Область техники
Изобретение относится к области генетики и молекулярной биологии клетки, в частности к молекулярной диагностике процессов, происходящих в концевых областях теломер хромосом в ходе клеточного цикла. Заявленный способ используется для построения профилей терминальных нуклеотидов концевых областей G-цепи теломерной ДНК человека, отображающих их количественное процентное соотношение. Полученные профили являются пролиферативными маркерами, указывающими на активацию или угнетение процессов деления клетки и контролирующими продолжительность клеточного цикла.
Изобретение является новой разновидностью ДНК-диагностики пролиферативного статуса клеток, тканей и организма в целом. Способ может найти применение в диагностике и лечении онкологических заболеваний, в определении и контролировании иммунного статуса организма человека, в геронтологии, косметологии, дерматологии, трансплантологии, клеточной биотехнологии и других областях, где требуется контролирование и направленное воздействие на деление клетки.
Уровень техники
Объектом исследования данного изобретения служат концевые однонитевые участки G-цепи теломерной ДНК, именуемые «оверхенгами» (от англ. overhang «довесок»), так как они вызывают наибольший интерес в вопросах, связанных с делением клетки. До настоящего времени известны два способа идентификации терминальных нуклеотидов G-оверхенгов теломерной ДНК на основе использования принципа лигирования. Эти способы имеют существенные методологические ограничения, связанные с невозможностью прямого лигирования к свободному одноцепочечному 3'-концу оверхенга.
В 2001 году для исследования этого вопроса у простейшего организма Tetrahymena thermophila был предложен метод Ligation-mediated primer extension protocol, который позволял измерять длину и нуклеотидные окончания G-оверхенгов, содержащих теломерные повторы TTGGGG. Основные сведения приводятся в статье: Naduparambil К. Jacob, Rose Skopp and Carolyn M. Price. G-overhang dynamics at Tetrahymena telomeres. The EMBO Journal, Vol.20, No.15, 2001, pp. 4299-4308. Из-за невозможности прямого лигирования к одноцепочечному 3'-концу оверхенга в основе этого метода лежит лигирование с универсальной последовательностью Unique oligo, которая предварительно гибридизуется с комплементарной областью Guide oligo последовательности. Авторами статьи были разработаны шесть вариантов Guide oligos последовательностей, меченных с помощью 32P и содержащих помимо комплементарной Unique oligo области, пятинуклеотидный участок, соответствующий возможному варианту окончания G-оверхенга.
Согласно протоколу исследований сначала смешивались Unique oligo и Guide oligo последовательности в эквимолярной концентрации. Затем полученные дуплексы, несущие одноцепочечный участок из 5 нуклеотидов одного из шести вариантов теломерного повтора, добавляли к анализируемому образцу и осуществляли лигирование с помощью Т4 DNA лигазы. Лигирование проводили при 16°C в течение ночи. После удаления непролигированных дуплексов осуществляли процедуру достройки 3'-конца Guide oligo последовательности по всей длине матрицы оверхенга с помощью Т4 DNA полимеразы. Критерием присутствия того или иного варианта окончания оверхенга являлось наличие соответствующих фрагментов меченной ДНК, которые могли синтезироваться только при условии лигирования и полного соответствия 5'-конца Unique oligo последовательности и 3'-конца оверхенга. Присутствие полученных фрагментов оценивали по наличию метки в полиакриламидном геле.
Критическим моментом для достижения эффективности лигирования, а следовательно, и успешности рассматриваемого анализа, является образование ник-подобной (nick-like) структуры между 5'-концом Unique oligo последовательности и 3'-концом оверхенга, которая сшивается с помощью лигазы. Для образования ника необходимо, чтобы пятинуклеотидный участок Guide oligo последовательности, комплементарный теломерному повтору оверхенга, отжегся на самом его конце. Несмотря на короткие размеры G-оверхенгов Tetrahymena от 14 до 27 н.о., не исключена вероятность отжига левее терминального сайта праймирования. Последний фактор, учитывая пространственную двухцепочечную конфигурацию дуплекса, образованного Unique oligo и Guide oligo последовательностями, может мешать дальнейшему отжигу теломерного пятинуклеотидного участка на расположенном правее терминальном сайте праймирования матрицы оверхенга. Эта зависимость эффективности метода от особенностей отжига и лигирования является главным ограничивающим фактором этого метода.
Единственным способом, использовавшимся для идентификации терминальных нуклеотидов хромосом человека, содержащих теломерные повторы TTAGGG, является метод Single Telomere length analysis (STELA), предложенный Agnel J. Sfeir et al. (2005, 2006). Основные сведения приводятся в работе: Agnel J. Sfeir, Weihang Chai, Jerry W. Shay and Woodring E. Wright. Telomere-End Processing: the Terminal Nucleotides of Human Chromosomes. Molecular Cell, Vol.18, 2005, pp. 131-138.
Авторами статьи предложены методы, позволяющие исследовать терминальные нуклеотиды концов хромосом для обеих цепей ДНК. Для изучения терминальных нуклеотидов C-богатой цепи использовался метод Primer-Ligation Assay, а также модификация метода Single Telomere length analysis (C strand STELA), которые основаны на принципе лигирования к свободному 5'-концу C-цепи. Большую трудность вызывает исследование G-богатой цепи теломерной ДНК, так как непосредственное лигирование к одноцепочечному 3'-концу невозможно. В связи с этим в статье была предложена соответствующая модификация STELA метода, предназначенная для целей исследования G-богатой цепи теломерной ДНК (G strand STELA). Этот способ является самым близким аналогом заявленного изобретения.
В рассматриваемом методе из-за невозможности непосредственного лигирования к одноцепочечному 3'-концу оверхенга так же, как и в предыдущем способе, предложена процедура отжига искусственной последовательности Guide platform [3'-AATCCC-5']10 на матрице комплементарной G-богатой цепи. Но в отличие от способа предложенного Jacob et al., отжиг платформы длиной 60 нуклеотидов на матрице 3'-оверхенга проходил сразу на первом этапе до гибридизации с другими дополнительными последовательностями. По мнению авторов это обеспечивало образование искусственных 5'-оверхенгов, которые позволяли произвести дальнейшее лигирование синтетических 5'-фосфорилированных кассет (G telorettes) непосредственно к 3'-концу оверхенга.
Каждая кассета включала участок длиной 7 н.о., соответствующий одному из шести вариантов теломерных окончаний, и последовательность, комплементарную универсальному обратному G teltail праймеру. Лигирование каждой из шести вариантов кассет осуществлялось только при полном соответствии ее 5'-конца и 3'-конца исследуемой G-богатой цепи. Для последующей амплификации использовался обратный G teltail праймер и прямой универсальный subtelomeric праймер, специфичный для участка субтеломерной области хромосом.
Амплификация пролигированных образцов приводила к образованию продуктов различной длины, которые анализировались в геле с помощью Саузерн-блоттинга и служили мерой присутствия и количественного измерения определенных вариантов окончаний G-богатой цепи теломерной ДНК. Для этого разделенные в денатурирующем агарозном геле фрагменты ДНК переносили на нейлоновый фильтр и гибридизовали с 32P-меченной субтеломерной пробой, образованной в ходе ПЦР с использованием прямого и обратного субтеломерного праймеров.
Существенным ограничением рассматриваемого метода, как и метода предложенного для исследования Tetrahymena, является зависимость его эффективности от условий отжига последовательности, комплементарной G-оверхенгу. Необходимость терминального отжига Guide platform последовательности на конце оверхенга предполагает обязательное формирование искусственных 5'-оверхенгов, образование которых зависит от условий гибридизации, длины платформы и размеров матрицы оверхенга.
Размеры G-оверхенгов теломерной ДНК человека варьируют от 12 до 624 н.о. При прочих равных условиях гибридизации 3'-оверхенгов и платформы успешность адекватного анализа разнородной по длине популяции оверхенгов, представляется затруднительным.
Эффективность анализа определяется условием, при котором хотя бы одна единица платформы должна образовывать искусственный 5'-оверхенг оптимальной длины не менее 6-7 н.о., что определяется размерами теломерной последовательности в каждой кассете. При сокращении размеров образующегося 5'-оверхенга до 1-3 н.о. эффективность последующего лигирования при заявленной температуре в 35°C снижается вплоть до отсутствия. Если предположить, что при условии преимущественного отжига полноразмерных последовательностей Guide platform при заявленной температуре в 70°C сайт праймирования крайнего теломерного повтора ААТССС-5' платформы приходится на самый конец 3'-оверхенга, то возникают следующие возможные варианты критически коротких 5'-оверхенгов:
В связи с этим преимущество при лигировании получат те кассеты, которые отжигаются с образованием 5'-оверхенгов длиной 5 н.о. и более. При таких условиях гибридизации платформы в наиболее неблагоприятных условиях оказываются оверхенги, оканчивающиеся на триплеты GGG, AGG и TAG, сигнал лигирования которых может быть занижен.
При этом нужно учитывать, что при первоначальной гибридизации платформы и оверхенга при заявленной температуре в 70°C образующийся 5'-оверхенг должен быть длиной не более 30 н.о., так как больший размер создаст условия для нестабильного отжига платформы. В этом случае большее преимущество в лигировании получат более короткие оверхенги длиной 40-50 нуклеотидов, чем более длинные. Только такая длина 3'-оверхенга обеспечивает однозначное формирование 5'-оверхенга, достаточного для дальнейшего успешного анализа.
Затруднительным моментом в рассматриваемой методике представляется анализ самых коротких оверхенгов меньше 30 нуклеотидов из-за образования нестабильного дуплекса при гибридизации с высокой температурой в 70°C. Это ограничение частично снимается при снижении температуры первоначальной гибридизации до комнатной температуры, что было осуществлено авторами гораздо позднее, в работе 2009 года. Однако в этом исследовании метод G strand STELA использовался только для определения длины теломер без идентификации терминальных нуклеотидов (Yong Zhao, Agnel J.Sfeir et al. Telomere extension occurs at most chromosome ends and is uncoupled from fill-in in human cancer cells. Cell. Vol.138, No.3, 2009, pp. 463-475).
Рассмотрим возможные случаи, при которых отжиг платформы может приводить к образованию остаточного 3'-оверхенга. Вероятность его формирования повышается за счет преимущественного отжига полноразмерных последовательностей платформы при заявленной высокой температуре гибридизации в 70°C. При этом всегда будут присутствовать такие варианты гибридизации, при которых сайт праймирования крайнего теломерного повтора ААТССС-5' платформы приходится на комплементарный ему последний повтор TTAGGG оверхенга. Возникают следующие возможные комбинации гибридизации в зависимости от окончания оверхенга:
G-оверхенги, особенно оканчивающиеся на триплеты GGG, в результате образования тупого 3'-конца, вообще рискуют остаться не пролигированными, а следовательно, могут полностью выпасть из анализа. Возможно, с этим связана заниженная частота встречаемости этого варианта окончания оверхенга, которую получили авторы в результате образования артефактов лигирования. В таких же жестких условиях находятся оверхенги, оканчивающиеся на GGT, GTT и ТТА триплеты, так как могут образовывать слишком маленький остаточный 3'-участок, недостаточный для отжига еще одной единицы платформы и последующего лигирования.
Значительная часть популяции теломерных G-оверхенгов представлена одноцепочечными фрагментами длиной больше ста и даже несколько сотен нуклеотидов. При длине свыше 60 н.о. вероятен отжиг нескольких единиц платформы по всей протяженности оверхенга. При этом всегда не исключены возможности гибридизации:
а) с образованием тупого конца, особенно если исследуемый оверхенг оканчивается на триплет GGG-3', что определяется особенностями окончания платформы на триплет 5'-ССС;
б) с формированием остаточного 3'-оверхенга с минимальной длиной и с температурой плавления гораздо меньше, чем выбранная авторами для гибридизации и лигирования;
в) с образованием критически малого 5'-оверхенга, длиной в 1-3 н.о., недостаточного для последующего отжига G telorettes и лигирования.
Перечисленные факторы могут привести к образованию очень короткого, недостаточного для последующего анализа, 5'-оверхенга, либо к его полному отсутствию. При этом дальнейшее исследование для части оверхенгов становится невозможным и сигнал лигирования оказывается существенно заниженным.
Таким образом, главный недостаток анализируемого способа заключается в том, что различные по длине матрицы оверхенгов лигируются с разной эффективностью. При прочих равных условиях гибридизации одни матрицы получают преимущество, а лигирование других может быть затруднено.
Следующей особенностью G strand STELA метода, которая может привести к существенному искажению результата анализа, является зависимость эффективности финальной ПЦР от сайта праймирования прямого субтеломерного праймера XpYpE2. Последовательность 5'-TTGTCTCAGGGTCCTAGTG-3' этого праймера по данным программы анализа нуклеотидных последовательностей BLAST Nucleotide NCBI присутствует, прежде всего, в субтеломерной области хромосом X и Y. Другие хромосомы человека могут иметь сайты праймирования в субтеломерной области, которые отличаются по степени гомологии с последовательностью праймера, а следовательно, и своими конкурентными способностями в ходе ПЦР. В связи с этим преимущество при амплификации получит теломерная ДНК тех хромосом, которые обладают самым большим количеством наиболее гомологичных сайтов. В самых выгодных условиях амплификации оказывается ДНК хромосом X и Y (100% гомологии в субтеломерной области). Теломерная ДНК остальных хромосом, имеющая более низкую гомологию с последовательностью XpYpE2 праймера, рискует хуже амплифицироваться на фоне довольно высокой температуры отжига в 58°C. Кроме того, обнаружены многочисленные сайты праймирования в генных последовательностях всех хромосом человека, за исключением 12-й хромосомы. Эти сайты праймирования также отличаются степенью гомологии с последовательностью праймера и своей конкурентоспособностью в ходе ПЦР. Они могут приводить к появлению дополнительных продуктов амплификации при условии сохранения целостности соответствующих им участков геномной ДНК после обработки рестриктазой EcoRI на первоначальном этапе анализа. При этом хромосома 12 может полностью выпасть из анализа, так как у нее не найдена соответствующая последовательность. Из этого следует, что G-оверхенги, принадлежащие разным хромосомам, амплифицируются с разной эффективностью, что не может не отразиться на конечном результате анализа.
