ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Изобретение относится к ферментной технологии и, более точно, к целлюлазным белкам слияния, содержащим каталитический домен и целлюлозосвязывающий домен. Белки слияния могут быть получены с помощью рекомбинантной технологии при использовании полинуклеотидов, экспрессирующих векторов и клеток-хозяев, которые также входят в объем изобретения. Белки слияния и ферментные препараты на их основе полезны для обработки целлюлозного материала, например текстильного материала. Кроме того, белки слияния могут использоваться для получения целлюлозной массы и в бумажной промышленности, для экстрагирования масел из растений, приготовления детергентных композиций или для улучшения качества кормов для животных. Изобретение также относится к способу обработки целлюлозного материала белками слияния, в частности, к способу биологической абразивной обработки (биостоунингу) или окончательной биологической отделки (биофинишингу) тканей или одежды, в особенности денима. Изобретение также относится к детергентным композициям и кормам для животных, содержащим белки слияния,
УРОВЕНЬ ТЕХНИКИ
Целлюлоза представляет собой основной структурный компонент высших растений и существует в природе в практически очищенной форме только в волокнах хлопка. Она придает растительным клеткам высокую прочность при растяжении, обеспечивая их устойчивость к механическим стрессам и осмотическому давлению. Целлюлоза представляет собой линейный полисахарид из остатков глюкозы, соединенных β-1,4-связями. В природе целлюлоза обычно ассоциирована с лигнином и гемицеллюлозой. Целлюлозный материал разлагается в природе под действием различных организмов, включая бактерии и грибы. Биологическая конверсия целлюлозы в глюкозу обычно осуществляется тремя основными группами ферментов: целлобиогидралазами (СВН), эндоглюканазами (EG) и бета-глюкозидазами (BG).
Целлюлазы имеют широкое промышленное применение. В текстильной промышленности целлюлазы используются для окончательной отделки денима с целью придания одежде из денима модного потертого вида, который обычно обеспечивается при стирке денима с камнями пемзы. Кроме того, целлюлазы используются, например, для ликвидации мшистости тканей и предотвращения образования узелков на поверхности одежды из хлопка. В качестве детергентов целлюлазы используются для осветления цветов и предотвращения посерения и скатывания одежды. Целлюлазы также применяются в пищевой промышленности и в производстве кормов для животных и имеют большой потенциал в производстве целлюлозной массы и в бумажной промышленности, например, в удалении чернил с волокнистых поверхностей и улучшении дренажа целлюлозной массы.
Промышленно используемые целлюлазы часто представляют собой смеси ферментов с различной активностью и субстратной специфичностью. Коммерческие ферментные препараты часто обладают активностью всех трех целлюлаз: СВН, EG и BG. Кроме того, уникальные свойства каждой целлюлазы более подходят для одних целей, нежели других, и поэтому предпринимались определенные попытки создания и использования целлюлаз, имеющих только нужную активность. Наиболее широко используемые целлюлазы получают из грибов Trichoderma reesei. Однако, для получения целлюлаз могут использоваться и другие грибы (см., например, US 5457046).
Целлюлазы, используемые для обработки денима, обычно делятся на две основные группы: кислые и нейтральные. Кислые целлюлазы обычно действуют при рН от 4, 0 до 5, 5, а нейтральные целлюлазы действуют при рН от 6 до 8. Целлюлазы, имеющие свойства как кислых, так и нейтральных целлюлаз, можно назвать гибридными целлюлазами. Кислые целлюлазы, применяемые главным образом для биостоунинга, получают из Trichoderma reesei (половая форма Hypocrea jecorina), а нейтральные целлюлазы получают из различных грибов, включая Melanocarpus, Myceliophthora, Fusarium, Acremonium и Chrysosporium (Haakana et al. 2004). Ферменты, получаемые из Т. reesei, включают, например, целлюлазы семейства гликозилгидролаз 5 (эндоклюканазу II, EG II), семейства 7 (целлобиогидролазу I, CBHI) и семейства 12 (эндоклюканазу III, EGIII; Ward et al. 1992), и нейтральные целлюлазы, большей частью эндоглюканазы семейства 45 и семейства 7 (Henrissat, 1991; Henrissat и Bairoch, 1993, 1996).
В патенте US 5874293 раскрываются улучшенная целлюлазная композиция, содержащая повышенные количества эндоклюканазы EG II из Т. reesei, для обработки содержащих целлюлозу тканей. Композиция улучшает цветовые свойства тканей, увеличивает их легкость и внешний вид, а также снижает образование узелков на поверхности тканей. В заявке WO 97/14804 описаны целлюлазы с мол. массой 20 кДа и 50 кДа с эндоглюканазной активностью, полученные из Melanocarpus sp., которые особенно эффективны в текстильной промышленности и в производстве детергентов. Предполагается, что для создания новых ферментных свойств, будут использоваться белки слияния, содержащие целлюлазы с мол. массой 20 кДа и 50 кДа, соединенные с целлюлозосвязывающим доменом, полученные из Trichoderma reesi. Однако в WO 97/14804 не приведено никаких специальных примеров получения таких белков и не описаны их свойства.
Все еще существует необходимость получения целлюлаз с улучшенными свойствами, включая эндоглюканазы, которые наиболее эффективны для обработки тканей и для применения в тех областях, где обычно используются целлюлазы. В частности, существует постоянная потребность в более эффективных целлюлазах для улучшения экономических процессов. Настоящее изобретение служит для удовлетворения этих потребностей.
В текстильной промышленности в последние годы большой интерес со стороны производителей денима вызывает технология создания внешнего вида ткани, называемая «стиркой с камнями пемзы» (stone washing) или «вывариванием» ткани. Традиционная стирка с камнями пемзы уменьшает прочность ткани и повышает нагрузку на стиральные машины. В окончательной обработке денима все больше используются ферменты, и целлюлазы заменяют камни пемзы или используются вместе с ними для придания тканям желаемого «потертого» вида. Контролируемая обработка ферментами приводит к меньшему повреждению одежды и машинного оборудования и устраняет необходимость использования камней.
Основная проблема, связанная с применением ферментной абразивной стирки, заключается в восстановлении окраски ткани за счет перераспределения удаленного красителя Индиго во время или после биостоунинга. Перераспределение красителя Индиго снижает нужный контраст между белыми и окрашенными нитями, что может быть более легко замечено на обратной стороне денима и внутренней поверхности карманов (как увеличение голубизны). На лицевой стороне это может выглядеть как уменьшенный контраст между окрашенными участками и участками, из которых краситель был удален в процессе биостоунинга. Восстановление окраски может быть снижено использованием агентов, препятствующих такому восстановлению, например, неионных этоксилированных спиртов во время обработки ткани или добавлением отбеливающих агентов на стадиях отполаскивания. Природа фермента оказывает влияние на восстановление окраски. В целом, нейтральные целлюлазы способствуют восстановлению окраски в меньшей степени, чем кислые целлюлазы.
В заявке WO 97/09410 указано, что добавление определенного типа целлюлазы к другим целлюлазам, способствующим «истиранию» ткани, снижает восстановление окраски. Дополнительная целлюлаза относится к семейству 5 или 7, но сама по себе не имеет значительной «истирающей» активности. Предпочтительно, указанная дополнительная целлюлаза происходит из Bacillus или Clostridium.
В патенте US 5916799 описана целлюлазная композиция, содержащая целлобиогидролазы и эндоглюканазы, которые используются для ограничения протеолиза за счет разделения ядра и связывающих доменов ферментов. Полученные ферментные композиции, как обнаружено, уменьшают восстановление окраски. В заявке WO 94/07983 отмечается, что перераспределение красителя в ткани во время процесса биостоунинга может быть уменьшено при использовании композиции целлюлаз из грибов, которая по существу свободна от целлобиогидролазного компонента. В заявке WO 96/23928 описана обработка тканей, содержащих целлюлозу, усеченными целлюлазами. Такие усеченные ферменты, не имеющие целлюлозосвязывающего домена (CBD), как обнаружено, уменьшают перераспределение красителя и повышают «истирание».
Главный вывод, который можно сделать при анализе приведенного ссылочного материала, заключается в том, что целлюлозосвязывающий домен способствует восстановлению окраски. Таим образом, настоящее изобретение обеспечивает целлюлазу, обладающую низкой активностью в отношении восстановления окраски, несмотря на присутствие целлюлозосвязывающего домена.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение базируется на неожиданном обнаружении того факта, что эффект эндоглюканазы может быть значительно усилен при соединении ее с определенным целлюлозосвязывающим доменом без влияния на активность в отношении восстановления окраски.
Изобретение относится к новым белкам слияния эндоглюканазы, имеющим улучшенные гидролазные свойства, для применения в текстильной индустрии, особенно в производстве денима с помощью биостоунинга. Заявленные белки слияния имеют то преимущество, что активны как при кислых, так и при нейтральных значениях рН, и поэтому более эффективны при использовании в целях биошлифования. При обработке денима белки слияния обеспечивают низкий эффект восстановления окраски. Использование более эффективных эндоглюканаз в соответствии с изобретением является экономичным. Дополнительные преимущества заключаются в том, что облегчаются перевозка и хранение ферментных продуктов, поскольку они используются в меньших количествах. Белки слияния также полезны для применения в детергентных композициях и в других областях, например, в пищевой промышленности, экстрагировании масел из растений или в производстве целлюлозной массы и бумаги.
Объектом изобретения также являются полинуклеотиды, кодирующие новые белки слияния эндоглюканазы.
Объектом изобретения также является способ получения белков слияния.
Еще один объект изобретения относится к новым экспрессирующим векторам, содержащим указанные полинуклеотиды, полезным для получения белков слияния эндоглюканазы, и новым хозяевам, трансформированным такими экспрессирующими векторами.
Еще один объект изобретения относится к ферментным композициям, содержащим один или более белков слияния эндоглюканазы.
Объектом изобретения являются также способы обработки целлюлозного материала белком слияния, например, для использования в текстильной промышленности, для приготовления детергентных композиций, кормов для животных, для экстрагирования масел из растений, для производства целлюлозной массы и бумаги и, в частности, для окончательной обработки тканей, в особенности, для биостоунинга и биофинишинга денима.
Еще один объект изобретения относится к корму для животных и детергентным композициям, содержащим белки слияния.
Изобретение также относится к целлюлазному белку слияния, содержащему первую аминокислотную последовательность эндоглюканазного ядра и вторую аминокислотную последовательность, содержащую линкер и целлюлозосвязывающий домен (CBD), имеющую по крайней мере 75%-ную идентичность с SEQ ID NO:15, или варианту, или фрагменту указанного белка, имеющему целлюлозосвязывающую активность.
Изобретение также относится к выделенному полинуклеотиду, выбранному из группы, включающей:
а) нуклеотидную последовательность SEQ ID NO:3 или 5, или последовательность, кодирующую белок слияния целлюлазы по п.1;
б) последовательность, комплементарную, указанной в а);
в) последовательность, полученную в соответствии с вырожденностью генетического кода, на основе последовательностей а) и б).
Изобретение также относится к экспрессирующему вектору, содержащему указанную нуклеотидную последовательность, и клетке-хозяину, содержащей экспрессирующий вектор, и к ферментным препаратам, содержащим белок слияния.
Изобретение также относится к способу получения белка слияния, включающему трансформацию клетки-хозяина экспрессирующим вектором, кодирующим указанный белок слияния, и культивирование указанной клетки-хозяина в условиях, обеспечивающих экспрессию указанного белка слияния, и, при необходимости, выделение и очистку белка слияния.
Изобретение также относится к способам обработки целлюлозного материала, предусматривающим обеспечение контактирования целлюлозного материала с белком слияния.
Изобретение также относится к способам биологической абразивной обработки тканей, включающим стадию обеспечения контактирования целлюлазного белка слияния или ферментного препарата с тканью денима или одеждой из денима, и к способам окончательной биологической обработки тканей, включающим стадию обеспечения контактирования целлюлазного белка слияния или ферментного препарата с текстильными материалами такими, как ткань, или одежда, или нити.
