Область изобретения
Изобретение относится к способам увеличения молекулярной массы изолированных капсульных полисахаридов Streptococcus pneumoniae, имеющих фосфодиэфирную связь между повторяемыми единицами сахарида.
Предшествующий уровень техники
При получении поливалентных конъюгированных пневмококковых вакцин, направленных на профилактику инвазивных заболеваний, вызванных микроорганизмом Streptococcus pneumoniae (также известный как пневмококк), выбранные серотипы Streptococcus pneumoniae выращивали, чтобы получить полисахариды, необходимые для производства вакцины. Клетки выращивали в ферментаторе с индуцированным лизисом, завершая ферментацию прибавлением деоксихолата натрия или альтенативного лизирующего агента.
Бульон лизата потом собирали для очистки выходного потока и восстановления капсульных полисахаридов, которые окружают бактериальные клетки. После конъюгации с белком-носителем полисахарид был включен в заключительный продукт вакцины и обладал иммуногенными свойствами в вакцинируемой популяции в отношении выбранных серотипов Streptococcus pneumoniae.
Размер полисахарида - качественный признак, который оценивается в каждой партии препарата и должен соответственно контролироваться. Традиционная обработка включает использование NaOH (гидроксид натрия) в качестве базового титранта во время ферментации. Использование NaOH имеет преимущество понижения pH деоксихолата лизата без вспенивания, чтобы удалить белок и улучшить фильтрацию. Этот материал подвергают центрифугированию, сопровождаемому фильтрацией, чтобы удалить большинство твердых частиц ниже 0,45 µм к номинальным размерам. Однако такие традиционные способы обработки приводят к более низкой молекулярной массе полисахарида (<450 кДа) для серотипов, имеющих фосфодиэфирную связь между повторяемыми единицами сахарида (например, 6A, 6B, 19A и 19F).
Соответственно, улучшенные способы выделения высокомолекулярного капсульного полисахарида из клеточных лизатов Streptococcus pneumoniae, в частности лизатов, содержащих полисахариды серотипов Streptococcus pneumoniae 6A, 6B, 19A или 19F, необходимы.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Обеспечиваются улучшенные способы выделения высокомолекулярных капсульных полисахаридов из клеточных лизатов Streptococcus pneumoniae, содержащих серотипы, имеющих фосфодиэфирную связь между повторяемыми единицами сахарида. В одном способе CO2 вводится в ферментативную культуру серотипа Streptococcus pneumoniae, содержащего фосфодиэфирную связь между повторяемыми единицами сахарида.
Соответственно, в одном варианте реализации изобретения способ включает стадии: 1) получение ферментационной культуры бактериальных клеток Streptococcus pneumoniae, которые продуцируют капсульные полисахариды, включающие фосфодиэфирную связь между повторяемыми единицами; 2) введение CO2 в ферментационную культуру; 3) лизис бактериальных клеток в ферментационной культуре; и 4) выделение капсульных полисахаридов Streptococcus pneumoniae из ферментационной культуры, где получают жидкую фракцию, содержащую изолированные высокомолекулярные капсульные полисахариды Streptococcus pneumoniae, содержащие фосфодиэфирные связи между повторяемыми единицами.
В специфическом варианте реализации получали ферментационные культуры бактериальных клеток Streptococcus pneumoniae, которые продуцируют серотипы полисахаридов 19A, 6A, 19F, 6B и их комбинации. В другом специфическом варианте реализации введение CO2 в ферментационную культуру включает прибавление бикарбонатного иона  к ферментационной культуре, например прибавление NaHCO3 (бикарбонат натрия) к ферментационной культуре. В дополнительном варианте реализации введение CO2 в ферментационную культуру включает прибавление карбонатного иона
к ферментационной культуре, например прибавление NaHCO3 (бикарбонат натрия) к ферментационной культуре. В дополнительном варианте реализации введение CO2 в ферментационную культуру включает прибавление карбонатного иона 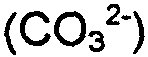 к ферментационной культуре, например прибавление Na2CO3 (карбонат натрия) к ферментационной культуре.
к ферментационной культуре, например прибавление Na2CO3 (карбонат натрия) к ферментационной культуре.
В другом варианте реализации введение CO2 в ферментационную культуру включает первое прибавление NaHCO3 и второе прибавление Na2CO3. В еще одном варианте реализации введение CO2 в ферментационную культуру включает покрытие ферментационной культуры CO2. В другом варианте реализации молекулярная масса изолированных капсульных полисахаридов Streptococcus pneumoniae является, по крайней мере, 700 кДа. В другом варианте реализации данное изобретение относится к жидкой фракции, содержащей изолированные высокомолекулярные капсульные полисахариды Streptococcus pneumoniae, где полисахариды включают фосфодиэфирные связи между повторяемыми единицами, где жидкая фракция получена способом, описанным выше.
В другом варианте реализации данного изобретения предоставлен способ получения жидкой фракции, содержащей изолированные высокомолекулярные капсульные полисахариды Streptococcus pneumoniae серотипа 19А. Способ включает стадии: 1) получение ферментационной культуры бактериальных клеток Streptococcus pneumoniae, которые продуцируют капсульные полисахариды серотипа 19A; 2) введение CO2 в ферментационную культуру; 3) лизис бактериальных клеток в ферментационной культуре; и 4) выделение капсульных полисахаридов Streptococcus pneumoniae серотипа 19A из ферментационной культуры; посредством чего была получена жидкая фракция, содержащая изолированные высокомолекулярные капсульные полисахариды Streptococcus pneumoniae серотипа 19А. В другом варианте реализации данное изобретение относится к жидкой фракции, содержащей изолированные высокомолекулярные капсульные полисахариды Streptococcus pneumoniae серотипа 19А, где жидкая фракция получена способом, описанным выше.
