Испрашиваемый приоритет
По настоящей заявке испрашивается приоритет предварительной заявки США № 60/877518, поданной 27 декабря 2006 г., которая включена в настоящее описание в качестве ссылки.
Родственные заявки
Раскрытый объект заявки также относится к предварительной заявке США № 60/688872, поданной 8 июня 2005 г., заявке № 11/449919, поданной 8 июня 2006 г., и заявке PCT № PCT/US2006/22423. Эти предшествующие заявки также включены в настоящее описание в качестве ссылки в полном объеме.
Заявление о государственной поддержке
Это изобретение выполнено при поддержке правительства США грантами NIH AI39671 и CA84500. Правительство имеет определенные права на изобретение.
Область техники, к которой относится изобретение
Эта заявка относится к использованию антагонистов, конкретно к использованию антагонистов PD-1, для лечения устойчивых инфекций и опухолей.
Известный уровень техники
Иммуносупрессия иммунного ответа хозяина играет роль в устойчивой инфекции и иммуносупрессии опухолей. Устойчивые инфекции представляют собой инфекции, при которых вирус не выводится, а остается в конкретных клетках инфицированных индивидуумов. Устойчивые инфекции часто включают стадии как латентной, так и продуктивной инфекции без быстрого уничтожения или даже с возникновением чрезмерного повреждения клеток хозяина. Существует три типа устойчивого взаимодействия вирус-хозяин: латентная, хроническая и медленная инфекции. Латентная инфекция характеризуется отсутствием выявляемого инфицирующего вируса между эпизодами рецидивов заболевания. Хроническая инфекция характеризуется продолжительным присутствием инфицирующего вируса после первичной инфекции и может включать хроническое или рецидивирующее заболевание. Медленная инфекция характеризуется продолжительным инкубационным периодом с последующим прогрессированием заболевания. В отличие от латентной и хронической инфекций медленная инфекция может не начинаться с острого периода размножения вируса. В процессе устойчивых инфекций вирусный геном может либо стабильно интегрироваться в ДНК клетки, либо поддерживаться эписомально. Устойчивая инфекция возникает при инфицировании такими вирусами, как вирусы Т-клеточного лейкоза, вирус Эпштейна-Барр, цитомегаловирус, вирусы герпеса, вирус ветряной оспы, вирус кори, паповавирусы, прионы, вирусы гепатита, аденовирусы, парвовирусы и вирусы папилломы.
Механизмы, с помощью которых поддерживаются устойчивые инфекции, могут включать модуляцию экспрессии генов вируса и клетки и модификацию иммунного ответа хозяина. Реактивация латентной инфекции может быть запущена различными стимулами, включая изменения в клеточной физиологии, суперинфекцию другим вирусом и физический стресс или травму. Иммуносупрессия у хозяина часто связана с реактивацией ряда устойчивых вирусных инфекций.
Во многих исследованиях продемонстрированы дефектные иммунные ответы у больных с диагностированным раком. Идентифицирован ряд опухолевых антигенов, которые связаны с конкретными типами рака. Многие опухолевые антигены определены как характерные для солидных опухолей: MAGE 1, 2 и 3, определенные иммунологически; MART-1/Melan-A, gp100, раковоэмбриональный антиген (CEA), HER-2, муцины (т.е. MUC-1), простатспецифический антиген (PSA) и кислая фосфатаза простаты (PAP). Кроме того, вирусные белки, такие как белки вирусов гепатита B (HBV), Эпштейна-Барр (EBV) и папилломы человека (HPV), как показано, имеют важное значение для развития гепатоклеточной карциномы, лимфомы и рака шейки матки соответственно. Однако из-за иммуносупрессии у больных с диагностированным раком, врожденная иммунная система этих больных часто не способна отвечать на опухолевые антигены.
Как пассивная, так и активная иммунотерапия, как предполагается, может использоваться при лечении опухолей. Пассивный иммунитет снабжает компонентом иммунного ответа, таким как антитела или цитотоксические Т-клетки, интересующего индивидуума. Активная иммунотерапия использует терапевтический агент, такой как цитокин, антитело или химическое соединение, для активации эндогенного иммунного ответа, когда иммунная система примирована для узнавания опухоли как чужеродной. Индукция как пассивного, так и активного иммунитета является успешной при лечении конкретных типов рака.
В целом существует потребность в обеспечении безопасными и эффективными терапевтическими методами лечения заболеваний, например аутоиммунных заболеваний, воспалительных нарушений, аллергий, отторжения трансплантата, рака, иммунодефицита и других нарушений, связанных с иммунной системой.
Краткое изложение сущности изобретения
В настоящем описании раскрыто, что антиген-специфичные CD8+ Т-клетки становятся функционально толерантными («истощенными») в отношении инфекционного агента или опухолевого антигена после индукции полипептида-1 программируемой смерти (PD-1). Соответственно, с помощью снижения экспрессии или активности PD-1 иммунный ответ, специфичный в отношении инфекционного агента или опухолевых клеток, может быть увеличен. Индивидуумов с инфекциями, такими как устойчивые инфекции, можно лечить с использованием антагонистов PD-1. Индивидуума с опухолями также можно лечить с использованием антагонистов PD-1. Дополнительно индивидуумов можно лечить с помощью трансплантации терапевтически эффективного количества активированных Т-клеток, которые узнают интересующий антиген, в сочетании с терапевтически эффективным количеством антагониста PD-1.
В некоторых вариантах осуществления раскрыты способы индукции иммунного ответа на интересующий антиген у индивидуума, являющегося млекопитающим. Способ включает введение индивидууму терапевтически эффективного количества активированных Т-клеток, где Т-клетки специфически распознают интересующий антиген, и терапевтически эффективного количества антагониста полипептида-1 программируемой смерти (PD)-1. Индивидуум может представлять собой любого интересующего индивидуума, включая индивидуума с вирусной инфекцией, такой как устойчивая вирусная инфекция, или индивидуума с опухолью.
В дополнительных вариантах осуществления раскрыты способы индукции иммунного ответа на интересующий антиген у реципиента млекопитающего. Способы включают введение в контакт популяции донорских клеток от того же вида млекопитающих, включающей Т-клетки, с антигенпрезентирующими клетками (APC), и предварительный отбор интересующего антигена, где предварительно отобранный антиген представляется с помощью APC Т-клеткам с получением популяции донорских активированных Т-клеток в присутствии антагониста PD-1. Терапевтически эффективное количество популяции донорских активированных Т-клеток трансплантируется реципиенту. Реципиенту также вводят терапевтически эффективное количество антагониста PD-1.
В некоторых вариантах осуществления раскрыты способы лечения индивидуума, инфицированного патогеном, такие как способы лечения устойчивой инфекции. Способы включают введение индивидууму терапевтически эффективного количества антагониста полипептида-1 программируемой смерти (PD-1) и терапевтически эффективного количества антигенной молекулы из патогена. Примеры патогенов включают вирусные и грибковые патогены.
В дополнительных вариантах осуществления раскрыты способы лечения индивидуума с опухолью. Способы включают введение индивидууму терапевтически эффективного количества антагониста полипептида-1 программируемой смерти (PD-1) и терапевтически эффективного количества опухолевого антигена или нуклеиновой кислоты, кодирующей опухолевый антиген.
Представленные ниже и другие характеристики и преимущества станут более очевидными из последующего подробного описания некоторых вариантов осуществления, которые представлены со ссылкой на сопровождающие чертежи.
Краткое описание чертежей
Фиг.1A представляет собой гистограмму, показывающую уровни мРНК PD-1 в специфичных по DbGP33-41 и/или DbGP276-286 Т-клетках из незараженных трансгенных мышей, мышей, иммунизированных лимфоцитарным вирусом хориоменингита (LCMV) Армстронга (приблизительно через 30 дней после инфицирования), или мышей с недостаточностью CD-4, инфицированных LCMV-C1-13 (приблизительно через 30 дней после инфицирования), по результатам измерения с помощью метода генных матриц. Фиг.1B представляет собой серию изображений эксперимента с проточной цитометрией, демонстрирующую поверхностную экспрессию PD-1 на содержащих CD8+тетрамер+ Т-клетках у мышей, иммунизированных LCMV Армстронга, и мышей с недостаточностью CD-4, инфицированных LCMV-C1-13, приблизительно через 60 дней после инфицирования. CD8+ Т-клетки с анергией экспрессируют высокие уровни полипептида PD-1 на клеточной поверхности приблизительно через 60 дней после хронического инфицирования вирусом LCMV-C1-13 (помечено «хроническое»), но специфичные в отношении вируса CD8+ Т-клетки не экспрессируют полипептид PD-1 после освобождения от острой инфекции LCMV Армстронга (помечено «иммунные»). Фиг.1C представляет собой серию изображений эксперимента с проточной цитометрией, демонстрирующую присутствие PD-L1 на спленоцитах от хронически инфицированных и неинфицированных мышей. Это показывает, что экспрессия PD-L1 является наиболее высокой на спленоцитах, которые инфицированы вирусом.
Фиг.2A представляет собой серию диаграмм рассеяния, демонстрирующих, что когда мышей, инфицированных C1-13, лечат с 23 по 27 день после инфицирования, наблюдается приблизительно 3-кратное увеличение количества специфичных по DbNP396-404 и специфичных по DbGP33-41 CD8+ Т-клеток по сравнению с контрольными, не подвергаемыми лечению. Для того чтобы определить любые изменения в функции, измеряли продукцию IFN-γ и TNF-α в ответ на 8 различных эпитопов LCMV. Фиг.2B представляет собой диаграмму рассеяния, демонстрирующую, что когда все известные специфичные маркеры CD8+ Т-клеток измерены, наблюдается 2, 3-кратное увеличение суммарного количества специфичных по LCMV СВ8+ Т-клеток. Фиг.2C представляет собой серию схем проточной цитометрии, демонстрирующую продукцию IFN-γ и TNF-α в ответ на восемь различных эпитопов LCMV. Фиг.2D представляет собой диаграмму рассеяния, демонстрирующую, что у мышей, подвергнутых лечению, более высокое количество вирус-специфичных CD8+ Т-клеток обладает способностью продуцировать TNF-α. Фиг.2E представляет собой серию столбчатых гистограмм, демонстрирующих, что блокада PD-L1 также ведет к повышенному вирусному контролю в селезенке, печени, легком и сыворотке.
Фиг.3A представляет собой график, показывающий увеличение специфичных по DbGP33-41 и DbGP276-286 Т-клеток (отмечено «GP33» и «GP276») у мышей с недостаточностью CD-4, инфицированных C1-13, подвергавшихся лечению анти-PD-L1 (отмечено «αPD-L1») с 46 дня по 60 день после инфицирования по сравнению с контрольными (отмечено «untx»), что указывает на то, что мыши, подвергавшиеся лечению анти-PD-L1, содержали приблизительно в 7 раз больше специфичных по DbGP276-286 CD8+ Т-клеток селезенки и приблизительно в 4 раза больше специфичных по DbGP33-41 CD8+ Т-клеток селезенки, чем мыши, не подвергавшиеся лечению. Фиг.3B представляет собой серию изображений эксперимента с проточной цитометрией, демонстрирующую повышенную встречаемость специфичных по DbGP33-41 и DbGP276-286 CD8+ Т-клеток в селезенке у мышей с недостаточностью CD-4, инфицированных C1-13, подвергавшихся лечению анти-PD-L1 (отмечено «αPD-L1 Tx») с 46 дня по 60 день после инфицирования по сравнению с контролем (отмечено «untx»). Фиг.3C представляет собой серию изображений эксперимента с проточной цитометрией, демонстрирующую повышенную пролиферацию специфичных по DbGP276-286 CD8+ Т-клеток у мышей, подвергавшихся лечению анти-PD-L1, при измерении по включению BrdU и экспрессии Ki67. Фиг.3D представляет собой график, демонстрирующий, что мыши, обладающие высокими уровнями экспансии CD8+ Т-клеток, дают заметный ответ в мононуклеарных клетках периферической крови (PBMC), что показано путем сравнения специфичных по DbGP276-286 CD8+ Т-клеток в PBMC по сравнению со специфичными по DbGP276-286 CD8+ Т-клетками в селезенке.
Фиг.4A представляет собой серию графиков, показывающих увеличение продукции IFN-γ специфичными по DbGP276-286 и DbGP33-41 CD8+ Т-клетками у мышей, подвергавшихся лечению анти-PD-L1, по сравнению с контрольными. У мышей, подвергавшихся лечению анти-PD-L1, также наблюдалась более высокая встречаемость специфичных по DbNP396-404, KbNP205-212, DbNP166-175 и DbGP92-101 CD8+ Т-клеток. Фиг.4B представляет собой график, показывающий, что у мышей, подвергавшихся лечению анти-PD-L1, 50% специфичных по DbGP276-286 CD8+ Т-клеток продуцируют IFN-γ по сравнению с 20% специфичных по DbGP276-286 CD8+ Т-клеток у контрольных мышей. Фиг.4C представляет собой серию изображений эксперимента с проточной цитометрией, демонстрирующую, что у подвергавшихся лечению анти-PD-L1 хронически инфицированных мышей продуцируются более высокие уровни TNF-α, чем у иммунизированных мышей, инфицированных вирусом LCMV Армстронга. Фиг.4D представляет собой график, демонстрирующий, что лечение анти-PD-L1 мышей, инфицированных LCMV-C1-13, восстанавливает литическую активность ex vivo вирус-специфичных Т-клеток по сравнению с не подвергавшимися лечению инфицированными мышами при измерении с использованием теста высвобождения 51Cr. Фиг.4E представляет собой серию графиков, демонстрирующих снижение титра вирусов в различных органах после лечения α-PD-L1 мышей, инфицированных LCMV-C1-13. Титры вирусов снижались приблизительно в 3 раза в селезенке, в 4 раза в печени, в 2 раза в легком и в 2 раза в сыворотке после лечения анти-PD-L1 в течение 2 недель по сравнению с не подвергавшимися лечению мышами.
Фиг.5A представляет собой серию изображений эксперимента с проточной цитометрией, демонстрирующую поверхностную экспрессию PD-1 с использованием 10 тетрамеров ВИЧ, специфичных по доминантным эпитопам, направленным на таксон С хронической инфекции ВИЧ. Проценты отражают процент тетрамер+ клеток, которые являются PD-1+. Фиг.5B представляет собой серию графиков, демонстрирующих, что процент и MFI PD-1 существенно положительно регулируется на ВИЧ-специфичных CD8+ Т-клетках по сравнению с суммарной популяцией CD8+ Т-клеток (p<0,0001) при антиретровирусной терапии не подвергнутых лечению индивидуумов, и PD-1 увеличивается на суммарной популяции CD8+ Т-клеток у ВИЧ-инфицированных по отношению к ВИЧ-серонегативному контролю (p=0,0033 и p<0,0001 соответственно). В анализ включены 120 тетрамерных штаммов ВИЧ от 65 ВИЧ-инфицированных индивидуумов и 11 ВИЧ-серонегативных контролей. Фиг.5C представляет собой серию графиков, показывающих средний процент и MFI экспрессии PD-1 на тетрамер+ клетках по специфичности эпитопов. Фиг.5D представляет собой диаграмму, изображающую вариацию процента PD-1+ клеток на различных специфичных по эпитопам популяциях у индивидуумов с множественными определяемыми ответами. Горизонтальные отметки указывают медиану процента ВИЧ тетрамер+ PD-1+ клеток у каждого индивидуума.
Фиг.6А представляет собой серию графиков, показывающих, что не наблюдается корреляции между количеством ВИЧ-специфичных CD8+ Т-клеток при измерении с помощью окрашивания тетрамеров и вирусной нагрузкой плазмы, в то же время существует положительная корреляция между как процентом, так и MFI PD-1 на тетрамер+ клетках и вирусной нагрузкой плазмы (p=0,0013 и p<0,0001 соответственно). Фиг.6B представляет собой серию графиков, показывающих, что не наблюдается корреляции между количеством ВИЧ-тетрамер+ клеток и количеством CD4, в то же время существует обратная корреляция между процентом и MFI PD-1 на ВИЧ тетрамер+ клетках и количеством CD4 (p=0,0046 и p=0,015 соответственно). Фиг.6C представляет собой серию графиков, показывающих, что процент и MFI PD-1 на суммарной популяции CD8+ Т-клеток положительно коррелирует с вирусной нагрузкой плазмы (p=0,0021 и p<0,0001 соответственно). Фиг.6D представляет собой серию графиков, показывающих, что процент и MFI экспрессии PD-1 на суммарной популяции CD8+ Т-клеток отрицательно коррелирует с количеством CD4 (p=0,0049 и p=0,0006 соответственно).
Фиг.7A представляет собой серию изображений эксперимента с проточной цитометрией, демонстрирующую типичное фенотипическое окрашивание B*4201 TL9-специфичных CD8+ Т-клеток от индивидуума SK222, у которого 98% B*4201 TL9-специфичных CD8+ Т-клеток являются PD-1+. Фиг.7B представляет собой график, иллюстрирующий суммарные фенотипические данные от индивидуумов, у которых >95% ВИЧ-специфичных CD8+ Т-клеток являются PD-1+. От семи до 19 образцов было проанализировано для каждого из указанных фенотипических маркеров. Горизонтальная отметка указывает медиану процента тетрамер+ PD-1+ клеток, которые были позитивными для указанного маркера.
Фиг.8A представляет собой серию изображений эксперимента с проточной цитометрией, демонстрирующую типичные данные теста на пролиферацию от B*4201 позитивного индивидуума. После 6-дневной стимуляции пептидом процент B*4201 TL9-специфичных CD8+ Т-клеток повышался от 5,7% до 12,4% в присутствии анти-PD-L1 блокирующего антитела. Фиг.8B представляет собой линейный график, отражающий данные теста на суммарную пролиферацию, указывающий на значительное увеличение пролиферации ВИЧ-специфичных CD8+ Т-клеток в присутствии анти-PD-L1 блокирующего антитела (n=28, p=0,0006, парный t-тест). Фиг.8C представляет собой гистограмму, показывающую различные эффекты блокады PD-1/PD-L1 на пролиферацию ВИЧ-специфичных CD8+ Т-клеток, основанную на индивидуальных больных. Белые столбики указывают степень повышения тетрамер+ клеток в присутствии только пептида, черные столбики указывают степень повышения тетрамер+ клеток в присутствии пептида плюс анти-PD-L1 блокирующее антитело. Индивидуумы, у которых выполнялись тесты CFSE для более одного эпитопа, указаны символами в виде звездочки, квадрата или треугольника.
Фиг.9a-9d представляют собой диаграмму и серию графиков, демонстрирующих синергичный эффект терапевтической вакцины, объединенной с блокатором PD-L1, на частоту антиген-специфичных CD8-Т-клеток и титр вирусов у хронически инфицированных мышей. Фиг.9a представляет собой схематическую диаграмму экспериментального протокола. Инфицированных клоном-13 LCMV (CL-13) мышей вакцинировали вирусом коровьей оспы дикого типа (VV/WT) или вирусом коровьей оспы, экспрессирующим эпитоп LCMV GP33-41 (VV/GP33), через 4 недели после инфицирования. В то же время мышей лечили 5 раз каждые три дня анти-PD-L1 или без него. Фиг.9b представляет собой серию изображений эксперимента с проточной цитометрией, демонстрирующую частоту GP33- и GP276-специфичных CD8-Т-клеток в PBMC через 1 неделю после лечения. Число представляет собой частоту тетрамер-позитивных клеток по отношению к CD8-Т-клеткам. Данные представляют результаты трех экспериментов. Фиг.9c-9d представляют собой графики частоты GP33- и GP276-специфичных CD8-Т-клеток (фиг.9c) и титры вирусов (фиг.9d) в крови после лечения. Изменения количества тетрамер-позитивных CD8-Т-клеток и титров вирусов прослеживали в крови с помощью окрашивания тетрамеров и анализа бляшкообразования соответственно в указанные временные точки. Количество тетрамер-позитивных CD8-Т-клеток и титры вирусов представлены у мышей индивидуально (верхние четыре панели) и в сводном виде (нижняя панель) после введения VV/WT или VV/GP33 (прямая линия) и лечения анти-PD-L1 (затененная область). Пунктирные линии представляют собой предел определения вирусов. Объединены результаты трех экспериментов.
Фиг.10a-10d представляют собой графики и цифровые изображения, показывающие повышение антиген-специфичных CD8-Т-клеток и повышенный контроль вирусов в различных тканях мышей, получавших терапевтическую вакцину в сочетании с блокадой PD-L1. Фиг.10a представляет собой серию изображений эксперимента с проточной цитометрией, демонстрирующую частоту GP33-специфичных CD8-Т-клеток в различных тканях через 4 недели после лечения. Число представляет собой частоту тетрамер-позитивных клеток по отношению к CD8-Т-клеткам. Данные представляют результаты двух экспериментов. Фиг.10b представляет собой график зависимости числа GP33-специфичных CD8-Т-клеток в различных тканях через 4 недели после лечения. Фиг.10c представляет собой набор гистограмм, демонстрирующих титры вирусов в указанных тканях через 2 (закрашенные столбики) и через 4 (незакрашенные столбики) недели после лечения. Пунктирные линии представляют собой предел определения вирусов. N=6 мышей на группу. Объединены результаты двух экспериментов. Фиг.10d представляет собой цифровое изображение иммуноокрашивания селезенки антигенами aLCMV (красный) через 2 недели после лечения. Увеличение ×20.
Фиг.11a-11d представляют собой диаграммы и графики, показывающие усиленное восстановление функции истощенных CD8-Т-клеток с помощью терапевтической вакцины в сочетании с блокадой PD-L1. Фиг.11a представляет собой серию изображений эксперимента с проточной цитометрией, демонстрирующую продукцию IFN-γ и дегрануляцию спленоцитов вакцинированных мышей через 4 недели после лечения. Спленоциты стимулировали указанными пептидами в присутствии αCD107a/b антител и затем совместно окрашивали на IFN-γ. Представленные диаграммы ограничены окном для CD8-Т-клеток и представляют два независимых эксперимента. Фиг.11b представляет собой график, показывающий процент IFN-γ+CD107+ клеток по отношению к CD8-Т-клеткам, конкретно для каждого из пептидов LCMV из фиг.11a, суммированных для многих мышей (n=6 для каждого ответа). Суммированы результаты двух экспериментов. Фиг.11c представляет собой серию диаграмм, демонстрирующих продукцию TNF-α CD8-Т-клетками, способными продуцировать IFN-γ у вакцинированных мышей. После стимуляции спленоцитов пептидом GP33-41 или GP276-286 происходила сортировка с окном для продуцирующих IFN-γ CD8-Т-клеток, и затем строилась диаграмма IFN-γ (ось x) против TNF-α (ось y). Верхние и нижние цифры на диаграммах указывают частоту TNF-α+ клеток среди IFN-γ+ клеток и среднюю интенсивность флуоресценции (MFI) IFN-γ+ клеток, соответственно. Данные представляют результаты двух независимых экспериментов. Фиг.11d представляет собой график, показывающий процент TNF-α+ клеток по отношению к TNF-γ+ клеткам для пептида GP33-41 или GP276-286 из фиг.11c, суммированный для многих мышей (n=6 для каждого ответа).
Фиг.12a-12b представляют собой серию диаграмм, показывающих, что действие терапевтической вакцины в сочетании с блокадой PD-L1 изменяет фенотип антиген-специфичных CD8-Т-клеток хронически инфицированных мышей. Фиг.12a представляет собой серию диаграмм, показывающих фенотип GP33 тетрамер-специфичных CD8-Т-клеток в PBMC в указанные временные точки после лечения. Гистограммы ограничивались окном для GP33+ CD8-Т-клеток. Частота популяции, экспрессирующей высокий уровень CD27 или CD127, указана в виде процента на диаграммах. Числа на гистограммах для Granzyme B представляют MFI экспрессии. Данные представляют результаты трех независимых экспериментов. Фиг.12b представляет собой серию диаграмм, показывающих фенотипические изменения GP33 тетрамер-специфичных CD8-Т-клеток в различных тканях через 4 недели после лечения. Гистограммы ограничивались окном для GP33+ CD8-Т-клеток. Частота популяции, экспрессирующей высокий уровень CD127 или PD-1, указана в виде процента на диаграммах. Числа на гистограммах для Granzyme B и Bcl-2 представляют MFI экспрессии. Данные представляют результаты двух независимых экспериментов.
Фиг.13a-13e представляют собой схематическую диаграмму, диаграммы и графики, демонстрирующие синергичный эффект терапевтической вакцины в сочетании с блокатором PD-L1 на восстановление функции «бесполезных» истощенных CD8 Т-клеток. Фиг.13a представляет собой схематическую диаграмму протокола. У мышей истощали CD4 Т-клетки и затем их инфицировали клоном-13 LCMV. Некоторых мышей вакцинировали вирусом коровьей оспы дикого типа (VV/WT) или вирусом коровьей оспы, экспрессирующим эпитоп LCMV GP33-41 (VV/GP33), через 7 недели после инфицирования. В то же время мышей лечили 5 раз каждые три дня αPD-L1 или его изотипом. Через две недели после первоначального лечения антителами мышей забивали для анализа. Фиг.13b представляет собой серию изображений эксперимента с проточной цитометрией и гистограмму, демонстрирующие частоту GP33-специфичных CD8-Т-клеток в указанных тканях через 4 недели после лечения. Число представляет собой частоту GP33 тетрамер-позитивных клеток по отношению к CD8-Т-клеткам. Суммирована также частота GP33-специфичных клеток по отношению к CD8-Т-клеткам в различных тканях через 2 недели после лечения. Фиг.13c представляет собой серию изображений эксперимента с проточной цитометрией и гистограмму, демонстрирующие результаты экспериментов, в которых спленоциты стимулировали пептидом GP33 в присутствии антител αCD107a/b и затем совместно окрашивали на IFN-γ. Представленные диаграммы ограничивались окном для CD8-Т-клеток. Процент IFN-γ+CD107+ клеток по отношению к CD8-Т-клеткам, специфичным для пептида GP33, суммирован для многих мышей. Фиг.13d представляет собой гистограмму процента IFN-γ+ клеток после стимуляции пептидом GP33 по отношению к клеткам, позитивным по Db-ограниченному GP33-41 тетрамеру, суммированному для многих мышей. Фиг.13e представляет собой гистограмму титров вирусов в указанных тканях через 2 недели после лечения. Все графики представляют результаты двух экспериментов, и все суммированные результаты двух экспериментов объединены (n=6 мышей на группу).
Фиг.14a-14b представляют собой диаграммы и графики, показывающие, что блокада сигнального пути PD1/PD-L1 повышает суммарное количество специфичных для антигена Т-клеток с последующим адоптивным переносом мышам - врожденным носителям. Целые спленоциты адаптивно переносили мышам - врожденным носителям с или без лечения анти-PD-L1. Фиг.14a представляет собой серию диаграмм типичной проточной цитометрии при конкретных временных точках с ограничением окна для CD8+ Т-клеток. Фиг.14b представляет собой графики, демонстрирующие кинетику экспансии Db GP33-специфичных CD8 Т-клеток в периферической крови из двух независимых экспериментов (n=4 животных на группу).
Фиг.15a-15e представляют собой диаграммы и графики, показывающие, что блокада пути PD-1/PDL1 после адоптивной Т-клеточной иммунотерапии повышает продукцию цитокинов антиген-специфичными CD8 Т-клетками. Спленоциты выделяли на 17 день после переноса и анализировали на экспрессию цитокинов при стимуляции антигенным пептидом. Фиг.15a представляет собой серию диаграмм типичной проточной цитометрии, показывающих экспрессию IFNγ, определяемую с помощью внутриклеточного окрашивания цитокина через 5 часов после стимуляции определенными эпитопами CD8 или в контролях без пептида. Фиг.15b и 15d представляют собой диаграммы, показывающие двойную экспрессию TNFα или 107ab и IFNγ (статистические значения в квадрантах представляют собой процент окна для CD8). Фиг.15c и 15e представляют собой гистограммы процента клеток, продуцирующих IFNγ и также продуцирующих TNFα или 107ab (n=3 животных на группу).
Фиг.16A-16B представляют собой график и диаграммы, показывающие повышенные уровни клеток, секретирующих антитела, у мышей, инфицированных клоном-13 LCMV. Суммарные уровни ASC измеряли у мышей, хронически инфицированных LCMV, после лечения αPD-L1 с помощью ELISPOT и окрашивания CD138. Фиг.16A представляет собой график суммарного количества ASC селезенки, объединенные результаты трех независимых экспериментов. Фиг.16B представляет собой серию диаграмм, демонстрирующих, что увеличение клеток, секретирующих антитела (ASC), в селезенке может быть измерено с помощью маркера CD138. Показана одна типичная диаграмма, ASC характеризуются как CD138+ и B220 низкие/промежуточные (окно ограничено лимфоцитами).
Фиг.17 представляет собой график, демонстрирующий, что лечение анти-PD-L1 мышей, хронически инфицированных LCMV, не ведет к повышенным уровням ASC костного мозга. Суммарное количество ASC подсчитано в селезенке и костном мозге мышей, хронически инфицированных LCMV, через 14 дней после лечения анти(α)PD-L1 с помощью ELISPOT. Линия представляет собой среднее геометрическое в группе.
Фиг.18 представляет собой график, демонстрирующий, что совместное введение αPD-L1 и αCTLA-4 ведет к синергичному увеличению ASC селезенки. Мышам, хронически инфицированным LCMV, вводили αPD-L1, αCTLA-4 или оба пептида в течение 14 дней и ASC в селезенке подсчитывали с помощью ELISPOT. Линия представляет собой среднее геометрическое в группе.
Фиг.19A-19B представляют собой диаграммы, показывающие повышенную пролиферацию и активность герминативного центра В-клеток и CD4 Т-клеток у мышей, подвергавшихся лечению αPD-L1. Фиг.19A представляет собой диаграмму анализа с помощью проточной цитометрии CD4 Т-клеток и В-клеток, показывающую повышенные уровни Ki-67 после лечения αPD-L1. Результаты ограничены окнами либо для CD4, либо для В-клеток, как показано вверху каждой колонки. Фиг.19B представляет собой серию диаграмм, показывающих увеличенную частоту В-клеток, экспрессирующих PNA, и высокие уровни FAS, что указывает на повышенную активность герминативного центра у мышей, подвергавшихся лечению αPD-L1. Диаграммы представляют собой одну типичную диаграмму, суммирующую результаты двух отдельных экспериментов.
Фиг.20A-20C представляют собой диаграммы и графики, показывающие экспрессию PD-1 на субпопуляциях CD8 и CD4 Т-клеток. Фиг.20A представляет собой серию изображений эксперимента с проточной цитометрией, демонстрирующую поверхностную совместную экспрессию PD-1 и различных фенотипических маркеров среди CD8+/CD3+ лимфоцитов крови. Фиг.20B представляет собой серию графиков процента различных CD8+/CD3+ и (D) CD4+/CD3+ субпопуляций Т-клеток, которые экспрессируют PD-1. Горизонтальные линии указывают средний процент PD-1 на Т-клетках, которые позитивны (незакрашенные кружочки) и негативны (закрашенные треугольники) по указанному маркеру. Фиг.20C представляет собой серию графиков, представляющих фенотипические данные CD4+ Т-клеток, экспрессирующих PD-1, из одной субпопуляции.
Фиг.21A-B представляют собой диаграммы и графики, показывающие, что PD-1 экспрессируется на более высоком уровне на CD8 Т-клетках, специфичных для хронических инфекций. Фиг.21A представляет собой серию изображений эксперимента с проточной цитометрией, демонстрирующую типичное окрашивание на PD-1 CD8 Т-клеток, специфичных для вируса Эпштейна-Барр (EBV), цитомегаловируса (CMV), вируса гриппа и вируса коровьей оспы. Указано среднее геометрическое интенсивности флуоресценции (GMFI) экспрессии PD-1 среди тетрамер+ клеток. Фиг.21B представляет собой график, показывающий суммарные данные о GMFI PD-1 на CD8 Т-клетках от здоровых добровольцев (n=35), специфичных для EBV, CMV, вируса гриппа и вируса коровьей оспы.
Фиг.22A-C представляют собой диаграммы и графики, показывающие, что блокада анти-PD-L1 повышает пролиферацию in vitro CD8 Т-клеток, специфичных для хронических инфекций. Фиг.22A представляет собой серию изображений эксперимента с проточной цитометрией, демонстрирующую лимфоциты, которые были помечены CFSE, затем культивировались в течение 6 дней в указанных условиях. В изображениях отражено типичное окрашивание от индивидуумов, позитивных в отношении EBV и CMV. Фиг.22B представляет собой гистограмму ответов, специфичных для антигенов EBV, CMV, вируса гриппа и вируса коровьей оспы, после блокады с помощью анти-PD-L1 блокирующего антитела. Столбики указывают степень увеличения тетрамер+ клеток в присутствии пептида плюс анти-PD-L1 блокирующее антитело по сравнению только с пептидом. Фиг.22C представляет собой линейный график, показывающий взаимоотношения между степенью увеличения тетрамер+ клеток после блокады анти-PD-L1 антителом и экспрессией PD-1 (перед культивированием).
Фиг.23B-23C представляют собой диаграммы и графики, показывающие, что CD8+ Т-клетки, специфичные для вируса гепатита С (HCV), экспрессируют PD-1 при хронической инфекции HCV человека. Фиг.23A представляет собой типичные диаграммы от пяти больных с хронической инфекцией HCV, демонстрирующие экспрессию PD-1 на CD8+ Т-клетках, специфичных для HCV. Цифры жирным шрифтом показывают частоту экспрессии PD-1 (ось x) на CD8+ Т-клетках, специфичных для HCV (ось y). На диаграммах цифры курсивом показывают частоту тетрамер-позитивных клеток среди суммарных CD8+ Т-клеток. На оси y 1073 и 1406 указывают специфичные эпитопы HCV тетрамера. Больные указаны как «Pt» с последующим номером больного. Клетки ограничивали окном для CD8+ лимфоцитов. Диаграммы построены в логарифмическом масштабе. Фиг.23B представляет собой сравнение экспрессии PD-1 на CD8+ Т-клетках от здоровых доноров (CD8 здоровые), больных, инфицированных HCV (CD8 HCV), и на CD8+ Т-клетках, специфичных для HCV (HCV tet+). Фиг.23C представляет собой график экспрессии PD-1 на CD8+ Т-клетках, специфичных для вируса гриппа (Flu tet+) от инфицированных HCV (HCV+) и здоровых доноров (здоровые), в сравнении с экспрессией PD-1 на CD8+ Т-клетках, специфичных для HCV (HCV tet+). Для сравнения различий экспрессии PD-1 у одного и того же больного на суммарных CD8+ Т-клетках по отношению к CD8+ Т-клеткам, специфичным для HCV, использовали непарный t-тест.
Фиг.24A-24D представляют собой диаграммы и графики, показывающие, что частота CD8+ Т-клеток, экспрессирующих PD-1, в печени выше, чем в периферической крови. Фиг.24A представляет собой типичные диаграммы от пяти больных с хронической инфекцией HCV, демонстрирующие экспрессию PD-1 на суммарных CD8+ Т-клетках из периферической крови против них в печени. Цифры жирным шрифтом на диаграммах показывают частоту клеток с экспрессией PD-1 среди суммарных CD8+ Т-клеток в окне, ограниченном для лимфоцитов. Диаграммы построены в логарифмическом масштабе. Фиг.24B представляет собой сравнение экспрессии PD-1 на CD8+ Т-клетках из периферической крови против них в печени у больных, хронически инфицированных HCV. Для сравнения различия экспрессии PD-1 у одних и тех же больных использовали парный t-тест. Фиг.24C представляет собой сравнение экспрессии PD-1 на CD8+ эффекторных клетках памяти (TEM) из периферической крови против них в печени. Субпопуляции памяти были идентифицированы по дифференциальной экспрессии CD62L и CD45RA. Цифры жирным шрифтом наверху диаграмм указывают частоту клеток в каждом квадранте. Клетки ограничивали окном для CD8+ лимфоцитов. Субпопуляция TEM была ограничена окном (боксами), и экспрессия PD-1 представлена на гистограммах ниже. Пунктирная линия отражает экспрессию PD-1 на наивных CD8+ Т-клетках (используемых в качестве негативной популяции). Числа на гистограммах указывают частоту клеток, экспрессирующих PD-1. Сравнение частоты экспрессии PD-1 на CD8+ TEM клетках десяти больных с хронической инфекцией HCV суммировано ниже гистограмм. Для сравнения различия экспрессии PD-1 на CD8+ TEM из периферической крови и печени одного и того же больного использовали парный t-тест. Фиг.24D представляет собой типичные диаграммы от двух больных с хронической инфекцией HCV, показывающие различие в экспрессии CD127 на суммарных CD8+ Т-клетках из периферической крови и печени. Цифры жирным шрифтом указывают на частоту экспрессии CD127 на суммарных CD8+ Т-клетках. Клетки ограничены окном для CD8+ лимфоцитов. Диаграммы представлены в логарифмическом масштабе. Суммарное сравнение экспрессии CD127 на суммарных CD8+ Т-клетках в периферической крови по отношению к ним в печени показано ниже диаграмм FACS. Для статистического анализа использовали парный t-тест.
Фиг.25 представляет собой серию графиков и диаграмм, демонстрирующих, что CD8+ Т-клетки, специфичные для HCV, в печени экспрессируют истощенный фенотип. Типичные диаграммы экспрессии PD-1 и CD127 на CD8+ Т-клетках, специфичных для HCV, из периферической крови и печени двух больных хронической инфекцией HCV. Первый ряд диаграмм иллюстрирует HCV тетрамер-позитивную популяцию (боксы). Цифры над боксами указывают частоту тетрамер-позитивных клеток среди CD3+ лимфоцитов. Специфичность эпитопа тетрамера HCV указана на оси y (1073). Второй и третий ряды диаграмм демонстрируют экспрессию PD-1 и CD127 на CD8+ Т-клетках, специфичных для HCV, из периферической крови и печени двух больных хронической инфекцией HCV. Цифры жирным шрифтом указывают частоту экспрессии PD-1 или CD127 на CD8+ Т-клетках, специфичных для HCV. Диаграммы представлены в логарифмическом масштабе и ограничены окном для CD3+ CD8+ лимфоцитов. Ниже FACS диаграмм показаны суммарные данные по сравнению экспрессии PD-1 (слева) и экспрессии CD127 (справа) на суммарных CD8+ Т-клетках по сравнению с CD8+ Т-клетками, специфичными для HCV, из периферической крови (HCV tet+ PBMC) и по сравнению с CD8+ Т-клетками, специфичными для HCV, из печени. Для сравнения экспрессии у одного и того же больного использовали парные t-тесты.
Фиг.26 представляет собой серию диаграмм, демонстрирующих, что блокада пути PD-1/PD-L1 повышает экспансию антиген-стимулированных Т-клеток, специфичных для HCV. PBMC, меченные CFSE, от двух отдельных больных с HLA-A2 стимулировали с использованием родственного пептидного антигена в течение 6 дней в присутствии ИЛ-2 и анти-PD-L1 антитела (верхняя панель) или анти-PD-1 антитела (нижняя панель). Показан также нестимулированный контроль. В каждом квадранте представлен процент пролиферирующих CFSE-низко- и CFSE-высокоспецифичных для HCV HLA-A2+ CD8+ Т-клеток.
Фиг.27A-27D представляют собой диаграммы и графики, демонстрирующие повышенную экспрессию PD-1 на CD8 Т-клетках, специфичных для вируса иммунодефицита обезьян (SIV) после инфицирования SIV239. Фиг.27A представляет собой диаграмму, показывающую экспрессию PD-1 на суммарных CD8 Т-клетках от нормальной макаки. Фиг.27B представляет собой диаграмму, показывающую экспрессию PD-1 на суммарных и специфичных для капсидных белков SIV CD8 Т-клетках от макаки, инфицированной SIV239. Анализ выполнен на PBMC через 12 недель после инфицирования SIV. Фиг.27C представляет собой график, демонстрирующий сумму PD-1 позитивных клеток от суммарных и специфичных для SIV CD8 Т-клеток нормальных и инфицированных SIV макак. Данные для макак, инфицированных SIV, представлены через 12 недель после инфицирования. Фиг.27D (последняя панель) представляет собой график, демонстрирующий среднюю суммарную интенсивность флуоресценции (MFI) экспрессии PD-1 на суммарных и специфичных для SIV CD8 Т-клеток нормальных и инфицированных SIV макак.
Фиг.28A-28B представляют собой диаграмму и график соответственно, демонстрирующие, что блокада PD-1 in vitro ведет к увеличенной экспансии SIV-специфичных CD8 Т-клеток. PBMC от Mamu A*01 позитивных макак, которые были инфицированы SHIV89.6P, стимулировали пептидом P11C (0,1 мкг/мл) в отсутствие и в присутствии анти-PD-1 блокирующего Ab (10 мкг/мл) в течение 6 дней. Через три дня стимуляции добавляли ИЛ-2 (50 единиц/мл). По окончании стимуляции клетки окрашивали на поверхностную экспрессию CD3, CD8 и Gag-CM9 тетрамера. Нестимулированные клетки (nostim) служили в качестве негативных контролей. Клетки ограничивали окном для лимфоцитов, основываясь больше на рассеянии, чем на CD3, и анализировали на экспрессию CD8 и тетрамера. Фиг.28A представляет собой типичные FASC диаграммы. Цифры на диаграмме указывают частоту тетрамер-позитивных клеток в виде процента от суммарных CD8 Т-клеток. Фиг.28B представляет собой график, демонстрирующий суммарные данные от шести макак. Анализы выполнялись с использованием клеток, полученных через 12 недель после инфицирования. Степень повышения рассчитывали как отношение частоты тетрамер-позитивных клеток в P11C стимулированных культурах и в нестимулированных клетках.
Фиг.29 представляет собой серию диаграмм, показывающих кинетику экспрессии PD-L1, PD-L2 и PD-1 на различных клеточных типах после инфицирования LCMV. Мышей инфицировали 2×106 БОЕ клона-13 (CL-13). Экспрессия PD-L1, PD-L2 и PD-1 на различных клеточных типах показана в виде гистограммы в указанные временные точки после инфицирования. Представлена средняя интенсивность флуоресценции (MFI) для экспрессии PD-1 на указанном типе клеток.
Перечень последовательностей
Последовательности нуклеиновых кислот и аминокислотные последовательности, перечисленные в прилагаемом перечне последовательностей, показаны с использованием стандартных буквенных аббревиатур для нуклеотидных оснований и трехбуквенного кода для аминокислот, как определено в 37 C.F.R. 1.822. Показана только одна цепь последовательности каждой нуклеиновой кислоты, но комплементарная цепь подразумевается включенной путем любой ссылки на представленную цепь. В прилагаемом перечне последовательностей:
SEQ ID NO:1 представляет собой пример аминокислотной последовательности PD-1 человека.
SEQ ID NO:2 представляет собой пример аминокислотной последовательности PD-1 мыши.
SEQ ID NO:3 представляет собой пример аминокислотной последовательности PD-L1 человека.
SEQ ID NO:4 представляет собой пример аминокислотной последовательности PD-L2 человека.
SEQ ID NO:5-12 представляют собой примеры аминокислотных последовательностей областей каркаса человека.
SEQ ID NO:13-35 представляют собой примеры аминокислотных последовательностей антигенных пептидов.
SEQ ID NO:36-43 представляют собой аминокислотные последовательности пептидов главного комплекса гистосовместимости.
SEQ ID NO:44 и SEQ ID NO:45 представляют собой аминокислотные последовательности эпитопов Т-клеток.
SEQ ID NO:46 представляет собой примеры аминокислотных последовательностей вариантов PD-L2 человека.
SEQ ID NO:47-52 представляют собой примеры аминокислотных последовательностей антигенных пептидов.
Подробное описание
Это раскрытие относится к использованию антагонистов PD-1 для индукции иммунного ответа, такого как направленный на опухоль или хроническую вирусную инфекцию.
Термины
Если не указано иначе, технические термины используются в соответствии с общепринятым применением. Определения общих терминов молекулярной биологии можно найти в Benjamin Lewin, Genes V, опубликованной Oxford University Press, 1994 (ISBN 0-19-854287-9); Kendrew et al. (eds.), The Encyclopedia of Molecular Biology, опубликованной Blackwell Science Ltd., 1994 (ISBN 0-632-02182-9); и Robert A. Meyers (ed.), Molecular Biology and Biotechnology: a Comprehensive Desk Reference, опубликованной VCH Publishers, Inc., 1995 (ISBN 1-56081-569-8).
Для облегчения обзора различных вариантов осуществления этого раскрытия предлагаются следующие объяснения конкретных терминов.
Изменение уровня продукции или экспрессии: изменение путем либо увеличения, либо снижения уровня продукции или экспрессии последовательности нуклеиновой кислоты или аминокислотной последовательности (например, полипептида, миРНК, микроРНК, мРНК, гена) по сравнению с контрольным уровнем продукции или экспрессии.
Антисмысловые, смысловые молекулы и противоген: ДНК имеет две антипараллельные цепи, 5'→3' цепь обозначается как плюс цепь, а 3'→5' цепь обозначается как минус цепь. Так как РНК-полимераза наращивает нуклеиновые кислоты в направлении 5'→3', минус цепь ДНК служит в качестве матрицы для РНК в процессе транскрипции. Таким образом, транскрипт РНК должен иметь последовательность, комплементарную минус цепи и идентичную плюс цепи (за исключением замены T на U).
Антисмысловые молекулы представляют собой молекулы, которые специфически гибридизуются или специфически комплементарны либо РНК, либо плюс цепи ДНК. Смысловые молекулы представляют собой молекулы, которые специфически гибридизуются или специфически комплементарны минус цепи ДНК. Молекулы противогена представляют собой либо антисмысловые молекулы, либо смысловые молекулы, направленные на ДНК-мишень. Антисмысловая РНК (асРНК) представляет собой молекулу РНК, комплементарную смысловой (кодирующей) молекуле нуклеиновой кислоты.
Амплификация: когда используется при ссылке на нуклеиновую кислоту, это обозначает способ, с помощью которого увеличивается число копий молекулы нуклеиновой кислоты в образце или пробе. Примером амплификации является полимеразная цепная реакция, при которой биологический образец, отобранный у индивидуума, вводят в контакт с парой олигонуклеотидных праймеров в условиях, которые позволяют проходить гибридизации праймеров с матрицей нуклеиновой кислоты в образце. Праймеры при определенных условиях удлиняются, диссоциируют от матрицы и затем заново отжигаются, удлиняются и диссоциируют с амплификацией числа копий нуклеиновой кислоты. Продукт амплификации in vitro может быть охарактеризован с помощью электрофореза, паттернов рестрикции эндонуклеазами, гибридизации олигонуклеотидов или лигирования и/или секвенирования нуклеиновой кислоты с использованием стандартных методов. Другие примеры способов амплификации in vitro включают амплификацию с замещением цепи (см. патент США № 5744311); бестранскрипционную изотермическую амплификацию (см. патент США № 6033881); амплификацию с помощью цепной реакции репарации (см. заявку WO90/01069); амплификацию с помощью лигазной цепной реакции (см. патент EP-A-320308); амплификацию с помощью лигазной цепной реакции с заполнением пробелов (см. патент США № 5427930); амплификацию с помощью спаренных лигазной детекции и ПЦР (см. патент США № 6027889); и бестранскрипционную амплификацию РНК NASBATM (см. патент США № 6025134).
Антитело: полипептидный лиганд, включающий, по меньшей мере, вариабельную область легкой цепи или тяжелой цепи иммуноглобулина, который специфически распознает и связывает эпитоп (например, антиген, такой как опухолевый или вирусный антиген, или его фрагмент). Антитело включает интактные иммуноглобулины и их варианты и части, хорошо известные в данной области техники, такие как Fab' фрагменты, F(ab)'2 фрагменты, одноцепочечные Fv белки («scFv») и стабилизированные дисульфидной связью Fv белки («dsFv»). Белок scFv представляет собой гибридный белок, в котором вариабельная область легкой цепи иммуноглобулина и вариабельная область тяжелой цепи иммуноглобулина связаны линкером, в то время как в dsFvs цепи подвергнуты мутации для введения дисульфидной связи с целью стабилизации связи цепей. Термин также включает генетически сконструированные формы, такие как гибридные антитела (например, гуманизированные мышиные антитела), гетероконъюгированные антитела (например, биспецифические антитела). См. также Pierce Catalog and Handbook, 1994-1995 (Pierce Chemical Co., Rockford, IL); Kuby, J., Immunology, 3rd Ed., W.H. Freeman & Co., New York, 1997.
