Настоящее изобретение относится к области продуцирования полипептидов и сред для культуры клеток. В данном документе описаны применение мета-тирозина в среде для культуры клеток для увеличения удельной продуктивности (qP), способ продуцирования полипептида в среде для культуры клеток, содержащей мета-тирозин, и среда для культуры клеток, содержащая мета-тирозин.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Культуры клеток используются в ферментационных способах для продуцирования веществ, в частности белков. Делается различие между способами, в которых культуры клеток являются генетически немодифицированными и образуют их собственные метаболические продукты, и способами, в которых организмы генетически модифицируются таким образом, что они либо продуцируют большее количество их собственных веществ, таких как белки, либо продуцируют чужеродные (гетерологичные) вещества. Организмам, продуцирующим вещества, дают питательную среду, которая гарантирует выживание организмов и обеспечивает продуцирование желательного целевого соединения. Для этих целей известны многочисленные культуральные среды, которые обеспечивают оптимальное культивирование специфического хозяина.
Белковые биотерапевтические средства, подобные моноклональным антителам, считаются общепринятыми лекарственными средствами для лечения серьезных расстройств и заболеваний, таких как рак, рассеянный склероз и ревматоидный артрит (см. обзор от Leader et al. (Leader et al. 2008)). Для обеспечения безопасности пациента и функциональной эффективности должны тщательно отслеживаться критические характеристики качества молекул данных высокоактивных соединений. Непреднамеренные химические модификации целевого белка могут происходить на протяжении всего процесса продуцирования, начиная с синтеза микроорганизмами и культурами клеток, на протяжении очистки белка, приготовления препарата и хранения. Чаще всего наблюдаемыми путями химической деградации для белковых терапевтических средств являются дезамидирование аспарагина, изомеризация аспартата (Wakankar and Borchardt, 2006; Diepold et al. 2012; Dengl et al. 2013) и окисление (Li et al. 1995; Ji et al. 2009; Hensel et al. 2011). Недавно в нескольких исследованиях был описан дополнительный релевантный, при этом генерированный ферментативно, побочный продукт, так называемый белок SV синтезированных рекомбинантно биотерапевтических средств (Khetan et al. 2010; Wen et al. 2009; Feeney et al. 2013). SV представляют собой непреднамеренные замены аминокислот в предполагаемых генетически положениях, которые возникают либо в результате собственных мутаций нуклеотидов (Bridges, 2001), либо неправильного включения альтернативной аминокислоты во время трансляции (Zeck et al. 2012). Считается, что неправильное трансляционное включение вызвано неразборчивостью аминоацил-тРНК-синтазы (aaRS) в отношении структурно родственных аминокислот при ограничении эндогенного субстрата (Feeney et al. 2013; Jakubowski, 2001). Неправильное включение свободного мета-тирозина в клеточные белки в качестве потенциального цитотоксического механизма для окисленных аминокислот описано у Gurer-Urhan et. al., (2006).
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Обнаружили, что дополнение культуральной среды мета-тирозином обеспечивает повышенную удельную продуктивность (qP) эукариотической клетки(-хозяина), которая продуцирует (неэндогенный) полипептид. Обнаружили, что добавление мета-тирозина в культуру эукариотических клеток не приводит к значимому уменьшению жизнеспособности клеток или титра конечного продукта. Рост клеток в такой культуре уменьшается/подвергается негативному влиянию (представленному пониженной плотностью жизнеспособных клеток (VCD) и общей продукцией биомассы (указанной как CTI)). В настоящем способе для увеличения удельной продуктивности культивируемых клеток нет необходимости осуществлять сдвиг температуры, осмоляльности или рН. Также нет необходимости модулировать удельную продуктивность добавлением препаратов, подобных вальпроевой кислоте или бутирату натрия. Встречаемость вариантов аминокислотной последовательности (SV) из-за неправильного включения альтернативной аминокислоты во время трансляции мРНК в присутствии мета-тирозина контролируется дополнительной подпиткой фенилаланина в нелимитирующих концентрациях.
Одним аспектом, как описано в данном документе, является применение мета-тирозина для увеличения удельной продуктивности эукариотической клетки-хозяина, которая продуцирует/экспрессирует/секретирует (неэндогенный/экзогенный) полипептид.
В одном воплощении данного аспекта эукариотическая клетка-хозяин представляет собой клетку млекопитающего. В одном воплощении данного аспекта эукариотическая клетка-хозяин представляет собой клетку яичника китайского хомяка (СНО). В одном воплощении данного аспекта клетка СНО представляет собой растущую в суспензии клетку/линию клеток СНО (суспензионная клетка/линия клеток СНО). В одном воплощении данного аспекта клетка СНО представляет собой клетку СНО-K1.
В одном воплощении данного аспекта мета-тирозин добавляют с получением (конечной) концентрации от 0,2 мМ до 0,7 мМ. В одном воплощении данного аспекта мета-тирозин добавляют с получением (конечной) концентрации от 0,25 мМ до 0,6 мМ. В одном воплощении данного аспекта мета-тирозин добавляют с получением (конечной) концентрации от 0,3 мМ до 0,5 мМ. В одном воплощении данного аспекта мета-тирозин добавляют с получением (конечной) концентрации от 0,3 мМ до 0,4 мМ.
В одном воплощении данного аспекта удельная продуктивность увеличивается по меньшей мере на 5% по сравнению с идентичным способом продуцирования без добавления мета-тирозина. В одном воплощении данного аспекта удельная продуктивность увеличивается по меньшей мере на 10% по сравнению с идентичным способом продуцирования без добавления мета-тирозина. В одном воплощении данного аспекта удельная продуктивность увеличивается по меньшей мере на 20% по сравнению с идентичным способом продуцирования без добавления мета-тирозина. В одном воплощении данного аспекта удельная продуктивность увеличивается по меньшей мере на 25% по сравнению с идентичным способом продуцирования без добавления мета-тирозина.
В одном воплощении данного аспекта применение осуществляется в культуральной среде, не содержащей белок. В одном воплощении данного аспекта применение осуществляется в химически определенной культуральной среде. В одном воплощении данного аспекта применение осуществляется в химически определенной культуральной среде, не содержащей белок.
В одном воплощении данного аспекта применение осуществляется в культуральной среде, которая дополнительно содержит фенилаланин в нелимитирующей концентрации. В одном воплощении данного аспекта применение осуществляется в культуральной среде, которая дополнительно содержит фенилаланин в нелимитирующей концентрации, где фенилаланин добавляют посредством непрерывной подпитки или последством одного или более чем одного индивидуального болюсного впрыскивания маточных растворов Phe в начале или на протяжении процесса ферментации.
В одном воплощении данного аспекта молярное отношение мета-тирозин/фенилаланин меньше чем или равно 1,25. В одном воплощении данного аспекта точность последовательности конечного белка в отношении неправильного включения м-Tyr (мета-тирозин) вместо Phe выше чем или равна 95,0%. В одном воплощении данного аспекта молярное отношение мета-тирозин/фенилаланин меньше чем или равно 1,25, и точность последовательности конечного белка в отношении неправильного включения м-Tyr вместо Phe выше чем или равна 95,0%. Это означает то, что 5% или менее остатков мета-тирозина неправильно включаются в последовательность белка относительно правильно включенных остатков фенилаланина. В одном воплощении данного аспекта молярное отношение мета-тирозин/фенилаланин меньше чем или равно 0,25. В одном воплощении данного аспекта точность последовательности конечного белка в отношении неправильного включения м-Tyr вместо Phe выше чем или равна 99,0%. В одном воплощении данного аспекта молярное отношение мета-тирозин/фенилаланин меньше чем или равно 0,25, и точность последовательности конечного белка в отношении неправильного включения м-Tyr вместо Phe выше чем или равна 99,0%. В одном воплощении данного аспекта молярное отношение мета-тирозин/фенилаланин меньше чем или равно 0,125. В одном воплощении данного аспекта точность последовательности конечного белка в отношении неправильного включения м-Tyr вместо Phe выше чем или равна 99,5%. В одном воплощении данного аспекта молярное отношение мета-тирозин/фенилаланин меньше чем или равно 0,125, и точность последовательности конечного белка в отношении неправильного включения м-Tyr вместо Phe выше чем или равна 99,5%. В одном воплощении данного аспекта молярное отношение мета-тирозин/фенилаланин меньше чем или равно 0,025. В одном воплощении данного аспекта точность последовательности конечного белка в отношении неправильного включения м-Tyr вместо Phe выше чем или равна 99,9%. В одном воплощении данного аспекта молярное отношение мета-тирозин/фенилаланин меньше чем или равно 0,025, и точность последовательности конечного белка в отношении неправильного включения м-Tyr вместо Phe выше чем или равна 99,9%.
В одном воплощении данного аспекта применение осуществляется при постоянной температуре.
В одном воплощении данного аспекта применение осуществляется при температуре, которая снижается по ходу применения.
В одном воплощении данного аспекта применение осуществляется при постоянном рН.
В одном воплощении данного аспекта полипептид представляет собой иммуноглобулин или его вариант, или его фрагмент, или его гибрид. В одном воплощении данного аспекта полипептид представляет собой человеческий или гуманизированный иммуноглобулин или его вариант, или его фрагмент, или его гибрид. В одном воплощении данного аспекта полипептид представляет собой гуманизированное антитело. В одном воплощении данного аспекта полипептид представляет собой гуманизированное моноклональное антитело.
Одним аспектом, как описано в данном документе, является способ продуцирования полипептида в эукариотической клетке-хозяине, экспрессирующей нуклеиновую кислоту, кодирующую полипептид, включающий культивирование эукариотической клетки-хозяина в культуральной среде, содержащей мета-тирозин.
В одном воплощении данного аспекта мета-тирозин добавляют с получением (конечной) концентрации от 0,2 мМ до 0,7 мМ. В одном воплощении данного аспекта мета-тирозин добавляют с получением (конечной) концентрации от 0,25 мМ до 0,6 мМ. В одном воплощении данного аспекта мета-тирозин добавляют с получением (конечной) концентрации от 0,3 мМ до 0,5 мМ. В одном воплощении данного аспекта мета-тирозин добавляют с получением (конечной) концентрации от 0,3 мМ до 0,4 мМ.
В одном воплощении данного аспекта удельная продуктивность увеличивается по меньшей мере на 5% по сравнению с идентичным способом продуцирования без добавления мета-тирозина. В одном воплощении данного аспекта удельная продуктивность увеличивается по меньшей мере на 10% по сравнению с идентичным способом продуцирования без добавления мета-тирозина. В одном воплощении данного аспекта удельная продуктивность увеличивается по меньшей мере на 20% по сравнению с идентичным способом продуцирования без добавления мета-тирозина. В одном воплощении данного аспекта удельная продуктивность увеличивается по меньшей мере на 25% по сравнению с идентичным способом продуцирования без добавления мета-тирозина.
В одном воплощении данного аспекта эукариотическая клетка-хозяин представляет собой клетку млекопитающего. В одном воплощении данного аспекта эукариотическая клетка-хозяин представляет собой клетку яичника китайского хомяка (СНО). В одном воплощении данного аспекта клетка СНО представляет собой суспензионную клетку СНО/клетку СНО, растущую в суспензии. В одном воплощении данного аспекта клетка СНО представляет собой клетку СНО-K1.
В одном воплощении данного аспекта способ осуществляется в культуральной среде, не содержащей белок. В одном воплощении данного аспекта способ осуществляется в химически определенной культуральной среде. В одном воплощении данного аспекта способ осуществляется в химически определенной культуральной среде, не содержащей белок.
