Изобретение в области биотехнологии относится к микробиологической промышленности и представляет собой способ получения ферментных препаратов с карбогидразной активностью путем культивирования мультикопийных штаммов мицелиальных грибов рода Penicillium verruculosum, трансформированных фрагментами ДНК с клонированными гомологичными и гетерологичными генами.
Энергетические потребности человечества на настоящем этапе его развития требуют привлечения возобновляемых источников энергии, в первую очередь растительной биомассы. Основа процесса биоконверсии растительной биомассы заключается в ферментативном гидролизе целлюлозы и гемицеллюлозы до сахаров с их последующей переработкой путем сбраживания в этанол или превращения в другие продукты микробного синтеза.
В глобальном процессе разложения растительной биомассы основную роль выполняют бактерии и мицелиальные грибы. Глубокая деструкция целлюлозы и гемицеллюлозы осуществляется под действием полиферментных систем целлюлаз и гемицеллюлаз (ксиланаз). В настоящее время во многих странах ведется поиск новых грибных продуцентов целлюлаз и гемицеллюлаз. Наибольшее распространение получили грибные продуценты, представители рода Trichoderma, которые обладают высокой секреторной способностью. В свою очередь грибы рода Penicillium синтезируют ферментные комплексы целлюлаз и гемицеллюлаз более сбалансированного состава и эффективнее расщепляют целлюлозу и содержащее ее растительное сырье. При этом индивидуальные ферменты Penicillium обладают высокой каталитической активностью и операционной стабильностью.
В последнее десятилетие получили широкое распространение мультикопийные грибные суперпродуценты ферментов, у которых амплификация генов, кодирующих секретируемые ферменты, позволила резко увеличить продуктивность штаммов микроорганизмов для получения дешевых промышленных ферментов. К известным мультикопийным продуцентам эндо-β1,4-глюканаз относятся штаммы на основе Trichoderma longibrachiatum [Clarkson et al., 1995, US Patent 5,419,778], Tr.reesei [Saloheimo et al., 1994, WO Patent 94/28117] и Humicola insolens [Rasmussen et al., 1991, Patent WO 91/17243]. Известны различные грибные мультикопийные продуценты ксиланаз - Aspergillus niger [van den Broek et al., 1994, US Patent 5,358,864], Tr.reesei [Nevalainen et al., 1994, US Patent 5,298,405], Thermoascus auranticus [Yu et al., 1990, US Patent 4,966,850], H.insolens [Schulein et al., 1997, US Patent 5,610,048].
Из грибов рода Penicillium большой интерес представляют Penicillium verruculosum.
Прототипом разработанного технического решения может служить патент РФ №2288267 '"Способ получения кормового комплексного ферментного препарата (варианты) и штамм Penicillium canescens (варианты)", в котором охарактеризован способ получения препаратов с высокой эндоглюканазной, ксиланазной, фитазной и пектин-лиазной активностью при ферментации мультикопийных штаммов P.canescens, в которых у включенных копий генов регуляторная область была представлена промоторами либо гена β-галактозидазы или гена ксиланазы P.canescens.
Техническая задача, решаемая группой разработанных технических решений, состоит в разработке способа получения ферментных препаратов с различными типами целлюлазной и гемицеллюлазной (ксиланазной) активности при ферментации мультикопийных штаммов, полученных при трансформации реципиентных высокопродуктивных штаммов P.verruculosum клонированными участками ДНК с гомологичными и гетерологичными генами, кодирующими ферменты целлюлазы и гемицеллюлазы (ксиланазы).
Технический результат, получаемый при реализации разработанных технических решений, состоит в получении промышленных ферментных препаратов с высокой активностью индивидуальных ферментов целлюлаз и гемицеллюлаз (ксиланаз) при ферментации новых мультикопийных штаммов грибов рода Penicillium, полученных с привлечением технологии рекомбинантных молекул ДНК.
Для получения указанного технического результата предложено использовать генетическую конструкцию для обеспечения экспрессии целевых гомологичных и гетерологичных генов в клетках мицелиального гриба P.verruculosum, используемого в качестве хозяина, содержащую целевую кодирующую последовательность, функционально связанную с регуляторными элементами гена целлобиогидролазы I P.verruculosum, которые представляют собой промотор, сигнальный пептид и терминатор гена целлобиогидролазы I.
Также для получения указанного технического результата предложено использовать способ получения штамма гриба P.verruculosum - продуцента фермента, кодируемого гомологичной или гетерологичной нуклеотидной последовательностью, предусматривающей трансформацию клетки гриба P.verruculosum генетической конструкцией, содержащей целевую кодирующую последовательность, функционально связанную с регуляторными элементами гена целлобиогидролазы I P.verruculosum, которые представляют собой промотор, сигнальный пептид и терминатор гена целлобиогидролазы I.