В другом варианте реализации данного изобретения предоставлен способ получения жидкой фракции, содержащей высокомолекулярные капсульные полисахариды изолированного серотипа 19F Streptococcus pneumoniae. Способ включает стадии: 1) получение ферментационной культуры бактериальных клеток Streptococcus pneumoniae, которые продуцируют капсульные полисахариды серотипа 19F; 2) введение CO2 в ферментационную культуру; 3) лизис бактериальных клеток в ферментационной культуре; и 4) выделение капсульных полисахаридов Streptococcus pneumoniae серотипа 19F из ферментационной культуры; посредством чего была получена жидкая фракция, содержащая высокомолекулярные капсульные полисахариды Streptococcus pneumoniae серотипа 19F. В другом варианте реализации данное изобретение относится к жидкой фракции, содержащей высокомолекулярные капсульные полисахариды Streptococcus pneumoniae серотипа 19F, где жидкая фракция получена способом, описанным выше.
В другом варианте реализации данного изобретения предоставлен способ получения жидкой фракции, содержащей высокомолекулярные капсульные полисахариды серотипа 6А Streptococcus pneumoniae. Способ включает стадии: 1) получение ферментационной культуры бактериальных клеток Streptococcus pneumoniae, которые продуцируют капсульные полисахариды серотипа 6А; 2) введение CO2 в ферментационную культуру; 3) лизис бактериальных клеток в ферментационной культуре; и 4) выделение капсульных полисахаридов Streptococcus pneumoniae серотипа 6A из ферментационной культуры; посредством чего была получена жидкая фракция, содержащая высокомолекулярные капсульные полисахариды Streptococcus pneumoniae серотипа 6A. В другом варианте реализации данное изобретение относится к жидкой фракции, содержащей изолированные высокомолекулярные капсульные полисахариды Streptococcus pneumoniae серотипа 6A, где жидкая фракция получена способом, описанным выше.
В другом варианте реализации данного изобретения предоставлен способ получения жидкой фракции, содержащей изолированные высокомолекулярные капсульные полисахариды серотипа 6B Streptococcus pneumoniae. Способ включает стадии: 1) получение ферментационной культуры бактериальных клеток Streptococcus pneumoniae, которые продуцируют капсульные полисахариды серотипа 6B; 2) поставку CO2 ферментационной культуре; 3) лизис бактериальных клеток в ферментационной культуре; и 4) выделение капсульных полисахаридов серотипа 6B Streptococcus pneumoniae из ферментационной культуры; посредством чего была получена жидкая фракция, содержащая изолированные высокомолекулярные капсульные полисахариды Streptococcus pneumoniae серотипа 6B. В другом варианте реализации данное изобретение относится к жидкой фракции, содержащей изолированные высокомолекулярные капсульные полисахариды Streptococcus pneumoniae серотипа 6B, где жидкая фракция получена способом, описанным выше.
КРАТКОЕ ОПИСАНИЕ ФИГУР
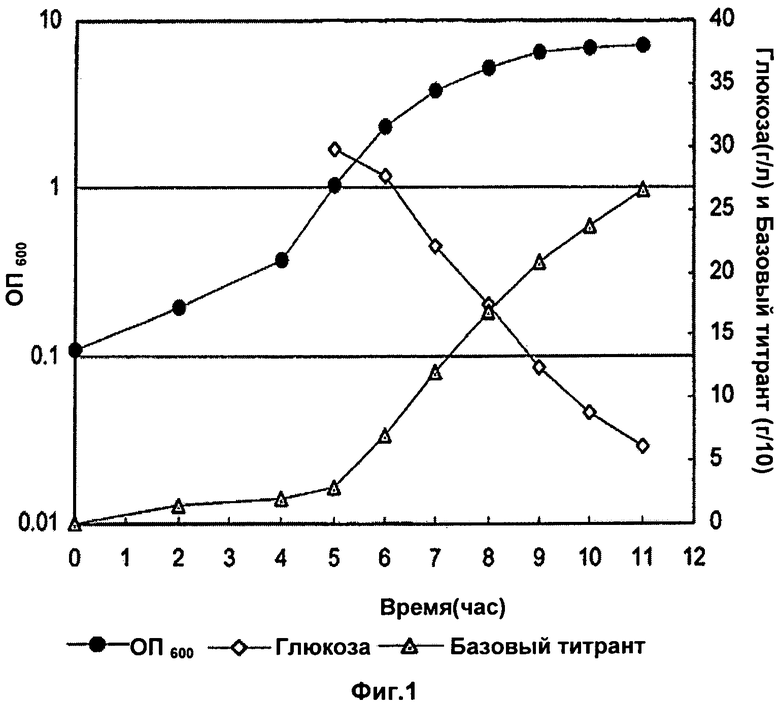
Фигура 1 показывает оптическую плотность (ОП), базовый и уровень глюкозы во время фазы ферментации с Na2CO3 в качестве базового титранта из лабораторных исследований до 3 л масштаба. Базовый титрант в граммах разделен на 10 для построения графика.
