Область техники, к которой относится изобретение
Изобретение относится к композиции, содержащей экстракт рибосомного белка (RPE) и, необязательно, Th1-активирующий адъювант, для получения лекарственного средства для лечения или профилактики паразитарных заболеваний и к ее применению.
Предшествующий уровень техники
Лейшманиоз включает несколько заболеваний, вызываемых внутриклеточными простейшими паразитами, принадлежащими к роду Leishmania, которые преимущественно инфицируют макрофаги различных млекопитающих, включающих человека и собак. В зависимости в значительной степени от видов паразита и иммунокомпетентного статуса человека-хозяина, спектр заболеваний распространяется от самоизлечивающегося кожного лейшманиоза (CL) до летального висцерального лейшманиоза (VL) или висцерального индийского лейшманиоза (индусская черная болезнь) (18). Висцерально-кожный лейшманиоз собак (VCL), вызываемый Leishmania infantum и L. Chagasi, является важным возникающим зоонозом, обнаруженным в странах Средиземноморского бассейна, Ближнего Востока и Латинской Америки (16), причем собаки являются основным резервуаром данных паразитов, играющим центральную роль в передаче человеку посредством феботоминовых мошек (47). Исход инфекции определяется посредством взаимодействий между иммунной системой хозяина и различными видами паразита, хотя патогенез лейшманиоза еще остается неясным и знание о механизмах, вовлеченных в иммунный ответ на Leishmania у людей и собак, еще является ограниченным. Обычно защитный иммунитет связывают с классическим клеточно-опосредованным иммунным ответом, который индуцирует активацию макрофагов посредством цитокинов, производимых Т-клетками, в то время как неизлечимое заболевание связывают с генерацией сильных гуморальных ответов (15, 26). Исследование для разработки вакцин второго поколения, основанных на неочищенных фракциях паразита или основанных на определенных антигенах паразита, было направлено на идентификацию различных поверхностных или секретируемых паразитом молекул, которые были протестированы как кандидаты для производства вакцин в нескольких экспериментальных моделях, с применением различных адъювантов (1, 17, 22, 46, 48, 49, 52, 54). Скрининг библиотек экспрессии с сыворотками инфицированных животных или людей также дал возможность выбора некоторых антигенов как кандидатов для производства вакцин (обзор в (9)). Среди них те, которые первоначально вызывают иммунный ответ Th1-типа у клеток инфицированных мышей или клеток человеческих пациентов, независимо от их клеточного расположения, были вовлечены в генерацию защитных ответов в различных моделях на животных (51, 55, 56). С другой стороны, некоторые из выделенных антигенов представляют собой внутриклеточные консервативные белки, которые преимущественно стимулируют гуморальные ответы у людей или собак, страдающих VL, или Th2-опосредованные гуморальные ответы у мышей, инфицированных в порядке эксперимента (3, 36, 38, 40, 42). Полагают, что неадекватный гуморальный ответ, индуцированный против них у собак, страдающих лейшманиозом, приводит к иммунопатологии, преимущественно обусловленной вредными эффектами иммунных комплексов, в частности увеиту (13), повреждениям центральной нервной системы (14) или нефриту (23, 24, 33, 34). Также недавно было показано, что присутствие иммунных комплексов IgG у людей с VL коррелирует с неспособностью к противодействию инфекциям, демонстрируя, что иммунные комплексы могут наносить вред инфицированному хозяину (30).
Несмотря на то, что они не считались вначале хорошими кандидатами для производства вакцин, белки, которые индуцируют высокие гуморальные ответы во время инфекционного процесса, ассоциировались с индукцией защитных ответов. Например, паразитарные тубулины и гистон Н2В распознавались клонами Т-клеток, полученных от иммунного донора (39) и rK39, служит причиной пролиферации и выработки IFN-γ Т-клетками иммунной мыши (25). Также было показано, что генетическая иммунизация генами паразита H2B, H3 и H4 вызывает защиту в мышиных моделях висцерального лейшманиоза (27). Также иммунизация рецепторами к активированной С киназе (LACK) (32), некоторым паразитарным цистеинпротеиназам (38, 41) или паразитарным гистонам, образующим гистоны (11, 20), вводимая с Th1-активирующими адъювантами, вызывает иммунный ответ, который коррелирует с защитой против кожного лейшманиоза на мышиных моделях.
В числе эволюционно консервативных антигенов Leishmania, некоторые очевидные линии позволяют предположить, что рибосомные белки являются иммунологически значимыми молекулами во время заражения Leishmania. В некоторых случаях рибосомные составляющие могут способствовать нарушению функционирования иммунной системы хозяина через их способность модулировать клеточную активность и высвобождение цитокинов во время инфекции. Так, инъекция рибосомного белка S3a L. major мышам линии BALB/c вызывала поликлональную экспансию клонов В-клеток и ингибировала пролиферацию Т-клеток (10). Также генетическая иммунизация ДНК вакциной, кодирующей предполагаемый 60S рибосомный белок L31, обостряет заболевание в модели на мышах посредством индукции IL-10 и Th2 цитокинов (44, 53). В дополнение, некоторые паразитарные рибосомные белки, подобные паразитарным кислым P белкам, связывали с генерацией сильных гуморальных ответов у собак и людей, страдающих лейшманиозом (рассмотрено в (42)).
Несмотря на продолжающиеся попытки, до сих пор еще отсутствуют полезные вакцины против паразитарных заболеваний, таких как лейшманиоз. Поэтому до сих пор имеется необходимость в такой вакцине.
Описание изобретения
В данной работе авторы изобретения показывают, что RPE, в особенности RPE Leishmania (LRPE), представляет собой цель для иммунного ответа у собак, естественно инфицированных L. Infantum, и мышей, экспериментально инфицированных L. major. Далее авторы демонстрируют, что сильный защитный Th1-опосредованный иммунный ответ индуцируется, когда LRPE вводят совместно с Th1-активирующим адъювантом, таким как CpG олигодезоксинуклеотид (ODN). Такие композиции (LRPE, в сочетании с Th1-активирующим адъювантом) являются очень привлекательными для применения в качестве вакцины. Изобретение далее описано ниже.
Применение
В первом аспекте изобретения предоставлено применение RPE и необязательно Th1-активирующего адъюванта для получения лекарственного средства для лечения или профилактики паразитарного заболевания у субъекта.
В предпочтительном варианте осуществления RPE получают посредством выполнения следующих стадий с использованием клеток паразита, служащего причиной паразитарного заболевания, когда он представлен у субъекта:
a. смешивание клеток паразита с лизирующим буфером,
b. центрифугирование полученной смеси для получения цитозольного экстракта,
c. получение RPE из полученного цитозольного экстракта.
На стадии a под паразитом предпочтительно подразумевается простейшее. Предпочтительные паразиты определяются здесь далее. Более предпочтительно простейшее находится в промастиготной стадии развития. Специалистам в данной области техники будет известно количество клеток паразита, приблизительно необходимое для получения желательного количества RPE. Типично, для получения приблизительно 500 микрограммов RPE будет использовано приблизительно 3×109 клеток паразита. Лизирующий буфер представляет собой буфер, который будет разрушать по меньшей мере некоторую часть клеток паразита. Предпочтительный лизирующий буфер содержит неионогенное поверхностно-активное вещество. Хорошие результаты были получены с Nonidet P 40 (NP40) в качестве неионогенного поверхностно-активного вещества. Однако может применяться другое неионогенное поверхностно-активное вещество. Предпочтительным применяемым лизирующим буфером раствором является следующий (Буфер A): 10 мМ Трис HCl, pH 8,0, 150 мМ NaCl, 1,5 мМ MgCl2 и 0,5% NP40 (Roche) и предпочтительно дополненный ингибиторами протеаз, такими как PMSF 1 мМ, лейпептин 8 мкг/мл, апротинин 4 мкг/мл и пентатин 8 мкг/мл. Подходящее количество паразитарных клеток (приблизительно 109 клеток/мл буфера А) обычно осторожно смешивают с этим лизирующим буфером, используя пипетку Эппендорфа.
На стадии b по меньшей мере одну стадию центрифугирования при 4°C проводят с полученной смесью стадии а. Обычно первую стадию центрифугирования осуществляют при приблизительно 3000 g в течение приблизительно 2 минут. Полученный супернатант предпочтительно снова центрифугируют при приблизительно 13000 g в течение приблизительно 15 минут при 4°C однократно или дважды.
На стадии с полученный супернатант применяют для получения RPE, как описано в (45). Кратко, полученный супернатант подвергают центрифугированию с высокой скоростью при приблизительно 90000 об/мин в течение приблизительно 30 мин при 4°C в роторе Beckman TL 100.3. Полученный осадок после центрифугирования представляет собой неочищенный рибосомный осадок, который ресуспендируют в буфере B (20 мМ Tris-HCl, pH 7,4, 500 мМ AcNH4, 100 мМ MgCl2, 5 мМ β-меркаптоэтанола) и центрифугируют со ступенчатым градиентом сахарозы (20/40%) в буфере B при приблизительно 90000 об/мин при 4°C в роторе TLl00.3. Полученный осадок содержит рибосомы. Этот осадок предпочтительно растворяют в PBS (физиологический раствор с фосфатным буфером), обрабатывают ультразвуком и хранят при -70°C.
Рибосомные белки представляют собой хорошо сохраняемые цитозольные белки. Следовательно, RPE, как определено здесь, может быть получен из любого эукариотического организма, будь он растением или животным, будь он из млекопитающих, рептилий, рыб, насекомых или любого другого организма, несущего хромосомы, такого так простейшее. Предпочтительно RPE получают из организма, который находится близко к заболеванию, предпочтительно организма на эволюционном дереве, вызывающего паразитарное заболевание. Следовательно, особенный интерес в качестве источника RPE для применения в лечении паразитарного заболевания представляют простейшие, такие как плазмодии, и в особенности члены трипаносоматидного семейства, более конкретно, различные виды трипаносомных простейших Leishmania. Существует более 20 известных видов Leishmania, включая виды подрода Leishmania, включающие комплекс L. major, включая L. major, комплекс L. Donovani, включая L. chagasi, L. donovani и L. infantum, комплекс L. Mexicana, включая L. amazonensis и L. mexicana, а также подвиды Viannia, включающие комплекс L. braziliensis, включая L. braziliensis и L. peruviana и комплекс L. guyanensis, включая L. guyanensis и L. panamensis. Видами Plasmodium, представляющих особенный интерес, являются Plasmodium falciparum и Plasmodium vivax. В предпочтительном варианте осуществления RPE получают из видов Leishmania, предпочтительно Leishmania major и/или Leishmania infantum. В еще одном предпочтительном варианте осуществления RPE получают из видов Plasmodium. Квалифицированный специалист поймет, что RPE может также быть получен посредством смешивания RPE из нескольких различных организмов, как идентифицировано здесь. Применение RPE в вакцине вместо использования данного белка является весьма привлекательным, поскольку RPE содержит большое количество различных антигенов. Каждый из этих антигенов может потенциально вызывать иммунный защитный ответ у обрабатываемого субъекта. Более того, существуют субъекты, которые отвечают на антиген А, а не на В и наоборот. Следовательно, вакцина, как определена здесь, предназначена для применения на широкой популяции субъектов, так как она содержит большое число различных антигенов. В предпочтительном варианте осуществления RPE включает в себя по меньшей мере один рибосомный белок, и/или по меньшей мере один антиген рибосомного белка, и/или по меньшей мере один белковый фрагмент рибосомного белка. В более предпочтительном варианте осуществления RPE включает в себя по меньшей мере два рибосомных белка, и/или по меньшей мере два антигена рибосомного белка, и/или по меньшей мере два белковых фрагмента рибосомного белка. Белковый фрагмент, как определен здесь, представляет собой предпочтительно фрагмент, содержащий по меньшей мере 2, 3, 5, 7, 10, 15, 20, 25, 30 или более соседних аминокислот соответствующего рибосомного белка. В варианте осуществления RPE, как определен здесь, не включает в себя или не состоит из кислого рибосомного белка P0 Leishmania infantum и/или рибосомного антигена LbeF4A из Leishmania braziliensis.
