Изобретение относится к генной инженерии и может найти применение в биотехнологии и в пищевой промышленности при производстве и переработке молочных продуктов.
Промышленная переработка молочной сыворотки является одной из важнейших проблем пищевой промышленности. Присутствие в ней лактозы является существенным фактором, затрудняющим ее промышленную переработку и возможность дальнейшего ее использования в пищевых целях. Это определяется, в первую очередь, незначительной сладостью лактозы, плохой растворимостью, низкой усваиваемостью человеческим организмом и явлением интолерантности (непереносимости), слабой сбраживаемостью микроорганизмами, в особенности дрожжами. Промышленная переработка сыворотки необходима и с экологической точки зрения, поскольку при ее сбрасывании в окружающую среду наряду с потерей ценных пищевых веществ возникает проблема загрязнения сточных вод.
Традиционные методы переработки сыворотки, основанные на кислотном гидролизе, не удовлетворяют современным требованиям ее полной утилизации. Улучшить технологические и диетические свойства сыворотки возможно путем гидролиза лактозы на составляющие моносахариды - глюкозу и галактозу ферментативным способом. Сладость продуктов гидролиза увеличивается в три раза, растворимость - в 2,5 раза, они легко сбраживаются микроорганизмами и усваиваются человеком. Полученные в результате ферментативной обработки гидролизаты лактозы могут служить заменителями сахарозы во многих отраслях молочной и пищевой промышленности.
Существенный интерес представляет получение безлактозного молока, что связано с интолерантностью (непереносимостью) молока значительной частью населения. Интолерантность в значительной степени определяется отсутствием в организме этих людей ферментов, расщепляющих лактозу.
Наиболее перспективными с точки зрения технологичности процесса и качества лактозных гидролизатов являются ферментативные способы гидролиза лактозы растворимыми и иммобилизованными бета-галактозидазами. Они позволяют перерабатывать любое лактозосодержащее сырье, а мягкие условия реакции позволяют исключить возможность образования нежелательных продуктов гидролиза, снижающих качество гидролизатов и требующих дополнительной очистки.
Иммобилизация бета-галактозидазы позволяет снизить производственные затраты и улучшить экономические показатели процесса гидролиза лактозы в сыворотке за счет многократного использования одной и той же партии фермента, обеспечения непрерывности процесса гидролиза, улучшения качества гидролиза при стабильно высоком уровне конверсии лактозы, снижения возможности образования побочных соединений и отсутствия фермента в конечном продукте.
Для иммобилизации бета-галактозидаз обычно используют химические способы, основанные на образовании ковалентной связи между матрицей носителя и ферментом, а также физические, заключающиеся в адсорбции бета-галактозидазы, или же включению ее в нерастворимую или полупроницаемую микрокапсулу. Срок службы одной партии иммобилизованной бета-галактозидазы может достигать нескольких тысяч часов, что значительно снижает стоимость процесса гидролиза лактозы по сравнению с использованием растворимых ферментов.
Для использования в молочной промышленности перспективны бета-галактозидазы, обладающие повышенной термостабильностью, высоким сродством к субстрату, способностью эффективно работать при рН молочной сыворотки. Получить такой ферментативный препарат можно, используя штаммы бактерий, трансформированные плазмидными ДНК, кодирующими синтез бета-галактозидазы с улучшенными свойствами.
Известен рекомбинантный белок, обладающий активностью бета-галактозидазы (патент RU 2059725 С1, класс 6 С 12 N 15/00, 15/56, 9/38, опубл. 10.05.1996 г.) [1].
Известна рекомбинантная плазмидная ДНК pVT1, кодирующая синтез рекомбинантного белка, обладающего активностью бета-галактозидазы [1].
Известен штамм Escherichia coli, трансформированный данной известной рекомбинантной плазмидной ДНК, - продуцент рекомбинантного белка, обладающего активностью бета-галактозидазы [1].
Однако для использования данного известного рекомбинантного белка, обладающего активностью бета-галактозидазы, в иммобилизованном виде применимы только известные приемы иммобилизации с присущими им недостатками, а использование неиммобилизованного фермента требует больших количеств препарата и экономически не оправдано.
Известен способ иммобилизации бета-галактозидазы на пористом кремнеземе (Авторское свидетельство СССР 1317024 А1, класс С 12 N 11/14, опубл. 15.06.87) [2], способ иммобилизации ферментов на активированной перекисью водорода титановой губке (Авторское свидетельство СССР 1472505 А1, класс С 12 N 11/14, опубл. 15.04.89) [3], способ химической иммобилизации ферментов на активированной диметилацетамидом целлюлозе (Авторское свидетельство СССР 1567625 А1, класс С 12 N 11/04, 11/12, опубл. 30.05.90) [4].
Однако все данные способы иммобилизации ферментов требуют предварительного получения высокоочищенных препаратов фермента, получаемых в результате многостадийной очистки. При химической иммобилизации происходит частичная инактивация фермента. Кроме того, при получении химически иммобилизованных препаратов бета-галактозидазы применяются токсичные реагенты и носители, типа фенол-формальдегидных смол, перекиси водорода и т.д., что затрудняет их применение в пищевой промышленности.
Известен способ создания слитых белков с целлюлозосвязывающими доменами для иммобилизации и очистки полипептидов (патент US 5137819; Kilburn, et al., 11.08.1992) [5].
Однако данный способ не позволяет получать слитые белки, способные к необратимой иммобилизации на целлюлозе в широком диапазоне рН и при высоких температурах.