Кроме того, успешность применения анализируемого метода зависит от степени деградации теломерной ДНК. Конечная амплификация может быть осуществима только при условии, что район между прямым субтеломерным праймером и обратным G teltail праймером не содержит двухцепочечных разрывов. Эти области, достигающие длины нескольких тысяч нуклеотидов, могут подвергаться разрушению в процессе экстракции ДНК. Не исключена также возможность разрушения целостности субтеломерной области хромосом в ходе обработки геномной ДНК рестриктазой EcoRI на первоначальном этапе анализа.
Свидетельством того, что лигирование и амплификация в ходе G strand STELA метода каждый раз происходят по-разному, являются полученные авторами фореграммы, в которых наблюдались различные наборы полос при повторах анализа от 2 до 8 раз на каждый вариант окончания G-оверхенга.
Вышеперечисленные недостатки описанного метода, связанные с возникновением артефактов в ходе лигирования и амплификации, могут существенным образом отразиться на конечном результате анализа и привести к неверному определению частот встречаемости вариантов нуклеотидных окончаний G-оверхенга. В итоге это может привести к построению искаженного профиля терминальных нуклеотидов G-цепи теломерной ДНК, играющего ключевую роль в оценке пролиферативного статуса клетки.
Раскрытие изобретения
Основной технической задачей изобретения является разработка нового эффективного метода исследования терминальных нуклеотидов G-богатой цепи теломерной ДНК для устранения недостатков существующих методик.
Техническим результатом изобретения является повышение специфичности и чувствительности метода, выражающихся в возможности успешного исследования всей фракции G-оверхенгов разной длины и получения достоверного профиля терминальных нуклеотидов теломерной ДНК.
Поставленная задача решается применением нового подхода к изучению терминальных нуклеотидов ДНК, заключающегося в сочетании амплификации теломерной ДНК с последующим дуплекс-специфическим анализом.
Сущность изобретения заключается в создании синтетических флуоресцентно-меченных зондов, соответствующих шести возможным вариантам теломерных окончаний. Присутствие или отсутствие тех или иных вариантов терминальных нуклеотидов выявляют в ходе гибридизации зондов с изучаемой теломерной ДНК и расщеплении их с помощью фермента дуплекс-специфической нуклеазы. Расщепление зонда происходит только при условии его полного комплементарного соответствия с исследуемой последовательностью. Это обеспечивает высокую специфичность анализа и приводит к разгоранию специфического флуоресцентного сигнала в пробирках, который регистрируется флуориметрически. Степень разгорания зондов, соответствующих возможным вариантам окончаний теломерной ДНК, служит не только критерием их присутствия, но и мерой их количественного соотношения.
Заявленный способ включает возможные способы регистрации и интерпретации результатов анализа. Результат оценки уровня сигналов предлагается представлять в виде столбчатой диаграммы, отражающей процентное содержание определенных вариантов нуклеотидных окончаний G-оверхенгов и именуемой «профилем терминальных нуклеотидов G-цепи теломерной ДНК».
Основная характеристика заявленного способа определяется спецификой осуществления дуплекс-специфического анализа применительно к объекту исследования теломерной ДНК. Дуплекс-специфический анализ или DSNP-анализ (от англ. Duplex-specific nuclease preference) основан на использовании фермента дуплекс-специфической нуклеазы (ДСН, DSN) из гепатопанкреаса камчатского краба, обладающего специфической особенностью расщеплять двухцепочечную ДНК и оставаться неактивным в отношении одноцепочечной ДНК. DSNP-метод был разработан российскими учеными для анализа однонуклеотидных полиморфизмов генов и успешно используется для изучения точечных мутаций генов (Shagin D.A. et al. A Novel Method for SNP Detection Using a New Duplex-Specific Nuclease From Crab Hepatopancreas. Genome Research, 2002, Vol.12, pp. 1935-1942).
Было показано, что эффективность ДСН-гидролиза совершенных ДНК-дуплексов значительно выше, чем несовершенных, содержащих хотя бы одно неспаренное основание. Эта особенность дуплекс-специфической нуклеазы легла в основу разработанного метода исследования полиморфизма терминальных нуклеотидов G-цепи теломерной ДНК человека с помощью DSNP-анализа, который лег в основу заявленного способа.
Общий принцип DSNP-анализа сводится к использованию коротких, длиной 10-12 нуклеотидов, синтетических зондов (сигнальных проб), несущих донор флуоресценции на 5'-конце и гаситель флуоресценции на противоположном 3'-конце. При полном комплементарном взаимодействии зонда и исследуемой ДНК-мишени нуклеазная активность ДСН приводит к разгоранию детектируемого флуоресцентного сигнала. При этом показана высокая чувствительность метода в связи со способностью дуплекс-специфической нуклеазы различать вариации в один нуклеотид (Shagin D.A. et al., 2002). При исследовании однонуклеотидных полиморфизмов при точечных мутациях генов исследуемый участок ДНК амплифицируют с помощью полимеразной цепной реакции, а затем аликвоту ПЦР продукта смешивают с сигнальными пробами и инкубируют в присутствии фермента ДСН.
Существенным отличием заявляемого способа от классического дуплекс-специфического анализа нуклеотидных полиморфизмов является то, что он применяется для исследования не гетеронуклеотидной последовательности ДНК генов, а концевых участков тандемно-повторяющейся ДНК теломер. Последнее свойство объекта исследования определило принципиальную модификацию этапов амплификации исследуемой теломерной ДНК методом полимеразной цепной реакции с использованием специального набора праймеров, представленного в таблице 1 (SEQ ID NO: 1-5).
Крайнее 3'-концевое расположение G-оверхенга не позволяет использовать естественный обратный праймер для его амплификации. Последний факт вызвал необходимость использования искусственного наращивания 3'-конца с помощью терминальной дезоксинуклеотидилтрансферазы и мононуклеотидов, что отразилось на структуре сигнальных проб. Разработан набор из шести сигнальных проб, представленных в таблице 2 (SEQ ID NO: 6-11), в которых позиция мисматчевой дискриминации располагается на расстоянии 4-5 нуклеотидов от 3'-конца в области естественного окончания G-оверхенга и начала мононуклеотидного поли(dC) хвоста, достроенного трансферазой.
Выбор dCTP в качестве строительного материала был обусловлен тем, что это единственный нуклеотид, непредставленный в естественном теломерном повторе TTAGGG человека, и обеспечивающий однозначную дискриминацию всех шести возможных вариантов нуклеотидных окончаний G-цепи теломерной ДНК. В примерах осуществления изобретения предложено использование комбинации карбоксифлуоресцеинового красителя (FAM) на 5'-конце и его эффективного гасителя RTQ1 на 3'-конце, что позволяло осуществлять количественный анализ всех возможных вариантов окончания G-оверхенгов по интенсивности роста зеленой флуоресценции во времени. При конструировании зондов возможно использование любых красителей и гасителей в соответствующих им областях флуоресценции, в том числе и мультиканально, что отображено в формуле изобретения в п.6 и 15.
При этом эффективность использования того или иного варианта мечения будет определяться спектральными характеристиками флуорофор-связанной флуоресценции ДНК на текущий момент исследования. При разработке заявляемого способа было установлено, что нуклеотидные последовательности ДНК обладают специфической флуоресценцией в видимом диапазоне волн света, связанной с установлением общего квантово-связанного спектра с молекулой флуорофора (красителя). Согласно полученным данным ДНК обладает не только известной ранее флуоресценцией в УФ-диапазоне, но и имеет мультицветовой интегрированный спектр флуорофор-связанной флуоресценции в видимом диапазоне волн света. Специфика этой флуоресценции в каждом из цветов видимого спектра зависит от нуклеотидной последовательности ДНК. В связи с этим нуклеотидная последовательность зонда, меченного флуоресцентным красителем, определяет специфику его флуоресценции в соответствующем спектре видимого диапазона, а также скорость его разгорания при проведении DSNP-анализа.
Основной характеристикой флуорофор-связанных флуоресцентных свойств ДНК является спектральный код, который представляет собой соотношение уровней флуоресценции основных цветовых каналов в видимом диапазоне волн света. На момент проведенных в 2010-2011 гг. исследований специфика фоновой флуоресценции и расщепления используемых FAM-RTQ1 зондов соответствовала текущему на тот момент спектральному коду флуоресценции ДНК. Для него было характерно ярко выраженное испускание флуоресценции теломерной ДНК в зеленом и красном спектрах видимого света. При этом четко проявлялась градация увеличения энергетического потенциала в зеленом спектре для теломерных повторов, входящих в состав зондов, оканчивающихся на различное число остатков гуанина. Это привело к необходимости использования в примерах осуществления изобретения различной концентрации зондов, меченных карбоксифлуоресцеином для выравнивания их фоновой флуоресценции. Кроме этого, при обработке результатов DSNP-анализа использовались зонд-специфические коэффициенты. Они отражали специфику разгорания зондов при их расщеплении в ходе DSNP-анализа. Она определялась особенностями флуорофор-связанной флуоресценции ДНК-последовательности зонда в используемом спектре и напрямую зависела от его нуклеотидной последовательности (см. осуществление DSNP-анализа в примере 1).
Исследования показали, что спектральный код флуоресценции ДНК динамичен во времени и соотношение уровней флуоресценции основных цветовых каналов подвержено колебаниям (флуктуациям). Эти изменения должны своевременно контролироваться, так как напрямую затрагивают уровни фоновой флуоресценции зондов и специфику их разгорания при проведении DSNP-анализа. Адекватность использования концентраций зондов и зонд-специфических коэффициентов при проведении анализа должна проверяться контрольным исследованием эквимолярной смеси синтетических аналогов окончаний оверхенгов, аналогично примеру 5.
Принципиальная схема идентификации терминальных нуклеотидов G-богатой цепи теломерной ДНК с помощью ПЦР и DSNP-анализа.
На первом этапе исследования из образца геномной ДНК, полученного любым способом, сохраняющим ее нативность, отделяют фракцию одноцепочечных оверхенгов теломерной ДНК. Для этих целей используется фермент дуплекс-специфическая нуклеаза, эффективность которой была показана в работе по исследованию длины теломерных оверхенгов (Yong Zhao et al. Quantitative telomeric overhang determination using a double-strand specific nuclease. Nucleic Acids Research, 2008, Vol.36, No. 3). Исследования авторов статьи показали, что использование ДСН в концентрации 0,2 ед. на каждые 5 мкг геномной ДНК позволяет эффективно расщепить двухцепочечную ДНК до фрагментов длиной не более 10 пар оснований и сохранить нативную фракцию одноцепочечных оверхенгов.
Для концентрирования и очищения оверхенгов от остатков геномной ДНК в заявляемом способе предложена модификация методики с использованием покрытых стрептавидином парамагнитных частиц, описанной в статье Woodring Е. Wright et al. (Normal human chromosomes have long G-rich telomeric overhangs at one end. Genes&Development. 1997, Vol.11, No. 21, pp. 2801-2809). По данным авторов 40 мкг геномной ДНК содержит порядка 1 фемтомоля (fmole) теломерных концов. В соответствии с этим для преципитации даже самых длинных оверхенгов достаточно не более 1 пикомоля (pmole) биотинилированного олигонуклеотидного зонда (СССТАА)4 и 3 мкл покрытых стрептавидином магнитных частиц в концентрации 10 мг/мл.
В примерах осуществления изобретения предложено использование магнитных частиц со стрептавидином в концентрации 1 мг/мл производства фирмы «Силекс» (Россия) с эффективностью связывания в среднем 8 пмоль трехкратно биотинилированного продукта на 10 мкл частиц. В связи с тем, что преципитация фракции оверхенгов разной длины идет более медленно, чем связывание коротких олигонуклеотидов с комплексами частица-зонд, предлагается использовать 10 мкл частиц на каждые 20 мкг исходной геномной ДНК. Такого количества достаточно, даже учитывая некоторую потерю магнитных частиц при отмывках.
Для предварительного расщепления геномной ДНК, вместо рестриктазы Hinfl используют дуплекс-специфическую нуклеазу, а связыванию нативных оверхенгов с комплементарными зондами предшествует предварительная инкубация последних с магнитными частицами, покрытыми стрептавидином. При этом использование отмывок в низкосолевом буфере обеспечивает эффективное удаление не связавшихся с магнитными частицами зондов на первом этапе и остатков двухцепочечной геномной ДНК на следующем этапе. Такая последовательность проведения этапов обеспечивает сокращение времени преципитации оверхенгов до 1,5 часов по сравнению с исходной методикой Wright et al., где финальный этап инкубации суспензии ДНК и магнитных частиц осуществлялся в течение ночи. Повышение температуры финальной элюции до 70-75°С позволяет более эффективно сконцентрировать фракцию исследуемых оверхенгов в объеме, подходящем для дальнейшей стадии достраивания поли(dC) хвоста. При этом использование тройного биотина для модификации зонда СТААСССТААСССТААСССТА обеспечивает более прочную связь со стрептавидином на магнитных частицах при проведении высокотемпературной элюции. Для получения очищенной фракции одноцепочечных оверхенгов подходит любой другой способ из группы гибридизационных или сорбционных, в том числе с использованием различных мембран, колонок, частиц или других поверхностей с целью иммобилизации и последующего снятия с их поверхности исследуемых молекул, что отображено в п.2 формулы изобретения.