В одном воплощении данного аспекта способ осуществляется в культуральной среде, которая дополнительно содержит фенилаланин в нелимитирующей концентрации. В одном воплощении данного аспекта способ осуществляется в культуральной среде, которая дополнительно содержит фенилаланин в нелимитирующей концентрации, где фенилаланин добавляют непрерывной подпиткой или одним или более чем одним индивидуальным впрыскиванием болюса маточных растворов Phe в начале или во время процесса ферментации.
В одном воплощении данного аспекта молярное отношение мета-тирозин/фенилаланин меньше чем или равно 1,25. В одном воплощении данного аспекта точность последовательности конечного белка в отношении неправильного включения м-Tyr вместо Phe выше чем или равна 95,0%. В одном воплощении данного аспекта молярное отношение мета-тирозин/фенилаланин меньше чем или равно 1,25, и точность последовательности конечного белка в отношении неправильного включения м-Tyr вместо Phe выше чем или равна 95,0%. Это означает то, что 5% или менее остатков мета-тирозина неправильно включаются в последовательность белка относительно правильно включенных остатков фенилаланина.
В одном воплощении данного аспекта молярное отношение мета-тирозин/фенилаланин меньше чем или равно 0,25. В одном воплощении данного аспекта точность последовательности конечного белка в отношении неправильного включения м-Tyr вместо Phe выше чем или равна 99,0%. В одном воплощении данного аспекта молярное отношение мета-тирозин/фенилаланин меньше чем или равно 0,25, и точность последовательности конечного белка в отношении неправильного включения м-Tyr вместо Phe выше чем или равна 99,0%.
В одном воплощении данного аспекта молярное отношение мета-тирозин/фенилаланин меньше чем или равно 0,125. В одном воплощении данного аспекта точность последовательности конечного белка в отношении неправильного включения м-Tyr вместо Phe выше чем или равна 99,5%. В одном воплощении данного аспекта молярное отношение мета-тирозин/фенилаланин меньше чем или равно 0,125, и точность последовательности конечного белка в отношении неправильного включения м-Tyr вместо Phe выше чем или равна 99,5%.
В одном воплощении данного аспекта молярное отношение мета-тирозин/фенилаланин меньше чем или равно 0,025. В одном воплощении данного аспекта точность последовательности конечного белка в отношении неправильного включения м-Tyr вместо Phe выше чем или равна 99,9%. В одном воплощении данного аспекта молярное отношение мета-тирозин/фенилаланин меньше чем или равно 0,025, и точность последовательности конечного белка в отношении неправильного включения м-Tyr вместо Phe выше чем или равна 99,9%.
В одном воплощении данного аспекта способ осуществляется при постоянной температуре. В одном воплощении данного аспекта способ осуществляется при температуре, которая снижается в ходе применения.
В одном воплощении данного аспекта способ осуществляется при постоянном рН.
В одном воплощении данного аспекта полипептид представляет собой иммуноглобулин или его вариант, или его фрагмент, или его гибрид. В одном воплощении данного аспекта полипептид представляет собой человеческий или гуманизированный иммуноглобулин или его вариант, или его фрагмент, или его гибрид. В одном воплощении данного аспекта полипептид представляет собой гуманизированное антитело. В одном воплощении данного аспекта полипептид представляет собой гуманизированное моноклональное антитело.
Одним аспектом, как описано в данном документе, является культуральная среда, содержащая мета-тирозин и фенилаланин в молярном отношении, которое меньше чем или равно 1,25. Одним аспектом, как описано в данном документе, является культуральная среда, содержащая мета-тирозин и фенилаланин в молярном отношении, которое меньше чем или равно 1,25, и точность последовательности конечного белка в отношении неправильного включения м-Tyr вместо Phe выше чем или равна 95,0%. Это означает то, что 5% или менее остатков мета-тирозина неправильно включаются в последовательность белка относительно правильно включенных остатков фенилаланина.
Одним аспектом, как описано в данном документе, является культуральная среда, содержащая мета-тирозин и фенилаланин в молярном отношении, которое меньше чем или равно 0,25. Одним аспектом, как описано в данном документе, является культуральная среда, содержащая мета-тирозин и фенилаланин в молярном отношении, которое меньше чем или равно 0,25, и точность последовательности конечного белка в отношении неправильного включения м-Tyr вместо Phe выше чем или равна 99,0%.
Одним аспектом, как описано в данном документе, является культуральная среда, содержащая мета-тирозин и фенилаланин в молярном отношении, которое меньше чем или равно 0,125. Одним аспектом, как описано в данном документе, является культуральная среда, содержащая мета-тирозин и фенилаланин в молярном отношении, которое меньше чем или равно 0,125, и точность последовательности конечного белка в отношении неправильного включения м-Tyr вместо Phe выше чем или равна 99,5%.
Одним аспектом, как описано в данном документе, является культуральная среда, содержащая мета-тирозин и фенилаланин в молярном отношении, которое меньше чем или равно 0,025. Одним аспектом, как описано в данном документе, является культуральная среда, содержащая мета-тирозин и фенилаланин в молярном отношении, которое меньше чем или равно 0,025, и точность последовательности конечного белка в отношении неправильного включения м-Tyr вместо Phe выше чем или равна 99,9%.
В одном воплощении данного аспекта культуральная среда представляет собой культуральную среду, не содержащую белок. В одном воплощении данного аспекта культуральная среда представляет собой химически определенную культуральную среду. В одном воплощении данного аспекта культуральная среда представляет собой химически определенную культуральную среду, не содержащую белок.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Полагают, что модуляция удельной продуктивности (qP) эукариотической клетки является мощным инструментом для того, чтобы затрагивать изменения в характеристиках качества продукта, таких как микрогетерогенность N-гликозилирования белка, варианты заряда, агрегаты белка и профили фрагментов. Например, считается, что умеренная qP поддерживает посттрансляционное N-гликозилирование, поскольку белок-мишень может иметь более продолжительное время нахождения в компартментах образования гликанов - эндоплазматическом ретикулуме (ER) и аппарате Гольджи (Hossleret al., 2009).
В настоящее время модуляция qP в биотехнологической практике осуществляется либо посредством препаратов, подобных ингибиторам гистондеацетилазы (HDAC) - вальпроевой кислоте и бутирату натрия (Lee et al. 2009; Murray-Beaulieu et al. 2009), или посредством физико-химических параметров, таких как сдвиг температуры (Hendrick et al. 2001), сдвиг рН (Yoon et al. 2005) и увеличение осмоляльности (Han et al. 2009). Данные подходы имеют значительные недостатки. Ингибиторы HDAC, с одной стороны, сильно уменьшают жизнеспособность клеток и индуцируют апоптоз, что может вызывать вторичные проблемы, подобные фрагментации целевого белка и плохой эффективности устранения белка и ДНК клетки-хозяина во время очистки белка. С другой стороны, физико-химические параметры сложно контролировать в производственных помещениях соответствующих GMP (надлежащая медицинская практика), и они часто варьируют по кинетике сдвига разных биореакторов из-за ограничений, накладываемых сосудами, и разных переносов массы. В данном документе описано то, что специфическое дополнение мета-тирозином в биотехнологических способах является многообещающей альтернативой существующей методики увеличения qP эукариотических клеток, особенно клеток СНО, без влияния на жизнеспособность клеток и без необходимости физико-химических корректировок.
Однако Gurer-Orhan et al., (2006) было описано то, что определенные уровни концентраций мета-тирозина индуцируют цитотоксические эффекты в клетках СНО, растущих прикрепленно.
Изобретение в том виде, как оно описано в данном документе, по меньшей мере частично основано на данных о том, что дополнение культуральной среды мета-тирозином приводит к повышенной удельной продуктивности (qP) эукариотической суспензионной клетки, особенно СНО, которая продуцирует экзогенный полипептид.
В отличие от данных Gurer-Orhan et al., добавление мета-тирозина не приводило к цитотоксическим эффектам (т.е. жизнеспособность клеток и титр конечного продукта не подвергаются значимому влиянию), несмотря на то, что рост клеток подвергался негативному влиянию, что можно видеть по пониженной плотности жизнеспособных клеток (VCD) и общей продукции биомассы (указанной в виде CTI) (см. Фиг. 1-5).
Следовательно, одним аспектом, как описано в данном документе, является применение мета-тирозина для увеличения удельной продуктивности эукариотической клетки-хозяина, которая продуцирует полипептид.
Одним аспектом, как описано в данном документе, является способ продуцирования полипептида в эукариотической клетке-хозяине, экспрессирующей нуклеиновую кислоту, кодирующую полипептид, включающий культивирование эукариотической клетки-хозяина в культуральной среде, содержащей мета-тирозин.
В одном воплощении всех аспектов эукариотическая клетка-хозяин представляет собой клетку млекопитающего. В одном воплощении клетка млекопитающего выбрана из клеток млекопитающих, содержащих клетки СНО (например, СНО-K1 или СНО DG44), клетки BHK (клетки почки новорожденного хомяка), клетки NS0, клетки SP2/0, клетки HEK 293 (клетки человеческой эмбриональной почки 293), клетки HEK 293 EBNA, клетки PER.C6 и клетки COS. В одном воплощении всех аспектов клетка млекопитающего представляет собой клетку яичника китайского хомяка (СНО). В одном воплощении всех аспектов клетка СНО представляет собой суспензионную клетку СНО. В одном воплощении всех аспектов клетка СНО представляет собой клетку СНО-K1.
Обнаружили то, что увеличение удельной продуктивности может достигаться в интервале концентраций мета-тирозина, добавленного в культуральную среду.
В одном воплощении всех аспектов мета-тирозин добавляют с получением концентрации от 0,2 мМ до 0,7 мМ. В одном воплощении всех аспектов мета-тирозин добавляют с получением концентрации от 0,25 мМ до 0,6 мМ. В одном воплощении всех аспектов мета-тирозин добавляют с получением концентрации от 0,3 мМ до 0,5 мМ. В одном воплощении всех аспектов мета-тирозин добавляют с получением концентрации от 0,3 мМ до 0,4 мМ.
В одном воплощении всех аспектов удельная продуктивность увеличивается по меньшей мере на 5% по сравнению с идентичным способом продуцирования без добавления мета-тирозина. В одном воплощении всех аспектов удельная продуктивность увеличивается по меньшей мере на 10% по сравнению с идентичным способом продуцирования без добавления мета-тирозина. В одном воплощении всех аспектов удельная продуктивность увеличивается по меньшей мере на 20% по сравнению с идентичным способом продуцирования без добавления мета-тирозина. В одном воплощении всех аспектов удельная продуктивность увеличивается по меньшей мере на 25% по сравнению с идентичным способом продуцирования без добавления мета-тирозина.
Обнаружили то, что для достижения повышенной удельной продуктивности не требуется осуществлять сдвиг температуры, осмоляльности или рН, и также не требуется модулировать удельную продуктивность посредством добавления препаратов, подобных вальпроевой кислоте или бутирату натрия, как описывается в современном уровне техники. Тем не менее, специалист в данной области понимает то, что данные модификации способа культивирования также могут быть дополнительно сделаны/включены в способ, описанный в данном документе.
В одном воплощении всех аспектов применение или способ осуществляются при постоянной температуре. В одном воплощении всех аспектов применение или способ осуществляются при температуре, которая снижается в ходе применения.
В одном воплощении всех аспектов применение или способ осуществляются при постоянном рН.
Возможные варианты последовательности (SV) из-за неправильного включения альтернативной аминокислоты во время трансляции, которое может происходить при добавлении мета-тирозина в культуральную среду, контролируются дополнительной подпиткой фенилаланина в нелимитирующих концентрациях.
В Таблице 1 показано количество/частота/доля образования варианта последовательности с заменой Phe на xTyr (х означает мета-Tyr и/или орто-Tyr) для индикаторного пептида (максимальный уровень неправильного включения xTyr вместо Phe посредством добавления 0,1 мМ, 0,3 мМ и 0,4 мМ орто-Tyr и мета-Tyr в сутки 14).