Известен способ ферментативного расщепления лактозы путем обработки ее раствора препаратом бета-галактозидазы (Weetall H.H., Havewala N.B., Pitcher W.H. Jr., Detar C.C., Vann W.P., Yaverbaum S. Preparation of immobilized lactase. Continued studies on the enzymatic hydrolysis of lactose. - Biotechnol. Bioeng. 1974, v.16, №5, p.689-696) [6].
Однако при этом иммобилизация бета-галактозидазы также достигалась достаточно трудоемким химическим способом. Кроме того, используемая бета-галактозидаза обладала недостаточно высокой термостабильностью, что не позволяло использовать эффективную схему высокотемпературного ферментативного гидролиза лактозы.
Технический результат, достигаемый настоящим изобретением, заключается в повышении прочности связывания с целлюлозой в широком диапазоне рН и при высоких температурах слитого белка по п.1, обладающего термостабильной бета-галактозидазной активностью; в повышении выхода белка за счет придания плазмиде по п.2 свойства экспрессировать белок по п.1 в бактериальных клетках и включения плазмиды по п.2 в состав штамма-продуцента по п.3 белка по п.1, в упрощении процесса иммобилизации и повышении прочности иммобилизованного способом по п.4 на целлюлозе белка по п.1, в совмещении процесса его очистки с процессом его иммобилизации на целлюлозе, в упрощении процесса ферментативного расщепления лактозы по п.5 с использованием белка по п.1 и в обеспечении экологической безопасности этого процесса.
Указанный технический результат достигается тем, что слитый рекомбинантный белок LACspCBD, состоящий из бета-галактозидазы Thermoanaerobacter ethanolicus, Gly-Ser спейсера PGSGSGSGSGSGA и целлюлозосвязывающего домена
GSNAPLGEKVLPSTFEDDTRQGWDWDGPSGVKGPITIESANGSKALSFNVEYPEKKPQDG
WATAARLILKDINVERGNNKYLAFDFYLKPDRASKGMIQMFLAFSPPSLGYWAQVQDSFN
IDLGKTVKCKKDRRTEVYKFNVFFDLDKIQDNKVLSPDTLLRDIIVVIADGNSDFKGKMY
IDNVRSY эндоглюканазы CelD из Anaerocellum thermophilum
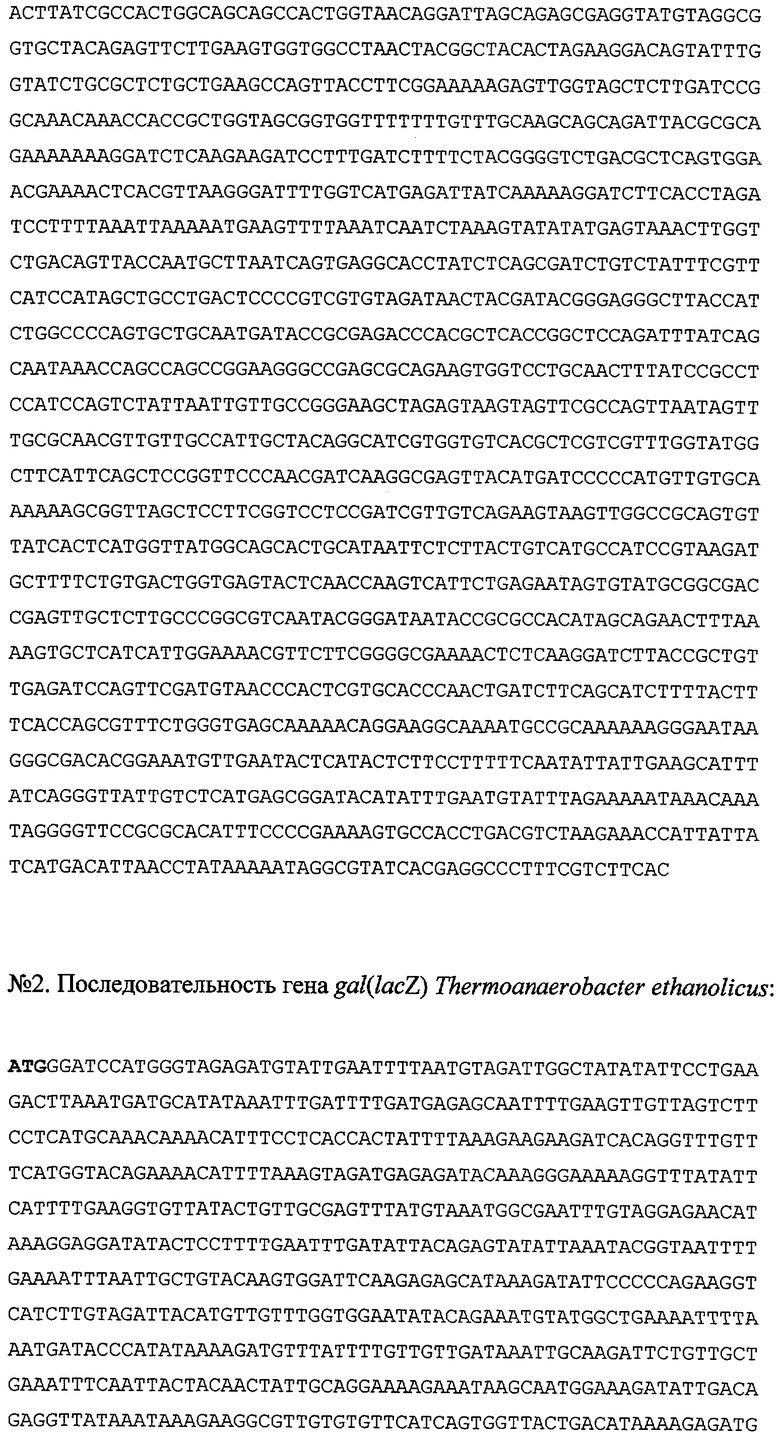
Достигается указанный технический результат также тем, что рекомбинантная плазмидная ДНК pLACspCBD размером 6233 н.п. (№1 в перечне последовательностей), содержащая:
- искусственный бактериальный оперон слитого белка бета-галактозидазы Thermoanaerobacter ethanolicus и целлюлозосвязывающего домена эндоглюканазы CelD Anaerocellum thermophilium, включающий промоторную область раннего промотора бактериофага Т5 (7-87 н.п.), нуклеотидную последовательность, кодирующую указанный слитный белок (115-2958 н.п.) и терминатор транскрипции (2979-3073 н.п.);
- бактериальный оперон бета-лактамазы (6028-5168 н.п. комплементарной цепи);
- бактериальный участок инициации репликации типа ColE1, обеспечивающий репликацию плазмидной ДНК в клетках Е.coli (4405 н.п.);
обеспечивает экспрессию слитного белка бета-галактозидазы Thermoanaerobacter ethanolicus и целлюлозосвязывающего домена эндоглюканазы CelD Anaerocellum thermophilium (белок LACspCBD) в клетках Е.coli.