На втором этапе исследования водный элюент, содержащий одноцепочечные G-оверхенги, используют в качестве субстрата для фермента терминальной дезоксинуклеотидилтрансферазы, которая достраивает мононуклеотиды к 3'-концу цепи. На этапе достройки полидезоксицитидиловой последовательности (полидезоксицитидилирования) в качестве строительного материала используют dCTP мононуклеотиды. При этом областью проведения DSNP-анализа является район контакта естественного окончания 3'-оверхенга и начала поли(dC) последовательности. Продолжительность синтеза подбирается эмпирически и осуществляется в течение времени, достаточного для достраивания последовательности длиной 100-200 нуклеотидов, что необходимо для обеспечения эффективности последующего осаждения коротких оверхенгов при их очистке.
На третьем этапе после стандартной процедуры переосаждения и очистки от не включившихся в цепь dCTP мононуклеотидов образец используют для аналогичного достраивания поли(dA)/поли(dT) хвоста с использованием dATP/dTTP мононуклеотидов и фермента терминальной дезоксинуклеотидилтрансферазы. Сочетание последовательных этапов достраивания полидезоксицитидиловой и полидезоксиадениловой (политимидиловой) последовательностей позволяет отделить исследуемую границу контакта окончания оверхенга и поли(dC) хвоста от сайта праймирования при последующем синтезе копий минусовой цепи. Сохранение нативности этой границы является главным условием получения адекватных результатов анализа.
На четвертом этапе осуществляют синтез минусовой цепи с использованием термостабильной ДНК полимеразы и oligo dT25 Adapter праймера (SEQ ID NO: 1) в случае проведения концевого полидезоксиаденилирования на предыдущем этапе. В другом варианте осуществления способа синтез минусовой цепи проводят с использованием oligo dA25 Adapter праймера (SEQ ID NO: 2), если на третьем этапе осуществлялось присоединение политимидиловой последовательности (концевое политимидилирование). Оба варианта праймеров содержат на своем 5'-конце адаптерную последовательность (SEQ ID NO: 3). В качестве адаптера может использоваться любая последовательность, не комплементарная теломерным повторам и соответствующая оптимальной температуре отжига при проведении синтеза минусовой цепи, что отражено в формуле изобретения в п.4. Количество циклов синтеза минусовой цепи доводят до 100-200 циклов, что важно для получения достаточного уровня флуоресцентного сигнала при финальном DSNP-анализе. Последнее условие особенно важно при небольшом исходном количестве анализируемой ДНК.
На пятом этапе после очистки и осаждения фракции минусовой цепи осуществляют достройку ее 3'-конца с помощью терминальной трансферазы и dATP мононуклеотидов, если на третьем этапе использовалось концевое полидезоксиаденилирование цепи оверхенга, или dTTP мононуклеотидов, если применялось ее политимидилирование.
На шестом этапе очищенную и переосажденную фракцию минусовых цепей с достроенным поли(dA) хвостом подвергают финальной асимметричной амплификации с использованием адаптерной последовательности (SEQ ID NO: 3), либо аналогичной ей, в качестве прямого праймера и oligo dT30 праймера (SEQ ID NO: 4) в качестве обратного. Во втором варианте осуществления способа минусовые цепи с поли(dT) хвостом амплифицируют с использованием той же адаптерной последовательности в качестве прямого праймера, но в сочетании с oligo dA30 праймером (SEQ ID NO: 5) в качестве обратного праймера. Количество циклов подбирается экспериментально, не более 18-20, так чтобы обеспечить сохранение исходного соотношения вариантов окончания G-оверхенгов.
Использование асимметричной ПЦР с соотношением прямого и обратного праймеров 10:1 обеспечивает преимущественный синтез минусовой цепи и условия ее последующей гибридизации с сигнальными пробами. При этом доминирование адаптерного праймера приводит к вытеснению oligo dT30(oligo dA30) праймера, который может частично отжигаться как прямой на поли(dA/dT) последовательности, расположенной непосредственно за адаптерной.
На седьмом этапе проводят DSNP-анализ. Очищенный ПЦР продукт разделяют на шесть равных частей и смешивают с сигнальными пробами в присутствии фермента дуплекс-специфической нуклеазы.
Конструкция сигнальных проб представлена в таблице 2 (SEQ ID NO: 6-11) и подобрана с учетом специфики исследуемого района, а именно границы между окончанием 3'-оверхенга и началом поли(dC) последовательности. Сигнальная проба представляет собой синтетический олигонуклеотидный зонд длиной 10 пар нуклеотидов, представленный плюсовой цепью G-оверхенга и соответствующий одному из шести вариантов его окончания.
Структура зонда, предназначенного для дуплекс-специфического анализа, предполагает наличие молекулы донора флуоресценции на 5'/3'-конце и соответствующего ему гасителя флуоресценции на противоположном 3'/5'-конце. В примерах осуществления изобретения использовалась комбинация красителя карбоксифлуоресцеина (FAM) и гасителя RTQ1. Возможно использование других красителей в комбинации с гасителем и соответствующих им каналов флуоресценции, в том числе и мультиканально, что отображено в формуле изобретения в п.6 и 15.
После внесения сигнальных проб в соответствующую пробирку, содержащую исследуемый образец, до добавления фермента ДСН, проводят измерение фонового сигнала, который затем вычитается при нормировании значений прироста флуоресценции при расщеплении сигнальных проб.
Расщепление сигнальной пробы осуществляется в присутствии дуплекс-специфической нуклеазы только при ее полном комплементарном соответствии участку амплифицированной минусовой цепи G-оверхенга. При этом происходит расщепление пробы преимущественно между четвертым и пятым или пятым и шестым нуклеотидами (Anisimova V.E. et al. Isolation, characterization and molecular cloning of Duplex-Specific Nuclease from the hepatopancreas of the Kamchatka crab. BMC Biochemistry, 2008, Vol.9, №14). Пространственное разобщение красителя и гасителя приводит к разгоранию флуоресценции, соответствующей определенному каналу детекции. Для красителя FAM это длина волны 520 нм в зеленом диапазоне видимого света.
Разгорание сигнала в пробирке с известной сигнальной пробой свидетельствует о присутствии соответствующего ей окончания G-оверхенга. При этом возможно не только качественное определение присутствия того или иного окончания, но и полуколичественное измерение их содержания. Это очень важно для отслеживания изменений соотношения различных вариантов окончания G-оверхенгов в ходе клеточного цикла.
Измерение флуоресцентного сигнала осуществляют в пробирках любым известным способом с помощью специально предназначенных для этой цели приборов (спектрофлуориметров, фотометров-флуориметров, флуоресцентных сканеров и детекторов, флуоресцентных стереомикроскопов и т.п.), что отображено в п.7 формулы изобретения.
Интерпретация результатов расщепления зондов включает этап нормирования сигналов, заключающийся в вычитании исходного фона (background), установленного до проведения анализа, и умножении полученных числовых значений на зонд-специфический коэффициент, эмпирически рассчитанный для каждого флуоресцентного зонда. Коэффициенты вычисляют при построении кривых роста флуоресценции во времени для каждой сигнальной пробы в ходе контрольного исследования эквимолярной смеси оверхенгов. Для этой цели используют синтетические аналоги G-оверхенгов, прошедшие все этапы анализа, начиная с синтеза поли(dC) хвоста и заканчивая финальной амплификацией минусовой цепи, аналогично примеру 5.
Коэффициенты специфичны для каждой сигнальной пробы в отдельности и для каждой комбинации «краситель-гаситель». Они отображают скорость расщепления зонда, которая является константной величиной для заданного промежутка времени проведения анализа и соответствуют текущему спектральному коду флуорофор-связанной флуоресценции ДНК в видимом диапазоне. Как уже отмечалось ранее, коэффициенты отражают специфику энергетического потенциала теломерных повторов, входящих в состав зондов. В основном он определяется спектральным кодом флуоресценции гуанина на момент исследования. В приведенных ниже примерах осуществления изобретения показаны коэффициенты нормирования для шести вариантов сигнальных проб, меченных комбинацией красителя FAM и гасителя RTQ1, после одного и двух часов проведения анализа (табл.3).
Изменения спектрального кода флуоресценции ДНК, другие концентрационные соотношения зондов, как и другие варианты мечения, требуют нового вычисления коэффициентов нормирования значений сигналов для каждого зонда. Поэтому периодическое исследование контрольного образца эквимолярной смеси синтетических аналогов окончаний оверхенгов с целью верификации коэффициентов нормирования сигналов является обязательным. Особую значимость это имеет в условиях нестабильных спектральных свойств ДНК.
Итоговое значение сигнала для каждого зонда вычисляется с использованием коэффициентов нормирования для одного и двух часов параллельно, а затем усредняется. Результат приводится в виде столбчатой диаграммы, отображающей усредненное процентное содержание полученных сигналов каждого зонда от суммарного уровня сигнала, полученного от всех зондов. Значения используют для построения диаграммы процентного соотношения вариантов нуклеотидных окончаний G-оверхенгов теломерной ДНК, именуемого профилем терминальных нуклеотидов G-цепи теломерной ДНК.
Таким образом, первым аспектом изобретения является набор специфических праймеров, представленных в таблице 1 и используемых для стадии амплификации минусовой цепи теломерной ДНК, предшествующей DSNP-анализу.
Вторым аспектом изобретения является набор специфических олигонуклеотидных зондов, приведенных в таблице 2, используемых в качестве сигнальных проб при проведении дуплекс-специфического анализа терминальных нуклеотидов теломерной ДНК.
Третьим аспектом изобретения является непосредственно способ идентификации терминальных нуклеотидов G-богатой цепи теломерной ДНК, включающий амплификацию минусовой цепи G-оверхенгов теломерной ДНК и ее последующий дуплекс-специфический анализ. Заявленный способ отличается от известных ранее тем, что: а) финальной амплификации подвергают полидезоксиаденилированные (политимидилированные) по 3'-концу копии минусовой цепи, содержащие адаптерную последовательность, используемую в качестве прямого праймера, взятого в избытке по отношению к обратному поли(dT)/поли(dA) праймеру; б) результаты анализируют с помощью гибридизации с флуоресцентно-меченными зондами в присутствии фермента дуплекс-специфической нуклеазы; в) результаты регистрируют по степени расщепления зондов и уровню разгорания флуоресценции с помощью любого аппаратно-программного комплекса, позволяющего отображать флуоресценцию пробирок с зондами на экране компьютера и количественно определять уровень сигнала в каждой из них.
Сопоставительный анализ с прототипом позволяет сделать вывод о том, что заявленный способ исследования терминальных нуклеотидов G-цепи теломерной ДНК принципиально отличается от вышеописанного метода Single Telomere length analysis (G strand STELA), предложенного Agnel J. Sfeir et al., и основывается на совершенно ином подходе с применением фермента дуплекс-специфической нуклеазы без использования процедуры лигирования. Заявленный способ отличается возможностью эффективного анализа всей фракции G-оверхенгов любой длины и разных окончаний. Этот факт является главным техническим результатом заявленного изобретения, выражающимся в повышении специфичности и чувствительности анализа по сравнению с прототипом. В связи с этим заявленный способ позволяет определить истинное соотношение вариантов окончаний теломерных оверхенгов. Построение на основе этого адекватного профиля терминальных нуклеотидов G-цепи теломерной ДНК является своеобразным маркером процессов, связанных с делением клетки. Таким образом, заявленный способ может стать новым инструментом фундаментальных исследований в области биологии клетки и решения множества прикладных задач в сфере медицины и биотехнологии.
Краткое описание чертежей
Заявленное изобретение поясняется следующими фигурами.
Фигура 1. Результат DSNP-анализа эквимолярной смеси синтетических аналогов окончаний теломерных оверхенгов в виде графического файла (пример 5). Представляет собой ряды последовательных снимков пробирок в зеленом спектре, полученных с помощью прибора флуоресцентного детектора FDG-001. Изображения пробирок в виде круглых областей соответствуют зонам измерений оптической плотности в программе LabWorks (UVP, England) и расположены построчно для каждого варианта зонда. Первая строка соответствует TAG варианту окончания G-цепи теломерной ДНК, а последняя - ТТА варианту. В каждой строке первая зона соответствует стартовому значению фона, вторая и третья зоны - уровням сигнала после одного и двух часов проведения DSNP-анализа соответственно.
Фигура 2. Результат конечной обработки данных DSNP-анализа образца 1 в виде построения профиля терминальных нуклеотидов G-цепи теломерной ДНК. В дальнейшем при описании чертежей под профилем понимают процентное соотношение различных вариантов нуклеотидных окончаний (триплетов) исследуемой популяции G-оверхенгов теломерной ДНК. Образец 1 представлен геномной ДНК, выделенной из культуры лимфоцитов периферической крови человека, не стимулированной к делению с помощью фитогемагглютинина (пример 1).
Фигура 3. Результат конечной обработки данных DSNP-анализа образца 2 в виде построения профиля терминальных нуклеотидов G-цепи теломерной ДНК. Образец 2, представлен геномной ДНК, выделенной из культуры лимфоцитов периферической крови человека, стимулированной к делению обработкой ФГА в течение 2 часов (пример 2).
Фигура 4. Результат конечной обработки данных DSNP-анализа образца 3 в виде построения профиля терминальных нуклеотидов G-цепи теломерной ДНК. Образец 3, представлен геномной ДНК, выделенной из культуры лимфоцитов периферической крови человека, стимулированной к делению обработкой ФГА в течение 4 часов (пример 3).
Фигура 5. Результат конечной обработки данных DSNP-анализа образца 4 в виде построения профиля терминальных нуклеотидов G-цепи теломерной ДНК. Образец 4, представлен геномной ДНК, выделенной из культуры лимфоцитов периферической крови человека, стимулированной к делению обработкой ФГА в течение 11 часов (пример 4).