Из данных супернатанта мета-, орто-Tyr и L-Phe и соответствующего неправильного включения xTyr вместо Phe эксперимента с добавлением могут быть рассчитаны пороговые отношения мета-Tyr/Phe и орто-Tyr/Phe, что приводит к конечной точности продуцируемой последовательности 99,9%, 99,5%, 99,0% и 95,0% (относящейся к неправильному включению орто- и мета-Tyr).

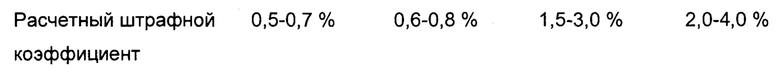
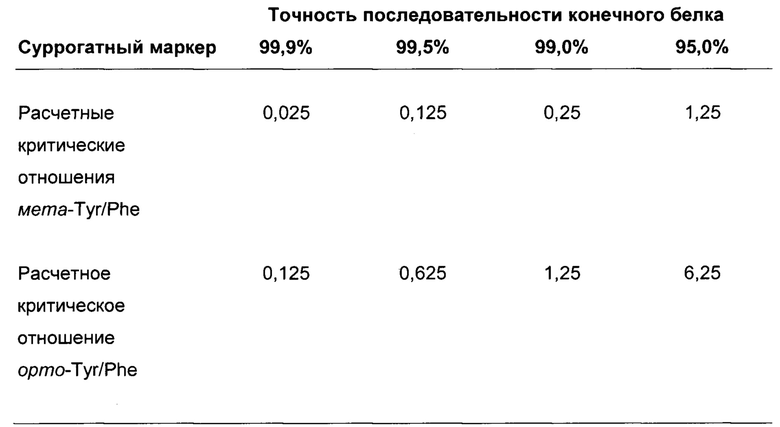
Обнаружили то, что максимальное пороговое отношение мета-Tyr/Phe 1,25, 0,25, 0,125 или 0,025, доведенное в культуральной среде, может контролировать/устранять нежелательные SV в продуцированных полипептидах (95,0%, 99,0%, 99,5% или 99,9% точности последовательности соответственно) с увеличенной, в то же самое время, удельной продуктивностью. Она остается увеличенной, независимо от дополнительного добавления Phe (см. Фиг. 8).
В одном воплощении всех аспектов культуральная среда дополнительно содержит фенилаланин в нелимитирующей концентрации.
В одном воплощении всех аспектов молярное отношение мета-тирозин/фенилаланин меньше чем или равно 1,25. В одном воплощении всех аспектов точность последовательности конечного белка в отношении неправильного включения м-Tyr вместо Phe выше чем или равна 95,0%. В одном воплощении данного аспекта молярное отношение мета-тирозин/фенилаланин меньше чем или равно 1,25, и точность последовательности конечного белка в отношении неправильного включения м-Tyr вместо Phe выше чем или равна 95,0%.
В одном воплощении всех аспектов молярное отношение мета-тирозин/фенилаланин меньше чем или равно 0,25. В одном воплощении всех аспектов точность последовательности конечного белка в отношении неправильного включения м-Tyr вместо Phe выше чем или равна 99,0%. В одном воплощении данного аспекта молярное отношение мета-тирозин/фенилаланин меньше чем или равно 1,25, и точность последовательности конечного белка в отношении неправильного включения м-Tyr вместо Phe выше чем или равна 99,0%.
В одном воплощении всех аспектов молярное отношение мета-тирозин/фенилаланин меньше чем или равно 0,125. В одном воплощении данного аспекта точность последовательности конечного белка в отношении неправильного включения м-Tyr вместо Phe выше чем или равна 99,5%. В одном воплощении данного аспекта молярное отношение мета-тирозин/фенилаланин меньше чем или равно 1,25, и точность последовательности конечного белка в отношении неправильного включения м-Tyr вместо Phe выше чем или равна 99,5%.
В одном воплощении всех аспектов молярное отношение мета-тирозин/фенилаланин меньше чем или равно 0,025. В одном воплощении данного аспекта точность последовательности конечного белка в отношении неправильного включения м-Tyr вместо Phe выше чем или равна 99,9%. В одном воплощении данного аспекта молярное отношение мета-тирозин/фенилаланин меньше чем или равно 1,25, и точность последовательности конечного белка в отношении неправильного включения м-Tyr вместо Phe выше чем или равна 99,9%.
Определения
Термин «биомасса» в том виде, как он используется в данном документе, относится к количеству или массе культивируемых клеток в культуральной среде. Биомассу можно измерять непосредственно или опосредованно посредством определения плотности жизнеспособных клеток, общей плотности клеток, интеграла количества клеток по времени (для плотности жизнеспособных клеток и общей плотности клеток), интеграла объема клеток по времени (для плотности жизнеспособных клеток и общей плотности клеток), объема осажденных клеток, сухой массы или сырой массы.
Термин «биореактор» в том виде, как он используется в данном документе, относится к любому сосуду, используемому для роста культуры клеток млекопитающего. Типично биореактор будет иметь объем по меньшей мере 1 литр и может иметь объем 10, 100, 250, 500, 1000, 2500, 5000, 8000, 10000, 12000 литров или более, или любой объем между ними. На протяжении периода культивирования типично контролируются внутренние условия биореактора, включающие рН, уровень растворенного кислорода и температуру, но не ограничивающиеся ими. Биореактор может состоять из любого материала, который подходит для содержания культур клеток млекопитающих, суспендированных в средах, при культуральных условиях по настоящему изобретению, включая стекло, пластмассу или металл.
Термин «плотность клеток» в том виде, как он используется в данном документе, относится к числу клеток, присутствующих в данном объеме среды.
Термин «жизнеспособность клеток» относится к той доле клеток, которые являются живыми в конкретное время относительно общего числа клеток - живых и мертвых - в культуре в данное время.
Термин «культура клеток» относится к. клеткам, растущим в суспензии или прикрепленным, во вращающихся флаконах, колбах, стеклянных сосудах для культивирований или сосудах для культивирований из нержавеющей стали и тому подобных. Термином «культура клеток» также охватываются крупномасштабные подходы, такие как биореакторы. Настоящим изобретением охватываются методики культуры клеток как для крупно-, так и для маломасштабного продуцирования полипептидов. Можно использовать методики, включающие биореактор с псевдоожиженным слоем, культуру во встряхиваемой колбе или систему биореактора на основе емкости с перемешиванием, но не ограничивающиеся ими, и работающие, в качестве альтернативы, в периодическом, периодическом с делением, подпиточном или перфузионном режиме.
Термины «среда для культуры клеток», «культуральная среда» или «среда», используемые в настоящем изобретении взаимозаменяемо, обозначают питательный раствор, используемый для выращивания клеток млекопитающих. Такой питательный раствор обычно включает разные факторы, необходимые для роста и поддержания клеточной среды. Например, типичный питательный раствор может включать композицию базальной среды, разные добавки, в зависимости от типа культивирования, и, время от времени, агенты для селекции. Типично в таких растворах предоставлены незаменимые и заменимые аминокислоты, витамины, источники энергии, липиды и микроэлементы, необходимые для минимального роста и/или выживания клетки. Такой раствор также может содержать дополнительные компоненты, которые увеличивают рост и/или выживание выше минимальных показателей, включающие гормоны и/или другие факторы роста, конкретные ионы, такие как ион натрия, хлорид, ион кальция, ион магния и фосфат, буферные компоненты, витамины, нуклеозиды или нуклеотиды, микроэлементы, аминокислоты, липиды и/или глюкозу или другой источник энергии, но не ограничивающиеся ими. Среду преимущественно готовят с оптимальными рН и концентрацией соли для выживания и пролиферации клеток. Среда может представлять собой среду, не содержащую белок, т.е. она не будет содержать полноразмерный белок, но будет содержать неопределенные пептиды, например, из растительных гидролизатов. Среда могла бы включать человеческий сывороточный альбумин и человеческий трансферрин, но потенциально и инсулин животного происхождения и липиды, или представляет собой среду, не содержащую чужеродных компонентов, содержащую человеческий сывороточный альбумин, человеческий трансферрин, человеческий инсулин и химически определенные липиды. В качестве альтернативы, среда может представлять собой химически определенную среду, которая представляет собой среду, в которой все вещества являются определенными и присутствуют в определенных концентрациях. Данные среды могли бы содержать только рекомбинантные белки и/или гормоны, или представлять собой химически определенную среду, не содержащую белок, т.е. содержащую только низкомолекулярные компоненты и синтетические пептиды/гормоны, если это необходимо. Химически определенные среды также могли бы быть без какого-либо белка.
Термин «клетка» или «клетка-хозяин» относится к клетке, в которую может может быть введена/трансфицирована или вводится/трансфицируется нуклеиновая кислота, например, кодирующая гетерологичный полипептид. Клетки-хозяева включают и прокариотические клетки, которые используются для размножения векторов/плазмид, и эукариотические клетки, которые используются для экспрессии нуклеиновой кислоты. В одном вопощении эукариотические клетки представляют собой клетки млекопитающих. В другом воплощении клетка-хозяин млекопитающего выбрана из клеток млекопитающих, содержащих клетки СНО (например, СНО-K1 или СНО DG44), клетки BHK, клетки NS0, клетки SP2/0, клетки HEK 293, клетки HEK 293 EBNA, клетки PER.C6 и клетки COS. Для ферментации клетки-хозяина и, таким образом, для экспрессии интересующего полипептида используется культуральная среда. Обычно для экспрессии фармацевтических полипептидов либо в малом масштабе в лаборатории, либо в большом масштабе в производственных процессах, широко используются клетки СНО. Характерные свойства и генетический фон клеток СНО хорошо известны из-за их широкого распространения и применения. Следовательно, клетки СНО одобряются надзорными органами для продуцирования терапевтических белков для применения у людей. В одном воплощении клетка млекопитающего представляет собой клетку СНО. В одном воплощении клетка млекопитающего представляет собой суспензионную линию клеток СНО/линию клеток СНО, растущую в суспензии.
Способ согласно настоящему изобретению подходит для продуцирования секретируемого гетерологичного полипептида в большом масштабе, т.е. промышленно.
Культивирование клетки для продуцирования желательного полипептида в большом масштабе обычно состоит из последовательности индивидуальных культивирований, где все культивирования, за исключением конечного, т.е. крупномасштабного культивирования, т.е. последнего культивирования в последовательности, проводятся до достижения определенной плотности клеток в культуральном сосуде. При достижении заданной плотности клеток всю культуру или ее часть используют для инокуляции следующего культивационного сосуда, который имеет больший объем, вплоть до 100 раз больше объема предыдущей культуры. Все культивирования, которые служат в качестве основы для по меньшей мере одного дальнейшего культивирования в большем объеме, обозначаются как «посевная ферментация» или «посевное культивирование». Только при крупномасштабном культивировании, т.е. при культивировании, которое не предназначено для того, чтобы служить в качестве основы для дальнейшего куьтивирования в большем объеме, которое также обозначается как «главная ферментация», определяется конечный момент культивирования, в зависимости от концентрации продуцированного секретированного гетерологичного иммуноглобулина в культивационной среде или времени культивирования. Термин «крупномасштабное» в том виде, как он используется в данной заявке, обозначает конечное культивирование способа промышленного производства. В одном воплощении крупномасштабное культивирование проводится в объеме по меньшей мере 100 л, в другом воплощении - по меньшей мере 500 л, в другом воплощении - по меньшей мере 1000 л, и вплоть до объема 25000 л. В одном воплощении среда конечного, т.е. крупномасштабного культивирования, не содержит агент для селекции эукариотических клеток.