Также указанный технический результат достигается тем, что штамм Escherichia coli ВКПМ В-8695 является продуцентом слитного белка бета-галактозидазы Thermoanaerobacter ethanolicus и целлюлозосвязывающего домена эндоглюканазы CelD Anaerocellum thermophilium (белка LACspCBD).
Достигается указанный технический результат также тем, что способ получения иммобилизованного на целлюлозе слитного белка LACspCBD, заключается в обработке супернатанта гидролизата клеток Escherichia coli M15 [pRep4, pLACspCBD] целлюлозой.
А также указанный технический результат достигается тем, что способ ферментативного расщепления лактозы путем обработки ее раствора препаратом бета-галактозидазы, имеет ту отличительную особенность, что в качестве препарата бета-галактозидазы используют рекомбинантный термостабильный белок LACspCBD, полученный способом по п.1-3 и иммобилизованный на целлюлозе по п.4.
На чертеже представлена схема рекомбинантной плазмидной ДНК pLACspCBD.
Ниже приведены примеры, иллюстрирующие изобретение.
Пример 1. Получение плазмиды pLACspCBD.
а) Химический синтез олигодезоксирибонуклеотидов.
Олигодезоксирибонуклеотиды были синтезированы твердофазным амидофосфитным методом с помощью синтезатора АСМ 100-2 (Новосибирск) и очищены методом электрофореза в 12%-ном ПАА геле.
б) Получение гена gal (lacZ) с последующим его клонированном.
Копию гена gal (lacZ) получали методом полимеразной цепной реакции, для чего были выбраны праймеры, соответствующие началу и концу данной области ДНК. Используя в качестве ДНК-матрицы хромосомную ДНК Thermoanaerobacter ethanolicus, методом ПЦР с использованием олигонуклеотидных праймеров состава (подчеркнуты сайты рестриктаз BamHI, XbaI и BglII)
1. 5'-ggaggcgggatccatgggtagagatgtattg-3'
2. 5'-aacccatctagagggtgggtttttaagatcttattgact-3',
соответствующих начальной и концевой части ДНК, амплифицировали последовательность нуклеотидов. Реакцию проводили в 25 мкл реакционной смеси, содержащей 2 мкг ДНК, 10 пкМ каждого праймера, 67 мМ трис-HCl (рН 8,8 при 25°С), 15 мМ сульфата аммония, 2,5 мМ хлористого магния, 0,01% Твин-20, смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ) и 1 ед. Tag-ДНК-полимеразы. Реакцию амплификации проводили под вазелиновым маслом: 5 циклов: 93°С - 2 мин, 57°С - 5 мин и 72°С - 30 сек; 30 циклов: 93°С - 1 мин, 63°С - 1 мин и 72°С - 30 сек; затем 63°С - 5 мин и 72°С -10 мин. Продукт амплификации размером 2267 н.п. обрабатывали хлороформом, переосаждали этиловым спиртом и растворяли в воде. Для клонирования данного продукта полученную ДНК PCRgal(lacZ), а также ДНК плазмиды pQE16 (Quagene, USA) гидролизовали рестрикционными эндонуклеазами BamHI и XbaI при 37°С в буфере, содержащем 66 мМ Трис-ацетат (рН 7,9 при 37°С), 20 мМ ацетата магния, 132 мМ ацетата калия и 0,2 мг/мл BSA в течение 1,5 часов. Выделенные из геля рестрикционные фрагменты: фрагмент ДНК соответствующий гену gal(lacZ) размером 2248 н.п. и фрагмент плазмиды pQE16 размером 2419 н.п. лигировали с помощью ДНК-лигазы бактериофага Т4. Полученной лигированной смесью трансформировали клетки Е.coli M15[pREP4] методом электропорации. Трансформированные клетки отбирали на агаризованной среде LB с антибиотиками ампициллином и канамицином. Плазмидную ДНК выделяли методом щелочного лизиса, анализировали с помощью рестриктаз BamHI и XbaI, а также рестриктаз, находящихся внутри клонируемых фрагментов, и отбирали клоны pLAC, плазмидная ДНК которых (размером 4667 н.п.) содержит последовательность гена gal(lacZ) (№2 в перечне последовательностей).
