Изобретение относится к области физиологии растений, в частности биотехнологии, а именно к способам регуляции цветения растений с помощью олигогалактуроновых кислот, являющихся олигомерными производными пектина клеточных стенок, содержащими от 5 до 8 сахаридных радикалов, связанных друг с другом гликозидными связями.
Известен способ регуляции цветения с помощью воздействия разнообразных физических и биохимических факторов, например фотопериода, низкой температуры, фитогормонов, в частности гиббереллинов (Бернье Ж., Кине Ж-М, Сакс Р. Физиология цветения, Т. 1,2. М.: Агропромиздат, 1985).
Кроме того, известно, что длиннодневные розеточные растения (белена, рудбекия и др.), растущие на неблагоприятной для зацветания длине дня, образуют цветоносный стебель и зацветают после обработки почек гибберелловой кислотой в концентрации 100 мг/л (Миляева Э.Л., Чайлахян М.Х. Изменения в стеблевых апексах при переходе от вегетативного роста к цветению. Известия АН СССР. Серия биология. 1974, 3, с.342-352).
Скорость зацветания зависит от концентрации гиббереллина, нанесенного на почки, от продолжительности обработок, а также от способа воздействия.
Однако в тех случаях, когда в качестве регулятора цветения использовали синтетический гиббереллин, такие обработки экологически небезопасны; в случаях использования природных гиббереллинов, получаемых обычно из богатых ими грибов, такие обработки, проводимые в промышленных масштабах, будут чрезвычайно дороги.
Известен также способ регуляции цветения пектиновыми фрагментами клеточных стенок в культуре in vitro (Tran Thanh Van, Cousson A.G., Gollin D.J. , Chelf P., Albersheim P. Nature, 1985, v.314, p.615-617).
В этих работах тонкие эпидермальные слои клеток листьев или стеблей табаков (Nicotiana tabacum) культивировали на питательной среде Мурасиге и Скуга. После добавления в среду культивирования олигогалактуронидов соотношение образующихся в культуре цветочных и вегетативных почек увеличивается в пользу цветочных.
Однако описанный способ воздействия проведен на фрагментах растительных тканей (тонких слоях эпидермиса), выращенных на искусственной питательной среде в стерильных условиях культуры in vitro. Эти условия существенно отличаются от условий культивирования целых растений in vivo в почве.
Задача исследования состоит в том, чтобы с помощью природных недорогих легко доступных веществ, являющихся производными стенок растительных клеток, регулировать цветение растении, выращиваемых в почве в природных условиях.
Эта задача была решена созданием нового способа регуляции цветения растений in vivo, заключающегося в обработке растений раствором действующего вещества, причем в качестве действующего вещества используют олигогалактуроновые кислоты со степенью полимеризации n=5-8, в концентрации (5-50)•10-7 моль/л, а обработку растений проводят в течение 5-10 дней.
Сущность изобретения состоит в том, что разработан совершенно новый способ регуляции цветения растений, позволяющий с помощью веществ, являющихся олигомерными производными пектинов, присутствующих в клеточной стенке растений, получить зацветание фотопериодически чувствительных растений на неблагоприятной для зацветания длине дня. Обнаружено, что заявляемое вещество - новый регулятор цветения участвует в запуске морфогенетической программы цветения растений.
Изобретение иллюстрируется следующими материалами:
На фиг. 1-4 представлена диаграмма, показывающая зависимость количества зацветающих растений (в %) от количества дней обработки при различной концентрации обрабатывающих растворов.