Th1-активирующий адъювант (как адъювант, содержащий мотив CpG ODN) определен в литературе (Liu N., et al. (2003), Nature Immunology, 687-693) как адъювант, который способен активировать или запускать Th1 иммунный ответ против данного антигена, когда используется вместе с этим антигеном (здесь RPE), как обнаружено в супернатантах спленоцитов обработанного субъекта при культивировании с антигеном. В качестве контроля активацию или запуск Th1-иммунного ответа оценивают в популяции спленоцитов того же субъекта, который не был обработан антигеном и адъювантом, или с той же популяцией, обработанной только антигеном. Запуск или активация Th1 иммунного ответа предпочтительно определяется посредством индукции IFNγ, как обнаруживают при культивировании спленоцитов обработанного субъекта с антигеном и/или посредством индуцирования выработки антигенспецифичных IgG2a иммуноглобулинов. Оценку индукции этого цитокина предпочтительно осуществляют посредством ELISA на спленоцитах, как описано в примере. Оценку индукции IgG2a предпочтительно осуществляют посредством ELISA или вестерн-блоттинга, как описано в примере. Индукция IFNγ и/или IgG2a при стимуляции спленоцитов RPE и адъювантом предпочтительно означает, что адъювант квалифицируют в качестве Th1-активирующего адъюванта. Альтернативно или в сочетании с первым определением запуска или активации Th1 иммунного ответа, данным выше, запуск или активация Th1 иммунного ответа может дополнительно определяться по отсутствию (или отсутствию индукции) Th2 иммунного ответа. Th2 иммунный ответ характеризуется по обнаруживаемому увеличению индукции IL-4, IL-10 и/или выработки обнаруживаемых IgG1 иммуноглобулинов при сравнении с необработанными спленоцитами. Оценку индукции IL-4 и/или IL-10 предпочтительно осуществляют посредством ELISA на спленоцитах, как описано в примере. Оценку индукции IgG1 предпочтительно осуществляют посредством ELISA или вестерн-блоттинга, как описано в примере.
Альтернативно или в сочетании с двумя первыми определениями запуска или активации Th1 иммунного ответа, данными выше, запуск или активация Th1 иммунного ответа могут дополнительно быть определены по генерации увеличения отношения IFNγ/IL-10 и/или отношения IFNγ/IL-4 и/или уменьшения отношения IgG1/IgG2a против определенного антигена, в этом случае RPE. В предпочтительном варианте осуществления изменение (увеличение или уменьшение, как указано выше) любого из этих отношений более чем в 2 раза указывает на то, что адъювант обладает свойствами Th1. Оценку индукции каждого из упомянутых цитокинов предпочтительно осуществляют посредством ELISA на спленоцитах, как описано в примере. Оценку индукции иммуноглобулина IgG1 или IgG2a предпочтительно осуществляют посредством ELISA или вестерн-блоттинга, как описано в примере.
В предпочтительном варианте осуществления Th1-активирующий адъювант представляет собой, или содержит, или состоит из олигодезоксинуклеотида. Более предпочтительно олигодезоксинуклеотид (ODN) содержит или состоит из CpG, в котором C представляет собой неметилированный (CpG ODN): 3'пурин-CpG-5'пиримидин. Предпочтительный олигодезоксинуклеотид представляет собой, или содержит, или состоит из фосфортиоат-модифицированной ODN последовательности. Применение олигодезоксинуклеотидов, имеющих такую модификацию, является преимущественным, так как олигодезоксинуклеотиды, используемые таким образом, являются более стабильными, чем немодифицированные олигонуклеотиды, и не будут легко разрушаться, как только они попадут в систему крови. Предпочтительный Th1-активирующий адъювант состоит из или включает по меньшей мере один мотив CpG, по меньшей мере два или по меньшей мере три. Предпочтительными последовательностями иммуностимулирующих ODN (5' до 3') были TCAACGTTGA и GCTAGCGTTAGCGT. Квалифицированный специалист не ограничен последовательностями, явным образом описанными здесь. Он может разработать другие последовательности и далее тестировать их на предмет их Th1-активирующего свойства, как определено здесь ранее. Этот предпочтительный идентифицированный адъювант CpG ODN является очень привлекательным, так как в примере продемонстрировано, что соинокуляция LRPE с этим Th1-активирующим адъювантом индуцирует защиту против стимула паразитами L. major у обеих линий мышей BALB/c и C57BL/6. В обеих моделях защита коррелирует со специфичной выработкой IFN-γ. У BALB/c было также обнаружено ограничение по выработке IL-4 и IL-10.
Одно преимущество настоящего изобретения состоит в том, что оно обеспечивает получение лекарственного средства для лечения широкого спектра паразитарных заболеваний, т.е. лекарственного средства с межвидовой специфичностью. При многих паразитарных заболеваниях вакцина, полученная против конкретных видов, работает только против этих конкретных видов. Один пример паразитарного заболевания, в котором это имеет место, представляет собой лейшманиоз. В настоящее время заболевание контролируют посредством лекарств, но лечение лекарствами не предотвращает распространение заболевания и во многих случаях является не очень эффективным. В предпочтительном варианте осуществления паразитарное заболевание представляет собой лейшманиоз или малярию. Более предпочтительно, паразитарное заболевание вызывается видами Leishmania или Plasmodium. В дополнительном предпочтительном варианте осуществления паразитарное заболевание вызывается различными видами, из которых получают RPE. В частности, лейшманиоз, вызванный одним видом из рода Leishmania, можно лечить посредством применения композиции на основе RPE из еще одного вида Leishmania. В одном варианте осуществления лейшманиоз, вызванный L. Major, успешно лечат композицией, содержащей RPE из L. infantum. Альтернативно, другие паразитарные заболевания, такие как малярия, можно успешно лечить композицией на основе RPE из еще одного вида, например на основе RPE L. infantum.
В контексте изобретения субъект означает человека или животное. Животное, которое охватывается объемом притязаний изобретения, включает млекопитающее, предпочтительно собаку.
В предпочтительном варианте осуществления лекарственное средство, как определено здесь, применяют для увеличения способности иммунной системы человека или животного бороться против инфекции и/или заболевания, более предпочтительно паразитарной инфекции и/или паразитарного заболевания. Конкретно, его можно применять для введения человеческому или животному субъекту. Лекарственное средство, как определено здесь, предпочтительно вводят парентерально, например, посредством инъекции или перфузии посредством внутривенного, подкожного, интраперитонеального, внутримышечного, интраартериального пути или введением внутрь пораженной ткани. Предпочтительным способом введения является подкожный. Лекарственное средство может быть сочетано с фармацевтически приемлемыми средой или системой доставки посредством общепринятых методов, известных в данной области. Например, RPE и необязательно Th1-активирующий адъювант могут быть растворены в физиологическом растворе, забуференном фосфатом (PBS). Способы получения парентерально вводимых композиций хорошо известны в данной области и описаны более подробно в различных источниках, включая, например, Remington's Pharmaceutical Sciences, Ed. AR Gennaro, 20th edition, 2000, Williams & Wilkins, PA, USA. Лекарственное средство предпочтительно вводят в терапевтически эффективной дозе, т.е. такой дозе, которая увеличит способность иммунной системы человека или животного бороться с инфекцией и/или заболеванием, как определено здесь. Предпочтительно терапевтически эффективная доза лекарственного средства изобретения будет предотвращать и/или замедлять развитие кожного повреждения и/или индуцировать значительное снижение паразитарной нагрузки в ухе и/или в дренирующем лимфатическом узле (ДЛУ). Оценка наличия кожного повреждения описана в пояснениях к фигуре 6. Оценка паразитарной нагрузки описана в примере. Терапевтически эффективная доза лекарственного средства изобретения будет предпочтительно предотвращать развитие кожного повреждения и/или будет предпочтительно индуцировать снижение паразитарной нагрузки в ухе приблизительно на 3 порядка величины и/или приблизительно сходной величины в ДЛУ после временного периода, включающего в себя первую одну вакцинацию с использованием композиции изобретения, сопровождаемую одной последующей инфекцией паразитом и временем ожидания, равным приблизительно ±6 недель. В предпочтительном варианте осуществления лекарственное средство, как определено здесь, представляет собой вакцину. В более предпочтительном варианте осуществления в вакцине используют по меньшей мере 12 мкг RPE, применяемого в вакцине. В даже более предпочтительном варианте осуществления должны использоваться по меньшей мере 12-20 мкг RPE для обеспечения иммунного ответа необязательно в сочетании с по меньшей мере 50 мкг Th1-активирующего адъюванта, такого как, например, CpG ODN. Вакцина, как определена здесь, может быть профилактической или терапевтической вакциной. Объем, в котором могут быть растворены RPE и необязательно Th1-активирующий адъювант, может изменяться от 100-500 микролитров.
Композиция
В дополнительном аспекте предоставлена композиция, содержащая RPE и необязательно Th1-активирующий адъювант. RPE и Th1-активирующий адъювант уже были определены в данном документе. В предпочтительном варианте осуществления композиция содержит RPE и Th1-активирующий адъювант. Предпочтительный Th1-активирующий адъювант представляет собой CpG ODN. Предпочтительная композиция содержит или состоит из RPE и необязательно Th1-активирующего адъюванта, растворенных в PBS. В дополнительном предпочтительном варианте осуществления настоящим изобретением также охватывается, что RPE и Th1-активирующий адъювант вводятся последовательно. Следовательно, для обоих компонентов нет необходимости физически присутствовать в одной единственной композиции, поскольку оба они вводятся субъекту. Такая композиция может дополнительно содержать фармацевтически приемлемые адъювант и/или носитель.
Такая композиция предпочтительно предназначена для применения в качестве лекарственного средства. Лекарственное средство, предпочтительно вакцина, уже было подробно определено здесь.
Способ
В еще одном аспекте изобретение предоставляет способ профилактики, и/или лечения паразитарного заболевания, и/или задержки его развития, и/или профилактики, и/или задержки развития кожного повреждения, и/или индукции существенного снижения паразитарной нагрузки в ухе, и/или в дренирующем лимфатическом узле (ДЛУ), все, как определены здесь. В этом способе вакцина изобретения функционирует в качестве терапевтической вакцины. Обычно существует временной период между инфекцией и заболеванием. В этом случае вакцина могла бы действовать как фармакологический иммунный продукт, который предотвращал бы, и/или лечил заболевание, и/или задерживал его развитие посредством вызывания у хозяина иммунного ответа, который противодействует патологическому эффекту инфекции. Терапевтическая вакцина отличается от профилактической вакцины тем, что терапевтическая вакцина будет индуцировать защиту у пациента, который уже имеет инфекцию или заболевание.
В данном документе и в его формуле изобретения глагол "содержать" и его сочетания применяют в его неограничивающем смысле для обозначения того, что элементы, следующие за словом, включены, но элементы, конкретно не упомянутые, не исключены. Дополнительно, глагол "состоять из" может быть замещен выражением "состоять по существу из", означая, что продукт, или композиция, или сохраненная смесь, как определены здесь, могут содержать дополнительные компонент(ы), отличные от конкретно идентифицированных, причем указанные дополнительные компонент(ы) не изменяют уникальные характеристики изобретения.
Дополнительно, ссылки на элемент посредством термина в единственном числе не исключает возможности того, что присутствует более чем один элемент, если контекст ясно не требует, что есть один и только один из элементов. Таким образом, термин в единственном числе обычно означает "по меньшей мере один".
Все патентные и литературные ссылки, цитируемые в настоящем описании, включены в данное описание, таким образом, во всей их полноте.
Изобретение далее иллюстрировано следующим примером, который не следует рассматривать как ограничивающий объем настоящего изобретения.
