ОБЛАСТЬ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к композициям и методам, используемым для получения противоопухолевой вакцины.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Позвоночные обладают способностью вырабатывать иммунный ответ в качестве защиты от окружающей среды, а также от аберрантных клеток, таких как опухолевые клетки, развивающиеся в организме. Иммунный ответ является результатом сложных взаимодействий различных клеток и факторов, но обычно включает два основных аспекта. Один из них является клеточным иммунным ответом, характеризующимся тем, что специализированные клетки непосредственно атакуют агент-нарушитель (несущий антиген), тогда как второй является гуморальным ответом, при котором молекулы антитела специфически связываются с антигеном и способствуют его элиминации. Действуя совместно, отдельные элементы очень эффективно ограничивают первичную атаку вторгающихся патогенов и выделяют их из организма-хозяина.
Первичными клетками, участвующими в выработке иммунного ответа, являются лимфоциты, которые в целом делятся на два основных класса. Клетки первого из них, называемые В клетками или В лимфоцитами, обычно образуются в костном мозге и, среди других выполняемых функций, ответственны за продуцирование и секретирование антител. Продукты В-клеточных антител имеют тенденцию реагировать непосредственно с чужеродными антигенами и нейтрализовать их или активировать другие компоненты иммунной системы, которая затем их элиминирует. В частности, опсонизирующие антитела связываются с внеклеточными чужеродными агентами, тем самым делая их чувствительными к фагоцитозу и последующему внутриклеточному киллингу. С другой стороны, Т клетки или Т лимфоциты, которые обычно образуются или созревают в тимусе, ответственны за опосредование клеточного иммунного ответа. Эти клетки не распознают весь антиген, но, вместо этого, реагируют на его короткие пептидные фрагменты, которые обнаруживаются на поверхности клетки-мишени, а также антигенпрезентирующей клетки. Более конкретно, очевидно, что белки, продуцированные в клетке или захваченные клеткой из внеклеточного пространства, при нормальном метаболизме непрерывно расщепляются до пептидов. Образующиеся в результате короткие фрагменты ассоциируются с молекулами внеклеточного главного комплекса гистосовместимости (МНС) и комплексы МНС-пептид переносятся на поверхность клетки и распознаются Т-клетками. Таким образом, клеточная иммунная система непрерывно контролирует полный спектр белков, продуцируемых или усваиваемых клетками, и вынуждает элиминировать любые клетки, презентирующие чужеродные антигены или опухолевые антигены, т.е. клетки, инфицированные вирусами, или раковые клетки.
Структура иммуноглобулина G (IgG) представляет собой сложный тетрамерный белок, содержащий две идентичных тяжелых (Н) цепи и две идентичных легких (L) цепи иммуноглобулина. Эти цепи соединяются вместе дисульфидными связями с образованием Y-образного антитела. Однако, в растворе молекула принимает очертания глобул и легко связывается с чужеродными антигенами, присутствующими в биологических жидкостях. Анализ аминокислотных последовательностей иммуноглобулинов привел к определению специфических областей с различными функциональными активностями внутри цепей. Каждая легкая и каждая тяжелая цепь содержит вариабельную область (VL и VH, соответственно), содержащую первые 110 аминокислотных концевых остатков. Трехмерный участок соединения областей VL и VH составляет участок распознавания "антигенсвязывающего сайта" ("ACS") молекулы иммуноглобулина. Вследствие тетрамерной природы иммуноглобулинов в молекуле имеется два идентичных антигенсвязывающих сайта. Вариабельные домены этих цепей содержат высокогетерогенные последовательности и сообщают разнообразие антигенсвязывающим сайтам, в высшей степени специфическим в отношении большого ряда антигенных структур. Гетерогенность вариабельных доменов неравномерно распределена по вариабельным областям, но расположена в трех сегментах, называемых гипервариабельными областями ("CDR"), обозначенными CDR1, CDR2 и CDR3. Подробнее об этих структурах см. монографию Watson et al., 1987, Molecular Biology of the Gene, Forth Edition, Benjamin/Cummings Publishing Co., Inc. Menio Park, Calif.
Каждая из тяжелых цепей включает также константную область, определяющую конкретный изотип и позволяющую отнести иммуноглобулин к одному из классов и подклассов иммуноглобулинов. Константная область содержит сегменты, называемые доменами (т.е. CH1, CH2 и т.д.), которые незначительно отличаются в антителах одного класса. Константная область не участвует в антигенном связывании, но может ассоциироваться с рядом биологических активностей, известных как "эффекторные функции", такими как связывание с Fc рецепторами на клеточной поверхности, а также связывание с белками комплемента. Антигенпрезентирующие клетки, такие как дендритные клетки и макрофаги, помимо других особенностей, отличаются обычно присутствием Fc рецептора. Соответственно, если антитело связано с патогеном, оно может затем связываться с фагоцитом через Fc участок. Это позволяет фагоцитам заглатывать и разрушать патоген, патогенные антигены могут процессироваться и выявляться с использованием АРС для дальнейшей стимуляции иммунного ответа.
В отличие от тяжелых цепей легкие цепи содержат единственную константную область (CL). Легкая цепь соединяется с тяжелой цепью дисульфидной связью, которая связывает константную область тяжелой цепи СН с CL. Кроме того, тяжелые цепи также содержат шарнирную область, отделяющую константные области CH1 и СН2 от остальной части молекулы. Эта та шарнирная область, которая в значительной степени отвечает за гибкость тетрамера. Две тяжелых цепи молекулы связываются вместе дисульфидными связями в точке между шарнирной областью и CH2.
Чтобы создать такой обширный спектр, получают гены иммуноглобулина таким образом, чтобы можно было образовывать большое число различных белков иммуноглобулина из ограниченного количества генов, т.е. внутренний полиморфизм. Благодаря внутреннему полиморфизму млекопитающие способны продуцировать антитела к, предположительно, бесконечному множеству антигенов. Обзор по генетике иммуноглобулинов и структуре белков см. Lewin, "Genes III", John Wiley & Sons, N.Y. (1987) и Benjamin and Leskovitz, 1988, Immunology, Alan R. Liss. Inc., New York.
За последние несколько лет антитела приобрели чрезвычайно важное значение в диагностике и терапии благодаря их многообразию и специфичности. Методы молекулярной биологии все больше используются для расширения разнообразия и доступности антител для научных исследований. Например, единичная продуцирующая антитело В клетка может быть иммортализована путем слияния с опухолевой клеткой и размножена с целью получения in vitro источника единичной специфичности, известной как "моноклональное антитело" (mAb). Такая бессмертная линия В клеток называется "гибридомой".
До последнего времени источником большинства mAb были мышиные гибридомы, культивируемые in vitro. To есть, мыши обычно инъецировали выбранный антиген или иммуноген. Затем животное умерщвляли и выделенные клетки селезенки сливали с бессмертными клетками миеломы. Мышиные клетки, хотя их широко использовали для диагностики, не очень подходят для применения в терапии большинства млекопитающих, включая человека. Частично вследствие того, что мышиные антитела другими млекопитающими распознаются как чужеродные и вызывают иммунный ответ, который сам по себе может вызвать заболевание. Для того, чтобы преодолеть, по меньшей мере, некоторые из проблем, связанных с иммунным ответом, вызываемым чужеродным mAb, и недостаток подходящего человеческого mAb, для создания (конструкции) химерных молекул гуманизированного иммуноглобулина, которые содержат антигенсвязывающие гипервариабельные области мышиных антител, при этом остальная часть молекулы состоит из последовательностей человеческого антитела, которые не распознаются как чужеродные, используют методы рекомбинантной ДНК. Такие антитела используются для лечения опухолей, так как мышиная вариабельная область распознает опухолевый антиген, и гуманизированный участок молекулы способен опосредовать иммунный ответ и при этом не выделяться быстро из организма. См., например, статью Jones et al., Nature, 321: 522-525 (1986).
Другие применения таких антител подробно описаны в опубликованной международной заявке WO 94/14847. В этих случаях эпитопы чужеродных антигенов, такие как вирусные или бактериальные эпитопы, "пересаживают" в гипервариабельную область иммуноглобулина с целью вызвать ответ. То есть созданные антитела используют в качестве вакцины для того, чтобы вызвать иммунный ответ и наделить долговременной иммуногенной памятью, тем самым способствовать победе субъекта над будущими инфекциями.
Эти и более традиционные вакцины эффективны потому, что они стимулируют обе составляющих иммунной системы. Несмотря на трудности, связанные с гуморальным компонентом иммунного ответа, он не может, по своей природе и без связи с другими явлениями, эффективно защитить животное от несметного числа патогенных атак, которым оно подвергается каждый день. Скорее, только наличие сильного клеточного ответа позволяет высшим организмам выживать и размножаться.
Как указывается выше, Т лимфоциты, или Т клетки, которые образуются из предшественников в костном мозге, являются главными участниками при создании иммунного ответа против поражающих вирусов и других микробов. Предшественники-стволовые клетки мигрируют в тимус (вилочковую железу), где они становятся специализированными в качестве так называемых тимоцитов. В частности, они начинают визуализировать рецепторные молекулы, которые позже позволяют зрелым Т клетками обнаруживать инфекцию. Для того чтобы быть полезными, Т клетки должны быть способны с помощью рецепторов связываться с антигенами (белковые маркеры передают сигнал о присутствии возбудителя ("интервента")). В то же время они не должны реагировать на вещества, образующиеся в организме, так как аутореактивные Т клетки могут разрушать нормальные ткани. Обычно только те тимоциты, которые образуют полезные рецепторы, созревают полностью и попадают в кровоток, стоят на страже здоровья организма. Другие тимоциты, неэффективные или те, которые атакуют собственные ткани организма, у здоровых людей элиминируются по механизму апоптоза перед тем, как оставить тимус.
Зрелые Т клетки, которые в конечном счете попадают в кровоток, либо в виде цитолитических Т лимфоцитов, либо Т-хелперных клеток, покоящихся, если они не встречают антигены, которые могут распознать их рецепторы. При столкновении со специфическим антигеном, в отношении которого лимфоциты проявляют аффинность, они пролиферируют и осуществляют эффекторные функции, результатом чего является элиминация чужеродных антигенов.