Обычно иммуноглобулин имеет тяжелую и легкую цепь. Каждая тяжелая и легкая цепь содержит константную область и вариабельную область (области известны также как «домены»). В сочетании вариабельные области тяжелой и легкой цепи специфически связывают антиген. Вариабельные области легкой и тяжелой цепи содержат область «каркаса», прерываемую тремя гипервариабельными областями, также называемыми «областями, определяющими комплементарность» или «CDR». Длина области каркаса и CDR определена (см. работу Kabat et al., Sequences of Proteins of Immunological Interest, U.S. Department of Health and Human Services, 1991, включенную в настоящее описание в качестве ссылки). База данных Кабата в настоящее время поддерживается в режиме прямого доступа. Последовательности областей каркаса различных легких и тяжелых цепей относительно консервативны внутри вида. Область каркаса антитела, которая представляет собой объединенные области каркасов составляющих легких и тяжелых цепей, служит для позиционирования и выравнивания CDR в трехмерном пространстве.
CDR в первую очередь отвечают за связывание с эпитопом антигена. CDR каждой цепи обычно обозначаются как CDR1, CDR2 и CDR3, с нумерацией последовательно, начиная с N-конца, и также обычно идентифицируются по цепи, в которой локализуется конкретная CDR. Таким образом, CDR3 VH локализована в вариабельном домене тяжелой цепи антитела, в котором она встречается, в то время как CDR1 VL представляет собой CDR1 из вариабельного домена легкой цепи антитела, в котором она встречается.
Ссылки на «VH» или «VH» относятся к вариабельной области тяжелой цепи иммуноглобулина, включая, такие как Fv, scFv, dsFv или Fab. Ссылки на «VL» или «VL» относятся к вариабельной области легкой цепи иммуноглобулина, включая, такие как Fv, scFv, dsFv или Fab.
«Моноклональное антитело» представляет собой антитело, продуцируемое одним клоном В-лимфоцитов или клеткой, в которую трансфицированы гены легкой и тяжелой цепей одного антитела. Моноклональные антитела продуцируются с помощью методов, известных специалистам в данной области техники, например, с помощью создания гибридомных продуцирующих антитело клеток путем слияния клеток миеломы с иммунными клетками селезенки. Моноклональные антитела включают гуманизированные моноклональные антитела.
«Гуманизированный» иммуноглобулин представляет собой иммуноглобулин, включающий область каркаса от человека и одну или более CDR иммуноглобулина не от человека (от животного, такого как мышь, крыса, или синтетическую). Иммуноглобулин не от человека, снабжающий CDR, называется «донором», а иммуноглобулин человека, снабжающий каркасом, называется «акцептором». В одном варианте осуществления в гуманизированном иммуноглобулине все CDR происходят от иммуноглобулина-донора. Присутствие константных областей не является обязательным, но если они присутствуют, они должны быть по существу идентичными константным областям иммуноглобулина человека, т.е. идентичными, по меньшей мере, приблизительно на 85-90%, приблизительно на 95% или более. Следовательно, все части гуманизированного иммуноглобулина за исключением, может быть, CDR, являются по существу идентичными соответствующим частям природных последовательностей иммуноглобулина человека. «Гуманизированное антитело» представляет собой антитело, включающее гуманизированную легкую цепь и гуманизированную тяжелую цепь иммуноглобулина. Гуманизированное антитело связывается с тем же антигеном, что и антитело-донор, которое снабжает CDR. Каркас акцептора гуманизированного иммуноглобулина или антитела может иметь ограниченное число замен аминокислот, взятых из каркаса донора. Гуманизированные или другие моноклональные антитела могут иметь дополнительные консервативные замены аминокислот, которые по существу не влияют на связывание антигена или другие функции иммуноглобулина. Гуманизированные иммуноглобулины могут быть созданы с помощью способов генной инженерии (например, см. патент США № 5585089).
«Нейтрализующее антитело» представляет собой антитело, которое препятствует любой биологической активности полипептида, такого как полипептид PD-1. Например, нейтрализующее антитело может мешать способности полипептида PD-1 снижать иммунный ответ, такой как цитотоксичность Т-клеток. В некоторых примерах нейтрализующее антитело может снижать способность полипептида PD-1 снижать иммунный ответ на приблизительно 50%, приблизительно 70%, приблизительно 90% или более. Для оценки потенциально нейтрализующих антител может быть использован любой стандартный метод измерения иммунных ответов, включая те, которые описаны в настоящем документе.
Антиген: соединение, композиция или вещество, которые могут стимулировать продукцию антител или Т-клеточный ответ у животного, включая композиции, которые вводятся животному или всасываются у него. Антиген взаимодействует с продуктами специфического гуморального или клеточного компонента иммунной системы, включая те, которые индуцируются гетерологичными иммуногенами. Термин «антиген» включает все родственные антигенные эпитопы. «Эпитоп» или «антигенная детерминанта» относится к сайту на антигене, на который отвечают В- и/или Т-клетки. В одном варианте осуществления Т-клетки отвечают на эпитоп, когда эпитоп презентируется в связи с молекулой MHC. Эпитопы могут быть образованы или из соседних аминокислот, или из аминокислот, не являющихся соседними, размещающимися рядом в результате третичной укладки белка. Эпитопы, образованные из соседних аминокислот, обычно сохраняются при экспозиции с денатурирующими растворителями, тогда как эпитопы, образованные при третичной укладке, обычно теряются при обработке денатурирующими растворителями. Эпитоп обычно включает по меньшей мере 3 и более обычно по меньшей мере 5, приблизительно 9 или приблизительно 8-10 аминокислот в уникальной пространственной конформации. Методы определения пространственной конформации эпитопов включают, например, рентгеновскую кристаллографию и 2-мерный ядерный магнитный резонанс.
Антиген может быть тканеспецифическим антигеном или антигеном, специфичным для заболевания. Эти термины не являются исключающими, так как тканеспецифический антиген может быть также антигеном, специфичным для заболевания. Тканеспецифический антиген экспрессируется в ограниченном количестве тканей, таком как одна ткань. Конкретными неограничивающими примерами тканеспецифического антигена являются простатспецифический антиген, специфический антиген матки и/или специфический антиген семенников. Тканеспецифический антиген может экспрессироваться более чем в одной ткани, такой как, но не ограничиваясь этим, антиген, который экспрессируется в более чем одной репродуктивной ткани, такой как и ткань простаты, и ткань матки. Антиген, специфичный для заболевания, экспрессируется сопутствуя развитию заболевания. Конкретным неограничивающим примером антигена, специфичного для заболевания, является антиген, чья экспрессия коррелирует с образованием опухоли или прогнозирует ее. Антиген, специфичный для заболевания, может быть антигеном, узнаваемым Т-клетками или В-клетками.
Антигенпрезентирующая клетка (APC): клетка, которая может презентировать антиген, связанный с молекулами MHC класса I или класса II, Т-клеткам. APC включают, но не ограничиваются этим, моноциты, макрофаги, дендритные клетки, В-клетки, Т-клетки и клетки Лангерганса. Т-клетка, которая представляет антиген другим Т-клеткам (включая CD4+ и/или CD8+ Т-клетки), представляет собой антигенпрезентирующую Т-клетку (T-APC).
Связанный или стабильно связанный (олигонуклеотид): олигонуклеотид связывается или стабильно связывается с нуклеиновой кислотой-мишенью, если достаточное количество олигонуклеотида образует пары оснований или гибридизуется со своей нуклеиновой кислотой-мишенью, так что позволяет определить это связывание. Связывание может быть определено с помощью либо физических, либо функциональных свойств мишень:олигонуклеотидный комплекс. Связывание между мишенью и олигонуклеотидом может быть определено с помощью любого метода, известного специалисту в данной области техники, включая как функциональные методы, так и методы оценки физического связывания. Например, связывание может быть определено функционально путем определения, обладает ли связывание выявляемым эффектом на процесс биосинтеза, такой как экспрессия гена, репликация ДНК, транскрипция, трансляция и тому подобное.
Физические методы определения связывания комплементарных цепей ДНК или РНК хорошо известны в данной области техники и включают такие методы, как методы ДНКазы I или химического футпринтинга, задержки в геле и аффинного расщепления, процедуры нозерн-блоттинга, дот-блоттинга и определение поглощения света. Например, один метод, который широко используется из-за простоты и надежности, включает выявление изменения поглощения света раствора, содержащего олигонуклеотид (или аналог) и нуклеиновую кислоту-мишень, при от 220 до 300 нм, в то время как температура медленно повышается. Если олигонуклеотид или аналог связывается со своей мишенью, возникает внезапное увеличение поглощения при характерной температуре, поскольку олигонуклеотид (или аналог) и мишень диссоциируют один от другого или плавятся.
Связывание между олигомером и его нуклеиновой кислотой-мишенью часто характеризуется температурой (Tm), при которой 50% олигомера диссоциирует от мишени. Более высокая (Tm) означает наличие более прочного или более стабильного комплекса по сравнению с комплексом с более низкой (Tm).
Рак или опухоль: малигнизированная неоплазма, которая подверглась характерной анаплазии с потерей дифференцировки, повышением скорости роста, инвазией в окружающую ткань со способностью к метастазированию. Рак репродуктивной системы представляет собой рак, первично возникающий в репродуктивной ткани, такой как матка, семенники, яичники, простата, фаллопиева труба или пенис. Например, рак простаты представляет собой малигнизированную неоплазму, которая возникает в или из ткани простаты, а рак матки представляет собой малигнизированную неоплазму, которая возникает в или из ткани матки, и рак семенников представляет собой малигнизированную неоплазму, которая возникает в семенниках. Остаточный рак представляет собой рак, который остается у индивидуума после любой формы лечения, получаемого индивидуумом для снижения или истребления рака щитовидной железы. Метастазирующий рак представляет собой рак в одном или более мест организма, отличных от места происхождения исходного (первичного) рака, из которого происходит метастазирующий рак.
Химиотерапия; химиотерапевтические агенты: при использовании в настоящем описании любой химический агент с терапевтической пригодностью для лечения заболеваний, характеризуемых аномальным клеточным ростом. Такие заболевания включают опухоли, неоплазмы и рак, а также заболевания, характеризуемые гиперпластическим ростом, такие как псориаз. В одном варианте осуществления химиотерапевтический агент представляет собой агент для использования при лечении неоплазм, таких как солидные опухоли. В одном варианте осуществления химиотерапевтический агент представляет собой радиоактивную молекулу. Специалист в данной области техники легко может идентифицировать химиотерапевтический агент для использования (например, см. Slapak and Kufe, Principles of Cancer Therapy, Chapter 86 in Harrison's Principles of Internal Medicine, 14th edition; Perry et al., Chemotherapy, Ch. 17 in Abeloff, Clinical Oncology 2nd ed., © 2000 Churchill Livingstone, Inc; Baltzer L, Berkery R (eds): Oncology Pocket Guide to Chemotherapy, 2nd ed. St. Louis, Mosby-Year Book, 1995; Fischer DS, Knobf MF, Durivage HJ (eds): The Cancer Chemotherapy Handbook, 4th ed. St. Louis, Mosby-Year Book, 1993). Иммуногенные полипептиды, раскрытые в настоящем описании, могут быть использованы в сочетании с дополнительными химиотерапевтическими агентами.
Контрольный уровень: уровень молекулы, такой как полипептид или нуклеиновая кислота, обычно выявляемый в природе при определенных условиях и/или для конкретного генетического фона. В определенных вариантах осуществления контрольный уровень молекулы может быть измерен в клетке или в образце, которые не подвергались, ни прямо, ни непрямо, обработке. В некоторых примерах контрольный уровень может представлять собой уровень в клетке, не контактировавшей с агентом, таким как антагонист PD-1. В дополнительных примерах контрольный уровень может представлять собой уровень у индивидуума, которому не вводили антагонист PD-1.
ДНК (дезоксирибонуклеиновая кислота): ДНК представляет собой длинноцепочечный полимер, который включает генетический материал у большинства живых организмов (некоторые вирусы имеют гены, включающие рибонуклеиновую кислоту (РНК)). Повторяющимися единицами в полимерах ДНК являются четыре различных нуклеотида, каждый из которых включает одно из четырех оснований, аденин, гуанин, цитозин и тимин, связанные с сахаром дезоксирибозой, к которой присоединена фосфатная группа. Триплеты нуклеотидов (обозначаемые как кодоны) кодируют каждую аминокислоту в полипептиде или стоп-сигнал. Термин «кодон» также используется для соответствующих (и комплементарных) последовательностей из трех нуклеотидов в мРНК, в которую транскрибируется последовательность ДНК.
Если не указано иначе, любая ссылка на молекулу ДНК предназначена для включения обратной комплементарной молекулы ДНК. Кроме тех случаев, когда в тексте настоящего описания требуется одноцепочечная молекула, молекулы ДНК, хотя пишутся с изображением только одной цепи, охватывают обе цепи двуспиральной молекулы ДНК.
Кодировать: говорят, что полинуклеотид кодирует полипептид, если он в своем природной состоянии или после манипуляций с помощью методов, хорошо известных специалистам в данной области техники, может транскрибироваться и/или транслироваться с получением мРНК для полипептида и/или полипептида или его фрагмента. Антисмысловая цепь комплементарна такой нуклеиновой кислоте, и кодирующая последовательность может быть выведена из нее.
Экспрессия: процесс, с помощью которого информация, кодируемая генами, превращается в структуры, присутствующие и работающие в клетке. Экспрессированные гены включают те, которые транскрибируются в мРНК и затем транслируются в белок, и те, которые транскрибируются в РНК, но не транслируются в белок (например, миРНК, транспортную РНК и рибосомальную РНК). Таким образом, экспрессия последовательности-мишени, такой как ген или промоторная область гена, может приводить к экспрессии мРНК, белка или обоих. Экспрессия последовательности-мишени может быть ингибирована или усилена (понижена или повышена).
Последовательности, контролирующие экспрессию: последовательности нуклеиновой кислоты, которые регулируют экспрессию гетерологичной последовательности нуклеиновой кислоты, к которой они оперативно присоединены. Последовательности, контролирующие экспрессию, оперативно связаны с последовательностью нуклеиновой кислоты, когда последовательности, контролирующие экспрессию, контролируют и регулируют транскрипцию и, когда это предназначено, трансляцию последовательности нуклеиновой кислоты. Таким образом, последовательности, контролирующие экспрессию, могут включать подходящие промоторы, энхансеры, терминаторы транскрипции, инициирующий кодон (т.е. ATG) перед геном, кодирующим белок, сигналы сплайсинга, элементы для поддержания правильной рамки считывания этого гена для разрешения корректной трансляции мРНК и стоп-кодоны. Термин «контролирующие последовательности» предназначен для включения как минимум тех компонентов, чье присутствие может повлиять на экспрессию, и может также включать дополнительные компоненты, чье присутствие является выгодным, например лидирующие последовательности и последовательности партнеров по гибридизации. Последовательности, контролирующие экспрессию, могут включать промотор.
Промотор представляет собой минимальную последовательность, достаточную для прямой транскрипции. Включенными также являются те промоторные элементы, которые достаточны для предоставления зависимой от промотора экспрессии гена возможности контролироваться специфичными для клеточного типа, специфичными для ткани или индуцибельными внешними сигналами или агентами; такие элементы могут быть локализованы в 5'- или 3'-областях гена. Включаются как конститутивные, так и индуцибельные промоторы (см., например, Bitter et al., Methods in Enzymology 153:516-544, 1987). Например, когда клонирование происходит в бактериальных системах, могут быть использованы индуцибельные промоторы, такие как pL бактериофага лямбда, plac, ptrp, ptac (гибридный промотор ptrp-lac) и тому подобное. В одном варианте осуществления при клонировании в клеточных системах млекопитающих могут быть использованы промоторы, происходящие из генома клеток млекопитающих (такой как промотор металлотионеина) или из вирусов млекопитающих (такие как длинный концевой повтор ретровируса; поздний промотор аденовируса; 7,5K промотор вируса коровьей оспы). Для обеспечения транскрипции последовательностей нуклеиновой кислоты могут быть также использованы промоторы, продуцируемые рекомбинантной ДНК или синтетическими методами.
Гетерологичный: происходящий от отдельных генетических источников или видов. Обычно антитело, которое специфически связывается с интересующим белком, не будет специфически связываться с гетерологичным белком.
Клетки-хозяева: клетки, в которых вектор может размножаться и его ДНК экспрессироваться. Клетка может быть прокариотной или эукариотной. Клетка может быть клеткой млекопитающего, такой как клетка человека. Термин также включает любых потомков подлежащей клетки-хозяина. Понятно, что все потомки могут не быть идентичными родительской клетке, так как могут происходить мутации, которые возникают в процессе репликации. Однако такие потомки включаются, когда используется термин «клетка-хозяин».
Иммунный ответ: ответ клетки иммунной системы, такой как В-клетка, Т-клетка или моноцит на стимул. В одном варианте осуществления ответ является специфичным в отношении конкретного антигена («антиген-специфичный ответ»). В одном варианте осуществления иммунный ответ представляет собой Т-клеточный ответ, такой как CD4+ ответ или CD8+ ответ. В другом варианте осуществления ответ представляет собой В-клеточный ответ и ведет к продукции специфических антител.
«Отсутствие ответа» в отношении иммунных клеток включает рефрактерность иммунных клеток к стимуляции, такой как стимуляция через активирующий рецептор или цитокин. Отсутствие ответа может возникать, например, из-за экспозиции с иммуносупрессорами или из-за экспозиции с высокими дозами антигена. Применяемый в настоящем описании термин «анергия» или «толерантность» включает рефрактерность к стимуляции, опосредованной активирующим рецептором. Такая рефрактерность является обычно антиген-специфичной и продолжается после прекращения экспозиции с толеризующим антигеном. Например, анергия у Т-клеток (как противоположность отсутствию ответа) характеризуется отсутствием продукции цитокинов (таких как ИЛ-2). Анергия Т-клеток возникает, когда Т-клетки экспонируются с антигеном и получают первый сигнал (сигнал, опосредуемый рецептором Т-клеток или CD-3) в отсутствие второго сигнала (костимулирующего сигнала). В этих условиях повторная экспозиция клеток с тем же самым антигеном (даже если экспозиция осуществляется в присутствии костимулирующей молекулы) ведет к невозможности продукции цитокинов и, таким образом, невозможности пролиферировать. Т-клетки в состоянии анергии могут, однако, генерировать ответ на неродственные антигены и могут пролиферировать, если культивируются с цитокинами (такими как ИЛ-2). Например, Т-клетки в состоянии анергии могут также выявляться по отсутствию продукции ИЛ-2 Т-лимфоцитами при измерении с помощью ИФА или теста на пролиферацию с использованием индикаторной клеточной линии. Альтернативно может быть использован репортерный генный конструкт. Например, Т-клетки в состоянии анергии не способны инициировать транскрипцию гена ИЛ-2, индуцируемую гетерологичным промотором под контролем 5'-энхансера гена ИЛ-2 или мультимерной последовательности AP1, которая может быть обнаружена в энхансере (Kang et al. Science 257:1134, 1992). Антиген-специфичные Т-клетки в состоянии анергии могут обладать пониженной цитотоксической активностью по меньшей мере на 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95% или даже 100% по сравнению с соответствующей контрольной антиген-специфичной Т-клеткой.
Иммуногенный пептид: пептид, который включает аллель-специфичный мотив или другую последовательность, так что пептид будет связываться с молекулой MHC и индуцировать цитотоксический ответ Т-лимфоцитов («CTL») или ответ В-клеток (например, продукцию антител) против антигена, из которого произошел иммуногенный пептид.
В одном варианте осуществления иммуногенные пептиды идентифицируют, используя мотивы последовательности или другие методы, такие как определение с помощью компьютерной нейронной сети или полиномного анализа, известные в данной области техники. Обычно алгоритмы для определения «порога связывания» пептидов используют для выбора пептидов с очками, которые дают им высокую вероятность связывания с определенной аффинностью и которые должны быть иммуногенными. Алгоритмы основаны либо на эффектах связывания MHC конкретной аминокислоты в определенном положении, эффектах связывания антителом конкретной аминокислоты в определенном положении, либо на эффектах связывания конкретной замены в пептиде, содержащем мотив. В контексте иммуногенного пептида «консервативный остаток» представляет собой такой остаток, который появляется с существенно более высокой частотой, чем это должно ожидаться при рандомизированном распределении в определенном положении в пептиде. В одном варианте осуществления консервативный остаток представляет собой такой, с помощью которого структура MHC может обеспечить точку контакта с иммуногенным пептидом.
Иммуногенные пептиды могут быть также идентифицированы с помощью измерения их связывания со специфичным белком MHC (например, HLA-A02.01) и по их способности стимулировать CD4 и/или CD8 при презентации в контексте белка MHC.
Иммуногенная композиция: композиция, включающая иммуногенный полипептид или нуклеиновую кислоту, кодирующую иммуногенный полипептид, которая индуцирует измеряемый ответ CTL против клеток, экспрессирующих полипептид, или индуцирует измеряемый ответ В-клеток (такой как продукция антител, которые специфически связывают полипептид) против полипептида. При использовании in vitro иммуногенная композиция может состоять из выделенной нуклеиновой кислоты, вектора, включающего нуклеиновую кислоту, или иммуногенного пептида. При использовании in vivo иммуногенная композиция должна обычно включать нуклеиновую кислоту, вектор, включающий нуклеиновую кислоту, и/или иммуногенный полипептид в фармацевтически приемлемых носителях, и/или другие агенты. Иммуногенная композиция может необязательно включать адъювант, антагонист PD-1, костимулирующую молекулу или нуклеиновую кислоту, кодирующую костимулирующую молекулу. Полипептид или нуклеиновая кислота, кодирующая полипептид, могут быть легко протестированы на их способность индуцировать CTL с помощью методов, известных в данной области техники.
Ингибирование или лечение заболевания: ингибирование заболевания, такого как рост опухоли или устойчивая инфекция, относится к ингибированию полного развития заболевания или снижению физиологических эффектов заболевания в процессе развития. В некоторых примерах ингибирование или лечение заболевания относится к снижению симптомов опухоли или инфекции патогеном. Например, лечение рака может предотвращать развитие паранеопластического синдрома у индивидуума, для которого известно, что он болеет раком, или вызывать снижение признака или симптома опухоли. В другом варианте осуществления лечение инфекции может относиться к ингибированию развития или уменьшению симптома инфекции. «Лечение» относится к терапевтическому вмешательству, которое облегчает признак или симптом заболевания или патологического состояния, относящегося к заболеванию. Терапевтическая вакцинация относится к введению агента индивидууму, уже инфицированному патогеном. Индивидуум может не иметь симптомов, так что лечение предотвращает развитие симптома. Терапевтическая вакцина может также снижать тяжесть одного или более существующих симптомов или снижать нагрузку патогеном.
Инфекционное заболевание: любое заболевание, вызываемое инфекционным агентом. Примеры инфекционных патогенов включают, но не ограничиваются этим, вирусы, бактерии, микоплазму и грибы. В конкретном примере это заболевание, вызываемое по меньшей мере одним типом инфекционного патогена. В другом примере это заболевание, вызываемое по меньшей мере двумя различными типами инфекционных патогенов. Инфекционные заболевания могут влиять на любую систему организма, быть острыми (короткодействующими) или хроническими/устойчивыми (долгодействующими), возникать с или без лихорадки, поражать любую возрастную группу и перекрывать друг друга.
Вирусные заболевания обычно возникают после иммуносупрессии из-за реактивации вирусов, уже присутствующих у реципиента. Конкретные примеры устойчивых вирусных инфекций включают, но не ограничиваются этим, цитомегаловирусные (CMV) пневмонию, энтерит и ретинит; лимфопролиферативное заболевание, вызываемое вирусом Эпштейна-Барр (EBV); ветрянку/опоясывающий лишай (вызываемый вирусом ветряной оспы, VZV); микозит, вызываемый NSV-1 и -2; энцефалит, вызываемый HSV-6; геморрагический цистит, вызываемый BK-вирусом; вирусный грипп; пневмонию, вызываемую респираторным синцитиальным вирусом (RSV); СПИД (вызываемый ВИЧ) и гепатит A, B или C.
Дополнительные примеры инфекционных вирусов включают: Retroviridae; Picornaviridae (например, полиовирусы, вирус гепатита A; энтеровирусы, вирусы коксаки человека, риновирусы, ECHO-вирусы); Calciviridae (такие как штаммы, которые вызывают гастроэнтерит); Togaviridae (например, вирусы энцефалита лошадей, рубелла вирусы); Flaviridae (например, вирусы лихорадки Денге, вирусы энцефалита, вирусы желтой лихорадки); Coronaviridae (например, коронавирусы); Rhabdoviridae (например, вирусы везикулярного стоматита, вирусы бешенства); Filoviridae (например, вирусы лихорадки Эбола); Paramyxoviridae (например, вирусы парагриппа, вирус свинки, вирус кори, респираторный синцитиальный вирус); Orthomyxoviridae (например, вирусы гриппа); Bungaviridae (например, вирусы Хантаан, бунгавирусы, флебовирусы и наировирусы); Arena viridae (вирусы геморрагической лихорадки); Reoviridae (например, реовирусы, орбивирусы и ротавирусы); Birnaviridae; Hepadnaviridae (вирус гепатита B); Parvoviridae (парвовирусы); Papovaviridae (вирусы папилломы, вирусы полиомы); Adenoviridae (большинство аденовирусов); Herpesviridae (вирус простого герпеса (HSV) 1 и HSV-2, вирус ветряной оспы, цитомегаловирус (CMV), вирусы герпеса); Poxviridae (вирус оспы человека, вирусы коровьей оспы, поксвирусы); и Iridoviridae (такой как вирус лихорадки африканских свиней); и неклассифицированные вирусы (например, этиологические агенты губчатых энцефалопатий, агент гепатита дельта (как предполагается, являющийся дефектным сателлитом вируса гепатита B), агенты гепатита не-A, не-B (класс 1=передающийся внутренним путем; класс 2=передающийся парентерально (т.е. гепатит C); вирус Норволк и родственные вирусы и астровирусы).
Примеры грибковых инфекций включают, но не ограничиваются этим: аспергиллез; молочницу (вызываемую Candida albicans); криптококкоз (вызываемый Cryptococcus); и гистоплазмоз. Таким образом, примеры инфекционных грибов включают, но не ограничиваются этим, Cryptococcus neoformans, Histoplasma capsulatum, Coccidioides immitis, Blastomyces dermatitidis, Chlamydia trachomatis, Candida albicans.
Примеры инфекционных бактерий включают: Helicobacter pyloris, Borelia burgdorferi, Legionella pneumophilia, Mycobacteria sps (такие как M. tuberculosis, M. avium, M. intracellular, M. kansaii, M. gordonae), Staphylococcus aureus, Neisseria gonorrhoeae, Neisseria meningitidis, Listeria monocytogenes, Streptococcus pyogenes (группа A Streptococcus), Streptococcus agalactiae (группа B Streptococcus), Streptococcus (группа Viridans), Streptococcus faecalis, Streptococcus bovis, Streptococcus (анаэробные sps.), Streptococcus pneumoniae, патогенную Campylobacter sp., Enterococcus sp., Haemophilus influenzae, Bacillus anthracis, corynebacterium diphtheriae, corynebacterium sp., Erysipelothrix rhusiopathiae, Clostridium perfringers, Clostridium tetani, Enterobacter aerogenes, Klebsiella pneumoniae, Pasturella multocida, Bacteroides sp., Fusobacterium nucleatum, Streptobacillus moniliformis, Treponema pallidium, Treponema pertenue, Leptospira и Actinomyces israelli. Другие инфекционные организмы (такие как простейшие) включают: Plasmodium falciparum и Toxoplasma gondii.
«Устойчивая инфекция» представляет собой инфекцию, при которой инфекционный агент (такой как вирус, микоплазма, бактерия, паразит или гриб) не выводится или не элиминируется из инфицированного хозяина даже после индукции иммунного ответа. Устойчивые инфекции могут представлять собой хронические инфекции, латентные инфекции или медленные инфекции. Латентная инфекция характеризуется отсутствием доказуемого наличия инфекционного вируса между эпизодами рецидивирующего заболевания. Хроническая инфекция характеризуется продолжительным присутствием инфекционного вируса после первичной инфекции и может включать хроническое или рецидивирующее заболевание. Медленная инфекция характеризуется продолжительным инкубационным периодом с последующим прогрессированием заболевания. В отличие от латентной и хронической инфекций медленная инфекция может не начинаться с острого периода размножения вируса. В то время как острые инфекции являются относительно короткими (продолжающимися от нескольких дней до нескольких недель) и вирусы устраняются из организма с помощью иммунной системы, устойчивые инфекции могут продолжаться, например, в течение месяцев, лет или даже пожизненно. Эти инфекции могут также часто рецидивировать в течение длительного периода времени, включая стадии молчащей и продуктивной инфекции, без уничтожения клеток или даже с индукцией избыточного повреждения клеток хозяина. Устойчивые инфекции часто включают стадии как молчащей, так и продуктивной инфекции без быстрого уничтожения или даже с индукцией избыточного повреждения клеток хозяина. В ходе устойчивых вирусных инфекций вирусный геном может быть либо стабильно интегрирован в клеточную ДНК, либо может поддерживаться эписомально. Устойчивая инфекция возникает при инфицировании такими вирусами, как вирусы Т-клеточного лейкоза, вирус Эпштейна-Барр, цитомегаловирус, вирусы герпеса, вирус ветряной оспы, вирус кори, паповавирусы, прионы, вирусы гепатита, аденовирусы, парвовирусы и вирусы папилломы.
Вызывающие заболевание инфекционные агенты (такие как имеющиеся внутри специфических клеток инфицированных индивидуумов) могут быть также определены у хозяина с помощью стандартных методов даже после разрешения иммунного ответа. Млекопитающие диагностируются как имеющие устойчивую инфекцию в соответствии с любым стандартным методом, известным в данной области техники и описанным, например, в патентах США №№ 6368832, 6579854 и 6808710 и в публикациях патентных заявок США №№ 20040137577, 20030232323, 20030166531, 20030064380, 20030044768, 20030039653, 20020164600, 20020160000, 20020110836, 20020107363 и 20020106730, все включены в настоящее описание в качестве ссылки.
«Облегчение симптома устойчивой инфекции» представляет собой облегчение любого состояния или симптома, связанного с устойчивой инфекцией. Альтернативно облегчение симптома устойчивой инфекции может включать снижение инфекционной микробной (такой как вирусная, бактериальная, грибковая или паразитарная) нагрузки у индивидуума относительно такой нагрузки у контроля, не подвергавшегося лечению. По сравнению с эквивалентным контролем, не подвергавшимся лечению, такое снижение или степень предотвращения составляет по меньшей мере 5%, 10%, 20%, 40%, 50%, 60%, 80%, 90%, 95% или 100% при измерении с помощью любого стандартного метода. Желательно, чтобы устойчивая инфекция полностью исчезала при определении любым стандартным методом, известным в данной области техники, в таком случае устойчивая инфекция рассматривается как излеченная. Больной, который подвергается лечению в отношении устойчивой инфекции, является таким, у которого врачом диагностировано такое состояние. Диагноз может быть установлен с помощью любых подходящих средств. Диагностика и мониторинг могут включать, например, определение уровня микробной нагрузки в биологическом образце (например, в биопсийном материале ткани, определение в крови или определение в моче), определение уровня суррогатного маркера микробной инфекции в биологическом образце, определение симптомов, связанных с устойчивыми инфекциями, или определение иммунных клеток, вовлеченных в иммунный ответ, типичный для устойчивых инфекций (например, определение антиген-специфичных Т-клеток, которые находятся в состоянии анергии и/или функционально ослаблены). У больного, у которого предотвращается развитие устойчивой инфекции, может быть поставлен или не поставлен такой диагноз. Специалист в данной области техники должен понимать, что эти больные могут быть подвергнуты тем же самым стандартным тестам, что и описанные выше, или могут быть идентифицированы без проверки, как имеющие высокий риск, обусловленный присутствием одного или более факторов риска (таких как семейный анамнез или экспозиция с инфекционным агентом).
Выделенный: «выделенный» биологический компонент (такой как нуклеиновая кислота, или белок, или органелла) по существу отделен или очищен от других биологических компонентов клетки организма, в которой компонент находится в природе, т.е. от других хромосомных и внехромосомных ДНК и РНК, белков и органелл. Нуклеиновые кислоты и белки, которые «выделены», включают нуклеиновые кислоты и белки, очищенные с помощью стандартных методов очистки. Термин также охватывает нуклеиновые кислоты и белки, полученные с помощью рекомбинантной экспрессии в клетке-хозяине, а также химически синтезированные нуклеиновые кислоты.
«Очищенное антитело» составляет по меньшей мере 60% по массе от белков и природных органических молекул, с которыми он связан в природе. В некоторых примерах препарат содержит антитело, такое как специфическое антитело против PD-1, PD-L1 или PD-L2, в количестве по меньшей мере приблизительно 75%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95% по меньшей мере приблизительно 99% по массе. Очищенное антитело может быть получено, например, с помощью аффинной хроматографии с использованием рекомбинантно продуцируемого белка или консервативных мотивов пептида и стандартных способов.
Меченое: определяемое соединение или композиция, которое конъюгировано прямо или непрямо с другой молекулой, для облегчения определения этой молекулы. Конкретные, неограничивающие примеры меток включают флуоресцентные метки, конъюгаты с ферментами и радиоактивные изотопы.
Лимфоциты: тип белых клеток крови, которые вовлечены в иммунную защиту организма. Существует два главных типа лимфоцитов: В-клетки и Т-клетки.
Главный комплекс гистосовместимости (MHC): групповое обозначение, предназначенное для охвата систем гистосовместимости антигенов, описанных у разных видов, включая антигены лейкоцитов человека («HLA»).
Млекопитающее: этот термин включает как человека, так и млекопитающих, не являющихся человеком. Сходно, термин «индивидуум» включает как человека, так и объектов ветеринарии.
Неоплазма: аномальная клеточная пролиферация, которая включает доброкачественные и злокачественные опухоли, а также другие пролиферативные нарушения.
Олигонуклеотид: линейная полинуклеотидная последовательность до приблизительно 100 нуклеотидных оснований в длину.
Открытая рамка считывания (ORF): последовательность нуклеотидных триплетов (кодонов), кодирующая аминокислоты без каких-либо внутренних кодонов терминации. Эти последовательности обычно способны транслироваться в пептид.
Оперативно связанный: первая последовательность нуклеиновой кислоты оперативно связана со второй последовательностью нуклеиновой кислоты, когда первая последовательность нуклеиновой кислоты расположена в функциональной связи со второй последовательностью нуклеиновой кислоты. Например, промотор оперативно связан с кодирующей последовательностью, если промотор влияет на транскрипцию или экспрессию кодирующей последовательности. Обычно оперативно связанные последовательности ДНК являются примыкающими и, когда необходимо соединить две области, кодирующие белок в одну и ту же рамку считывания.
Фармацевтически приемлемые носители: используемые фармацевтически приемлемые носители являются общепринятыми. В Remington's Pharmaceutical Sciences, by E. W. Martin, Mack Publishing Co., Easton, PA, 15th Edition (1975) описываются композиции и составы, подходящие для фармацевтической доставки раскрытых в настоящем описании гибридных белков.
В целом природа носителя должна зависеть от используемого конкретного способа введения. Например, парентеральные составы обычно включают вводимые жидкости, которые включают в качестве носителя фармацевтически и физиологически приемлемые жидкости, такие как вода, физиологический раствор, сбалансированные солевые растворы, водную декстрозу, глицерин или тому подобное. Для твердых составов (таких как порошковая, пилюльная, таблеточная или капсульная формы) общепринятые нетоксичные твердые носители могут включать, например, маннит, лактозу, крахмал или стеарат магния фармацевтической чистоты. В дополнение к биологически нейтральным носителям, фармацевтические композиции, предназначенные для введения, могут содержать минорные количества нетоксичных вспомогательных веществ, таких как увлажняющие или эмульгирующие агенты, консерванты и буферирующие рН агенты и тому подобное, например ацетат натрия или сорбитмонолаурат.
«Терапевтически эффективное количество» представляет собой количество композиции или клеток для достижения желаемого эффекта у индивидуума, подвергаемого лечению. Например, это может быть количество антагониста PD-1, необходимое для индукции иммунного ответа, ингибирования опухолевого роста или для измеряемого изменения внешних симптомов опухоли или устойчивой инфекции. При введении индивидууму обычно должна использоваться дозировка, которая в ткани-мишени (например, в лимфоцитах) будет достигать концентраций, которые, как показано, вызывают эффект in vitro.
Полинуклеотид: термин полинуклеотид или последовательность нуклеиновой кислоты относится к полимерной форме нуклеотида по меньшей мере из 10 оснований в длину. Рекомбинантный полинуклеотид включает полинуклеотид, который прямо не примыкает к обеим кодирующим последовательностям, с которыми он непосредственно граничит (одной на 5'-конце и одной на 3'-конце) в природном геноме организма, из которого он происходит. Термин, следовательно, включает, например, рекомбинантную ДНК, которая включена в вектор; в автономно реплицирующуюся плазмиду или вирус; или в геномную ДНК прокариота или эукариота, или которая существует в виде отдельной молекулы (например, кДНК), независимо от других последовательностей. Нуклеотиды могут представлять собой рибонуклеотиды, дезоксирибонуклеотиды или модифицированные формы любого нуклеотида. Термин включает одно- и двухцепочечные формы ДНК.
Полипептид: любая цепь аминокислот независимо от длины или посттрансляционной модификации (например, гликозилирования или фосфорилирования). Полипептид может составлять от 3 до 30 аминокислот в длину. В одном варианте осуществления полипептид составляет от приблизительно 7 до приблизительно 25 аминокислот в длину. В еще одном варианте осуществления полипептид составляет от приблизительно 8 до приблизительно 10 аминокислот в длину. В еще одном варианте осуществления пептид составляет приблизительно 9 аминокислот в длину. В отношении полипептидов «включает» указывает на то, что в молекулу может быть включена дополнительная аминокислотная последовательность или другие молекулы, «состоит по существу из» указывает на то, что в молекулу не включены дополнительные аминокислотные последовательности, но могут быть включены другие агенты (такие как метки или химические соединения), и «состоит» указывает на то, что в молекулу не включены дополнительные аминокислотные последовательности и дополнительные агенты.
Белок программируемой смерти (PD)-1: белок, который образует комплекс с белком PD-L1 или PD-L2 и вовлечен в иммунный ответ, такой как костимуляция Т-клеток. Обычно белок PD-1 по существу идентичен природному PD-1 (дикого типа) (см., например, Ishida et al. EMBO J. 11:3887-3895, 1992, Shinohara et al. Genomics 23:704-706, 1994; и патент США № 5698520, все включены в настоящее описание в качестве ссылки в полном объеме). В некоторых примерах при передаче сигнала PD-1 снижается, например, цитотоксичность CD8+ Т-клеток в результате снижения пролиферации Т-клеток, продукции цитокинов или клиренса вирусов. Таким образом, полипептид PD-1 может снижать цитотоксическую активность CD8+ Т-клеток по меньшей мере на 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или более 100% по сравнению с контрольными уровнями при измерении с помощью любого стандартного метода.
Применяемый в настоящем описании термин «активность» в отношении полипептида или белка PD-1 включает любую активность, которая присуща природному белку PD-1, такую как способность модулировать ингибиторный сигнал в активированной иммунной клетке таким способом, как с помощью модуляции контактирования природного лиганда с антигенпрезентирующей клеткой. Такая модуляция ингибиторного сигнала в иммунной клетке ведет к модуляции пролиферации и/или выживания иммунной клетки, и/или секреции цитокинов иммунной клеткой. Белок PD-1 может также модулировать костимуляторный сигнал путем конкуренции с костимуляторным рецептором за связывание молекулы B7. Таким образом, термин «активность PD-1» включает способность полипептида или белка PD-1 связываться со своим(ми) природным(ми) лигандом(ами), способность модулировать костимуляторные или ингибиторные сигналы иммунной клетки и способность модулировать иммунный ответ.
«Снижение экспрессии или активности PD-1» относится к снижению уровня или биологической активности PD-1 по отношению к уровню или биологической активности белка PD-1 в контроле, таком как не подвергаемый лечению индивидуум или необработанный образец. В конкретных примерах уровень или активность снижена по меньшей мере на 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 100% или даже выше 100% по отношению к необработанному контролю. Например, биологическая активность белка PD-1 снижается, если снижается связывание белка PD-1 с PD-L1, PD-L2 или обоими, тем самым вызывая снижение сигнализации PD-1 и, следовательно, приводя к увеличению цитотоксичности CD8+ Т-клеток.
«Ген PD-1» представляет собой нуклеиновую кислоту, которая кодирует белок PD-1. «Гибридный ген PD-1» представляет собой кодирующую область PD-1, оперативно связанную со второй, гетерологичной последовательностью нуклеиновой кислоты. Гибридный ген PD-1 может включать промотор PD-1 или может включать гетерологичный промотор. В некоторых вариантах осуществления вторая гетерологичная последовательность нуклеиновой кислоты представляет собой репортерный ген, т.е. ген, чья экспрессия может быть определена; репортерные гены включают без ограничения гены, кодирующие глюкуронидазу (GUS), люциферазу, хлорамфениколтрансацетилазу (CAT), зеленый флуоресцентный белок (GFB), щелочную фосфатазу и бета-галактозидазу.
Специфический связывающий агент: агент, который связывается по существу только с определенной мишенью. Таким образом, агент, специфически связывающийся с PD-1, представляет собой агент, который специфически связывается с полипептидом PD-1, но не с неродственными полипептидами. В одном варианте осуществления специфически связывающий агент представляет собой моноклональное или поликлональное антитело, которое специфически связывается с полипептидом PD-1, PD-L1 или PD-L2.
Термин «специфически связывается» обозначает в отношении антигена, такого как PD-1, предпочтительную ассоциацию антитела или другого лиганда целиком или частично с клеткой или тканью, несущей этот антиген, но не с клетками или тканями, у которых этот антиген отсутствует. Конечно, признается, что может возникать определенная степень неспецифического взаимодействия между молекулой и клеткой или тканью, не являющихся мишенью. Тем не менее, специфическое связывание может быть определено как опосредуемое специфическим узнаванием антигена. Хотя селективно взаимодействующие антитела связывают антиген, они могут делать это с низкой аффинностью. Специфическое связывание приводит к намного более сильной ассоциации между антителом (или другим лигандом) и клетками, несущими антиген, по сравнению с ассоциацией между антителом (или другим лигандом) и клетками, не несущими антиген. Специфическое связывание обычно приводит к более чем 2-кратному, такому как более чем 5-кратное, более чем 10-кратное или более чем 100-кратное, увеличению количества связанного антитела или другого лиганда (в единицу времени) с клеткой или тканью, несущими полипептид PD-1, по сравнению с клеткой или тканью с отсутствием полипептида. Специфическое связывание с белком в таких условиях требует антитела, которое отобрано по его специфичности к определенному белку. Разнообразные форматы иммунотестов подходят для отбора антител или других лигандов, специфически иммунореактивных на определенный белок. Например, твердофазные иммунотесты ИФА обычно используются для отбора моноклональных антител, специфически иммунореактивных на белок. См. Harlow & Lane, Antibodies, A Laboratory Manual, Cold Spring Harbor Publications, New York (1988) для описания форматов иммунотестов и условий, которые могут использоваться для определения специфической иммунореактивности.
Т-клетка: белая клетка крови, необходимая для иммунного ответа. Т-клетки включают, но не ограничиваются этим, CD4+ Т-клетки и CD8+ Т-клетки. CD4+ Т-лимфоцит представляет собой иммунную клетку, которая на своей поверхности несет маркер, известный как «кластер дифференцировки 4» (CD4). Эти клетки, также известные как хелперные Т-клетки, помогают организовывать иммунный ответ, включая ответы антител, а также ответы Т-клеток-киллеров. CD8+ Т-клетки несут маркер «кластер дифференцировки 8» (CD8). В одном варианте осуществления CD8+ Т-клетка представляет собой цитотоксический лимфоцит. В другом варианте осуществления CD8+ клетка представляет собой супрессорную Т-клетку. Т-клетка «активирована», когда она может отвечать на специфический интересующий антиген, презентированный на антигенпрезентирующих клетках.
Трансдуцированная/трансфицированная: трансдуцированная клетка представляет собой клетку, в которую введена молекула нуклеиновой кислоты с помощью методов молекулярной биологии. Применяемый в настоящем описании термин трансдукция охватывает все способы, с помощью которых молекула нуклеиновой кислоты может быть введена в такую клетку, включая трансфекцию вирусных векторов, трансформацию плазмидными векторами и введение депротеинизированной ДНК с помощью электропорации, липофекцию и бомбардировку частицами с ускорением.
Вектор: молекула нуклеиновой кислоты, которая в результате введения в клетку-хозяина создает тем самым трансформированную клетку-хозяина. Вектор может включать последовательности нуклеиновой кислоты, которые позволяют ему реплицироваться в клетке-хозяине, такие как ориджин репликации. Вектор может также включать одну или более нуклеиновых кислот, кодирующих селектируемый маркер и другие генетические элементы, известные в данной области техники. Векторы включают плазмидные векторы, включая плазмиды для экспрессии в грамотрицательных и грамположительных бактериальных клетках. Примеры векторов включают векторы для экспрессии в E. coli и Salmonella. Векторы также включают вирусные векторы, такие как, но не ограничиваясь этим, ретровирусные, ортопоксвирусные, авипоксвирусные, фоулпоксвирусные, каприпоксвирусные, суипоксвирусные, аденовирусные векторы, векторы на основе вируса герпеса, альфа вируса, бакуловируса, вируса Синдбиса, вируса коровьей оспы и полиовируса.
Если не объясняется иначе, все технические и научные термины, используемые в настоящем описании, имеют то же самое значение, что и обычно понимаемое специалистом в той области техники, к которой принадлежит это раскрытие. Термины единственного числа включают ссылки на множественное, если в контексте ясно не указано иначе. Сходно, слово «или» предназначено для включения «и», если в контексте ясно не указано иначе. Дополнительно должно быть понятно, что все размеры оснований или размеры аминокислот и все величины молекулярного веса или молекулярной массы, данные для нуклеиновых кислот или полипептидов, являются приблизительными и представлены для описания. Хотя на практике или при проверке этого раскрытия могут быть использованы методы и материалы, сходные или эквивалентные описанным в настоящем документе, подходящие методы и материалы описаны ниже. Термин «заключает в себе» означает «включает». Все публикации, патентные заявки, патенты и другие ссылки, указанные в настоящем описании, включены в полном объеме в качестве ссылок. В случае конфликта заявители будут руководствоваться настоящим описанием, включая объяснения терминов. Кроме того, материалы, методы и примеры являются только иллюстративными и не рассматриваются как лимитирующие.
Антагонисты PD-1
Методы, раскрытые в настоящем описании, включают использование ингибиторов пути PD-1 (антагонистов PD-1). Молекулы PD-1 являются членами надсемейства генов иммуноглобулинов. PD-1 человека обладает внеклеточной областью, содержащей домен надсемейства иммуноглобулинов, трансмембранным доменом и внутриклеточной областью, включающей иммунорецепторный ингибиторный мотив на основе тирозина (ITIM) (Ishida et al., EMBO J. 11:3887, 1992; Shinohara et al., Genomics 23:704, 1994; патент США № 5698520). Эти характеристики также характерны для большого семейства молекул, называемых иммуноингибиторными рецепторами, которое также включает gp49B, PIR-B и рецепторы, ингибирующие киллеров (KIR) (Vivier and Daeron (1997) Immunol. Today 18:286). Не будучи привязанным к теории, считается, что фосфорилированный по тирозиловым остаткам мотив ITIM этих рецепторов взаимодействует с фосфатазой, содержащей S112-домен, что ведет к ингибиторным сигналам. Подкласс этих иммуноингибиторных рецепторов связывается с молекулами главного комплекса гистосовместимости (MHC), так KIR и CTLA4 связываются с B7-1 и B7-2.
У человека PD-1 представляет собой 50-55 кДа трансмембранный рецептор типа I, который первоначально был идентифицирован в линии Т-клеток, подвергшихся индуцированному активацией апоптозу. PD-1 экспрессируется на Т-клетках, В-клетках и макрофагах. Лигандами PD-1 являются члены семейства B7 - PD-лиганд 1 (PD-L1, также известный как B7-H1) и PD-L2 (также известный как B7-DC).
In vivo PD-1 экспрессируется на активированных Т-клетках, В-клетках и моноцитах. Экспериментальные данные связывают взаимодействия PD-1 со своими лигандами с негативной регуляцией центрального и периферического иммунного ответа. В частности, пролиферация Т-клеток дикого типа, но не Т-клеток с недостаточностью PD-1, ингибируется в присутствии PD-L1. Кроме того, мыши с недостаточностью PD-1 проявляют аутоиммунный фенотип.