Изобретение также относится к детергентным композициям, содержащим белок слияния и детергентные добавки, корму для животных, содержащему белок слияния, и штамму Escherichia coli, депонированному под номером Е.coli DSM 18159.
Специфические воплощения изобретения нашли свое отражение в формуле изобретения.
Другие объекты, детали и преимущества изобретения станут ясными из представленных иллюстраций, подробного описания изобретения и примеров.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
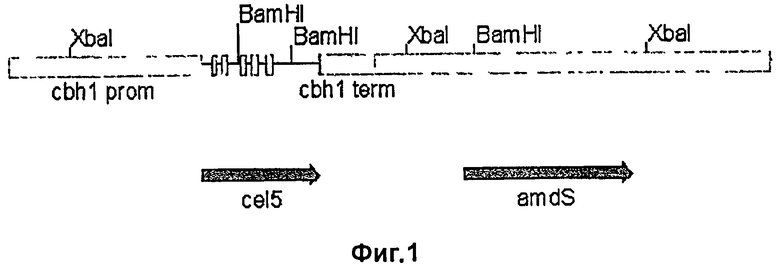
На Фиг.1 приведена схема экспрессионной кассеты, используемой для трансформации протопластов Trichoderma reesi для получения целлюлазы EG28 или EG28+CtCBD Thermoascus aurantiacus. Ген cel5A или cel5A_Ctcel7A линкер CBD находится под контролем промотора cbhI Trichoderma reesi (cbhI prom) и терминация транскрипции обеспечивается добавлением терминатора (cbhI term). В кассету встроен ген amdS (amdS) для селекции трансформантов. Экспрессионную кассету для получения EG28 и EG28+CICBD выделяют в виде фрагмента NotI размером 8,6 kb из pALK1930 или в виде фрагмента NotI размером 8,9 kb из pALK1948, соответственно.
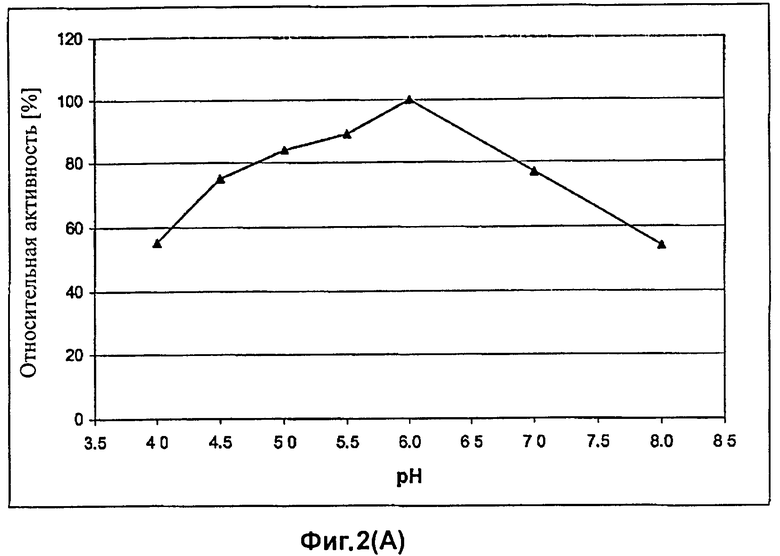
На Фиг.2 показана зависимость активности гетерологично продуцируемой целлюлазы EG28 T. aurantiacus, выделенной из культурального супернатанта, от величины рН при использовании CMC в качестве субстрата в течение 10 мин при 150°C (A). Температурный оптимум целлюлазы EG28 определяют при оптимальном рН (6,0). Реакционную смесь, содержащую CMC в качестве субстрата, тестируют в течение 10 мин (B). Температурный оптимум и оптимум рН слитого белка EG28+CtCBD совпадают с таковыми целлюлазы EG28 дикого типа.
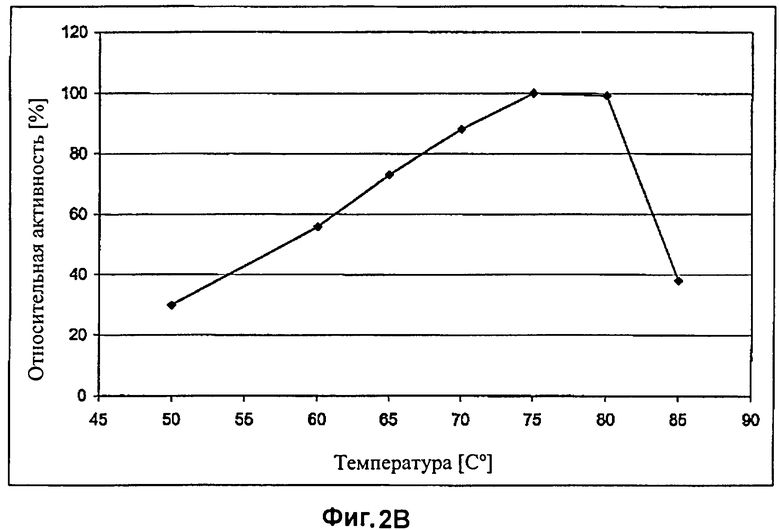
На Фиг.3 показан эффект абразивной обработки целлюлазой EG28, определяемый путем измерения интенсивности цвета, при различных температурах и рН 6.
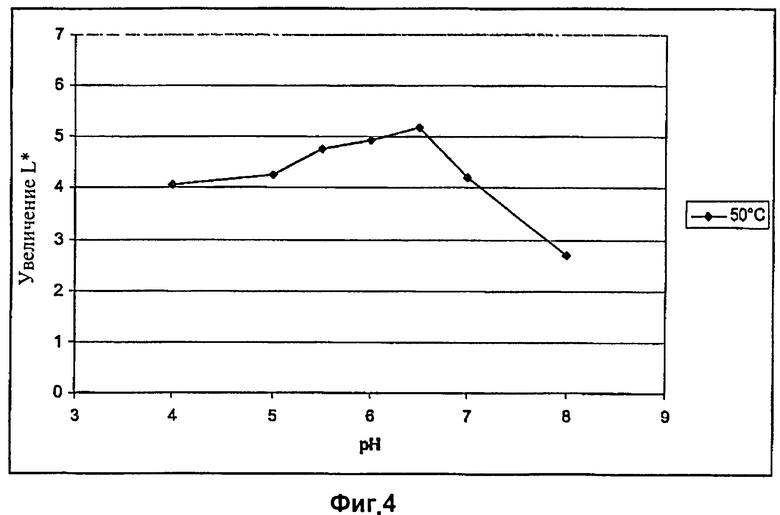
На Фиг.4 показан эффект абразивной обработки целлюлазой EG28+CtCBD, определяемый путем измерения интенсивности цвета, при различных рН и температуре 50°C.
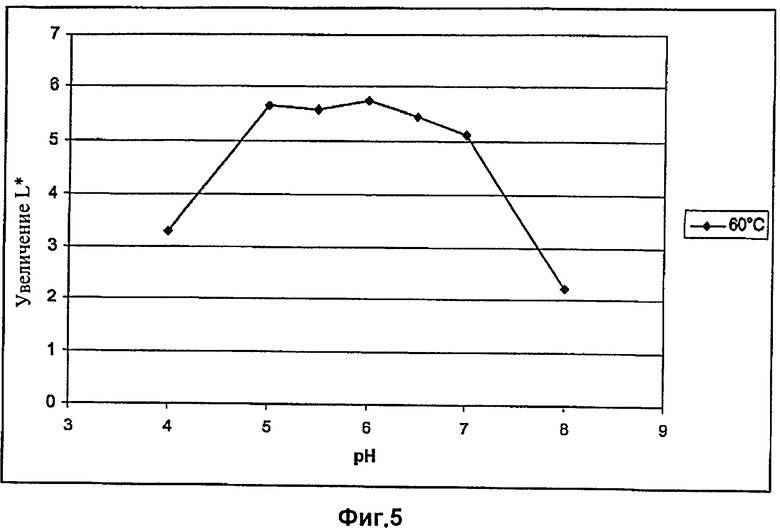
На Фиг.5 показан эффект абразивной обработки целлюлазой EG28+CtCBD, определяемый путем измерения интенсивности цвета, при различных рН и температуре 60°C.
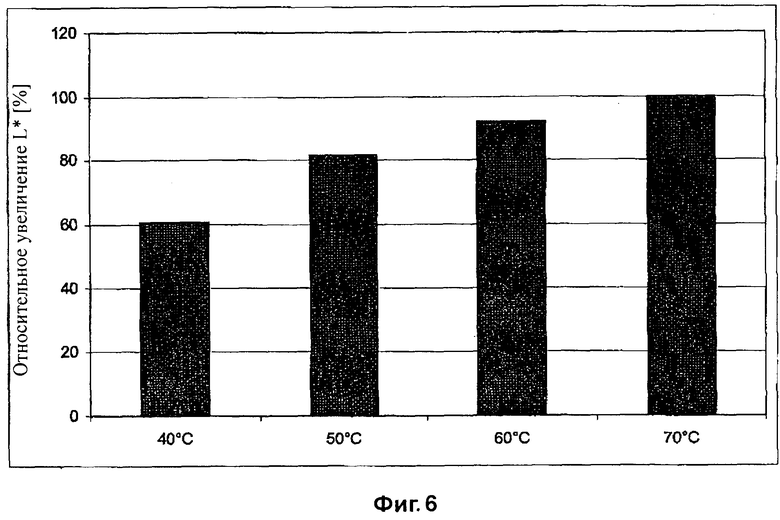
На Фиг.6 показан эффект абразивной обработки целлюлазой EG28+CtCBD, определяемый путем измерения интенсивности цвета, при различных температурах и рН 6.
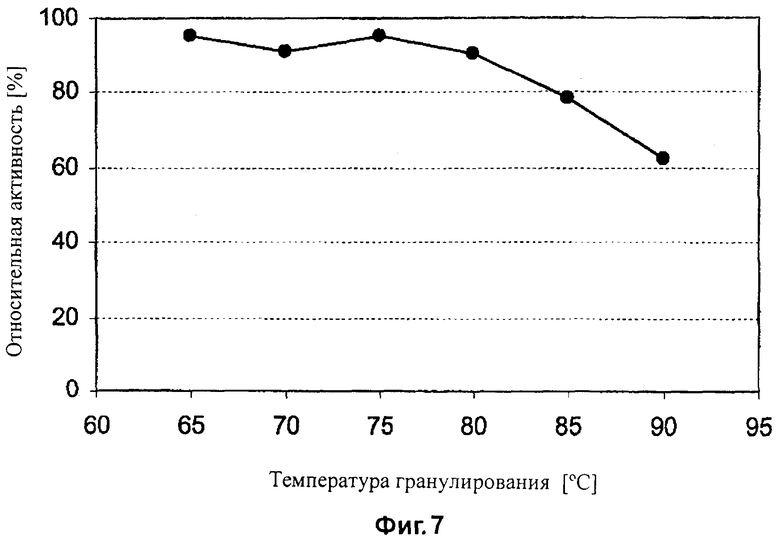
На Фиг.7 показано влияние температуры гранулирования на бета-глюканазную активность, определяемую в корме, в который была добавлена целлюлаза EG28.