Первичную структуру полученной плазмиды подтверждали секвенированием. Аминокислотная последовательность Gal(LacZ) приведена в перечне последовательностей под №5.
в) Получение последовательности, кодирующей Gly-Ser спейсер, с последующим ее клонированием.
Для получения последовательности спейсера, состоящего из повторов аминокислотных остатков Gly-Ser, были синтезированы два комплементарных олигонуклеотида следующего состава (подчеркнут сайт эндонуклеазы рестрикции BglII, под нуклеотидной последовательностью первого олигонуклеотида приведена кодируемая последовательность аминокислот):
1.5'-gatcc ccg ggt tct ggc tcc ggc tct ggt tcc ggt tct ggc gcc aga tct a-3'
P G S G S G S G S G S G A
2. 5'-agcttagatctggcgccagaaccggaaccagagccggagccagaacccggg-3'.
Для получения двуцепочечного фрагмента ДНК смешивали по 20 пкМ каждого олигонуклеотида, прогревали смесь 10 мин при 95°С и охлаждали в течение 4 часов до 25°С. Для клонирования данного продукта ДНК плазмиды pQE13 гидролизовали рестрикционными эндонуклеазами BamHI и HindIII при 37°С в буфере, содержащем 66 мМ Трис-ацетата (рН 7,9 при 37°С), 20 мМ ацетата магния, 132 мМ ацетата калия и 0,2 мг/мл BSA в течение 1,5 часов. Выделенный из геля рестрикционный фрагмент плазмиды pQE13 размером 3420 н.п. объединяли с двуцепочечным фрагментом ДНК, кодирующим спейсер, и лигировали с помощью ДНК-лигазы бактериофага Т4. Полученной лигированной смесью трансформировали клетки Е.coli M15[pREP4] методом электропорации. Трансформированные клетки отбирали на агаризованной среде LB с антибиотиками ампициллином и канамицином. Плазмидную ДНК выделяли методом щелочного лизиса, анализировали с помощью рестриктаз BamHI и HindIII, а также рестриктаз, находящихся внутри клонируемых фрагментов, и отбирали клоны pRsp, плазмидная ДНК которых (размером 3471 н.п.) содержит последовательность Gly-Ser спейсера (№3 в перечне последовательностей). Первичную структуру полученной плазмиды подтверждали секвенированием.
г) Получение участка гена celD, содержащего последовательность целлюлозосвязывающего домена, с последующим его клонированием в плазмидную конструкцию pRsp.
Копию участка гена celD получали методом полимеразной цепной реакции, для чего были выбраны праймеры, соответствующие началу и концу данной области ДНК. Используя в качестве ДНК-матрицы хромосомную ДНК Anaerocellum thermophilum, методом ПЦР с использованием олигонуклеотидных праймеров состава (подчеркнуты сайты рестриктаз BamHI и HindIII и BglII):
1. 5'-aaagaaggatccaatgcacctttaggcg-3'
2. 5'-cctcaaaaaagctttaggtagatctaacattatctatatac-3',
соответствующих начальной и концевой части ДНК, амплифицировали последовательность нуклеотидов. Реакцию проводили в 25 мкл реакционной смеси, содержащей 2 мкг ДНК, 10 пкМ каждого праймера, 67 мМ трис-HCl (рН 8,8 при 25°С), 15 мМ сульфата аммония, 2,5 мМ хлористого магния, 0,01% Твин-20, смесь дезоксинуклеотидтрифосфатов (дАТФ, дЦТФ, дТТФ, дГТФ по 2,5 мМ) и 1 ед. Taq-ДНК-полимеразы. Реакцию амплификации проводили под вазелиновьм маслом: 5 циклов: 93°С - 2 мин, 57°С - 5 мин и 72°С - 30 сек; 30 циклов: 93°С - 1 мин, 63°С - 1 мин и 72°С - 30 сек; затем 63°С - 5 мин и 72°С - 10 мин. Продукт амплификации размером 583 н.п. обрабатывали хлороформом, переосаждали этиловым спиртом и растворяли в воде. Для клонирования данного продукта полученную ДНК PCRCBD гидролизовали рестрикционными эндонуклеазами BamHI и HindIII при 37°С в буфере, содержащем 66 мМ Трис-ацетата (рН 7,9 при 37°С), 20 мМ ацетата магния, 132 мМ ацетата калия и 0,2 мг/мл BSA в течение 1,5 часов. Также ДНК плазмиды pRsp гидролизовали рестрикционными эндонуклеазами BglII и HindIII. Выделенные из геля рестрикционные фрагменты: фрагмент ДНК, соответствующий участку гена CelD размером 563 н.п., и фрагмент плазмиды pRsp размером 3465 п.н., содержащий селективный маркер, участок инициации репликации, промоторную область и последовательность, кодирующую Gly-Ser спейсер, лигировали с помощью ДНК-лигазы фага Т4. Полученной лигированной смесью трансформировали клетки Е.coli M15 [pREP4] методом электропорации. Трансформированные клетки отбирали на агаризованной среде LB с антибиотиками ампициллином и канамицином. Плазмидную ДНК выделяли методом щелочного лизиса, анализировали с помощью рестриктаз BamHI и HindIII, а также рестриктаз, находящихся внутри клонируемых фрагментов, и отбирали клоны pspCBD (размером 4028 н.п.), плазмидная ДНК которых содержит последовательность участка гена celD со спейсером на N-конце, состоящим из Gly-Ser повторов. Первичную структуру полученной плазмиды подтверждали секвенированием.
