ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
В настоящей заявке испрашивается приоритет предварительной заявки на патент США №60/849927, поданной 6 октября 2006 года, содержание которой включено в настоящее описание в качестве ссылки.
ПРАВА ПРАВИТЕЛЬСТВА
Это изобретение было сделано при поддержке правительства Соединенных Штатов, гранты № DAMD17-02-0137 и DAMD17-02-0562 от Министерства Обороны и гранты № W81XWH-05-1-0091 и W81XWH-05-1-0318, контролируемые конгрессом программы медицинских исследований (Congressionally Directed Medical Research Program). Правительство имеет определенные права на данное изобретение.
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к композициям дезоксирибонуклеиновых кислот (ДНК), кодирующим пригодные молекулы, эффективные для вызова иммунного ответа против опухолеассоциированных макрофагов. Более конкретно, настоящее изобретение относится к композициям ДНК, кодирующим, по меньшей мере, один эпитоп эндопептидазы, такой как легумаин (legumain), который экспрессируется в опухолеассоциированной клетке, такой как опухолеассоциированный макрофаг. Настоящее изобретение также относится к способам использования композиции ДНК для ингибирования опухолевого роста и опухолевых метастазов.
УРОВЕНЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
Опухолеассоциированные макрофаги (TAM) связаны с развитием опухоли и метастазированием. Новая противоопухолевая стратегия состоит в иммунизации против молекул, сверхэкспрессируемых ТАМ, и, посредством этого, в изменении окружающей микросреды опухоли, которая привлекает эти макрофаги и опосредует их функцию (см. Oosterling et al. 2005 J. Pathol. 207:147-155; Emens et al. 2005 Endocr. Relat Cancer 12:1-17). ТАМ состоят предпочтительно из популяции поляризованных макрофагов М2 (CD206+, F4/80+) с низкой цитотоксичностью по отношению к опухолевым клеткам из-за ограниченного продуцирования ими оксида азота и провоспалительных цитокинов (см. Mills et al. 2000 J. Immunol. 164:6166-6173).
ТАМ также обладают низкой антиген-презентирующей способностью и эффективно подавляют активацию Т-клеток. Фактически, эти макрофаги фенотипа М2 действительно способствуют пролиферации опухолевых клеток и метастазированию, секретируя широкий спектр факторов роста и про-ангиогенных факторов, а также металлопротеиназ, с вовлечением в сигнальную циркуляцию, что регулирует функцию фибробластов опухолевой стромы (Mantovani et al. 2004, Novartis. Found. Symp. 256:137-145).
В настоящее время анти-ТАМ эффекты, вызываемые низкомолекулярными ингибиторами, по сообщениям, способствуют подавлению опухоли (см. Lewis et al. 2005 Am. J. Pathol. 167:627-635; и Mantovani et al. 2004, Eur. J. Cancer 40:1660-1667). Например, «yondelis», антинеопластический агент, оказывает избирательный цитотоксический эффект на ТАМ, тем самым значительно снижая продуцирование ими IL6 и CCL2, что способствует угнетению роста опухолей человека, связанных с воспалением (см. Allavena et al. 2005, Cancer Res. 65:2964-2971). Другой подобный пример иллюстрируется бифосфонатным соединением, золедроновой кислотой, которая подавляет секрецию MMP9 ТАМ, тем самым ингибируя опухолевую металлопротеиназную активность и уменьшая связывание VEGF c его тирозинкиназными рецепторами на пролиферирующих эндотелиальных клетках (см. Giraudo et al. 2004, J. Clin. Invest. 114:623-633).
Как было показано в различных экспериментальных моделях, хемокин CCL5 очень важен в пополнении ТАМ. Антагонист этого хемокина уменьшал опухолевый инфильтрат и замедлял рост опухоли (см. Robinson et al. 2003, Cancer Res. 63:8360-8365). Следовательно, хотя терапевтическое нацеливание на ТАМ все еще довольно новый подход, начальные клинические результаты обнадеживают, так как они позволяют предположить, что нацеливание на ТАМ может дополнить более традиционные противораковые терапевтические схемы.
Легумаин - это абсолютно новое эволюционное ответвление семейства С13 цистеиновых протеаз (см. Ishii 1994, Methods Enzymol. 244:604-615),и он хорошо сохранился у растений и у млекопитающих, включая человека. Впервые легумаин был выявлен в растениях как фермент процессинга запасных белков во время прорастания семян и впоследствии обнаружен у паразитов, а затем у млекопитающих. Легумаин - это кислотоустойчивая цистеиновая эндопиптедаза с необычайно узкой специфичностью, обязательно требующая наличия аспарагина в сайте P1 субстратной последовательности (см. Chen et al. 1997, J. Biol. Chem. 272:8090-8098).
В настоящей заявке выбор легумаина в качестве мишени для противоопухолевой терапии основан на том факте, что ген, кодирующий эту эндопептидазу, как выяснилось, высоко активен во многих мышиных и человеческих опухолевых тканях, но отсутствует или представлен на очень низких уровнях во всех нормальных тканях, из которых эти опухоли развились. Важно, что сверхэкспрессия легумаина возникает в таких стрессовых условиях, как опухолевая гипоксия, которая ведет к усиленному росту опухоли, ангиогенезу и метастазированию.
Легумаин представляет собой особенно предпочтительную целевую эндопептидазу для композиций и способов по настоящему изобретению благодаря наблюдению, что легумаин интенсивно сверхэкспрессируется ТАМ в опухолевых тканях молочных желез у мышей, что подтверждается анализом экспрессии гена и иммуногистохимическими исследованиями. ТАМ имеют чрезвычайно высокую экспрессию легумаина в строме опухоли. В противоположность, классические макрофаги фенотипа М1, которые выполняют важные функции иммунного надзора и презентирования антигена, не экспрессируют легумаин. Следовательно, нацеливание на ТАМ, которые сверхэкспрессируют легумаин, не мешает биологическим функциям макрофагов М1, включая цитотоксичность и презентирование антигена. Настоящее изобретение представляет композиции ДНК для индуцирования иммунного ответа против ТАМ, сверхэкспрессирующих легумаин или другие эндопептидазы, также сверхэкспрессируемые в ТАМ, и которые пригодны для лечения опухолей и опухолевых метастазов.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Композиция ДНК по настоящему изобретению содержит конструкцию ДНК, которая кодирует полипептид, включающий, по меньшей мере, иммуногенный фрагмент цистеиновой эндопептидазы, сверхэкспрессируемой в опухолеассоциированных клетках (например, опухолеассоциированных макрофагах). Конструкция ДНК включает структурные элементы, которые способствуют экспрессии полипептида в иммунных клетках субъекта, которому ввели конструкцию ДНК. Предпочтительно, эндопептидаза включает легумаин или, по меньшей мере, один его иммуногенный фрагмент (например, эпитоп). Конструкция ДНК заключена в фармацевтически приемлемый носитель, чтобы она могла быть введена пациенту. Композиция может кодировать единственный иммуногенный фрагмент эндопептидазы (например, эпитоп легумаина), полипептид, включающий два или более иммуногенных фрагментов эндопептидазы (т.е. иммуногенный полипептид), целый белок эндопептидазы или любую его часть, которая будет вызывать иммунный ответ у субъекта. Предпочтительно, чтобы конструкция ДНК кодировала человеческий легумаин, имеющий последовательность аминокислотных остатков, состоящих из SEQ ID NO: 2, белок, имеющий по меньшей мере 80% идентичность последовательности с SEQ ID NO: 2 (например, человеческий легумаин, свиной легумаин или мышиный легумаин), или его иммуногенный фрагмент, экспрессируемый в иммуногенных клетках субъекта, которому введена конструкция ДНК. Конструкции ДНК по настоящему изобретению пригодны для ингибирования роста опухоли и опухолевых метастазов.
Конструкция ДНК может быть «оголенной», предпочтительно в форме плазмиды. Такие «оголенные» конструкции ДНК могут быть заключены в липосомный носитель, полимерный носитель или вводиться посредством электропорации, генной пушки и тому подобных, при желании. В некоторых предпочтительных вариантах конструкция ДНК встроена в аттенуированный вирусный вектор или аттенуированный бактериальный вектор.
В предпочтительном варианте осуществления конструкция ДНК встроена в аттенуированный бактериальный вектор, такой как аттенуированная Salmonella typhimurium, например дважды аттенуированный (AroA-, dam-) штамм Salmonella typhimurium.
Необязательно, композиция ДНК может также включать конструкцию ДНК, кодирующую иммунный эффекторный белок, такой как цитокин. Предпочтительные цитокины включают CCL21, IL-2 и CD40LT.
В особенно предпочтительном варианте осуществления композиция ДНК по изобретению включает минигенную конструкцию ДНК, которая кодирует иммуногенный полипептид, включающий множество иммуногенных фрагментов (например, эпитопов) цистеиновой эндопептидазы (например, легумаина), который активно экспрессируется в опухолеассоциированных клетках. Иммуногенный полипептид, способный вызывать иммунный ответ против опухолеассоциированных клеток, экспрессируется в иммунных клетках и заключен в фармацевтически приемлемый носитель. Иммуногенные фрагменты соединены вместе последовательно пептидом-линкером с каждым последующим фрагментом в полипептиде. Пептиды-линкеры, в основном, длиной по меньшей мере три аминокислотных остатка и, предпочтительно, включают аминокислотную последовательность ААА или AAY. Использующийся в настоящем описании термин «пептид-линкер» относится к последовательности по меньшей мере из двух аминокислотных остатков, предпочтительно, по меньшей мере трех аминокислотных остатков, которые образуют последовательность аминокислотных остатков, связанную с иммуногенными фрагментами эндопептидазы и отличающуюся от природной эндопептидазы. Типично, комбинация пептидов-линкеров и иммуногенных фрагментов эндопептидазы будет включать полипептид длиной меньше чем приблизительно 100 аминокислотных остатков, более предпочтительно, длиной, приблизительно, от 19 до 62 аминокислот (например, от двух до, приблизительно, пяти иммуногенных фрагментов по 8-10 аминокислот каждый, объединенных вместе одним-четырьмя пептидами-линкерами по три аминокислоты каждый). Предпочтительно, минигенная конструкция ДНК кодирует иммуногенные фрагменты легумаина человека (SEQ ID NO: 2).
Композиции ДНК по настоящему изобретению могут действовать как вакцины, нацеленные на опухолеассоциированные макрофаги, которые экспрессируют цистеиновую эндопептидазу, такую как легумаин, обеспечивая высоко селективную цель для Т-клеточно-опосредованной противораковой иммунотерапии. Подход нацеливания на эндопептидазу, такую как легумаин, экспрессируемую опухолеассоциированными макрофагами, имеет несколько преимуществ над способами лечения, направленными против антигенов, которые экспрессируются исключительно самими опухолевыми клетками. Так, например, легумаин сверхэкспрессируется в ТАМ и, следовательно, не повреждается подавляемой экспрессией МНС-антигена, как зачастую случается в опухолевых клетках. Кроме того, опухолевые клетки часто становятся повышенно устойчивыми к опосредованному Т-клетками уничтожению из-за дефектов в апоптозных сигнальных путях, повышенной регуляции антиапоптозных белков или иммуносуппрессивных воздействий на цитотоксические Т-лимфоциты (CTL). Нацеливание на ТАМ, экспрессирующие легумаин, позволяет терапевтической композиции лечить ряд различных злокачественных образований, в противоположность способам лечения, вовлекающим антигены, экспрессирующиеся исключительно специфичными типами опухолей.
В одном предпочтительном варианте осуществления композиции ДНК по настоящему изобретению нарушают периферическую Т-клеточную устойчивость к аутоантигену легумаина, передавая свои кДНК ДНК, кодирующей один или более их иммуногенных фрагментов, как пероральная композиция ДНК с атеннуированным бактериальным вектором доставки (например, аттенуированный штамм Salmonella typhimurium). В таких вариантах осуществления композиция ДНК контактировала с антигенпрезентирующими клетками (APC) во вторичном лимфоидном органе, т.е. Пейеровых бляшках тонкой кишки. С профилактической целью, Т-клеточно-опосредованный противоопухолевый иммунный ответ, индуцированный вакцинацией композицией ДНК по изобретению, ингибировал рост опухоли в многочисленных мышиных опухолевых моделях. Настоящие композиции ДНК также значительно подавляют диссеминацию установленных легочных метастазов в терапевтической модели карциномы толстой кишки CT26.
Предпочтительная композиция ДНК включает аттенуированный носитель Salmonella, такой как дважды аттенуированный штамм S. typhimurium, например штамм, называемый RE 88, который включает dam- и AroA- мутации и поставляется Remedyne Corporation (Goleta, CA). В настоящем изобретении аттенуированный носитель Salmonella трансфицирован так, что он включает конструкцию ДНК, кодирующую эндопептидазу (например, легумаин) или полипептид, содержащий ее иммуногенный фрагмент. Эндопептидаза или полипептид экспрессируются в иммунных клетках млекопитающих, которым они вводятся. Сама бактерия не экспрессирует легумаин или полипептид, а только доставляет ДНК в такие иммунные клетки, как макрофаги или дендритные клетки (DC), которые в свою очередь экспрессируют эндопептидазу или полипептид, включающий ее иммуногенный фрагмент. Такие композиции могут обеспечить пролонгированные противоопухолевые эффекты в мышиных моделях. К тому же, эксперименты in vivo по истощению Т-клеток показали вовлечение CD8+, а не CD4+ Т-клеток в иммунный ответ, ассоциированный с композициями, кодирующими легумаин и полипептиды, включающие иммуногенные фрагменты легумаина. Наблюдавшийся цитотоксический эффект, опосредованный CD8+ Т-клетками in vitro, был специфически направлен против ТАМ-мишеней, которые сверхэкспрессируют антиген легумаина.
Композиции ДНК по настоящему изобретению также могут включать конструкции ДНК, которые кодируют иммунные эффекторные молекулы как адъюванты для композиции. Такие иммунные эффекторные молекулы включают, например, IL-2, индуктор пролиферации Т-клеток, CCL21, хемокин, который химически притягивают зрелые дендритные клетки, и наивные Т-клетки, а также CD40LT, известный индуктор созревания дендритных клеток. Нуклеиновые кислоты, кодирующие иммунные эффекторные белки, предпочтительно встроены в плазмиду. Конструкции ДНК легумаина и иммунного эффекторного белка могут быть встроены в одну и ту же плазмиду или в две отдельные плазмиды. CTL-ответ, индуцируемый против ТАМ, может замедлять рост ряда опухолей и не специфичен по отношению к конкретному типу опухолей.
Настоящее изобретение также предоставляет способ ингибирования опухолевого роста и опухолевых метастазов у млекопитающих, включая введение млекопитающему композиции ДНК по изобретению в количестве, достаточном для вызова иммунного ответа против ТАМ, экспрессирующих легумаин.
Еще один аспект настоящего изобретения - это эффективная комбинация схемы лечения, которая объединяет химиотерапию и терапию композицией ДНК по изобретению. В этом способе осуществления настоящего изобретения различные химиотерапевтические агенты, такие как доксорубицин, паклитаксел и/или циклофосфамид, которые не вызывают угнетения костного мозга при введении в максимальной допустимой дозе (МДД/MTD), вводились пациенту в сочетании с композицией ДНК по изобретению, содержащей конструкцию ДНК, кодирующую ТАМ-экспрессируемую эндопептидазу, такую как легумаин или полипептид, кодирующий ее иммуногенную часть, предпочтительно содержащую минигенную конструкцию, которая включает, по меньшей мере, два иммуногенных фрагмента эндопептидазы, последовательно соединенных вместе пептидами-линкерами между каждыми из последовательных иммуногенных фрагментов в полипептиде.
Другим предпочтительным способом осуществления является способ ингибирования роста опухоли или опухолевых метастазов у млекопитающих (например, человека), включающий стадии введения млекопитающим композиции ДНК по изобретению в количестве, достаточном, чтобы вызвать у них иммунный ответ против ТАМ, сверхэкспрессирущих эндопептидазу, такую как легумаин, с последующим введением млекопитающим эффективного противоопухолевого количества противоопухолевого химиотерапевтического средства.
Предпочтительно, чтобы млекопитающим, подвергшимся лечению способами по настоящему изобретению, являлся человек.
В способе осуществления настоящего изобретения, композиции ДНК могут вводиться энтерально посредством перорального введения или парентерально посредством инъекции или внутривенной инфузии. Предпочтительным является пероральное введение композиций. Композиции могут быть упакованы в герметичные контейнеры и снабжены информацией для клиницистов по эффективному введению композиции.
Композиции ДНК по настоящему изобретению являются пригодными для лечения и профилактики ряда болезненных состояний. Например, больной, страдающий колоректальным раком, раком молочной железы, раком легких и тому подобное, может получить пользу от иммунизации композициями по настоящему изобретению. Композиции по настоящему изобретению также полезны для исследования роли легумаина в различных формах рака.
Краткое описание чертежей
ФИГ.1. Легумаин экспрессируется в высокой степени на опухолеассоциированных макрофагах в строме опухоли. (А) Экспрессия легумаина в ТАМ была явно выражена, как показано на панели А. Макрофаги, инфильтрирующие опухоль, были визуализированы с помощью Г/Э окрашивания, как показано стрелками. Экспрессия легумаина выявлена двойным окрашиванием антилегумаиновых антител в сочетании с анти-CD68 антителами. (Увеличение ×35) (B) Повышенная экспрессия легумаина на ТАМ была подтверждена проточными цитометрическими анализами дважды положительных популяций CD206+/F4/80+ макрофагов M2, которые были выделены из свежей опухолевой ткани. (С) Многоцветовая проточная цитометрия продемонстрировала повышающую регуляцию маркера CD206 макрофага M2 на RAW-клетках после их культивирования с IL-4, IL-10 и IL-13 (10 нг/мл). Было показано, что легумаин активно экспрессируется на F4/80+/CD206+ положительных RAW клетках, культивировавшихся с IL-4, IL-10 и IL-13, как показано на верхнем фото, тогда как сравниваемые с диким типом RAW клетки изображены на нижнем фото. (D) Подтверждение экспрессии легумаина на RAW клетках с помощью вестерн-блоттинга, следующей за стимуляцией с IL-4, IL-13 и IL-10, отдельно или комбинированно.
ФИГ.2. Выделение легумаинэкспрессирующих клеток ведет к подавлению роста опухоли. (А) Схема композиции ДНК по изобретению, построенная с pCMV/myc/cyto векторным остовом, где ген легумаина был присоединен к C-концу мутантного полиубиквитина. Был включен целый фрагмент, и экспрессия белка была продемонстрирована вестерн-блоттингом. (В) Профилактическая модель: график вакцинации был рассчитан на три иммунизации с однонедельными интервалами, следуемыми за внутривенными введениями приблизительно 2×105 клеток немелкоклеточного рака легкого D121, приблизительно 50×104 клеток рака толстой кишки CT26, и инъекцией приблизительно 7×103 клеток рака молочной железы 4Т1 в жировую ткань молочной железы. Массы легких были измерены через 24 дня (D121 или CT-26) или через 30 дней (4Т1) после введения опухолевых клеток и проанализированы в каждой группе. Разница между двумя контрольными группами (PBS и/или пустой вектор) и лечебной группой была статистически значимой **P<0,005. Нормальный вес легкого = 0,2 г. (С) Терапевтическая модель: группам мышей BALB/c (n=8) в жировую ткань молочной железы изначально ввели приблизительно 7×103 клеток рака молочной железы 4T1 и в дальнейшем вакцинировали три раза на 3, 7 и 11 дни PBS, пустым вектором или pLegumain композицией ДНК, соответственно, и первичную опухоль удаляли на 12 день. Графики выживания отображают результаты для лечебной и контрольных групп по 8 мышей в каждой. Разница между контрольной группой пустого вектора и лечебной группой была статистически значима **P<0,005.