Кроме того, для получения указанного технического результата предложено использовать способ получения ферментного препарата, включающего ферменты целлюлазы и гемицеллюлазы (ксиланазы), путем культивирования микроорганизмов, причем указанным выше способом получают штамм гриба P.verruculosum, осуществляют культивирование полученного штамма и выделяют целевой продукт из культуральной жидкости гриба. В первом варианте реализации указанного способа для получения ферментного препарата получают штамм P.verruculosum PV-Cbh1P (BKM F-3973D), мультикопийный по гомологичному гену cbhI - целлобиогидролазы I P.verruculosum. Во втором варианте реализации указанного способа для получения ферментного препарата получают штамм P.verruculosum PV-Egl4T (BKM F-3976D), мультикопийный по гетерологичному гену eglIV - эндоглюканазы IV Tr.longibrachiatum. В третьем варианте реализации указанного способа для получения ферментного препарата получают штамм P.verruculosum PV-Xyl3T (BKM F-3979D), мультикопийный по гетерологичному гену xylIII - ксиланазы III Tr.longibrachiatum.
Указанные варианты не исчерпывают возможности разработанного способа.
Схема разработки способа получения каждого из ферментных препаратов, обладающего одной из целлюлазной или ксиланазной активностей, складывется из выполнения одного подготовительного и трех типовых этапов.
Подготовительный этап.Выделяют ген, кодирующий целлобиогидролазу I P.verruculosum и конструируют экспрессионный вектор для трансформации клеток P.verruculosum, в котором ген целевого белка сопряжен с регуляторными последовательностями (промотор и терминатор) гена, кодирующего целлобиогидролазу I Р.verruculosum.
Этап 1. Путем клонирования в фаговом векторе или ПЦР-синтезом получают фрагмент ДНК, кодирующий ген каждого из перечисленных ферментов с полной регуляторной областью или только структурную часть последовательности, кодирующую зрелый белок. Включают этот фрагмент ДНК в структуру векторной молекулы. Получают экспрессионную плазмиду, в которой нуклеотидная последовательность структурной части целевого гена функционально совмещена с нуклеотидной последовательностью, кодирующей промоторную область и сигнальный пептид гена cbhI целлобиогидролазы I P.verruculosum (или линейный фрагмент ДНК, содержащий нуклеотидную последовательность гена и указанные выше регуляторные области).
Этап 2. Каждой экспрессионной плазмидой с включенным геном (или фрагментом ДНК в линейной форме) проводят трансформацию реципиентного высокопродуктивного штамма P.verruculosum 537(niaD-) в условиях котрансформации вместе с плазмидой (или фрагментом ДНК в линейной форме), несущей последовательность niaD гена, как маркера селекции, и осуществляют отбор трансформантов, секретирующих в культуральную жидкость искомую активность. Проводят ферментацию отобранных трансформантов в качалочных колбах и среди них выбирают наиболее продуктивный вариант(ы) штамма-продуцента.
Этап 3. Проводят процесс ферментации отобранного наиболее активного варианта штамма-продуцента в ферментерах, получают и характеризуют ферментные препараты с высокой активностью целевого фермента.
Изобретение иллюстрируется следующими примерами.
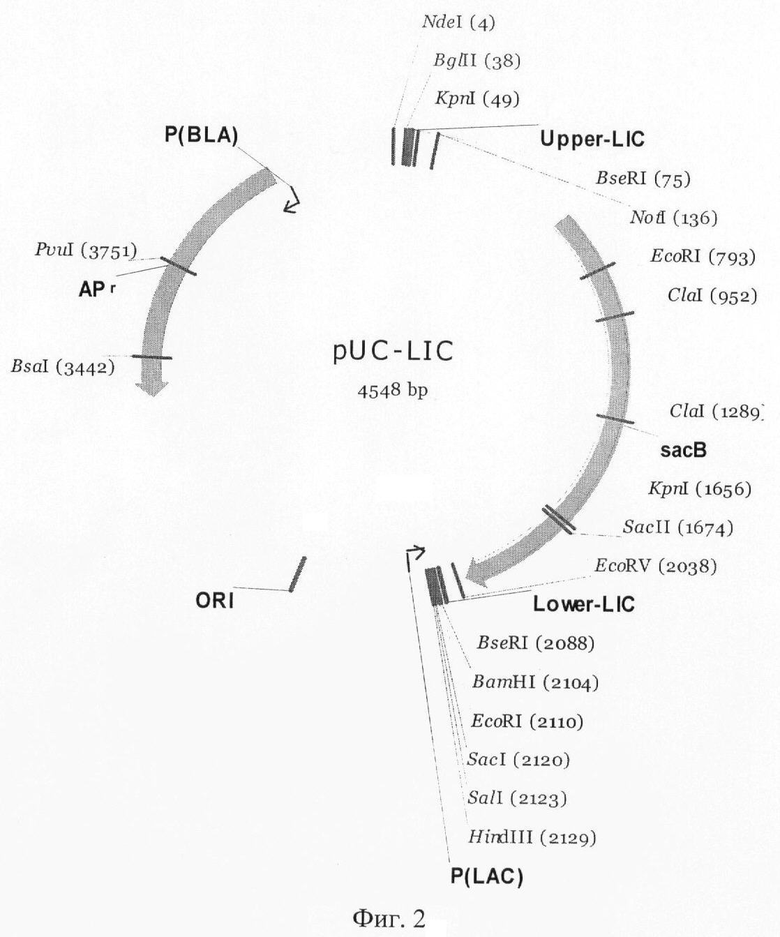
Пример 1. Выделение и секвенирование фрагмента ДНК, несущего cbhI ген целлобиогидролазы P.verruculosum (целлобиогидролаза I 7-го семейства гликозид-гидролаз, молекулярная масса 66 кДа), и получение векторной плазмиды pUC-LIC на основе структурных элементов этого гена.