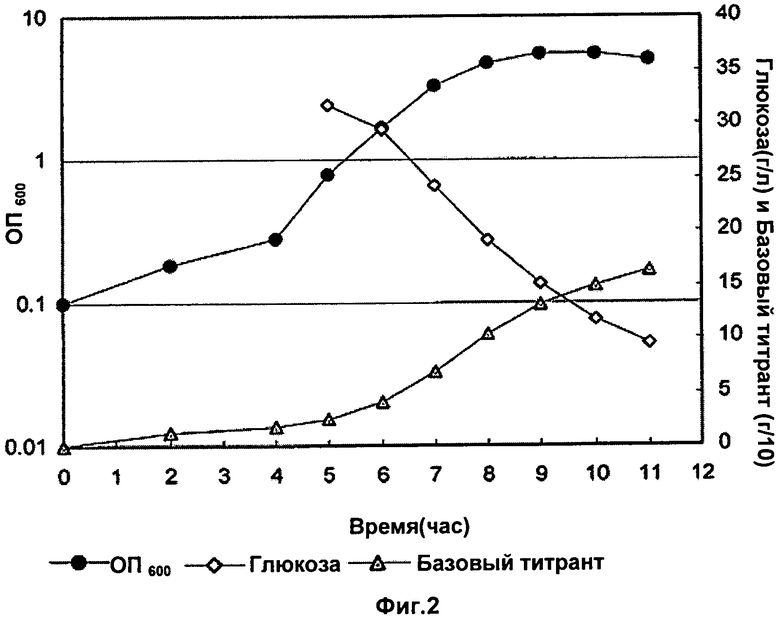
Фигура 2 показывает оптическую плотность (ОП), базовый и уровень глюкозы во время фазы ферментации с NaOH в качестве базового титранта из лабораторных исследований до 3 л масштаба. Базовый титрант в граммах разделен на 10 для построения графика.
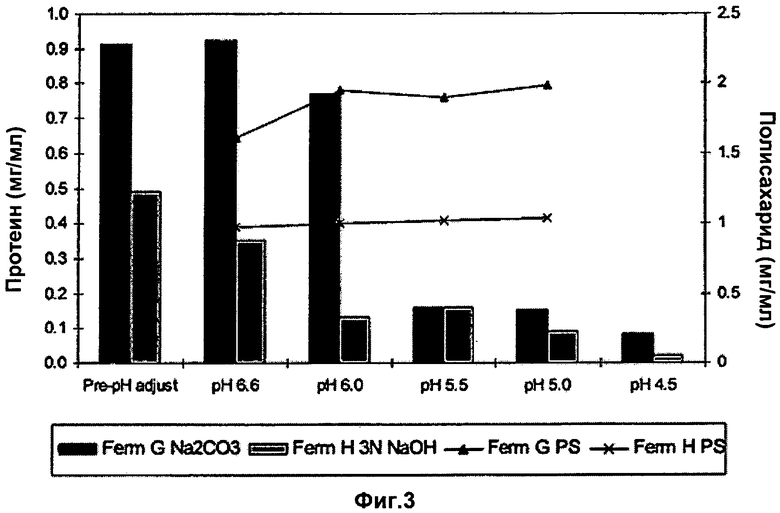
Фигура 3 показывает общий уровень белка и полисахарида при различных корректировках pH к дополнительной основной подаче Na2CO3 или NaOH.
ДЕТАЛЬНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Streptococcus pneumoniae - грам-положительные, ланцетоподобные кокки, которые обычно находятся в парах (диплококки), но также и в коротких цепях или как отдельные клетки. Они легко растут на чашке с кровяным агаром блестящими колониями и показывают альфа-гемолиз, если не выращены анаеробно, где они показывают бета-гемолиз. Клетки большинства серотипов пневмококков имеют капсулу, которая покрывает каждую клетку. Эта капсула - детерминанта вирулентности у людей, поскольку интерферирует с фагоцитозом, препятствуя антителам присоединяться к бактериальным клеткам. В настоящее время идентифицировано более чем 90 известных серотипов капсульных полисахаридов, с 23 общими серотипами, вызывающими приблизительно 90% инвазивных заболеваний во всем мире.
Как вакцина, полисахаридная оболочка пневмококков может предоставить необходимую степень иммунности к Streptococcus pneumoniae у людей с развитой или неослабленной иммунной системой, но конъюгированный белок с полисахаридом допускает иммунную реакцию у детей и пожилых, которые имеют риск пневмококковой инфекции. Пневмококковые клетки выращивали в ферментаторе с индуцированным лизисом в конце ферментации. Бульон лизата потом собирали для очистки выходного потока и восстановления капсульных полисахаридов.
Размер полисахарида - качественный признак, оцененный для каждой партии препарата, и должен соответственно контролироваться. Молекулярная масса серотипов, имеющих фосфодиэфирную связь между повторяемыми единицами сахарида (например, 6A, 6B, 19A и 19F), зависит от параметров процесса ферментации. Способы данного изобретения позволяют выделить капсульные полисахариды с высокой молекулярной массой из клеточных лизатов Streptococcus pneumoniae, содержащих серотипы, имеющие фосфодиэфирную связь между повторяемыми единицами сахарида, такие как серотип 6A, серотип 6B, серотип 19A, серотип 19F и их комбинации.
В развитии настоящих способов концентрация HySoy и выбор базового титранта были модифицированы в попытке изменить заключительный выход полисахаридов и молекулярные массы. Были проверены четыре схемы ферментации. В первой использовался базовый NaOH процесс с 28 г/л HySoy. Во второй использовался 20% карбонат натрия как базовый титрант и 20 г/л HySoy. В третьей скомбинированы преимущества первых двух подходов введением карбоната путем порционирования бикарбоната натрия и используя смешанный NaOH/карбонат в качестве базового титранта. В четвертом подходе использовался карбонат в качестве базового титранта с прибавлением 10 мМ бикарбоната, чтобы поддержать рост.
Использование NaOH в качестве базового титранта во время ферментации давало возможность снизить деоксихолат лизата до pH 5,0 без вспенивания, чтобы удалить белок и улучшить фильтрацию, но привело к более низкой молекулярной массе полисахарида (<450 кДа). Na2CO3 обеспечил более высокую молекулярную массу, но имел результатом вспенивание, если pH деоксихолата лизата был снижен. На стадии удержания более высокого pH - 6,6, ферментация с использованием Na2CO3 привела к образованию гелеподобного материала с последующими проблемами фильтрации.