Пример аминокислотной последовательности PD-1 человека представлен ниже (см. также Ishida et al., EMBO J. 11:3887, 1992; Shinohara et al. Genomics 23:704, 1994; патент США № 5698520):

Пример аминокислотной последовательности PD-1 мыши представлен ниже:

Дополнительные аминокислотные последовательности раскрыты в патенте США № 6808710 и публикациях патентных заявок США №№ 2004/0137577, 2003/0232323, 2003/0166531, 2003/0064380, 2003/0044768, 2003/0039653, 2002/0164600, 2002/0160000, 2002/0110836, 2002/0107363 и 2002/0106730, которые включены в настоящее описание в качестве ссылки. PD-1 является членом надсемейства иммуноглобулинов (Ig), которое содержит единственный Ig V-подобный домен в своей внеклеточной области. Цитоплазматический домен PD-1 содержит два тирозина с наиболее проксимальным по отношению к мембране тирозином (VAYEEL (см. аминокислоты 223-228 SEQ ID NO:2) у PD-1 мыши), локализованным в пределах ITIM (иммунорецепторного ингибиторного мотива на основе тирозина). Присутствие ITIM в PD-1 указывает на то, что функцией этой молекулы является ослабление сигнализации антиген-рецептор с помощью вовлечения цитоплазматических фосфатаз. Белки PD-1 человека и мыши характеризуются приблизительно 60% идентичностью аминокислот с четырьмя консервативными сайтами потенциального N-гликозилирования и остатками, которые определяют домен Ig-V. ITIM в цитоплазматической области и ITIM-подобный мотив, окружающий карбоксиконцевой тирозин (TEYATI (см. аминокислоты 166-181 SEQ ID NO:2) у человека и мыши соответственно), также являются консервативными ортологами у мыши и человека.
PD-1 по своей способности связывать PD-L1 относится к членам семейства молекул CD28/CTLA-4. In vivo, как и CTLA4, PD-1 быстро индуцируется на поверхности Т-клеток в ответ на анти-CD3 (Agata et al. Int. Immunol. 8:765, 1996). Однако в отличие от CTLA4 PD-1 индуцируется также на поверхности В-клеток (в ответ на анти-IgM). PD-1 экспрессируется также на субпопуляции тимоцитов и миелоидных клеток (Agata et al. (1996) выше; Nishimura et al. (1996) Int. Immunol. 8:773).
Анергия Т-клеток сопутствует индукции экспрессии PD-1. В настоящем описании раскрыто, что цитотоксичность Т-клеток может быть увеличена в результате введения в контакт Т-клетки с агентом, который снижает экспрессию или активность PD-1. Более конкретно, в настоящем описании раскрыто, что агент, который снижает экспрессию или активность PD-1, может быть использован для повышения иммунного ответа, такого как на вирусный антиген или опухолевый антиген.
Не будучи привязанными к теории, снижение экспрессии или активности PD-1 ведет к увеличению активности цитотоксических Т-клеток, увеличивая специфический иммунный ответ на инфекционный агент. Для того чтобы Т-клетки отвечали на чужеродные белки, антигенпрезентирующие клетки (APC) должны подать два сигнала покоящимся Т-лимфоцитам. Первый сигнал, который придает специфичность иммунному ответу, передается через рецептор Т-клеток (TCR) после узнавания чужеродного антигенного пептида, презентированного в контексте главного комплекса гистосовместимости (MHC). Второй сигнал, называемый костимулирующим, индуцирует пролиферацию Т-клеток и становление их функциональности. Костимуляция не является ни антиген-специфичной, ни ограниченной MHC и обеспечивается одним или более особыми полипептидами клеточной поверхности, экспрессируемыми APC. Если Т-клетки стимулируются только через Т-клеточный рецептор без получения дополнительного костимулирующего сигнала, они становятся не реагирующими, переходят в состояние анергии или гибнут, что ведет к негативной модуляции иммунного ответа.
Белки CD80 (B7-1) и CD86 (B7-2), экспрессируемые на APC, являются необходимыми костимуляторными полипептидами. В то время как B7-2 играет преобладающую роль в процессе первичных иммунных ответов, B7-1 позитивно регулируется позже в процессе иммунного ответа на пролонгированные первичные ответы Т-клеток или вторичные костимуляторные ответы Т-клеток. Полипептиды B7 способны обеспечивать иммунные клетки костимуляторными или ингибиторными сигналами для стимуляции или ингибирования ответов иммунных клеток. Например, в состоянии связывания с костимуляторным рецептором PD-L1 (B7-4) индуцирует костимуляцию иммунных клеток или, когда присутствует в растворимой форме, ингибирует костимуляцию иммунных клеток. В состоянии связывания с ингибиторным рецептором молекулы PD-L1 могут передавать иммунной клетке ингибиторный сигнал. Примеры членов семейства B7 включают B7-1, B7-2, B7-3 (узнаваемый антителом BB-1), B7h (PD-L1) и B7-4, а также их растворимые фрагменты или производные. Члены семейства B7 связываются с одним или более рецепторов на иммунной клетке, таким как CTLA4, CD28, ICOS, PD-1 и/или другими рецепторами, и в зависимости от рецептора обладают способностью передавать ингибиторный сигнал или костимуляторный сигнал иммунной клетке.
CD28 представляет собой рецептор, который конститутивно экспрессируется на покоящихся Т-клетках. После сигнализации через Т-клеточный рецептор связывание CD28 и передача костимуляторного сигнала индуцируют пролиферацию Т-клеток и секрецию ИЛ-2. CTLA4 (CD152), рецептор, гомологичный CD28, отсутствует на покоящихся Т-клетках, но его экспрессия индуцируется после активации Т-клеток. CTLA4 играет роль в негативной регуляции Т-клеточных ответов. ICOS, полипептид, родственный CD28 и CTLA4, вовлечен в продукцию ИЛ-10. PD-1, рецептор, с которым связываются PD-L1 и PD-L2, также быстро индуцируется на поверхности Т-клеток. PD-1 также экспрессируется на поверхности В-клеток (в ответ на анти-IgM) и на субпопуляции тимоцитов и миелоидных клеток.
Контактирование PD-1 (например, с помощью поперечной сшивки или агрегации) ведет к передаче ингибиторного сигнала в иммунной клетке, приводя к снижению иммунных ответов, сопровождающихся ростом анергии иммунных клеток. Члены семейства PD-1 связываются с одним или более рецепторов, таких как PD-L1 и PD-L2 на антигенпрезентирующих клетках. PD-L1 и PD-L2, оба из которых являются полипептидными лигандами PD-1 человека, являются членами семейства B7 полипептидов (см. выше). Каждый лиганд PD-1 содержит сигнальную последовательность, домен IgV, домен IgC, трансмембранный домен и короткий цитоплазматический хвост. In vivo эти лиганды, как показано, экспрессируются в плаценте, селезенке, лимфатических узлах, тимусе и сердце. PD-L2 экспрессируется также в поджелудочной железе, легких и печени, тогда как PD-L1 экспрессируется в печени плодов, активированных Т-клетках и эндотелиальных клетках. Оба лиганда PD-1 положительно регулируются на активированных моноцитах и дендритных клетках.
Пример аминокислотной последовательности PD-L1 (GENBANK®, № депозита AAG18508, доступный с 4 октября 2000 г.) представлен ниже:

Пример аминокислотной последовательности предшественника PD-L2 (GENBANK®, № депозита AAK153370, доступный с 8 апреля 2002 г.) представлен ниже:

Пример варианта аминокислотной последовательности предшественника PD-L2 (GENBANK®, № депозита Q9BQ51, доступный с 12 декабря 2006 г.) представлен ниже:

Антагонисты PD-1 включают агенты, которые снижают экспрессию или активность лиганда 1 PD (PD-L1) или лиганда 2 PD (PD-L2) или снижают взаимодействие между PD-1 и PD-L1 или взаимодействие между PD-1 и PD-L2. Примеры соединений включают антитела (такие как антитело против PD-1, антитело против PD-L1 и антитело против PD-L2), молекулы РНКи (такие как молекулы РНКи анти-PD-1, РНКи анти-PD-L1 и РНКи анти-PD-L2), антисмысловые молекулы (такие как антисмысловая РНК анти-PD-1, антисмысловая РНК анти-PD-L1 и антисмысловая РНК анти-PD-L2), доминантно негативные белки (такие как доминантно негативный белок PD-1, доминантно негативный белок PD-L1 и доминантно негативный белок PD-L2) и низкомолекулярные ингибиторы.
Антагонист PD-1 представляет собой любой агент, обладающий способностью снижать экспрессию или активность PD-1 в клетке. Экспрессия или активность PD-1 снижается по меньшей мере приблизительно на 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или 100% по сравнению с такой экспрессией или активностью у контроля. Примеры снижения активности составляют по меньшей мере приблизительно на 50%, по меньшей мере приблизительно на 60%, по меньшей мере приблизительно на 70%, по меньшей мере приблизительно на 80%, по меньшей мере приблизительно на 90%, по меньшей мере приблизительно на 95% или до полного отсутствия определяемой активности. В одном примере контроль представляет собой клетку, которая не обрабатывалась антагонистом PD-1. В другом примере контроль представляет собой стандартную величину или клетку, контактировавшую с агентом, таким как носитель, известный как не обладающий влиянием на активность PD-1. Экспрессия или активность PD-1 может быть определена с помощью любого стандартного метода, известного в данной области техники, включая описанные в настоящем документе. Необязательно антагонист PD-1 ингибирует или снижает связывание PD-1 с PD-L1, PD-L2 или с обоими.
А. Антитела
Антитела, которые специфически связывают PD-1, PD-L1 или PD-L2 (или их сочетание), используются в раскрытых в настоящем описании методах. Антитела включают моноклональные антитела, гуманизированные антитела, деиммунизированные антитела и гибридные белки-иммуноглобулины (Ig). Поликлональные антитела анти-PD-1, анти-PD-L1 или анти-PD-L2 могут быть получены специалистом в данной области техники, например, с помощью иммунизации подходящего индивидуума (такого как объект ветеринарии) лигандом PD-1 или PD-1 иммуногеном. Титр антител анти-PD-1, анти-PD-L1 или анти-PD-L2 у иммунизированного индивидуума может быть прослежен с течением времени с помощью стандартных методов, таких как твердофазный иммуноферментный анализ (ИФА) с использованием иммобилизованного лиганда PD-1 или полипептида PD-1.
В одном примере молекулы антител, которые специфически связывают PD-1, PD-L1 или PD-L2 (или их сочетание), могут быть выделены от млекопитающего (например, из сыворотки) и дополнительно очищены с помощью методов, известных специалисту в данной области техники. Например, антитела могут быть очищены с использованием хроматографии с протеином А для выделения антител IgG.
Клетки, продуцирующие антитела, могут быть получены от индивидуума и использованы для получения моноклональных антител с помощью стандартных методов (см. Kohler and Milstein Nature 256:495-49, 1995; Brown et al., J. Immunol. 127:539-46, 1981; Cole et al., Monoclonal Antibodies and Cancer Therapy, Alan R. Liss, Inc., pp.77 96, 1985; Gefter, M. L. et al. (1977) Somatic Cell Genet. 3:231 36; Kenneth, R. H. in Monoclonal Antibodies: A New Dimension In Biological Analyses. Plenum Publishing Corp., New York, N.Y. (1980); Kozbor et al. Immunol. Today 4:72, 1983; Lerner, E. A. (1981) Yale J. Biol. Med. 54:387-402; Yeh et al., Proc. Natl. Acad. Sci. 76:2927-31, 1976). В одном примере иммортализованную клеточную линию (обычно миелому) сливают с лимфоцитами (обычно спленоцитами) от млекопитающего, иммунизированного PD-1, PD-L1 или PD-L2, и проводят скрининг культуральных супернатантов полученных гибридомных клеток для идентификации гибридомы, продуцирующей моноклональное антитело, которое специфически связывается с интересующим полипептидом.
В одном варианте осуществления при получении гибридомы иммортализованная клеточная линия (такая как миеломная клеточная линия) происходит от того же вида млекопитающих, что и лимфоциты. Например, мышиные гибридомы могут быть созданы путем слияния лимфоцитов от мыши, иммунизированной пептидом PD-1, PD-L1 или PD-L2, с иммортализованной клеточной линией мыши. В одном примере используется миеломная клеточная линия мыши, которая чувствительна к культуральной среде, содержащей гипоксантин, аминоптерин и тимидин («HAT-среда»). Любое количество миеломных клеточных линий может использоваться в качестве партнера по слиянию в соответствии со стандартными методами, включая, например, миеломные линии P3-NS1/1-Ag4-1, P3-x63-Ag8.653 или Sp2/O-Agl4, которые доступны от американской коллекции типов культур (ATCC), Rockville, Md. HAT-чувствительные миеломные клетки мыши могут быть слиты с мышиными спленоцитами с использованием полиэтиленгликоля («ПЭГ»). Затем проводят селекцию гибридомных клеток, полученных в результате слияния, с использованием HAT-среды, которая уничтожает неслитые (и непродуктивно слитые) миеломные клетки. Гибридомные клетки, продуцирующие интересующее моноклональное антитело, могут быть определены, например, с помощью скрининга супернатантов гибридомных культур на продукцию антител, которые связывают молекулу PD-1, PD-L1 или PD-L2, например, с использованием иммунологического теста (такого как твердофазный иммуноферментный анализ (ИФА) или радиоиммуноанализ (РИА)).
В качестве альтернативы получения гибридом скринингом моноклональных антител, моноклональное антитело, которое специфически связывает PD-1, PD-L1 или PD-L2, может быть идентифицировано и выделено путем скрининга рекомбинантной комбинаторной библиотеки иммуноглобулинов (такой как библиотека антител в формате фагового дисплея) с PD-1, PD-L1 или PD-L2 для выделения членов библиотеки иммуноглобулинов, которые специфически связывают полипептид. Наборы для создания и скрининга библиотек фагового дисплея имеются в продаже (например, но не ограничиваясь этим, у Pharmacia и Stratagene). Примеры методов и реагентов, особенно подходящих для использования при создании и скрининге библиотеки фагового дисплея, могут быть найдены, например, в патенте США № 5223409; публикации PCT № WO 90/02809; публикации PCT № WO 91/17271; публикации PCT № WO 92/18619; публикации PCT WO 92/20791; публикации PCT № WO 92/15679; публикации PCT № WO 92/01047; публикации PCT WO 93/01288; публикации PCT № WO 92/09690; Barbas et al., Proc. Natl. Acad. Sci. USA 88:7978-7982, 1991; Hoogenboom et al., Nucleic Acids Res. 19:4133-4137, 1991.
Аминокислотная последовательность антител, которые связывают PD-1, раскрыта, например, в патентной публикации США № 2006/0210567, которая включена в настоящее описание в качестве ссылки. Антитела, которые связывают PD-1, раскрыты также в патентной публикации США № 2006/0034826, которая также включена в настоящее описание в качестве ссылки. В некоторых примерах антитело специфически связывает PD-1 или лиганд PD-1 или PD-2 с константой сродства по меньшей мере 107 М-1, такой как по меньшей мере 108 М-1, по меньшей мере 5×108 М-1 или по меньшей мере 109 М-1.
В одном примере определяют последовательность областей, определяющих специфичность каждой CDR. Остатки, находящиеся вне CDR (сайты, не контактирующие с лигандом), заменяют. Например, в любой из последовательностей, как в таблице выше, не менее чем одна, две или три аминокислоты могут быть заменены. Получение гибридных антител, которые включают область каркаса от одного антитела и области CDR от отличного антитела, хорошо известно в данной области техники. Например, гуманизированные антитела могут быть получены обычным способом. Антитело или фрагмент антитела может представлять собой гуманизированный иммуноглобулин, обладающий областями, определяющими комплементарность (CDR) от донорского моноклонального антитела, которое связывает PD-1, PD-L1 или PD-L2, и каркасами вариабельных областей тяжелой и легкой цепей от каркасов тяжелой и легкой цепей акцепторного иммуноглобулина человека. Обычно гуманизированный иммуноглобулин специфически связывается с PD-1, PD-L1 или PD-L2 с константой сродства по меньшей мере 107 М-1, такой как по меньшей мере 108 М-1, по меньшей мере 5×108 М-1 или по меньшей мере 109 М-1.
Гуманизированные моноклональные антитела могут быть получены с помощью переноса донорских областей, определяющих комплементарность (CDR), из вариабельных областей тяжелой и легкой цепей донорского иммуноглобулина мыши (такого как PD-1, PD-L1 или PD-L2) в вариабельный домен человека и последующей замены остатков человека в областях каркаса, когда требуется сохранить аффинность. Использование компонентов антител, происходящих от гуманизированных моноклональных антител, избавляет от потенциальных проблем, связанных с иммуногенностью константных областей донорского антитела. Методы получения гуманизированных моноклональных антител описаны, например, Jones et al., Nature 321:522, 1986; Riechmann et al., Nature 332:323, 1988; Verhoeyen et al., Science 239:1534, 1988; Carter et al., Proc. Natl. Acad. Sci. U.S.A. 89:4285, 1992; Sandhu, Crit. Rev. Biotech. 12:437, 1992; и Singer et al., J. Immunol. 150:2844, 1993. Антитело может быть любого изотипа, но в некоторых вариантах осуществления антитело представляет собой IgG, включая, но не ограничиваясь этим, IgG1, IgG2, IgG3 и IgG4.
В одном варианте осуществления последовательность каркаса вариабельной области тяжелой цепи гуманизированного иммуноглобулина может быть по меньшей мере приблизительно на 65% идентична последовательности каркаса вариабельной области тяжелой цепи донорского иммуноглобулина. Таким образом, последовательность каркаса вариабельной области тяжелой цепи гуманизированного иммуноглобулина может быть по меньшей мере приблизительно на 75%, по меньшей мере приблизительно на 85%, по меньшей мере приблизительно на 90% или по меньшей мере приблизительно на 95% идентична последовательности каркаса вариабельной области тяжелой цепи донорского иммуноглобулина. Области каркаса человека и мутации, которые могут быть сделаны областях каркаса гуманизированного антитела, известны в данной области техники (см., например, в патенте США № 5585089, который включен в настоящее описание в качестве ссылки).
Примерами антител человека являются LEN и 21/28 CL. Последовательности каркасов тяжелой и легкой цепей известны в данной области техники. Примеры каркасов легких цепей Mab LEN человека имеют следующие последовательности:

Примеры каркасов тяжелых цепей Mab 21/28' CL человека имеют следующие последовательности:

Антитела, такие как мышиные моноклональные антитела, гибридные антитела и гуманизированные антитела, включают полноразмерные молекулы, а также их фрагменты, такие как Fab, F(ab')2 и Fv, которые включают вариабельную область тяжелой цепи и легкой цепи и способны связывать специфические эпитопные детерминанты. Эти фрагменты антител сохраняют некоторую способность селективно связываться с их антигеном или рецептором. Эти фрагменты включают:
(1) Fab, фрагмент, который содержит одновалентный антигенсвязывающий фрагмент молекулы антитела, может быть получен путем гидролиза целого антитела ферментом папаином с получением интактной легкой цепи и части одной тяжелой цепи;
(2) Fab', фрагмент молекулы антитела, который может быть получен путем обработки целого антитела пепсином с последующим восстановлением с получением интактной легкой цепи и части одной тяжелой цепи; на молекулу антитела получают два Fab' фрагмента;
(3) (Fab')2, фрагмент антитела, который может быть получен путем обработки целого антитела ферментом пепсином без последующего восстановления; (Fab')2 представляет собой димер двух Fab' фрагментов, удерживаемых вместе двумя дисульфидными связями;
(4) Fv, фрагмент, созданный с помощью генной инженерии, содержащий вариабельную область легкой цепи и вариабельную область тяжелой цепи, экспрессируемых как две цепи; и
(5) одноцепочечное антитело (такое как scFv), определяемое как молекула, созданная с помощью генной инженерии, содержащая вариабельную область легкой цепи, вариабельную область тяжелой цепи, связанные подходящим полипептидным линкером, в виде генетически созданной гибридной одноцепочечной молекулы.
Методы создания таких фрагментов известны в данной области техники (см., например, Harlow and Lane, Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory, New York, 1988). В некоторых примерах вариабельная область включает вариабельную область легкой цепи и вариабельную область тяжелой цепи, экспрессируемые как индивидуальные полипептиды. Антитела Fv обычно составляют 25 кДа и содержат полный антигенсвязывающий сайт с тремя CDRs на каждую тяжелую цепь и каждую легкую цепь. Для получения этих антител VH и VL могут быть экспрессированы с двух индивидуальных конструктов нуклеиновых кислот в клетке-хозяине. Если VH и VL экспрессируются не в контакте, цепи антитела Fv обычно удерживаются вместе с помощью нековалентных взаимодействий. Однако эти цепи имеют тенденцию к диссоциации при разведении, поэтому разработаны методы поперечной сшивки цепей с помощью глутаральдегида, внутримолекулярных дисульфидных связей или пептидного линкера. Таким образом, в одном примере Fv может представлять собой Fv, стабилизированный дисульфидными связями (dsFv), где вариабельная область тяжелой цепи и вариабельная область легкой цепи химически соединены дисульфидными связями.
В дополнительном примере фрагменты Fv включают цепи VH и VL, связанные пептидным линкером. Эти одноцепочечные антигенсвязывающие белки (scFv) получают с помощью конструирования структурного гена, включающего последовательности ДНК, кодирующие домены VH и VL, связанные олигонуклеотидом. Структурный ген вставляют в экспрессионный вектор, который затем вводят в клетку-хозяина, такую как E. coli. Рекомбинантные клетки-хозяева синтезируют одну полипептидную цепь с линкерным пептидом, соединяющим два домена V. Методы получения scFvs известны в данной области техники (см. Whitlow et al., Methods: a Companion to Methods in Enzymology, Vol.2, page 97, 1991; Bird et al., Science 242:423, 1988; патент США № 4946778; Pack et al., Bio/Technology 11:1271, 1993; и Sandhu, выше).
Фрагменты антител могут быть получены с помощью протеолитического гидролиза антитела или путем экспрессии ДНК, кодирующей фрагмент, в E. coli. Фрагменты антител могут быть получены путем гидролиза целых антител пепсином или папаином с помощью общепринятых методов. Например, фрагменты антител могут быть получены с помощью ферментативного расщепления антител пепсином с получением 5S фрагмента, обозначаемого (Fab')2. Этот фрагмент может быть дополнительно расщеплен с использованием тиолового восстанавливающего агента и, необязательно, блокирующей группы для сульфгидрильных групп, возникающих в результате расщепления дисульфидных связей, с получением одновалентных 3,5S Fab' фрагментов. Альтернативно ферментативное расщепление с использованием пепсина прямо дает два одновалентных Fab' фрагмента и Fc фрагмент (см. патент США № 4036945 и патент США № 4331647, а также содержащиеся там ссылки; Nisonhoff et al., Arch. Biochem. Biophys. 89:230, 1960; Porter, Biochem. J. 73:119, 1959; Edelman et al., Methods in Enzymology, Vol.1, page 422, Academic Press, 1967; и Coligan et al. в разделах 2.8.1-2.8.10 и 2.10.1-2.10.4).
Могут использоваться другие методы расщепления антител, такие как разделение тяжелых цепей с образованием одновалентных фрагментов легкая-тяжелая цепь, дополнительное расщепление фрагментов или другие ферментативные, химические или генетические методы, до тех пор, пока фрагменты связывают антиген, который узнается интактным антителом.
Специалист в данной области техники должен сознавать, что могут быть получены консервативные варианты антител. Такие консервативные варианты, используемые во фрагментах антител, таких как dsFv фрагменты, или в scFv фрагментах, должны сохранять аминокислотные остатки, необходимые для правильной укладки областей VH и VL и стабилизации между ними, и должны сохранять зарядные характеристики остатков, для того чтобы сохранить низкую pI и низкую токсичность молекул. Для увеличения выхода в областях VH и VL могут быть сделаны аминокислотные замены (такие как не больше одной, не больше двух, не больше трех, не больше четырех или не больше пяти аминокислотных замен). Таблицы консервативных аминокислотных замен, представляющие функционально сходные аминокислоты, хорошо известны специалисту в данной области техники. Следующие шесть групп представляют собой примеры аминокислот, которые рассматриваются как являющиеся консервативными заменами друг для друга:
1) аланин (A), серин (S), треонин (T);
2) аспарагиновая кислота (D), глютаминовая кислота (E);
3) аспарагин (N), глутамин (Q);
4) аргинин (R), лизин (K);
5) изолейцин (I), лейцин (L), метионин (M), валин (V); и
6) фенилаланин (F), тирозин (Y), триптофан (W).
Таким образом, специалист в данной области техники может с легкостью изучить аминокислотную последовательность интересующего антитела, определить положение одной или более аминокислот из краткой таблицы, представленной выше, идентифицировать консервативную замену и получить консервативный вариант с использованием хорошо известных методов молекулярной биологии.
Эффекторные молекулы, такие как терапевтические, диагностические части или части для определения, могут быть присоединены к антителу, которое специфически связывает PD-1, PD-L1 или PD-L2, с использованием ряда способов, известных специалистам в данной области техники. Могут быть использованы как ковалентные, так и нековалентные способы присоединения. Процедура присоединения эффекторной молекулы к антителу варьируется в соответствии с химической структурой эффектора. Полипептиды обычно содержат разнообразные функциональные группы; такие как группы карбоновой кислоты (COOH), свободного амина (-NH2) или сульфгидрильные (-SH) группы, которые доступны для взаимодействия с подходящей функциональной группой на антителе с результирующим привязыванием эффекторной молекулы. Альтернативно антитело дериватизируют для экспонирования или присоединения дополнительных, реакционно-способных функциональных групп. Дериватизация может включать присоединение любой из ряда линкерных молекул, таких как доступные от Pierce Chemical Company, Rockford, IL. Линкером может быть любая молекула, используемая для соединения антитела с эффекторной молекулой. Линкер способен образовывать ковалентные связи как с антителом, так и с эффекторной молекулой. Подходящие линкеры хорошо известны специалистам в данной области техники и включают, но не ограничиваются этим, углеродные линкеры с линейной или разветвленной цепью, гетероциклические углеродные линкеры или пептидные линкеры. Когда антитело и эффекторная молекула являются полипептидами, линкеры могут быть присоединены к составляющим аминокислотам через их боковые группы (например, через дисульфидную связь к цистеину) или к амино- и карбоксильным группам альфа углерода концевых аминокислот.
Последовательности нуклеиновых кислот, кодирующие антитела, могут быть получены с помощью любого подходящего метода, включая, например, клонирование соответствующих последовательностей или с помощью прямого химического синтеза такими методами, как фосфотриэфирный метод Narang et al., Meth. Enzymol. 68:90-99, 1979; фосфодиэфирный метод Brown et al., Meth. Enzymol. 68:109-151, 1979; диэтилфосфорамидитный метод Beaucage et al., Tetra. Lett. 22:1859-1862, 1981; твердофазный фосфорамидиттриэфирный метод, описанный Beaucage & Caruthers, Tetra. Letts. 22(20):1859-1862, 1981, например, с использованием автоматического синтезатора, как описано, например, в Needham-VanDevanter et al., Nucl. Acids Res. 12:6159-6168, 1984; и метода с твердой подложкой из патента США № 4458066. При химическом синтезе продуцируется одноцепочечный олигонуклеотид. Он может быть превращен в двухцепочечную ДНК с помощью гибридизации с комплементарной последовательностью или путем полимеризации ДНК-полимеразой с использованием одной цепи в качестве матрицы. Специалист в данной области техники должен знать, что в то время как химический синтез ДНК обычно лимитирован последовательностями из приблизительно 100 оснований, более длинные последовательности могут быть получены с помощью лигирования более коротких последовательностей.
Иллюстративные последовательности, кодирующие нуклеиновые кислоты, для кодирования антитела, которое специфически связывает PD-1, PD-L1 или PD-L2, могут быть получены с помощью способов клонирования. Примеры подходящих методов клонирования и секвенирования и инструкции, достаточные для указаний специалистам в данной области техники, для проведения многих задач по клонированию представлены в Sambrook et al. выше, Berger and Kimmel (eds.) выше и Ausubel выше. Информация о продукте от производителей биологических реагентов и лабораторного оборудования также дает полезные сведения. Такие производители включают SIGMA Chemical Company (Saint Louis, MO), R&D Systems (Minneapolis, MN), Pharmacia Amersham (Piscataway, NJ), CLONTECH Laboratories, Inc. (Palo Alto, CA), Chem Genes Corp., Aldrich Chemical Company (Milwaukee, WI), Glen Research, Inc., GIBCO BRL Life Technologies, Inc. (Gaithersburg, MD), Fluka Chemica-Biochemika Analytika (Fluka Chemie AG, Buchs, Switzerland), Invitrogen (San Diego, CA) и Applied Biosystems (Foster City, CA), а также многие другие коммерческие источники, известные специалисту в данной области техники.
Нуклеиновые кислоты могут быть также получены с помощью методов амплификации. Методы амплификации включают полимеразную цепную реакцию (ПЦР), лигазную цепную реакцию (ЛЦР), систему амплификации на основе транскрипции (TAS), систему самоподдерживаемой репликации последовательности (3SR). Большое разнообразие методов клонирования, клеток-хозяев и методов амплификации in vitro хорошо известно специалистам в данной области техники.
В одном примере используемое антитело получают путем вставки кДНК, которая кодирует вариабельную область от антитела, которое специфически связывает PD-1, PD-L1 или PD-L2, в вектор, который включает кДНК, кодирующую эффекторную молекулу (EM). Вставку делают так, что вариабельная область и EM считываются в рамке так, что продуцируется один непрерывный полипептид. Таким образом, кодируемый полипептид содержит функциональную область Fv и функциональную область EM. В одном варианте осуществления кДНК, кодирующую определяемый маркер (такой как фермент), лигируют с scFv так, что маркер локализуется на карбоксильном конце scFv. В другом примере определяемый маркер локализуется на аминоконце scFv. В дополнительном примере кДНК, кодирующую определяемый маркер, лигируют с вариабельной областью тяжелой цепи антитела, которое специфически связывает PD-1, PD-L1 или PD-L2, так, что маркер локализуется на карбоксильном конце вариабельной области тяжелой цепи. Вариабельная область тяжелой цепи может затем лигироваться с вариабельной областью легкой цепи антитела, которое специфически связывает PD-1, PD-L1 или PD-L2, с использованием дисульфидных связей. В еще одном примере кДНК, кодирующую маркер, лигируют с вариабельной областью легкой цепи антитела, которое специфически связывает PD-1, PD-L1 или PD-L2, так, что маркер локализуется на карбоксильном конце вариабельной области легкой цепи. Вариабельная область легкой цепи может затем лигироваться с вариабельной областью тяжелой цепи антитела, которое специфически связывает PD-1, PD-L1 или PD-L2, с использованием дисульфидных связей.
После выделения и клонирования нуклеиновых кислот, кодирующих антитело или его функциональный фрагмент, белок может быть экспрессирован в созданной рекомбинантной клетке, такой как бактерии, клетки растений, дрожжи, клетки насекомых и млекопитающих. Одна или более последовательностей ДНК, кодирующих антитело или его функциональный фрагмент, могут быть экспрессированы in vitro с помощью переноса ДНК в подходящую клетку-хозяина. Клетка может быть прокариотной или эукариотной. Термин также включает любых потомков подлежащей клетки-хозяина. Понятно, что все потомки могут не быть идентичными родительской клетке, так как могут происходить мутации, которые возникают в процессе репликации. Методы стабильного переноса, означающего, что чужеродная ДНК постоянно поддерживается в хозяине, известны в данной области техники.
Полинуклеотидные последовательности, кодирующие антитело или его функциональный фрагмент, могут быть оперативно связаны с последовательностями, контролирующими экспрессию. Последовательность, контролирующую экспрессию, оперативно связанную с кодирующей последовательностью, лигируют так, что экспрессия кодирующей последовательности достигается в условиях, совместимых с последовательностями, контролирующими экспрессию. Последовательности, контролирующие экспрессию, включают, но не ограничиваются этим, подходящие промоторы, энхансеры, терминаторы транскрипции, инициирующий кодон (т.е. ATG) впереди гена, кодирующего белок, сигнал сплайсинга для интронов, элементы, поддерживающие правильную рамку считывания этого гена для возможности корректной трансляции мРНК, и стоп-кодоны.
Полинуклеотидные последовательности, кодирующие антитело или его функциональный фрагмент, могут быть вставлены в экспрессионный вектор, включая, но не ограничиваясь этим, плазмиду, вирус или другой носитель, с которым можно манипулировать для возможности вставки или включения последовательностей и который может экспрессироваться либо в прокариотах, либо в эукариотах. Хозяева могут включать организмы микробов, дрожжей, насекомых и млекопитающих. Методы экспрессии последовательностей ДНК, имеющих эукариотные или вирусные последовательности, в прокариотах хорошо известны в данной области техники. Биологически функционирующие вирусные и плазмидные ДНК векторы, способные экспрессироваться и реплицироваться в хозяине, хорошо известны в данной области техники.
Трансформация клетки-хозяина рекомбинантной ДНК может быть осуществлена с помощью традиционных методов, которые хорошо известны в данной области техники. Когда хозяин является прокариотом, таким как E. coli, компетентные клетки, которые способны захватывать ДНК, могут быть получены из клеток, собранных после фазы экспоненциального роста и затем обработанных методом с CaCl2 с использованием процедур, хорошо известных в данной области техники. Альтернативно могут быть использованы MgCl2 или RbCl. Трансформация может быть также осуществлена после образования протопласта клетки-хозяина, если это желательно, или с помощью электропорации.
Когда хозяином является эукариот, могут быть использованы такие методы трансфекции ДНК, как совместная преципитация с фосфатом кальция, общепринятые механические процедуры, такие как микроинъекция, электропорация, вставка плазмиды, инкапсулированной в липосомы, или вирусные векторы. Клетки эукариот могут также котрансформироваться с полинуклеотидными последовательностями, кодирующими антитело или его функциональный фрагмент, и второй чужеродной молекулой ДНК, кодирующей селектируемый фенотип, такой как ген тимидинкиназы простого герпеса. Другим методом является использование эукариотного вирусного вектора, такого как вирус 40 обезьян (SV40) или вирус папилломы быка, для временного инфицирования или трансформирования эукариотных клеток и экспрессии белка (см., например, Eukaryotic Viral Vectors, Cold Spring Harbor Laboratory, Gluzman ed., 1982). Специалист в данной области техники может легко использовать экспрессионные системы, такие как используемые плазмиды и векторы, для продукции белков в клетках, включая клетки высших эукариот, таких как клеточные линии COS, CHO, HeLa и миеломная клеточная линия.
Выделение и очистка рекомбинантно экспрессированного полипептида может быть осуществлена с помощью общепринятых методов, включая препаративную хроматографию и иммунологические разделения. После экспрессии рекомбинантные антитела могут быть очищены в соответствии со стандартными процедурами данной области техники, включая осаждение сульфатом аммония, аффинную колоночную хроматографию, колоночную хроматографию и тому подобное (см. в целом R. Scopes, Protein Purification, Springer-Verlag, N. Y., 1982). В настоящем описании раскрыты по существу чистые композиции с гомогенностью, по меньшей мере приблизительно от 90 до 95%, и от 98 до 99% или более гомогенности может быть использовано в фармацевтических целях. После очистки, частичной или до гомогенности, как это желательно, если полипептиды будут использоваться терапевтически, они должны быть по существу свободными от эндотоксина.
Методы экспрессии одноцепочечных антител, включая одноцепочечные антитела из бактерий, таких как E. coli, и/или их повторной укладки до подходящей активной формы описаны и хорошо известны и применимы к антителам, раскрытым в настоящем описании. См. Buchner et al., Anal. Biochem. 205:263-270, 1992; Pluckthun, Biotechnology 9:545, 1991; Huse et al., Science 246:1275, 1989, и Ward et al., Nature 341:544, 1989, все включены в настоящее описание в качестве ссылки.
Часто функционально гетерологичные белки из E. coli или других бактерий выделяют из телец включений, и это требует солюбилизации с использованием сильных денатурирующих агентов и последующей повторной укладки. При стадии солюбилизации, как хорошо известно в данной области техники, для разобщения дисульфидных связей должен присутствовать восстанавливающий агент. Примером буфера с восстанавливающим агентом является: 0,1 М трис pH 8, 6M гуанидин, 2 мМ ЭДТА, 0,3М DTE (дитиоэритритол). Повторное окисление дисульфидных связей может быть осуществлено в присутствии тиоловых реагентов низкой молекулярной массы в восстановленной и окисленной форме, как описано в статье Saxena et al., Biochemistry 9:5015-5021, 1970, включенной в настоящее описание в качестве ссылки, и особенно как описано Buchner et al., выше.
Ренатурация обычно достигается разведением (например, 100-кратным) денатурированного и восстановленного белка в буфере для повторной укладки. Примером буфера является 0,1 М трис, pH 8,0, 0,5 М L-аргинин, 8 мМ окисленный глутатион (GSSG) и 2 мМ ЭДТА.
При модификации протокола очистки двухцепочечного антитела области тяжелой и легкой цепей отдельно солюбилизируются и восстанавливаются и затем объединяются в растворе для повторной укладки. Примером получаемого выхода является такой, когда при смешивании этих двух белков в молярном отношении не превышается такой молярный избыток одного белка над другим, как 5-кратный. Желательно после завершения окислительно-восстановительной перетасовки добавлять в раствор для повторной укладки избыток окисленного глутатиона или других окисляющих соединений низкой молекулярной массы.
Кроме рекомбинантных методов раскрытые в настоящем описании антитела и их функциональные фрагменты могут быть также созданы полностью или частично с использованием стандартного синтеза пептидов. Твердофазный синтез полипептидов менее приблизительно 50 аминокислот в длину может быть осуществлен путем присоединения C-концевой аминокислоты последовательности к нерастворимой подложке с последующим последовательным добавлением оставшихся аминокислот в последовательности. Методы твердофазного синтеза описаны Barany & Merrifield, The Peptides: Analysis, Synthesis, Biology. Vol.2: Special Methods in Peptide Synthesis, Part A. pp.3-284; Merrifield et al., J. Am. Chem. Soc. 85:2149-2156, 1963, и Stewart et al., Solid Phase Peptide Synthesis, 2nd ed., Pierce Chem. Co., Rockford, 111, 1984. Белки большей длины могут быть синтезированы путем конденсации амино- и карбоксильных концов более коротких фрагментов. Методы образования пептидных связей путем активации карбокситерминального конца (такие как с помощью использования присоединяющего реагента N,N'-дициклогексилкарбодиимид) хорошо известны в данной области техники.
B. Ингибиторные нуклеиновые кислоты
Ингибиторные нуклеиновые кислоты, которые снижают экспрессию и/или активность PD-1, PD-L1 или PD-L2, также могут быть использованы в методах, раскрытых в настоящем описании. В одном варианте осуществления это малая ингибиторная РНК (миРНК) для интерференции или ингибирования экспрессии гена-мишени. Последовательности нуклеиновой кислоты, кодирующие PD-1, PD-L1 или PD-L2, раскрыты в GENBANK®, №№ депозитов NM_005018, AF344424, NP_079515 и NP_054862.
Обычно миРНК создают путем расщепления относительно длинных молекул двухцепочечной РНК ферментами Dicer или DCL (Zamore, Science, 296:1265-1269, 2002; Bernstein et al., Nature, 409:363-366, 2001). У животных и растений миРНК собираются в RISC и управляют рибонуклеолитической активностью RISC, специфичной в отношении последовательности, тем самым приводя к расщеплению мРНК или других РНК молекул-мишеней в цитоплазме. В ядре миРНК также управляют гистоном, ассоциированным с гетерохроматином, и метилированием ДНК, что приводит к транскрипционному молчанию индивидуальных генов или больших доменов хроматина. миРНК PD-1 имеется в продаже, например, от Santa Cruz Biotechnology, Inc.
В настоящем раскрытии предлагается РНК, которая подходит для интерференции или ингибирования экспрессии гена-мишени, эта РНК включает двухцепочечную РНК от приблизительно 15 до приблизительно 40 нуклеотидов, содержащую от 0 до 5-нуклеотидных 3'- и/или 5'-выступов на каждой цепи. Последовательность РНК по существу идентична части мРНК или транскрипта гена-мишени, такого как PD-1, PD-L1 или PD-L2, для которого желательна интерференция или ингибирование. Для целей этого раскрытия последовательность РНК, «по существу идентичная» специфической части мРНК или транскрипта гена-мишени, для которого желательна интерференция или ингибирование, отличается не более чем на приблизительно 30 процентов и в некоторых вариантах осуществления не более чем на приблизительно 10 процентов от конкретной части мРНК или транскрипта гена-мишени. В конкретных вариантах осуществления последовательность РНК целиком идентична конкретной части мРНК или транскрипта гена-мишени.
Таким образом, миРНК, раскрытые в настоящем описании, включают двухцепочечную РНК от приблизительно 15 до приблизительно 40 нуклеотидов и с 3'- или 5'-выступами, имеющими в длину от 0 до 5-нуклеотидов на каждой цепи, где последовательность двухцепочечной РНК по существу идентична (см. выше) части мРНК или транскрипта нуклеиновой кислоты, кодирующей PD-1, PD-L1 или PD-L2. В конкретных примерах двухцепочечная РНК содержит от приблизительно 19 до приблизительно 25 нуклеотидов, например, 20, 21 или 22 нуклеотида, по существу идентичных нуклеиновой кислоте, кодирующей PD-1, PD-L1 или PD-L2. В дополнительных примерах двухцепочечная РНК содержит от приблизительно 19 до приблизительно 25 нуклеотидов, на 100% идентичных нуклеиновой кислоте, кодирующей PD-1, PD-L1 или PD-L2. Следует отметить, что в этом контексте «приблизительно» относится только к целому числу. В одном примере «приблизительно» 20 нуклеотидов относится к нуклеотиду от 19 до 21 нуклеотида в длину.
В отношении выступа на двухцепочечной РНК длина выступа является независимой у двух цепей, так что длина одного выступа не зависит от длины выступа на другой цепи. В конкретных примерах длина 3'- или 5'-выступа составляет 0 нуклеотидов по меньшей мере на одной цепи и в некоторых случаях 0 нуклеотидов на обеих цепях (следовательно, дцРНК с тупыми концами). В других примерах длина 3'- или 5'-выступа составляет от 1 нуклеотида до 5 нуклеотидов на по меньшей мере одной цепи. Более конкретно, в некоторых примерах длина 3'- или 5'-выступа составляет 2 нуклеотида на по меньшей мере одной цепи или 2 нуклеотида на обеих цепях. В конкретных примерах молекула дцРНК имеет 3'-выступы из 2 нуклеотидов на обеих цепях.
Таким образом, в одном варианте осуществления предлагаемой РНК двухцепочечная РНК содержит 20, 21 или 22 нуклеотида и длина 3'-выступа составляет 2 нуклеотида на обеих цепях. В вариантах осуществления, предлагаемых в настоящем описании РНК, двухцепочечная РНК содержит приблизительно 40-60% аденина+урацила (AU) и приблизительно 60-40% гуанина+цитозина (GC). Более конкретно, в характерных примерах двухцепочечная РНК содержит приблизительно 50% AU и приблизительно 50% GC.
В настоящем документе описываются также РНК, которые дополнительно включают по меньшей мере один модифицированный рибонуклеотид, например, в смысловой цепи двухцепочечной РНК. В конкретных примерах модифицированный рибонуклеотид находится в 3'-выступе по меньшей мере одной цепи или более конкретно в 3'-выступе смысловой цепи. Конкретно рассматривается, что примеры модифицированных рибонуклеотидов включают рибонуклеотиды, которые включают определяемую метку (например, флуорохром, такой как родамин или ФИТЦ), аналог тиофосфата нуклеотида, дезоксирибонуклеотид (рассматриваемый как модифицированный, так как базовой молекулой является рибонуклеиновая кислота), 2'-фторурацил, 2'-аминоурацил, 2'-аминоцитидин, 4-тиоурацил, 5-бромурацил, 5-йодурацил, 5-(3-аминоаллил)урацил, инозин или 2'O-Me-нуклеотидный аналог.
Антисмысловые молекулы и рибозимы для PD-1, PD-L1 и PD-L2 также используются в раскрытом в настоящем описании методе. Антисмысловые нуклеиновые кислоты представляют собой молекулы ДНК или РНК, которые комплементарны, по меньшей мере, части специфической молекулы мРНК (Weintraub, Scientific American 262:40, 1990). В клетке антисмысловые нуклеиновые кислоты гибридизуются с соответствующей мРНК, образуя двухцепочечную молекулу. Антисмысловые нуклеиновые кислоты мешают трансляции мРНК, так как клетка не будет транслировать мРНК, которая является двухцепочечной. Предпочтительны антисмысловые олигомеры из приблизительно 15 нуклеотидов, так как они легко синтезируются и с меньшей вероятностью вызывают проблемы, чем более крупные молекулы, при введении в клетку-мишень, продуцирующую PD-1, PD-L1 или PD-L2. Использование методов с антисмысловыми последовательностями для ингибирования трансляции генов in vitro хорошо известно в данной области техники (см., например, Marcus-Sakura, Anal. Biochem. 172:289, 1988).
Антисмысловой олигонуклеотид может состоять, например, из 5, 10, 15, 20, 25, 30, 35, 40, 45 или 50 нуклеотидов в длину. Антисмысловая нуклеиновая кислота может быть создана с использованием химического синтеза и реакций ферментативного лигирования с применением процедур, известных в данной области техники. Например, молекула антисмысловой нуклеиновой кислоты может быть химически синтезирована с использованием природных нуклеотидов или различным образом модифицированных нуклеотидов, созданных для увеличения биологической стабильности молекул или для повышения физической стабильности дуплекса, образованного между антисмысловой и смысловой нуклеиновыми кислотами, например могут быть использованы фосфоротиоатные производные, и замещенные акридином нуклеотиды. Примеры модифицированных нуклеотидов, которые могут быть использованы для создания антисмысловой нуклеиновой кислоты, включают среди прочего 5-фторурацил, 5-бромурацил, 5-хлорурацил, 5-йодурацил, гипоксантин, ксантин, 4-ацетилцитозин, 5-(карбоксигидроксилметил)урацил, 5-карбоксиметиламинометил-2-тиоуридин, 5-карбоксиметиламинометилурацил, дигидроурацил, бета-D-галактозилквеозин, инозин.
Использование олигонуклеотида для глушения транскрипции известно как триплексная стратегия, так как олигонуклеотид наматывается вокруг двуспиральной ДНК, образуя трехцепочечную спираль. Следовательно, эти триплексные соединения могут создаваться для узнавания уникального сайта на выбранном гене (Maher, et al., Antisense Res. and Dev. 1(3):227, 1991; Helene, C, Anticancer Drug Design 6(6):569, 1991). Этот тип ингибиторного олигонуклеотида используется также в раскрытых в настоящем описании методах.
Используются также рибозимы, которые представляют собой молекулы РНК, обладающие способностью специфически расщеплять другие одноцепочечные РНК способом, аналогичным рестрикции ДНК эндонуклеазами. Путем модификации нуклеотидных последовательностей, которые кодируют эти РНК, можно создать молекулы, которые узнают специфические нуклеотидные последовательности в молекуле РНК, и расщепить их (Cech, J. Amer. Med. Assn. 260:3030, 1988). Главным преимуществом этого подхода является то, что, так как они являются специфичными в отношении последовательности, инактивируются только мРНК с конкретными последовательностями.
Существует два основных типа рибозимов, а именно рибозимы типа тетрагимена (Hasselhoff, Nature 334:585, 1988) и типа «головки молота». Рибозимы типа тетрагимена узнают последовательности, которые составляют четыре основания в длину, в то время как рибозимы типа «головки молота» узнают последовательности оснований из 11-18 оснований в длину. Чем длиннее узнаваемая последовательность, тем больше вероятность того, что последовательность будет встречаться исключительно в типе мРНК-мишени. Следовательно, рибозимы типа головки молота предпочтительнее рибозимов типа тетрагимена для инактивации специфического типа мРНК, и узнаваемые последовательности из 18 оснований предпочтительнее более коротких узнаваемых последовательностей.
Различные системы доставки известны и могут использоваться для введения миРНК и других молекул ингибиторных нуклеиновых кислот в качестве терапевтических агентов. Такие системы включают, например, инкапсулирование в липосомы, микрочастицы, микрокапсулы, наночастицы, рекомбинантные клетки, способные экспрессировать терапевтическую(ие) молекулу(лы) (см., например, Wu et al, J. Biol. Chem. 262, 4429, 1987), конструкцию терапевтической нуклеиновой кислоты в виде части ретровирусного или другого вектора и тому подобное.