Нуклеотидная последовательность CBD из гена celD приведена под №4 в перечне последовательностей.
Аминокислотная последовательность CBD из CelD приведена под №7.
е) Получение плазмиды, содержащей последовательность гена gal (lacZ), Gly-Ser спейсера, и целлюлозосвязывающего домена CBD.
Для получения плазмиды, содержащей последовательность гена gal(lacZ), Gly-Ser спейсера, и целлюлозосвязывающего домена CBD, плазмиду pLAC гидролизовали рестрикционными эндонуклеазами BglII и Bgll при 37°С в буфере 66 мМ Трис-ацетат (рН 7,9 при 37°С), 20 мМ ацетат магния, 132 мМ ацетат калия и 0,2 мг/мл BSA в течение 1 часа, а плазмиду pspCBD гидролизовали рестрикционными эндонуклеазами BamHI и Bgll при 37°С в буфере, содержащем 50 мМ Трис-HCl (рН 7,5 при 37°С), 10 мМ хлорида магния, 100 мМ хлорида натрия и 0,1 мг/мл BSA, в течение 1 часа. Выделенные из геля рестрикционные фрагменты: фрагмент ДНК плазмиды pLAC, включающий участок гена gal (lacZ) размером 3222 н.п., а также фрагмент ДНК плазмиды pspCBD, включающий Gly-Ser спейсер и последовательность целлюлозосвязывающего домена CBD, размером 3011 н.п., лигировали с помощью ДНК-лигазы бактериофага Т4. Полученной лигированной смесью трансформировали клетки Е.coli M15 [pREP4] методом электропорации. Трансформированные клетки отбирали на агаризованной среде LB с антибиотиками ампициллином и канамицином. Плазмидную ДНК выделяли методом щелочного лизиса, анализировали с помощью рестриктаз BamHI, BglI а также рестриктаз, находящихся внутри клонируемых фрагментов, и отбирали клоны pLACspCBD (размером 6233 н.п.), плазмидная ДНК которых содержит последовательность гена gal (lacZ), Gly-Ser спейсера, и целлюлозосвязывающего домена CBD. Первичную структуру полученной плазмиды подтверждали секвенированием. Нуклеотидная последовательность рекомбинантной плазмидной ДНК pLACspCBD приведена в перечне последовательностей под №1.
Пример 2. Получение штамма Е.coli - продуцента рекомбинантного белка LACspCBD, состоящего из бета-галактозидазы, Gly-Ser спейсера, целлюлозосвязывающего домена.
Для получения штамма Е.coli - продуцента рекомбинантного белка LACspCBD, клетки штамма Е.coli M15 [pREP4] трансформировали плазмидой pLACspCBD. Трансформированные клетки выращивали в 500 мл среды LB при 37°С до оптической плотности, соответствующей 1 ед. поглощения при длине волны 600 нм, индуцировали 150 мкл 0,1 М раствора изопропил-бета-О-тиогалактопиранозида и выращивали в течение 8 часов. Для контроля продукции рекомбинантного белка LACspCBD клетками Е.coli M15 [pREP4, pLACspCBD] применяли метод электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия. Разделение белков проводили в 12% полиакриламидном геле в стандартной системе буферов (электродный буфер: 25 мМ Tris-HCl, 192 мМ глицин, 0,1% додецилсульфат натрия, рН 8,3; буфер для геля: 375 мМ трис-хлоридный буфер, рН 8,8). По окончании электрофореза гели окрашивали 0,15% раствором Кумасси G250 в 25% изопропаноле и 10% уксусной кислоте и отмывали в 10% уксусной кислоте. При сравнении спектра белков у штаммов Е.coli M15 [pREP4] и Е.coli M15 [pREP4, pLACspCBD] обнаруживали появление дополнительной белковой полосы с молекулярной массой 109 кДа, что соответствует молекулярной массе рекомбинантного белка LACspCBD.
Пример 3. Определение бета-галактозидазной активности рекомбинантного белка в лизатах клеток штамма Е.coli M15 [pREP4, pLACspCBD].
Клетки Е.coli M15 [pREP4], трансформированные плазмидой pLACspCBD, выращивали в 3,5 мл среды LB с ампициллином и канамицином при 37°С до оптической плотности, соответствующей 1 ед. поглощения при длине волны 600 нм, добавляли 3 мкл 0,1 М раствора изопропил-бета-D-тиогалактопиранозида и выращивали в течение 4 ч. Культуру разводили до оптической плотности 0,7 при 530 нм, отбирали 400 мкл суспензии, осаждали клетки центрифугированием при 5000 об/мин в течение 10 мин. Клетки ресуспендировали в 500 мкл фосфатно-цитратного буфера (ФЦБ, 50 мМ, рН 6.0), лизировали ультразвуком, клеточный дебрис удаляли центрифугированием при 16000 об/мин в течение 10 мин.
В полученных лизатах определяли активность бета-галактозидазы по способности гидролизовать пара-нитро-фенил-бета-D-галактозид (ПНФГ) с образованием пара-нитро-фенила (ПНФ) и галактозы. За единицу активности принимали количество мкМ ПНФ, образующегося за 1 мин.
Для определения активности бета-галактозидазы к 0,4 мл 0,04 М раствора ПНФГ добавляли 1 мл ФЦБ, 100 мкл лизата клеток и инкубировали реакционную смесь при 70°С в течение 15 мин. Останавливали реакцию добавлением 1,0 мл раствора гидрокарбоната натрия (1 М). Концентрацию ПНФ определяли спектрофотометрически при 400 нм.