Т клетки разделяются на несколько субпопуляций с учетом различных задач, которые они выполняют. Эти субпопуляции включают хелперные Т клетки (Th), которые необходимы для стимулирования или повышения Т- и В-клеточного ответа; цитотоксические (или цитолитические) Т лимфоциты (CTL), которые непосредственно убивают их клетки-мишени с помощью лизиса клеток, и Т-супрессоры, или регуляторные Т клетки (Ts или Tr), которые подавляют иммунный ответ. В каждом случае Т клетки распознают антигены, но только когда присутствуют на поверхности клетки в виде специализированного белкового комплекса, связанного с поверхностью антигенпрезентирующей клетки. Более конкретно, Т клетки используют специфический рецептор, так называемый специфический к антигену Т-клеточный рецептор (TCR), который является трансмембранным белком, способным распознавать антиген в ассоциации с группой белков, в целом называемых главным комплексом гистосовместимости (МНС, ГКГ). Тысячи идентичных TCR экспрессируются на каждой клетке. TCR относится, как по функции, так и по структуре, к поверхностному антителу (несекретированным), которое В клетки используют в качестве рецепторов антигена. Кроме того, различные субпопуляции Т клеток также экспрессируют ряд белков клеточной поверхности, некоторые из которых называются "маркерными белками", так как они характерны для конкретных субпопуляций. Например, большинство Th клеток экспрессируют белок клеточной поверхности CD4, тогда как большинство клеток CTL экспрессируют белок клеточной поверхности CD 8, а клетки Tr экспрессируют молекулы CD25 и CD4. Эти поверхностные белки играют важную роль в инициации и сохранении иммунных ответов, которые зависят от распознавания конкретных белков или белковых комплексов на поверхности АРС или взаимодействия между этими белками или белковыми комплексами.
Недавно стало известно, что главный комплекс гистосовместимости, или MHC, на самом деле содержит ряд гликозилированных белков, имеющих определенные четвертичные структуры. Вообще структуры относятся к двум типам: класса I MHC, который визуализует пептиды белков, полученных внутри клетки (такие как аутобелки или белки, продуцируемые после вирусной репликации), и класса II MHC, который в целом визуализует пептиды белков, попавших в клетку извне, со стороны (растворимые антигены, такие как бактериальные токсины). Распознавание различных антигенов обеспечивается внутренним полиморфизмом, который непрерывно создает многообразный пул MHC молекул, способных связывать любые могущие образовываться патогенные пептиды. По существу, все ядросодержащие клетки продуцируют и экспрессируют белки класса I MHC, который может проявлять природные пептиды, пептиды, ассоциированные с опухолью, или пептиды, продуцированные вирусным "захватчиком". Напротив, некоторые другие ядросодержащие клетки, и среди них специализированные лимфоидные клетки, известные как антигенпрезентирующие (антигенпредставляющие) клетки, продуцируют и экспрессируют белки класса II MHC. Вне зависимости от типа клеток, оба класса MHC несут пептиды к клеточной поверхности и представляют их покоящимся Т лимфоцитам. Обычно Th клетки распознают комплексы класса II МНС-антиген, тогда как CTL имеют тенденцию распознавать комплексы класса I МНС-антиген, хотя также происходит перекрестная (кросс-) презентация антигенов.
Когда покоящаяся Т клетка, несущая подходящий TCR, встречает АЗС, визуализующую пептид на своей поверхности, TCR связывается с комплексом пептид-МНС. Более конкретно, сотни TCR связываются с многочисленными комплексами пептид-МНС. Когда контактирует достаточное количество TCR, кумулятивный эффект активирует Т клетки. Рецепторы на Т клетках, отвечающие за специфическое распознавание комплекса МНС-антиген и за ответ на него, состоят из комплекса нескольких интегральных белков плазматической мембраны. Как и ранее обсуждавшийся MHC комплекс, разнообразный пул TCR обеспечивается внутренним полиморфизмом, приводящим к соматической реаранжировке. Следует подчеркнуть, что хотя пул TCR может быть разно(много)образным, каждая индивидуальная Т клетка экспрессирует только единственный специфический TCR. Однако, каждая Т клетка обычно экспонирует тысячи копий этого рецептора, специфического в отношении только одного пептида, на поверхности каждой клетки. Кроме того, несколько других типов мембраносвязанных белков участвуют в связывании и активации Т клеток.
Активация Т клеток влечет за собой появление ряда химических сигналов (главным образом цитокинов), которые непосредственно заставляют клетки действовать или стимулировать действие других клеток иммунной системы. В случае активации комплекса белков МНС класса 1-антиген CTL пролиферируют и разрушают инфицированные клетки, презентирующие тот же антиген. Киллинг инфицированных клеток лишает вирус жизненно важной поддержки и делает его доступным для антител, которые, наконец, их удаляют. Напротив, активация Th клеток комплексами белок МНС класса П-антиген не разрушает антигенпрезентирующую клетку (которая является частью защитной системы хозяина), но, скорее, стимулирует Th клетку размножаться и генерировать сигналы (опять же главным образом цитокины), которые влияют на многие клетки. Помимо этих результатов, передача сигнала вызывает В-клеточную стимуляцию, активацию макрофагов, дифференцировку CTL и стимуляцию воспаления. Этот согласованный ответ является относительно специфическим и направлен на чужеродные элементы, несущие пептид, представляемый системой класса II МНС.
Постоянный контроль эпитопов во всех этих структурах в организме, находящемся под воздействием системы иммунного контроля, дает очень эффективные способы распознавания и сохранения "своих" и уничтожения эпитопов и их носителей, поражающих организм или вызывающих патологию. При правильном действии иммунный ответ вызывает поразительно эффективное элиминации микроскопических патогенов и неопластических (опухолевых) клеток, которые, как полагают, постоянно возникают в организме и, по большей части, удаляются иммунной системой ранее, чем их можно обнаружить. Некоторые части тела, такие как мозг, глаз и яички, защищены от иммунного контроля, про эти области также говорится, что они обладают иммунной привилегией. Обычно сложные механизмы распознавания "своего" очень эффективны и способствуют направлению сильного ответа на чужеродные антигены. К сожалению, иммунная система иногда работает неправильно (отказывает) и направляет свое действие против клеток хозяина, вызывая аутоиммунный ответ. Как правило, аутоиммунная реакция происходит, когда антигенные рецепторы на иммунных клетках распознают специфические антигены на здоровых клетках и вызывают гибель клеток, несущих эти конкретные вещества. Во многих случаях аутоиммунные реакции являются самоограниченными, так как они прекращаются, когда вызвавшие их антигены исчезают. Однако, в некоторых случаях аутореактивные лимфоциты живут дольше, чем должны были бы, и продолжают индуцировать апоптоз или иным способом элиминировать нормальные клетки.
Последние данные показывают, что иммунная защита против всех видов рака требует генерирования сильного клеточного иммунного ответа на уникальный опухолевый антиген, экспрессируемый в клетках злокачественных опухолей. В результате для успешной иммунной защиты, во-первых, требуется уникальный антиген, экспрессируемый в опухолевых клетках (опухолеспецифический антиген), и, во-вторых, индукция мощного Т-клеточного иммунного ответа, нацеленного на опухолевый антиген.
В настоящее время известно несколько ассоциированных с опухолями антигенов, они используются в преклинических и клинических исследованиях для получения вакцин. Например, PSMA, РАР и PSA представляют собой антигены, экспрессирующиеся в опухолевых клетках простаты. Her2/neu и MUC1, экспрессируемые раковыми клетками молочной железы и другими раковыми клетками, включая клетки рака легкого, яичников, толстой кишки и поджелудочной железы. MAGE и MART-1 являются антигенами, связанными с опухолевыми клетками меланомы, а СЕА представляет собой антиген, ассоциированный с поджелудочной железой или колоректальным раком. Описаны также другие ткане- и/или опухолеспецифические антигены. Однако, хотя все эти антигены экспрессируются в опухолевых клетках в нормальной или аберрантной форме, они также экспрессируются в ряде нормальных клеток и, следовательно, не могут использоваться для профилактической вакцинации. Другими словами, эти опухолевые антигены распознаются также иммунными клетками как "свои" молекулы, и, следовательно, не происходит настоящей активации иммунной системы. Это создает, по меньшей мере, два препятствия для применения этих ассоциированных с опухолями молекул в качестве основы для вакцины. Первым препятствием является иммунологическая неотвечаемость (толерантность) иммунной системы на "свои" молекулы, что ограничивает ее способность вырабатывать мощный клеточный иммунный ответ. Вторая трудность заключается в том, что любой выработанный мощный клеточный иммунный ответ не должен быть направлен на нормальные клетки, которые экспрессируют антиген-мишень. По этой причине все обсуждавшиеся выше опухолевые антигены предлагается использовать только в качестве мишеней для терапевтической вакцинации.
Недавно описан новый белок, который позволит преодолеть трудности, связанные с известными опухолевыми антигенами. Близкий родственник ("брат") регулятора импринтных сайтов (BORIS) впервые был описан как ДНК-связывающий белок, обнаруженный в яичке. Этот белок содержит 11 доменов цинковых пальцев (ZF), общих с CCCNC-связывающим фактором (CTCF), который является поливалентным нуклеарным фактором, содержащим 11 цинковых пальцев. CTCF является консервативным, убиквитарным и в высшей степени изменчивым фактором, участвующим в различных аспектах регуляции гена и образующим подверженные метилированию инсуляторы, которые регулируют инактивацию Х хромосомы и экспрессию импринтных генов. Однако, BORIS отличается от CTCF на N и на С конце и экспрессируется взаимоисключающе с CTCF в процессе развития клеток эмбриона мужского пола. Экспрессия BORIS ограничивается яичком, а затем только выбранной клеточной субпопуляцией сперматоцитов, которые вместе с регуляцией (восстановлением) меток метилирования участвуют в процессе развития клеток эмбриона мужского пола. Эта субпопуляция клеток яичек также является единственным типом нормальных клеток, о котором известно, что он не экспрессирует CTCF. Так как ингибирование экспрессии CTCF в культивированных клетках ведет к апоптозу, резонно предположить, что BORIS активируется, сохраняя некоторые жизненно важные функции BORIS в клетках яичек (Loukinov et al. (2002) Proc. Natl. Acad. Sci. 99 (10): 6806-6811).
