ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Данное изобретение относится к способу продуцирования полипептида фактора VIII, включающему применение лиганда С2-домена, в частности O-фосфо-L-серина (OPLS).
УРОВЕНЬ ТЕХНИКИ
Классическая гемофилия, или гемофилия А, является наследственным заболеванием крови. Оно является результатом дефицита фактора свертывания крови VIII, связанного с Х-хромосомой, и поражает почти исключительно мужчин с частотой от одного до двух случаев на 10000 человек. Дефект Х-хромосомы передается женщинами-носителями, которые сами не больны гемофилией. Клиническим проявлением гемофилии А является повышенная склонность к кровотечению. До лечения концентратами фактора VIII средняя продолжительность жизни для лиц с тяжелой формой гемофилии составляла менее 20 лет. Применение концентратов фактора VIII из плазмы значительно улучшило ситуацию для пациентов с гемофилией, экстенсивно увеличивая среднюю продолжительность их жизни, давая большинству из них возможность жить более или менее нормальной жизнью. Однако существуют некоторые проблемы, связанные с полученными из плазмы концентратами и их использованием, наиболее серьезной из которых является передача вирусов. До сих пор вирусы, вызывающие СПИД, гепатит В и гепатит ни-А, ни-В, серьезно поражают население. Поэтому в последнее время были разработаны различные способы инактивации вирусов и новые высокоочищенные концентраты фактора VIII, которые установили очень высокий стандарт безопасности и для фактора VIII, полученного из плазмы.
Известно, что фактор VIII (FVIII) экспрессируется в клетках млекопитающих на очень низком уровне. Также известно, что фактор VIII является неустойчивым белком в среде без сыворотки или без белка. Для повышения стабильности и титра фактора VIII применяли добавление различных веществ.
WO 9743436 раскрывает добавление ингибиторов металлозависимых ингибиторов и/или химотрипсинов.
WO 88/08035 и WO 87/04187 раскрывают добавление фосфолипидов к культуральной среде фактора VIII. Также описана коэкспрессия фактора Виллебранда (vWF).
US 20050227913 A1 раскрывает OPLS в качестве ингибитора агрегации фактора VIII путем связывания с С2-доменом (2303-2332). Утверждается, что менее агрегированный фактор VIII является менее иммуногенным.
K.Hansen, M.Kjalke, Р.В.Rasmussen, L.Kongerslev and M.Ezban, Cytotechnol. 24 (3), 227-234, 1997 раскрывает применение бацитрацина А и фосфатидилсерина для предотвращения деградации фактора VIII в среде.
WO 90/02175 A1 раскрывает процессы продуцирования рекомбинантного полипептида(ов) путем культивирования эукариотических клеток в присутствии ингибиторов протеазы для предотвращения деградации полипептида(ов).
ЕР 1707634 A1 раскрывает, что значительное количество фактора VIII ассоциировано с клеточной поверхностью и может быть удалено путем промывания буферами с высокой ионной силой.
В то же время по-прежнему существует необходимость совершенствования способов продуцирования для того, чтобы увеличить общий выход полипептидов фактора VIII и/или уменьшить производственные затраты.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Первый аспект изобретения относится к способу продуцирования полипептида фактора VIII, включающему этапы а) культивирования клеток млекопитающих, экспрессирующих полипептид фактор VIII, в условиях, подходящих для экспрессии указанного полипептида фактора VIII, в том числе включающих клеточную культуральную среду с содержанием лиганда С2-домена, и b) выделения экспрессированного полипептида фактора VIII из клеток млекопитающих с помощью подходящих средств.
Второй аспект изобретения относится к способу продуцирования полипептида фактора VIII, включающему этапы а) культивирования клеток млекопитающих, экспрессирующих полипептид фактор VIII, в условиях, подходящих для экспрессии указанного полипептида фактора VIII, в том числе включающих клеточную культуральную среду, и b) выделения экспрессированного полипептида фактора VIII из клеток млекопитающих с помощью подходящих средств, включающих добавление к указанным клеткам лиганда С2-домена.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фиг.1. Последовательность гена (кДНК) фактора VIII (SEQ ID №1).
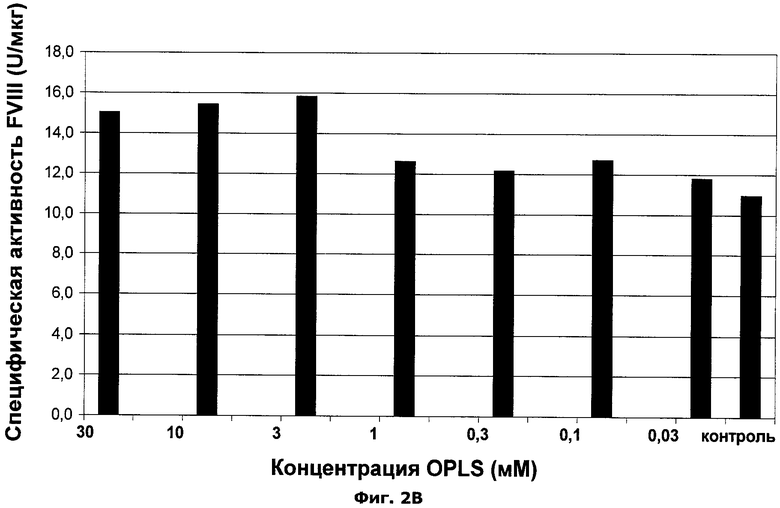
Фиг.2А-В. Влияние О-фосфо-L-серина на производительность FVIII и специфическую активность белка FVIII.
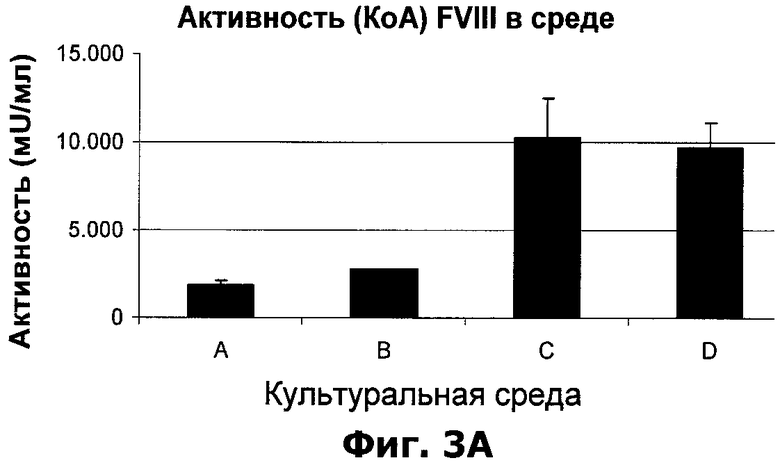
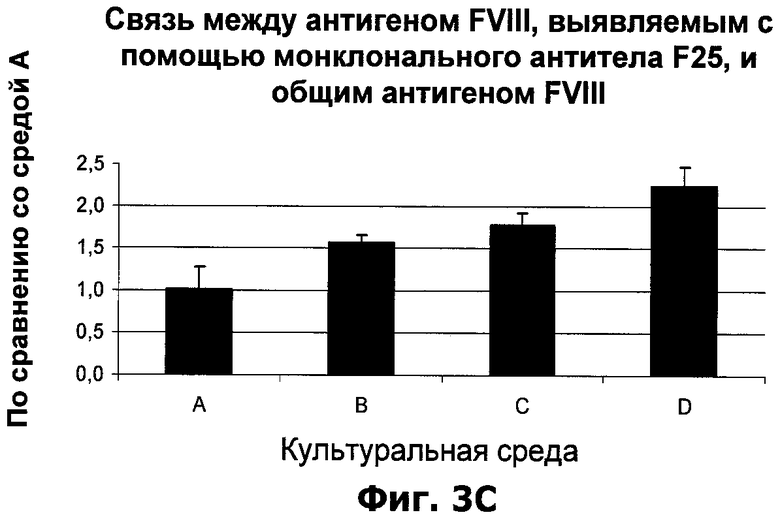
Фиг.3А-С. Влияние О-фосфо-L-серина и/или гидролизата растительного белка на фактор VIII в среде клеток, продуцирующих фактор VIII. Особенность условий A-D указана в таблице 3.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Как указывалось выше, первый аспект изобретения относится к способу продуцирования полипептида фактора VIII, включающему этапы а) культивирования клеток млекопитающих, экспрессирующих полипептид фактор VIII, в условиях, подходящих для экспрессии указанного полипептида фактора VIII, в том числе включающих клеточную культуральную среду с содержанием лиганда С2-домена, и b) выделения экспрессированного полипептида фактора VIII из клеток млекопитающих с помощью подходящих средств.
Второй аспект изобретения относится к способу продуцирования полипептида фактора VIII, включающему этапы а) культивирования клеток млекопитающих, экспрессирующих полипептид фактор VIII, в условиях, подходящих для экспрессии указанного полипептида фактора VIII, в том числе включающих клеточную культуральную среду, и b) выделения экспрессированного полипептида фактора VIII из клеток млекопитающих с помощью подходящих средств, включающих добавление к указанным клеткам лиганда С2-домена.
В обоих случаях лиганд С2-домена играет важную роль в способствовании повышению уровней титра полипептида фактора VIII в клеточной культуральной среде.