Термин «деление» в том виде, как он используется в данном документе, также известен как пассирование или субкультура клеток. Он включает перенос малого числа клеток в свежую среду, при этом отделенными клетками завевают новую культуру. В суспензионных культурах малое количество культуры, содержащей несколько клеток, разводят в большем объеме свежей среды.
Термин «титр» в том виде, как он используется в данном документе, относится к общему количеству рекомбинантно экспрессированного полипептида, продуцированного культурой клеток млекопитающего в данном количестве объма среды. Титр типично выражают в единицах миллиграммов полипептида на миллилитр среды.
Термин «ген» обозначает нуклеиновую кислоту, которая представляет собой отрезок, например, на хромосоме или на плазмиде, который может осуществлять экспрессию пептида, полипептида или белка. Помимо кодирующей области, т.е. структурного гена, ген содержит другие функциональные элементы, например, сигнальную последовательность, промотор(ы), интроны и/или терминаторы.
Термин «структурный ген» обозначает область гена без сигнальной последовательности, т.е. кодирующую область.
Термин «экспрессия» в том виде, как он используется в данном документе, относится к транскрипции и/или трансляции, происходящей в клетке. Уровень транскрипции желательного продукта в клетке-хозяине может быть определен на основе количества соответствующей мРНК, которая присутствует в клетке. Например, мРНК, транскрибируемую от выбранной нуклеиновой кислоты, можно количественно определять посредством ПЦР (полимеразная цепная реакция) или посредством нозерн-гибридизации (см. Sambrook, et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press (1989)). Белок, кодируемый выбранной нуклеиновой кислотой, можно количественно определять посредством разных способов, например, посредством ELISA (твердофазный иммуноферментный анализ), посредством анализа биологической активности белка или посредством применения анализов, которые не зависят от такой активности, таких как вестерн-блоттинг или радиоиммуноанализ, посредством применения антител, которые распознают и связываются с белком (см. Sambrook, et al., выше).
«Полипептид» представляет собой полимер из аминокислотных остатков, соединенных пептидными связями, независимо от того, продуцируется ли он в природе или синтетически. Полипептиды из менее чем примерно 20 аминокислотных остатков могут быть названы «пептидами». Полипептиды, содержащие две или более чем две аминокислотные цепи, или содержащие аминокислотную цепь длиной 100 аминокислот или более, могут называться «белками». Полипептид или белок также могут содержать непептидные компоненты, такие как углеводные группы или ионы металлов. Углеводы и другие непептидные заместители могут быть добавлены к белку клеткой, в которой продуцируется белок, и могут варьировать, в зависимости от типа клеток. Белки и полипептиды определяются в данном документе в показателях структуры их аминокислотного остова; присоединения, такие как углеводные группы, обычно не определяются, но, тем не менее, могут присутствовать. В одном воплощении полипептид представляет собой иммуноглобулин или фрагмент иммуноглобулина, или конъюгат иммуноглобулина. В одном воплощении полипептид представляет собой тяжелую цепь иммуноглобулина или легкую цепь иммуноглобулина, или его фрагмент, гибрид или конъюгат. «Экзогенный» или «неэндогенный» полипептид представляет собой полипептид, который не происходит из используемой клетки-хозяина.
Термин «нуклеиновая кислота» в том виде, как он используется в данном документе, представляет собой полимер, состоящий из индивдуальных нуклеотидов, т.е. полинуклеотид. Он относится к встречающейся в природе или частично или полностью не встречающейся в природе нуклеиновой кислоте, которая, например, кодирует полипептид, который может быть продуцирован рекомбинантно. Нуклеиновая кислота может быть построена из фрагментов ДНК, которые либо выделяются, либо синтезируются химическими способами. Нуклеиновая кислота может быть интегрирована в другую нуклеиновую кислоту, например, в экспрессионной плазмиде или геноме/хромосоме клетки-хозяина. Плазмида включает челночный и экспрессионный векторы. Типично плазмида также будет содержать прокариотический элемент размножения, содержащий репликатор (например, репликатор ColE1) и селектируемый маркер (например, ген устойчивости к ампициллину или тетрациклину) для репликации и селекции вектора в бактерии соответственно.
Термин «иммуноглобулин» обозначает молекулу, содержащую по меньшей мере два полипептида так называемой легкой цепи (легкую цепь) и два полипептида так называемой тяжелой цепи (тяжелую цепь). Каждый из полипептидов тяжелой и легкой цепи содержит вариабельный домен (вариабельную область) (обычно аминоконцевая часть полипептидной цепи), содержащий связывающие области, которые способны взаимодействовать с антигеном. Каждый из полипептидов тяжелой и легкой цепи также содержит константную область (обычно карбоксиконцевая часть). Константная область тяжелой цепи опосредует связывание иммуноглобулина i) с клетками, несущими рецептор Fc гамма (FcγR), такими как фагоцитарные клетки, или ii) с клетками, несущими неонатальный рецептор Fc (FcRn), также известный как рецептор Брамбелла. Она также опосредует связывание с некоторыми факторами, включающими факторы классической системы комплемента, такими как компонент C1q.
Термин «иммуноглобулин» используется в данном документе в самом широком смысле и охватывает разные структуры иммуноглобулинов, включающие моноклональные антитела, поликлональные антитела, мультиспецифичные антитела (например, биспецифичные антитела) и фрагменты иммуноглобулинов, но не ограничивающиеся ими, при условии, что они демонстрируют желательную антигенсвязывающую активность.
В зависимости от аминокислотной последовательности константной области тяжелых цепей иммуноглобулины делятся на разные классы: класс IgA, класс IgD, класс IgE, класс IgG и класс IgM. Некоторые из данных классов дополнительно делятся на подклассы (изотипы), т.е. IgG - на IgG1, IgG2, IgG3 и IgG4, или IgA - на IgA1 и IgA2. Согласно классу, к которому принадлежит иммуноглобулин, константные области тяжелой цепи называются α (IgA), δ (IgD), ε (IgE), γ (IgG) и μ (IgM) соответственно. В одном воплощении иммуноглобулин представляет собой иммуноглобулин класса IgG. В другом воплощении иммуноглобулин имеет человеческую константную область или константную область человеческого происхождения. В другом воплощении иммуноглобулин принадлежит к подклассу IgG4 или подклассу IgG1, IgG2 или IgG3, который модифицирован таким образом, что не может быть выявлено связывание с рецептором Fcγ (например, с FcγRIIIa) и/или с C1q. В одном воплощении иммуноглобулин принадлежит к человеческому подклассу IgG4 или мутировавшему человеческому подклассу IgG1. В одном воплощении иммуноглобулин принадлежит к человеческому подклассу IgG1 с мутациями L234A и L235A. В другом воплощении иммуноглобулин относится к связыванию с рецептором Fey подкласса IgG4 или подкласса IgG1 или IgG2 с мутацией L234, L235 и/или D265, и/или содержит мутацию PVA236. В другом воплощении иммуноглобулин имеет мутацию, выбранную из S228P, L234A, L235A, L235E, SPLE (S228P и L235E) и/или PVA236 (PVA236 означает то, что аминокислотная последовательность ELLG (приведенная однобуквенным кодом аминокислот) от положения аминокслоты 233 до 236 IgG1 или EFLG IgG4 заменена на PVA). В одном воплощении иммуноглобулин принадлежит к подклассу IgG4 и имеет мутацию S228P IgG4, или иммуноглобулин принадлежит к подклассу IgG1 и имеет мутации L234A и L235A.
Вариабельный домен легкой или тяжелой цепи иммуноглобулина, в свою очередь, содержит разные отрезки, т.е. четыре каркасные области (FR) и три гипервариабельные области (CDR).
Термин «фрагмент иммуноглобулина» обозначает полипептид, содержащий по меньшей мере один домен из группы доменов, содержащей вариабельный домен, домен СН1, каркасную область, домен СН2, домен СН3, домен СН4 тяжелой цепи иммуноглобулина, или вариабельный домен, или домен CL легкой цепи иммуноглобулина. Также включены его производные и варианты. Дополнительно может присутствовать вариабельный домен, в котором подвергнуты делеции одна или более чем одна аминокислота или аминокислотная область.
Термин «конъюгат иммуноглобулина» обозначает полипептид, содержащий по меньшей мере один домен тяжелой или легкой цепи иммуноглобулина, конъюгированный посредством пептидной связи с другим полипептидом. Другой полипептид представляет собой неиммуноглобулиновый пептид, такой как гормон, рецептор гормона роста, антифузогенный пептид или тому подобное.
Термин «нелимитирующая концентрация Phe» или «неограничивающая концентрация Phe» в том виде, как он используется в данном документе, означает то, что подпитку фенилаланином осуществляют в избыточных количествах, т.е. что имеет место дополнение культуры, например, 0,6 мМ Phe в сутки, начиная с суток 6 до суток 14. Без дополнительной подпитки Phe, в общем, будет лимитирующим к суткам 10 или 11 («лимитирование по Phe»). Дополнение фенилаланином может добавляться посредством непрерывной подпитки или, в качестве альтернативы, посредством одного или более чем одного индивидуального болюсного впрыскивания маточных растворов Phe в начале или на протяжении процесса ферментации.
Термин «повышенная удельная продуктивность» означает то, что удельная продуктивность соответствующей клетки-хозяина выше при условиях, описанных в данном документе, относительно идентичного способа продуцирования без добавления мета-тирозина. Удельная продуктивность (qP) в качестве меры эффективности продуцирования (количество продуцированного полипептида/белка, например, в пикограммах) клетки в сутки рассчитывается, как отражено в примерах.
ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
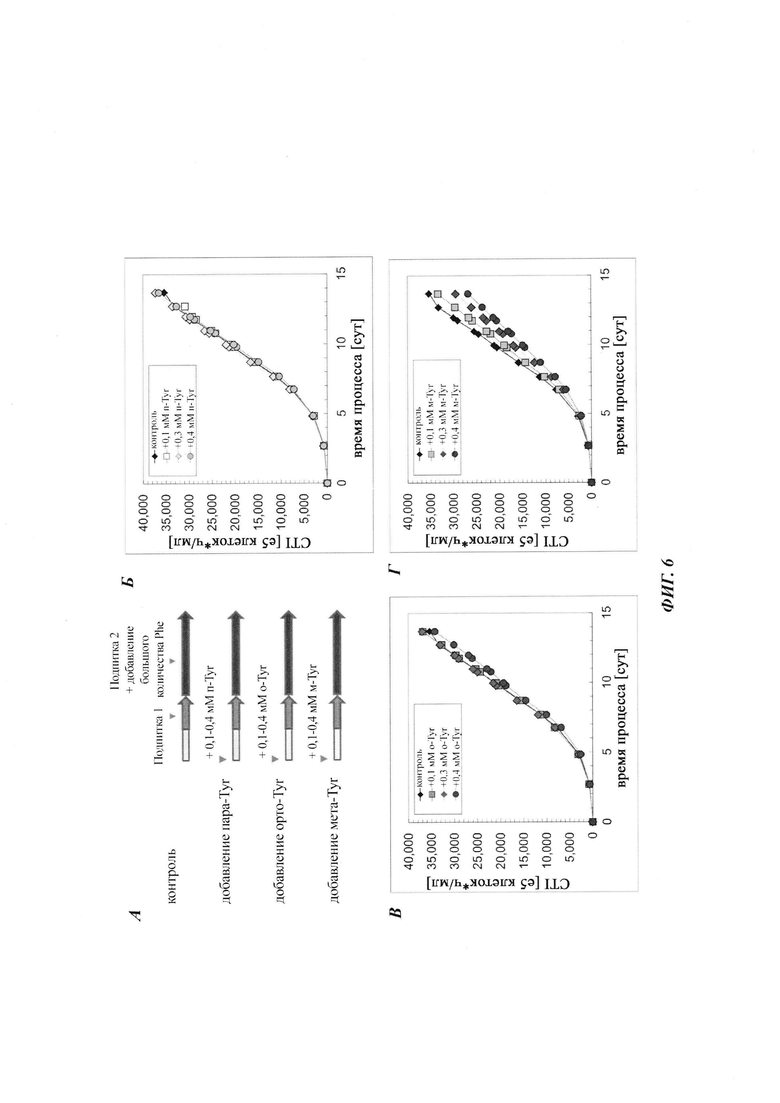
Фиг. 1. Мета-Tyr модулирует генерацию биомассы СНО при условиях лимитирования по Phe. (А) Для способа с подпиткой использовали две последовательные непрерывные подпитки, подпитку 1 и подпитку 2. Подпиточные культуры СНО либо не дополняли (контроль), либо дополняли 0,1 мМ, 0,3 мМ или 0,4 мМ пара-Tyr, орто-Tyr или мета-Tyr в начале процесса. Интеграл числа клеток по времени (CTI) как мера генерации биомассы клеток СНО показан для дополнения пара-Tyr (Б), орто-Tyr (В) или мета-Tyr (Г).