Позднее было показано, что хотя сверхэкспрессия CTCF также блокирует клеточную пролиферацию, экспрессия BORIS в нормальных BORIS-негативных клетках стимулирует рост клеток, который может привести к трансформации (Klenova et al. (2002) Cancer Biol. 12: 399-414). Человеческий BORIS картирован в области 20q13, которая общеизвестна по частым конверсиям и/или амплификациям, наблюдаемым во многих опухолях тех же типов, в которых также часто наблюдается потеря гетерозиготности (LOH) в паралогичных генах на 16q22, где расположен CTCF. Эти области ассоциированы с "горячими точками", связанными с раком молочной железы, простаты, яичника, желудка, печени, эндометриального рака, злокачественной глиомы, рака толстой (ободочной) кишки и эзофагального рака, а также опухолей Вилмса. Важно, что аномальная активация экспрессии BORIS обнаруживается в значительном числе широкого ряда новообразований. Используя Нозерн-блоттинг или RT-PCR, Klenova et al. (2002) проанализировали уровни мРНК BORIS в более чем 200 линиях клеток, представляющих большинство основных форм человеческих опухолей, и обнаружили транскрипты в более чем половине испытанных линий клеток. Последующий анализ первичного рака, для выборки рака молочной железы, подтвердил результаты, полученные с этими клеточными линиями.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к нефункциональным мутантным полинуклеотидам, кодирующим опухолевый антиген "брата" регулятора импринтных сайтов (BORIS), и применению таких полинуклеотидов для профилактической вакцинации и иммунотерапии первичного или метастатического рака. Полинуклеотид может представлять собой либо ДНК, либо РНК. В одном предпочтительном варианте изобретения опухолевый антиген представляет собой нефункциональную мутантную форму молекулы BORIS, лишенной способности связываться с ДНК. В другом предпочтительном варианте изобретения, по меньшей мере, одни домен "цинковый палец" (ZF) является нефункциональным вследствие мутации или делеции, и функция BORIS элиминируется. В другом предпочтительном варианте изобретения любая комбинация доменов "цинковый палец" мутирует или удаляется, и функция белка, полипептида или пептида BORIS элиминируется. Еще в одном предпочтительном варианте изобретения удаляются (делегируются) все ZF-связывающие сайты. Еще в одном предпочтительном варианте полинуклеотид, кодирующий мутантную форму BORIS, сливается с молекулярным адъювантом. Еще в одном предпочтительном варианте полинуклеотид, кодирующий нефункциональную мутантную форму BORIS, смешивается, по меньшей мере, с одним отличным полинуклеотидом, кодирующим молекулярный адъювант. Можно использовать любой молекулярный адъювант, который повышает клеточный иммунный ответ. Цитокины, хемокины и молекулы костимуляторов являются особенно предпочтительными. Особенно предпочтительными хемокинами, цитокинами и молекулами костимуляторов являются бета-дефензин2, IL12, IL18, MIPα3, IFNγ и CD80/86.
Настоящее изобретение относится также к вектору, содержащему полинуклеотид, кодирующий нефункциональную мутантную форму BORIS. В предпочтительном варианте изобретения вектор направляет экспрессию в системе бактериальных клеток, клеток млекопитающих, клеток дрожжей или в вирусной системе.
Кроме того, настоящее изобретение относится к нефункциональной
модифицированной (мутантной) форме белка, полипептида или пептида BORIS. Нефункциональный мутант можно получать любым известным методом, который вводит в последовательность делеции, замены или добавления, что приводит в результате к нефункциональному белку. В предпочтительном варианте изобретения мутантный BORIS белок, полипептид или пептид лишен ДНК-связывающей способности. В другом предпочтительном варианте изобретения мутантный BORIS белок, полипептид или пептид смешивается с обычным адъювантом. Еще в одном предпочтительном варианте изобретения нефункциональный мутантный BORIS белок, полипептид или пептид связывается с фармацевтически приемлемым носителем (остов). Еще в одном предпочтительном варианте изобретения нефункциональный мутантный BORIS белок, полипептид или пептид связывается с пептидом, который модифицирует BORIS и сохраняет его антигенные свойства. Еще в одном предпочтительном варианте изобретения нефункциональный мутантный BORIS белок, полипептид или пептид связывается с областью белковой трансдукции (PTD).
Настоящее изобретение также относится к дендритным клеткам, экспрессирующим молекулу нефункционального мутантного BORIS. В предпочтительном варианте изобретения дендритные клетки трансфецируются при использовании ДНК, кодирующей молекулу мутантного BORIS. Еще в одном предпочтительном варианте изобретения дендритные клетки инфицируются вирусным вектором, который кодирует молекулу нефункционального мутантного BORIS. Еще в одном предпочтительном варианте изобретения дендритные клетки "нагружают" нефункциональным мутантным BORIS белком, полипептидом или пептидом, или любой нефункциональной модифицированной белковой формой BORIS.
Настоящее изобретение охватывает клеточные иммунные ответы, вырабатываемые против нефункциональной мутантной формы BORIS белка, полипептида или пептида или любой нефункциональной модифицированной белковой формы BORIS. Настоящее изобретение охватывает антитела к нефункциональной мутантной форме BORIS белка, полипептида или пептида или любой модифицированной белковой форме BORIS.
Настоящее изобретение также охватывает профилактическую или терапевтическую вакцину против рака, содержащую полинуклеотид, кодирующий нефункциональную мутантную форму BORIS, нефункциональную мутантную BORIS белка, полипептида или пептида, или дендритные клетки, экспрессирующие нефункциональную мутантную молекулу BORIS.
Настоящее изобретение также относится к способу лечения рака, заключающемуся во введении пациенту (нуждающемуся в профилактической вакцине), эффективного количества полинуклеотида, кодирующего нефункциональную мутантную форму BORIS, нефункциональной мутантной формы BORIS белка, полипептида или пептида, или дендритных клеток, экспрессирующих или содержащих нефункциональную мутантную молекулу BORIS. Введение может быть осуществлено внутримышечным, подкожным, интрадермальным, внутривенным, назальным, ректальным, вагинальным или перитонеальным способом. Рак может представлять собой первичный или метастатический рак. У пациента может быть рак нескольких различных типов. В предпочтительном варианте изобретения рак представляет собой рак молочной железы, простаты, яичника, желудка, печени, эндометриальный рак, злокачественную глиому, рак толстой (ободочной) кишки или эзофагальный рак.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
На Фигурах 1a, b представлены результаты вакцинации мышей (n=10) с помощью BORIS (иммунизация ДНК) pIL12/IL18 (молекулярный адъювант). Это приводит к защите мышей от контрольного заражения опухолевыми клетками 104 4Т1, естественно экспрессирующими мышиный BORIS. На Фигуре 1а по оси Y показана степень выживания, а по оси Х откладываются дни после контрольного заражения для pBORIS/pIL12/IL18, pIL12/IL18 и одного лишь вектора. На Фигуре 1b показано соотношение между объемом опухоли и количеством дней после контрольного заражения опухолью для pBORIS/pIL12/IL18 по сравнению с pIL12/IL18 и одним лишь вектором (*Р<0.001).
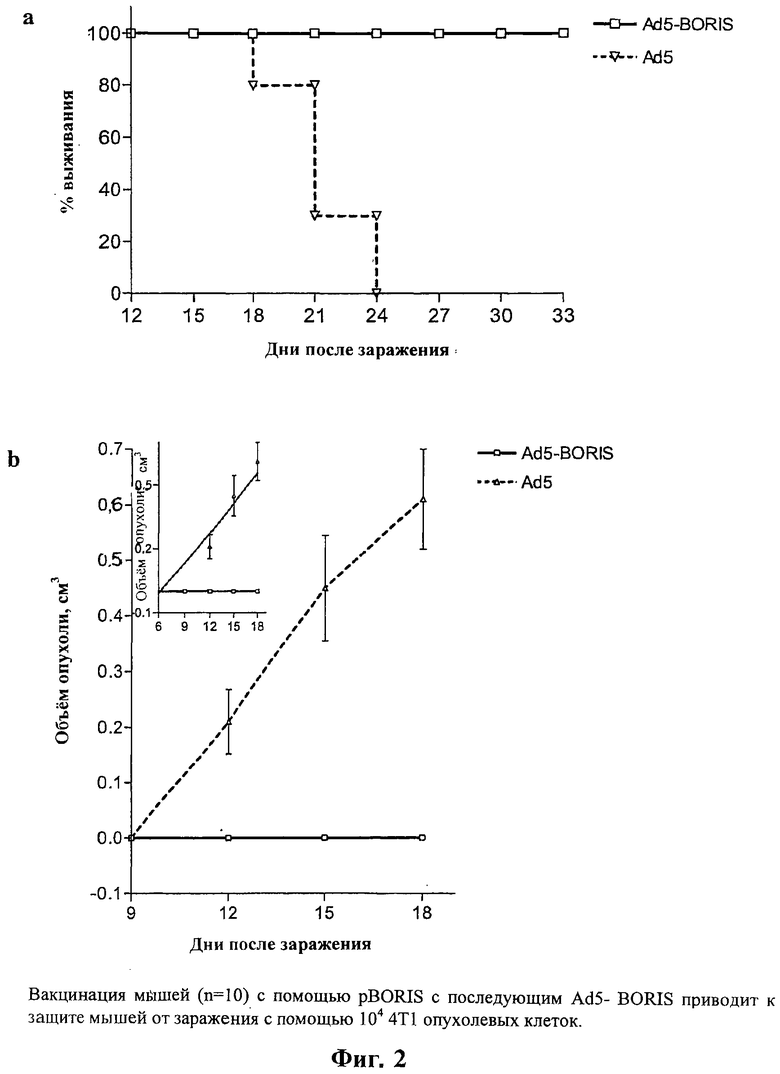
На Фигуре 2а и b показано соотношение между процентом выживших и числом дней после контрольного заражения клетками опухоли (а) и между объемом опухоли и числом дней после контрольного заражения для мышей, вакцинированных pBORIS (ДНК-иммунизация) с последующим Ad5-BORIS (вирусоподобные частицы), и зараженных с помощью 104 4Т1 клеток. Данные демонстрируют полную защиту против контрольного заражения клетками опухоли, по меньшей мере, через 33 дня после контрольного заражения.
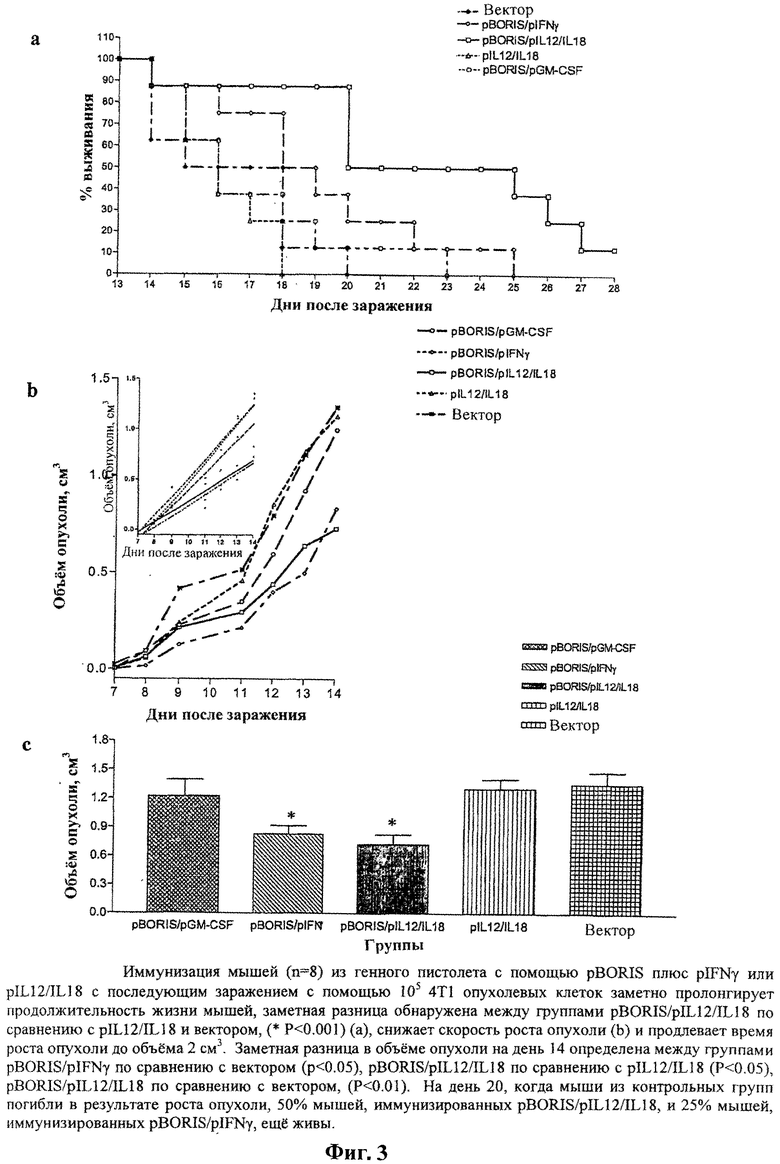
На Фигуре 3а, b и с показаны результаты иммунизации мышей pBORIS плюс pIFNγ или pIL12/IL18 с помощью генного пистолета с последующим контрольным заражением 105 клетками опухоли 4Т1. На Фигуре 3а показано продолжительное время роста опухоли до объема 2 см3, а на Фигуре 3b показано более низкая степень роста опухоли. На Фигуре 3с показана значительная разница объема опухоли на день 14 между группами pBORIS/pIFNγ по сравнению с вектором (р<0.05), pBORIS/pIL12/IL18 по сравнению с pIL12/IL18 (Р0.05), и pBORIS/pIL12/IL18 по сравнению с вектором (Р<0.01).