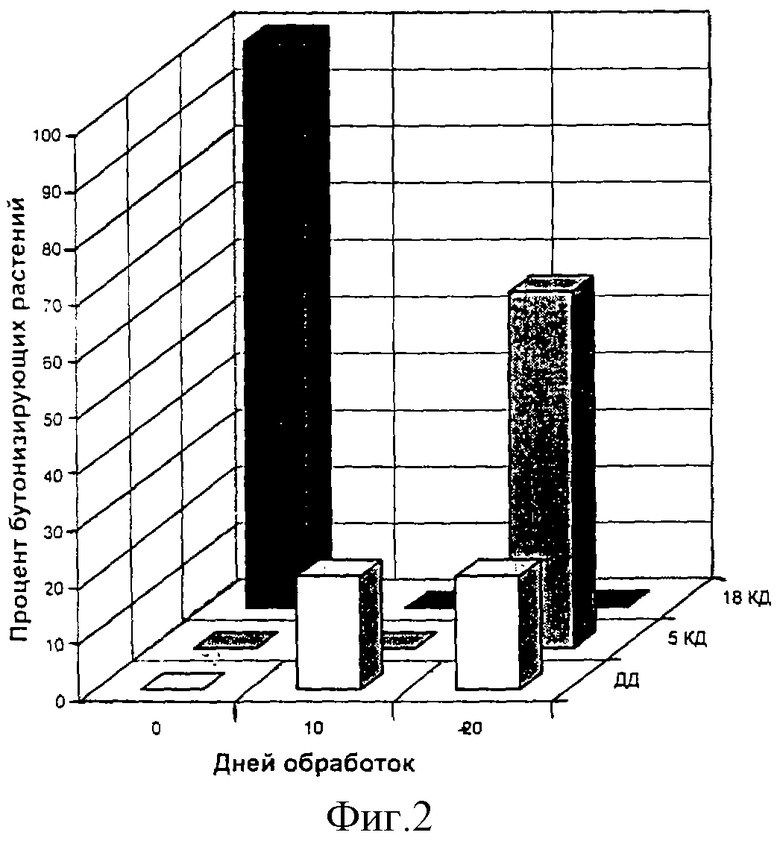
Пример 1. Растения рудбекии, выращенные на неиндуктивном коротком дне (КД) и находящиеся в розеточном состоянии, обрабатывают октагалактуроновой кислотой (ОГК) в течение 5 и 10 дней (n=8). После обработки в течение 5 дней раствором концентрации 5•10-7 обнаружено начало стеблевания у 73% обработанных растений (фиг.1, 5 дней обработок, внутренний горизонтальный ряд) после обработки в течение 10 дней - у 51% растений (фиг.1, 10 дней обработок, внутренний горизонтальный ряд). Ни одно не обработанное октагалактуроновой кислотой растение не перешло из розеточного состояния к стеблеванию в неиндуктивных условиях короткого дня (фиг.1, 0 дней обработок, внутренний горизонтальный ряд)*.
Примерно такие же результаты были получены после обработки ПГК в концентрации 5•10-6 моль/л (фиг.1, внутренний горизонтальный ряд)
*Стеблевание розеточных длиннодневных растений является показателем их перехода к цветению (Чайлахян М.Х. Регуляция цветения высшых растений. М.: Наука, 1988, с.599).
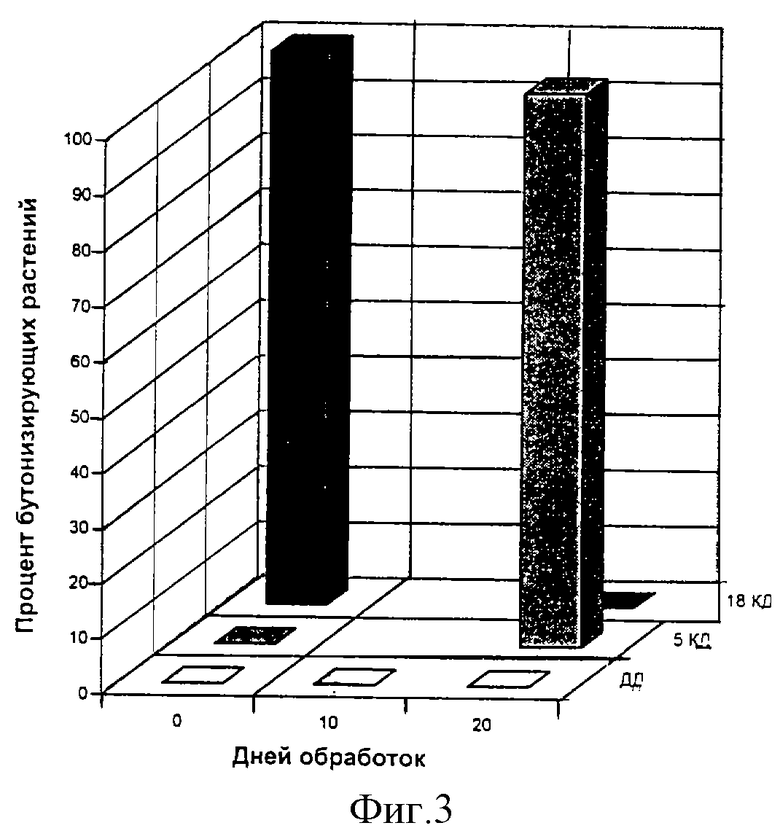
Пример 2. Короткодневные фотопериодически чувствительные растения периллы красной, выращивавшиеся на длинном дне (ДД), обрабатывали пентагалактуроновой кислотой (ПГК) раствором концентрации 25•10-7 в неиндуктивных условиях длинного дня (n=5). 35% растений перешли к бутонизации после 10 и 20 дней обработок, в то время как необработанные растения оставались вегетативными (фиг.2, наружный горизонтальный ряд).
При обработке периллы ПГК в сочетании с недостаточной для цветения фотопериодической индукцией в 5 КД число бутонизирующих растений через 40 дней после начала обработки составило 63% по сравнению с контролем (фиг.1, 5 КД, средний горизонтальный ряд).