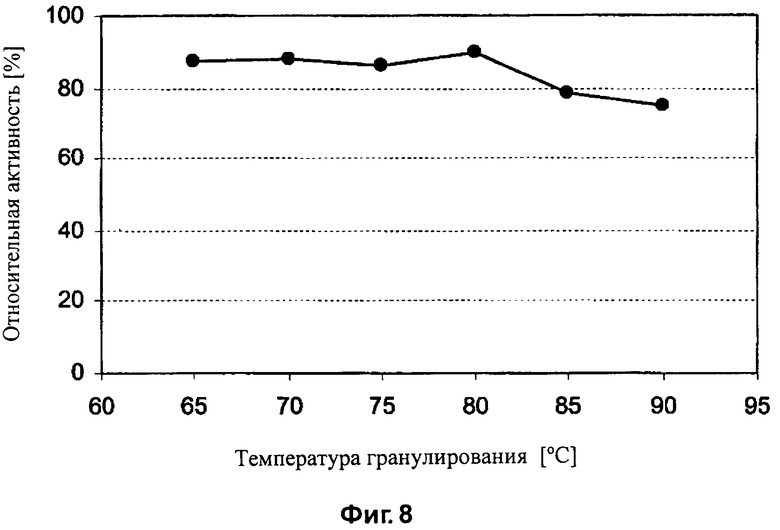
На Фиг.8 показано влияние температуры гранулирования на бета-глюканазную активность, определяемую в корме, в который была добавлена целлюлаза EG28+CtCBD.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Целлюлозы, содержащие каталитический домен/ядро (CD) обнаруживают целлюлазную активность. Кроме того, каталитический домен в молекуле целлюлазы может включать один или более «целлюлозосвязывающих доменов» (CBDs), также называемых «углеводосвязывающими доменами/модулями» (CBD/CBM), которые могут быть локализованы на N- или C-концевых участках каталитического домена CBDs имеют углеводосвязывающую активность и способны опосредовать связывание целлюлазы с кристаллической целлюлозой, но не обладают гидролитической активностью или имеют очень низкую гидролитическую активность на растворимых субстратах. Два указанные выше домена обычно соединены посредством гибкого и высоко гликозилированного линкерного участка, здесь называемого линкером. Такие конструкции описаны, например, Srisodsuk et al., 1993. Некоторые из природных эндоглюканаз и целлобиогидролаз имеют целлюлозосвязывающий домен (CBD), в то время как другие этого домена не имеют.
Эндоглюканазы (EGs) являются одним из трех типов целлюлаз, обычно необходимых для биологического разложения целлюлозы до глюкозы. Эндоглюканазы расщепляют внутренние бета-1,4-глюкозидные цепи, в то время как целлобиогидролазы отщепляют дисахарид целлобиозу от конца целлюлозной полимерной цепи, а бета-1,4-глюкозидазы гидролизуют целлобиозу и другие короткие целлоолигосахариды до глюкозы. Эндоглюканаза (EG) в соответствии с настоящим изобретением классифицируется как Е.С.3.2.1.4. Она представляет собой бета-D-глюкан-4-глюканогидролазу и катализирует эндогликолиз 1,4-бета-D-гликозидных связей в полимерах глюкозы таких, как целлюлоза. Некоторые эндоглюканазы способны гидролизовать, например, и 1,4-связи в бета-D-глюканах, также содержащих 1,3-связи. Они могут быть также классифицированы как эндо-1,3(4)-бета-глюканазы (Е.С.3.2.1.6). Таким образом, фермент может катализировать реакции на различных субстратах и принадлежать к различным классам.
Целлюлазы, включая эндоглюканазы, могут быть также классифицированы по принадлежности к различным семействам гликозилгидролаз в соответствии с первичной последовательностью, что подтверждается анализом трехмерной структуры некоторых членов семейства (Henrissat 1991, Henrissat и Bairoch, 1993, 1996). Например, семейство 45 (ранее celK) содержит эндоглюканазы (Е.С.3.2.1.4) и семейство 5 (ранее celA) содержит главным образом эндоглюканазы (Е.С.3.2.1.4). Семейство 7 (ранее семейство целлюлаз celC) содержит как эндоглюканазы, так и целлобиогидролазы. Некоторые гликозилгидролазы являются многофункциональными ферментами, содержащими каталитические домены, принадлежащие к различным семействам гликозилгидролаз. Для целей настоящего изобретения эндоглюканазная часть белка слияния, предпочтительно, принадлежит гликозилгидролазному семейству 45 или семейству 5 и, более предпочтительно, семейству 5.
В соответствии с настоящим изобретением эндоглюканазная часть белка слияния получена из грибов, предпочтительно, рода Thermoascus, и, более предпочтительно, Thermoascus aurantiacus. Такая эндоглюканаза описана, например, Hong et al., 2003. В заявке WO 03/062409 предполагается использование этого фермента в пищевых целях, поскольку помимо эндоглюканазной он также имеет бета-глюканазную активность. Более предпочтительно, эндоглюканаза происходит из штамма ALKO4242 Thermoascus aurantiacus, депонированного в CBS под номером 116239. Эндоглюканазный ген указанного штамма встроен в плазмиду pALK1926, которая депонирована в DSM под номером 17326. Белок, кодируемый данным геном, обозначен как «Та EG28» или просто «EG28».
Альтернативно, эндоглюканазная часть белка слияния может быть получена из Acremonium sp. предпочтительно, A. thermophilum, и, более предпочтительно, из штамма ALKO4245, депонированного в CBS под номером 116240. Эндоглюканаза, получаемая из этого штамма и кодируемая геном cel45A, обозначена как «At EG40» или просто «EG40».
Используемый здесь термин «эндоглюканазное ядро» означает каталитический домен/ядро (CD) фермента, обладающего по крайней мере эндоглюканазной активностью. Такой каталитический домен может быть представлен природной (интактной) формой или же может быть модифицирован.
В соответствии с одним вариантом воплощения изобретение относится к эндоглюканазному ядру, имеющему, по крайней мере, 75, 80, 85, 90, 95, 98 или 99% идентичности с SEQ ID NO:2 (Та EG28). Предпочтительно, ядро содержит, по крайней мере, зрелый белок, который соответствует аминокислотам от 19 до 334 SEQ ID NO:2. Сигнальная последовательность предсказана с помощью программы SignalP V3.0 (Nielsen et al., 1997; Bendtsen et al., 2004); значение NN получено при использовании нейральных сетей и значение НММ при использовании скрытых моделей Маркова. Альтернативно, эндоглюканазная часть белка слияния имеет, по крайней мере, 75, 80, 85, 90, 95, 98 или 99%-ную идентичность с эндоглюканазным ядром At EG40, кодируемым полинуклеотидом, содержащим SEQ ID NO:8. Предпочтительно, ядро содержит, по крайней мере, зрелый белок, который соответствует аминокислотам от 22 до 297 EG40.
Используемый здесь термин «целлобиогидролаза» или «СВН» относится к ферментам, которые отщепляют целлюлозу от конца глюкозной цепи и продуцируют целлобиозу. Их также называют 1,4-бета-D-глюкан целлюбиогидролазами или целлюлозе 1,4-бета-целлобиозидазами. Указанные ферменты гидролизуют 1,4-бета-D-глюкозидные связи на восстанавливающем или невосстанавливающем концах полимера, содержащего такие связи, например, целлюлозы, с образованием целлобиозы.
CBD, включая линкер, предпочтительно получают из Chaetomium thermophilum и, в особенности, из целлобиогидролазы (CBHI/Cel7A), кодируемой геном из штамма ALKO4265, депонированного в CBS под номером 730.95. Этот CBD, включая линкер, обозначают как «CtCBD». Согласно предпочтительному варианту осуществления изобретения линкер, соединенный с целлюлозосвязывающим доменом, имеет последовательность, по крайней мере, на 80, 85, 90, 95, 98 или 99% идентичную SEQ ID NO:15 (которая соответствует аминокислотам 335-415 SEQ ID NO:4). Согласно другому предпочтительному варианту осуществления изобретения вторая аминокислотная последовательность содержит аминокислоты 335-379 SEQ ID NO:4.
Термин «происходящий из» применительно к микроорганизму означает, что полипептид может естественным образом продуцироваться указанным специфическим микроорганизмом или же полинуклеотид, кодирующий полипептид, может быть выделен из данного микроорганизма. Данный термин также относится к клетке-хозяину, в которую встроен полинуклеотид из указанного микроорганизма, кодирующий полипептид. Однако, при этом не исключаются незначительные модификации последовательности, например, замещения, делении, вставки и/или инверсии нескольких аминокислот/кодонов так, что сохраняется биологическая активность кодируемого полипептида.
Ядро и линкер + CBD, соответственно, могут быть фрагментом или вариантом указанных последовательностей, причем указанный фрагмент или вариант имеют целлюлазную активность и/или целлюлозосвязывающую активность. Например, первая аминокислотная последовательность может содержать фрагмент или вариант аминокислотной последовательности, имеющей, по крайней мере, 75%-ную идентичность с SEQ ID NO:2 или 8, а вторая аминокислотная последовательность может содержать фрагмент или вариант аминокислотной последовательности, имеющей, по крайней мере, 75%-ную идентичность с SEQ ID NO:15.
Используемое здесь понятие «целлюлазная активность» означает каталитическую способность гидролизовать целлюлозу или ее производные, например, эндоглюконазную или бета-глюканазную активность. Кроме того, эндоглюканазная и/или бета-глюканазная активность некоторых целлюлаз может дополняться гемицеллюлазной и/или ксиланазной активностью.
Используемый здесь термин «идентичность» относится к глобальной идентичности между аминокислотными последовательностями при их сравнении друг с другом, начиная с первой аминокислоты, кодируемой соответствующим геном, и заканчивая последней аминокислотой. Идентичность полноразмерных последовательностей измеряют с использованием глобальной программы выравнивания Needleman-Wunsch при обеспечении EMBOSS (European Molecular Biology Open Software Suite; Rice et al., 2000), версия 3.0.0 со следующими параметрами: EMBLOSUM62, штрафной пробел 10.0, штраф протяженности 0.5. Алгоритм описан Needleman-Wunsch (1970). Специалисту ясно, что результаты, полученные при использовании алгоритма Needleman-Wunsch, являются сравнимыми только при выравнивании соответствующих доменов последовательности. Таким образом, сравнение, например, целлюлазных последовательностей, включающих CBD или сигнальные последовательности, с последовательностями, не содержащими указанных элементов, не является достоверным.
В соответствии с одним вариантом осуществления изобретения белок слияния содержит эндоглюканазное ядро, кодируемое геном, эквивалентным гену, встроенному в Е.Coli DSM 17326. Предпочтительно, белок слияния кодируется слитым геном, эквивалентным гену, встроенному в Е.Coli DSM 18159. «Эквивалентность» означает здесь практическое сходство или сходство. В соответствии со специфическим вариантом осуществления изобретения белок слияния содержит эндоглюканазное ядро, включающее последовательность SEQ ID NO:2 и линкер и CBD, включающие SEQ ID NO:15. В особенности, ядро и линкер + CBD включают последовательность SEQ ID NO:4 или 6 или вариант или фрагмент указанных последовательностей, обладающих целлюлазной и целлюлозосвязывающей активностью.
Используемый здесь термин «фрагмент» относится к части специфической аминокислотной последовательности, который имеет длину, достаточную для сохранения заданной биологической активности. Другими словами, фрагмент может быть, например, только зрелой частью аминокислотной последовательности или даже фрагментом зрелой части. Вариант специфической аминокислотной последовательности относится к аминокислотной последовательности, которая не идентична специфической аминокислотной последовательности, а содержит по крайней мере некоторые аминокислотные изменения, т.е. делеции, замещения, инверсии, вставки и т.д., которые не имеют существенного влияния на биологическую активность белка по сравнению с активностью специфической аминокислотной последовательности при использовании в нужных целях. Биологическая активность в контексте настоящего изобретения относится к целлюлазной активности, способности связывать целлюлозу или обеим указанным активностям.
Белок слияния, согласно заявленному изобретению, может быть получен путем прикрепления эндоглюканазного ядра к линкеру и участку CBD в подходящей кодирующей ДНК при использовании хорошо известной технологии рекомбинантных ДНК для продуцирования нужного рекомбинантного белка. Коротко говоря, полинуклеотиды, кодирующие участки слияния, амплифицируют и клонируют; нуклеотиды могут быть синтезированы. Слитый полинуклеотид встраивают в вектор экспрессии, вектором трансформируют клетку-хозяин и экспрессируют белок. Предпочтительно, линкер и CBD прикрепляют к C-концу эндоглюканазного ядра.
Вектор экспрессии представляет собой клонирующую плазмиду или вектор, способный экспрессировать ДНК, кодирующую эндоглюканазные белки слияния, после введения вектора в подходящую клетку-хозяин. При использовании грибов в качестве хозяев представляющий интерес ген предпочтительно вводят в грибную клетку-хозяин в составе клонирующего или экспрессирующего вектора, который интегрируется в грибную хромосому или обеспечивает интеграцию представляющего интерес гена в хромосому хозяина. Другие последовательности, которые являются частью клонирующего или экспрессирующего вектора, также могут быть интегрированы вместе с указанной ДНК в процессе интеграции. Кроме того, в клетках грибов экспрессирующий вектор или его части могут быть направлены в предопределенный локус. Альтернативно, нужный ген слияния может быть представлен в виде автономно реплицирующейся плазмиды.