Минимизация количества Na2CO3 при использовании смеси NaOH и Na2CO3 в качестве pH титранта обеспечила преимущества величины молекулярной массы при Na2CO3, устраняя вспенивание и проблемы фильтрации из-за непредвиденного высвобождения большого количества CO2. Использование 20% Na2CO3 (мас./об.) в качестве базового титранта с прибавлением 10 мМ NaHCO3, чтобы поддержать рост (четвертый подход), привело к выходу однородных высокомолекулярных полисахаридов.
Данное изобретение таким образом обеспечивает улучшенные способы выделения высокомолекулярных капсульных полисахаридов из клеточных лизатов Streptococcus pneumoniae, содержащих серотипы, имеющих фосфодиэфирную связь между повторяемыми единицами сахарида. В одном способе CO2 вводится в ферментативную культуру серотипа Streptococcus pneumoniae, содержащего фосфодиэфирную связь между повторяемыми единицами сахарида. Примеры серотипов Streptococcus pneumoniae, содержащих фосфодиэфирную связь между повторяемыми единицами сахарида, включают серотипы 6A, 6B, 19A и 19F. Соответственно, в одном варианте реализации изобретения предоставлен способ получения жидкой фракции, содержащей изолированные капсульные полисахариды высокой молекулярной массы Streptococcus pneumoniae, которые включают фосфодиэфирные связи между повторяемыми единицами, который включает стадии: 1) получение ферментационной культуры бактериальных клеток Streptococcus pneumoniae, которые продуцируют капсульные полисахариды, включающие фосфодиэфирную связь между повторяемыми единицами; 2) введение CO2 в ферментационную культуру; 3) лизис бактериальных клеток в ферментационной культуре; и 4) выделение капсульных полисахаридов Streptococcus pneumoniae из ферментационной культуры; посредством чего была получена жидкая фракция, содержащая изолированные высокомолекулярные капсульные полисахариды Streptococcus pneumoniae с фосфодиэфирными связями между повторяемыми единицами. В другом варианте реализации данное изобретение относится к жидкой фракции, содержащей изолированные высокомолекулярные капсульные полисахариды Streptococcus pneumoniae с фосфодиэфирными связями между повторяемыми единицами, где жидкая фракция получена способом, описанным выше.
Способы изобретения приводят к капсульным полисахаридам с высокой молекулярной массой Streptococcus pneumoniae, которые включают фосфодиэфирные связи между повторяемыми единицами (например, серотипы 6A, 6B, 19A и 19F). Как используется здесь, ″высокая молекулярная масса″ относится к молекулярным массам, которые составляют по крайней мере приблизительно 480 кДа, приблизительно 490 кДа, приблизительно 500 кДа, приблизительно 510 кДа, приблизительно 520 кДа, приблизительно 525 кДа, приблизительно 550 кДа, приблизительно 575 кДа, приблизительно 600 кДа, приблизительно 625 кДа, приблизительно 650 кДа, приблизительно 675 кДа, приблизительно 700 кДа, приблизительно 725 кДа, приблизительно 750 кДа, приблизительно 775 кДа, приблизительно 800 кДа, приблизительно 825 кДа, приблизительно 850 кДа, приблизительно 875 кДа, приблизительно 900 кДа, приблизительно 925 кДа, приблизительно 950 кДа, приблизительно 975 кДа или приблизительно 1000 кДа.
В определенных способах введение CO2 в ферментационную культуру включает прибавление бикарбонатного иона к ферментационной культуре например, прибавление NaHCO3 к ферментационной культуре. Прибавление NaHCO3 5-50 мМ может использоваться, такое как 5 мМ, 10 мМ, 15 мМ, 20 мМ, 25 мМ, 30 мМ, 35 мМ, 40 мМ, 45 мМ, или 50 мМ. В других способах введение CO2 в ферментационную культуру включает прибавление карбонатного иона к ферментационной культуре, например прибавление Na2CO3 к ферментационной культуре.
Прибавления Na2CO3 0,1 М-2,0 М могут использоваться, такие как 0,1 М, 0,2 М, 0,4 М, 0,6 М, 0,7 М, 0,9 М, 1,0 М, 1,5 М, 1,8 М или 2,0 М. Массообъемный (мас./об.) эквивалент может также использоваться, такой как 5% (мас./об.) Na2CO3, 10% (мас./об.) Na2CO3 или 20% (мас./об.) Na2CO3. В остальных других способах, введение CO2 в ферментационную культуру включает первое прибавление NaHCO3 и второе прибавление Na2CO3 к ферментационной культуре. В дальнейших способах, введение CO2 в ферментационную культуру включает покрытие ферментационной культуры CO2. Покрытие CO2 5-100% может использоваться, например, 5%, 10%, 15%, 20%, 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% или 100%.
В пределах способов данного изобретения бактериальные клетки могут быть подвергнуты лизису, используя любой литический агент. ″Литический агент″ является любым агентом, который способствует деструкции клеточной оболочки и высвобождению автолизина, который вызывает клеточный лизис, включая, например, детергенты. Как используется здесь, термин ″детергент″ относится к любому анионному или катионному детергенту, способному индуцировать лизис бактериальных клеток. Примеры таких детергентов для использования в пределах способов данного изобретения включают деоксихолат натрия (DOC), N-лаурил саркозин (NLS), хенодеоксихолат натрия и сапонины.
В одном варианте реализации данного изобретения литическим агентом, используемым для лизиса бактериальных клеток, является DOC. DOC - натриевая соль желчной кислоты - деоксихолевой кислоты, которую обычно получают из биологических источников, таких как коровы или волы. DOC активизирует LytA протеин, который является автолизином, который вовлечен в рост клеточной оболочки, и деление Streptococcus pneumoniae. LytA протеин имеет холин связывающие домены в его части C-терминала, и мутации lytA гена, как известно, приводят к LytA мутантам, которые являются резистентными к лизису DOC.