C. Низкомолекулярные ингибиторы
Антагонисты PD-1 включают молекулы, которые идентифицированы из больших библиотек как природного продукта, так и синтетических (или полусинтетических) экстрактов или химических библиотек, в соответствии с методами, известными в данной области техники. Методы скрининга, которые определяют снижение активности PD-1 (такие как определение гибели клеток), пригодны для идентификации соединений из различных источников на активность. Первоначальный скрининг может быть осуществлен с использованием смешанной библиотеки соединений, других разнообразных соединений и библиотек соединений. Таким образом, могут быть идентифицированы молекулы, которые связывают PD-1, PD-L1 или PD-L2, молекулы, которые ингибируют экспрессию PD-1, PD-L1 и/или PD-L2, и молекулы, которые ингибируют активность PD-1, PD-L1 и/или PD-L2. Эти низкомолекулярные соединения могут быть идентифицированы из комбинаторных библиотек, библиотек природных продуктов или других библиотек низкомолекулярных соединений. Кроме того, антагонист PD-1 может быть идентифицирован из соединений из коммерческих источников, а также из коммерчески доступных аналогов идентифицированных ингибиторов.
Точный источник тестируемых экстрактов или соединений не является критичным для идентификации антагонистов PD-1. Соответственно, антагонисты PD-1 могут быть идентифицированы фактически из любого числа химических экстрактов или соединений. Примеры таких экстрактов или соединений, которые могут быть антагонистами PD-1, включают, но не ограничиваются этим, экстракты на основе растений, грибов, прокариот или животных, бульоны ферментации и синтетические соединения, а также модификации существующих соединений. Доступны также многочисленные методы осуществления произвольного или прямого синтеза (например, полусинтеза или тотального синтеза) любого числа химических соединений, включая, но не ограничиваясь этим, соединения на основе сахаридов, липидов, пептидов и нуклеиновых кислот. Библиотеки синтетических соединений коммерчески доступны от Brandon Associates (Merrimack, N.H.) и Aldrich Chemical (Milwaukee, Wis.). Антагонисты PD-1 могут быть идентифицированы из библиотек синтетических соединений, которые коммерчески доступны от ряда компаний, включая Maybridge Chemical Co. (Trevillet, Cornwall, UK), Comgenex (Princeton, N.J.), Brandon Associates (Merrimack, N.H.) и Microsource (New Milford, Conn.). Антагонисты PD-1 могут быть идентифицированы из библиотеки редких химических соединений, такой как библиотека, которая доступна от Aldrich (Milwaukee, Wis.). Антагонисты PD-1 могут быть идентифицированы из библиотек природных соединений в форме бактериальных, грибковых, растительных и животных экстрактов, коммерчески доступных от ряда источников, включая Biotics (Sussex, UK), Xenova (Slough, UK), Harbor Branch Oceangraphics Institute (Ft. Pierce, FIa.) и PharmaMar, U.S.A. (Cambridge, Mass.). Природные и полученные синтетически библиотеки и соединения легко модифицируются с помощью общепринятых химических, физических и биохимических способов.
Используемые соединения могут быть найдены в многочисленных химических классах, хотя обычно они являются органическими соединениями, включая низкомолекулярные органические соединения. Низкомолекулярные органические соединения, обладающие молекулярной массой более 50, хотя менее приблизительно 2500 дальтон, например менее приблизительно 750 или менее приблизительно 350 дальтон, могут быть использованы в раскрытых в настоящем описании методах. Примеры классов включают гетероциклы, пептиды, сахариды, стероиды и тому подобное. Соединения могут быть модифицированы для увеличения эффективности, стабильности, фармацевтической совместимости и тому подобное. В некоторых вариантах осуществления используемые соединения обладают Kd для PD-1, PD-L1 или PD-L2 менее 1 нМ, менее 10 нМ, менее 1 мкМ или менее 1 мМ.
D. Варианты пептида PD-1 в качестве антагонистов
В одном варианте осуществления варианты белка PD-1, которые функционируют в качестве антагониста, могут быть идентифицированы путем скрининга комбинаторных библиотек мутантов, таких как точечные мутанты или укороченные мутанты белка PD-1, для идентификации белков с антагонистической активностью. В одном примере антагонист представляет собой растворимый белок PD-1.
Таким образом, библиотека вариантов PD-1 может быть создана с помощью комбинаторного мутагенеза на уровне нуклеиновой кислоты и закодирована библиотекой генного разнообразия. Библиотека вариантов PD-1 может быть получена, например, с помощью ферментативного лигирования смеси синтетических олигонуклеотидов в генетические последовательности, такие как набор вырожденных потенциальных последовательностей PD-1, экспрессируемых в индивидуальные полипептиды, или, альтернативно, как набор более крупных гибридных белков (таких как для фагового дисплея), содержащий набор последовательностей PD-1.
Существуют разнообразные методы, которые могут быть использованы для получения библиотек потенциальных вариантов PD-1 из вырожденной олигонуклеотидной последовательности. Химический синтез вырожденной генной последовательности может быть осуществлен в автоматическом ДНК-синтезаторе, и синтетический ген затем лигирован в подходящий экспрессионный вектор. Использование набора вырожденных генов позволяет заготовить в одной смеси все последовательности, кодирующие желаемый набор потенциальных последовательностей антагонистов PD-1. Методы синтеза вырожденных олигонуклеотидов известны в данной области техники (см., например, Narang, et al., Tetrahedron 39:3, 1983; Itakura et al. Annu. Rev. Biochem. 53:323, 1984; Itakura et al. Science 198:1056, 1984).
Кроме того, библиотеки фрагментов кодирующей последовательности белка PD-1 могут быть использованы для создания совокупности фрагментов PD-1 для скрининга и последующей селекции вариантов антагониста PD-1. В одном варианте осуществления библиотека кодирующих последовательностей фрагментов может быть создана путем обработки двухцепочечных фрагментов ПЦР кодирующей последовательности PD-1 нуклеазой в условиях, когда одноцепочечный разрыв возникает только примерно один раз на молекулу, денатурации двухцепочечной ДНК, ренатурации ДНК с образованием двухцепочечной ДНК, которая может включать смысловую/антисмысловую пары из различных продуктов, возникающих после разрывов, удаления одноцепочечных частей из заново сформированных дуплексов путем обработки нуклеазой S1 и лигирования библиотеки полученных фрагментов в экспрессионный вектор. С помощью этого метода может быть получена экспрессионная библиотека, которая кодирует N-концевые, C-концевые и внутренние фрагменты PD-1 различных размеров.
В данной области техники известно несколько методов скрининга генных продуктов комбинаторных библиотек, созданных с помощью точечных мутаций или усечения, и скрининга библиотек кДНК для генных продуктов, имеющих выбранное свойство. Такие способы адаптируемы для быстрого скрининга генных библиотек, созданных с помощью комбинаторного мутагенеза белков PD-1. Наиболее широко используемые методы, которые подходят для анализа с высокой пропускной способностью при скрининге генных библиотек, обычно включают клонирование генной библиотеки в реплицируемых экспрессионных векторах, трансформацию подходящих клеток полученной библиотекой векторов и экспрессию комбинаторных генов в условиях, при которых определение желаемой активности облегчает выделение вектора, кодирующего тот ген, чей продукт был определен. Для идентификации антагонистов PD-1 может быть использован возвратный групповой мутагенез (REM) в сочетании с методами скрининга (Arkin and Youvan, Proc. Natl. Acad. Sci. USA 89:7811-7815, 1992; Delagrave et al., Protein Eng. 6(3):327-331, 1993).
В одном варианте осуществления для анализа библиотеки вариантов PD-1 могут быть использованы тесты на основе клеточных технологий. Например, библиотека экспрессионных векторов может быть трансфицирована в клеточную линию, которая обычно синтезирует и секретирует PD-1. Трансфицированные клетки затем культивируют так, что секретируется PD-1 или конкретный вариант PD-1. Влияние экспрессии мутанта на активность PD-1 в супернатантах клеток может быть определено, например, с помощью любого функционального теста. Плазмидная ДНК может быть затем извлечена из клеток, в которых активность PD-1 заингибирована, и индивидуальные клоны дополнительно охарактеризованы.
В качестве антагонистов PD-1 могут быть также использованы пептидомиметики. Пептидные аналоги обычно используются в фармацевтической промышленности в качестве непептидных лекарств со свойствами, аналогичными таковым матричного пептида. Эти типы непептидных соединений обычно разрабатываются с помощью компьютерного молекулярного моделирования. Пептидные миметики, которые структурно сходны с терапевтически используемыми пептидами, могут быть использованы для получения эквивалентного терапевтического или профилактического эффекта. Обычно пептидомиметики структурно сходны с полипептидом-образцом (например, с полипептидом, который обладает биологической активностью PD-1), но обладает одной или более пептидными связями, необязательно замещенными связями -CH2NH-, -CH2S-, -CH2-CH2-, -CH.=.CH- (цис и транс), -COCH2-, -CH(OH)CH2 и -CH2SO-. Эти пептидные связи могут замещаться с помощью методов, известных в данной области техники (см., например, Morley, Trends Pharm. Sci. pp. 463-468, 1980; Hudson et al. Int. J. Pept. Prot. Res. 14:177-185, 1979; Spatola, Life Sci. 38:1243-1249, 1986; Holladay, et al. Tetrahedron Lett. 24:4401-4404, 1983). Пептидные миметики могут быть получены экономично, быть стабильными и могут иметь повышенный период полужизни или всасывания. Промечивание пептидомиметиков обычно включает ковалентное присоединение одной или более меток, прямо или через спейсер (такой как амидная группа) к невзаимодействующему(им) положению(ям) на пептидомиметике, которое предсказано данными по количественной оценке структуры-активности и/или молекулярным моделированием. Такие невзаимодействующие положения обычно представляют собой положения, которые не образуют прямые контакты с макромолекулой(ами), с которой(ыми) связывается пептидомиметик для индукции терапевтического эффекта. Дериватизация пептидомиметиков не должна существенно вмешиваться в желаемую биологическую или фармацевтическую активность пептидомиметика.
В раскрытых в настоящем описании методах может также использоваться доминантно негативный белок или нуклеиновая кислота, кодирующая доминантно негативный белок, который вмешивается в биологическую активность PD-1 (т.е. в связывание PD-1 с PD-L1, PD-L2 или обоими). Доминантно негативный белок представляет собой любую аминокислотную молекулу, обладающую последовательностью, которая по меньшей мере на 50%, 70%, 80%, 90%, 95% или даже 99% идентична последовательности по меньшей мере из 10, 20, 35, 50, 100 или более 150 аминокислот белка дикого типа, которому соответствует доминантно негативный белок. Например, доминантно негативный PD-L1 имеет такую мутацию, что он связывается с PD-1 более прочно, чем нативный PD-1 (дикого типа), но не активирует какой-либо путь клеточной сигнализации через PD-1.
Доминантно негативный белок может быть введен в экспрессионный вектор. Экспрессионный вектор может представлять собой невирусный вектор или вирусный вектор (например, ретровирусный, рекомбинантный связанный с аденовирусом вирусный или рекомбинантный аденовирусный вектор). Альтернативно доминантно негативный белок может быть прямо введен в виде рекомбинантного белка системно или в инфицированную область, например, с помощью методов микроинъекций.
Полипептидные антагонисты могут продуцироваться в прокариотных или эукариотных клетках-хозяевах в результате экспрессии полинуклеотидов, кодирующих аминокислотную последовательность, часто в виде части более крупного полипептида (гибридного белка, например, с ras или с ферментом). Альтернативно такие пептиды могут быть синтезированы химическими методами. Методы экспрессии гетерологичных белков в рекомбинантных хозяевах, методы химического синтеза полипептидов и трансляции in vitro хорошо известны в данной области техники (см. Maniatis et al. Molecular Cloning: A Laboratory Manual (1989), 2nd Ed., Cold Spring Harbor, N.Y.; Berger and Kimmel, Methods in Enzymology, Volume 152, Guide to Molecular Cloning Techniques (1987), Academic Press, Inc., San Diego, Calif.; Kaiser et al., Science 243:187, 1989; Merrifield, Science 232:342, 1986; Kent, Annu. Rev. Biochem. 57:957, 1988).
Пептиды могут быть получены, например, путем прямого химического синтеза и использованы в качестве антагонистов взаимодействия PD-1 с лигандом. Пептиды могут быть получены в виде модифицированных пептидов с непептидными частями, присоединенными с помощью ковалентной связи к N-концу и/или C-концу. В определенных предпочтительных вариантах осуществления, либо карбоксиконец, либо аминоконец, либо оба химически модифицированы. Наиболее обычные модификации концевых амино- и карбоксильных групп представляют собой ацетилирование и амидирование соответственно. В различные варианты осуществления могут быть введены аминоконцевые модификации, такие как ацилирование (например, ацетилирование) или алкилирование (например, метилирование), и карбоксиконцевые модификации, такие как амидирование, также как и другие модификации концевых частей, включая циклизацию. Определенные аминоконцевые и/или карбоксиконцевые модификации и/или удлинения пептида по отношению к последовательности остова могут придать выгодные физические, химические, биохимические и фармакологические свойства, такие как повышенная стабильность, усиленная действенность и/или эффективность, устойчивость к протеазам сыворотки, желаемые фармакокинетические свойства и другие.
Методы лечения: введение антагониста PD-1 индивидууму
В настоящем описании предлагаются методы лечения разнообразных инфекций и типов рака. В этих методах лечат или предотвращают инфекцию или рак, или облегчают их симптом путем введения индивидууму терапевтически эффективного количества антагониста PD-1. Индивидуумом может быть любое млекопитающее, такое как человек, примат, мышь, крыса, собака, кошка, корова, лошадь и свинья. В некоторых примерах индивидуум представляет собой примата, такого как человек. В дополнительных примерах индивидуумом являются мышевидные, такие как мышь.
В некоторых вариантах осуществления у индивидуума есть риск развития инфекции. Индивидуум с риском развития инфекции представляет собой индивидуума, у которого еще нет инфекции, но он может быть инфицирован интересующим инфекционным агентом. В дополнительных примерах у индивидуума есть инфекция, такая как устойчивая инфекция. Индивидуум с устойчивой инфекцией может быть идентифицирован стандартными методами, подходящими для специалиста в данной области техники, такого как врач.
В некоторых примерах индивидуум имеет устойчивую инфекцию при инфицировании бактериями, вирусом, грибком или паразитом. Обычно устойчивые инфекции в отличие от острых инфекций эффективно не исчезают при индукции иммунного ответа хозяина. Инфекционный агент и иммунный ответ достигают равновесия так, что инфицированный индивидуум остается с инфекцией в течение длительного периода времени без обязательного проявления симптомов. Устойчивые инфекции включают, например, латентную, хроническую и медленную инфекции. Устойчивая инфекция возникает при инфицировании такими вирусами, как вирусы Т-клеточного лейкоза, вирус Эпштейна-Барр, цитомегаловирус, вирусы герпеса, вирус ветряной оспы, вирус кори, паповавирусы, прионы, вирусы гепатита, аденовирусы, парвовирусы и вирусы папилломы.
При хронической инфекции инфекционный агент может быть определен в организме во все периоды времени. Однако признаки и симптомы заболевания могут присутствовать или отсутствовать в течение продолжительного периода времени. Примеры хронических инфекций включают гепатит B (вызываемый вирусом гепатита B (HBV)) и гепатит C (вызываемый вирусом гепатита C (HCV)), инфекции, вызываемые аденовирусом, цитомегаловирусом, вирусом Эпштейна-Барр, вирусом простого герпеса 1, вирусом простого герпеса 2, вирусом герпеса 6 человека, вирусом ветряной оспы, вирусом гепатита B, вирусом гепатита D, вирусом папилломы, парвовирусом B19, вирусом полиомы BK, вирусом полиомы JC, вирусом кори, вирусом краснухи, вирусом иммунодефицита человека (ВИЧ), вирусом I Т-клеточного лейкоза человека и вирусом II Т-клеточного лейкоза человека. Паразитарные устойчивые инфекции могут возникать в результате инфицирования Leishmania, Toxoplasma, Trypanosoma, Plasmodium, Schistosoma и Encephalitozoon.
При латентной инфекции инфекционный агент (такой как вирус) является, по-видимому, неактивным и дремлющим, так что у индивидуума не всегда проявляются признаки или симптомы. При латентной вирусной инфекции вирус остается в равновесии с хозяином в течение продолжительных периодов времени перед повторным появлением симптомов; однако действующие вирусы не могут быть определены до наступления реактивации заболевания. Примеры латентных инфекций включают инфекции, вызываемые вирусом простого герпеса (HSV)-1 (герпетические лихорадки), HSV-2 (генитальный герпес) и вирусом VZV ветряной оспы-опоясывающего лишая (ветрянки/опоясывающий лишая).
При медленных инфекциях инфекционные агенты градуально растут в количестве в течение очень длительного периода времени, в течение которого не наблюдается существенных признаков или симптомов заболевания. Примеры медленных инфекций включают СПИД (вызываемый ВИЧ-1 и ВИЧ-2), опухоли у животных, вызываемые лентивирусами и прионами.
Кроме того, устойчивые инфекции часто возникают как поздние осложнения острых инфекций. Например, подострый склерозирующий панэнцефалит (SSPE) может возникать после острой коревой инфекции, или регрессивный энцефалит может возникать в результате инфекционного заболевания краснухой.
В одном неограничивающем примере у индивидуума может быть диагностирована устойчивая инфекция хламидиями после определения вида хламидии в биологическом образце от этого индивидуума с использованием анализа ПЦР. Млекопитающие, у которых не диагностирована устойчивая инфекция, не нуждаются в лечении в соответствии с этим раскрытием. Микробные агенты, способные вызвать устойчивую инфекцию, включают вирусы (такие как вирус папилломы, вирус гепатита, вирус иммунодефицита человека и вирус герпеса), бактерии (такие как Escherichia coli и Chlamydia spp.), паразиты (такие как Leishmania spp., Schistosoma spp., Trypanosoma spp., Toxoplasma spp.) и грибы.
В дополнение к соединению, которое снижает экспрессию или активность PD-1, индивидууму, подвергаемому лечению, может также вводиться вакцина. В одном примере вакцина может включать адъювант. В другом примере вакцина может включать примирующую бустерную иммунизацию. Вакцина может представлять собой убитую нагреванием вакцину, ослабленную вакцину или субъединичную вакцину. Индивидуума, уже инфицированного патогеном, можно лечить терапевтической вакциной, такой как с антагонистом PD-1 и антигеном. Индивидуум может иметь асимптоматическое заболевание, так что лечение предотвращает развитие симптома. Терапевтическая вакцина может также снижать тяжесть одного или более существующих симптомов или снижать нагрузку патогеном.
В некоторых примерах методов лечения индивидууму вводят терапевтически эффективное количество антагониста PD-1 в сочетании с вирусным антигеном. Неограничивающие примеры подходящих вирусных агентов включают среди прочего: антигены HA, NA, M, NP и NS вируса гриппа; p24, pol, gp41 и gpl20 ВИЧ; белки F и G метапневмовируса (hMNV); белки El, E2 и белки каркаса вируса гепатита С (HCV); El, E2 и белки каркаса вируса лихорадки Денге (DEN 1-4); белок Ll вируса папилломы человека; пептид gp220/350 и EBNA-3A вируса Эпштейна-Барр; гликопротеин gB, гликопротеин gH, pp65, IEl (экзон 4) и ppl50 цитомегаловируса (CMV); эпитопы пептида IE62 и гликопротеина E вируса ветряной оспы (VZV); эпитопы гликопротеина D вируса простого герпеса. Антигенные полипептиды могут соответствовать полипептидам, существующим в природных вирусных изолятах животных или человека, или могут быть созданы для включения одной или более аминокислотных замен по сравнению с природным (патогенным или непатогенным) изолятом. Примеры антигенов перечислены ниже:
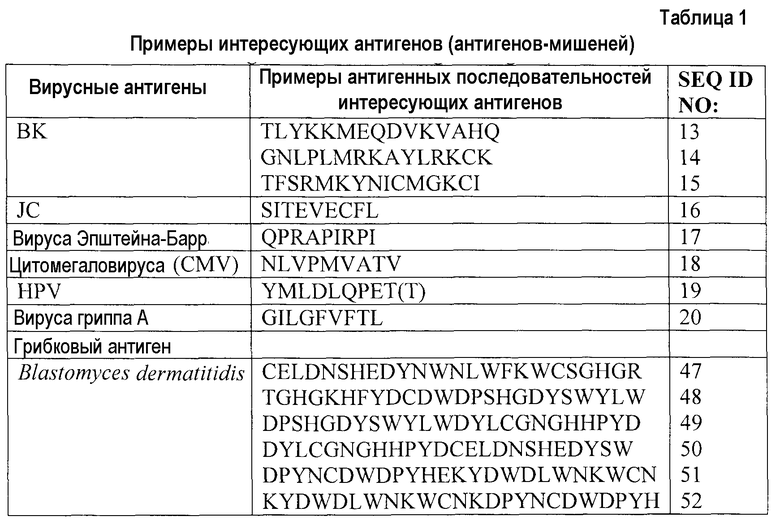
В дополнительных вариантах осуществления у индивидуума есть опухоль. Метод включает введение индивидууму терапевтически эффективного количества антагониста PD-1, тем самым проводя лечение опухоли. В некоторых примерах терапевтически эффективное количество опухолевого антигена или нуклеотида, кодирующего опухолевый антиген, также вводится индивидууму. Антагонист PD-1 и опухолевый антиген или нуклеотид, кодирующий опухолевый антиген, могут вводиться одновременно или последовательно.
Введение антагониста PD-1 ведет к снижению размера, распространения или метастазирующего потенциала опухоли у индивидуума. Эффективность определяется в связи с любым известным методом диагностики или лечения конкретной опухоли.
Опухоли (также называемые «раком») включают солидные опухоли и лейкозы. Примеры опухолей включают опухоли, перечисленные в таблице 2 (вместе с известными опухолевыми антигенами, связанными с этими типами рака).
Примеры опухолей и их опухолевых антигенов
Примеры интересующих опухолевых антигенов включают перечисленные ниже в таблице 3:

Конкретными неограничивающими примерами являются ангиоиммунобластная лимфома или нодулярная лимфома Ходжкина с преобладанием лимфоцитов. Ангиоиммунобластная лимфома (AIL) представляет собой агрессивный (быстро прогрессирующий) тип Т-клеточной неходжкинской лимфомы, характеризующийся увеличенными лимфатическими узлами и гипергаммаглобулинемией (повышенными антителами в крови). Другие симптомы могут включать кожную сыпь, лихорадку, потерю веса, позитивный тест Кумба или ночную потливость. Этот рак обычно возникает у взрослых. Больные обычно имеют возраст 40-90 лет (медиана около 65) и чаще являются мужчинами. При прогрессии AIL могут развиваться гепатоспленомегалия, гемолитическая анемия и поликлональная гипергаммаглобулинемия. Кожа вовлечена приблизительно у 40-50% больных.
Нодулярная лимфома Ходжкина с преобладанием лимфоцитов представляет собой В-клеточную неоплазму, которая, очевидно, происходит из герминативного центра В-клеток с мутированными нефункционирующими генами иммуноглобулинов. Сходно с ангиоиммунобластной лимфомой неопластические клетки связаны с сетью фолликулярных дендритных клеток. Экспрессия PD-1 наблюдается в Т-клетках, тесно связанных с неопластической CD20+ клеткой в нодулярной лимфоме Ходжкина с преобладанием лимфоцитов, с паттерном, сходным с наблюдаемым в CD57+ Т-клетках. CD57 идентифицирован как другой маркер Т-клеток, связанных с герминативным центром, как и CXCR5, данные, которые поддерживают заключение о том, что неопластические клетки в нодулярной лимфоме Ходжкина с преобладанием лимфоцитов имеют тесную связь с Т-клетками, связанными с герминативным центром.
Экспрессия интересующего опухолевого антигена может быть определена на уровне белка или нуклеиновой кислоты с использованием любого метода, известного в данной области техники. Например, для определения экспрессии гена может быть использован анализ нозерн-гибридизацией с применением зондов, которые специфически узнают одну или более из этих последовательностей. Альтернативно экспрессию измеряют с использованием тестов на основе ПЦР с обратной транскрипцией, например, используя праймеры, специфичные для дифференциально экспрессируемой последовательности генов. Экспрессия определяется также на уровне белка, например, с помощью измерения уровней пептидов, кодируемых описанными в настоящем документе генными продуктами, или измерения их активности. Такие методы хорошо известны в данной области техники и включают, например, иммунотесты на основе антител к белкам, кодируемым этими генами. Любой биологический материал может быть использован для определения/количественной оценки белка или активности.
В одном примере у индивидуума ранее был диагностирован рак. В дополнительных примерах индивидуум подвергался предшествующему лечению в связи с раком. Однако в некоторых примерах у индивидуума ранее не был диагностирован рак. Диагноз солидной опухоли может быть сделан посредством идентификации исследуемой массы, хотя он может быть сделан с помощью других средств, таких как радиологическая диагностика или ультразвук. Лечение рака может включать хирургическую операцию или может включать использование химиотерапевтических агентов, таких как доцетаксель, винорелбин гемцитабин, капецитабин, или сочетаний циклофосфамида, метотрексата и фторурацила; циклофосфамида, доксорубицина и фторурацила; доксорубицина и циклофосфамида; доксорубицина и циклофосфамида с паклитакселем; доксорубицина и затем CMF (циклофосфамид, эпирубицин и фторурацил). Кроме того, лечение может включать использование облучения.
В некоторых примерах индивидууму вводят терапевтически эффективное количество антагониста PD-1. Индивидууму также вводят терапевтически эффективное количество опухолевого антигена или нуклеиновой кислоты, кодирующей антиген. Введение может быть одновременным или последовательным.
Для лечения индивидуума с устойчивой инфекцией или раком заинтересованному индивидууму вводят терапевтически эффективное количество антагониста PD-1. В одном примере терапевтически эффективное количество антагониста PD-1 представляет собой биологически активную дозу, такую как доза, которая должна индуцировать повышение CD8+ T-клеточной цитотоксической активности с повышением специфичного иммунного ответа против инфекционного агента. Желательно, чтобы антагонист PD-1 обладал способностью снижать экспрессию или активность PD-1 в специфичных в отношении антигена иммунных клетках (например, T-клетках, таких как CD8+ T-клетки) по меньшей мере на 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или более 100% ниже контрольного уровня без лечения. Уровень активности PD-1 в иммунных клетках измеряют любым методом, известным в данной области техники, включая, например, анализ иммуноблотингом, иммуногистохимию, ELISA и анализ Нозерн-блотингом. Альтернативно биологическую активность PD-1 измеряют путем оценки связывания PD-1 с PD-L1, PD-L2 или обоими. Биологическую активность PD-1 определяют в соответствии с его способностью повышать CD8+ T-клеточную цитотоксичность, включая, например, продукцию цитокинов, клиренс инфекционного агента и пролиферацию специфичных в отношении антигена CD8+ T-клеток. Предпочтительно, чтобы агент, который снижает экспрессию или активность PD-1, мог повышать иммунный ответ, специфичный в отношении инфекционного агента или опухоли по меньшей мере на 5%, 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или более 100% выше контрольного уровня без лечения. Агент настоящего изобретения, таким образом, представляет собой любой агент, обладающий одной или более из этих форм активности. Хотя агент предпочтительно экспрессируется в CD8+ T-клетках, следует понимать, что любая клетка, которая может оказывать влияние на иммунный ответ против устойчивых инфекций, также включается в методы изобретения и включает, например, B-клетки.
Необязательно индивидууму вводят один или более дополнительных терапевтических агентов. Дополнительные терапевтические агенты включают, например, антивирусные соединения (например, видарабин, ацикловир, ганцикловир, валганцикловир, нуклеозидный аналог - ингибитор обратной транскриптазы (NRTI) ((например, AZT (зидовудин), ddI (диданозин), ddC (зальцитабин), d4T (ставудин) или 3TC (ламивудин)), ненуклеозидные ингибиторы обратной транскриптазы (NNRTI) (например, невирапин или делавирдин), протеазный ингибитор (саквинавир, ритонавир, индинавир или нельфинавир), рибавирин или интерферон), антибактериальные соединения, противогрибковые соединения, противопаразитные соединения, противовоспалительные соединения, антибластомное средство (химиотерапевтические агенты) или анальгетики.
Дополнительный терапевтический агент вводят до, одновременно или после введения антагониста PD-1. Например, антагонист PD-1 и дополнительный агент вводят в раздельных составах, разделенных друг от друга во времени по меньшей мере 1, 2, 4, 6, 10, 12, 18 или более 24 часами. Необязательно дополнительный агент составлен вместе с антагонистом PD-1. Когда дополнительный агент находится в другой композиции, можно использовать разные пути введения. Агент вводят в дозах, о которых известно, что они эффективны для лечения, подавления или профилактики инфекции таким агентом.
Концентрации антагониста PD-1 и дополнительного агента зависят от ряда факторов, включая средства введения, локализацию объекта лечения, физиологическое состояние млекопитающего и другие вводимые лекарства. Таким образом, терапевтические дозы можно титровать для оптимизации безопасности и эффективности и находятся в пределах компетенции специалиста в данной области техники. Определение подходящей дозы и режима введения в конкретной ситуации находится в пределах компетенции специалиста в данной области техники.
Необязательно индивидууму дополнительно вводят вакцину, которая вызывает иммунный ответ против вируса иммунодефицита человека (ВИЧ), туберкулеза, гриппа или гепатита C. Типичные вакцины описаны, например, в Berzofsky et al. (J. Clin. Invest. 114:456-462, 2004). Если это желательно, вакцину вводят с примирующей бустерной инъекцией или с адъювантами. Вакциной также может быть опухолевая вакцина, такая как терапевтически эффективное количество опухолевого антигена. В некоторых вариантах осуществления индивидууму вводят терапевтически эффективное количество антигенного полипептида, такого как вирусный или опухолевый антиген.
Индивидууму можно вводить терапевтически эффективное количество опухолевого антигена или нуклеиновой кислоты, кодирующей опухолевый антиген. Полинуклеотиды включают рекомбинантную ДНК, которая включена в вектор автономно реплицирующейся плазмиды или вируса или в геномную ДНК прокариота или эукариота или находится в виде отдельной молекулы (такой как кДНК), независимой от других последовательностей. Нуклеотиды могут представлять собой рибонуклеотиды, дезоксирибонуклеотиды или модифицированные формы любого нуклеотида. Термин включает одноцепочечные и двухцепочечные формы ДНК.
Был сконструирован ряд вирусных векторов, включая вирус полиомы, т.е. SV40 (Madzak et al., 1992, J. Gen. Virol, 73:1533-1536), аденовирус (Berkner, 1992, Cur. Top. Microbiol. Immunol, 158:39-6; Berliner et al., 1988, Bio Techniques, 6:616-629; Gorziglia et al., 1992, J. Virol, 66:4407-4412; Quantin et al., 1992, Proc. Nad. Acad. Sci. USA, 89:2581-2584; Rosenfeld et al., 1992, Cell, 68:143-155; Wilkinson et al., 1992, Nucl. Acids Res., 20:2233-2239; Stratford-Perricaudet et al., 1990, Hum. Gene Ther., 1:241-256), вирус оспы (Mackett et al., 1992, Biotechnology, 24:495-499), аденоассоциированный вирус (Muzyczka, 1992, Curr. Top. Microbiol. Immunol, 158:91-123; On et al., 1990, Gene, 89:279-282), вирусы герпеса, включая HSV и EBV (Margolskee, 1992, Curr. Top. Microbiol. Immunol., 158:67-90; Johnson et al., 1992, J. Virol, 66:2952-2965; Fink et al., 1992, Hum. Gene Ther. 3:11-19; Breakfield et al., 1987, Mol. Neurobiol, 1:337-371; Fresse et al., 1990, Biochem. Pharmacol, 40:2189-2199), вирусы синдбис (H. Herweijer et al., 1995, Human Gene Therapy 6:1161-1167; патенты США №№ 5091309 и 52217879), альфавирусы (S. Schlesinger, 1993, Trends Biotechnol. 11:18-22; I. Frolov et al., 1996, Proc. Natl. Acad. Sci. USA 93:11371-11377) и ретровирусы птиц (Brandyopadhyay et al., 1984, Mol. Cell Biol, 4:749-754; Petropouplos et al., 1992, J. Virol., 66:3391-3397), мыши (Miller, 1992, Curr. Top. Microbiol. Immunol., 158:1-24; Miller et al., 1985, Mol. Cell Biol., 5:431-437; Sorge et al., 1984, Mol. Cell Biol., 4:1730-1737; Mann et al., 1985, J. Virol., 54:401-407) и человека (Page et al., 1990, J. Virol., 64:5370-5276; Buchschalcher et al., 1992, J. Virol., 66:2731-2739). В данной области техники известны также бакуловирусные (многоядерный полиэдроидный вирус Autographa californica; AcMNPV) векторы, и они имеются в продаже (например, от PharMingen, San Diego, Calif.; Protein Sciences Corp., Meriden, Conn.; Stratagene, La Jolla, Calif.).
В одном варианте осуществления в вирусный вектор включен полинуклеотид, кодирующий опухолевый антиген или вирусный антиген. Подходящие векторы включают ретровирусные векторы, ортопоксвирусные векторы, авипоксные векторы, фаулпоксные векторы, каприпоксные векторы, суипоксные векторы, аденовирусные векторы, векторы вируса герпеса, альфавирусные векторы, бакуловирусные векторы, векторы вируса синдбис, векторы вируса оспы и полиовирусные векторы. Конкретными иллюстративными векторами являются поксвирусные векторы, такие как вирус оспы, фаулпоксный вирус и крайне ослабленный вирус оспы (MVA), аденовирус, бакуловирус и тому подобное.
Используемые поксвирусы включают ортопоксный, суипоксный, авипоксный и капрпоксный вирусы. Ортопокс включает оспу, эктромелию и раккунпокс. Одним из примеров используемого ортопокса является коровья оспа. Авипокс включает фаулпокс, оспу канареек и оспу свиней. Каприпокс включает оспу коз и оспу овец. В одном примере суипокс представляет собой оспу свиней. Примеры поксвирусных векторов для экспрессии описаны, например, в патенте США № 6165460, который включен в настоящее описание в качестве ссылки. Другие вирусные векторы, которые могут быть использованы, включают другие ДНК-вирусы, такие как вирус герпеса и аденовирусы, и РНК-вирусы, такие как ретровирусы и вирус полиомиелита.
В некоторых вариантах осуществления антагонисты PD-1 вводят в количестве, достаточном для повышения цитотоксичности T-клеток, таких как CD8+ T-клетки. Повышение T-клеточной цитотоксичности ведет к повышению иммунного ответа и подавлению устойчивой инфекции или к снижению признака или симптома опухоли. Повышенный иммунный ответ можно измерить, например, по повышению пролиферации иммунных клеток, таких как T- или B-клетки, повышению продукции цитокинов и усилению клиренса инфекционного агента или снижению тяжести опухоли. Таким образом, метод может вести к смягчению одного или более симптомов, связанных с устойчивой инфекцией или опухолью. Таким образом, введение антагониста PD-1 подавляет устойчивую инфекцию, ингибирует рост/размер опухоли или смягчает один или более симптомов, связанных с устойчивой инфекцией или опухолью по меньшей мере на 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или 100% по сравнению с индивидуумом без лечения.
Лечение является эффективным, если лечение ведет к клинически полезному результату, такому как снижение количества инфекционного агента или снижение тяжести опухоли у индивидуума. При профилактическом лечении «эффективное» означает, что лечение задерживает или предотвращает инфекцию из источника. Эффективность может быть определена с помощью любого известного метода диагностики или лечения конкретной инфекции или опухоли.
Так, методы включают введение индивидууму фармацевтической композиции, которая включает терапевтически эффективное количество антагониста PD-1. Эффективное количество терапевтического соединения, такого как антитело, может составлять, например, от приблизительно 0,1 мг/кг до приблизительно 150 мг/кг. Эффективные дозы варьируют, как ясно специалистам в данной области техники, в зависимости от пути введения, использования наполнителя и совместного введения с другими терапевтическими средствами, включая использование других противоинфекционных агентов или терапевтических агентов для лечения, профилактики или облегчения симптома конкретной инфекции или опухоли. Для больного человека, страдающего от (или имеющего риск развития) инфекции или опухоли, используют лечебный режим с использованием стандартных методов.
Антагонист PD-1 вводят такому индивидууму с помощью методов, известных в данной области техники. Можно использовать любой антагонист PD-1, такой как раскрытые в настоящем описании. Кроме того, можно использовать более одного антагониста PD-1. Антагонист PD-1 можно вводить местно или системно. Например, антагонист PD-1 вводят перорально, ректально, назально, местно парентерально, подкожно, внутрибрюшинно, внутримышечно или внутривенно. Антагонист PD-1 можно вводить профилактически или после выявления инфекции или опухоли. Антагонист PD-1 необязательно составляют в виде компонента смеси терапевтических лекарств для лечения инфекции. Примеры составов, пригодных для парентерального введения, включают водные растворы активного агента в изотоническом солевом растворе, 5% раствор глюкозы или другой стандартный фармацевтически приемлемый наполнитель. В качестве фармацевтических наполнителей для доставки терапевтических соединений используют также стандартные солюбилизирующие агенты, такие как PVP или циклодекстрины.
Описанные в настоящем описании терапевтические соединения составляют в композиции для других путей введения с использованием традиционных методов. Например, антагонист PD-1 составляют в капсулу или таблетку для перорального введения. Капсулы могут содержать любые стандартные фармацевтически приемлемые вещества, такие как желатин или целлюлоза. Таблетки могут быть составлены в соответствии с традиционными процедурами путем прессования смесей терапевтического соединения с твердым носителем и смазывающим агентом. Примеры твердых носителей включают крахмал и сахар бентонит. Антагонист PD-1 можно вводить в форме таблетки с твердой оболочкой или капсулы, содержащей связующий агент, такой как лактоза или манит, традиционный наполнитель и таблетирующий агент. Другие составы включают мазь, суппозиторий, пасту, спрей, пластырь, крем, гель, рассасывающийся тампон или пену. Такие составы получают с помощью методов, хорошо известных в данной области техники.
Кроме того, антагонисты PD-1 можно вводить путем имплантации (либо прямо в орган (например, кишечник или печень), либо подкожно) твердого или рассасывающегося матрикса, который медленно высвобождает соединение в соседние и окружающие ткани индивидуума. Например, для лечения желудочно-кишечной инфекции соединение можно вводить системно (например, внутривенно, ректально или перорально) или местно (например, прямо в ткань желудка). Альтернативно импрегнированный антагонистом PD-1 брикет или рассасывающийся тампон помещают в непосредственном контакте с тканью желудка. Антагонист PD-1 медленно высвобождается in vivo путем диффузии лекарства из тампона и разрушения полимерного матрикса. Другим примером служит лечение инфекции печени (т.е. гепатита) путем инфузии в сосудистое русло печени раствора, содержащего антагонист PD-1.
Когда терапевтическое соединение представляет собой нуклеиновую кислоту, кодирующую антагонист PD-1, нуклеиновую кислоту можно вводить in vivo для стимуляции экспрессии кодируемого белка путем конструирования содержащего ее в виде части подходящего экспрессионного вектора нуклеиновой кислоты и его введения таким образом, чтобы он становился внутриклеточным (как при использовании ретровирусного вектора, путем прямой инъекции, путем бомбардировки микрочастицами, путем покрытия липидами или рецепторами клеточной поверхности или трансфицирующими агентами или путем его введения в соединении с гомеобокс-подобным пептидом, о котором известно, что он поступает в ядро (см., например, Joliot, et al., Proc. Natl. Acad. Sci USA 88:1864-1868, 1991), и тому подобное. Альтернативно терапевтическую нуклеиновую кислоту вводят внутриклеточно и включают в ДНК клетки хозяина для экспрессии путем гомологичной рекомбинации или оставляют эписомальной.
Для местного введения ДНК можно использовать стандартные векторы генной терапии. Такие векторы включают вирусные векторы, включая векторы, берущие начало от вирусов гепатита с дефектом репликации (таких как HBV и HCV), ретровирусы (см. публикацию PCT № WO 89/07136; Rosenberg et al., N. Eng. J. Med. 323(9):570-578, 1990, аденовирус (см. Morsey et al., J. Cell. Biochem., Supp. 17E, 1993), аденоассоциированный вирус (Kotin et al., Proc. Natl. Acad. Sci. USA 87:2211-2215, 1990), вирусы простого герпеса с дефектом репликации (HSV; Lu et al., Abstract, page 66, Abstracts of the Meeting on Gene Therapy, Sept. 22-26, Cold Spring Harbor Laboratory, Cold Spring Harbor, New York, 1992) и любые модифицированные версии этих векторов. Можно использовать любую другую систему доставки, которая обеспечивает перенос нуклеиновых кислот в эукариотные клетки in vivo. Например, нуклеиновые кислоты могут быть упакованы в липосомы, такие как катионные липосомы (липофектин), опосредуемые рецепторами системы доставки, отличные от вирусных векторы на основе нуклеиновых кислот, оболочки эритроцитов или микросферы (такие как микрочастицы; см., например, патент США № 4789734; патент США № 4925673; патент США № 3625214). Можно вводить также голую ДНК.
Что касается ингибирующих нуклеиновых кислот, то терапевтически эффективное количество представляет собой количество, которое способно вызывать клинически желаемый результат, например снижение продукта гена PD-1 у подвергнутого лечению животного. Такое количество может быть определено специалистом в данной области техники. Доза для любого конкретного пациента зависит от многих факторов, включая размеры больного, площадь поверхности тела, возраст, конкретное соединение, подлежащее введению, пол, время и путь введения, общее состояние здоровья и одновременное введение других лекарств. Дозы могут варьировать, но предпочтительная доза для внутривенного введения ДНК составляет приблизительно от 106 до 1022 копий молекулы ДНК.
Обычно плазмиды вводят млекопитающему в количестве от приблизительно 1 нанограмма до приблизительно 5000 микрограмм ДНК. Желательно, чтобы композиция содержала от приблизительно 5 нанограмм до приблизительно 1000 микрограмм ДНК, от 10 нанограмм до 800 микрограмм ДНК, от 0,1 микрограмм до 500 микрограмм ДНК, от 1 микрограмма до 350 микрограмм ДНК, от 25 микрограмм до 250 микрограмм ДНК или от 100 микрограмм до 200 микрограмм ДНК. Альтернативно введение рекомбинантных аденовирусных векторов, кодирующих антагонист PD-1, млекопитающему может осуществляться при концентрации по меньшей мере 105, 106, 107, 108, 109, 1010 или 1011 бляшкообразующих единиц (БОЕ).
В некоторых вариантах осуществления для лечения неврологических инфекций антагонист PD-1 можно вводить внутривенно или интратекально (например, путем прямой инфузии в спинномозговую жидкость). Для местного введения импрегнированный соединением брикет или рассасывающийся тампон помещают в непосредственный контакт с тканью центральной нервной системы (ЦНС). Соединение или смесь соединений медленно высвобождается in vivo посредством диффузии лекарства из брикета и разрушения полимерного матрикса. Альтернативно соединение вводят инфузией в мозг или спинномозговую жидкость с помощью стандартных методов. Например, в просверленном отверстии черепа размещают полое кольцо буравчика с катетером для использования в качестве ввода. Доступ жидкости из резервуара, соединенного с катетером, обеспечивается иглой или зондом, вставленными через верхнюю часть полого кольца буравчика. Сборка катетера (описанная, например, в патенте США № 5954687) обеспечивает продвижение потока жидкости, пригодного для переноса жидкостей в определенное место или из него, на мозге, рядом с ним или внутри мозга для обеспечения введения лекарства на протяжении определенного периода времени.
В дополнительных вариантах осуществления при сердечных инфекциях антагонист PD-1 может доставляться, например, в ткань сердца (такую как миокард, перикард или эндокард) путем прямой внутрисердечной инъекции через грудную клетку или путем использования стандартных методов на основе чрескожного катетера под флуороскопическим контролем. Таким образом, антагонист PD-1 можно прямо инъецировать в ткань или можно инфузировать через стент или катетер, который вставлен в полость тела. Для введения соединения можно использовать множество коронарных катетеров или перфузионных катетеров. Альтернативно антагонистом PD-1 покрывают или пропитывают стент, который помещают в сосуд сердца.
Легочные инфекции можно лечить, например, путем введения антагониста PD-1 с помощью ингаляции. Соединения доставляют в форме аэрозольного спрея из контейнера под давлением или дозатора, который содержит подходящий вытеснитель, такой как газ, такой как диоксид углерода, или распылителя.
Специалист в данной области техники должен понимать, что лечащиеся пациенты могли подвергаться тем же тестам для диагностики устойчивой инфекции у индивидуума или могли быть выявлены без проверки в качестве индивидуума с высоким риском благодаря наличию одного или более факторов риска (таких как экспозиция с инфекционным агентом, экспозиция с инфицированным индивидуумом, генетическая предрасположенность или наличие патологического состояния, предрасполагающего к вторичным инфекциям). Снижение симптомов устойчивой инфекции или повреждения может также включать, не ограничиваясь этим, смягчение симптомов, снижение степени заболевания, стабилизацию (без ухудшения) состояния заболевания, отсрочку или замедление развития заболевания и смягчение или временное облегчение болезненного состояния. Лечение может производиться дома под тщательным наблюдением сотрудника службы здравоохранения или в медицинском стационаре.
Методы измерения иммунного ответа после лечения с использованием раскрытых в настоящем описании методов хорошо известны в данной области техники. Активность T-клеток можно оценивать, например, с помощью тестов, которые выявляют продукцию цитокинов, тестов измерения пролиферации T-клеток, тестов измерения клиренса микробного агента и тестов измерения цитотоксичности CD8+ T-клеток. Эти тесты описаны, например, в патенте США № 6808710 и публикациях патентных заявок США №№ 20040137577, 20030232323, 20030166531, 20030064380, 20030044768, 20030039653, 20020164600, 20020160000, 20020110836, 20020107363, и 20020106730, каждая из которых включена в настоящее описание в качестве ссылки.
Необязательно способность антагониста PD-1 повышать цитотоксичность CD8+ T-клеток оценивают с помощью тестов, в которых измеряют пролиферацию CD8+ T-клеток (например, по включению тимидина, с помощью анализа BrdU и окрашивания маркерами клеточного цикла (например, Ki67 и CFSE), описанных, например, Dong et al. (Nature 5:1365-1369, 1999). В одном примере клеточную пролиферацию отслеживают при культивировании очищенных T-клеток, экспрессирующих PD-1, с антагонистом PD-1 по первичному сигналу активации, как описано выше, и 3H-тимидину. Уровень пролиферации T-клеток определяют по включению тимидина.
Цитотоксичность CD8+ T-клеток может быть также оценена в тестах лизиса (таких как тесты выделения 51Cr или тесты, определения выделения перфорина или гранзима), тестах, выявляющих активацию каспаз, или в тестах, в которых измеряется клиренс микробного агента из инфицированного индивидуума. Например, количество вируса в биологическом образце от инфицированного индивидуума (например, сыворотке, селезенке, печени, легком или ткани, в отношении которой вирус является тропным) может быть измерено до и после лечения.
Может быть также измерена продукция цитокинов, таких как IFNγ, TNF-α и ИЛ-2. Например, очищенные T-клетки культивируют в присутствии антагониста белка PD-1 и сигнала первичной активации. Уровень разных цитокинов в супернатанте может быть определен с помощью сэндвичных твердофазных иммуноферментных анализов или других традиционных тестов, описанных, например, в Dong et al. (Nature 5:1365-1369, 1999).
Если это желательно, эффективность антагониста PD-1 оценивают по его способности индуцировать костимуляцию T-клеток. Например, метод костимуляции T-клеток in vitro включает контакт очищенных T-клеток, которые экспрессируют PD-1, с первым или первичным сигналом активации в форме моноклонального антитела против CD3 или форболового эфира или антигена в комплексе с MHC класса II. Способность рассматриваемого соединения в качестве агента снижать экспрессию или активность PD-1 и, следовательно, обеспечивать вторичный или костимулирующий сигнал, необходимый для модуляции иммунной функции этих T-клеток, может быть проанализирована с помощью любого из традиционных тестов, хорошо известных в данной области техники.
Ответ B-клеток на антагонист PD-1 может быть оценен с помощью специфичного в отношении LCMV ELISA, ELISPOT для плазматических клеток, теста для B-клеток памяти, фенотипирования B-клеток и анализа центров размножения с помощью иммуногистохимии.
Методы лечения: адоптивная иммунотерапия
В настоящем описании раскрыты методы лечения заинтересованного индивидуума, такого как индивидуум с устойчивой вирусной инфекцией или опухолью. Методы включают введение терапевтически эффективного количества цитотоксичных T-клеток, специфичных в отношении интересующего антигена, такого как вирусный антиген или опухолевый антиген, и терапевтически эффективного количества антагониста PD-1.