ДНК, кодирующую эндоглюканазные белки слияния, предпочтительно помещают под контроль (т.е. функционально связывают) с определенными контролирующими последовательностями, такими как промоторные последовательности, в составе вектора. Во время трансформации указанные контролирующие последовательности интегрируют в геном хозяина вместе с представляющим интерес геном. Альтернативно, контролирующие последовательности могут быть встроены в сайт интеграции.
Контролирующие экспрессию последовательности в составе экспрессирующего вектора варьируют в зависимости от того, сконструирован ли вектор для экспрессии определенного гена в прокариотическом или эукариотическом хозяине (например, вектор-переносчик может содержать ген для селекции в бактериальном хозяине). Последовательности, контролирующие экспрессию, могут содержать регуляторные элементы транскрипции, такие как промоторы, энхансеры, и последовательности терминации транскрипции и/или регуляторные элементы трансляции, такие как сайты инициации трансляции и терминации.
Полинуклеотидная молекула, такая как ДНК, способна экспрессировать полипептид, если она содержит последовательности, контролирующие экспрессию, включающие информацию о регулировании транскрипции, и функционально связаны с нуклеотидной последовательностью, кодирующей полипептид.
Функциональная связь представляет собой связь, в которой последовательность соединена с регуляторной последовательностью (или последовательностями) таким образом, что экспрессирующая последовательность находится в таком участке, где подвергается влиянию или контролю со стороны регуляторной последовательности. Две ДНК-последовательности (такие, как последовательность промоторного участка, связанная с 5'-концом последовательности, кодирующей белок) считаются связанными функциональным образом, если промотор оказывает влияние на транскрипцию.
Векторы в соответствии с настоящим изобретением могут содержать и другие функционально связанные регуляторные элементы такие, как энхансерные последовательности.
В предпочтительном варианте осуществления изобретения конструируют генетически стабильные трансформанты таким образом, что ДНК, кодирующую белки слияния, встраивают в хромосому хозяина путем трансформации с помощью вектора, который может содержать последовательности, обеспечивающие интеграцию указанного вектора в хромосому.
Клетки, в хромосомы которых стабильно интегрирована ДНК, кодирующая эндоглюканазные белки слияния, могут быть отобраны, например, путем включения в них маркера (маркеров), гомологичного или гетерологичного, который обеспечивает селекцию клеток-хозяев, содержащих вектор экспрессии в составе хромосомы. Маркер может обеспечивать устойчивость к биоцидам, например, устойчивость к антибиотикам или тяжелым металлам, таким, как медь. Кроме того, маркеры могут комплементировать ауксотрофную мутацию в хромосоме хозяина и т.д. Селектируемый маркер может представлять собой, например, селектируемый ген, непосредственно связанный с последовательностью ДНК, которая является подлежащим экспрессии геном, или же он может быть введен в ту же самую клетку путем совместной трансформации. Могут использоваться и другие системы селекции.
После конструирования вектор экспрессии, содержащий ДНК, кодирующую белок слияния, его вводят в подходящую клетку-хозяин любым пригодным способом, включая трансформацию, известную из уровня техники. После трансформации реципиентные клетки выращивают в подходящей селективной среде, которая отбирает трансформированные клетки по их росту.
Подходящими хозяйскими системами для экспрессии и продукции белка являются, например, системы продукции на основе грибов Trichoderma (ЕР 244 234) или Aspergillus такие, как A. oryzae или A. niger (WO 97/08325, WO 95/33386, патент США 5843745, патент США 5770418), или системы продукции на основе грибов Fusarium такие, как F. oxysporum (Malardier et al., 1989). Подходящие системы продукции на основе бактерий включают системы, основанные на Bacillus, например, В. subtilis, В. licheniformis, В. amyloliquefaciens, или Е.coli, или Streptomyces.). Подходящие системы продукции на основе дрожжей включают системы, основанные на Saccharomyces, Shizosaccharomyces или Pichia pastoris. Могут использоваться и системы продукции на основе других микроорганизмов или клеток млекопитающих или растений.
Экспрессия клонированных генных последовательностей приводит к продукции нужного белка или фрагмента белка. Такая экспрессия может происходить постоянно в трансформированных клетках или контролируемым образом.
Для получения ферментных препаратов согласно заявленному изобретению хозяева, имеющие нужные свойства (хозяева, способные экспрессировать экономически значимые количества эндоглюканазных белков слияния), культивируют в подходящих условиях и нужные ферменты предпочтительно секретируются из организма-хозяина в культуральную среду. Затем белки при необходимости выделяют из указанной культуральной среды известными способами. Предпочтительно, хозяевами для такого продуцирования являются нитчатые грибы такие, как Trichoderma или Aspergillus и, в особенности, Т. reesei.
Используемое здесь понятие «ферментный препарат» относится к любому ферментному продукту, который содержит, по крайней мере, один из заявленных эндоглюканазных белков слияния. Таким образом, ферментный препарат может представлять собой истощенную культуральную среду или фильтрат. Истощенная культуральная среда означает среду, в которой выращивался хозяин, и которая содержит продуцируемые хозяином ферменты. Предпочтительно, клетки-хозяева отделяют от указанной культуральной среды после продуцирования ферментов. При необходимости ферментные препараты могут быть лиофилизированы или же ферментная активность может быть сконцентрирована и/или стабилизирована для сохранения. Если требуется, нужный фермент может быть в дальнейшем очищен в соответствии с известными подходящими методами такими, как экстракция, преципитация, хроматография, аффинная хроматография, электрофорез и другие подобные методы.
Однако, преимущество заявленного изобретения заключается в том, что культуральная среда, содержащая или не содержащая клеток-хозяев, может быть использована в качестве ферментного препарата как таковая без дальнейшей очистки, поскольку эндонуклеазные белки слияния могут секретироваться в культуральную среду и проявлять активность в этой культуральной среде. Ферментные препараты являются чрезвычайно экономичными с точки зрения их получения и использования, поскольку выделение специфического фермента из культуральной среды не является обязательным.
В дополнение к эндоглюканазному белку слияния, ферментные препараты могут содержать один или более других ферментов, которыми могут быть, например, другие целлюлазы, амилазы, липазы, протеазы, гемицеллюлазы, ксиланазы, пектиназы и/или оксидазы такие, как лакказы и пероксидазы. Альтернативно, до, в процессе или после обработки эндоглюканазным белком слияния может осуществляться обработка другим ферментом. Обработка ферментом может включать, например, обработку одной или более амилазой (например, для расшлихтовки денима), или одной или более целлюлазой, или одной или более пероксидазой, или одной или более лактазой. Это зависит от того, какие ферменты входят в ферментный препарат или используются для обработки.
В дополнение к белку слияния, ферментный препарат может содержать различные добавки такие, как стабилизаторы, буферы, консерванты, сурфактанты и/или компоненты культуральной среды. Предпочтительными добавками являются такие добавки, которые обычно используются в ферментных препаратах в определенных целях.
Ферментные препараты могут быть получены в жидкой или твердой форме, например, в форме сухого порошка или гранул, в особенности, в форме устойчивых гранул и стабилизированной жидкости. Следует иметь в виду, что ферментные препараты могут быть дополнительно обогащены, или частично или полностью лишены определенной специфической ферментной активности для того, чтобы отвечать требованиям специфического использования в различных случаях, например, в текстильной промышленности. Смесь ферментов с разной активностью, секретируемая хозяином, может иметь преимущество при определенных промышленных применениях, например, в биостоунинге и биофинишинге.
Эндоглюканазные белки слияния и препараты, содержащие такие белки, используются, например, в для производства текстиля, пищевых и кормовых продуктов, растительных масел, целлюлозосодержащей массы и в бумажной промышленности. Указанные белки могут применяться для обработки любого целлюлозосодержащего материала такого, как текстильный материал, растения, используемые в кормах для животных, растительный материал для экстракции масла, целлюлозосодержащая масса, полученная из древесины путем механической или химической обработки, или вторичные волокна. Они могут также использоваться в детергентах, которые в норме содержат дополнительные компоненты, например, поверхностно-активные вещества, сурфактанты, отбеливатели и/или наполнители. В контексте настоящего изобретения понятие «целлюлозосодержащий материал» относится к любому материалу, содержащему целлюлозу или ее производные в качестве основного компонента. Целлюлозосодержащий материал вводят в контакт с эффективным количеством белка слияния в подходящих условиях, например, при соответствующих рН и температуре, и осуществляют реакцию в течение времени, необходимого для ее протекания.
Слитые ферменты особенно полезны для обработки текстильных материалов, таких как ткани и одежда (детали одежды). Текстильные материалы могут быть произведены из природных целлюлозных волокон или из сделанных человеком целлюлозных волокон или их смесей, или же из смеси синтетических волокон и целлюлозосодержащих волокон. Предпочтительно, целлюлозный материал представляет собой хлопок, в особенности деним. Термин «деним» в контексте настоящего изобретения относится к тканям из денима, в особенности, одежде из денима, в том числе к джинсам. Преимущественным образом, деним представляет собой материал, окрашенный Индиго. Деним также может быть обработан производными Индиго или Индиго вместе с другими красителями, например, деним окрашивают Индиго вместе с серосодержащей протравой.
Эндоглюканазные белки слияния особенно полезны для биостоунинга и биофинишинга.
Стирка с камнями пемзы включает три этапа: расшлихтовку, абразивную обработку и заключительную обработку. Первый этап обычно включает влажную обработку джинсов и сопровождается удалением крахмала и других шлихтующих агентов, обычно применяемых для обработки основы пряжи с целью предотвращения повреждений при плетении. Альфа-амилазы используются для удаления шлихтующих агентов на основе крахмала для улучшения и универсальности процесса влажной обработки. После расшлихтовки джинсы обычно отполаскивают в воде или подвергают непосредственной абразивной обработке.
Второй этап, абразивная обработка, проводится при использовании ферментов или камней пемзы или с обоими указанными средствами. Во всех случаях для удаления красителя необходимо механическое воздействие и обработку обычно осуществляют в стиральных машинах, подобных машинам с барабанами. Термин «состаренная» («вареная») относится к внешнему виду ткани денима, которая прошла обработку целлюлазными ферментами или камнями пемзы или обоими указанными средствами. Синонимами данного термина являются термины «потертый вид» и «вид стирки с камнями». В результате неравномерного удаления красителя существует контраст между окрашенными участками и участками, с которых краситель был удален.
За абразивной обработкой следует третий этап, заключительная обработка, которая включает стадии стирки и отполаскивания, в процессе которых могут использоваться детергенты, оптические осветлители, отбеливающие агенты или размягчители. После обработки ферментами реакцию останавливают для предотвращения повреждения обработанного материала, например, инактивацией температуры и/или рН, во время которой происходит окончательное отмывание и/или удаление детергентов Это означает, что механическая прочность уже не подвергается воздействию ферментов.
Используемый здесь термин «биостоунинг» ткани или одежды означает использование ферментов вместо, или в дополнение, к использованию камней пемзы для обработки ткани или одежды, в особенности денима.
Как отмечалось выше, обработка целлюлазами может полностью заменять стирку с камнями пемзы (например, 1 кг коммерческого фермента заменяет использование 100 кг камней). Однако, обработка целлюлазами может быть скомбинирована с обработкой камнями пемзы, когда нужно получить сильно истертый продукт. Эффект персиковой кожи, когда создается внешний вид, напоминающий тонкий выступающий пушок персика, также достигается комбинированной стиркой с нейтральной целлюлазой и камнями пемзы. Описанные белки слияния особенно полезны для эффективного обеспечения потертого внешнего вида и минимизации обратного окрашивания в процессе биостоунинга.