На базе гомологии аминокислотных и нуклеотидных последовательностей грибных целлобиогидролаз семейства 7 глюкозид-гидролаз синтезируют два олигонуклеотидных праймера:
(1) 5'-ACCAACTGCTACACCGGCAACACCTGG-3'
(2) 5'CGGTAAGGGTTGAA(G/A)TCACA(G/A)TC(G/A)CAICCATC-3',
где I - инозин.
Далее с помощью ПЦР полимеразной цепной реакции (ПЦР) синтезируют внутренний фрагмент ДНК гена целлобиогидролазы I размером 660 п.н. В качестве матрицы для ПЦР используют геномную ДНК штамма P.verruculosum, выделенную из мицелия. Нуклеотидную последовательность полученного фрагмента определяют методом Сэнгера по обеим цепям. Сравнительный анализ нуклеотидных последовательностей ДНК пробы и генов грибных целлобиогидролаз показывает, что максимальная гомология между последовательностью выделенной ДНК пробы и последовательностями целлобиогидролаз из P.funiculosum и P.occitanis составляет 90% и 89% соответственно, что позволяет отнести полученную последовательность к фрагменту целлобиогидролазы I семейства 7 глюкозид-гидролаз.
Для получения полноразмерного гена целлобиогидролазы I P.verruculosum применяют метод инверсного ПЦР. Для этого геномную ДНК P.verruculosum обрабатывают рестриктазами EcoRI, SalI, XbaI, PstI и циркулизируют при помощи Т4 ДНК лигазы, что являлось матрицей для инверсного ПЦР. Для амплифицирования оставшихся участков гена целлобиогидролазы используют два противоположно направленных олигонуклеотидных праймера.
(3) 5'-AACAACTCCAACACCGGAATCGGCAACCAC-3'
(4) 5'-AACCGGTGACGAAGTTGAGACGCAACGAGT-3'
На ДНК-матрице, полученной при гидролизе геномной ДНК с использованием рестриктазы EcoRI максимальная величина амплифицированного ПЦР фрагмента достигает 2000 п.н. Нуклеотидную последовательность ДНК гена определяют по обеим цепям методом Сэнгера. Ген целлобиогидролазы I P.verruculosum состоит из 1575 п.н. и не содержит интронов. Транслированная аминокислотная последовательность целлобиогидролазы I включает в себя 525 аминокислот.
Чтобы определить структуру промотора и терминатора гена целлобиогидролазы I применяют метод инверсного ПЦР. Используют праймеры (3) и (4), а также синтезируют новые праймеры (5), (6) и (7) для секвенирования ДНК промоторной части гена в 5'-нетранслируемой области:
(5) 5'-GATAATGTCTGATTCTTCAAGGGAA-3'
(6) 5'-GCTTCAGCCTCTGCTCTCCA-3'
(7) 5'-GACTGGAGACGAGCTGACAGCA-3'
В качестве матрицы используют лигированную геномную ДНК, предварительно обработанную эндонуклеазой рестрикции HindIII. Амплифицированный ПЦР фрагмент имеет размер в 3500 п.н. и его структуру определяют методом Сэнгера по обеим цепям при использовани праймеров для амплификации. Промоторный район из 1303 п.н. содержит структурные элементы, свойственные промоторам эукариотических генов - ТАТА бокс (-90 п.н. от стартового кодона), транскрипционную область (-65 п.н. от стартового кодона) и CreI участок (-492 п.н. от стартового кодона). Аналогичным способом изучают последовательность гена целлобиогидролазы в 3'-нетранслируемой области, что позволило идентифицировать ее как терминаторную часть гена. Общая структура cbhI гена P.verruculosum - промотора, участка гена cbhI, кодирующего сигнальный пептид и зрелый белок целлобиогидролазы I, и терминатора представлена на фиг.1.
Для облегчения дальнейших шагов по клонированию гомологичных и гетерологичных белков штамма-продуцента P.verruculosum конструируют универсальный вектор на основе двух известных плазмид pUC19 и pLIC-SGC. NdeI-HindIII фрагмент плазмиды pLIC выделяют и клонируют в плазмиду pUC19, разрезанную по тем же сайтам рестрикции. Полученный вектор был назван pUC-LIC. Линеаризацию плазмиды pUC-LIC проводят путем разрезания BseRI, что приводит к экспонированию двух сайтов для лигирования ПЦР фрагментов - 5'-ТАСТТССААТССАТ-3' и 5'-AGTAAAGGTGGAT-3'. После обработки вектора и клонируемой вставки Т4 ДНК полимеразой в условиях проявления ее эндонуклеазной активности (в смеси только с одним нуклеотидом) на векторе и на вставке образуются два комплементарных одноцепочечных участка, которые соответственно лигируются. Также в результате линеаризации вектора pUC-LIC из исходного вектора удаляется ген SacB, что обеспечивает дополнительную селекцию колоний на чашках Петри с сахарозой после клонирования в pUC-LIC. Схематическое изображение вектора pUC-LIC показано на фиг.2.