После обработок ПГК в сочетании с фотопериодической индукцией в 18 КД переход к цветению оказался заторможенным по сравнению с необработанными растениями. Так, к сороковому дню после начала обработок ни одно обработанное ПГК растение не бутонизировало (фиг.2, 18 КД), в то время как необработанные растения зацвели (фиг.2, 18 КД, внутренний горизонтальный ряд).
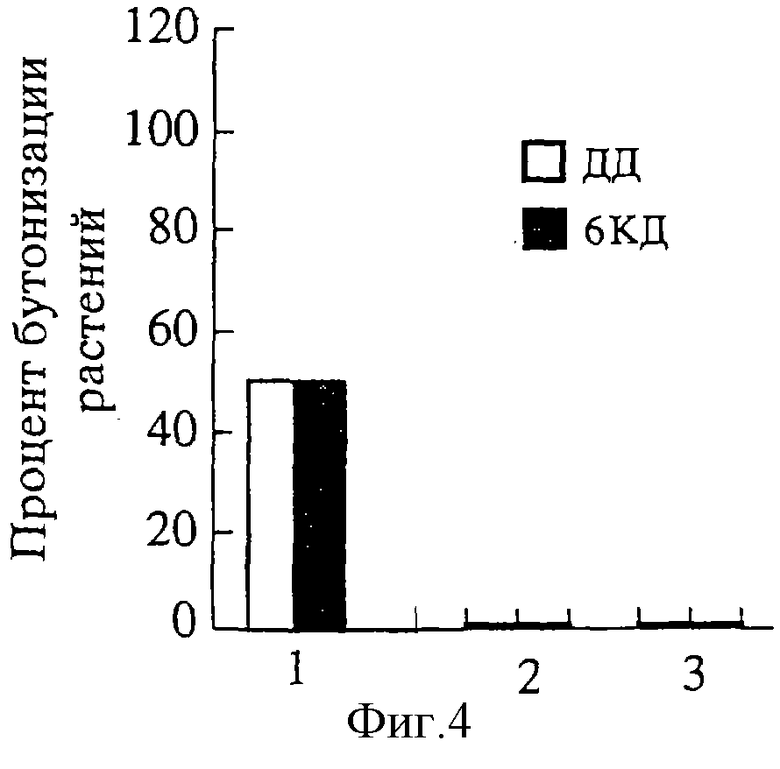
Пример 3. После обработки ОГК в сочетании с 5-дневной обработкой, недостаточной для зацветания периллы, и фотопериодической индукцией в 5 КД бутонизировало 90% растений (фиг.3, 5 КД, средний горизонтальный ряд), а в сочетании с 18-дневной индукцией КД зацветали только необработанные растения (фиг. 3, 18 КД, внутренний горизонтальный ряд). На ДД растения не цвели (наружный горизонтальный ряд).
Пример 4. В опытах с обработками периллы красной, получившей недостаточную для зацветания фотопериодическую индукцию в 6 КД с одновременной обработкой смесью олигогалактуроновых кислот со степенью полимеризации от 2 до 14, через 50 суток после окончания обработки бутонизировали 50% растений (фиг. 4, 1). Обработка дистилированной водой (фиг.4, 2), ацетатным буфером (фиг. 4, 3) в сочетании с индукцией 6 КД не привело к цветению. Однако уменьшенная степень бутонизации по сравнению с примерами 1-3 предполагает сделать вывод о том, что регуляция цветения происходит недостаточно из-за использования смеси олигогалактуроновых кислот со степенью полимеризации 2-14, а не 5-8. Очевидно, оптимальная регуляция цветения происходит при степени полимеризации олигогалактуроновых кислот 5-8.