Биостоунинг обычно осуществляют при рН от 3,0 до 8,0, предпочтительно, при рН от 5,0 до 7,0. Температура может варьировать от около 30°C до 80°C и, предпочтительно, от 50°C до 60°C. Отношение объема жидкости к весу ткани может варьировать от около 3:1 до 20:1, предпочтительно, от 5:1 до 10:1. Время обработки составляет от 15 мин до 90 мин, предпочтительно, от 30 мин до 60 мин. Следует иметь в виду, что доза фермента существенным образом зависит от типа ткани, стиральной машины, условий стирки (рН, температура, отношение объема жидкости к весу ткани, нагрузка денима, скорость процесса) и типа используемого ферментного препарата и т.д. Если необходимо, могут использоваться камни пемзы в сочетании с эндоглюканазными белками слияния. Доза ферментов может быть значительно снижена. Специалисту в данной области понятно, каким образом можно подобрать подходящие дозы ферментов и условия стирки.
Заявленные эндоглюканазные белки слияния обеспечивают неожиданные преимущества, поскольку ферменты обладают высокой активностью при минимальном обратном окрашивании ткани и хорошем контрасте окрашенных и неокрашенных участков. Кроме того, белки слияния легко получить и они могут быть использованы при относительно высоком разбросе температуры и рН.
Эндоглюканазные белки слияния полезны для биофинишинга тканей и одежды. Биофинишинг относится к использованию ферментов в процессе контролируемого гидролиза целлюлозных волокон для модификации поверхности тканей или нитей таким образом, чтобы предотвратить перманентное скатывание, улучшить мягкость и гладкость ткани, когда ее пробуют на ощупь, очистить поверхностную структуру с удалением катышек, что приводит к более ясному цвету, улучшает драпируемость ткани, впитывание влаги и поглощение красителя.
Ферментный депилинг может осуществляться на любой стадии влажной обработки ткани, предпочтительно, после необязательной расшлихтовки и/или отбеливания при условиях, сходных с условиями биостоунинга.
Используемые эндоглюканазные белки слияния, включающие также бета-глюканазы, гемицеллюлазы или ксиланазы, могут оказаться полезными и для улучшения качества кормов для животных, в особенности для обработки растительного материала ферментами.
Крахмал, белки и липиды могут легко расщепляться в пищеварительной системе моногастрических животных, таких как домашняя птица и свиньи, в то время как большая часть некрахмальных полисахаридов (NSP), в том числе связанные смешанным образом бета-глюканы, например из ячменя и овса, остаются нерасщепленными, поскольку у указанных животных отсутствуют ферменты с необходимой активностью. Более того, перевариваемость других компонентов, в особенности животных жиров, снижается в присутствии NSP.
Бета-глюканазы коммерчески использовались для решения проблем, имеющих отношение к связанным смешанным образом бета-глюканам из ячменя и овса. Известно, что бета-глюканазы снижают вязкость кишечного содержимого, вызываемую растворимыми бета-глюканами, а также высвобождают питательные вещества, инкапсулированные клеточными стенками, богатыми бета-глюканами. Использование бета-глюканаз улучшает самочувствие животных, что определяется прибавкой в весе и показателем отношения переработки корма. Кроме того, у домашней птицы уменьшается липкость помета.
Гранулирование с применением пара является основной технологической процедурой приготовления пищевых продуктов во всем мире. Преимущества ее при производстве продуктов из пищевой массы заключаются в более легкой обработке, уменьшении токсических компонентов и организмов и в перечисленных выше улучшениях качества продукта. Термоустойчивость и высокая активность белков слияния, описанные выше, делают их подходящими для использования в производстве пищевых продуктов.
Изобретение иллюстрируется приведенными ниже неограничивающими примерами. Следует понимать, что варианты осуществления изобретения, приведенные в описании, так же, как и примеры, служат исключительно в иллюстративных целях и в объем изобретения входят его различные изменения и модификации.
Пример 1. Получение целлюлазы Thermoascus aurantiacus ALKO4242 EG 28 в Trichoderma reesei
Для выделения и ферментной обработки ДНК (плазмиды, ДНК-фрагменты), трансформации Е.coli и т.д. используются стандартные биологические методы. Используемые базовые методы описаны в известных руководствах по молекулярной биологии, например, в руководстве Sambrook et al. (1989) и Sambrook и Russel (2001).
Ген cel5A Thermoascus aurantiacus (SEQ ID NO:1), кодирующий целлюлазу EG28 (SEQ ID NO:2), амплифицируют с помощью ПЦР непосредственно из геномной ДНК Thermoascus aurantiacus ALKO4242, выделенной методом Raeder и Broda (1985). Прямой (SEQ ID NO:9) и обратный (SEQ ID NO:10) праймеры конструируют на основе опубликованной последовательности эндоглюканазы Thermoascus aurantiacus (AF487830). Амплифицированный продукт размером 1,3 кб, включающий полный ген (от старт-кодона до стоп-кодона), клонируют в виде SacII-PstI-фрагмента в векторе pBluescript II KS+. Секвенируют два независимых клона и отбирают один клон, который обозначают pALK.1926. Штамм Е.coli, содержащий pALK1926, депонирован в немецкой коллекции микроорганизмов и клеточных культур GmbH под номером DSM 17326.
Плазмиду экспрессии (pALK1930, Фиг.1) конструируют для получения рекомбинантной целлюлазы Thermoascus aurantiacus EG28/Cel5A (SEQ ID NO:2). Ген cel5A, включающий собственную сигнальную последовательность, точно сливают с промотором cbhl Trichoderma reesei с помощью ПЦР. Промотор cbhl, терминатор cbhl и маркерный ген amdS включают в состав плазмиды, как описано Paloheimo et al. (2003). Линейную кассету экспрессии (Фиг.1) выделяют из остова вектора расщеплением ферментами рестрикции, вводят трансформацией в Trichoderma reesei A96, и отбирают трансформанты с помощью ацетамида как единственного источника азота. Хозяйский штамм лишен четырех основных эндогенных целлюлаз: CBHI/Cel7A, CBHII/Cel6A EGI/Cel7B и EGII/Cel5A. Трансформанты выращивают в соответствии с методами Penttilä et al. (1987) с модификациями, описанными Karhunen et al. (1993) Трансформанты очищают на селекционных планшетах до стадии конидий и споруляции на PD.
Продуцирование целлюлазы трансформантами анализируют в культуральных супернатантах при культивировании во встряхиваемых флаконах (50 мл). Трансформанты выращивают в течение 7 дней на сложной среде с целлюлозой (Joutsjoki et al. 1993), забуференной 5%-ным KH2PO4 при рН 5,5. Ферментная активность рекомбинантного белка измеряют в культуральном супернатанте по высвобождению редуцирующих Сахаров из карбоксиметилцеллюлозы (2% CMC) при 50°C в 50 мМ ацетатного буфера (рН 4,8), как описано Bailey и Nevalainen, 1981; Haakana et al. 2004. Активность по отношению к бета-глюкану ячменя (1%) также определяют измерением высвобождения редуцирующих сахаров 50°C в 50 мМ ацетатного буфера (рН 4,8), как описано Stálbrand et al. 1993. Продуцирование рекомбинантного белка также определяют в культуральном супернатанте путем SDS-полиакриламидного гель-электрофореза. Генотипы отобранных трансформантов анализируют Саузерн-блоттингом при использовании кассеты экспрессии в качестве пробы.
Оптимум рН гетерологично продуцируемой целлюлазы EG28/Cel5A определяют в универсальном буфере Mcllvaine при диапазоне рН от 4,0 до 8,0 при использовании карбоксиметилцеллюлозы в качестве субстрата. Как показано на Фиг.2А, оптимум рН для активности целлюлазы EG28/Cel5A составляет 6,0. Оптимальная температура ферментной активности целлюлазы EG28/Cel5A составляет 75°C (Фиг.2B).
Отобранный трансформант RF6188 культивируют в биореакторе с получением материала для тестирования возможных применений (см. Пример 4).
Пример 2. Получение рекомбинантного белка слияния Thermoascus aurantiacus ALKO4242 EG28+CtCBD
Для получения рекомбинантного белка слияния Thermoascus aurantiacus ALK04242 EG 28+CtCBD (SEQ ID NO:4) целлюлозо-связывающий домен (CBD) целлюлазы CBHI/Cel7A Chaetomium thermophilum ALKО4265 соединяют с целлюлазой EG28/Cel5A. Конструкцию, содержащую каталитический домен EG28 (аминокислоты 1-334 полноразмерного полипептида), прикрепляют к линкеру и CBD Chaetomium thermophilum CBHI/Cel7A CtCBD (SEQ ID NO:15).
Используют стандартные молекулярно-биологические методы, описанные в Примере 1. Сначала встраивают уникальный рестрикционный сайт SnaBI около C-концевого участка EG28/Cel5A путем ПЦР. Это обеспечивает непосредственное слияние любой ДНК со слепыми концами после аминокислоты Y334 полипептида EG28/Cel5A. Линкер и CBD-домен CBHI/Cel7A Chaetomium thermophilum кодирующего гена (Cel7A, SEQ ID NO:17) амплифицируют с помощью ПЦР при использовании прямого (SEQ ID NO:11) и обратного (SEQ ID NO:12) праймеров и геномной ДНК Chaetomium thermophilum ALKО4265 в качестве матрицы. Амплифицированный 1,6 кб продукт лигируют с геном Cel5A (после Y334) с получением Та Cel5A_Ct Cel7линкерCBD (SEQ ID NO:3). Полученную плазмиду обозначают pALK1946. Штамм E.coli, содержащий pALK1946, депонирован в немецкой коллекции микроорганизмов и клеточных культур GmbH под номером DSM 18159.
Плазмиду экспрессии (pALK1948, Фиг.1) для получения целлюлазы EG28+CtCBD конструируют, как описано в Примере 1. Линейную кассету размером 8,9 кб (Фиг.1) выделяют из остова вектора расщеплением рестрикционным ферментом Notl, вводят трансформацией в Trichoderma reesei A33 (штамм лишен генов, кодирующих четыре основные эндогенные целлюлазы: CBHI/Cel7A, CBHII/Cel6A EGI/Cel7B и EGII/Cel5A) и отбирают трансформанты, как описано в Примере 1. Оптимум рН и температуры для белка слияния определяют так же, как и для белка дикого типа EG28.
Отобранный трансформант RF6377 культивируют в биореакторе с получением материала для тестирования возможных применений (см. Примеры 5-10).
Пример 3. Получение рекомбинантного белка слияния Acremonium thermophilum ALKO4245 EG40+CtCBD
Для получения рекомбинантного белка слияния Acremonium thermophilum ALKO4245 EG40+CtCBD (SEQ ID NO:6) целлюлозо-связывающий домен (CBD) целлюлазы EG40 замещают таковым Chaetomium thermophilum ALKO4265 CBHI/Cel7A. Конструкцию, содержащую каталитический домен EG40 (аминокислоты 1-234 полноразмерного полипептида), прикрепляют к линкеру и CBD Chaetomium thermophilum (CtCBD, SEQ ID NO:15).
Используют стандартные молекулярно-биологические методы, описанные в Примере 1. Ген Cel45A Acremonium thermophilum (SEQ ID NO:8) амплифицируют с помощью ПЦР из геномной ДНК Acremonium thermophilum ALKO4245 при использовании праймеров (SEQ ID NO:13) и (SEQ ID NO:14). ПЦР-фрагмент 1,1 кб клонируют в векторе pBluescript II KS+ в виде фрагмента SacII-PstI. После этого встраивают уникальный рестрикционный сайт NruI около-С-концевого участка EG40 путем ПЦР. Это обеспечивает непосредственное слияние любой ДНК со слепыми концами после аминокислоты S234 полипептида EG40. Линкер и CBD-домен CBHI/Cel7A Chaetomium thermophilum кодирующего гена (Cel7A) амплифицируют с помощью ПЦР, как описано в Примере 2. Рестрикционный фрагмент лигируют с геном Cel45A (после S234) с получением At Cel45A_Ct Cel7линкерCBD (SEQ ID NO:5). Далее конструируют плазмиду экспрессии для продуцирования целлюлазы EG40+CtCBD и нарабатывают рекомбинантный белок (SEQ ID NO:6) в Trichoderma, как описано в Примере 1.