Активность бета-галактозидазы в экстрактах из клеток Е.coli M15 [pREP4, pLACspCBD] составила 5 ед/мг белка при 70°С, рН 6,0.
Пример 4. Метод получения препарата белка LACspCBD иммобилизованного на целлюлозе и раствора белка LACspCBD.
В культуру клеток Е.coli M15 [pREP4, pLACspCBD], выросших в L-бульоне с ампициллином и канамицином при 37°С до оптической плотности, соответствующей 1 ед. поглощения при длине волны 600 нм, добавляли изопропил-бета-D-тиогалактопиранозид до концентрации 0,2 мМ и выращивали в течение 8 часов. Клетки осаждали центрифугированием при 7000 об/мин в течение 20 мин, ресуспендировали в растворе, содержащем 1% тритона Х-100 и 1 мкг/мл лизоцима, инкубировали при 4°С в течение 30 мин и разрушали ультразвуком; клеточный дебрис удаляли центрифугированием при 16000 об/мин в течение 20 мин.
Для иммобилизации белка LACspCBD к супернатанту добавляли 2 мл гранулированной целлюлозы. Целлюлозу со связавшимся белком 5 раз отмывали от клеточного лизата в 10-кратном объеме 1% раствора тритона X-100.
Для получения раствора белка LACspCBD целлюлозу с иммобилизованным белком вносили в колонку, снимали белок 100% формамидом и диализовали в течение ночи против раствора, содержащего 10 мМ трис-HCl, рН 7,0 и 150 мМ NaCl.
Для получения термостабильной бета-галактозидазы, не содержащей целлюлозосвязывающего домена, проводили выделение белка из бактерий Е.coli, трансформированных плазмидой pVT1.
По результатам электрофореза в ПААГ в присутствии ДСН концентрация белка составляла 0,8-1,0 мг на 1 мл сорбента для иммобилизованного белка LACspCBD, 1,0-2,0 мг/мл для раствора белка LACspCBD и 1,0-2,0 мг/мл для раствора термостабильной бета-галактозидазы, не содержащей целлюлозосвязывающего домена. Степень чистоты всех препаратов составляла 95%. Удельная активность бета-галактозидазы препаратов иммобилизованного слитного белка, раствора слитного белка и раствора термостабильной бета-галактозидазы составляла соответственно 100000,150000 и 200000 ед/мл.
Для трех ферментных препаратов: иммобилизованного препарата белка LACspCBD, раствора белка LACspCBD и раствора термостабильной бета-галактозидазы, не содержащей целлюлозосвязывающего домена, определяли константу Михаэлиса, температурный оптимум, оптимальные значения рН и действие на активность бета-галактозидазы галактозы, ионов Са++, Mg++, Mn++, дитиотрейтола и меркаптоэтанола.
Константа Михаэлиса полученного иммобилизованного препарата белка LACspCBD на лактозе составляла 29 мМ (в диапазоне концентраций лактозы 1-80 мМ) при температуре 75°С, рН 5,7, что соответствует константам Михаэлиса препаратов раствора белка LACspCBD и бета-галактозидазы, не содержащей целлюлозосвязывающего домена.
Температурный оптимум иммобилизованного препарата фермента LACspCBD и препарата фермента LACspCBD в растворе составляет 65-70°С, что несколько ниже температурного оптимума термостабильной бета-галактозидазы, не содержащей целлюлозосвязывающего домена, соответствующего 75-80°С. При 85°С в течение 1 часа сохраняется соответственно 25% и 35% активности в обоих (иммобилизованном и неиммобилизованном) ферментных препаратах LACspCBD и в препарате бета-галактозидазы без целлюлозосвязывающего домена.
Оптимальные значения рН для всех трех ферментных препаратов одинаковы и составляют 5,7-6,0 (ФЦБ, 75°С) на субстрате ПНФГ.
Для выявления температурного и рН-оптимума определение активности ферментов проводили при различных температурах и оптимальной величине рН (6,0) или при различных значениях рН и оптимальной температуре (65 и 75°С для бета-галактозидаз с целлюлозосвязывающим доменом и без него) соответственно.
Ингибирующим действием на активность всех трех препаратов бета-галактозидазы обладает галактоза в концентрации 5% в ФЦБ, ионы Са++ и Mg++ в концентрации 5-10 мМ не оказывали ингибирующего действия на активность бета-галактозидазы, ионы Mn++ (10 мМ) оказывали небольшое активирующее действие (125% активности) (условия те же). Дитиотрейтол и меркаптоэтанол в концентрации 5-10 мМ не ингибировали бета-галактозидазу.
Для характеристики целлюлозосвязывающих свойств белка LACspCBD через колонку с иммобилизованным белком LACspCBD пропускали по 100 объемов колонки буферных растворов с различными значениями рН и разной концентрацией NaCl при различных температурах. После чего анализировали концентрацию иммобилизованного белка и активность бета-галактозидазы. Белок LACspCBD проявляет способность к необратимому связыванию с целлюлозой (не смывается с сорбента) в диапазоне значений рН 4-11, температуры 0-75°С и концентраций соли 0-5 М NaCl. После протекания через колонку буферов с различными значениями рН и разной концентрацией NaCI при различных температурах иммобилизованный белок LACspCBD сохраняет 100% бета-галактозидазной активности при оптимальных условиях.
Пример 5. Применение иммобилизованного на целлюлозе химерного белка LACspCBD для гидролиза лактозы.