ФИГ.3. Популяция ТАМ в опухолевой строме сократилась благодаря специфическими CD8+ CTL, индуцированными композицией ДНК, основанной на легумаине. (А) RAW макрофаговые клетки активно экспрессируют легумаин после культивирования с IL-4, IL-10, IL-13 10 нг/мл и служат клетками-мишенями в 4-часовом анализе высвобождения 51Cr. Спленоциты, выделенные у мышей, иммунизированных pLegumain вакциной, эффективно уничтожали RAW клетки, обработанные in vitro этими цитокинами в различных соотношениях клеток эффектор-мишень, но не были способны вызвать цитотоксическое уничтожение нестимулированных RAW клеток, лишенных экспрессии легумаина. **P<0,005 по сравнению с контрольными группами. (В) Проточная цитометрия детектирует процент популяций ТАМ со специфическими макрофаговыми маркерами (CD206 и F4/80) в опухолевой ткани после вакцинации. Было показано снижение процентного соотношения популяций ТАМ среди клеток опухолевой ткани, выделенной у мышей, пролеченных композицией ДНК по изобретению. Не было уменьшения популяций ТАМ, выделенных у мышей, пролеченных либо пустым вектором, либо pLegumain вслед за истощением CD8+ Т-клеток (**P<0,005). (С) Результаты проточной цитометрии были подтверждены иммуногистохимическим окрашиванием, оцененным конфокальной микроскопией. Популяция ТАМ в опухолевой строме значительно сократилась после вакцинации. Увеличение ×5 (Г/Э), ×35 (Контроль, Пустой Вектор и pLegumain).
ФИГ.4. Ответ специфичных CD8+ Т-клеток, рестриктированных по МНС-антигенам первого класса против клеток, экспрессирующих легумаин. (А) Графики FACS показывают, что ДНК терапия увеличивает экспрессию ко-стимулирующих молекул благодаря DC. Лимфоциты из Пейеровых бляшек, полученные спустя 3 дня после вакцинации, были окрашены меченым FITC антителом к CD11c в комбинации с PE конъюгированным антителом к CD80, анти-MHC первого класса или анти-CD40 антителами. (*P<0,05, в сравнении с контрольными группами). (В) Внутриплазматическое высвобождение INF-гамма CD8+ Т-клетками было оценено FACS анализом. **P<0,005 по сравнению с контрольными группами. (С) Продуцирование специфического INF-гамма контролировалось на уровне отдельных клеток с помощью ELISPOT. Отображено количество иммунных пятен на лунку для лимфоцитов из иммунизированных мышей, рестимулированных либо legumain+ клетками опухолевой ткани 4T1, либо legumain- клетками 4T1. **P<0,005 по сравнению с лечебной группой без стимуляции. ##P<0,005 по сравнению с контрольными группами. (D) Спленоциты, выделенные из пролеченных мышей, эффективно уничтожали ТАМ, как показало применение теста с высвобождением 51Cr (*P<0,01 по сравнению с контрольными группами). Ингибирующие эксперименты с антителами против MHC-антигенов первого класса H2Kd/H-2Dd показали, что T-клетка, опосредовавшая лизис опухолевой клетки, была рестриктирована по MHC антигену первого класса. Истощение in vivo Т-клеток CD4+ или CD8+ показало, что лимфоциты, выделенные у вакцинированных мышей, у которых были истощены Т-клетки CD8+, не смогли вызвать цитотоксический лизис клеток-мишеней, истощение Т-клеток CD4+ не подавляло цитотоксический лизис этих клеток-мишеней. *P<0,01 по сравнению с группой PBS или пустого вектора.
ФИГ.5. Нейтрализация ТАМ ведет к уменьшению высвобождения факторов роста, миграции опухолевых клеток и метастазов. (А) Композиция ДНК по изобретению уменьшает высвобождение факторов роста ТАМ. Ткань опухоли молочной железы 4Т1 и мышиная сыворотка были собраны через 12 дней после вакцинаций и введения опухолевых клеток. После 24 часов или 48 часов культивирования супернатанты клеток опухолевых тканей были собраны, и концентрации TGF-бета, TNF-альфа и VEGF сыворотки или супернатантов были измерены с помощью ELISA. Разницы между лечебной и контрольной группами были значительными. *Р<0,01, **Р<0,005. (В) Было проведено иммуногистохимическое окрашивание, с целью обнаружить экспрессию этих факторов роста в микросреде опухоли. Вакцинированные лечебные группы показали, что после сокращения ТАМ уменьшилось высвобождение VEGF, TGF-бета и ММР-9, по сравнению с группами пустого вектора. (С) Был проведен трансвелл-анализ миграции (transwell migration assay), с целью определить миграцию опухолевых клеток после вакцинации. Число мигрирующих клеток явно уменьшилось после вакцинации. ***Р<0,001 по сравнению с группой пустого вектора. (D) Были проведены эксперименты in vivo, чтобы определить возможность формирования опухолевых метастазов у мышей. Мыши были пролечены вакциной по терапевтической схеме, как описано выше. Показатели опухолевого метастазирования и массы легких измерялись через 25 дней после эксцизии первичной опухоли. Показатель метастазирования выражался как % легочной поверхности, покрытой слившимися метастатическими очагами: 0 = отсутствовали; 1 = <5%, 2 = от 5% до 50% и 3 = >50%. Разницы в массе легких между группой мышей, пролеченной вакциной, и всеми контрольными группами были статистически значимыми (**Р<0,005).
ФИГ.6. Элиминация ТАМ приводит к снижению опухолевого ангиогенеза. Суппрессия VEGF-индуцированного ангиогенеза: мыши BALB/с были вакцинированы S.typhimurium, трансфицированной либо пустым вектором, либо pLegumain, либо pLegumain после элиминации либо CD8+, либо CD4+ Т-клеток in vivo, соответственно. Спустя неделю после последней вакцинации подкожно в среднюю линию живота мышам был введен Матригель (Matrigel). Васкуляризация была индуцирована VEGF или bFGF. (А) Изображения были сделаны с помощью цифровой камеры спустя 6 дней после имплантации пробки Матригеля. Кроме того, срез пробок Матригеля, окрашенных трихромом по Массону, показывает увеличение кровеносных сосудов в пробках Матригеля, как выделено стрелками (увеличение ×5). (В) Количественный анализ сосудистого роста был выполнен после окрашивания эндотелия in vivo FITC-меченным изолектином В4 и оценки с помощью флуорометрии. Уменьшение VEGF-индуцированной неоваскуляризации происходило только после вакцинации вектором, кодирующим легумаин, но не после вакцинации пустым вектором или pLegumain после истощения CD8+ Т-клеток. **Р<0,005, *P<0,01 по сравнению с легумаиновой лечебной группой. (С) Иммуногистохимическое окрашивание было выполнено и исследовано с помощью конфокальной микроскопии. Поперечные срезы пробок Матригеля были окрашены для определения типа клеток, проросших или мигрировавших в эти пробки. Изображения показывают, что эндотелиальные клетки с маркером CD31 или макрофаги с маркером CD68 проросли или мигрировали в пробки Матригеля, как показано стрелками (увеличение ×35). Контролем служило Г/Э окрашивание (увеличение ×5).
ФИГ.7. Клеточная линия 4Т1 была устойчиво трансфицирована ретровирусом, содержащим легумаиновую плазмиду, а затем использована в качестве клеток-мишеней для спленоцитов из иммунизированных мышей (Панель А, левое фото имеет 5× увеличение; правое фото имеет 35× увеличение), изображения были сделаны спустя 2 дня после трансфекции, и положительные клетки помечены стрелками. Данные анализа высвобождения 51Cr показаны на Части В. Спленоциты, выделенные из мышей, иммунизированных pLegumain композицией ДНК, эффективно уничтожали 4Т1 клетки, трансфицированные легумаином (*P<0,01 по сравнению с группами пустого вектора). Опухолеассоциированное опосредованное Т-клетками уничтожение было специфично для легумаина, поскольку нормальные 4Т1 клетки без легумаиновой экспрессии не были лизированы.
ФИГ.8 показывает нуклеотидную последовательность нуклеиновой кислоты, кодирующей легумаин человека (SEQ ID NO: 1).
ФИГ.9 показывает последовательность аминокислотных остатков (SEQ ID NO: 2) легумаина человека.
ФИГ.10 показывает нуклеотидную последовательность (SEQ ID NO: 3) нуклеиновой кислоты, кодирующей легумаин мыши.
ФИГ.11 показывает последовательность аминокислотных остатков (SEQ ID NO: 4) легумаина мыши.
ФИГ.12 показывает нуклеотидную последовательность (SEQ ID NO: 3) нуклеиновой кислоты, кодирующей IL-2 человека.
ФИГ.13 показывает последовательность аминокислотных остатков IL-2 человека (SEQ ID NO: 6).
ФИГ.14 показывает нуклеотидную последовательность (SEQ ID NO: 7) нуклеиновой кислоты, кодирующей CCL21 человека.
ФИГ.15 показывает последовательность аминокислотных остатков CCL21 человека (SEQ ID NO: 8).
ФИГ.16 показывает нуклеотидную последовательность (SEQ ID NO: 9) нуклеиновой кислоты, кодирующей CD40L человека.
ФИГ.17 показывает последовательность аминокислотных остатков CD40L человека (SEQ ID NO: 10).
ФИГ.18 показывает нуклеотидную последовательность убиквитинилированного легумаина мыши (SEQ ID NO: 11).
ФИГ.19 показывает последовательность аминокислотных остатков убиквитинилированного легумаина мыши (SEQ ID NO: 12).
ФИГ.20 показывает последовательность аминокислотных остатков эпитопных последовательностей мышиного легумаина.
ФИГ.21 показывает схематичное изображение плазмидных кодирующих легумаин минигенов, содержащих иммуногенные фрагменты легумаина.
ФИГ.22 показывает, что pCMV-Kb/Kd минигенная композиция защищает мышей от заражения клетками D2F2 карциномы молочной железы. Группы мышей BALB/c (n=8) были иммунизированы три раза с недельным интервалом дважды аттенуированной Salmonella typhimurium RE-88, несущей указанные векторы. Спустя одну неделю после последней иммунизации мышей заразили с помощью в/в инъекции клетками карциномы молочной железы D2F2 в количестве 2×105. А. Схема протокола эксперимента. В. Значение объема опухоли мышей спустя 5-25 дней после заражения опухолевыми клетками. С. Вес опухоли мышей спустя 25 дней после заражения, *P<0,05 по сравнению с контрольной группой пустого вектора.
ФИГ.23 показывает, что pKd минигенная вакцина предотвращает метастазирование карциномы молочной железы D2F2 у изогенных мышей BALB/c. Группы мышей (n=8) были иммунизированы 3 раза с 1-недельными интервалами с помощью принудительного кормления аттенуированной Salmonella typhimurium RE-88, несущей указанные векторы. Спустя 2 недели после последней иммунизации мышей заразили с помощью в/в инъекции клетками карциномы молочной железы D2F2 в количестве 1×105. А. Схема протокола эксперимента. В. Средний показатель мышиного легочного метастазирования каждой экспериментальной группы спустя 25 дней после заражения опухолевыми клетками. Показатели легочного опухолевого метастазирования устанавливали, определяя % покрытой метастазами площади поверхности: 0, метастазы отсутствовали, 1, <20%, 2, от 20 до 50% и 3, >50% представляли индивидуальными символами для каждой лечебной группы. *P<0,05 по сравнению с контрольной группой пустого вектора.
ФИГ.24 иллюстрирует высвобождение IFN-гамма в легумаин-специфических Т-клетках, вызванное pCMV-Kb/Kd минигенной композицией. А. Экспрессия легумаина свежеполученными из опухолевых тканей 4Т1 клетками, использованными в качестве клеток-стимуляторов. С целью показать степень экспрессии легумаина на этих клетках была проведена проточная цитометрия. В. Продуцирование IFN-гамма контролировалось с помощью ELISPOT на уровне отдельных клеток, таких как лимфоциты из иммунизированных мышей, рестимулированных или legumain+ 4Т1 опухолевыми клетками, свежеполученными из опухолевой ткани, или legumain- 4Т1 культуральными клетками. Высвобождение INF-гамма показано количеством иммунных пятен на лунку. *P<0,05 по сравнению с группами мышей, чьи лимфоциты не были стимулированы legumain+ опухолевых клеток. #P<0,05 по сравнению с контрольными группами.
ФИГ.25 иллюстрирует специфическое CTL уничтожение легумаин положительных макрофаговых клеток, вызванное применением pCMV-Kb/Kd минигенной композиции. А. Показана экспрессия легумаина макрофаговыми клетками RAW линии после культивирования с IL-4, IL-10 и IL-13. С целью показать экспрессию легумаина на этих клетках был проведен Вестерн-блотт анализ. В. Спустя 2 недели после последней иммунизации были умерщвлены группы мышей BALB/c (n=4), а извлеченные у них спленоциты были стимулированы облучаемыми в течении 5 дней 4Т1 legumain+ клетками. После этого был проведен цитотоксический анализ либо с диким типом RAW legumain- клеток (нижняя панель), либо с RAW legumain+ клетками в качестве клеток-мишеней (нижняя панель). *<0,05 по сравнению с группой пустого вектора, где в качестве мишени использовались RAW legumain+ клетки.
ФИГ.26 иллюстрирует подавление ангиогенеза у изогенных мышей BALB/c, вызванное pCMV-Kb/Kd минигенной композицией. А. Результат матригельного анализа. Матригель был имплантирован мышам, вакцинированным либо пустым вектором, либо pCMV-Db/Dd, либо pCMV-Kb/Kd вакцинами. Измерение концентрации гемоглобина (Hb) в матригелевых пробках производилось для измерения роста кровеносных сосудов. Средняя концентрация Hb матригелевых пробок в каждой группе мышей отображена на гистограмме (n=4; среднее + стандартное отклонение). *P<0,05 по сравнению с контрольной группой пустого вектора. В. Окрашивание трихромом по Масону участка Матригеля проводили спустя 7 дней после имплантации матригелевой пробки. Стрелки показывают кровеносные сосуды в матригелевой пробке.
ПОДРОБНОЕ ОПИСАНИЕ ПРЕДПОЧТИТЕЛЬНЫХ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ
Настоящее изобретение предоставляет композицию ДНК, которая нацелена на опухолеассоциированные клетки, такие как опухолеассоциированные макрофаги (ТАМ). Композиция ДНК включает заключенную в фармацевтически приемлемый носитель конструкцию ДНК, кодирующую цистеиновую эндопептидазу, которая сверхэкспрессируется в опухолеассоциированных клетках, или по меньшей мере, один ее иммуногенный фрагмент, способный вызвать иммунный ответ против опухолеассоциированных клеток. Предпочтительно, чтобы композиция ДНК включала минигенную конструкцию ДНК, кодирующую экспрессируемый в иммунных клетках иммуногенный полипептид, включающий множество иммуногенных фрагментов цистеиновой эндопептидазы (например, легумаина), которая интенсивно экспрессируется в опухолеассоциированных клетках, где полипептид способен вызывать иммунный ответ против опухолеассоциированных клеток. Минигенный вариант осуществления - множество иммуногенных фрагментов эндопептидазы, объединенных вместе последовательно пептидами-линкерами между каждыми из последовательных фрагментов в полипептиде.
Предпочтительно, цистеиновая эндопептидаза - легумаин (например, легумаин человека; SEQ ID NO: 2). Конструкция ДНК может кодировать одиночный иммуногенный фрагмент цистеиновой эндопептидазы (например, эпитоп), но предпочтительно кодирует полипептидное включение двух или более иммуногенных фрагментов эндопептидазы.
Термин «конструкция ДНК», используемый в настоящем описании и в приложенной формуле изобретения, относится к структуре ДНК, которая кодирует представляющий интерес белок или полипептид, такой как легумаин или иммуногенный фрагмент легумаина (эпитоп), обобщенно называемой «ДНК легумаина», а также белки, такие как IL-2, CCL21, CD40L и им подобные. Предпочтительно, чтобы каждый иммуногенный фрагмент включал приблизительно 8-10 аминокислотных остатков в длину. Конструкции ДНК включают любую ДНК, которая может быть перенесена в меченые клетки, включая линейную ДНК и плазмидную ДНК, так же как ДНК, которая была включена в генетический материал клетки или вируса. Предпочтительно, конструкция ДНК - это ДНК, которая была включена в вирусный или бактериальный вектор доставки, например непатогенный аттенуированный вирусный или бактериальный вектор. В процессе лечения субъекта композицией по изобретению, ДНК легумаина доставляется в иммунные клетки (например, макрофаги и дендрические клетки), которые затем экспрессируют белок или пептид, включая в его иммуногенный фрагмент. Вирусный или бактериальный носители ДНК легумаина сами не экспрессируют легумаин.
В некоторых предпочтительных вариантах осуществления конструкция ДНК - это минигенная конструкция, которая кодирует иммуногенный полипептид, включающий множество (например, 2-5) иммуногенных фрагментов эндопептидазы, которая активно экспрессируется в ТАМ (например, легумаин). Используемый в настоящем описании термин «миниген» относится к конструкциям ДНК, кодирующим множество частей (фрагментов) интересующего протеина, которые связаны вместе, предпочтительно небольшими пептидами по меньшей мере из трех аминокислот. Такие минигены кодируют полипептиды, включающие иммуногенные фрагменты представляющего интерес белка, но не кодируют целый представляющий интерес белок. Предпочтительно, чтобы минигенная конструкция кодировала полипептид, включающий приблизительно 2-5 иммуногенных фрагментов (например, пептидные последовательности из участков эпитопа белка), предпочтительно соединенных пептидными связями. Миниген может также включать последовательности, которые кодируют лидерные последовательности и/или другие последовательности, используемые для облегчения экспрессии или транспортировки полипептида.
Как отмечено выше, иммуногенные фрагменты в полипептидах соединены между собой предпочтительно пептидами линкерами или спейсерами между каждым фрагментом. Пептид линкер предпочтительно состоит из по меньшей мере трех аминокислотных остатков (например, ААА или AAY). Предпочтительно, конструкция ДНК, кодирующая иммуногенный фрагмент легумаина, также кодирует лидерную последовательность, такую как лидерная последовательность эндоплазматического ретикулума (ER), присоединенную пептидом линкером к N-концу полипептида, кодируемого конструкцией ДНК. Когда конструкция ДНК кодирует полипептид, включающий два или более иммуногенных фрагментов легумаина, лидерная последовательность предпочтительно связывается с первым иммуногенным фрагментом от его N-конца. Предпочтительно, иммуногенные фрагменты эндопептидазы состоят из 8-10 смежных аминокислотных остатков одного или более ее эпитопов.