Описание чертежей
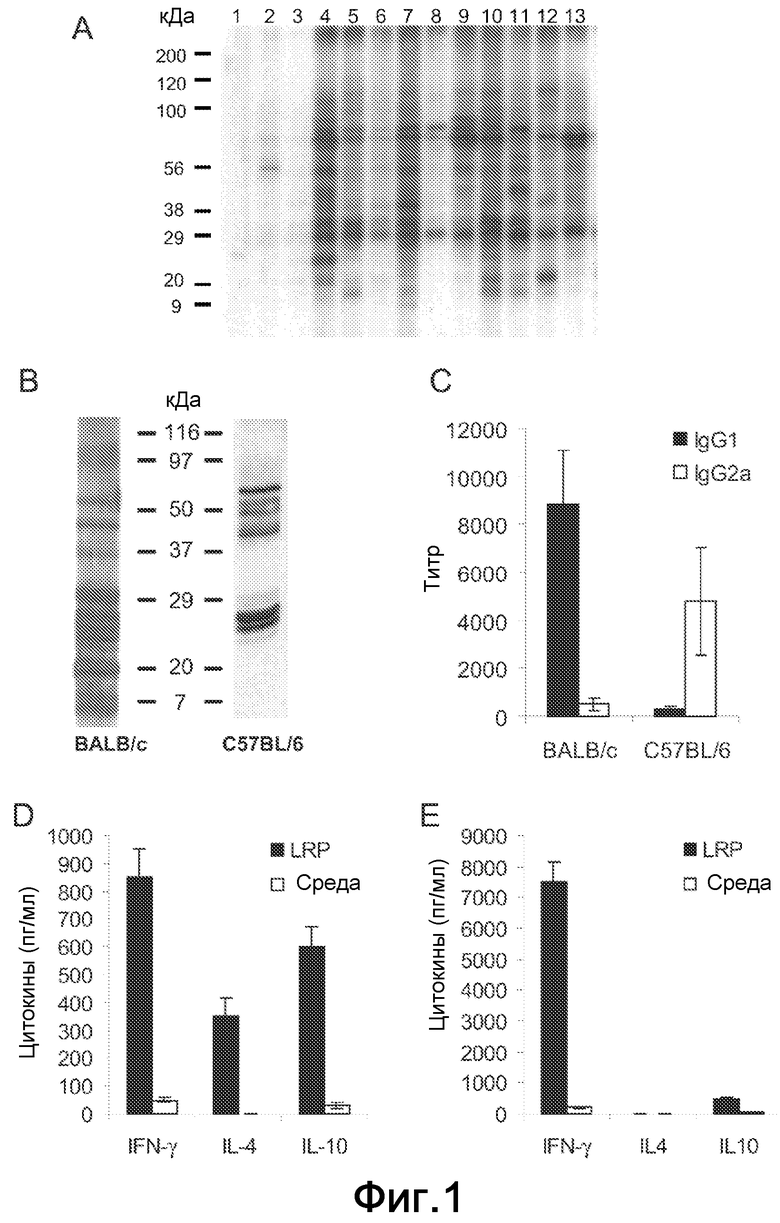
Фиг.1. (A) Рибосомные белки L. infantum подвергали электрофорезу на SDS-PAGE геле с линейным 10-14% градиентом, переносили на нитроцеллюлозные блоты и инкубировали с сывороткой здоровых собак (полосы 1-3) и сывороткой собак, страдающих VCL (полосы 4-13). Сыворотку использовали при разбавлении 1/200. В качестве вторичного реагента применяли пероксидазу хрена, конъюгированную с антителами против собачьих IgG (B-E). Четыре BALB/c мыши были инфицированы под кожу 5×104 промастиготами в стационарной фазе L. major в левую подушечку стопы и сыворотки были получены через восемь недель после стимуляции. Четыре C57BL/6 мыши были внутрикожно инфицированы 300 метациклическими промастиготами L. major в дерму уха, и сыворотки были получены на 14 неделе после стимуляции. Предынфекционные сыворотки были также получены для обеих линий перед стимуляцией паразитами. (B) Рибосомные белки L. major подвергали электрофорезу на SDS-PAGE геле с линейным 10-14% градиентом, переносили на нитроцеллюлозные блоты и инкубировали с вытяжками сывороток из BALB/c или C57BL/6 инфицированных мышей. Сыворотку использовали при разбавлении 1/200. Ни одна из предынфекционных сывороток не показала реакционноспособности против LRP (не показано). (C) Титры для IgG1 и IgG2a антител против LRP в обеих линиях определяли индивидуально посредством ELISA. (D) Через восемь недель после инфицирования BALB/c мышей умерщвляли и их клетки подколенного ДЛУ культивировали in vitro в течение 48 ч в присутствии 12 мкг мл-1 L. major LRP или только в среде. Уровни IFN-γ, IL-4 и IL-10 оценивали посредством ELISA в супернатантах культуры. (E) Через четырнадцать недель после инфекции C57BL/6 мышей умерщвляли и их ретромаксилярные ДЛУ обрабатывали как в D.
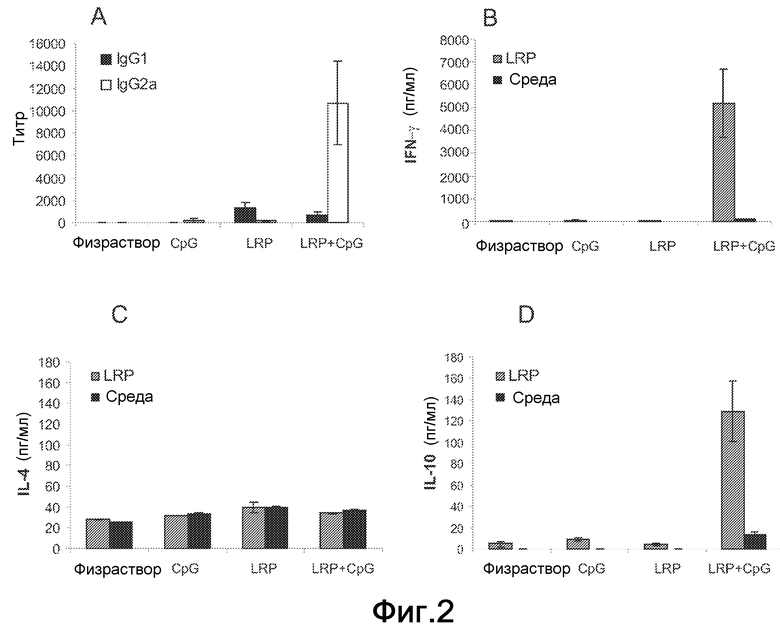
Фиг.2. (A) Анализ специфичного гуморального ответа, индуцированного у BALB/c мышей. BALB/c мышей (шесть на группу) иммунизировали под кожу в правую подушечку стопы тремя дозами рибосомных белков по отдельности (LRP), или с добавкой адъюванта CpG ODN (LRP+CpG), с CpG ODN адъювантом по отдельности (CpG) или с PBS (физраствор). Через четыре недели после третьей иммунизации у мышей забирали кровь и сыворотки оценивали посредством ELISA на предмет специфичных ответов против-LRP антител обоих IgG1 (черные столбцы) и IgG2a (белые столбцы) изотипов. Ни одна из предыммунных сывороток не показала реакционноспособности. (B-D) Через четыре недели после вакцинации мышей умерщвляли, и их селезенки были получены и культивированы in vitro в течение 48 ч в присутствии LRP (серые столбцы) или среды по отдельности (черные столбцы). Уровни IFN-γ (B), IL-4 (C) и IL-10 (D) оценивали посредством ELISA в супернатантах культуры.
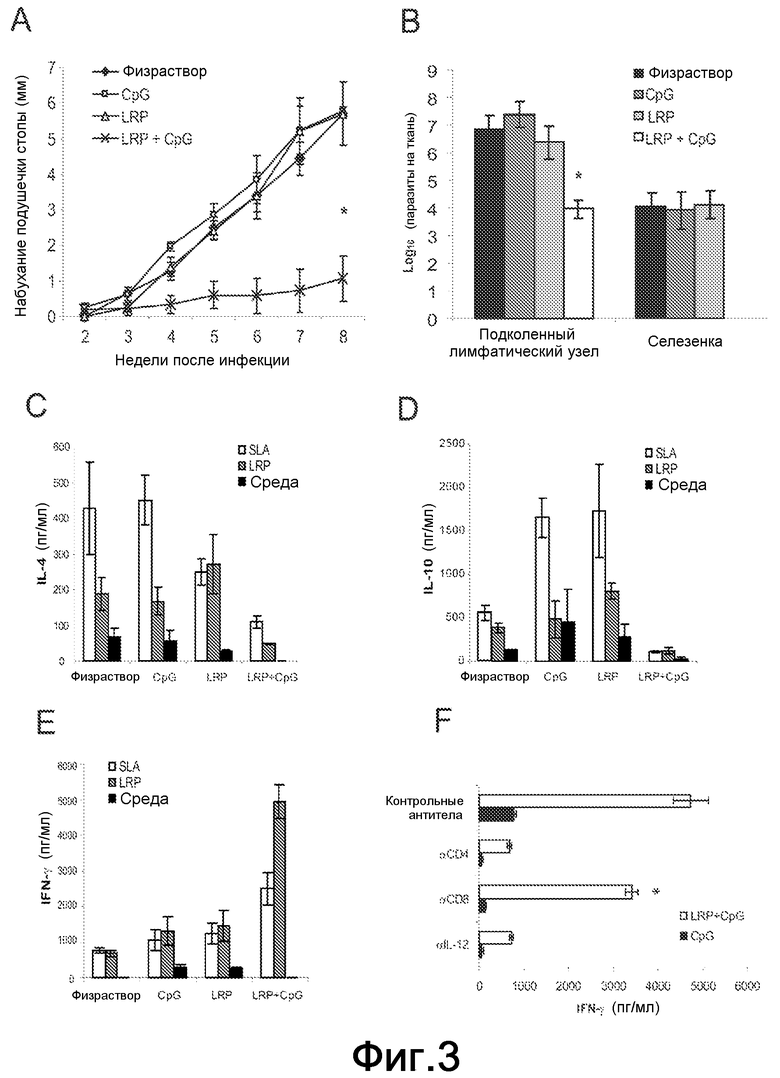
Фиг.3. (A) Протекание инфекции L. major у вакцинированных BALB/c мышей. Мышей (шесть на группу) иммунизировали под кожу, как указано на Фиг.2. Через один месяц после последней иммунизации животных инфицировали в левую заднюю подушечку стопы 5×104 промастиготов L. major в стационарной фазе. Набухание подушечки стопы дано в виде различия в толщине между инфицированной и неинфицированной контралатеральными стопами. Результаты представляют среднее и стандартное отклонение для двух независимых экспериментов. *P<0,001 значимые различия в воспалении для мышей, вакцинированных LRP+CpG ODN, при сравнении с мышами группы CpG ODN на восьмой неделе после стимуляции. (B) На восьмой неделе после инфицирования количество жизнеспособных паразитов в подколенных ДЛУ инфицированных ноги и селезенки индивидуально определяли посредством ограничивающего разбавления. Результаты представляют среднее и стандартное отклонение для двух независимых экспериментов. * P<0,01 значимые различия в подколенной паразитарной нагрузке для мышей, вакцинированных LRP+CpG ODN, по сравнению с мышами группы CpG ODN на неделе 8 после стимуляции. (C-D) Выработку цитокинов у вакцинированных и инфицированных мышей определяли через восемь недель после стимуляции L. major. Были получены подколенные ДЛУ инфицированной ноги и культивированы in vitro в течение 48 ч в присутствии SLA (белые столбцы), LRP (серые столбцы) или среды по отдельности (черные столбцы). Уровни IL-4 (C), IL-10 (D) и IFN-γ (E) в супернатантах культуры тестировали посредством ELISA. Этот эксперимент повторяли со сходным результатом. (F) Анализ включения IL-12 и T клеток в выработку IFNγ, ассоциированную с защитой, придаваемой вакцинацией LRP+CpG ODN. Подколенные ЛУ (лимфатические узлы) LN от мышей, вакцинированных CpG ODN (черные столбцы) и LRP+CpG ODN (белые столбцы), были получены через восемь недель после стимуляции 5×104 промастиготов L. major в стационарной фазе и стимулированы в культуре LRP в присутствии либо против-IL-12, против-CD4 или против-CD8, и контрольных моноклональных антител. Уровни IFN-γ оценивали посредством ELISA после 78 ч инкубации. Различия в выработке IFN-γ между обработкой против-CD8 моноклональными антителами и обработкой контрольными антителами были статистически значимыми (*P<0,05). Данные соответствовали одному представительному эксперименту из двух независимых экспериментов со сходным результатом.