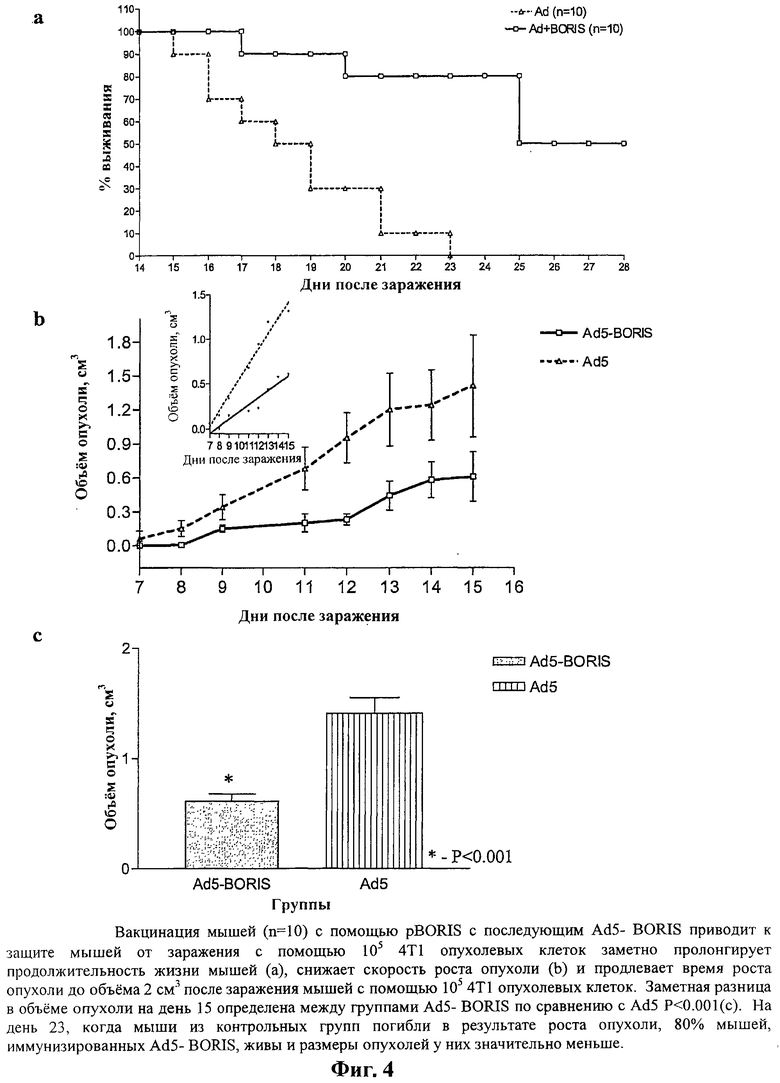
На Фигуре 4а, b и с показаны результаты вакцинации с помощью pBORIS (ДНК-иммунизация) с последующей инъекцией Ad5-BORIS (вирусоподобные частицы). На Фигуре 4а показана значительно пролонгированная продолжительность жизни вакцинированных мышей, тогда как на Фигуре 4b показано, что у этих мышей наблюдается более низкая скорость роста опухоли и более продолжительное время роста опухоли до объема 2 см3 после контрольного заражения мышей опухолевыми клетками 4Т1 105. На Фигуре 4 с показана значительная разница в объеме опухоли на день 15 между группами Ad5-BORIS по сравнению с Ad5 (P<0.001).
На Фигуре 5 показано выравнивание методом наилучшего приближения человеческого и мышиного BORIS полипептидов, осуществляемое с помощью пакета программ GCG, по умолчанию расширение гэпов принимается равным нулю, причем консервативные области цинковых пальцев выделены и указаны как ZF-11.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Хотя настоящее изобретение может быть воплощено во многих различных формах, в данном описании раскрываются конкретные иллюстрирующие варианты изобретения, которые поясняют на примерах принципы изобретения. Следует подчеркнуть, что настоящее изобретение не ограничивается конкретными ограничивающими примерами.
Настоящее изобретение включает применение антигена, экспрессируемого только в иммунологически привилегированных клетках яичка и появляющегося во многих трансформированных опухолевых клетках, для предупреждения эффекта толерантности, который может индуцировать другие опухолевые антигены. Данное изобретение также включает введение специфических изменений в ДНК, кодирующую антиген, для элиминации побочных эффектов и аутоиммунных реакций. В данном контексте применяются следующие определения.
Термин "опухоль", "рак", "новообразование", "неоплазия" и их этимологические родственники применяются попеременно (на равных правах, взаимозаменяемо) в контексте данной заявки для обозначения в целом аномальных пролиферативных заболеваний и сопутствующих пораженных клеток или клеточных масс. Предпочтительно, клетки с аномальной пролиферацией относятся в данном описании к антигену с иммунной привилегией.
Цитотоксические Т лимфоциты (CTL) представляют собой эффекторные Т клетки, обычно CD8+, которые могут опосредовать лизис клеток-мишеней, несущих антигенные пептиды, ассоциированные с молекулой МНС. Другие цитотоксические клетки включают гамма/дельта и CD4+ NK.1.1+ клетки.
Выражения "антиген с иммунной привилегией" и "иммунологически привилегированный антиген" относятся к выделению определенных сайтов и антигенов в организме из иммунной системы и, следовательно, связаны с антигенами, в отношении которых вырабатывается аномальный иммунный ответ. Антигены с иммунной привилегией, экспрессируемые эктопически (т.е. вне их обычно иммунологически привилегированных сайтов), могут вызвать аутоиммунитет или иммунитет к опухоли. Иммунологически привилегированные антигены экспрессируются в некоторых опухолях, что приводит к выработке иммунного ответа как в отношении опухолевого, так и неопухолевого сайта, экспрессирующего те же иммунологически привилегированные антигены.
Антигенпрезентирующие клетки (АРС) представляют собой клетки, включая дендритные клетки, макрофаги и В клетки, которые могут процессировать и презентировать (представлять) антигенные пептиды совместно с молекулами МНС класса I или класса II и доставлять костимулирующий сигнал, необходимый для активации Т клеток.
"Область (домен) цинкового пальца" ("цинковые пальцы") относится к малому независимо складывающемуся ("выпетливающемуся") домену, которому для стабилизации его структуры необходима координация с одним или более ионов цинка. Пальцы связываются с субсайтами из трех пар оснований, а специфические контакты опосредуются аминокислотами в положениях - 1, 2, 3 и 6 относительно начала альфа-спирали.
"Нефункциональная мутантная форма BORIS" относится к BORIS белку, полипептиду или пептиду с потерей функции. "Потеря функции" предполагает обозначать невозможность осуществить любую из важных активностей молекулы дикого типа BORIS, такую как ДНК-связывание, восстановление "отцовского" ("родительского") паттерна ДНК-метилирования и т.д.
"Нефункциональный мутант" относится к изменениям уровня ДНК или белка, которые нарушают активность дикого типа полученного белка. Такие изменения могут представлять собой аминокислотные замены, делеции или добавления в областях молекулы, ведущих себя как каталитические сайты и/или принимающих участие в связывании ДНК или белка. Примерами изменений, способных нарушить (уничтожить) активность, являются делеции или замены важных аминокислот, принимающих участие в каталитическом или связывающем взаимодействии, добавления аминокислот, которые изменяют необходимую трехмерную структуру сайта, участвующего в каталитических и/или связывающих взаимодействиях, или добавления или делении нуклеотидов, которые вызывают сдвиги рамки считывания, тем самым нарушая необходимую трехмерную структуру. Мутации можно осуществлять обычными молекулярными методами, такими как ПЦР, применение олигонуклеотидов и т.д. (см., например, Sambrook, Maniatis и Fritsch). Природные мутации также можно выделить из популяции клеток (см., например, Sambrook, Maniatis и Fritsch).
"Пептид" относится к молекуле, содержащей, по меньшей мере, 2 аминокислоты, соединенные пептидной связью. "Полипептид" относится к молекуле, содержащей, по меньшей мере, 10 аминокислот, соединенных пептидными связями, а "белок" относится к молекуле, содержащей, по меньшей мере, 20 аминокислот.
"Полинуклеотид, кодирующий нефункциональную мутантную форму BORIS" относится к любому полинуклеотиду, последовательность которого, по меньшей мере, на 50%, 60% или 70% идентична последовательности человеческого (SEQ ID NO:1) или мышиного (SEQ ID NO:3) полинуклеотида BORIS, более предпочтительно, на 75%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентична последовательности человеческого (SEQ ID NO:1) или мышиного (SEQ ID NO:3) полинуклеотида BORIS.
"Нефункциональный мутантный BORIS пептид, полипептид или белок" относится к молекуле BORIS, которая лишена возможности осуществить любую из важных активностей молекулы дикого типа BORIS, такую как ДНК-связывание, восстановление "отцовского" ("родительского") паттерна ДНК-метилирования и т.д. "Нефункциональный мутантный BORIS пептид, полипептид или белок" имеет последовательность, которая, по меньшей мере, на 50%, 60% или 70% идентична последовательности человеческого (SEQ ID NO:2) или мышиного (SEQ ID NO:4) пептида, полипептида или белка BORIS, более предпочтительно, на 75%, 80%, 90%, 95%, 96%, 97%, 98% или 99% идентична последовательности человеческого (SEQ ID NO:2) или мышиного (SEQ ID NO:4) пептида, полипептида или белка BORIS.