Пример 2. Получение ферментных препаратов с увеличенной целлобиогидролазной активностью при ферментации штаммов P.verruculosum, мультикопийных по cbhI гену P.verruculosum (ген целлобиогидролазы I, 7-е семейство гликозид-гидролаз, молекулярная масса 66 кДа).
На первом этапе получают экспрессионную плазмиду pCBH1-12, в которой клонированная нуклеотидная последовательность полного cbhI гена P.verruculosum соединена с гомологичными районами промоторной области, сигнального пептида и терминаторной области. Для этой цели синтезируют следующие олигонуклеотиды:
(8) 5'-TACTTCCAATCCATGAAGCTTGGAAACGACGCCGACA-3'
(9) 5'-TATCCACCTTTACTGCGGCCGCATCCATTCAATCTCGTATG-3'
Полученный ПЦР фрагмент клонируют в вектор pUC-LIC по указанным выше сайтам лигирования (см. пример 1). Таким образом, экспрессионная плазмида pCBH1-12 содержит все регуляторные части для экспрессии гомологичного гена целлобиогидролазы I. Для возможности линеаризации экспрессионной плазмиды перед трансформацией в штамм реципиент между сайтом лигирования и вектором вводят рестрикционный сайт NotI.
Реципиентный штамм P.verruculosum 537(niaD-) получают путем отбора клонов, дефектных по нитрат-редуктазе и не способных расти на минимальной среде, где в качестве единственного источника азота используется нитрат натрия - NaNO3.
На втором этапе проводят котрансформацию реципиентного штамма P.verruculosum 537(niaD-) 10-20 мкг экспрессионной векторной плазмиды pCBH1-12 и 1-3 мкг маркерной плазмиды pSTA (niaD+) и осуществляют скрининг клонов на наличие трансформантов с повышенной секрецией целлобиогидролазы.
Полученные niaD+ трансформанты пересевают на чашки с минимальной средой и источником азота NaNO3 и инкубируют при 30°С в течение 140 часов до образования конидий. Затем трансформанты ферментируют в качалочных колбах, используя водную ферментационную среду следующего состава, в г/л: микрокристаллическая целлюлоза (МКЦ) - 40, пшеничные отруби - 10, дрожжевой экстракт - 10, (NH4)2SO4 - 5, KH2PO4 - 5, MgSO4×7H2O - 0,3, CaCl2×2H2O - 0,3, рН 4,5. Колбы инкубируют на качалке при 32°С и 200 об/мин в течение 144 ч. Далее отбирают трансформанты с увеличенной по сравнению с реципиентным штаммом P.verruculosum целлобиогидролазной активностью. Целлобиогидролазную активность в культуральной жидкости измеряют по гидролизу МКЦ.
При анализе в ДДС-электрофорезе в денатурирующих условиях обнаружено увеличение размера полосы, соответствующей зоне, занимаемой целлобиогидролазой I (66 кДа).
На третьем этапе проводят ферментацию наиболее продуктивного из трансформированных штамма P.verruculosum PV-Cbh1P (BKM F-3973D) в 10-литровом ферментере, используя питательную среду того же состава, что в качалочных колбах, и проводя подпитку глюкозой, при рН 4,5 и 32°С в течение 144 часов. После окончания ферментации с помощью ультрафильтрации культуральной жидкости на полых волокнах (с пределом отсечения 10 kDа) и последующего лиофильного высушивания получают сухие ферментные препараты и характеризуют их по уровню целлобиогидролазной активности.
Активность целлобиогидролзы в культуральной жидкости наиболее активного трансформанта P.verruculosum PV-Cbh1P (BKM F-3973D) составлялет 35 ед/мл, активность целлобиогидролзы в сухом препарате - 1200 ед/г.
Целлобиогидролаза I обладает высокой удельной гидролитической (осахаривающей) активностью по отношению к нерастворимым кристаллическим целлюлозным субстратам (см. таблицу).
Пример 3. Получение ферментных препаратов с эндоглюканазной активностью при ферментации штаммов P.verruculosum мультикопийных по eglIV гену Tr.longibrachiatum (ген эндо-β1,4-глюканазы IV, 61-е семейство гликозид-гидролаз, молекулярная масса 36 кДа).