Пример 4. Активность целлюлазы EG28 при окончательной обработке денима при различных температурах
Целлюлазу Thermoascus aurantiacus EG28, полученную в Trichoderma (штамм RF6188), как описано в Примере 1, тестируют на способность создавать потертый вид денима (биостоунинг), который обеспечивается традиционной стиркой с камнями пемзы, при различных температурах. В качестве контроля сравнения используют эффективный фермент для окончательной обработки денима, представляющий собой коммерческий препарат EGII, обогащенный кислыми целлюлазами, полученный в Trichoderma (патент США 5874293).
В качестве тестируемого материала используют джинсы, сделанные из окрашенного Индиго денима, после расшлихтовки альфа-амилазой ECOSTONE® A200. Обработку целлюлазой проводят в стиральной машине Electrolux Wascator FOM 71 CLS при условиях, описанных в Таблице 1.
Эндоглюканазную активность (ECU) препарата ферментов определяют по высвобождению редуцирующих сахаров из гидроэтилцеллюлозы, как ранее описано Bailey и Nevalainen, 1981. Препарат EGII, обогащенный кислыми целлюлазами, и EG28 дозируют как 220 ECU/г и около 380 ECU/г ткани, соответственно. Ферменты тестируют при их оптимальном рН, а именно препарат EGII при рН 5 и препарат EG28 при рН 6. После сушки ферменты инактивируют (10 мин при 40°C) повышением рН до 11 с помощью гидроксида натрия. Затем джинсы трижды отполаскивают водой и высушивают в барабане.
Эффект биостоунинга/уровень потертости определяют измерением цвета по значениям отражения в спектрофотометре Minolta CM 2500 при использовании координат цветовой шкалы L*a*b* (освещение D65/2°). Интенсивность цвета лицевой и обратной сторон денима и карманов измеряют после расшлихтовки (т.е. до обработки целлюлазами) и после обработки целлюлазами. Каждое измерение цвета лицевой и обратной сторон денима и карманов представляют как среднее примерно 40, 20 или 12 измерений, соответственно. Для каждого тестирования используют по две пары джинсов и конечный результат представляют как среднее значение. Результаты экспериментов приведены в Таблице 2 и на Фиг.3
Контроль рН (рН 6,1-6,3)
35, 5 г Na2HPO42H2O+15 г лимонной кислоты
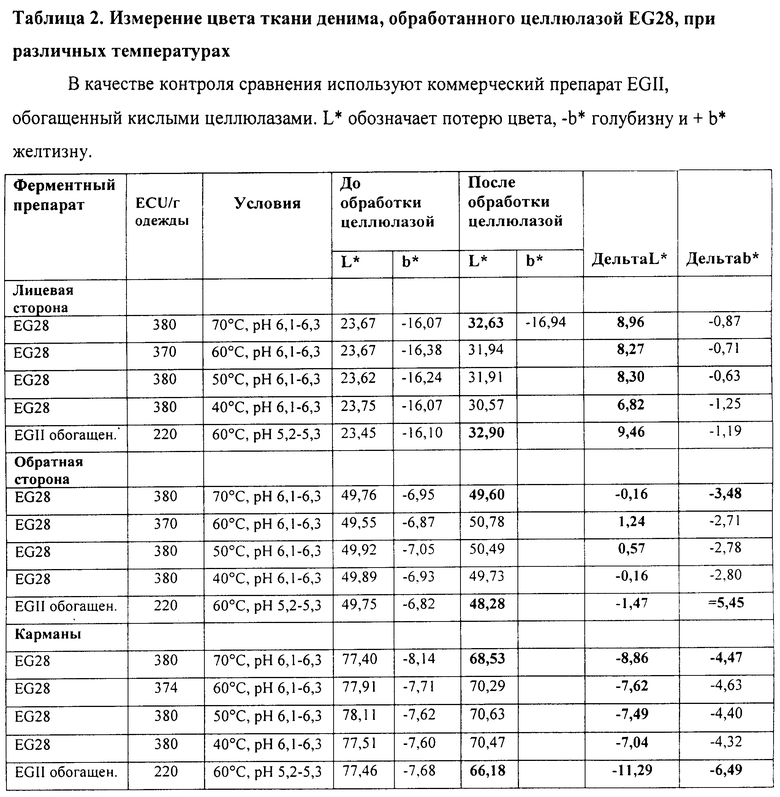
Результаты, приведенные в Таблице 2 и на Фиг.3, свидетельствуют о том, что наилучший эффект потертости при использовании препарата EG28 (штамм RF6188), обеспечивается при температуре от 50 до 70°C. При использовании дозы 380 ECU/г ткани препарата EG28 при 70°C наблюдается сходный уровень потертости (потеря цвета L*) при сравнении с использованием дозы 220 ECU/г ткани препарата EGII при 60°C. Однако, эффект обратного окрашивания (перераспределение красителя Индиго) на обратной стороне денима и карманах ниже при использовании препарата EG28 (более высокая потеря цвета, меньше голубого) при сравнении с использованием препарата EGII. Также контраст окраски на лицевой стороне денима более ярко выражен.
Пример 5. Активность целлюлазного белка слияния EG28+CtCBD по сравнению с целлюлазой EG28 при окончательной обработке денима
Рекомбинантный белок слияния Thermoascus aurantiacus EG28+CtCBD, полученный в Trichoderma (штамм RF6377), как описано в Примере 2, сравнивают с препаратом EG28 из штамма RF6188 в процессе биостоунинга денима при рН 6 и температуре 60°C. Деним и тестирующая система для биостоунинга такая же, как и в Примере 4, за исключением того, что вес денима достигает 1430 г вместе с дополнительными различными кусками денима, которые не включаются в измерения. Эффект обработки целлюлазами оценивают так же, как и в Примере 4.
Результаты, приведенные в Таблице 3, свидетельствуют о том, что эффект биостоунинга препарата EG28+CtCBD является очень хорошим при низкой дозе. При сравнении продуктов штамма RF6377 и штамма RF6188 наблюдается сходный эффект потертости (потеря цвета L*), однако для этого требуется в 6,5 раз меньшая доза продукта RF6377 по сравнению с продуктом RF6188. Присоединение целлюлозо-связывающего домена (CBD) к целлюлазе EG28 не увеличивает нежелательного эффекта обратного окрашивания, но существенно повышает активность, направленную на истирание (вываривание) ткани.
Пример 6. Активность целлюлазного белка слияния EG28+CtCBD по сравнению с целлюлазой EGII при окончательной обработке денима
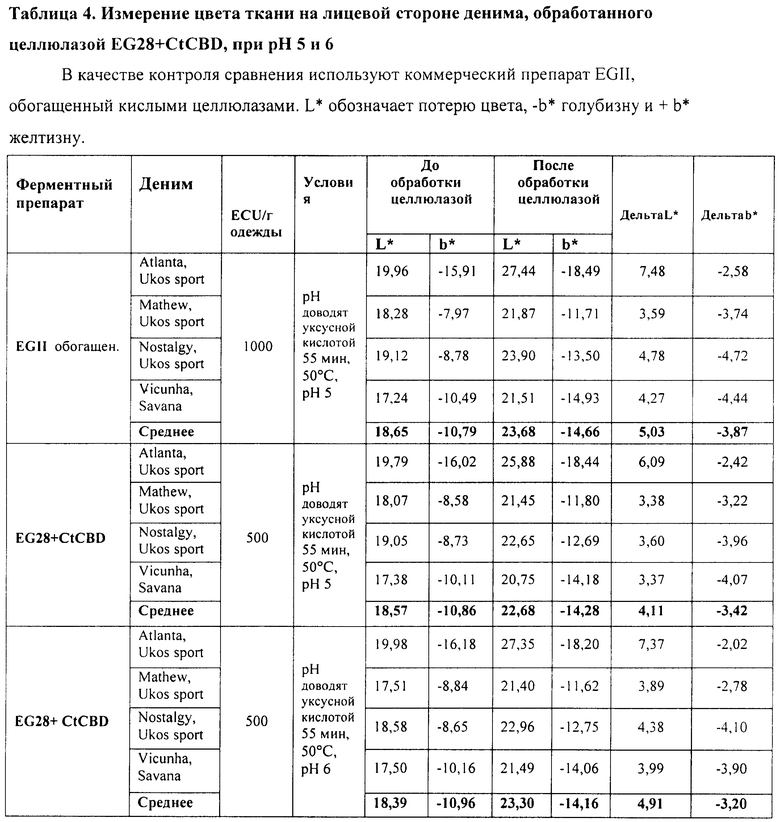
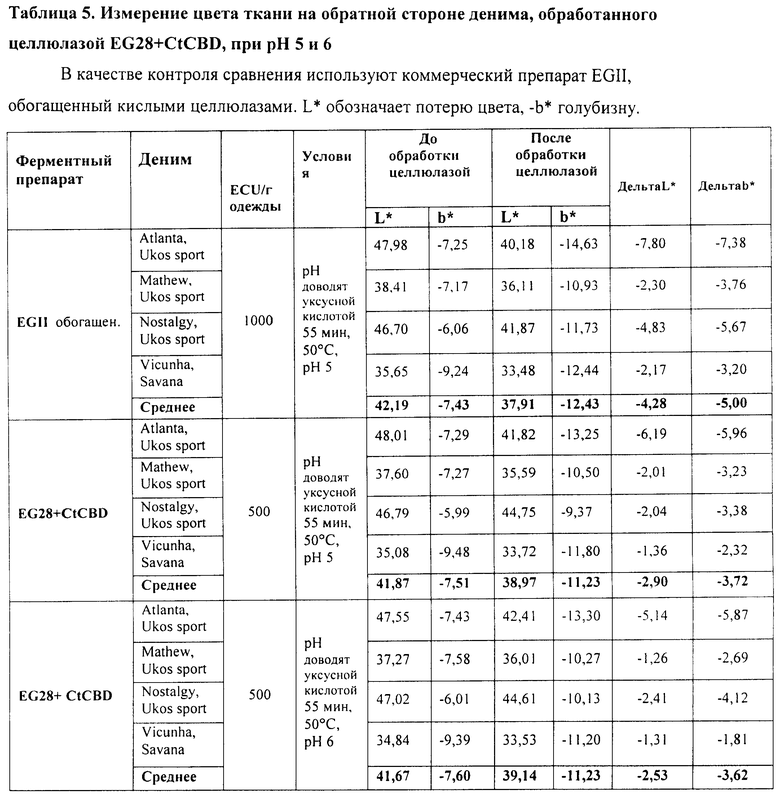
Рекомбинантный белок слияния Thermoascus aurantiacus EG28+CtCBD, полученный в Trichoderma (штамм RF6377), как описано в Примере 2, сравнивают с коммерческим препаратом EGII, обогащенным кислыми целлюлазами (патент США 5874293), при биостоунинге денима.
Тестирующая система для биостоунинга такая же, как и в Примере 4, за исключением того, что используют куски различных типов денима Ukos Sport (Бельгия) и Vicunha (Бразилия) при общем весе 1, 2 кг. Условия обработки EG28+CtCBD и EGII приведены в Таблице 4. Эффект обработки целлюлазами оценивают так же, как и в Примере 4.
Результаты экспериментов по обработке четырех различных типов денима приведены в Таблицах 4 и 5. При использовании дозы 500 ECU/г ткани препарата EG28+CtCBD наблюдается сходный уровень потертости при сравнении с использованием дозы 1000 ECU/г ткани препарата EGII. При использовании препарата EG28+CtCBD требуется примерно только половина ECU-дозы препарата EGII, обогащенного кислыми целлюлазами. Кроме того, для процесса биостоунинга культуральная среда, в которой выращивался рекомбинантный хозяин, продуцирующий EG28+CtCBD, по объему в два раза эффективнее среды, в которой выращивался рекомбинантный хозяин, продуцирующий EGII. Соответственно, обратное окрашивание в меньшей степени наблюдалось на обратной стороне денима при использовании препарата EG28+CtCBD. Активность препарата EG28+CtCBD при 50°C была лучшей при рН 6, чем при рН 5.