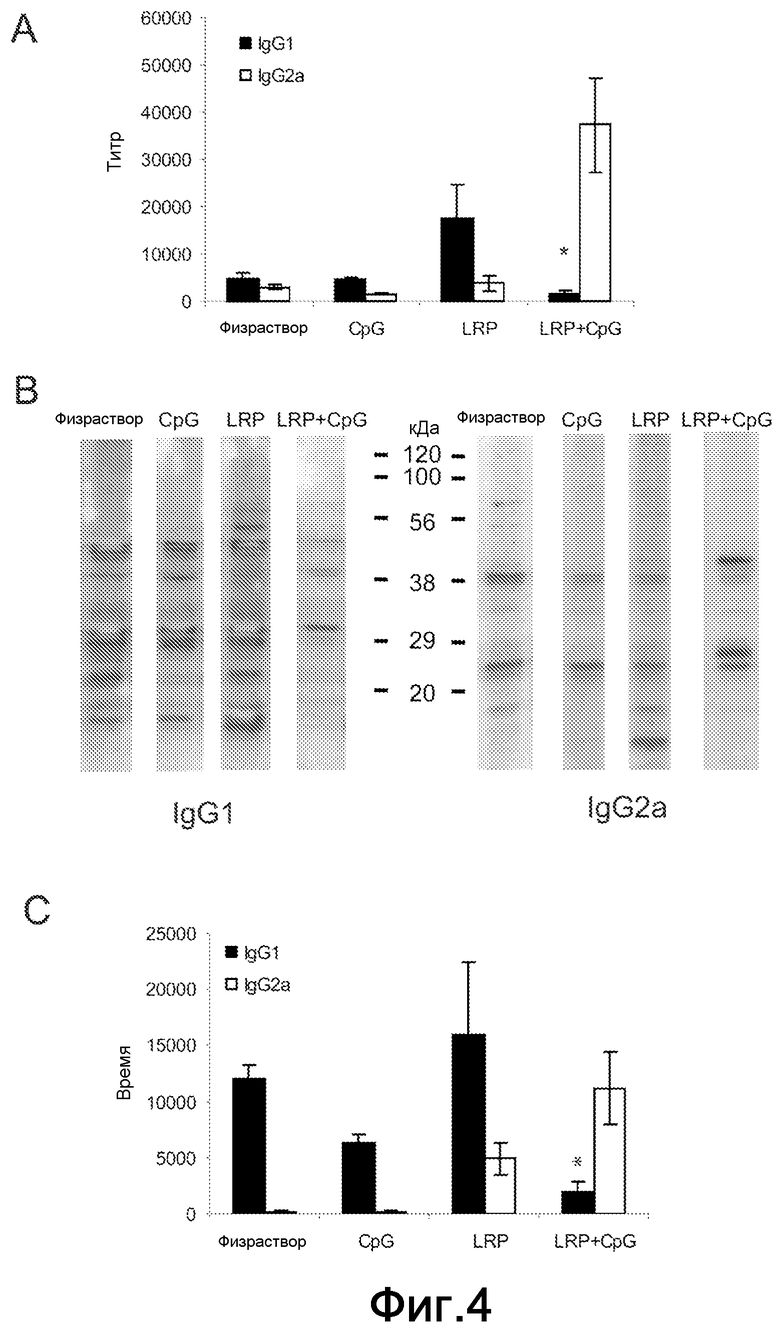
Фиг.4. Анализ поляризации IgG1/IgG2a. (A) Образцы сыворотки были получены через восемь недель после стимуляции и титр для IgG1 и IgG2a антител против LRP определяли индивидуально посредством ELISA. Различия в титре IgG1 между мышами, вакцинированными LRP+CpG ODN, и другими тремя группами были статистически значимыми (*P<0,01). (B) LRP L. major разделяли на SDS-PAGE геле с линейным 10-14% градиентом, переносили на нитроцеллюлозные блоты и инкубировали полученными сыворотками от указанных групп мышей при разбавлении 1/200. Показаны ответы антител обоих изотипов IgG1 и IgG2a. (C) Те же сыворотки использовали для определения титров IgG1 и IgG2a против SLA. Различия в титре IgG1 между мышами, вакцинированными LRP+CpG ODN, и другими тремя группами были статистически значимыми (*P<0,02).
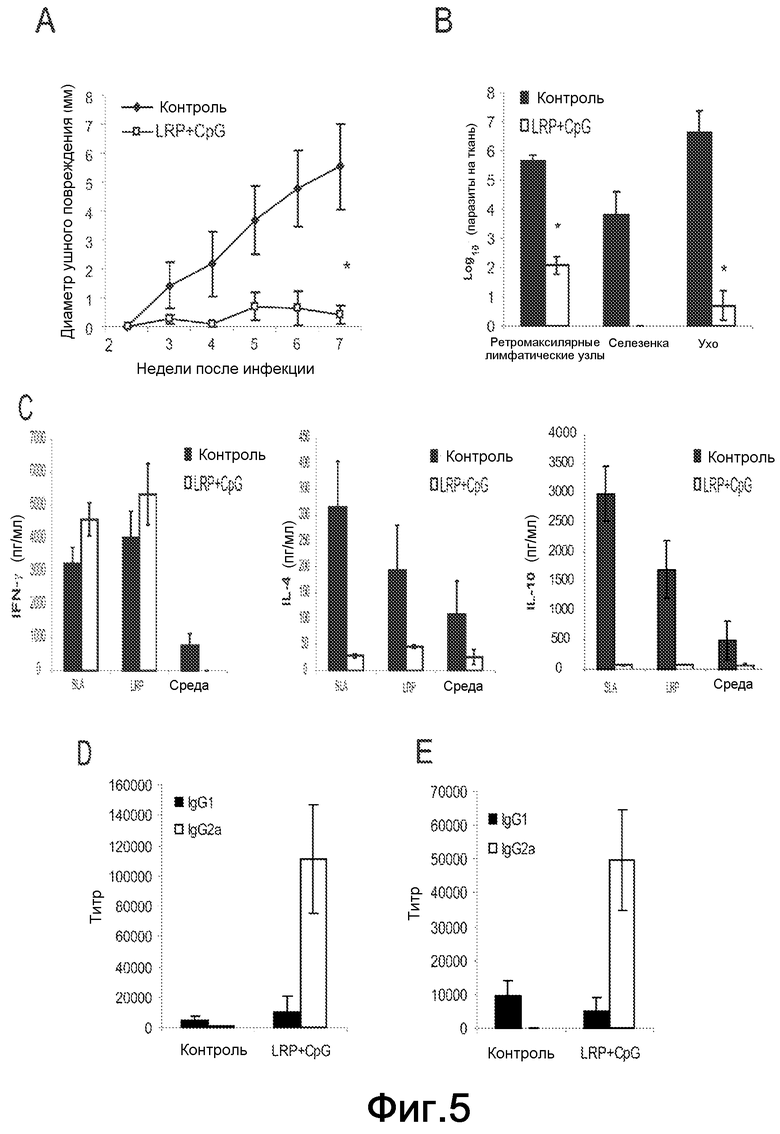
Фиг.5. Шесть BALB/c мышей были вакцинированы LRP+CpG ODN и инфицированы в левую подушечку стопы, как указано на Фиг.3. Через восемнадцать недель после первой стимуляции паразитами мыши были внутрикожно реинфицированы в ухо 300 метациклическими промастиготами L. major. В качестве контроля шесть необработанных мышей были также внутрикожно стимулированы в ухо. (A) Протекание инфекции L. major у защищенных и реинфицированных BALB/c мышей. Значения представляют средний диаметр повреждения + стандартное отклонение (СО). *P<0,0001 значимые различия в воспалении для реинфицированных по сравнению с контрольными инфицированными мышами на неделю 7 после стимуляции. (B) Через семь недель после реинфекции, мышей умерщвляли и паразитарная нагрузка в дерме уха, селезенке и локальном ДЛУ индивидуально количественно оценивалась. Результаты выражены как среднее ±СО для двенадцати ушей и ДЛУ. *P<0,001 значимое снижение для реинфицированных по сравнению с инфицированными контрольными мышами. (C) После умерщвления ретромаксилярные ДЛУ от контрольных (черные столбцы) и реинфицированных мышей (белые столбцы) были получены и культивированы in vitro в течение 48 ч в присутствии SLA, LRP или среды по отдельности. Уровни IFN-γ, IL-4 и IL-10 оценивали посредством ELISA в супернатантах культуры. (D-E) Образцы сыворотки были получены через семь недель после рестимуляции и титр для IgG1 и IgG2a антител против LRP (D) и SLA (E) определяли индивидуально посредством ELISA.
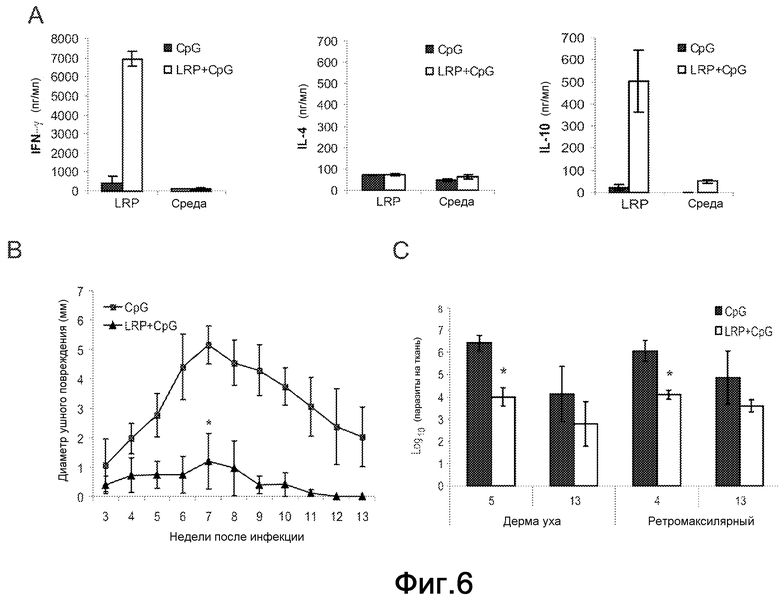
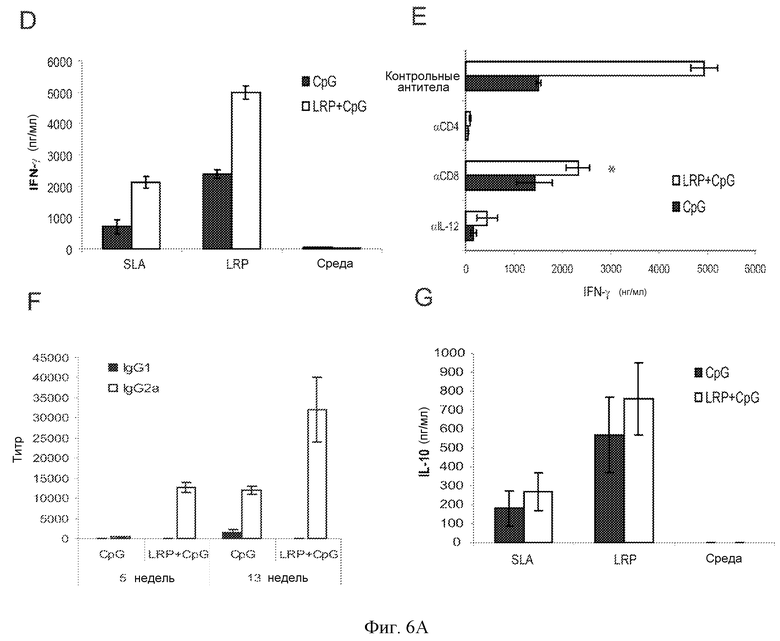
Фиг.6 и 6a. Защита против инфекции L. major у C57BL/6 мышей. Мышей (шесть на группу) подкожно иммунизировали в правую подушечку стопы тремя дозами LRP+CpG ODN и CpG ODN по отдельности. (A) Выработка IFN-γ, IL-4 и IL-10 спленоцитами C57BL76 вакцинированных мышей. Через четыре недели после вакцинации CpG ODN (черные столбцы) или LRP+CpG ODN (белые столбцы) мышей умерщвляли и получали их селезенки, и их культивировали in vitro в течение 48 ч в присутствии LRP или среды по отдельности. Уровень цитокинов оценивали посредством ELISA в супернатантах культур. (B) Протекание инфекции L. major у C57BL/6 вакцинированных мышей. Двенадцать мышей на группу иммунизировали, как описано выше, и через четыре недели после последней инокуляции они были инфицированы посредством внутрикожной инокуляции в ухо 300 метациклическими промастиготами L. major. Значения представляют средний диаметр повреждения + стандартное отклонение (СО). *P<0,001 значимое снижение в воспалении между двумя группами мышей. (C) Паразитарную нагрузку в дерме уха и в местном ДЛУ от мышей, вакцинированных CpG ODN (черные столбцы) или LRP+CpG ODN (белые столбцы), количественно определяли на пятую неделю (шесть мышей на группу) и тринадцатую неделю (шесть мышей на группу) после инфицирования. Результаты выражены как среднее ±СО для двенадцати ушей и ДЛУ. *P<0,01 значимое снижение между двумя группами мышей. (D-E) Выработка IFN-γ у мышей, вакцинированных CpG (черные столбцы) или LRP+CpG ODN (белые столбцы). Ретромаксилярные клетки были получены через 5 недель после инфицирования и стимулированы в культуре в присутствии SLA, LRP и среды по отдельности (D) или LRP в присутствии антител против-IL-12, против-CD4 или против-CD8, и контрольных моноклональных антител. Уровень IFN-γ оценивали посредством ELISA после 78 ч инкубации. Различия в выработке IFN-γ между обработкой моноклональными антителами против-CD8 и обработкой контрольными антителами были статистически значимыми (*P<0,01). (F) Образцы сыворотки были получены на недели 5 и 13 после стимуляции, и титры IgG1 и IgG2a антител против LRP определяли индивидуально посредством ELISA. (G) Выработка IL-10 у мышей, вакцинированных CpG ODN (черные столбцы) или LRP+CpG ODN (белые столбцы). Ретромаксилярные клетки были получены через 5 недель после инфицирования и стимулированы в культуре в присутствии SLA, LRP или среды по отдельности. Уровень IL-10 оценивали посредством ELISA после 78 ч инкубации.