Раскрытые в настоящем описании методы предназначены для повышения иммунного ответа, такого как усиление иммунной системы у индивидуума. Введение очищенных специфичных в отношении антигена T-клеток и антагониста PD-1, как раскрыто в настоящем описании, должно повышать способность индивидуума к преодолению патологических состояний, таких как инфекционное заболевание или опухоль, путем направления иммунного ответа против патогена (такого как вирус или грибок) или неоплазмы. Таким образом, путем очистки и создания очищенной популяции отобранных специфичных в отношении антигена T-клеток индивидуума ex vivo и введения терапевтического количества таких клеток повышают иммунный ответ индивидуума-реципиента. Введение терапевтически эффективного количества антагониста PD-1 также повышает иммунный ответ реципиента.
В настоящем описании предлагаются методы индукции у реципиента иммунного ответа на интересующий антиген. Реципиентом может быть любой заинтересованный индивидуум, включая индивидуума с хронической инфекцией, такой как вирусная или грибковая инфекция, или индивидуума с опухолью. Такие инфекции описаны выше.
Инфекции у людей с иммунодефицитом являются обычной проблемой для реципиентов аллотрансплантатных стволовых клеток и реципиентов трансплантатов органов с постоянной иммунодепрессией. Возникающие в результате недостаточности T-клеток инфекции у таких индивидуумов обычно связаны с повторной активацией вирусов, уже имеющихся у реципиента. Например, после заражения большинство вирусов группы герпеса (такие как CMV, EBV, VZV, HSV) являются дремлющими и остаются в подавленном состоянии благодаря T-клеткам. Однако при подавлении иммунитета больных с помощью традиционных режимов дремлющие вирусы могут вновь активироваться. Например, в результате иммунодепрессии у больных может происходить повторная активация CMV, повторная активация вируса Эпштейна-Барр (EBV), которая вызывает опухоль B-клеток (EBV лимфопролиферативное заболевание), и повторная активация вируса BK, которая вызывает геморрагический цистит. Дополнительными примерами T-клеточного иммунодефицита служат ВИЧ-инфекция и врожденный иммунодефицит. Эти вирусные инфекции и повторные активации могут быть результатом подавления иммунитета у индивидуумов.
В нескольких вариантах осуществления реципиенту трансплантата костного мозга обеспечивается иммунный ответ против опухоли. Противоопухолевый иммунитет может быть обеспечен индивидууму путем введения специфичных в отношении антигена T-клеток, которые узнают опухолевый антиген. Такое введение реципиенту должно повышать иммунный ответ реципиента на опухоль за счет снабжения T-клетками, которые направлены на интересующий опухолевый антиген, узнают его и иммунологически взаимодействуют с ним.
В одном примере метод включает выделение из донора популяции донорских клеток, включая T-клетки (такие как мононуклеарные клетки периферической крови), и контактирование популяции донорских клеток, включающей T-клетки, с популяцией антигенпрезентирующих клеток (APC) от донора, которые презентируют интересующий антиген, необязательно в присутствии PD-1 с получением в результате популяции донорских клеток, включающей активированные донорские CD4+ и/или CD8+ T-клетки при сниженном количестве аллореактивных T-клеток, которые узнают интересующий антиген. Реципиенту вводят терапевтически эффективное количество популяции активированных донорских CD4+ и/или CD8+ T-клеток, обеспечивая тем самым у реципиента иммунный ответ на интересующий антиген. Введение очищенных специфичных в отношении антигена T-клеток может повышать способность индивидуума к преодолению патологических состояний, таких как инфекционное заболевание или опухоль, путем направления иммунного ответа против патогена (такого как вирус или грибок) или неоплазмы. В результате у реципиента обеспечивается иммунный ответ против интересующего антигена.
В нескольких осуществлениях метод также включает введение индивидууму терапевтически эффективного количества антагониста PD-1. Введение антагонистов PD-1 подробно описано выше.
Для создания популяции T-клеток, специфичных в отношении данного интересующего антигена, может быть использован любой антигенный пептид (такой как иммуногенный фрагмент) из интересующего антигена. В данной области техники известно множество таких антигенных пептидов, таких как вирусные и опухолевые антигены (см., например, таблицы 1-2). Настоящее раскрытие не ограничивается использованием специфичных антигенных пептидов. Конкретные примеры антигенных пептидов из интересующих антигенов включают, но не ограничиваются этим, антигены, которые являются вирусными, грибковыми или опухолевыми антигенами, такими как показано в таблице 1. В данной области техники известны дополнительные антигенные пептиды (например, см. Novellino et al., Cancer Immunol. Immunother. 54(3):187-207, 2005, и Chen et al., Cytotherapy, 4:41-8, 2002, обе включены в настоящее описание в качестве ссылки).
Хотя в таблице 1 раскрыты конкретные фрагменты полноразмерных интересующих антигенов, специалист в данной области техники должен понимать, что в раскрытых в настоящем описании методах могут быть также использованы другие фрагменты или полноразмерный белок. В одном примере интересующий антиген представляет собой «иммуногенный фрагмент» полноразмерной последовательности антигена. «Иммуногенный фрагмент» относится к части белка, который при презентации клеткой в контексте молекулы MHC может в тесте активации T-клеток активировать T-клетки против экспрессирующей белок клетки. Обычно такие фрагменты, которые связываются молекулами MHC класса I, представляют собой последовательность из от 8 до 12 аминокислот из полноразмерного антигена, хотя, разумеется, можно использовать более длинные фрагменты. В некоторых примерах иммуногенным фрагментом является такой фрагмент, который способен специфически связываться молекулой MHC на поверхности APC без дополнительного процессинга последовательности эпитопа. В отдельных примерах иммуногенный фрагмент представляет собой последовательность из 8-50 аминокислот из последовательности полноразмерного антигена, например из 8-20 аминокислот из 8-15 аминокислот, из 8-12 аминокислот, из 8-10 аминокислот, или 8, 9, 10, 11, 12, 13, 14, 15 или 20 последовательно расположенных аминокислот из последовательности полноразмерного антигена. В некоторых примерах клетки APC инкубируют с иммуногенным фрагментом в условиях, достаточных для специфического связывания иммуногенного фрагмента с молекулами MHC на поверхности APC, без потребности во внутриклеточном процессинге.
В одном примере антиген включает пептид из интересующего антигена с аминокислотной последовательностью, содержащей мотив связывания молекулой HLA индивидуума. Такие мотивы хорошо известны в данной области техники. Например, HLA-A2 является обычным аллелем в человеческой популяции. Связывающий мотив для этой молекулы включает пептиды из 9 или 10 аминокислот, содержащие лейцин или метионин во втором положении и валин или лейцин в последних положениях (см. примеры выше). Пептиды, которые включают такие мотивы, могут быть получены любыми методами, известными в данной области техники (такими как рекомбинантные, химические и т.д.). Зная аминокислотную последовательность интересующего антигена, можно определить последовательности иммуногенных фрагментов с предсказанным связыванием с MHC с помощью опубликованных программ. Например, для предсказания эпитопов любого связанного с опухолью, вирусом или грибком антигена можно с помощью обычных методов использовать программу для связываемого HLA мотива в Интернете (Bioinformatics and Molecular Analysis Section-BIMAS). Интересующие антигены (либо полноразмерные белки, либо их иммуногенные фрагменты) можно затем получить и очистить с помощью стандартных методов. Например, интересующие эпитопы или полноразмерные антигены можно получить рекомбинантным путем или с помощью химического синтеза с использованием стандартных методов. В значительной мере очищенный пептидный препарат должен давать единственную главную полосу в невосстанавливающем полиакриламидном геле. В других примерах интересующий антиген включает сырой вирусный лизат.
В одном примере интересующий антиген представляет собой связанный с опухолью антиген, а аминокислотные последовательности, содержащие мотивы связывания с HLA, представляют собой такие последовательности, которые определяют субдоминантные или скрытые эпитопы. Такие эпитопы можно выявить по более низкому относительному сродству связывания к молекуле HLA по сравнению с другими эпитопами в молекуле или по сравнению с другими молекулами, которые связываются с молекулой HLA.
Посредством изучения аналогов антигена с одиночными заменами аминокислот и секвенирования эндогенно связанных, подвергнутых естественному процессингу пептидов, были идентифицированы критические остатки, которые соответствуют мотивам, необходимым для специфического связывания молекул антигена с HLA (см., например, Southwood et al., J. Immunol. 160:3363, 1998; Rammensee et al., Immunogenetics 41:178, 1995; Rammensee et al., J. Curr. Opin. Immunol. 10:478, 1998; Engelhard, Curr. Opin. Immunol. 6:13, 1994; Sette and Grey, Curr. Opin. Immunol. 4:79, 1992). Более того, рентгеноструктурный кристаллографический анализ комплексов HLA-пептид выявил карманы внутри пептидсвязывающей щели молекул HLA, которые обеспечивают размещение остатков пептидных лигандов специфичным для аллеля способом; эти остатки в свою очередь определяют способность HLA связывать пептиды, в которых они находятся. (См., например, Madden, Annu. Rev. Immunol. 13:587, 1995; Smith et al., Immunity 4:203, 1996; Fremont et al., Immunity 8:305, 1998; Stern et al., Structure 2:245, 1994; Jones, Curr. Opin. Immunol. 9:75, 1997; Brown et al., Nature 364:33, 1993.)
Интересующий антиген выбирают на основе индивидуума, подлежащего лечению. Например, если индивидуум нуждается в повышенном противовирусном или противогрибковом иммунитете, выбирают один или более связанных с вирусом или грибком антигенов-мишеней. Иллюстративные интересующие антигены из вирусов включают, наряду с другими, антигены из вируса Эпштейна-Барр (EBV), вируса гепатита C (HCV), цитомегаловируса (CMV), простого вируса герпеса (HSV), вируса BK, вируса JC и вируса иммунодефицита человека (ВИЧ). Иллюстративные интересующие антигены из грибков включают антигены из Candida albicans, Cryptococcus, Blastomyces и Histoplasma или другого инфекционного агента. В другом примере индивидуум нуждается в повышенном противоопухолевом иммунитете. Иллюстративные интересующие антигены из опухолей включают WT1, PSA, PRAME. Иллюстративные интересующие антигены перечислены в таблицах 1 и 2. В некоторых примерах интересующий антиген включает, и вирусный антиген, и опухолевый антиген, и грибковый антиген, и опухолевый антиген, или вирусный антиген, грибковый антиген и опухолевый антиген.
Для лечения индивидуума с опухолью интересующий опухолевый антиген выбирают на основании экспрессии белка опухолью реципиента. Например, если у реципиента имеется опухоль молочной железы, выбирают антиген опухоли молочной железы, а если у реципиента имеется опухоль простаты, то выбирают антиген опухоли простаты, и т.д. В таблице 2 перечислены иллюстративные опухоли и соответствующие связанные с опухолью антигены, которые можно использовать для получения очищенных специфичных в отношении антигена T-клеток, которые можно ввести индивидууму с конкретной опухолью. Однако специалист в данной области техники должен понимать, что те же самые или другие опухоли можно лечить с использованием дополнительных опухолевых антигенов.
В одном примере специфичные в отношении антигена T-клетки, которые узнают опухолевый антиген, вводят в терапевтически эффективном количестве индивидууму, которому был или должен быть введен аллотрансплантат или аутотрансплантат стволовых клеток или которого вакцинировали опухолевым антигеном. Например, можно вводить терапевтически эффективное количество специфичных в отношении антигена T-клеток, которые распознают один или более связанных с опухолью антигенов, например, по меньшей мере, интересующих антигенов, перечисленных в таблицах 1 и 2.
В конкретных примерах, когда у реципиента имеется опухоль или когда у него имеется или когда им должен быть получен аллотрансплантат, донорные специфичные в отношении антигена опухоли T-клетки и антагонист PD-1 вводят в терапевтически эффективном количестве после пересадки аллотрансплантата стволовых клеток для предотвращения, снижения или задержки рецидива опухоли или для лечения злокачественного рецидива. Очищенные специфичные в отношении антигена T-клетки можно вводить индивидууму после снижения объема. Еще в одном примере реципиента вакцинируют интересующим опухолевым антигеном, очищенными специфичными в отношении антигена T-клетками, очищенными из реципиента и затем повторно введенными реципиенту с терапевтически эффективным количеством антагониста PD-1 для усиления реакции иммунной системы реципиента против опухоли.
Введение терапевтически эффективного количества специфичных в отношении антигена T-клеток и терапевтически эффективного количества антагониста PD-1 можно использовать профилактически для предотвращения рецидива опухоли у реципиента или для лечения рецидива опухоли. Такие специфичные в отношении антигена T-клетки могут вызывать гибель клеток, содержащих связанный с опухолью антиген, или помогать другим иммунным клеткам.
В отдельном примере у реципиента имеется опухоль и у него имеется или он должен получить аллотрансплантат стволовых клеток для восстановления иммунитета. После облучения костного мозга или введения цитотоксичного лекарства, которое разрушило или иным образом нарушило функцию костного мозга, вводят по меньшей мере два типа донорских специфичных в отношении антигена T-клеток в терапевтически эффективном количестве; специфичные в отношении антигена T-клетки, которые узнают связанный с вирусом антиген (или связанный с грибком антиген), и специфичные в отношении антигена T-клетки, которые специфически узнают связанный с опухолью антиген. Кроме того, индивидууму вводят терапевтически эффективное количество антагониста PD-1. Такое введение можно использовать для индукции противоопухолевого эффекта и противовирусного эффекта (такого как противовирусный эффект).
Для получения популяции специфичных в отношении антигена T-клеток для введения заинтересованному индивидууму популяция клеток, включающая T-клетки, может контактировать с антигенпрезентирующими клетками (APC), такими как дендритные клетки или T-APC, для презентации интересующего антигена. В некоторых вариантах осуществления отвечающие T-клетки (такие как лимфоциты или PBMC) обрабатывают антагонистом PD-1 и добавляют к APC, презентирующим один или более интересующих антигенов, и инкубируют в условиях, достаточных для обеспечения взаимодействия между презентирующими антиген APC и T-клетками, с получением специфичных в отношении антигена T-клеток. Обработка отвечающих T-клеток антагонистом PD-1 может производиться одновременно с контактированием или APC. Обработка антагонистом PD-1 может также производиться непосредственно перед контактом с APC.
Таким образом, в настоящем описании предлагаются методы получения обогащенной популяции специфичных в отношении антигена T-клеток. Обычно T-APC презентируют антигены T-клеткам и индуцируют ограниченный MHC ответ класса I (CD8+ T-клетки) и класса II (CD4+ T-клетки) ограниченного типа. Типичный ответ T-клеток представляет собой активацию и пролиферацию. Таким образом, образуется популяция, которая включает T-клетки, которые специфически узнают интересующий антиген. Таким образом, индивидууму, такому как индивидуум с хронической инфекцией или опухолью, можно вводить терапевтически эффективное количество этой популяции клеток для получения иммунного ответа.
Обычно клетки APC и T-клетки являются аутологичными. В конкретных неограничивающих примерах клетки APs и отвечающие T-клетки берут из одного и того же индивидуума. Однако APC и отвечающие T-клетки могут быть синергичными. APC можно использовать для презентации антигена популяции аутологичных T-клеток. Специалисту в данной области техники должно быть ясно, что антигенные пептиды, которые связываются с молекулами MHC класса I и II, могут быть получены ex vivo (например, вместо процессинга из полноразмерного белка в клетке) и обеспечивать взаимодействие (такое как связывание) с молекулами MHC класса I и II на клеточной поверхности. Обычно APC презентируют антиген в контексте обоих классов I и II MHC.
В одном примере интересующий антиген, инкубированный с APC, представляет собой гибридный белок, который включает аминокислотную последовательность из интересующего антигена (такую как последовательность из 8-50 аминокислот, например последовательность из 8-15 или 8-12 аминокислот из интересующего антигена). Так, ряд связывающихся MHC эпитопов может быть включен в один антигенный полипептид, или может быть использован тример в одной цепи, где каждый тример содержит молекулу MHC класса I, b2 микроглобулин и интересующий антигенный пептид (см. Nature 2005; V.436, р.578). В некоторых примерах используют только один антиген, но в других вариантах осуществления используют более одного антигена, например по меньшей мере 2 разных антигена, по меньшей мере 3 разных антигена, по меньшей мере 4 разных антигена, по меньшей мере 5 разных антигенов, по меньшей мере 10 разных антигенов, по меньшей мере 15 разных антигенов, по меньшей мере 20 разных антигенов или даже по меньшей мере 50 разных антигенов.
В еще одних примерах интересующий антиген представляет собой полноразмерную антигенную аминокислотную последовательность (такую как полноразмерный грибковый антиген, опухолевый антиген или вирусный антиген, например вирусный лизат или полноразмерный катепсин G). В дополнительных примерах можно использовать один или более антигенов из инфицирующего агента. В некоторых примерах интересующий полноразмерный антиген экспрессируется APC.
APC могут быть получены с помощью методов, известных специалисту в данной области техники (см. Melenhorst et al., Cytotherapy 7, supp.1, 2005; Melenhorst et al., Blood 106:671a, 2005; Gagliardi et al., Int. Immunol. 7:1741-52, 1995, включенные в настоящее описание в качестве ссылки). В одном примере для получения T-APC моноциты периферической крови донора активируют с помощью ИЛ-2 и антитела, которое специфически связывает CD3 (такого как OKT3) в течение приблизительно трех или более дней, например приблизительно от одной до двух недель, например приблизительно в течение от семи до десяти дней.
Наблюдалось, что в присутствии презентируемого антигена T-клетки, которые узнают антиген, связываются с антигенпрезентирующими клетками (APC), презентирующими интересующий антиген, сильнее, чем клетки, которые не являются специфичными в отношении антигена (и таким образом не связываются специфичным для антигена образом). В отдельном примере специфичные в отношении антигена T-клетки отбирают путем экспозиции APCs с целевым пептидным антигеном (таким как целевой вирусный или связанный с опухолью антиген), против которого должны быть направлены желаемые T-клетки, в присутствии антагониста PD-1, так что APC презентирует антиген в связи с главным комплексом гистосовместимости (MHC) класса I и/или класса II. Например, APC могут быть экспонированы с достаточным количеством интересующего антигена для достаточной оккупации молекул MHC на поверхности APC (например, оккупируется по меньшей мере 1% молекул MHC, например по меньшей мере 5%, по меньшей мере 7,5% или по меньшей мере 10%) и могут стимулировать предпочтительное связывание T-клеток-мишеней в присутствии антагониста PD-1 с APC, презентирующими интересующий антиген (по сравнению с APC, которые не презентируют интересующий антиген). Популяцию T-клеток, такую как популяция, которая была примирована интересующим антигеном, затем инкубируют с APC необязательно в присутствии антагониста PD-1, такого как антитело, которое специфически связывает PD-1, для предпочтительной активации клеток с получением таким образом популяции клеток, обогащенной желаемыми T-клетками, которые узнают интересующий антиген.
T-клетки, такие как находящиеся в популяции PBMC или лимфоцитов, можно инкубировать с одним или более интересующими антигенами необязательно в присутствии антагониста PD-1 для получения популяции T-клеток, которая примирована в отношении одного или более интересующих антигенов. T-клетки можно примировать с помощью любого метода, известного в данной области техники. В отдельных примерах PBMC или лимфоциты инкубируют в присутствии очищенного целевого пептидного антигена необязательно в присутствии антагониста PD-1. В некоторых примерах интересующий антиген представляет собой вирусный или опухолевый антиген, такой как, но не ограничиваясь этим, один или более интересующих антигенов, перечисленных в таблице 1. Интересующий антиген может находиться в очищенной форме, такой как химически синтезированный пептид. В других примерах интересующий антиген находится в неочищенной форме, такой как сырой лизат, например вирусный лизат.
Количество антигена, используемое для примирования T-клеток, может быть легко определено с помощью методов, известных в данной области техники. Обычно, если антиген используют в очищенной форме, используют приблизительно 1-10 мкг/мл пептида. При использовании вирусного лизата можно использовать приблизительно 0,1-100 мкл лизата, например приблизительно 75 мкл. При использовании T-APC можно использовать приблизительно 4-6 миллионов T-APC, презентирующих интересующий антиген, на каждые 40-60 миллионов T-клеток (или лимфоцитов, или PBMC).
В отдельном примере лимфоциты примируют in vitro путем их инкубации с растворимым антигеном или вирусным лизатом в течение 5-7 дней в условиях, которые обеспечивают примирование T-клеток. Жизнеспособные T-клетки выделяют, например, с помощью центрифугирования в фиколле-гипаке с получением в результате примированных T-клеток. Если это желательно, жизнеспособные примированные T-клетки можно вновь примировать один или более раз, например, путем инкубации с антигеном еще в течение 5-7 дней в тех же условиях, которые использовались для первого примирования, и выделить жизнеспособные T-клетки.
В другом примере лимфоциты примируют in vivo путем инокуляции индивидууму антигена, например, в форме вакцины. В этом примере T-клетки, полученные из индивидуума после иммунизации, уже примированы. Например, лимфоциты или PBMC, полученные из индивидуума, затем инкубируют с APC в присутствии антагониста PD-1, как описано в настоящем описании, без необходимости в дополнительном примировании.
Метод может дополнительно включать получение APC, которые презентируют интересующий антиген. Например, APC можно инкубировать с достаточным количеством одного или более разных пептидных антигенов в условиях, достаточных для презентации целевого пептида (пептидов) на поверхности APC. Это создает популяцию APC, которые презентируют интересующий антиген на молекулах MHC на поверхности APC. Раскрытые методы не ограничены конкретными методами презентации интересующего антигена на поверхности APC.
Антигены могут также экспрессироваться APC либо естественным образом, либо благодаря вставке гена, содержащей последовательность ДНК, кодирующей целевой белок (антиген). Нуклеиновая кислота, кодирующая интересующий антиген, может быть введена в T-клетки в виде матричной РНК или с помощью вектора, такого как экспрессионный вектор для млекопитающих, или вирусного вектора (например, аденовирусного, поксвирусного или ретровирусного векторов). Полинуклеотиды, кодирующие интересующий антиген, включают рекомбинантную ДНК, которая представляет собой автономно реплицируемую плазмиду или вирус, или которая включена в геномную ДНК эукариота, или которая существует в виде отдельной молекулы, независимой от других последовательностей. Нуклеиновую кислоту, кодирующую интересующий антиген, можно также вводить с помощью электропорации, липофекции или основанной на фосфате кальция трансфекции.
Был сконструирован ряд вирусных векторов, включая полиому, т.е. SV40 (Madzak et al., 1992, J. Gen. Virol., 73:1533-1536), аденовирус (Berkner, 1992, Cur. Top. Microbiol. Immunol., 158:39-6; Berliner et al., 1988, Bio Techniques, 6:616-629; Gorziglia et al., 1992, J. Virol., 66:4407-4412; Quantin et al., 1992, Proc. Nad. Acad. Sci. USA, 89:2581-2584; Rosenfeld et al., 1992, Cell, 68:143-155; Wilkinson et al., 1992, Nucl. Acids Res., 20:2233-2239; Stratford-Perricaudet et al., 1990, Hum. Gene Ther., 1:241-256), вирус оспы (Mackett et al., 1992, Biotechnology, 24:495-499), аденоассоциированный вирус (Muzyczka, 1992, Curr. Top. Microbiol. Immunol., 158:91-123; On et al., 1990, Gene, 89:279-282), вирусы герпеса, включая HSV, CMV и EBV (Margolskee, 1992, Curr. Top. Microbiol. Immunol., 158:67-90; Johnson et al., 1992, J. Virol., 66:2952-2965; Fink et al., 1992, Hum. Gene Ther. 3:11-19; Breakfield et al., 1987, Mol. Neurobiol., 1:337-371; Fresse et al., 1990, Biochem. Pharmacol., 40:2189-2199), вирусы синдбис (H. Herweijer et al., 1995, Human Gene Therapy 6:1161-1167; патенты США №№ 5091309 и 52217879), альфавирусы (S. Schlesinger, 1993, Trends Biotechnol. 11:18-22; I. Frolov et al., 1996, Proc. Natl. Acad. Sci. USA 93:11371-11377) и ретровирусы птиц (Brandyopadhyay et al., 1984, Mol. Cell Biol., 4:749-754; Petropouplos et al., 1992, J. Virol., 66:3391-3397), мыши (Miller, 1992, Curr. Top. Microbiol. Immunol., 158:1-24; Miller et al., 1985, Mol. Cell Biol., 5:431-437; Sorge et al., 1984, Mol. Cell Biol., 4:1730-1737; Mann et al., 1985, J. Virol., 54:401-407) и человека (Page et al., 1990, J. Virol., 64:5370-5276; Buchschalcher et al., 1992, J. Virol., 66:2731-2739). В данной области техники известны также бакуловирусные (Autographa californica многоядерный полиэдроидный вирус; AcMNPV) векторы, и они могут быть получены из коммерческих источников (таких как PharMingen, San Diego, Calif.; Protein Sciences Corp., Meriden, Conn.; Stratagene, La Jolla, Calif.).
В одном варианте осуществления полинуклеотид, кодирующий интересующий антиген, включают в вирусный вектор для транспортировки в APC. Подходящие векторы включают ретровирусные векторы, ортопоксные векторы, авипоксные векторы, фаулпоксные векторы, каприпоксные векторы, суипоксные векторы, аденовирусные векторы, векторы вируса герпеса, альфавирусные векторы, бакуловирусные векторы, векторы вируса синдбис, векторы вируса оспы и полиовирусные векторы. Конкретными иллюстративными векторами являются поксвирусные векторы, такие как вирус оспы, фаулпоксный вирус и крайне ослабленный вирус оспы (MVA), аденовирус, бакуловирус и тому подобное.
Используемые поксвирусы включают ортопоксный, суипоксный, авипоксный и капрпоксный вирусы. Ортопокс включает оспу, эктромелию и раккунпокс. Одним из примеров используемого ортопокса является оспа. Авипокс включает фаулпокс, оспу канареек и оспу свиней. Каприпокс включает оспу коз и оспу овец. В одном примере суипокс представляет собой оспу свиней. Примеры поксвирусных векторов для экспрессии описаны, например, в патенте США № 6165460, который включен в настоящее описание в качестве ссылки. Другие вирусные векторы, которые могут быть использованы, включают другие ДНК-вирусы, такие как вирус герпеса и аденовирусы, и РНК-вирусы, такие как ретровирусы и вирус полиомиелита.
Подходящие векторы раскрыты, например, в патенте США № 6998252, который включен в настоящее описание в качестве ссылки. В одном примере рекомбинантный поксвирус, такой как рекомбинантный вирус оспы, синтетическим путем модифицируют путем вставки гибридного гена, содержащего регуляторные последовательности оспы или ДНК последовательности, функционально эквивалентные фланкирующим последовательностям ДНК, которые в естественных условиях не соседствуют с фланкирующими регуляторными последовательностями ДНК оспы, которые кодируют интересующий антиген. Рекомбинантный вирус, содержащий такой гибридный ген, эффективен для экспрессии антигена. В одном примере вектор вируса оспы включает (A) сегмент, состоящий из (i) первой последовательности ДНК, кодирующей антиген, и (ii) промотора поксвируса, где промотор поксвируса находится рядом с последовательностью ДНК, кодирующей антигенный полипептид, и осуществляет ее транскрипционный контроль; и рядом с указанным сегментом (B) ДНК из несущественной области поксвируса. Вирусный вектор может кодировать маркер селекции. В одном примере поксвирус включает, например, ген тимидинкиназы (см. патент США № 6998252, который включен в настоящее описание в качестве ссылки).
Популяцию APC, которые презентируют антиген(ы) в достаточном количестве, инкубируют с T-клетками (такими как лимфоциты или PBMC) необязательно в присутствии эффективного количества антагониста PD-1 в условиях, достаточных для обеспечения связывания презентирующих антиген APC с T-клетками, которые могут специфически иммунологически взаимодействовать с антигеном (специфичными в отношении антигена T-клетками). Используют достаточное количество APC, экспрессирующих антиген вместе с MHC в количестве, достаточном для стимуляции повышения связывания желаемой T-клетки с APC. В отдельных примерах по меньшей мере 20% APC презентируют желаемый антиген на молекулах MHC на поверхности APC, например по меньшей мере 30% APC, по меньшей мере 40% APC, по меньшей мере 50% APC или по меньшей мере 60% APC. Оптимальное количество добавленных T-клеток может варьировать в зависимости от использованного количества APC. В некоторых примерах используют отношение T-клетки: APC, равное по меньшей мере 6:1, например по меньшей мере 8:1, по меньшей мере 10:1, по меньшей мере 12:1, по меньшей мере 15:1, по меньшей мере 16:1, по меньшей мере 20:1 или даже по меньшей мере 50:1.
Для повышения количества специфичных в отношении антигена T-клеток можно стимулировать пролиферацию клеток, например, путем инкубации в присутствии цитокина, такого как ИЛ-2, ИЛ-7, ИЛ-12 и ИЛ-15. Количество добавленного цитокина является достаточным для стимуляции получения и пролиферации T-клеток и может быть определено с помощью традиционных методов. В некоторых примерах количество добавленных ИЛ-2, ИЛ-7, ИЛ-12 или ИЛ-15 составляет приблизительно 0,1-100 МЕ/мл, например по меньшей мере 1 МЕ/мл, по меньшей мере 10 МЕ/мл или по меньшей мере 20 МЕ/мл.
После связывания достаточного количества специфичных в отношении антигена T-клеток с APC получают T-клетки, которые специфично узнают интересующий антиген. Это создает популяцию обогащенных (например, очищенных) специфичных в отношении антигена T-клеток, которые специфичны в отношении интересующего антигена. В некоторых примерах полученная популяция T-клеток, которые специфичны в отношении интересующего антигена, имеет по меньшей мере 30% чистоту, например по меньшей мере 40% чистоту или даже по меньшей мере 50% чистоту. Чистота популяции T-клеток, специфичных в отношении антигена, может быть оценена с помощью методов, известных специалисту в данной области техники.
В одном примере во время стимуляции пролиферации специфичных в отношении антигена T-клеток клетки можно подсчитать для определения количества клеток. После достижения желаемого количества клеток определяют чистоту. Чистота может быть определена, например, с использованием маркеров, находящихся на поверхности специфичных в отношении антигена T-клеток, одновременно с оценкой продукции цитокинов после распознавания антигена, таких как интерферон (IFN)γ, фактор некроза опухолей (TNF)α, интерлейкин (ИЛ)-2, ИЛ-10, трансформирующий фактор роста (TGF)β1 или ИЛ-4. Обычно специфичные в отношении антигена T-клетки являются позитивными по маркеру CD3, наряду с маркером CD4 или CD8 и IFNγ (который специфичен для активированных T-клеток). Например, для идентификации (и сортировки, если это желательно) популяций клеток, которые позитивны по CD3, CD4 или CD8 и IFN-γ, можно использовать сортинг флуоресцентно активированных клеток (FACS) с использованием дифференциально окрашенных анти-CD3, анти-CD4, анти-CD-8 и анти-IFN-γ. Вкратце, стимулированные специфичные в отношении антигена T-клетки инкубируют в присутствии анти-CD3, анти-CD4, анти-CD-8 и анти-IFN-γ (причем каждое содержит различающийся присоединенный флуорофор) в течение времени, достаточного для связывания антитела с клетками. После удаления несвязанного антигена клетки анализируют с помощью FACS с использованием традиционных методов. Специфичными в отношении антигена T-клетками являются те клетки, которые являются позитивными по IFN-γ и позитивными по CD8 или позитивными по CD4. В отдельных примерах полученная популяция антигенных T-клеток имеет чистоту, составляющую, по меньшей мере, 30% относительно суммарной популяции CD4+ или CD8+ позитивных клеток, например по меньшей мере 40% чистоту, по меньшей мере 50% чистоту, по меньшей мере 60% чистоту или даже по меньшей мере 70% чистоту по сравнению с суммарной популяцией CD4 позитивных или CD8 позитивных клеток.
В другом примере метод дополнительно включает определение цитотоксичности специфичных в отношении антигена T-клеток. Методы определения цитотоксичности известны в данной области техники, например тест выделения 51Cr (см., например, Walker et al. Nature 328:345-8, 1987; Qin et al. Acta Pharmacol. Sin. 23(6):534-8, 2002; включенные в настоящее описание в качестве ссылки).
Специфичные в отношении антигена T-клетки могут быть подвергнуты одному или более раундам селекции для повышения чистоты специфичных в отношении антигена T-клеток. Например, очищенные специфичные в отношении антигена T-клетки, полученные, как описано выше, вновь инкубируют с APC, презентирующими интересующий антиген, в присутствии антагониста PD-1 в условиях, достаточных для обеспечения связывания APC с очищенными специфичными в отношении антигена T-клетками. Полученные специфичные в отношении антигена T-клетки можно стимулировать к пролиферации, например, с помощью ИЛ-2. Обычно полученные специфичные в отношении антигена T-клетки, которые специфически иммунологически взаимодействуют с интересующим антигеном, являются более чистыми после последующих раундов стимуляции с помощью APC по сравнению с клетками, полученными в результате лишь одного раунда селекции. В одном примере полученная популяция очищенных специфичных в отношении антигена T-клеток имеет по меньшей мере 90% чистоту относительно всех имеющихся CD3+ клеток, например по меньшей мере 95% чистоту или по меньшей мере 98% чистоту. В отдельном примере полученная популяция очищенных специфичных в отношении антигена T-клеток имеет по меньшей мере 95% чистоту относительно всех имеющихся CD4+ клеток, например по меньшей мере 98% чистоту. В другом примере полученная популяция очищенных специфичных в отношении антигена T-клеток имеет по меньшей мере 90% чистоту относительно всех имеющихся CD3+ клеток, например по меньшей мере 93% чистоту.
В настоящем раскрытии также предлагаются терапевтические композиции, которые включают обогащенные (например, очищенные) специфичные в отношении антигена T-клетки и антагонист PD-1. В отдельных примерах полученную обогащенную популяцию специфичных в отношении антигена T-клеток (специфичных в отношении интересующего антигена) помещают в лекарственную форму для введения нуждающемуся в этом индивидууму. Антагонист PD-1 также находится в лекарственной форме для введения нуждающемуся в этом индивидууму.
В одном примере полученная популяция очищенных специфичных в отношении антигена T-клеток имеет по меньшей мере 30% чистоту относительно всех имеющихся CD3+ клеток, например по меньшей мере 40% чистоту, по меньшей мере 50% чистоту, по меньшей мере 80% чистоту или даже по меньшей мере 90% чистоту. В отдельном примере полученная популяция очищенных специфичных в отношении антигена T-клеток имеет по меньшей мере 30% чистоту относительно всех имеющихся CD3+ клеток, например по меньшей мере 40% чистоту, по меньшей мере 50% чистоту, по меньшей мере 80% чистоту, по меньшей мере 90% чистоту, по меньшей мере 95% чистоту или даже по меньшей мере 98% чистоту. В другом примере полученная популяция очищенных специфичных в отношении антигена T-клеток имеет по меньшей мере 50% чистоту относительно всех имеющихся CD3+ клеток, например по меньшей мере 60% чистоту, по меньшей мере 75% чистоту, по меньшей мере 80% чистоту, по меньшей мере 90% чистоту или даже по меньшей мере 93% чистоту. Размноженные и отобранные специфичные в отношении антигена T-клетки можно проверить на наличие микоплазмы, стерильность, эндотоксин и качество, контролируемое по функции и чистоте, перед сохранением при низкой температуре или перед инфузией реципиенту.
Индивидууму вводят терапевтически эффективное количество специфичных в отношении антигена T-клеток. Конкретные неограничивающие примеры терапевтически эффективного количества очищенных специфичных в отношении антигена T-клеток включают очищенные специфичные в отношении антигена T-клетки, вводимые в дозе от приблизительно 1×105 клеток на килограмм индивидуума до приблизительно 1×109 клеток на килограмм индивидуума, например от приблизительно 1×106 клеток на килограмм до приблизительно 1×108 клеток на килограмм, например от приблизительно 5×106 клеток на килограмм до приблизительно 75×106 клеток на килограмм, например, приблизительно 25×106 клеток на килограмм или приблизительно 50×106 клеток на килограмм.
Очищенные специфичные в отношении антигена T-клетки можно вводить в одной или множестве доз по решению врача. Например, клетки можно вводить с интервалами, равными приблизительно двум неделям, в зависимости от желаемого ответа и полученного ответа. В некоторых примерах при получении желаемого ответа дополнительное введение специфичных в отношении антигена T-клеток не производят. Однако если у реципиента проявляются один или более симптомов, связанных с инфекцией или наличием или ростом опухоли, в этот момент можно ввести терапевтически эффективное количество специфичных в отношении антигена T-клеток. Введение может быть местным или системным.
Раскрытые в настоящем описании очищенные специфичные в отношении антигена T-клетки можно вводить с фармацевтически приемлемым носителем, таким как физиологический раствор. Антагонист PD-1 может быть составлен в фармацевтически приемлемом носителе, как описано выше. В некоторых примерах со специфичными в отношении антигена T-клетками и антагонистом PD-1 вводят другие терапевтические агенты. Другие терапевтические агенты можно вводить до, во время и после введения специфичных в отношении антигена T-клеток в зависимости от желаемого эффекта. Иллюстративные терапевтические агенты включают, но не ограничиваются этим, противомикробные агенты, иммуностимуляторы, такие как интерферон-альфа, химиотерапевтические агенты или пептидные вакцины того же антигена, который используется для стимуляции T-клеток in vitro. В отдельном примере композиции, содержащие очищенные специфичные в отношении антигена T-клетки, также включают один или более терапевтических агентов.
Раскрытие иллюстрируется следующими неограничивающими примерами.
Примеры
Пример 1
Ингибирование пути PD-1 у мышей с хронической инфекцией с помощью антител против PD-L1
Для изучения влияния хронической вирусной инфекции на функцию CD8+ T-клеток использовали мышей, инфицированных разными штаммами вируса лимфоцитарного хориоменингита (LCMV). Штамм Армстронга LCMV вызывает острую инфекцию, которая прекращается в пределах 8 дней, оставляя после себя долгоживущую популяцию высокофункциональных покоящихся CD8+ T-клеток памяти. Штамм C1-13 LCMV, напротив, вызывает устойчивую инфекцию у хозяина, отличающуюся виремией, которая длится до 3 месяцев. Вирус остается в некоторых тканях неограниченно долго, и специфичные в отношении антигена T-клетки становятся функционально неполноценными. CD8+ T-клетки DbNP396-404 физически исчезают, а CD8+ T-клетки DbGP33-41 и DbGP276-286 сохраняются, но теряют способность пролиферировать или секретировать противовирусные цитокины, такие как IFN-γ и TNF-α.
Мыши C57BL/6 были получены из Национального института рака (Frederick, MD). Мышей инфицировали внутривенно (в/в) 2×106 БОЕ LCMV-Cl-13. Истощение CD4 производили инъекцией 500 мкг GK1.5 в ЗФР в день инфицирования и на следующий день после инфицирования. Иммунизированных LCMV получают инфицированием мышей в/бр 2×105 БОЕ LCMV Армстронга.
Генетический анализ с помощью микроматриц проводили на очищенных FACS наивных специфичных в отношении DbGP33-41 P14- трансгенных CD8+ T-клетках, специфичных в отношении DbGP33-41 CD8+ T-клетках памяти, полученных из иммунизированных LCMV Армстронга мышей, и специфичных в отношении DbGP33-41 или DbGP276-286 CD8+ T-клетках, полученных из мышей, инфицированных LCMV Cl-13 с истощенным CD4+. Выделение РНК и генетический анализ с помощью микроматриц проводили, как описано в Kaech et al., (Cell 111:837-51, 2002). мРНК PD-1 экспрессировалась на высоком уровне в истощенных CD8+ T-клетках по сравнению с CD8+ T-клетках памяти (фиг.1A). Более того, PD-1 экспрессировался на поверхности CD8+ T-клеток у инфицированных LCMV C1-13 мышей, но отсутствовал на поверхности CD8+ T-клеток после клиренса LCMV Армстронга (фиг.1B). У хронически инфицированных мышей также на более высоком уровне экспрессировался один из лигандов PD-1, PD-L1, на большинстве лимфоцитов и APC по сравнению с неинфицированными мышами. Таким образом, сохранение вирусного антигена и истощение CD8+ T-клеток происходят одновременно с индукцией экспрессии PD-1.
Для проверки гипотезы о том, что блокада пути PD-1/PD-L1 может восстанавливать функцию T-клеток и повышать контроль над вирусом во время хронической инфекции LCMV, коингибиторный путь PD-1/PD-L1 разрушали во время хронической инфекции LCMV с помощью блокирующих антител против αPD-L1. Блокирующее моноклональное антитело против PD-L1 вводили внутрибрюшинно (в/бр) на каждый 3 день мышам, инфицированным LCMV C1-13 (200 мкг моноклональных антител IgG2b крысы против PD-L1 мыши (клон 10F.5C5 или 10F.9G2)), с 23 по 37 день после инфицирования. На 37 день у мышей, подвергнутых лечению, было приблизительно в 2,5 раз больше специфичных в отношении DbNP396-404 CD8+ T-клеток и в 3 раза больше специфичных в отношении DbGP33-41 CD8+ T-клеток по сравнению с контрольными мышами без лечения (фиг.2A). Индукция пролиферации была специфична для CD8+ T-клеток, поскольку количество CD4+ T-клеток в печени было приблизительно одинаковым у мышей, подвергнутых и не подвергнутых лечению (~6×104 IAbGP61-80 CD4+ T-клеток на селезенку).
Кроме повышения пролиферации CD8+ T-клеток, ингибирование сигнализации PD-1 также вызывало повышение продукции противовирусных цитокинов в специфичных в отношении вируса CD8+ T-клетках. Была определена продукция IFN-γ и IFN-α CD8+ T-клетками в ответ на восемь разных эпитопов CTL. Усредненный ответ был в 2,3 раза выше у мышей, подвергнутых лечению, по сравнению с мышами без лечения (фиг.2B и 2C). После лечения также наблюдалось 2-кратное повышение частоты встречаемости продуцирующих TNF-α клеток (фиг.2D). Удаление вируса также ускорялось, поскольку удалялся из сыворотки, селезенки и печени подвергнутых лечению мышей. Пониженные титры вируса наблюдались в легком и почке (~10-кратно) к 37 дню после инфицирования (через 14 дней после начала лечения) у подвергнутых лечению мышей. Однако у мышей, не подвергнутых лечению, выявлялись значительные уровни вируса в этих тканях (фиг.2E). Титры вируса в сыворотке и гомогенатах тканей определяли с помощью клеток Vero, как описано в Ahmed et al. (J. Virol. 51:34-41, 1984). Результаты, показывающие, что антагонист PD-1 повышает пролиферацию CD8+ T-клеток и удаление вируса, таким образом, указывают на то, что ингибирование сигнализации PD-1 восстанавливает функцию CD8+ T-клеток. Более того, ингибирование сигнализации PD-1 также повышает ответы B-клеток, поскольку количество секретирующих специфичное в отношении LCMV антитело клеток в селезенке также увеличивалось (>10-кратно) после лечения.
CD4+ T-клетки играют ключевую роль в формировании и поддержании ответов CD8+ T-клеток. В этом смысле CD8+ T-клетки, примированные в отсутствие CD4+ T-клеток (так называемые «беспомощные» CD8+ T-клетки), не способны формировать нормальные иммунные ответы. Более того, хроническая инфекция LCMV является более тяжелой в отсутствие CD4+ T-клеток. Соответственно, беспомощные T-клетки, возникшие во время инфекции LCMV-C1-13, проявляют даже более глубокое функциональное нарушение по сравнению с T-клетками, возникшими в присутствии CD4+ T-клеток. Специфичные в отношении DbNP396-404 CD8+ T-клетки удаляются до неопределимого уровня, а DbGP33-41 и DbGP276-286 CD8+ T-клетки полностью теряют способность секретировать IFN-γ и IFN-α.
CD4+ T-клетки истощались во время инфекции LCMV-C1-13, и мышей лечили обработкой антителами против PD-L1 с 46 по 60 день после инфицирования. Специфичные в отношении LCMV CD4+ T-клетки не выявлялись по данным внутриклеточного окрашивания IFN-γ до или после обработки. У мышей, лечившихся обработкой, было приблизительно в 7 раз больше DbGP276-286 CD8+ T-клеток и в 4 раза больше DbGP33-41 CD8+ T клеток в селезенке по сравнению с контрольными не подвергнутыми лечению мышами (фиг.3A). Количество специфичных в отношении вируса CD8+ T-клеток в селезенке также возрастало (фиг.3B). Это увеличение специфичных в отношении вируса CD8+ T-клеток у подвергнутых лечению мышей было объяснено усилением пролиферации, выявленным по включению BrdU. У мышей, не подвергнутых лечению, 43% DbGP276-286 CD8+ T-клеток включали BrdU на промежуточном уровне и 2% включали BrdU на высоком уровне, в то время как у мышей, подвергнутых лечению, 50% DbGP276-286 CD8+ T-клеток включали BrdU на промежуточном уровне и 37% включали BrdU на высоком уровне. Анализ с помощью BrdU проводили путем введения 1 мг/мл BrdU в питьевую воду во время обработки, и окрашивание производили в соответствии с протоколом производителя (BD Biosciences, San Diego, CA). Более того, у подвергнутых лечению мышей имелся более высокий процент CD8+ T-клеток, экспрессирующих связанный с клеточным циклом белок Ki67 (60% против 19% у мышей без лечения, фиг.3C). Ответ на лечение CD8+ T-клеток в PBMC был характерен только для мышей с высоким уровнем экспансии CD8+ T-клеток.
Ингибирование PD-1 также повышало продукцию противовирусного цитокина в беспомощных, истощенных специфичных в отношении вируса CD8+ T-клетках. После обработки количество DbGP33-41 и DbGP276-286 CD8+ T-клеток, продуцирующих IFN-γ, значительно увеличивалось (фиг.4A), хотя у подвергнутых лечению мышей было также выявлено большее количество CD8+ T-клеток, специфичных в отношении DbNP396-404, KbNP205-212, DbNP166-175 и DbGP92-101 (фиг.4A). 50% специфичных в отношении DbGP276-286 CD8+ T-клеток из подвергнутых лечению мышей могут продуцировать IFN-γ по сравнению с 20% специфичных в отношении DbGP276-286 CD8+ T-клеток у контрольных не подвергнутых лечению мышей (фиг.4B). Уровни IFN-γ и TNF-α, продуцируемых специфичными в отношении DbGP276-286 CD8+ T-клетками из подвергнутых лечению мышей, были, однако, ниже по сравнению с полностью функциональными специфичными в отношении DbGP276-286 клетками памяти (фиг.4C).
Ингибирование PD-1 также повышало лизирующую активность беспомощных, истощенных специфичных в отношении вируса CD8+ T-клеток. Лизирующую активность специфичных в отношении вируса CD8+ T-клеток ex vivo определяли после лечения с использованием теста выделения 51Cr (Wherry et al., 2003. J. Virol. 77:4911-27). Титры вируса снижались приблизительно в 3 раза в селезенке, в 4 раза в печени, в 2 раза в легком и в 2 раза в сыворотке через 2 недели лечения по сравнению с мышами, не подвергнутыми лечению (фиг.4E).
Эти результаты, таким образом, демонстрируют, что блокада пути PD-1 нарушает толерантность периферических CTL к хронической вирусной инфекции и что истощенные CD8+ T-клетки, лишенные поддержки со стороны CD4+ T-клеток, не являются необратимо инактивированными.
Пример 2
Введение противовирусной вакцины и антагониста PD-1
Один подход к поддержанию T-клеточных ответов во время устойчивой инфекции заключается в терапевтической вакцинации. Обоснование для этого подхода заключается в том, что эндогенные антигены могут находиться не в оптимальной или иммуногенной форме во время хронической вирусной инфекции, и что обеспечение иммуногеном в форме вакцины может обеспечивать более эффективный стимул для специфичных в отношении вируса T- и B-клеток. При использовании модели хронического LCMV мышам вводили рекомбинантный вирус оспы, экспрессирующий эпитоп LCMV GP33, в качестве терапевтической вакцины (VVGP33), что приводило к умеренному повышению ответов CD8+ T-клеток у некоторых хронически инфицированных мышей. У четырех из девяти хронически инфицированных мышей, которые получали терапевтическую вакцину, проявлялся положительный ответ, в то время как ни у одной из контрольных мышей не было значительного повышения иммунного ответа против GP33. При сочетании этой терапевтической вакцинации с ингибитором PD-1 ответы специфичных в отношении LCMV T-клеток поддерживались на более высоком уровне по сравнению с любым раздельным лечением, и действие сочетанного лечения превосходило аддитивное действие.