Пример 7. Влияние рН на активность целлюлазного препарата EG28+CtCBD при 50°C и 60°C
Рекомбинантный белок слияния Thermoascus aurantiacus EG28+CtCBD, полученный в Trichoderma (штамм RF6377), как описано в Примере 2, тестируют на активность при биостоунинге денима при 50°C и 60°C.
Окрашенный Индиго деним (отличный от используемого в ранее приведенных примерах) после расшлихтовки и разрезания на куски используют в качестве тестового материала. Обработку целлюлазой осуществляют в устройстве LP-2 Launder Ometer, как описано ниже. Около 7,2 г ткани денима (лоскут размером примерно 12 см × 12 см), 200 мл цитрат-фосфатного буфера Mcllvaine и 90 стальных шариков (диаметром 0,6 см) загружают в 1, 2 л контейнеры. Устройство Launder Ometer работает в течение 120 мин при различных значениях рН от 4 до 9 и температуре 50°C и 60°C. После удаления лоскутов из контейнеров их отполаскивают водой и вымачивают в воде, содержащей NaOH (рН больше 11) в течение 10 мин при перемешивании. Затем лоскуты вымачивают в теплой воде, содержащей жидкий детергент (ОМО Color) и перемешивают в течение 10 мин и далее осторожно отполаскивают теплой водой несколько раз. Лоскуты высушивают при комнатной температуре. Эффект обработки целлюлазой оценивают так же, как и в Примере 4. Каждое измерение на лицевой стороне лоскута денима определяют как среднее из 20 измерений.
Результаты экспериментов, приведенные в Таблицах 6 и 7 и на Фиг.4 и 5, показывают, что оптимальное значение рН для целлюлазы EG28+CtCBD при 50°C составляет от 5,5 до 6,5 и при 60°C от 5 до 7.
Пример 8. Активность целлюлазного препарата EG28+CtCBD при окончательной обработке денима различных температурах
Рекомбинантный белок слияния Thermoascus aurantiacus EG28+CtCBD, полученный в Trichoderma (штамм RF6377), как описано в Примере 1, тестируют на способность создавать потертый вид ткани, сходный с тем, который обеспечивается при стирке денима с камнями пемзы, при различных температурах. В качестве контроля сравнения используют эффективные ферменты для окончательной обработки денима, а именно коммерческий препарат EGII, обогащенный кислыми целлюлазами (патент США 5874293), и препарат на основе нейтральных целлюлаз ECOSTONE® С1. Тестирующая система для биостоунинга такая же, как и в Примере 4, за исключением того, что используют куски различных типов денима и время стирки составляет 55 мин. Ферментную активность (эндоглюканазная единица, ECU) препаратов EG28+CtCBD и EGII определяют, как описано в Примере 4. Активность (единица нейтральной целлюлазы, NCU) препарата на основе нейтральных целлюлаз ECOSTONE® C1 определяют по высвобождению редуцирующих сахаров из карбоксиметилцеллюлозы, как описано Bailey и Nevalainen, 1981; Haakana et al. 2004. Дозы ECOSTONE® C1, EG28+CtCBD и EGII составляют 250 NCU/г ткани, 500 ECU/г ткани или 1000 ECU/г ткани, соответственно. Эффект обработки целлюлазой оценивают так же, как и в Примере 4.
Активность EG28+CtCBD при температуре от 40 до 70°C и рН 6 приведена в Таблице 8 и на Фиг.6. Предпочтительный температурный интервал для фермента составляет от 50 до 70°C. Более низкие температуры такие, как 40°C, могут использоваться для получения эффекта потертости с большей интенсивностью окраски, если желательно, чтобы ткань выглядела более темной.
Результаты исследований согласуются с таковыми, полученными в Примере 6, где активность EG28+CtCBD при биостоунинге примерно в два раза выше активности коммерческого препарата EGII, обогащенного кислыми целлюлазами. Кислые целлюлазы обычно способствуют обратному окрашиванию. Нейтральные целлюлазы, подобные ECOSTONE® C1, обычно вызывают слабое обратное окрашивание, обеспечивая хороший контраст. Результаты, полученные в этом примере, показывают, что сходный эффект выявляется при использовании целлюлазы EG28+CtCBD и нейтральных целлюлаз (Таблица 8).
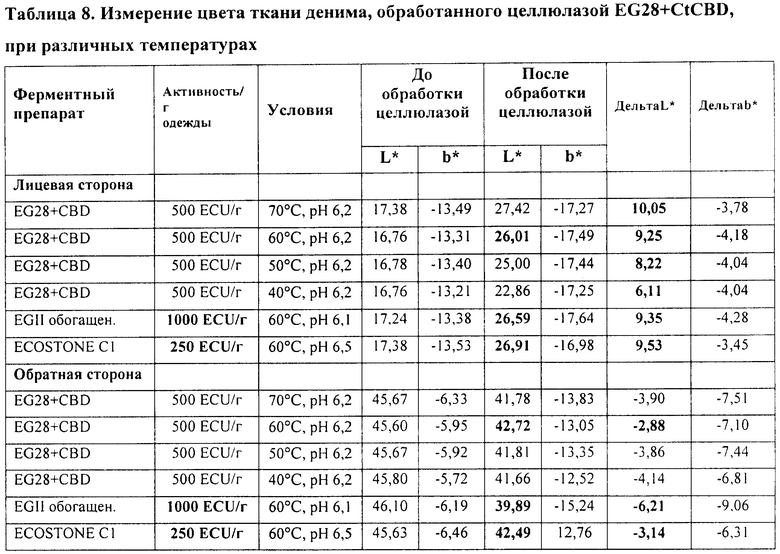
В качестве контроля сравнения используют коммерческий препарат EGII, обогащенный кислыми целлюлазами, и препарат на основе нейтральных целлюлаз ECOSTONE® C1. L* обозначает потерю цвета, -b* голубизну и +b* желтизну.
Пример 9. Активность целлюлазных препаратов EG28 и EG28+CtCBD при биофинишинге (удалении катышек)
Тестируют активность целлюлазы EG28 из штамма RF6188 и препарата EG28+CtCBD из штамма RF6377 при удаления катышек с хлопчато-бумажного трикотажа. Обработку целлюлазами осуществляют в стиральной машине Electrolux Wascator FOM 71 CLS при условиях, описанных в Таблице 9.
Лоскуты двух низкого качества джемперов с воротником «поло», на поверхности которых имеются многочисленные катышки, сделанных из 100%-ной хлопчато-бумажной ткани типа джерси или 95%-ной хлопчато-бумажной ткани типа репса и 5%-ной лайкры используют в качестве тестируемого материала и контроля. Образцы сначала предварительно стирают в течение 10 мин при 60°C и 1 мл/л сурфактанта/увлажняющего агента (Sandoclean PCJ, Sandos, и Imacol CN, Clariant) и три раза отполаскивают водой. После этого хлопчато-бумажный трикотаж обрабатывают целлюлазой при 60°C в течение 60 мин в присутствии тех же самых текстильных добавок, который используются при предварительной стирке. Ферменты инактивируют, как описано в Примере 4, за исключением того, что температура в процессе щелочного отполаскивания составляет 60°C. Лоскуты трикотажа три раза отполаскивают и высушивают в барабане.
Эффект обработки целлюлазами оценивают визуально невооруженным глазом и с помощью лупы. В качестве контроля используют предварительно выстиранные образцы ткани без фермента.
Результаты экспериментов показывают, что препарат EG28+CtCBD повышает устойчивость ткани к скатыванию. Число катышек на поверхности трикотажа значительно снижается после обработки EG28+CtCBD по сравнению с контролем (стирка без фермента). Препарат EG28 также имеет аналогичный эффект.
Пример 10. Тест на стабильность гранулированного пищевого продукта
Два целлюлазных препарата EG28 из штамма RF6188 и препарата EG28+CtCBD из штамма RF6377 по отдельности тестируют в эксперименте, напоминающем промышленный способ производства пищевого продукта. До начала тестирования препараты EG28 и EG28+CtCBD, высушенные распылением, грунтуют с пшеничной мукой для улучшения гомогенности. Ферменты EG28 и EG28+CtCBD, предварительно смешанные с мукой, добавляют в дозах 200 г/тонну и 500 г/тонну пищевого продукта, соответственно. Сверхдозы ферментов используются для обеспечения анализа образцов, гранулированных при высоких температурах, когда их активность может быть существенным образом снижена.
Для получения пищевой смеси в процессе полупромышленного производства растительного пищевого продукта с номинальной способностью к грануляции 5 т/ч используют мельницу и миксер. Мельница представляет собой устройство Champion, оснащенное сеткой с отверстиями диаметром 3,0 мм. Загрунтованный сырой материал перемешивают в 2,000 л горизонтальном миксере при 27 rpm до помещения в устройство пищевой минимельницы. Устройство содержит горизонтальный миксер (объем 700 л, скорость 48 rpm, смешивающая емкость 300 кг), дозирующий винт Skjold TR и каскадный миксер Kahl (длина 130 см, диаметр 30 см, скорость 155 rpm), оснащенный 37 подвижными поддонами. Время перерыва для 300 кг/ч составляет около 30 с. На стороне каскадного миксера смонтирован коллектор с пусковым устройством для воды и три паровых вентеля для подачи пара на пищевой продукт. Пар образуется в бойлере под высоким давлением с максимальной емкостью 400 кг пара/час. Тестирование проводят при сверхдавлении в 2 атмосферы и пар подают из вентеля за счет уменьшения давления под контролем введения пара в каскадный миксер Три вентеля на коллекторе используют для получения нужной температуры пищевого продукта. Температуру пищевого продукта измеряют цифровым термометром, расположенным на выходе из каскадного миксера, прямо перед тем, как продукт попадает на сетку для гранулирования. Используют пресс для грануляции Simon Heesen типа монорольного с сеткой диаметром 3 мм и мотором 7,5 kW. Внутренний диаметр сетки 173 мм, высота валика пресса 50 мм, диаметр валика пресса 140 мм, скорость оборота 500 rpm и номинальная емкость 300 кг/ч. Образцы отбирают после того, как они были гранулированы в прессе и охлаждают в перегородчатом охлаждающем контейнере с перфорированным дном при скорости вентилирования 1500 м3 воздуха/ч.
Получают партию продукта из пшеничной массы весом 300 кг. Премикс получают из 10 кг указанного продукта и 200 г (или 500 г) тестируемого фермента в 70 л в миксере. Скорость вращения миксера составляет 45 rpm, а время перемешивания - 10 мин. Указанный премикс добавляют к 290 кг пищевого продукта и помещают в горизонтальный миксер в устройстве пищевой минимельницы и перемешивают 10 мин. После взятия пробы образовавшейся массы пищевой продукт гранулируют пропусканием через пресс (диаметр отверстий сетки 3 мм). Продукт нагревают до нужной температуры (от 65 до 90°C) введением пара в каскадный миксер. Для каждой температуры образец сначала отбирают через 10 мин после достижения нужной температуры гранулирования (которую измеряют в пищевом продукте точно перед началом гранулирования). Образцы отбирают в течение 1,0 мин, что соответствует с 5,0 кг гранулированного пищевого продукта. Суб-образец весом около 500 г помещают в охлаждающий контейнер на 10-15 с после того, как гранулы выйдут из пресса. Все образцы аэрируют и охлаждают при сходной температуре в течение 15 мин. Образцы однородно разделяют на специальном делящем устройстве и заполняют ими пластиковые пакеты.
Образцы анализируют следующим образом: 2,5 г хорошо загрунтованного образца и 20 мл ацетатного буфера (0,05 М, рН 5,0) перемешивают при комнатной температуре в течение 30 мин, центрифугируют (10 мин, 4000 rpm) и разводят. 1,0 мл очищенного образца в трех повторах уравновешивают в течение 5 мин в водяной бане при 40°C. Реакцию инициируют добавлением таблетки бета-глюказима (Megazyme, Ireland) без перемешивания. После 30 мин реакцию останавливают добавлением 5,0 мл Trizma Base 1% (вес на объем) (Sigma-Aldrich) с интенсивным перемешиванием на мешалке вортекс. Пробирку оставляют при комнатной температуре примерно в течение 5 мин; жидкость перемешивают снова и фильтруют через бумажный фильтр Whatman I. Поглощение фильтрата измеряют при 590 нм. Результаты анализируют при использовании стандартной кривой.
