Область техники
Настоящее изобретение относится к способам амплификации последовательности нуклеиновой кислоты-мишени, включающим внедрение в цепь. Изобретение также относится к наборам и композициям, подходящим для применения в данных способах.
Уровень техники
Способы амплификации последовательности нуклеиновой кислоты-мишени путем внедрения в цепь были описаны, например, в публикации международной заявки WO 2009/150467. Внедрение в последовательность нуклеиновой кислоты-мишени опосредуется одноцепочечным олигонуклеотидом для внедрения, который расплетает дуплекс-мишень, обеспечивая возможность связывания прямого и обратного праймера.
Краткое описание фигур
Фигура 1: Амплификация ДНК-мишени с использованием двух олигонуклеотидов для внедрения при их параллельной конфигурации, антипараллельной конфигурации или обратной антипараллельной конфигурации. IO1 - первый олигонуклеотид для внедрения; IO2 - второй олигонуклеотид для внедрения. F-праймер - прямой праймер или праймер, располагающийся в 3'-5' направлении; R-праймер - обратный праймер или праймер, располагающийся в 5'-3' направлении. Неудлиняемые концы IO1 и IO2 обозначены пунктирной линией.
Фигура 2: Амплификация ДНК-мишени с использованием двух олигонуклеотидов для внедрения. Показаны графики амплификации при (а) параллельной конфигурации олигонуклеотидов для внедрения, (с) антипараллельной конфигурации олигонуклеотидов для внедрения и (е) обратной антипараллельной конфигурации олигонуклеотидов для внедрения. Представлены результаты двух реакций. За процессом амплификации следили путем детекции красителя Sybr Green I. Ось X на графиках амплификации: время (минуты), ось Y: флуоресценция красителя Sybr Green I (интенсивность флуоресценции, относительные единицы). Специфичность реакций дополнительно оценивали с помощью анализа кривых плавления. Результаты анализа кривых плавления показаны для параллельной конфигурации олигонуклеотидов для внедрения (b), антипараллельной конфигурации олигонуклеотидов для внедрения (d) и антипараллельной обратной конфигурации олигонуклеотидов для внедрения (f). На оси X показан анализ кривых плавления: температура (градусы Цельсия), ось Y: -d(флуоресценция)/d(температура) (относительные единицы).
Фигура 3: Амплификация ДНК-мишени с использованием двух олигонуклеотидов для внедрения. Реакции проводили либо с использованием двух комплементарных олигонуклеотидов для внедрения, либо с использованием одного комплементарного олигонуклеотида для внедрения и одного некомплементарного олигонуклеотида для внедрения. Оси X и Y для графиков амплификации являются таким же, как на Фигуре 2. (а) - показаны результаты амплификации при параллельной конфигурации используемых олигонуклеотидов. (b) - показаны результаты амплификации при антипараллельной конфигурации олигонуклеотидов. Представлены результаты двух реакций.
Фигура 4: Специфичность праймеров в реакции амплификации с использованием двух олигонуклеотидов для внедрения. Реакции проводили либо с использованием комплементарных прямого и обратного праймеров, либо с использованием комплементарного прямого и некомплементарного обратного праймера. Концентрация ДНК-мишени составляла 1 пкМ. Оси X и Y для графика амплификации являются такими же, как на Фигуре 2.
Фигура 5: Совместимость реакции с использованием двух олигонуклеотидов для внедрения с мишень-специфичными зондами, (а) - показано схематичное изображение конфигураций, позволяющих использование мишень-специфичных зондов. (b) и (с) - показана амплификация и выявление в режиме реального времени ДНК-мишени при использовании (b) двух олигонуклеотидов для внедрения или (с) одного олигонуклеотида для внедрения (SIBA). За амплификацией следили в режиме реального времени либо с помощью красителя Sybr green I, либо с помощью мишень-специфичного зонда, как указано в подписях для кривых. Ось X для каждого графика: время (минуты). Ось Y: флуоресценция красителя Sybr green I или зонда (относительные единицы).
Фигура 6: Устойчивость (а) реакции с использованием двух олигонуклеотидов для внедрения в цепь и (b) стандартной реакции с использованием одноцепочечного олигонуклеотида для внедрения в цепь (SIBA) к выявлению неспецифической амплификации. Стандартная SIBA была менее устойчивой к выявлению неспецифической амплификации при использовании коротких праймеров по сравнению с реакцией амплификации с использованием двух олигонуклеотидов для внедрения в цепь. Концентрация ДНК-мишени составляла 1 пкМ для длинных праймеров и 1 фМ для коротких праймеров. Контроль амплификации осуществляли с помощью Sybr green I или зонда, имеющего сайт связывания, не перекрывающийся с сайтами связывания олигонуклеотидов для внедрения в цепь или праймеров. Ось X для каждого графика: время (минуты). Ось Y: флуоресценция красителя Sybr green I или зонда (относительные единицы), (а) - показаны результаты контроля за амплификацией с помощью красителя Sybr green I или зонда во время амплификации с использованием двух олигонуклеотидов для внедрения в цепь. (b) - показаны результаты контроля за амплификацией с помощью Sybr green I в реакции SIBA.
Фигура 7: Амплификация ДНК-мишени из плазмидной ДНК с использованием двух олигонуклеотидов для внедрения в цепь. Плазмидную ДНК использовали непосредственно или после обработки ферментом для рестрикции EcoRV-HF. Контроль амплификации осуществляли с использованием красителя Sybr green I. Ось X: время (минуты). Ось Y: флуоресценция Sybr green I (относительные единицы).
Фигура 8: Амплификация ДНК-мишени, имеющей два идентичных сайта внедрения. Реакции осуществляли с одним олигонуклеотидом для внедрения, который связывался с обоими сайтами для внедрения в ДНК-мишень, (а), (с), (е) и (g) - показаны графики амплификации по результатам мониторинга амплификации ДНК-мишени в режиме реального времени с использованием Sybr green I. (b), (d), (f) и (h) - показаны результаты соответствующих анализов кривых плавления, (i) - показаны результаты электрофоретического анализа продуктов реакции в неденатурирующих условиях, а) и b): параллельная конфигурация олигонуклеотидов для внедрения в цепь, используемых для амплификации дуплекса ДНК-мишени, содержащего 324 п.н. (с) и (d): параллельная конфигурация олигонуклеотидов для внедрения в цепь, используемых для амплификации ДНК-мишени, (е) и (f): антипараллельная конфигурация олигонуклеотидов для внедрения в цепь, используемых для амплификации ДНК-мишени, (g) и (h): обратная антипараллельная конфигурация олигонуклеотидов для внедрения в цепь, используемых для амплификации ДНК-мишени, (i) антипараллельная конфигурация олигонуклеотидов для внедрения в цепь, используемых для амплификации ДНК-мишени. Оси X и Y графиков амплификации и анализов кривых плавления являются такими же, как на Фигуре 2. Контроль реакции амплификации ДНК-мишени осуществляли в режиме реального времени с использованием красителя Sybr green I. (i) Дорожки на электрофореграмме: дорожка 1: линейка маркеров молекулярной массы BioRad EZ, нагрузка 20 п.н.; дорожки 2-6: 107, 106, 105, 104 и 103 копий соответственно; дорожка 7 - контроль (вода).
Фигура 9: Система на основе FRET (резонансного переноса энергии флуоресценции) для отслеживания внедрения и амплификации в режиме реального времени: (а) схематичное изображение меченых праймеров и олигонуклеотидов для внедрения в цепь. (b) Результаты мониторинга внедрения и амплификации в режиме реального времени и выявления ДНК-мишени с использованием FRET-меченных олигонуклеотидов при параллельной конфигурации. Ось X для (b): время (минуты). Ось Y: флуоресценция зонда (относительные единицы).
Фигура 10: Чувствительность амплификации на основе внедрения в цепь с использованием двух олигонуклеотидов. Чувствительность оценивали с помощью трех различных анализов с использованием серийных разведений ДНК-мишени (от 106 копий до 1 копии). Амплификацию ДНК-мишени контролировали в режиме реального времени с помощью красителя Sybr green I. Графики амплификации: (а) анализ 1, (b) анализ 2 и (с) анализ 3. Ось X: время (минуты). Ось Y: флуоресценция Sybr green I (относительные единицы).
Краткое описание последовательностей
SEQ ID NO: 1 представляет собой нуклеотидную последовательность олигонуклеотида для внедрения в цепь.
SEQ ID NO: 2 представляет собой нуклеотидную последовательность олигонуклеотида для внедрения в цепь.
SEQ ID NO: 3 представляет собой нуклеотидную последовательность ДНК-праймера.
SEQ ID NO: 4 представляет собой нуклеотидную последовательность ДНК-праймера.
SEQ ID NO: 5 представляет собой нуклеотидную последовательность ДНК-праймера.
SEQ ID NO: 6 представляет собой нуклеотидную последовательность некомплементарного олигонуклеотида для внедрения в цепь.
SEQ ID NO: 7 представляет собой нуклеотидную последовательность некомплементарного ДНК-праймера.
SEQ ID NO: 8 представляет собой нуклеотидную последовательность зонда.
SEQ ID NO: 9 представляет собой нуклеотидную последовательность зонда.
SEQ ID NO: 10 представляет собой нуклеотидную последовательность ДНК-праймера.
SEQ ID NO: 11 представляет собой нуклеотидную последовательность матрицы-мишени.
SEQ ID NO: 12 представляет собой нуклеотидную последовательность матрицы-мишени.
SEQ ID NO: 13 представляет собой нуклеотидную последовательность олигонуклеотида для внедрения в цепь.
SEQ ID NO: 14 представляет собой нуклеотидную последовательность ДНК-праймера.
SEQ ID NO: 15 представляет собой нуклеотидную последовательность ДНК-праймера.
SEQ ID NO: 16 представляет собой нуклеотидную последовательность матрицы-мишени.
SEQ ID NO: 17 представляет собой нуклеотидную последовательность матрицы-мишени.
SEQ ID NO: 18 представляет собой нуклеотидную последовательность матрицы-мишени.
SEQ ID NO: 19 представляет собой нуклеотидную последовательность матрицы-мишени.
SEQ ID NO: 20 представляет собой нуклеотидную последовательность матрицы-мишени.
SEQ ID NO: 21 представляет собой нуклеотидную последовательность меченого олигонуклеотида для внедрения в цепь.
SEQ ID NO: 22 представляет собой нуклеотидную последовательность меченого олигонуклеотида для внедрения в цепь.
SEQ ID NO: 23 представляет собой нуклеотидную последовательность меченого праймера.
SEQ ID NO: 24 представляет собой нуклеотидную последовательность меченого праймера.
SEQ ID NO: 25 представляет собой нуклеотидную последовательность ДНК-праймера.
SEQ ID NO: 26 представляет собой нуклеотидную последовательность матрицы-мишени.
SEQ ID NO: 27 представляет собой адаптерную нуклеотидную последовательность.
SEQ ID NO: 28 представляет собой адаптерную нуклеотидную последовательность.
SEQ ID NO: 29 представляет собой адаптерную нуклеотидную последовательность.
SEQ ID NO: 30 представляет собой адаптерную нуклеотидную последовательность.
SEQ ID NO: 31 представляет собой адаптерную нуклеотидную последовательность.
Краткое описание изобретения
Настоящее изобретение относится к системе для внедрения в цепь последовательности нуклеиновой кислоты-мишени по меньшей мере в двух положениях. В способах согласно изобретению используется один или более олигонуклеотидов для внедрения в цепь, связывающихся с 3'-5' и 5'-3' участками последовательности нуклеиновой кислоты-мишени и вызывающих внедрение в них, обеспечивая возможность связывания прямого и обратного праймеров для осуществления амплификации указанной последовательности нуклеиновой кислоты-мишени. Внедрение в цепь последовательности нуклеиновой кислоты-мишени как в 3'-5', так и в 5'-3' положении обеспечивает сочетание каждого события связывания праймера с независимым событием внедрения в цепь и повышает возможность использования последовательностей олигонуклеотидов для внедрения в цепь, не перекрывающихся с последовательностями праймеров для амплификации. Опосредованное внедрение в цепь в двух различных положениях также обеспечивает преимущества, которые заключаются в возможности амплификации более длинных последовательностей нуклеиновых кислот-мишеней по сравнению с последовательностями, которые обычно амплифицируют с помощью внедрения в цепь в одном положении.
Кроме того, один вид олигонуклеотида для внедрения в цепь может вызывать внедрение в цепь как в 5'-3', так и в 3'-5' положении, при условии что подходящие связывающие последовательности присутствуют в обоих участках последовательность-мишени. Подобным образом, можно использовать один вид праймера, когда подходящая связывающая последовательность присутствует в обоих участках последовательность-мишени. Указанные варианты реализации изобретения обеспечивают возможность амплификации и секвенирования неизвестных последовательностей, где в матрице, содержащей последовательность-мишень, присутствуют известные участки связывания (такие как адаптерные последовательности). Также могут быть созданы олигонуклеотиды для внедрения в цепь, связывающиеся с 3'-5' и 5'-3' участками связывания дуплекса последовательности нуклеиновой кислоты-мишени в альтернативных конфигурациях, что позволяет создавать различные последовательности, нацеленные на конкретный ампликон, для оптимизации параметров амплификации. Более того, олигонуклеотиды для внедрения в цепь и праймеры могут быть созданы таким образом, чтобы их участки связывания не перекрывались, в результате чего участок ампликона остается свободным для связывания зонда, что, таким образом, приводит к снижению конкуренции между разными видами олигонуклеотидов за связывание с ампликоном во время амплификации и предупреждает выявление продуктов неспецифической амплификации.
Таким образом, настоящее изобретение обеспечивает способ амплификации последовательности нуклеиновой кислоты-мишени, включающий приведение указанной последовательности нуклеиновой кислоты-мишени в контакт с по меньшей мере с одним прямым праймером, по меньшей мере одним обратным праймером и первым и вторым олигонуклеотидами для внедрения в цепь в условиях, обеспечивающих амплификацию указанной последовательности нуклеиновой кислоты-мишени, где первый олигонуклеотид для внедрения в цепь делает участок в 3'-5' направлении связывания последовательности нуклеиновой кислоты-мишени одноцепочечным для обеспечения возможности связывания прямого праймера, и второй олигонуклеотид для внедрения в цепь делает 5'-3' участок связывания последовательности нуклеиновой кислоты-мишени одноцепочечным для обеспечения возможности связывание обратного праймера.
Изобретение также обеспечивает способ амплификации последовательности нуклеиновой кислоты-мишени, содержащей 3'-5' и 5'-3' участки связывания олигонуклеотида для внедрения в цепь, включающий приведение указанной последовательности нуклеиновой кислоты-мишени в контакт с олигонуклеотидом для внедрения в цепь и одним или более праймерами, способными обеспечивать амплификацию последовательности нуклеиновой кислоты-мишени, где указанный олигонуклеотид для внедрения в цепь делает 3'-5' и 5'-3' участки связывания олигонуклеотидов для внедрения в цепь в последовательности нуклеиновой кислоты-мишени одноцепочечными для обеспечения возможности связывания указанного одного или более праймеров.
Изобретение также обеспечивает набор, включающий по меньшей мере один прямой и по меньшей мере один обратный праймер для последовательности нуклеиновой кислоты-мишени и первый и второй олигонуклеотиды для внедрения в цепь, которые имеют участки связывания в 3'-5' и 5'-3' направлении соответственно в последовательности нуклеиновой кислоты-мишени.
Изобретение также обеспечивает набор, включающий олигонуклеотид для внедрения в цепь и один или более праймеров и по меньшей мере один ДНК-адаптер, где указанный олигонуклеотид для внедрения в цепь может связываться с ДНК-адаптером при его наличии в 3'-5' и 5'-3' участке связывания последовательности нуклеиновой кислоты-мишени, и где указанные один или более праймеров способны обеспечивать амплификацию указанной последовательности нуклеиновой кислоты-мишени.
Изобретение также обеспечивает способ амплификации последовательности нуклеиновой кислоты-мишени, содержащей область неизвестной последовательности, включающий создание последовательности нуклеиновой кислоты-мишени, содержащей участки связывания олигонуклеотидов для внедрения в цепь, расположенные выше и ниже указанной обалсти неизвестной последовательности, и осуществление способа согласно изобретению с использованием олигонуклеотидов для внедрения в цепь и праймеров для амплификации указанной последовательности нуклеиновой кислоты-мишени.
Изобретение также обеспечивает способ определения последовательности нуклеиновой кислоты-мишени, содержащей область неизвестной последовательности, включающий создание последовательности нуклеиновой кислоты-мишени, содержащей участки связывания олигонуклеотидов для внедрения в цепь, расположенные выше и ниже указанной области неизвестной последовательности, осуществление способа согласно изобретению с использованием олигонуклеотидов для внедрения в цепь и праймеров для амплификации последовательности нуклеиновой кислоты-мишени и определение последовательности указанной неизвестной области.
Подробное описание изобретение
Необходимо понимать, что различные варианты применения описанных способов могут быть адаптированы для конкретных нужд в данной области техники. Также необходимо понимать, что используемые в настоящей заявке термины употребляются исключительно в целях описания конкретных вариантов реализации изобретения и не ограничивают настоящее изобретение. Кроме того, употребление предметов в единственном числе в настоящей заявке и прилагаемой формуле изобретения включает множественное число указанных предметов, если иное явно не следует из контекста. Таким образом, ссылка, например, на «полипептид» включает два или более таких полипептидов и т.п. Все публикации, патенты и заявки на патенты, цитированные в настоящей заявке выше и далее, полностью включены в настоящую заявку посредством ссылки.
Способы амплификации последовательности нуклеиновой кислоты-мишени
Способы согласно изобретению обеспечивают амплификацию последовательности нуклеиновой кислоты-мишени путем внедрения в цепь нуклеиновой кислоты в двух отдельных положениях. Внедрение в цепь в каждом положении, опосредованное олигонуклеотидом для внедрения в цепь, приводит к тому, что последовательность нуклеиновой кислоты-мишени становится одноцепочечной для обеспечения возможности связывания праймера. Праймеры, как правило, не способны амплифицировать последовательность нуклеиновой кислоты-мишени при взаимодействии с ней в отсутствие олигонуклеотида для внедрения в цепь (олигонуклеотидов). Другими словами, праймеры не способны связываться со своими участками связывания в последовательности нуклеиновой кислоты-мишени, если их участки связывания не были подвержены действию олигонуклеотидов для внедрения в цепь, которые делают указанные участкие связывания праймеров одноцепочечными. Олигонуклеотиды для внедрения в цепь также, как правило, не способны к удлинению при помощи ДНК-полимеразы. В частности, способы согласно изобретению предпочтительно позволяют амплифицировать последовательности нуклеиновой кислоты-мишени при изотермических условиях, при которых указанная последовательность нуклеиновой кислоты-мишень присутствует в виде дуплекса нуклеиновых кислот. Внедрение в цепь по меньшей мере в двух сайтах дуплекса приводит к тому, что последовательность нуклеиновой кислоты-мишени становится одноцепочечной при изотермических условиях, обеспечивая возможность опосредованной праймером амплификации.
Последовательности нуклеиновой кислоты-мишени
Последовательность нуклеиновой кислоты-мишени может иметь любое происхождение и может, например, быть искусственной или природной. Последовательность нуклеиновой кислоты-мишени может содержать известную последовательность или участки известной и неизвестной последовательности. Последовательность нуклеиновой кислоты-мишень может представлять собой последовательность нуклеиновой кислоты человека, млекопитающего, бактерий или вируса. Последовательность нуклеиновой кислоты-мишени может представлять собой участок гена или хромосомы. Последовательность нуклеиновой кислоты-мишени может быть специфичной для генотипа или организма (такого как патоген), подлежащего выявлению с помощью амплификации ДНК. Последовательность нуклеиновой кислоты-мишени может быть уникальной для генома конкретного вида. Таким образом, последовательность нуклеиновой кислоты-мишени для выявления конкретного вида, как правило, отличается от любой гомологичной последовательности нуклеиновой кислоты у родственного вида. Как правило, последовательность нуклеиновой кислоты-мишени содержит несколько несоответствий с гомологичной последовательностью нуклеиновой кислоты у родственного вида. Последовательность нуклеиновой кислоты-мишени может представлять собой последовательность, специфичную для конкретного штамма бактерий или конкретного серотипа, изолята или клады вируса.