На первом этапе получают фрагмент ДНК с нуклеотидной последовательнстью eglIV гена, кодирущей эндоглюканазу IV Tr.longibrachiatum и получают экспрессионную плазмиду pPrCBHI-EGIV_Trich. Для амплификации гена eglIV синтезируют пару праймеров:
(10) 5'-CAAACAGAAGCAACCGACACAATGATCCAGAAGCTTTCCAACCTC-3'
(11) 5'-GAGGAGAAGCCCGGTTAGTTAAGGCACTGGGCGTAGTAG-3'
Олигонуклеотиды (10) и (11) синтезируют для конструирования плазмиды pPrCBHI-EGIV_Trich. В качестве матрицы для первичных реакций ПЦР используют геномную ДНК Tr.longibrachiatum. Так как Tr.longibrachiatum является вариантом Tr.reesei, за основу для создания праймеров взята последовательность гена из кДНК эндоглюканазы IV из Tr.reesei, доступная в базе данных EMBL (AN:Y 11113). Нуклеотидная последовательность гена эндоглюканазы IV из Tr.longibrachiatum состоит из 1086 пар оснований и содержит 1 интрон (фиг.З)
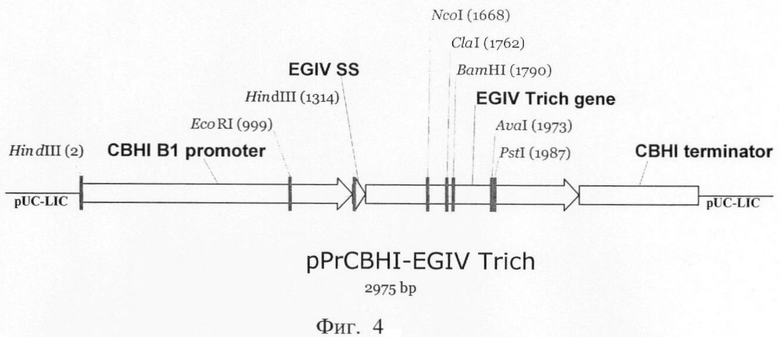
Плазмида pPrCBHI-EGIV_Trich содержит все структурные элементы, необходимые для экспрессии в P.verruculosum: cbhI промотор и cbhI терминатор. В данной конструкции клонируют полноразмерный ген эндоглюканазы IV Tr.longibrachiatum, включая последовательность, кодирующую сигнальный пептид.
Схематическая карта экспрессионной плазмиды pPrCBHI-EGIV_Trich представлена на фиг.4.
На втором этапе проводят котрансформацию реципиентного штамма P.verruculosum 537(niaD-) 10-20 мкг экспрессионной векторной плазмиды pPrCBHI-EGIV_Trich и 1-3 мкг маркерной плазмиды pSTA(niaD+) и осуществляют скрининг клонов на наличие трансформантов с повышенной секрецией эндоглюканазы.
Полученные niaD+ трансформанты P.verruculosum пересевают на чашки с минимальной средой и источником азота NaNO3, инкубируют при 30°С в течение 140 часов до образования конидий. Затем трансформанты P.verruculosum ферментируют в качалочных колбах, как описано в примере 2, и далее отбирают трансформанты с увеличенной по сравнению с реципиентным штаммом P.verruculosum эндоглюканазной активностью. Эндоглюканазную активность в культуральной жидкости измеряют по гидролизу Na-соли карбоксиметилцеллюлозы (КМЦ).
При анализе в ДДС-электрофорезе в денатурирующих условиях обнаружено, что эндоглюканаза IV занимает зону, соответствующую 36 кДа.
На третьем этапе проводят ферментацию наиболее продуктивного из трансформированных штамма P.verruculosum PV-Egl4T (BKM F-3976D) в 10-литровом ферментере (как описано в примере 2).
Активность эндоглюканазы в культуральной жидкости штамма P.verruculosum PV-Egl4T (BKM F-3976D) в конце ферментации составляет 850 ед/мл.
Сухой ферментный препарат получают с помощью ультрафильтрации культуральной жидкости на полых волокнах (с пределом отсечения 10 кДа) и последующим лиофильным высушиванием. Активность эндоглюканазы в сухом ферментном препарате трансформанта P.verruculosum PV-Egl4T (BKM F-3976D) составляет 6500 ед/г.
Пример 4. Получение ферментных препаратов с ксиланазной активностью при ферментации штаммов P.verruculosum мультикопийных по xynIII гену Tr.longibrachiatum (ген эндо-β1,4-ксиланазы III, 10-го семейства гликозид-гидролаз, молекулярная масса 30 кДа).
На первом этапе получают фрагмент ДНК с нуклеотидной последовательностью xynIII гена, кодирущей ксиланазу III Tr.longibrachiatum и экспрессионную плазмиду pPrCBHI-XylIII_Trich. Для амплификации гена xynIII синтезируют пару праймеров:
(36) 5'-CAAACAGAAGCAACCGACACAATGAAAGCAAACGTCATCTTGT-3'
(37) 5'-GAGGAGAAGCCCGGTTATTGTAAGATGCCAACAATGCT-3'
В качестве матрицы для первичных реакций ПЦР используют геномную ДНК Tr.longibrachiatum. Так как Tr.longibrachiatum является вариантом Tr.reesei, за основу для написания праймеров используют последовательность гена из кДНК ксиланазы III из Tr.reesei, доступную в базе данных GeneBank (AN:AB036796). Нуклеотидная последовательность ксиланазного гена из Tr.longibranchiatum состоит из 1349 пар оснований, содержит 2 интрона и полностью идентична последовательности ксиланазного гена из Tr.reesei (фиг.5).