Фигура 6. Результат конечной обработки данных DSNP-анализа образца 5 в виде построения профиля терминальных нуклеотидов G-цепи теломерной ДНК. Образец 5, представлен смесью синтетических аналогов окончаний теломерных оверхенгов в эквимолярном соотношении (пример 5).
Фигура 7. Совмещенный график динамики изменений профиля терминальных нуклеотидов G-цепи теломерной ДНК лимфоцитов крови. График построен по шкале времени воздействия на культуру лимфоцитов стимулирующего деление фактора (ФГА) в течение первых одиннадцати часов.
Фигура 8. Профиль терминальных нуклеотидов G-цепи теломерной ДНК лейкемической культуры Jurkat, синхронизированной в митотической фазе.
Фигура 9. Профиль терминальных нуклеотидов G-цепи теломерной ДНК лейкемической культуры К-562, синхронизированной в митотической фазе.
Осуществление изобретения
Способ иллюстрируется следующими примерами.
Пример 1. Анализ терминальных нуклеотидов G-цепи теломерной ДНК человека, выделенной из культуры лимфоцитов периферической крови, не стимулированной к делению с помощью фитогемагглютинина (ФГА).
До проведения исследования был проведен анализ экспрессии генов циклинов методом стандартной ОТ-ПЦР для определения степени естественной активации деления лимфоцитов. Было установлено, что используемая культура лимфоцитов характеризовалась невысоким естественным уровнем экспрессии циклинов D2 и D3. Уровень мРНК этих генов по отношению к мРНК нормировочного гена GAPDH составил 0,54 и 0,73 соответственно. Это свидетельствовало о незначительной прогрессии G1 фазы клеточного цикла, соответствующей ранней подготовке к делению у незначительной части клеток. Экспрессия генов циклинов групп А, В и Е не была детектирована, что говорило об отсутствии митоза и предшествующей ему синтетической фазы. В связи с этим данная культура лимфоцитов вполне удовлетворяла условиям неделящегося состояния и слабой активации подготовки к делению у незначительной части клеток. Эти сведения важны для последующего установления взаимосвязи профиля терминальных нуклеотидов G-цепи теломерной ДНК и профессии клеточного цикла.
Для выделения фракции однонитевых 3'-оверхенгов G-цепи теломерной ДНК использовали образец геномной ДНК, выделенный с помощью набора Diatom™ DNA Prep100 («Биоком», Россия). Экстракцию проводили в соответствии с рекомендациями производителя в некоторой модификации. Инкубацию образца в лизирующем буфере на первом этапе экстракции и финальный этап элюции ДНК проводили при температуре 55°С, вместо 65°С, во избежание денатурации ДНК. Далее 20 мкг геномной ДНК вносили в реакционную смесь, состоящую из следующих компонентов:
4 мкл буфера для дуплекс-специфической нуклеазы («Евроген», Россия);
0,8 ед дуплекс-специфической нуклеазы («Евроген», Россия).
Объем реакционной смеси доводили до 40 мкл и деликатно перемешивали на вортексе.
Реакцию расщепления двухцепочечной ДНК проводили при 37°С в течение 2,5 часов. Затем фермент ДСН удаляли из реакционной смеси 3-кратной обработкой раствором сорбента BlueSorb («Клоноген», Санкт-Петербург). Для этого к образцу каждый раз добавляли по 6 мкл сорбента, смесь тщательно встряхивали на вортексе и осаждали сорбент кратковременным центрифугированием с последующим отделением жидкой фазы, содержащей ДНК.
После удаления нуклеазы образец инкубировали с магнитными частицами, несущими на своей поверхности олигонуклеотидные зонды, комплементарные повторам G-цепи теломерной ДНК. Для этих целей использовали суперпарамагнитные частицы, покрытые стрептавидином в концентрации 1 мг/мл производства фирмы «Силекс» (Россия) и синтетические олигонуклеотидные зонды, состоящие из последовательности СТААСССТААСССТААСССТА, содержащие на своем 5'-конце три молекулы биотина («Синтол», Россия). Эксперименты показали, что в среднем 100 мкл магнитных частиц присоединяли 80 пикомоль 3xBiotin-зонда. Соответственно 10 мкл магнитных частиц связывали около 8 пикомоль зонда. Этого количества более чем достаточно для преципитации однонитевых оверхенгов. Избыточность используемого количества частиц и зонда по отношению к количеству теломерных оверхенгов необходима, во-первых, для обеспечения эффективности связывания магнитных частиц и протяженных теломерных последовательностей, а, во-вторых, для удобства работы с объемом магнитных частиц во избежание их потери при отмывках.
Все манипуляции с магнитными частицами производили согласно рекомендациям производителя, используя оригинальные отмывочные растворы и магнитный штатив. Ресуспендированные магнитные частицы в количестве 10 мкл трижды отмывали 0,5х отмывочным раствором по 200 мкл, суспендировали в 100 мкл того же раствора и инкубировали с 10 пикомолями 3xBiotin-зонда. Конъюгацию стрептавидина и биотина проводили при комнатной температуре в течение 30 минут. При этом образец аккуратно суспендировали с помощью пипетки-дозатора каждые 5-7 минут. Затем магнитные частицы снова отмывали 3 раза 0,5х отмывочным раствором от несвязавшегося зонда, суспендировали в 50 мкл 1х раствора и добавляли исследуемый образец ДНК после обработки нуклеазой и сорбентом.
Связывание зондов с нативной фракцией одноцепочечных G-оверхенгов осуществляли при комнатной температуре в течение 40 минут, аккуратно перемешивая содержимое пробирки с помощью пипетки-дозатора каждые 5-7 минут. После этого магнитные частицы с иммобилизованными G-оверхенгами отмывали от остатков двухцепочечной геномной ДНК с помощью 0,1х отмывочного раствора по 200 мкл четыре раза. После тщательного удаления остатков отмывочного буфера магнитные частицы суспендировали в дистиллированной очищенной воде в объеме 20 мкл и инкубировали в термостате при 70-75°С в течение 5 минут. Температурную элюцию проводили в два этапа, каждый раз собирая магнитные частицы с помощью магнитного штатива и отбирая раствор элюента пипеткой-дозатором в новую пробирку. Второй раз элюцию осуществляли в объеме 15 мкл. Объединенный элюент использовали для реакции синтеза поли(dC) хвоста (концевого полидезоксицитидилирования) в объеме 40 мкл реакционной смеси, содержащей:
водный элюент оверхенгов;
8 мкл 5х буфера для дезоксинуклеотидилтрансферазы («Силекс», Россия);
2 мкл 5 мМ раствора dCTP («Силекс», Россия);
2,5 ед терминальной дезоксинуклеотидилтрансферазы («Силекс», Россия).
Объем реакционной смеси при необходимости доводили до 40 мкл, используя очищенную дистиллированную воду.
Реакцию проводили при температуре 37°С в течение 40 минут, осуществляя деликатное перемешивание на вортексе каждые 10 минут.
Без специальной инактивации фермента осуществляли переосаждение образца ДНК, для чего в реакционную смесь последовательно добавляли:
1 мкл раствора соосадителя гликогена с концентрацией 20 мг/мл («Силекс», Россия);
25 мкл 7М раствора ацетата аммония;
165 мкл 96% охлажденного этилового спирта.
Смесь тщательно перемешивали на вортексе и высаживали в морозильной камере при температуре -20°С в течение не менее 12 часов.
Осаждение осуществляли центрифугированием при 15 тысячах об/мин в течение 60 минут. После осторожной промывки осадка охлажденным 70% спиртом в объеме 300 мкл осуществляли повторное центрифугирование при тех же оборотах в течение 30 минут. Подсушенный при открытой пробирке осадок растворяли в 25 мкл очищенной воды.
Далее в этой же пробирке, аналогично описанному выше способу, осуществляли реакцию полидезоксиаденилирования, с единственным отличием, что вместо раствора dCTP добавляли 2 мкл 5 мМ раствора dATP. Реакцию проводили при температуре 37°С в течение 30 минут и осуществляли переосаждение образца ДНК описанным выше методом. После финального центрифугирования осадок растворяли в 25 мкл очищенной дистиллированной воды.
Реакцию синтеза минусовых цепей проводили в объеме 40 мкл на программируемом термоциклере «Терцик» («ДНК-Технология», Россия). Образец после тщательного перемешивания на вортексе помещали в 0,5 мл ПЦР пробирку, содержащую следующие компоненты:
4 мкл 10х буфера для Taq ДНК-полимеразы, содержащего 25 mM MgCl2 («Силекс», Россия);
3 мкл 5 мМ раствора dNTP («Силекс», Россия);
4 мкл 20 мкМ раствора праймера SEQ ID NO: 1;
2,5 ед термостабильной Taq ДНК-полимеразы («Силекс», Россия).
Объем реакционной смеси доводили до 40 мкл и покрывали минеральным маслом во избежание испарения при проведении реакции. Применение амплификатора с греющейся крышкой позволяет избежать необходимости использования масла.
Синтез проводили в 4 раунда по 50 циклов каждый со следующим температурным режимом: 94°С - 40 с, 43°С - 30 с, 72°С - 40 с. В первом раунде первоначальная денатурация при 95°С составляла 30 секунд, а в последующих раундах увеличивалась до двух минут. Завершающая достройка цепи при 72°С увеличивалась до 3 минут в первых трех раундах и до 5 минут в последнем раунде. Перед проведением каждого последующего раунда добавлялась новая порция фермента термостабильной Taq ДНК-полимеразы в количестве 2,5 ед. Суммарное количество циклов синтеза минусовой цепи составило 200.
После проведения последнего раунда образец тщательно отбирали из-под масла и переливали в микропробирку объемом 1,5 мл, куда добавляли равный объем хлороформа. После интенсивного встряхивания на вортексе проводили центрифугирование при 10 тысячах об/мин в течение 3 минут. К отобранному супернатанту добавляли:
1 мкл раствора соосадителя тРНК с концентрацией 10 мг/мл («Силекс», Россия);
25 мкл 7М раствора ацетата аммония;
165 мкл 96% охлажденного этилового спирта.
Смесь тщательно перемешивали на вортексе и высаживали в морозильной камере при температуре -20°С в течение не менее 12 часов. Использование в качестве соосадителя раствора тРНК позволило избежать осаждения не включившихся в цепь «холостых» праймеров. Применение для этих целей гликогена или линейного полиакриламида не приемлемо, так как они осаждают короткие олигонуклеотиды.
Осаждение осуществляли центрифугированием при 15 тысячах об/мин в течение 60 минут. После деликатной промывки осадка охлажденным 70% спиртом в объеме 300 мкл осуществляли повторное центрифугирование при тех же оборотах в течение 30 минут. Подсушенный при открытой пробирке осадок растворяли в 25 мкл очищенной дистиллированной воды.
Реакцию полидезоксиаденилирования 3'-конца полученной фракции минусовой цепи проводили в этой же микропробирке, куда последовательно вносили:
8 мкл 5х буфера для дезоксинуклеотидилтрансферазы («Силекс», Россия);
4 мкл 5 мМ раствора dATP («Силекс», Россия);
15 ед терминальной дезоксинуклеотидилтрансферазы («Силекс», Россия).
Объем реакционной смеси доводили до 40 мкл, используя очищенную дистиллированную воду. Реакцию проводили при температуре 37°С, осуществляя деликатное перемешивание смеси на вортексе каждые 10 минут. Синтез осуществляли в течение 30 минут без температурной инактивации фермента.
Сразу после реакции для проведения переосаждения ДНК последовательно добавляли:
1 мкл раствора соосадителя гликогена с концентрацией 20 мг/мл («Силекс», Россия);
25 мкл 7М раствора ацетата аммония;
165 мкл 96% охлажденного этилового спирта.
Смесь тщательно перемешивали на вортексе и высаживали в морозильной камере при температуре -20°С в течение не менее 12 часов.
Осаждение осуществляли центрифугированием при 15 тысячах об/мин в течение 60 минут. После деликатной промывки осадка 70% спиртом в объеме 300 мкл осуществляли повторное центрифугирование при тех же оборотах в течение 30 минут. Подсушенный при открытой пробирке осадок растворяли в 50 мкл очищенной воды.
Реакцию финальной асимметричной амплификации минусовой цепи проводили в двух микропробирках объемом 0,5 мл в конечном объеме реакции 40 мкл на программируемом термоциклере «Терцик» («ДНК-Технология», Россия). Образец после тщательного перемешивания на вортексе разделяли на две части и переносили в две ПЦР пробирки, каждая из которых содержала смесь из следующих компонентов:
4 мкл 10х буфера для Taq ДНК-полимеразы, содержащего 25 mM MgCl2 («Силекс», Россия);
4 мкл 5 мМ раствора dNTP («Силекс», Россия);
4 мкл раствора, содержащего смесь адаптерного праймера (SEQ ID NO: 3) и Oligo dT30 праймера (SEQ ID NO: 4) в концентрации 20 мкМ и 2 мкМ соответственно;
5 ед термостабильной Taq ДНК-полимеразы («Силекс», Россия).
Объем реакционной смеси в каждой пробирке доводили до 40 мкл и после деликатного смешивания смесь покрывали минеральным маслом во избежание испарения в ходе ПЦР. Применение амплификатора с греющейся крышкой позволяет избежать необходимости использования масла.
Амплификацию осуществляли в течение 20 циклов со следующим температурным режимом: 94°С - 40 с, 48°С - 30 с, 72°С - 60 с. Первоначальная денатурация при 95°С составляла 3 минуты, финальная достройка цепи при 72°С увеличивалась до 5 минут. Увеличение количества циклов не рекомендуется, так как может привести к искажению исходного соотношения вариантов окончания G-оверхенгов.
