Изобретение относится к области физиологии растений, в частности биотехнологии и сельскому хозяйству, а именно к способам регуляции цветения растений, и может быть использовано в кормопроизводстве, технологиях ремедиации загрязненных водоемов, мелиорации заболоченных территорий и цветоводстве для массового производства семенного материала.
Известен способ регуляции цветения растений температурным режимом. Выдержка растений в условиях низких положительных температур в течение определенного времени (вернализация) приводит к цветению при переводе в нормальные температурные условия (Физиология растений: Учебник для студ. ВУЗов / Н.Д.Алехина. Ю.В.Балнокин, В.Ф.Гавриленко и др. Под ред. И.П.Ермакова. - М.: Издательский центр «Академия», 2005. - 640 с.)
Известны способы регуляции цветения условиями питания. Если в растении много соединений азота, то оно будет расти, а если углеводы преобладают над азотом, то растение цветет.Выращивание растений на богатой азотом среде в условиях недостаточного освещения обусловливает вегетативное развитие. На бедной среде в условиях достаточной освещенности в растении баланс смещается в сторону углеводов и растение начинает цвести. Другой вариант реализации такого способа - использование «плодового пояса», затрудняющего отток сахаров из листьев растения и вызывающего его цветение (Физиология растений: Учебник для студ. ВУЗов / Н.Д.Алехина. Ю.В.Балнокин, В.Ф.Гавриленко и др. Под ред. И.П.Ермакова. М.: Издательский центр «Академия», 2005. - 640 с.).
Известен способ регуляции цветения стресс-факторами. Для стимуляции цветения растения некоторое время вообще не поливают или снижают степень увлажнения почвы. Для разных растений может потребоваться период засухи от 1 до 6 месяцев (Физиология растений: Учебник для студ. ВУЗов / Н.Д.Алехина, Ю.В.Балнокин, В.Ф.Еавриленко и др. Под ред. И.П.Ермакова. - М.: Издательский центр «Академия», 2005. - 640 с.).
Известны способы регуляции цветения путем обработки растений раствором действующего вещества - фитогормонами, пектиновыми фрагментами клеточных стенок, липохитоолигосахаридами, олигогалактуроновые кислоты, нитробензолом и др. (Патент РФ 2197816, А01Н 4/00. опубл. 10.02.2003).
Известны способы управления цветением растений, основанные на генетической трансформации растений, изменяющей реакцию растений на внешние факторы, и использовании специальных внешних индукторов цветения (Патент РФ 2241753, C12N 15/82, C12N 15/11, опубл. 10.12.2004).
Наиболее близким к заявленному изобретению, взятым за прототип, является способ управления цветением растений, включающий изменение режима освещения растений, в котором переход из вегетативной фазы к репродуктивной вызывается заданием определенного соотношения темновой и световой фазы, последовательности изменения этого соотношения или увеличением интенсивности освещения в световой фазе, компенсирующем недостаточную длину дня (Thomas В.Light signals and flowering. J.Exp. Botany, 2006. Vol.57. №13. P.3387-3393). Этот способ основан на актиноритмизме растений (Мошков Б.С. Актинометризм растений. М.: Агропромиздат, 1987. - 272 с.), т.е. способности растений качественно и количественно оценивать режим освещения. Реакция растений на соотношение световой и темновой фазы и его (соотношения) динамику называется фотониктопериодизмом (Чайлахян М.Х. Регуляция цветения высших растений. М.: Наука, 1988. - 560 с.).
Недостатком всех известных способов, включая прототип, является то, что при таких способах воздействия на водный гиацинт (Eichhornia crassipes) может наблюдаться лишь спорадическое зацветание одиночных растений.
Задачей настоящего изобретения является разработка способа перевода растения водного гиацинта (Eichhornia crassipes) из вегетативной фазы в репродуктивную с целью повышения синхронности и массовости зацветания растений водного гиацинта в заранее известные сроки, в том числе для использования в технологии производства его семян.