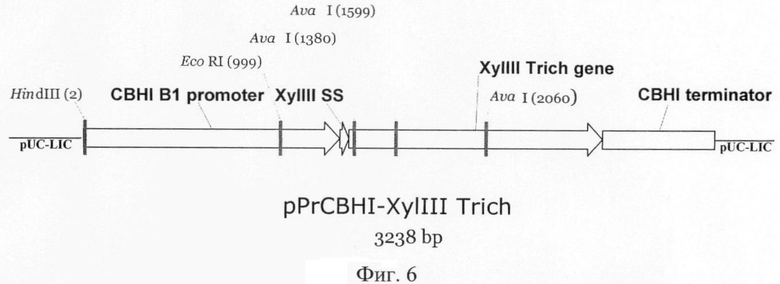
Плазмида pPrCBHI-XylIII_Trich содержит все структурные элементы, необходимые для экспрессии в P.verruclosum: cbhI промотор и cbhI терминатор. В данной конструкции клонируют полноразмерный ген ксиланазы III Tr.longibrachiatum, включая последовательность, кодирующую сигнальный пептид. Схематическая карта экспрессионной плазмиды pPrCBHI-XylIII_Trich представлена на фиг.6.
На втором этапе проводят котрансформацию штамма реципиентного P.verruculosum 537(niaD-) 10-20 мкг экспрессионной векторной плазмиды pPrCBHI-XylIII_Trich и 1-3 мкг маркерной плазмиды pSTA(niaD+) и осуществляют скрининг клонов на наличие трансформантов с повышенной секрецией ксиланазы.
Полученные niaD+ трансформанты P.verruculosum пересевают на чашки с минимальной средой и источником азота NaNO3, инкубируют при 30°С в течение 140 часов до образования конидий.
Затем трансформанты P.verruculosum ферментируют в качалочных колбах, как описано в примере 2, и далее отбирают трансформанты с увеличенной по сравнению с реципиентным штаммом P.verruculosum ксиланазной активностью. Ксиланазную активность в культуральной жидкости измеряют по гидролизу березового ксилана.
При анализе в ДДС-электрофорезе в денатурирующих условиях обнаружено, что ксиланаза III занимает зону, соответствующую 30 кДа.
На третьем этапе проводят ферментацию наиболее продуктивного из трансформированных штамма P.verruculosum PV-Xyl3T (ВКМ F-3979D) в 10-литровом ферментере (как описано в примере 2).
Ксиланазная активность в культуральной жидкости штамма P.verruculosum PV-Xyl3T (BKM F-3979D) в конце ферментации составляет 1950 ед/мл.
Сухой ферментный препарат получают с помощью ультрафильтрации культуральной жидкости на полых волокнах (с пределом отсечения 10 кДа) и последующим лиофильным высушиванием. Активность ксиланазы в сухом ферментном препарате трансформанта P.verruculosum PV-Xyl3T (В KM F-3979D) составляет 43600 ед/г.
Методы определения активности. Целлобиогидролазную (авицелазную) активность измеряют, проводя гидролиз МКЦ (5 мг/мл) при рН 5,0 (0,1 М ацетатный буфер) и 50°С в течение 60 минут. После этого определяют в реакционной смеси концентрацию восстанавливающих сахаров (ВС) методом Нельсона-Сомоджи. За единицу целлобиогидролазной активности принимают такое количество фермента, которое в течение 1 минуты при температуре 50°С и рН 5,0 при гидролизе суспензии МКЦ концентрацией 5 мг/мл освобождает количество ВС, эквивалентных 1 микромолю глюкозы, определяемых методом Сомоджи-Нельсона.
Эндоглюканазную (КМЦ-азную) активность измеряют, проводя гидролиз КМЦ (5 мг/мл) при рН 5,0 (0,1 М ацетатный буфер) и 50°С в течение 10 минут. За единицу эндоглюканазной активности принимают такое количество фермента, которое в течение 1 минуты при температуре 50°С и рН 5,0 при гидролизе раствора КМЦ концентрацией 5 мг/мл приводит к образованию количества ВС, эквивалентного 1 микромолю глюкозы, определяемых методом Сомоджи-Нельсона (А.П.Синицын, А.В.Гусаков, И.М.Черноглазов. Биоконверсия лигноцеллюлозных материалов, Учебное пособие. - М.: Изд-воМГУ, 1995, с.144-156).
Ксиланазную активность измеряют, проводя гидролиз березового ксилана (5 мг/мл) при рН 5,0 (0,1 М ацетатный буфер) и 50°С в течение 10 минут. За единицу ксиланазной активности принимают такое количество фермента, которое в течение 1 минуты при температуре 50°С и рН 5,0 при гидролизе раствора ксилана концентрацией 5 мг/мл приводит к образованию количества ВС, эквивалентных 1 микромолю ксилозы, определяемых методом Сомоджи-Нельсона (А.П.Синицын, А.В.Гусаков, И.М.Черноглазов. Биоконверсия лигноцеллюлозных материалов, Учебное пособие. - М.: Изд-во МГУ, 1995, с.144-156).
