Изобретение относится к области биотехнологии, а именно генной инженерии, и может быть использовано для создания микробиологического дрожжевого продуцента капсидного белка L1 вируса папилломы человека типа 18.
Вирус папилломы человека (HPV - Human Papilloma Virus), поражая эпителиальные клетки, вызывает различные заболевания у человека. В зависимости от типа инфицирующего вируса, который на основании данных о гомологии последовательности ДНК классифицирован более чем в 70 типов, папилломавирусная инфекция проявляется в виде образования доброкачественных бородавок, незлокачественных кондилом или папиллом на слизистой половых органов или дыхательных путей. Вирус папилломы человека типа 16 и 18 вызывает эпителиальную дисплазию слизистой половых органов, именно эти типы вируса папилломы ассоциированы с большей частью преинвазивных и инвазивных карцином аногенитальной сферы (рак шейки матки, влагалища, вульвы и анального канала), из которых рак шейки матки - один из самых распространенных и опасных видов новообразований у женщин. По данным ВОЗ ежегодно раком шейки матки заболевают около 500 тысяч женщин во всем мире, причем более половины этих случаев заканчиваются летальным исходом. Клинические проявления рака шейки матки не обнаруживаются длительное время, в то же время ранняя диагностика папилломавирусной инфекции довольно сложна. Именно поэтому в борьбе с распространением такого тяжелого заболевания предпочтение отдается профилактике папилломавирусной инфекции, в частности, путем индуцирования иммунного ответа у человека при вакцинации.
Вирус папилломы относится к семейству паповирусов (Papoviridae), имеет диаметр 40-50 нм. Капсид вируса образован из 72 белковых капсомеров, состоящих из основного белка L1 и минорного белка L2. Геном вируса представлен в виде кольцевой двуцепочечной ДНК, которая содержит 8 ранних генов (Е1-Е7) и два поздних гена, кодирующих белки капсида L1 и L2. Главный белок L1 (55-60 кДа) способен сам собираться в вирусоподобные частицы (VLP - virus-like particle), сходные по строению с вирионами, но не содержащие вирусную ДНК. Белок L1 обладает высокой иммуногенностью и индуцирует образование антител, нейтрализующих инфекционный вирус.
В настоящее время рекомбинантные белки LI HPV представляют собой перспективные иммуногенные компоненты для создания профилактических вакцин, направленных против опасных типов папилломавируса. Затруднения, препятствующие созданию эффективных папилломавирусных вакцин, прежде всего, связаны с разработкой оптимальных экспрессионных систем для продукции рекомбинантных белков HPV, обладающих иммуногенными свойствами природных белков HPV L1.
Известно получение белков L1 и L2 HPV типа 16 путем культивирования клеток CV-1 млекопитающих, инфицированных рекомбинантным вектором вируса коровьей оспы [Zhou J et al., Virology v.185, p.251-257, 1991]. Известна экспрессия папилломавирусного белка L1, способного спонтанно собираться в вирусоподобные частицы и обладающего иммуногенностью, в эукариотической системе на основе культивируемых клеток насекомых (бакуловирусная система) [Kimbauer R. et al., Proc. Natl. Acad. Sci USA, v.89, р.12180-12184, 1992]. В научной и патентной литературе представлены примеры использования для получения рекомбинантных белков HPV системы экспрессии на основе дрожжей Saccharomyces cerevisiae, которым отдается предпочтение благодаря международно признанной безопасности дрожжей, способности к наработке больших количеств белка в нативной конформации. Так, в патенте RU 2206608, 2003, C12N 7/ 00 описывается создание дрожжевого продуцента белка L1 HPV или L2 HPV разных типов, в том числе HPV 16 и 18, на примере Saccharomyces cerevisiae. Клонирование генов L1 и L2 HPV 16 осуществляли с использованием геномной ДНК, экстрагированной из клеток Caski (ATCC №CRL 1550). Конструировали экспрессионный плазмидный дрожжевой вектор, содержащий последовательность ДНК, кодирующую соответствующий полипептид, под контролем промотора GAL10 и трансформировали им клетки S. cerevisiae. Способность рекомбинантных клеток S. cerevisiae экспрессировать белки L1 или L2 HPV анализировали иммуноблотингом.