Хотя нет никакого свидетельства, что использование DOC во время очистки полисахарида Streptococcus pneumoniae представляет риск для здоровья, использование таких биологических реагентов может затронуть потенциальные регулирующие вопросы. Соответственно, в одном варианте реализации данного изобретения литический агент, используемый для лизиса бактериальных клеток, является литическим агентом неживотного происхождения. Литические агенты неживотного происхождения для использования в пределах способов данного изобретения включают агенты от неживотных источников с механизмом активности, подобным DOC (т.е. влияние LytA функции, приводящее к лизису клеток Streptococcus pneumoniae).
Такие литические агенты неживотного происхождения включают, но не ограничиваются следующими: аналоги DOC, сурфактанты, детергенты и структурные аналоги холина, и могут быть определены, используя процедуры, как описано в Экспериментальном разделе здесь ниже. В одном варианте реализации литический агент неживотного происхождения выбран из группы, состоящей из следующих: декансульфоновая кислота, трет-октилфенокси поли (оксиэтилен) этанолы (например, Igepal® CA-630, CAS #: 9002-93-1, доступный от Sigma Aldrich, St. Louis, МО), конденсаты октилфенол этиленоксида (например, Triton® Х-100, доступный от Sigma Aldrich, St. Louis, МО), N-лаурил саркозин натрий (NLS), лаурил иминодипропионат, додецилсульфат натрия, хенодеоксихолат, гиодеоксихолат, гликодеоксихолат, тауродеоксихолат, таурохенодеоксихолат и холат. В другом варианте реализации литический агент неживотного происхождения - NLS.
В пределах способов данного изобретения - выделение капсульных полисахаридов Streptococcus pneumoniae, используя стандартные способы, известные специалистам в данной области. Например, после ферментации бактериальных клеток, которые продуцируют капсульные полисахариды Streptococcus pneumoniae, бактериальные клетки подвергали лизису, чтобы получить клеточный лизат. Капсульные полисахариды могут быть потом выделены из клеточного лизата, используя способы очистки, известные в данной области, включая центрифугирование, осаждение, ультрафильтрацию и колоночную хроматографию (см., например, опубликованные патентные заявки США №20060228380, 20060228381, 20070184071, 20070184072, 20070231340 и 20080102498).
Изменения процесса, описанные выше, позволяют выделить капсульные полисахариды с высокой молекулярной массой из клеточных лизатов Streptococcus pneumoniae, содержащих серотипы, имеющих фосфодиэфирную связь между повторяемыми единицами сахарида, такими как серотип 6A, серотип 6B, серотип 19A, серотип 19F и их комбинации. Это - существенное усовершенствование процесса ферментации/выделения, который может значительно увеличить производство пневмококковых полисахаридов.
Следующие примеры предлагаются как иллюстративные, а не ограничительные.
ПРИМЕРЫ
Выбранные серотипы Streptococcus pneumoniae были выращены, чтобы получить полисахариды, необходимые для производства вакцины для активной иммунизации против инвазивных заболеваний, вызванных Streptococcus pneumoniae, благодаря капсульным серотипам, включенным в вакцину. Клетки выращивали в ферментаторе с индуцированным лизисом в конце ферментации. Бульон лизата потом собирали для очистки выходного потока и восстановления капсульных полисахаридов. Поскольку размер полисахарида - качественный признак, оцененный для каждой партии препарата, размер полисахарида должен контролироваться. Молекулярная масса серотипов, имеющих фосфодиэфирную связь между повторяемыми единицами сахарида (например, 6A, 6B, 19A и 19F), как находили, зависела от параметров процесса ферментации. Следующий пример описывает исследования, касающиеся поставки CO2 во время ферментации серотипов Streptococcus pneumoniae, имеющих фосфодиэфирную связь между повторяемыми единицами, чтобы улучшить молекулярную массу полисахарида.
Пример 1: Влияние введения диоксида углерода на молекулярную массу полисахарида
Ферментация
Лабораторные работы были выполнены в 3 л ферментаторах Braun Biostat В (В. Braun Biotech, Allentown, PA). Они были наполнены средой HYS 1,8 л (20 г/л HySoy, 2,5 г/л NaCl, 0,5 г/л KH2PO4, 0,013 г/л CaCl2·2H2O, 0,15 г/л L-цистеина HCl). Ферментаторы потом автоклавировали в течение 60 мин при 121°C. После охлаждения добавляли 40 или 60 мл/л раствора 50% Глюкозы + 1% Сульфата Магния (мас./об.) (DMS) к каждой единице. Если необходимо, бикарбонат натрия прибавляли перед инокуляцией.
Две 2 л колбы для посева, содержащие 1 L HYS среды, были инокулированы Типом 19A или Типом 6A семенного фонда и инкубированы при 36°C без встряхивания в течение приблизительно 6-8 часов. Инокуляцию ферментатора выполняли с объемом 100 мл (~5,2% об./об.) аликвотированного от колбы с ОП600 между 0,3-0,9 и pH между 4,75-5,60. Температуру ферментации и pH контролировали при заданных значениях. Стандартные условия - 36°C, воздушный поток 1 л/мин, контролированный pH до 7 и взбалтывание 75 об/мин, использовались. Два импеллера были помещены в низкие и средние положения на вал мешалки. Колба, содержащая соответствующий базовый титрант (3 N NaOH, 3 N NaOH, смешанный с различными концентрациями NaHCO3, 3 N NaOH, смешанный с различными концентрациями Na2CO3 и NaHCO3, и 20% Na2CO3), была присоединена для автоматического контроля за pH. Образцы изымали в различные периоды времени для определения pH, ОП600, глюкозы, полисахарида и белка. Процесс был закончен, когда концентрация глюкозы была близка к истощению или никакое увеличение ОП в течение долгого времени не было отмечено.