После проведения полимеразной цепной реакции образцы тщательно отбирали из-под масла и переносили в общую микропробирку объемом 1,5 мл, куда добавляли равный объем хлороформа. После интенсивного встряхивания на вортексе проводили центрифугирование при 10 тысячах об/мин в течение 3 минут. К отобранному супернатанту добавляли:
1,5 мкл раствора соосадителя гликогена с концентрацией 20 мг/мл («Силекс», Россия);
50 мкл 7М раствора ацетата аммония;
329 мкл 96% охлажденного этилового спирта.
Смесь тщательно перемешивали на вортексе и высаживали в морозильной камере при температуре -20°С в течение 1,5 часов. Инкубирование в течение большего количества времени может привести к нежелательному чрезмерному осаждению «холостых» праймеров.
Осаждение осуществляли центрифугированием при 15 тысячах об/мин в течение 40 минут. После промывки осадка 70% спиртом в объеме 400 мкл осуществляли повторное центрифугирование при тех же оборотах в течение 20 минут. Подсушенный при открытой пробирке осадок растворяли в 90 мкл очищенной воды.
Для проведения дуплекс-специфического (DSNP) анализа готовили 6 тонкостенных ПЦР пробирок фирмы «Axygen» объемом 0,5 мл, в каждую из которых добавляли:
2 мкл 10х буфера для дуплекс-специфической нуклеазы («Евроген», Россия);
3 мкл раствора одного из 6 зондов, представленных в последовательностях SEQ ID NO: 6-11 (табл.2) в следующей модификации. Олигонуклеотиды были синтезированы компанией «Синтол» (Россия) и содержали молекулу карбоксифлуоресцеина (FAM) на 5'-конце и гасителя флуоресценции RTQ1 на противоположном 3'-конце.
Исходные стоковые концентрации растворов для каждого зонда подбирались эмпирически. Они соответствовали следующим концентрациям: 10 мкМ для SEQ ID NO: 10 и SEQ ID NO: 11; 7,5 мкМ для SEQ ID NO: 6; 5 мкМ для SEQ ID NO: 7 и SEQ ID NO: 9; 2,5 мкМ для SEQ ID NO: 8. Следует отметить, что приведенные концентрации зондов, а также условия проведения и обработки результатов DSNP-анализа удовлетворяли условиям существовавшего на момент исследования спектрального кода флуорофор-связанной флуоресценции ДНК (декабрь 2010 - октябрь 2011 гг.).
Существенные изменения спектрального кода требуют перерасчета значений концентраций стоковых растворов и коэффициентов нормирования сигналов для каждого зонда в таблице 3. Использование других красителей в отличных от зеленого спектрах флуоресценции также требует отдельного установления концентраций растворов зондов и коэффициентов нормирования сигналов в соответствии с текущим спектральным кодом флуоресценции ДНК. Их определение осуществляется в ходе исследования контрольного образца, представляющего собой эквимолярную смесь различных окончаний синтетических аналогов теломерных оверхенгов (пример 5). В связи с возможностями спектральной нестабильности флуоресцентных свойств ДНК периодическое исследование контрольного образца является крайне целесообразным и необходимым.
Для проведения DSNP-анализа в каждую пробирку, содержащую ДСН-буфер и зонд, вносили по 15 мкл тщательно перемешанной аликвоты образца. Сразу после смешивания образца и зондов проводилось измерение фонового сигнала (background) каждой пробирки с использованием аппаратно-программного комплекса, состоящего из прибора флуоресцентного детектора FDG-001 (совместная разработка автора патента и института теоретической и экспериментальной биофизики, Пущино, Россия) и программы обработки данных LabWorks 4.6 (UVP Biolmaging Systems, Англия).
Прибор FDG-001 предназначен для выявления сигналов флуоресценции в разных спектрах видимого света. Зеленый канал детекции флуоресценции прибора соответствовал спектральным характеристикам карбоксифлуоресцеина (FAM) и имел диапазон возбуждающего излучения, равный 450-490 нм, а регистрируемого излучения - 510-560 нм. Для возбуждения флуоресценции оптической метки пробирка с зондами равномерно освещалась светодиодами синего света соответствующей длины волны. Для получения изображения флуоресцирующих пробирок свет, проходя через зеленый интерференционный светофильтр, попадал в объектив монохромной камеры WAT-120N (Watec Со Ltd, Япония). Плата видеозахвата StudioTMiMovieBox (Pinnacle Systems GmbH, Германия) преобразовывала аналоговый телевизионный сигнал в цифровую форму и позволяла отображать изображения пробирок на экране компьютера и сохранять в виде графического файла.
После сохранения изображения фонового сигнала для каждого зонда в каждую пробирку добавляли по 0,125 ед фермента дуплекс-специфической нуклеазы (ДСН). Расщепление зондов осуществляли в течение двух часов при 37°С, деликатно перемешивая на вортексе каждые 30 минут. После одного и двух часов проведения анализа осуществляли видеозахват каждой пробирки в виде окружности фиксированной площади подобно изображению фиг.1, полученному для примера 5.
Ряды последовательных снимков пробирок в зеленом спектре анализировали в программе LabWorks, позволяющей производить измерения уровня оптической плотности заданной области изображения. Полученные с помощью программы величины значимой оптической плотности (Mean Density) в условных единицах служили мерой интенсивности флуоресцентного сигнала зонда в соответствующей пробирке. Для измерения уровня флуоресцентного сигнала возможно применение любых флуориметрических приборов и аппаратно-программных комплексов, что отображено в п.7 формулы изобретения.
Обработка полученных данных включала следующие этапы:
1-й этап: вычитание фонового сигнала и получение уровня сигнала для каждого зонда после 1-го и 2-х часов проведения анализа;
2-й этап: нормирование уровня сигнала умножением на соответствующий каждому зонду коэффициент из таблицы 3 (осуществляется параллельно для 1-го и 2-х часов проведения анализа);
3-й этап: определение уровней сигналов каждого зонда в процентах после 1-го и 2-х часов проведения анализа.
Этапы 1-3 обработки полученных данных представлены в таблице 4.
4-й этап: усреднение процентного содержания уровней сигналов, полученных после одного и двух часов проведения анализа. Результатом конечной обработки данных явилось построение столбчатой диаграммы, отображающей усредненное процентное соотношение вариантов нуклеотидных окончаний G-оверхенгов, и именуемой «профилем терминальных нуклеотидов G-цепи теломерной ДНК» (фиг.2).
Пример 2. Анализ терминальных нуклеотидов G-цепи теломерной ДНК человека, выделенной из культуры лимфоцитов периферической крови, стимулированной к делению в присутствии фитогемагглютинина (ФГА) в течение 2 часов. Стимуляцию деления лимфоцитов осуществляли добавлением фитогемагглютинина к культуральной среде RPMI-1640 в конечной концентрации 10 мкг/мл.
Изучение экспрессии генов циклинов в культуре лимфоцитов методом стандартной ОТ-ПЦР выявило резкое возрастание количества мРНК циклина D2 на фоне отсутствия циклинов А2, В1, D3 и Е1. Уровень мРНК циклина D2 по отношению к мРНК нормировочного гена GAPDH составил 1,32, что свидетельствовало о значительной прогрессии ранней G1 фазы клеточного цикла у значительной части клеток. Отсутствие детектируемой экспрессии генов циклинов групп А, В и Е свидетельствовало об отсутствии митоза и предшествующей ей синтетической фазы. В связи с этим данная культура лимфоцитов удовлетворяла условиям неделящегося состояния и значительной активации ранней подготовки к делению.
Все этапы анализа проводили аналогично методике, описанной в примере 1.
Последовательные этапы обработки данных, полученных в ходе дуплекс-специфического анализа, представлены в таблице 5.
Конечный результат обработки данных представлен столбчатой диаграммой усредненных процентных значений флуоресцентных сигналов зондов, соответствующих вариантам нуклеотидных окончаний G-оверхенгов в изученной популяции лимфоцитов. Данная диаграмма представлена фиг.3 и соответствует профилю терминальных нуклеотидов G-цепи теломерной ДНК, выделенной из культуры лимфоцитов человека, стимулированной к делению в присутствии ФГА в течение 2 часов.
Пример 3. Анализ терминальных нуклеотидов G-цепи теломерной ДНК человека, выделенной из культуры лимфоцитов периферической крови, стимулированной к делению в присутствии фитогемагглютинина (ФГА) в течение 4 часов.
Изучение экспрессии генов циклинов в этой культуре лимфоцитов так же, как в предыдущем примере, выявило высокий уровень мРНК циклина D2 (1,19) на фоне отсутствия циклинов А2, В1, D3 и Е1. В связи с этим данная культура лимфоцитов, как и культура в предыдущем примере, удовлетворяла условиям неделящегося состояния и значительной активации ранней подготовки к делению.
Все этапы анализа проводили аналогично методике, описанной в примере 1. Последовательные этапы обработки данных, полученных в ходе дуплекс-специфического анализа, представлены в таблице 6.
Конечный результат обработки данных представлен столбчатой диаграммой усредненных процентных значений флуоресцентных сигналов зондов, соответствующих вариантам нуклеотидных окончаний G-оверхенгов в изученной популяции лимфоцитов. Данная диаграмма представлена фиг.4 и соответствует профилю терминальных нуклеотидов G-цепи теломерной ДНК, выделенной из культуры лимфоцитов человека, стимулированной к делению в присутствии ФГА в течение 4 часов.
Пример 4. Анализ терминальных нуклеотидов G-цепи теломерной ДНК человека, выделенной из культуры лимфоцитов периферической крови, стимулированной к делению в присутствии фитогемагглютинина (ФГА) в течение 11 часов.
Изучение экспрессии генов циклинов в культуре лимфоцитов методом стандартной ОТ-ПЦР выявило присутствие всех детектируемых циклинов. Были получены следующие значения уровней мРНК циклинов по отношению к мРНК нормировочного гена GAPDH: А2 (0,25), В1 (0,95), D2 (1,47), D3 (0,91) и Е1 (0,17). Высокие уровни мРНК циклинов D2 и D3 свидетельствовали о существенной прогрессии G1 фазы клеточного цикла у значительной части клеток. В то же время другая часть клеток находилась на синтетическом этапе подготовки к делению (циклины Е1 и А2) и этапе митоза (циклин В1). Видимо, изначально данная культура лимфоцитов представляла собой смешанную популяцию клеток, находящихся на разных этапах клеточного цикла. Часть клеток еще до стимуляции РНА уже находилась в G1 и S фазах. Дальнейшая прогрессия клеточного цикла привела через 11 часов воздействия ФГА к появлению синтетической и митотической фаз соответственно.
Исследуемая культура представляет особый интерес, так как позволяет установить характер профиля терминальных нуклеотидов, существующий большую часть клеточного цикла. Она позволяет определить, какой профиль существует в течение поздней G1 фазы, синтетической и постсинтетической фаз, а также во время самого митоза, за исключением периода первых часов вхождения в раннюю G1 фазу, составляющего триггерный механизм начала активации деления клетки.
Все этапы анализа проводили аналогично методике, описанной в примере 1.
Последовательные этапы обработки данных, полученных в ходе дуплекс-специфического анализа, представлены в таблице 7.
Конечный результат обработки данных представлен профилем терминальных нуклеотидов G-цепи теломерной ДНК, выделенной из культуры лимфоцитов человека, стимулированной к делению в присутствии ФГА в течение 11 часов (фиг.5).
Пример 5. Анализ терминальных нуклеотидов теломерной ДНК образца, представленного смесью синтетических аналогов окончаний G-оверхенгов в эквимолярной концентрации. Данный образец использовался в качестве контрольного для установления концентрационных соотношений зондов и коэффициентов нормирования сигнала их расщепления при проведении DSNP-анализа.
В эппендорфовской микропробирке объемом 1,5 мл готовили смесь анализируемой ДНК, для чего добавляли по 0,5 пикомоль каждого из шести вариантов окончаний G-оверхенга. Синтетические аналоги оверхенгов представляли собой синтетические дезоксирибоолигонуклеотиды длиной 25 нуклеотидов, синтезированные стандартным фосфорамидитным методом в компании «Евроген» (Россия) и заканчивающиеся на своем 3'-конце триплетами: TAG, AGG, GGG, GGT, GTT и ТТА.
Смесь использовали для реакции полидезоксицитидилирования в конечном объеме 40 мкл. Далее все последующие этапы полидезоксиаденилирования, синтеза минусовой цепи и т.д. осуществляли согласно методике, описанной в примере 1.
Фиг.1 представляет собой графическое изображение флуоресценции каждого зонда в отдельности после одного и двух часов проведения DSNP-анализа. Обработку полученных изображений проводили согласно описанной в примере 1 процедуре анализа с помощью программы LabWorks. Последовательные этапы обработки полученных значений флуоресценции для каждого зонда и соответствующего ему терминального окончания представлены в таблице 8.
Конечный результат обработки данных представлен столбчатой диаграммой усредненных процентных значений прироста сигнала при расщеплении зондов, изображенной на фиг.6. Полученная диаграмма соответствует профилю терминальных нуклеотидов исследуемой эквимолярной смеси синтетических аналогов окончаний теломерных оверхенгов.
Как видно из диаграммы, полученный профиль указывает на присутствие всех вариантов триплетных нуклеотидных окончаний в равном соотношении приблизительно по 16,7%, что полностью соответствует исходному эквимолярному соотношению в смеси синтетических аналогов окончаний G-оверхенгов. Максимальное отклонение полученных в ходе этого анализа значений от теоретически ожидаемых составляет не более 0,5%. Проведение подобных исследований контрольных образцов с известным заранее количественным соотношением окончаний теломерных оверхенгов выявил предел погрешности заявляемого способа не более 1%. Этот факт позволил применить подобранные концентрации растворов зондов и коэффициенты нормирования сигналов с высокой степенью достоверности результатов при исследовании нативных фракций теломерных оверхенгов в примерах 1-4.