Иммуногенные фрагменты эндопептидазы, такой как легумаин, включая легумаин человека, могут быть идентифицированы хорошо известными в данной области способами, такими как программа HLA Binding Predictions, представленная на сайте NIH www website Отделом Биоинформатики и Молекулярных Анализов (BIMAS) Национального Института Здоровья (NIH), описание которой приведено в настоящем описании в качестве ссылки.
Композиции ДНК по настоящему изобретению стимулируют образование CTL, которые эффективны против опухолеассоциированных макрофагов, экспрессирующих эндопептидазу. На такие опухолеассоциированные макрофаги активно нацелены CTL, которые продуцируются в ответ на иммунизацию композициями ДНК по изобретению. Элиминация или нейтрализация ТАМ изменяет окружающую микросреду опухоли, что приводит к подавлению роста опухоли и опухолевых метастазов.
Используемый в настоящем описании термин «иммунитет» относится к долговременным терминам иммунологической защиты против вирусных форм возбудителей инфекции или опухолевого антигена. Термин «иммунизация» относится к воздействию антигена патогенного возбудителя, полученного из невирулентного источника, что приводит иммунитет субъекта, подвергшегося лечению, к патогенному фактору.
Конструкция ДНК, используемая в композиции ДНК по настоящему изобретению, включает предпочтительно нуклеиновую кислоту, кодирующую полипептид, содержащий легумаин (например, легумаин человека) или иммуногенные фрагменты легумаина, функционально связанные с регуляторными элементами, необходимыми для экспрессии гена в иммунных клетках. В предпочтительном варианте осуществления, конструкция ДНК кодирует полноразмерный белок легумаина человека (SEQ ID NO: 2) или полипептид, поддерживая при этом высокую степень гомологии, по меньшей мере 80% (например, легумаин свиньи, легумаин мыши и подобные), или его иммуногенный фрагмент, который может вызвать иммунный ответ против клеток, сверхэкспрессирующих легумаин.
В особенно предпочтительном варианте осуществления конструкция ДНК включает минигенное кодирование 2-5 иммуногенных фрагментов легумаина (например, легумаина человека), связанных тремя аминокислотными пептидами-линкерами (например, AAA или AAY), с одним пептидом-линкером между каждым иммуногенным фрагментом.
Пригодные конструкции ДНК, включая минигенные конструкции, предпочтительно включают регуляторные элементы, необходимые для экспрессии нуклеотидов. Такие элементы включают, например, промотор, стартовый кодон, стоп-кодон и сигнал полиаденилирования. К тому же энхансеры часто требуются для экспрессии последовательности, кодирующей иммуногенный белок-мишень. Как известно в данной области, эти элементы предпочтительно функционально связаны с последовательностью, кодирующей необходимый протеин. Регуляторные элементы отобраны предпочтительно такие, какие будут функциональны для видов, в которые они будут вводиться. Желательно, чтобы конструкция ДНК была в форме плазмид или встроена в вирусный или бактериальный вектор. Конструкция ДНК, кодирующая легумаин, изначально может быть встроена в бактериальный вектор трансфекционно, с использованием способов, хорошо известных в данной области. Затем трансформированная бактерия может быть культивирована, чтобы обеспечить готовый бактериальный запас для включения ДНК легумаина в генетический материал бактерии. Культуры таких трансформированных бактерий обеспечивают готовый источник композиций ДНК по настоящему изобретению.
Стартовые и стоп-кодоны желательно включать в виде части нуклеотидной последовательности, кодирующей эндопептидазу или иммуногенный полипептид в композиции ДНК по настоящему изобретению. В матрице с кодирующей последовательностью должны быть стартовый и терминирующий кодоны.
Промоторы и сигналы полиаденилирования, включенные в композицию по настоящему изобретению, отбирались предпочтительно с сохранением функциональности в клетках иммунизированных субъектов.
Примеры промоторов, применимых в композициях по настоящему изобретению, особенно при производстве композиции ДНК генетической вакцины для человека, включают, но не ограничиваются промоторами из Simian Virus 40 (SV40), Mouse Mammary Tumor Virus (MMTV) промотор, Human Immunodeficiency Virus (HIV) такой как HIV Long Terminal Repeat (LTR) промотор, вирус Moloney, Cytomegalovirus (CMV), такой как CMV предранний промотор, Epstein Barr Virus (EBV), Rous Sarcoma Virus (RSV), так же как промоторы из человеческих генов, человеческий актин, человеческий миозин, человеческий гемоглобин, человеческий мышечный креатин или человеческий металлотионеин.
Примеры сигналов полиаденилирования, применимых в композициях ДНК по настоящему изобретению, особенно в производстве композиции ДНК для человека, включают, но не ограничиваются, SV40 сигналами полиаденилирования и LTR сигналами полиаденилирования.
В дополнение к регуляторным элементам, необходимым для экспрессии ДНК, другие элементы также могут быть включены в молекулы ДНК. Такие дополнительные элементы включают энхансеры. Энхансерами могут быть, например, человеческий актин, человеческий миозин, человеческий гемоглобин, человеческий мышечный креатин и вирусные энхансеры как CMV, RSV и EBV.
Регуляторные последовательности и кодоны в целом видозависимы, поэтому с целью максимизировать производство белка регуляторные последовательности и кодоны предпочтительно отбираются по эффективности для иммунизируемых видов. Любой средний специалист сможет произвести конструкции ДНК, функциональные для данного вида, подвергающегося воздействию.
Конструкции ДНК, используемые в настоящих композициях, могут быть «оголенной» ДНК, по определению Restifo et al. Gene Therapy 2000; 7:89-92, соответствующее сообщение которых приведено в настоящем описании в качестве ссылки. Предпочтительно, конструкция ДНК находится в плазмидной форме или ДНК, включенной в генетический материал аттенуированного вируса или аттенуированной бактерии. Используемые носители или переносчики включают биодеградирующие микрокапсулы, иммуностимулирующие комплексы (ISCOM) и липосомы для оголенных конструкций ДНК и различные физиологически доступные буферы для генетически сконструированных живых вирусов или бактерий.
Примеры подходящих аттенуированных живых бактериальных векторов, которые могут быть трансформированы для включения FAP конструкции ДНК, включают Salmonella typhimurium, Salmonella typhi, виды Shigella, виды Bacillus, виды Lactobacillus, Bacille Calmette-Guerin (BCG), Escherichia coli, Vibrio cholerae, виды Campylobacter, виды Listeria или другие подходящие бактериальные векторы, известные в данной области. Предпочтительный вектор - это аттенуированный вектор живой Salmonella typhimurium, особенно когда композиция предназначена для перорального введения. Предпочтительная аттенуированная живая Salmonella typhimurium включает AroA- штаммы, такие как SL7207, или дважды аттенуированная AroA-, dam- штаммы, такие как RE88.
Способы трансформации живых бактериальных векторов с экзогенной конструкцией ДНК хорошо описаны в данной области. См., например, Joseph Sambrook and David W. Russell, Molecular Cloning, A Laboratory Manual, 3rd Ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York (2001) (Sambrook and Russell). После трансформации экзогенный генетический материал встраивается в генетический материал бактерии так, что бактерия репродуцируется, экзогенная ДНК реплицируется вместе с нативной ДНК организма. Таким образом, у однажды трансформированной бактерии нормальные репродуктивные процессы обеспечивают готовый запас экзогенной ДНК.
Представленные вирусные векторы включат Bacteriophages, Herpes вирус, Adenovirus, Adeno ассоциированный вирус, Sindbis вирус, Polio вирус, Vaccinia вирус и Avipox. Способы трансформации вирусных векторов с экзогенной конструкцией ДНК также хорошо описаны в данной области. См. Sambrook и Rassell выше.
Используемые липосомные носители являются однослойными или многослойными везикулами, имеющими мембранную часть, сформированную из липофильного вещества, и внутреннюю водную часть. Водная часть используется в настоящем изобретении с целью содержания полинуклеотидного вещества, предназначенного для доставки в выделенную клетку. В целом предпочтительно, чтобы вещество, формирующее липосомы, имело катионную группу, как четвертичную аммониевую группу, и одну или более липофильные группы, как насыщенные или ненасыщенные алкильные группы, имеющие приблизительно 6-30 углеродных атомов. Одна группа подходящих материалов описана в Европейской Патентной Публикации №0187702 и в дальнейшем обсуждалась в Патенте США №6228844 Wolff et. al., соответствующие сообщения которых приведены в настоящем описании в качестве ссылки. В литературе описано множество других подходящих липосомформирующих катионных липидных соединений. См., например, L. Stamatatos, et al., Biochemistry 1988; 27:3917-3925; и H. Eibl, et al., Biophysical Chemistry 1979; 10:261-271. Альтернативно, может быть использована микросфера, такая как полилактид-когликолидная биодеградируемая микросфера. Как известно в данной области, полинуклеотидная конструкция заключена в капсулу или, другими словами, образует комплекс с липосомой или микросферой для доставки полинуклеотида в ткань.
Другие пригодные носители включают полимерные микросферы, содержащие биодеградируемые поли(ортоэфирные) материалы, такие как описываемые Wang et al., Nat. Mater., 2004; 3(3):190-6. Epub 2004 Feb. 15, соответствующие сообщения которых приведены в настоящем описании в качестве ссылки.
Композиции, реализующие настоящее изобретение, предпочтительно включают конструкции ДНК, такие как иммуногенные фрагменты легумаина человека или его функционального гомолога. Функциональные гомологи легумаина предпочтительно охватывают по меньшей мере 80% идентичность последовательности аминокислотного остатка с человеческим легумаином, более предпочтительно, по меньшей мере приблизительно 90%, наиболее предпочтительно, по меньшей мере 95% идентичности с человеческим легумаином.
ГенБанк (GenBank) - это база данных генетических последовательностей Национального Института Здоровья (NIH), которая представляет снабженную комментариями коллекцию всех доступных публикаций последовательностей ДНК. ГенБанк - это часть Международного Сотрудничества по Базе Данных Нуклеотидных Последовательностей, объединяющего усилия ДНК Базы Данных Японии (DDBJ), Европейской Молекулярной Биологической Лаборатории (EMBL) и ГенБанка Национального Центра Биотехнологической Информации.
Нуклеотидная последовательность нуклеиновой кислоты, кодирующей человеческий легумаин, SEQ ID NO: 1 (ФИГ.8) была опубликована в Каталоге ГенБанка № BC026250, сообщение о котором приведено в настоящем описании в качестве ссылки. Соответствующая последовательность аминокислотных остатков человеческого легумаина - SEQ ID NO: 2 (ФИГ.9).
Последовательность нуклеиновой кислоты ДНК, кодирующей мышиный легумаин, SEQ ID NO: 3 показана на ФИГ.10. Соответствующая аминокислотная последовательность мышиного легумаина - SEQ ID NO: 4 (ФИГ.11).
Chen et al., J. Biological Chem. 1997, 272(12): 8090-8098 (приведено в настоящем описании в качестве ссылки) опубликовал данные о структуре и характеристике свиного легумаина, который имеет примерно 83% идентичность последовательности с человеческим легумаином.
Вследствие природной вырожденности генетического кода, другие последовательности ДНК, кодирующие аминокислотную последовательность человеческого легумаина, могут использоваться в практическом осуществлении изобретения. Такие последовательности ДНК включают также те, что способны гибридизоваться с человеческим легумаином.
Последовательности ДНК, кодирующие человеческий легумаин и которые могут быть использованы согласно изобретению, включают нуклеиновые кислоты, имеющие делеции, добавления или замены различных нуклеотидных остатков, для таких в SEQ ID NO: 1, которые приводят к последовательности, кодирующей тот же генный продукт легумаина. Молекулы ДНК, кодирующие функционально эквивалентные гомологи человеческого легумаина, также могут быть использованы в композициях ДНК по настоящему изобретению.
Генный продукт, кодируемый нуклеиновой кислотой, может также содержать делеции, добавления или замены аминокислотных остатков внутри последовательности аминокислотных остатков легумаина, которые приводят к «молчащему» изменению, приводя таким образом к функционально эквивалентному легумаину. Такие аминокислотные последовательности могут быть сделаны на основе сходства в полярности, заряде, растворимости, гидрофобности, гидрофильности и/или амфипатической природе вовлеченных остатков. Например, отрицательно заряженные аминокислоты включают аспарагиновую и глутаминовую кислоты; положительно заряженные аминокислоты включают лизин и аргинин; аминокислоты с незаряженными полярными концевыми группами, имеющими схожее значение гидрофильности, включают: лейцин, изолейцин, валин, глицин, аланин, аспарагин, глутамин, серин, треонин, фенилаланин, тирозин. Как указывалось в настоящем описании, функционально эквивалентный легумаин относится к белку, который включает один или более эпитопов, которые, когда распознаются Т-клетками, позволяют этим самым Т-клеткам распознать эпитопы легумаина, указывающие на клетки, экспрессирующие легумаин. В предпочтительных вариантах осуществления функционально эквивалентный легумаин имеет последовательность аминокислотных остатков, обладающую по меньшей мере 80% идентичностью последовательности с последовательностью аминокислотных остатков человеческого легумаина (SEQ ID NO: 2), например, по меньшей мере примерно 90% идентичность последовательности или по меньшей мере примерно 95% идентичность последовательности.
Конструкции ДНК, кодирующие легумаин, могут быть сконструированы с целью изменения последовательности, кодирующей легумаин (относительно к ДНК нативного легумаина, SEQ ID NO: 1) для разнообразия окончаний, включая, но не ограничиваясь, изменениями, которые модифицируют процессинг и экспрессию генного продукта легумаина. Например, мутации могут быть введены в ДНК с помощью технологий, которые широко известны в данной области, например сайт-направленный мутагенез, чтобы включить новые сайты рестрикции, изменить характер гликозилирования, фосфорилирования и т.д.
В предпочтительном варианте осуществления композиция ДНК по настоящему изобретению включает конструкцию ДНК, кодирующую иммуногенный полипептид, включающий множество иммуногенных фрагментов человеческого легумаина, и конструкцию ДНК, функционально кодирующую по меньшей мере один иммунный эффекторный белок, обе из которых экспрессируются в иммунных клетках. Используемая в описании и в прилагаемой формуле изобретения фраза «иммунный эффекторный белок» означает белок, вовлеченный в регуляцию пути иммунной системы. Предпочтительно, иммунный эффекторный белок представляет собой цитокин.
Цитокины - это белки и полипептиды, продуцируемые клетками, которые могут влиять на поведение других клеток, такое как клеточная пролиферация, клеточная дифференциация, регуляция иммунных ответов, гемопоэз и воспалительные ответы. Цитокины классифицированы на множество семейств, включая хемокины, гемопоэтины, иммуноглобулины, факторы некроза опухоли, и множества не относящихся к какому-либо семейству молекул. См. основной Оксфордский Словарь Биохимии и Молекулярной Биологии, Переработанное Издание, Типография Оксфордского Университета, 2000; и С.А.Janeway, P.Travers, M.Walport and M.Schlomchik, Immunobiology, Fifth Edition, Garland Publishing, 2001 (здесь и далее «Janeway and Travers»). Краткая классификация цитокинов представлена в «Janeway and Travers», в приложении III, страницы 677-679, соответствующие описания которых включены в настоящем описании в качестве ссылки.
Гемопоэтины включают, например, эритропоэтин, интерлейкин-2, (IL-2, белок из 133 аминокислот, продуцируемый Т-клетками и участвующий в Т-клеточной пролиферации), IL-3, IL-4, IL-5, IL-6, IL-7, IL-9, IL-11, IL-13, IL-15 (белок из 114 аминокислот, подобный IL-2, который стимулирует рост эпителия тонкого кишечника, Т-клеток и NK-клеток), гранулоцитарный колониестимулирующий фактор (G-CSF), гранулоцитарно-макрофагиальный колониестимулирующий фактор (GM-CSF), онкостатин М (OSM) и фактор, ингибирующий лейкемию (LIF).
Интерфероны включают, например, IFN-альфа, IFN-бета и IFN-гамма (гомодимерный белок из 143 аминокислот, продуцируемый Т-клетками и NK-клетками, вовлеченный в активацию макрофагов, повышает экпрессию молекул МНС и антиген-процессирующих компонентов, переключение класса IG и супрессию TH2).
Иммуноглобулины включают, например, B7.1 (CD80) и B7.2 (CD86), каждый из которых совместно стимулирует T-клеточные ответы.
Семейство факторов некроза опухоли (TNF) включает, например, TNF-альфа, TNF-бета (лимфотоксин), лимфотоксин-бета (LT-бета), лиганды CD40, лиганд Fas, лиганд CD27, лиганд CD30, лиганд 4-1BB, Trail и лиганд OPG.
Биологические роли лиганда CD40 (CD40L), в частности его взаимодействие с CD40, экспрессируемых на антигенпрезентирующих клетках во время совместной стимуляции Т-клеточной активации, хорошо известны в данной области. CD40 - это гликопротеин 48 кДа, экспрессируемый на поверхности всех зрелых В-клеток, большинства злокачественных образований из зрелых В-клеток и некоторых ранних В-клеточных острых лейкоцитарных лейкозов, но не экспрессируется на плазматических клетках, Clark, Tissue Antigens 1990, 35:33-36. CD40L, мембранный белок II типа приблизительно 35 кДа, экспрессируется на поверхности Т-клеток в результате распознавания антигена. Члены семейства TNF наиболее биологически активны, когда они экспрессируются как гомотримеры. Не исключено, что при этом CD40L может экспрессироваться как гомотример (CD40LT) путем модификации мотива лейциновой «молнии» из 33 аминокислот, слитой с N-концом всего внеклеточного домена этого лиганда. ДНК CD40LT по сообщениям Gurunathan et al. J.Immunol. 1998, 161:4563 стимулирует клеточные иммунные ответы, такие как индукция IFN-гамма и цитолитическую активность Т-клеток, в результате вакцинации мышей ДНК, кодирующей высокоиммуногенную модель антигена бета-галоктозидазы.
CD40LT - это важный фактор в активации Т-клеток, необходимый для выработки эффективного защитного иммунитета против опухолевых аутоантигенов. Как только МНС-антигены 1 класса:пептидные комплексы поглощаются дендритными клетками (DC) и презентируются наивным Т-клеткам, первый антигенный сигнал доставляется посредством Т-клеточных рецепторов (TCR), за которым следует активация CD40LT. На поверхности Т-клеток CD40LT затем вызывают костимулирующую активность DC посредством взаимодействия CD40-CD40LT. Примированные таким образом, эти антигенпрезентирующие клетки могут экспрессировать костимулирующие молекулы В7.1 (CD80) и В7.2 (CD86), которые посылают второй костимулирующий сигнал Т-клеткам посредством взаимодействия с CD28, событие, обязательное для полной активации Т-клеток для одновременного продуцирования провоспалительных цитокинов INF-гамма и IL12 и осуществления эффекторных функций.
Разнообразные цитокины, не выделенные в отдельную группу, включают, например, фактор роста опухоли-бета (TGF-бета), IL-1альфа, IL-1бета, IL-1RA, IL-10, IL-12 (стимулирующий фактор природных клеток-киллеров; гетеродимер, имеющий цепочку из 197 аминокислот и цепочку из 306 аминокислот, которые задействованы в активацию NK-клеток и индукцию дифференцировки Т-клеток в TH1-подобные клетки), фактор, ингибирующий макрофаги (MIF), IL-16, IL-17 (фактор, индуцирующий продукцию цитокинов, который индуцирует продукцию цитокинов в эпителии, эндотелии и фибробластах) и IL-18.