Без привязи к какой-либо конкретной теории считается, что увеличение уровней титра полипептида фактора VIII в клеточной культуральной среде обусловлено лигандом С2-домена (в частности О-фосфо-L-серином (OPLS)), который либо самостоятельно, либо в сочетании с соевым ингибитором трипсина (SBTI) и/или гидролизатом растительного белка или (i) увеличивает количество полипептида фактора VIII, секретируемого клетками, и/или (ii) конкурирует со связанным с клеткой полипептидом фактором VIII вне клеток, и/или (iii) уменьшает деградацию полипептида фактора VIII и тем самым увеличивает количество функционального полипептида фактора VIII, присутствующего в супернатанте.
Ниже изобретение будет рассмотрено более детально.
Лиганд С2-домена представляет собой лиганд, способный к связыванию или связанный с С2-доменом (см. ниже) полипептида фактора VIII. Предпочтительно лиганд С2-домена должен быть способен к вытеснению (конкурентному выключению) полипептида фактора VIII с клеточной мембраны.
В настоящее время в наиболее предпочтительном воплощении лиганд С2-домена представляет собой O-фосфо-L-серин (OPLS), т.е. молекулу формулы (HO)2P(O)OCH2CH(NH2)CO2H.
Подходящими альтернативными лигандами С2-домена считаются те, которые имеют формулу (XO)(HO)P(O)OCH2CH(NH2)CO2H, где Х выбирают среди возможно замещенного C1-6-алкила, возможно замещенного С2-6-алкенила, возможно замещенного фенила, возможно замещенного гетероарила, возможно замещенного гетероциклила и возможно замещенного бензила. В одном из его воплощений Х выбирают среди возможно замещенного C1-6-алкила, возможно замещенного бензила и возможно замещенного С2-6-алкенила.
В данном контексте термин "C1-6-алкил" используется для обозначения линейной, циклической и разветвленной углеводородной группы, имеющей от 1 до 6 атомов углерода, такой как метил, этил, пропил, изопропил, пентил, циклопентил, гексил, циклогексил.
Аналогичным образом термин "С2-6-алкенил" используется для обозначения линейной, циклической или разветвленной углеводородной группы, имеющей от 2 до 6 атомов углерода и содержащей по меньшей мере одну ненасыщенную связь. Примерами алкенильных групп являются винил, аллил, бутенил, пентенил и гексенил. Предпочтительными примерами алкенила являются винил, аллил, бутенил, особенно аллил.
В данном контексте, т.е. в связи с терминами "алкил" и "алкенил", термин "возможно замещенный" используется для обозначения того, что группа в данном случае может быть замещена один или несколько раз, предпочтительно 1-3 раза, группой(ами), выбранной среди гидрокси (которая при связывании с ненасыщенным атомом углерода может присутствовать в таутомерной кето-форме), C1-6-алкокси (т.е. C1-6-алкил-окси), С2-6-алкенилокси, карбокси, оксо (формирующей кетоновую или альдегидную функциональность), C1-6-алкилкарбонил, формил, арил, арилокси, ариламино, арилкарбонил, гетероарил, гетероарилокси, гетероариламино, гетероарилкарбонил, гетероциклил, гетероциклилокси, гетероциклиламино, гетероциклилкарбонил, амино, моно- и ди-(С1-6-алкил)амино; карбамоил, моно- и ди-(С1-6-алкил)аминокарбонил, амино-C1-6-алкил-аминокарбонил, моно- и ди-(С1-6-алкил)амино-С1-6-алкил-аминокарбонил, C1-6-алкилкарбониламино, гуанидино, карбамидо, C1-6-алкил-сульфонил-амино, C1-6-алкил-сульфонил, C1-6-алкил-сульфинил, С1-6-алкилтио, галоген, где любой арил, гетероарил и гетероциклил может быть замещен, как конкретно описано ниже для арила, гетероарила и гетероциклила.
Термин "галоген" включает фтор, хлор, бром и йод.
В данном контексте термин "арил" используется для обозначения полностью или частично ароматического карбоциклического кольца или кольцевой системы, такой как фенил, нафтил, 1,2,3,4-тетрагидронафтил, антрацил, фенантрацил, пиренил, бензопиренил, флуоренил и ксантенил, среди которых фенил является предпочтительным примером.
Термин "гетероарил" используется для обозначения полностью или частично карбоциклического ароматического кольца или кольцевой системы, где один или более атом углерода заменен на гетероатом, например на атом азота (=N- или -NH-), серы и/или кислорода. Примерами таких гетероарильных групп являются бензимидазолил, оксазолил, изоксазолил, тиазолил, изотиазолил, пирролил, имидазолил, пиразолил, пиридинил, пиримидинил, пиразинил, пиридазинил, фурил, тиенил, хинолил, триазолил, тетразолил, изохинолил, индолил, в частности бензимидазолил, пирролил, имидазолил, пиридинил, пиримидинил, фурил, тиенил, хинолил, тетразолил и изохинолил.
Термин "гетероциклил" используется для обозначения неароматического карбоциклического кольца или кольцевой системы, где один или более атом углерода заменен на гетероатом, например на атом азота (=N- или -NH-), серы и/или кислорода. Примерами таких гетероциклических групп (названных в соответствии с кольцами) являются тетрагидрофуран, имидазолидин, пиперазин, гексагидропиридазин, гексагидропиримидин, диазепан, пирролидин, пиперидин, азепан, оксазинан (морфолин) и тиазинан.
В данном контексте, т.е. в связи с терминами "арил", "бензил", "гетероарил", "гетероциклил" и т.п. (например, "арилокси", "гетероарилкарбонил" и др.), термин "возможно замещенный" используется для обозначения того, что группа в данном случае может быть замещена один или несколько раз, предпочтительно 1-5 раз, в частности 1-3 раза, группой(ами), выбранной среди групп гидрокси, C1-6-алкил, C1-6-алкокси, оксо (который может быть представлен в таутомерной енольной форме), карбокси, C1-6-алкилкарбонил, формил, амино, моно- и ди-(С1-6-алкил)амино; карбамоил, моно- и ди-(С1-6-алкил)аминокарбонил, амино-С1-6-алкил-аминокарбонил, С1-6-алкилкарбониламино, гуанидино, карбамидо, C1-6-алкилсульфониламино, арилсульфониламино, гетероарилсульфониламино, C1-6-алкилсульфонил, C1-6-алкилсульфинил, C1-6-алкилсульфонилокси, сульфанил, амино, аминосульфонил, моно- и ди-(С1-6-алкил)аминосульфонил или галогенов, где любые алкил, алкокси и т.п. представленные заместители могут быть замещены на гидрокси, C1-6-алкокси, С2-6-алкенилокси, амино, моно- и ди-(С1-6- алкил)амино, карбокси, C1-6-алкилкарбониламино, галоген, C1-6-алкилтио, C1-6-алкилсульфониламино или гуанидино.
В наиболее интересных воплощениях (касающихся как первого, так и второго аспекта изобретения) лиганд С2-домена (например OPLS) присутствует в клеточной культуральной среде в концентрации 0,1-100 мМ, такой как 5-30 мМ, в частности 10-20 мМ.
Также интересными являются воплощения (касающиеся как первого, так и второго аспекта изобретения), в которых лиганд С2-домена добавляли к клеткам в этапе b) в концентрации 1-200 мМ, такой как 50-150 мМ, в частности 70-130 мМ.
Подробная информация об этапах продуцирования будет в деталях раскрыта ниже.
Как говорилось ранее, было установлено, что соевый ингибитор трипсина (SBTI) можно выгодно сочетать с лигандом С2-домена в клеточной культуральной среде на этапе а). Таким образом, в настоящее время в предпочтительном воплощении клеточная культуральная среда дополнительно содержит соевый ингибитор трипсина.
Соевый ингибитор трипсина выделяют из Glycine max. Соевый ингибитор трипсина из соевых бобов является мономерным белком, содержащим 181 аминокислотный остаток в одноцепочечной полипептидной цепи, связанной двумя дисульфидными мостиками. Молекулярный вес, определенный по аминокислотной последовательности, составляет 20,1 кДа. Соевый ингибитор трипсина ингибирует свою целевую протеазу путем формирования стехиометрического комплекса 1:1.
В наиболее типичных воплощениях концентрация соевого ингибитора трипсина в клеточной культуральной среде составляет 0,01-100 мг/мл, например 0,1-10 мг/мл, в частности 0,3-3 мг/мл.
Кроме того, было установлено, что гидролизат растительного белка (иногда называемый "продуктом расщепления растительного происхождения" и т.п.) можно выгодно сочетать с лигандом С2-домена (а возможно, также и с соевым ингибитором трипсина) в клеточной культуральной среде на этапе а). Таким образом, в настоящее время в одинаково предпочтительном воплощении клеточная культуральная среда дополнительно содержит гидролизат растительного белка.
Гидролизат растительного белка может быть получен от одного из различных источников, например из коммерческих источников. Типичными типами гидролизатов являются гидролизат соевого белка, гидролизаты пшеничного белка, гидролизат белка гороха, гидролизат белка риса и т.д. WO 01/23527 А1, который настоящим включен путем ссылки, раскрывает получение и общее применение гидролизата соевого белка.
В наиболее типичных воплощениях концентрация гидролизата растительного белка в клеточной культуральной среде составляет 0,1-100 мг/мл, например 1-10 мг/мл, в частности 2-7 мг/мл.