Ферментную активность определяют до гранулирования и после гранулирования. Данные представлены в Таблице 10. Влияние температуры гранулирования на активность бета-глюканазы также и показано на Фиг.7 и 8. Целлюлазы EG28 и EG28+CtCBD стабильны при температуре до 80°C. Целлюлаза EG28+CtCBD более стабильна, чем EG28 при температуре гранулирования 90°C.
Перечень депонированных организмов


| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ОБРАБОТКИ ЦЕЛЛЮЛОЗНОГО МАТЕРИАЛА И ИСПОЛЬЗУЕМЫЕ В НЕМ ФЕРМЕНТЫ | 2006 |
|
RU2458128C2 |
| РЕКОМБИНАНТНЫЙ БЕЛОК LACspCBD, ОБЛАДАЮЩИЙ БЕТА-ГАЛАКТОЗИДАЗНОЙ АКТИВНОСТЬЮ И СПОСОБНОСТЬЮ САМОПРОИЗВОЛЬНО СВЯЗЫВАТЬСЯ С ЦЕЛЛЮЛОЗОСОДЕРЖАЩИМИ СОРБЕНТАМИ, РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, КОДИРУЮЩАЯ СИНТЕЗ РЕКОМБИНАНТНОГО БЕЛКА LACspCBD, ШТАММ Escherichia coli M15 [pREP4, pLACspCBD] - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО БЕЛКА LACspCBD, СПОСОБ ПОЛУЧЕНИЯ ИММОБИЛИЗОВАННОГО РЕКОМБИНАНТНОГО БЕЛКА LACspCBD НА ЦЕЛЛЮЛОЗЕ И СПОСОБ ФЕРМЕНТАТИВНОГО РАСЩЕПЛЕНИЯ ЛАКТОЗЫ | 2004 |
|
RU2278160C2 |
| СПОСОБЫ ПОВЫШЕНИЯ УСИЛИВАЮЩЕЙ ЦЕЛЛЮЛОЛИТИЧЕСКОЙ АКТИВНОСТИ ПОЛИПЕПТИДА | 2008 |
|
RU2510417C2 |
| ПОЛИПЕПТИДЫ, ОБЛАДАЮЩИЕ ЦЕЛЛЮЛАЗНОЙ АКТИВНОСТЬЮ | 2010 |
|
RU2597289C2 |
| ФЕРМЕНТНАЯ КОМПОЗИЦИЯ, СПОСОБНАЯ ЭФФЕКТИВНО РАЗЛАГАТЬ ЦЕЛЛЮЛОЗНЫЙ МАТЕРИАЛ | 2009 |
|
RU2529949C2 |
| СПОСОБ ЗАТИРАНИЯ | 2004 |
|
RU2376347C2 |
| НОВЫЕ РЕГУЛИРУЮЩИЕ ЭКСПРЕССИЮ ПОСЛЕДОВАТЕЛЬНОСТИ И ПРОДУКТ ЭКСПРЕССИИ В ОБЛАСТИ ГИФОМИЦЕТОВ | 2001 |
|
RU2272835C2 |
| РЕКОМБИНАНТНЫЙ БЕЛОК Collbd-CBD, РЕКОМБИНАНТНАЯ ПЛАЗМИДА pОС-Collbd, ШТАММ Escherichia coli - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО БЕЛКА Collbd-CBD, СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО БЕЛКА Collbd-CBD | 2009 |
|
RU2408726C1 |
| КОМПОЗИЦИЯ ФЕРМЕНТНОЙ СМЕСИ ДЛЯ ГИДРОЛИЗА СМЕСИ ЦЕЛЛЮЛОЗНЫХ И ГЕМИЦЕЛЛЮЛОЗНЫХ МАТЕРИАЛОВ (ВАРИАНТЫ) И СПОСОБЫ ЕЕ ИСПОЛЬЗОВАНИЯ (ВАРИАНТЫ) | 2009 |
|
RU2536256C2 |
| ГЕНЕТИЧЕСКАЯ КОНСТРУКЦИЯ ДЛЯ ОБЕСПЕЧЕНИЯ ЭКСПРЕССИИ ЦЕЛЕВЫХ ГОМОЛОГИЧНЫХ И ГЕТЕРОЛОГИЧНЫХ ГЕНОВ В КЛЕТКАХ МИЦЕЛИАЛЬНОГО ГРИБА PENICILLIUM VERRUCULOSUM, ИСПОЛЬЗУЕМОГО В КАЧЕСТВЕ ХОЗЯИНА, СПОСОБ ПОЛУЧЕНИЯ ШТАММА ГРИБА PENICILLIUM VERRUCULOSUM И СПОСОБ ПОЛУЧЕНИЯ ФЕРМЕНТНОГО ПРЕПАРАТА | 2008 |
|
RU2378372C2 |











Изобретение относится к области биотехнологии и касается целлюлазных слитых белков. Представленные слитые белки содержат аминокислотную последовательность эндоглюканазного ядра, имеющую, по крайней мере, 95%-ную идентичность с SEQ ID NO:2, слитую с аминокислотной последовательностью, содержащей линкер и целлюлозосвязывающий домен (CBD), имеющую, по крайней мере 95%-ную идентичность с SEQ ID NO:15. Такие слитые белки могут быть получены с помощью рекомбинантной технологии при использовании подходящих полинуклеотидов, экспрессирующих векторов и клеток-хозяев. Представленное изобретение обеспечивает целлюлазу, обладающую низкой активностью в отношении восстановления окраски, и может быть использовано для обработки целлюлозного материала, например, текстильного материала и для биологической абразивной обработки денима. Кроме того, представленные слитые белки и ферментные препараты на их основе могут использоваться для приготовления детергентных композиций или для улучшения качества кормов для животных. 12 н. и 14 з.п. ф-лы, 8 ил., 10 табл., 10 пр.
1. Целлюлозный слитый белок, содержащий первую аминокислотную последовательность эндоглюканазного ядра, имеющую, по крайней мере, 95%-ную идентичность с SEQ ID NO:2 и целлюлазную активность, или фрагмент указанного белка, имеющий целлюлазную активность и вторую аминокислотную последовательность, содержащую линкер и целлюлозосвязывающий домен (CBD), имеющую, по крайней мере, 95%-ную идентичность с SEQ ID NO:15 и целлюлозосвязывающую активность, или фрагмент указанного белка, имеющий целлюлозосвязывающую активность.
2. Слитый белок по п.1, где эндоглюканаза принадлежит семейству 5 гликозилгидролаз.
3. Слитый белок по любому из пп.1 или 2, где эндоглюканазное ядро происходит из Thermoascus aurantiacus, в особенности из Т. aurantiacus CBS 116239.
4. Слитый белок по п.1, где эндоглюканазное ядро имеет, по крайней мере, 98%-ную идентичность с SEQ ID NO:2, или фрагмент указанного белка, имеющий целлюлазную активность.
5. Слитый белок по п.1, где эндоглюканазное ядро имеет, по крайней мере, 99%-ную идентичность с SEQ ID NO:2, или фрагмент указанного белка, имеющий целлюлазную активность.
6. Слитый белок по п.4, где эндоглюканазное ядро содержит аминокислоты 19-334 SEQ ID NO:2.
7. Слитый белок по п.1, где линкер и CBD происходят из целлобиогидролазы Chaetomium thermophilum.
8. Слитый белок по п.7, где линкер и CBD происходят из целлобиогидролазы С. thermophilum CBS 730.95.
9. Слитый белок по п.1, где эндоглюканазное ядро содержит последовательность SEQ ID NO:2, а линкер и CBD содержат последовательность SEQ ID NO:15.
10. Слитый белок по п.1, где эндоглюканазное ядро кодируется геном, который включен в штамм E.coli DSM 17326.
11. Слитый белок по п.1, который кодируется слитым геном, который включен в штамм E.coli DSM 18159.
12. Выделенный полинуклеотид, кодирующий целлюлазный слитый белок и выбранный из группы, включающей:
а) нуклеотидную последовательность SEQ ID NO:3;
б) последовательность, полученную в соответствии с вырожденностью генетического кода, на основе последовательности а) и
в) нуклеотидную последовательность, кодирующую белок слияния, содержащий первую аминокислотную последовательность эндоглюканазного ядра, имеющую, по крайней мере, 95%-ную идентичность с SEQ ID NO:2 и целлюлазную активность, или фрагмент указанного белка, имеющий целлюлазную активность и вторую аминокислотную последовательность, содержащую линкер и целлюлозосвязывающий домен (CBD), имеющую, по крайней мере, 95%-ную идентичность с SEQ ID NO:15 и целлюлозосвязывающую активность, или фрагмент указанного белка, имеющий целлюлозосвязывающую активность.
13. Полинуклеотид по п.12, представляющий собой слитый ген, кодирующий целлюлазный слитый белок по п.1.
14. Вектор экспрессии целлюлазного слитого белка по п.1, несущий полинуклеотид по п.12.
15. Клетка-хозяин, относящаяся к нитевидным грибам, содержащая вектор экспрессии по п.14 и продуцирующая слитый белок по п.1.
16. Клетка-хозяин по п.15, относящаяся к виду грибов рода Trichoderma, содержащая экспрессирущий вектор по п.14 и продуцирующая слитый белок по п.1.
17. Способ получения слитого белка по любому из пп.1-11, включающий трансформацию клетки-хозяина экспрессирующим вектором, кодирующим указанный белок слияния, и культивирование указанной клетки-хозяина в условиях, обеспечивающих экспрессию указанного белка слияния, и при необходимости выделение и очистку продуцируемого клеткой белка слияния.
18. Ферментный препарат, имеющий целлюлазную и целлюлозосвязывающую активность и содержащий слитый белок по любому из пп.1-11, полученный способом, включающим трансформацию клетки-хозяина экспрессирующим вектором, кодирующим указанный слитый белок, и культивирование указанной клетки-хозяина в условиях, обеспечивающих экспрессию указанного слитого белка, и при необходимости выделение и очистку продуцируемого клеткой белка слияния.
19. Способ обработки целлюлозного материала, предусматривающий обеспечение контактирования целлюлозного материала со слитым белком по любому из пп.1-11 или ферментным препаратом по п.18.
20. Способ по п.19, где целлюлозный материал представляет собой текстильный материал, растение, используемое для кормления животных, или полученную из древесины целлюлозную массу, или вторичное волокно.
21. Способ биологической абразивной обработки тканей (биостоунинг), включающий стадию обеспечения контактирования целлюлазного белка слияния по любому из пп.1-11 или ферментного препарата по п.18 с тканью денима или одеждой из денима.
22. Способ окончательной биологической обработки тканей (биофинишинг), включающий стадию обеспечения контактирования целлюлазного белка слияния по любому из пп.1-11 или ферментного препарата по п.18 с текстильными материалами, такими, как ткань, или одежда, или нити.
23. Детергентная композиция, содержащая белок слияния по любому из пп.1-11 и детергентные добавки для обработки целлюлозного материала, такого, как текстильный материал.
24. Корм для животных, содержащий белок слияния по любому из пп.1-11 для улучшения пищеварения и степени усвояемости корма, содержащий целлюлозный материал, такой, как растительный материал.
26. Штамм Escherichia coli, содержащий ген, кодирующий слитый белок по п.1, депонированный под номером DSM 18159.
| Прибор, замыкающий сигнальную цепь при повышении температуры | 1918 |
|
SU99A1 |
| Прибор, замыкающий сигнальную цепь при повышении температуры | 1918 |
|
SU99A1 |
| US 7138263 А, 21.11.2006 | |||
| GARRAD G | |||
| et al., Cellulose binding domains promoter hydrolysis of different sites on crystalline cellulose, PNAS, 2000, v.97, №19, p.10342-10347 | |||
| POOLE D.M | |||
| et al., Characterization of hybrid proteins consisting of the catalytic domain of Clostridium and | |||