Молекула нефункционального мутантного BORIS распознается как "чужой" антиген, экспрессируемый только в трансформированных опухолевых клетках, и используется в качестве антигена для преодоления ограничений предыдущего уровня техники. Мутантная форма BORIS используется в качестве идеальной нетоксической вакцины, так как она не должна вызывать нежелательных побочных эффектов, вызываемых его активностью ДНК-связывания и/или нативной функцией. Другими словами, мутантный BORIS, применяемый для вакцинации, не обладает функциональной активностью и присутствует только как иммуноген (антиген). В отличие от других опухолеспецифических антигенов, BORIS не экспрессируется в нормальных тканях у женщин. Кроме того, даже хотя BORIS экспрессируется в процессе полового созревания в нормальном яичке у мужчин, введение и/или экспрессия нефункционального мутантного BORIS должна быть безвредной, так как яичко является иммунологически привилегированным органом (недоступным для иммунных клеток). Другими словами, анти-BORIS иммунный ответ, вырабатывающийся после иммунизации, не является опасным для нормальных клеток, а вакцина BORIS не вызывает аутоиммунитет. Помимо этого, гарантируется вырабатывание мощного иммунного ответа, так как BORIS, в отличие от других опухолеспецифических антигенов, распознается как чужеродный антиген. BORIS-специфические Т клетки не уничтожаются в тимусе, распознают мутантный BORIS как "чужой" антиген и вырабатывают иммунный ответ.
В одном варианте изобретения кДНК, кодирующую мышиный BORIS (mBORIS), получают методом RT-PCR на мРНК, выделенной из мышиного или человеческого яичка. ДНК-связывающий домен молекулы делегируется и заменяется малым спейсером, о котором известно, что он хорошо работает при создании антител-одноцепочечных Fv доменов. Корректная последовательность подтверждается автоматизированным анализом нуклеотидной последовательности. В полученной в результате молекуле отсутствуют 11 ZF доменов, и она состоит из N-концевой области m BORIS (аминокислоты 1-258), связанной с С-концевой областью (аминокислоты 573-636) через спейсер, содержащий 18 аминокислот.
Мутантную кДНК клонируют в вектор pORF под контролем промотора hEF1-HTLV, однако можно использовать другие векторы экспрессии. Мутантная кДНК функционально связывается с промотором и/или регуляторными молекулами, способными вызывать экспрессию в клетке-хозяине. Можно применять вирусные векторы, включая векторы а вирусной ДНК или РНК, аденовирусы и ретровирусы (см. Vasilevko, V. et al. (2003) Clin. Exp.Metastas. 20: 489-98; Leitner, W.W. et al. (2003) Nat Med 9: 33-39; Ribas, A et al (2002) Curr. Gene Ther 2: 57-58).
Помимо приведенного выше, изобретение охватывает применение вирусоподобных частиц, кодирующих молекулы нефункционального мутантного BORIS, таких как аденовирус, вирус человеческого гепатита В, вирус человеческого гепатита С, вирус осповакцины, полиовирус и т.д.. Рекомбинантные вирусные белки различных вирусов обладают полезным свойством самосборки в вирусоподобные частицы (VLP). Эти частицы не содержат вирусных нуклеиновых кислот, и, следовательно, являются нерепликативными, неинфекционными и сохраняют конформационно корректные антигенные эпитопы. Продуцирование VLP показано во многих экспериментальных системах, таких как клетки млекопитающих, инфицированные бакуловирусом клетки насекомых, дрожжей, Е. coli, бесклеточные системы и трансгенные растения. Важно отметить, что вакцинация с помощью VLP вызывает вырабатывание не только гуморального, но также и клеточного иммунных ответов. VLP инфицируют "профессиональные" АРС, а затем индуцируют защитные клеточные иммунные реакции, включая CD4+Th1 (тип CD4+T клеток, помогающих клеткам CD8+T) и CD8+CTL ответы. Таким образом, VLP четко показали исключительную способность активировать клеточный иммунный ответ (Т-клеточный ответ). Возможное применение VLP в качестве профилактических вакцин в настоящее время проверяют в ряде различных клинических испытаний. Результаты этих испытаний обнадеживают вследствие прекрасной переносимости и высокой иммуногенности, о которых сообщается в этих исследованиях. Получение VLP вакцины, состоящей из переносимого BORIS антигена, будет стимулировать сильный клеточный иммунный ответ против раковых клеток, экспрессирующих этот опухолевый антиген. Коровый антиген (HBcAg) вируса гепатита В (HBV) и VSV являются примерами подходящих VLP.
Для получения более сильного (устойчивого) клеточного иммунного ответа усеченный или мутантный mBORIS перед клонированием в вектор объединяют с молекулярными адъювантами, такими как молекулы костимулятора В7, бета-дефензин 2/3, MIP3α, IFNγ, цитокины, хемокины и т.д. Другие подходящие молекулярные адъюванты перечислены в нижеприведенной Таблице 1.
Или же для ДНК-иммунизации можно использовать обычные адъюванты, такие как Tween 80, 5% этанол или бупивокаин. Другие примеры традиционных адъювантов включают минеральные соли (такие как гели гидроксида алюминия или фосфата алюминия), масляные эмульсии и составы на основе поверхностно-активных веществ,такие как MF59, QS21, AS08 [SBAS2] (эмульсия масло- в воде+MPL+QS21), Монтанид ISA-51 и ISA-720, адъюванты из макрочастиц, такие как виросомы, AS04 [SBAS4] соль А1 с MPL), ISCOMS, сополимер лактида с гликолидом (PLG), соединения микробиального происхождения (природные и синтетические), включая монофосфориллипид А (MPL), Detox (MPL+M.Phlei скелета клеточных стенок), AGP [RC-529], DC_Chol, ОМ-174 (производное липида А), мотивы CpG, модифицированные LT и СТ (генетически модифицированные бактериальные токсины), эндогенные иммуномодуляторы, такие как GM-CSF, IL-12, иммудаптин, а также все другие хемокины, цитокины и костимулирующие молекулы, перечисленные в Таблице, приведенной выше, и инертные носители, такие как частицы золота.
Адъюванты можно смешивать с полинуклеотидом, кодирующим нефункциональную мутантную форму белка, полипептида или пептида родственника регулятора импринтных сайтов (BORIS), нефункциональным мутантным BORIS белком, полипептидом или пептидом и дендритной клеткой, экспрессирующей нефункциональный мутантный BORIS пептид, полипептид или белок.
Для повышения/промотирования презентации нефункционального мутантного BORIS "профессиональными" антигенпрезентирующими клетками (АРС) метаболического пути МНС класса в конструкции нефункционального мутантного BORIS можно включать дополнительные пептидные молекулы. Одним из таких примеров является конструкция, создаваемая с помощью трансдуцирующего пептид домена (PTD). В целом, иммунный ответ основывается на процессировании и презентации нативного антигена. Опухолевые антигены могут экспрессироваться в клетках бактерий, дрожжей и млекопитающих, однако белковые антигены, экспрессируемые в этих системах, по-видимому, не стимулируют максимальный Т-клеточный иммунный ответ (ответы, склонные либо к CTL, либо к Тh1 ответам), так как растворимые экзогенные белки процессируются, в основном, с использованием метаболического пути МНС класса II. На самом деле, многие противоопухолевые вакцины основаны на индукции CD8+CTL, но для этого обычно требуется, чтобы белок синтезировался в цитозоле АРС. К сожалению, в целом плазматические мембраны эукариотических клеток непроницаемы для большинства белков. Недавно было показано, что чужеродные белки, соединенные с белок-традуцирующим доменом (PTD), могут проникать через плазматическую мембрану, что позволяет аккумулировать белок внутри клеток. Это повышает презентацию чужеродных пептидов молекулами МНС класса II АРС антигенспецифическим CD8+Т клеткам.
Вакцинация/иммунизация
Препараты (рецептуры, композиции) вакцин до настоящему изобретению содержат иммуногенное количество полинуклеотида, кодирующего нефункциональную мутантную форму белка, полипептида или пептида BORIS, нефункционального мутантного BORIS белка, полипептида или пептида, или дендритных клеток, экспрессирующих нефункциональный мутантный BORIS пептид, полипептид или белок, в комбинации с фармацевтически приемлемым носителем. Можно использовать мимеотопы, которые представляют собой полипептиды с неродственной последовательностью, но с трехмерной структурой, соответствующей нефункциональному мутантному BORIS белку, полипептиду или пептиду, и которые функционируют идентичным образом. Мимеотипы, которые представляют собой любую биологическую молекулу, не родственную структуру BORIS, но имеющую идентичный(е) 3х-мерный(е) эпитоп(ы), могут распознаваться анти-BORIS клетками.
"Иммуногенное количество" представляет собой количество полинуклеотида, кодирующего нефункциональный мутантный BORIS, белок, полипептид или пептид; нефункционального мутантного BORIS, белка, полипептида или пептида; или дендритных клеток, экспрессирующих нефункциональный мутантный BORIS, белок, полипептид или пептид, достаточное для того, чтобы вызвать иммунный ответ у субъекта, которому вводится вакцина. Вводимое количество представляет собой количество, которое индуцирует заданный иммунный ответ и заданную степень защиты. Примеры фармацевтически приемлемых носителей включают, но без ограничения, стерильную апирогенную воду и стерильный апирогенный физиологический солевой раствор.
Рецептуры вакцин по настоящему изобретению пригодны для пациентов, у которых диагностирован, по меньшей мере, один тип рака, включая, но без ограничения, рак молочной железы, простаты, яичника, желудка, печени, эндометриальный рак, злокачественную глиому, рак толстой (ободочной) кишки или эзофагальный рак. Рецептуры вакцин по данному изобретению также годятся для пациентов с известной генетической предрасположенностью к раку. Кроме того, рецептуры вакцин по настоящему изобретению пригодны для всей популяции в целом, включая представителей популяции, не болеющих раком или не имеющих генетической предрасположенности к раку, желающих прибегнуть к защите от контакта, по меньшей мере, с одним типом раковых опухолей, в которых экспрессируется BORIS, белок, полипептид или пептид.
Введение вакцины можно осуществлять любым подходящим способом, включая парентеральную инъекцию (такую как интраперитонеальная, подкожная или внутримышечная инъекция), интрадермальную, внутривенную инъекцию, назальное, ректальное, вагинальное применение или введение через дыхательные пути. Местное применение вируса для введения через дыхательные пути можно осуществлять с помощью интраназального введения (например, с помощью пипетки, тампона (турундочки) или ингалятора, которые вводят в нос фармацевтический препарат). Местное применение вируса для введения в дыхательные пути также можно вводить ингаляцией, например, создавая фармацевтический препарат в виде респирабельных частиц (включая как твердые частицы, так и частицы жидкости), содержащих репликон, а затем побуждая субъекта вдыхать респирабельные частицы. Способы и аппаратура для введения респирабельных частиц фармацевтических препаратов общеизвестны, и может применяться любой обычный метод. "Иммуногенное количество" представляет собой количество частиц репликона, достаточное для того, чтобы вызвать иммунный ответ у субъекта, которому вводится вакцина.