В заключении отметим, что рассмотренные выше примеры иллюстрируют возможность применения заявленного способа не только для качественного определения присутствия вариантов нуклеотидных окончаний G-цепи теломерной ДНК, но и для точной количественной оценки их соотношения в ходе клеточного цикла. Заявленный способ подходит как для исследования очень коротких оверхенгов, так и для более протяженных. Показана эффективность его применения как для анализа синтетических аналогов теломерных оверхенгов, так и для естественных фракций G-оверхенгов, представленных разнообразными по длине полинуклеотидами.
В заявленном способе за счет полного ухода от процедуры лигирования, снимается зависимость результативности анализа от эффективности ее прохождения, а именно от длины матрицы G-оверхенга, которая наблюдается в способе, предложенном Agnel J. Sfeir et al. Этот факт демонстрирует бесспорное техническое преимущество заявленного способа перед существующими аналогами. Оно позволило автору патента выявить неизвестные ранее изменения терминальных нуклеотидов теломерных оверхенгов, происходящие в первые часы после активации клеточного деления. Было установлено существование и взаимный переход равновесного и неравновесного состояния сбалансированности профиля терминальных нуклеотидов G-цепи теломерной ДНК человека, который определяет временную продолжительность клеточного цикла.
Активация клеточного деления сопровождается возникновением несбалансированного неравновесного профиля терминальных нуклеотидов, характерной особенностью которого является резкое возрастание содержания ТТА варианта окончания G-цепи теломерной ДНК. При этом его количество достигает уровня 60-70%, что наблюдалось во втором и третьем примерах осуществления изобретения, где исследовались культуры лимфоцитов после 2-х и 4-х часов воздействия ФГА (фиг.3 и 4).
Таким образом, были получены ярко выраженные различия между профилем терминальных нуклеотидов в 1-м примере и профилями во 2-м и 3-м примерах. В первом примере профиль соответствовал культуре, не стимулированной к делению с помощью фитогемагглютинина, в основном неделящегося состояния со слабой естественной активацией на раннем этапе подготовки к делению. Для этого профиля характерно присутствие в достаточно большом количестве окончаний на триплеты TAG и AGG, при этом количество ТТА окончания было не выше 30%. Такой профиль по процентному соотношению вариантов нуклеотидных окончаний относится к частично сбалансированным.
С помощью заявленного способа были проанализированы другие культуры лимфоцитов, не стимулированные с помощью ФГА, для которых была характерна задержка в G0 фазе большинства клеток на фоне отсутствия видимой прогрессии G1 фазы. Анализ таких культур, в полной мере соответствующих требованиям отсутствия активации подготовки к делению, выявил наличие в них полностью сбалансированного равновесного профиля терминальных нуклеотидов. Для него характерно выполнение следующего соотношения процентного содержания терминальных триплетов:
(TAG=TTA=16-20%)+(GGG=GGT=5-7%)=(AGG=GTT=25-27%).
При этом сам профиль напоминал фигуру «симметричной короны» подобно профилю, изображенному на фиг.5. Сравнение такого симметричного профиля с профилем, полученным в 1-м примере, доказывает тот факт, что в исследованной культуре лимфоцитов уже запущены механизмы активации деления клеток. Об этом свидетельствует падение содержания AGG варианта окончания теломерных оверхенгов на фоне умеренного прироста ТТА окончания. Дальнейшую тенденцию изменения профиля терминальных нуклеотидов при активации деления в результате воздействия стимула (ФГА) в течение 2-х и 4-х часов демонстрируют примеры 2 и 3.
Таким образом, возрастание ТТА варианта окончания теломерной ДНК служит признаком начала активации пролиферации клетки и условием нормального прохождения этапов подготовки к делению и самого деления. В ходе подготовки клетки к делению, при условии ее немалигнизации, неравновесный профиль постепенно переходит в равновесное состояние сбалансированности. Этот процесс наглядно демонстрирует 4-й пример осуществления изобретения, в котором исследовалась культура лимфоцитов после 11 часов воздействия фитогемагглютинина. При этом содержание ТТА варианта окончания теломерной ДНК снизилось до 18% и приблизительно сравнялось с количеством TAG варианта. Содержание AGG и GTT вариантов составило приблизительно в равном соотношении по 26% каждый. Как видно из диаграммы фиг.5, полученный профиль полностью соответствует профилю сбалансированного равновесного состояния терминальных нуклеотидов, полученному для культур, не активированных к делению лимфоцитов, покоящихся в G0 фазе.
Впервые с помощью заявленного способа было показано, что инициация клеточного деления под воздействием какого-либо стимула, например ФГА, осуществляется мгновенно и сопровождается перестройками терминальных нуклеотидов теломерной ДНК. При этом переход большинства нуклеотидных окончаний G-цепи теломерной ДНК в окончание ТТА является триггерным механизмом запуска подготовки клетки к делению, а именно, перехода G0 фазы в раннюю G1 фазу.
Использование заявленного способа предоставляет уникальную возможность динамического исследования изменений профиля терминальных нуклеотидов за счет точного установления их процентного соотношения. Возможность построения графиков, отображающих изменения количественного содержания вариантов окончаний теломерных оверхенгов в ходе клеточного цикла выгодно отличает заявляемый способ от всех существующих аналогов. Отслеживание подобной динамики изменений профиля терминальных нуклеотидов позволило установить характер их возникновения.
При рассмотрении фиг.7, где изображена подобная динамика видно, что кривые, соответствующие определенным вариантам нуклеотидных окончаний (триплетов) имеют волнообразный характер. Изменение количества определенного варианта окончания сопряжено с изменениями соседних вариантов. Это наталкивает на мысль о взаимном переходе одних вариантов окончаний оверхенгов в другие в результате экзонуклеазного воздействия на их терминальные нуклеотиды. Такой характер возможного перехода одного варианта окончания оверхенга в другой четко прослеживался в первые 11 часов воздействия стимула к делению и осуществлялся по направлению: AGG→TAG→ТТА→GTT→GGT→GGG→AGG.
В первые два часа сигнал ТТА окончания стремительно возрастал за счет TAG и AGG окончаний в результате последовательного понуклеотидного отщепления AGG→TAG→ТТА. В этот же интервал времени незначительно колебалось содержание триплетов GTT и GGT и GGG за счет последовательных переходов окончаний ТТА→GTT→GGT→GGG. Подобные переходы соседних вариантов окончаний друг в друга отображены на протяжении всего графика фиг.7. При этом четко соблюдались их количественные соотношения. Если снижался уровень одного терминального триплета, то возрастал уровень соседнего с ним триплета со сдвигом в один-два нуклеотида. Существующий механизм понуклеотидного смещения окончаний теломерных оверхенгов является своеобразными «нуклеотидными часами», которые с одной стороны обеспечивают триггерный механизм активации деления, а с другой стороны, определяют существование сбалансированного равновесного профиля терминальных нуклеотидов в течение большего времени клеточного цикла.
Существование сбалансированного равновесного профиля терминальных нуклеотидов на протяжении большей части клеточного цикла является защитным молекулярным механизмом здоровых клеток. Он предотвращает преждевременное начало нового клеточного деления и определяет временную продолжительность клеточного цикла. Для здоровых лимфоцитов периферической крови, стимулированных к делению с помощью ФГА, он составляет приблизительно 48 часов (Morice et al., 1993). Следующий несбалансированный неравновесный профиль не может возникнуть ранее, чем через установленное количество часов после начала первичного воздействия стимулирующего агента. Наступление сбалансированного равновесного состояния, начиная с 10-12 часов после начала стимуляции деления, и его поддержание в ходе большей части клеточного цикла являются важным диагностическим признаком здоровых клеток.
Сокращение продолжительности клеточного цикла за счет более раннего наступления следующего неравновесного состояния профиля терминальных нуклеотидов и его существование в течение более длительного времени, чем фаза G1 клеточного цикла, является характерной особенностью малигнизации клеток. При этом нарушения в образовании сбалансированного равновесного профиля терминальных нуклеотидов, а именно блокировка цикла понуклеотидных переходов окончаний G-цепи теломерной ДНК, могут приводить к длительному преобладанию ТТА варианта ее окончания. Предположительно ТТА окончание G-цепи теломерной ДНК, находясь на конце большинства хромосом, ответственно за активацию в них экспрессии генов, определяющих начало подготовки клетки к делению. В результате длительного преобладания ТТА окончания G-цепи теломерной ДНК создаются условия постоянной активации деления клетки и сокращения продолжительности клеточного цикла, что может являться причиной ракового перерождения.
С помощью заявленного способа было проведено исследование лейкемических культур лимфоцитов человека Jurkat и К-562, синхронизированных в митотической фазе клеточного цикла. Синхронизация клеток осуществлялась в течение 12 часов при добавлении колхицина в культуральную среду RPMI-1640 в конечной концентрации 0,5 мкг/мл. Большая длительность воздействия колхицина обеспечила максимальное накопление клеток в митотической фазе.
Синхронизация лейкемических клеток в метафазе митотического деления выявила отсутствие сбалансированного равновесного профиля терминальных нуклеотидов. При этом степень нарушения сбалансированности профиля коррелировала со степенью злокачественности клеток и скоростью их размножения. Для культуры Jurkat, характеризующейся в два раза большей скоростью деления по сравнению с другой исследуемой культурой, был характерен крайне несбалансированный профиль терминальных нуклеотидов (фиг.8). Этот профиль был представлен в основном окончанием ТТА (82,3%) и очень незначительным присутствием GTT (15,9%) и GGT (1,8%) терминальных триплетов. Отсутствие других вариантов терминальных окончаний, особенно AGG и TAG триплетов, свидетельствовало о блокировке цикла понуклеотидных переходов окончаний G-цепи теломерной ДНК. Результатом частичной блокировки переходов терминальных триплетов TTA→GTT→GGT и полной блокировки переходов GGT→GGG→AGG→TAG явилось отсутствие образования сбалансированного равновесного профиля.
Культуре К-562 также соответствовал несбалансированный профиль терминальных нуклеотидов, но с меньшей степенью несбалансированности (фиг.9). На фоне преобладания ТТА триплета (57,5%) присутствовали окончания AGG (22,1%), GGT (14,5%) и GTT (5,9%), которые свидетельствовали о снижении прогрессии цикла понуклеотидных переходов окончаний в сторону образования сбалансированного равновесного профиля. Характерной особенностью обоих культур явилось отсутствие TAG варианта окончания, что, видимо, связано с его полным переходом в ТТА триплет.
Таким образом, установление наличия или отсутствия сбалансированного равновесного профиля терминальных нуклеотидов, а также степени его несбалансированности при синхронизации клеток в специфичных для него фазах клеточного цикла (например, митотической), является важным диагностическим критерием в онкологии. Изучение изменений терминальных нуклеотидов в ходе клеточного цикла раковых клеток, а именно, продолжительности существования неравновесного профиля и нарушений, связанных с возникновением его равновесного состояния, может служить признаком, определяющим степень малигнизации клеток.
Применение заявленного способа позволяет уловить минимальные изменения в процентном содержании тех или иных вариантов нуклеотидных окончаний теломерной ДНК и может служить универсальным инструментом исследования клеточного цикла любых клеток и тканей. Профили терминальных нуклеотидов G-цепи теломерной ДНК могут использоваться для определения пролиферативного статуса разнообразных клеток и тканей и являться критерием оценки их способности к активации деления. Например, составление профилей терминальных нуклеотидов теломерной ДНК иммунных клеток может выявлять степень их активации и готовности к делению и являться критерием иммунного статуса организма. Исследование пролиферативного потенциала стволовых клеток, а также любых способных к делению клеток, с использованием заявленного способа может найти применение в трансплантологии как критерий приживаемости клеток, в косметологии, дерматологии и геронтологии как фактор регенерационных способностей тканей и потенциала организма к омоложению в целом.