Примеры
МАТЕРИАЛЫ И МЕТОДЫ
Линии мышей и штаммы паразитов
Самки BALB/c мышей имели возраст 6-8 недель к моменту начала экспериментов и были приобретены у Harlan Interfauna Iberica S. A. (Barcelona, Spain). Паразиты L. major (клон WHOM/IR/-173) и клон V1 (MHOM/IL/80 (Friedlin) поддерживали в вирулентном состоянии посредством пассажа у BALB/c мышей. Амастиготы L. major были получены из подколенных дренирующих лимфатических узлов (ДЛУ) и трансформированы в промастиготы посредством культивирования при 26°C в среде Шнейдера (Gibco, BRL), дополненной 20% фетальной телячьей сывороткой (FCS), пока они не достигали поздней стационарной фазы. Промастиготы обоих клонов культивировали при 26°C в среде Шнейдера (Gibco, BRL), дополненной 20% FCS. Промастиготы инфективной стадии (метациклики) L. major (клон V1) выделяли из стационарных культур посредством отрицательной селекции с применением агглютинина арахиса (Vector Laboratories, Burlingame, CA.). Промастиготы L. infantum (MCAN/ES/96/BCN/l50, MON-1) культивировали при 26°C в среде RPMI (Gibco, BRL), дополненной 10% FCS.
CpG ODN и антигены лейшманиоза
Для получения экстрактов рибосомного белка Leishmania (LRP), L. major и L. infantum промастиготы были собраны, промыты дважды в предварительно охлажденной PBS и ресуспендированы в 1 мл NP40 (Roche Diagnostics, GmbH, Manheim Germany, cat. N 11332473001) лизирующем буфере (10 мМ Tris ClH, pH 8,0, 150 мМ NaCl, 1,5 мМ MgCl2 и 0,5% NP40, PMSF 1 мМ, лейпептин 8 мкг/мл, апротинин 4 мкг/мл и пентатин 8 мкг/мл) и пипетировали вверх и вниз 10 раз. После лизиса образцы были микроцентрифугированы при 3000 g в течение 2 мин при 4°C для отделения ядер в осадок. Супернатант был дважды микроцентрифугирован при 13000 g в течение 15 мин при 4°C и рибосомы были получены из цитозольного супернатанта, как описано в (45). Кратко, цитозоль подвергают центрифугированию с высокой скоростью при приблизительно 90000 об/мин в течение 30 мин при 4°C в роторе Beckman TL 100.3. Неочищенный рибосомный осадок ресуспендируют в буфере B (20 мМ Tris-HCl, pH 7,4, 500 мМ AcNH4, 100 мМ MgCL2, 5 мМ β-меркаптоэтанола) и центрифугируют со ступенчатым градиентом сахарозы (20/40%) в буфере B при приблизительно 90000 об/мин при 4°C в роторе TL100.3. Осадок промытых рибосом растворяют в PBS, обрабатывают ультразвуком и хранят при -70°C.
Суммарные белки L. major (растворимый антиген Leishmania [SLA]) получали посредством трех циклов замораживания и оттаивания стационарных промастигот L. major, суспендированных в PBS. После лизиса клеток растворимые антигены отделяли от нерастворимой фракции посредством центрифугирования в течение 15 мин при 12000 g, используя микроцентрифугу, и хранили при -70°C. Фосфортиоат-модифицированные последовательности ODN, содержащие CpG мотивы (CpG ODN), были синтезированы Isogen (The Netherlands). Последовательностями иммуностимулирующих ODN (от 5' до 3') являлись TCAACGTTGA и GCTAGCGTTAGCGT.
Иммунизации и паразитарная стимуляция
BALB/c мыши были подкожно (s.c.) инокулированы в правую подушечку стопы либо 12 мкг L. major LRP по отдельности или с добавлением 50 мкг CpG ODN (25 мкг каждого иммуностимулирующего ODN), CpG ODN (50 мкг) адъюванта по отдельности или забуференного фосфатом физиологического раствора (PBS). Каждую группу активизировали через 2 и 4 недели, используя такой же режим. Первую стимуляцию паразитами осуществляли посредством подкожной инокуляции 5×104 промастиготами L. major в стационарной фазе (клон MHOM/IR/-173) в левую (необработанную) подушечку стопы через четыре недели после последней инокуляции. Развитие инфекции сопровождалось измерением толщины с помощью метрического калибра. Контрлатеральная подушечка стопы каждого животного представляла контрольное значение, и набухание рассчитывали следующим образом: толщина левой подушечки стопы минус толщина правой подушечки стопы. Животных умерщвляли, когда повреждения становились некротическими. Для реинфекции шесть BALB/c мышей были вакцинированы и инфицированы, как описано выше. После восемнадцати недель 300 метациклических промастигот L. major (клон V1) были введены инъекцией в дерму обоих ушей каждой мыши. Эволюцию инфекции регистрировали посредством измерения диаметра уплотнений ушных повреждений с помощью метрического калибра. В качестве контроля группа из шести необработанных BALB/c мышей была также инфицирована в дерму уха.
C57BL/6 мышам инъецировали подкожно в подушечку стопы 12 мкг L. major LRP + 50 мкг CpG ODN (25 мкг каждого иммуностимулирующего ODN), и 50 мкг CpG ODN (50 мкг) адъюванта по отдельности. Этих мышей активизировали через 2 и 4 недели, используя такой же режим иммунизации. Инфицирование осуществляли через 4 недели после последней вакцинации посредством внутрикожной (i.d.) инокуляции 300 метациклических промастигот L. major (клон V1) в дерму обоих ушей мыши. Эволюцию инфекции регистрировали посредством измерения диаметра уплотнений ушных повреждений с помощью метрического калибра.
Количественная оценка паразитов
Количество паразитов определяли в ушах посредством анализа с ограничивающим разбавлением (6). Кратко, уши были извлечены от инфицированных мышей. Вентральные и дорсальные фрагменты инфицированных ушей были отделены. Фрагменты ушей помещали в модифицированную по Дульбекко среду Игла, содержащую ферментную смесь либеразы (50 мкг/мл). После 2 часов инкубации при 37°C ткани были разрезаны на маленькие кусочки, гомогенизированы и отфильтрованы с использованием сеточного фильтра для клеток (размер пор 70 мкм). Гомогенизированную ткань серийно разбавляли в 96-луночном плоскодонном планшете для микротитрования, содержащем среду Шнейдера с добавкой 20% FCS. Количество жизнеспособных паразитов определяли из наивысшего разбавления, при котором промастиготы могут быть выращены вплоть до 7 дней инкубации при 26°C. Количество паразитов также определяли в местных дренирующих лимфатических узлах (ДЛУ) инфицированных ушей (ретромаксилярных), и подушечке стопы (подколенных), и в селезенке. Органы были извлечены, механически разъединены и затем серийно разбавлены, как описано выше. Паразитарную нагрузку выражали в виде количества паразитов в целом органе.
Измерение цитокинов в супернатантах
Селезенки и соответствующие местные ДЛУ удаляли асептически, механически разъединяли и высевали в полную среду RPMI (RPMI 1640, дополненная 10% FCS, 2 мМ глутамина и 10 мМ 2-меркаптоэтанола). 5×106 клеток/мл высевали в 48-луночных планшетах в течение 48 ч при 37°C в присутствии LRP (12 мкг/мл) или SLA (12 мкг/мл). Высвобождение IFN-γ, IL-10 и IL-4 измеряли в супернатантах клеточных культур спленоцитов и ДЛУ с применением промышленных наборов ELISA (Diaclone, Besançon, France). В некоторых случаях клетки ДЛУ, стимулированные 12 мкг/мл LRP, инкубировали в присутствии 10 мкг/мл моноклональных антител (mAb) против мышиного CD4 (GK 1.5), мышиного IL-12 (C17.8), мышиного CD8 (53-6.7). Соответствующие изотип-спаренные контроли также анализировали в тесте. Антитела (без азида/низкий эндотоксин™) были приобретены у BD (PharMingen).
Анализ гуморальных ответов
Образцы сыворотки были проанализированы на предмет специфичных антител против LRP или SLA посредством ELISA или вестерн-блоттинга. Кратко, стандартные плашки ELISA были покрыты в течение ночи при комнатной температуре 100 мкл LRP (5 мкг/мл в PBS) или SLA (2 мкг/мл в PBS). Титр определяли посредством серийного разбавления сыворотки, и он был определен как обращение наивысшего фактора разбавления сыворотки, давая поглощение >0,2. Изотип-специфичные анализы были проведены с соответствующими против-мышиными иммуноглобулинами, конъюгированными с пероксидазой хрена (Nordic Immunological Laboratories, Tilburg, Netherlands): против-IgG1 (1/1000) и против-IgG2a (1/500). Ортофенилендиамин дигидрохлорид - OPD- (Dako, A/S, Glostrup, Denmark) применяли в качестве субстрата пероксидазы для ELISA анализов. Через 15 мин реакцию останавливали посредством добавления 100 мкл H2SO4 1 M и поглощение считывали при 450 нм.
Для анализа методом вестерн-блоттинга рибосомные белки L. infantum и L. major получали, ресуспендировали в буфере Laemmli, разделяли посредством SDS-PAGE и переносили на нитроцеллюлозные мембраны (Amersham, Aylesbury, UK). Блоты испытывали с сыворотками для контрольных собак, собак, страдающих VCL лейшманиозом, или использовали сыворотки от различных групп мышей в этой работе при указанных разбавлениях. В качестве вторичных антител применяли конъюгированные с пероксидазой хрена против-мышиные против-IgG (1/1000), против-IgG1 (1/1000), против-IgG2a (1/500) иммуноглобулины и против-собачьи-IgG (1/2000), приобретенные от Nordic Immunological Laboratories (Tilburg, Netherlands).
Статистический анализ
Статистический анализ проводили посредством t-теста Стьюдента. Различия считали значимыми, когда P<0,05.
РЕЗУЛЬТАТЫ
Антигенность LRP во время инфекции
Чтобы провести анализ антигенности рибосомных белков Leishmania (LRP), реакционноспособность против L. infantum LRP сывороток от собак, естественным образом инфицированных этим паразитом, анализировали посредством вестерн-блоттинга. Наблюдали, что сыворотки от собак, страдающих активной формой заболевания, распознавали большое число белковых полос в экстракте LRP (Фиг.1A). Сыворотки от C57BL/6 и BALB/c мышей, экспериментально инфицированных L. major, также распознавали много белковых полос в L. major LRP, причем количество рибосомных белков, распознаваемых IgG антителами, присутствующими в сыворотках от BALB/c-подверженных мышей, было выше, чем количество белков, распознаваемых этими антителами в сыворотках C57BL/6-резистентных мышей (Фиг.1B).
Поскольку индукцию IgG1 и IgG2a антител можно применять в качестве маркера иммунных ответов Th2-типа и Th1-типа (8), авторы изобретения провели анализ IgG1/IgG2a поляризации против LRP у мышей, инфицированных L. major. У BALB/c мышей против-LRP ответ был преобладающим образом IgG1 изотипа, в то время как он был IgG2a изотипа у C57BL/6 мышей (Фиг.1C). Также определяли выработку IFN-γ, IL-4 и IL-10 после in vitro стимуляции клеток ДЛУ LRP у обеих линий мышей. У BALB/c мышей, страдающих CL, выработку LRP-специфичных IFN-γ обнаруживали, но также была сильно стимулирована выработка IL-4 и IL-10, причем отношение IFN-γ/IL-4 ≈2,4 и отношение IFNγ/IL-10 ≈1,4 (Фиг.1D). Напротив, было получено отсутствие IL-4 (<7,5 пг/мл) и высокое отношение IFN-γ/IL-10 (≈15), когда клетки ДЛУ от излеченных C57BL/6 мышей были стимулированы LRP (Фиг.1E).
Иммуногенность LRP у BALB/c мышей
Иммунные ответы на LRP оценивали у BALB/c мышей после введения рибосомных белков в отсутствие и в присутствии CpG ODN. После вакцинации LRPE+CpG ODN против-LRPE гуморальный ответ был преобладающим образом IgG2a изотипа, в то время как более низкий титр антител IgG1 изотипа обнаруживали в сыворотках от мышей, иммунизированных LRPE по отдельности (Фиг.2A). После in vitro стимуляции LRPE клетки селезенки от мышей, иммунизированных LRPE+GpG ODN, секретировали более высокие уровни IFN-γ, чем уровни, секретируемые клетками от контролей и от мышей, иммунизированных LRPE по отдельности (Фиг.2B). Никакого увеличения выработки IL-4 не наблюдали после стимуляции LRPE в любой группе (Фиг.2C).