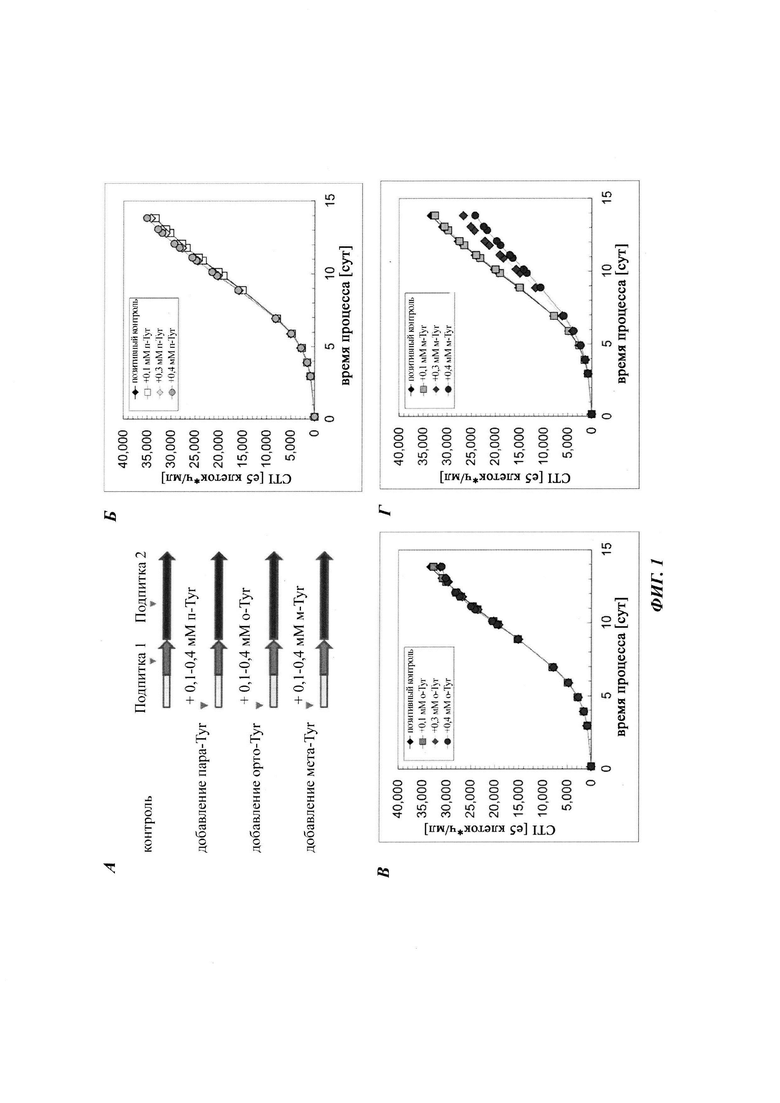
Фиг. 2. Разные роли мета-Tyr и орто-Tyr в регуляции роста клеток СНО при условиях лимитирования по Phe. Плотность жизнеспообных клеток показана для дополнения пара-Tyr (А), орто-Tyr (Б) или мета-Tyr (В). Культивирование с подпиткой без дополнения пара-Tyr, орто-Tyr или мета-Tyr показано в качестве контроля.
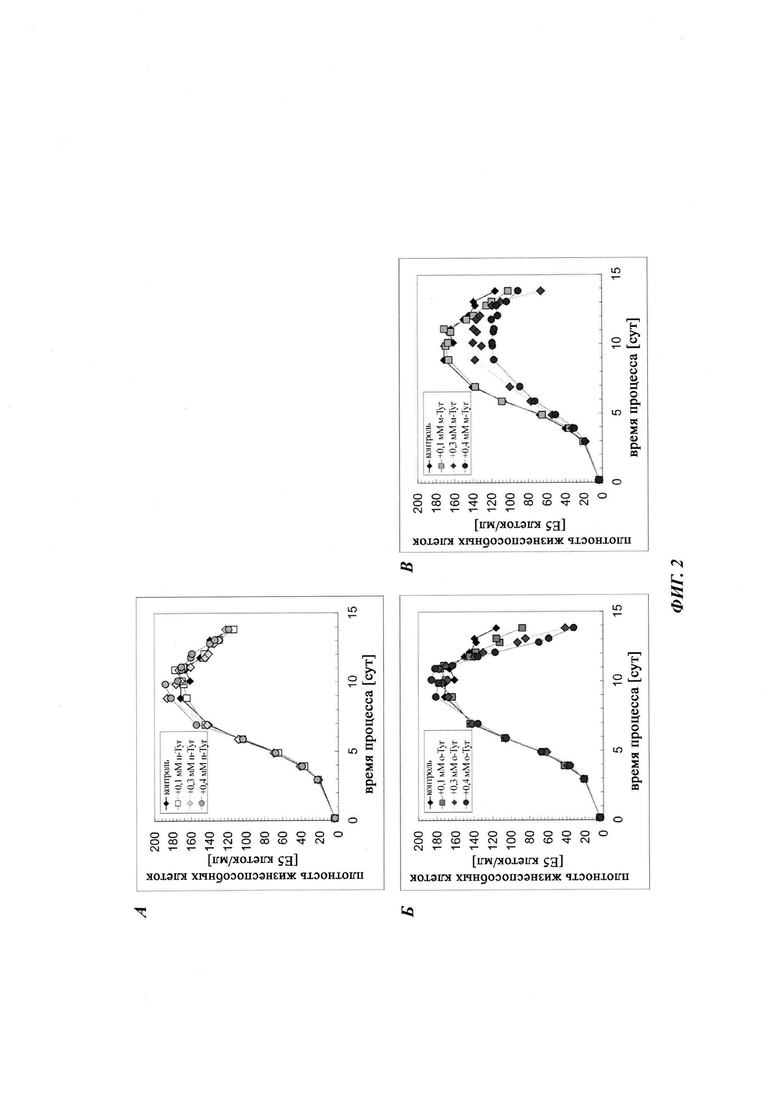
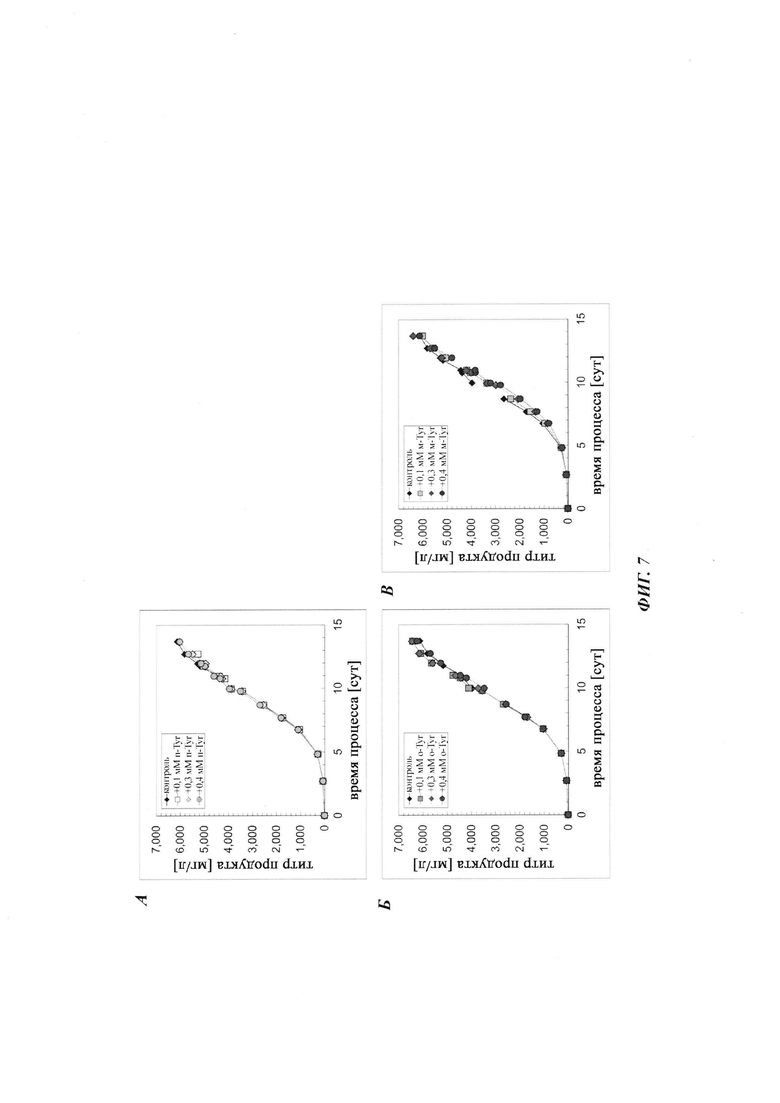
Фиг. 3. Дополнение мета-Tyr и орто-Tyr в культивированиях СНО с подпиткой не изменяет выход продукта при условиях лимитирования по Phe. Концентрация продукта показана для дополнения пара-Tyr (А), орто-Tyr (Б) или мета-Tyr (В). Культивирование с подпиткой без дополнения пара-Tyr, орто-Tyr или мета-Tyr показано в качестве контроля.