Пример 3
Ингибирование пути PD-1 у хронически инфицированных мышей с использованием РНКи PD-1
Интерферирующая РНК (РНКи) способна подавлять экспрессию гена в клетках млекопитающего. Длинные двухцепочечные РНК (дцРНК) вводят в клетки, и затем они подвергаются процессингу до малых сайленсинговых РНК (миРНК), которые направлены против конкретных молекул мРНК или небольшой группы мРНК. Эта методология особенно пригодна в ситуациях, когда антитела не функционируют. Например, РНКи можно использовать в ситуации, когда уникальные сплайсинговые варианты продуцируют растворимые формы PD-1 и CTLA-4.
Сайленсные РНК для PD-1 вставляют в имеющийся в продаже экспрессионный вектор для миРНК, такой как экспрессионные векторы pSilencerTM или аденовирусные векторы (Ambion, Austin, TX). Эти векторы затем вводят в контакт с истощенными T-клетками-мишенями in vivo или ex vivo (см. пример 4 ниже).
Пример 4
Ex vivo омоложение истощенных T-клеток
Специфичные в отношении вируса CD8+ T-клетки выделяют из хронически инфицированных LCMV-Cl-13 мышей с помощью магнитных шариков или центрифугирования в градиенте плотности. Трансфицированные CD8+ T-клетки вводят в контакт с моноклональным антителом, направленным против PD-L1, PD-L2 или PD-1. Как описано в примере 1, ингибирование пути PD-1 ведет к омоложению CD8+ T-клеток. Соответственно, происходит, например, повышение пролиферации CD8+ T-клеток и продукции цитокинов. Эти омоложенные CD8+ T-клетки вновь вводят инфицированным мышам, и количество вируса измеряют, как описано в примере 1.
Пример 5
In vitro скрининг новых омолаживающих CD8+ T-клетки соединений
Соединения, которые модулируют путь PD-1, можно идентифицировать с помощью скрининговых тестов in vivo и in vitro на основании их способности обращать истощение CD8+ T-клеток в результате хронической вирусной инфекции.
Истощенные CD8+ T-клетки получают от мышей, хронически инфицированных LCMV-C1-13, и затем вводят в контакт с тестируемым соединением. Количество противовирусных цитокинов (например, IFN-γ или TNF-α), выделенных из контактировавших T-клеток, измеряют, например, с помощью ELISA или другого количественного метода и сравнивают с количеством, если оно имеется, противовирусного цитокина, выделенного из истощенных T-клеток, не контактировавших с тестируемым соединением. Повышение количества противовирусного цитокина, выделенного обработанными клетками, по сравнению с его количеством в необработанных клетках идентифицирует соединение в качестве антагониста PD-1, пригодного для модуляции активности T-клеток.
Пример 6
In vivo скрининг новых омолаживающих CD8+ T-клетки соединений
Истощенные CD8+ T-клетки получают из мышей, хронически инфицированных LCMV-C1-13. Инфицированным мышам внутривенно вводят тестируемое соединение. Количество противовирусных цитокинов (таких как IFN-γ или TNF-α), которое выделяется в сыворотку подвергнутых и не подвергнутых лечению мышей, измеряют, например, с помощью ELISA или другого количественного метода и сравнивают. Повышение количества противовирусного цитокина, обнаруженного в сыворотке подвергнутых лечению мышей по сравнению с его количеством у не подвергнутых лечению мышей, идентифицирует тестируемое соединение как антагонист PD-1. Альтернативно до и после обработки тестируемым соединением может быть определен титр вируса (например, сывороточный титр вируса).
Пример 7
Шимпанзе в качестве модели для иммунотерапии устойчивой инфекции HCV
Шимпанзе служат моделью устойчивости HCV у человека. Дефекты в T-клеточном иммунитете, ведущие к пожизненной персистенции вируса, включают и недостаточность специфичных в отношении HCV CD4+ T-хелперных клеток, и нарушенную или измененную активность CD8+ T-эффекторных клеток. Устойчиво инфицированные шимпанзе лечат антителами против CTLA-4, PD-1 или сочетанием обоих. Определяют эффективность блокады ингибирующих путей в сочетании с вакцинацией с использованием рекомбинантных структурных и отличных от структурных белков HCV и способность таких стратегий повышать частоту встречаемости и продолжительность жизни специфичных в отношении вируса T-клеток памяти. Недостаточность T-клеточного иммунитета является исключительно специфичной в отношении HCV у устойчиво инфицированных людей и шимпанзе. Исследуют кровь и печень инфицированных шимпанзе на экспрессию CTLA-4, PD-1, BTLA и их лигандов и на наличие клеток Treg. Противовирусная активность может быть затем восстановлена путем введения шимпанзе гуманизированных моноклональных антител, которые блокируют сигнализацию через эти молекулы.
Устойчиво инфицированных шимпанзе лечат гуманизированными антителами αCTLA-4 (MDX-010, Medarex) или антителами αPD-1. Первоначальная доза MDX-010 составляет 0,3 мг/кг, затем через 2 недели 1,0 мг/кг и затем с трехнедельными интервалами 3, 10, 30 мг/кг. После лечения антителами против коингибирующих молекул необходимо определить гуморальный и клеточный иммунные ответы, а также уровень РНК HCV. Образцы собирают на 1, 2, 3, 5 и 8 неделях и затем с месячными интервалами. Образцы включают: 1) сыворотку для анализа трансаминаз, аутоантител, нейтрализующих антител против HCV и ответов цитокинов, 2) плазму на содержание вирусов и эволюцию генома, 3) PBMC для измерений in vitro иммунитета, экспрессии и функции костимулирующих/ингибирующих рецепторов, 4) свежую (не фиксированную) печень для выделения внутрипеченочных лимфоцитов и РНК и 5) фиксированную (формалин/заключенную в парафин) печень для гистологического и иммуногистохимического анализа. Собирают также региональные лимфоузлы в 2 или 3 временные точки для оценки экспрессии коингибирующих молекул и сплайсинговых вариантов с помощью иммуногистохимических и молекулярных методов. Тесты для оценки эффективности и безопасности такого лечения следует проводить, как описано в настоящем описании.
Для определения того, потенцирует ли вакцинация антигенами HCV терапевтический эффект антител против PD-1, шимпанзе лечат следующим образом: 1) внутримышечная иммунизация рекомбинантными гликопротеинами E1 и E2 оболочки (в адъюванте MF59) и другими белками (центр плюс NS 3, 4 и 5, составленными с ISCOMS) на 0, 4 и 24 неделях; 2) внутримышечная иммунизация вакциной, используемой в 1), но вводимой совместно с антителами αCTLA-4 (30 мг каждого на кг массы тела, внутривенно на 0, 4 и 24 неделях, когда вводят вакцину); 3) идентично 2) за исключением того, что вместо антител CTLA-4 используют антитела αPD-1 (или BTLA); 4) идентично группам 2 и 3 за исключением того, что в дополнение к вакцине используют сочетание антител CTLA-4 и PD-1 (или BTLA). После иммунизации на протяжении периода 1 год отслеживают специфичные в отношении HCV T- и B-клеточные ответы.
Маркеры, проверявшиеся на HCV-тетрамер+ и суммарных T-клетках в этом анализе, включают маркеры дифференцировки (например, CD25, CD69, CD38 и HLA-DR), выживания/пролиферации (например, bcl-2 и Ki67), потенциала цитотоксичности (например, гранзимы и перфорин) и рецепторы цитокинов (CD122 и CD127). Между уровнем хемокина IP-10 до лечения и ответом на ПЭГ IFN-γ/рибавирин существует интересная корреляция. Уровень IP-10 измеряют для исследования потенциальной корреляции между негативными регуляторными путями или специфичными в отношении HCV T-клеточными ответами и уровнем IP-10. Экспрессию ингибиторных рецепторов и лигандов на PBMC оценивают с помощью проточной цитометрии.
Пример 8
Иммуноокрашивание PD-1 в реактивной лимфоидной ткани
Материалы судебной практики были получены из Brigham & Women's Hospital, Boston, MA в соответствии с установленными в учреждении правилами. Все диагнозы основывались на гистологических и иммунофенотипических особенностях, описанных в системе классификации лимфом Всемирной организации здравоохранения (Jaffe ES, et al. 2001), и во всех случаях диагностический материал проверялся гематопатологом.
Иммуноокрашивание PD-1 производили на фиксированных формалином заключенных в парафин срезах ткани после восстановления антигена обработкой в микроволновой печи в 10 мМ цитратном буфере, pH 6,0 ранее описанным моноклональным антителом против PD-1 человека (2H7; 5) с использованием непрямого метода авидин-биотин-пероксидаза хрена и проявления с помощью окрашивания диаминобензидином, как описано ранее (Jones D, et al. 1999; Dorfman DM, et al. 2003). Случаи рассматривались как иммунореактивные в отношении PD-1, если по меньшей мере 25% неопластических клеток проявляли позитивную окраску. Для подтверждения специфичности окрашивания окрашивание PD-1 сравнивали с окрашиванием контрольным антителом мыши изотипа IgG, разбавленным до той же концентрации белка, во всех исследованных случаях.
Для окрашивания фиксированных в формалине заключенных в парафин образцов реактивной лимфоидной ткани, тимуса и ряда случаев В-клеточных и T-клеточных лимфопролиферативных нарушений использовали моноклональное антитело 2H7 против PD-1. В образцах миндалины, проявляющих реактивные изменения, включая фолликулярную гиперплазию, в субпопуляции преимущественно мелких лимфоцитов в центрах размножения выявлялось цитоплазматическое окрашивание PD-1 с редкими PD-1-позитивными клетками, выявляемыми в межфолликулярных T-клеточных зонах. Характер окрашивания PD-1 в центрах размножения был практически идентичен характеру окрашивания антителом против CD-3, маркера всех T-клеток, тогда как антитело против CD-20, маркера всех B-клеток, окрашивало подавляющее большинство B-клеток центров размножения. Сходные результаты наблюдались на гистологических срезах реактивных лимфатических узлов и селезенки. Окрашивание PD-1 не наблюдалось в тимусе взрослых.
Пример 9
Иммуноокрашивание PD-1 в заключенных в парафин срезах ткани при B-клеточных и T-клеточных лимфопролиферативных нарушениях
Исследовали ряд B-клеточных и T-клеточных лимфопролиферативных нарушений на предмет экспрессии PD-1; результаты суммированы в таблице 4. Обследовали сорок два случая B-клеточных лимфопролиферативных нарушений на предмет экспрессии PD-1, включая типичные случаи лимфобластного лейкоза/лимфобластной лимфомы предшественников B-клеток, а также ряд лимфопролиферативных нарушений зрелых B-клеток, включая ряд B-клеточных неходжкинских лимфом фолликулярного происхождения, включая 6 случаев фолликулярной лимфомы и 7 случаев лимфомы Беркитта. Ни при одном B-клеточном лимфопролиферативном нарушении не наблюдалось окрашивание PD-1. В некоторых случаях имелась ненеопластическая реактивная лимфоидная ткань, в которой наблюдался паттерн окрашивания PD-1, выявлявшийся в миндалине и других реактивных лимфоидных тканях, отмечавшийся выше.
Сходным образом в 25 случаях ходжкинской лимфомы, включая 11 случаев классической ходжскинской лимфомы и 14 случаев ходжкинской лимфомы с преобладанием лимфоцитов, не было продемонстрировано окрашивание PD-1 неопластических клеток. Интересно, что во всех 14 случаях ходжкинской лимфомы с преобладанием лимфоцитов T-клетки, окружающие неопластические CD20-позитивные L&H клетки, были иммунопозитивными в отношении PD-1, сходно с характером окрашивания, отмечавшимся для CD57+ T-клеток в ходжкинской лимфоме с преобладанием лимфоцитов. Эти PD-1-позитивные клетки представляли собой субпопуляцию имеющейся суммарной популяции CD3+ T-клеток.
Исследовали ряд T-клеточных лимфопролиферативных нарушений на предмет экспрессии PD-1; результаты суммированы в таблице 4. Случаи лимфобластного лейкоза/лимфобластной лимфомы предшественников T-клеток, неоплазмы незрелых T-клеток были негативными в отношении PD-1, как и случаи неоплазмы периферических, посттимусных T-клеток, включая случаи T-клеточного пролимфоцитарного лейкоза, лимфомы периферических T-клеток, неустановленной анапластической крупноклеточной лимфомы и T-клеточного лейкоза/лимфомы взрослых. Напротив, во всех 19 случаях ангиоиммунобластной лимфомы имелись очаги PD-1-позитивных клеток, которые были также иммунореактивными в отношении маркеров для всех T-клеток, таких как CD-3. PD-1-позитивные клетки постоянно обнаруживались в очагах сетей размножающихся CD21+ фолликулярных дендритных клеток (FDC), являющихся характерной особенностью ангиоиммунобластной лимфомы.
Иммуноокрашивание PD-1 при лимфопролиферативных нарушениях
*количество иммунореактивных случаев/общее количество случаев
**PD-1-позитивные клетки образуют розетки вокруг неопластических L&H клеток в 14/14 случаях.
Пример 10
Общие методы изучения экспрессии PD-1 на специфичных в отношении ВИЧ CD8+ T-клетках
Для проведения экспериментов, подробно изложенных в примерах 11-14, использовали следующие методы.
Индивидуумы: Участники исследования с инфекцией ВИЧ монофилетического таксона C были набраны из клиник для амбулаторных больных McCord Hospital, Durban, South Africa, и St. Mary's Hospital, Mariannhill, South Africa. Периферическая кровь была получена от 65 индивидуумов этой группы, причем все субъекты до момента анализа не получали противоретровирусную терапию. Индивидуумов отбирали для включения в исследование на основе экспрессии у них аллелей HLA, соответствующих десяти тетрамерам класса I, которые были сконструированы (см. ниже). Среднее количество вирусов в когорте составляло 42800 копий РНК ВИЧ-1/мл плазмы (диапазон 163-750000), а медиана абсолютного количества CD4 равнялась 362 (диапазон 129-1179). Информация, касающаяся продолжительности инфекции, была недоступна. Все индивидуумы дали письменное информированное согласие на исследование, которое было утверждено наблюдательными комиссиями местных учреждений.
Конструирование антител против PD-1 и PD-L1: Моноклональные антитела против PD-L1 (29E.2A3, IgG2b мыши) и PD-1 человека (EH12, IgGl мыши) были получены, как описано ранее, и было показано, что они блокируют взаимодействие PD-1:PD-L1.
Тетрамеры MHC класса I: В настоящем исследовании использовали десять тетрамеров ВИЧ MHC класса I, синтезированных, как описано ранее (Altman JD, et al. 1996): A*0205 GL9 (p24, GAFDLSFFL; SEQ ID NO:1), A*3002 KIY9 (интеграза, KIQNFRVYY; SEQ ID NO:2), B*0801 DI8 (p24, DIYKRWII; SEQ ID NO:3), B*0801 FL8 (Nef, FLKEKGGL; SEQ ID NO:4), B*4201 RM9 (Nef, RPQVPLRPM; SEQ ID NO:5), B*4201 TL9 (p24, TPQDLNTML; SEQ ID NO:6), B*4201 TL10 (Nef, TPGPGVRYPL; SEQ ID NO:7), B*4201 YL9 (RT, YPGIKVKQL; SEQ ID NO:8), B*8101 TL9 (p24, TPQDLNTML; SEQ ID NO:9) и Cw0304 YL9 (p24, YVDRFFKTL; SEQ ID NO:10).
Окрашивание тетрамеров HLA класса I и фенотипический анализ: Свежевыделенные мононуклеарные клетки периферической крови (PBMC, 0,5 миллиона) окрашивали на тетрамеры в течение 20 минут при 37°С. Клетки затем однократно промывали забуференным фосфатом физиологическим раствором (ЗФР), осаждали и прямо окрашивали флуоресцеинизотиоцианатом (ФИТЦ), конъюгированным с анти-CD8 (Becton Dickinson), конъюгированным с фикоэритрином анти-PD-1 (клон EH 12) и ViaProbe (Becton Dickinson). Клетки затем инкубировали в течение 20 минут при комнатной температуре, однократно промывали ЗФР и ресуспендировали в 200 мкл ЗФР с 1% параформальдегида и извлекали на активируемом флуоресценцией сортере (FACSCaliburTM, Becton Dickinson). На FACSCaliburTM получали, как минимум, 100000 событий.
Тесты пролиферации CFSE: Один миллион свежевыделенных PBMC промывали дважды в ЗФР, осаждали и ресуспендировали в 1 мл 0,5 мкМ карбоксифлуоресцеина диацетата сукцинимидилового сложного эфира (CFSE, Molecular Probes) в течение 7 минут при 37°С. Клетки промывали дважды в ЗФР, ресуспендировали в 1 мл среды R10 (RPMI 1640 с добавлением глутатиона, пенициллина, стрептомицина и 10% сыворотки эмбрионов телят (FCS)) и помещали в одну лунку 24-луночного планшета. Пилотные исследования показали, что конечная концентрация пептида 0,2 мкг/мл обеспечивает оптимальные пролиферативные ответы, поэтому в каждом тесте использовали эту конечную концентрацию пептида в лунке. Лунки отрицательного контроля содержали только PBMC в среде или PBMC в среде с очищенным анти-PD-L1 (10 мкг/мл), а лунки положительного контроля стимулировали 10 мкг/мл фитогемаглюттинина (PHA). После 6-дневной инкубации в инкубаторе при 37°С клетки промывали 2 мл ЗФР и окрашивали конъюгированными с PE тетрамерами MHC класса I, ViaProbe (Becton Dickinson) и антителами анти-CD8-APC. Клетки извлекали на FACSCalibur и анализировали с помощью программы CellQuest® (Becton Dickinson). Клетки пропускали на ViaProbe с ограничением окна для CD8+ лимфоцитов. Кратность увеличения тетрамер+ клеток рассчитывали путем деления процента клеток CD8+ тетрамер+ в присутствии пептида на процент клеток CD8+ тетрамер+ в отсутствие пептидной стимуляции.
Статистический анализ: Анализ корреляции Спирмана, тест Манна-Уитни и парный t-критерий производили с помощью GraphPad Prism Version 4.0a. Все тесты были двусторонними, и в качестве значимых считались величины p с p<0,05.
Пример 11
Экспрессия PD-1 на специфичных в отношении ВИЧ CD8+ T-клетках
Была синтезирована панель из 10 тетрамеров MHC класса I, специфичных в отношении доминирующего монофилетического таксона C вируса ВИЧ-1 эпитопов CD8+ T-клеток, на основе преобладающих аллелей HLA и частоты атакуемых эпитопов в Gag, Nef, интегразе и RT, обеспечивающая прямую визуализацию экспрессии PD-1 на поверхности этих клеток. Было проведено типирование HLA с высоким разрешением на всей группе, и для исследования была отобрана подгруппа из 65 индивидуумов, не получавших противоретровирусную терапию, на основе экспрессии подходящих аллелей HLA. Было обследовано в общей сложности 120 индивидуальных эпитопов, и на фиг.5A показано репрезентативное окрашивание ex vivo PD-1 на ВИЧ тетрамер+ клетках. Экспрессия PD-1 легко различима на этих тетрамер+ клетках и она была значительно выше, чем в общей популяции CD8 T-клеток от тех же индивидуумов (p<0,0001); в свою очередь экспрессия PD-1 и на тетрамер+ CD8+ T-клетках, и на клетках общей популяции CD8+ T-клеток была значительно выше, чем у серологически ВИЧ-отрицательных контролей (фиг.5B). Для восьми из десяти тестированных тетрамеров был выявлен, по меньшей мере, один индивидуум, у которого уровень экспрессии специфичных в отношении антигена CD8+ клеток составлял 100% (фиг.5C). PBMC от 3 до 25 индивидуумов окрашивали для каждого ответа тетрамера ВИЧ с медианными уровнями экспрессии в диапазоне от 68% до 94% тетрамер+ клеток (фиг.5C). Эти данные были дополнительно подтверждены анализом средней интенсивности флуоресценции (MFI) PD-1 и на тетрамер+ клетках, и на общей популяции CD8+ T-клеток (фиг.5B, C).
Затем было определено, имеется ли доказательство специфичных для эпитопов различий в плане уровней экспрессии PD-1 у индивидуумов с множественными выявляемыми ответами. Из 65 обследованных индивидуумов у 16 индивидуумов было от 3 до 5 тетрамер-положительных ответов у каждого. Экспрессия PD-1 была почти идентичной и приближалась к 100% для каждого проанализированного ответа для трех из шестнадцати индивидуумов; однако у остальных 13 индивидуумов были выявлены различающиеся паттерны экспрессии PD-1 в зависимости от эпитопа (фиг.5D). Эти данные указывают на то, что экспрессия PD-1 может быть дифференцированной на имеющихся одновременно специфичных в отношении эпитопов CD8+ T-клетках от одного индивидуума, что, возможно, соответствует недавним данным, указывающим на эпитоп-специфичные различия в противовирусной эффективности (Tsomides TJ, et al. 1994; Yang O, et al. 1996; Loffredo JT, et al. 2005).
Пример 12
Связь между экспрессией PD-1 и прогрессированием заболевания ВИЧ
Была определена связь между экспрессией PD-1 на специфичных в отношении ВИЧ CD8+ T-клетках и содержанием вируса в плазме и количеством CD4+ клеток, каждый из которых является прогностическим фактором прогрессирования заболевания ВИЧ. В соответствии с предшествующими исследованиями соотношение между количеством тетрамер-положительных клеток и содержанием вируса или количеством CD4+ клеток не показало сколь либо значимой корреляции (фиг.6A, B). Напротив, имелись значимые положительные корреляции с содержанием вируса и процентом и MFI экспрессии PD-1 на позитивных по тетрамеру ВИЧ клетках (p=0,0013 и p<0,0001 соответственно, фиг.6A). Имелись также обратные корреляции между показателем CD4 и процентом и MFI PD-1 на позитивных по тетрамеру ВИЧ клетках (p=0,0046 и p=0,0150 соответственно, фиг.6B). Поскольку тестированные тетрамеры, по-видимому, представляют собой лишь часть специфичной в отношении популяции CD8+ T-клеток у этих индивидуумов, исследовали также связь между экспрессией PD-1 на всех CD8+ клетках и этими параметрами. Имелись значимые положительные корреляции между содержанием вируса и процентом и MFI экспрессии PD-1 на общей популяции CD8+ T-клеток (p=0,0021 и p<0,0001 соответственно, фиг.6C), и наблюдались также обратные корреляции между показателем CD4+ клеток и процентом и MFI экспрессии PD-1 на общей популяции CD8+ T-клеток (p=0,0049 и p=0,0006, соответственно, фиг.6D). В этой же группе тестировали экспрессию PD-1 на специфичных в отношении CMV CD8+ T-клетках у 5 индивидуумов, и на этих клетках экспрессировалось значительно меньше PD-1 по сравнению со специфичными в отношении ВИЧ CD8 T-клетками (медиана 23% CMV тетрамер+ PD-1+, p=0,0036), и эта экспрессия не отличалась от экспрессии в основной части CD8+ T-клеток у этих индивидуумов, что указывает на то, что высокая экспрессия PD-1 не является одинаковой особенностью всех специфичных в отношении вируса CD8+ T-клеток. Эти данные позволяют предполагать, что повышенное количество антигена при хронической инфекции ВИЧ вызывает повышенную экспрессию PD-1 на CD8+ T-клетках, и они соответствуют данным для мышей с хронической инфекцией LCMV, у которых экспрессия PD-1 связана с функциональным истощением CD8+ T-клеток (Barber DL, et al. 2005). Более того, они дают первую четкую связь в широком исследовании, включающем анализ множества эпитопов, между специфичными в отношении ВИЧ CD8+ T-клетками и содержанием вируса или показателем CD4.
Пример 13
Связь между экспрессией PD-1 и состоянием и функцией CD8 T-клеток памяти
Экспрессию PD-1 затем анализировали в контексте ряда дополнительных фенотипических маркеров, связанных с состоянием и функцией CD8 T-клеток памяти, включая CD27, CD28, CD45RA, CD57, CD62L, CD127, CCR7, перфорин, гранзим B и Ki67 (фиг.7). Репрезентативная окраска на эти маркеры на B*4201 TL9 тетрамер+ клетках от одного индивидуума показана на фиг.7A, а обобщенные данные для 13 индивидуумов показаны на фиг.7B. Эти исследования были ограничены теми ответами тетрамеров, которые превышали 95% позитивности PD-1, поскольку мультипараметрная проточная цитометрия с более чем 4 цветами не была доступна в KwaZulu Natal. ВИЧ тетрамер+ PD-1+ клетки экспрессируют на высоком уровне CD27 и гранзим B, на очень низком уровне CD28, CCR7 и внутриклеточный Ki67, на низком уровне CD45RA и перфорин и на промежуточном уровне CD57 и CD62L (фиг.7B). Эти данные указывают на то, что специфичные в отношении ВИЧ PD-1+ T-клетки проявляют эффектор/эффекторный фенотип памяти и согласуются с предшествующими сообщениями о смещенном созревании специфичных в отношении ВИЧ CD8+ T-клеток. Кроме того, было проведено секвенирование вируса для определения того, определяют ли эти клетки ускользание от иммунной реакции. Из проверенных 45 этих тетрамер-положительных ответов только у 5 вирусные эпитопы отличались от консенсусной последовательности монофилетического таксона C Южной Африки, что указывает на то, что эти клетки испытывают слабое давление отбора in vivo.
Предшествующие эксперименты на мышах с использованием модели LCMV показали, что блокада in vivo взаимодействия PD-1/PD-L1 путем инфузии анти-PD-L1 блокирующего антитела ведет к повышению функциональности специфичных в отношении LCMV CD8+ T-клеток по результатам измерения продукции цитокинов, киллерной способности, способности к пролиферации и, что наиболее примечательно, снижения количества вируса. Краткая (12-часовая) специфичная в отношении антигена стимуляция in vitro свежевыделенных PBMC из 15 ВИЧ+ индивидуумов в присутствии или в отсутствие 1 мкг/мл очищенного анти-PD-L1 антитела не вызывала повышения продукции IFN-γ, TNF-α или ИЛ-2.
Пример 14
Влияние блокады пути PD-1/PD-L1 на пролиферацию специфичных в отношении ВИЧ CD8+ T-клеток
Поскольку специфичные в отношении ВИЧ CD8+ T-клетки также проявляют нарушенную способность к пролиферации (2004), было определено, может ли блокада PD-1/PD-L1 повышать эту функцию in vitro. Репрезентативные данные для B*4201-позитивного индивидуума показаны на фиг.8A. Инкубация свежевыделенных меченных CFSE PBMC только со средой или со средой с анти-PD-L1 антителом приводила к поддержанию популяции специфичных в отношении B*4201-TL9 CD8+ T-клеток (1,2% CD8+ T-клеток), которые оставались CFSEhi через шесть дней в культуре. Стимуляция меченных CFSE PBMC в течение 6 дней пептидом TL9 приводила к 4,8-кратной экспансии CFSElo B*4201 TL9 тетрамер+ клеток, тогда как стимуляция меченных CFSE PBMC пептидом TL9 в присутствии анти-PD-L1 блокирующего антитела дополнительно повышала пролиферацию специфичных в отношении TL9 клеток, что приводило к 10,3-кратному повышению тетрамер+ клеток. Тесты пролиферации CFSE были проведены на 28 образцах в присутствии и в отсутствие очищенного блокирующего антитела против PD-L1 человека. Наблюдалось значительное повышение пролиферации специфичных в отношении ВИЧ CD8+ T-клеток в присутствии пептида плюс блокирующее антитело против PD-L1 по сравнению с показателем пролиферации после стимуляции одним пептидом (фиг.8B; p=0,0006, парный t-тест). Кратность увеличения тетрамер+ клеток в присутствии блокирующего антитела против PD-L1 в зависимости от индивидуума и эпитопа у данного индивидуума (фиг.8C), что вновь позволяет предполагать наличие специфичных от эпитопа различий в степени функционального истощения этих ответов.
Пример 15
Терапевтическая вакцинация в сочетании с блокадой ингибиторного пути PD-1 синергично улучшает иммунологический контроль хронической вирусной инфекции: концептуальное исследование комбинаторной терапевтической вакцины
Функциональное повреждение T-клеток, включая продукцию цитокинов, цитолиз и пролиферацию специфичных в отношении антигена T-клеток, служит определяющей характерной особенностью многих хронических инфекций. Инактивированный T-клеточный иммунный ответ наблюдается во время множества разных устойчивых инфекций патогенов, включая ВИЧ, HBV, HCV и TB у человека. Инактивация T-клеток во время хронической инфекции могла бы коррелировать с величиной и устойчивостью содержания антигена и возникать из-за нарушения сигналов, проксимальных по отношению к рецептору T-клеток, увеличения ингибирующих белков или снижения костимулирующих белков и дефектов во вспомогательных или цитокиновых сигналах. Дефект в истощенных T-клетках служит первичной причиной неспособности хозяина уничтожить устойчивый патоген. Во время хронической инфекции в истощенных специфичных в отношении вируса CD8 T-клетках повышаются два ключевых ингибиторных белка: PD-1 и CTLA-4. Блокада in vivo PD-1 повышает количество и функцию специфичных в отношении вируса CD8 T-клеток и вызывает снижение количества вируса.
У современных стратегий вакцинации хронических вирусных инфекций имеется несколько недостатков. Конкретно, эффективная поддержка противовирусных ответов CD8 T-клеток не наблюдается после терапевтической вакцинации. Кроме того, высокое содержание вируса и низкий пролиферативный потенциал отвечающих T-клеток во время хронической инфекции, вероятно, ограничивают эффективность терапевтической вакцинации. Таким образом, важно разработать стратегию терапевтической вакцинации для эффективной поддержки эндогенных T-клеточных ответов хозяина для контроля хронической инфекции.
Для определения эффективности использования антагониста PD-1 в сочетании с терапевтической вакциной использовали хорошо известную модель хронической инфекции, индуцированной инфицированием клоном-13 LCMV. В качестве терапевтической вакцины использовали вирус коровьей оспы, экспрессирующий эпитоп GP33 LCMV, для мониторинга специфичного в отношении эпитопа иммунного ответа CD8 T-клеток. Терапевтическую вакцину соединяли с антителом против PD-L1 для блокирования ингибиторного пути для исследования синергичного действия в отношении пролиферации специфичных в отношении антигена CD8 T-клеток и удаления устойчивого вируса.
В этих экспериментах использовали следующие методы:
Мыши и инфекции: Мыши C57BL/6 (самки в возрасте от 4 до 6 недель) были получены из The Jackson Laboratory (Bar Harbor, ME). Мышей содержали в свободном от патогенов виварии в соответствии с правилами NIH по уходу за животными. Для инициации хронических инфекций мышей инфицировали 2×106 БОЕ клона-13 LCMV (CL-13), как описано ранее. Выращивание вируса и тесты с бляшками для определения титров вируса были описаны ранее.
In vivo блокада антителом и терапевтическая вакцинация: Двести микрограмм крысиного антитела против PD-L1 мыши (10F:9G2) вводили внутрибрюшинно на каждый третий день, начиная с 4 недель после инфицирования CL-13. В момент первого введения анти-PD-L1 внутрибрюшинно вводили 2×106 БОЕ рекомбинантного вируса коровьей оспы, экспрессирующего эпитоп GP33-41 (VV/GP33), в качестве терапевтической вакцины или вирус коровьей оспы дикого типа (VV/WT) в качестве контрольной вакцины.
Выделение лимфоцитов: лимфоциты выделяли из тканей и крови, как описано ранее. Печень и легкое перфузировали охлажденным на льду ЗФР удалением для выделения лимфоцитов.
Проточная цитометрия: пептидные тетрамеры класса I MHC получали и использовали, как описано ранее. Все антитела были получены от BD Pharmingen за исключением антител для гранзима B (Caltag), Bcl-2 (R&D Systems) и CD127 (eBioscience). Все окрашивание поверхностных и внутриклеточных цитокинов проводили, как описано (Barber et al., Nature 439:682, 2006). Для выявления дегрануляции спленоциты стимулировали в течение 5 ч в присутствии брефельдина, монензина, анти-CD107a-ФИТЦ и анти-CD107b-ФИТЦ.
Конфокальная микроскопия: из мышей выделяли селезенку и замораживали в OCT (TissueTek). Из этих блоков готовили 10-20 мм криостатные срезы и фиксировали в охлажденном на льду ацетоне в течение 10 минут. Для иммунофлуоресценции срезы окрашивали следующими антителами: ER-TR7 для выявления ретикулярных клеток (Biogenesis, Kingston, NH) и поликлональную сыворотку морской свинки против LCMV. Окраску визуализировали с помощью Alexa Fluor-488 козы против Ig крысы и Alexa Fluor-568 козы против Ig морской свинки (Molecular Probes) и анализировали с помощью конфокальной микроскопии (Leica Microsystems AG, Germany). Изображения получали с помощью ImageJ (National Institutes of Health) и Photoshop (Adobe Systems Inc.).
Результаты показали, что сочетание терапевтической вакцины с антителом против PD-L1 обеспечивает синергичный эффект на пролиферацию специфичных в отношении антигена CD8 T-клеток и удаление устойчивого вируса. Терапевтическая вакцина могла эффективно поддерживать функционально восстановленную популяцию CD8 T-клеток путем блокады ингибиторного пути PD-1/PD-L1. При сочетанной терапевтической вакцинации систематически достигались повышенная пролиферация специфичных в отношении антигена CD8 T-клеток и ускоренный контроль над вирусом (фиг.9A-9D и фиг.10A-10D). Сочетанная терапевтическая вакцина стимулирует резкое увеличение функционально активных CD8 T-клеток (фиг.11A-D). Кроме того, терапевтическая вакцина с использованием вектора, экспрессирующего специфический эпитоп, во время блокады пути PD-1/PD-L1 повышает пролиферацию CD8 T-клеток, специфичных в отношении эпитопа, кодируемого вектором (фиг.9 и 11). Повышенный уровень экспрессии CD127, наблюдаемый на специфичных в отношении антигена CD8 T-клетках в группе, лечившейся сочетанной вакциной, отражает возникновение продолжительных ответов клеток памяти, в то время как сниженные уровни экспрессии PD-1 и гранзима B коррелируют с удалением устойчивого вируса (фиг.12A-12B).
Имелся синергичный эффект терапевтической вакцины, соединенной с блокадой PD-L1, на восстановление функции «беспомощных» истощенных CD8 T-клеток (см. фиг.13A-13E). У мышей снижали количество CD4 T-клеток и затем их инфицировали клоном-13 LCMV. Некоторых мышей вакцинировали вирусом коровьей оспы дикого типа (VV/WT) или вирусом коровьей оспы, экспрессирующим эпитоп LCMV GP33-41 (VV/GP33) на 7 недель, после инфицирования. В тот же период мышей лечили αPD-L1 5 раз каждые три дня или его изотипом. Через две недели после начала лечения антителами мышей забивали для анализа. Результаты показаны на фиг.13A. Исследовали также частоту встречаемости специфичных в отношении GP33 CD8 T-клеток (фиг.13b). Спленоциты стимулировали пептидом GP33 в присутствии антител αCD107a/b и затем совместно окрашивали на IFN-γ. Показанные графики были получены с ограничением окна для CD8 T-клеток (фиг.13c). Определяли также процент IFN-γ+ клеток после стимуляции пептидом GP33 от клеток, позитивных в отношении ограниченного Db GP33-41 тетрамера (фиг.13d), а также титр вируса (фиг.13e). Результаты демонстрируют синергичный эффект вакцины, соединенной с блокадой PD-1.
Эти результаты показывают, что сочетания блокады негативного регуляторного пути и поддержания CD8 T-клеток во время хронической инфекции можно использовать в создании терапевтических вакцин для улучшения ответов T-клеток у больных с хроническими инфекциями или злокачественными новообразованиями. Терапевтические интервенции, такие как использование антагониста PD-1, которые поддерживают ответы T-клеток и снижают содержание вируса, могли бы увеличивать выживание в отсутствие болезни и снижать передачу вируса. Эффективная терапевтическая вакцинация могла бы использоваться при хронических вирусных инфекциях и устойчивых бактериальных, паразитарных инфекциях. Эта стратегия также пригодна для лечения злокачественных новообразований.
Пример 16
Усиление T-клеточной иммунотерапии посредством блокады пути PD1/PDL1
Важно разработать стратегии для лечения и уничтожения хронических вирусных инфекций, таких как вирус иммунодефицита человека и гепатит С. CDC недавно сообщила, что более одного миллиона американцев живут с ВИЧ, что служит примером потребности в более эффективной терапии. Важно определить, как ингибиторная сигнализация в отношении лимфоцитов может вносить вклад в способность патогена устойчиво ускользать от иммунного ответа хозяина.
Было показано, что ингибиторный иммунорецептор PD-1 (член семейства B7/CD28 костимулирующих рецепторов) и его лиганд (PD-L1) резко увеличиваются во время состояний хронической инфекции вирусом лимфоцитарного хориоменингита (LCMV). Дополнительные исследования с использованием модели LCMV показали, что блокада пути PD1/PDL1 значительно повышает эндогенный противовирусный ответ CD8 T-клеток во время поздних фаз хронической инфекции, когда CD8 T-клетки истощены. Истощенные T-клетки функционально неполноценны и не организуют эффективные иммунные ответы при встрече с антигеном. Однако блокада пути PD1/PDL1, видимо, обращает истощение и восстанавливает их функциональную способность. Данные позволяют предполагать, что эти эффекты хорошо сохраняются за пределами непосредственного периода лечения анти-PDL1.
Следующие эксперименты были проведены для того, чтобы (1) оценить способность анти-PDL1 повышать пролиферацию и жизнеспособность противовирусных CD8 T-клеток после адоптивного переноса иммунных (клеток памяти) спленоцитов мышам (носителям) с врожденной инфекцией, (2) оценить функциональную активность специфичных в отношении вируса CD8 T-клеток памяти, которые были размножены в условиях блокады PD1/PDL1, и (3) определить экспрессию различных маркеров дифференцировки в специфичных в отношении вируса CD8 T-клетках, которые были размножены в условиях блокады PD1/PDL1.
Роль пути PD-1 оценивали на хорошо разработанной модели цитоиммунотерапии хронической вирусной инфекции. Описанная в настоящем описании модель параллельна модели T-клеточной цитоиммунотерапии опухолей в отношении иммунологических барьеров, которые ограничивают применимость этих видов терапии (таких как искаженные или подавленные T-клеточные/противоопухолевые ответы). У мышей, инфицированных неонатально или in utero LCMV, не организуются эндогенные специфичные в отношении LCMV иммунные ответы, и на протяжении всей их жизни у них продолжают сохраняться высокие уровни инфекционного LCMV в крови и во всех тканях. Эти животные являются врожденными носителями и в значительной степени толерантны к патогену. При адоптивном переносе спленоцитов от LCMV иммунных мышей врожденному носителю перенесенные иммунные клетки памяти быстро подвергаются размножению и обеспечивают мощный иммунный ответ против вируса. Приблизительно 2/3 животных, получающих адоптивную цитоиммунотерапию, справляются с задачей полного удаления инфекции при переносе высоких доз спленоцитов.
В этих экспериментах использовали следующие материалы и методы:
Мыши и инфекции: самок мышей B57BL/6 в возрасте 4-6 недель получали из Jackson Laboratory (Bar Harbor, Maine). Острую инфекцию вызывали внутрибрюшинной инъекцией 2×105 БОЕ LCMV Армстронга. Мышей-врожденных носителей размножали в Emory University (Atlanta, GA) из колоний, берущих начало от неонатально инфицированных мышей (104 БОЕ клона-13 LCMV, внутрь мозга).
Адоптивная иммунотерапия и in vivo блокада антителом. Выделяли 40×106 целых спленоцитов от LCMV иммунных мышей (через 30-90 дней после инфицирования) и переносили их внутривенно 6-12-недельным мышам-носителям LCMV. В течение 15 дней после адоптивной иммунотерапии на каждый 3-й день вводили 200 микрограмм антитела крысы против PD-L1 мыши (10F.9G2).
Проточная цитометрия и окрашивание тетрамера. Тетрамеры H-2Db класса I MHC, комплексированные с GP33-41 LCMV, получали, как описано ранее. Все антитела были получены от BD/Pharmingen (San Diego, CA). Мононуклеарные клетки периферической крови и спленоциты выделяли и окрашивали, как описано ранее. Данные получали с помощью проточного цитометра FACSCaliburTM (BD) и анализировали с использованием программы FlowJoe (Tree Star Inc. Ashland, OR).
Окрашивание внутриклеточных цитокинов: Для окрашивания внутриклеточных цитокинов 106 спленоцитов культивировали в присутствии или в отсутствие указанного пептида (0,2 мкг/мл) и брефельдина A в течение 5-6 часов при 37°С. После окрашивания на маркеры поверхности клетки пермеабилизовывали и окрашивали на внутриклеточные цитокины с использованием препарата Cytofix/Cytoperm (BD/Pharmigen).
Были получены следующие результаты:
Терапия антителом против PD-L1 повышает количество специфичных в отношении вируса CD8 T-клеток: Мононуклеарные клетки периферической крови (PBMC) выделяли из лечившихся и не лечившихся животных на 7, 11, 15, 22 и 35 дни. Клетки, специфичные в отношении эпитопа Db GP33, определяли окрашиванием тетрамером. В двух независимых экспериментах было обнаружено, что у животных, получавших терапию антителом против PD-L1 в течение первых 15 дней после адоптивного переноса, появлялось значительно большее количество специфичных в отношении LCMV CD8 T-клеток после нормирования к числу Db GP33-позитивных клеток на миллион PBMC (фиг.14). Эти данные подтверждают роль пути PD-1/PD-L1 в обеспечении определенной части подавления пролиферации T-клеток памяти. Более того, эти результаты позволяют предполагать, что терапевтическое ингибирование этого пути могло бы усилить развитие и поддержание вторичного иммунного ответа, возникающего после адоптивного переноса в условия хронической инфекции с высоким содержанием антигена.
Блокада PD-1/PD-L1 повышает функциональность специфичных в отношении антигена CD8 T-клеток: Спленоциты выделяли из лечившихся и не лечившихся животных на 17 день после адоптивного переноса и анализировали на предмет экспрессии воспалительных цитокинов (IFN-гамма и TNF-альфа) или CD107ab (ассоциированный с лизосомами мембранный белок, LAMP). Было обнаружено, что экспрессия IFN-гамма увеличивается у животных, получавших блокирующее антитело против PD-L1, по сравнению с животными без лечения при всех определенных эпитопах CD8 (фиг.15a). Кроме того, после терапии антитело против PD-L1 возрастала также совместная экспрессия IFN-гамма и TNF-альфа (фиг.15B-15E). Эти данные указывают на то, что адоптивно перенесенные спленоциты памяти, размножающиеся в условиях блокады PD-L1, функционально превосходны в плане продукции воспалительных цитокинов и высвобождения цитолитических гранул по сравнению со спленоцитами от животных, не получавших лечение.
Пример 17
Ответы мышиных B-клеток во время блокады PD-1
Для определения того, повышает ли блокада PD-1 ответы B-клеток во время хронической инфекции LCMV, были проведены следующие эксперименты. Ответы и B-клеток, и T-клеток важны для контроля хронической инфекции LCMV, поэтому улучшение ответов B-клеток у хронически инфицированных LCMV мышей может способствовать снижению содержания вируса и повышению функции T-клеток.
В этих экспериментах использовали следующие материалы и методы:
Мыши и вирус: самок мышей B57Bl/6 в возрасте от четырех до шести недель получали из Jackson Laboratory (Bar Harbor, Maine). Перед инфицированием у мышей с хроническим LCMV снижали CD4 T-клетки введением антитела gk1.5. Предшествующие данные показывают, что введение 500 мкг gk1.5 в дни -2 и 0 перед введением вируса ведет к снижению количества CD4 T-клеток в селезенке и лимфатических узлах на 95-99% с медленным восстановлением количества CD4 T-клеток на протяжении от 2 до 4 недель. Мыши получали 2×106 БОЕ штамма клон-13 LCMV внутривенно в день 0 для инициации хронической инфекции. Титры вируса определяли с помощью 6-дневного бляшкового теста на клетках Vero.
Определение ASC с помощью ALISPOT: из суспензий одиночных клеток селезенки и костного мозга удаляли эритроциты обработкой 0,84% NH4Cl и ресуспендированием в RPMI с добавкой 5% FCS. Секретирующие антитела клетки выявляли помещением клеток в 96-луночные планшеты для фильтрации нитроцеллюлозным дном Multiscreen HA (Millipore). Планшеты предварительно покрывали 100 мкл 5 мкг/мл антителами козы против IgG+IgM+IgA мыши (Caltag/Invitrogen) в течение ночи при 4°C. Планшеты затем промывали 3×ЗФР/0,2% твин, затем 1×ЗФР и блокировали в течение 2 часов RPMI+10% FCS для предотвращения неспецифического связывания. Блокирующую среду заменяли 100 мкл RPMI 5% FCS, и в планшет добавляли 50 мкл последовательных трехкратных разведений 1×107 клеток/мл. Планшеты инкубировали в течение 6 часов при 37°С и 5% CO2. Клетки удаляли, и планшеты промывали 3× ЗФР и 3× ЗФР/0,2% твин. Лунки затем покрывали биотинилированными козьими антителами против IgG мыши (Caltag/Invitrogen), разбавленными 1/1000 в ЗФР/0,2% твин/1% FCS и инкубировали в течение ночи при 4°С. Второе антитело удаляли, и планшеты промывали 3×ЗФР/0,2% твин. Инкубировали с авидин-D-HRP (Vector), разбавленным 1/1000 в ЗФР/0,2% твин/1% FCS, в течение одного часа при комн. темп. Планшеты промывали 3×ЗФР/0,2% твин и 3×ЗФР, и проявление проводили добавлением 100 мкл хромогенного субстрата пероксидазы хрена-H2O2. Субстрат получали добавлением 150 мкл свежеприготовленного раствора AEC (10 мг 3-амино-9-этилкарбазол (ICN) на мл, растворенный в диметилформамиде (Sigma)) к 10 мл 0,1 М натрий-ацетатного буфера pH 4,8, фильтрацией этого через мембрану с размером пор 0,2 мм и непосредственно перед использованием добавлением 150 мл 3% H2O2. Через 3-5 минут появлялись зернистые красные пятна, и реакцию прекращали промывкой водопроводной водой. Пятна пересчитывали с помощью стереомикроскопа, снабженного вертикальным белым освещением.
Определение суммарных клеток костного мозга: Для расчета суммарного ответа ASC в костном мозге ответ клеток костного мозга двух бедер умножали на коэффициент 7,9, поскольку исследования распределения 59Fe показали, что 12,6% всего костного мозга мыши локализовано в двух объединенных бедрах. Не было выявлено различий в ASC активности клеток костного мозга между бедром, большеберцовой костью, плечевой костью, ребром или грудиной. Обычно из двух бедер получают от 2×107 до 2,5×107 суммарных клеток костного мозга.
Проточная цитометрия: Прямо конъюгированные антитела были получены от Pharmingen (анти-B220, анти-CD4, анти-CD138, анти-CD95, анти-Ki67, анти-IgD биотинилированные) или Vector labs (PNA). Стрептавидин-APC был получен от Molecular Probes. Все окрашивание проводили при 4°С в ЗФР с добавлением 1% FCS и 0,1% азида натрия. Клетки затем фиксировали в 2% формальдегиде (в ЗФР) и анализировали на FACS Calibur с использованием программы CellQuest (BD Biosciences).
Статистический анализ: Тесты проводили с использованием Prism 4.0 (GraphPad, San Diego, CA). Статистический анализ осуществляли с использованием двустороннего непарного T-теста с 95% доверительным интервалом.
Суммарное количество секретирующих антитело клеток в селезенке возрастает после in vivo блокады PD-1: мышей, инфицированных клоном-13 LCMV, лечили антителом (α) против PD-L1 приблизительно в течение 60 дней после инфицирования. Мышам вводили 200 мкг αPD-L1 на каждый третий день в течение двух недель. На 14 день лечения αPD-L1 мышей забивали, и количество секретирующих антитело клеток в селезенке измеряли с помощью ELISPOT и окрашивания с проточной цитометрией. В трех отдельных экспериментах у мышей, лечившихся αPD-L1, были показаны значительно увеличенные уровни секретирующих антитело клеток (ASC) в селезенке (p=0,011) по сравнению с мышами без лечения (фиг.16a). ASC могут дифференцироваться из B-клеток в селезенке путем подавления маркера B-клеток B220 и экспрессии CD138 (синдекам-1). В соответствии с результатами ELISPOT у инфицированных мышей, лечившихся αPD-L1, наблюдалось повышенное количество B220low/int CD138+ клеток (фиг.16b).