Петров P.M. и др. [WO 2006/065166 A1, 2006] получали капсидные белки L1 вирусных серотипов 16, 18, 31 HPV, вызывающих наиболее агрессивные формы заболеваний у человека, из рекомбинантных штаммов дрожжей. Эти штаммы получали трансформацией дрожжевых клеток S. cerevisiae плазмидами с генами, кодирующими белки L1 указанных серотипов. Фрагменты ДНК, кодирующие эти белки, были выделены из клинического материала. Созданные дрожжевые продуценты при культивировании были способны экспрессировать белки L1 в виде вирусоподобных частиц. Белки L1 использовали для приготовления профилактических и терапевтических вакцин.
К недостаткам использования системы экспрессии на основе Saccharomyces cerevisiae можно отнести нестабильность рекомбинантных клеток, связанную с относительно высокой частотой потерь автономной плазмиды при делении клеток. При культивировании штамма в неселективных для плазмиды условиях это приводит к накоплению клеток, не содержащих плазмиды и не способных к синтезу целевого белка. Это существенно осложняет культивирование рекомбинантного штамма и увеличивает себестоимость целевого продукта. Для индукции экспрессии рекомбинантных белков используется достаточно дорогой реагент - галактоза, что увеличивает себестоимость целевого продукта.
Известно использование метилотрофных дрожжей Pichia pastoris для получения основного капсидного белка HPV [Bazan S.B. et al., Arch. Virol. 2009, 154(10), 1609-17]. В геном Р. pastoris был введен ген, кодирующий HPV-L1, под контролем регулируемого метанолом промотора. Этот ген имел оптимизированный кодоновый состав. Очистку белка L1 выполняли с использованием гепарин-сефарозной хроматографии с последующей стадией сборки вирусоподобных частиц. Образование биологически активных VLP подтверждали методами иммуноэлектронной микроскопии и гемагглютинации.
Ближайшим аналогом является RU 2445358, 2012, в котором раскрыт рекомбинантный штамм-продуцент капсидного белка L1 HPV типа 18 Pichia angusta (синоним Hansenula polymorphd) BKM Y-2989D, полученный путем интеграции в геном экспрессионной плазмиды, содержащей фрагмент ДНК с рекомбинантным геном, кодирующим L1 HPV типа 18, под контролем промотора DAK (ген дигидроксиацетон киназы) Н. polymorpha и селективный маркер (ген LEU2 Н. polymorphd). Описанный рекомбинантный штамм-продуцент, имея высокую продуктивность, тем не менее, не обеспечивает получение антигена, обладающего оптимальной для использования в вакцине иммуногенностью. Для повышения продуктивности штамма-продуцента, полученного в соответствии с ближайшим аналогом, экспрессионная кассета рекомбинантного белка вводится в геном реципиентного штамма в большом количестве копий. Однако в некоторых копиях экспрессионной кассеты в ходе получения и культивирования штамма могут возникать мутации, изменяющие свойства целевого белка. При этом контроль за возникновением мутаций в случае высокой копийности гена практически невозможен. Более того, при превышении определенного уровня синтеза может нарушаться процесс укладки целевого белка, что приводит к снижению его выхода, ухудшению биологической активности и усложняет процесс очистки.
Задачей изобретения является создание рекомбинантного штамма дрожжей Н. polymorpha - стабильного эффективного микробиологического дрожжевого продуцента главного капсидного белка вируса папилломы человека типа 18, обладающего необходимыми биологическими свойствами (правильная конформация, иммуногенность, чистота) и низкой себестоимостью. Полученный дрожжевой продуцент должен обеспечивать возможность контролировать отсутствие мутаций в экспрессируемом чужеродном гене, иметь контролируемый уровень экспрессии рекомбинантного гена, чтобы обеспечить высокую эффективность укладки синтезируемого полипептида с образованием природной конформации получаемого продукта, иметь достаточно высокий выход целевого белка, обеспечивающий экономическую целесообразность использования продуцента.