Если в качестве вакцины используют РНК или ДНК, РНК и ДНК можно вводить непосредственно методами, известными специалистам в данной области техники, такими как доставка на золотых гранулах (генный пистолет), доставка с помощью липосом, или непосредственная инъекция. Одна или более конструкция или реплицирующая РНК может использоваться в любой комбинации, эффективно вызывая иммуногенный ответ у субъекта. Обычно количество нуклеиновых кислот во вводимой вакцине может быть таким, чтобы индуцировать заданный иммунный ответ и заданную степень защиты. Точные количества вводимой вакцины могут зависеть от решения практикующего врача и могут быть индивидуальными для каждого субъекта и антигена.
Вакцину можно вводить по схеме, предусматривающей однократный прием, или, предпочтительно, по схеме, предусматривающей многократный прием, когда первичный курс вакцинации может состоять из 1-10 раздельных доз, а затем, через последовательные временные интервалы дают другие дозы с целью сохранения или усиления иммунного ответа, например, 1-4 месяца для второй дозы и, при необходимости, последующая(ие) доза(ы) через несколько месяцев. Примеры подходящих схем иммунизации включают: (ш) 0, 1 месяц и 6 месяцев, (ii) 0, 7 дней и 1 месяц, (iii) 0 и 1 месяц, (iv) 0 и 6 месяцев, или другие схемы, пригодные для выявления заданных иммунных ответов, предположительно, обеспечивающих защитный иммунитет или уменьшающих симптомы заболевания или уменьшающих тяжесть заболевания.
Человеческий hBORIS можно выделять из человеческого яичка и манипулировать с ним подобным образом. Аналогично, BORIS можно выделять из яичка любого млекопитающего или позвоночного и использовать подобным образом.
ПРИМЕРЫ
1. Получение плазмиды, кодирующей ZF-делегированную форму молекулы mBORIS под контролем промотора hEF1-HTLV
Реакцию RT-PCR осуществляют, используя РНК поли-А мышиного яичка и следующие праймеры:
MB1F 5'-CGTCACCATGGCI GCCGCTGAGGTCCCTG
MB1R 5'-AAGCTTCTGAAAGCTCTGAGGCTITCCCTTOG
MB2F 5'-GGATCCGAGACGTTAGCCCCCAACAAGGACAGG
MB2R 5'-GAATTCTCACTTATCCATCATGTTAAAGATCATCTCGCAGG
SpF 5'-AGCTTGGTGGAGGCGGTTCAGGCGGAGGTGGCTCTGGCGGTGGCGGATCGG
SpR 5'-OATCCCOATCCOCCACCOCCAOAOCCACCTCCOCCTOAACCOCCTCCACCA
Условия ПЦР следующие: 94°С 30 сек, 60°С 30 сек, 72°С 2 мин. Проводят тридцать (30) циклов.
Продукты ПЦР субклонируют в клонирующий вектор PCRII-TOPO (Invitrogen). С-концевую кДНК в PCRII-TOPO подвергают рестрикции с помощью фермента BamHI, a позитивные клоны, содержащие инсерт, собирают и затем разрезают при использовании фермента HindIII. Праймеры (SpF и SpR) для амплификации спейсера отжигают, получают выступающие липкие концы и лигируют в вектор, разрезанный с помощью BamHI-HindIII. N-концевой кодируемый фрагмент разрезают, используя HindIII и инсерты (вставки), теперь отделенные от вектора, собирают и лигируют в конструкцию, разрезанную с помощью HindIII, содержащую С-конец и спейсер. Затем клоны с подходящей ориентацией отбирают, секвенируют (например, см. ниже последовательность молекулы ZF-делетированного BORIS) и субклонируют в плазмиду pORF под контролем промотора hEF1-HTLV (Invitrogen).
Клетки СНО трансфецируют с помощью полученной конструкции, используя стандартные методы молекулярной генетики (Sambrook J, Fritsch EF and Maniatis Т (1989) Molecular Cloning: A Laboratory Manual, Cold Spring Harbor).
Экспрессию конструкции mBORIS без цинковых пальцев (с делениями цинковых пальцев) анализируют Нозерн-блоттингом мРНК, выделенной из клеток СНО стандартным методом молекулярной генетики Sambrook et al. 1989).
2. Иммунизация мышей с помощью ДНК. кодирующей ZF-делетированную форму молекулы BORIS
Плазмиду, кодирующую ZF-делетированную конструкцию mBORIS, выделяют, используя набор EndoFree Plasmid maxi (Qiagen). Чистоту плазмиды ДНК подтверждают УФ-спектром (соотношение оптический плотностей при 260 нм/280 нм >1.7) и гель-электрофорезом.
Золотые гранулы покрывают ДНК (1 мкг/0.5 мг золота) и 5-7-недельных мышей Balb/c иммунизируют с помощью генного пистолета Helios. Мышей иммунизирую (обстреливают) подобным образом трижды за две недели. Через десять дней после последнего "обстрела" у мышей отбирают пробу крови и заражают, вводя 1.0×104 или 1.0×105 клеток 4Т1 рака молочной железы. Размер опухоли измеряют каждый день или каждые два-три дня с помощью штангенциркуля.
3. Иммунные исследования
Иммунизация мышей плазмидной вакциной:
Одним из примеров метода с применением профилактической противораковой вакцины является применение ДНК, кодирующей делегированную форму молекулы мышиного BORIS, в которой отсутствуют домены цинкового пальца и, следовательно, отсутствует свойство ДНК-связывания.
Очищенную плазмиду используют для покрытия золотых гранул (2 мкг плазмиды/0.5 мг золотых гранул), как описано нами ранее (Ghochikyan et al. (2003) Eur. J. hnmunol 33: 3232-41). Иммунизацию мышей BALB/с осуществляют через выбритую кожу живота с помощью генного пистолета Helios (Bio-Rad, Hercules, CA), как описано Ross et al. (2000, Nat. hnmunol 1: 127-131). Коротко говоря, мышей "бомбардируют" 3 раза дозами, содержащими 2 мкг ДНК на 0.5 мг золотых гранул размером ~1 мкм (DeGussa-Huls Corp., Ridefield Park, NJ) под давлением гелия 400 psi (2758 кПа). Мышей иммунизируют и обогащают (обстреливают) тем же методом раз в две недели и контрольно заражают двумя различными дозами клеток рака молочной железы (105 или 104) через десять дней после последнего обогащения, как описано (Vasilevko, V. et al. (2003). Различные группы мышей иммунизируют плазмидой, кодирующей модифицированный BORIS, смешивают с ДНК, кодирующей определенный(е) молекулярный(е) адъювант(ы) (подробнее см. Таблицу 2). Известно, что такие молекулярные адъюванты повышают клеточный иммунный ответ на различные антигены.
Получение аденовирусного вектора, кодирующего ZF- делетированную фоуму молекулы мышиного BORIS (Ad5-BORIS), и иммунизация мышей Рекомбинантный вирус Ad5-BORIS получают, используя аденовирусную векторную систему AdEasy XL от Stratagene. Шаттл-вектор конструируют субклонированием фрагмента ZF-делетированного mBORIS в плазмиду pShnttle-CMV. Для этой цели фрагмент BORIS синтезируют методом ПЦР, используя в качестве матрицы плазмиду pORF-mBORIS и следующие праймеры:
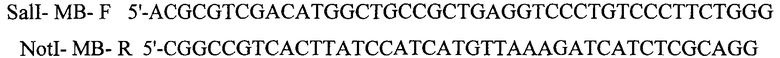
Продукт ПЦР субклонируют в клонирующий вектор PCR4-TOPO (Invitrogen). Фрагмент BORIS разрезают, используя рестриктазы Sail и Notl. Полученный продукт очищают на агарозном геле и субклонируют, используя сайты клонирования SalI-NotI, в вектор pShnttle-CMV.
Экспрессию in vitro ZF-делетированного mBORIS анализируют в клетках СНО иммуноблоттингом (на Фигуре 6, см. ниже).

Шаттл-вектор, несущий делегированный BORIS, линеаризуют с помощью Pmel и очищают на агарозном геле. Электрокомпетентные клетки BJ-5183-Ad-1 трансформируют с использованием плазмиды pShuttle-mBORIS, расщепляемой с помощью Рте, получают рекомбинантную Ad плазмиду. Клетки AD-293 трансфецируют при использовании ДНК выбранного рекомбинантного Ad-BORIS и готовят первичные исходные вирусные растворы. Полученный первичный вирусный раствор (10 бое/мл) амплифицируют в клетках AD-293, а затем очищают в CsCl-градиенте. Очищенный вирус диализуют против PBS - 5% сахароза и используют для иммунизации мышей.
Мышам Balb/c, иммунизированным с помощью pBORIS четыре раза через две недели, один раз вводят в.м. бустерную инъекцию Ad5-BORIS (109 БОЕ). Контрольным животным инъецируют векторы и проводят бустерную иммунизацию Ad5. Через десять дней после последней бустерной инъекции проводят контрольное заражение двумя различными дозами клеток 4Т1 рака молочной железы (105 или 104), как описано (Vasilevko, V. et al. (2003).
Линии опухолевых клеток
Используют опухолевые клетки млекопитающих, предоставленные доктором F. Miller (Karmanos Cancer Institute, Detroit, MI). Клетки 4Т1.2 представляют собой устойчивый к тиогуанину вариант, полученный из клеток 410.4 (клеточная линия млекопитающих, первоначально выделенная из единичной спонтанно выросшей опухоли млекопитающих у мышей BALB/c fC3H) без воздействия мутагена. Клетки культивируют (37°С, 10% СО2) в основной среде Игла, модифицированной по способу Дульбекко (DMEM), с низким содержанием глюкозы и с добавлением 5% фетальной бычьей сыворотки, 5% сыворотки новорожденных телят, 2 мМ глутамина, 100 Ед./мл пенициллина, 100 мг/мл стрептомицина, 0.1 мМ заменимых аминокислот и 1 мМ пирувата натрия (D10) (Life Technologies, Inc.).
Определение объема опухоли
Объемы опухолей определяют ежедневно, делая измерения по двум направлениям и рассчитывая по формуле L×(W2)/2, где L обозначает длину, a W обозначает ширину опухоли. Эксперименты заканчиваются со смертью мыши или по достижении опухолью размера около 1.5 см3 в экспериментах, включающих контрольное заражение с помощью 104 клеток, и 2 см3 в экспериментах, включающих контрольное заражение с помощью 105 клеток 4Т1.
Время образования (латентный период) обозначается как время до момента, когда опухоль достигнет объема более 0.1 см3. Для определения скорости роста опухолей анализируют кривые рассеяния в периоды почти линейного роста опухоли.
Статистический анализ
Результаты, полученные при определении среднего времени появления опухолевых узлов (латентный период) и роста опухоли (объем опухоли), а также время поствакцинального иммунитета, изучают методом анализа вариантов (ANOVA) и после испытания используют критерий Тьюки для множественных сравнений. Среднее и стандартное отклонение (девиацию) (SD) рассчитывают, используя программное обеспечение GraphPad Prism 3.0.