Полипептид фактор VIII
Изобретение адаптировано для продуцирования полипептида фактора VIII в клетке млекопитающих.
Зрелая молекула фактора VIII состоит из 2332 аминокислот, которые могут быть сгруппированы в три гомологичные А-домена, два гомологичные С-домена и В-домен, которые расположены в следующем порядке: А1-А2-В-А3-С1-С2. В ходе его секретирования в плазму фактор VIII превращается внутриклеточно в ряд гетеродимеров, связанных с ионами металлов, в результате чего одноцепочечный фактор VIII расщепляется на В-А3 границе и на различных участках, входящих в В-домен. В результате такого процессинга образуется тяжелая цепь, состоящая из А1, А2 и различных частей В-домена, имеющая молекулярный размер от 90 кДа до 200 кДа. Тяжелые цепи связаны через ион металла с легкой цепью, которая состоит из A3-, С1- и С2-доменов (Saenko et al., 2002). В плазме этот гетеродимерный фактор VIII связан с высокоаффинным к нему фактором Виллебранда, который защищает его от преждевременного катаболизма. Время полужизни в плазме неактивированного фактора VIII, связанного с фактором Виллебранда, составляет около 12 часов.
В процессе свертывания крови фактор VIII активируется с помощью FXa и тромбина путем протеолитического расщепления в аминокислотах Arg372 и Arg740 в тяжелой цепи и в Arg1689 в легкой цепи, в результате чего высвобождается фактор Виллебранда и образуется активированный гетеродимер фактор VIII, который будет формировать теназный комплекс с FIXa и FX на фосфолипидных поверхностях в присутствии Са2+. Гетеродимер состоит из домена А1, 50 кДа фрагмента, домена А2, 43 кДа фрагмента, и легкой цепи (А3-С1-С2), 73 кДа фрагмента. Таким образом, активная форма фактора VIII (фактор Villa) состоит из А1-субъединицы, ассоциированной через двухвалентную связь иона металла с отщепленной тромбином А3-С1-С2 легкой цепью, и свободной А2-субъединицей, относительно слабо ассоциированной с доменами А1 и A3.
Молекула фактора VIII, состоящая из тяжелой цепи (НС) и легкой цепи (LC) фактора VIII, связанная с малым линкером, полученным из В-домена (фактор VIII с удаленным В-доменом, или BDD-FVIII) сохраняет биологическую активность фактора VIII полной длины (нативного).
Практикуя способ данного изобретения, любой полипептид фактор VIII, который является терапевтически полезным, например эффективным в профилактике или лечении кровотечений, можно сделать релевантным. Он включает человеческий фактор VIII дикого типа, гибридный человеческий/свиной фактор VIII и фактор VIII с удаленным В-доменом, но не ограничивается ими.
Используемый в данном документе термин "полипептид фактор VIII" охватывает фактор VIII, а также полипептиды, связанные с фактором VIII, но не ограничивается ими.
Термин "фактор VIII" используется для обозначения полипептидов, имеющих аминокислотную последовательность, описанную в Toole et al., Nature 1984, 312: 342-347 (человеческий фактор VIII дикого типа), а также фактор VIII дикого типа, полученный от других видов, такой как, например, фактор VIII от крупного рогатого скота, свиньи, собаки, мыши и лосося, но не ограничивается этим. Он также охватывает природные аллельные вариации фактора VIII, которые могут существовать и переходить от одного индивидуума к другому. Кроме того, степень и место гликозилирования или других посттрансляционных модификаций могут варьировать в зависимости от выбранных принимающих клеток (клеток-хозяев) и характера принимающей клеточной среды. Термин "фактор VIII" также используется для обозначения полипептидов фактора VIII в их нерасщепленной (зимогенной) форме, а также тех, которые были протеолитически обработаны для получения их соответствующих биологически активных форм, которые могут быть обозначены как фактор VIIIa.
Термин "полипептиды, связанные с фактором VIII", включает полипептиды фактора VIII, которые или химически модифицированы по сравнению с человеческим фактором VIII (т.е. производные фактора VIII), и/или содержат одно или более изменение аминокислотной последовательности по сравнению с человеческим фактором VIII (т.е. варианты фактора VIII), и/или содержат усеченные аминокислотные последовательности по сравнению с человеческим фактором VIII (т.е. фрагменты фактора VIII), но не ограничивается ими. Такие полипептиды, связанные с фактором VIII, могут обладать различными свойствами по сравнению с человеческим фактором VIII, включая стабильность, связывание с фосфолипидами, измененную специфическую активность и т.п. Термин "полипептиды, связанные с фактором VIII" используется для обозначения таких полипептидов в их нерасщепленной (зимогенной) форме, а также таких, которые были протеолитически обработаны для получения их биологически активных форм, которые могут быть обозначены как "полипептиды, связанные с фактором VIII" или "активированные полипептиды, связанные с фактором VIII"
Используемый в данном документе термин "полипептиды, связанные с фактором VIII" также включает в себя, без ограничения, полипептиды, демонстрирующие большей частью такую же или повышенную биологическую активность по сравнению с человеческим фактором VIII дикого типа, а также полипептиды, в которых биологическая активность фактора VIII была большей частью изменена или уменьшена по сравнению с активностью человеческого фактора VIII дикого типа. Эти полипептиды включают фактор VIII или фактор VIIIa, который был химически модифицирован, и варианты фактора VIII, в которые были внесены специфические изменения аминокислотной последовательности, которые изменили или нарушили биологическую активность полипептида, но не ограничиваются ими.
Кроме того, он охватывает полипептиды со слегка модифицированной аминокислотной последовательностью, например полипептиды, имеющие модифицированный N-конец, включая N-концевые аминокислотные делеции или добавления, и/или полипептиды, которые были химически модифицированы по сравнению с человеческим фактором VIII.
Полипептиды, связанные с фактором VIII, в том числе варианты фактора VIII, демонстрирующие большей частью такую же или повышенную биологическую активность по сравнению с фактором VIII дикого типа, или, наоборот, демонстрирующие большей частью модифицированную или уменьшенную биологическую активность по сравнению с фактором VIII дикого типа, включают полипептиды с аминокислотной последовательностью, которая отличается от последовательности фактора VIII дикого типа вставкой, удалением или заменой одной или более аминокислот, но не ограничиваются ими.
Полипептиды, связанные с фактором VIII, включая варианты, охватывают те, которые демонстрируют специфическую активность, которая составляет по меньшей мере около 10%, по меньшей мере около 20%, по меньшей мере около 30%, по меньшей мере около 40%, по меньшей мере около 50%, по меньшей мере около 60%, по меньшей мере около 70%, по меньшей мере около 80%, по меньшей мере около 90%, по меньшей мере около 100%, по меньшей мере около 110%, по меньшей мере около 120% и по меньшей мере около 130% от специфической активности фактора VIII дикого типа, произведенного клетками того же типа, при этом анализ активности фактора VIII проводят в соответствии с описанием в данной инструкции.
Полипептиды, связанные с фактором VIII, включая варианты, имеющие большей частью такую же или повышенную биологическую активность по сравнению с фактором VIII дикого типа, охватывают те, которые демонстрируют специфическую активность, которая составляет по меньшей мере около 25%, такую как по меньшей мере 50%, по меньшей мере 75% или по меньшей мере 90% от специфической биологической активности фактора VIII дикого типа, произведенного клетками того же типа, при этом анализ активности фактора VIII проводят в соответствии с описанием в данной инструкции ("Материалы и методы").
Полипептидами, связанными с фактором VIII, включая варианты, имеющие большей частью уменьшенную биологическую активность по сравнению с фактором VIII дикого типа, являются те, которые демонстрируют менее около 25%, например менее около 10% или менее около 5% от специфической активности фактора VIII дикого типа, произведенного клетками того же типа, при этом анализ активности фактора VIII был проведен в соответствии с описанием в данной инструкции ("Материалы и методы").
Примеры полипептидов фактора VIII включают полученный из плазмы человеческий фактор VIII, как описано например в Fulcher et al.; Proc. Acad. Nat. Sci. USA 1982; 79: 1648-1652 и Rotblat et al.; Biochemistry 1985; 24: 4294-4300, и FVIII, полученный из плазмы свиньи, как описано например в Fass et al.; Blood 1982; 59:594-600 и Knutson et al.; Blood 1982; 59: 615-624, но не ограничиваются ими. Примеры вариантов последовательностей фактора VIII описаны например в Lollar et al.; Blood 2000; 95(2): 564-568 (гибридные свиные/человеческие полипептиды FVIII) и Lollar et al.; Blood 2001; 97(1): 169-174, но не ограничиваются ими.
Клонирование кДНК фактора VIII (Wood, W.I., et al. (1984) Nature 312, 330-336; Vehar, G.A., et al. (1984) Nature 312, 337-342) сделало возможным экспрессию фактора VIII, которая позволила рекомбинантно получить несколько продуктов рекомбинантного фактора VIII, которые были одобрены регулирующими органами в период между 1992 и 2003 годами. Кодирующая последовательность фактора VIII (кДНК) показана на фиг.1. Тот факт, что центральный В-домен полипептидной цепи фактора VIII, находящийся между аминокислотами Arg-740 и Glu-1649, не представляется необходимым для полной биологической активности, привел также к разработке фактора VIII с удаленным В-доменом. См. также Kjalke M, Heding A, Talbo G, Persson E, Thomsen J and Ezban M (1995), "Amino acid residues 721-729 are required for full Factor VIII activity". Eur. J. Biochem: 234: 773-779.