Задача решена тем, что согласно изобретению для получения рекомбинантного штамма-продуцента главного капсидного белка LI HP VI 8 в геном реципиентного штамма DLT2 H. polymorpha последовательно интегрируют две экспрессионные кассеты, содержащие по одной копии нужного гена. Первая кассета содержит фрагмент ДНК с рекомбинантным геном, кодирующим HPV18-L1, под контролем промотора гена МОХ (метанол оксидаза) и селективный маркер (ген TRP3 Н. polymorpha). Вторая экспрессионная кассета содержит аналогичный ген HPV18-L1, но под контролем промотора гена DAK (ген кодирует дигидроксиацетон киназу) дрожжей Н. polymorpha и селективный маркер (ген LEU2 S. cerevisiae). В результате трансформации получают рекомбинантный штамм, содержащий одну копию гена HPV18-L1 под контролем промотора DAK и одну копию гена HPV18-L1 под контролем промотора МОХ. При этом искусственные гены интегрированы непосредственно в геном штамма-продуцента, т.е. находятся в составе хромосом дрожжей Н. polymorpha. Это обеспечивает высокую митотическую стабильность штамма, давая возможность оптимизировать условия его культивирования без учета риска накопления клеток, потерявших способность синтезировать чужеродный белок. Кроме того, наличие только одной копии рекомбинантного гена в каждой экспрессионной кассете с уникальной промоторной и терминальной областью позволяет контролировать отсутствие мутаций этого гена в процессе его введения в геном и при культивировании штамма. Контроль за отсутствием мутаций осуществляют путем анализа нуклеотидной последовательности продуктов полимеразной цепной реакции с использованием праймеров, уникальных к специфическим последовательностям в кодирующей области каждой экспрессионной кассеты.
Преимуществом предложенного изобретения также является возможность получения стабильно высокого уровня синтеза целевого белка без нарушения эффективности его укладки. Использование экспрессионных кассет с двумя разными промоторами (МОХ и DAK), максимальная активность которых достигается при разных условиях культивирования, обеспечивает большую равномерность синтеза продукта в процессе культивирования штамма. При этом не превышается уровень синтеза белка, который может привести к возможному нарушению эффективности укладки и, как следствие, снижению продуктивности штамма.
В результате отбора клонов, способных синтезировать белок HPV18-L1, был выделен трансформант, наиболее отвечающий требуемым свойствам. Полученный рекомбинантный штамм позволяет выделять белок HPV18-L1 в виде VLP с правильной конформацией, обладающий необходимыми антигенными и иммуногенными свойствами, с достаточно высоким выходом. Образование VLP подтверждали с использованием метода гель-фильтрационной хроматографии. Штамм депонирован 21.04.2011 в коллекции ЗАО НПК «Комбиотех» под номером КБТ-11/pPV-182. Штамм является новым и ни в патентной, ни в научно-технической литературе не описан.
Технический результат предложенного изобретения заключается в повышении иммуногенности (улучшении биологических свойств: антигенность, иммуногенность, правильная сборка VLP) рекомбинантного белка LI HPV типа 18, а также снижении себестоимости.
Изобретение может быть проиллюстрировано следующими примерами.
В приводимых примерах все генно-инженерные операции производили согласно стандартным методикам и инструкциям компаний производителей ферментов и наборов для манипуляций с ДНК in vitro. Трансформацию клеток Escherichia coli и Hansenula polymorpha осуществляли согласно ранее описанным методам (Inoue et al, 1990, Gene, 96:23-28. и Bogdanova et al., 1995, Yeast 11:343-353, соответственно). Синтез фрагментов ДНК, а также определение последовательности нуклеотидов производились ЗАО "Евроген" г. Москва. Были использованы синтетические фрагменты ДНК, нуклеотидные последовательности которых приведены в Списке последовательностей.