4. Иммунология
В- и Т-клеточный иммунный ответ против BORIS анализируют, применяя два различных протокола иммунизации.
1. Получение мышиных BORIS белков и иммунизация мышей.
ZF-делетированный фрагмент мышиного BORIS субклонируют в бактериальный экспрессирующий вектор pET24d(+), используя NcoI-XhoI сайты клонирования в рамке считывания с С-концевым 6His "хвостом". Вводят оба сайта и удаляют стоп-кодон на стадии клонирования ПЦР. Кроме того, создают плазмиды, кодирующие молекулу делегированного BORIS, слитую с доменом белковой трансдукции (PTD). Домен белковой трансдукции HIV-Tat (Tat47-57 YGRKKRRQRRR) соединяют с N-концом делегированного BORIS методом ПЦР, а затем клонируют в сайты клонирования NcoI-XhoI вектора pET24d(+). Штамм E.coli BL21(DE3), трансформированный при использовании полученных плазмид pET-mBORIS или pET-TATmBORIS, выращивают в LB с канамицином при 28°С до достижения А600 0-8. Синтез белка индуцируют, добавляя IPTG до конечной концентрации 1 мМ. Клетки собирают через три-пять часов центрифугированием и используют для очистки белка аффинной хроматографией на колонке с никель-NTA (нитрилотриуксусная кислота) (Qiagen). См. Фигуру 7.

Кроме того, ZF-делетированный фрагмент BORIS, слитый с PTD, субклонируют в экспрессирующий вектор дрожжей рОАРгальфа в рамке считывания с сигнальной последовательностью в сайты клонирования EcoRI-XbaI. Оба сайта вводят, а инициирующий кодон ATG удаляют в ходе ПЦР стадии клонирования. Штамм Pichia pastoris X33 трансформируют электропорацией с pGAPZ-BORIS, линеаризуют с использованием рестриктазы AvrII и позитивные клоны отбирают на среде YPD, содержащей 100 мкг/мл Zeocin. Для анализа экспрессии отобранные клоны выращивают в бульоне YPD/Zeocin и анализируют в супернатанте в различные временные точки иммуноблоттингом.
2. Иммунизация мышей дендритными клетками (DC).
Первичные DC костного мозга получают из клеток-предшественников в костном мозге мышей следующим образом. Истощенные эритроцитами клетки костного мозга из бедренных и большеберцовых костей засевают в полной среде RPMI-10 с добавлением рекомбинантного мышиного GM-CSF (100 Ед./мл). На день 3 не прикрепленные к субстрату гранулоциты осторожно удаляют и прибавляют свежую среду. Не прикрепленные к субстрату DC собирают на день 7 и очищают, используя набор для позитивной селекции (Miltenyi) с микрогранулами CD11c.
DC собирают на день 7 культивирования, очищают позитивной селекцией и инфицируют с помощью Ad5-BORIS, инкубируя при плотности клеток 107 клеток/мл в RPMI-1640 при множественности заражения 1000-2000. Через 1 час полную среду добавляют для разбавления DC до конечной концентрации 1×106-2×106 клеток/мл. Через 24 часа клетки собирают, интенсивно отмывают, чтобы исключить любой перенос аденовирусных частиц, и используют для иммунизации. Кроме того, DC, которые собраны в день 7 культивирования и очищены позитивной селекцией, инкубируют с 10 мкг/мл ZF-делетированного белка mBORIS при 37°С, 5% СО2 в течение 24 часов, дважды отмывают PBS. Поглощение белка клетками DC анализируют в аликвотах методом проточной цитометрии с применением антител против мышиного BORIS и соответствующих вторичных антител, меченных FITC. Мышей Balb/c иммунизируют и.п.(интраперитонеально) трижды каждые три недели с помощью 106 DC и Т-клеточный ответ анализируют через 10 дней после последнего введения в культуры (бустер) культуры спленоцитов.
5. Результаты
Результаты иммунизации представлены на Фигурах 1-4.
ДНК, кодирующую мутантную форму специфического в отношении раковых клеток мышиного BORIS антигена без ДНК-связывающей функции (делеция 11-цинковых пальцев), создают, используя экспрессирующие векторы млекопитающих pORF (hivivogen) и аденовирусную векторную систему AdEasy XL (Stratagene). Эти вакцины используют в качестве профилактической противораковой вакцины на мышиной модели рака молочной железы. Для этой модели используют мышей BALB/c (гаплотип H-2d) и нативную линию опухолевых клеток млекопитающих 4Т1, которая представляет собой устойчивый к тиогуанину вариант, образованный из клеток 410.4 без мутагенной обработки. Важно отметить, что эти мышиные клетки молочной железы экспрессируют полноразмерную молекулу мышиного BORIS, как было продемонстрировано методом RT-PCR. Следовательно, имеется идеальная модель для изучения способности молекулы BORIS применяться в качестве иммунной вакцины против рака.
Проводят два различных типа экспериментов. Первый тип экспериментов включает группу мышей, вакцинированных pBORIS (плазмидой, кодирующей делегированную молекулу мышиного BORIS), смешанный с ДНК, кодирующей различные мышиные цитокины (pGM-CSF; pIL12/IL18; pIFNγ) в качестве молекулярных адъювантов. Мышей инъецируют вектором (pORF) или pIL12/IL18 в качестве контроля. Мышей иммунизируют и проводят бустерную иммунизацию с помощью генного пистолета, а затем проводят контрольное заражение с помощью 104 или 105 клеток 4Т1.
Второй тип эксперимента включает группу мышей, вакцинированных с помощью pBORIS, буферную иммунизацию которых проводят дефектным по репликации аденовирусным вектором (Ad5), который модифицирован таким образом, чтобы экспрессировать ZF-делетированную молекулу мышиного BORIS (Ad5-BORIS). В качестве контрольной используют группу мышей, инъецированных вектором с последующей бустерной иммунизацией с помощью Ad5. Контрольное заражение животных проводят с помощью 104 или 105 клеток 4Т1 и анализируют появление и рост опухоли. Заметим, ранее было найдено, что инъекция такого малого количества как 104 клеток 4Т1.2 в молочные железы мышей BALB/c приводит к локальному росту опухолей молочной железы у 100% животных с контрольным заражением.
Вакцинация с помощью pBORIS плюс pIL12/IL18 или pBORIS с последующим Ad5-BORIS приводит к защите мышей от контрольного заражения с помощью 104 немодифицированных 4Т1 опухолевых клеток. Хотя у 50% мышей из группы, иммунизированной с помощью pBORIS в смеси с pIL12/IL18, образуются небольшие опухоли (0.2-0.4 см3), они все жизнеспособны на день 39. Все подопытные мыши умерли, примерно, десятью днями ранее.
Результаты, полученные на мышах, иммунизированных с помощью Ad5-BORIS, являются выдающимися. На день 24, когда мыши в контрольной группе умерли от опухоли, 100% мышей, иммунизированных вакциной Ad5-BORIS, были не только живы, но у них совсем не образовалась опухолей. На самом деле у них не образуется опухоль вплоть до дня 33 после контрольного заражения. Эти результаты показывают, что вакцина ZF-делетированного BORIS эффективно защищает при введении 104 опухолевых клеток молочной железы.
Второй ряд экспериментов проводят в более жестких условиях и с контрольным заражением мышей 105 4Т1 опухолевых клеток. Вакцинация плазмидой pBORIS плюс pIFNy или pIL12/IL18 значительно продлевает время роста опухоли до объема 2 см3 и повышает выживаемость мышей BALB/c. Вакцинация также снижает скорость роста опухоли при контрольном заражении с помощью 105 опухолевых клеток 4Т1. Более глубокий эффект обнаруживают у мышей, вакцинированных с помощью pBORIS с последующей бустерной иммунизацией вакциной Ad5-BORIS перед заражением с помощью 105 немодифицированных клеток 4Т1. В этом случае на день 23, когда все мыши в контрольной группе умерли в результате роста опухоли, 80% мышей, иммунизированных вакциной Ad5-BORIS, остаются живыми и у выживших мышей опухоли значительно меньше по размеру.
Отдельные группы мышей BALB/c иммунизируют делегированным мышиным белком BORIS, очищенным от Е. coli системы. В данном случае пяти мышам подкожно инъецируют белок (50 мкг/мышь), смешанный с обычным адъювантом типа Qui1 A Th1 (Sigma). После четырех иммунизации у всех животных наблюдается появление заметного титра антител против BORIS. Другую группу из 5 мышей одновременно иммунизируют интраперитонеально выделенными дендритными клетками, инфицированными вакциной Ad5-BORIS. После трех инъекций у мышей вырабатывается Т-клеточный ответ против mBORIS, который обнаруживается in vitro в культуре спленоцитов, активированных белком mBORIS. Следовательно, иммунизация с помощью BORIS индуцирует В- и Т-клеточный иммунный ответ у мышей и этот иммунный ответ предохраняет животное от (контрольного) заражения.
6. Усеченный BORIS, связанный с PTD, как субъединичная вакцина
PTD связывается с нефункциональным усеченным или мутантным белком BORIS, и продукт слияния образует дрожжевую экспрессирующую систему. Гены, кодирующие PTD и нефункциональный мутантный BORIS, субклонируют в дрожжевой экспрессирующий вектор, такой как pGAPZα. Экспрессированный и секретированный белок очищают стандартными методами молекулярной генетики. Мышей иммунизируют антигеном, приготовленным в двух различных обычных адъювантах, и анализируют иммунный ответ, а также защиту от опухолевого антигена.
7. Вирусоподобные частицы, кодирующие нефункциональный мутантный BORIS, как субъединичная вакцина
Получают субъединичную вакцину VLP-BORIS на основе корового антигена (HBcAg) вируса гепатита В (HBV). Этот антиген "самособирается" в VLP после экспрессии в клетках дрожжей. Чужеродные последовательности можно встраивать в некоторые области HBcAg, не нарушая процесс сборки. Соответственно, получают частицы химерного HBcAg-BORIS, которые используют для иммунизации мышей.
8. Анализ на мышах BALB/c и р53 КО
Анализируют иммунный ответ у мышей BALB/c без заражения и молодых мышей с нокаутом гена р53, у которых к этому возрасту не появляется опухоль. Определяют как гуморальный, так и клеточный иммунный ответ у мышей, иммунизированных различными вакцинами BORIS. Анализируют сыворотки иммунизированных мышей с целью обнаружения продукции антитела против BORIS в течение 3х-месячного эксперимента. Определяют пролиферацию CD4+ и CD8+ Т клеток и активацию регуляторных Т клеток до и после заражения мышей BALB/c. Одновременно анализируют активацию NK клеток, которые могут непосредственно вызывать киллинг опухолевых клеток молочной железы. Показана функциональная активность BORIS-специфических цитотоксических Т-лимфоцитов (CTL) до и после заражения опухолевыми клетками молочной железы 4Т1. Для обнаружения NK и CTL активности в качестве клеток-мишеней используют опухолевые клетки Р815, которые естественно (в природе) экспрессируют молекулы BORIS дикого типа, наряду с 4Т1 клетками.