Этап а) - Трансфекция и культивирование клеток
Клетки
Клетки млекопитающих, экспрессирующие полипептид фактор VIII, как правило, выбирали из группы, состоящей из клеток млекопитающих, которые эндогенно экспрессировали полипептид фактор VIII, и клеток млекопитающих, в которые был трансфицирован ген полипептида фактора VIII.
В одном интересном воплощении последнего клетки млекопитающих были трансфицированы вектором экспрессии, содержащим молекулу нуклеиновой кислоты, кодирующей полипептид фактор VIII, и области контроля экспрессии, функционально с ней связанные.
Экспрессия белка в клетках хорошо известна специалистам в области продуцирования белков. В практическом применении данного изобретения клетками являются клетки млекопитающих, более предпочтительно установленная линия клеток млекопитающих, включая клеточные линии СНО (например АТСС CCL 61), COS-1 (например АТСС CRL 1650), почек хомячков (BHK) и HEK293 (например АТСС CRL 1573; Graham et al., J. Gen. Virol. 36: 59-72, 1977), но не ограничиваясь ими. Предпочтительной клеточной линией ВНК является клеточная линия ВНК tk-ts13 (Waechter and Baserga, Proc.Natl.Acad.Sci.USA 79:1106-1110, 1982), далее именуемая клетками ВНК 570. Клеточную линию ВНК 570 можно получить из American Type Culture Collection, 12301 Parklawn Dr., Rockville, MD 20852, под регистрационным номером АТСС CRL 10314. Также клеточную линию ВНК tk-ts13 можно получить из АТСС под регистрационным номером CRL 1632. Предпочтительными клеточными линиями СНО являются клеточная линия СНО K1, доступная в АТСС под регистрационным номером СС161, а также клеточные линии CHO-DXB11 и CHO-DG44.
Другие подходящие клеточные линии включают клетки Rat Hep I (гепатомы крысы; АТСС CRL 1600), Rat Hep II (гепатомы крысы; АТСС CRL 1548), ТСМК (АТСС CCL 139), легкого человека (АТСС НВ 8065), NCTC 1469 (АТСС CCL 9.1); DUKX (клеточная линия СНО) (Urlaub and Chasin, Proc. Natl. Acad. Sci. USA 77:4216-4220, 1980) (клетки DUKX также именуются клетками DXB11) и DG44 (клеточная линия СНО) (Cell, 33:405, 1983, и Somatic Cell and Molecular Genetics 12:555, 1986), но не ограничиваются ими. Также могут использоваться клетки 3Т3, клетки Namalwa, миеломы и слияния миелом с другими клетками. В некоторых воплощениях клетки могут быть мутантными или рекомбинантными, такими как, например, клетки, которые экспрессируют спектры ферментов, катализирующих посттрансляционную модификацию белков (например ферменты гликозилирования, такие как гликозилтрансферазы и/или гликозидазы, или ферменты процессинга, такие как пропептиды), качественно или количественно отличающиеся от того типа клеток, из которых они были получены. Особенно предпочтительны клетки DUKX (клеточная линия СНО).
В настоящее время предпочтительными клетками являются клетки HEK293, COS, почек хомячков (BHK) и клетки миеломы, в частности клетки яичников китайского хомячка (СНО).
Культивирование клеток
В некоторых воплощениях клетки, используемые в практике изобретения, способны расти в суспензионных культурах. Используемыми в данном случае подходящими для суспензии клетками являются такие клетки, которые могут расти в суспензии без образования больших плотных агрегатов, т.е. клетки, которые являются монодисперсными или растут в виде неплотных агрегатов с небольшим количеством клеток в агрегате. Подходящие для суспензии клетки включают клетки, которые растут в суспензии без адаптации или манипуляций (такие как, например, гемопоэтические клетки или лимфоидные клетки), и клетки, которые стали подходящими для суспензии путем постепенной адаптации таких клеток, которым необходимо прикрепление (таких как, например, эпителиальные клетки или фибробласты) к росту в суспензии, но не ограничиваются ими.
Клетки, используемые в практике изобретения, могут быть адгезивными клетками (также известными как клетки, зависимые от адгезии или зависимые от прикрепления). Используемыми в данном случае адгезивными клетками являются такие клетки, которым для размножения и роста нужно присоединиться или заякориться на подходящей поверхности. В одном воплощении изобретения используемыми клетками являются адгезивные клетки. В этих воплощениях и фазы размножения, и фаза продуцирования включают применение микроносителей. Используемые адгезивные клетки должны быть способны к миграции на носителях (и во внутренней структуре носителей, если используется макропористый носитель) во время фаз(ы) размножения и к миграции на новые носители после их переноса в продукционный биореактор. Если адгезивные клетки не обладают достаточной способностью мигрировать на новые носители самостоятельно, их можно освободить от носителей путем связывания содержащего клетки микроносителя с протеолитическими ферментами или ЭДТА. Используемая среда (особенно без компонентов животного происхождения) должна также содержать компоненты, пригодные для поддержки адгезивных клеток; подходящую среду для культивирования адгезивных клеток можно получить от коммерческих поставщиков, таких как, например, Sigma.
Клетки также могут быть адаптированными к суспензии или подходящими для суспензии. При использовании таких клеток размножение клеток может происходить в суспензии, так что микроносители используются только в заключительной фазе размножения в культуральном сосуде для продуцирования и в стадии продуцирования. В случае адаптированных к суспензии клеток используемыми микроносителями являются, как правило, макропористые носители, в которых клетки закрепляются посредством физического захвата во внутренней структуре носителей. Однако в случае таких адаптированных к суспензии клеток и размножение клеток, и продукция могут происходить в суспензии.
В таких воплощениях клетки млекопитающих, как правило, выбирают среди СНО, BHK, HEK293, клеток миеломы и т.д.
Клеточная культуральная среда
Помимо компонентов, упомянутых выше, а именно лиганда С2-домена (необходим для изобретения, относящегося к первому аспекту изобретения), возможно соевого ингибитора трипсина, а также возможно гидролизата растительного белка, клеточная культуральная среда содержит ряд других компонентов, которые, как известно специалисту, необходимы для размножения клеток и продуцирования полипептида фактора VIII.
Термин "клеточная культуральная среда" (или просто "среда") относится к питательному раствору, используемому для выращивания клеток млекопитающих, который обычно содержит по меньшей мере один компонент из одной или более следующих категорий: (1) соли, например натрий, калий, магний и кальций, способствующие осмоляльности среды; (2) источник энергии, обычно в форме углеводов, таких как глюкоза; (3) все незаменимые аминокислоты, и обычно основной набор из двадцати аминокислот; (4) витамины и/или другие органические соединения, необходимые в низких концентрациях; и (5) микроэлементы, которые определяются как неорганические соединения, которые обычно требуются в очень низких концентрациях, обычно в микромолярных. Питательный раствор возможно может быть дополнен одним или более компонентом из любой из следующих категорий: (а) гормоны и другие факторы роста, такие как, например, инсулин, трансферрин и эпидермальный фактор роста; и (b) гидролизаты белков и тканей. Предпочтительно, чтобы клеточная культуральная среда не содержала компонентов животного происхождения.
Данное изобретение охватывает культивирование клеток млекопитающих в среде без компонентов животного происхождения. Используемый в данном случае термин "компоненты животного происхождения" подразумевает любые компоненты, которые производятся в интактных животных (такие как, например, белки, выделенные и очищенные от сыворотки), или производятся с использованием компонентов, производимых в интактных животных (такие как, например, аминокислота, сделанная с помощью фермента, выделенного и очищенного из животного для гидролиза материала растительного происхождения). Напротив, белок, который имеет последовательность животного белка (т.е. имеет геномное происхождение от животного), но который производится in vitro в культуре клеток (такой как, например, рекомбинантная дрожжевая или бактериальная клетка или установленная непрерывная линия клеток млекопитающих, рекомбинантная или нет), в средах без компонентов, производимых, а также выделенных и очищенных из интактных животных, не является компонентом "животного происхождения" (такой как, например, инсулин, производимый в дрожжевой или бактериальной клетке, или инсулин, производимый в установленной линии клеток млекопитающих, такой как, например, клетки СНО, ВНК или НЕК, или интерферон, производимый в клетках Namalwa). Например, белок, который имеет последовательность животного белка (т.е. имеет геномное происхождение от животного), но который производится в рекомбинантной клетке в средах без компонентов животного происхождения (такой как, например, инсулин, производимый в дрожжевой или бактериальной клетке) не является "компонентом животного происхождения". Соответственно, клеточной культурной средой без компонентов животного происхождения является такая среда, которая может содержать животные белки, которые получены рекомбинантно; однако такая среда не содержит, например, животную сыворотку или белки или другие продукты, очищенные от животной сыворотки. Такая среда может, например, содержать один или более компонент, полученный из растений. Может быть использована любая клеточная культуральная среда, в частности не содержащая компонентов животного происхождения, которая поддерживает рост и существование клеток в условиях изобретения. Как правило, среда содержит воду, регулятор осмоляльности, буфер, источник энергии, аминокислоты, неорганический или рекомбинантный источник железа, один или более синтетический или рекомбинантный фактор роста, витамины и кофакторы. В одном воплощении среда не содержит компонентов животного происхождения и белков ("безбелковая"). Среды без компонентов животного происхождения и/или белков доступны от коммерческих поставщиков, таких как, например, Sigma, JRH Biosciences, Gibco, Hyclone и Gemini.