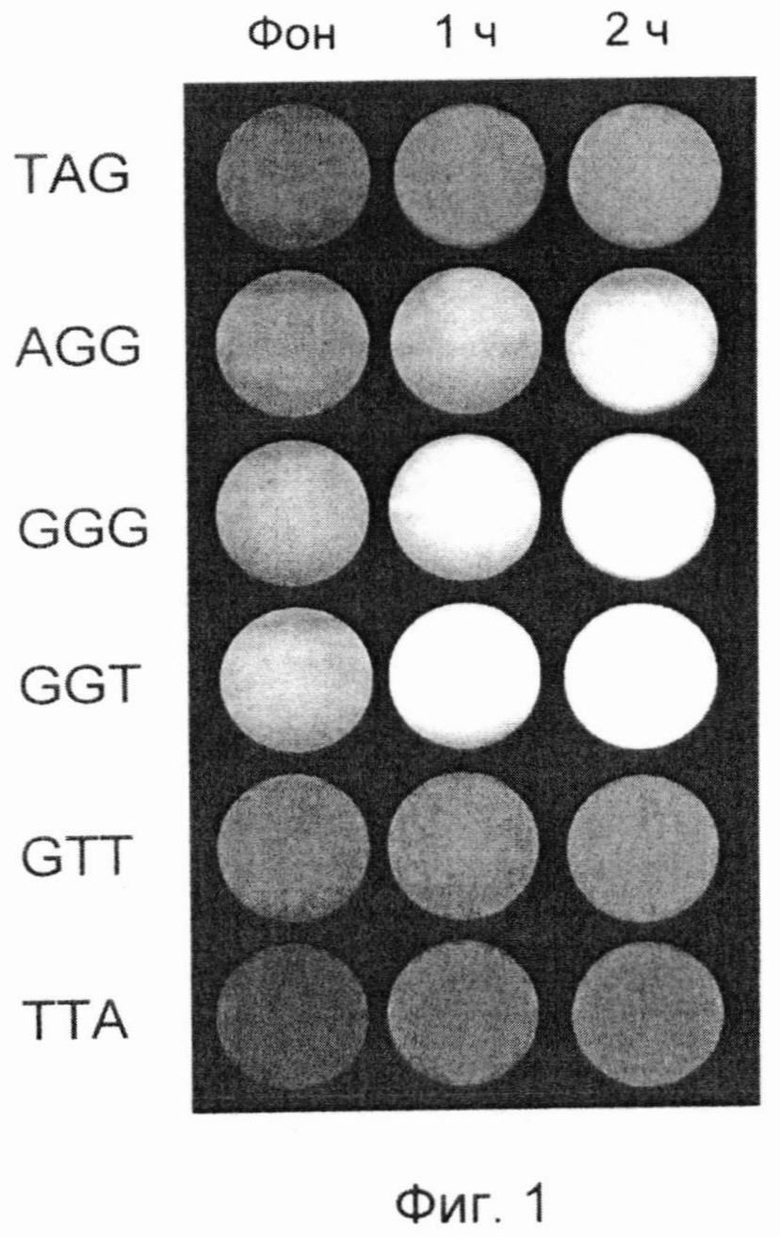

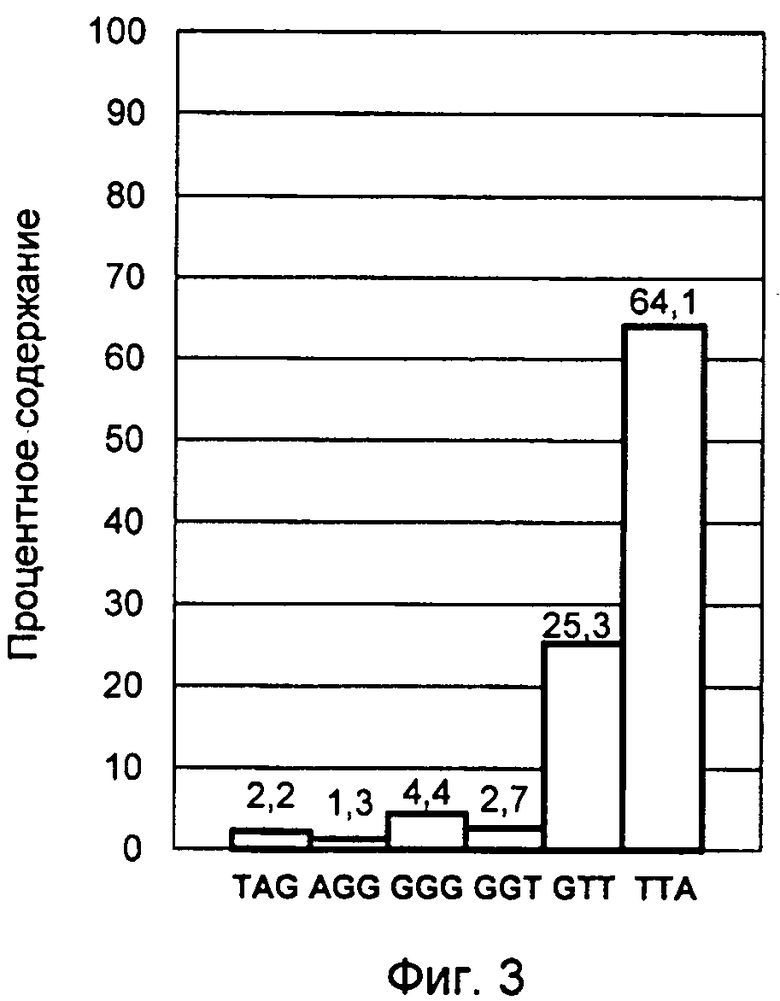
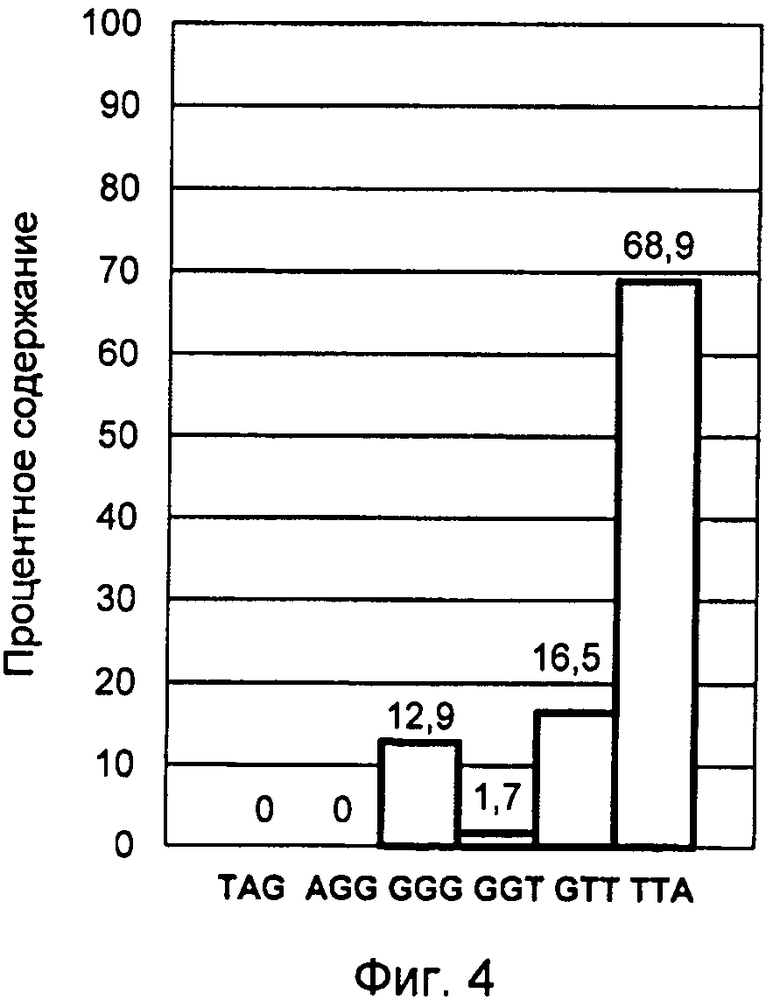
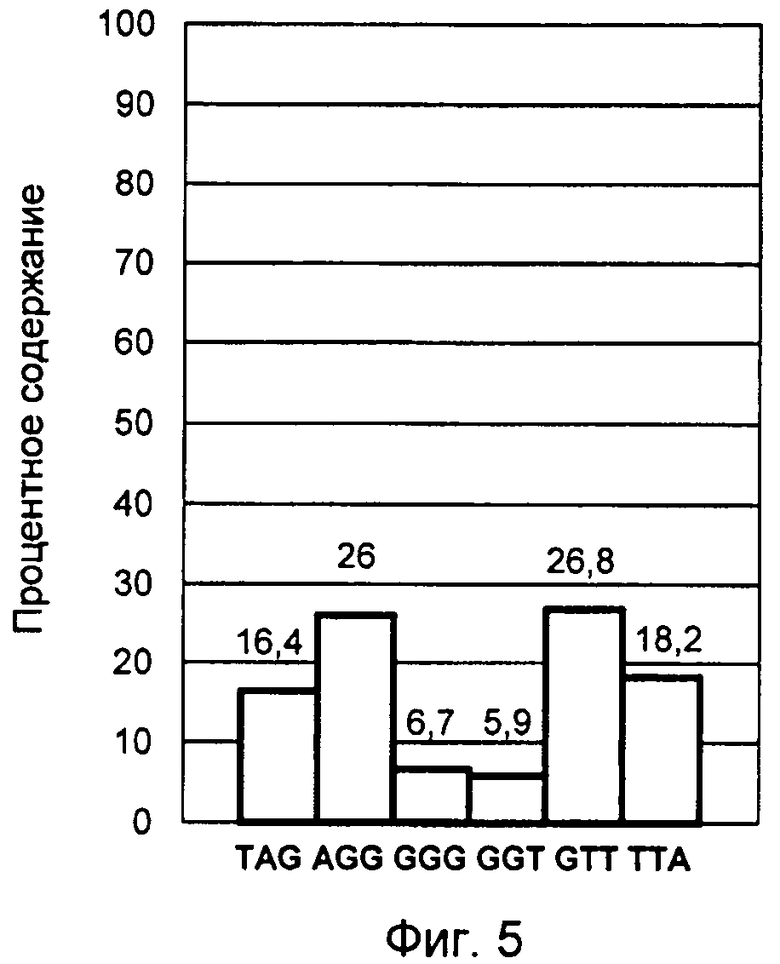
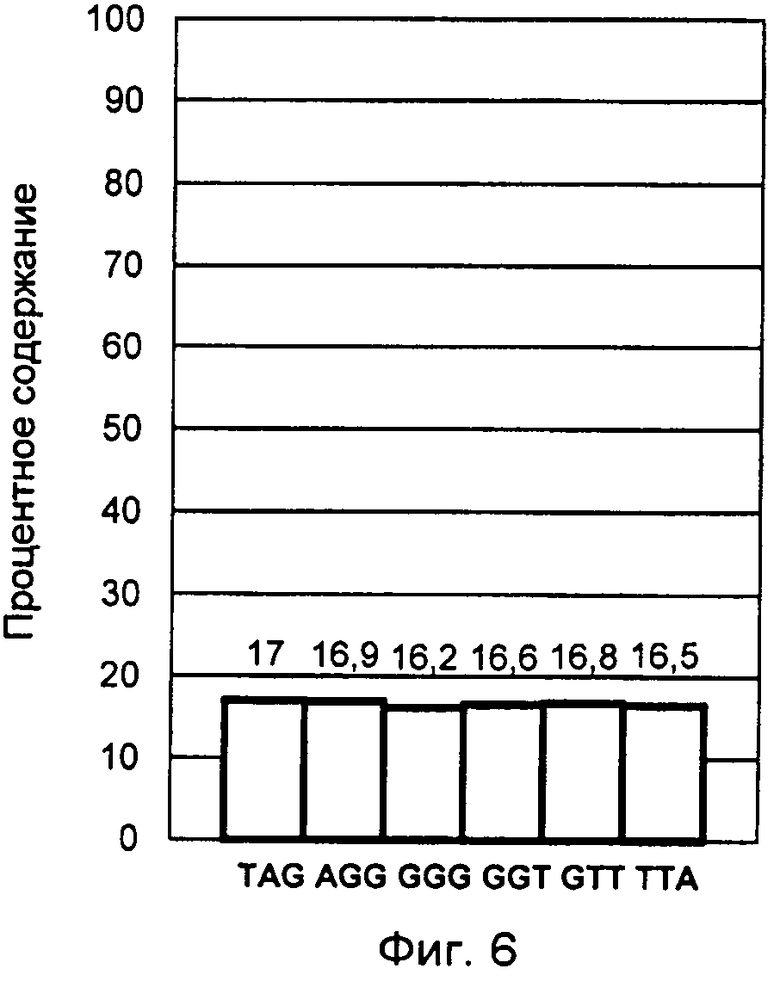
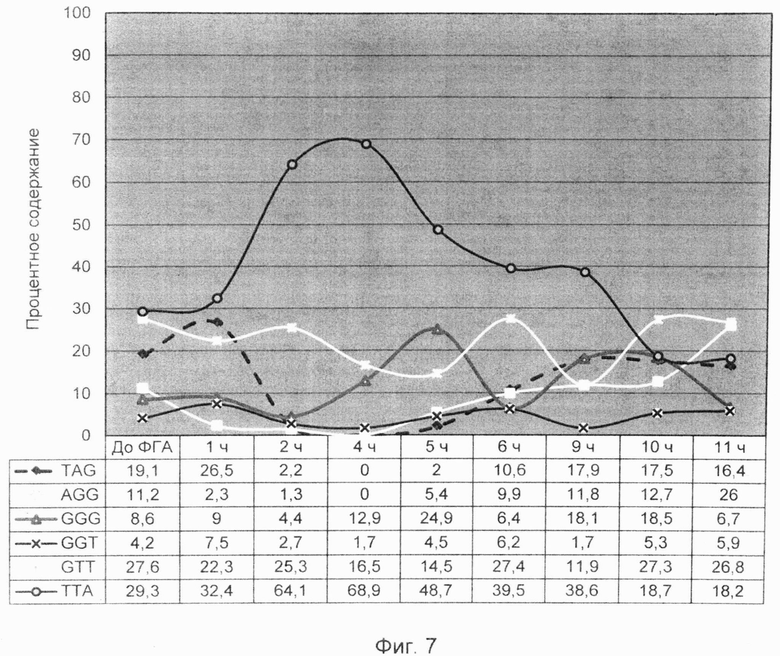
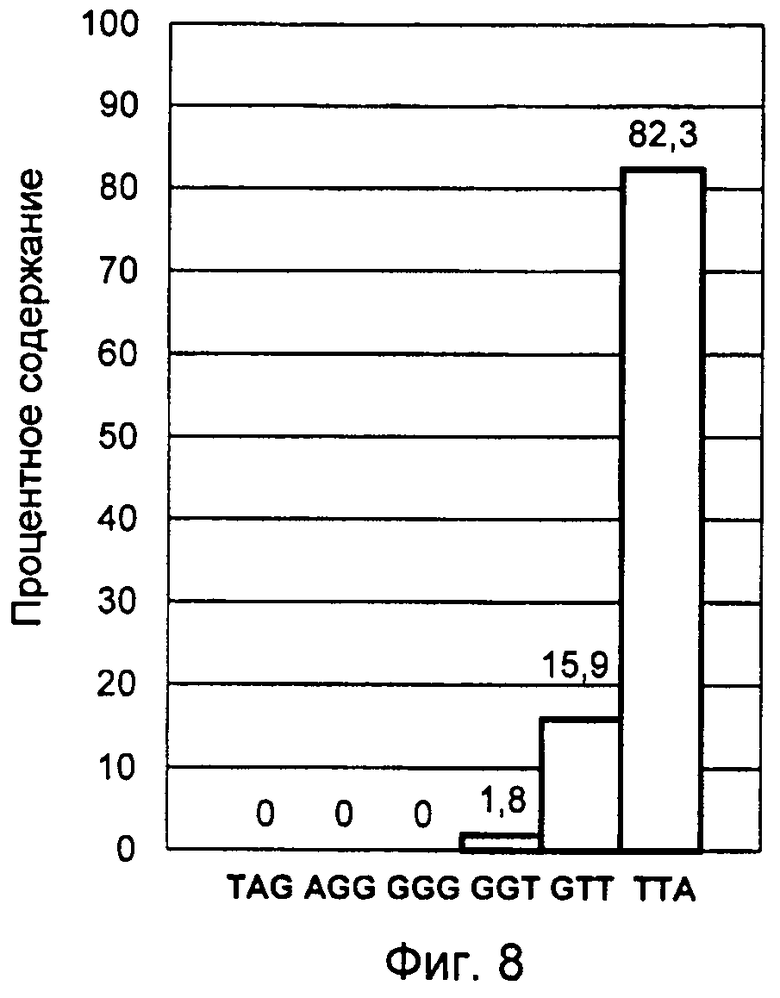
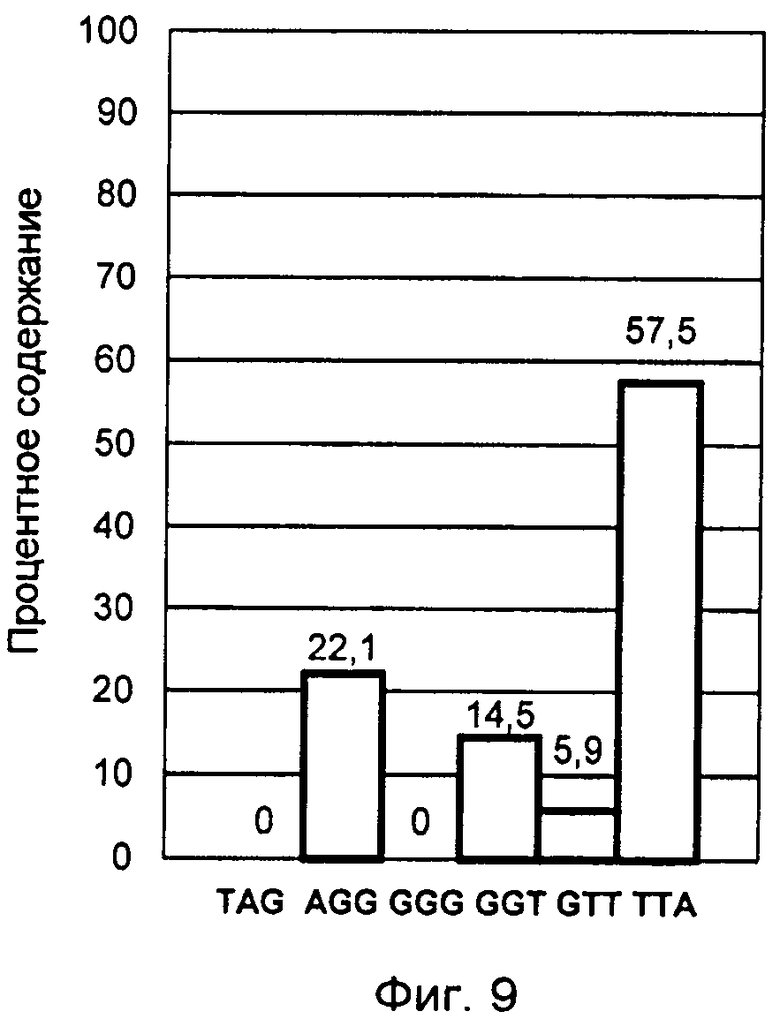
Изобретение относится к молекулярной биологии и генетике клетки. Предложен способ, включающий этапы предварительной экстракции геномной ДНК, выделения специфической фракции однонитевых G-оверхенгов теломерной ДНК и последующей амплификации их минусовой цепи с дуплекс-специфическим анализом, причем этапы амплификации включают модификацию 3'-концов теломерных оверхенгов с помощью терминальной дезоксинуклеотидилтрансферазы и осуществляются с использованием набора праймеров SEQ ID NO: 1-5. Дуплекс-специфический анализ основан на гибридизации минусовых цепей оверхенгов с набором специфических флуоресцентно-меченных зондов SEQ ID NO: 6-11, соответствующих шести возможным вариантам нуклеотидных окончаний G-цепи теломерной ДНК человека. Степень разгорания зондов в присутствии фермента дуплекс-специфической нуклеазы служит критерием присутствия соответствующего варианта нуклеотидного окончания и мерой его количественного содержания. Полученные профили терминальных нуклеотидов G-цепи теломерной ДНК служат пролиферативными маркерами, указывающими на степень активации деления клетки и определяющими временную продолжительность клеточного цикла. Изобретение является новой разновидностью ДНК-диагностики, предназначено для выявления изменений нуклеотидных окончаний ДНК теломерных областей хромосом, происходящих в ходе клеточного цикла, и может быть использовано в медицине для диагностики и лечения онкологических заболеваний, в иммунологии, трансплантологии, косметологии, дерматологии, геронтологии, клеточной биотехнологии и других областях, заинтересованных в оценке пролиферативного статуса клетки и методах его направленного регулирования. 3 н. и 12 з.п. ф-лы, 9 ил., 8 табл., 5 пр.
1. Способ количественной оценки терминальных нуклеотидов G-цепи теломерной ДНК человека, включающий следующие этапы:
а) экстракция геномной ДНК с последующим выделением фракции 3'-выступающих однонитевых концов теломерной ДНК (G-оверхенгов) при помощи магнитных частиц с иммобилизованными на их поверхности зондами, комплементарными повторам G-цепи теломерной ДНК;
б) амплификация полученной фракции G-оверхенгов, осуществляемая в следующие подэтапы: 1) 3'-модифицирование за счет присоединения полидезоксицитидиловой и расположенной за ней полидезоксиадениловой последовательностей с помощью фермента терминальной дезоксинуклеотидилтрансферазы; 2) амплификация с использованием праймера SEQ ID NO:1 с целью получения минусовых копий, содержащих адаптерную последовательность SEQ ID NO:3; 3) 3'-модифицирование минусовых копий за счет присоединения полидезоксиадениловой последовательности с помощью фермента терминальной дезоксинуклеотидилтрансферазы; 4) финальная ПЦР амплификация минусовых цепей с использованием пары праймеров, указанных в SEQ ID NO:3 и SEQ ID NO:4, в условиях избытка адаптерной последовательности;
в) проведение дуплекс-специфического анализа полученной фракции минусовых цепей G-оверхенгов с использованием олигонуклеотидных зондов, флуоресцентно-меченных красителем FAM и гасителем RTQ1, соответствующих шести возможным вариантам триплетных нуклеотидных окончаний G-цепи теломерной ДНК человека и указанных в SEQ ID NO: 6-11;
г) регистрация результатов дуплекс-специфического анализа с помощью детектора FDG-001 и программы LabWorks 4.6, включающая измерение уровней флуоресценции зондов в ходе гидролиза в присутствии фермента дуплекс-специфической нуклеазы, математическую обработку данных, заключающуюся в нормировании сигналов за счет вычитания догидролизного фона и умножения на специфические коэффициенты, представленные в табл. 3, и конечную оценку количественного соотношения уровней сигналов зондов, соответствующих терминальным триплетам G-оверхенгов в виде построения профиля терминальных нуклеотидов G-цепи теломерной ДНК в процентах (фиг. 2).
2. Способ по п.1, отличающийся тем, что для выделения фракции теломерных G-оверхенгов на этапе (а) используют способ, сохраняющий нативность одноцепочечной ДНК из группы гибридизационных или сорбционных методов с применением различных мембран, колонок, частиц или других поверхностей с целью иммобилизации и последующего снятия с их поверхности исследуемых молекул.