500 мкл иммобилизованного на целлюлозе химерного белка LACspCBD внесли в термостатируемую хроматографическую колонку с внутренним диаметром 4,6 мм и высотой слоя сорбента 7 мм. Через колонку пропускали раствор субстрата, состоящего из 10 мМ ацетата аммония (рН 6,5) и лактозы в различных концентрациях. Степень расщепления лактозы определяли хроматографически с использованием прямофазового метода ВЭЖХ при различных исходных концентрациях лактозы и различных температурах. При 70°С реактор, полученный на основе белка LACspCBD, гидролизует 70% исходного раствора лактозы с концентрацией 150 мМ (5%) (что соответствует концентрации лактозы в коровьем молоке) при скорости потока раствора субстрата 1,2 объема колонки в минуту. 90% исходного раствора лактозы в тех же условиях гидролизуется при скорости потока раствора субстрата 0,64 объема колонки в минуту. Пропускание через реактор 10000 объемов колонки при скорости потока 1 объем колонки в минуту и температуре 70°С не приводит к снижению эффективности расщепления лактозы. Хранение реактора при комнатной температуре в течение 6 месяцев не приводит к изменению параметров реактора.
Перечень последовательностей приведен в конце описания.




| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНАЯ ПЛАЗМИДНАЯ ДНК, КОДИРУЮЩАЯ СИНТЕЗ РЕКОМБИНАНТНОГО БЕЛКА TUL4spCBD, ШТАММ ESCHERICHIA COLI M15 [pREP4, pTUL4spCBD] - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО БЕЛКА TUL4spCBD, РЕКОМБИНАНТНЫЙ БЕЛОК TUL4spCBD И СПОСОБ ЕГО ПОЛУЧЕНИЯ, СПОСОБ ПОЛУЧЕНИЯ СПЕЦИФИЧЕСКИХ АНТИТЕЛ К БЕЛКУ TUL4spCBD | 2004 |
|
RU2270249C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА (ВАРИАНТЫ), ШТАММ ESCHERICHIA COLI (ВАРИАНТЫ) - ПРОДУЦЕНТ ХИМЕРНЫХ БЕЛКОВ, ХИМЕРНЫЙ БЕЛОК (ВАРИАНТЫ), СПОСОБ ИММОБИЛИЗАЦИИ, КОНЦЕНТРИРОВАНИЯ И ОЧИСТКИ РЕКОМБИНАНТНЫХ БЕЛКОВ НА ЦЕЛЛЮЛОЗЕ, СПОСОБ ИММОБИЛИЗАЦИИ РЕКОМБИНАНТНЫХ БЕЛКОВ НА ПОЛИСТИРОЛЬНЫХ НОСИТЕЛЯХ | 2008 |
|
RU2378371C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА, ШТАММ Escherichia coli, ХИМЕРНЫЙ БЕЛОК И ИХ ПРИМЕНЕНИЕ | 2010 |
|
RU2422524C1 |
| РЕКОМБИНАНТНЫЙ БЕЛОК Collbd-CBD, РЕКОМБИНАНТНАЯ ПЛАЗМИДА pОС-Collbd, ШТАММ Escherichia coli - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО БЕЛКА Collbd-CBD, СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО БЕЛКА Collbd-CBD | 2009 |
|
RU2408726C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА, ШТАММ Escherichia coli, ХИМЕРНЫЙ БЕЛОК И ИХ ПРИМЕНЕНИЕ | 2010 |
|
RU2422525C1 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pESAT6-CFP10-DBD, РЕКОМБИНАНТНЫЙ ШТАММ Escherichia coli M15 [pREP4, pESAT6-CFP10-DBD], СПОСОБ ПОЛУЧЕНИЯ, ИММОБИЛИЗАЦИИ, КОНЦЕНТРИРОВАНИЯ И ОЧИСТКИ РЕКОМБИНАНТНОГО БЕЛКА ESAT6-CFP10-DBD НА ДЕКСТРАНЕ, РЕКОМБИНАНТНЫЙ БЕЛОК ESAT6-CFP10-DBD И ИММУНОГЕННАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ БЕЛОК ESAT6-CFP10-DBD | 2013 |
|
RU2539026C1 |