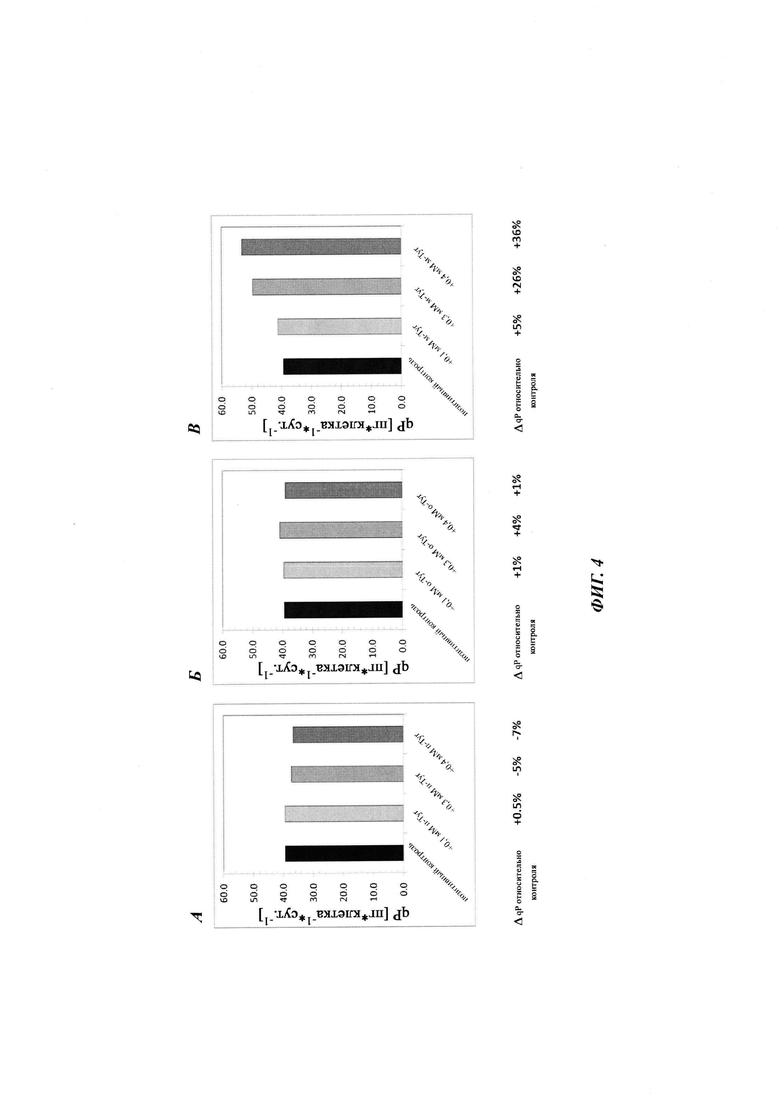
Фиг. 4. Дополнение мета-Tyr увеличивает удельную клеточную скорость образования продукта - qP - при условиях лимитирования по Phe. Удельная клеточная скорость образования продукта - qP - показана для дополнения пара-Tyr (А), орто-Tyr (Б) или мета-Tyr (В). Культивирование с подпиткой без дополнения пара-Tyr, орто-Tyr или мета-Tyr показано в качестве контроля.
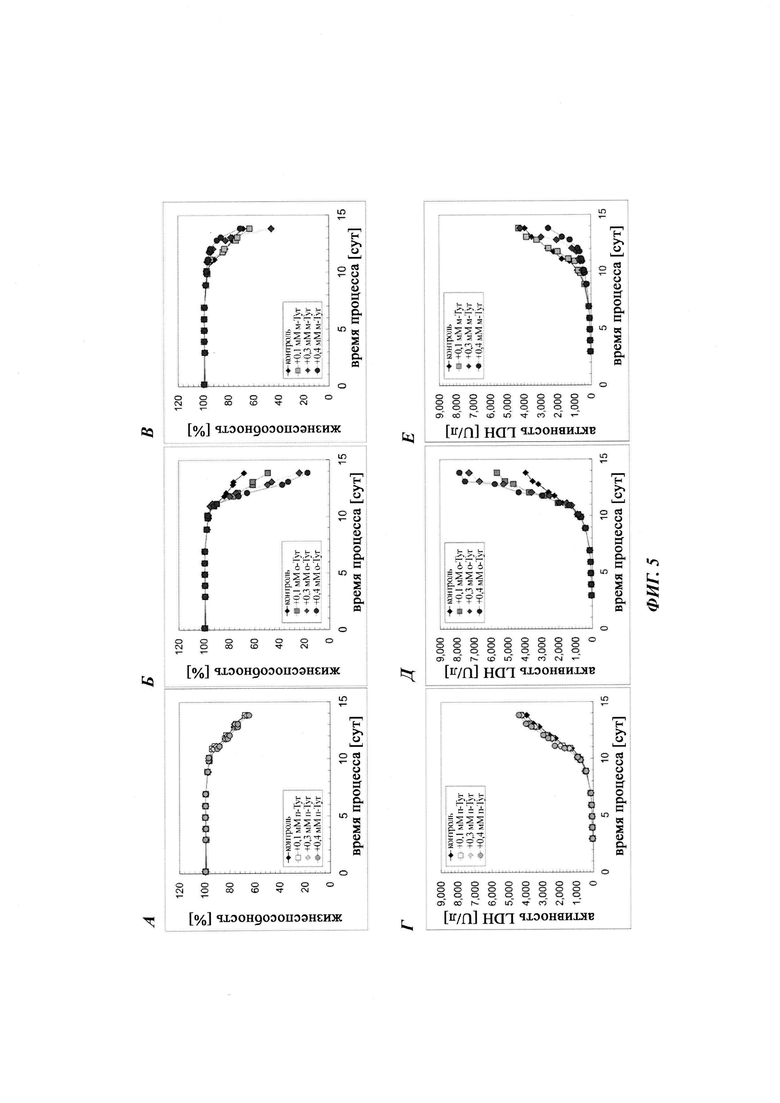
Фиг. 5. Разные роли мета-Tyr и орто-Tyr для жизнеспособности клеток СНО при условиях лимитирования по Phe. Жизнеспособность клеток и активность LDH (лактатдегидрогеназа) в супернатанте показана для дополнения пара-Tyr (А, Г), орто-Tyr (Б, Д) и мета-Tyr (В, Е). Культивирование с подпиткой без дополнения пара-Tyr, орто-Tyr или мета-Tyr показано в качестве контроля.
Фиг. 6. Мета-Tyr модулирует генерацию биомассы СНО при условиях, не лимитирующих по Phe. (А) Для способа с подпиткой использовали две последовательные непрерывные подпитки, подпитку 1 и подпитку 2 с высокой концентрацией Phe. Подпиточные культуры СНО либо не дополняли (контроль), либо дополняли 0,1 мМ, 0,3 мМ или 0,4 мМ пара-Tyr, орто-Tyr или мета-Tyr в начале процесса. Интеграл числа клеток по времени (CTI) как мера генерации биомассы клеток СНО показан для дополнения пара-Tyr (Б), орто-Tyr (В) или мета-Tyr (Г).
Фиг. 7. Дополнение мета-Tyr и орто-Tyr при культивированиях СНО с подпиткой не изменяет выход продукта при условиях, не лимитирующих по Phe. Концентрация продукта показана для дополнения пара-Tyr (А), орто-Tyr (Б) и мета-Tyr (В). Культивирование с подпиткой без дополнения пара-Tyr, орто-Tyr или мета-Tyr показано в качестве контроля.
Фиг. 8. Дополнение мета-Tyr увеличивает удельную клеточную скорость образования продукта - qP - при условиях, не лимитирующих по Phe. Удельная клеточная скорость образования продукта - qP - показана для дополнения пара-Tyr (А), орто-Tyr (Б) или мета-Tyr (В). Культивирование с подпиткой без дополнения пара-Tyr, орто-Tyr или мета-Tyr показано в качестве контроля.
Следующие примеры и графические материалы приведены для того, чтобы помочь пониманию настоящего изобретения, истинный объем которого изложен в приложенной формуле изобретения. Понятно, что в изложенных методиках можно сделать модификации без отступления от сущности изобретения.
Реактивы и материал
DL-орто-тирозин (2-гидрокси-DL-фенилаланин), DL-мета-тирозин (3-гидрокси-DL-фенилаланин), L-пара-тирозин, L-фенилаланин и гуанидиния гидрохлорид приобретали у Sigma-Aldrich (Мюнхен, Германия). L-орто-тирозин (2-гидрокси-L-фенилаланин) и L-мета-тирозин (3-гидрокси-L-фенилаланин) приобретали у RSP Amino Acids, LLC (Shirley, MA, США). Все другие реактивы приобретали у Merck (Дармштадт, Германия) и Sigma-Aldrich (Мюнхен, Германия).
Клетки и культивирование клеток
Для всех экспериментов с культурой клеток использовали рекомбинантную клеточную линию СНО-K1, названную клон 1, экспрессирующую гуманизированное моноклональное антитело. Рекомбинантная клеточная линия СНО-K1 была получена с использованием клеточной линии-хозяина СНО-K1, чувствительной к L-метионина сульфоксимину (Lonza, Кельн, Германия). Данные клетки культивировали в химически определенной среде CD-CHO, не содержащей белок (Life Technologies, Дармштадт, Германия), дополненной 50 мкМ L-метионина сульфоксимином (Sigma-Aldrich, Мюнхен, Германия), на протяжении посевного культивирования. Посевное культивирование проводили во встряхиваемых колбах с использованием увлажненного инкубатора, 7% CO2 и установки на 37°С. Клетки делили каждые трое-четверо суток для субкультивирования и увеличения объема культуры. Для всех экспериментов использовали клетки с идентичным возрастом в культуре (приблизительно 21 поколение) до начала эсперимента.
Эксперименты по дополнению мета-Tyr и орто-Tyr
Все эксперименты по дополнению проводили с использованием систем культивирования во встряхиваемых колбах, химически определенной среды CD-СНО без селективного давления L-метионина сульфоксимина и двух подходящих применяемых последовательных непрерывных подпиток (подпитки 1 и подпитки 2). Рекомбинантные клетки СНО-K1, продуцирующие гуманизированное моноклональное антитело, инокулировали с использованием 3×105 жизнеспособных клеток/мл и культивировали в течение 14 суток. Базовую среду CD-CHO дополняли в стерильных условиях перед инокуляцией 0,1 мМ, 0,3 мМ или 0,4 мМ орто-Tyr, мета-Tyr или пара-Tyr. Контрольное культивирование без дополнения орто-Tyr, мета-Tyr или пара-Tyr использовали в качестве эталона. Условия, не лимитирующие по фенилаланину (Phe), осуществляли посредством увеличения концентрации Phe в подпитке 2.
Плотности жизнеспособных клеток, жизнеспособность и интеграл количества клеток по времени
Для анализа плотности жизнеспособных клеток и общей плотности клеток использовали автоматизированную систему Cedex HiRes (Roche Diagnostics, Мангейм, Германия). Различение плотности жизнеспособных клеток и общей плотности клеток осуществляли с использованием способа окрашивания с исключением трипанового синего и анализа более чем 10 изображений на образец и сутки согласно спецификациям изготовителя. Плотность жизнеспособных клеток (VCD) и жизнеспособность клеток рассчитывали, как описано в уравнении 1 (Ур. 1) и уравнении 2 (Ур. 2) соответственно.
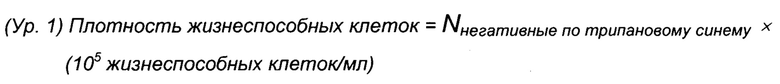
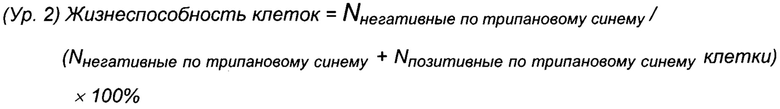
В качестве индикатора общей генерации биомассы в способе рассчитывали интеграл совокупного числа клеток по времени (CTI) (Ур. 3).
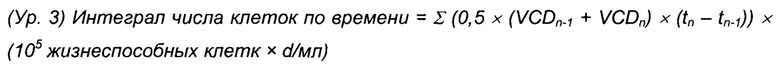
Активность лактатдегидрогеназы (LDH) в бесклеточном супернатанте анализировали с использованием системы Cobas Integra 400 plus (Roche Diagnostics, Мангейм, Германия).
Количественное измерение титра IgG и расчет qP
Титр продукта количественно измеряли либо системой Cobas Integra 400 plus (Roche, Мангейм, Германия) согласно протоколу изготовителя, либо способом ВЭЖХ (высокоэффективная жидкостная хроматография) PorosA, как описано ранее (Zeck et al. 2012). Для анализа эффективности клеточной продукции рассчитывали общую удельную продуктивость qP согласно уравнению 4.