Последовательность нуклеиновой кислоты-мишени, подлежащая выявлению, может иметь любой размер и последовательность. Длина последовательности нуклеиновой кислоты-мишени или ампликона является достаточной для обеспечения гибридизации с прямым и обратным праймерами и связывания соответствующим образом с олигонуклеотидом (олигонуклеотидами) для внедрения в цепь в 3'-5' и 5'-3' области указанной последовательность-мишени. Длина ампликона, как правило, составляет по меньшей мере 60 нуклеотидов, более предпочтительно, по меньшей мере 65 или по меньшей мере 70 нуклеотидов при измерении от 5'-сайта связывания прямого праймера до 5'-сайта связывания обратного праймера. Длина ампликона может составлять от примерно 60 до примерно 80 нуклеотидов. Согласно некоторыми вариантам реализации изобретения, длина ампликона может составлять больше 80, например, больше 100 нуклеотидов, например, больше 150, 200, 300, 400, 500, 1000 или более нуклеотидов. Длина ампликона может составлять от примерно 70 до примерно 1000 нуклеотидов, например, от примерно 70 до примерно 800, от примерно 70 до примерно 600, от примерно 70 до примерно 500 нуклеотидов, от примерно 70 до примерно 400, от примерно 100 до примерно 400 или от примерно 100 до примерно 200 нуклеотидов.
Примеры подходящих последовательностей нуклеиновых кислот-мишеней для способов согласно изобретению включают последовательности SEQ ID NO 11, 12, 16, 17, 18, 19, 20 и 26.
Последовательность нуклеиновой кислоты-мишень имеет 3'-5' (верхнюю, 5') и 5'-3' (нижнюю, 3') область, каждая из которых содержит участки связывания олигонуклеотида для внедрения в цепь и праймера. Участки 3'-5' связывания олигонуклеотида для внедрения в цепь и праймера могут перекрываться в последовательности или быть неперекрывающимися. Подобным образом, участки 5'-3' связывания олигонуклеотида для внедрения в цепь и праймера могут перекрываться в последовательности или быть неперекрывающимися. Последовательность нуклеиновой кислоты-мишени также может содержать участки связывания одного или более олигонуклеотидных зондов. Участки связывания зонда могут перекрываться в последовательности с 3'-5' или 5'-3' участками связывания олигонуклеотида для внедрения в цепь и/или праймера или быть неперекрывающимися с участком связывания для любого олигонуклеотида для внедрения в цепь или праймера. Участок связывания зонда может предпочтительно располагаться между 3'-5' и 5'-3' участками связывания олигонуклеотидов для внедрения в цепь в последовательности нуклеиновой кислоты-мишени. Выбор участков связывания и создание соответствующих последовательностей олигонуклеотидов для внедрения в цепь, праймеров и зондов обсуждается более подробно ниже.
Длина участков связывания олигонуклеотидов для внедрения в цепь, праймеров и зондов определяется длиной включенных в них последовательностей, комплементарных мишени, как описано ниже более подробно. Как описано ниже, олигонуклеотид для внедрения в цепь, как правило, содержит по меньшей мере 25 нуклеотидов в комплементарной мишени последовательности, и праймер содержит по меньшей мере 10 нуклеотидов. Таким образом, каждый участок связывания олигонуклеотида для внедрения в цепь в последовательности-мишени может составлять по меньшей мере 25 нуклеотидов в длину, и каждый участок связывания праймера может составлять по меньшей мере 10 нуклеотидов в длину. Последовательность-мишень может дополнительно содержать участок связывания зонда, составляющий, как правило, по меньшей мере 10 нуклеотидов в длину.
Участки связывания олигонуклеотидов для внедрения в цепь в 3'-5' и 5'-3' направлении могут присутствовать в одной цепи последовательности нуклеиновой кислоты-мишени или могут быть расположены в противоположных цепях дуплекса, включающего последовательность нуклеиновой кислоты-мишени. Олигонуклеотид (олигонуклеотиды) для внедрения в цепь может, таким образом, связываться с одной и той же цепью последовательности нуклеиновой кислоты-мишени в параллельной конфигурации, сохраняя направление от 5' к 3'-концу. Альтернативно, олигонуклеотид (олигонуклеотиды) для внедрения в цепь может связываться с противоположными цепями последовательности нуклеиновой кислоты-мишени в антипараллельной конфигурации, поворачивая направление 5' к 3' в дуплексе-мишени в противоположную сторону. При антипараллельной конфигурации 3'-концы олигонуклеотидов для внедрения в цепь могут быть направлены друг к другу или друг от друга. Таким образом, 3'-концы каждого олигонуклеотида для внедрения в цепь могут быть направлены к центру ампликона (антипараллельная конфигурация) или в сторону соответствующего конца ампликона (обратная антипараллельная конфигурация). Связывание 3'-конца или 5'-конца олигонуклеотида для внедрения в цепь может происходить вблизи участка связывания соответствующего праймера. Указанные выше конфигурации связывания показаны на Фигуре 1.
Применение конкретных конфигураций связывания может обеспечивать разные эффекты на параметры амплификации. Например, когда при связывании олигонуклеотида для внедрения в цепь его 5'-конец располагается вблизи участка связывания соответствующего праймера, специфичность и кинетический профиль связывания праймера может отличаться от этих показателей при связывании праймера вблизи 3'-конца олигонуклеотида для внедрения в цепь. 3'-конец олигонуклеотида для внедрения в цепь, как правило, содержит ряд модифицированных нуклеотидов (таких как 2'-O-метилРНК-нуклеотиды), которые могут влиять на параметры связывания связывающегося вблизи от них праймера. Предполагается, что при параллельной и обратной антипараллельной конфигурации специфичность амплификации также может быть усилена в связи с необходимостью миграции ветви с 3'-конца (содержащего, как правило, модифицированные нуклеотиды) олигонуклеотидов для внедрения в цепь для возможности связывания праймера. Таким образом, способы согласно изобретению обеспечивают возможность изменять скорость и специфичность амплификации путем использования разных конфигураций связывания олигонуклеотида (олигонуклеотидов) для внедрения в цепь.
Участки связывания олигонуклеотидов для внедрения в цепь в 3'-5' и 5'-3' направлении в последовательности нуклеиновой кислоты-мишени могут связываться с одним типом олигонуклеотида для внедрения в цепь. Таким образом, может быть предложен один тип олигонуклеотида для внедрения в цепь для инициации внедрения в цепь в двух положениях последовательности нуклеиновой кислоты-мишени, как дополнительно обсуждается ниже. Согласно указанному варианту реализации изобретения, каждый их 3'-5' и 5'-3' участков связывания содержит последовательность, комплементарную по меньшей мере части олигонуклеотида для внедрения в цепь. Участки связывания в 3'-5' и 5'-3' направлении, как правило, гомологичны или идентичны друг другу. Участки связывания в 3'-5' и 5'-3' направлении могут быть по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 95% гомологичными или идентичными друг другу или полностью являться идентичными. Между 3'-5' и 5'-3' участками связывания может быть 1, 2, 3, 4, 5, 6, 7 или 8, например, от 1 до 5 или от 1 до 3, несоответствий. Кроме того, может быть предложен один вид праймера для инициации амплификации в двух точках последовательности нуклеиновой кислоты-мишени, как дополнительно обсуждается ниже. Последовательность нуклеиновой кислоты-мишени в этом случае имеет участки связывания в 3'-5' и 5'-3' направлении, каждый из которых содержит последовательность, комплементарную по меньшей мере части праймера, и которые могут быть гомологичными или идентичными друг другу, как описано выше.
Способ согласно изобретению может обеспечивать выявление более чем одной последовательность нуклеиновой кислоты-мишени с помощью множества комбинаций олигонуклеотидов для внедрения в цепь, праймеров (и необязательно зондов), все из которых специфичны к разным последовательностям нуклеиновой кислоты-мишени. Как правило, олигонуклеотид для внедрения в цепь/пары праймеров и/или зонды, связывающиеся с различными последовательностями нуклеиновой кислоты-мишеней, метят различными парами флуорофор/гаситель, обеспечивая, таким образом, мультиплексную систему. Можно выявлять по меньшей мере две, три, четыре, пять, десять или более различных последовательностей-мишеней. Можно выявлять более чем одну последовательность нуклеиновой кислоты-мишень из одного и того же организма. Альтернативно, можно выявлять последовательности нуклеиновых кислот-мишени, специфичные по меньшей мере для двух, трех, четырех, пяти, десяти или более различных генотипов, организмов или патогенов.
Прямой и обратный праймеры
Подходящие прямой и обратный праймеры выбирают на основе интересующей последовательности нуклеиновой кислоты-мишени с учетом сайта связывания соответствующего олигонуклеотида для внедрения в цепь, делающего 3'-5' или 5'-3' участок связывания последовательности нуклеиновой кислоты-мишени одноцепочечными для обеспечения возможности связывания соответствующего праймера.
Прямой и обратный праймеры содержат последовательность, которая частично или полностью комплементарна последовательности-мишени, и необязательно 5'- и/или 3'-фланкирующую некомплементарную последовательность. Альтернативно, прямой и обратный праймеры могут полностью состоять из последовательности, частично или полностью комплементарной последовательности-мишени. Длина последовательности праймера, комплементарной последовательности-мишени, является достаточной для обеспечения специфичной гибридизации с последовательностью нуклеиновой кислоты-мишенью. Длина комплементарной последовательности, как правило, составляет по меньшей мере 10 нуклеотидов, более предпочтительно, по меньшей мере 15, по меньшей мере 16 или по меньшей мере 17 нуклеотидов. Длина комплементарной последовательности может составлять 10-25, 15-25, 10-30 или 15-30 нуклеотидов.
Следует понимать, что указанные выше длины последовательности относятся к фрагментам праймеров, которые могут быть частично или полностью комплементарны последовательности нуклеиновой кислоты-мишени. Между последовательностями праймеров и последовательностью-мишенью возможны несоответствия в конкретных положениях при сохранении возможности специфичной амплификации и выявления последовательности-мишени, в частности, в случае комбинированного применения прямого и обратного праймеров и связывания олигонуклеотида (олигонуклеотидов) для внедрения в цепь с 3'-5' и 5'-3' участками последовательности нуклеиновой кислоты-мишени для обеспечения амплификации. Между комплементарным участком праймера и соответствующей областью последовательности-мишени может быть 1, 2, 3, 4 или 5 несоответствий.
Как правило, длина прямого и обратного праймера в общем составляет менее 30, более предпочтительно, менее 25 нуклеотидов, например, от 15 до 25 или от 15 до 23 нуклеотидов. Особо предпочтительным является использование праймеров, длина которых составляет менее 30 нуклеотидов, в случаях, когда для внедрения в цепь используется рекомбиназа. Такие праймеры не способны действовать в качестве субстратов для рекомбиназ. Согласно некоторыми вариантам реализации изобретения, можно использовать праймеры длиной менее 15 нуклеотидов, например, праймеры длиной от примерно 8 до примерно 14, от примерно 10 до примерно 14 или от примерно 12 до примерно 14 нуклеотидов. Применение таких коротких праймеров является предпочтительным в комбинации с зондом, участок связывания которого в последовательности нуклеиновой кислоты-мишени не перекрывается с участком связывания для праймера или олигонуклеотида для внедрения в цепь. Вероятность выявления продуктов неспецифической амплификации, полученных в результате использования коротких праймеров, можно снизить или устранить путем использования зонда с неперекрывающимся сайтом связывания.
Прямой праймер (или праймер в направлении 3'-5') связывается с 3'-участком одной цепи дуплекса последовательности нуклеиновой кислоты-мишени в положении, близком или перекрывающимся с сайтом связывания олигонуклеотида для внедрения в цепь. Обратный (или 5'-3') праймер связывается с 3'-участком противоположной цепи дуплекса последовательности нуклеиновой кислоты-мишени (по отношению к прямому праймеру) в положении, близком или перекрывающимся с сайтом связывания олигонуклеотида для внедрения в цепь. 5'-сайты связывания прямого и обратного праймеров, как правило, находятся на расстоянии по меньшей мере 60 нуклеотидов, более предпочтительно, по меньшей мере 65 или по меньшей мере 70 нуклеотидов в длину друг от друга в дуплексе последовательности-мишени.
В зависимости от конфигурации связывания олигонуклеотидов для внедрения в цепь, как показано на Фигуре 1, прямой праймер может иметь область перекрывания последовательности или область комплементарности с последовательностью соответствующего олигонуклеотида для внедрения в цепь. Область перекрывания или комплементарности последовательности может составлять 1-8 нуклеотидов в длину и может составлять по меньшей мере 5 или по меньшей мере 6 нуклеотидов в длину. Обратный праймер, подобным образом, может иметь область перекрывания или комплементарности последовательности с последовательностью соответствующего олигонуклеотида для внедрения в цепь, составляющую 1-8 нуклеотидов, например, по меньшей мере 5 или по меньшей мере 6 нуклеотидов в длину.
Альтернативно, последовательность прямого праймера может не перекрываться или не являться комплементарной последовательности соответствующего олигонуклеотида для внедрения в цепь, и/или последовательность обратного праймера может не перекрываться или не являться комплементарной последовательности соответствующего олигонуклеотида для внедрения в цепь, при этом праймер связывается в соответствующем положении вблизи сайта связывания олигонуклеотида для внедрения в цепь в последовательности-мишени.
Применение одного или более праймеров, которые имеют участки связывания в последовательности-мишени, не перекрывающиеся с участками связывания олигонуклеотидов для внедрения в цепь, может обеспечить различные преимущества. Согласно вариантам реализации изобретения, когда в способах согласно изобретению используются олигонуклеотидные зонды для выявления амплификации ДНК, последовательности олигонуклеотидов для внедрения в цепь и зондов и/или прямого и/или обратного праймера и зонда могут не перекрываться или не быть комплементарными. Последовательности участков связывания прямого праймера, обратного праймера, каждого олигонуклеотида для внедрения в цепь и любого зонда в последовательности нуклеиновой кислоты-мишени могут не перекрываться. Кроме того, последовательности любых праймеров, олигонуклеотидов для внедрения в цепь или зондов могут не быть комплементарными. Создание последовательностей различных видов олигонуклеотидов, способных связываться с последовательностью нуклеиновой кислоты-мишенью в независимых неперекрывающихся участках может обеспечить снижение конкуренции между разными видами олигонуклеотидов за связывание с последовательностью нуклеиновой кислоты-мишени, и также снизить формирование и/или предупредить выявление нежелательных продуктов амплификации.
Более конкретно, праймеры, имеющие длину между 16 и 23 основаниями, как правило, используют в способах амплификации на основе внедрения в цепь с использованием одного олигонуклеотида для внедрения в цепь (способы амплификации с одним олигонуклеотидом для внедрения в цепь, SIBA). Обычно примерно 8 оснований на 3'-конце последовательностей праймеров перекрываются или комплементарны последовательности олигонуклеотида для внедрения в цепь (последовательность прямого праймера перекрывается с последовательностью олигонуклеотида для внедрения в цепь, тогда как последовательность обратного праймера комплементарна последовательности олигонуклеотида для внедрения в цепь). Такая конфигурация обеспечивает эффективную амплификацию ДНК-мишени и минимизирует риск неспецифичной амплификации. Также возможно использовать короткие праймеры длиной ≤14 оснований, не перекрывающиеся с олигонуклеотидом для внедрения в цепь. Короткие праймеры, которые не содержат последовательностей, перекрывающихся с последовательностью олигонуклеотида для внедрения в цепь, способны обеспечивать более эффективную амплификацию ДНК-мишени по сравнению с длинными перекрывающимися праймерами. Это связано с тем, что 3'-конец более длинного перекрывающегося праймера конкурирует с олигонуклеотидом для внедрения в цепь за сайт связывания матрицы-мишени. Например, прямой праймер должен сначала переместиться в результате миграции ветви в дуплексе до перемещения олигонуклеотида для внедрения в цепь.
Однако короткие праймеры (≤14 оснований) могут приводить к получению продуктов неспецифической амплификации. Во избежание этих проблем в реакции SIBA, как правило, используются более длинные праймеры (16-23 оснований), 3'-концы которых перекрываются или комплементарны последовательности олигонуклеотида для внедрения в цепь. При такой конфигурации область, периферическая по отношению к олигонуклеотиду для внедрения в цепь, все еще составляет около 14 оснований в длину, в связи с чем остается только короткая периферическая область, которая диссоциирует при амплификации ДНК-мишени.
В способах согласно изобретению, включающих внедрение в цепь в двух сайтах последовательности-мишени (3'-5' и 5'-3'), можно использовать более короткие праймеры по сравнению с SIBA. Более того, можно использовать более эффективно неперекрывающиеся праймеры. Это связано с возможностью встраивания сайта связывания зонда на ДНК-мишени, независимого от сайтов связывания олигонуклеотидов для внедрения в цепь и праймеров. Более того, возможность использовать в способах согласно изобретению различные конфигурации праймера и олигонуклеотида для внедрения в цепь, такие как обратная антипараллельная конфигурация, позволяет минимизировать или устранить риск вызванной использованием короткого праймера неспецифической амплификации.
Когда праймер связывается вблизи соответствующего олигонуклеотида для внедрения в цепь (без перекрывания или комплементарности последовательности), расстояние между ближайшей границей участка связывания олигонуклеотида для внедрения в цепь и участка связывания соответствующего праймера, как правило, составляет 15 нуклеотидов или менее, предпочтительно 10 нуклеотидов или менее, например, от примерно 1 до примерно 15 нуклеотидов, от примерно 5 до примерно 15 нуклеотидов, от примерно 5 до примерно 10 нуклеотидов или от примерно 3 до примерно 8 нуклеотидов. Это обеспечивает гарантию того, что праймер способен гибридизоваться с одноцепочечной областью, образованной в результате связывания олигонуклеотида для внедрения в цепь.
Предпочтительно, каждый праймер создается таким образом, чтобы обеспечить возможность специфичного выявления конкретной последовательности нуклеиновой кислоты-мишени, например, конкретного генотипа, или последовательности нуклеиновой кислоты, присутствующей в конкретной мишени, такой как конкретный организм или конкретный патоген. Таким образом, каждый праймер, как правило, специфично или селективно гибридизуется с комплементарной последовательностью, обнаруживаемой только в мишени. Однако каждый праймер также может гибридизоваться с другими последовательностями, такими как последовательности, обнаруживаемые в других видах, при условии что при использовании в комбинации со вторым праймером, олигонуклеотидом (олигонуклеотидами) для внедрения в цепь и необязательно олигонуклеотидным зондом достигается специфичное выявление последовательности нуклеиновой кислоты-мишени.
Любой прямой или обратный праймер, используемый согласно изобретению, может содержать один или более модифицированных нуклеотидов и/или выявляемую метку, например, флуоресцентный краситель. Согласно некоторыми вариантам реализации изобретения, прямой или обратный праймер может образовывать пару резонансного переноса энергии флуоресценции (fluorescence resonance energy transfer, FRET) с соответствующим олигонуклеотидом для внедрения в цепь, и таким образом, содержит флуорофор или гаситель, как обсуждается ниже.
Следует понимать, что способы согласно изобретению могут включать применение более чем одной пары прямого и обратного праймеров, как правило, в тех случаях, когда более чем одна последовательность-мишень подлежит одновременному выявлению в системе мультиплексного анализа.
Олигонуклеотид (олигонуклеотиды) для внедрения в цепь
Один или более подходящих олигонуклеотидов для внедрения в цепь выбирают на основе интересующей последовательности-мишени нуклеиновой кислоты с учетом сайтов связывания прямого и обратного праймеров и требуемой способности олигонуклеотида (олигонуклеотидов) для внедрения в цепь делать последовательность нуклеиновой кислоты-мишени одноцепочечной в соответствующих участках для обеспечения возможности связывания прямого и обратного праймера. Когда последовательность нуклеиновой кислоты-мишени содержит гомологичные или идентичные участки связывания олигонуклеотидов для внедрения в цепь в 3'-5' и 5'-3' направлении, может быть предложен один вид олигонуклеотида для внедрения в цепь для осуществления амплификации. Альтернативно, могут быть предложены два разных вида олигонуклеотидов для внедрения в цепь (первый и второй), которые связываются с разными последовательностями в 3'-5' и 5'-3' области последовательности нуклеиновой кислоты-мишени. Следующее далее описание признаков олигонуклеотида для внедрения в цепь применимо как к первому, так и ко второму вызывающему внедрение в цепь олигонуклеотиду (при их использовании).