Хемокины представляют собой семейство цитокинов, которые являются относительно небольшими хемоаттрактантными белками и полипептидами, которые стимулируют миграцию и активацию различных клеток, такую как миграция лейкоцитов (например, фагоцитов и лимфоцитов). Хемокины играют роль при воспалении и других иммунных ответах. Хемокины разделены на множество семейств, включая хемокины C, хемокины CC, хемокины CXC и хемокины CX3C. В названии указано количество остатков цистеина (с) и расстояние между ними в молекулах; хемокины C содержат один цистеин, хемокины CC содержат два следующих друг за другом цистеина, CXC содержат два цистеина, отделенных друг от друга одним аминокислотным остатком, и хемокины CX3C содержат два цистеина, отделенных друг от друга тремя аминокислотными остатками. Хемокины взаимодействуют с большим количеством хемокиновых рецепторов, присутствующих на поверхности клеток. См. Janeway и Travers, приложение IV, страницу 680, которые включены в настоящее описание в качестве ссылки.
Кроме того, хемокины могут обладать иммуномодулирующей активностью и быть вовлечены в иммунные ответы при раке. Например, сообщалось, что мышиный 6Ckine/SLC, мышиный аналог хемокина вторичной лимфоидной ткани человека (SLC), в настоящее время обозначаемый CCL21, вызывает противоопухолевый ответ в опухолевой клеточной линии карциномы толстой кишки C-26. См. Vicari, et al. J. Immunol. 2000; 165 (4): 1992-2000. CCL21 человека и его мышиный аналог, 6Ckine/SLC, относятся к хемокинам СС, которые взаимодействуют с хемокиновым рецептором CCR7. Vicari et al. также сообщалось, что мышиный 6Ckine/SLC (muCCL21) представляет собой лиганд хемокинового рецептора CXCR3. CCL21 человека, muCCL21 мыши и множество других хемокинов вовлечены в регуляцию различных клеток иммунной системы, таких как дендритные клетки, T-клетки и природные клетки-киллеры (NK).
Mig и IP-10 относятся к хемокинам CXC, которые взаимодействуют с рецептором CXCR3, ассоциированным с активированными T-клетками. Лимфотактин представляет собой хемокин C, который взаимодействует с рецептором XCR1, ассоциированным с T-клетками и NK-клетками. Фракталкин представляет собой хемокин CX3C, который взаимодействует с рецептором CX3CR1, ассоциированным с T-клетками, моноцитами и нейтрофилами.
Особенно предпочтительные иммунные эффекторные белки, кодируемые композициями ДНК по настоящему изобретению, включают цитокины IL-2 (гемопоэтин), CCL21 (хемокин), а также лиганды CD40, такие как лигандовый тример (CD40LT), цитокин семейства TNF.
ДНК и белковые последовательности для IL-2 человека были опубликованы в GenBank, Accession № BC070338, описание которой включено в настоящее описание в качестве ссылки. ДНК и белковые последовательности IL-2 мыши были опубликованы в GenBank, Accession № NM 008366, описание которой включено в настоящее описание в качестве ссылки.
Последовательность нуклеиновой кислоты, кодирующей IL-2 человека, представлена на ФИГ.12 (SEQ ID NO: 5), а соответствующая ей аминокислотная последовательность (SEQ ID NO: 6) показана на ФИГ.13.
ДНК и белковые последовательности для CCL21 человека были опубликованы в GenBank, Accession № AB002409, описание которой включено в настоящее описание в качестве ссылки.
Последовательность нуклеиновой кислоты, кодирующей CCL21 человека, представлена на ФИГ.14 (SEQ ID NO: 7), а соответствующая ей аминокислотная последовательность (SEQ ID NO: 8) показана на ФИГ.15.
Лиганд человека CD40 (CD40L) представляет собой белок из 261 аминокислот, в своей наиболее активной форме существующий как триммер (CD40LT). Последовательность ДНК, кодирующая CD40L человека (также известный как CD154), была опубликована в GenBank, Accession № NM 000074, описание которой включено в настоящее описание в качестве ссылки (ФИГ.16, SEQ ID NO: 9). Соответствующая последовательность белка CD40L показана на ФИГ.17 (SEQ ID NO: 10).
Аспекты способов по настоящему изобретению предполагают введение млекопитающему композиции ДНК, содержащей конструкцию ДНК, кодирующую легумаин, который экспрессируется в иммунных клетках млекопитающего. Предпочтительно, млекопитающее является человеком. Композиция может быть введена перорально, внутримышечно, интраназально, внутрибрюшинно, подкожно, внутрикожно или местно в зависимости от особенности лекарственной формы, в которой выпущена композиция. Предпочтительно, композиция выпускается в перорально применяемой лекарственной форме, такой как раствор, суспензия, эмульсия, капсула, таблетка и тому подобное.
Композиция ДНК по изобретению может быть использована, чтобы обеспечить долговременное ингибирование роста опухоли и/или опухолевых метастазов при лечении пациента композицией. В предпочтительном варианте осуществления композиция ДНК вводилась в сочетании с противоопухолевым химиотерапевтическим средством. Композиция ДНК может вводиться вместе с химиотерапевтическим средством в комбинированной лекарственной форме, или композиция и химиотерапевтическое средство могут вводиться в отдельных лекарственных формах и с отдельными интервалами между приемами лекарственных средств, разработанными для фармакологии вводимого химиотерапевтического средства.
Химиотерапевтические средства, используемые в комбинации с композициями ДНК по настоящему изобретению, включат противоопухолевые средства, такие как доксорубицин, паклитаксол, циклофосфамид, этопозид, 5-фторурацил, метотрексат и так далее.
Композиции ДНК по настоящему изобретению приготавливают предпочтительно с фармацевтически приемлемыми носителями или наполнителями, такими как вода, физиологический раствор, декстроза, глицерин и так далее, и их комбинациями для целей приготовления и введения композиции. Композиции могут также содержать вспомогательные вещества, такие как смачивающие компоненты, эмульгирующие вещества, буферы, и другие вспомогательные вещества, которые хорошо известны в фармацевтической отрасли.
Композиции по настоящему изобретению предпочтительно перорально вводят млекопитающим, например человеку, в виде раствора или суспензии в фармацевтически приемлемом носителе с концентрациями ДНК в пределах от приблизительно 1 до приблизительно 10 микрограмм на миллилитр, в зависимости от массы ДНК, кодирующей легумаин. Наиболее предпочтительной лекарственной формой для композиции ДНК по изобретению является суспензия аттенуированной легумаин-трансфицированной бактерии в подходящем буферном растворе, которая может быть приготовлена для перорального введения. Подходящая дозировка композиции будет зависеть от субъекта, получающего вакцину, активности композиции и частично от решения лечащего врача, осуществляющего введение или назначение композиции.
Вводимая млекопитающему дозировка и режим введения, в случае назначения более одного введения, будет зависеть от млекопитающего и лекарственного средства. Эффективные количества дозировки и режимы введения могут быть определены эмпирически через клинические изучения эффекта-дозы, как хорошо известно в данной области. Дозировку и режим введения выбирают, чтобы обеспечить достаточное количество экспрессии легумаина в иммунных клетках с целью вызвать иммунный ответ у млекопитающего против опухолеассоциированных макрофагов, экспрессирующих легумаин. Предпочтительно, чтобы дозировка композиции, вводимой млекопитающему, вызывала экспрессию антигена легумаина в иммунных клетках млекопитающего в количестве, достаточном для поддержания иммунного ответа против легумаин-презентирующих опухолеассоциированных макрофагов, который будет продолжаться в течение периода по меньшей мере месяц, например, по меньшей мере 6 месяцев или по меньшей мере приблизительно 1 год. В некоторых предпочтительных вариантах осуществления, дозировка легумаин-трансформированных клеток, введенных пациенту, составляет приблизительно 1×108 клеток, трансфицированных с приблизительно 0,3-0,8 мкг легумаина ДНК.
Композиции по настоящему изобретению могут быть упакованы в подходящие стерильные емкости, такие как ампулы, флаконы или пузырьки, в виде либо многократной, либо однократной лекарственной формы. После заполнения композицией ДНК, емкости предпочтительно герметизируют. Предпочтительно, композиции упаковывают в емкости с прикрепленной к ним этикеткой, которая маркирует композицию и на которой представлена запись в форме, предписанной правительственным учреждением, таким как Управление по контролю за качеством пищевых продуктов и медикаментов Соединенных Штатов, отражающая одобрение композиции согласно соответствующим законам, информацию о дозировке и тому подобное. Этикетка предпочтительно содержит информацию о композиции, которая может использоваться для введения композиции пациенту медицинским работником. Упаковка также предпочтительно содержит напечатанные информационные материалы, касающиеся введения композиции, инструкции, указания и любые обязательно необходимые предупреждения.
Следующие примеры предназначены для дополнительной иллюстрации характерных особенностей и осуществлений настоящего изобретения и не предназначены для его ограничения.
Материалы и способы
Животные, штаммы бактерий и клеточные линии. Самок мышей BALB/с и C57BL/6, 6-8 недельного возраста получали от Scripps Research Institute Rodent Breeding Facility. Дважды аттенуированный штамм RE88 (aroA-, dam-) S. typhimurium был получен от Remedyne Corporation, Goleta, CA. Клеточная линия СТ-26 рака толстой кишки мыши была любезно предоставлена Dr. I. J. Fidler (MD Anderson Cancer Center), а клетки немелкоклеточной карциномы легкого мыши D121 - подарок от Dr. L. Eisenbach (Weizmann Institute of Science, Rehovot, Israel). Клетки карциномы молочной железы мыши 4Т1 были любезно предоставлены Dr. Suzanne Ostrand-Rosenberg (University of Maryland).
Иммуногистохимические анализы. Иммуногистохимические анализы были выполнены на опухолевых тканях 4Т1 и срезах матригелевых пробок. Экспрессия легумаина макрофагов определялась на срезах опухолевой ткани 4Т1 с помощью биотинилированных крысиных противомышиных CD68 mAb (BD Bioscience Pharmingen) с GFP-конъюгированным стрептавидином в качестве вторичного репортерного реагента. Кроличья антилегумаиновая антисыворотка была приготовлена с помощью иммунизации очищенным человеческим легумаином, продуцированным в E.coli. (Ishii, Methods Enzymol. 1994; 244:604-615). Реакция была визуализирована с помощью конъюгированного с техасовым красным стрептовидина. Дополнительно, срезы опухолевой ткани 4Т1 и срезы матригелевой пробки фиксировались и окрашивались MMP-9, VEGF, TGF-бета и антителами F4/80 (eBioscience, San Diego, CA) для срезов опухолевой ткани 4Т1, в то время как антитела CD68 и CD31 (BD Bioscince Pharmingen) использовались для срезов матригелевой пробки. Все тканевые срезы были визуализированы с помощью техасского красного или конъюгированного с GFP стрептавидина как вторичного репортерного реагента, и препараты анализировались лазерным сканированием с помощью конфокальной микроскопии (Bio-Rad Laboratories). Все снимки были зафиксированы SPOT™ охлажденной цветной цифровой камерной системой (Diagnostic Instruments. Inc).
Иммунизация и введение опухолевых клеток. Профилактическая модель: мыши BALB/с или C57BL/6 каждые были разделены на три экспериментальные группы (n=8) и иммунизированы PBS, пустым вектором или S. typhimurium, трансфицированной pUb-легумаином. Всем мышам внутривенно (i.v.) вводилось приблизительно 5×104 клеток СТ-26 (BALB/с), приблизительно 2×105 клеток D121 (C57BL/6) или инъекция жира молочных желез с приблизительно 7×103 клетками 4Т1 (BALB/с), 1 неделю после последней иммунизации, чтобы вызвать либо экспериментальные, либо спонтанные легочные метастазы. Массы легких в экспериментальных и контрольных группах были определены через 24 дня после введения опухолевых клеток. Терапевтическая модель: мыши BALB/с были разделены на три экспериментальные группы (n=8), которым сначала в жировое тело ввели приблизительно 7×103 клеток 4Т1 в нулевой день и затем трижды иммунизировали композицией ДНК по изобретению на 3, 7 и 11 день. После 24 дней первичная опухоль была иссечена для определения масс легких мышей и оценки метастазирования, или коэффициента выживаемости мышей.
Истощение клеток CD4 + или CD8 + in vivo, цитотоксичность и данные анализов ELISPOT. Истощение клеток CD4+ или CD8+ in vivo было представлено, как ранее описывалось (Ceredig et al. 1985, Nature 314:98-100). Цитотоксичность была измерена и просчитана c помощью стандартного теста с радиоактивным 51Cr, как уже сообщалось (Zhou et al. 2005, Blood, 106:2026-2032). Анализы ELISPOT были выполнены с помощью набора ELISPOT (BD Bioscience Pharmingen) согласно инструкциям, данными производителем.
Анализ ангиогенеза в матригеле in vivo. Матригель был использован для оценки суппрессии ангиогенеза после вакцинации. Коротко говоря, мышам BALB/с через 2 недели после последней вакцинации подкожно (s.c.) в область грудины был введен ростзамедляющий фактор Матригель (BD Bioscience), содержащий VEGF или bFGF-2 (приблизительно 200 нг/пробку) и опухолевые клетки 4Т1 (приблизительно 5×103/пробку), которые были предварительно облучены гамма-радиацией в 1000 Гр (приблизительно 100000 рад). Эндотелий был окрашен через 6 дней после имплантации Матригеля с помощью в/в инъекции с помощью лектина I Bandiera simplofica (Изолектин В4), конъюгированного с флуоресцеином (Vector Laboratories). Это производилось параллельно с окрашиванием эндотелия контрольных животных. Спустя приблизительно 30 минут мыши были умерщвлены, матригелевые пробки извлечены, и флуоресценция оценена с помощью флуориметрии. Дополнительно, матригелевые пробки были удалены спустя 6 дней после имплантации матригеля, фиксированы в растворе Буэна в течение 24 часов и затем залиты в парафин. Из всех тканей были сделаны срезы по количеству препаратов и окрашены трихромом Массона. Все снимки были произведены SPOT™ охлажденной цветной цифровой камерной системой, как было описано выше.
Проточная цитометрия (FACS). Активированные маркеры Т-клеток были оценены с помощью анализа двухцветовой проточной цитометрии на анализаторе BD Biosciences FASCALIBUR@. Маркеры DC-клеток были определены окрашиванием свежекультивированных лимфоцитов от успешно вакцинированной мыши и контрольной мыши с анти-CD11c Ab в комбинации с FITC-конъюгированными анти-CD40, CD-80 и Ab против МНС-антигенов II класса. Макрофаги, имеющие высокие уровни CD206 и F4/80, были подсчитаны с помощью двухцветового проточного анализа. Опухолевые клетки были культивированы из успешно вакцинированной мыши BALB/с и затем окрашены анти-CD206 антителами, конъюгированными с РЕ (Cell Science, Inc.), антителами анти-F4/80, конъюгированными с АРС, и антилегумаиновыми антителами, конъюгированными с FITC, за чем следовал FACS-анализ. Все антитела были закуплены в Pharmingen, San Diego, СА. IFN-гамма, высвобожденный с внутриклеточного уровня, был определен в лимфоцитах пейеровых бляшек, полученных спустя 3 дня после периода иммунизации и окрашенных антителами анти-CD8, меченными АРС. Клетки были фиксированы, пермеабилизированы и в дальнейшем окрашены РЕ-мечеными анти-IFN-гамма антителами для определения внутриклеточной экспрессии IFN-гамма.
Анализ миграции. Анализ клеточной миграции был произведен с использованием модифицированных камер Бойдена (Transwell, Corning Inc., NY). Опухолевые клетки были получены из опухолевой ткани либо пролеченной, либо контрольной групп мышей для выполнения трансвелл-анализа миграции. После культивирования в течение 4 часов клетки на нижней поверхности мембраны были фиксированы 1% параформальдегидом, окрашены 1% кристальным фиолетовым и подсчитаны (Shi et al. 2004, Mol. Cancer Res. 2:395-402).
Статистический анализ. Статистическая значимость разницы результатов между экспериментальными и контрольными группами была определена тестом Стьюдента. Результаты были расценены как значимые, если двусторонние величины Р были <0,05. Анализ Каплана-Мейера использовался для оценки выживаемости мышей.
ПРИМЕР 1
Векторная конструкция, экспрессия белка и трансформация S. typhimurium плазмидами ДНК. Две конструкции были приготовлены, основываясь на векторе pCMV, который имеется в продаже от Invitrogen, Carlsbad, CA. Конструкция pUb-легумаина включала полиубиквитинированный непроцессированный мышиный легумаин. ДНК непроцессированного мышиного легумаина имеет нуклеотидную последовательность, показанную на ФИГ.10, SEQ ID NO: 3 (последовательность аминокислотных остатков мышиного легумаина показана на ФИГ.11, SEQ ID NO: 4). Контролем служила конструкция пустого вектора. Мышиный легумаин был получен из клеток 4Т1 рака молочной железы с использованием общей РНК как матрицы способом ПЦР. Экспрессионный вектор был получен на основе pCMV-клеточного вектора (Invitrogen), содержащего полиубиквитинированную последовательность, клонированную перед легумаиновой последовательностью. Последовательность нуклеиновой кислоты полиубиквитинированного мышиного легумаина показана на ФИГ.18 (SEQ ID NO: 11). Последовательность аминокислотных остатков убиквитированного мышиного легумаина показана на ФИГ.19 (SEQ ID NO: 12). Экспрессия белка легумаина была продемонстрирована с помощью вестерн-блоттинга с поликлональными кроличьими антителами против мышиного легумаина, а также противомышиными антителами к бета-актину (Santa Cruz Biotechnology, Inc.) в качестве loading контроля. Специфический белок был выявлен с помощью козьих противокроличьих HPR-антител, конъюгированных с IgG (Bio-Red Laboratories). Аттенуированные Salmonella typhimurim были трансдуцированы плазмидами ДНК вакцины с помощью электропорации, как описано в Luo et al. 2003, Proc. Natl. Acad. Sci. U.S.A. 100:8850-8855 и Xiang et al. 2000, Proc. Natl. Acad. Sci. U.S.A. 97:5492-5497.