В одном воплощении клеточная культуральная среда практически не содержит сыворотку. В другом воплощении среда не содержит компонентов животного происхождения. В другом воплощении среда не содержит белки ("безбелковая"), а также компоненты животного происхождения.
В одном воплощении среда является коммерчески доступной безбелковой средой СНО без компонентов животного происхождения, такой как, например, EXCELL™ (SAFC Biosciences), PF-CHO, PF-CHO-LS, SFM4CHO или CDM4CHO (все из Hyclone), а клеточной линией являются клетки СНО.
В некоторых воплощениях клетки, используемые в практике данного изобретения, адаптированы к суспензионному росту в среде без компонентов животного происхождения, такой как, например, среда без сыворотки. Такие адаптационные процедуры описаны например в Scharfenberg, et al., Animal Cell Technology Developments towards the 21st Century, E. C. Beuvery et al. (Eds.), Kluwer Academic Publishers, pp.619-623, 1995 (клетки BHK и СНО); Cruz, Biotechnol. Tech. 11: 117-120, 1997 (клетки насекомых); Keen, Cytotechnol. 17: 203-211, 1995 (клетки миеломы); Berg et al., Biotechniques 14: 972-978, 1993 (клетки человеческой почки 293). В частном предпочтительном воплощении принимающими клетками являются клетки BHK 21 или СНО, которые были разработаны для экспрессии человеческого фактора VIII и адаптированы к росту в отсутствие сыворотки или компонентов животного происхождения.
Клеточные культуральные процедуры
Способы изобретения обычно выполняли во встряхиваемом культуральном сосуде, и обычно использовали полунепрерывный тип процесса. В ходе этого процесса после инокуляции выращивали клетки, и при достижении определенной плотности собирали около 70% культуры, а к оставшейся культуре добавляли свежую клеточную культуральную среду до ее первоначального объема. Это, как правило, повторяли примерно 2-10 раз.
Кроме того, может быть использован процесс типа микроносителя. В процессе, основанном на микроносителе, клетки мигрировали во внутреннюю структуру носителей (макропористые носители), либо присоединялись к поверхности носителей (твердые носители), либо происходило и то, и другое. В процессе, основанном на микроносителе, вначале в культуральный сосуд помещали клетки млекопитающих, микроносители и клеточную культуральную среду. В последующие дни добавляли дополнительную клеточную культуральную среду, если культуральный объем не доходил до конечного рабочего объема сосуда вначале. В течение следующего периода выполняли периодический сбор культурального супернатанта, содержащего продукт, и заменяли его новой жидкой средой, до тех пор, пока культура окончательно не заканчивалась. При сборе супернатанта, содержащего продукт, перемешивание культуры, например встряхивание, останавливали и давали осесть носителям, содержащим клетки, после чего удаляли часть клеточного культурального супернатанта, содержащего продукт. В целях улучшения общего результата процедуры предпочтительно может быть применен этап охлаждения до сбора супернатанта, содержащего продукт, см. например WO 03/029442. В некоторых воплощениях перед тем, как дать осесть носителям, клеточную культуральную среду охлаждали до температуры от примерно 18°C до примерно 32°C, или от примерно 20°C до примерно 28°C, или от примерно 22°C до примерно 30°C.
Другие применимые варианты клеточной культуральной процедуры описаны в WO 02/29084 (Novo Nordisk A/S).
Прежде чем наступала фаза продуцирования, на которой проводился регулярный сбор содержащего продукт культурального супернатанта для дальнейшей обработки, клетки размножались стандартным образом или по любой схеме, подходящей для конкретной клетки в данном случае. Фаза размножения может содержать один этап или несколько поэтапных процедур. В одноэтапной процедуре размножения клетки извлекали из сосуда для хранения и инокулировали непосредственно в культуральный сосуд (возможно содержащий микроносители), где и происходила продукция. В многоэтапной процедуре размножения клетки извлекали из сосуда для хранения и размножали с помощью большого числа культуральных сосудов, постепенно увеличивая размер до достижения окончательного культурального сосуда (возможно содержащего микроносители), где и происходила продукция. Во время этапов размножения клетки выращивали в условиях, оптимизированных для роста. Культуральные условия, такие как температура, рН, давление растворенного кислорода, концентрация растворенного CO2 и т.п., являются такими, о которых известно, что они оптимальны для конкретной клетки, и будут очевидны специалисту в данной области (см. например Animal Cell Culture: A Practical Approach 2nd Ed., Rickwood, D. and Hames, B.D., eds., Oxford University Press, New York (1992)).
В одном подходе клеточный культуральный процесс производили в одном культуральном сосуде: клетки инокулировали непосредственно в культуральный сосуд (возможно содержащий микроносители), где происходила продукция; клетки размножались до достижения подходящей клеточной плотности, и начиналась стадия продуцирования.
В другом подходе клеточный культуральный процесс производили по меньшей мере в двух различных культуральных сосудах: одном или более сосуде для посева (первый этап(ы) размножения), а затем в культуральном сосуде для продуцирования (последний этап размножения, после которого следует фаза продуцирования). На первом этапе размножения клетки, экспрессирующие желаемый полипептид, инокулировали в культуральный сосуд для посева, содержащий клеточную культуральную среду, и они размножались до тех пор, пока не достигали минимальной плотности для следующего посева. Впоследствии размножившуюся посеянную культуру переносили в культуральный сосуд для продуцирования, содержащий клеточную культуральную среду и (возможно) микроносители. В случае процесса с использованием микроносителей клетки культивировали в этом культуральном сосуде в условиях, при которых клетки мигрировали на поверхность твердых носителей или на внешние и внутренние поверхности макропористого носителя, и они продолжали расти на этом последнем этапе размножения до тех пор, пока полностью не занимали носители. Во время этого последнего этапа размножения осуществляли смену среды, позволяя микроносителям оседать на дно культурального сосуда, после чего удаляли заранее определенную долю объема резервуара, и соответствующую долю объема резервуара восполняли свежей средой. Затем ресуспендировали микроносители в среде и этот процесс удаления и замены среды повторяли с заданным интервалом, например каждые 24 часа. Количество замен среды зависит от плотности клеток и, как правило, может составлять от 10 до 95%, предпочтительно от 25% до 80% объема резервуара.
В случае суспензионного процесса, например перфузии, периодического или полунепрерывного процесса, клетки выращивали, свободно суспендировав их без иммобилизации на носителях. В суспензионном клеточно-перфузионном процессе клетки инокулировали в культуральный сосуд для посева, содержащий культуральную среду без компонентов животного происхождения, и они размножались до тех пор, пока не достигали минимальной плотности для следующего посева. Впоследствии размножившуюся посеянную культуру переносили в крупный культуральный сосуд, содержащий культуральную среду без компонентов животного происхождения, и клетки размножались до тех пор, пока не достигали по меньшей мере заранее определенной плотности. В этой фазе клетки выращивали в суспензии, позволяя количеству клеток в культуральном сосуде возрасти до заранее определенного или критического значения. Замену среды осуществляли путем постоянной перфузии культурального сосуда свежей средой.
Количество перфузируемой среды зависит от плотности клеток и обычно может составлять от 10 до 95%, предпочтительно от 25% до 80% объема резервуара в сутки (24 часа). 60-95% среды резервуара, например около 80%, меняли, когда плотность клеток достигала значения, подходящего для начала фазы продуцирования, обычно каждые 24 часа. Замену 80% среды также предпочтительно проводить в фазе продуцирования.
В простом периодическом процессе клетки инокулировали в культуральный сосуд для посева, содержащий питательную среду без компонентов животного происхождения, и они размножались до тех пор, пока не достигали минимальной плотности для следующего посева. Впоследствии размножившуюся посеянную культуру переносили в крупный культуральный сосуд, содержащий культуральную среду без компонентов животного происхождения.
Периодический процесс, такой как этот, может быть удлинен путем подачи концентрированного раствора питательных веществ в резервуар. Это увеличивает время процесса и в конечном итоге приводит к увеличению продукции FVIII в культуральном сосуде. Время сбора должно быть определено как баланс между максимально возможно длинным процессом в резервуаре и риском лизиса клеток.
Процесс полунепрерывного культивирования (Draw-Fill) похож на повторяющуюся периодическую ферментацию. При периодической ферментации клетки выращивали в культуральном сосуде и среду собирали в конце цикла. В процессе полунепрерывного культивирования культуральный сосуд использовали для сбора до того, как любое из питательных веществ исчерпывалось. Вместо удаления всего содержимого сосуда удаляли только часть объема резервуара (обычно 80% объема резервуара). После сбора в сосуд добавляли такой же объем свежей среды. Затем клеткам позволяли расти в сосуде еще раз и через определенное количество дней снова забирали 80%. При повторных периодических процессах клетки, оставшиеся в сосуде после сбора, могут быть использованы в качестве посевного материала для следующей партии.