Каждый олигонуклеотид для внедрения в цепь содержит последовательность, которая комплементарна мишени, и необязательно дополнительную фланкирующую некомплементарную последовательность (последовательности). Длина последовательности, которая комплементарна мишени, может быть определена специалистом в данной области техники опытным путем и является достаточной для обеспечения эффективного внедрения в цепь последовательности нуклеиновой кислоты-мишени, необязательно при изотермических условиях. Комплементарная последовательность может содержать РНК или ДНК-нуклеотиды комплементарного спаривания и модифицированные нуклеотиды. Как правило, длина комплементарной последовательности составляет по меньшей мере 25 или по меньшей мере 27 нуклеотидов, как правило, по меньшей мере 30 нуклеотидов, например, по меньшей мере 32, по меньшей мере 33 или по меньшей мере 35 нуклеотидов, более предпочтительно, по меньшей мере 36, 37, 38, 39 или 40 нуклеотидов в длину или более. Длина комплементарной последовательности может составлять 30-50, 32-50, 35-50, 40-50, 35-48, 35-46, 38-45 или 40-45 нуклеотидов в длину.
Следует понимать, что указанные выше длины последовательностей относятся к части олигонуклеотида для внедрения в цепь, которая может быть частично или полностью комплементарной последовательности нуклеиновой кислоты-мишени. Между последовательностью олигонуклеотида для внедрения в цепь и последовательностью-мишенью могут быть несоответствия в конкретных положениях при сохранении возможности специфической амплификации и выявления последовательности-мишени, в частности, в случае комбинированного применения прямого и обратного праймеров и олигонуклеотида для внедрения в цепь для достижения амплификации. Может быть 1, 2, 3, 4, 5, 6, 7, или 8, например, от 1 до 5 или от 1 до 3 несоответствий между комплементарной областью олигонуклеотида для внедрения в цепь и соответствующей областью последовательности-мишени в зависимости от общей длины комплементарной последовательности.
Комплементарная последовательность олигонуклеотида для внедрения в цепь гибридизуется с частью последовательности-мишени, которая может перекрываться или может не перекрываться с частью последовательность-мишени, образующей участок связывания праймера. Олигонуклеотид для внедрения в цепь может содержать область перекрывания или комплементарности последовательности соответствующего прямого или обратного праймера, составляющую 1-8 нуклеотидов, например, по меньшей мере 5 или по меньшей мере 6 нуклеотидов в длину. Альтернативно, последовательность олигонуклеотида для внедрения в цепь может не содержать область перекрывания с последовательностью прямого или обратного праймера. Согласно указанному варианту реализации изобретения, как обсуждается выше, олигонуклеотид для внедрения в цепь связывается в положении вблизи участка связывания прямого или обратного праймера таким образом, что может делать указанный участок связывания праймера одноцепочечным.
Ближайшие границы 3'-5' и 5'-3' участка связывания олигонуклеотидов для внедрения в цепь в последовательности нуклеиновой кислоты-мишени могут быть расположены на расстоянии по меньшей мере 15, например, по меньшей мере 20 или по меньшей мере 25 нуклеотидов в последовательности нуклеиновой кислоты-мишени, однако, согласно некоторыми вариантам реализации изобретения, также могут использоваться более короткие расстояния между участками связывания.
Связывание 5'-области комплементарной последовательности олигонуклеотида для внедрения в цепь, как правило, происходит в пределах 25 или менее нуклеотидов, более предпочтительно, 20 или менее нуклеотидов от соответствующей границы дуплекса нуклеотидной последовательности-мишени, подлежащей денатурации (ампликон).
Олигонуклеотид для внедрения в цепь необязательно также содержит не комплементарную мишени область (области) последовательности, которая фланкирует комплементарную область последовательности. Олигонуклеотид для внедрения в цепь может содержать некомплементарную 5'-область, которая может состоять из любой нуклеотидной последовательности. 5'-некомплементарная область, как правило, составляет по меньшей мере 3 нуклеотида в длину, чаще по меньшей мере 6, по меньшей мере 8, предпочтительно по меньшей мере 10, по меньшей мере 12 или по меньшей мере 14 нуклеотидов в длину. 5'-некомплементарная область может способствовать связыванию рекомбиназы, так как рекомбиназы связываются кооперативно. Олигонуклеотид для внедрения в цепь может содержать 3'-некомплементарную область, составляющую, как правило, 1-3 нуклеотида в длину, которая содержит нуклеотиды, блокирующие полимеразное удлинение, например, 3'-инвертированный дТ.
Длина олигонуклеотида для внедрения в цепь, как правило, составляет по меньшей мере 30 нуклеотидов, если в способе амплификации вместе с указанным олигонуклеотидом для внедрения в цепь используется рекомбиназа для внедрения в цепь. Длина олигонуклеотида для внедрения в цепь предпочтительно составляет по меньшей мере 35, по меньшей мере 40 или по меньшей мере 45 нуклеотидов, более предпочтительно, по меньшей мере 50 нуклеотидов и может составлять по меньшей мере 55 нуклеотидов или более. Длина олигонуклеотида для внедрения в цепь может составлять 40-70, 45-70, 45-70, 50-70, 55-70, 45-65, 50-65, 50-60 или 55-65 нуклеотидов.
Как правило, олигонуклеотид для внедрения в цепь имеет неудлиняемый 3'-конец и, таким образом, не может служить субстратом для ДНК-полимеразы, и амплификация последовательности-мишени в таком случае происходит только при дальнейшем связывании специфичных прямого и обратного праймеров. Это позволяет избежать образования продуктов неспецифической амплификации. Олигонуклеотид для внедрения в цепь может содержать один, два, три, четыре, пять, шесть, семь, восемь или более модифицированных нуклеотидов в 3'-области, например, в пределах 10-15 или 10-20 нуклеотидов от 3'-конца. Олигонуклеотид для внедрения в цепь может содержать 3'-модификацию 3'-концевого нуклеотида, который может представлять собой дидезоксинуклеотид, или содержать 3'-аминоаллильную группу, 3'-углеродный спейсер, 3'-фосфат, 3'-биотин, 3'-сиалил или 3'-тиол. 3'-нуклеотид может представлять собой нуклеотид, встроенный с помощью 3'-3'-инвертированной связи. Альтернативно или дополнительно, 3'-область олигонуклеотида для внедрения в цепь может содержать нуклеотиды со слабой способностью служить субстратом для ДНК-полимераз, такие как нуклеотиды ПНК (пептидной нуклеиновой кислоты), ЗНК (закрытой нуклеиновой кислоты), 2'-5'-сшитой ДНК, 2'-фторРНК или 2'-O-метилРНК, или их комбинации.
Если олигонуклеотид для внедрения в цепь представляет собой ПНК-олигомер, содержащий, состоящий из или по существу состоящий из ПНК-нуклеотидов, то указанный олигонуклеотид может дестабилизировать и вызывать внедрение в цепь дуплекса ДНК в отсутствие фермента рекомбиназы. Таким образом, если используется ПНК-олигонуклеотид, способы согласно изобретению можно осуществлять в отсутствие фермента рекомбиназы. ПНК-олигонуклеотид может содержать ПНК-нуклеотиды и другие нуклеотиды, такие как ДНК-нуклеотиды, при условии что указанный олигонуклеотид содержит достаточное количество ПНК-нуклеотидов для способности опосредовать внедрение в цепь дуплекса. Специалист в данной области техники может опытным путем определить, какое количество ПНК необходимо встроить в олигонуклеотид, путем исследования ее способности влиять на внедрение в цепь и обеспечивать возможность амплификации ДНК.
Олигонуклеотид для внедрения в цепь может содержать выявляемую метку, например, флуоресцентный краситель. Согласно некоторыми вариантам реализации изобретения, олигонуклеотид для внедрения в цепь может образовывать пару FRET с прямым или обратным праймером и, таким образом, содержать флуорофор или гаситель, как обсуждается ниже.
Способы согласно изобретению включают внедрение в цепь по меньшей мере в двух сайтах последовательности нуклеиновой кислоты-мишени, опосредованную первым и вторым олигонуклеотидами для внедрения в цепь или одним видом олигонуклеотида для внедрения в цепь, где последовательность нуклеиновой кислоты-мишени содержит два сайта связывания для одного олигонуклеотида для внедрения в цепь. Следует понимать, что способы согласно изобретению могут также включать внедрение в цепь с помощью дополнительных олигонуклеотидов для внедрения в цепь в дополнительных сайтах последовательности нуклеиновой кислоты-мишени, например, в 3 или более, 4 или более, 5 или более, 8 или более, или 10 или более сайтах. Кроме того, в мультиплексной системе способы согласно изобретению могут включать применение дополнительных олигонуклеотидов для внедрения в цепь, направленных на участки связывания дополнительных последовательностей-мишеней в 3'-5' и 5'-3' направлении.
Амплификация последовательности нуклеиновой кислоты-мишени
Способ амплификации ДНК включает амплификацию на основе внедрения в цепь. Амплификация на основе внедрения в цепь включает внедрение в цепь по меньшей мере в двух сайтах последовательностей нуклеиновой кислоты-мишеней. Внедрение в цепь происходит как в 3'-5', так и в 5'-3' участке последовательности нуклеиновой кислоты-мишени.
Последовательность нуклеиновой кислоты-мишени инкубируют с прямым праймером, обратным праймером и одним или более (например, первым и вторым) олигонуклеотидами для внедрения в цепь, способными делать как 3'-5', так и 5'-3' участки связывания для соответствующих праймеров одноцепочечными, в условиях, обеспечивающих амплификацию указанной последовательности нуклеиновой кислоты-мишени. Согласно некоторыми вариантам реализации изобретения, один вид праймера может служить в качестве как прямого, так и обратного праймера.
Такие условия, как правило, включают присутствие фермента ДНК-полимеразы. Подходящие условия включают любые условия, известные в данной области техники, используемые для обеспечения активности полимеразных ферментов. Условия, как правило, включают присутствие дезоксинуклеотидтрифосфатов (дНТФ), выбранных из дАТФ, дТТФ, дЦТФ, дГТФ, дУТФ и их аналогов, подходящих буферных агентов/изменяющих рН агентов и других факторов, которые требуются для активности или стабильности ферментов. Как правило, присутсвуют все четыре из дезоксинуклеотидтрифосфатов дАТФ, дТТФ, дЦТФ, дГТФ. Условия могут включать присутствие детергентов и стабилизирующих агентов. Используемые температурные условия, как правило, являются изотермическими, т.е. температура остается постоянной на протяжении всего процесса амплификации. Используемая температура, как правило, зависит от природы полимеразного фермента и других ферментативных компонентов и также отражает температуру гибридизации, требуемую для праймеров и олигонуклеотидов для внедрения в цепь.
Используемая полимераза, как правило, обладает активностью замещения цепи. Термин «замещение цепи» используется в настоящей заявке для описания способности ДНК-полимеразы необязательно совместно со вспомогательными белками замещать комплементарные цепи на области двуцепочечной ДНК, встречающиеся во время синтеза ДНК. Подходящие ДНК-полимеразы включают полимеразу polI из Е. coli, В. subtilis или В. stearothermophilus и их функциональные фрагменты и варианты, а также ДНК-полимеразы Т4 и Т7 и их функциональные фрагменты и варианты. Предпочтительно полимераза представляет собой ДНК-полимеразу Bsu или ее функциональный фрагмент или вариант.
Условия амплификации предпочтительно включают присутствие рекомбиназы. В способе согласно изобретению можно использовать любую систему рекомбиназ. Система рекомбиназ может быть прокариотической или эукариотической и может быть бактериальной, дрожжевой, фаговой или системой рекомбиназ млекопитающих. Рекомбиназа может обеспечивать полимеризацию одноцепочечного олигонуклеотида в 5'-3' или 3'-5' направлении. Рекомбиназа может происходить из фага семейства Myoviridae, такого как фаг Т4, Т2, Т6, Rb69, Aeh1, KVP40, фага акинетобактерий 133, фага аэромонад 65, цианофага P-SSM2, цианофага PSSM4, цианофага S-PM2, Rbl4, Rb32, фага аэромонад 25, вибриофага nt-1, phi-1, Rbl6, Rb43, фага 31, фага 44RR2,8t, Rb49, фага Rb3 или фага LZ2. Согласно предпочтительному варианту реализации изобретения, используют рекомбиназу фага Т4 UvsX (номер доступа: Р04529) или ее функциональный вариант или фрагмент. Также могут использоваться эукариотические системы Rad, система recA-Reco Е. coli или другие прокариотические системы. Рекомбиназа может представлять собой RecA Е. coli.
Условия могут дополнительно включать присутствие вспомогательных белков системы рекомбинации, такие как белок, связывающийся с одноцепочечной ДНК (например, gp32 Т4, номер доступа Р03695) и белок-грузчик рекомбиназы (например, UvsY, номер доступа NP_049799.2). Согласно предпочтительному варианту реализации изобретения, условия включают присутствие белков gp32 Т4, UvsX и UvsY. Рекомбиназа (такая как UvsX), а также белок-грузчик рекомбиназы (такой как UvsY) и белок, связывающийся с одноцепочечной ДНК (такой как gp32) в случае их использования могут представлять собой нативные, гибридные или мутантные белки из одного и разных источников, представляющих собой фаги семейства Myoviridae. Нативный белок может представлять собой белок дикого типа или природный вариант белка.
Условия могут дополнительно включать другие факторы, используемые для усиления эффективности рекомбиназы, такие как соединения, используемые для контроля ДНК-взаимодействий, например, пролин, ДМСО, БСА, ПЭГ или другие агенты, изменяющие свойства вещества в растворе, которые, как известно повышают нагрузку рекомбиназы на ДНК (Lavery P. et al. J. Biol. Chem. 1992, 267, (13), 9307-9314).
Условия могут также включать присутствие системы восстановления АТФ. Различные системы восстановления АТФ известны специалисту в данной области техники и включают гликолитические ферменты. Подходящие компоненты системы восстановления АТФ могут включать один или более из фосфокреатина, креатинкиназы, миокиназы, пирофосфатазы, сахарозы и фосфорилазы сахарозы. Условия могут включать присутствие АТФ.
Дополнительные компоненты, такие как ионы магния, ДТТ или другие восстанавливающие агенты и соли также могут быть включены.
Также компоненты могут включать один или более ферментов для рестрикции (таких как одна или более эндонуклеаз рестрикции) для расщепления нуклеиновой кислоты, содержащей последовательность нуклеиновой кислоты-мишени, до или одновременно с приведением последовательности нуклеиновой кислоты-мишени в контакт с другими реагентами для амплификации. Скорость амплификации последовательности нуклеиновой кислоты-мишени, содержащейся в ДНК-плазмиде, может быть повышена путем расщепления плазмиды с помощью фермента для рестрикции для получения, таким образом, линейной исходной матрицы. Таким образом, способы согласно изобретению могут включать приведение нуклеиновой кислоты, содержащей нуклеиновую кислоту-мишень, подлежащую амплификации, в контакт с ферментом для рестрикции. Любые подходящие ферменты для рестрикции, имеющие подходящий сайт распознавания в нуклеиновой кислоте, содержащей последовательность нуклеиновой кислоты-мишень, могут использоваться для расщепления. Сайт распознавания, как правило, расположен в области нуклеиновой кислоты, отличной от последовательности нуклеиновой кислоты-мишени.
Различные компоненты, описанные выше, могут быть предложены в различающихся концентрациях для обеспечения амплификации ДНК. Специалист в данной области техники может выбрать подходящие рабочие концентрации различных компонентов на практике.
Выявление амплифицированной ДНК
За присутствием амплифицированной ДНК, которая является результатом приведения последовательности нуклеиновой кислоты-мишени в контакт с праймерами и олигонуклеотидом для внедрения в цепь (олигонуклеотидами) в условиях, обеспечивающих амплификацию ДНК, можно следить с помощью любого подходящего способа.
Один или оба праймера или один или более олигонуклеотидов для внедрения в цепь (олигонуклеотид) (таких как первый и/или второй олигонуклеотиды для внедрения в цепь (олигонуклеотид)) могут содержать метку или другой выявляемый фрагмент. Можно использовать любую метку или выявляемый фрагмент. Примеры подходящих меток включают флуоресцентные фрагменты и пары FRET флуорофора и акцепторного фрагмента. Например, прямой праймер может образовывать пару FRET с олигонуклеотидом для внедрения в цепь, имеющим участок связывания в 3'-5' направлении в последовательности нуклеиновой кислоты-мишени, и/или обратный праймер может образовывать пару FRET с олигонуклеотидом для внедрения в цепь, имеющим 5'-3' участок связывания в последовательности нуклеиновой кислоты-мишени. Праймер (праймеры) может быть мечен флуорофором или гасителем, при этом олигонуклеотид для внедрения в цепь (олигонуклеотиды) метят соответствующим членом пары FRET, гасителем или флуорофором. Подходящие метки и сайты присоединения описаны ниже. Применение таких пар FRET может обеспечивать способы, позволяющие выявлять внедрение в цепь и амплификацию последовательности нуклеиновой кислоты-мишени. Также могут использоваться другие системы гашения, выявляющие изменения во взаимодействии двух выявляемых фрагментов, включая контактное гашение.
Более предпочтительно или дополнительно, можно использовать один или более зондов, выявляющих амплифицированную ДНК, путем повторного включения метки или другого выявляемого фрагмента. Предпочтительно, сигнал зонда отслеживают в режиме реального времени вместе с амплификацией последовательности нуклеиновой кислоты-мишени. Зонд может связываться в любом подходящем положении в последовательности нуклеиновой кислоты-мишени. Особо предпочтительно зонд может связываться с областью последовательности нуклеиновой кислоты-мишени, которая не перекрывается с участком связывания праймера и/или олигонуклеотида для внедрения в цепь. Таким образом, особо предпочтительно сайт связывания зонда может располагаться в пределах последовательности нуклеиновой кислоты-мишени независимо от сайта (сайтов) связывания одного или более других видов олигонуклеотида. Выбор неперекрывающихся участков связывания зонда может снизить конкуренцию за связывание зонда во время амплификации. Применение способов связывания зонда в независимом положении в последовательности нуклеиновой кислоты-мишени также может снизить или предотвратить выявление продуктов неспецифической амплификации, таких как праймер-димер, обеспечивая более точное выявление амплификации последовательности нуклеиновой кислоты-мишени.
Зонды, выявляющие различные амплифицированные последовательности-мишени, могут испускать флуоресцентные сигналы с разной длиной волны для обеспечения мультиплексной системы выявления. Два или более, например, три, четыре, пять, шесть, восемь, десять или более различных зондов, можно использовать для мульплексного выявления нескольких различных последовательностей-мишеней в одной реакции. Олигонуклеотидный зонд для применения в способах согласно изобретению, как правило, составляет примерно от 8 до примерно 25 нуклеотидов в длину, например, от примерно 10 до примерно 20, от примерно 12 до примерно 25 или от примерно 15 до примерно 25 нуклеотидов в длину. Согласно некоторыми вариантам реализации изобретения, зонд может также использоваться в качестве олигонуклеотида для внедрения в цепь (и, соответственно, обладать признаками, описанными выше для олигонуклеотидов для внедрения в цепь). Например, может быть предложен дополнительный меченый олигонуклеотид для внедрения в цепь, действующий в качестве зонда, который имеет участок связывания в последовательности нуклеиновой кислоты-мишени вблизи 3'-5' или 5'-3' участка связывания олигонуклеотида для внедрения в цепь таким образом, что он может образовывать пару FRET с соответствующим олигонуклеотидом для внедрения в цепь, связывающимся с 3'-5' или 5'-3' участком. Согласно указанному варианту реализации изобретения, олигонуклеотид для внедрения в цепь, связывающийся с 3'-5' или 5'-3' участком, можно метить флуорофором или гасителем, и дополнительный олигонуклеотид для внедрения в цепь можно метить соответствующим взаимодействующим выявляемым фрагментом (гасителем или флуорофором).
Зонд может содержать последовательность, которая полностью комплементарна последовательности нуклеиновой кислоты-мишени, или может иметь одно или более несоответствий, например, 2 или 3 несоответствия последовательности-мишени, при условии что он способен специфично выявлять последовательность-мишень при использовании в комбинации с олигонуклеотидом для внедрения в цепь (олигонуклеотидами) и праймером (праймерами). Олигонуклеотидный зонд для применения согласно изобретению может представлять собой гибридизационный зонд, показывающий конформационные изменения при связывании мишени (как описано, например, в патенте США US 7241596), молекулярный маяк (как описано, например, в патенте США US 5925517) или расщепляемый зонд, такой как расщепляемый эндонуклеазой зонд (как описано, например, в патентах США US 7435561 и US 20050214809), или расщепляемый ферментом для рестрикции зонд.