Таким образом, предлагаемое изобретение позволяет осущеставить получение ферментных препаратов с высокой целлобиогидролазной, эндоглюканазной и ксиланазной активностями путем культивирования мультикопийных штаммов мицелиального гриба Penicillium verruculosum, трансформированных фрагментами ДНК с клонированными гомологичными и гетерологичными генами.
| название | год | авторы | номер документа |
|---|---|---|---|
| ГЕНЕТИЧЕСКАЯ ФЬЮЖН-КОНСТРУКЦИЯ ДЛЯ ОБЕСПЕЧЕНИЯ ЭКСПРЕССИИ МУЛЬТИФЕРМЕНТНОГО КОМПЛЕКСА КАРБОГИДРАЗ В КЛЕТКАХ МИЦЕЛИАЛЬНОГО ГРИБА Penicillium verruculosum, ИСПОЛЬЗУЕМОГО В КАЧЕСТВЕ ХОЗЯИНА, СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ШТАММА ГРИБА Penicillium verruculosum И СПОСОБ ПОЛУЧЕНИЯ ФЕРМЕНТНОГО ПРЕПАРАТА НА ЕГО ОСНОВЕ | 2013 |
|
RU2550044C2 |
| ШТАММ ГРИБА Penicillium verruculosum B10 EGII ПРОДУЦЕНТ ЭНДО-1.3/1.4-β-ГЛЮКАНАЗЫ, ЦЕЛЛЮЛАЗЫ, β-ГЛЮКОЗИДАЗЫ И КСИЛАНАЗЫ И СПОСОБ ПОЛУЧЕНИЯ КОРМОВОГО КОМПЛЕКСНОГО ФЕРМЕНТНОГО ПРЕПАРАТА | 2012 |
|
RU2532840C2 |
| НОВЫЙ РЕКОМБИНАНТНЫЙ ШТАММ (ВАРИАНТЫ) МИЦЕЛИАЛЬНОГО ГРИБА PENICILLIUM VERRUCULOSUM И ФЕРМЕНТНЫЙ ПРЕПАРАТ (ВАРИАНТЫ), ПРЕДНАЗНАЧЕННЫЙ ДЛЯ ГИДРОЛИЗА ПЛОДОВО-ЯГОДНОГО СЫРЬЯ, И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2014 |
|
RU2574206C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ МИЦЕЛИАЛЬНОГО ГРИБА PENICILLIUM VERRUCULOSUM ( ВАРИАНТЫ) И СПОСОБ ПОЛУЧЕНИЯ ФЕРМЕНТНОГО ПРЕПАРАТА С ЕГО ИСПОЛЬЗОВАНИЕМ (ВАРИАНТЫ) | 2015 |
|
RU2646136C2 |
| СПОСОБ ПОЛУЧЕНИЯ КОРМОВОГО КОМПЛЕКСНОГО ФЕРМЕНТНОГО ПРЕПАРАТА (ВАРИАНТЫ) И ШТАММ PENICILLIUM CANESCENS (ВАРИАНТЫ) | 2004 |
|
RU2288267C2 |
| ГЕНЕТИЧЕСКАЯ КОНСТРУКЦИЯ ДЛЯ ОБЕСПЕЧЕНИЯ ЭКСПРЕССИИ КОМПЛЕКСА ФЕРМЕНТОВ ЭНДОГЛЮКАНАЗ И КСИЛАНАЗ В КЛЕТКАХ ГРИБА PENICILLIUM VERRUCULOSUM И СПОСОБ ПОЛУЧЕНИЯ КОМПЛЕКСНЫХ ФЕРМЕНТНЫХ ПРЕПАРАТОВ НА ЕГО ОСНОВЕ, ПРЕДНАЗНАЧЕННЫХ ДЛЯ КОРМОПРОИЗВОДСТВА | 2017 |
|
RU2653429C1 |
| ШТАММ ГРИБА PENICILLIUM VERRUCULOSUM MX-73 ПРОДУЦЕНТ МОДИФИЦИРОВАННОЙ КСИЛАНАЗЫ Е С ПОВЫШЕННОЙ ТЕРМОСТАБИЛЬНОСТЬЮ, ФЕРМЕНТНЫЙ ПРЕПАРАТ НА ЕГО ОСНОВЕ ДЛЯ ИСПОЛЬЗОВАНИЯ В ПИЩЕВОЙ И КОРМОВОЙ ПРОМЫШЛЕННОСТИ И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2018 |
|
RU2711578C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ МИЦЕЛИАЛЬНОГО ГРИБА PENICILLIUM CANESCENS CL14, ПРОДУЦИРУЮЩИЙ КОМПОНЕНТ ЦЕЛЛЛЮЛОСОМЫ CLOSTRIDIUM THERMOCELLUM, И СПОСОБ ЕГО КУЛЬТИВИРОВАНИЯ | 2016 |
|
RU2646132C1 |
| ФРАГМЕНТ ДНК МИЦЕЛИАЛЬНОГО ГРИБА PENICILLIUM VERRUCULOSUM, КОДИРУЮЩИЙ СИНТЕЗ СЕКРЕТИРУЕМОЙ ЭНДОГЛЮКАНАЗЫ III И ГРУППА ШТАММОВ PENICILLIUM CANESCENS, СИНТЕЗИРУЮЩИХ ЭНДОГЛЮКАНАЗУ III PENICILLIUM VERRUCULOSUM, СКОНСТРУИРОВАННЫХ МЕТОДАМИ ТРАНСФОРМАЦИИ И ГЕНЕТИЧЕСКОЙ ИНЖЕНЕРИИ НА ОСНОВЕ ЭТОГО ФРАГМЕНТА ДНК | 2001 |
|
RU2238974C2 |
| СПОСОБ ПОЛУЧЕНИЯ ФЕРМЕНТНОГО ПРЕПАРАТА ДЛЯ РАСЩЕПЛЕНИЯ ГЕМИЦЕЛЛЮЛОЗНЫХ ГЕТЕРОПОЛИСАХАРИДОВ КЛЕТОЧНОЙ СТЕНКИ РАСТЕНИЙ И ФЕРМЕНТНЫЙ ПРЕПАРАТ (ВАРИАНТЫ) | 2007 |
|
RU2358756C1 |