Поставленная задача решается тем, что способ перевода растения водного гиацинта (Eichhornia crassipes) из вегетативной фазы в репродуктивную включает изменение режима освещения растений, но в отличие от прототипа для растений гиацинта, выращенных со сменой темновой и световой фазы и освещением от 7 до 120 тыс.лк, в период световой фазы устанавливают уровень освещенности в последовательности, высокий (4-120 тыс.лк) - низкий (7-3000 лк) - высокий (4-120 тыс.лк), причем уровень освещенности меняют однократно или ежесуточно, а для растений, выращенных при непрерывном освещении интенсивностью 7-15 тыс.лк, уровень освещенности снижают однократно или ежесуточно до 600-2000 лк на 3-5 часов или до 7-20 лк на 10-12 часов. Для разового зацветания растений уровень освещенности в течение световой фазы можно менять однократно в последовательности: высокий - низкий - высокий, а затем следуют дни с высоким уровнем освещенности.
При непрерывном освещении для разового зацветания уровень освещенности снижают однократно.
Предлагаемый способ осуществляется следующим образом.
Растения водного гиацинта растут на 10-24-часовом световом дне при постоянной освещенности от 7 до 120 тыс.лк и температуре 18-25°С в течение 2-3 месяцев. Уровень освещенности в лабораторных условиях задается мощностью, количеством ламп и расстоянием от ламп до растений, на открытом воздухе, под солнцем, наличием или отсутствием тени. В лабораторных условиях опытах использовались светильники с натриевыми лампами ДНаТ-125 и люминесцентные лампы низкого давления ЛБ-40. Длительность световой фазы задается таймером. Освещенность определяется люксметром. При изменении мощности постоянного светового потока, падающего на растения, соотношения световой и темновой фаз. последовательности изменения (увеличения или уменьшения) соотношения световой и темновой фаз, временного охлаждения растений до 5-15°С - все растения оставались в вегетативной фазе.
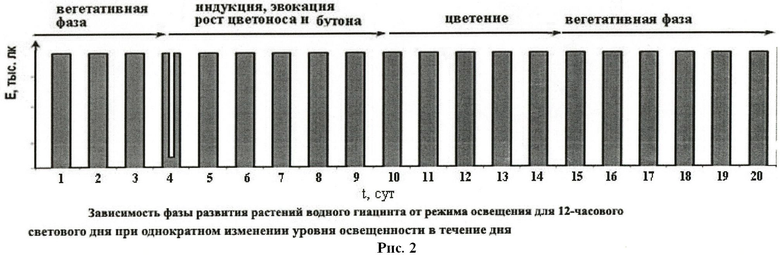
В опытах с наличием темновой фазы для перехода растений к цветению после 1/3 длительности световой фазы интенсивность освещения снижают до 600-2000 лк путем отключения части ламп, увеличения расстояния от ламп до растений или укрыванием растений полупрозрачным материалом на время, равное 1/3 длительности светового периода, а затем восстанавливают прежний уровень освещенности. Процедуру выполняют ежесуточно или однократно (см. рис.1 и рис.2 для 12-часового светового дня).
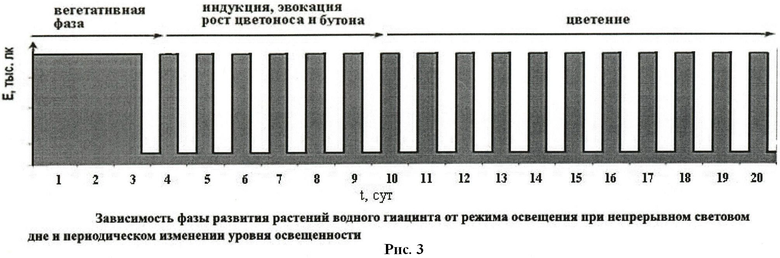
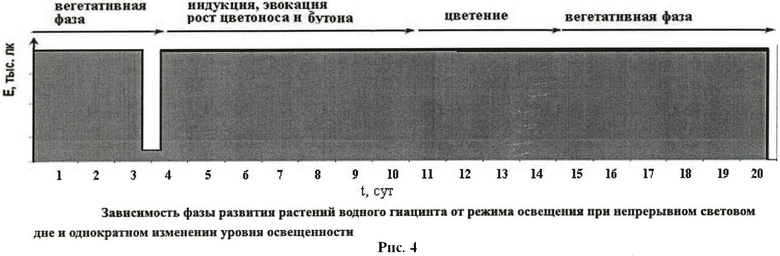
При непрерывном (24-часовом) освещении интенсивностью 7-15 тыс.лк для перехода растений к цветению возможны два варианта воздействия. В первом интенсивность освещения снижают до 600-2000 лк на время 3-5 часов однократно или ежесуточно. Другой вариант - снижение интенсивности освещения до 5-20 лк на 10-12 часов однократно или периодически раз в сутки (см. рис 3 и 4). Примеры конкретного применения изобретения приведены ниже.