Измерение оптической плотности (ОП600)
Определяли клеточную плотность бульона ферментации, считывая спектральную поглощательную способность образцов при 600 нм, используя Shimadzu (Columbia, MD) UV-1601 (полоса пропускания 2 нм) или Spectronics (Westbury, NY) Genesys 5 спектрофотометр (полоса пропускания 5 нм). Единица была заполнена средой HYS, разбавленной деионизированной водой (ДИ) в количестве, необходимом для растворения образца. Образец был растворен, чтобы удерживать спектральную поглощательную способность ниже считывания 0,4, который в пределах линейного диапазона спектрофотометра.
Концентрация Глюкозы
Определяли уровни глюкозы, центрифугируя клетки и используя супернатант непосредственно или 3× растворенный с ДИ водой. Образцы были проанализированы на Nova Biomedical (Waltham, MA) BioProfile 400.
Анализ Полисахарида
Отбирали образцы при снятии показаний по окончании ферментации и обрабатывали 12% деоксихолатом натрия (DOC) до концентрации 0,13% (мас./об.) и осторожно перемешивали. Их выдерживали 8-24 час при 5°C потом pH доводили до 5,0 50% уксусной кислотой, чтобы осадить большую часть DOC и белка. После другого интервала выдержки 12-24 часов при 5°C, образцы центрифугировали (14000 об/мин, Son/all (Thermo Fisher Scientific, Waltham, MA) ротор SS34, 10 минут при 15°C). pH супернатанта доводили до 6,0. Супернатант потом фильтровали через 0,45 мкм Pall (East Hills, NY) HT Tuffryn Membrane шприц-фильтр (низкое связывание белка). Фильтрованный продукт анализировали высокоэффективной эксклюзионной хроматографией (HPLC-SEC), используя стандартную методику, известную данной области (см., например, Aquilar, M. «HPLC of Peptides and Proteins: Methods and Protocols» Totowa, NJ: Humana Press (2004)).
Анализ Белка
Уровни белка анализировали электрофорезом на полиакриламидном геле натрий додецилсульфата (SDS-PAGE) способом, известным в данной области (см., например, Walker, J.M. ″The Protein Protocols Handbook″ Totowa, NJ: Humana Press (2002)). Фильтрованный клеточный лизат (супернатант) как был получен выше, помещали аликвотами в пробирки микроцентрифуги 65 мкл/пробирку. Добавки восстанавливающего агента (10 мкл дитиотреитола (DTT)) и NuPAGE® (Invitrogen, Carlsbad, CA) 4× литий додецилсульфат (LDS) буфер для образца (25 мкл) были сделаны к каждому образцу. Образцы встряхивали и нагревали в течение 10 минут перед выгрузкой 10 мкл/ряд на 4-12% NuPAGE® Bis-Tris 12 лунки с гелем. Гели запускали в буфер NuPAGE® MES-SDS при 150 V, ограничиваясь примерно 60 минутами, и впоследствии окрашивали, используя протокол окрашивания Zoion (Zoion Biotech, Worcester, MA). Анализ образцов выполняли, используя визуализатор UVP (UVP Inc, Upland, CA) с LabWorks™ (UVP Inc) с программным обеспечением V.3, чтобы получить приблизительные концентрации определенных полос интересующего белка. Бычий сывороточный альбумин (BSA) Фракция V использовался, чтобы построить калибровочную кривую белка, чтобы вычислить приблизительные значения белка в образцах (в бульоне клеточного лизата).
Анализ Молекулярной массы
Ферментированные образцы 1-2 л обрабатывали 12% DOC натрия до концентрации 0,13% (мас./об.) при встряхивании 200 об/мин. Образцы выдерживали 8-24 час или при 5°C или при 20°C. рН образцов потом корректировали до 5,0 или 6,6 50% уксусной кислотой, чтобы осадить большую часть DOC и белка. После другого интервала выдержки 12-24 часов при 5°C образцы центрифугировали (11000 об/мин, Sorvall (Thermo Fisher Scientific, Waltham, MA) ротор SLA-3000, 15 минут при 10°C). pH образцов супернатантов потом корректировали до 6,0 3 N NaOH и фильтровали, используя фильтры МР60 0,45 мкм Millipore (Billerica, MA). Образцы затем подвергали модифицированному процессу очистки, состоящему из диафильтрации обрезанной молекулярной массы 100 К (MWCO) (5× концентрация с последующей 7,5× диафильтрацией ДИ водой), 0,1% НВ осаждения, углеродной фильтрации. Очищенный материал затем подвергали анализу Multi Angle Laser Light Scattering (MALLS).
Исследование Процесса Ферментации
Базируясь на предыдущих исследованиях, процесс ферментации был оптимизирован, заменяя Na2CO3 на NaOH в качестве базового титранта. Использование NaOH позволило восстановить pH до сниженного 5,0, что привело к существенному осаждению белка. Na2CO3 высвобождает CO2 при низком pH (<6,6), создавая пенообразование. Воздействие базового титранта на Тип 19A полисахарида и уровни белка было исследовано. Два 3 л ферментатора устанавливали с одним ферментатором, служащим в качестве контроля над процессом, используя 20% раствор Na2CO3 (маc./об.) в качестве основной подачи. В другом ферментаторе использовался 3 N NaOH в качестве основной подачи.
