ЕЁ ПРИМЕНЕНИЕ | 2009 |
|
RU2559527C2 |
| ВИРУС ГРИППА, СПОСОБНЫЙ ИНФИЦИРОВАТЬ СОБАЧЬИХ, И ЕГО ПРИМЕНЕНИЕ | 2006 |
|
RU2520081C2 |
| ВИРУС ГРИППА, СПОСОБНЫЙ ИНФИЦИРОВАТЬ СОБАЧЬИХ, И ЕГО ПРИМЕНЕНИЕ | 2014 |
|
RU2711807C2 |
| ВАКЦИНА ДЛЯ ЗАЩИТЫ ПТИЦЫ ОТ QХ-ПОДОБНОГО ВИРУСА ИНФЕКЦИОННОГО БРОНХИТА И СПОСОБ ЕЕ ИСПОЛЬЗОВАНИЯ | 2009 |
|
RU2568053C2 |
| РЕКОМБИНАНТНЫЙ ВИРУС БОЛЕЗНИ НЬЮКАСТЛА, ЭКСПРЕССИРУЮЩИЙ Н5 ГЕМАГЛЮТИНИН ВИРУСА ПТИЧЬЕГО ГРИППА (AVIAN INELUENZA) | 2007 |
|
RU2441070C2 |
| ВЕКТОРЫ НА ОСНОВЕ РЕКОМБИНАНТНЫХ ВИРУСОВ ОТРЯДА Mononegavirales | 2007 |
|
RU2435857C2 |
| МАТЕРИАЛЫ И МЕТОДЫ, ИСПОЛЬЗУЕМЫЕ ДЛЯ ЛЕЧЕНИЯ РЕСПИРАТОРНЫХ ЗАБОЛЕВАНИЙ У СОБАК | 2011 |
|
RU2711791C2 |
| ШТАММ ВИРУСА ГРИППА А СОБАК (ВАРИАНТЫ), ИММУНОГЕННЫЙ ПОЛИПЕПТИД, ПОЛИНУКЛЕОТИД, КОДИРУЮЩИЙ ЕГО, ВЕКТОР ЭКСПРЕССИИ ПОЛИПЕПТИДА, ИММУНОГЕННАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ ПОЛИПЕПТИД, И СПОСОБ ИНДУКЦИИ ИММУННОГО ОТВЕТА У ЖИВОТНОГО | 2006 |
|
RU2449014C2 |



















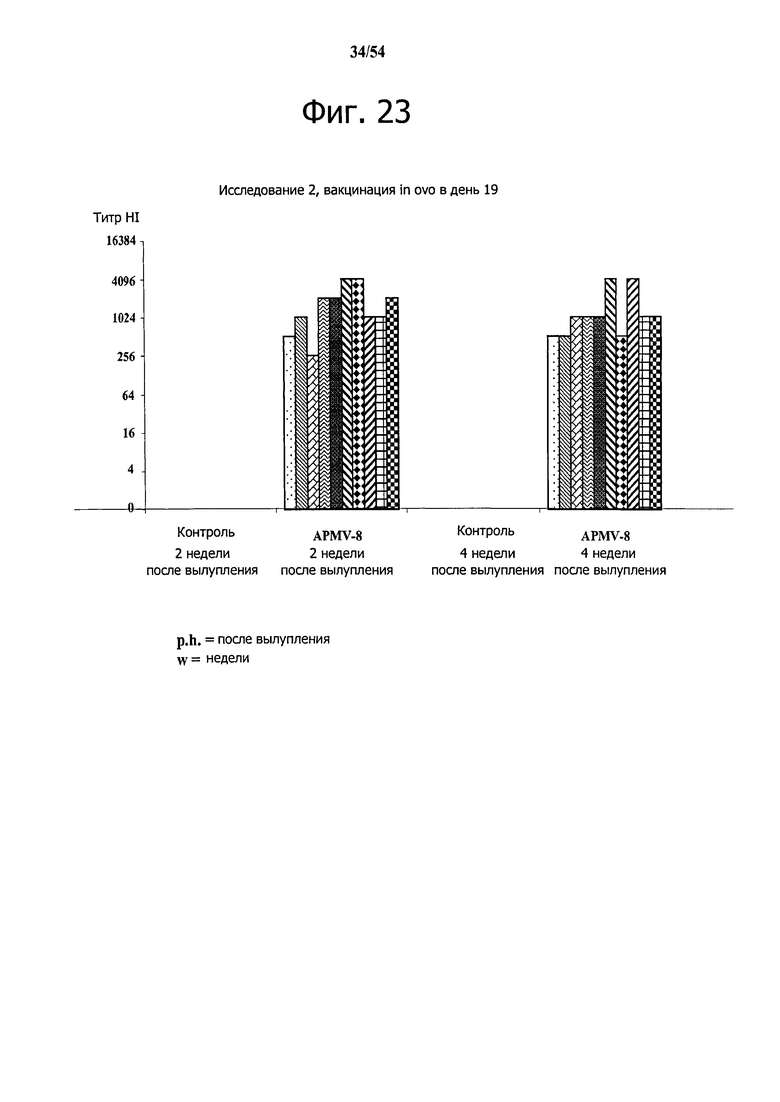






































































































































































































































Настоящее изобретение относится к области генной инженерии и касается способа получения рекомбинантного вирусного вектора парамиксовируса птиц - 8 (APMV-8). Способ включает введение в несущественный участок генома APMV-8 гетерологичной последовательности ДНК, где вирусный вектор APMV-8 включает полинуклеотид с последовательностью SEQ ID NO: 1 или полинуклеотид, комплементарный полинуклеотиду с последовательностью SEQ ID NO: 1, причем несущественный участок выбирается из группы, включающей нетранслируемый участок, расположенный перед открытой рамкой считывания NP, межгенные участки между двумя открытыми рамками считывания генома APMV-8 и нетранслируемый участок, расположенный после открытой рамки считывания L. Представленное изобретение может быть использовано в качестве вакцины для животных. 3 з.п. ф-лы, 35 ил., 9 табл., 7 пр.