Процесс полунепрерывного культивирования проводили в два этапа. Первую фазу процесса проводили идентично простому периодическому процессу. После первого сбора культуральный сосуд снова действовал как при простом периодическом процессе; однако продолжительность партии была короче по сравнению с первой из-за высокой начальной плотности клеток. Эти короткие "повторные периодические фазы" могут продолжаться бесконечно.
Подпитываемый процесс полунепрерывного культивирования представляет собой ферментацию при полунепрерывном культивировании с концентрированным питательным веществом, похожим на тип, предлагаемый в подпитываемом процессе. Дело при простом процессе полунепрерывного культивирования заключается в том, что добавленной свежей среды может быть недостаточно для поддержания клеток для повторных периодических ферментаций. Включение питания снимает эту проблему. Питание также позволяет использовать культуральный сосуд с длительными периодическими промежутками в процессе полунепрерывного культивирования.
Культуральный сосуд можно использовать в широком диапазоне временных циклов и широком диапазоне объемов полунепрерывного культивирования. Диапазоны и предпочтительные значения можно увидеть в таблице 1 ниже.
Следует понимать, что в процессе, где фаза размножения представляет собой многоэтапную процедуру, размножение может иметь место в культуральных сосудах постепенно увеличивающегося размера, пока не будет получено достаточное количество клеток для введения в последний культуральный сосуд. Например, может последовательно использоваться один или более культуральный сосуд для посева объемом 5 л, 50 л, 100 л или 500 л. Культуральный сосуд для посева, как правило, имеет вместимость от 5 л до 1000 л. Обычно клетки инокулируют в культуральный сосуд для посева с начальной плотностью около 0,2-0,4×106 клеток/мл, и они размножаются до тех пор, пока культура не достигнет плотности клеток около 1,0×106 клеток/мл. Как правило, минимальная плотность для последующего посева составляет от примерно 0,8×106 до примерно 1,5×106 клеток/мл.
Некоторые из заданных величин, которые пригодны для продуцирования фактора VIII, не обязательно подходят для первоначального роста клеток либо в посеянной культуре, либо на микроносителях. Например, температура, давление растворенного кислорода и/или рН могут быть различными для двух фаз. Замены среды во время размножения проводят для того, чтобы сохранить клетки живыми и растущими, а не для того, чтобы собрать культуральный супернатант для дальнейшей обработки.
Возможно, заданная величина перепада температуры культивирования может быть использована при внесении и во время фазы продуцирования. Кроме того, при внесении температура фазы продуцирования, действующий рН и частота замены среды, как правило, изменяются на величины, которые являются оптимальными для продуцирования.
Микроносители
Используемые в данном случае микроносители представляют собой частицы, которые достаточно малы, чтобы использоваться в суспензионных культурах (с таким темпом встряхивания, который не вызывает существенное повреждение клеток). Они являются твердыми, пористыми или имеют твердый остов с пористым покрытием на поверхности. Микроносители могут быть, например, на основе целлюлозы или декстрана, и их поверхности (наружные и внутренние поверхности в случае пористых носителей) могут быть положительно заряжены, но они не ограничиваются этими примерами. Другие подробности можно найти в WO 02/29083 и в "Microcarrier cell culture, principles and methods. Amersham Pharmacia Biotech. 18-1140-62. Edition AA".
Используемые твердые микроносители включают Cytodex 1™ и Cytodex 2™ (Amersham Pharmacia Biotech, Piscataway NJ), но не ограничиваются ими. Твердыми носителями являются, в частности, носители, подходящие для адгезии клеток (субстратзависимые клетки). Используемые макропористые носители включают Cytopore 1™ и Cytopore 2™ (Amersham Pharmacia Biotech, Piscataway NJ), но не ограничиваются ими. В частности, предпочтительными являются носители Cytopore 1™ со средним диаметром частиц 230 мкм, средним размером пор 30 мкм и плотностью положительного заряда 1,1 мг-экв/г.
Культуральные условия для крупномасштабного культивирования
Изобретение, в частности, актуально для крупномасштабного продуцирования. Под термином "крупномасштабная продукция" понимается продукция с участием культурального сосуда объемом по меньшей мере 100 л. Однако в предпочтительных воплощениях масштаб, как правило, не менее 250 л, такой как по меньшей мере 500 л, например не менее 1000 л или даже 5000 л или более. Термин "крупномасштабная" может быть использован наравне с терминами "в промышленных масштабах" и "производственного масштаба".
Способ для крупномасштабного продуцирования полипептида, как правило, проводят в течение не менее 120 часов, например 1-26 недель.
В случае, если клеточной культуральный процесс проводится по меньшей мере в двух различных культуральных сосудах, например в одном или более культуральном сосуде(ах) для посева (первый этап(ы) размножения), затем в культуральном сосуде для продуцирования (последний этап размножения и последующая стадия продуцирования), то этот процесс обычно включает перенос примерно 50 л размножившейся посеянной культуры (примерно 1,0×106 клеток/мл) в культуральный сосуд объемом 500 л, содержащий 150 л клеточной культуральной среды. Для крупномасштабной культуры поддерживают соответствующие условия, например температуру, рН, давление растворенного кислорода (DOT) и темп перемешивания, а объем постепенно увеличивают путем добавления среды в культуральный сосуд. В случае процесса с микроносителем культуральный сосуд также содержит количество микроносителей, соответствующее конечной концентрации микроносителя в диапазоне от 1 до 10 г/л. После переноса клетки обычно переходят на поверхность носителей или во внутреннюю часть носителей в течение первых 24 часов.
Культуральный сосуд
Культуральные сосуды, подходящие для данного изобретения, могут быть размещены, например, на стандартных реакторах с мешалкой (CSTR), где перемешивание осуществляется с помощью импеллеров обычных типов или с помощью воздушных реакторов путем введения воздуха в нижнюю часть сосуда. В числе других параметров, которые обычно контролируются в рамках указанных пределов, имеются рН, давление растворенного кислорода (DOT), концентрация растворенного CO2 и температура. Давление растворенного кислорода можно поддерживать, например, путем барботажа чистым кислородом. Концентрацию растворенного CO2 можно поддерживать путем барботажа воздухом. Средой для температурного контроля является, как правило, вода, при необходимости подогретая или охлажденная. Вода может быть пропущена через рубашку вокруг сосуда или через спиральную трубку, погруженную в культуру.
Термин "культуральный сосуд" может быть использован наравне с терминами "резервуар", "реактор", "ферментер" и "биореактор".
Этап b) - Выделение экспрессированного полипептида
На этом этапе b) полипептид фактор VIII выделяли из клеток млекопитающих с помощью подходящих средств. В типичном воплощении клетки могут быть удалены из среды, и среда может быть очищена путем последовательной фильтрации собранного материала через фильтры 1,0 мкм и 0,2 мкм.
Затем можно повысить концентрацию фактора VIII в среде (клеточном культуральном супернатанте) с помощью катионообменной хроматографии, где объединяются фракции, богатые фактором VIII. Полипептид фактор VIII затем можно очистить на колонке путем связывания с антителами анти-фактор VIII (например на колонке с антителами F25, см. например WO 95/13301 и/или Nordfang et al. 1995 (Thromb. Haemostas. 54:586-590)), а затем элюировать в условиях, сохраняющих активность полипептида фактора VIII. Другие примеси можно удалить путем буферного обмена с помощью гель-фильтрации.
Согласно второму аспекту изобретения, но также применимо к первому аспекту изобретения, лиганд С2-домена добавляли для облегчения выделения полипептида фактора VIII из клеток, т.е. лиганд С2-домена (например OPLS) добавляли для освобождения связанного с клеткой полипептида фактора VIII.
Особенностью данного изобретения является то, что полипептид фактор VIII может быть выделен из клеток без инактивации или разрушения клеток млекопитающих. Таким образом, в частном воплощении экспрессированный полипептид фактор VIII добывают из клеточной культуральной среды практически без снижения жизнеспособности клеток. Кроме того, это выгодно, если продукцию можно продолжить с использованием той же партии клеток.
После того как среду, содержащую полипептид фактор VIII, выделили из клеток, ее можно подвергнуть одному или более этапам обработки, чтобы очистить желаемый белок, включая аффинную хроматографию, хроматографию с гидрофобным взаимодействием; ионообменную хроматографию, эксклюзионную хроматографию; электрофоретические процедуры (например препаративное изоэлектрическое фокусирование), дифференциальную растворимость (например осаждение сульфатом аммония) или выделение и т.п., но не ограничиваясь ими. См., в целом, Scopes, Protein Purification, Springer-Verlag, New York, 1982; и Protein Purification, J.-C. Janson and Lars Ryden, editors, VCH Publishers, New York, 1989.
Очистка полипептидов фактора VIII, в частности, может включать аффинную хроматографию на колонке с антителами против фактора VIII и активацию протеолитического расщепления.
Следующие примеры используются для иллюстрации данного изобретения, не ограничивая его.
ПРИМЕРЫ
Материалы и методы
Клеточная линия: клеточная линия, используемая для трансфекции, dhfr-СНО клетки DUKX-B11 клетки (Urlaub, G. & Chasin, L.A. (1980) Proc. Natl. Acad. Sci. USA 77, 4216-4220), была адаптирована к росту в суспензионной культуре в среде без сыворотки с добавленим рибонуклеозидов и дезоксирибонуклеозидов.