Лечение хронически инфицированных LCMV мышей αLD-L1 не ведет к повышенному уровню ASC костного мозга. Определяли, увеличиваются ли также секретирующие антитело клетки в костном мозге по время лечения αPD-L1. Большинство долгоживущих плазматических клеток находится в костном мозге, и эти плазматические клетки важны для долговременного поддержания уровня антитела в сыворотке. Хронически инфицированных LCMV мышей лечили αPD-L1 приблизительно в течение 60 дней после инфицирования. На 14 день лечения αPD-L1 измеряли уровни ASC в селезенке и костном мозге с помощью ELISPOT. Хотя через две недели после лечения в селезенке имелось повышенное количество ASC, изменений в количестве ASC в костном мозге в этот момент времени не было (фиг.17).
Совместное лечение хронически инфицированных LCMV мышей αPD-L1 и 118αCTLA-4 ведет к синергичному повышению уровня ASC в селезенке. Далее исследовали, будет ли блокада сигнализации другой молекулы негативной регуляции, CTLA-4, повышать эффект, наблюдаемый при блокаде PD-1. Полагают, что связывание CTLA-4 с B7 и конкурирует с позитивной костимулирующей молекулой CD28, и/или дает прямо сигналы, противодействующие TCR. Мышей, инфицированных клоном-13 LCMV, лечили, либо αPD-L1, либо αCTLA-4, либо обоими, или оставляли без лечения, и через две недели лечения измеряли уровни секретирующих антитело клеток с помощью ELISPOT. Хотя лечение αCTLA-4 не оказало выявляемого влияния на уровень ASC, совместное лечение αPD-L1 и αCTLA-4 вело к синергичному увеличению ASC по сравнению с увеличением, наблюдаемым при изолированном лечении αPD-L1 (фиг.18).
Повышенные пролиферация B-клеток и CD4 T-клеток и активность центров размножения у лечившихся αPD-L1 мышей: Анализ проточной цитометрией популяций селезенки у хронических мышей, лечившихся αPD-L1, показал повышенные уровни пролиферации по усиленному окрашиванию Ki-67, и CD4 T-клеток, и B-клеток. B-клетки в активной зоне центра размножения можно идентифицировать в селезенке по высокому уровню окрашивания PNA и FAS. После лечения αPD-L1 имелось значительное увеличение частоты встречаемости PNA+FAS+ B-клеток по сравнению с контролем без лечения (фиг.19a-19b).
Пример 17
Экспрессия PD-1 на T-клетках человека
CD8 T-клетки существенны для контроля многих хронических инфекций. Как раскрыто в настоящем описании, эти CD8 T-клетки становятся истощенными после хронической антигенной стимуляции, что характеризуется индукцией гипопролиферативного состояния и утратой способности продуцировать противовирусные цитокины. Истощенные Т-клетки имеют высокую экспрессию белка 1 программированной смерти (PD-1), и PD-1 также стимулируется активацией T-клеток и может быть включен лигандами PD-1, PD-L1 и PD-L2. В настоящем описании раскрыто, что ингибиторный путь PD-1 служит важным посредником истощения CD8 T-клеток во время хронической вирусной инфекции у мышей. Специфичные в отношении вируса CD8 T-клетки поддерживали высокий уровень экспрессии PD-1 в ответ на хроническую инфекцию, но не в ответ на инфекцию, которая успешно ликвидируется. Блокада взаимодействия PD-1/PD-L1 приводила к повышенной пролиферации T-клеток, продукции противовирусных цитокинов и снижению содержания вируса.
Была произведена оценка того, экспрессируют ли специфичные в отношении хронических инфекций CD8 T-клетки человека PD-1 и повышает ли блокада PD-1 ответы CD8 T-клеток. В этом исследовании (1) определен паттерн экспрессии PD-1 на субпопуляциях мононуклеарных клеток периферической крови человека (PBMC): CD4, CD8, B-клетках, NK, моноцитах, DC; (2) определен фенотип CD4 и CD8 T-клеток, которые экспрессируют PD-1; (3) определена экспрессия PD-1 на специфичных в отношении хронически сохраняющегося антигена [вируса Эпштейна-Барр (EBV) и цитомегаловируса (CMV)] и быстро удаляемого антигена (грипп и коровья оспа); и (4) определен эффект блокады взаимодействия PD-1/PD-L1 на пролиферацию специфичных в отношении антигена клеток.
В этих исследованиях использовали следующие материалы и методы:
Образцы крови: Образцы периферической крови были получены от 36 здоровых индивидуумов, которые были серопозитивными в отношении вирусов EBV, CMV, гриппа или коровьей оспы. Эти индивидуумы были отобраны на основе экспрессии их аллеля HLA, соответствующего тетрамерам класса I HLA, специфичных в отношении белков вирусов EBV, CMV, гриппа или коровьей оспы. PBMC выделяли из образцов крови на среде отделения лимфоцитов (Cellgro, Herndon, VA).
Антитела, пептиды и тетрамеры: Были получены конъюгированное с фикоэритрином антитело против PD-1 человека (EH12, IgG1 мыши) и неконъюгированное антитело против PD-L1 человека (29E.2A3, IgG2b мыши). Прямо конъюгированные антитела были получены от Beckman Coulter, San Diego, CA (анти-CD3, CD11a, CD27, CD28, CD38, CD45RA, CD57, CD62L и гранзима-B), BD Pharmingen, San Diego, CA (CD8, CD95, CD195, HLA-DR, Ki-67 и перфорина) и R&D systems, Minneapolis, MA (CCR7). Пептиды были получены в лаб. пептидного синтеза в Emory University, Atlanta, GA. Плазмидные конструкты, экспрессирующие HLA-A2, -B7 и -B8, были любезно предоставлены Tetramer Core Facility, Atlanta, GA, а меченные APC пептидные тетрамеры класса I MHC, несущие эпитопы CTL EBV (HLA-A2-GLCTLVAML (SEQ ID NO:36), HLA-B8-RAKFKQLL (SEQ ID NO:37) и FLRGRAYGL (SEQ ID NO:38)), CMV (HLA-A2-NLVPMVATV (SEQ ID NO:39), HLA-B7-TPRVTGGGAM (SEQ ID NO:40)), гриппа (HLA-A2-GILGFVFTL (SEQ ID NO:41)) и коровьей оспы (HLA-A2-CLTEYILWV (SEQ ID NO:42) и KVDDTFYYV (SEQ ID NO:43)).
Иммунофенотипирование и пролиферация CFSE: Образцы цельной гепаринизированной крови человека (200 мкл) окрашивали антителами или тетрамерами и затем анализировали (Ibegbu et al., J Immunol. 174:6088-6094, 2005) на FACS Calibur с использованием программы CellQuest или на проточном цитометре LSRII с использованием программы FACSDiva (BD Immunocytometry Systems). Для анализов CFSE PBMC (2x106/мл) тщательно промывали и метили 3 мкМ карбоксифлуоресцеина диацетата сукцинимидильным эфиром (CFSE, Molecular Probes) при комнатной температуре в темноте в течение 5 мин (см., например, Weston and Parish, J Immunol. Methods 133:87-97, 1990). Меченные CFSE PBMC стимулировали либо одним пептидом (1 мкг/мл), либо пептидом с антителом против PD-L1 (10 мкг/мл). Контрольные культуры содержали либо только PBMC, PBMC с антителом против PD-L1 или PBMC с контрольным по изотипу антителом (IgG2b, 10 мкг/мл). После 6-дневной инкубации при 37°С клетки промывали и окрашивали внеклеточно на тетрамеры, наряду с антителами против CD3 и CD8.
Были получены следующие результаты:
Паттерн экспрессии PD-1 на субпопуляциях PBMC: Экспрессию PD-1 исследовали на субпопуляциях PBMC здоровых индивидуумов. Было обнаружено, что CD8+ T-клетки, CD4+ T-клетки и моноциты (CD14+) экспрессируют PD-1 на высоком уровне, B-клетки (CD20+) экспрессируют PD-1 на низком уровне, а клетки NK (CD56+) и DC (CDl1c+) не экспрессируют PD-1.
PD-1 преимущественно экспрессируется эффекторными T-клетками памяти CD8 и CD4: CD8 T-клетки от нормальных здоровых индивидуумов исследовали на предмет совместной экспрессии PD-1 и различных фенотипических маркеров, связанных с состоянием дифференцировки и функцией (фиг.20A). В целом наивные и имеющие фенотип центральной памяти CD8 T-клетки экспрессируют PD-1 лишь на низком уровне, в то время как CD8 T-клетки, которые экспрессируют различные маркеры, связанные с эффекторным/эффекторной памяти/или истощения фенотипом, также экспрессируют PD-1 на высоком уровне (фиг.20B). Эти данные позволяют предполагать, что PD-1 преимущественно экспрессируется CD8 T-клетками эффекторной памяти. Сходная тенденция была обнаружена при исследовании заявителями CD4 T-клеток (фиг.20C).
PD-1 увеличивается на устойчивых специфичных в отношении антигена CD8 T-клетках памяти: Для оценки того, проявляют ли специфичные в отношении хронических инфекций CD8 T-клетки повышенную экспрессию PD-1, было произведено сравнение экспрессии PD-1 на CD8 T-клетках памяти, специфичных в отношении хронически устойчивых вирусов (EBV и CMV), и специфичных в отношении острой вирусной инфекции (гриппа и коровьей оспы) T-клетках у 36 здоровых индивидуумов путем окрашивания на специфичные в отношении вирусов EBV, CMV, гриппа и коровьей оспы тетрамеры (фиг.21A-21B). На фиг.21A представлены репрезентативные PD-1 GMF1 CD8 T-клеток, специфичных в отношении вирусов EBV, CMV, гриппа и коровьей оспы. Было обнаружено, что экспрессия PD-1 на специфичных в отношении EBV CD8 T-клетках повышена по сравнению со специфичными в отношении вирусов гриппа (p=0,0335) и коровьей оспы (p=0,0036) CD8 T-клетками (фиг.21A-21B). Сходным образом, специфичные в отношении CMV CD8 T-клетки экспрессировали PD-1 чаще, чем клетки, специфичные в отношении вирусов гриппа (p=0,0431) и коровьей оспы (p=0,019), (фиг.21A-21B). Эти результаты позволяют предполагать наличие корреляции между экспрессией PD-1 и воздействием антигена.
Блокада анти-PD-L1 повышает пролиферацию хронически устойчивых специфичных в отношении вируса CD8 T-клеток: была произведена оценка того, повышает ли блокада PD-1 ответы устойчивых специфичных в отношении вируса CD8 T-клеток сходно с результатами, наблюдавшимися на мышах. Меченные CFSE клетки стимулировали каждым из пептидов, специфичных в отношении вирусов EBV, CMV, гриппа и коровьей оспы, в присутствии или в отсутствие антител против PD-L1. Через 6 дней сравнивали процент клеток тетрамер+ CFSElo и клеток CD8+ CFSElo в культурах, которые стимулировали только пептидом, и культурах, которые стимулировали пептидом и затем блокировали анти-PD-L1. На фиг.22A показаны репрезентативные графики проточной цитометрии с пролиферацией специфичных в отношении CMV и EBV CD8 T-клеток. Объединенные данные для CMV - (n=5), EBV - (n=6), грипп - (n=2) и коровья оспа - (n=2) серопозитивных индивидуумов показаны на фиг.22B. Блокада взаимодействия PD-1/PD-L1 антителом против PD-L1 приводила к повышению пролиферации специфичных в отношении EBV и CMV CD8 T-клеток, в то время как специфичные в отношении вирусов гриппа и коровьей оспы CD8 T-клетки не проявляли пролиферации после блокады анти-PD-L1. Эти результаты показывают, что в присутствии пептида плюс блокирующее антитело против PD-L1 происходит 3,5-кратное повышение частоты встречаемости специфичных в отношении EBV и CMV CD8 T-клеток по сравнению со стимуляцией одним пептидом. Была произведена оценка того, связана ли пролиферация специфичных в отношении антигена CD8 T-клеток после блокады антителом против PD-L1 с экспрессией PD-1 на этих клетках. Данные показывают положительную корреляцию между экспрессией PD-1 и пролиферацией специфичных в отношении антигена CD8 T-клеток (p=0,0083) (фиг.22C).
Пример 18
Инфильтрованные в печень лимфоциты при хронической инфекции HCV человека проявляют истощенный фенотип с высокой экспрессией PD-1 и низкой экспрессией CD127
Описанные ниже эксперименты документируют, что при хронической инфекции HCV периферические специфичные в отношении HCV T-клетки экспрессируют PD-1 на высоком уровне и что блокада взаимодействия PD-1/PD-L1 ведет к повышению способности к пролиферации. Важно, что внутрипеченочные специфичные в отношении HCV T-клетки не только экспрессируют PD-1 на высоком уровне, но и экспрессируют пониженное количество рецептора альфа ИЛ-7 (CD127) - фенотип истощения, который был специфичен для антигена HCV и локализовался в печени, являющейся местом репликации вируса.
В настоящее время не существует вакцины для профилактики инфекции HCV, а единственная лицензированная терапия, альфа-интерферон (IFNα), отдельно или в сочетании с нуклеозидным аналогом рибавирином, дорогостоящая, связана в лучшем случае лишь с 50% скоростью клиренса для самого преобладающего генотипа (генотипа 1) и осложняется значительными побочными эффектами. Недостаточность эффективных вариантов терапии против HCV подчеркивает необходимость эффективных вмешательств, направленных на усиление или дополнение естественного иммунного ответа, которые сами по себе или в сочетании с лекарственной противовирусной терапией могут предотвращать пагубные последствия инфекции HCV.
В настоящее время мало известно об экспрессии PD-1 и его роли в истощении T-клеток при хронической инфекции HCV, особенно в месте активной инфекции, печени. Настоящее исследование было предпринято для лучшего понимания фенотипа T-клеток при инфекции HCV путем измерения экспрессии PD-1 на специфичных в отношении антигена CD8+ T-клетках и в печени, и в периферической крови больных с хронической инфекцией HCV.
В этих исследованиях использовали следующие материалы и методы:
Индивидуумы: В исследование было включено семнадцать больных с хронической инфекцией HCV (позитивных по антителам HCV и ПЦР HCV), негативных по ВИЧ по скринингу антителом. До включения в исследование все больные не получали противовирусной терапии HCV. Семь из пятнадцати больных были позитивными по HLA-A2 по данным анализа FACS. Характеристики больных суммированы в таблице 5.
Демографические и клинические данные группы больных
Тестирование антител HCV, определение содержания вируса и генотипирование: Тестирование антител HCV с помощью ELISA проводили с использованием инструкций производителя набора (Abbott Diagnostics, Abbott Park, Ill; Bio-Rad Laboratories, Hercules, CA). Количественное определение содержания вируса HCV производили с помощью анализа ОТ-ПЦР в реальном времени (Roche Molecular Systems, Alameda CA). Генотипирование HCV производили с помощью анализа ОТ-ПЦР в реальном времени (Abbott Diagnostics, Abbott Park, Ill) и с помощью анализа зондовыми полосками (LIPA) (Bayer Diagnostics, Research Triangle Park, NC).
Мононуклеарные клетки периферической крови: У каждого больного собирали кровь (50-70 мл) с антикоагулянтами ЭДТА и гепарином и использовали либо прямо для окрашивания FACS, либо для выделения PBMC. PBMC выделяли с помощью градиента плотности Ficoll-Paque PLUS (Amersham, Oslo, Norway), промывали дважды ЗФР и либо анализировали немедленно, либо хранили замороженными в среде, содержащей 90% сыворотки эмбрионов телят (Hyclone) и 10% диметилсульфоксида (Sigma-Aldrich, St. Louis, MO).
Биопсия печени: Ткань печени получали либо игольчатой биопсией под ультразвуковым контролем, либо посредством флуороскопического метода через яремную вену и немедленно помещали в среду RPMI-1640 (Gibco), содержащую 10% сыворотку эмбрионов телят (Hyclone, Logan, UT) для иммунологических анализов. Другой фрагмент фиксировали в формалине для гистологического исследования.
Выделение внутрипеченочных T-клеток: образец печеночного биоптата, полученный в среде RPMI-1640 (Gibco, Carlsbad, CA), содержащей 10% сыворотку эмбрионов телят (Hyclone, Logan, UT), промывали три раза этой же средой для удаления обломков клеток и RBC. Выделение инфильтрованных в печень лимфоцитов проводили с использованием автоматизированной механической системы дезагрегации (Medimachine, Becton Dickinson, San Jose, CA). Образец вставляли в 50 мкм Medicon и вставляли в Medimachine и обрабатывали в течение 15 секунд. Дезагрегированные клетки удаляли с помощью шприца через порт для шприца. Medicon промывали дважды средой RPMI (Gibco, Carlsbad, CA), содержащей 10% сыворотку эмбрионов телят (Hyclone, Logan, UT), для обеспечения максимального выхода клеток. Клетки немедленно использовали для окрашивания FACS.
Антитела, тетрамеры HLA-A2 и проточная цитометрия: Клетки окрашивали меченными ФИТЦ, PE, PerCP и APC промечивали моноклональными антителами или тетрамерами в соответствии с инструкциями производителя и осуществляли проточную цитометрию с помощью FACS Calibur (Becton Dickinson, San Jose, CA). Данные FACS анализировали с помощью программы FlowJo (Treestar). Использовали следующие моноклональные антитела от BD Pharmingen (BD Biosciences, San Jose, CA): анти-CD8 PerCP и анти-CD45RA APC. Анти-CD62L ФИТЦ, CD3 ФИТЦ и CD127 PE были получены от Beckman Coulter (Fullerton, CA). Анти-PD-1 PE конъюгированное антитело (клон EH12) было получено, как описано (Dorfman et al., Am. J. Surg. Pathol. 30:802-810, 2006). HLA-A2 тетрамеры были специфичны для следующих эпитопов CD8+ T-клеток: HCV 1073: CINGVCWTV (SEQ ID NO:44); HCV-1406: KLVALGINAV (SEQ ID NO:45). Сбор проточной цитометрией производили на FACSCaliberTM (BD Biosciences, San Jose, CA), а анализ осуществляли с помощью программы FlowJo (v8.1.1).
Промечивание CFSE и блокада антителом: 10×06 PBMC промывали ЗФР и метили 3 мкМ CFSE (Molecular Probes). Концентрацию клеток доводили до 1×106 клеток/мл и культивировали в присутствии 2 мкг/мл пептида A2-HCV 1073 (CINGVCWTV, SEQ ID NO:44). На 3 день после стимуляции добавляли 10 Е/мл ИЛ-2. В каждый тест включали нестимулированный контроль. Специфические блокирующие антитела (анти-PD-Ll; клон # 29E и анти-PD-1; клон # EH12 (Dofman et al., выше)) добавляли к клеточным культурам в концентрации 10 мкг/мл в момент стимуляции. Клетки инкубировали в течение 6 дней, собирали и окрашивали антителами поверхности и тетрамерами и анализировали проточной цитометрией.
Статистический анализ: результаты представляли в виде графиков и анализировали с помощью GraphPad Prism (v4). Внутренние сравнения для одного и того же больного проводили с использованием парных t-тестов. Сравнения между больными проводили с использованием непарных t-тестов.
Были получены следующие результаты:
Экспрессия PD-1 на специфичных в отношении антигена HCV CD8+ T-клетках: Исследовали семнадцать больных с инфекцией HCV (все ВИЧ-негативные) (таблица 5). У пятнадцати больных отбирали образцы и крови, и печени для фенотипирования с помощью анализа проточной цитометрией, и все больные до включения в исследование не подвергались лечению с использованием фармакологической противовирусной терапии. Семь больных из группы были позитивными по HLA-A2 с выявляемой популяцией специфичных в отношении HCV CD8+ T-клеток на периферии по окрашиванию на HLA тетрамеры (таблица 5). Эти специфичные в отношении HCV CD8+ T-клетки анализировали на предмет экспрессии PD-1 (фиг.23A). Уровень экспрессии PD-1 на суммарных CD8+ T-клетках периферической крови от здоровых доноров не отличался значительно от уровня в общем пуле периферических CD8+ T-клеток от инфицированных HCV больных (фиг.23B). Напротив, большинство специфичных в отношении HCV тетрамер-позитивных CD8+ T-клеток, отобранных из периферической крови, были PD-1-позитивными (среднее 85%, SEM, стандартная ошибка средней, 3,6) (фиг.23A) при значительно более высокой экспрессии по сравнению с общей популяцией CD8+ T-клеток (p<0,0001) (фиг.23B). Также исследовали экспрессию молекул дифференцировки, костимуляции, передвижения и эффекторной функции на специфичных в отношении антигена CD8+ T-клетках. Специфичные в отношении HCV тетрамер-позитивные клетки проявляют фенотип памяти (высокий CD11a, низкий CD45RA), маркеры ранней дифференцировки (высокий CD27, высокий CD28, промежуточная экспрессия CCR7 и CD62L) и низкие уровни посредников эффекторной функции гранзима B и перфорина. Интересно, что эти HCV тетрамер-позитивные T-клетки периферической крови экспрессировали на высоком уровне CD127 (α-цепь рецептора ИЛ-7), фенотипического маркера, экспрессия которого на низком уровне указывает на нарушенную дифференцировку T-клеток памяти.
Для определения того, отличается ли фенотип CD8+ T-клеток при протекании нехронической инфекции, Flu-специфичные T-клетки исследовали у пяти здоровых HLA-A2+ доноров, которые не были инфицированы HCV. Процент периферических Flu тетрамер+ CD8+ T-клеток, которые экспрессируют PD-1, равнялся 49% (SEM 14,1) (фиг.23C). С помощью тетрамерного анализа у пяти из семи HLA-A2-позитивных хронических больных HCV были выявлены Flu-специфичные CD8+ T-клетки. Процент Flu-специфичных T-клеток, экспрессирующих PD-1, у этих хронически инфицированных HCV больных не отличался значительно от той же популяции у здоровых доноров (фиг.23C). Важно, что поскольку у пяти из семи HLA-A2 больных HCV также имелись выявляемые Flu-специфичные CD8+ T-клетки, можно было провести внутреннее сравнение для каждого больного PD-1 для T-клеток, специфичных для нехронической (Flu) и хронической (HCV) инфекции. Разница между экспрессией PD-1 Flu-специфичными и HCV-специфичными T-клетками была значительной (фиг.23C). Процент специфичных в отношении HCV CD8+ T-клеток, экспрессирующих PD-1 (среднее 83%, SEM 6,4), был выше, чем процент PD-1+ специфичных в отношении Flu CD8+ T-клеток (49%, SEM 12,3) (p=0,048) (фиг.23C).
Экспрессия PD-1 на лимфоцитах периферической крови и лимфоцитах, инфильтрованных в печень человека: Периферическую кровь и биоптаты печени от пятнадцати больных, хронически инфицированных HCV, анализировали на экспрессию PD-1. Репрезентативный анализ проточной цитометрией у пяти больных показан на фиг.24A. В то время как в периферической крови 27% (SEM 3,4) CD8+ T-клеток были PD-1+, в печени частота таких клеток была увеличена вдвое (57%, SEM 3,6) (фиг.24B). Таким образом, печень обогащена клетками, экспрессирующими PD-1 на высоком уровне. В то время как наивные клетки должны экспрессировать на высоком уровне и CD62L, и CD45RA, в печени большинство CD8+ T-клеток было CD62L низкий/CD45RA низкий, что соответствует фенотипу памяти (фиг.24C). Специальный анализ этой популяции памяти и в печени, и на периферии показал, что экспрессия PD-1 в печени была увеличена по сравнению с периферией (фиг.24C). Эти данные позволяют предполагать, что увеличение процента клеток, экспрессирующих PD-1 на внутрипеченочных T-клетках, обусловлено не только отсутствием наивной популяции в этом компартменте. Вместо этого имеется преимущественное обогащение печени PD-1+CD8+ T-клетками эффекторной памяти (CD62L низкий/CD45RA низкий) по сравнению с периферической кровью (фиг.23C).
Экспрессия CD127 на лимфоцитах периферической крови и лимфоцитах, инфильтрованных в печень человека: ИЛ-7 необходим для поддержания CD8+ T-клеток памяти (Kaech et al., Nat Immunol. 4:1191-8, 2003), и альфа-цепь его рецептора, CD127, снижается на специфичных в отношении антигена T-клетках при устойчивых инфекциях LCMV и вируса герпеса гамма (см., например, Fuller et al., J. Immunol 174:5926-30, 2005). Эта утрата CD127 во время хронической инфекции коррелирует с нарушенной продукцией цитокинов, повышенной подверженности апоптозу и снижением способности специфичных в отношении вируса CD8+ T-клеток памяти сохраняться у хозяина. Соответственно, избавление от острой инфекции вируса гепатита B (HBV) коррелирует с повышением экспрессии CD127 и одновременной утратой экспрессии PD-1 (Boettler et al., J. Virol 80:3532-40, 2006). Интересно, что у больных хроническим HCV лишь 20% (SEM 4,8) суммарных периферических CD8+ T-клеток было CD127-негативным, но в печеночных CD8+ T-клеточных инфильтратах этот процент был значительно увеличен до 58% (SEM 4,4) (фиг.24D). Таким образом, печень обогащена клетками, экспрессирующими истощенный фенотип с преобладанием клеток с высоким PD-1 и низким CD127. Эти данные позволяют предполагать, что инфильтрация в печень CD8+ T-клеток у больных хроническим HCV не отражает фенотипически популяцию периферических CD8+ T-клеток. При протекании ВИЧ инфекции, когда вирус инфицирует T-клетки и моноциты в периферической крови, низкий уровень CD127 ассоциирован с дефектами функциональных T-клеток или T-клеток памяти (Boutboul et al., Aids 19:1981-6, 2005). В этом исследовании компартментализация клеток в печени, указывающая на их истощенный фенотип, позволяет предполагать, что фенотип тесно связан с местом устойчивой репликации вируса.
Экспрессия PD-1 и CD127 на специфичных в отношении антигена HCV CD8+ T-клетках в печени: у двоих исследованных больных с HLA-A2 из группы также имелась выявляемая окрашиванием на тетрамеры специфичная в отношении HCV популяция в печени (фиг.25). Экспрессию PD-1 и CD127 прямо сравнивали на специфичных в отношении HCV тетрамер-позитивных CD8+ T-клетках в печени и на периферии у этих индивидуумов. Специфичные в отношении HCV CD8+ T-клетки с периферии были преимущественно PD-1-позитивными (среднее 85%, SEM 3,6) и CD127-позитивными (среднее 84%, SEM 4,0), в то время как специфичные в отношении HCV CD8+ T-клетки были преимущественно PD-1-позитивными (среднее 92%), но лишь редко CD127-позитивными (среднее 13%) (фиг.25). В месте репликации вируса, видимо, происходит размножение CD127-негативных клеток, экспрессирующих PD-1 на высоком уровне. То, что периферические специфичные в отношении антигена CD8+ T-клетки экспрессируют CD127 иначе, чем во внутрипеченочном компартменте, могло бы быть связано с уровнем или временем экспозиции антигена, необходимым для снижения CD127. При инфекции LCMV мышей в условиях экспозиции с сохраняющимся уровнем антигена при хронической инфекции CD127 устойчиво снижался, в то время как краткая экспозиция с антигеном LCMV при использовании GP33 лишь временно подавляла экспрессию CD127 и не вызывала истощение T-клеток (Lang et al., Eur J. Immunol. 35:738-45, 2005). Наблюдалась также зависимость нарушения экспрессии CD62L и CD127 от доступности антигена и времени экспозиции, в то время как сохраняющийся антиген вызывал стойкое снижение и CD62L, и CD127 (Bachmann et al., J. Immunol 175:4686-96, 2005). Вне связи с теорией, при хронической инфекции HCV немногочисленные специфичные в отношении HCV CD8+ T-клетки, выявляемые на периферии, возможно, не экспонируются постоянно с достаточным для поддержания низкого уровня CD127 количеством антигена. Таким образом, T-клетки могут «полагать», что вирус был удален.
Блокада PD-1/PD-L1 ведет к усиленному размножению специфичных в отношении HCV тетрамер-позитивных CD8+ T-клеток: Сведения, полученные на популяции больных, позволяют предполагать, что блокада взаимодействия PD-1/PD-L1 антителом против PD-L1 или PD-1 повышает пролиферативную способность специфичных в отношении HCV T-клеток (фиг.26). Добавление блокирующих антител в присутствии ИЛ-2 и HCV-специфичного пептида приводило к четырехкратному повышению размножения специфичных в отношении HCV T-клеток, как показано отслеживанием частоты CD8+ T-клеток, меченных карбоксифлуоресцеина сукцинимидиловым сложным эфиром (CFSE)low тетрамером, после стимуляции родственным пептидом в течение 6 дней.
Результаты показывают, что в месте инфекции, печени, частота специфичных в отношении HCV CD8+ T-клеток, экспрессирующих PD-1, высока. Во-вторых, большинство специфичных в отношении HCV CD8+ T-клеток из периферической крови больных с хронической инфекцией HCV экспрессирует CD127 на высоком уровне. Фенотип T-клеток при хронической инфекции HCV был охарактеризован путем изучения экспрессии молекулы PD-1, связанной с нарушением эффекторной функции и истощением T-клеток. Результаты показывают, что большинство специфичных в отношении HCV T-клеток во внутрипеченочном компартменте экспрессирует PD-1, но лишено CD127, фенотип, соответствующий истощению T-клеток. Таким образом, антагонисты PD-1 полезны в качестве терапевтических агентов для лечения инфекции HCV.
Пример 19
Блокада PD-1 индуцирует размножение SIV-специфичных CD8 T-клеток in vitro
Противовирусные CD8 T-клетки играют решающую роль в контроле инфекций ВИЧ/SIV. Центральная роль CD8 T-клеток была показана повторным появлением вируса при временных снижениях in vivo у инфицированных SIV макак. В соответствии с этим современные стратегии вакцин, разработанные для индукции высокой частоты противовирусных CD8 T-клеток, включали введение макакам патогенных SHIV и SIV (см., например, Barouch et al., Science 290, 486-92 (2000); Casimiro et al., J. Virol 79, 15547-55 (2005)).
И функция, и частота противовирусных CD8 T-клеток важны для контроля хронических вирусных инфекций, таких как ВИЧ (Migueles et al. Nat Immunol. 3, 1061-8, 2002) и вирус лимфоцитарного хориоменингита (LCMV). Эффективные противовирусные CD8 T-клетки обладают рядом функциональных свойств, включая способность продуцировать различные цитокины, цитотоксический потенциал и высокий пролиферативный потенциал и низкий апоптоз. При хронических вирусных инфекциях специфичные в отношении вируса CD8 T-клетки подвергаются истощению, которое связано с утратой многих из этих функций (Zajac et al., J. Exp. Med. 188, 2205-13, 1998). Сходным образом было показано, что функция специфичных в отношении ВИЧ CD8 T-клеток от индивидуумов с прогрессирующим заболеванием повреждена. Эти CD8 T-клетки могут продуцировать цитокины, такие как IFN-γ, но нарушены продукция ИЛ-2, цитокина, который важен для пролиферации и выживания T-клеток; экспрессия перфорина (Appay et al., J. Exp. Med. 192, 63-75, 2000), молекулы, которая важна для цитолитической функции; и пролиферативная способность, свойство, которому приписывают решающую роль в контроле ВИЧ (см., например, Harari et al., Blood 103, 966-72, 2004) и SIV. Специфичные в отношении ВИЧ CD8 T-клетки экспрессируют PD-1 на высоком уровне, и эта экспрессия прямо пропорциональна уровню виремии. Временная блокада взаимодействия между PD-1 и PD-L1 in vitro восстанавливает функцию специфичных в отношении ВИЧ T-клеток.
Исследовали экспрессию PD-1 на специфичных в отношении SIV CD8 T-клетках после инфицирования макак патогенным SIV239. Результаты показывают, что специфичные в отношении SIV CD8 T-клетки экспрессируют PD-1 на высоком уровне и что блокада пути PD-1:PDL-1 in vitro приводит к усиленному размножению этих клеток.
Были получены следующие результаты:
Повышенная экспрессия PD-1 на специфичных в отношении SIV CD8 T-клетках после инфицирования SIV239: Для понимания роли экспрессии PD-1 и ее связи с контролем инфекции SIV исследовали уровень экспрессии PD-1 на CD8 T-клетках от нормальных здоровых и инфицированных SIV макак. Значительная доля (40-50%) суммарных CD8 T-клеток от нормальных здоровых макак экспрессировала PD-1 (фиг.27A). Экспрессия PD-1 была преимущественно ограничена клетками памяти и отсутствовала на наивных CD8 T-клетках. Сходный паттерн экспрессии PD-1 наблюдался также на суммарных CD8 T-клетках от инфицированных SIVmac239 макак (фиг.27B и C). Однако большинство (>95%) специфичных в отношении SIV Gag CM9 CD8 T-клеток было позитивным по экспрессии PD-1, и в значительной части этих клеток экспрессия PD-1 была дополнительно повышена (MFI 580) по сравнению с суммарными CD8 T-клетками (MFI 220) (фиг.27D). Совместно эти результаты показывают, что значительная доля CD8 T-клеток памяти от нормальных и инфицированных SIV макак экспрессирует PD-1 и что уровень экспрессии PD-1 на специфичных в отношении SIV CD8 T-клетках дополнительно увеличен.
In vitro блокада PD-1 ведет к усиленному размножению специфичных в отношении SIV CD8 T-клеток: Для исследования влияния блокады PD-1 на функцию специфичных в отношении SIV CD8 T-клеток проводили тесты пролиферации в присутствии и в отсутствие блокирующего антитела против молекулы PD-1 человека, которое перекрестно взаимодействует с PD-1 макаки. PBMC от Mamu A*01-позитивных макак-резус, которых инфицировали патогенным вирусом иммунодефицита обезьян и человека 89.6P (SHIV 89.6P), стимулировали пептидом Pl1C (эпитоп Gag-CM9) в отсутствие и в присутствии блокирующего Ab против PD-1 в течение шести дней. В конце стимуляции оценивали частоту Gag CM-9 тетрамер-позитивных клеток. Нестимулированные клетки служили в качестве отрицательных контролей. Как можно видеть на фиг.28A-28B, стимуляция пептидом P11C приводила к приблизительно 4-80-кратному увеличению частоты тетрамер-позитивных клеток. Кроме того, у четырех из шести тестированных макак стимуляция пептидом P11C в присутствии блокирующего Ab против PD-1 приводила к приблизительно 2-4-кратному дополнительному увеличению частоты тетрамер-позитивных клеток по сравнению со стимуляцией пептидом P11C в отсутствие блокирующего антитела.
Эти результаты показывают, что блокада PD-1 повышает пролиферативную способность специфичных в отношении SIV CD8 T-клеток у инфицированных SIV макак.
Пример 20
Роль PD-L2
Два лиганда PD-1 различаются по их паттернам экспрессии: PD-L1 конститутивно экспрессируется и повышается до больших величин и на гематопоэтических, и на негематопоэтических клетках, тогда как PD-L2 индуцибельно экспрессируется только на дендритных клетках (DC) и макрофагах. Хотя некоторые исследования по оценке роли, которую играет PD-L2 в активации T-клеток, показали ингибиторную функцию PD-L2, в других исследованиях сообщалось, что PD-L2 стимулирует пролиферацию T-клеток и продукцию ими цитокинов. Для оценки роли PD-L2 в иммунном ответе T-клеток исследовали кинетику экспрессии PD-L2 клетками разного типа ex vivo после инфицирования LCMV Армстронга (фиг.29). В отличие от экспрессии PD-L1, экспрессия PD-L2 ограниченной на DC в течение очень короткого периода (день 1-4 после инфицирования). Этот результат позволяет предполагать, что экспрессия PD-L2 тесно связана с регуляцией DC и ведет к регуляции активации T-клеток.
Пример 21
PD-1 экспрессируется большинством CD8 T-клеток эффекторной памяти в крови здоровых людей
Исследовали экспрессию PD-1 на CD3+/CD8+ T-клетках из крови здоровых взрослых людей. В крови человека 20-60% CD8 T-клеток экспрессировали PD-1. Проверяли связь между состоянием дифференцировки T-клеток и экспрессией PD-1. На основании паттернов экспрессии CD45RA и CCR7 CD3+/CD8+ T-клетки были поделены на субпопуляции наивных, центральной памяти (TCM), эффекторной памяти (TEM) и окончательно дифференцированных эффекторных (TEMRA). PD-1 не экспрессировался наивными T-клетками и приблизительно одной третью TCM и TEMRA. Напротив, 60% TEM экспрессировали PD-1. Эти данные показывают, что большинство TEM, выделенных из крови здоровых взрослых людей, экспрессирует PD-1.
На основе этих анализов T-клетки были разделены на множество популяций на базе экспрессии CD45RA и CCR7. Была обнаружена дополнительная связь между экспрессией CD45RA и экспрессией PD-1. Конкретно, CCR7-/CD8+ T-клетки с наименьшей экспрессией CD45RA содержали наибольшую долю PD-1+ клеток. В заключение, PD-1 преимущественно экспрессировался TEM, в меньшей степени TEMRA и TCM и не экспрессировался наивными CD8 T-клетками. Эти данные иллюстрируют то, что большая часть TEM CD8 T-клеток у здоровых взрослых людей экспрессирует PD-1.
Для дополнительной характеристики свойств PD-1+ CD8 T-клеток изучали совместную экспрессию PD-1 и нескольких маркеров дифференцировки T-клеток. Большинство PD-1+ CD8 T-клеток несло маркеры, связанные со знакомством с антигеном и эффекторной/эффекторной памяти дифференцировкой. Например, CD11a+/CCR7-/CD62L-/CD45RA-/KLRGl+/гранзим B+/перфорин+ CD8 T-клетки были обогащены по экспрессии PD-1. Напротив, CD8 T-клетки наивного фенотипа (CD11a-/CCR7+/CD62L+/CD45RA+/KLRG1-) экспрессировали PD-1 на низком уровне. Таким образом, PD-1 преимущественно экспрессировался на знакомых с антигеном CD8 T-клетках с эффекторными/эффекторной памяти свойствами.
Пример 22
PD-1 экспрессируется большинством CD4 T-клеток эффекторной памяти в крови здоровых людей
Затем исследовали экспрессию PD-1 среди CD3+ CD4+ T-клеток. В крови здоровых взрослых людей тридцать процентов CD4 T-клеток экспрессировали PD-1. Подобно CD8 T-клеткам, наивные CD4 T-клетки слабо экспрессировали PD-1. В то время как меньшинство TCM CD4 T-клеток экспрессировало PD-1, экспрессия PD-1 была преимущественно обогащена среди TEM CD4 T-клеток (50%).
Для дополнительной характеристики свойств CD4 T-клеток, которые экспрессировали PD-1, исследовали CD4+/CD3+ T-клетки из крови здоровых индивидуумов на предмет совместной экспрессии PD-1 и нескольких маркеров дифференцировки T-клеток. Сходно с CD8 T-клетками, экспрессия PD-1 была обогащена на CD4 T-клетках с эффекторным/эффекторной памяти фенотипом, включая CD62L-, CD95+, CD45RA-, CCR7- и CCR5+ клетки.
Пример 23
PD-1 сильнее экспрессируется на CD8 T-клетках, специфичных в отношении инфекций EBV и CMV у человека
Для проверки того, коррелирует ли экспрессия PD-1 с продолжительным наличием вирусного антигена, сравнивали экспрессию PD-1 на специфичных в отношении вирусов EBV, CMV, гриппа и коровьей оспы CD8 T-клетках. Специфичные в отношении EBV и CMV CD8 T-клетки экспрессировали PD-1 на высоком уровне. Напротив, специфичные в отношении вируса гриппа CD8 T-клетки памяти экспрессировали PD-1 на промежуточном уровне, а специфичные в отношении вируса коровьей оспы CD8 T-клетки экспрессировали PD-1 на низком уровне. Таким образом, CD8 T-клетки памяти, специфичные в отношении хронических инфекций (EBV и CMV), экспрессировали PD-1 на более высоком уровне, чем при острых (грипп и коровья оспа) инфекциях. Эти результаты показывают, что CD8 T-клетки, специфичные в отношении хронических инфекций (EBV и CMV), экспрессировали PD-1 на более высоком уровне, чем при острых инфекциях (вирусы гриппа и коровьей оспы). CD8 T-клетки, специфичные в отношении самых распространенных хронических инфекций, могут экспрессировать PD-1 на высоком уровне.
Пример 24
Блокада анти-PD-L1 повышает пролиферацию CD8 T-клеток, специфичных в отношении инфекций EBV и CMV у человека
Блокада ингибиторного пути PD-1 ведет к повышенной клональной экспансии специфичных в отношении ВИЧ CD8 T-клеток после стимуляции in vitro. Поскольку CD8 T-клетки, специфичные в отношении распространенных хронических инфекций, также экспрессируют PD-1, проверяли, может ли блокада пути PD-1/PD-L1 повышать пролиферацию CD8 T-клеток, специфичных в отношении EBV, CMV, а также вируса коровьей оспы (острая инфекция, дающая PD-1 CD8 T-клетки памяти). Лимфоциты, выделенные из крови индивидуумов, имеющих CD8 T-клетки, специфичные в отношении EBV, CMV или VV, метили CFSE и культивировали в течение 6 дней в разных условиях. Как ожидалось, инкубация свежевыделенных мононуклеарных клеток периферической крови (PBMC) только со средой или средой с антителом против PD-L1 не вызывала пролиферацию специфичных в отношении вируса CD8 T-клеток. Стимуляция PBMC в течение 6 дней пептидами из вирусов приводила к делению тетрамер+ CD8 T-клеток. Однако пептидная стимуляция PBMC в присутствии блокирующего антитела против PD-L1 дополнительно повышала деление специфичных в отношении EBV и CMV CD8 T-клеток, что приводило к большей кратности экспансии по сравнению с одним пептидом. Повышенное деление, индуцированное блокирующим антителом против PD-L1, варьировало между индивидуумами и даже между эпитопами у данного индивидуума. Более того, блокада PD-1 не приводила к повышенной экспансии специфичных в отношении коровьей оспы и гриппа CD8 T-клеток. Степень повышения деления, индуцированного блокирующим антителом против PD-L1 в культуре, могла быть связана с количеством PD-1, экспрессируемым специфичными в отношении антигена CD8 T-клетками перед стимуляцией. Эти данные позволяют предполагать, что экспрессия PD-1 на CD8 T-клетках, специфичных в отношении хронических инфекций, ингибирует их пролиферативную способность при антигенной стимуляции.
Пример 25
Продолжительная блокада PD-L1 дополнительно повышает пролиферацию CD8 T-клеток, специфичных в отношении хронических инфекций
При стимуляции in vitro добавление блокирующего PD-L1 антитела вело к повышенному делению среди CD8 T-клеток, специфичных в отношении EBV и CMV. mAb против PD-L1 добавляли один раз (день 0), и пролиферацию оценивали в конце шестидневного периода культивирования. Лечение мышей анти-PD-L1 in vivo включало множественные инъекции блокирующего антитела. Более того, в этих исследованиях на мышах блокада PD-L1 in vivo приводила к быстрому повышению экспрессии PD-1 среди CD8 T-клеток, специфичных в отношении хронического вирусного антигена. По этим причинам была проведена проверка того, будут ли повторные добавки анти-PD-L1 к стимулированным культурам T-клеток дополнительно повышать пролиферацию. Добавление mAb против PD-L1 в 0, 2 и 4 дни культивирования приводило даже к большему накоплению CD8 T-клеток, специфичных в отношении EBV, чем однократное добавление mAb в день 0. Сходные данные наблюдались для CD8 T-клеток, специфичных в отношении CMV. Эти данные позволяют предполагать, что продолжительная блокада сигнализации PD-1 может оптимизировать способность к увеличению количества CD8 T-клеток, специфичных в отношении хронических антигенов.