ПРИМЕР 2
Иммуногенные фрагменты мышиного легумаина. Две плазмиды, включающие каждая миниген легумаина, кодирующий три иммуногенных фрагмента легумаина, соединенные вместе спейсером из 3-х аминокислот (AAY) между каждым фрагментом, были получены включением минигена легумаина в плазмиду pCMV/myc/ER MCS, которая имеется в продаже от Invitrogen, Carlsbad, CA (см. ФИГ.20 и 21). Вектор включает сегмент, кодирующий ER-сигнальный пептид, myc-эпитоп и ER-удерживающий сигнал (см. ФИГ.21). Вставка минигена была произведена между сайтом BssH II в части ER-сигнального пептида и сайтом Xho I, как показано в ФИГ.21. Первая легумаиновая минигенная плазмида (pCMV-Db/Dd; обозначаемая также pH-2Dd на фигурах) включает спейсер AAY, иммуногенный легумаиновый фрагмент legu137, спейсер AAY, иммуногенный легумаиновый фрагмент legu238, спейсер AAY и иммуногенный легумаиновый фрагмент legu223. Вторая легумаиновая минигенная плазмида (pCMV-Kb/Kd; обозначаемая также pH-2Kd на фигурах) включает спейсер AAY, иммуногенный легумаиновый фрагмент legu405, спейсер AAY, иммуногенный легумаиновый фрагмент legu180, спейсер AAY и иммуногенный легумаиновый фрагмент legu229. На ФИГ.20 показаны последовательности аминокислотных остатков для иммуногенных легумаиновых фрагментов, а также показатели связывания MHC первого класса и идентификационный номер последовательности (SEQ ID NO) для каждого фрагмента. Экспрессия пептида проверялась с помощью Вестерн-блоттинга клеток COS-7, трансфицированных с моноклональным anti-myc антителом (Invitrogen). Как только экспрессия пептида подтверждалась, перед myc эпитопными последовательностями вектора немедленно вставлялся стоп-кодон. Каждый из полученных минигенных плазмидных векторов (pCMV-Db/Dd и pCMV-Kb/Kd) проверялся определением нуклеотидных последовательностей, а затем трансфицировался в аттенуированную бактерию S. typhimurium RE88 с помощью электропорации, чтобы доставить в композиции ДНК по изобретению минигены, кодирующие легумаин. Как описывается в деталях ниже, эти композиции были эффективны для борьбы с карциномой молочной железы 4Т1 и D2F2 у мышей BALB/c. Наблюдаемый опухолезащитный ответ был опосредован CD8 Т-клетками, которые выборочно уничтожали legumain+ опухолеассоциированные клетки макрофагов, что приводило к значительной супрессии опухолевого ангиогенеза.
Пероральная иммунизация и введение опухолевых клеток. Группам мышей BALB/c (n=8) 3 раза с однонедельным интервалом принудительно перорально вводили 100 мкл PBS, содержащей приблизительно 5×108 CFU дважды аттенуированной S.typhimurium, содержащую или пустой вектор, pCMV-Db/Dd плазмиду, или pCMV-Kb/Kd плазмиду. Спустя 2 недели после последней вакцинации мышам вводили внутривенно или подкожно клеточные линии D2F2 карциномы.
Цитотоксический анализ. Цитотоксичность определяли посредством стандартного анализа высвобождения 51Cr, как описывалось выше. Процент специфического лизиса клеток-мишеней вычисляли по формуле [(E-S)/(T-S)]×100, где E представляет собой среднюю величину экспериментального высвобождения, S представляет собой среднюю величину самопроизвольного высвобождения и T представляет собой среднюю величину полного высвобождения.
Анализ ELISPOT. Спленоциты выделяли у всех экспериментальных групп мышей BALB/c 2 недели спустя после введения D2F2 опухолевых клеток и культивировали в течение 24 ч с облученными (1000Гр) 4Т1 клетками или с 4Т1 клетками, свежеполученными из 4Т1 опухолевой ткани молочной железы. Исследование проводилось в соответствии с инструкциями производителя (BD Bioscience, San Jose, CA).
Анализ антиангиогенной активности. Супрессия ангиогенеза определялась анализом Матригеля, как описано выше. Рост сосудов в Матригеле определялся измерением концентрации гемоглобина с использованием реагента Драбкина (водный раствор, содержащий 1 г/л NaHCO3, 0,05 г/л KCN и 0,2 г/л K3Fe(CN)6). Пробки Матригеля были извлечены через 6 дней после имплантации Матригеля, фиксированы в растворе Буэна (15 весовых частей насыщенного водного раствора пикриновой кислоты, 5 весовых частей 37% водного раствора формалина и 1 весовая часть ледяной уксусной кислоты) в течение 24 часов и залиты в парафин. Из всех тканей были сделаны срезы по количеству препаратов и окрашены трихромом Массона. Все снимки были произведены SPOT™ охлажденной цветной цифровой камерной системой. Используя минигенный подход по настоящему изобретению, изогенным мышам BALB/c минигенные анти-ТАМ композиции ДНК подавляли как опухолевый рост, так и ангиогенез. Шесть иммуногенных пептидов легумаина были определены как Н-2Dd-1,2,3, или Н-2Kd-1,2,3 рестриктированные минигены, основываясь на предсказании связывания этих молекул МНС-антигена 1 класса HLA пептидом с помощью Программы предсказания связывания пептида HLA, обеспеченной Отделом Биоинформатики и Молекулярных Анализов (BIMAS) на вебсайте NIH. Аминокислотные последовательности этих пептидов и их связывающие активности, как прогнозировано программой DNASTAR@ (DNAStar, Inc., Madison, WI), перечислены на ФИГ.20.
Минигенная вакцина pCMV-Kb/Kd защищает мышь от введения клеток опухоли молочной железы D2F2 и предотвращает легочное метастазирование. Первоначально, минигенные композиции ДНК были опробованы в профилактической модели карциномы молочной железы, когда мыши были сначала вакцинированы минигенной композицией, и затем подкожно им были введены клетки карциномы молочной железы мышей D2F2 (ФИГ.22). Заметное замедление опухолевого роста наблюдалось у изогенных мышей BALB/c, вакцинированных pCMV-Kb/Kd, но не pCMV-Db/Dd. В противоположность, у всех мышей, вакцинированных только пустым контрольным вектором, был выявлен быстрый подкожный опухолевый рост.
Минигенная вакцина легумаина pCMV-Kb/Kd подавляет легочное метастазирование D2F2, разрушая ТАМ, как доказано заметным подавлением экспериментальных метастазов, выявленным у мышей BALB/c с внутривенно введенными клетками карциномы молочной железы D2F2 через 2 недели после третьей вакцинации. В этой модели метастазы предотвращались, в отличие от pCMV-Db/Dd. Мыши, вакцинированные только пустым контрольным вектором, обнаруживали одинаковый быстрый метастазированный рост легочной опухоли (ФИГ.23).
Минигенная вакцина вызывает CTL ответ, который способен уничтожать legumain + клетки. С целью изобразить специфичный ответ Т-клеток, получаемый после минигенных вакцинаций, Т-клеточная активация была продемонстрирована с помощью специфичного высвобождения INF-гамма активированными Т-клетками (ФИГ.24, панель В). Эти клетки были стимулированы клетками, полученными из свежей 4Т1 опухолевой ткани, обильно экспрессирующей легумаин, как было показано анализами с помощью проточной цитометрии (ФИГ.24, панель В). Важно, что цитотоксические анализы ясно продемонстрировали значительную CTL активность только у иммунизированных мышей. Клетками-мишенями были клетки мышиной макрофаговой клеточной линии RAW, культивировавшейся с IL-4, IL-10 и IL-13 цитокинами, чтобы вызвать экспрессию легумаина. Это было подтверждено также с помощью Вестерн-блотт анализов (ФИГ.25, Панель А), когда было показано, что дикий тип RAW клеток является легумаин негативным. Спленоциты, выделенные у контрольных мышей, которых лечили пустым вектором, показали аналогичное фоновое уничтожение RAW клеток, позитивное или негативное по отношению к экспрессии легумаина (ФИГ.25, Панель В). Однако спленоциты из мышей, которых лечили pCMV-Kb/Kd, вызывали значительно более сильное действие против legumain+ клеток-мишеней, чем против таких же клеток, выделенных у мышей, которых лечили с помощью pCMV-Db/Dd или пустым вектором (ФИГ.25, Панель В). Эти данные демонстрируют избирательность pCMV-Kb/Kd минигениндуцированной активности CTL и их способности избирательно уничтожать legumain+ макрофаги.
pCMV-Kb/Kd минигенная вакцина вызывает антиангиогенные эффекты. С целью проанализировать степень, в которой атиогенезис играет ключевую роль в противоопухолевой защите, индуцированной вакциной pCMV-Kb/Kd, были проведены матригельные анализы, в которых вызывалось образование кровеносных сосудов внутри Матригеля с помощью рекомбинантной bFGF. Разница в образовании кровеносных сосудов между различными лечебными группами определялась измерением соответствующей концентрации гемоглобина (Hb) в матригельной пробке, полученной от иммунизированных или контрольных мышей. Так, мыши, которых лечили вакциной pCMV-Kb/Kd, показывали в среднем четкое снижение соответствующей концентрации Hb (ФИГ.26, панель А). Более того, когда участки Матригеля от иммунизированных и контрольных мышей анализировались с помощью трехцветного окрашивания по Масону, участки от мышей контрольной группы пустого вектора обнаруживали обильные многочисленные кровеносные сосуды. Напротив, кровеносные сосуды были явно снижены в секции Матригеля от мышей, которых лечили PCMV-Kb/Kd (ФИГ.26, Панель B). Эти данные демонстрируют, что иммунизация вакциной PCMV-Kb/Kd приводит к снижению опухолевой васкуляризации. Взятые вместе, эти открытия свидетельствуют, что pCMV-Kb/Kd минигенные композиции вызывали значительные антиангиогенные эффекты, которые способствовали защите BALB/c мышей от введения D2F2 опухолевых клеток молочной железы в профилактической модели.
В отдельных экспериментах с немелкоклеточными клетками D121 карциномы изогенных C57/BL мышей, как pCMV-Kb/Kd, так и pCMV-Db/Dd вакцины обеспечивали эффективные анти-ТАМ иммунные ответы.
ОБСУЖДЕНИЕ РЕЗУЛЬТАТОВ
Легумаин служит в качестве цели для уничтожения сверхэкспрессирующих ТАМ во время развития опухоли. Хорошо известно, что ТАМ играют важную роль в развитии опухоли и матастазировании. Таким образом, выделение этих М2 макрофагов является новой противоопухолевой стратегией. Легумаин был изначально определен как достоверный маркер молекул ТАМ по причине его высоких уровней экспрессии на этих клетках в окружающей микросреде опухоли и строме. ТАМ были выделены из мышиной 4Т1 опухолевой ткани молочной железы. Проточная цитометрия (FACS) селезенки показала, что легумаин сверхэкспрессируется на CD206 и F4/80 дважды положительных М2 макрофагах, особенно при сравнении с нормальными М1 макрофагами (ФИГ.1, Панель B). Результат был также подтвержден с помощью иммуногистохимических анализов, показавших, что ТАМ могут быть визуализированы H/E окрашиванием, и сверхэкспрессия легумаина была в дальнейшем обнаружена двойным окрашиванием антилегумаиновых антител (Ab) (зеленый) в сочетании с анти-CD68 Ab (красный) (ФИГ.1, Панель А). Эти данные показывают, что инфильтрированные ТАМ являются непропорционально большой клеточной субпопуляцией в 4Т1 опухолевой ткани и что легумаин является потенциально эффективной мишенью для уничтожения ТАМ.
Индукция экспрессии легумаина на ТАМ с помощью Th2 цитокинов. Мышиная макрофаговая клеточная линия, RAW, совместно культивируемая с этими цитокинами, была использована для оценки степени, в которой экспрессия легумаина на ТАМ была вызвана Th2 цитокинами, такими как IL-4, IL-10 и IL-13. Одновременно с повышенной регуляцией легумаина наблюдалось значительное увеличение экспрессии CD206+, F4/80+ этими RAW клетками (ФИГ.1, Панель С). Эти результаты были подтверждены с помощью Вестерн-блоттинга (ФИГ.1, Панель D). Не было получено данных об экспрессии легумаина опухолевыми клеточными линиями, культивируемыми с этими цитокинами. Эти результаты свидетельствуют, что Th2 цитокины, такие как IL-4, IL-10 и IL-13, высвобождаются опухолевыми и другими опухолевыми стромальными клетками и накапливаются в окружающей микросреде опухоли. В этой окружающей среде, цитокины могут потенциально вызывать пролиферацию и трансформацию от М1 макрофагов до популяции с фенотипом М2, сверхэкспрессирущей легумаин.
Нацеливание на ТАМ подавляет развитие опухоли. Рост опухоли и опухолевые метастазы сильно коррелируют с присутствием ТАМ, и таким образом нацеливание на эту субпопуляцию макрофагов ведет к подавлению опухолевого роста и метастазов. С целью вызвать иммунный ответ против этих ТАМ был подготовлен экспрессирующий вектор для композиции ДНК, кодирующий легумаин. Аттенуированная бактерия Salmonella typhimurium (штамм RE88) была трансфицирована плазмидным вектором, кодирующим легумаин с целью обеспечить средство доставки ДНК легумаина в иммунные клетки. Контрольная композиция была приготовлена путем включения пустого плазмидного вектора в такой же штамм аттенуированной бактерии. ФИГ.2, Панель А схематично показывает карту конструкции вектора, основанную на pCMV/myc/cyto векторном остове, включающем кодирующую легумаин ДНК. Ген, кодирующий легумаин, был присоединен к C-концу мутантного полиубиквитина (pLegumain) с целью улучшить процессирование антигена в протеасоме. Затем целый фрагмент был вставлен между PstI и NotI сайтами рестрикции плазмиды. Экспрессия легумаина была продемонстрирована с помощью Вестерн-блоттинга. В профилактической, так же как и в терапевтической моделях снижение количества ТАМ с помощью Композиции ДНК, основанной на легумаине, по настоящему изобретению, эффективно ингибирует спонтанные метастазы 4Т1 опухоли молочной железы и метастазы немелкоклеточных карцином легкого D121 и толстой кишки CT26 на мышиных моделях.
В профилактической модели, мыши C57BL/6J иммунизировались три раза физиологическим раствором с фосфатным буфером (PBS; контрольная группа 1), аттенуированной S.typhimurium, со встроенным пустым вектором (контрольная группа 2) или композицией ДНК по изобретению (pLegumain-трансфицированная аттенуированная S.typhimurium; лечебная группа). Спустя одну неделю после последней иммунизации этим мышам внутривенно (i.v.) ввели приблизительно 2×105 клеток немелкоклеточной D121 опухоли легкого. Через 24 дня после этого измеряли и анализировали легочные метастазы. В двух контрольных группах средний вес легкого был значительно больше, чем у лечебной группы (ФИГ.2, Панель В). Похожие результаты были получены в модели CT26 опухоли толстой кишки и в модели 4Т1 раковой опухоли молочной железы изогенных BALB/c мышей (ФИГ.2, Панель В).
В более сложной терапевтической модели, BALB/c мышам сначала вводили клетки 4Т1 рака молочной железы, а затем, как описывалось выше, иммунизировали три раза pLegumain основанной Композицией ДНК (лечебная группа), PBS (контроль) или композицией с пустым вектором (контроль). Спустя двенадцать дней после введения 4Т1 опухолевых клеток, первичную опухоль хирургически удаляли. График продолжительности жизни для контрольных и лечебной групп показал, что 75% (6/8) мышей, иммунизированных pLegumain, оставались в живых в течении 3 месяцев. С другой стороны, мыши из контрольных групп все погибли в течение одного месяца (ФИГ.2, Панель С). Эти данные показывают, что легумаин композиции ДНК, основанные на легумаине, по настоящему изобретению эффективно подавляют рост опухолевых клеток и метастазы в мышиных моделях 4Т1 рака молочной железы, немелкоклеточном D121 раке легкого и СТ-26 карциноме толстой кишки. В сочетании с хирургическим вмешательством, это лечение способно, в самом деле, значительно увеличить продолжительность жизни мышей путем ингибирования опухолевых клеточных метастазов в этих очень серьезных (перспективных) терапевтических мышиных опухолевых моделях.
Нацеливание на легумаин вызывает специфичный CD8+ CTL ответ, снижающий популяции ТАМ в опухолевой строме. Противолегумаиновая иммунизация вызывает специфичный Т-клеточный ответ против ТАМ, обильно экспрессирующих эту аспарагиновую эндопептидазу. Специфичный Т-клеточный ответ демонстрировали посредством анализа высвобождения 51Cr, где спленоциты, выделенные из мышей, успешно иммунизированных композицией ДНК по изобретению, эффективно уничтожали RAW макрофаги, экспрессирующие высокий уровень легумаина после культивирования с цитокинами IL-4, IL-10 и IL-13. Эти же спленоциты не смогли вызвать цитотоксичное уничтожение клеток, не экспрессировавших легумаин (ФИГ.3, Панель А), что говорит о высоком уровне специфичности этого Т-клеточного ответа против легумаина. Кроме того, такой же результат был получен с использованием легумаин трансфицированных клеток как мишеней в анализе высвобождения 51Cr (ФИГ.7). Более того, результаты, отображенные на ФИГ.3, Панель В, демонстрируют резкое сокращение популяции F4/80+/CD206+ макрофагов после лечения композицией ДНК по изобретению, основанной на легумаине. Эти данные были также подтверждены иммуногистохимическим окрашиванием, как показано на ФИГ.3, панель С.
Рестриктированный по MHC первого класса CD8 + CTL специфично активен против ТАМ. Не ограничиваясь какой-либо теорией, предполагают, что трансфицированные бактерии поглощаются Пейеровыми бляшками кишечника, где ДНК легумаина затем встраивается в иммунные клетки, такие как макрофаги и дендритные клетки (ДК), которые потом экспрессируют легумаин ДНК. Было обнаружено, что ДК в Пейеровых бляшках успешно иммунизированных мышей активируются спустя 3 дня после вакцинации pLegumain, как показали маркеры активации, CD40, CD80 и MHC-I, активирующие DC (ФИГ.4, панель А). Было также обнаружено, что CD8+ Т-клеточная активация является специфичной к легумаину, о чем свидетельствуют двойное окрашивание INF-гамма и CD8 на спленоцитах, полученных от успешно иммунизированных мышей (ФИГ.4, Панель В), и специфичное высвобождение INF-гамма активированными Т-клетками, стимулированными легумаин-положительными клетками (ФИГ.4, панель С). К тому же, in vivo иммунное истощение CD4+ или CD8+ Т-клеток показало, что только CD8+ Т-клетки играют значительную роль в специфичном цитотоксическом уничтожении ТАМ, так как только их истощение полностью аннулировало уничтожающий эффект. Специфичная цитотоксичность была рестриктирована по антигенам МНС первого класса, так как уничтожение было специфично ингибировано анти-H-2Dd/H-2Kd антителами, (ФИГ.4, панель D). Рассмотренные вместе эти результаты показывают, что композиции ДНК по изобретению, основанные на легумаине, сначала активировали DC в Пейеровых бляшках, после чего эти клетки презентировали легумаиновые пептиды по пути MHC антигена первого класса к Т-клеточным рецепторам (TCR) на активированных CD8+ Т-клетках, что приводило к специфичному цитотоксичному ответу CD8+ Т-клеток, нейтрализующему ТАМ.
Нейтрализация ТАМ в опухолевой строме снижает высвобождение факторов опухолевого роста и факторов проангиогенеза, а также ингибирует миграцию опухолевых клеток и метастазы. ТАМ могут оказывать влияние на метастазы опухоли несколькими путями, поскольку они секретируют большое разнообразие факторов роста опухоли, факторов проангиогенеза и опухолеассоциированные ферменты, стимулирующие опухолевый ангиогенез, а также опухолевый рост и метастазы. С целью оценить степень, в которой элиминация ТАМ реально снижает высвобождение некоторых из этих факторов, от вакцинированных мышей и подходящих контрольных животных были получены сыворотка и клетки опухолевой стромы. Свежеполученные опухолевые клетки были культивированы, и их кондиционированные среды (супернатанты) собирались через 24 и 48 часов, соответственно. ELISA, выполненный с целью определения количества TNF-альфа, VEGF и TGF-бета, показал значительное снижение TNF-альфа и VEGF в обеих кондиционированных средах опухолевых клеток и сыворотке. TGF-бета был понижен только в клеточных кондиционированных средах, но не в сыворотке (ФИГ.5, Панель А).