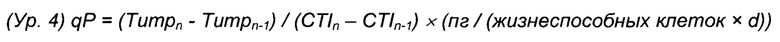
Определение вариантов последовательности пептидов и идентификация вариантов последовательности с мета- и орто-Tyr с использованием синтетических пептидов
Количественное измерение вариантов последовательности пептидов осуществляли, как описано ранее (Zeck et al. 2012). Вкратце, образцы антител (250 мкг) денатурировали добавлением денатурирующего буфера (0,4 М Tris, 8 М гуанидиния гидрохлорид, рН 8) до конечного объема 240 мкл. Восстановление достигалось добавлением 20 мкл 0,24 М DTT (дитиотрейтол), свежеприготовленного в денатурирующем буфере, и инкубированием при 37°С в течение 60 мин. Затем образец алкилировали добавлением 20 мкл 0,6 М йодуксусной кислоты в воде в течение 15 мин при комнатной температуре в темноте. Избыток алкилирующего реактива инактвировали добавлением 30 мкл раствора DTT. В образце затем заменяли буфер на приблизительно 480 мкл 50 мМ Tris/HCl, рН 7,5, с использованием колонок уровня качества для ДНК NAP 5 Sephadex G-25 (GE Healthcare, Мюнхен, Германия). Расщепление проводили трипсином в течение 5 ч при 37°С (отношение 1:37). Полученную смесь пептидов инъецировали и разделяли без предобработки с использованием ВЭЖХ с обращенной фазой (Agilent 1100 Cap LC, Agilent Technologies, Бёблинген, Германия). Для разделения использовали С18-эфирную колонку Polaris 3 (1×250 мм; диаметр частиц 3 мкм, размер пор 180  ) от Varian (Дармштадт, Германия). Растворителями были 0,1%-ная муравьиная кислота в воде (А) и в ацетонитриле (Б) (Sigma Aldrich, Мюнхен, Германия). Линейный градиент от 2 до 38% Б прогоняли за 80 мин при 37°С. Элюат ВЭЖХ делили с использованием Triversa NanoMate (Advion, Ithaca, NY), и 380 нл/мин инфундировали в классический тандемный масс-спектрометр LTQ Orbitrap (Thermo Fisher Scientific, Драйайх, Германия), работающий в режиме положительных ионов. Для подтверждения пиков орто- и мета-Tyr в выделенных хроматограммах ионов авторы изобретения использовали синтетические пептиды mAb НС66-72 DQFTISR (немодифицированный), DQpYTISR, DQmYTISR и DQoYTISR. Синтетические пептиды приобретали у Biosyntan GmbH (Берлин, Германия).
) от Varian (Дармштадт, Германия). Растворителями были 0,1%-ная муравьиная кислота в воде (А) и в ацетонитриле (Б) (Sigma Aldrich, Мюнхен, Германия). Линейный градиент от 2 до 38% Б прогоняли за 80 мин при 37°С. Элюат ВЭЖХ делили с использованием Triversa NanoMate (Advion, Ithaca, NY), и 380 нл/мин инфундировали в классический тандемный масс-спектрометр LTQ Orbitrap (Thermo Fisher Scientific, Драйайх, Германия), работающий в режиме положительных ионов. Для подтверждения пиков орто- и мета-Tyr в выделенных хроматограммах ионов авторы изобретения использовали синтетические пептиды mAb НС66-72 DQFTISR (немодифицированный), DQpYTISR, DQmYTISR и DQoYTISR. Синтетические пептиды приобретали у Biosyntan GmbH (Берлин, Германия).
Расчет штрафных коэффициентов и суррогатных маркеров для прогнозирования вариантов последовательности с заменой Phe на орто-Tyr и Phe на мета-Tyr
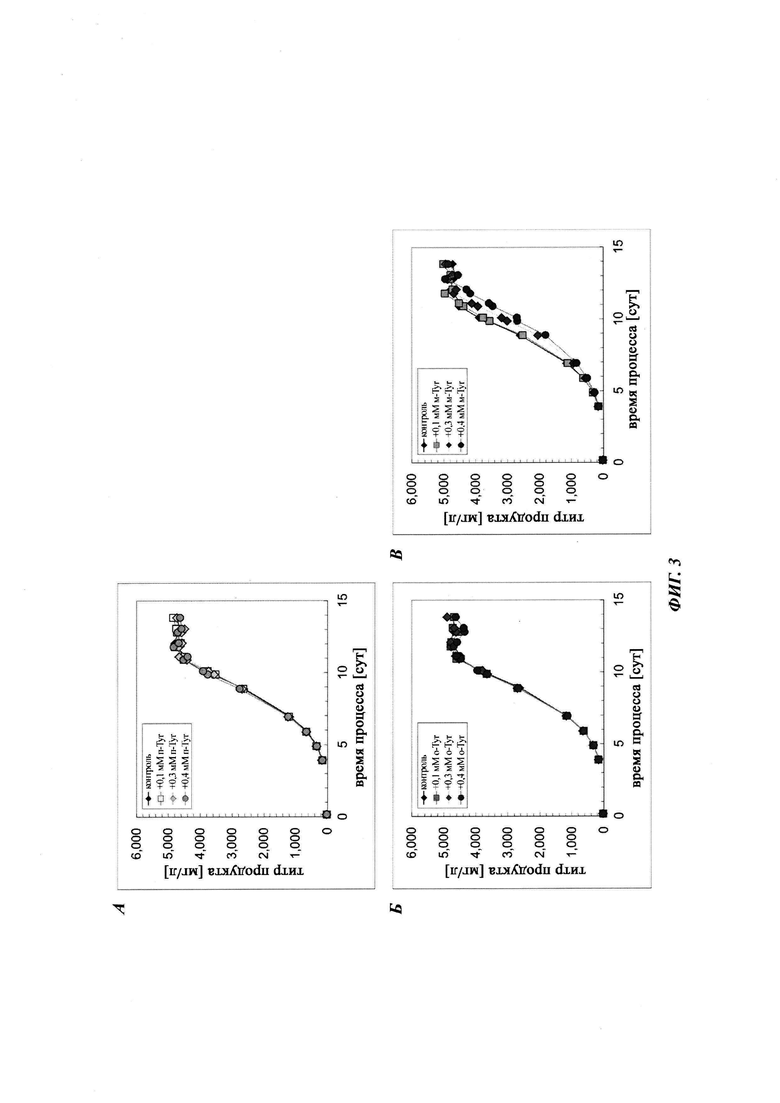
Авторы изобретения предположили, что включение мета- и/или орто-Tyr вместо Phe может быть описано упрощенной моделью, в которой предполагается, что за использование мета- и/или орто-Tyr вместо Phe накладывается штраф. Данный штрафной коэффициент может возникать из разных источников, таких как лучший транспорт L-Phe в клетки и/или механизм редактирования во время синтеза белка, который старается предотвратить использование мета- и/или орто-Tyr. Данное предположение приводит к уравнению:
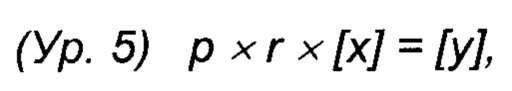
где р представляет собой штрафной коэффициент, r представляет собой отношение средних концентраций мета- или орто-Tyr к Phe (на протяжении данного интервала времени), [х] представляет собой концентрацию продуцированного белка (за интервал времени), и [у] представляет собой концентрацию продуцированного белка, который имеет вариант последовательности. Заметьте, что данная модель не включает каких-либо зависимостей от времени, стадии процесса, отношений концентраций Phe и мета- или орто-Tyr, которые приводят, например, к фазовым сдвигам. Расчет штрафного коэффициента является прямым (Ур. 6).
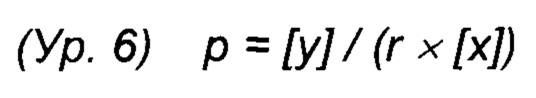
Аналогично, зная штрафной коэффициент, также возможно рассчитать отношения концентраций мета- или орто-Tyr к Phe для определения желательной процентной доли продукта без вариантов поледовательности.
Пример 1
Эффекты добавления мета-тирозина на модуляцию удельной продуктивности (qP) при условиях лимитирования по фенилаланину
Ранее Gurer-Orhan et al. сообщали о том, что добавление к клеткам СНО мета-Tyr демонстрировало дозозависимую клеточную цитотоксичность. В скринингах концентрации наблюдали 50%-ное уменьшение способности клеток СНО к восстановлению МТХ (метотрексат) при добавлении 0,5 мМ мета-Tyr (Gurer-Orhan et al., (2006)). До настоящего времени не было приведено данных или концентрации для добавления орто-Tyr в культуры клеток. С использованием подхода дозозависимого культивирования была поставлена цель определить релевантность и переносимые концентрации мета-Tyr и орто-Tyr для эффективности роста клеток СНО. Для этого модель культивирования СНО, описанную в материалах и методах, дополняли 0,1 мМ, 0,3 мМ или 0,4 мМ пара-Tyr, орто-Tyr или мета-Tyr. Авторы изобретения использовали стандартный способ культивирования без добавления в качестве эталона после так называемого «контроля» или «позитивного контроля». В данном случае Phe будет лимитирущим к суткам 10/11.
В первом подходе анализировали влияние мета- и орто-Tyr на маркеры макроскопического роста клеток, плотность жизнеспособных клеток (VCD), жизнеспособность клеток и интеграл количества клеток по времени (CTI) в качестве маркера общей продукции биомассы. В сутки 9/10 все протестированные культуры, за исключением культуры с добавлением мета-Tyr, достигали максимальной VCD приблизительно 180×105 клеток/мл, тогда как клон 1 СНО, обработанный мета-Tyr, демонстрировал дозозависимую пониженную максимальную VCD (Фиг. 2). Наблюдали пониженный совокупный CTI для культур СНО, дополненных мета-Tyr (Фиг. 1).
В отличие от данных, опубликованных ранее Gurer-Orhan et al., не наблюдали значимого влияния добавления мета-Tyr на жизнеспособность клеток в модели культивирования СНО с подпиткой авторов изобретения. Однако дополнение орто-Tyr демонстрировало меньшую жизнеспособность клеток - меньше чем 60% в сутки 14, и более высокую конечную активность LDH в супернатанте по сравнению с контролем и мета-Tyr, а также добавлением пара-Tyr (Фиг. 5).
Общая продуктивность культур, определенная посредством анализа концентрации продукта, не выявила различий между тестируемыми вариантами (Фиг. 3). Все культуры демонстрировали стагнацию титра, начиная с суток 11/12. Дополнительно, один мета-Tyr демонстрировал в целом более высокую удельную продуктивность qP (Фиг. 4). Добавление 0,1 мМ, 0,3 мМ и 0,4 мМ мета-Tyr при условиях лимитирования по Phe увеличивало qP по сравнению с контролем на +5%, +26% и +36% соответственно.
Пример 2
Эффекты добавления мета-тирозина на модуляцию удельной продуктивности (qP) при условиях, не лимитирующих по фенилаланину
Во втором подходе анализировали роль добавления мета- и орто-Tyr при условиях, не лимитирующих по Phe, в культивированиях СНО с подпиткой. Для этого для предотвращения лимитирования по Phe увеличивали количество Phe в подпитке 2 (Фиг. 6).
Вновь все протестированные культуры, за исключением культуры с добавлением мета-Tyr, достигали аналогичных CTI приблизительно от 35000 до 40000×105 клеток-ч/мл, тогда как клон 1 СНО, обработанный мета-Tyr, демонстрировал дозозависимое уменьшение CTI (Фиг. 6). Для всех протестированных условий не наблюдали различий в титре продукта (Фиг. 7). Однако, по сравнению с условиями лимитирования по Phe, описанными выше, предоставление культурам СНО достаточного количества Phe предотвращает стагнацию титра продукта. Дополнение мета-Tyr демонстрировало общую более высокую удельную продуктивность qP (Фиг. 4), даже при условиях, не лимитирующих по Phe (Фиг. 8). Добавление 0,1 мМ, 0,3 мМ и 0,4 мМ мета-Tyr при условиях, не лимитирующих по Phe, увеличивало qP по сравнению с контролем на +3%, +26% и +28% соответственно.