1. Способ получения рекомбинантного вирусного вектора APMV-8, который включает введение в несущественный участок генома APMV-8 гетерологичной последовательности ДНК, где вирусный вектор APMV-8 включает полинуклеотид с последовательностью SEQ ID NO: 1 или полинуклеотид, комплементарный полинуклеотиду с последовательностью SEQ ID NO: 1.
2. Способ по п. 1, где несущественный участок выбирается из группы, включающей нетранслируемый участок, расположенный перед открытой рамкой считывания NP, межгенные участки между двумя открытыми рамками считывания генома APMV-8 и нетранслируемый участок, расположенный после открытой рамки считывания L.
3. Способ по п. 1 или 2, который включает стадии:
а) получение экспрессионных плазмид, экспрессирующих гены NP, Р и L;
б) получение транскрипционной плазмиды, включающей гетерологичную последовательность ДНК в несущественном участке полноразмерного генома APMV-8;
в) трансфекцию экспрессионных плазмид и транскрипционной плазмиды в клетку-хозяина;
г) выделение/получение инфекционного вируса APMV-8 из клетки-хозяина.
4. Способ по п. 1 или 2, который включает стадии:
а) получение экспрессионных плазмид, экспрессирующих гены NP, Р и L;
б) получение транскрипционной плазмиды, включающей гетерологичную последовательность ДНК в несущественном участке полноразмерного генома APMV-8;
в) получение экспрессионной плазмиды, экспрессирующей полимеразу Т7;
г) трансфекцию экспрессионных плазмид и транскрипционной плазмиды в клетку-хозяина;
д) восстановление/получение инфекционного вируса APMV-8 из клетки-хозяина.
| WO 2009101149 A2, 20.08.2009 | |||
| PALDURAI A., at al., Complete genome sequences of avian paramyxovirus type 8 strains goose/Delaware/1053/76 and pintail/Wakuya/20/78, VIRUS RESEARCH, 25.03.2009, Vol.142, no | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Аппарат для электрической передачи изображений без проводов | 1920 |
|
SU144A1 |
| СУХАЯ ЖИВАЯ ВАКЦИНА ПРОТИВ НЬЮКАСЛСКОЙ БОЛЕЗНИ И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2000 |
|
RU2162341C1 |
| US 0007547442 B2, 16.06.2009 | |||
| US 20090175826 A1, 09.07.2009 | |||
| US 20090041852 A1, 12.02.2009 | |||
| EP 0000974659 A3, 26.01.2000. | |||