Праймер, олигонуклеотид для внедрения в цепь или зонд, используемые в способах согласно изобретению, могут быть помечены любым флуорофором или гасителем. Флуорофор и гаситель выбирают таким образом, чтобы спектр поглощения гасителя перекрывался со спектром испускания флуорофора. Флуорофор и гаситель также выбирают и располагают таким образом, чтобы при гибридизации с матрицей-мишенью происходило повышение сигнала флуорофора благодаря сниженному эффекту гашения.
Гаситель может являться не флуоресцентным, например, не флуоресцентным хромофором. Гаситель может представлять собой темновой гаситель. Альтернативно, гаситель может иметь другой спектр флуоресценции по сравнению с флуорофором таким образом, что при специфическом отслеживании флуоресценции флуорофора или гасителя изменение любого сигнала может сообщать о гибридизации с матрицей-мишенью. Флуорофор или гаситель может быть расположен на 5' или 3'-конце молекулы меченного олигонуклеотида. 3'-концевое расположение может применяться, в частности, в вариантах реализации изобретения, в которых зависимое от полимеразы удлинение является нежелательным. Флуорофор или гаситель также может быть расположен внутри молекулы, например, на расстоянии десяти или менее нуклеотидов от 5' или 3'-конца меченной молекулы.
Флуорофор может представлять собой любой флуоресцентный фрагмент, как правило, флуоресцентный органический краситель. Гаситель может представлять собой любой фрагмент, который гасит флуоресценцию флуорофора, и, как правило, хромогенной молекулы, такой как органический краситель. Специалист в данной области техники способен выбрать подходящие пары флуорофор-гаситель для олигонуклеотидного зонда на основе общего известного уровня техники. Образование подходящих пар обсуждается, например, в следующих источниках: Marras SE: Selection of Fluorophore and Quencher Pairs for Fluorescent Nucleic Acid Hybridization Probes. In: Fluorescent Energy Transfer Nucleic Acid Probes. Edited by Didenko V, vol. 335: Humana Press; 2006: 3-16; Didenko VV: DNA probes using fluorescence resonance energy transfer (FRET): designs and applications. Biotechniques 2001, 31(5): 1106-1116, 1118, 1120-1101.
Подходящие флуорофоры включают, но не ограничиваются указанными, флуоресцеин и производные флуоресцеина, такие как карбоксифлуоресцеины (FAM, включая 6-FAM, 5-FAM, dT FAM), VIC, гексахлор-6-карбоксифлуоресцеин (HEX) и JOE, 5-(2'-аминоэтил)аминонафталин-1-сульфоновую кислоту (EDANS), кумарин и производные кумарина, такие как 3-фенил-7-изоцианатокумарин, Lucifer yellow, NED, Texas red, тетраметилродамин, карбокситетраметилродамин (TAMRA), 6-карбокси-Х-родамин (ROX), 5 карбоксиродамин, N-(р-2-бензоксазолил)фенил)малеимид, цианиновые красители, такие как CY5, родаминовые красители, ксантеновые красители, нафтиламины, акридины, бензоксадиазолы, стильбены и пирены. Подходящие гасители включают, но не ограничиваются указанными, DABSYL, 4'-(4-диметиламинофенилазо)бензойную ксилоту (DABCYL), 4-диметиламинофенилазофенил-4'-малеимид (DABMI), тетраметилродамин, карбокситетраметилродамин (TAMRA), гаситель Black Hole 1, гаситель Black Hole 2, гаситель Black Hole 3, гаситель Dark 1, гаситель Dark 2, гаситель Iowa Black RQ и Iowa Black FQ.
Предпочтительные пары флуорофор/гаситель включают:
- TAMRA и Black Hole 2;
- ROX и Black Hole 2;
- ROX и DABCYL;
- FAM (такой как dT-FAM) и Iowa Black FQ;
- FAM (такой как dT-FAM) и DABCYL;
- ROX и Iowa Black FQ;
- CY5 и Iowa Black RQ.
Флуорофор или гаситель, как правило, ковалентно присоединен к меченой молекуле олигонуклеотида. Флуорофор или гаситель может быть присоединен с помощью любого подходящего линкера к одному или более нуклеотидам, присутствующим в последовательности молекулы олигонуклеотида. Специалист в данной области техники способен выбрать любой подходящий линкер на основе общего известного уровня техники. Подходящие линкеры обсуждаются, например, в источнике: Agrawal S (ed.): Protocols for Oligonucleotides and Analogs: Synthesis and Properties: Humana Press; 1993.
Согласно некоторыми вариантам реализации изобретения, способы согласно изобретению могут включать применение одного или более зондов, содержащих область, комплементарную последовательности нуклеиновой кислоты-мишени, флуорофор и гаситель. Последовательность такого олигонуклеотидного зонда может содержать по меньшей мере 20% РНК-нуклеотидов, модифицированных РНК-нуклеотидов и/или ПНК-нуклеотидов. Применение таких зондов обеспечивает преимущество, которое заключается в предупреждении флуоресцентного сигнала от зонда в присутствии белка, способного связываться с одноцепочечными ДНК (такого как рекомбиназа), в отсутствие комплементарной матричной последовательности. Другими словами, по меньшей мере 20% нуклеотидов, присутствующих в олигонуклеотидном зонде, представляют собой РНК-нуклеотиды, модифицированные РНК-нуклеотиды и/или ПНК-нуклеотиды. Более предпочтительно, последовательность олигонуклеотидного зонда может содержать по меньшей мере 25%, по меньшей мере 30%, по меньшей мере 35%, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80% или по меньшей мере 90% РНК-нуклеотидов, модифицированных РНК-нуклеотидов и/или ПНК-нуклеотидов. Если зонд содержит РНК-основания, фермент РНКаза Н, такой как РНКаза Н2, может быть предложен в способе согласно изобретению для усиления сигнала от зонда путем расщепления дуплекса зонд-мишень и снижения гашения. Предпочтительный фермент РНКаза Н2 представляет собой РНКазу Н2 Thermococcus gammatolerans. Альтернативно, могут использоваться другие формы расщепляемого зонда, такие как фермент для рестрикции или расщепляемые эндонуклеазой зонды, как описано выше.
Когда используют зонд, меченный флуорофором, и гаситель, указанные флуорофор и гаситель, как правило, располагаются на расстоянии по меньшей мере восьми нуклеотидов друг от друга в последовательности зонда, более предпочтительно, на расстоянии по меньшей мере десяти или по меньшей мере двенадцати нуклеотидов, в зависимости от длины зонда. Флуорофор и гаситель могут быть расположены на 5'- и 3'-концах, и таким образом, на максимально возможном расстоянии в зонде. Расстояние между флуорофором и гасителем выбирают таким образом, что при гибридизации зонда с последовательностью нуклеиновой кислоты-мишени (в открытой или линейной конформации) интенсивность гашение флуорофора гасителем снижается, приводя к выявляемому сигналу, указывающему на присутствие последовательности нуклеиновой кислоты-мишени. Подходящее расстояние между флуорофором и гасителем может быть оптимизировано опытным путем.
Для выявления амплифицированной ДНК также могут использоваться красители, выстраивающиеся в амплифицированную ДНК, такие как краситель Sybr green I и тиазол оранжевый.
Выявление сигнала от амплифицированной ДНК может осуществляться с помощью любой подходящей системы, включая способы выявления в режиме реального времени.
Применение способов амплификации
Способы амплификации согласно изобретению можно применять в любых случаях, когда желательна специфичная амплификация последовательности нуклеиновой кислоты-мишени.
Способы согласно изобретению могут использоваться для выявления последовательности нуклеиновой кислоты-мишени, и, например, для диагностики наличия в клиническом образце последовательности нуклеиновой кислоты-мишени. Настоящее изобретение обладает особыми преимуществами при применении в медицинских учреждениях. Способы выявления согласно изобретению обеспечивают высокоспецифичный анализ, позволяющий определять присутствие последовательности нуклеиновой кислоты-мишени. Способ может применяться при ряде заболеваний. Изобретение обеспечивает способ диагностики заболевания у субъекта, включающий осуществление способа амплификации последовательности нуклеиновой кислоты-мишени согласно изобретению в образце от указанного субъекта для выявления последовательности нуклеиновой кислоты-мишени, связанной с указанным заболеванием.
Любой образец может использоваться для выявления последовательности нуклеиновой кислоты-мишени, при условии что из указанного образца может быть извлечена или получена нуклеиновая кислота. Образец может представлять собой, например, природный образец, референсный образец или клинический образец. Когда способы согласно изобретению используют для диагностики заболевания путем выявления последовательности нуклеиновой кислоты-мишени, образец обычно представляет собой клинический образец, например, образец, полученный от пациента, страдающего заболеванием, или пациента с подозрением на заболевание. Подходящие типы клинических образцов различаются в соответствии с конкретным типом заболевания или инфекции, которая присутствует или предположительно присутствует у пациента. Образец может представлять собой образец слюны, мокроты, крови, плазмы крови, сыворотки крови, мочи или стула. Образец может представлять собой клеточный или тканевой образец. Согласно предпочтительным вариантам реализации изобретения, образцы отбирают у субъектов, представляющих собой животных, таких как субъекты, представляющие собой млекопитающих. Образцы в целом отбирают у субъектов, представляющих собой людей. Однако настоящее изобретение также применимо в целом для домашних животных, домашнего скота, птиц и рыб. Например, изобретение можно применять в ветеринарных или сельскохозяйственных учреждениях. Образец содержит нуклеиновую кислоту, которая может представлять собой ДНК или РНК. Если нуклеиновая кислота присутствует в образце в подходящем виде, позволяющем ее выявление в соответствии с изобретением, указанный образец может использоваться непосредственно. Однако, как правило, нуклеиновая кислота происходит, получена или выделена из образца. Способы обработки образцов, содержащих нуклеиновые кислоты, выделения нуклеиновых кислот и/или очистки нуклеиновых кислот для применения в способах выявления хорошо известны в данной области техники. Можно выделять тотальную нуклеиновую кислоту или выделять раздельно ДНК и РНК.
Как правило, образец обрабатывают соответствующим образом для того, чтобы нуклеиновая кислота быть предоставлена в удобной форме для приведения ее в контакт с праймерами и олигонуклеотидом для внедрения в цепь (олигонуклеотидами) и необязательно дополнительными реагентами. Если нуклеиновая кислота представляет собой ДНК, то она, как правило, предложена в двуцепочечной форме. Если нуклеиновая кислота представляет собой РНК, ее, как правило превращают в кДНК с использованием обратной транскриптазы или полимеразы с обратной транскриптазной активностью. РНК может быть применима для выявления бактерий по причине очень большого числа рибосом в бактериальных клетках, эффективно повышающих концентрацию последовательностей-мишеней. Помимо рибосомальной РНК (рРНК) для выявления прокариот и эукариот могут использоваться другие формы РНК, например, транспортные РНК (тРНК), информационные РНК (иРНК), малые интерферирующие РНК (миРНК), малая ядерная РНК (мяРНК), микроРНК.
Способ согласно изобретению может использоваться для диагностики инфицирования субъекта патогеном, включая выявление последовательности нуклеиновой кислоты-мишени от указанного патогена. Определение наличия патогена может осуществляться при любом присутствующем заболевании или расстройстве или при подозрении на заболевание или расстройство у пациента. Такие заболевания могут включать заболевания, вызванные присутствием патогена, связанные с присутствием патогена или усугубляющиеся присутствием патогена. Таким образом, пациент может проявлять симптомы, указывающие на присутствие патогена, и образец может быть получен от указанного пациента для определения присутствия патогена с помощью способа, описанного выше.
Можно выявлять любой патоген. Патоген может представлять собой вирус или бактерию или паразита. Патоген может представлять собой патоген, включая, но не ограничиваясь указанными, грибы, вирусы, включая вирусы папилломы человека (ВПЧ), ВИЧ, ВПГ2/ВПГ1, вирус гриппа (типа А, В и С), полиовирус, вирус саркомы Рауса, риновирусы, ротавирусы, вирус гепатита А, группу норовирусов, энтеровирусы, астровирусы, вирус кори, вирус парагриппа, вирус эпидемического паротита, вирус ветряной оспы, цитомегаловирус, вирус Эпштейна-Барра, аденовирусы, вирус краснухи, вирус Т-клеточной лимфомы человека I типа (ВТЛЧ-I), вирус гепатита В, вирус гепатита С, вирус гепатита D, поксвирус, вирус геморрагической лихорадки Марбург и Эбола; бактерии, включая следующие виды бактерий: Mycobacterium tuberculosis, Neisseria gonorrhoeae, Vibrio cholerae, Treponema pallidum, Bordetella pertussis, Franciscella tularensis, Helicobacter pylori, Leptospira interrogans, Legionella pneumophila, Yersinia pestis, Haemophilus influenza (тип b), Toxoplasma gondii, Moraxella catarrhalis, а также хламидии, шигеллы, сальмонеллы, псевдомонады, бруцеллы, стрептококки (типа А и В), пневмококки, менингококки и возбудителей кампиллобацилярного энтерита, донованоза и актиномикоза; грибковые патогены, включая возбудителей кандидоза и аспергиллеза; паразитарные патогены, включая вид Pneumocystis carinii, а также цепней, сосальщиков, круглых червей, криптоспоридий, шистосом, возбудителей амебиоза, лямблиоза, трихомониаза и трихинеллеза.
Кроме того, применение способов согласно изобретению включает анализ, клонирование фрагментов и выявление с помощью однонуклеотидного полиморфизма (single-nucleotide polymorphism, SNP).
Согласно другому аспекту изобретения, последовательность нуклеиновой кислоты-мишени может быть амплифицирована для возможности определения ее последовательности. Согласно такому варианту реализации изобретения, последовательность нуклеиновой кислоты с частично или полностью неизвестной последовательностью можно амплифицировать путем создания подходящих участков связывания одного или более олигонуклеотидов для внедрения в цепь (олигонуклеотида), фланкирующих область, последовательность которой подлежит выявлению. Изобретение, таким образом, обеспечивает способ определения последовательности нуклеиновой кислоты-мишени, содержащей область неизвестной последовательности, включающий создание последовательности нуклеиновой кислоты-мишени, содержащей участки связывания олигонуклеотидов для внедрения в цепь, расположенные выше и ниже указанной области неизвестной последовательности, амплификацию указанной последовательности нуклеиновой кислоты-мишени в соответствии со способом амплификации согласно изобретению, описанным выше, и определение последовательности указанной область неизвестной последовательность. Изобретение также обеспечивает способ амплификации последовательности нуклеиновой кислоты-мишени, содержащей область неизвестной последовательности, включающий создание последовательности нуклеиновой кислоты-мишени, содержащей участки связывания олигонуклеотидов для внедрения в цепь, расположенные выше и ниже указанной области неизвестной последовательности, и амплификацию указанной последовательности нуклеиновой кислоты-мишени в соответствии со способом амплификации согласно изобретению, описанным выше. Последовательность нуклеиновой кислоты-мишени, содержащая участки связывания в 3'-5' и 5'-3' направлении олигонуклеотидов для внедрения в цепь, может быть создана путем лигирования олигонуклеотидов, содержащих участок связывания олигонуклеотидов для внедрения в цепь, к 5' и/или 3'-концам интересующей последовательности нуклеиновой кислоты. Альтернативно, интересующая последовательность нуклеиновой кислоты может быть встроена или лигирована в подходящую векторную нуклеиновую кислоту, такую как плазмида, содержащую участок связывания олигонуклеотидов для внедрения в цепь, фланкирующий сайт предполагаемого встраивания последовательности нуклеиновой кислоты, с получением, таким образом, последовательности нуклеиновой кислоты-мишени. Согласно другим вариантам реализации изобретения, последовательность, подлежащая определению, может быть частично известна. Таким образом, можно создать один вид олигонуклеотида для внедрения в цепь (и его соответствующего праймера), связывающийся с областью известной последовательности, и другие виды олигонуклеотидов для внедрения в цепь и праймеров, связывающиеся с введенной адаптерной последовательностью, фланкирующей область неизвестной последовательности. Способ амплификации на основе внедрения в цепь в положении, расположенном выше известной последовательности и ниже неизвестной последовательности, можно в дальнейшем использовать для амплификации области неизвестной последовательности с возможностью определения ее последовательности.
Подходящие олигонуклеотиды, содержащие участки связывания олигонуклеотидов для внедрения в цепь, представляют собой ДНК-адаптеры, как правило, представленные в виде двуцепочечной молекулы с липкими концами или без липких концов. В целом, если адаптер представлен двуцепочечным олигонуклеотидом, он имеет тупые концы для возможности его лигирования с интересующим фрагментом ДНК. Олигонуклеотиды, содержащие участок связывания олигонуклеотидов для внедрения в цепь, могут дополнительно содержать участок связывания праймеров. Может быть предложен один вид олигонуклеотида, содержащего участок связывания олигонуклеотида для внедрения в цепь (и необязательно также участок связывания праймера), где один и тот же вид олигонуклеотида для внедрения в цепь (и необязательно один и тот же вид праймера) используется для обеспечения внедрения в последовательность нуклеиновой кислоты-мишени в 3'-5'и 5'-3'положениях.
Определение последовательности нуклеиновой кислоты-мишени можно осуществлять с использованием любого подходящего способа секвенирования. Подходящие способы секвенирования включают секвенирование по Сэнгеру или способы секвенирования нового поколения типа секвенирования Ion Torrent, SOLiD, Illumina и 454 секвенирование. Фрагменты, подлежащие секвенированию, можно предварительно амплифицировать непосредственно с присоединенных к ним адаптеров или сначала клонировать в плазмиды для секвенирования. В последнем случае клонированный фрагмент может содержать адаптерную последовательность (последовательности), или же указанная адаптерная последовательность может быть представлена плазмидой, в которую лигирован фрагмент.
Может использоваться любая подходящая адаптерная последовательность, которая обеспечивает возможность связывания олигонуклеотида для внедрения в цепь и/или праймера при встраивании в положении, фланкирующем последовательность нуклеиновой кислоты-мишени, или внутри последовательности нуклеиновой кислоты-мишени для обеспечения возможности амплификации указанной последовательности нуклеиновой кислоты-мишени. Адаптерная последовательность, встроенная в положении, фланкирующем или находящемся в пределах 3'-5' области последовательности нуклеиновой кислоты-мишени, как правило, является идентичной адаптерной последовательности, встроенной в положении, фланкирующем или находящемся в пределах 5'-3' области последовательности нуклеиновой кислоты-мишени. Однако могут использоваться различные адаптерные последовательности для 3'-5' и 5'-3' области последовательности нуклеиновой кислоты-мишени, при условии что предложены один или более олигонуклеотидов для внедрения в цепь и один или более праймеров, способных амплифицировать последовательность нуклеиновой кислоты-мишени на основе различных адаптеров. Специалист в данной области техники способен выбрать подходящую адаптерную последовательность для конкретной последовательности-мишени. Адаптерные последовательности можно выбирать на свое усмотрение таким образом, чтобы они не мешали каким-либо возможно присутствующим последовательностям. Адаптерные последовательности также могут быть выбраны с учетом предпочтения рекомбиназы к пиримидину. Адаптерные последовательности могут содержать метку для очистки или разделения до или после амплификации. Можно добавлять сайты рестрикции или сайты распознавания никирующего фермента, способствующие дополнительной обработке ампликонов.
Наборы и композиции
Изобретение обеспечивает набор или композицию, содержащую по меньшей мере один прямой и по меньшей мере один обратный праймер для последовательности нуклеиновой кислоты-мишени и первый и второй олигонуклеотиды для внедрения в цепь, которые имеют участки связывания в 3'-5' и 5'-3' направлении в указанной последовательности нуклеиновой кислоты-мишени.
Изобретение также обеспечивает набор или композицию, содержащую олигонуклеотид для внедрения в цепь, который может связываться как с 3'-5' участком связывания, так и с 5'-3' участком связывания в последовательности нуклеиновой кислоты-мишени, один или более праймеров, способных обеспечивать амплификацию указанной последовательности нуклеиновой кислоты-мишени, и по меньшей мере один ДНК-адаптер. ДНК-адаптер, как правило, является двуцепочечным. Набор или композиция может дополнительно содержать ДНК-лигазу, которую можно использовать для лигирования ДНК-адаптера с интересующей нуклеиновой кислотой. Набор или композиция может дополнительно содержать один или более ферментов для рестрикции. Как правило, участки связывания в 3'-5' и 5'-3' направлении для олигонуклеотида для внедрения в цепь включают последовательность ДНК-адаптера, и таким образом, олигонуклеотид для внедрения в цепь способен связываться по меньшей мере с частью ДНК-адаптера. Олигонуклеотид для внедрения в цепь способен делать участки связывания в 3'-5' и 5'-3' направлении одного или более праймеров одноцепочечными. Один или более праймеров, таким образом, связываются с участками в последовательности нуклеиновой кислоты-мишени, расположенными вблизи участков связывания олигонуклеотида для внедрения в цепь. Один или более праймеров могут также связываться с последовательностью ДНК-адаптера, такой как ДНК-адаптер, обеспечивая возможность как внедрения в цепь, так и амплификации неизвестной последовательности. Набор или композиция может содержать один вид праймера, который может связываться как с 3'-5', так и с 5'-3' участком связывания в последовательности нуклеиновой кислоты-мишени (как правило, адаптерной последовательностью), или прямой и обратный праймеры для указанной последовательности нуклеиновой кислоты-мишени.