Иммуногистологическое окрашивание подтвердило снижение экспрессии этих факторов в опухолевой ткани (ФИГ.5, Панель В). К тому же значительное снижение миграции опухолевых клеток наблюдалось при сравнении лечебной и контрольной групп (ФИГ.5, Панель С) в анализах миграции и инвазии. Эти результаты показывают, что характеристики опухолевых клеток изменились после вызванного вакциной изменения окружающей микросреды опухоли, послужившего причиной подавления ТАМ. Способность контролировать опухолевые метастазы была подтверждена анализом in vivo с помощью определения показателя метастазирования и массы легких спустя 24 дня после удаления первичной опухоли в терапевтической модели. Показатель метастазирования и массы легких у пролеченных мышей значительно снизились по сравнению с двумя контрольными группами (ФИГ.5, Панель D).
Элиминация ТАМ в опухолевой строме приводит к снижению опухолевого ангиогенеза. Значительный антиангиогенный эффект наблюдался после элиминации ТАМ в опухолевой строме, что могло частично происходить благодаря тому факту, что эти макрофаги М2 производят широкий ряд факторов проангиогенеза. Анализы Матригеля выявили новые кровеносные сосуды, разросшиеся in vivo, что было определено количественно с помощью окрашивания эндотелия меченым FITC изолектином В4. Эти результаты четко показывают, что рост сосудов был значительно снижен после вакцинации композицией ДНК по настоящему изобретению (ФИГ.6, Панель В). Наблюдалось, что наибольшее количество кровеносных сосудов вырастало в матригелевой пробке у контрольных мышей, иммунизированных пустым вектором, по сравнению с мышами, иммунизированными композицией ДНК, что было выявлено цифровым изображением и окрашиванием с помощью окраски трихромом по Массону (ФИГ.6, Панель А). Более того, был выполнен иммунохимический гистологический анализ с целью оценить тип клеток, которые реально мигрировали в матригелевых пробках. Снимки конфокальным микроскопом показали, что эндотелиальные клетки, экспрессирующие CD31, или макрофаги, экспрессирующие CD68, росли и мигрировали в значительной степени в матригелевых пробках в контрольной группе пустого вектора, но это происходило в значительно меньшей степени в лечебной группе (ФИГ.6, Панель С).
Трансгенные опухолевые клетки, экспрессирующие легумаин, помечаются спленоцитами мышей, иммунизированных композицией ДНК по изобретению. В изучаемой модели клеточная линия 4Т1 была стабильно трансфицирована ретровирусом, содержащим легумаиновую плазмиду, и затем использована в выделении клеток (ФИГ.7, панель А, левое фото имеет 5-кратное увеличение, правое фото имеет 35-кратное увеличение). Микрофотографические снимки были сделаны спустя 2 дня после трансфекции. Положительные клетки выделены стрелками (ФИГ.7, панель А). Был представлен анализ высвобождения 51Cr; данные показаны на ФИГ.7, Панель В. Спленоциты, выделенные у мышей, иммунизированных pLegumain композицией ДНК, были эффективны в уничтожении клеток 4Т1, трансфицированных легумаином (*Р<0,01 по сравнению с контрольными группами пустого вектора). Опухолеассоциированное опосредованное Т-клетками уничтожение было специфичным для легумаина, так как клетки 4Т1, недостаточно экспрессирующие легумаин, не лизировали.
Настоящее изобретение представляет новую парадигму (образец) в противоопухолевом лечении, т.е. снижение плотности ТАМ в опухолевой строме снижает высвобождение факторов, усиливающих рост опухоли и ангиогенез, что изменяет окружающую микросреду опухоли и значительно подавляет пролиферацию опухолевых клеток, васкуляризацию и метастазирование. Выделение ТАМ в строме опухоли могло усилить опасения, что нейтрализация этих клеток способна помешать нормальным иммунологическим функциям этих важных компонентов природной иммунной системы. Однако, не в данном случае.
Циркулирующие моноциты - это универсальные предшественники, способные дифференцироваться в различные формы специализированных макрофагов. Фактически, цитокинная окружающая среда оказывает сильное воздействие на дифференциацию и функцию тканевых макрофагов. Макрофаги, активируемые бактериальными продуктами и цитокинами Th1, рассматриваются как проявление фенотипа М1, т.е. классические активированные макрофаги с повышенной бактериологической активностью и цитотоксическим действием против опухолевых клеток. С другой стороны, макрофаги, активированные цитокинами Th2, такими как IL-4 и IL-13 или иммуносупрессорами, такими как витамин D3 и IL-10, классифицируются, как имеющие фенотип М2, характеризуемый низкой цитотоксической функцией, но высокой тканеизменяющей активностью.
Клетки М1 имеют иммуностимулирующие качества и защищают организм от патогенных инфекций, в то время как М2 клетки ослабляют острые воспалительные реакции, активно уничтожают продукты распада клеток и производят множество факторов, стимулирующих рост и ангиогенез, особенно для восстановления поврежденных тканей. Кроме того, макрофаги, происходящие из здоровой или воспаленной ткани, способны лизировать опухолевые клетки, экспрессировать иммуностимулирующие цитокины и презентировать опухолеассоциированные антигены для стимуляции пролиферации и противоопухолевых функций Т- и NK-клеток. М2 макрофаги, такие как ТАМ, демонстрируют сниженные уровни этих активностей. Это может быть результатом их воздействия на противовоспалительные молекулы, происходящие из опухоли, такие как IL-4, IL-10, TGF-бета1 и простагландин Е2. В самом деле, Mantovani с коллегами высказали мнение, что воздействие на IL-4 и IL-10 может побудить моноциты в опухоли развиться в поляризованный тип II или М2 макрофагов (Mantovani et al. 2002, Trends Immunol. 23:549-555).
В той степени, в какой они были до сих пор изучены, дифференцированные зрелые ТАМ имеют фенотип и функции, подобные макрофагам типа II (Mantovani et al. 2004, Eur. J. Cancer 40:1660-1667). Таким образом, цитокины, присутствующие в окружающей микросреде опухоли, имеют возможность вызывать и определять дифференциацию образующихся мононуклеарных фагоцитов. В самом деле, растущая совокупность данных показывает, что ТАМ склоняются в сторону М2 макрофагов в окружающей микросреде опухоли и продуцируют множество факторов, способствующих росту опухоли и ангиогенезу, равно как иммуносупрессивные молекулы. Так, наличие ТАМ в месте локализации опухоли и продолжение экспрессии и высвобождения их продуктов может скорее содействовать, чем противодействовать развитию опухоли и метастазированию.
ТАМ экспрессирует многочисленные линии CD206, маннозный рецептор, активированный на М2 макрофагах с последующим воздействием на IL-4 и IL-13 (Porcheray et al. 2005, Clin. Exp. Immunol. 142:481-489). Как здесь было показано, эта популяция макрофагов экспрессирует высокий уровень легумаина. Важно, что цитокины Th2 IL-4, IL-10 и IL-13 активируют экспрессию CD206 и легумаина на макрофаговой клеточной линии RAW. Это открытие может быть легче понято, если считать, что макрофаги из периферической крови дифференцируются в макрофаги М2, как только они поступают в место локализации опухоли, где IL-4, IL-13 и IL-10 высвобождаются опухолевыми клетками и клетками опухолевой стромы (Stein et al. 1992, J. Exp. Med. 176:287-292). Так, выделение М2 макрофагов, экспрессирующих легумаин, не только начинает подавлять опухолевый рост и метастазы, но также поддерживает нормальное функционирование макрофагов фенотипа М1.
На взаимосвязь между инфильтрацией ТАМ и прогнозированием в отношении опухолевых пациентов также указывалось в нескольких исследованиях, из которых следовало, что увеличение инфильтрации макрофагами ухудшало прогнозы (см. например, Wyckoff et al. 2004, Cancer Res. 64:7022-7029). Несколько групп признаков показывают, что в опухолевой строме существует симбиотическая взаимосвязь между раковыми клетками и ТАМ, при которой раковые клетки привлекают ТАМ и повышают их выживаемость, в то время как ТАМ отвечают опухолепроизводным молекулам, обеспечивая важные факторы роста и внеклеточные матричные ферменты. Это, в свою очередь, стимулирует опухолевую пролиферацию, ангиогенез и инвазию окружающих тканей (см. например, Wyckoff et al. 2004, Cancer Res. 64:7022-7029). Таким образом, аттенуирование ТАМ в опухолевой среде обеспечивает эффективную стратегию изменения опухолевой стромы и изменяет окружающую микросреду опухоли.
Композиции ДНК по настоящему изобретению вызывают устойчивый CTL ответ против легумаина, представляющего собой аспарагиновую эндопептидазу, которая функционирует как белок стрессов, и обильно сверхэкспрессируется ТАМ. Было показано, что противолегумаиновый иммунный ответ является ответом специфичных CD8+ Т-клеток, рестриктированных по МНС-антигенам первого класса. Важно, что настоящее изобретение показывает, что после иммунизации композицией ДНК по изобретению, основанной на легумаине, значительно снижается плотность дважды положительных CD206+ и F4/80+ макрофагов, например ТАМ. Дополнительно, разнообразие факторов, таких как VEGF, ММР-9 и TGF-бета, которые высвобождаются ТАМ, как было показано, презентируются на низких уровнях как в супернатанте культивируемых опухолевых клеток, так и в мышиной сыворотке. Хорошо известно, что VEGF и металлопротеиназа ММР-9 играют важные роли в формировании сосудов опухолей и стимуляции опухолевого ангиогенеза. В этом отношении ТАМ важны, поскольку они продуцируют как VEGF, так и ММР-9. Все более усиливая ангиогенез в сочетании с активизированной экспрессией VEGF и внеклеточных протеаз, таких как ММР-9, тогда как TGF-бета известен как важный фактор роста, участвующий в миграции опухолевых клеток по кровеносным сосудам. Фактически, TGF-бета может давать пролиферативные и антиапоптозные сигналы опухолевым клеткам, так же как и активирующий активатор плазминогена урокиназного типа (uPa), который может содействовать разрушению внеклеточного матрикса, что необходимо для возникновения прорастания сосудов.
Важно, что настоящее изобретение демонстрирует, что как только ТАМ нейтрализовываются в опухолевой ткани специфическими цитотоксическими Т-лимфоцитами CD8+ (CTL), опухолевые клетки изменяют свой характер, становясь менее злокачественными и менее инвазивными. Образование новых сосудов в опухолевых тканях также значительно уменьшается при лечении композицией ДНК по настоящему изобретению. Дополнительно, по имеющимся данным, ТАМ участвуют в иммуносуппрессии и толерантности (выносливости) в опухолевой микросреде и могут ингибировать Т-клеточные ответы, индуцируя апоптоз активированных Т-клеток с помощью активации высвобождения NO, PGs, TNF-альфа и активности аргиназы (см. например, Saio et al. 2001, J. Immunol. 167:5583-5593). После нейтрализации ТАМ активность специфических Т-клеток CD8+ заметно усиливалась, еще более показывая, что подход против ТАМ по настоящему изобретению обеспечивает эффективную стратегию в резком изменении иммунологической толерантности против опухолей.
В мышиной модели спонтанного метастазирования карциномы молочных желез 4Т1 была достигнута поразительно большая продолжительность жизни, в которой 75% (6 из 8) мышей прожили 3 месяца после прививки опухолевых клеток 4Т1 в молочную железу, когда производилась хирургическая резекция первичной опухоли с последующим лечением композицией ДНК по настоящему изобретению, основанной на легумаине. Что более неожиданно, у 62% (5 из 8) мышей совсем не выявили легочных метастазов. Подобные результаты были получены в профилактических моделях в двух других опухолевых моделях, т.е. немелкоклеточной карциноме легкого D121 и карциноме толстой кишки СТ-26. Эти дополнительные подтверждающие данные показывают, что выделение ТАМ для изменения окружающей микросреды опухоли является потенциально универсальной противоопухолевой стратегией, подавляющей инвазию опухолевых клеток и метастазы путем понижения концентрации факторов высвобождающихся ТАМ, усиливающих опухолевый рост и метастазы.
В примере 2 для создания минигенной композиции ДНК по настоящему изобретению был использован экспрессирующий вектор млекопитающих с ER сигналом (pCMV/Myc/ER). ER сигнальный пептид направляет белок в отделение секреции. Вектор также включал С-концевой пептид, который удерживает белок в эндоплазматическом ретикулуме (ER). Пептиды, кодируемые композицией ДНК, преобразовывались в ER иммунных клетках кишечника и затем присоединялись к антигенному связывающему сайту MHC первого класса для окончательного презентирования к Т-клеточным рецепторам, которые индуцируют легумаинспецифический Т-клеточный ответ против мышиных ТАМ, экспрессирующих легумаин. Эти конструкции создают эффективные минигенные вакцины, которые могут вызывать эффективный CD8+ Т-клеточный ответ против опухолей различных органов в мышиных опухолевых моделях.
Дополнительные эксперименты с D121 клетками немелкоклеточной карциномы у изогенных C57/BL мышей показали, что обе минигенные конструкции pCMV-Db/Dd и pCMV-Kb/Kd обеспечивали эффективный иммунный ответ против ТАМ. С другой стороны, результаты с клетками D2F2 мышей BALB/c показали, что конструкция pCMV-Kb/Kd была значительно более эффективной, чем либо pCMV-Db/Dd, либо pCMV-Kb/Kd. Возможное объяснение этого открытия состоит в том, что существует намного большее количество легумаиновых пептидов, предназначенных присоединяться к H-2Dd/Kd с наивысшим сродством, чем таковых, присоединяющихся к H-2Dd/Kb молекулам. Согласно BioInformatics & Molecular Analysis Section (BIMAS) NIH, прогнозируемые величины пептидов легумаина, связывающихся с Н-2Dd и Н-2Kd, значительно больше, чем сообщаемые величины для Н-2Db и Н-2Kb, а связывающий показатель Н-2Kd наиболее высокий (ФИГ.20).
Полагая, что спектр рецепторов Т-клетки почти неограничен, то может быть гораздо большее число комплексов пептид-H-2Dd/Kd, которые могут быть распознаны CTL мышей BALB/с. Более того, пептид H-2Dd имел более высокий индекс антигенности, чем два или три пептида Н-2К (ФИГ.20). Основанная на этих сведениях, антигенность менее предсказуема, чем связывающий показатель МНС с отношением к иммунному ответу против ТАМ. Основанные на легумаине минигенные композиции по настоящему изобретению показали эффективную защиту против опухолей путем разрушения ТАМ в моделях опухоли молочных желез. Противоопухолевая защита была вызвана основанными на Kd минигенными вакцинами, приводящими к воздействию на ТАМ в окружающей микросреде карциномы молочной железы D2F2 у сингенных мышей BALB/с. Этот защитный иммунный ответ, как было обнаружено, опосредован Т-клетками CD8+, которые прицельно уничтожают legumain+ ТАМ, а также ведет к выраженному подавлению опухолевого ангиогенеза. Важно, что минигенные конструкции ДНК по изобретению обеспечивают эффективность, подобную вакцине, кодирующей целый ген легумаина. Точно так же как Dd, так и Kd-основанные конструкции обеспечивают ответы против ТАМ в модели рака D121 у мышей C57/BL. Вместе взятые, эти данные представляют первую противолегумаиновую минигенную композицию, эффективную против ТАМ, эта стратегия может быть особенно полезна для лиц с различными генетическими предпосылками и тем самым обеспечивает простую, более надежную и гибкую альтернативу стратегии цельногенных вакцин в лечении рака молочной железы.
Многочисленные изменения и модификации описанных выше осуществлений изобретения можно произвести, не отступая от сущности и объема новых признаков изобретения. Никоим образом не предполагается и не должно подразумеваться ограничение изобретения приведенными здесь иллюстративными вариантами его осуществления.