Должно быть ясно, что точные детали описанных методов или композиций можно варьировать или модифицировать без отхода от сути описанного изобретения. Заявители заявляют права на все такие модификации и вариации, которые попадают в объем и суть нижеследующей формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПОЗИЦИИ И СПОСОБЫ ЛЕЧЕНИЯ ИНФЕКЦИЙ И ОПУХОЛЕЙ | 2007 |
|
RU2478400C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ ПЕРСИСТИРУЮЩИХ ИНФЕКЦИЙ | 2006 |
|
RU2596491C2 |
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ ПЕРСИСТИРУЮЩИХ ИНФЕКЦИЙ | 2016 |
|
RU2746383C2 |
| СПОСОБ И КОМПОЗИЦИИ ДЛЯ ЛЕЧЕНИЯ ПЕРСИСТИРУЮЩИХ ИНФЕКЦИЙ | 2006 |
|
RU2434641C2 |
| СПОСОБЫ ЛЕЧЕНИЯ РАКА С ИСПОЛЬЗОВАНИЕМ АНТАГОНИСТОВ, СВЯЗЫВАЮЩИХ С ОСЬЮ PD-1, И ИНГИБИТОРОВ TIGIT | 2014 |
|
RU2702108C2 |
| АНТИТЕЛА К PD-L1 И ИХ ПРИМЕНЕНИЕ ДЛЯ УСИЛЕНИЯ ФУНКЦИИ Т-КЛЕТОК | 2009 |
|
RU2636023C2 |
| СПОСОБЫ ЛЕЧЕНИЯ РАКА С ИСПОЛЬЗОВАНИЕМ АНТАГОНИСТОВ, СВЯЗЫВАЮЩИХ С ОСЬЮ PD-1, И ИНГИБИТОРОВ TIGIT | 2014 |
|
RU2825390C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ИММУНОТЕРАПИИ | 2014 |
|
RU2680010C2 |
| СТАБИЛЬНЫЕ СОСТАВЫ АНТИТЕЛ ПРОТИВ РЕЦЕПТОРА ПРОГРАММИРУЕМОЙ СМЕРТИ PD-1 ЧЕЛОВЕКА И ОТНОСЯЩИЕСЯ К НИМ СПОСОБЫ ЛЕЧЕНИЯ | 2012 |
|
RU2633509C2 |
| ТЕРАПИЯ ДЛЯ ЛЕЧЕНИЯ РАКА С ПОМОЩЬЮ ВНУТРИОПУХОЛЕВОГО И/ИЛИ ВНУТРИВЕННОГО ВВЕДЕНИЯ РЕКОМБИНАНТНОГО MVA, КОДИРУЮЩЕГО 4-1BBL (CD137L) И/ИЛИ CD40L | 2019 |
|
RU2830601C2 |


















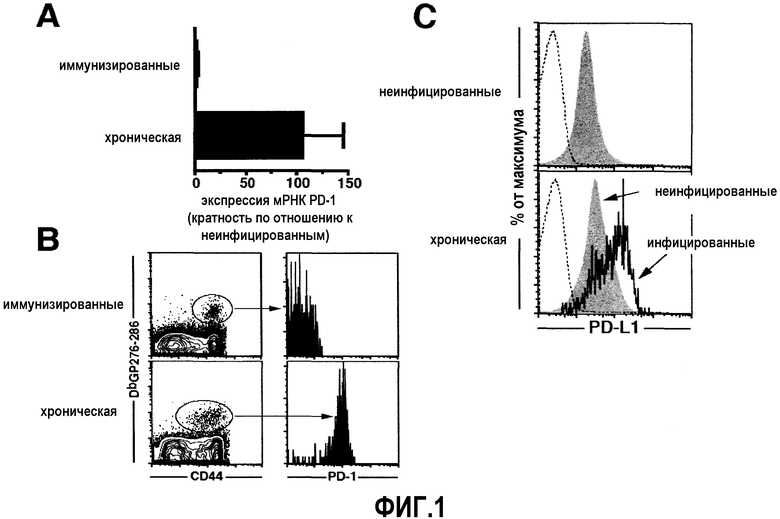
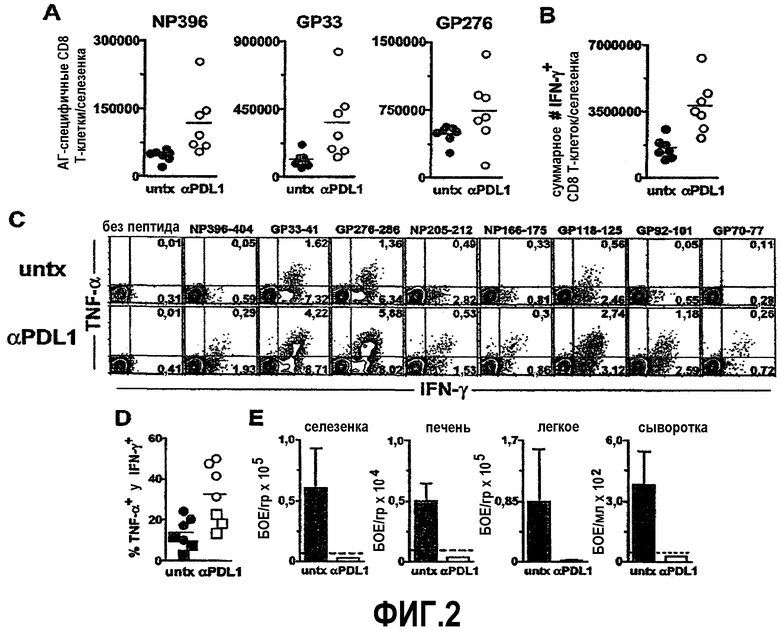
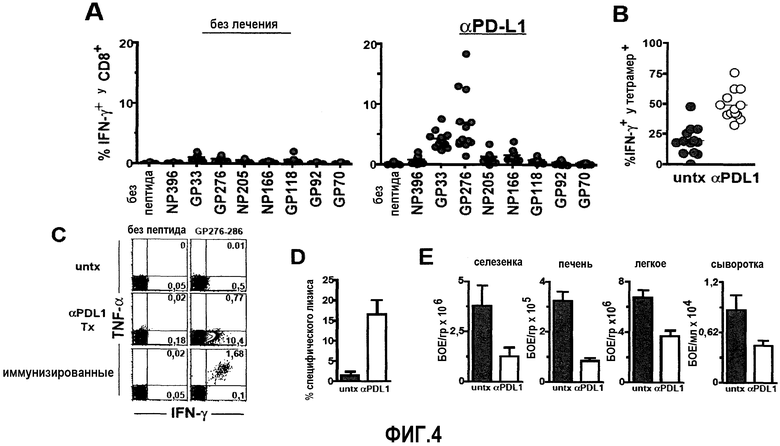
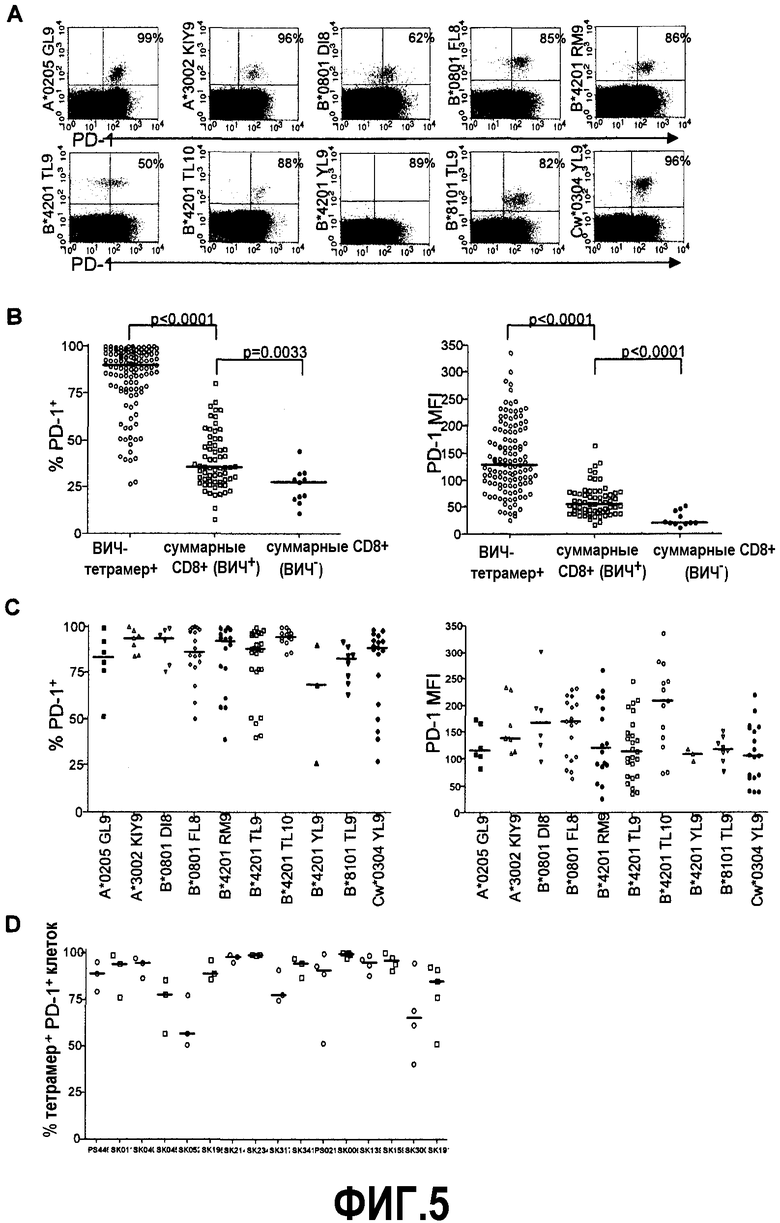
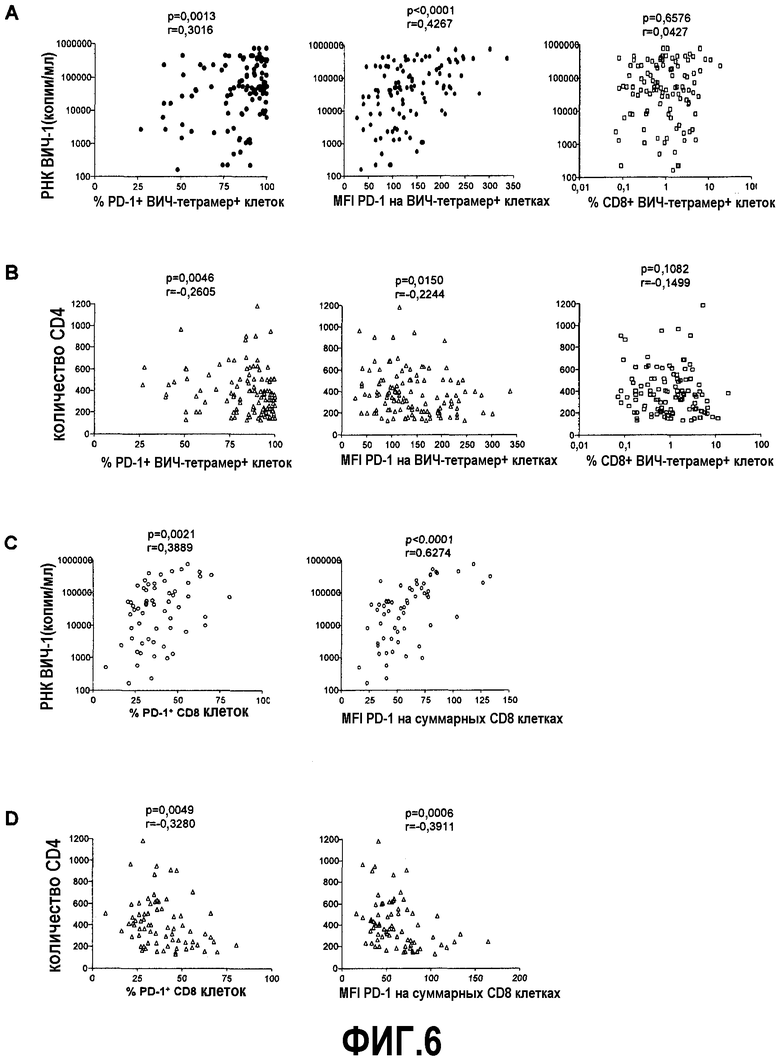
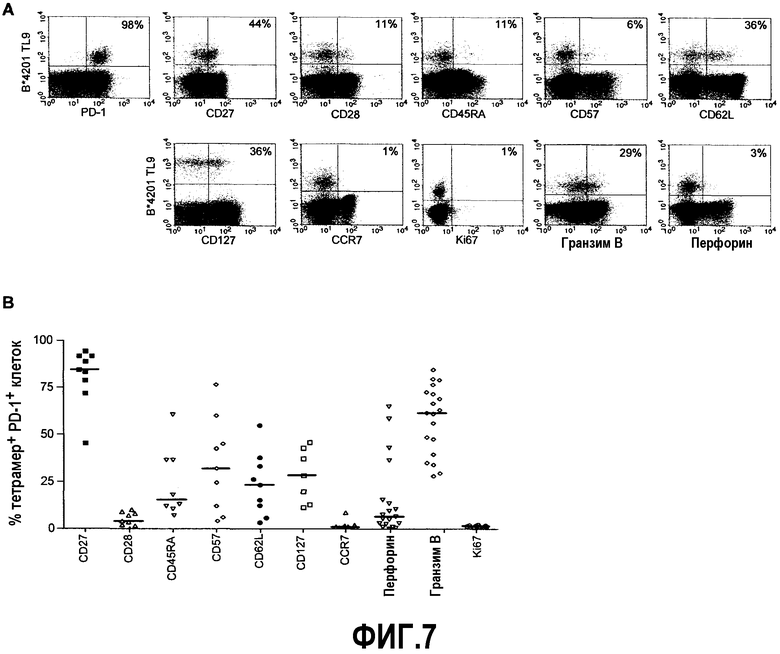
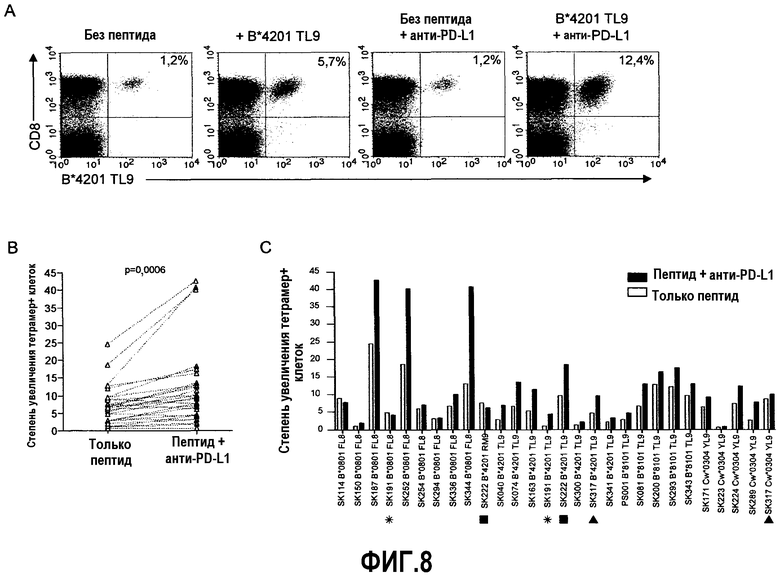
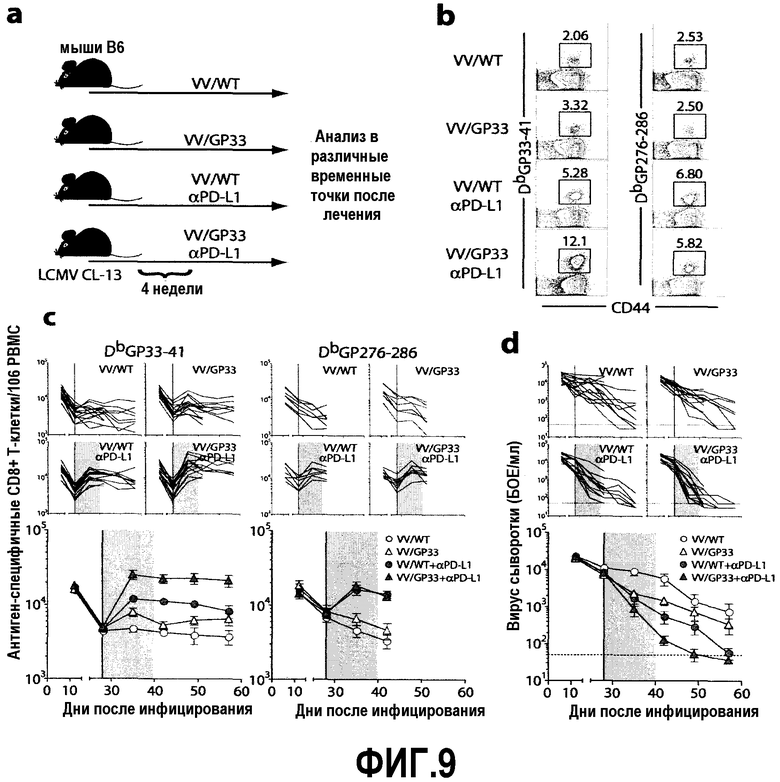
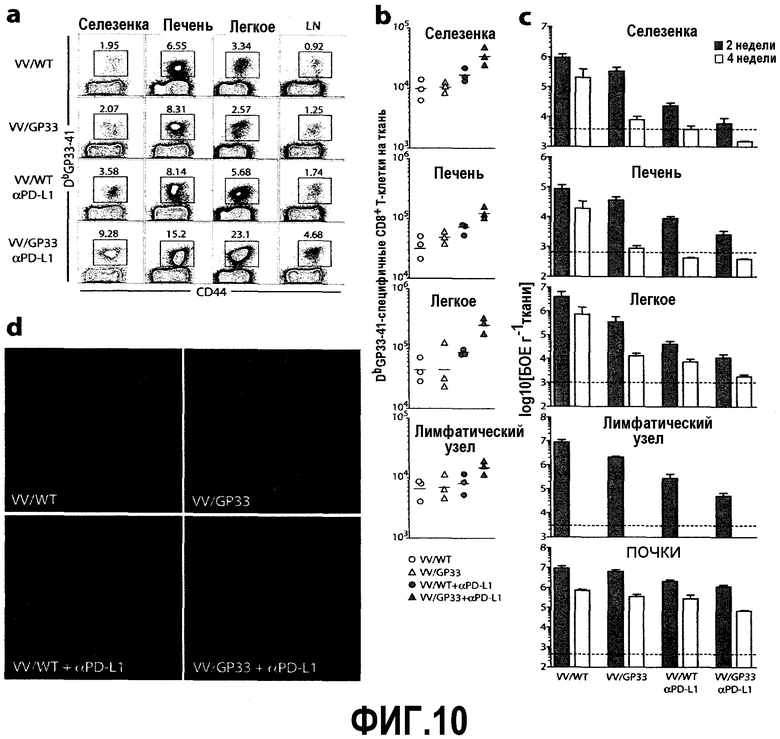
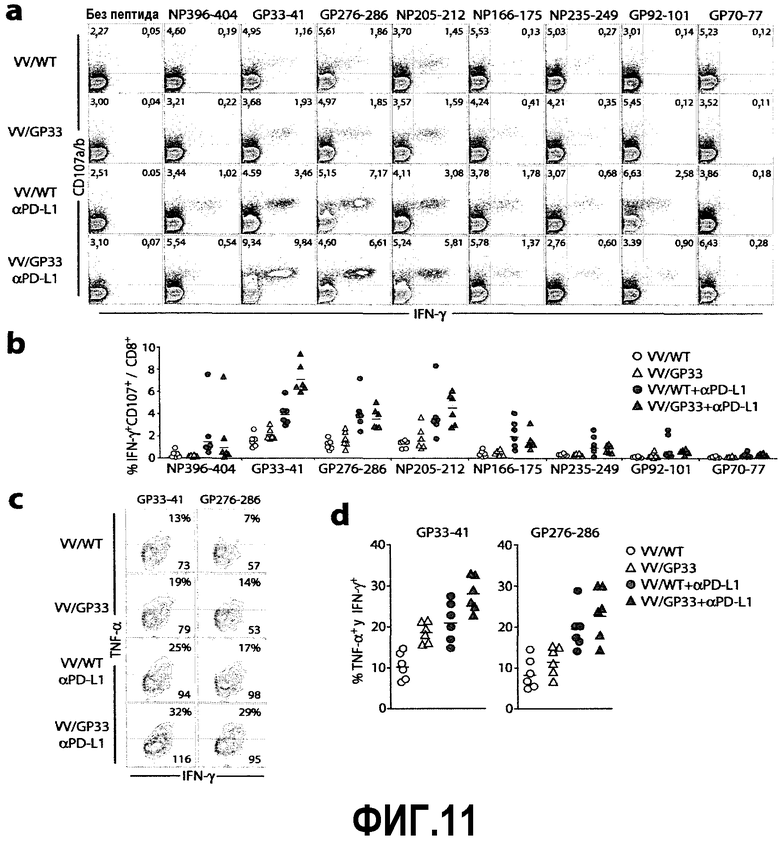
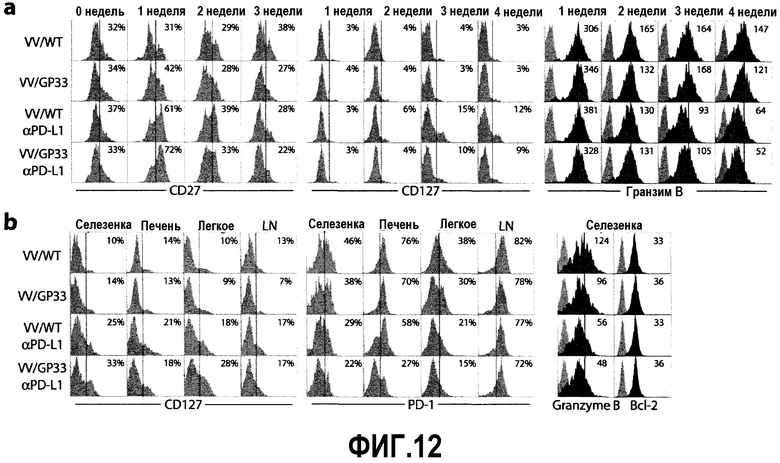
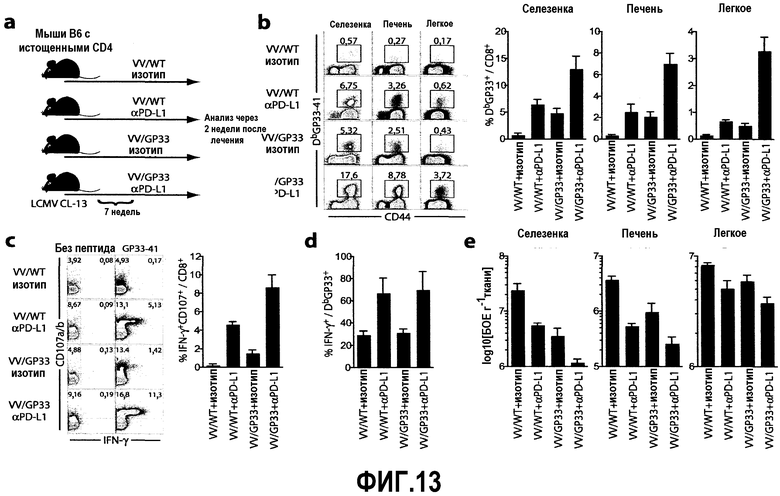
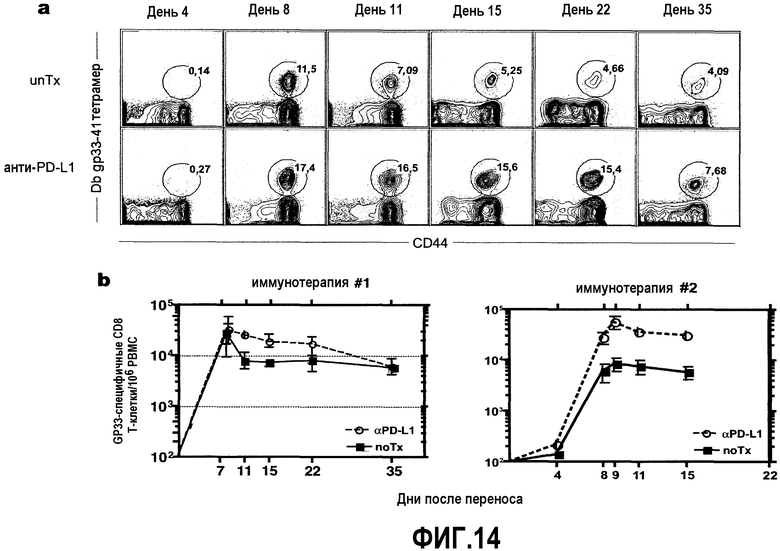
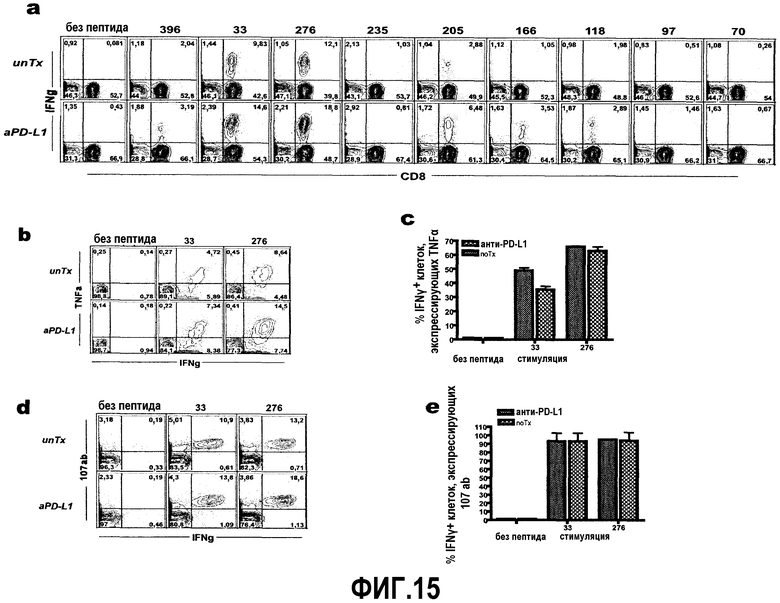
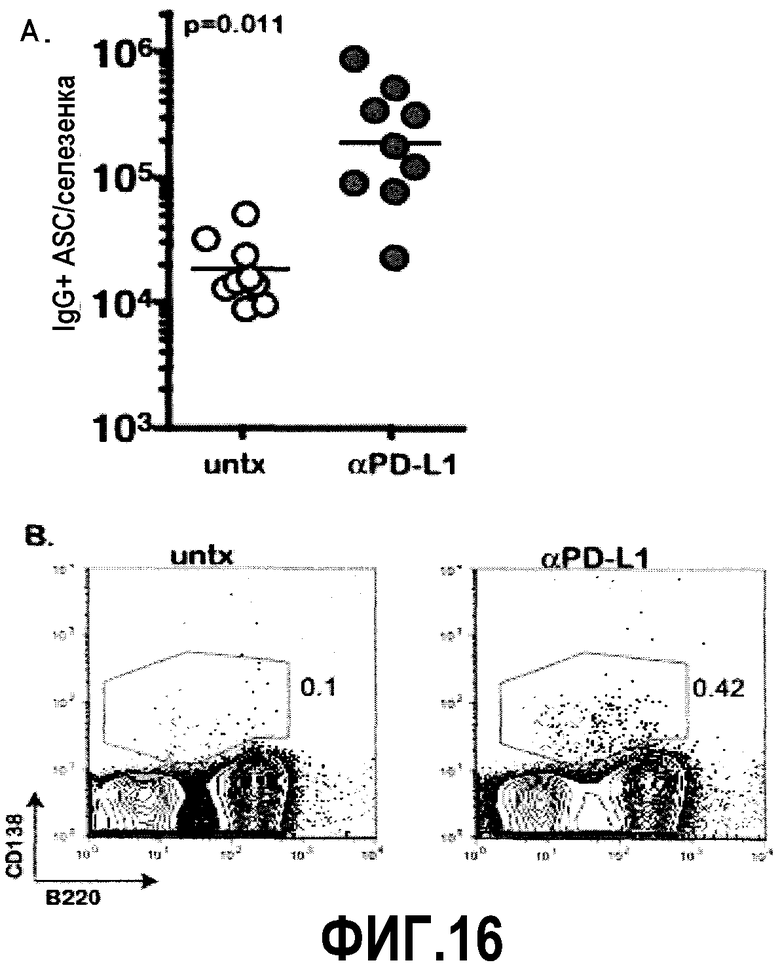
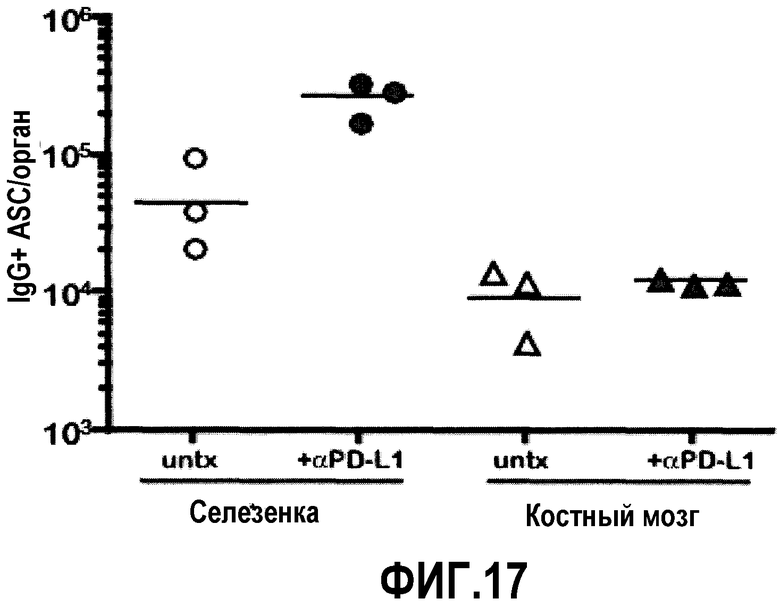
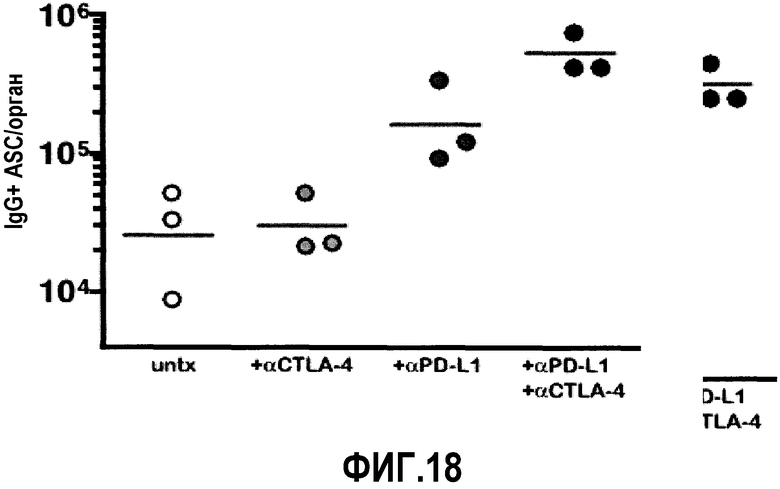
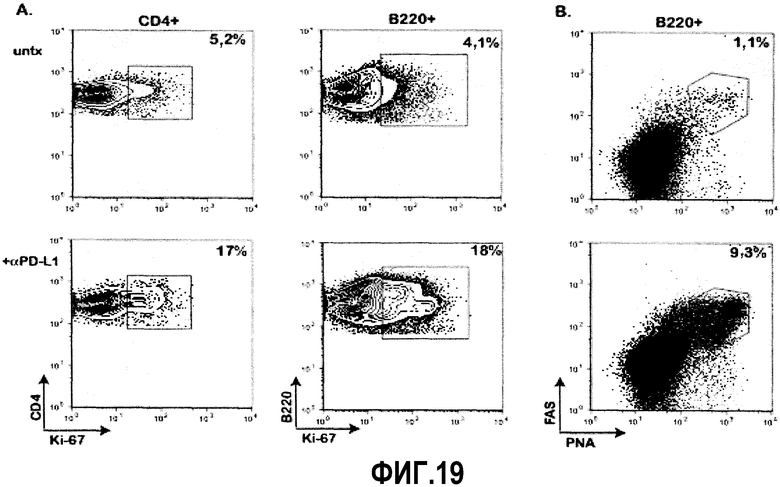
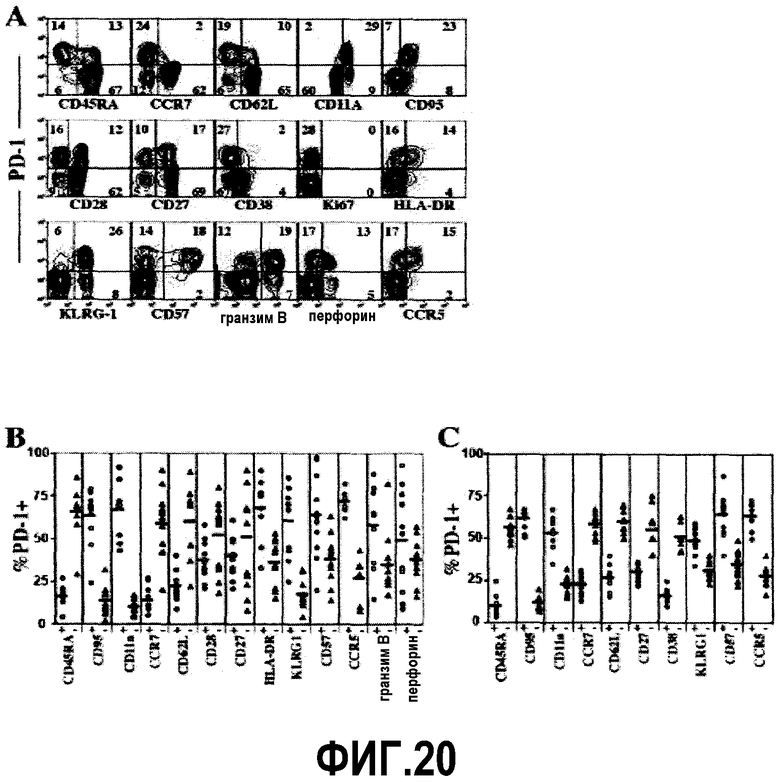
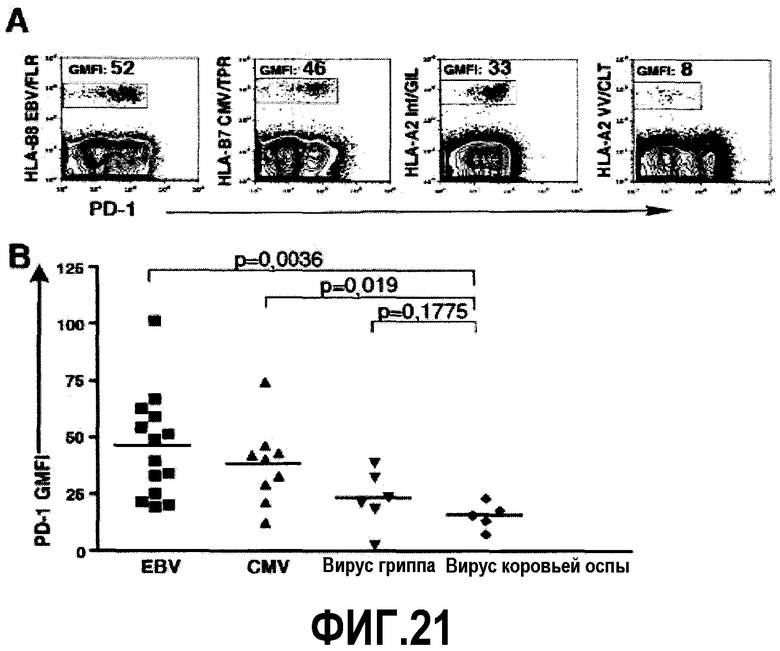
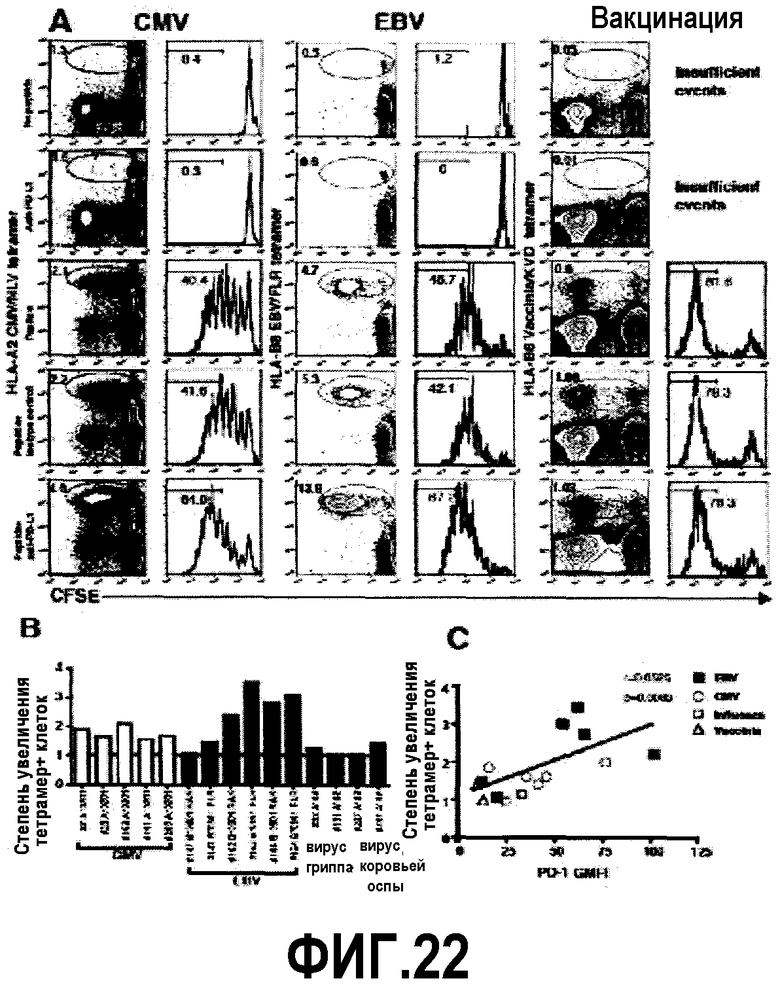
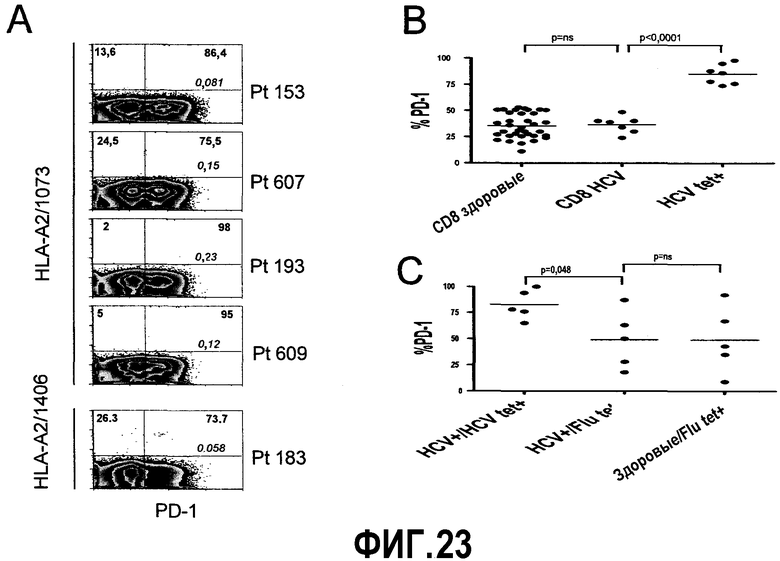
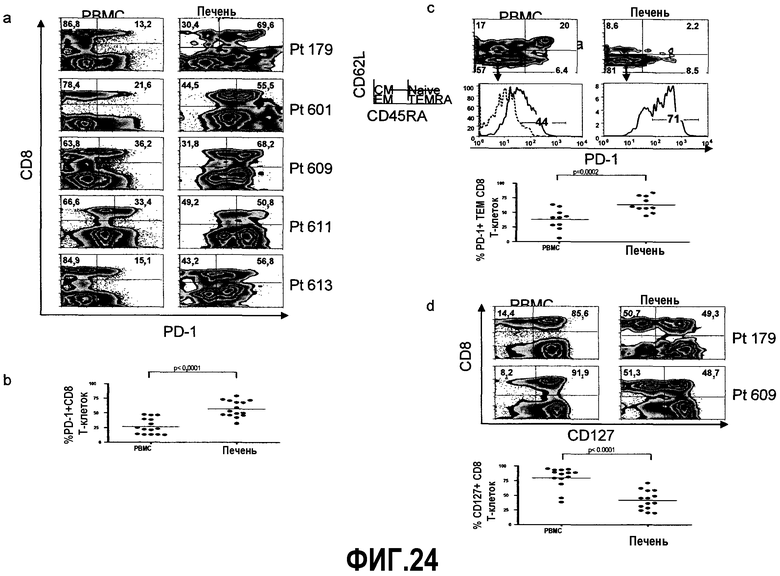
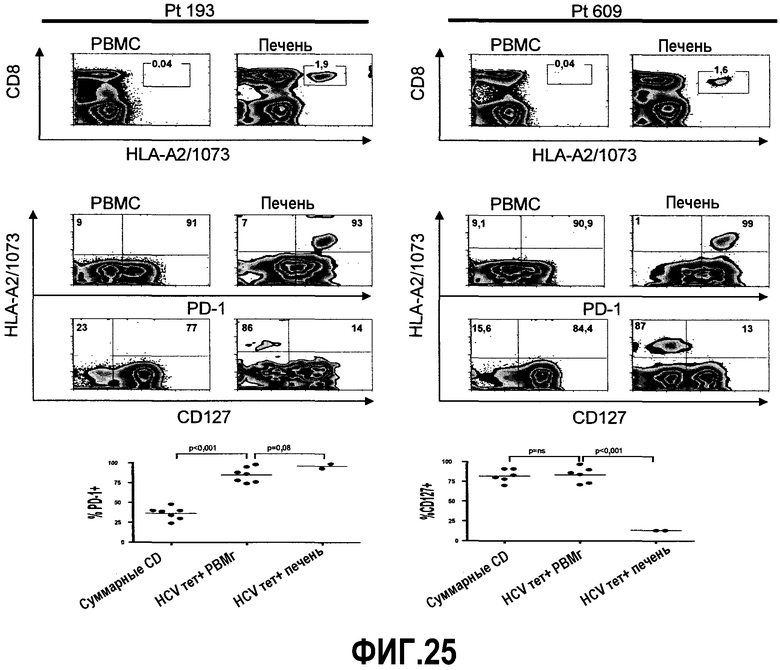
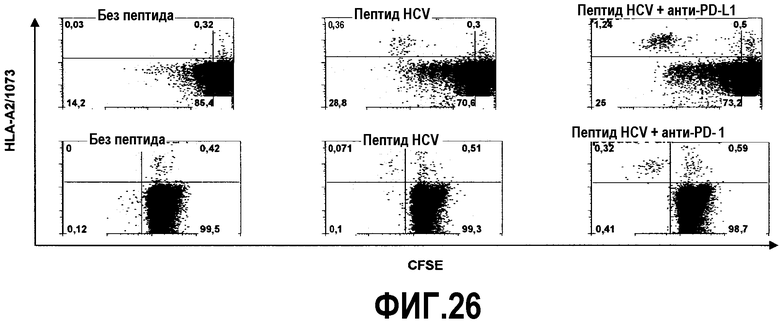
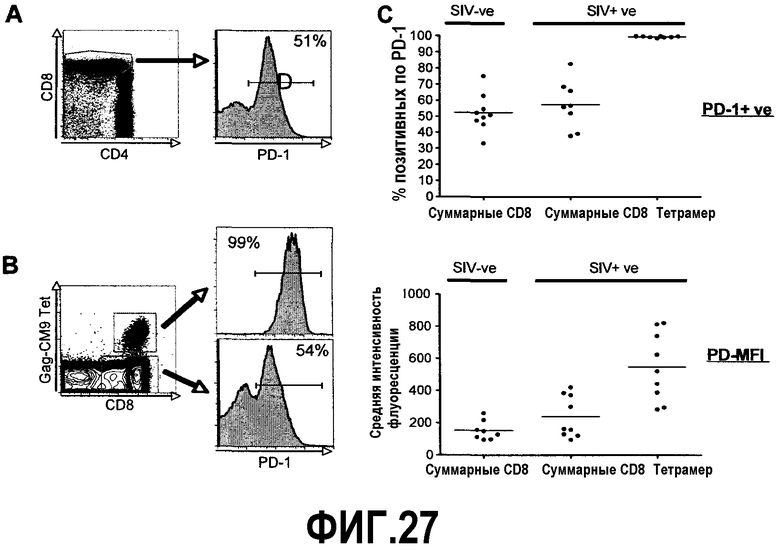
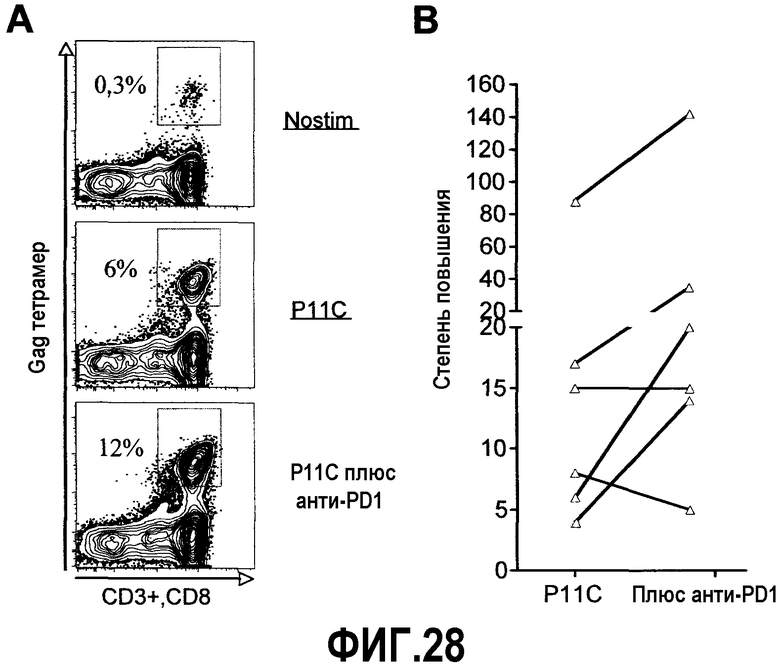
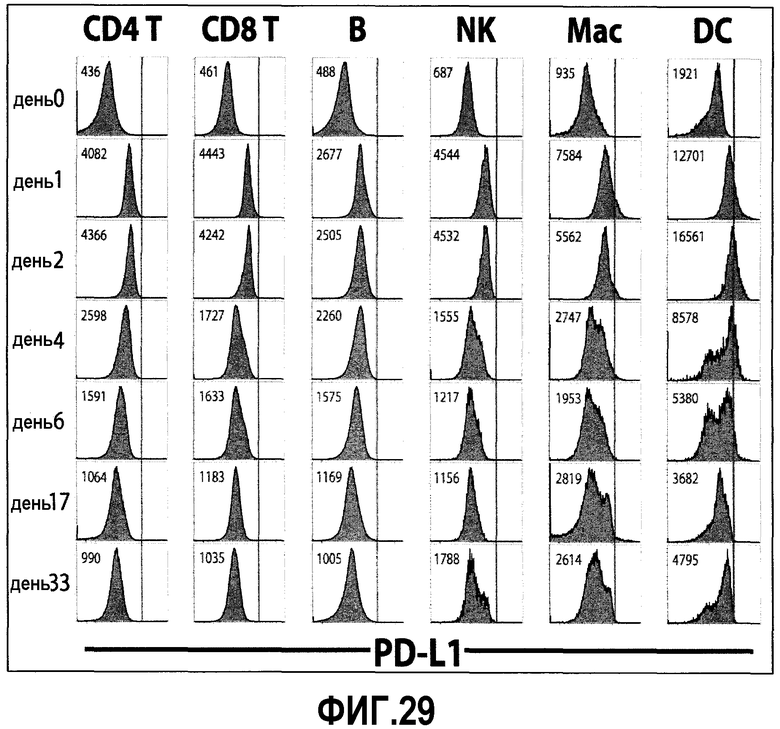
Изобретение относится к медицине, а именно к онкологии и иммунологии, и может быть использовано для лечения индивидуума с устойчивым инфицированием патогеном или опухолью. Для этого индивидууму вводят терапевтически эффективное количество антагониста пептида программируемой смерти (PD-1) и терапевтически эффективное количество антигенной молекулы из патогена или опухоли (вакцина). Изобретение обеспечивает синергическое действие комбинации антигена и антагониста PD-1(антитело против PD-L1) за счет усиления Т-клеточного иммунного ответа в отношении патогена или опухоли. 13 з.п. ф-лы, 29 ил., 5 табл., 25 пр.
1. Способ лечения индивидуума с устойчивым инфицированием патогеном или опухолью, включающий введение индивидууму терапевтически эффективного количества антагониста пептида программируемой смерти (PD-1) и терапевтически эффективного количества антигенной молекулы из патогена или опухоли соответственно,
где патоген представляет собой вирус и где антигенная молекула представляет собой вирусный полипептид или нуклеиновую кислоту, кодирующую вирусный полипептид,
где опухоль представляет собой острый миелогенный лейкоз, хронический миелогенный лейкоз, острый лимфобластный лейкоз, хронический лимфоцитарный лейкоз, неходжкинскую лимфому, множественную миелому, малигнизированную меланому, рак молочной железы, рак легких, рак ободочной кишки или почечно-клеточный рак и
где антигенная молекула представляет собой опухолевый полипептид или нуклеиновую кислоту, кодирующую опухолевый полипептид, и
где антагонист PD-1 представляет собой антитело против PD-1, антитело против PD-L1, малую ингибиторную РНКи против PD-1, малую ингибиторную РНК против PD-L1, антисмысловую РНК против PD-1, антисмысловую РНК против PD-L1, доминантно негативный белок PD-1, доминантно негативный белок PD-L1.
2. Способ по п.1, где антагонист PD-1 представляет собой антитело, которое специфически связывает PD-1 или антитело, которое специфически связывает лиганд-1 пептида программируемой смерти (PD-L1) или их сочетания.
3. Способ по п.1, где индивидуум характеризуется ослабленным иммунитетом.
4. Способ по п.1, где антагонист PD-1 представляет собой антитело против PD-1 или антитело против PD-L1.
5. Способ по п.4, где антитело, которое специфически связывает PD-1, представляет собой (1) моноклональное антитело или его функциональный фрагмент, (2) гуманизированное антитело или его функциональный фрагмент или (3) иммуноглобулиновый гибридный белок.
6. Способ по п.4, где антитело, которое связывает PD-L1, представляет собой (1) моноклональное антитело или его функциональный фрагмент, (2) гуманизированное антитело или его функциональный фрагмент или (3) иммуноглобулиновый гибридный белок.
7. Способ по п.1, где индивидуум имеет устойчивое инфицирование патогеном и способ включает введение вирусного антигена или нуклеиновой кислоты кодирующей вирусный антиген индивидууму, тем самым осуществляя лечение устойчивой инфекции у индивидуума.
8. Способ по п.1, где у индивидуума отсутствуют симптомы, так что лечение предотвращает развитие симптомов.
9. Способ по п.1, где индивидуум имеет опухоль и где способ включает введение индивидууму указанного опухолевого антигена или указанной нуклеиновой кислоты, кодирующей опухолевый антиген, тем самым осуществляя лечение опухоли.
10. Способ по п.9, где антагонист PD-1 представляет собой антитело, которое специфически связывает PD-1 или антитело, которое специфически связывает лиганд-1 пептида программируемой смерти (PD-L1) или их сочетания.
11. Способ по п.10, где антитело, которое специфически связывает PD-1, представляет собой (1) моноклональное антитело или его функциональный фрагмент, (2) гуманизированное антитело или его функциональный фрагмент или (3) иммуноглобулиновый гибридный белок.
12. Способ по п.10, где антитело, которое связывает PD-L1, представляет собой (1) моноклональное антитело или его функциональный фрагмент, (2) гуманизированное антитело или его функциональный фрагмент или (3) иммуноглобулиновый гибридный белок.
13. Способ по п.9, где опухолевый антиген представляет собой PRAME, WT1, сурвайвин, циклин D, циклин Е, протеиназу 3 и ее пептид PR1, эластазу нейтрофилов, катепсин G, MAGE, MART, тирозиназу, GP100, NY-Eso-1, херцептин, раковоэмбриональный антиген (СЕА) или простатспецифический антиген (PSA).
14. Способ по п.1, где вирус представляет собой вирус иммунодефицита человека или вирус гепатита.
| WO 2006133396 A2, 14.12.2006 | |||
| US 20030232323 А1, 18.12.2003 | |||
| YAO S | |||
| et al | |||
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| Печь для сжигания твердых и жидких нечистот | 1920 |
|
SU17A1 |
| TERRAZAS L.I | |||
| et al | |||
| Role of the | |||