Согласно связанному аспекту изобретения, предложен набор или композиция, содержащая олигонуклеотид для внедрения в цепь, который может связываться как с 3'-5', так и 5'-3' участком связывания в последовательности нуклеиновой кислоты-мишени, и один или более праймеров, способных обеспечивать амплификацию указанной последовательности нуклеиновой кислоты-мишени, где олигонуклеотид для внедрения в цепь способен делать участки связывания в 3'-5' и 5'-3' направлении одного или более праймеров одноцепочечными, и где каждый из олигонуклеотида для внедрения в цепь и одного или более праймеров связывается с ДНК-адаптерной последовательностью. Каждый из олигонуклеотида для внедрения в цепь и одного или более праймеров может, таким образом, быть способен связываться с идентичной ДНК-адаптерной последовательностью, присутствующей в 3'-5' и 5'-3' положении в последовательности нуклеиновой кислоты-мишени. Набор или композиция может содержать нуклеиновую кислоту-вектор, содержащую адаптерные последовательности, фланкирующие сайт клонирования.
Праймер (праймеры) и олигонуклеотид для внедрения в цепь (олигонуклеотиды), предложенные в описанных выше наборах или композициях, могут представлять собой любые из праймеров и олигонуклеотидов для внедрения в цепь, описанных выше для применения в соответствующих способах согласно изобретению. Наборы и композиции согласно изобретению могут также содержать один или более дополнительных олигонуклеотидов для внедрения в цепь.
Композиция может представлять собой, например, раствор, лиофилизат, суспензию или эмульсию в масляном или водном носителе.
В наборе согласно изобретению различные виды олигонуклеотидов (такие как праймер (праймеры) и олигонуклеотид для внедрения в цепь (олигонуклеотиды)) могут быть представлены в виде смеси или в раздельных емкостях. Набор может также необязательно содержать инструкции по применению в способах согласно изобретению. Таким образом, набор, содержащий первый и второй олигонуклеотид для внедрения в цепь, может содержать инструкции по применения для использования в способах амплификации последовательности нуклеиновой кислоты-мишени согласно изобретению, включающих применение первого и второго олигонуклеотида для внедрения в цепь. Набор может содержать устройство для выявления амплифицированной ДНК. Набор может содержать реагенты для секвенирования ДНК.
Набор или композиция согласно изобретению может необязательно содержать один или более зондов, выявляющих амплифицированную ДНК. Зонд, предложенный в наборе или композиции, может представлять собой любой из зондов, описанных выше для применения в способах согласно изобретению.
Набор или композиция может необязательно содержать одну или более из ДНК-полимеразы, рекомбиназы и вспомогательных белков системы рекомбинации. Предпочтительно, ДНК-полимераза представляет собой полимеразу Bsu. Предпочтительно, рекомбиназа представляет собой рекомбиназу UvsX бактериофага Т4, необязательно в комбинации со вспомогательными белками системы рекомбинации UvsY и gp32. Набор или композиция может дополнительно содержать дНТФ, подходящие буферы и другие факторы, которые требуются для амплификации ДНК в способах согласно изобретению, как описано выше.
Следующие далее примеры иллюстрируют настоящее изобретение.
Примеры
Пример 1. Амплификация ДНК-мишени с использованием двух олигонуклеотидов для внедрения в цепь
Способы применения двух олигонуклеотидов для внедрения в цепь для амплификации искусственной ДНК-мишени показаны на Фигурах 1 и 2. Все используемые олигонуклеотиды были получены из компании MWG Eurofins (Германия) или компании IDT ДНК Technologies (Бельгия). Все реагенты и буферы, включая креатинкиназу и фосфорилазу сахарозы, были получены из Sigma-Aldrich (Сент-Луис, Миссури, США). Белок бактериофага Т4, связывающийся с одноцепочечной ДНК, (gp32) и полимераза BSU были получены из компании New England Biolabs (Ипсвич, Массачусетс, США). Белки UvsX и UvsY были очищены, как описано ранее [1, 2]. РНКаза HII Thermococcus gammatolerans была получена из компании GeneSys Ltd (Великобритания).
Изотермические реакции амплификации ДНК проводили при температуре 40°С в течение по меньшей мере 90 минут. Реакционный объем составлял 20 мкл, если иное не указано. Буферный раствор для реакций состоял из 10 мМ Tris-ацетата, рН 8,0, 10 мМ ацетата магния, 5% ДМСО, 5% ПЭГ 1000, 4 мМ ДТТ, 0,5 мМ ЭДТА, 0,1 мг/мл БСА, 150 мМ сахарозы, 2 мМ АТФ, 200 мкМ дНТФ, красителя SybrGreen I в разведении 1:100000 и 60 мМ Tris-фосфокреатина. Белки, содержащие в буфере, представляли собой gp32 в концентрации 250 нг/мкл, 5 мкМ UvsX, 0,0625 ед/мкл BSU, 0,0125 ед/мкл фосфорилазы сахарозы и 0,025 ед/мкл креатинкиназы. Концентрация каждого праймера и олигонуклеотида для внедрения в цепь составляла 200 нМ, если иное не указано. Были созданы олигонуклеотиды для внедрения в цепь, связывающиеся либо в параллельной, либо в антипараллельной конфигурации с дуплексом-мишенью (Фигура 1). Реакции инициировали путем добавления ацетата магния отдельно или совместно с ДНК-мишенью, присутствующей в концентрации 10 фМ или 1 пкМ. Выявление амплификации в режиме реального времени осуществляли с использованием устройства Agilent MX Pro. Устройство было запрограммировано на циклы по 60 секунд при температуре 40°C с выявлением флуоресценции для каждого цикла. Специфичность продуктов реакции оценивали путем проведения анализа плавления сразу по окончании циклов. Анализ плавления осуществляли путем быстрого нагрева реакционной смеси до 95°С в течение 15 секунд с последующим этапом быстрого охлаждения до 25°С в течение 60 секунд. Затем реакционные смеси медленно нагревали от температуры 25°С до температуры 85°С, при этом измеряли флуоресценцию при каждом повышении температуры на 0,5°С.
В случае параллельной конфигурации использовали два вызывающих внедрение в цепь олигонуклеотида (SEQ ID NO: 1 и 2) и прямой (SEQ ID NO: 3) и обратный (SEQ ID NO: 4) праймеры. Амплификация выявлялась только в реакции, включающей ДНК-мишень, содержащую последовательность SEQ ID NO: 11. Реакция без ДНК-мишени (в отсутствие матричного контроля, ОМК) не приводила к амплификации, как показывает отсутствие сигнала от красителя Sybr Green I (Фигура 2а). Специфичность реакции при параллельной конфигурации также была показана путем добавления смеси геномной ДНК от 15 видов бактерий, не содержащих ДНК-мишень (добавляли 1000 копий геномной ДНК каждого вида на реакцию). В указанной смеси амплификация не выявлялась, что указывает на то, что указанная конфигурация обеспечивает выявление только ДНК-мишени. Анализ плавления с использованием красителя Sybr Green I также подтвердил, что в реакционных пробирках, содержащих ДНК-мишень, происходила специфичная реакция амплификации (Фигура 2b).
В случае антипараллельной конфигурации использовали два вызывающих внедрение в цепь олигонуклеотида (SEQ ID NO: 1 и 2) и прямой (SEQ ID NO: 3) и обратный (SEQ ID NO: 5) праймеры. Амплификация выявлялась только при добавлении в реакцию ДНК-мишени, содержащей последовательность-мишень SEQ ID NO: 12 (Фигура 2с). Кроме того, при использовании указанной конфигурации амплификация не наблюдалось в реакции ОМК и в реакции с использованием смеси геномной ДНК от видов бактерий. Анализ плавления с использованием красителя Sybr Green I подтвердил, что в реакционных пробирках, содержащих ДНК-мишень, происходила специфичная реакция (Фигура 2d).
Также проводили анализ реакции при обратной антипараллельной конфигурации, показанной на Фигуре 1с. Два вызывающих внедрение в цепь олигонуклеотида (SEQ ID NO: 1 и 13) и прямой (SEQ ID NO: 14) и обратный (SEQ ID NO: 25) праймеры использовали для амплификации ДНК матрицы-мишени, включающей последовательность SEQ ID NO: 26. Амплификация выявлялась только в случае добавления ДНК-мишени в реакцию (Фигура 2е). В отсутствие ДНК-мишени не наблюдали выявляемого сигнала в образце. Анализ плавления с использованием красителя Sybr Green I также подтвердил, что в реакционных пробирках, содержащих ДНК-мишень, происходила специфичная реакция (Фигура 2f).
Пример 2. Необходимость олигонуклеотидов для внедрения в цепь для амплификации
Оба вызывающих внедрение в цепь олигонуклеотида были необходимы для экспоненциальной амплификации ДНК-мишени. Это было показано с использованием двух олигонуклеотидов для внедрения в цепь, комплементарных последовательности ДНК-мишени, или одного комплементарного и одного некомплементарного олигонуклеотида для внедрения в цепь. В случае параллельной (Фигура 3а) и антипараллельной (Фигура 3b) конфигурации другой вызывающий внедрение в цепь олигонуклеотид (SEQ ID NO: 1), комплементарный последовательности ДНК-мишени, был замещен на некомплементарный вызывающий внедрение в цепь олигонуклеотид (SEQ ID NO: 6). Праймеры и реагенты, используемые для обеих конфигураций были такими, как описано в Примере 1. Реакции амплификации проводили в присутствии 1 пкМ ДНК-мишени. Амплификация ДНК-мишени происходила только тогда, когда оба вызывающих внедрение в цепь олигонуклеотида были комплементарны последовательности ДНК-мишени (Фигура 3). Эти данные также предполагают, что продукт амплификации содержит полноразмерную матрицу-мишень. Более того, реакция ОМК или реакция с замещением комплементарного олигонуклеотида для внедрения в цепь на некомплементарный олигонуклеотид не приводили к амплификации. Эти данные демонстрируют, что праймеры не приводят к амплификации ДНК-мишени в отсутствие олигонуклеотида для внедрения в цепь.
Пример 3. Необходимость праймеров для амплификации
Прямой и обратный праймеры необходимы для амплификации ДНК-мишени. Это было показано путем замещения одного из праймеров, комплементарных ДНК-мишени, на некомплементарный праймер. Реакцию амплификации проводили либо в присутствии обратного праймера последовательности SEQ ID NO: 4, либо в присутствии обратного праймера, не комплементарного ДНК-мишени (SEQ ID NO: 7). Концентрация ДНК-мишени составляла 1 пкМ. Амплификация выявлялась только в реакции, содержащей обратный праймер (SEQ ID NO: 4), комплементарный ДНК-мишени (Фигура 4). В реакции, в которой комплементарный обратный праймер (SEQ ID NO: 4) был замещен на некомплементарный обратный праймер (SEQ ID NO: 7), амплификация не выявлялась. Такие данные указывают на то, что олигонуклеотиды для внедрения в цепь сами по себе не приводят к амплификации последовательности ДНК-мишени из-за их неспособности служить в качестве субстратов для полимеразы. Более того, указанные данные показывают, что все олигонуклеотиды требуются для прохождения амплификации.
Пример 4. Совместимость мишень-специфичного зонда с двумя олигонуклеотидами для внедрения в цепь
Амплификация на основе внедрения в цепь с двумя олигонуклеотидами для внедрения в цепь обеспечивает улучшенные возможности для создания зондов, специфичных в отношении ДНК-мишени. Как при параллельной, так и при антипараллельной конфигурации ДНК-мишень содержит участки, не комплементарные ни вызывающим внедрение в цепь олигонуклеотидам, ни праймерам. Наличие указанных участков предполагает возможность создания дополнительных зондов для возможности их связывания без конкуренции с олигонуклеотидами для внедрения в цепь или праймерами (Фигура 5а). Анализ реакции амплификации при антипараллельной конфигурации проводили в присутствии мишень-специфичного зонда. Указанный зонд содержал флуорофор и гаситель, разделенные между собой РНК-основанием. Кроме того, зонд был заблокирован на 3'-конце для дополнительной гарантии отсутствия неспецифического удлинения. Концентрация олигонуклеотидов для внедрения в цепь (SEQ ID NO: 1 и 2) составляла 200 нМ, тогда как концентрация праймеров (SEQ ID NO: 3 и 4) и зонда (SEQ ID NO: 8) составляла 400 нМ. Стандартную реакцию амплификации проводили, как описано в Примере 1 в присутствии РНКазы Н2 Thermococcus gammatolerans в концентрации 10 мкг/мл. Продукты амплификации в режиме реального времени выявляли либо с помощью красителя Sybr Green I, либо с помощью зонда, меченного флуорофором и гасителем (Фигура 5b). Результаты показывают, что зонд эффективно связывался с ДНК-мишенью во время реакции и генерировал выявляемый сигнал.
Описанный ранее способ амплификации на основе внедрения в цепь с использованием только одного олигонуклеотида для внедрения в цепь (SIBA) предполагает сложные химические реакции с участием зонда, поскольку конфигурация не позволяет зонду свободно связываться с областью-мишенью. Это вызвано тем, что все участки ДНК-мишени, как правило, служат в качестве сайта связывания олигонуклеотида для внедрения в цепь или праймеров. Таким образом, мишень-специфичный зонд должен конкурировать с вызывающим внедрение в цепь олигонуклеотидом и праймерами за связывание, если он не сконструирован специальным образом. Результаты, показанные на Фигуре 5с, иллюстрируют проблемы, связанные с использованием мишень-специфичного зонда, специфичного для той же области-мишени, что и вызывающий внедрение в цепь олигонуклеотид в SIBA. В анализе SIBA концентрации олигонуклеотида для внедрения в цепь (SEQ ID NO: 1), прямого праймера (SEQ ID NO: 3) и обратного праймера (SEQ ID NO: 5) составляли 200 нМ, тогда как концентрация зонда (SEQ ID NO: 9) составляла 400 нМ. За реакцией амплификации в режиме реального времени следили либо с помощью красителя Sybr Green I, либо с помощью зонда (Фигура 5с). Повышение сигнала от красителя Sybr Green I в реакции, содержащей ДНК-мишень, показало, что происходила амплификация. Однако при использовании зонда, перекрывающегося с сайтом связывания олигонуклеотида для внедрения в цепь, сигнал не выявлялся, что дает основания предполагать, что зонд был неспособен связываться с ДНК-мишенью. Это, вероятно, вызвано конкуренцией между вызывающим внедрение в цепь олигонуклеотидом и зондом.
Пример 5. Применение мишень-специфичного зонда при использовании конфигурации с двумя олигонуклеотидами для внедрения в цепь приводит к снижению сигнала неспецифической амплификации, вызванной использованием коротких праймеров.
Применение очень коротких праймеров может приводить к неспецифической амплификации в реакции SIBA из-за возможного удлинения олигонуклеотида для внедрения в цепь. Такая неспецифическая амплификация выявлялась при использовании определенных коротких праймеров (14 оснований), которые потенциально могли удлинять область ДНК олигонуклеотида для внедрения в цепь (Фигура 6b). Таким образом, в стандартной SIBA праймеры создают таким образом, чтобы их длина составляла примерно 16-23 основания, а их 3'-концы были частично гомологичны вызывающему внедрение в цепь олигонуклеотиду. При конфигурации с двумя олигонуклеотидами для внедрения в цепь вероятность выявления в способе амплификации продуктов неспецифической амплификации, полученных в результате применения коротких праймеров, ниже, что связано с возможностью использования сайта связывания зонда, не перекрывающегося с сайтами связывания какого-либо из олигонуклеотидов для внедрения в цепь или праймеров.
На Фигуре 6 показана реакция амплификации, осуществляемая либо с использованием двух олигонуклеотидов для внедрения в цепь в антипараллельной конфигурации, либо с помощью стандартной SIBA. В обоих способах амплификации на основе внедрения в цепь применение коротких праймеров (14 оснований в указанном примере) и длинных праймеров (21 основание в указанном примере) приводило к повышению сигнала красителя Sybr Green I в присутствии матрицы-мишени. Однако при использовании коротких праймеров низкий сигнал также выявлялся в реакции ОМК, тогда как использование длинных праймеров не приводило к появлению сигнала в реакции ОМК. В то же время конфигурация с двумя олигонуклеотидами для внедрения в цепь позволяла зонду связываться с последовательностью-мишенью без конкуренции с вызывающим внедрение в цепь олигонуклеотидом и праймерами. Такая конфигурация приводила к предотвращению выявления сигнала, возникающего при использовании коротких праймеров в реакции ОМК (Фигура 6а).
Пример 6. Амплификация плазмидной ДНК с использованием двух олигонуклеотидов для внедрения в цепь
Способ амплификации с двумя олигонуклеотидами для внедрения в цепь можно использовать для амплификации различных типов ДНК-матриц. Дуплекс ДНК-мишень (мишень для анти/параллельной конфигурации, SEQ ID NO: 20) клонировали в коммерчески доступный вектор pCR2,1, MWG Eurofins (Германия). Вставку фланкировали сайтами рестрикции EcoRI. Плазмиду затем использовали в качестве матрицы в способе с антипараллельной конфигурацией двух олигонуклеотидов для внедрения в цепь, как описано в Примере 1. Концентрация каждого олигонуклеотида для внедрения в цепь, праймера и зонда составляла 200, 400 и 600 нМ соответственно. Соответствующее количество (100 фМ - 10 пкМ) расщеплений рестриктазой или нерасщепленной плазмиды добавляли в реакцию, как показано на Фигуре 7. Расщепленную плазмиду получали путем инкубирвоания плазмиды-мишени с ферментом для рестрикции EcoRV-HF (New England Biolabs, Ипсвич, Массачусетс, США) в течение 3 часов при температуре 37°С. Как показано на Фигуре 7, способ позволял амплифицировать ДНК-мишень как из разрезанной рестриктазой, так и из нерасщепленной плазмиды. Однако плазмидная ДНК, разрезанная с помощью фермента для рестрикции, выявлялась раньше по сравнению с нерасщепленной плазмидой. Это может быть вызвано задержкой, связанной с первым циклом амплификации, когда циклические плазмиды используются в качестве матрицы-мишени. Кроме того, такие данные предполагают, что ферменты для рестрикции можно использовать для минимизации задержки во время первого цикла амплификации при использовании комбинированной ДНК в качестве матрицы, что можно осуществить путем предварительного инкубирования комбинированной ДНК с подходящими ферментами для рестрикции или путем добавления фермента для рестрикции к реагентам амплификации.
Пример 7. Амплификация ДНК-мишени с несколькими сайтами внедрения в цепь
Может быть создан дуплекс-мишень, терминальные участки которого содержат идентичные сайты связывания для олигонуклеотидов для внедрения в цепь. В этом случае предполагается, что только один вызывающий внедрение в цепь олигонуклеотид требуется для диссоциации обоих концов дуплекса-мишени. Такой дуплекс-мишень имитирует библиотеку фрагментов неизвестной ДНК, которые лигированы с адаптерами (известной последовательностью), или фрагментов ДНК, в которых известна только часть последовательности. Указанные фрагменты неизвестной ДНК затем можно амплифицировать с использованием адаптер-специфичных праймеров. Продукты амплификации затем используются в качестве матрицы для последующих способов применения, таких как секвенирование ДНК. Такую систему также можно использовать для других последующих способов применения, таких как анализ фрагментов, клонирование, выявление однонуклеотидного полиморфизма (SNP). Фрагменты ДНК можно эффективно амплифицировать с использованием двух идентичных последовательностей-мишеней для одного вида олигонуклеотида для внедрения в цепь в параллельной, анти-параллельной или обратной антипараллельной конфигурации (Фигура 8). В указанном примере стандартную реакцию амплификации проводили, как описано в Примере 1, за исключением того, что 5% ПЭГ 1000 был замещен на 7,5% ПЭГ 400.