Вектор экспрессии: транскрипцию фактора VIII проводили с помощью промотора аденовируса SV40, а селекцию с помощью селективного маркера дигидрофолатредуктазы. Экспрессированная молекула фактора VIII состоит из тяжелой цепи (НС) и легкой цепи (LC) фактора VIII, связанных с коротким линкером, полученным из В-домена. В-домен удаляли, так как это позволяет сохранить высокую экспрессию фактора VIII и биологическую активность фактора VIII.
Трансфекция: β-лактамазный ген удаляли путем расщепления с помощью ферментов из плазмиды №815 F8-500B-pTSV7, а полученный фрагмент, содержащий ген фактора VIII, очищали в геле и использовали для трансфекции DUKX-B11 СНО-клеток с помощью реагента FuGENE 6 (Roche). Трансфекцию проводили в 6-луночных платах в среде α-МЕМ (Gibco), дополненной рибонуклеозидами и дезоксирибонуклеозидами и 10% диализированной FBS (фетальной бычьей сывороткой). Через два дня после трансфекции клетки переносили во флаконы ТС80 в среду α-МЕМ (Gibco) без рибонуклеозидов и дезоксирибонуклеозидов, но с 10% диализированной FBS. После отбора выживших трансфектантов в течение 15 дней начинали ступенчатую амплификацию с метотрексатом. Клетки амплифицировали с метотрексатом до 1000 нМ с несколькими субклонированиями, выполняемыми во время этого процесса.
SF-адаптация и культивирование клеток: клетки адаптировали для роста в среде без сыворотки путем ступенчатого снижения концентрации FBS в среде SF. Клетки адаптировали и поддерживали во флаконах объемом 125 мл на шейкере.
Культивирование клеток во время экспериментов с дополненной средой без сыворотки: Для экспериментов с дополненными средами клетки культивировали в соответствии с перфузионной моделью высокой плотности клеток в пробирках объемом 50 мл с вентилируемой крышкой на шейкере при температуре 35°C в среде без сыворотки, как описано ниже. Клетки культивировали в больших колбах на шейкере при 37°C. Жизнеспособность клеток оценивали по собранным клеткам, и она всегда составляла более 95%. Собранные клетки ресуспендировали в свежей среде. 2,5 мл собранных и ресуспендированных клеток добавляли к 2,5 мл свежей среды с добавкой, так чтобы получить общий объем 5 мл с концентрацией 1×107 клеток/мл. Пробирки для шейкера помещали в шейкер с температурой 35°C и скоростью 250 об/мин. Через 24 часа в образцах анализировали плотность клеток, жизнеспособность, КоА, ИФА и целостность белка фактора VIII с помощью Вестерн-блоттинга.
Жизнеспособность клеток: жизнеспособность клеточной культуры можно измерить, например, в соответствии с описанием Mammalian Cell Culture; essential techniques, 1997 (Wiley) Editors: A. Doyle and J. Bryan Griffiths (см. например протоколы 13 и 14).
Анализ КоА (анализ активности фактора VIII): В присутствии кальция и фосфолипидов фактор Х переходит в активную форму фактор Ха с помощью фактора IXa. Это образование в значительной степени стимулируется фактором VIII, который может рассматриваться в качестве кофактора этой реакции. При использовании оптимального количества Са2+ и фосфолипидов и избытка фактора IXa и X скорость активации фактора Х зависит только от количества фактора VIII. Фактор Ха гидролизует хромогенный субстрат S-2765 таким образом, что освобождается хромофорная группа, pNA. Цвет затем оценивают фотометрически при длине волны 405 нм. Полученный фактор Ха и, следовательно, интенсивность цвета, пропорциональны активности фактора VIII в образце. Гидролиз S-2765 образованным тромбином предотвращают путем добавления синтетического ингибитора тромбина, I-2581, вместе с субстратом (Chromogenix Coatest SP Factor VIII, diaPharma).
Другие тесты на активность фактора VIII: другие подходящие анализы для выявления активности фактора VIII могут быть проведены как простые тесты in vitro, описанные, например, в Kirkwood TBL, Rizza CR, Snape TJ, Rhymes IL, Austen DEG. Identification of sources of interlaboratory variation in factor VIII assay. В J Haematol 1981; 37; 559-68.; или Kessels et al., British Journal of Haematology, Vol.76 (Suppl.1) pp.16 (1990). Биологическую активность фактора VIII можно также количественно оценить путем измерения способности препарата приводить в норму время свертывания плазмы, дефицитной по фактору VIII, например как описано в Nilsson et al., 1959.(Nilsson IM, Blombaeck M, Thilen A, von Francken I., Carriers of haemophilia A - A laboratory study, Acta Med Scan 1959; 165:357). В этом анализе биологическая активность выражается в единицах/мл плазмы (1 единица соответствует количеству FVIII, находящемуся в нормальной пулированной плазме).
ИФА: Стрипованные лунки предварительно покрывали овечьими поликлональными антителами к человеческому фактору VIII. Образцы разводили и вносили в лунки. Имеющийся антиген фактор VIII связывался с прикрепленным антителом. После отмывки несвязанного материала вносили меченные пероксидазой овечьи обнаруживаемые антитела и давали им связаться с фиксированным фактором VIII. Лунки снова промывали, вносили раствор ТМБ (субстрат пероксидазы тетраметилбензидин) и давали прореагировать в течение определенного периода времени. Развивалась синяя окраска, которая изменялась на желтую при остановке реакции кислотой. Полученный цвет измеряли спектрофотометрически в микропланшетном фотометре при 450 нм. Абсорбция при 450 нм прямо пропорциональна количеству антигена фактора VIII, фиксированного в лунке (VisuLize, FVIII antigen kit, Affinity biologicals). Анализ калибровали с помощью очищенного фактора VIII с удаленным В-доменом.
F25 ИФА: ИФА: Стрипованные лунки предварительно покрывали овечьими поликлональными антителами к человеческому фактору VIII. Образцы разводили и вносили в лунки. Имеющийся антиген фактор VIII связывался с прикрепленным антителом. После отмывки несвязанного материала вносили разведенные мышиные моноклональные антитела F25 против фактора VIII, распознающие С-конец тяжелой цепи фактора VIII, и давали им связаться с фиксированным фактором VIII. Лунки снова промывали, вносили разведенные козлиные меченные пероксидазой противомышиные IgG (DAKO) и давали им связаться с фиксированными антителами F25. Лунки снова промывали, вносили раствор ТМБ (субстрат пероксидазы тетраметилбензидин) и давали прореагировать в течение определенного периода времени. Развивалась синяя окраска, которая изменялась на желтую при остановке реакции кислотой. Полученный цвет измеряли спектрофотометрически в микропланшетном фотометре при 450 нм. Абсорбция при 450 нм прямо пропорциональна количеству антигена фактора VIII, фиксированного в лунке. Анализ калибровали с помощью собственного стандарта тяжелых цепей фактора VIII, аффинно очищенного антителами F25.
(Антитело F25: См. например WO 95/13301 и/или Nordfang et al. 1995 (Thromb. Haemostas. 54:586-590).
Пример 1: Добавление OPLS в бессывороточную культуральную среду клеток, продуцирующих фактор VIII
OPLS добавляли в бессывороточную клеточную культуральную среду в указанной концентрации в соответствии с деталями эксперимента, описанными в материалах и методах. Результаты можно видеть ниже в таблице 2 и в фиг.2А и 2В.
Вывод:
Добавление OPLS увеличивает специфическую производительность клеток, продуцирующих фактор VIII (см. фиг.2А), а также добавление OPLS увеличивает специфическую активности фактора VIII (см. фиг.2В).
Пример 2: Добавление О-фосфо-L-серина и/или растительного гидролизата в бессывороточную культуральную среду клеток, продуцирующих фактор VIII
Клетки, продуцирующие BDD-фактор VIII (клеточная линия 1C5-SF), культивировали в 50 мл пробирках с крышками с фильтром (пробирки с фильтром 50 для биореактора, ТРР). 2,5×106 клеток в 5 мл среды CDM4CHO, дополненной О-фосфо-L-серином до концентрации 20 мМ и/или растительным гидролизатом до концентрации 5 мг/мл, как показано в Таблице 3. Каждое условие было протестировано в четырех культурах по 5 мл. Культуры инкубировали в шейкере-инкубаторе (37°C, 8% CO2 и 250 об/мин). Через четыре дня после посева 1,2 мл каждой культуры центрифугировали при 2000g в течение 5 минут и удаляли клеточный осадок. Супернатант стабилизировали добавлением имидазола рН 7,2 до конечной концентрации 20 мМ и Твина 80 до конечной концентрации 0,02%, и замораживали в аликвотах по 0,2 мл при температуре -80°C.
Общее содержание антигена фактора VIII каждой культуры определяли с помощью ИФА "сэндвич"-типа. Аликвоты стабилизированной и замороженной среды размораживали и анализировали в соответствии с описанием в Материалах и методах. Было определено содержание фактора VIII, распознаваемого антителами F25, которые селективно связывают фактор VIII с инактивированным С-концом тяжелой цепи. Аликвоты стабилизированной и замороженной среды размораживали и анализировали с помощью F25 ИФА в соответствии с описанием в Материалах и методах.
Для тестирования активности аликвоты стабилизированной и замороженной среды размораживали и анализировали с помощью анализа КоА в соответствии с описанием в Материалах и методах.