| РЕКОМБИНАНТНАЯ БЕЛКОВАЯ КОНСТРУКЦИЯ ДСД-сп-β-ГАЛ, ОБЛАДАЮЩАЯ АКТИВНОСТЬЮ ФЕРМЕНТА ТЕРМОСТАБИЛЬНОЙ β-ГАЛАКТОЗИДАЗЫ (ЛАКТАЗЫ) И СПОСОБНОСТЬЮ АФФИННО СВЯЗЫВАТЬСЯ С ДЕКСТРАНОМ, ПЛАЗМИДНАЯ ДНК pGD-10, ОПРЕДЕЛЯЮЩАЯ БИОСИНТЕЗ ДСД-сп-β-ГАЛ, И ШТАММ-ПРОДУЦЕНТ Escherichia coli DH5α/PGD-10 | 2009 |
|
RU2428477C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pD1spGBD, ШТАММ Escherichia coli - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО БЕЛКА D1-GBD, РЕКОМБИНАНТНЫЙ БЕЛОК D1-GBD И СПОСОБ ЕГО ПОЛУЧЕНИЯ, СПОСОБ ИССЛЕДОВАНИЯ СВЯЗЫВАНИЯ БЕЛКА D1-GBD С АНТИТЕЛАМИ СЫВОРОТОК БОЛЬНЫХ, СПОСОБ ПОЛУЧЕНИЯ СПЕЦИФИЧЕСКИХ АНТИТЕЛ К БЕЛКУ D1-GBD | 2008 |
|
RU2401302C2 |
| РЕКОМБИНАНТНАЯ ПЛАЗМИДА pD4spGBD, ШТАММ Escherichia coli - ПРОДУЦЕНТ РЕКОМБИНАНТНОГО БЕЛКА D4-GBD, РЕКОМБИНАНТНЫЙ БЕЛОК D4-GBD И СПОСОБ ЕГО ПОЛУЧЕНИЯ, СПОСОБ ИССЛЕДОВАНИЯ СВЯЗЫВАНИЯ БЕЛКА D4-GBD С АНТИТЕЛАМИ СЫВОРОТОК БОЛЬНЫХ, СПОСОБ ПОЛУЧЕНИЯ СПЕЦИФИЧЕСКИХ АНТИТЕЛ К БЕЛКУ D4-GBD | 2008 |
|
RU2401305C2 |
| СПОСОБ ПОЛУЧЕНИЯ ИММУНОГЕННОЙ КОМПОЗИЦИИ НА ОСНОВЕ ГИБРИДНОГО БЕЛКА CFP10-DBD И ДЕКСТРАНА, РЕКОМБИНАНТНАЯ ПЛАЗМИДА pCFP10-DBD, ШТАММ Escherichia coli [pREP4, pCFP10-DBD], ХИМЕРНЫЙ БЕЛОК CFP10-DBD И ИХ ПРИМЕНЕНИЕ | 2013 |
|
RU2546875C1 |
Изобретение относится к области генной инженерии и биотехнологии и может быть использовано в пищевой промышленности. Предложен слитый белок бета-галактозидазы и целлюлозосвязывающего домена эндоглюканазы CelD из Anaerocellum thermophilium. Сконструирована рекомбинантная плазмидная ДНК pLACspCBD, обеспечивающая его экспрессию в клетках Е.coli. Получен штамм Е.coli - продуцент указанного слитого белка. Разработан способ получения данного слитого белка в иммобилизованном виде путем обработки целлюлозы гидролизатом указанного штамма Е.coli. Иммобилизованный слитый белок используют для ферментативного расщепления лактозы. Применение изобретения позволяет упростить переработку молочных продуктов. 5 н.п. ф-лы, 1 ил.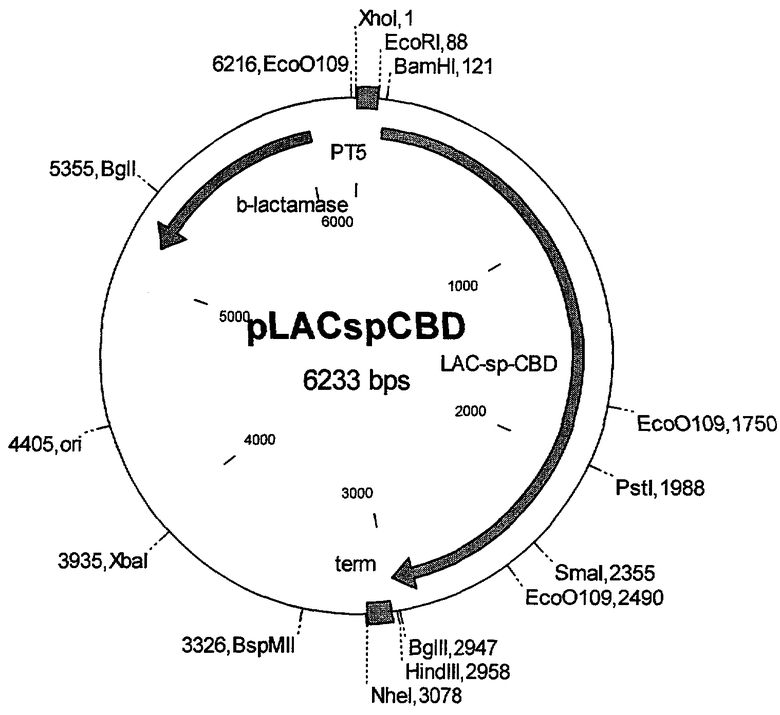
искусственный бактериальный оперон слитого белка бета-галактозидазы Thermoanaerobacter ethanolicus и целлюлозосвязывающего домена эндоглюканазы CelD Anaerocellum thermophilium, включающий промоторную область раннего промотора бактериофага Т5 (7-87 н.п.), нуклеотидную последовательность, кодирующую указанный слитый белок (115-2958 н.п.) и терминатор транскрипции (2979-3073 н.п.);
бактериальный оперон бета-лактамазы (6028-5168 н.п. комплементарной цепи);
бактериальный участок инициации репликации типа ColEl, обеспечивающий репликацию плазмидной ДНК в клетках Е.coli (4405 н.п.).
| US 5137819, 11.08.1992 | |||
| ENZYME MICROB TECHNOL | |||
| Циркуль-угломер | 1920 |
|
SU1991A1 |
| ENDOGLUCANASE [ANAEROCELLUM THERMOPHILUM], CAB01405.1 G[:1483210 accession Z 77855.1, 05.08.1996 | |||
| Приспособление для кидки челнока в ткацких станках | 1926 |
|
SU5182A1 |
| PROTEIN ENG | |||
| Пуговица для прикрепления ее к материи без пришивки | 1921 |
|
SU1992A1 |
| Способ производства сиропа гидролизованной лактозы из молочной сыворотки | 1990 |
|
SU1739951A1 |
| US 5670623, 23.09.1997 | |||
| WO 03070928, 28.08.2003. | |||