Примечательно, что специфичные IL-10 обнаруживали в супернатанте культур, установленных от селезенок мышей, вакцинированных LRPE+CpG ODN, причем отношение IFNγ/IL-10 ≈40 (Фиг.2D). Все вместе эти результаты демонстрируют, что LRPE, вводимый без адъювантов, индуцировал только слабые IgG1 гуморальные ответы, но совместное введение с CpG ODN активировал Th1-подобный ответ против этих антигенов у BALB/c мышей.
Вакцинация LRPE+CpG ODN защищает BALB/c мышей против стимуляции L. major
Поскольку перенаправление Th2 ответов, индуцированных антигенами, ассоциированных с заболеванием, в сторону Th1 ответа рассматривают как перспективный подход для разработки вакцин против Leishmania (7), авторы проанализировали, способна ли вакцинация LRPE+CpG ODN индуцировать защиту против инфекции L. major. Фиг.3A показывает, что LRPE+CpG OND индуцирует эффективную защиту, поскольку набухание подушечки стопы у этих мышей было снижено (среднее значение равно 0,7 мм на неделе 8), при сравнении с набуханием у контролей и мышей, вакцинированных LRPE по отдельности (среднее значение ≈5,5 мм). Авторы затем проанализировали паразитарную нагрузку в подколенном ДЛУ и в селезенках четырех групп мышей. ДЛУ от мышей, иммунизированных LRPE-CpG, показал снижение паразитарной нагрузки по отношению к другим группам приблизительно на 3 log. Дополнительно, в то время как сходные паразитарные нагрузки были обнаружены в селезенках контрольных мышей и мышей, иммунизированных LRPE по отдельности, никаких паразитов не смогли обнаружить в селезенках мышей, вакцинированных LRPE+CpG ODN (Фиг.3B).
Для определения иммунологических параметров, ассоциированных с защитой, индуцируемой LRPE+CpG ODN, аналитически определяли SLA или LRPE-управляемую выработку IL-4, IL-10 и IFN-γ. Специфически индуцированную выработку IL-4 и IL-10, управляемую SLA или LRPE, обнаруживали в клетках ДЛУ от контролей (физраствор и CpG) и от мышей, иммунизированных LRPE по отдельности (Фиг.3C-D). Напротив, клетки ДЛУ от мышей, иммунизированных LRPE+CpG ODN, вырабатывали более высокие количества IFN-γ, чем количества, обнаруженные в других трех группах (Фиг.3E). Вклад Т-клеток CD4+ и CD8+ и зависимость от IL-12 специфической выработки IFN-γ под действием LRPE также были проанализированы. Как показано на Фиг.3F, выработка IFN-γ полностью ингибировалась моноклональными антителами против-IL-12 или против-CD4. Добавление против-CD8 антител к клеточным культурам ДЛУ только частично снижало количества этого цитокина в супернатантах. Принимая во внимание, что у BALB/c мышей IL-4 зависимая выработка высоких титров антител ассоциирована с развитием заболевания, авторы проанализировали посредством ELISA гуморальные ответы, вызванные против LRPE на восьмой неделе после инфекции. Антитела против LRPE, вызываемые паразитарной стимуляцией у мышей, которых иммунизировали LRPE+CpG ODN, были в основном IgG2a изотипа. Также более низкий титр антител против-LRPE IgG1-изотипа был представлен в сыворотках защищенных мышей, при сравнении с мышами, иммунизированными LRPE по отдельности, и в двух контрольных группах (Фиг.4A). Снижение титра IgG1 против LRPE коррелировало со снижением количества полос рибосомных белков, распознаваемых IgG1 антителами из сывороток мышей, вакцинированных LRPE+CpG ODN. Как показано на Фиг.4B, IgG1 изотип антител из сывороток мышей, иммунизированных физраствором, CpG или LRPE по отдельности распознавал более высокое количество белковых полос в LRPE вестерн-блоттингах, в то время как только несколько полос распознавались IgG1 антителами защищенных мышей. Анализ методом вестерн-блоттинга также показал, что не было увеличения в количестве белковых полос, распознаваемых IgG2a антителами в сыворотках от мышей, вакцинированных LRPE+CpG ODN, при сравнении с другими тремя группами (Фиг.4B). Вакцинация LRPE+CpG ODN также кондиционировала глобальный гуморальный ответ против-Leishmania, индуцированный инфекцией L. major. Антитела против SLA, вызываемые паразитарной стимуляцией у мышей, которых иммунизировали LRPE+CpG ODN, были в основном IgG2a изотипа, причем титр против-SLA антител IgG1 изотипа был существенно ниже, чем титры, обнаруженные в других трех группах (Фиг.4C).
LRPE+CpG ODN вакцинированные и инфицированные мыши являются резистентными против реинфекции L. major в дерму уха
Для определения, способны ли вакцинированные LRPE+CpG ODN и инфицированные мыши бороться со второй паразитарной стимуляцией, шесть BALB/c мышей были вакцинированы и инфицированы в подушечку стопы, как описано выше. Набухание подушечки стопы этой группы мышей составляло <0,7 мм в течение 18 недель (данные не показаны). Далее, эти защищенные мыши были реинфицированы 300 метациклическими промастиготами L. major в дерму уха. Контрольная группа из шести необработанных мышей была также инфицирована. Наблюдали, что реинфицированные мыши, вакцинированные LRPE+CpG ODN, были защищены против развития кожных повреждений, так как у этих мышей не наблюдали никакой патологии, в то время как у контрольных мышей развивались явные повреждения уха на седьмой неделе (Фиг.5A). Паразитарная нагрузка в дерме уха и в ретромаксилярном ДЛУ была также существенно различной между двумя группами (Фиг.5B). Низкая паразитарная нагрузка в ухе и в ДЛУ у вакцинированных реинфицированных мышей коррелирует с отсутствием паразита в селезенке. Чтобы выяснить клеточный ответ, происходящий после реинфекции, анализировали секрецию IFN-γ, IL-4 и IL-10 клетками ретромаксилярного ДЛУ клетки после in vitro стимуляции LRPE или SLA (Фиг.5C). У контрольных мышей, что также происходит, когда мыши были инфицированы в подушечку стопы (Фиг.3C-E), была обнаружена специфическая выработка IFN-γ, но также выработка IL-4 и IL-10 была сильно стимулирована. Напротив, клетки ДЛУ вакцинированных-реинфицированных мышей вырабатывали высокие количества специфичного IFN-γ, причем IL-4 и IL-10 попросту не обнаруживались (Фиг.5C). Соответственно, их IgG гуморальный ответ против LRPE (Фиг.5D) и SLA (Фиг.5E) был IgG2a изотипа.
Вакцинация LRPE+CpG ODN придает защиту против кожной патологии вследствие стимуляции L. major у C57BL/6 мышей
Принимая во внимание то, что вакцинация LRPE+CpG ODN защищает против инфекции L. major у BALB/c мышей посредством перенаправления Th2 иммунного ответа против LRPE в сторону Th1 ответа, авторы проанализировали эффект введения этой вакцины у C57BL/6 мышей, модели, которая естественным образом развивает Th1 ответы против антигенов Leishmania. Группу из C57BL/6 мышей иммунизировали тремя дозами LRPE+CpG ODN, а контрольные мыши получили только адъювант CpG ODN. Инокуляция индуцировала Th1 ответ, продемонстрированный посредством выработки in vitro IFN-γ в супернатанте культур клеток селезенки, стимулированных LRPE. Присутствие специфичного IL-10 также обнаруживали в супернатанте культур клеток селезенки, установленных от мышей, вакцинированных LRPE+CpG ODN, в то время как никакой выработки специфичного IL-4 не наблюдали после стимуляции (Фиг.6A). Мыши, вакцинированные LRPE+CpG ODN, были защищены против развитий кожных повреждений, так как наблюдали небольшую патологию или никакой патологии не наблюдали (Фиг.6B). У мышей, иммунизированных CpG ODN, развивались повреждения, которые достигали пика на седьмой неделе и были почти полностью излечены на неделе 13. Так как в этой модели количество паразитов на инфицированном участке достигало пика как раз перед развитием повреждения (5), авторы определяли паразитарную нагрузку в ухе и в местном ДЛУ (ретромаксилярном) на пятой неделе. Количество паразитов в дерме уха вакцинированных мышей имело ≈300-кратное снижение (1,0×104 паразитов для LRPE+CpG ODN и 3×106 паразитов для мышей, иммунизированных CpG-ODN) и ≈40-кратное снижение в ДЛУ (5,0×104 паразитов для LRPE+CpG ODN и 2×106 паразитов для CpG ODN мышей). После излечивания у контрольных мышей (13 недель после стимуляции) снижение количества паразитов наблюдали во всех группах.
Для определения иммунологических параметров, ассоциированных с защитой, исследовали управляемую антигенами выработку IL-4, IL-10 и IFN-γ. На пятой неделе после стимуляции культуры клеток ДЛУ клетки были установлены и стимулированы LRPE или SLA. Как показано на Фиг.6D, клетки от вакцинированных мышей вырабатывали более SLA и LRPE специфичного IFN-γ, чем клетки от контрольных мышей. Также анализировали вклад Т-клеток CD4+ и CD8+ и зависимость от IL-12 для LRPE специфичной выработки IFN-γ. Секреция IFN-γ полностью ингибировалась моноклональными антителами против-IL-12 или против-CD4. Обработка антителами против-CD8 только частично снижала уровень этого цитокина. Эти данные демонстрировали, что у C57BL/6 вакцинация LRPE+CpG ODN индуцировала снижение патологии и паразитной нагрузки в коже и в местном ДЛУ, что коррелировало с индукцией более раннего специфичного Th1 ответа против LRPE. В соответствии с этим IgG2a специфичные антитела против-LRPE были обнаружены раньше и с более высокими титрами у вакцинированных мышей по сравнению с контрольными мышами (Фиг.6F). Авторы также обнаружили SLA или LRPE антигенспецифичную выработку IL-10, которая статистически не отличалась при сравнении между вакцинированными и контрольными мышами.
ОБСУЖДЕНИЕ
В этой работе авторы показали, что многие паразитарные рибосомные белки наблюдают в сыворотках от собак с VCL и у мышей, инфицированных L. major, указывая на то, что они являются сильными антигенами в процессе протекания природной Leishmania и экспериментальных инфекций. Ответ инфицированных BALB/c мышей против LRPE был Th2 типа, так как ответ антител против-LRPE был преобладающим образом IgG1 изотипа. Это наблюдение было подтверждено тем фактом, что после in vitro стимуляции клеток ДЛУ от инфицированных мышей с помощью LRPE, в супернатантах культуры были обнаружены высокие количества IL-4. Несмотря на то, что LRPE были также вовлечены в выработку in vitro INF-γ, были обнаружены сравнимые уровни IL-10, плейотропного противовоспалительного цитокина, который придает инфицированным макрофагам невосприимчивость к активационным сигналам для деструкции паразита (31). Кроме того, принимая во внимание то, что эффекты IL-4 и IL-10 в активации заболевания у BALB/c мышей, по-видимому, являются аддитивными (обзор дан в (37)), стимуляция этих цитокинов посредством LRPE может быть принята как указание, что ответы хозяина против рибосомных антигенов благоприятствуют распространению паразита и устойчивости у BALB/c мышей. Авторы могут предположить, что в ранний период после инфицирования Leishmania иммунная система хозяина подвергается первичному воздействию антигенов посредством широко распространенного LRPE, высвобождаемого при цитолизе паразитов, что впоследствии может быть увеличено в результате пролиферации паразита. Таким образом, сильная иммуногенность LRPE и их патоантигенная роль, вероятно, основаны на их высокой распространенности и атигенной специфичности (несмотря на их эволюционно консервативный характер). Напротив, ответы C57BL/6 мышей против LRPE были Th1 типа, с генерацией IgG2a специфичных антител и выработкой in vitro высоких уровней специфичных INF-γ, причем уровни IL-4 необнаружимы. Также авторы обнаружили LRPE специфичную выработку IL-10 на неделе 14 после инфекции. Принимая во внимание, что отношение IFN-γ/IL-10 было сходным с отношением, полученным, когда клетки были стимулированы SLA (данные не показаны), авторы предполагают, что выработка IL-10, индуцированная этими антигенами, могла быть ассоциирована с регуляторным ответом, который благоприятствует устойчивости паразита, которая была видна в этой модели (4). Данные авторов указывают на то, что ответ против LRPE находится в соответствии с уклоном в сторону индукции Th2-опосредованных гуморальных ответов, относящихся к патологии у подверженного хозяина, и Th1-ассоциированым защитным ответам у резистентного хозяина. Так как перенаправление Th2 ответов, индуцированных против некоторых эпитопов Leishmania, в сторону Th1 ответа, вероятно, является перспективной стратегией для индукции защиты против инфекции L. major (7), авторы сначала решили проанализировать у BALB/c мышей иммуногенность LRPE, совместно вводимых с CpG ODN, адъювантом, который придает Th1-относящийся долговременный иммунитет и защиту, при иммунизации различными антигенами Leishmania (43), и который также может подавлять некоторые паразит-специфичные Th2-ответы у мышей (12, 57). Как показано на Фиг.2, было обнаружено, что иммунный ответ, развивающийся у BALB/c, принадлежал к типу Th1, так как иммунизированные мыши развивали антитела против-LRPE IgG2a изотипа, и спленоциты из вакцинированных мышей продуцировали высокие количества IFN-γ, но не IL-4 после in vitro стимуляции LRPE. Сходные специфичные иммунные ответы были генерированы у C57BL/6 мышей, когда их иммунизировали LRPE+CpG ODN (Фиг.6A). У обеих линий мышей управляемую LRPE выработку IL-10 также наблюдали после иммунизации.