3. Способ по п.1, отличающийся тем, что для амплификации фракции теломерных G-оверхенгов на этапе (б) после полидезоксицитидилирования проводят политимидилирование с помощью фермента терминальной дезоксинуклеотидилтрансферазы, далее последовательно осуществляют амплификацию минусовой цепи с использованием праймера SEQ ID NO: 2, 3'-модифицирование полученных минусовых копий через присоединение политимидиловой последовательности с помощью фермента терминальной дезоксинуклеотидилтрансферазы и их финальную амплификацию с использованием пары праймеров, состоящей из указанных в SEQ ID NO: 3 и SEQ ID NO: 5 последовательностей, при условии избытка адаптерного праймера.
4. Способ по п.1 или 3, отличающийся тем, что для амплификации фракции G-оверхенгов на этапе (б) в качестве адаптерной последовательности, содержащейся в SEQ ID NO: 1-3, используется подходящая по структуре и температуре отжига олигонуклеотидная последовательность, не комплементарная повторам G-цепи теломерной ДНК человека.
5. Способ по п.1, отличающийся тем, что при проведении дуплекс-специфического анализа на этапе (в) вместо зондов, представленных в SEQ ID NO: 6-11 используются варианты длиной 10- 12 нуклеотидов.
6. Способ по п.1, отличающийся тем, что при проведении дуплекс-специфического анализа на этапе (в) используется один вид или сочетание нескольких видов флуоресцентного мечения зондов с комбинацией «флуорофор-гаситель» на любом из противоположных концов зонда, состоящей из красителей FITC, FAM, R110, ТЕТ, R6G, HEX, JOE, VIC, YakimaYellow, TAMRA, Cy3, ROX, SuperROX, Cy 3.5, Cy 5, Cy 5.5, Cy 7, Cy 7.5 и аналогичных им и соответствующих гасителей из ряда RTQ, BHQ, Dabcyl и подобных им.
7. Способ по п.1, отличающийся тем, что для регистрации результатов дуплекс-специфического анализа на этапе (г) используются виды аппаратно-программного обеспечения, позволяющие количественно определять уровень значимого флуоресцентного сигнала для каждого зонда, в том числе и мультиканально при сочетании нескольких видов мечения по п.6.
8. Способ по п.1, отличающийся тем, что математическая обработка уровней сигналов при регистрации результатов дуплекс-специфического анализа на этапе (г) заключается в нормировании значений умножением на коэффициенты, приведенные в таблице 3, для каждого зонда из группы FAM-RTQ1 после 1 и 2 часов проведения анализа.
9. Способ по п.1, отличающийся тем, что математическая обработка уровней сигналов при регистрации результатов дуплекс-специфического анализа на этапе (г) заключается в нормировании значений умножением на коэффициенты, отличные от таблицы 3 и рассчитанные для вариантов зондов в различных концентрационных соотношениях, включая указанные, для групп флуоресцентного мечения по п.6 и соответствующие текущему состоянию спектральных свойств флуорофор-связанной флуоресценции ДНК на момент исследования.
10. Способ по п.1, отличающийся тем, что полученные в ходе его осуществления результаты применяются для целей медицинской диагностики заболеваний и старения организма, оценки пролиферативных свойств и тканеспецифических функций клеток и тканей, определения прогноза их развития и выбора специфической терапии с использованием в качестве критерия определенного профиля терминальных нуклеотидов G-цепи теломерной ДНК человека.
11. Набор специфических дезоксирибоолигонуклеотидов, представленных в SEQ ID NO: 1-5, используемых в качестве праймеров для амплификации минусовых цепей теломерных G-оверхенгов в ходе ПЦР в способе по п.1.
12. Набор специфических дезоксирибоолигонуклеотидов по п.11, используемых в качестве праймеров для амплификации минусовых цепей теломерных G-оверхенгов в ходе ПЦР, отличающийся тем, что в качестве адаптерной последовательности, содержащейся в SEQ ID NO: 1-3, используется подходящая по структуре и температуре отжига дезоксирибоолигонуклеотидная последовательность, не комплементарная повторам G-цепи теломерной ДНК человека.
13. Набор специфических дезоксирибоолигонуклеотидов, представленных в SEQ ID NO: 6-11, используемых в качестве флуоресцентно-меченных зондов при проведении дуплекс-специфического анализа в способе по п.1.
14. Набор специфических дезоксирибоолигонуклеотидов по п. 13, используемых в качестве флуоресцентно-меченных зондов длиной 10- 12 нуклеотидов.
15. Набор специфических дезоксирибоолигонуклеотидов по п.13, используемых в качестве флуоресцентно-меченных зондов при проведении дуплекс-специфического анализа, в котором применяется один вид или сочетание нескольких видов флуоресцентного мечения с комбинацией «флуорофор-гаситель» на любом из противоположных концов зонда, состоящей из красителей FITC, FAM, R110, ТЕТ, R6G, HEX, JOE, VIC, Yakima Yellow, TAMRA, Cy 3, ROX, SuperROX, Cy 3.5, Cy 5, Cy 5.5, Cy 7, Cy 7.5 и аналогичных им и соответствующих гасителей из ряда RTQ, BHQ, Dabcyl и подобных им.
| US 20020091281 A1, 11.07.2002 | |||
| WO 2009021518 A1, 19.02.2009 | |||
| US 20090306182 A1, 10.12.2009 | |||
| СПОСОБ ИЗМЕРЕНИЯ ДЛИНЫ ТЕЛОМЕР КЛЕТОК ЛЕЙКОКОНЦЕНТРАТА ПУПОВИННОЙ КРОВИ | 2010 |
|
RU2434940C1 |