Пример 1. Растения водного гиацинта (Eichhornia crassipes), выращенные на 12-часовом световом дне при освещенности 15-120 тыс.лк, переводят на следующий режим освещения: 4 часа - 15-120 тыс.лк, 4 часа - 1-2 тыс.лк, 4 часа 15-20 тыс.лк. Через 5-8 суток растения зацветают.
Пример 2. Растения водного гиацинта, выращенные на 24-часовом световом дне (непрерывное освещение) при освещенности 7-10 тыс.лк, переводят на следующий режим освещения: 4 часа - 0.6-1,5 тыс.лк, 20 часов - 7-10 тыс.лк. Через 7-10 суток растения зацветают.
Пример 3. Растения водного гиацинта, выращенные на 14-часовом дне при освещенности 7-10 тыс.лк, переводят на следующий режим освещения: 14 часов - 7-10 тыс.лк, 10 часов - 5-20 лк. Через 9-12 суток все растения зацветают.
Пример 4. Растения водного гиацинта, выращенные на 14-часовом дне при освещенности 7-10 тыс.лк, переводят на одни сутки в режим освещения: 5 часов - 7-10 тыс.лк, 4 часа, 1-2 тыс.люкс, 5 часов - 7-10 тыс.лк, а в последующий период на 14-часовой световой день при освещенности 7-10 тыс.лк. Через 6-8 суток растения зацветают.
Пример 5. Растения водного гиацинта, выращенные на 16-17-часовом дне при освещенности 50-120 тыс.лк переводят на одни сутки в режим освещения: 7 часов - 50-120 тыс.лк, 4 часа - 3 тыс.лк, 5 часов - 50 тыс.лк. Через 6-8 суток растения зацветают.
Пример 6. Растения водного гиацинта, выращенные на 16-17-часовом дне при освещенности 7-9 тыс.лк, переводят на одни сутки в режим освещения: 5 часов - 4-5 тыс.лк, 4 часа - 0.6-1.5 тыс.лк. 5 часов - 4-5 тыс.лк, а в последующий период на 14-часовой световой день при освещенности 4-5 тыс.лк. Через 6-8 суток растения зацветают.
Таким образом, в результате разработки заявляемого изобретения выявлен новый тип внешнего фактора, регулирующего переход растения из вегетативной фазы в репродуктивную, и новый тип реакции растений на свет как сигнальный фактор - фотосциопериодизм. Растения отвечают постоянным цветением на периодически повторяющиеся изменения интенсивности освещения в порядке темнота - высокий - низкий - высокий или чередование высокого и низкого уровня интенсивности в условиях непрерывного освещения. Как и в известной форме актиноритмизма фотониктопериодизме. в фотосциопериодизме обнаружен эффект последействия. Достаточно одних или нескольких суток с переменной интенсивностью освещения, чтобы потом растения зацвели на освещении с постоянной интенсивностью (фотосциопериодическая индукция). Для фотосциопериодической индукции обязательно наличие периода низкого уровня освещенности между периодами интенсивного освещения.