Дополнительные ссылки
Filippova, G.N. et al. Tumor-associated Zinc Finger Mutations in the CTCF Transcription Factor Selectively Alter Its DNA- binding Specificity. Cancer Research 62: 48-52 (2002).
Kim, J.J. et al. In vivo engineering of a cellular immune response by co-administration of IL-12 expression vector with a DNA immunogen. J. Immunol 158: 816-826 (1997).
Kim, J.J. et al. CD8 positive Т cells influence antigen-specific immune responses through the expression of chemokines. Journal of Clinical Investigation 102: 1112-24 (1998).
Kim, J.J. et al. Modulation of amplitude and direction of n vivo immune responses by co-administration of cytokine gene expression cassettes with DNA immunogens. European Journal of Immunology 28: 1089-1103. (1998).
Kim, J.J. et al. Intracellular adhesion molecule-1 modulates beta-chemokines and directly costimulates Т cells in vivo. Journal of Clinical Investigation 103: 869-77 (1999).
Kim, J.J. et al. Macrophage Colony-Stimulating Factor Can Modulate Immune Responses and Attract Dendritic Cells in Vivo. Human Gene Therapy 11: 305-321 (2000).
Lutz, M.В. et al. An advanced culture method for generating large quantities of highly pure dendritic cells from mouse bone marrow. Immunol. Meth. 223: 77-92 (1999).
Nardelli, В. & Tarn, J. P. The MAP system. A flexible and unambiguous vaccine design of branched peptides. Pharm. Biotechnol. 6: 803-819 (1995).
Resko, J.E. J. et al. Cell Growth Inhibition by the Multivalent Transcription Factor CTCF. Cancer Research 61: 6002-7 (2001).
Ribas, A., Butterfield, L.H., Glaspy, J.A. & Economou, J.S. Cancer Immunotherapy Using Gene- modified Dendritic Cells. Curr. Gene Ther 2: 57-78 (2002).
Ross, R.M., Xu, Y., Bright, R.A. & Robinson, H.L. C3d enhancement of antibodies to hemagglutinin accelerates protection against influenza virus challenge. Nat. Immunol. 1, 127-131 (2000).
Smith, M., Burchell, J. M., Graham, R., E.P., C. & J., T.-P. Expression of B7.1 in a MUC1-expressing mouse mammary epithelial tumor cell line inhibits tumorigenicity but does not induce autoimmunity in MUC1 transgenic mice. Immunol. 97, 648-655 (1999).
Рядовому специалисту в данной области техники очевидно, что можно использовать различные модификации материалов и методов при применении вышеописанного изобретения. Такие модификации можно рассматривать как входящие в объем настоящего изобретения, определяемый нижеприведенной формулой изобретения.
Описание последовательности: нуклеотидная последовательность молекулы BORIS с ZF-делецией

Описание последовательности: нуклеотидная последовательность мышиного BORIS дикого типа

Описание последовательности: нуклеотидная последовательность человеческого BORIS дикого типа


Описание последовательности: аминокислотная последовательность человеческого BORIS

Описание последовательности: аминокислотная последовательность мышиного BORIS

Описание последовательности: аминокислотная последовательность мышиного BORIS с ZF-делецией

| название | год | авторы | номер документа |
|---|---|---|---|
| ПРОФИЛАКТИЧЕСКАЯ ПРОТИВОРАКОВАЯ ВАКЦИНА | 2004 |
|
RU2385163C2 |
| КОМПОЗИЦИЯ ДНК ДЛЯ ВЫЗОВА ИММУННОГО ОТВЕТА ПРОТИВ ОПУХОЛЕАССОЦИИРОВАННЫХ МАКРОФАГОВ | 2007 |
|
RU2459631C2 |
| ПРОТИВОРАКОВЫЕ ВАКЦИНЫ, НАЦЕЛЕННЫЕ НА BORIS, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2799786C2 |
| ПРОТИВОРАКОВЫЕ ВАКЦИНЫ, НАЦЕЛЕННЫЕ НА BORIS, И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2759681C1 |
| СОЧЕТАНИЕ РЕКОМБИНАНТНОЙ МИКРОБАКТЕРИИ И БИОЛОГИЧЕСКИ АКТИВНОГО СРЕДСТВА В КАЧЕСТВЕ ВАКЦИНЫ | 2005 |
|
RU2495677C2 |
| ИНДИВИДУАЛИЗИРОВАННЫЕ ПРОТИВООПУХОЛЕВЫЕ ВАКЦИНЫ | 2012 |
|
RU2670745C9 |
| ИНДИВИДУАЛИЗИРОВАННЫЕ ПРОТИВООПУХОЛЕВЫЕ ВАКЦИНЫ | 2012 |
|
RU2779946C2 |
| СЛИТЫЙ БЕЛОК ТЕЛОМЕРАЗНОЙ ОБРАТНОЙ ТРАНСКРИПТАЗЫ, КОДИРУЮЩИЕ ЕГО НУКЛЕОТИДЫ И ИХ ПРИМЕНЕНИЕ | 2007 |
|
RU2473691C2 |
| ПРОСТАТОАССОЦИИРОВАННЫЕ АНТИГЕНЫ И ИММУНОТЕРАПЕВТИЧЕСКИЕ СХЕМЫ НА ОСНОВЕ ВАКЦИН | 2013 |
|
RU2609651C2 |
| ПРОСТАТОАССОЦИИРОВАННЫЕ АНТИГЕНЫ И ИММУНОТЕРАПЕВТИЧЕСКИЕ СХЕМЫ НА ОСНОВЕ ВАКЦИН | 2013 |
|
RU2737765C2 |















Настоящее изобретение относится к биотехнологии и представляет собой иммуногенную композицию для предупреждения и лечения раковых заболеваний, которая содержит нефункциональный BORIS белок, в последовательности которого отсутствуют все «цинковые пальцы». Настоящее изобретение также раскрывает иммунотерапевтическую композицию против рака, содержащую указанный нефункциональный BORIS белок или клетку бактерии, млекопитающего или дрожжей или вирусную частицу, способную к экспрессии указанного нефункционального BORIS белка. Настоящее изобретение раскрывает также способ иммунизации пациента путём введения эффективного количества указанной иммунотерапевтической композиции, а также применение указанной иммунотерапевтической композиции для приготовления вакцины против рака. Изобретение позволяет повысить эффективность профилактической и терапевтической вакцины против рака. 4 н. и 18 з.п. ф-лы, 7 ил., 2 табл., 8 пр.
1. Иммуногенная композиция для предупреждения и лечения раковых заболеваний, содержащая нефункциональный BORIS белок, и фармацевтически приемлемый носитель, наполнитель или адъювант, где нефункциональный BORIS белок содержит SEQ ID NO:2, в которой отсутствуют все цинковые пальцы.
2. Композиция по п. 1, в которой нефункциональный мутантный BORIS не связывается с ДНК.
3. Композиция по п. 1, в которой нефункциональный BORIS белок присоединен к фармацевтически приемлемому носителю.
4. Композиция по п. 1, в которой BORIS белок связан с доменом белковой трансдукции (PTD).
5. Композиция по п. 1, в которой нефункциональный BORIS белок связан с пептидом, который модифицирует BORIS и сохраняет или повышает антигенность указанного нефункционального BORIS белка
6. Композиция по п. 1, где нефункциональный BORIS белок экспрессируется в бактериальной клетке, клетке млекопитающего (включая дендритную клетку), клетке дрожжей или в вирусных частицах.
7. Композиция по п. 1, где адъювант является обычным или молекулярным и усиливает ее иммуногенность.
8. Композиция по п. 7, в которой адъювант или молекулярный адъювант смешивается, сшивается или объединяется с указанным нефункциональным BORIS белком.
9. Композиция по п. 7, в которой адъювантом могут быть молекулы из группы цитокинов, хемокинов или костимулирующих молекул.
10. Композиция по п. 1, где нефункциональный BORIS белок экспрессируется дендритной клеткой млекопитающего.
11. Иммунотерапевтическая композиция против рака, содержащая нефункциональный BORIS белок или бактериальную клетку, клетку млекопитающего (включая дендритную клетку), клетку дрожжей или вирусную частицу, экспрессирующую нефункциональную форму BORIS белка, содержащего SEQ ID NO:2, в которой отсутствуют все цинковые пальцы, и дополнительно содержащая адъювант, молекулярный адъювант, фармацевтически приемлемое вещество, усиливающее антиопухолевый иммунный ответ.
12. Способ иммунизации пациента, заключающийся во введении ему эффективного количества иммунотерапевтической композиции по п. 11, который повышает иммунный ответ к опухолям.
13. Способ по п. 12, в котором эффективное количество иммунотерапевтической композиции содержит молекулярный адъювант из группы цитокинов, хемокинов или костимулирующих молекул.
14. Способ по п. 12, в котором эффективное количество иммунотерапевтической композиции содержит фармацевтически приемлемый носитель.
15. Способ по п. 12, в котором эффективное количество иммунотерапевтической композиции содержит нефункциональный BORIS белок, связанный с пептидом, который модифицирует BORIS и сохраняет или повышает его антигенное и противоопухолевое свойства.
16. Способ по п. 12, в котором эффективное количество иммунотерапевтической композиции содержит нефункциональный BORIS белок, который дополнительно содержит домен белковой трансдукции (PTD).
17. Способ по п. 12, в котором введение осуществляют внутримышечно, подкожно, интрадермально, внутривенно, назально, ректально, вагинально или перитонеально.
18. Способ по п. 12, в котором пациент болен более чем одним типом рака.
19. Способ по п. 12, в котором у пациента рак.
20. Способ по п. 12, в котором у пациента есть генетическая предрасположенность к раку.
21. Способ по п. 12, в котором иммунизация приводит к выработанному клеточному иммунному ответу, включающему Т клетки, которые распознают эпитоп нефункционального BORIS белка.
22. Применение иммунотерапевтической композиции по п. 11 для приготовления вакцины против рака, при котором иммунотерапевтическая композиция содержит эффективное количество нефункционального BORIS белка, содержащего SEQ ID NO:2, в которой отсутствуют все цинковые пальцы.
| НАСОС ДЛЯ ПЕРЕКАЧИВАНИЯ ВЛАЖНОЙ ВОЛОКНИСТОЙ МАССЫ | 1995 |
|
RU2083879C1 |
| US 5156841 A, 20.10.1992 | |||
| LOUKINOV DMITRI I., et al., BORIS, A novel male germ-line-spercific protein associated with epigenetic reprogramming events, shares the same 11-zinc-finger domain with CTCF, the insulator protein involved in reading imprinting marks in the soma, PNAS, Vol.99, No.10, 14.05.2002, pp.6806-6811. | |||