Несмотря на сообщения о том, что вакцинация генетическими вакцинами, кодирующими L31, может быть вовлечена в выработку специфичных IL-10 после генетической вакцинации (44), авторы полагают, что выработка IL-10 может в большей степени относиться к гомеостатическому контролю Th1 ответов, имеющих место после введения LRPE+CpG ODN. В действительности недавно сообщалось, что INF-γ продуцирующие Th1 клетки могут также быть вовлечены в выработку IL-10 по принципу механизма контроля с обратной связью (2). Более того, полученные высокие значения отношения IFNγ/IL-10 могут обеспечить хороший прогноз по выходу вакцины (44).
Данные, приведенные здесь, указывают на то, что совместное введение LRPE+CpG ODN индуцирует защиту в двух различных моделях кожного экспериментального лейшманиоза: высокая доза инокулята в подушечке стопы BALB/c мышей (широко применяется в исследованиях вакцин при кожном лейшманиозе) и низкая доза в ухе C57BL/6 мышей (модель, которая более близко имитирует заболевание у человека, с точки зрения пути и инфекционной дозы). Вакцинированные BALB/c мыши имели сниженную паразитарную нагрузку в подколенном ДЛУ при отсутствии диссеминации паразита в селезенке. Также очень низкое воспаление обнаруживали в инфицированных подушечках стопы. Вакцинированные C57BL/6 мыши были защищены против ушной кожной патологии, показывая и снижение паразитарной нагрузки в коже и в ДЛУ. Эта защита сравнима с защитой, проявляемой C57BL/6 мышами, вакцинированными убитым нагревом антигеном Leishmania плюс CpG ODN и многокомпонентной вакциной, составленной из LACK, LmST1l и TSA, также тестированной на этой модели (28, 29, 43).
Важно, что защита у обеих линий коррелирует с генерацией Th1 специфичных иммунных ответов против LRPE. Анализ клеточных ответов in vitro измеряли на неделе 8 после инфицирования у BALB/c мышей и на неделе 5 (совпадающей с пиком паразитной нагрузки (4)) у C57BL/6 мышей. Клетки ДЛУ клетки от обеих линий мышей, вакцинированных LRPE+CpG ODN, секретировали более высокие уровни IFN-γ, чем их соответствующие контрольные группы при стимуляции LRPE. Было обнаружено, что IFN-γ ответ был IL-12 зависимым и продуцировался Т-клетками CD4+ с меньшим вкладом для Т-клеток CD8+. Как ожидалось, SLA-специфичная выработка IFN-γ была выше для защищенных мышей обеих линий. У BALB/c мышей генерация Th1 ответов у защищенных мышей коррелирует с генерацией преобладающих IgG2a специфичных антител против LRPE (Фиг.4A). Некоторые различия между линиями наблюдали в выработке in vitro IL-10. Данные авторов показали, что защищенные BALB/c мыши продуцировали значительно более низкие уровни IL-10, чем контроли после in vitro стимуляции как LRPE, так и SLA, в соответствии с вовлечением этого цитокина в подверженность в этой модели (35). С другой стороны, как контрольные, так и вакцинированные C57BL/6 LNC продуцировали IL-10 после in vitro стимуляции LRPE или SLA во время острой фазы инфекции (Фиг.6F). Как и в случае с IFN-γ, уровни IL-10 были выше, когда клетки были стимулированы LRPE, чем SLA. Тот факт, что выработка IFN-γ и IL-10 следовала сходному профилю, может быть принят как указание на то, что выработка IL-10 после вакцинации отражает гомеостатические механизмы, которые контролируют вредные эффекты, которые сильный Th1 будет вызывать у хозяина (4). Примечательно, что авторы обнаружили, что защита, показанная у BALB/c мышей после вакцинации LRPE+CpG ODN, также имеет отношение к существенному снижению выработки управляемого антигеном IL-4 после стимуляции in vitro LRPE или SLA. Эти клеточные ответы коррелировали in vivo с обращением Th2-опосредованных ответов антител против рибосомных белков. Таким образом, сыворотки от защищенных BALB/c мышей представляли существенное снижение по титру и, особенно, по количеству антигенов, распознаваемых IgG1 антителами, специфичными для LRPE. В дополнение, иммунизация BALB/c мышей LRPE+CpG ODN также имела четкий эффект на глобальный гуморальный ответ, вызываемый у мышей инфекцией L. major (Фиг.5C). Таким образом, инфицирование вакцинированных мышей индуцирует ограниченные IgG1 против-Leishmania специфичные антитела, в то время как гуморальный ответ, индуцированный в других исследованных контрольных группах мышей, был выше и с преобладанием антител Th2 типа (т.e. IgG1 изотипа). В целом, защита, наблюдаемая у вакцинированных мышей из обеих линий мышей, коррелирует с генерацией Th1 ответов против LRPE, что также приводит к пониженной регуляции управляемых IL-4 Th2 и IL-10 ответов против SLA у BALB/c мышей. Данные авторов указывают, что защищенные BALB/c мыши приобретали иммунологический статус, который придает им способность сопротивляться дальнейшей инфекции (привлекающая характеристика для вакцины, которая может применяться в эндемических областях, где повторное воздействие паразита будет очень частым). После рестимуляции в дерму уха эти мыши показали сильную защиту против инфекции L. major. Были обнаружены очень слабое развитие кожных повреждений (в некоторых случаях обнаруживали полное отсутствие кожных повреждений) и существенное снижение количества паразитов в инфицированном ухе и в ДЛУ. Хотя клетки ДЛУ от защищенных мышей не продуцировали больших количеств IFN-γ, чем контроли (измеряли на неделе 7 после инфекции), титр IgG2a антител против LRPE и SLA может быть взят в качестве указания на то, что эти мыши устанавливали специфичный Th1 защитный ответ после вторичной стимуляции. Примечательно, что специфическая выработка IL-4 и IL-10 цитокинов, ассоциированных с заболеванием, у этих мышей была очень низкой. Эти данные указывают, что иммунное состояние, генерированное после первой стимуляции паразитами, является крайне активным, приводя к быстрому и эффективному устранению паразитов из участка реинфекции.
Данные, представленные здесь, демонстрируют, что вакцинация LRPE+CpG ODN имеет непосредственное влияние на решения иммунной системы во время инфекции Leishmania как у резистентных, так и у подверженных мышей. По мнению авторов, генерация вакцин против такого сложного паразита, как Leishmania, могла бы быть оптимизирована посредством внедрения различных целевых антигенов в готовую лекарственную форму вакцины, приобретая преимущество этих антигенов, которые индуцируют требуемый иммунитет (в основном CD4+ и CD8+ IFN-γ опосредованные ответы), и направляя в сторону Th1 уклона патоантиген-управляемые иммунные ответы, которые приводят к патологии (IL-4 Th2-управляемый и IL-10 дезактивирующий ответы). Несмотря на это, следует учитывать, что Th2-ответ против некоторых из этих антигенов может не быть перенаправлен посредством обычных Th1-индукторов, как происходит с мета-1 антигеном L. major (50).
ССЫЛКИ
1. Aguilar-Be, I. R. da Silva Zardo, E. Paraguai de Souza, G. P. Borja-Cabrera, M. Rosado-Vallado, M. Mut-Martin, R. Garcia-Miss Mdel, C. B. Palatnik de Sousa, and E. Dumonteil. 2005. Cross-protective efficacy of a prophylactic Leishmania donovani DNA vaccine against visceral and cutaneous murine leishmaniasis. Infect Immun 73:812-9.
2. Anderson, C. F., M. Oukka, V. J. Kuchroo, and D. Sacks. 2007. CD4(+)CD25(-)Foxp3(-) Th1 cells are the source of IL-10-mediated immune suppression in chronic cutaneous leishmaniasis. J Exp Med 204:285-97.
3. Badaro, R., D. Benson, M. C. Eulalio, M. Freire, S. Cunha, E. M. Netto, D. Pedral-Sampaio, C. Madureira, J. M. Burns, R. L. Houghton, J. R. David, and S. G. Reed. 1996. rK39: a cloned antigen of Leishmania chagasi that predicts active visceral leishmaniasis. J Infect Dis 173:758-61.
4. Belkaid, Y., K. F. Hoffmann, S. Mendez, S. Kamhawi, M. C. Udey, T. A. Wynn, and D. L. Sacks. 2001. The role of interleukin (IL)-10 in the persistence of Leishmania major in the skin after healing and the therapeutic potential of antiIL-10 receptor antibody for sterile cure. J Exp Med 194:1497-506.
5. Belkaid, Y., S. Kamhawi, G. Modi, J. Valenzuela, N. Noben-Trauth, E. Rowton, J. Ribeiro, and D. L. Sacks. 1998. Development of a natural model of cutaneous leishmaniasis: powerful effects of vector saliva and saliva preexposure on the long-term outcome of Leishmania major infection in the mouse ear dermis. J Exp Med 188:1941-53.
6. Buffet, P. A., A. Sulahian, Y. J. Garin, N. Nassar, and F. Derouin. 1995. Culture microtitration: a sensitive method for quantifying Leishmania infantum in tissues of infected mice. Antimicrob Agents Chemother 39:2167-8.
7. Campos-Neto, A. 2005. What about Th1/Th2 in cutaneous leishmaniasis vaccine discovery? Braz J Med Biol Res 38:979-84.
8. Coffman, R. L. 1993. Mechanisms of helper T-cell regulation of B-cell activity. Ann N Y Acad Sci 681:25-8.
9. Coler, R. N., and S. G. Reed. 2005. Second-generation vaccines against leishmaniasis. Trends Parasitol 21:244-9.
10. Cordeiro-Da-Silva, A., M. C. Borges, E. Guilvard, and A. Ouaissi. 2001. Dual role of the Leishmania major ribosomal protein S3a homologue in regulation of T- and B-cell activation. Infect Immun 69:6588-96.
11. Chenik, M., H. Louzir, H. Ksontini, A. Dilou, I. Abdmouleh, and K. Dellagi. 2006. Vaccination with the divergent portion of the protein histone H2B of Leishmania protects susceptible BALB/c mice against a virulent challenge with Leishmania major. Vaccine 24:2521-9.
12. Chiaramonte, M. G., M. Hesse, A. W. Cheever, and T. A. Wynn. 2000. CpG oligonucleotides can prophylactically immunize against Th2-mediated schistosome egg-induced pathology by an IL-12-independent mechanism. J Immunol 164:973-85.
13. Garcia-Alonso, M., A. Blanco, D. Reina, F. J. Serrano, C. Alonso, and C. G. Nieto. 1996. Immunopathology of the uveitis in canine leishmaniasis. Parasite Immunol 18:617-23.
14. Garcia-Alonso, M., C. G. Nieto, A. Blanco, J. M. Requena, C. Alonso, and I. Navarrete. 1996. Presence of antibodies in the aqueous humour and cerebrospinal fluid during Leishmania infections in dogs. Pathological features at the central nervous system. Parasite Immunol 18:539-46.
15. Gradoni, L. 2001. An update on antileishmanial vaccine candidates and prospects for a canine Leishmania vaccine. Vet Parasitol 100:87-103.
16. Gramiccia, M., and L. Gradoni. 2005. The current status of zoonotic leishmaniases and approaches to disease control. Int J Parasitol 35:1169-80.
17. Handman, E., A. H. Noormohammadi, J. M. Curtis, T. Baldwin, and A. Sjolander. 2000. Therapy of murine cutaneous leishmaniasis by DNA vaccination. Vaccine 18:3011-7.