| название | год | авторы | номер документа |
|---|---|---|---|
| ДНК-ВАКЦИНЫ ПРОТИВ ОПУХОЛЕВОГО РОСТА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2004 |
|
RU2343195C2 |
| ПРОФИЛАКТИЧЕСКАЯ ПРОТИВОРАКОВАЯ ВАКЦИНА | 2004 |
|
RU2556128C2 |
| ГУМАНИЗИРОВАННОЕ МОНОКЛОНАЛЬНОЕ АНТИТЕЛО, СПЕЦИФИЧНОЕ К ЛЕГУМАИНУ | 2013 |
|
RU2574203C2 |
| ПРОФИЛАКТИЧЕСКАЯ ПРОТИВОРАКОВАЯ ВАКЦИНА | 2004 |
|
RU2385163C2 |
| ИДЕНТИФИКАЦИЯ ОПУХОЛЕАССОЦИИРОВАННЫХ АНТИГЕНОВ ДЛЯ ДИАГНОСТИКИ И ТЕРАПИИ | 2013 |
|
RU2644686C2 |
| СЛИТЫЕ БЕЛКИ С FC-ФРАГМЕНТОМ ИММУНОГЛОБУЛИНА ДЛЯ ПОВЫШЕНИЯ ИММУНОГЕННОСТИ БЕЛКОВЫХ И ПЕПТИДНЫХ АНТИГЕНОВ | 2000 |
|
RU2248214C2 |
| ТЕРАПИЯ ДЛЯ ЛЕЧЕНИЯ РАКА С ПОМОЩЬЮ ВНУТРИОПУХОЛЕВОГО И/ИЛИ ВНУТРИВЕННОГО ВВЕДЕНИЯ РЕКОМБИНАНТНОГО MVA, КОДИРУЮЩЕГО 4-1BBL (CD137L) И/ИЛИ CD40L | 2019 |
|
RU2830601C2 |
| ПРОСТАТОАССОЦИИРОВАННЫЕ АНТИГЕНЫ И ИММУНОТЕРАПЕВТИЧЕСКИЕ СХЕМЫ НА ОСНОВЕ ВАКЦИН | 2013 |
|
RU2737765C2 |
| МОДИФИЦИРОВАННЫЕ ЭПИТОПЫ ДЛЯ УСИЛЕНИЯ ОТВЕТОВ CD4+ Т-КЛЕТОК | 2013 |
|
RU2724994C2 |
| КОНЪЮГАТЫ ДЛЯ ВВЕДЕНИЯ БИОЛОГИЧЕСКИ АКТИВНЫХ СОЕДИНЕНИЙ | 2009 |
|
RU2567667C2 |









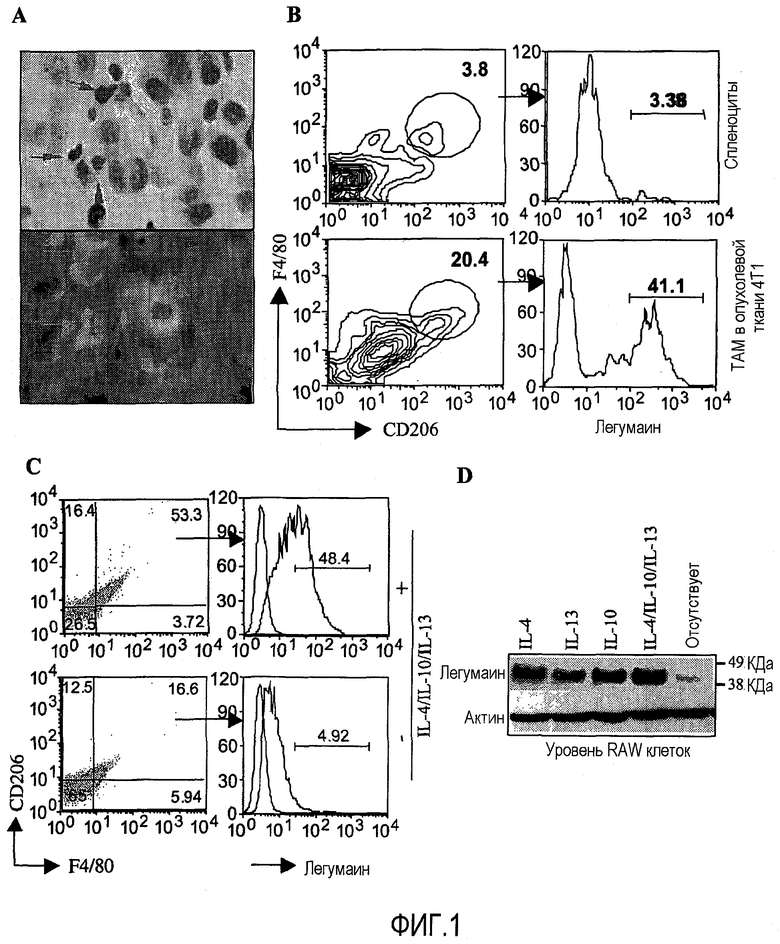
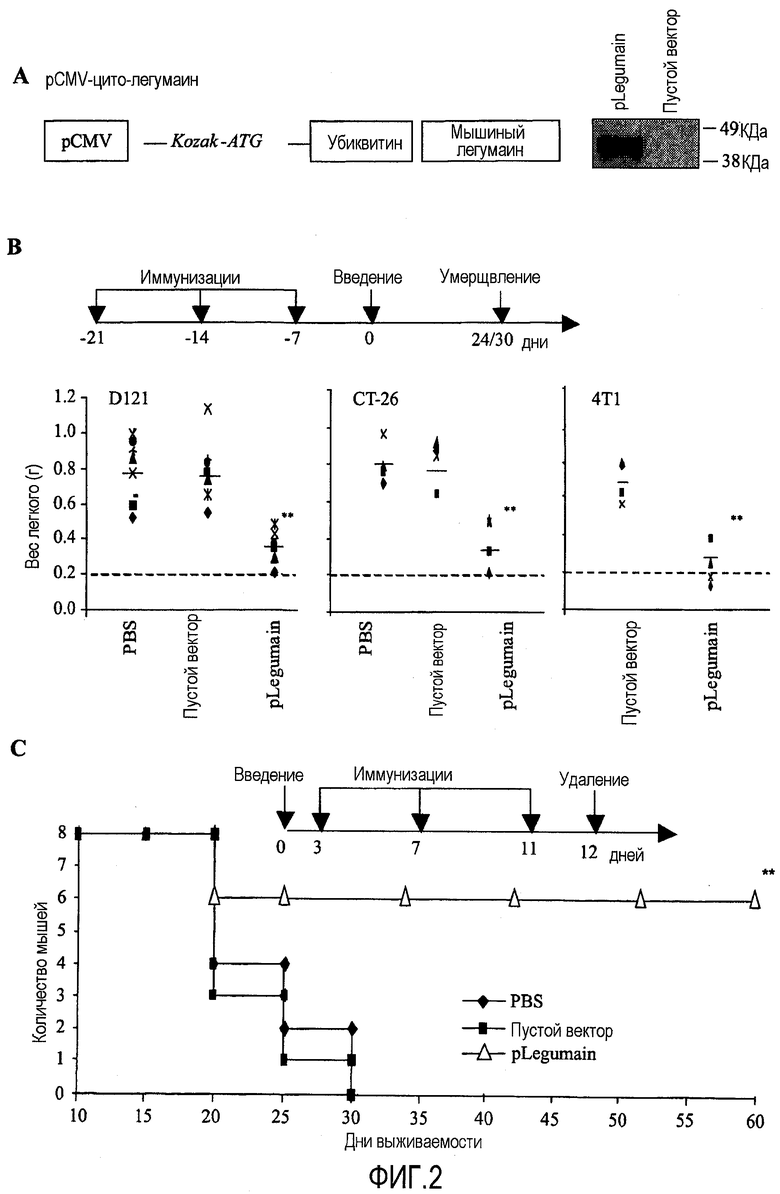
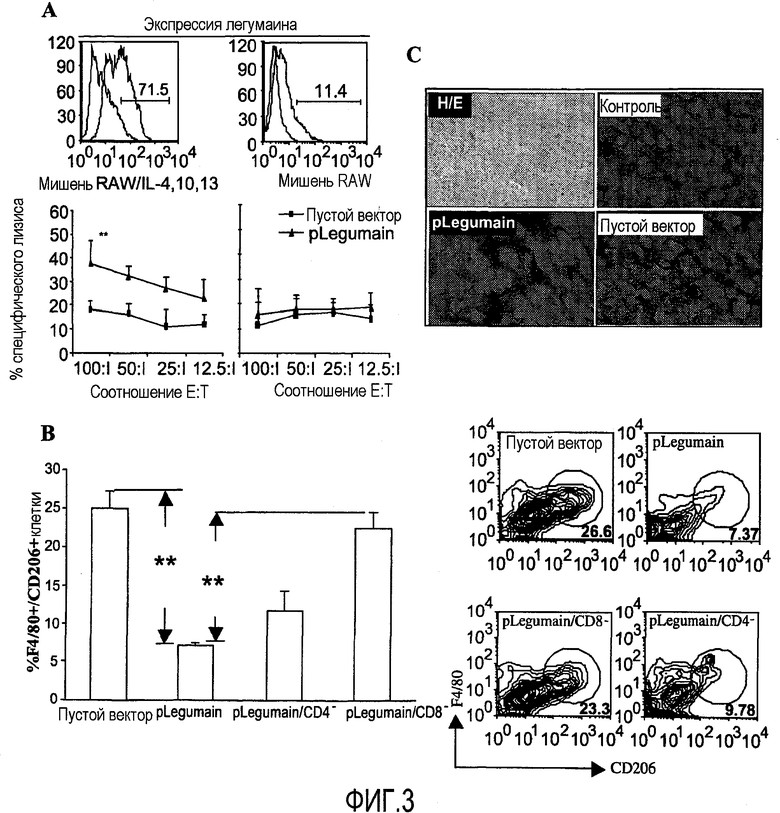
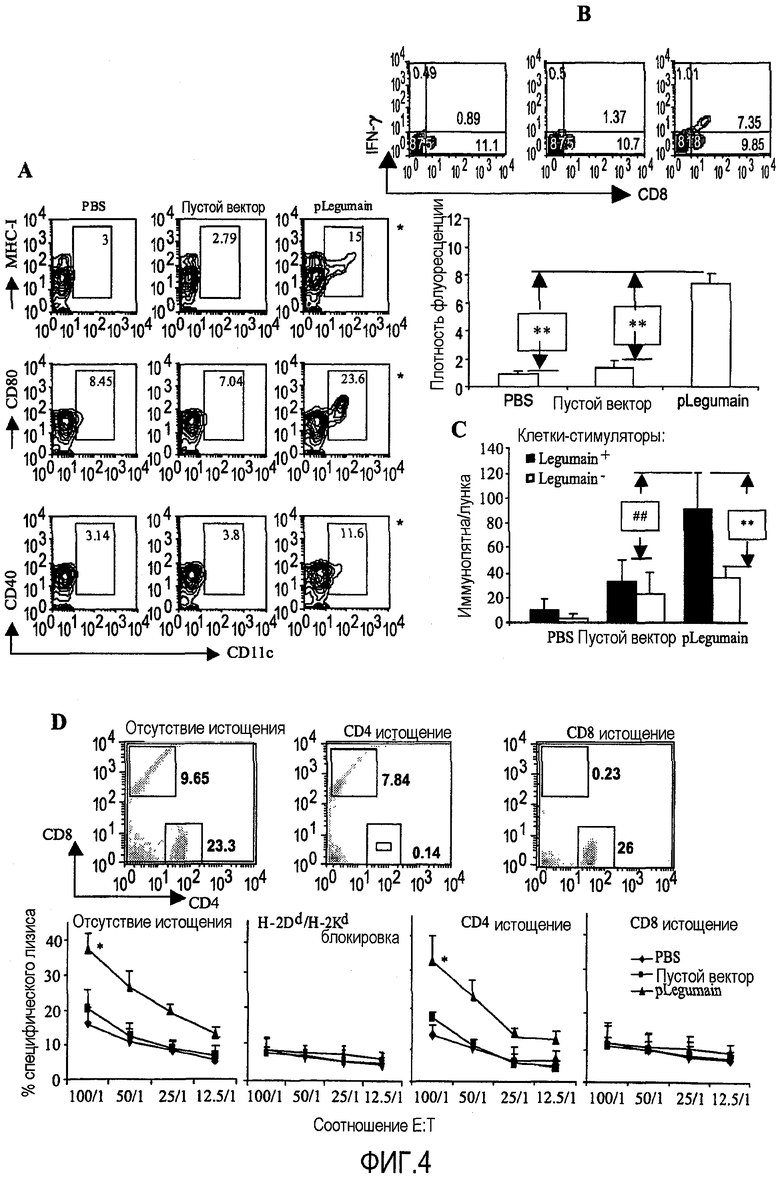
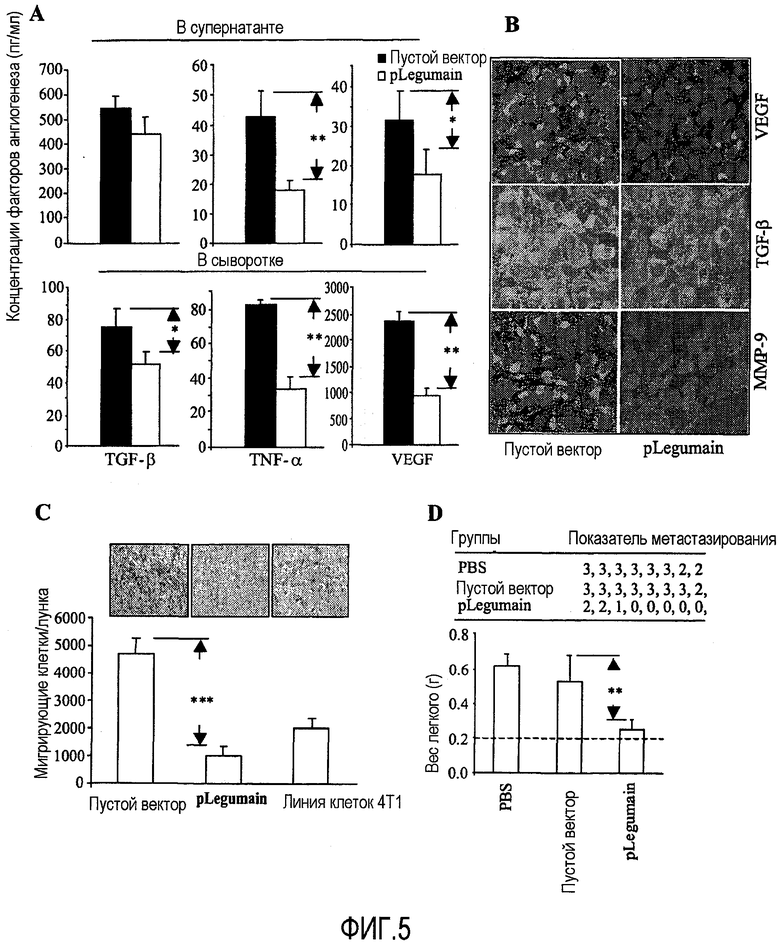
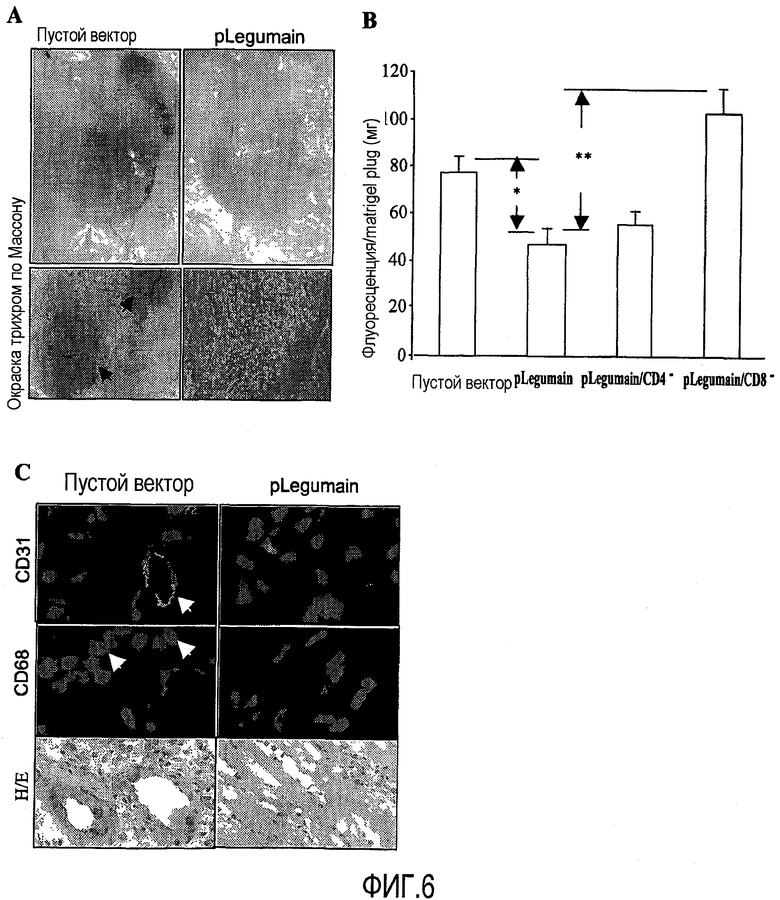
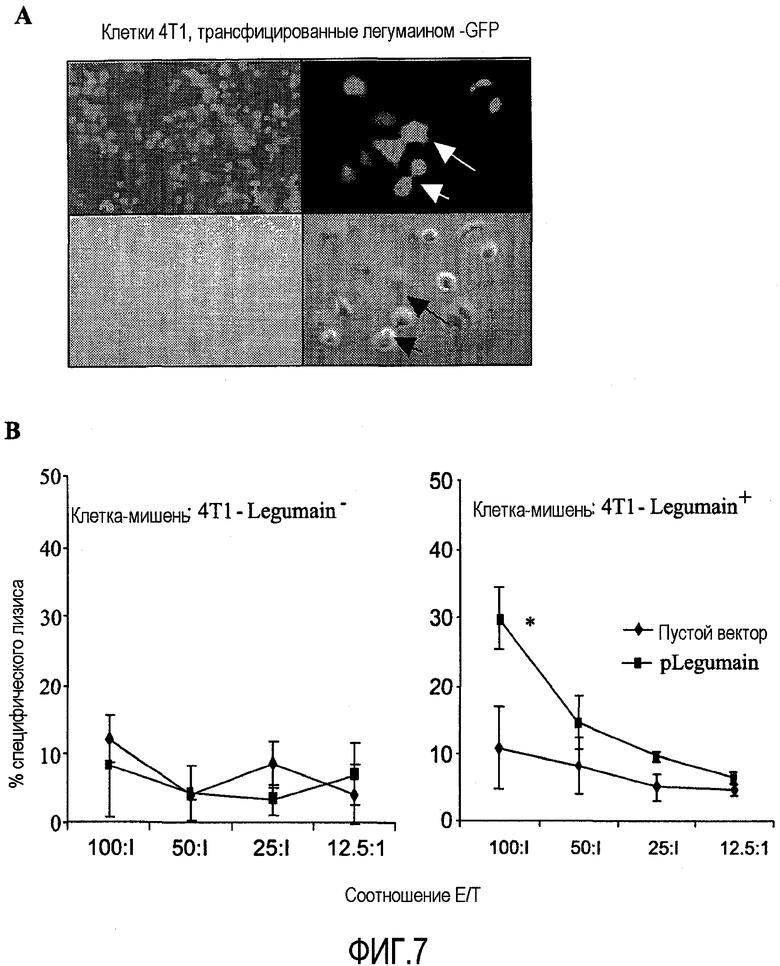






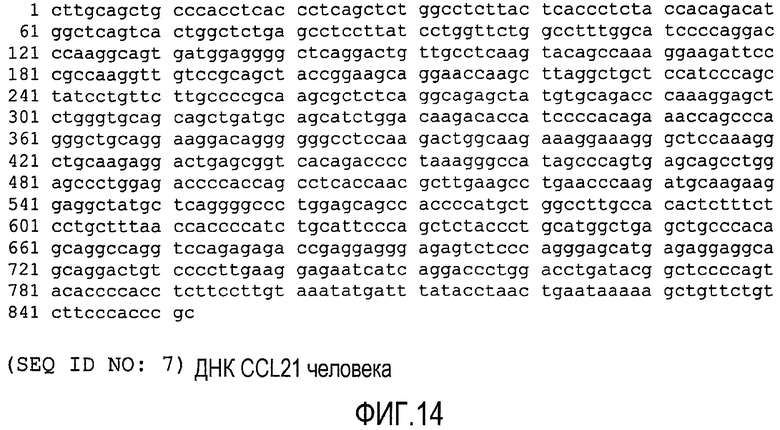

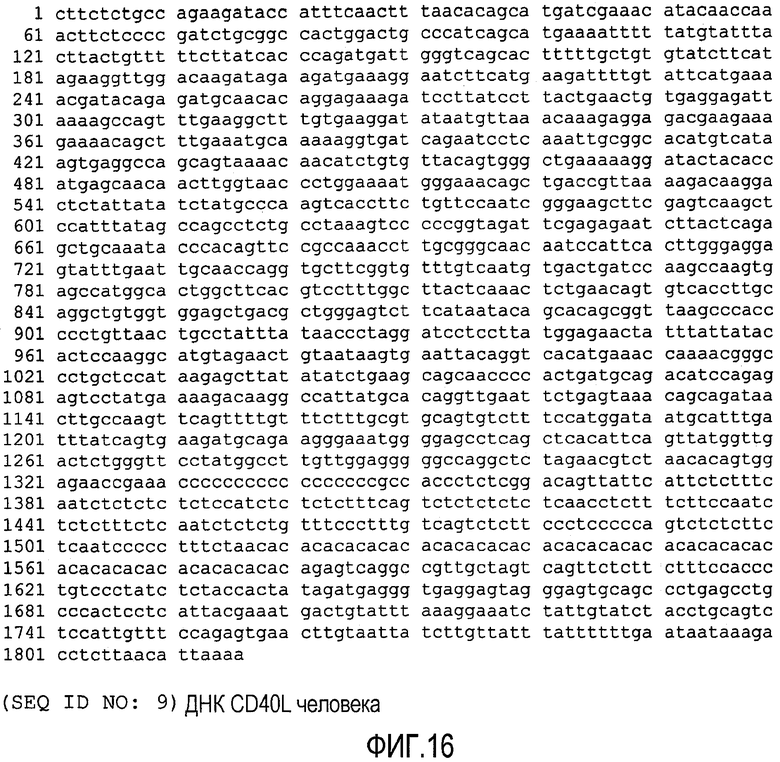

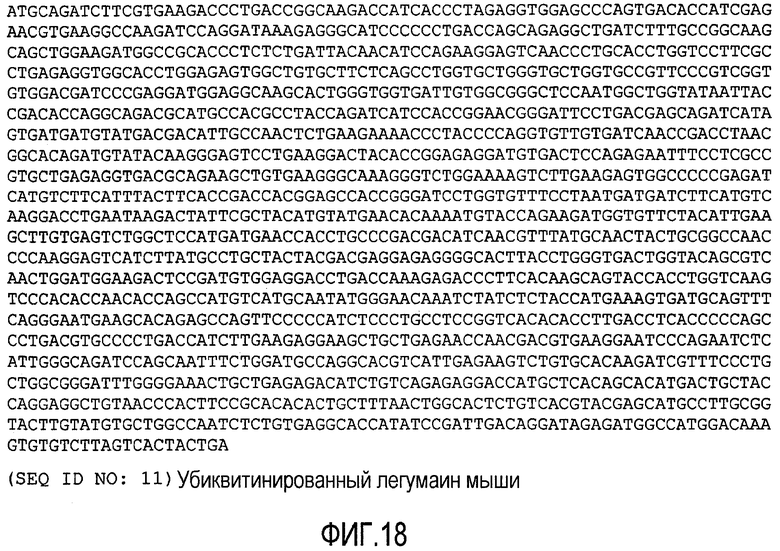


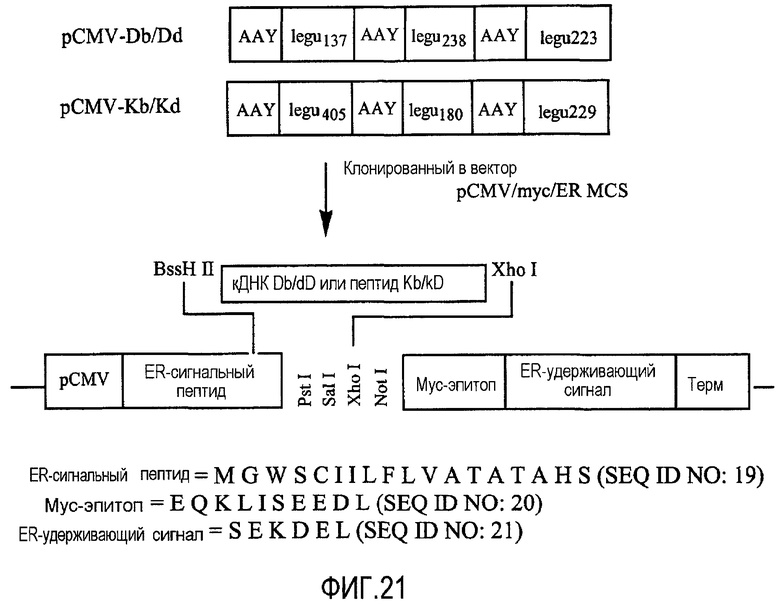
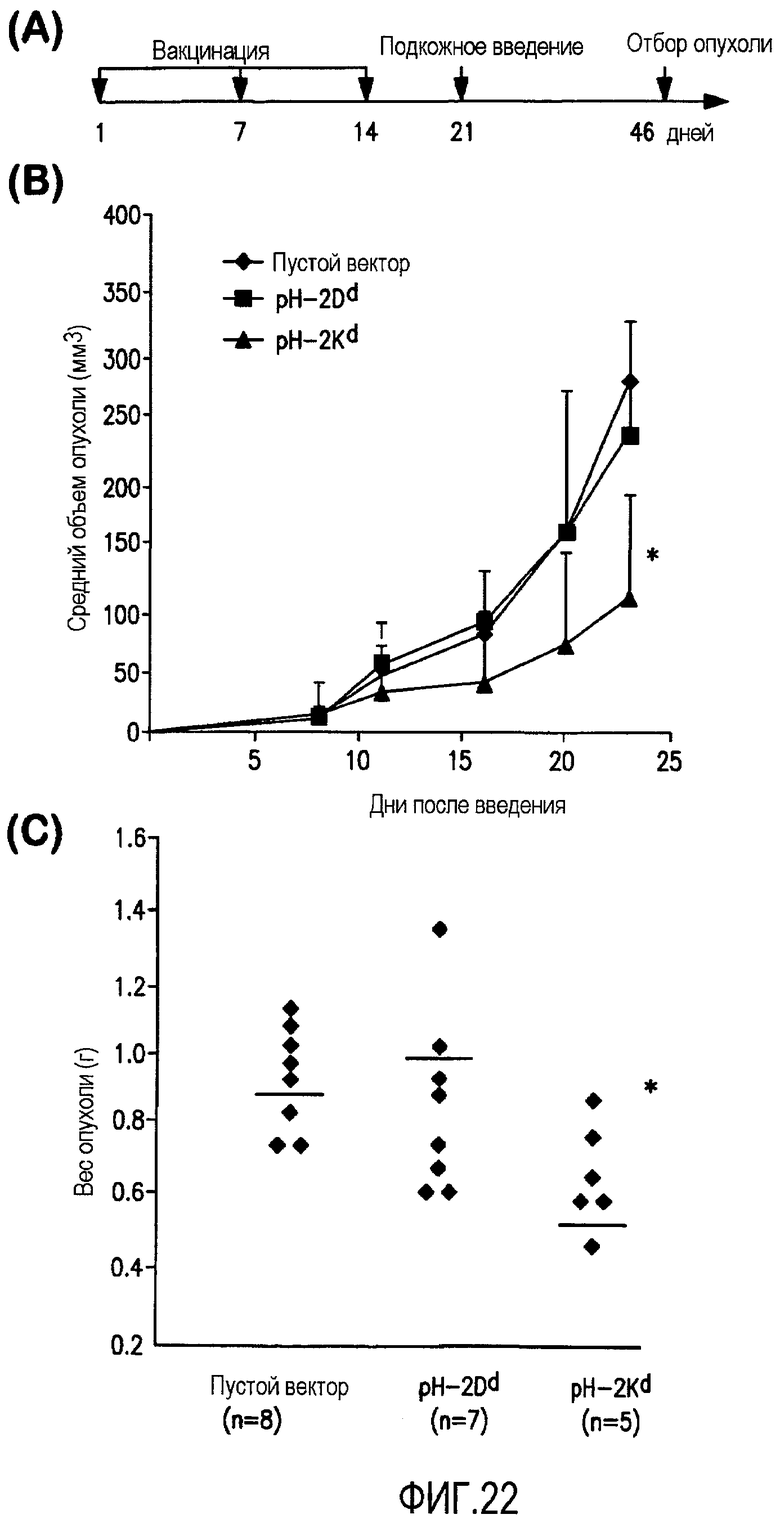
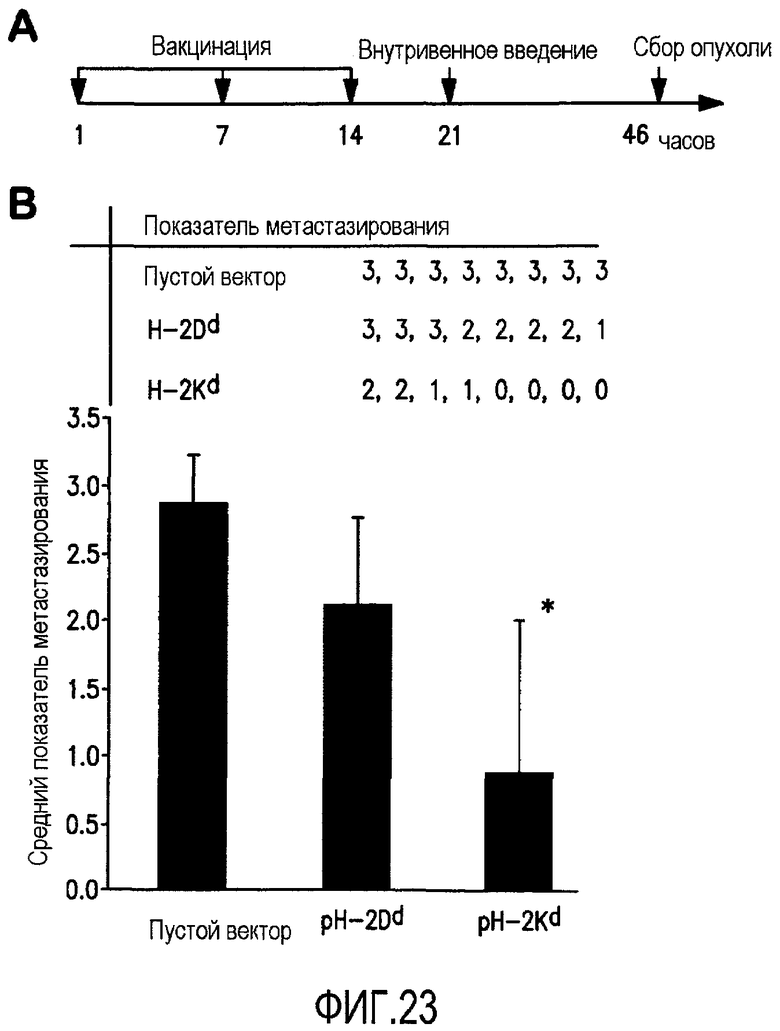
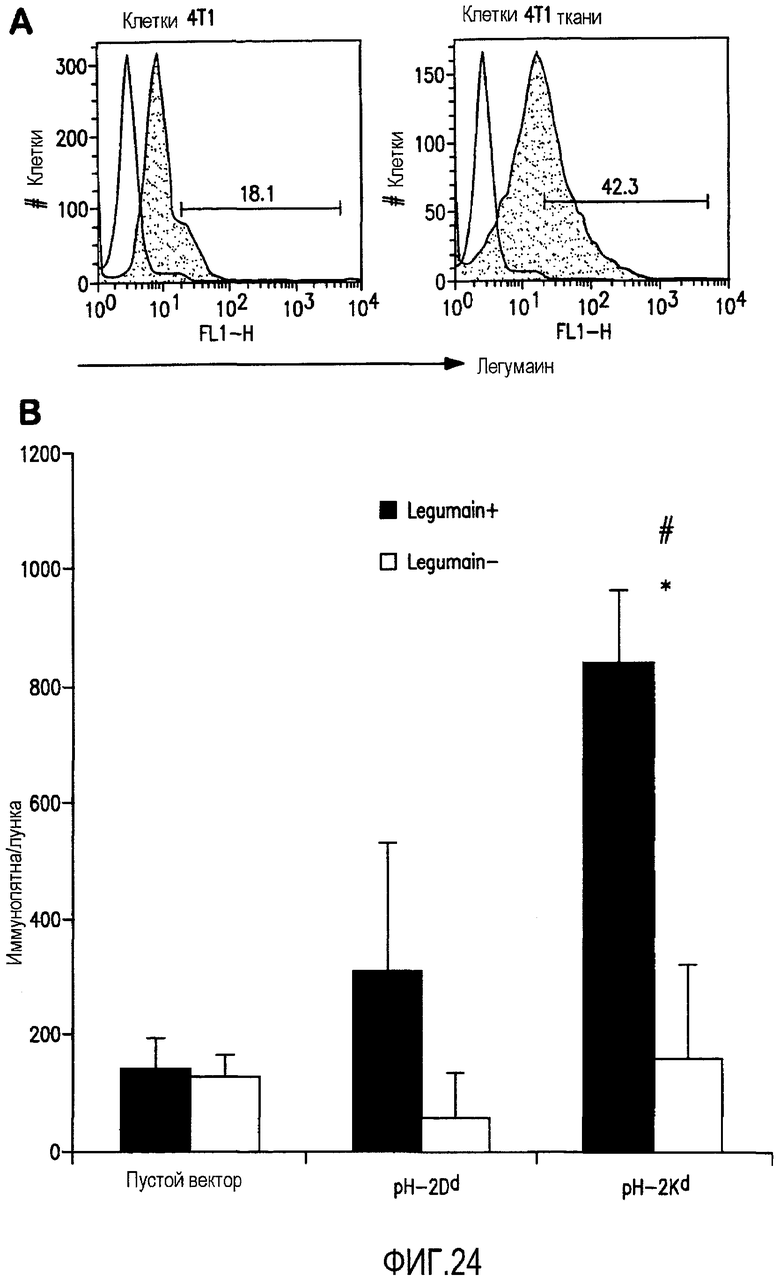
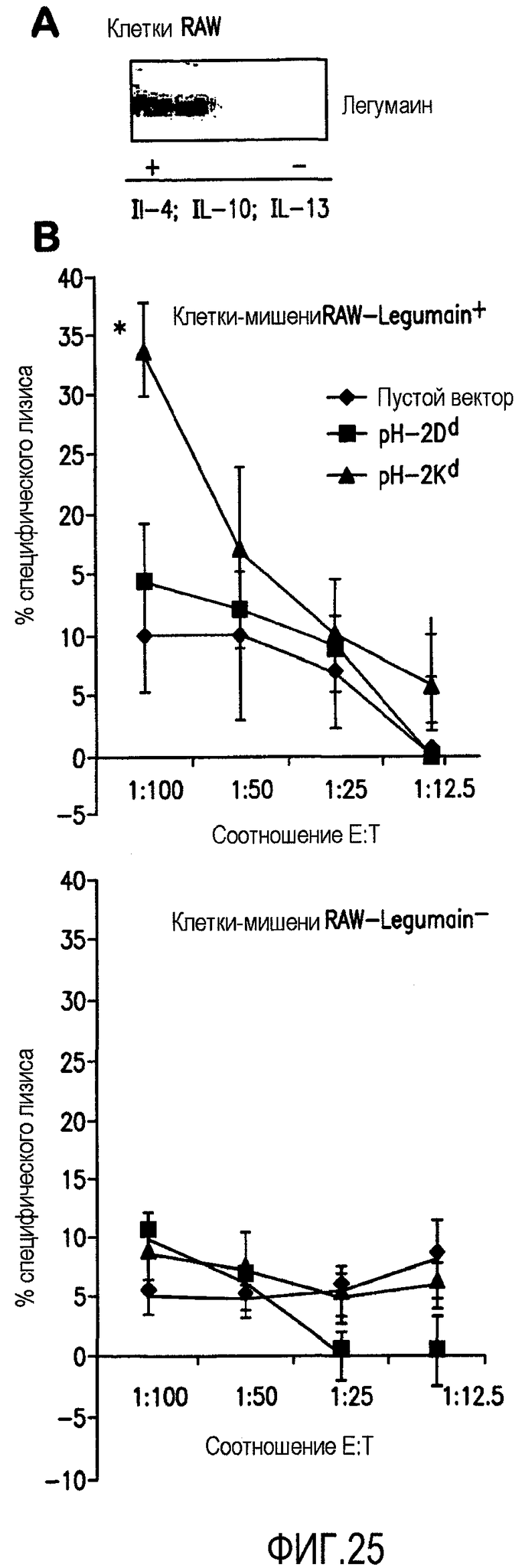
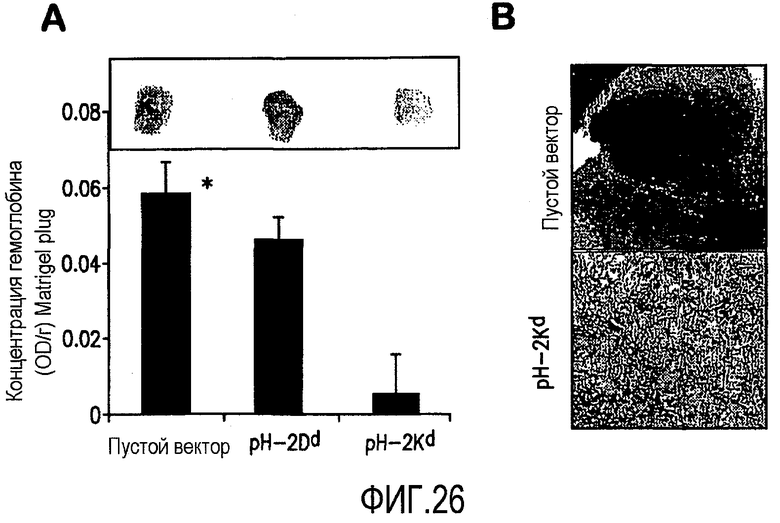
Изобретение относится к медицине и касается фармацевтической композиции для вызова иммунного ответа против опухолеассоциированных макрофагов, содержащей минигенную конструкцию ДНК, которая кодирует полипептид, в фармацевтически приемлемом носителе, где полипептид содержит три иммуногенных фрагмента легумаина, связанных вместе линкерными пептидами, и где каждый линкерный пептид состоит из последовательности ААА или AAY. Изобретение обеспечивает эффективность в борьбе с карциномой молочной железы, немелкоклеточным раком легкого и карциномой толстой кишки. 7 з.п. ф-лы, 26 ил., 2 пр.