Изобретение относится к биотехнологии и представляет собой способ получения фермента, предпочтительно целлюлазы или ксиланазы, расщепляющих целлюлозу или ксиланы соответственно, путем культивирования штамма гриба Penicillium verruculosum. При этом указанный штамм получают трансформацией клетки гриба Penicillium verruculosum генетической конструкцией, содержащей целевую кодирующую последовательность фермента, функционально связанную с регуляторными элементами гена целлобиогидролазы I Penicillium verruculosum, которые представляют собой промотор и терминатор гена целлобиогидролазы I, a сигнальный пептид при этом зависит от того, каков целевой фермент. Изобретение позволяет получать ферменты с высокой степенью эффективности. 3 н. и 3 з.п. ф-лы, 1 табл., 6 ил.
1. Генетическая конструкция для обеспечения экспрессии целевых гомологичных и гетерологичных генов ферментов в клетках мицелиального гриба Penicillium verruculosum, используемого в качестве хозяина, содержащая целевую кодирующую последовательность фермента, функционально связанную с регуляторными элементами гена целлобиогидролазы I Penicillium verruculosum, которые представляют собой промотор и терминатор гена целлобиогидролазы I, а сигнальный пептид при этом зависит от того, каков целевой фермент.
2. Способ получения штамма гриба Penicillium verruculosum - продуцента фермента, кодируемого нуклеотидной последовательностью, предусматривающий трансформацию клетки гриба Penicillium verruculosum генетической конструкцией, содержащей целевую кодирующую последовательность фермента, функционально связанную с регуляторными элементами гена целлобиогидролазы I Penicillium verruculosum, которые представляют собой промотор и терминатор гена целлобиогидролазы I, а сигнальный пептид при этом зависит от того, каков целевой фермент.
3. Способ получения фермента, предпочтительно целлюлазы или ксиланазы, расщепляющих целлюлозу или ксиланы соответственно, путем культивирования микроорганизмов, отличающийся тем, что способом по п.2 получают штамм гриба Penicillium verruculosum, осуществляют культивирование полученного штамма и выделяют целевой продукт из культуральной жидкости гриба.
4. Способ получения фермента по п.3, отличающийся тем, что получают штамм Penicillium verruculosum PV-CbhIP (BKM F-3973D), содержащий множество копий гена cbhI - высокоактивной целлобиогидролазы I Penicillium verruculosum.
5. Способ получения фермента по п.3, отличающийся тем, что получают штамм Penicillium verruculosum PV-Egl4T (BKM F-3976D), содержащий множество копий гена eglIV - эндоглюканазы IV Trichoderma longibrachiatum.
6. Способ получения фермента по п.3, отличающийся тем, что получают штамм Penicillium verruculosum PV-Xyl3T (BKM F-3979D), содержащий множество копий гена xylIII - ксиланазы III Trichoderma longibrachiatum.
| СПОСОБ ПОЛУЧЕНИЯ КОРМОВОГО КОМПЛЕКСНОГО ФЕРМЕНТНОГО ПРЕПАРАТА (ВАРИАНТЫ) И ШТАММ PENICILLIUM CANESCENS (ВАРИАНТЫ) | 2004 |
|
RU2288267C2 |
| ФРАГМЕНТ ДНК PCG 2,6, КОДИРУЮЩИЙ СИНТЕЗ СЕКРЕТИРУЕМОЙ БЕТА-ГАЛАКТОЗИДАЗЫ PENICILLIUM CANESCENS, И ШТАММ ГРИБА PENICILLIUM CANESCENS - ПРОДУЦЕНТ СЕКРЕТИРУЕМОЙ БЕТА-ГАЛАКТОЗИДАЗЫ | 1997 |
|
RU2126049C1 |
| US 2007148730 A1, 28.06.2007 | |||
| KOCH et | |||
| al | |||
| Cloning, sequencing, and heterologous expression of a cellulase- encoding cDNA (cbhl) from Penicilliam janthinellum, Gene, 1993 Feb 14; 124(1):57-65 | |||
| HOU Y et | |||
| al | |||
| Cloning, sequencing and expression analysis of the first cellulase gene encoding | |||