В случае параллельной конфигурации использовали 400 нМ олигонуклеотида для внедрения в цепь (SEQ ID NO: 13), 200 нМ прямого (SEQ ID NO: 14) и 200 нМ обратного праймера (SEQ ID NO: 15) для амплификации приемлемого количества дуплекса ДНК-мишени, содержащего 324 п.н. (SEQ ID NO: 16). Дуплекс-мишень (SEQ ID NO: 16) содержал 200 п.н. фрагмента гена лактазы человека (LCT), фланкированного последовательностями, которые служили в качестве сайтов связывания для олигонуклеотида для внедрения в цепь (SEQ ID NO 28 и 29). Амплификацию выявляли только в реакции, которая содержала дуплекс ДНК-мишени (SEQ ID NO: 16). В реакциях в отсутствие дуплекса ДНК-мишени (отсутствие матричного контроля, ОМК) не наблюдалось какого-либо выявляемого сигнала от красителя Sybr Green I (Фигура 8А). Наблюдалась очень высокая скорость и эффективность амплификации - 1000 копий ДНК-мишени выявлялось в течение 20 минут от начала реакции. Анализ плавления с использованием красителя Sybr Green I также подтвердил, что реакции были специфичными (Фигура 8В).
Было показано, что при параллельной конфигурации с использованием 400 нМ олигонуклеотида для внедрения в цепь (SEQ ID NO: 2), 200 нМ прямого (SEQ ID NO: 5) и 200 нМ обратного праймера (SEQ ID NO: 4) также наблюдалась амплификация другой ДНК-мишени (SEQ ID NO: 17), содержащей фланкирующие последовательности, которые служат в качестве сайтов связывания для олигонуклеотида для внедрения в цепь (SEQ ID NO 30 и 31). Эффективность реакции была сходной с показанной ранее эффективностью, как видно из Фигуры 8А. Было обнаружено, что амплификация ДНК-мишени (SEQ ID NO: 17) также была очень эффективна - 1000 копий было выявлено в течение 20 минут от начала реакции (Фигура 8С). Анализ плавления с использованием красителя Sybr Green I также подтвердил, что в реакционных пробирках, содержащих ДНК-мишень, происходила специфичная реакция амплификации (Фигура 8D).
В случае антипараллельной конфигурации использовали 400 нМ олигонуклеотида для внедрения в цепь (SEQ ID NO: 2) и 400 нМ праймера (SEQ ID NO: 5) для амплификации ДНК-мишени (SEQ ID NO: 18), содержащей адаптерную последовательность (SEQ ID NO: 27). Праймер (SEQ ID NO: 5) служил в качестве как прямого, так и обратного праймера. Таким образом, можно использовать только один IO и один вид праймера для амплификации ДНК-мишени, если последовательности связываются подходящем образом. Амплификация выявлялась только в реакциях, которые содержали ДНК-мишень (SEQ ID NO: 18). Реакции без ДНК-мишени (в отсутствие матричного контроля, ОМК) не приводили к какому-либо выявляемому сигналу от красителя Sybr Green I (Фигура 8Е). Анализ плавления с использованием красителя Sybr Green I также показал, что реакция была мишень-специфичной (Фигура 8F).
Подобные исследования проводили с использованием обратной антипараллельной конфигурации. Использовали 400 нМ олигонуклеотида для внедрения в цепь (SEQ ID NO: 13) и 400 нМ праймера (SEQ ID NO: 15) для амплификации ДНК-мишени (SEQ ID NO: 19). Праймер (SEQ ID NO: 15) служил в качестве как прямого, так и обратного праймера. Использование указанной конфигурации также обеспечивало возможность специфичной амплификации ДНК-мишени, при этом неспецифичные реакции не наблюдались в реакции ОМК (Фигура 8G). Анализ плавления с использованием красителя Sybr Green I также показал, что реакция была мишень-специфичной (Фигура 8Н).
Скорость амплификации при использовании обратной антипараллельной амплификации оказалась ниже по сравнению с параллельной и антипараллельной конфигурацией. Самая высокая скорость амплификации наблюдалась при параллельной конфигурации. Различия в скорости амплификации могут быть опосредованы различными факторами. Например, несмотря на то что во всех трех системах использовался сходный вызывающий внедрение в цепь олигонуклеотид, праймеры в указанных системах различались. Это может объяснять некоторые различия в скорости амплификации, поскольку температура плавления и длина праймеров может влиять на скорость амплификации.
Продукты реакции с использованием антипараллельной конфигурации также подвергали электрофоретическому анализу в полиакриламидном геле (ПААГ-электрофорезу) в неденатурирующих условиях (Фигура 8i). Стандартную реакцию амплификации проводили, как описано в Примере 1, за исключением того, что ацетат магния использовали в концентрации 20 мМ, 5% ПЭГ 1000 был замещен на 7,5% ПЭГ 400, и реакцию проводили при 44°C с использованием устройства для ПЦР Bio Rad CFX 96 в течение 120 минут. Для амплификации ДНК-мишени с получением подходящего количества (SEQ ID NO: 18) использовали 200 нМ олигонуклеотида для внедрения в цепь (SEQ ID NO: 2) и 400 нМ праймера (SEQ ID NO: 5). Праймер (SEQ ID NO: 5) использовали в качестве как прямого, так и обратного праймера. Таким образом, для амплификации ДНК-мишени использовали только один IO и праймер одного вида, поскольку были предложены соответствующее связывающие последовательности. Для ПААГ-электрофореза аликвоту реакционной смеси 5 мкл наносили на 8% гель в трис-боратном буфере (ТВЕ, Invitrogen, Великобритания). Электрофорез проводили при постоянном напряжении 150 V в течение 60 мин. Гели окрашивали флуоресцентным красителем для окрашивания нуклеиновых кислот в геле (GelRed; Biotium, США) и визуализировали с использованием системы Gel Doc™ EZ (BioRad, Великобритания). Отдельный банд, соответствующей ожидаемой длине продукта амплификации, наблюдался только в реакции, содержащей матрицу-мишень. В образцах, не содержавших матрицу-мишень, не было выявлено бандов, что указывает на отсутствие продуктов неспецифической амплификации.
Пример 8. Зонды на основе FRET для отслеживания внедрения в цепь и амплификации
В Примере 8 описаны химические реакции, лежащие в основе применения зонда на основе FRET для отслеживания внедрения в цепь и амплификации ДНК-мишени. Химия реакции позволяет одновременно следить за процессами внедрения, происходящими в обоих терминальных участках дуплекса-мишени, а также обеспечивает способ подтверждения амплификации полноразмерной ДНК-мишени. Праймеры и олигонуклеотиды для внедрения в цепь метили флуорофорами и гасителями соответственно для образования системы резонансного переноса энергии флуоресценции (fluorescence resonance energy transfer, FRET), как показано на Фигуре 9А. Праймеры, которые определяют терминальную область дуплекса-мишени, метят флуорофором на 5'-конце или на внутреннем участке (разные флуорофоры для прямого и обратного праймера). Вызывающий внедрение олигонуклеотид, который связывается с 3'-5' участком ДНК-мишени, метят гасителем на 3'-конце, тогда как вызывающий внедрение олигонуклеотид, который связывается с 5'-3' участком ДНК-мишени, метят гасителем на 5'-конце.
Проводили стандартную реакцию амплификации с использованием параллельной конфигурации, как описано в Примере 1, за исключением того, что 5% ПЭГ 1000 был замещен на 7,5% ПЭГ 400. Использовали 200 нМ олигонуклеотида для внедрения в цепь, меченного гасителем в непосредственной близости от его 5'-конца (SEQ ID NO: 21), и 200 нМ олигонуклеотида для внедрения в цепь, меченного гасителем на 3'-конце (SEQ ID NO: 22). Прямой праймер (SEQ ID NO: 23) и обратный праймер (SEQ ID NO: 24), меченные на внутреннем участке флуорофором ROX и Cy5 соответственно, использовали в концентрации 200 нМ. Указанные олигонуклеотиды использовали для амплификации ДНК-мишени (SEQ ID NO: 17). Выявление амплификации в режиме реального времени осуществляли с использованием устройства для ПЦР Bio Rad CFX 96. Сигналы измеряли для 120 циклов после каждого цикла (каждый цикл длится 30 секунд) и выражали в виде относительных единиц флуоресценции (relative fluorescence units, RFU).
На Фигуре 9В показана динамика сигнала системы FRET во время амплификации матрицы-мишени (SEQ ID NO: 17). В ходе реакции терминальные участки ампликона включают различные флуорофоры. Участок в 3'-5' направлении в этом случае включает флуорофор ROX, тогда как 5'-3' участок включает флуорофор Cy5. Во время внедрения в цепь ампликона интенсивность испускаемого флуорофором сигнал снижалась (поскольку олигонуклеотиды для внедрения в цепь, меченные гасителем, оказывались в непосредственной близости от терминального участка ампликона), как видно на Фигуре 9В. Одновременное снижение сигнала, испускаемого прямым и обратным праймерами, указывало на то, что оба праймера были включены в ампликон.
Пример 9. Чувствительность системы множественного внедрения
В указанном примере проводили оценку аналитической чувствительности трех разных анализов. Проводили стандартную реакцию амплификации, как описано в Примере 1, за исключением того, что 5% ПЭГ 1000 был замещен на 7,5% ПЭГ 400. Анализы 1, 2 и 3 использовали для амплификации ДНК-мишени 1 (SEQ ID NO: 16), ДНК-мишени 2 (SEQ ID NO: 17) и ДНК-мишени (SEQ ID NO: 18) соответственно. Используемые олигонуклеотиды, а также их концентрации были такими, как описано в Примере 7. Проводили анализ ДНК-мишени в десятикратных серийных разведениях (от 106 копий до 1 копии). Результаты представлены на Фигуре 10. Чувствительность всех трех анализов составляла по меньшей мере 100 копий ДНК-мишени (анализы 1, 2 и 3 показаны на Фигурах 10А, 10В и 10С соответственно). Для анализа 3, согласно некоторым вариантам реализации изобертения, была показана чувствительность, позволяющая выявлять даже одну копию ДНК-мишени (Фигура 11С).