1. Фармацевтическая композиция для вызова иммунного ответа против опухолеассоциированных макрофагов, содержащая минигенную конструкцию ДНК, которая кодирует полипептид, в фармацевтически приемлемом носителе, где полипептид содержит три иммуногенных фрагмента легумаина, связанных вместе линкерными пептидами, и где каждый линкерный пептид состоит из последовательности ААА или AAY, и каждый иммуногенный фрагмент легумаина выбирают из группы, состоящей из SEQ ID NO: 13, SEQ ID NO: 14, SEQ ID NO: 15, SEQ ID NO: 16, SEQ ID NO: 17 и SEQ ID NO: 18.
2. Композиция по п.1, где каждый линкерный пептид имеет последовательность AAY.
3. Композиция по п.1, где минигенная конструкция включена в цитомегаловирусный (CMV) плазмидный экспрессионный вектор, который кодирует последовательность ER-сигнального пептида, последовательность Мус-эпитопа и последовательность ER-удерживающего сигнала, где ER-сигнальный пептид состоит из SEQ ID NO: 19, Мус-эпитоп состоит из SEQ ID NO: 20, ER-удерживающий сигнал состоит из SEQ ID NO: 21.
4. Композиция по п.3, где минигенная конструкция включена в вектор между сайтом BssH II в последовательности ER-сигнального пептида и сайтом Xho I.
5. Композиция по п.1, которая дополнительно содержит конструкцию ДНК, кодирующую иммунный эффекторный белок, экспрессируемый в иммунных клетках.
6. Композиция ДНК по п.5, где иммунный эффекторный белок является цитокином.
7. Композиция ДНК по п.6, где цитокином является CCL21, IL-2 или CD40LT.
8. Композиция по п.1, где минигенная конструкция встроена в аттенуированный бактериальный вектор AroA-, dam- Salmonella typhimurium.
| D.P.Dickinson "Cysteine peptidases of mammals: their biological roles and potential effects in the oral cavity and other tissues in health and disease" Crit Rev Oral Biol Med | |||
| Топчак-трактор для канатной вспашки | 1923 |
|
SU2002A1 |
| Lode H.N | |||
| et al., "DNA minigene vaccination for adjuvant neuroblastoma therapy" Ann N Y Acad Sci | |||
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| Lin EY et al., "Macrophages: | |||