Во время фазы восстановления клетки были лизированы в ферментаторе с DOC (конечная концентрация 0,13% (мас./об.)), ферментатор выдерживали при 36°C в течение 30 минут. После этой стадии лизат оставляли на ночь при встряхивании при температуре окружающей среды (22°C). После выдержки лизата pH лизата титровали в диапазоне от неоткорректированного до 4,5 с образцами, взятыми при различных заданных значениях pH. Эти образцы выдерживали ночь при температуре окружающей среды перед обработкой и анализом полисахаридов и концентраций белка. ОП, базовый и уровень глюкозы во время фазы ферментации показаны на Фиг.1 и Фиг.2. Главным различием была более высокая заключительная ОП в случае использования карбоната.
Эффект пострегулирования pH DOC лизата на общие уровни белка был также исследован и показан на Фиг.3. Более низкие уровни pH уменьшали белковую нагрузку в клетке, свободной от бульона и при использовании NaOH, и при использовании Na2CO3. Более низкий pH (<6,6) не оказывал негативного воздействия на выход полисахаридов. Результаты анализа ферментации служили признаком, что основная подача NaOH была приемлемой альтернативой процессу, использующему основную подачу Na2CO3 во время ферментации, но привела к более низким выходам, чем при подаче Na2CO3.
Эффект базового титранта на молекулярную массу 19A и 6А
Выполняли ряд ферментации при 3 л процессе, чтобы определить влияние базового титранта, концентрации HySoy и pH стадии выдержки на молекулярную массу серотипа 19A. Определение молекулярной массы осуществляли, используя исследование MALLS с последующим модифицированным процессом очистки. Результаты показаны в Таблице 1. Для серотипа 6А только базовый титрант был оценен. Результаты показаны в Таблице 2.
Эффект Бикарбоната и Смешанного базового pH титранта
В первом исследовании (Процессы L29331-122 и -139) переменные уровни начального бикарбоната натрия и базовые смеси гидроксида натрия и карбоната натрия использовались в связи с pH 5,0 стадии выдержки после стадии выдержки DOC. Начальные прибавления бикарбоната колебались от 10-50 мМ, и карбонат натрия, прибавленный к 3N гидроксиду натрия в качестве базового титранта колебался от 0,2-1,8 М. Один процесс включал 50 мМ начального бикарбоната и NaOH использовался в качестве базового титранта. Уровни карбоната в конце ферментации колебались от 14-111 мМ. Молекулярная масса серотипа 19А колебалась от 520 до 713 кДа. Параметры ферментационного процесса и результаты показаны в Таблице 3.
Во втором исследовании (L29331-159 и -185) использовали начальные прибавления бикарбоната 15-30 мМ и базовые смеси, используя 0,4-1,0 М Na2CO3. Уровни карбоната в конце ферментации колебались от 24-62 мМ. Молекулярная масса серотипа 19A колебалась от 502 до 763 кДа. Параметры ферментационного процесса и результаты показаны в Таблице 4.
Сравнение смешанного и чистого карбонатного базового титрования ферментационного процесса
Эксперимент был выполнен, чтобы сопоставить процесс при базовой смеси (0,7 М Na2CO3/3 N NaOH) и процесс карбонатного титрования (20% раствор Na2CO3, мас./об.). Результаты (Таблица 5) подтвердили, что молекулярная масса от процесса карбонатного титрования была выше и более однородной (778, 781 кДа), чем молекулярная масса от процесса титрования базовой смесью (561-671 кДа). Выход полисахарида был также выше при процессе Na2CO3.
Пилотное производство
Осуществляли несколько экспериментальных процессов (100 л) с серотипом 19A с различными базовыми титрантами. Определение молекулярной массы осуществляли, используя исследование MALLS после полного процесса очистки, и данные предоставляли для очищенной заключительной партии. Результаты показаны в Таблице 6.
Влияние базового титранта и покрытия на молекулярную массу серотипа 19A
Выполняли ряд ферментации при 3 л процессе, чтобы определить, влияют ли базовый титрант и атмосферное покрытие на молекулярную массу. Определение молекулярной массы осуществляли, используя исследование MALLS с последующим модифицированным процессом очистки. Результаты показаны в Таблице 7.
Все публикации и заявки на патент, упомянутые описании, являются показательными, исходя из уровня специалистов в данной области, которым принадлежит это изобретение. Все публикации и заявки на патент здесь включены в отношении той же самой степени, как каждая индивидуальная публикация или заявка на патент были определенно и индивидуально указаны, чтобы быть включенными ссылкой.