В результате использования заявляемого изобретения обнаружен новый класс регуляторов роста и развития растений: олигогалактуроновые кислоты, которые выделены из клеточных стенок растений и являются экологически чистыми природными веществами. Заявляемое изобретение дало возможность получить 35-90% растений, перешедших к бутонизации даже для таких фотопериодически чувствительных растений, как рудбеккия и перилла красная, выращенных на неблагоприятной для зацветания длине дня.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ МИКРОРАЗМНОЖЕНИЯ СТЕВИИ STEVIA REBAUDIANA L. | 1993 |
|
RU2092036C1 |
| СПОСОБ ПЕРЕВОДА РАСТЕНИЯ ВОДНОГО ГИАЦИНТА (EICHHORNIA CRASSIPES) ИЗ ВЕГЕТАТИВНОЙ ФАЗЫ В РЕПРОДУКТИВНУЮ | 2011 |
|
RU2460280C1 |
| СПОСОБ ПОДГОТОВКИ РАСТЕНИЙ ЗЕМЛЯНИКИ САДОВОЙ (FRAGARIA L.), РАЗМНОЖЕННЫХ IN VITRO, К УСЛОВИЯМ КУЛЬТИВИРОВАНИЯ EX VITRO | 2006 |
|
RU2302106C1 |
| Способ выделения и очистки гормонов цветения растений короткодневных видов | 1977 |
|
SU1038343A1 |
| СПОСОБ ПОЛУЧЕНИЯ ОБОГАЩЕННОЙ СЕЛЕНОМ БИОМАССЫ СПИРУЛИНЫ (SPIRULINA PLATENSIS) | 2000 |
|
RU2199582C2 |
| СПОСОБ СТИМУЛИРОВАНИЯ РОСТА И РАЗВИТИЯ МАСЛИЧНЫХ КУЛЬТУР | 1996 |
|
RU2121272C1 |
| СПОСОБ ПОВЫШЕНИЯ УРОЖАЙНОСТИ КАРТОФЕЛЯ И ТОМАТОВ | 2003 |
|
RU2243658C1 |
| СПОСОБ ПОВЫШЕНИЯ УСТОЙЧИВОСТИ ОВОЩНЫХ КУЛЬТУР И КАРТОФЕЛЯ К БОЛЕЗНЯМ | 2001 |
|
RU2195823C2 |
| СПОСОБ СТИМУЛИРОВАНИЯ РОСТА И РАЗВИТИЯ МАСЛИЧНЫХ КУЛЬТУР | 1995 |
|
RU2099948C1 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСФОРМИРОВАННЫХ РАСТЕНИЙ КУКУРУЗЫ IN VITRO | 2001 |
|
RU2196421C1 |
Изобретение относится к физиологии растений, а именно к способам регуляции цветения растений. Растения обрабатывают раствором действующего вещества. В качестве действующего вещества используют олигогалактуроновые кислоты со степенью полимеризации n=5-8 в концентрации (5-50)•10-7 моль/л. Обработку растений проводят в течение 5-10 дней. Изобретение позволяет регулировать цветение растений легкодоступными природными веществами. 4 ил.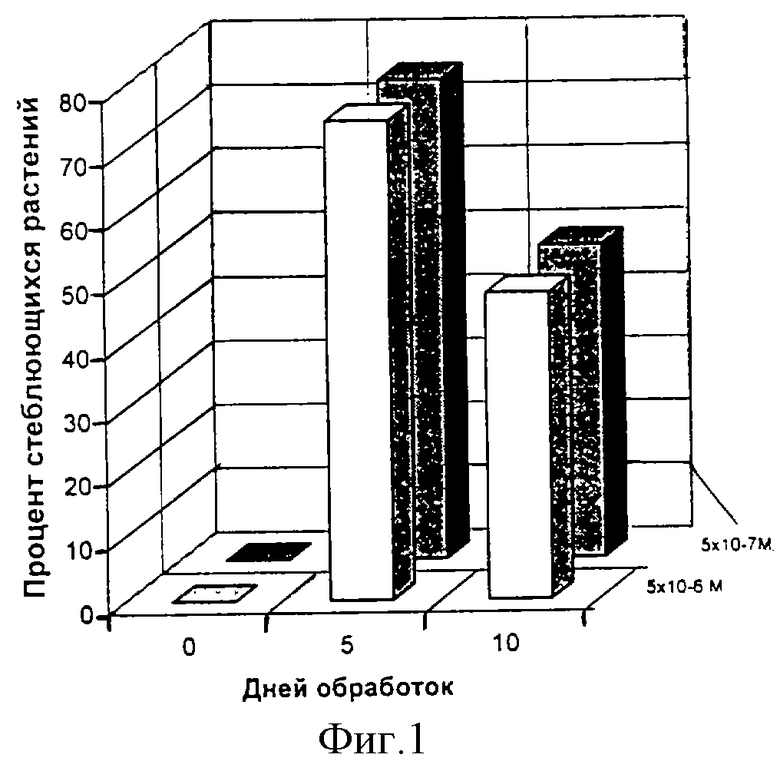
Способ регуляции цветения растений in vivo, заключающийся в обработке растений раствором действующего вещества, отличающийся тем, что в качестве действующего вещества используют олигогалактуроновые кислоты со степенью полимеризации n= 5-8 в концентрации (5-50)•10-7 моль/л, а обработку растений проводят в течение 5-10 дней.
| Kiem tran thanh van and et etc | |||
| Nature, vol 314 | |||
| Горизонтальный водяной двигатель с поворотными лопастями | 1927 |
|
SU6012A1 |
| СПОСОБ СТИМУЛИРОВАНИЯ РОСТА И РАЗВИТИЯ МАСЛИЧНЫХ КУЛЬТУР | 1995 |
|
RU2099948C1 |
| RU 94012735 A1, 20.12.1995 | |||
| 0 |
|
SU398313A1 | |
| GB 2052466 A1, 28.01.1981. | |||