Переход из репродуктивной фазы в вегетативную осуществляется длительным (2-3 недели) освещением с постоянной интенсивностью в течение световой фазы.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ АДАПТАЦИИ РАСТЕНИЙ-РЕГЕНЕРАНТОВ К УСЛОВИЯМ EX VITRO | 2023 |
|
RU2828836C1 |
| Способ выращивания костреца безостого в защищенном грунте | 1986 |
|
SU1416086A1 |
| УСТАНОВКА ДЛЯ ПЕРЕРАБОТКИ И ИСПОЛЬЗОВАНИЯ ЖИДКИХ ОТХОДОВ ЖИВОТНОГО ПРОИСХОЖДЕНИЯ, ВКЛЮЧАЯ МЕТАНИЗАЦИЮ, КУЛЬТИВИРОВАНИЕ МИКРОСКОПИЧЕСКИХ ВОДОРОСЛЕЙ И МАКРОФИТОВ И ВЕРМИКУЛЬТИВИРОВАНИЕ | 2014 |
|
RU2684594C2 |
| СПОСОБ ПОЛУЧЕНИЯ ГИБРИДА РАСТЕНИЯ ПОДСОЛНЕЧНИКА | 2005 |
|
RU2302105C1 |
| Способ выращивания озимых злаков | 1984 |
|
SU1537202A1 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ОКОНЧАНИЯ ЯРОВИЗАЦИИ МНОГОЛЕТНИХ КОРМОВЫХ ТРАВ | 1999 |
|
RU2176444C2 |
| Способ выращивания картофеля | 1981 |
|
SU1015868A1 |
| ПОСЛЕДОВАТЕЛЬНОСТИ НУКЛЕИНОВЫХ КИСЛОТ И ПЕПТИДЫ/БЕЛКИ СЕМЕЙСТВА FT, ОБУСЛОВЛИВАЮЩИЕ СВОЙСТВА ПОДАВЛЕНИЯ ЦВЕТЕНИЯ У ТРАНСФОРМИРОВАННЫХ ИМИ РАСТЕНИЙ ТАБАКА И ТРАНСГЕННЫХ РАСТЕНИЙ | 2013 |
|
RU2644685C2 |
| СИСТЕМА С РЕГУЛИРУЕМОЙ СРЕДОЙ И СПОСОБ БЫСТРОГО РАЗВЕДЕНИЯ СЕМЕННОГО КАРТОФЕЛЯ | 2006 |
|
RU2411715C2 |
| СПОСОБ ИНТЕНСИВНОГО ВЫРАЩИВАНИЯ КАРТОФЕЛЯ ПРИ КАПЕЛЬНОМ ОРОШЕНИИ | 2011 |
|
RU2462023C1 |

Изобретение относится к области физиологии растений, в частности биотехнологии. Способ перевода растения водного гиацинта (Eichhornia crassipes) из вегетативной фазы в репродуктивную включает изменение режима освещения растений, отличающийся тем, что для растений гиацинта, выращенных со сменой темновой и световой фазы и освещением от 7 до 120 тыс. лк, в период световой фазы устанавливают уровень освещенности в последовательности: высокий (4-120 тыс.лк) - низкий (7-3000 лк) - высокий (4-120 тыс.лк), причем уровень освещенности меняют однократно или ежесуточно, а для растений, выращенных при непрерывном освещении интенсивностью 7-15 тыс. лк, уровень освещенности снижают однократно или ежесуточно до 600-2000 лк на 3-5 часов или до 7-20 лк на 10-20 часов. Изобретение позволяет повысить синхронность и массовость зацветания растений водного гиацинта в заранее известные сроки. 4 ил., 6 пр.
Способ перевода растения водного гиацинта (Eichhornia crassipes) из вегетативной фазы в репродуктивную, включающий изменение режима освещения растений, отличающийся тем, что для растений гиацинта, выращенных со сменой темновой и световой фазы и освещением от 7 до 120 тыс лк, в период световой фазы устанавливают уровень освещенности в последовательности высокий (4-120 тыс.лк) - низкий (7-3000 лк) - высокий (4-120 тыс.лк), причем уровень освещенности меняют однократно или ежесуточно, а для растений, выращенных при непрерывном освещении интенсивностью 7-15 тыс лк, уровень освещенности снижают однократно или ежесуточно до 600-2000 лк на 3-5 ч или до 7-20 лк на 10-20 ч.
| СПОСОБ РЕГУЛЯЦИИ ЦВЕТЕНИЯ РАСТЕНИЙ IN VIVO | 2000 |
|
RU2197816C2 |
| ТАРАКАНОВ И.Г | |||
| Фоторегуляция в адаптивных стратегиях овощных растений, Автореферат, 2007 | |||
| KING RW, Regulation of flowering in the long-day grass Lolium temulentum by gibberellins and the FLOWERING LOCUC T gene, Plant Physiol, 2006, Jun, 141(2), 498-507 | |||
| ADAMS S, Interaction between the light quality and flowering time pathways in arabidopsis, Plant J, 2009, Oct, 60(2), 257-267. | |||