18. Herwaldt, B. L. 1999. Leishmaniasis. Lancet 354:1191-9.
20. Iborra, S., M. Soto, J. Carrion, C. Alonso, and J. M. Requena. 2004. Vaccination with a plasmid DNA cocktail encoding the nucleosomal histones of Leishmania confers protection against murine cutaneous leishmaniosis. Vaccine 22:3865-76.
22. Jaafari, M. R., A. Ghafarian, A. Farrokh-Gisour, A. Samiei, M. T. Kheiri, F. Mahboudi, F. Barkhordari, A. Khamesipour, and W. R. McMaster. 2006. Immune response and protection assay of recombinant major surface glycoprotein of Leishmania (rgp63) reconstituted with liposomes in BALB/c mice. Vaccine 24:5708-17.
23. Lopez, R., R. Lucena, M. Novales, P. J. Ginel, E. Martin, and J. M. Molleda. 1996. Circulating immune complexes and renal function in canine leishmaniasis. Zentralbl Veterinarmed B 43:469-74.
24. Mancianti, F., A. Poli, and A. Bionda. 1989. Analysis of renal immune-deposits in canine leishmaniasis. Preliminary results. Parassitologia 31:213-30.
25. Martins, D. R., S. M. Jeronimo, J. E. Donelson, and M. E. Wilson. 2006. Leishmania chagasi T-cell antigens identified through a double library screen. Infect Immun 74:6940-8.
26. McMahon-Pratt, D., and J. Alexander. 2004. Does the Leishmania major paradigm of pathogenesis and protection hold for New World cutaneous leishmaniases or the visceral disease? Immunol Rev 201:206-24.
28. Mendez, S., Y. Belkaid, R. A. Seder, and D. Sacks. 2002. Optimization of DNA vaccination against cutaneous leishmaniasis. Vaccine 20:3702-8.
29. Mendez, S., S. Gurunathan, S. Kamhawi, Y. Belkaid, M. A. Moga, Y. A. Skeiky, A. Campos-Neto, S. Reed, R. A. Seder, and D. Sacks. 2001. The potency and durability of DNA- and protein-based vaccines against Leishmania major evaluated using low-dose, intradermal challenge. J Immunol 166:5122-8.
30. Miles, S. A., S. M. Conrad, R. G. Alves, S. M. Jeronimo, and D. M. Mosser. 2005. A role for IgG immune complexes during infection with the intracellular pathogen Leishmania. J Exp Med 201:747-54.
31. Moore, K. W., R. de Waal Malefyt, R. L. Coffman, and A. O'Garra. 2001. Interleukin-10 and the interleukin-10 receptor. Annu Rev Immunol 19:683-765.
32. Mougneau, E., F. Altare, A. E. Wakil, S. Zheng, T. Coppola, Z. E. Wang, R. Waldmann, R. M. Locksley, and N. Glaichenhaus. 1995. Expression cloning of a protective Leishmania antigen. Science 268:563-6.
33. Nieto, C. G., R. Barrera, M. A. Habela, I. Navarrete, C. Molina, A. Jimenez, and J. L. Serrera. 1992. Changes in the plasma concentrations of lipids and lipoprotein fractions in dogs infected with Leishmania infantum. Vet Parasitol 44:175-82.
34. Nieto, C. G., I. Navarrete, M. A. Habela, F. Serrano, and E. Redondo. 1992. Pathological changes in kidneys of dogs with natural Leishmania infection. Vet Parasitol 45:33-47.
35. Noben-Trauth, N., R. Lira, H. Nagase, W. E. Paul, and D. L. Sacks. 2003. The relative contribution of IL-4 receptor signaling and IL-10 to susceptibility to Leishmania major. J Immunol 170:5152-8.
36. Pateraki, E., R. Portocala, H. Labrousse, and J. L. Guesdon. 1983. Antiactin and antitubulin antibodies in canine visceral leishmaniasis. Infect Immun 42:496-500.
37. Peters, N., and D. Sacks. 2006. Immune privilege in sites of chronic infection: Leishmania and regulatory T cells. Immunol Rev 213:159-79.
38. Pollock, K. G., K. S. McNeil, J. C. Mottram, R. E. Lyons, J. M. Brewer, P. Scott, G. H. Coombs, and J. Alexander. 2003. The Leishmania mexicana cysteine protease, CPB2.8, induces potent Th2 responses. J Immunol 170:1746-53.
39. Probst, P., E. Stromberg, H. W. Ghalib, M. Mozel, R. Badaro, S. G. Reed, and J. R. Webb. 2001. Identification and characterization of T cell-stimulating antigens from Leishmania by CD4 T cell expression cloning. J Immunol 166:498-505.
40. Rafati, S., A. Nakhaee, T. Taheri, A. Ghashghaii, A. H. Salmanian, M. Jimenez, M. Mohebali, S. Masina, and N. Fasel. 2003. Expression of cysteine proteinase type I and II of Leishmania infantum and their recognition by sera during canine and human visceral leishmaniasis. Exp Parasitol 103:143-51.
41. Rafati, S., A. H. Salmanian, T. Taheri, M. Vafa, and N. Fasel. 2001. A protective cocktail vaccine against murine cutaneous leishmaniasis with DNA encoding cysteine proteinases of Leishmania major. Vaccine 19:3369-75.
42. Requena, J. M., C. Alonso, and M. Soto. 2000. Evolutionarily conserved proteins as prominent immunogens during Leishmania infections. Parasitol Today 16:246-50.
43. Rhee, E. G., S. Mendez, J. A. Shah, C. Y. Wu, J. R. Kirman, T. N. Turon, D. F. Davey, H. Davis, D. M. Klinman, R. N. Coler, D. L. Sacks, and R. A. Seder. 2002. Vaccination with heat-killed Leishmania antigen or recombinant leishmanial protein and CpG oligodeoxynucleotides induces long-term memory CD4+ and CD8+ T cell responses and protection against Leishmania major infection. J Exp Med 195:1565-73.
44. Roberts, M. T., C. B. Stober, A. N. McKenzie, and J. M. Blackwell. 2005. Interleukin-4 (IL-4) and IL-10 collude in vaccine failure for novel exacerbatory antigens in murine Leishmania major infection. Infect Immun 73:7620-8.
45. Rodriguez-Gabriel, M. A., M. Remacha, and J. P. Ballesta. 2000. The RNA interacting domain but not the protein interacting domain is highly conserved in ribosomal protein P0. J Biol Chem 275:2130-6.
46. Rosa, R., C. Marques, O. R. Rodrigues, and G. M. Santos-Gomes. 2007. Immunization with Leishmania infantum released proteins confers partial protection against parasite infection with a predominant Th1 specific immune response. Vaccine 25:4525-32.
47. Santos-Gomes, G. M., R. Rosa, C. Leandro, S. Cortes, P. Romao, and H. Silveira. 2002. Cytokine expression during the outcome of canine experimental infection by Leishmania infantum. Vet Immunol Immunopathol 88:21-30.
48. Santos, W. R., V. M. de Lima, E. P. de Souza, R. R. Bernardo, M. Palatnik, and C. B. Palatnik de Sousa. 2002. Saponins, IL12 and BCG adjuvant in the FML-vaccine formulation against murine visceral leishmaniasis. Vaccine 21:30-43.
49. Saraiva, E. M., A. de Figueiredo Barbosa, F. N. Santos, G. P. Borja-Cabrera, D. Nico, L. O. Souza, C. de Oliveira Mendes-Aguiar, E. P. de Souza, P. Fampa, L. E. Parra, I. Menz, J. G. Dias, Jr., S. M. de Oliveira, and C. B. Palatnik-de-Sousa. 2006. The FML-vaccine (Leishmune) against canine visceral leishmaniasis: a transmission blocking vaccine. Vaccine 24:2423-31.
50. Serezani, C. H., A. R. Franco, M. Wajc, J. K. Umada Yokoyama-Yasunaka, G. Wunderlich, M. M. Borges, and S. R. Uliana. 2002. Evaluation of the murine immune response to Leishmania meta 1 antigen delivered as recombinant protein or DNA vaccine. Vaccine 20:3755-63.
51. Skeiky, Y. A., J. A. Guderian, D. R. Benson, O. Bacelar, E. M. Carvalho, M. Kubin, R. Badaro, G. Trinchieri, and S. G. Reed. 1995. A recombinant Leishmania antigen that stimulates human peripheral blood mononuclear cells to express a Th1-type cytokine profile and to produce interleukin 12. J Exp Med 181:1527-37.
52. Stager, S., D. F. Smith, and P. M. Kaye. 2000. Immunization with a recombinant stage-regulated surface protein from Leishmania donovani induces protection against visceral leishmaniasis. J Immunol 165:7064-71.
54. Tonui, W. K., J. S. Mejia, L. Hochberg, M. L. Mbow, J. R. Ryan, A. S. Chan, S. K. Martin, and R. G. Titus. 2004. Immunization with Leishmania major exogenous antigens protects susceptible BALB/c mice against challenge infection with L. major. Infect Immun 72:5654-61.
55. Webb, J. R., A. Campos-Neto, Y. A. Skeiky, and S. G. Reed. 1997. Molecular characterization of the heat-inducible LmSTI1 protein of Leishmania major. Mol Biochem Parasitol 89:179-93.
56. Webb, J. R., D. Kaufmann, A. Campos-Neto, and S. G. Reed. 1996. Molecular cloning of a novel protein antigen of Leishmania major that elicits a potent immune response in experimental murine leishmaniasis. J Immunol 157:5034-41.
57. Zimmermann, S., O. Egeter, S. Hausmann, G. B. Lipford, M. Rocken, H. Wagner, and K. Heeg. 1998. CpG oligodeoxynucleotides trigger protective and curative Th1 responses in lethal murine leishmaniasis. J Immunol 160:3627-30.
Группа изобретений относится к области медицины и ветеринарии. Применение RPE (экстракт рибосомного белка) совместно с Th1-активирующим адъювантом для получения лекарственного средства для лечения или профилактики паразитарного заболевания обеспечивает перекрестный иммунитет от различных видов Leishmania. 2 н. и 11 з.п. ф-лы, 7 ил.
1. Применение экстракта рибосомного белка из видов Leishmania и Th1-активирующего адъюванта для получения лекарственного средства для лечения или профилактики у пациента паразитарного заболевания, вызываемого видами Leishmania, где экстракт рибосомного белка может быть получен посредством осуществления следующих стадий с использованием клеток паразита, вызывающих паразитарное заболевание, когда они присутствуют у пациента:
а. смешивание клеток Leishmania с лизирующим буфером,
b. центрифугирование полученной смеси для получения цитозольного экстракта,
с. получение экстракта рибосомного белка из полученного цитозольного экстракта.
2. Применение по п.1, где лекарственное средство представляет собой вакцину.
3. Применение по п.1, где виды Leishmania представляют собой Leishmania major.
4. Применение по п.1, где Th1-активирующий адъювант включает олигодезоксинуклеотид.
5. Применение по п.1, где олигодезоксинуклеотид включает CpG ODN.
6. Применение по п.1, где паразитарное заболевание представляет собой лейшманиоз.
7. Применение по п.1, где паразитарное заболевание вызывается видами, отличающимися от видов, из которых производят экстракт рибосомного белка.
8. Композиция, содержащая экстракт рибосомного белка из видов Leishmania и Th1-активирующий адъювант, для применения в качестве лекарственного средства для лечения или профилактики у пациента паразитарного заболевания, вызываемого видами Leishmania, где экстракт рибосомного белка может быть получен посредством осуществления следующих стадий с использованием клеток паразита, вызывающих паразитарное заболевание, когда они присутствуют у пациента:
а. смешивание клеток Leishmania с лизирующим буфером,
b. центрифугирование полученной смеси для получения цитозольного экстракта,
с. получение экстракта рибосомного белка из полученного цитозольного экстракта.
9. Композиция по п.8, состоящая из экстракта рибосомного белка из видов Leishmania и Th1-активирующего адъюванта.
10. Композиция по п.8, где Th1-активирующий адъювант включает олигодезоксинуклеотид.
11. Композиция по п.8, в которой олигодезоксинуклеотид включает CpG ODN.
12. Композиция по п.8, дополнительно содержащая фармацевтически приемлемые адъювант и/или носитель.
13. Композиция по п.8, где лекарственное средство представляет собой вакцину.
| IBORRA S et al, Vaccination with the Leishmania infantum acidic ribosomal PO protein plus CpG oligodeoxynucleotides induces protection against cutaneous leishmaniasis in C57BL/6 mice but does not prevent progressive disease in BALB/c mice, Infect | |||
| Immun., 2005 sep., 73(9), abstract | |||
| IBORRA S et al, The Leishmania infantum acidic ribosomal protein |