Список ссылок
Leader В, Baca QJ, Golan DE. 2008. Protein therapeutics: a summary and pharmacological classification. Nat Rev Drug Discov 7:21-39.
Wakankar AA, Borchardt RT. 2006. Formulation considerations for proteins susceptible to asparagine deamidation and aspartate isomerization. J Pharm Sci 95:2321-2336.
Diepold K, Bomans K, Wiedmann M, Zimmermann B, Petzold A, Schlothauer T, Mueller R, Moritz B, Stracke JO, Molhoj M, Reusch D, Bulau P. 2012. Simultaneous assessment of Asp isomerization and Asn deamidation in recombinant antibodies by LC-MS following incubation at elevated temperatures. PLoS One 7:e30295.
Dengl S, Wehmer M, Hesse F, Lipsmeier F, Popp O, Lang K. 2013. Aggregation and chemical modification of monoclonal antibodies under upstream processing conditions. Pharm Res 30:1380-1399.
Li S, Schoneich C, Borchardt RT. 1995. Chemical instability of protein pharmaceuticals: Mechanisms of oxidation and strategies for stabilization. Biotechnol Bioeng 48:490-500.
Ji JA, Zhang B, Cheng W, Wang YJ. 2009. Methionine, tryptophan, and histidine oxidation in a model protein, PTH: mechanisms and stabilization. J Pharm Sci 98:4485-4500.
Hensel M, Steurer R, Fichtl J, Elger C, Wedekind F, Petzold A, Schlothauer T, Molhoj M, Reusch D, Bulau P. 2011. Identification of potential sites for tryptophan oxidation in recombinant antibodies using tert-butylhydroperoxide and quantitative LC-MS. PLoS One 6:e17708.
Khetan A, Huang YM, Dolnikova J, Pederson NE, Wen D, Yusuf-Makagiansar H, Chen P, Ryll T. 2010. Control of misincorporation of serine for asparagine during antibody production using CHO cells. Biotechnol Bioeng 107:116-123.
Wen D, Vecchi MM, Gu S, Su L, Dolnikova J, Huang YM, Foley SF, Garber E, Pederson N, Meier W. 2009. Discovery and investigation of misincorporation of serine at asparagine positions in recombinant proteins expressed in Chinese hamster ovary cells. J Biol Chem 284:32686-32694.
Feeney L, Carvalhal V, Yu XC, Chan B, Michels DA, Wang YJ, Shen A, Ressl J, Dusel B, Laird MW. 2013. Eliminating tyrosine sequence variants in CHO cell lines producing recombinant monoclonal antibodies. Biotechnol Bioeng 110:1087-1097.
Bridges BA. 2001. Hypermutation in bacteria and other cellular systems. Philos Trans R Soc Lond В Biol Sci 356:29-39.
Zeck A, , Regula JT, Larraillet V, Mautz В, Popp O, Gopfert U, Wiegeshoff F, Vollertsen UE, Gorr IHKoll H, Papadimitriou A. 2012. Low level sequence variant analysis of recombinant proteins: an optimized approach. PLoS One 7:-e40328.
Jakubowski H. 2001. Translational accuracy of aminoacyl-tRNA synthetases: implications for atherosclerosis. J Nutr 131.2983S-2987S.
Gurer-Orhan H, Ercal N, Mare S, Pennathur S, Orhan H, Heinecke JW. 2006. Misincorporation of free m-tyrosine into cellular proteins: a potential cytotoxic mechanism for oxidized amino acids. Biochem J 395:277-284.
Hossler P, Khattak SF, Li ZJ. 2009. Optimal and consistent protein glycosylation in mammalian cell culture. Glycobiology 19:936-949.
Lee S, Park JR, Seo MS, Roh KH, Park SB, Hwang JW, Sun B, Seo K, Lee YS, Kang SK, Jung JW, Kang KS. 2009. Histone deacetylase inhibitors decrease proliferation potential and multilineage differentiation capability of human mesenchymal stem cells. Cell Prolif 42:711-720.
Murray-Beaulieu V, Hisiger S, Durand C, Perrier M, Jolicoeur M. 2009. Na-butyrate sustains energetic states of metabolism and t-PA productivity of CHO cells. J Biosci Bioeng 108:160-167.
Hendrick V, Winnepenninckx P, Abdelkafi C, Vandeputte O, Cherlet M, Marique T, Renemann G, Loa A, Kretzmer G, Werenne J. 2001. Increased productivity of recombinant tissular plasminogen activator (t-PA) by butyrate and shift of temperature: a cell cycle phases analysis. Cytotechnology 36:71-83.
Yoon SK, Choi SL, Song JY, Lee GM. 2005. Effect of culture pH on erythropoietin production by Chinese hamster ovary cells grown in suspension at 32.5 and 37.0 degrees C. Biotechnol Bioeng 89:345-356.
Han YK, Koo TY, Lee GM. 2009. Enhanced interferon-beta production by CHO cells through elevated osmolality and reduced culture temperature. Biotechnol Prog 25:1440-1447.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ РЕКОМБИНАНТНОГО ПОЛИПЕПТИДА | 2011 |
|
RU2577972C2 |
| СПОСОБ ПОЛУЧЕНИЯ ПОЛИПЕПТИДОВ | 2015 |
|
RU2699715C2 |
| УЛУЧШЕННАЯ СРЕДА ДЛЯ КУЛЬТИВИРОВАНИЯ КЛЕТОК | 2011 |
|
RU2798069C2 |
| НОВЫЕ ЭУКАРИОТИЧЕСКИЕ КЛЕТКИ И СПОСОБЫ ИХ ПОЛУЧЕНИЯ ДЛЯ РЕКОМБИНАНТНОЙ ЭКСПРЕССИИ ПРЕДСТАВЛЯЮЩЕГО ИНТЕРЕС ПРОДУКТА | 2014 |
|
RU2720525C2 |
| Комплекс аналога инсулина со сниженной аффинностью к инсулиновому рецептору и его применение | 2018 |
|
RU2779462C2 |
| ПРИМЕНЕНИЕ НИЗКОЙ ТЕМПЕРАТУРЫ И/ИЛИ НИЗКОГО pН В КУЛЬТУРЕ КЛЕТОК | 2008 |
|
RU2478702C2 |
| СРЕДА ДЛЯ КУЛЬТИВИРОВАНИЯ КЛЕТОК | 2011 |
|
RU2644651C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА, СПЕЦИФИЧНЫЕ К АЛЬФА-ЭНОЛАЗЕ, И СПОСОБЫ ПРИМЕНЕНИЯ В ПРОТИВООПУХОЛЕВОЙ ТЕРАПИИ | 2014 |
|
RU2761662C2 |
| СПОСОБ СНИЖЕНИЯ ГЕТЕРОГЕННОСТИ АНТИТЕЛ И СПОСОБ ПОЛУЧЕНИЯ СООТВЕТСТВУЮЩИХ АНТИТЕЛ | 2012 |
|
RU2580020C2 |
| НОВЫЙ ВАРИАНТ ГАММА-АМИНОБУТИРАТПЕРМЕАЗЫ И СПОСОБ ПОЛУЧЕНИЯ ИЗОЛЕЙЦИНА С ЕГО ИСПОЛЬЗОВАНИЕМ | 2021 |
|
RU2826168C1 |

Изобретение относится к области биотехнологии, а именно к применению мета-тирозина для увеличения удельной продуктивности, представляющей собой количество продуцированного полипептида в сутки, клетки яичника китайского хомяка CHO-K1, которая продуцирует экзогенный полипептид, а также к продуцированию экзогенного полипептида в CHO-K1 и увеличению удельной продуктивности в CHO-K1. Способ включает культивирование CHO-K1 в культуральной среде, содержащей мета-тирозин. Изобретение позволяет увеличить выход экзогенного полипептида. 3 н. и 10 з.п. ф-лы, 8 ил., 1 табл., 2 пр.
1. Применение мета-тирозина для увеличения удельной продуктивности клетки яичника китайского хомяка (СНО), которая продуцирует экзогенный полипептид, где
удельная продуктивность представляет собой количество продуцированного полипептида в сутки, а
СНО представляет собой CHO-K1.
2. Применение по п. 1, в котором клетка СНО представляет собой суспензионную клетку СНО.
3. Применение по п. 1, в котором мета-тирозин добавляют с получением концентрации от 0,2 мМ до 0,7 мМ.
4. Применение по п. 1, где применение осуществляется в культуральной среде, которая дополнительно содержит фенилаланин в нелимитирующей концентрации.
5. Применение по п. 1, в котором молярное отношение мета-тирозин/фенилаланин меньше чем или равно 1,25.
6. Применение по любому из пп. 1-5, в котором полипептид представляет собой иммуноглобулин, или его вариант, или его фрагмент, или его гибрид.
7. Способ продуцирования экзогенного полипептида в эукариотической клетке-хозяине, экспрессирующей нуклеиновую кислоту, кодирующую этот полипептид, включающий культивирование эукариотической клетки-хозяина в культуральной среде, содержащей мета-тирозин, где
эукариотическая клетка-хозяин представляет собой клетку яичника китайского хомячка CHO-K1.
8. Способ по п. 7, в котором мета-тирозин добавляют с получением концентрации от 0,2 мМ до 0,7 мМ.
9. Способ по п. 7, в котором полипептид представляет собой иммуноглобулин, или его вариант, или его фрагмент, или его гибрид.
10. Способ по п. 7, в котором культуральная среда дополнительно содержит фенилаланин в нелимитирующей концентрации.
11. Способ по п. 7, в котором молярное отношение мета-тирозин/фенилаланин меньше чем или равно 1,25.
12. Способ по п. 7-11, в котором на протяжении данного способа поддерживается постоянная температура.
13. Способ увеличения удельной продуктивности эукариотической клетки-хозяина, которая продуцирует экзогенный полипептид, при котором в культуральную среду добавляют мета-тирозин, где
удельная продуктивность представляет собой количество продуцированного полипептида в сутки, а
эукариотическая клетка-хозяин представляет собой клетку яичника китайского хомячка CHO-K1.
| GURER-ORHAN H., et al | |||
| "Misincorporation of free m-tyrosine into cellular proteins: a potential cytotoxic mechanism for oxidized amino acids", Biochem J | |||
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| СПОСОБ ПОЛУЧЕНИЯ КЛЕТКИ, СПОСОБНОЙ ПРОДУЦИРОВАТЬ ГЕТЕРОПРОТЕИНЫ С ВЫСОКИМ ВЫХОДОМ | 2008 |
|
RU2486245C2 |
| КЛЕТКА ДЛЯ ПОЛУЧЕНИЯ ГЕТЕРОПРОТЕИНОВ И СПОСОБ ПОЛУЧЕНИЯ НА ЕЕ ОСНОВЕ | 2008 |
|
RU2494148C2 |
| СПОСОБ И УСТРОЙСТВО ДЛЯ ЗАПИСИ И ВОСПРОИЗВЕДЕНИЯ ВИДЕОДАННЫХ И ИНФОРМАЦИОННЫЙ НОСИТЕЛЬ ДАННЫХ, НА КОТОРОМ ЗАПИСЫВАЮТСЯ ВИДЕОДАННЫЕ ПОСРЕДСТВОМ ЭТОГО СПОСОБА И УСТРОЙСТВА | 2002 |
|
RU2266625C2 |