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ORION DIAGNOSTICA OY
<120> СПОСОБ АМПЛИФИКАЦИИ ДНК НА ОСНОВЕ ВНЕДРЕНИЯ В ЦЕПЬ
<130> N402749WO
<150> GB 1410022.6
<151> 2014-06-05
<160> 31
<170> PatentIn, версия 3.5
<210> 1
<211> 61
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ОЛИГОНУКЛЕОТИД ДЛЯ ВНЕДРЕНИЯ
<220>
<221> misc_feature
<222> (48)..(60)
<223> 2'-O-метилРНК
<220>
<221> misc_feature
<222> (61)..(61)
<223> ИНВЕРТИРОВАННЫЙ дТТФ
<400> 1
ttgtccatag actgctcgac ctgatacacg ttatcgtcca tacggatucg ggaucucaua 60
t 61
<210> 2
<211> 57
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ОЛИГОНУКЛЕОТИД ДЛЯ ВНЕДРЕНИЯ
<220>
<221> misc_feature
<222> (44)..(56)
<223> 2'-O-метилРНК
<220>
<221> misc_feature
<222> (57)..(57)
<223> ИНВЕРТИРОВАННЫЙ дТТФ
<400> 2
tcctcctgta cctcgttaca aacaggtgta tttagtacag aagauggauu uaaauat 57
<210> 3
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ПРАЙМЕР
<400> 3
aacaagaagg cgtactcgac c 21
<210> 4
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ПРАЙМЕР
<400> 4
tggggcaaaa tattta 16
<210> 5
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ПРАЙМЕР
<400> 5
aaccaaagtg gagtgttaca a 21
<210> 6
<211> 62
<212> ДНК
<213> Искусственная последовательность
<220>
<223> НЕКОМПЛЕМЕНТАРНЫЙ ОЛИГОНУКЛЕОТИД ДЛЯ ВНЕДРЕНИЯ
<220>
<221> misc_feature
<222> (47)..(61)
<223> 2'-O-метилРНК
<220>
<221> misc_feature
<222> (62)..(62)
<223> ИНВЕРТИРОВАННЫЙ дТТФ
<400> 6
ttgtccatag actgctgaaa aaaccgcatc atttatgata tgcttcuccu cgcauaaucu 60
at 62
<210> 7
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> НЕКОМПЛЕМЕНТАРНЫЙ ПРАЙМЕР
<400> 7
gtgtacagag catttaagat t 21
<210> 8
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ЗОНД
<220>
<221> misc_feature
<222> (8)..(8)
<223> КАРБОКСИТЕТРАМЕТИЛРОДАМИНОВАЯ МЕТКА
<220>
<221> misc_feature
<222> (16)..(16)
<223> МЕТКА-ГАСИТЕЛЬ BLACK HOLE 2
<220>
<221> misc_feature
<222> (27)..(27)
<223> ИНВЕРТИРОВАННЫЙ дТТФ
<400> 8
gtacacatca acugttagct ggggcat 27
<210> 9
<211> 31
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ЗОНД
<220>
<221> misc_feature
<222> (10)..(10)
<223> КАРБОКСИТЕТРАМЕТИЛРОДАМИНОВАЯ МЕТКА
<220>
<221> misc_feature
<222> (20)..(20)
<223> МЕТКА-ГАСИТЕЛЬ BLACK HOLE 2
<220>
<221> misc_feature
<222> (29)..(29)
<223> ИНВЕРТИРОВАННЫЙ дТТФ
<220>
<221> misc_feature
<222> (31)..(31)
<223> ИНВЕРТИРОВАННЫЙ дТТФ
<400> 9
tgatacacgt tatcguccat acggattcgg t 31
<210> 10
<211> 14
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ПРАЙМЕР
<400> 10
aacaagaagg cgta 14
<210> 11
<211> 129
<212> ДНК
<213> Искусственная последовательность
<220>
<223> МАТРИЦА-МИШЕНЬ
<400> 11
aacaagaagg cgtactcgac ctgatacacg ttatcgtcca tacggattcg ggatctcagt 60
acacatcaac tggttacaaa caggtgtatt tagtacagaa gatggattta aatattttgc 120
cccagctaa 129
<210> 12
<211> 143
<212> ДНК
<213> Искусственная последовательность
<220>
<223> МАТРИЦА-МИШЕНЬ
<400> 12
aacaagaagg cgtactcgac ctgatacacg ttatcgtcca tacggattcg ggatctcagt 60
acacatcaac tgttagctgg ggcaaaatat ttaaatccat cttctgtact aaatacacct 120
gtttgtaaca ctccactttg gtt 143
<210> 13
<211> 60
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ОЛИГОНУКЛЕОТИД ДЛЯ ВНЕДРЕНИЯ
<220>
<221> misc_feature
<222> (45)..(59)
<223> 2'-O-метилРНК
<220>
<221> misc_feature
<222> (60)..(60)
<223> ИНВЕРТИРОВАННЫЙ дТТФ
<400> 13
ttgtccatag acacgcaaat gaaagacgtt cttaacaagt ttcagaaaga uuugauggct 60
<210> 14
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ПРАЙМЕР
<400> 14
gctttaaccg aacagcaaat 20
<210> 15
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ПРАЙМЕР
<400> 15
gcagtacgct tagccatca 19
<210> 16
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> МАТРИЦА-МИШЕНЬ
<220>
<221> misc_feature
<222> (5)..(63)
<223> АДАПТЕРНАЯ ОБЛАСТЬ
<220>
<221> misc_feature
<222> (264)..(320)
<223> АДАПТЕРНАЯ ОБЛАСТЬ
<400> 16
aaaagcttta accgaacagc aaatgaaaga cgttcttaac aagtttcaga aagatttgat 60
ggctgtctgt gcttctgtgg tgccgagcat tagccccagg aactgcacct cctcctgtct 120
cacggggctg atggtgggtc cagcatctag gagagtgtga cacagggtgg ttggtgctct 180
ggcgctgatt tgagttctca gatctctgag accccagcag aggctgcacg gtaccccaaa 240
tccgcacagc cattgacggg atggcaaatg aaagacgttc ttaacaagtt tcagaaagat 300
ttgatggcta agcgtactgc aaaa 324
<210> 17
<211> 191
<212> ДНК
<213> Искусственная последовательность
<220>
<223> МАТРИЦА-МИШЕНЬ
<220>
<221> misc_feature
<222> (5)..(60)
<223> АДАПТЕРНАЯ ОБЛАСТЬ
<220>
<221> misc_feature
<222> (136)..(187)
<223> АДАПТЕРНАЯ ОБЛАСТЬ
<400> 17
tcagaaccaa agtggagtgt tacaaacagg tgtatttagt acagaagatg gatttaaata 60
acacatacat aacagacctt tatcttatat cccctcggca accctgaagc tgcaatgaat 120
tgatatctgt cgttcgttac aaacaggtgt atttagtaca gaagatggat ttaaatattt 180
tgccccatca g 191
<210> 18
<211> 195
<212> ДНК
<213> Искусственная последовательность
<220>
<223> МАТРИЦА-МИШЕНЬ
<220>
<221> misc_feature
<222> (5)..(60)
<223> АДАПТЕРНАЯ ОБЛАСТЬ
<400> 18
aaaaaaccaa agtggagtgt tacaaacagg tgtatttagt acagaagatg gatttaaata 60
cacccaaggc ctcacactcg atactgtttt gagcgcctct gtcatggttg atatggtcat 120
cagggccaag tggaatattt aaatccatct tctgtactaa atacacctgt ttgtaacact 180
ccactttggt taaaa 195
<210> 19
<211> 197
<212> ДНК
<213> Искусственная последовательность
<220>
<223> МАТРИЦА-МИШЕНЬ
<400> 19
tcaggcagta cgcttagcca tcaaatcttt ctgaaacttg ttaagaacgt ctttcatttg 60
cccggcaaat tgagtcacgg tcggtaacac tttcagcaaa gtccggattt gaagcgtaac 120
ccgcattaaa agaaatgcaa atgaaagacg ttcttaacaa gtttcagaaa gatttgatgg 180
ctaagcgtac tgctcag 197
<210> 20
<211> 140
<212> ДНК
<213> Искусственная последовательность
<220>
<223> МАТРИЦА-МИШЕНЬ
<400> 20
aagaaggcgt actcgacctg atacacgtta tcgtccatac ggattcggga tctcagtaca 60
catcaactgt tagctggggc aaaatattta aatccatctt ctgtactaaa tacacctgtt 120
tgtaacactc cactttggtt 140
<210> 21
<211> 56
<212> ДНК
<213> Искусственная последовательность
<220>
<223> МЕЧЕНЫЙ ОЛИГОНУКЛЕОТИД ДЛЯ ВНЕДРЕНИЯ
<220>
<221> misc_feature
<222> (13)..(13)
<223> МЕТКА-ГАСИТЕЛЬ BLACK HOLE 2
<220>
<221> misc_feature
<222> (44)..(56)
<223> 2'-O-метилРНК
<400> 21
tcctcctgta cctcgttaca aacaggtgta tttagtacag aagauggauu uaaaua 56
<210> 22
<211> 56
<212> ДНК
<213> Искусственная последовательность
<220>
<223> МЕЧЕНЫЙ ОЛИГОНУКЛЕОТИД ДЛЯ ВНЕДРЕНИЯ
<220>
<221> misc_feature
<222> (44)..(56)
<223> 2'-O-метилРНК
<220>
<221> misc_feature
<222> (56)..(56)
<223> МЕТКА-ГАСИТЕЛЬ BLACK HOLE 3
<400> 22
tcctcctgta cctcgttaca aacaggtgta tttagtacag aagauggauu uaaaua 56
<210> 23
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> МЕЧЕНЫЙ ПРАЙМЕР
<220>
<221> misc_feature
<222> (14)..(14)
<223> 6-КАРБОКСИ-Х-РОДАМИНОВАЯ МЕТКА
<400> 23
aaccaaagtg gagtgttaca a 21
<210> 24
<211> 16
<212> ДНК
<213> Искусственная последовательность
<220>
<223> МЕЧЕНЫЙ ПРАЙМЕР
<220>
<221> misc_feature
<222> (11)..(11)
<223> Cy5 TAG
<400> 24
tggggcaaaa tattta 16
<210> 25
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> ПРАЙМЕР
<400> 25
agttgatgtg tactgagatc 20
<210> 26
<211> 151
<212> ДНК
<213> Искусственная последовательность
<220>
<223> МАТРИЦА-МИШЕНЬ
<400> 26
cagttgatgt gtactgagat cccgaatccg tatggacgat aacgtgtatc aggtcgagaa 60
aagtacacat caactgttag ctggggcaaa aaagcaaatg aaagacgttc ttaacaagtt 120
tcagaaagat ttgatggcta gcgtactgct g 151
<210> 27
<211> 56
<212> ДНК
<213> Искусственная последовательность
<220>
<223> АДАПТЕР
<400> 27
aaccaaagtg gagtgttaca aacaggtgta tttagtacag aagatggatt taaata 56
<210> 28
<211> 59
<212> ДНК
<213> Искусственная последовательность
<220>
<223> АДАПТЕР
<400> 28
gctttaaccg aacagcaaat gaaagacgtt cttaacaagt ttcagaaaga tttgatggc 59
<210> 29
<211> 57
<212> ДНК
<213> Искусственная последовательность
<220>
<223> АДАПТЕР
<400> 29
gcagtacgct tagccatcaa atctttctga aacttgttaa gaacgtcttt catttgc 57
<210> 30
<211> 56
<212> ДНК
<213> Искусственная последовательность
<220>
<223> АДАПТЕР
<400> 30
aaccaaagtg gagtgttaca aacaggtgta tttagtacag aagatggatt taaata 56
<210> 31
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> АДАПТЕР
<400> 31
tggggcaaaa tatttaaatc catcttctgt actaaataca cctgtttgta ac 52
<---
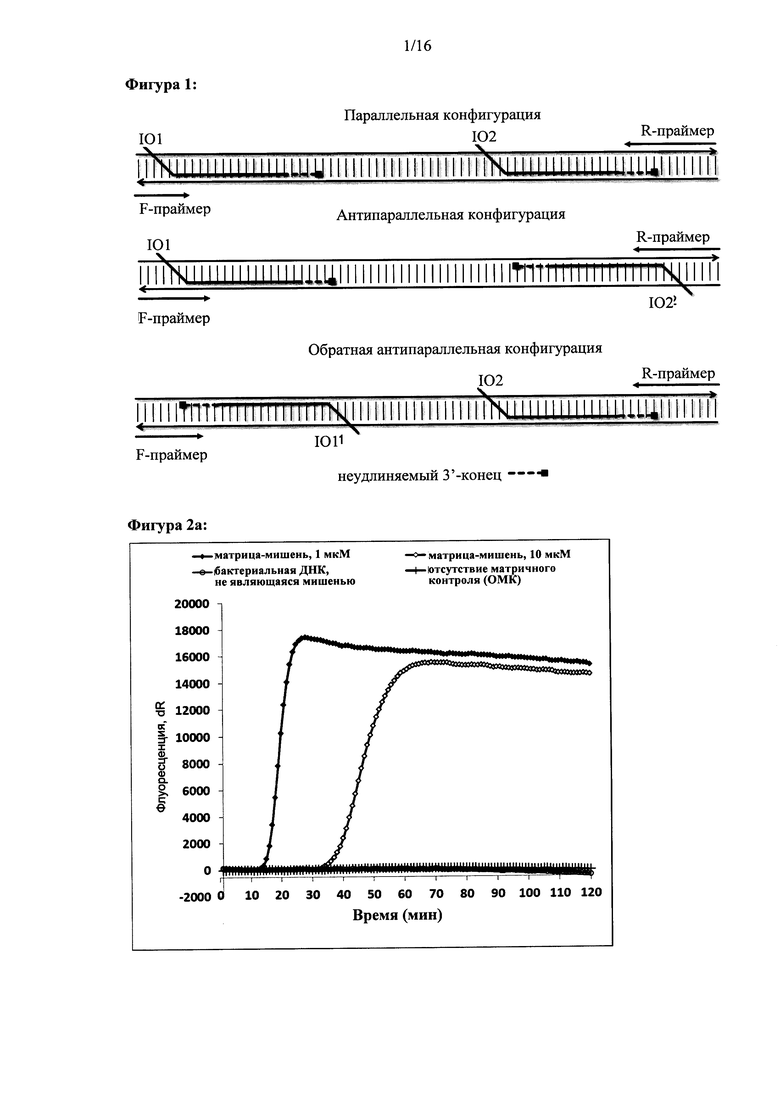


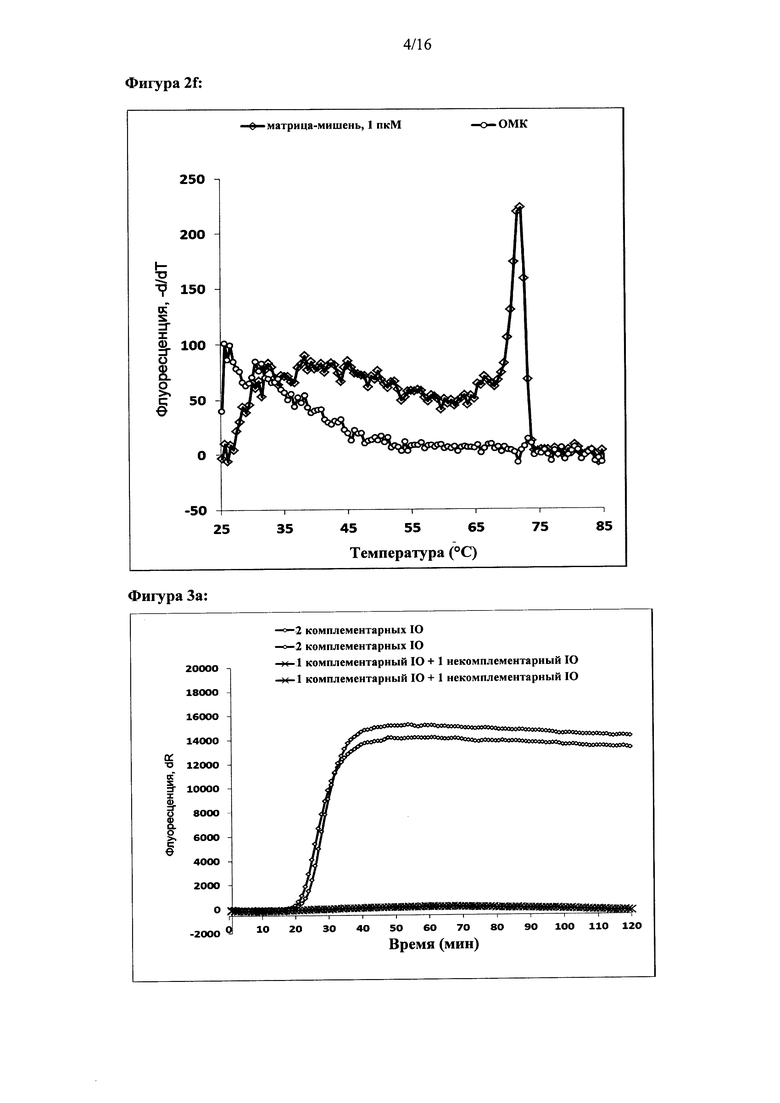
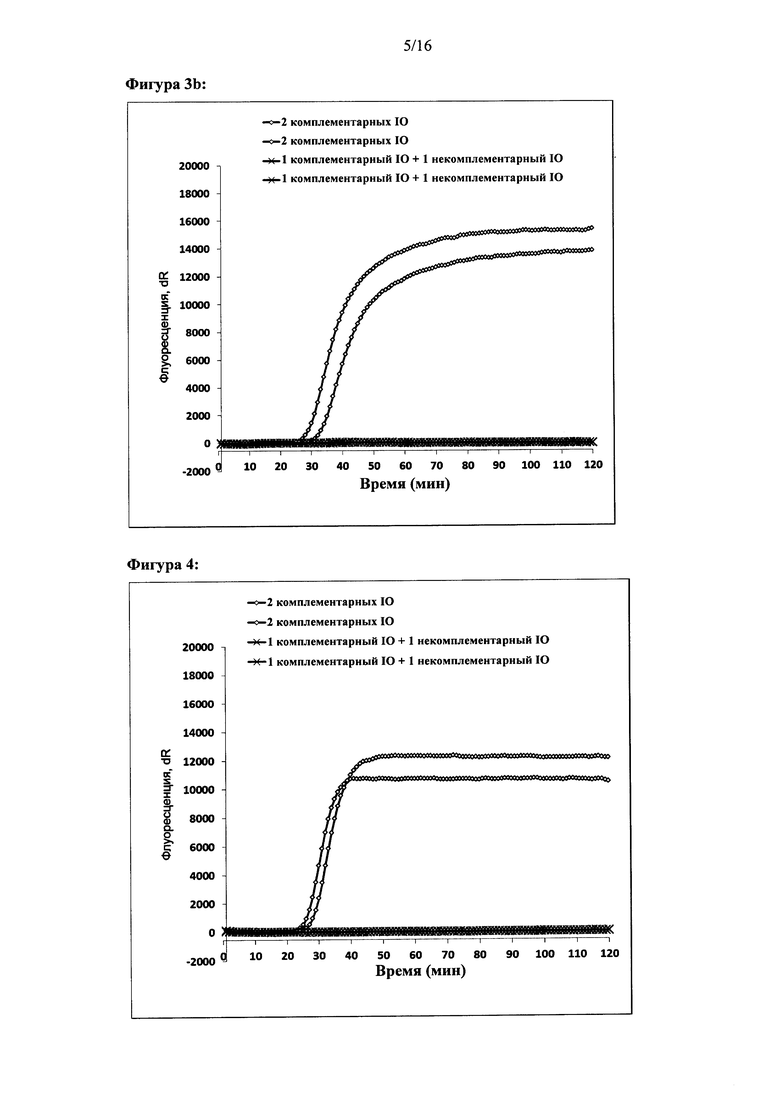

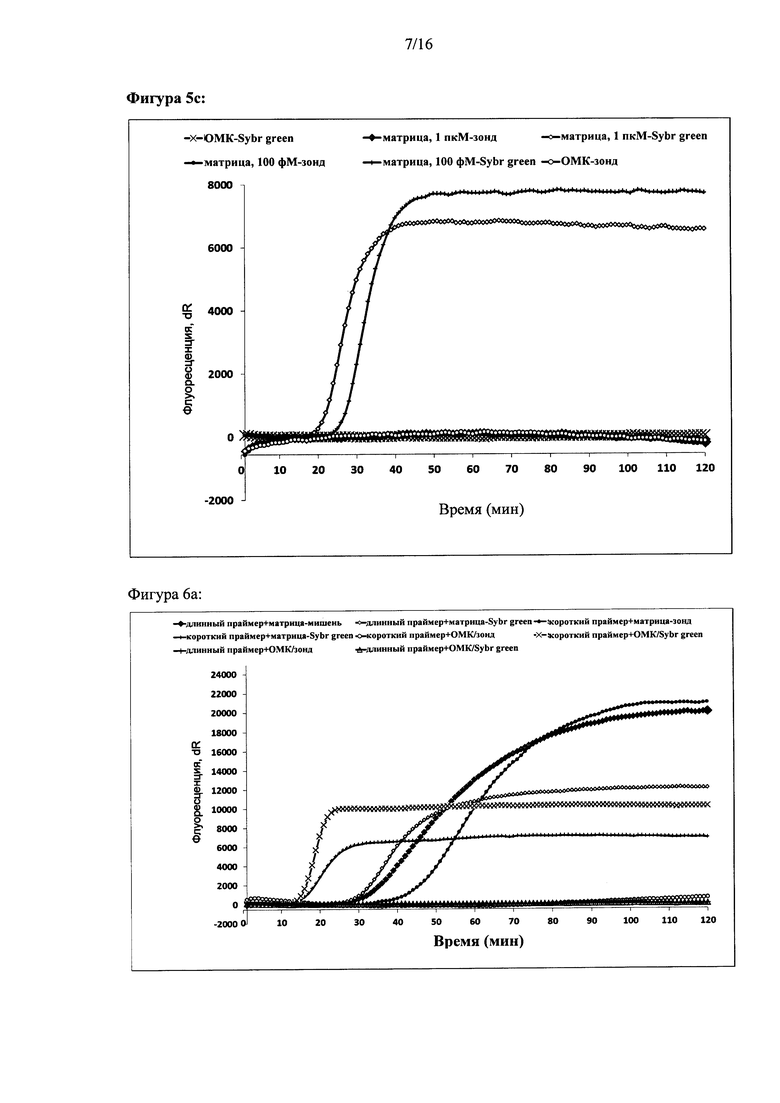

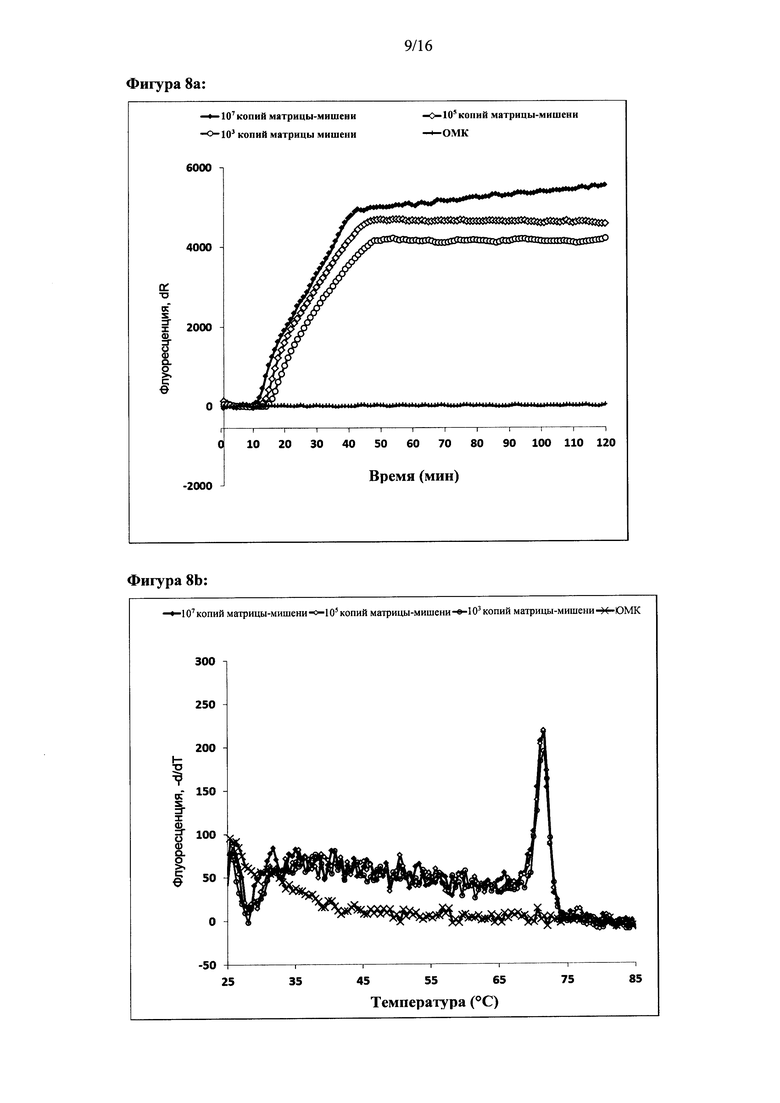
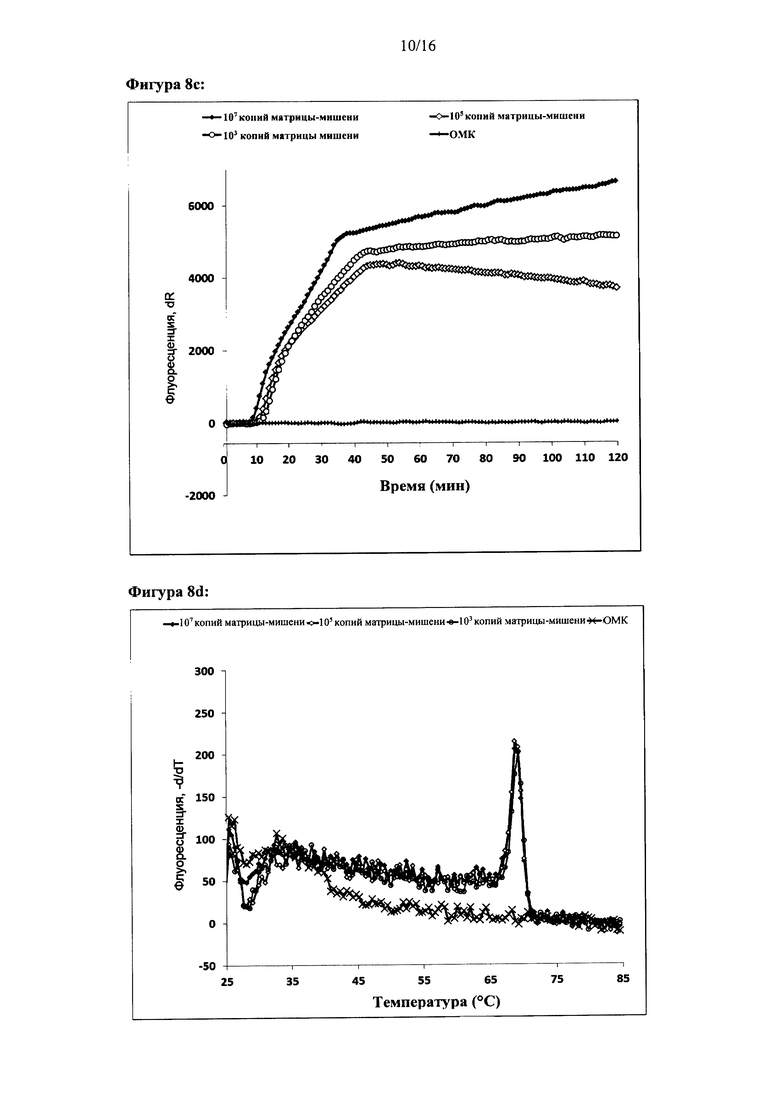
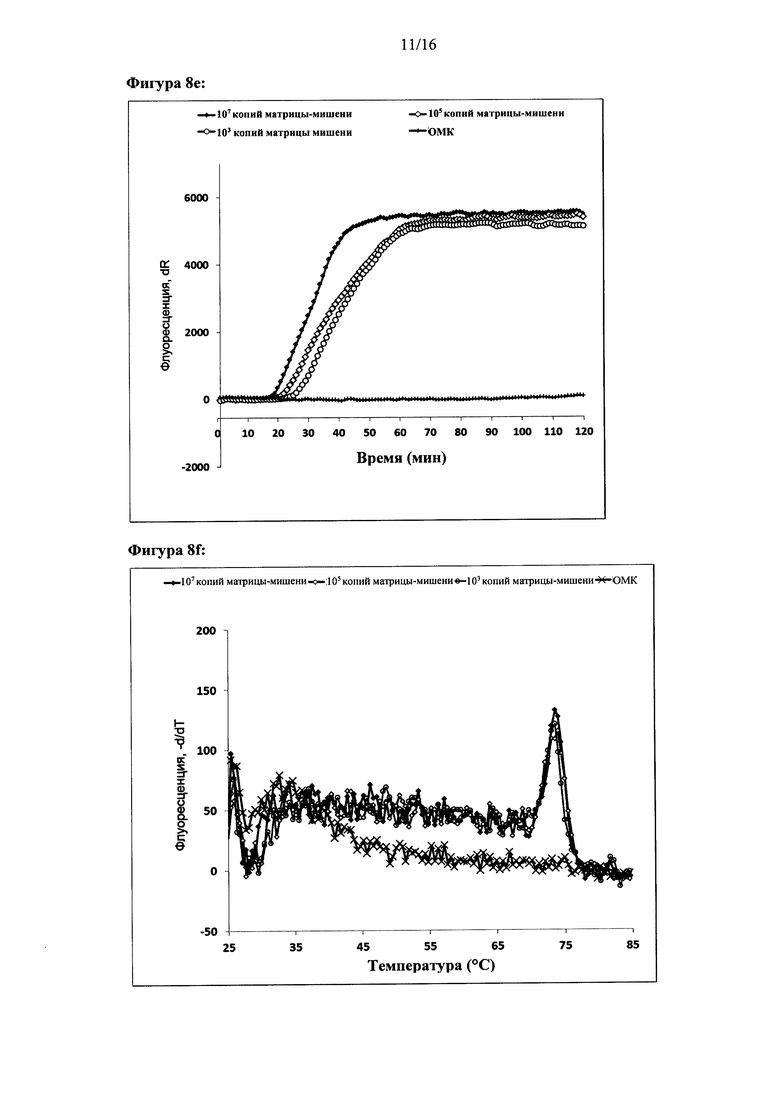

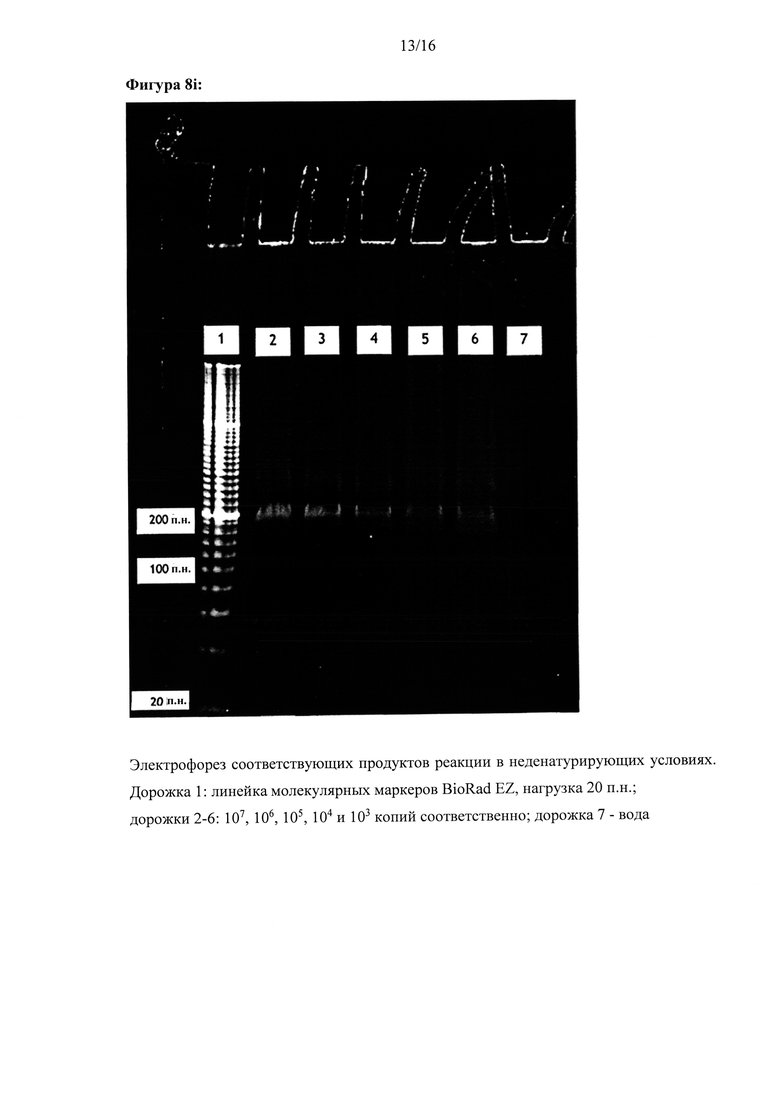
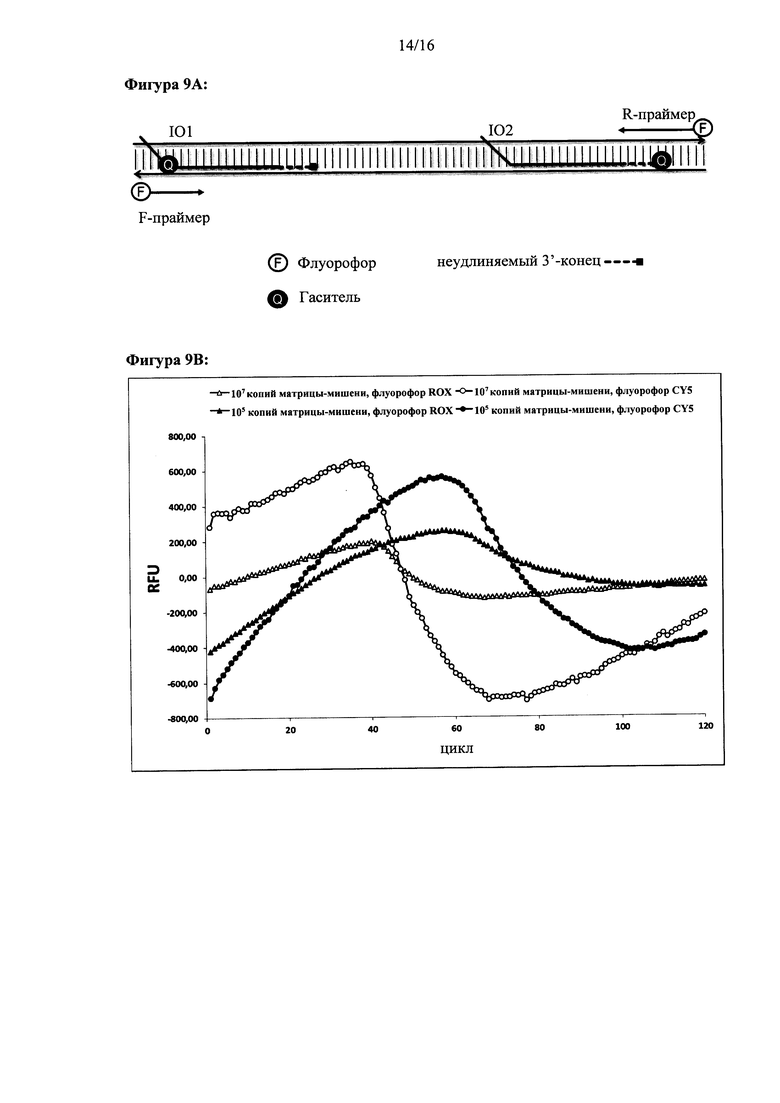
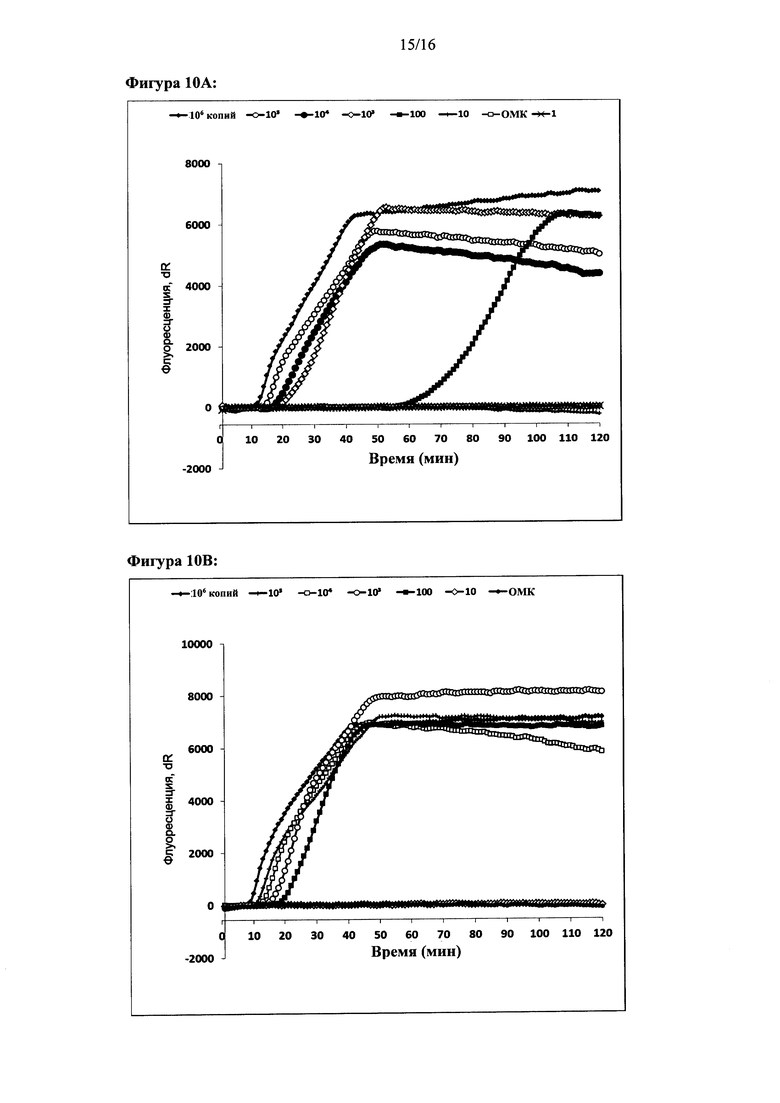
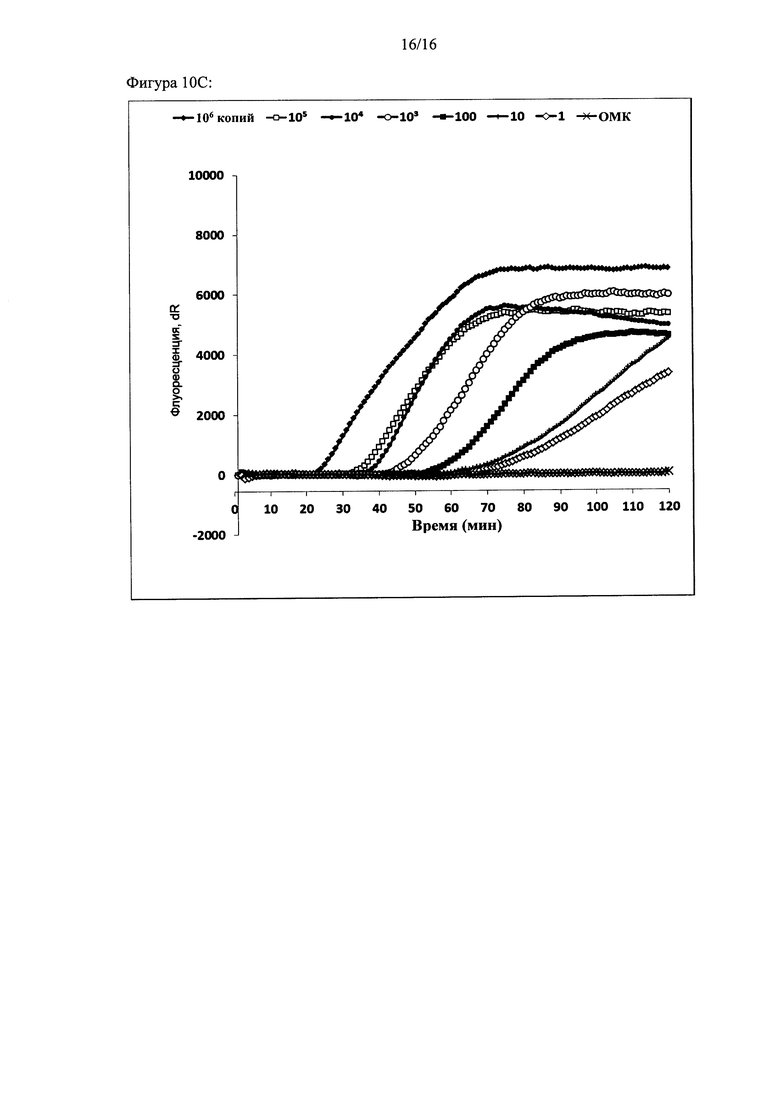
Изобретение относится к области биотехнологии. Описана группа изобретений, включающая способ амплификации последовательности нуклеиновой кислоты-мишени с неизвестной последовательностью (варианты), способ определения последовательности нуклеиновой кислоты-мишени, содержащей область с неизвестной последовательностью, и набор для изотермической амплификации нуклеиновой кислоты-мишени с неизвестной последовательностью. В одном из вариантов реализации способ включает приведение последовательности нуклеиновой кислоты-мишени в контакт с олигонуклеотидом для внедрения в цепь, который имеет неудлиняемый 3’-конец, и по меньшей мере одним праймером, способными обеспечивать амплификацию последовательности нуклеиновой кислоты-мишени, при этом олигонуклеотид для внедрения в цепь связывается с адаптерными последовательностями, чтобы сделать участки связывания олигонуклеотидов для внедрения в цепь, располагающиеся в 3’-5’ и 5’-3’ направлении, в последовательности нуклеиновой кислоты-мишени одноцепочечными для обеспечения связывания по меньшей мере одного праймера. Изобретение расширяет арсенал средств для амплификации последовательностей нуклеиновых кислот-мишеней с неизвестной последовательностью. 4 н. и 18 з.п. ф-лы, 30 ил., 9 пр.