Хотя предшествующее изобретение было описано в некоторых деталях посредством иллюстрации и примеров в целях ясности понимания, определенные изменения и модификации могут быть осуществлены в рамках прилагаемой формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ПОЛУЧЕНИЯ РАСТВОРА, СОДЕРЖАЩЕГО ВЫСОКОМОЛЕКУЛЯРНЫЕ ИЗОЛИРОВАННЫЕ КАПСУЛЬНЫЕ ПОЛИСАХАРИДЫ STREPTOCOCCUS PNEUMONIAE СЕРОТИПА 19А (ВАРИАНТЫ) | 2009 |
|
RU2511404C2 |
| УСКОРЕННЫЙ СПОСОБ ОЧИСТКИ ДЛЯ ПОЛУЧЕНИЯ КАПСУЛЬНЫХ ПОЛИСАХАРИДОВ STREPTOCOCCUS PNEUMONIAE | 2008 |
|
RU2516340C2 |
| Капсульные полисахариды Streptococcus pneumoniae и их конъюгаты | 2015 |
|
RU2743793C1 |
| Капсульные полисахариды Streptococcus pneumoniae и их конъюгаты | 2015 |
|
RU2688831C2 |
| КАПСУЛЬНЫЙ ПОЛИСАХАРИД Streptococcus pneumoniae И ЕГО ИММУНОГЕННЫЙ КОНЪЮГАТ | 2019 |
|
RU2803122C2 |
| КАПСУЛЬНЫЙ ПОЛИСАХАРИД Streptococcus pneumoniae И ЕГО ИММУНОГЕННЫЙ КОНЪЮГАТ | 2019 |
|
RU2839981C1 |
| МУЛЬТИВАЛЕНТНАЯ КОМПОЗИЦИЯ НА ОСНОВЕ КОНЪЮГАТА ПНЕВМОККОВЫЙ ПОЛИСАХАРИД-БЕЛОК | 2007 |
|
RU2484846C2 |
| Капсульные полисахариды Streptococcus pneumoniae и их конъюгаты | 2015 |
|
RU2805417C1 |
| МУЛЬТИВАЛЕНТНАЯ КОМПОЗИЦИЯ НА ОСНОВЕ КОНЪЮГАТА ПНЕВМОККОВЫЙ ПОЛИСАХАРИД-БЕЛОК | 2007 |
|
RU2493870C2 |
| Способ получения иммуногенного конъюгата капсульный полисахарид Streptococcus pneumoniae-белок-носитель | 2015 |
|
RU2710393C2 |
Изобретение относится к области биотехнологии и касается способа получения жидкой фракции, содержащей изолированные высокомолекулярные капсульные полисахариды Streptococcus pneumoniae. Представленный способ включает получение ферментационной культуры бактериальных клеток Streptococcus pneumoniae серотипов 19А, 6A, 19F или 6В, продуцирующих капсульные полисахариды, включающие фосфодиэфирную связь между повторяемыми единицами; введение в ферментационную культуру CO2; лизис бактериальных клеток с получением жидкой фракции, содержащей упомянутые полисахариды; выделение капсульных полисахаридов из клеточного лизата с получением жидкой фракции изолированных высокомолекулярных капсульных полисахаридов с молекулярной массой 480 кДа. Представленные изобретения могут быть использованы при производстве вакцин для иммунизации против заболеваний, вызванных Streptococcus pneumoniae. 2 н. и 12 з.п. ф-лы, 3 ил., 7 табл., 1 пр.
1. Способ получения жидкой фракции, содержащей изолированные высокомолекулярные капсульные полисахариды Streptococcus pneumoniae, где упомянутые полисахариды включают фосфодиэфирные связи между повторяемыми единицами, в котором:
a) получают ферментационную культуру бактериальных клеток Streptococcus pneumoniae, которые продуцируют капсульные полисахариды, включающие фосфодиэфирную связь между повторяемыми единицами;
b) вводят CO2 в упомянутую ферментационную культуру;
c) лизируют бактериальные клетки в упомянутой ферментационной культуре, получая клеточный лизат, где жидкая фракция содержит изолированные высокомолекулярные капсульные полисахариды Streptococcus pneumoniae и где упомянутые полисахариды включают фосфодиэфирные связи между повторяемыми единицами;
d) выделяют капсульные полисахариды Streptococcus pneumoniae из клеточного лизата, полученного на стадии c), с получением жидкой фракции изолированных высокомолекулярных капсульных полисахаридов Streptococcus pneumoniae;
таким образом, получая жидкую фракцию, содержащую изолированные высокомолекулярные капсульные полисахариды Streptococcus pneumoniae, где упомянутые полисахариды включают фосфодиэфирные связи между повторяемыми единицами; и
где упомянутыми капсульными полисахаридами Streptococcus pneumoniae являются серотип 19A, серотип 6A, серотип 19F или серотип 6B, и
где молекулярная масса упомянутых изолированных капсульных полисахаридов Streptococcus pneumoniae составляет, по крайней мере, 480 кДа.
2. Способ по п.1, где упомянутые капсульные полисахариды Streptococcus pneumoniae являются серотипом 19A.
3. Способ по п.1, где упомянутые капсульные полисахариды Streptococcus pneumoniae являются серотипом 6A.
4. Способ по п.1, где упомянутые капсульные полисахариды Streptococcus pneumoniae являются серотипом 19F.
5. Способ по п.1, где упомянутые капсульные полисахариды Streptococcus pneumoniae являются серотипом 6B.
6. Способ по п.1, где введение CO2 в упомянутую ферментационную культуру включает прибавление бикарбонатного иона к ферментационной культуре.
7. Способ по п.6, где прибавление к ферментационной культуре включает прибавление NaHCO3.
8. Способ по п.1, где введение CO2 в упомянутую ферментационную культуру включает прибавление карбонатного иона к ферментационной культуре.
9. Способ по п.8, где прибавление к ферментационной культуре включает прибавление Na2CO3.
10. Способ по п.9, где pH упомянутой ферментационной культуры находится между 6,0 и 6,6.
11. Способ по п.1, где введение CO2 в упомянутую ферментационную культуру включает первое прибавление NaHCO3 и второе прибавление Na2CO3.
12. Способ по п.1, где введение CO2 в упомянутую ферментационную культуру включает покрытие ферментационной культуры CO2.
13. Способ по п.1, где лизис Streptococcus pneumoniae в упомянутой ферментационной культуре включает прибавление деоксихолата натрия к упомянутой ферментационной культуре.
14. Жидкая фракция для производства вакцины для активной иммунизации против инвазивных заболеваний, вызванных Streptococcus pneumoniae, содержащая высокомолекулярные изолированные капсульные полисахариды Streptococcus pneumoniae, где упомянутые полисахариды включают фосфодиэфирные связи между повторяемыми единицами и где упомянутая жидкая фракция получена способом по любому одному из пп.1-13.
| US 20070184071 A1, 09.08.2007 | |||
| US 20080286838 A1, (Wyeth), 20.11.2008 | |||
| WO 2001068903 A1, 20.09.2001 |