Качество фактора VIII в среде каждой культуры оценивали по специфической активности, рассчитанной по активности и общему содержанию антигена фактора VIII. Долю фактора VIII с инактивированным С-концом тяжелой цепи оценивали по соотношению между количеством антигена фактора VIII, выявляемым с помощью F25 ИФА, и общим количеством антигена фактора VIII.
Результаты, полученные с двумя добавками, показаны на фиг.3А-С. Эти данные говорят о положительном влиянии добавленного O-фосфо-L-серина или растительного гидролизата к культурам клеток, продуцирующих фактор VIII. Обе добавки улучшали выход и качество рекомбинантного фактора VIII из культур клеток, и обе добавки увеличивали долю фактора VIII с инактивированным С-концом тяжелой цепи в среде. Кроме того, дополнительное положительное влияние на долю фактора VIII с инактивированным С-концом тяжелой цепи было отмечено, когда О-фосфо-L-серин и растительный гидролизат использовались в комбинации.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ КУЛЬТИВИРОВАНИЯ КЛЕТОК БЕЗ КОМПОНЕНТОВ ЖИВОТНОГО ПРОИСХОЖДЕНИЯ | 2004 |
|
RU2369634C2 |
| СПОСОБ ПОВЫШЕНИЯ УДЕЛЬНОЙ СКОРОСТИ И ПРОДУЦИРОВАНИЯ В ЭУКАРИОТИЧЕСКИХ КЛЕТКАХ | 2015 |
|
RU2725191C2 |
| СРЕДА ДЛЯ КУЛЬТИВИРОВАНИЯ КЛЕТОК БЕЗ БЕЛКОВ И БЕЗ СЫВОРОТКИ | 2009 |
|
RU2536244C2 |
| СРЕДА, НЕ СОДЕРЖАЩАЯ БЕЛКОВ И СЫВОРОТКИ, И СПОСОБ КУЛЬТИВИРОВАНИЯ КЛЕТОК МЛЕКОПИТАЮЩИХ В ТАКОЙ СРЕДЕ | 2000 |
|
RU2380412C2 |
| СРЕДА ДЛЯ КУЛЬТИВИРОВАНИЯ КЛЕТОК БЕЗ БЕЛКОВ И БЕЗ СЫВОРОТКИ | 2000 |
|
RU2266325C2 |
| НЕ СОДЕРЖАЩАЯ ЖИВОТНЫХ БЕЛКОВ СРЕДА ДЛЯ КУЛЬТИВИРОВАНИЯ КЛЕТОК | 2005 |
|
RU2383616C2 |
| ЭКСПРЕССИОННАЯ ГЕНЕТИЧЕСКАЯ КОНСТРУКЦИЯ pOptivec/F8BDD, КОДИРУЮЩАЯ РЕКОМБИНАНТНЫЙ ФАКТОР СВЕРТЫВАЕМОСТИ КРОВИ VIII ЧЕЛОВЕКА С ДЕЛЕЦИЕЙ В-ДОМЕНА, И КЛЕТОЧНАЯ ЛИНИЯ DG-OV-F8BDD-18, ПРОДУЦИРУЮЩАЯ РЕКОМБИНАНТНЫЙ ФАКТОР СВЕРТЫВАЕМОСТИ КРОВИ VIII ЧЕЛОВЕКА С ДЕЛЕЦИЕЙ В-ДОМЕНА | 2009 |
|
RU2429294C2 |
| СРЕДА ДЛЯ КУЛЬТУРЫ КЛЕТОК, НЕ СОДЕРЖАЩАЯ ОЛИГОПЕПТИДОВ | 2017 |
|
RU2758802C2 |
| СРЕДА ДЛЯ КУЛЬТУРЫ КЛЕТОК, НЕ СОДЕРЖАЩАЯ ОЛИГОПЕПТИДОВ | 2013 |
|
RU2642269C9 |
| СПОСОБ ЭКСПРЕССИИ БЕЛКА | 2007 |
|
RU2486236C2 |






Изобретение относится к области биотехнологии, конкретно к продукции терапевтически активных полипептидов с использованием клеток млекопитающих, и может быть использовано для продуцирования полипептида фактора VIII. Способ продуцирования полипептида фактора VIII предусматривает культивирование клеток млекопитающих, экспрессирующих полипептид фактор VIII, в условиях, подходящих для экспрессии указанного полипептида фактора VIII, которые включают клеточную культуральную среду с О-фосфо-L-серином (OPLS), и выделение экспрессированного полипептида фактора VIII из клеток млекопитающих с помощью подходящих средств. Изобретение позволяет получить полипептид фактор VIII с увеличенной специфической активностью. 2 н. и 9 з.п. ф-лы, 3 ил., 3 табл., 2 пр.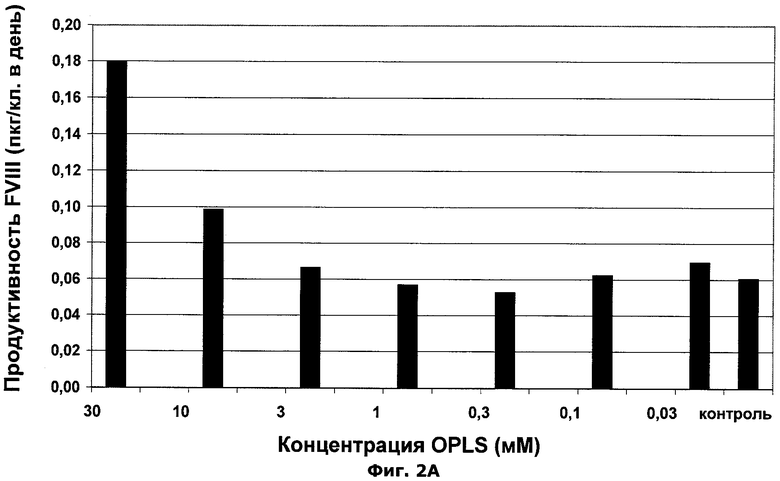
1. Способ продуцирования полипептида фактора VIII, содержащий этапы: а) культивирования клеток млекопитающих, экспрессирующих полипептид фактора VIII, в условиях, подходящих для экспрессии указанного полипептида фактора VIII, которые включают клеточную культуральную среду с О-фосфо-L-серином (OPLS), и b) выделения экспрессированного полипептида фактора VIII из клеток млекопитающих с помощью подходящих средств.
2. Способ продуцирования полипептида фактора VIII, содержащий этапы: а) культивирования клеток млекопитающих, экспрессирующих полипептид фактора VIII в условиях, подходящих для экспрессии указанного полипептида фактора VIII, которые включают клеточную культуральную среду, и b) выделения экспрессированного полипептида фактора VIII из клеток млекопитающих с помощью подходящих средств, включающих добавление к указанным клеткам О-фосфо-L-серина (OPLS).
3. Способ по п.1 или 2, где О-фосфо-L-серин (OPLS) находится в клеточной культуральной среде в концентрации 0,1-100 мМ.
4. Способ по п.1 или 2, где О-фосфо-L-серин (OPLS) добавляют к клеткам на этапе b) в концентрации 1-200 мМ.
5. Способ по п.1 или 2, где клеточная культуральная среда дополнительно содержит соевый ингибитор трипсина.
6. Способ по п.1 или 2, где клеточная культуральная среда дополнительно содержит гидролизат растительного белка.
7. Способ по п.1 или 2, где клетки млекопитающих выбраны из группы, состоящей из клеток млекопитающих, которые эндогенно экспрессируют полипептид фактора VIII, и клеток млекопитающих, в которые был трансфицирован ген полипептида фактора VIII.
8. Способ по п.1 или 2, где в клетки млекопитающих был трансфицирован вектор экспрессии, содержащий молекулу нуклеиновой кислоты, кодирующую полипептид фактора VIII, и области контроля экспрессии, функционально с ней связанные.
9. Способ по п.1 или 2, где клеточная культуральная среда практически не содержит сыворотку.
10. Способ по п.1 или 2, где экспрессированный полипептид фактора VIII получают из клеточной культуральной среды практически без снижения жизнеспособности клеток.
11. Способ по п.10, где продукция продолжается с использованием той же партии клеток.
| Захват манипулятора | 1978 |
|
SU745672A1 |
| GILBERT G.E | |||
| et al | |||
| Specific membrane binding of factor VIII is mediated by O-phospho-L-serine, a moiety of phosphatidylserine // Biochemistry | |||
| Способ изготовления фанеры-переклейки | 1921 |
|
SU1993A1 |
| WO 2004071420 А2, 26.08.2004 | |||
| РЕКОМБИНАНТНАЯ МОЛЕКУЛА ДНК, КОДИРУЮЩАЯ МОДИФИЦИРОВАННЫЙ СВИНОЙ ФАКТОР VIII (POL 1212), ЭКСПРЕССИРУЮЩИЕ ВЕКТОРЫ, МОДИФИЦИРОВАННЫЙ СВИНОЙ ФАКТОР VIII, ТЕРАПЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СПОСОБЫ ПОЛУЧЕНИЯ БЕЛКА МОДИФИЦИРОВАННОГО СВИНОГО ФАКТОРА VIII (ВАРИАНТЫ) И ЛИНИИ КЛЕТОК (ВАРИАНТЫ) | 2001 |
|
RU2285724C2 |