1. Способ амплификации последовательности нуклеиновой кислоты-мишени с неизвестной последовательностью, при этом указанная последовательность нуклеиновой кислоты фланкирована участками связывания по меньшей мере одного олигонуклеотида для внедрения в цепь в 3’-5’ и 5’-3’ направлении, при этом каждый из указанных участков связывания, располагающихся в 3’-5’ и 5’-3’ направлении, содержит адаптерную последовательность, при этом указанный способ включает приведение указанной последовательности нуклеиновой кислоты-мишени в контакт с олигонуклеотидом для внедрения в цепь, который имеет неудлиняемый 3’-конец, и по меньшей мере одним праймером, способными обеспечивать амплификацию указанной последовательности нуклеиновой кислоты-мишени, при этом указанный олигонуклеотид для внедрения в цепь связывается с указанными адаптерными последовательностями, чтобы сделать участки связывания олигонуклеотидов для внедрения в цепь, располагающиеся в 3’-5’ и 5’-3’ направлении, в последовательности нуклеиновой кислоты-мишени одноцепочечными для обеспечения связывания указанного по меньшей мере одного праймера, при этом условия, способствующие амплификации указанной последовательности нуклеиновой кислоты-мишени, включают присутствие рекомбиназы, причём указанный способ осуществляют при изотермических условиях.
2. Способ по п. 1, отличающийся тем, что указанные участки связывания олигонуклеотидов для внедрения в цепь, располагающиеся в 3’-5’ и 5’-3’ направлении, присутствуют в одной и той же цепи последовательности нуклеиновой кислоты-мишени.
3. Способ по п. 1, отличающийся тем, что указанные участки связывания олигонуклеотидов для внедрения в цепь, располагающиеся в 3’-5’ и 5’-3’ направлении, располагаются на противоположных цепях последовательности нуклеиновой кислоты-мишени.
4. Способ по п. 3, отличающийся тем, что указанные олигонуклеотиды для внедрения в цепь связаны с противоположными цепями последовательности нуклеиновой кислоты-мишени, при этом их 3’-концы направлены друг к другу или друг от друга.
5. Способ по любому из пп. 1-4, отличающийся тем, что указанный участок связывания олигонуклеотида для внедрения в цепь, располагающийся в 3’-5’ и/или 5’-3’ направлении, в последовательности нуклеиновой кислоты-мишени не перекрывается с участком связывания соответствующего прямого или обратного праймера.
6. Способ по любому из пп. 1-5, включающий приведение указанной последовательности нуклеиновой кислоты-мишени в контакт с по меньшей мере одним дополнительным олигонуклеотидом для внедрения в цепь.
7. Способ по любому из пп. 1-6, отличающийся тем, что олигонуклеотид для внедрения в цепь и праймер, которому указанный олигонуклеотид позволяет связываться с последовательностью нуклеиновой кислоты-мишени, образуют систему резонансного переноса энергии флуоресценции (FRET).
8. Способ по любому из пп. 1-7, включающий приведение указанной последовательности нуклеиновой кислоты-мишени в контакт по меньшей мере с одним прямым и по меньшей мере с одним обратным праймерами, при этом указанный олигонуклеотид для внедрения в цепь делает 3’-5’ и 5’-3’ участки связывания олигонуклеотидов для внедрения в цепь в последовательности нуклеиновой кислоты-мишени одноцепочечными для обеспечения возможности связывания указанных прямого и обратного праймеров.
9. Способ по любому из пп. 1-8, включающий приведение указанной последовательности нуклеиновой кислоты-мишени в контакт с одним видом праймера, который может связываться как с указанным 3’-5’, так и с указанным 5’-3’ участками связывания в последовательности нуклеиновой кислоты-мишени, при этом указанный олигонуклеотид для внедрения в цепь делает 3’-5’ и 5’-3’ участки связывания олигонуклеотидов для внедрения в цепь в последовательности нуклеиновой кислоты-мишени одноцепочечными для обеспечения возможности связывания указанного одного вида праймера с его как 3’-5’, так и 5’-3’ участком связывания.
10. Способ по любому из пп. 1-9, отличающийся тем, что один указанный олигонуклеотид для внедрения в цепь связывается с указанными адаптерными последовательностями как в 3’-5’, так и в 5’-3’ участке связывания в последовательности нуклеиновой кислоты-мишени, делая указанные участки однецепочечными.
11. Способ по любому из пп. 1-8, отличающийся тем, что указанные 3’-5’ и 5’-3’ участки связывания содержат разные адаптерные последовательности.
12. Способ амплификации последовательности нуклеиновой кислоты-мишени, содержащей участок с неизвестной последовательностью, включающий создание последовательности нуклеиновой кислоты-мишени, содержащей участки связывания олигонуклеотида для внедрения в цепь, расположенные в 3’-5’ и 5’-3’ направлении относительно указанного участка с неизвестной последовательностью, при этом каждая из указанных связывающих последовательностей содержит адаптерную последовательность и амплификацию указанной последовательности нуклеиновой кислоты-мишени путем осуществления способа по любому из пп. 1-6.
13. Способ определения последовательности нуклеиновой кислоты-мишени, содержащей область с неизвестной последовательностью, включающий создание последовательности нуклеиновой кислоты-мишени, содержащей участки связывания вызывающих внедрение в цепь олигонуклеотидов, расположенные в 3’-5’ направлении и 5’-3’ направлении относительно указанной области с неизвестной последовательностью, при этом каждая из указанных связывающих последовательностей содержит адаптерную последовательность, амплификацию указанной последовательности нуклеиновой кислоты-мишени путем осуществления способа по любому из пп. 1-6 и определение последовательности указанной области с неизвестной последовательностью.
14. Способ по п. 12 или 13, отличающийся тем, что олигонуклеотид для внедрения в цепь одного вида связывается с указанной ДНК-адаптерной последовательностью как в 3’-5’, так и в 5’-3’ участке связывания.
15. Набор для изотермической амплификации нуклеиновой кислоты-мишени с неизвестной последовательностью, содержащий олигонуклеотид для внедрения в цепь, который имеет неудлиняемый 3’-конец и по меньшей мере один праймер и по меньшей мере один ДНК-адаптер, при этом указанный олигонуклеотид для внедрения в цепь может связываться с указанным ДНК-адаптером, если указанный ДНК-адаптер присутствует в 3’-5’ и 5’-3’ участках связывания последовательности нуклеиновой кислоты-мишени, и при этом указанный по меньшей мере один праймер способен обеспечивать амплификацию указанной последовательности нуклеиновой кислоты-мишени, где указанный набор дополнительно содержит рекомбиназу.
16. Набор для амплификации нуклеиновой кислоты-мишени с неизвестной последовательностью по п. 15, который содержит по меньшей мере один прямой и по меньшей мере один обратный праймеры для указанной последовательности нуклеиновой кислоты-мишени.
17. Набор для амплификации нуклеиновой кислоты-мишени с неизвестной последовательностью по п. 15, содержащий один вид праймера, который может связываться как с 3’-5’, так и с 5’-3’ участком связывания в последовательности нуклеиновой кислоты-мишени.
18. Набор для амплификации нуклеиновой кислоты-мишени с неизвестной последовательностью по любому из пп. 15-17, отличающийся тем, что указанный 3’-5’ и/или 5’-3’ участок связывания олигонуклеотида для внедрения в цепь в указанной последовательности нуклеиновой кислоты-мишени не перекрывается с участком связывания соответствующего прямого или обратного праймера.
19. Набор для амплификации нуклеиновой кислоты-мишени с неизвестной последовательностью по любому из пп. 15-18, отличающийся тем, что олигонуклеотид для внедрения в цепь и праймер, связывающийся с 3’-5’ и/или 5’-3’ участком указанной последовательности нуклеиновой кислоты-мишени, образуют систему FRET.
20. Набор для амплификации нуклеиновой кислоты-мишени с неизвестной последовательностью по любому из пп. 15-19, дополнительно содержащий по меньшей мере одну эндонуклеазу рестрикции.
21. Набор для амплификации нуклеиновой кислоты-мишени с неизвестной последовательностью по любому из пп. 15-20, содержащий по меньшей мере один дополнительный олигонуклеотид для внедрения в цепь.
22. Набор для амплификации нуклеиновой кислоты-мишени с неизвестной последовательностью по любому из пп. 15-21, дополнительно содержащий по меньшей мере одну ДНК-лигазу.
| WO 2009150467 A1, 17.12.2009 | |||
| EP 1598427 A1, 23.11.2005 | |||
| Термостатирующее устройство | 1989 |
|
SU1634963A1 |
| WO 2015185655 A1, 10.12.2015 | |||
| RU 2015150098 A, 29.05.2017 | |||
| Приспособление для очистки от золы жаровых труб | 1926 |
|
SU4271A1 |