Пример 1. Получение плазмидного вектора рАМ131, содержащего рекомбинантный ген DAK-HPV18-L1 и селективный маркер ген LEU2 S. cerevisiae.
Фрагмент ДНК DAK-HPV18 (SEQ ID NO:1) и препарат плазмиды AMIpSLI (Agaphonov et al., 1999, Yeast 15:541-551), содержащей ген LEU2 S. cerevisiae в качестве дрожжевого селективного маркера, был гидролизован рестриктазами BsrGI и HindIII. Полученные препараты фрагмента ДНК с рекомбинантным геном и линеаризованной плазмиды AMIpSLI внесли в реакционную смесь для проведения лигирования в концентрации 30 нг/мкл. После инкубации с Т4 ДНК лигазой в течение 2 часов, 2 мкл реакционной смеси использовали для трансформации штамма Е. coli DH5a. Из нескольких выросших трансформантов выделили плазмидную ДНК. На основании рестрикционного анализа была отобрана плазмида, содержащая фрагмент DAK-HPV18. Определение последовательности нуклеотидов этой плазмиды подтвердило, что она содержит ген, кодирующий HPV18-L1 под контролем промотора DAK H. polymorpha. Полученную плазмиду обозначили рАМ131.
Пример 2. Получение плазмидного вектора рКАМ132, содержащего рекомбинантный ген MOX-HPV18-L1 и неполный ген TRP3 H. polymorpha в качестве селективного маркера.
Фрагмент ДНК MOX-HPV18 (SEQ ID NO:2) и препарат плазмиды pTZ-MOX (Agaphonov et al., 1995, Yeast, 11:1241-1247), содержащей неполный ген TRP3 H. polymorpha в качестве дрожжевого селективного маркера, были гидролизованы рестриктазами BglII и Xhol. Продукт гидролиза фрагмента MOX-HPV18 и продукт гидролиза плазмиды pTZ-MOX длиной 4.3 т.п.н. были выделены после электрофоретического разделения в агарозном геле. Полученные препараты ДНК внесли в реакционную смесь для проведения лигирования в концентрации 30 нг/мкл. После инкубации с ДНК лигазой Т4 в течение 2 часов, 2 мкл реакционной смеси использовали для трансформации штамма Е. coli DH5a. Из нескольких выросших трансформантов выделили плазмидную ДНК. На основании рестрикционного анализа была отобрана плазмида, содержащая фрагмент MOX-HPV18. Определение последовательности нуклеотидов этой плазмиды подтвердило, что она содержит ген, кодирующий HPV18-L1 под контролем промотора МОХ H. polymorpha. Полученную плазмиду обозначили рКАМ132.
Пример 3. Получение штамма H. polymorpha, содержащего экспрессионную кассету HPV18-L1 с промотором МОХ.
Штамм H. polymorpha DLT2 [Agaphonov et al., 1994, Yeast v10, pp.509-513; коллекция штаммов лаборатории молекулярной генетики РКНПК МЗ РФ] трансформировали плазмидой рКАМ132, гидролизованной рестриктазами Ecl136II и XhoI. Трансформантов отбирали на среде, содержащей 6.7 г/л смеси солей и витаминов "Yeast Nitrogen Base" (Difco), 20 г/л D-глюкозы, 20 г/л агара и 60 мг/л лейцина. Среди полученных трансформантов были отобраны клоны, способные продуцировать HPV18-L1. Один из таких трансформантов был обозначен DLT2/pKAM132.
Пример 4. Получение штамма H. polymorpha, содержащего две экспрессионные кассеты HPV18-L1, одна из которых с промотором МОХ, а другая с промотором DAK.
Штамм DLT2/pKAM132 трансформировали плазмидой рАМ131, гидролизованной рестриктазой BsrGI. Трансформантов отбирали на среде, содержащей 6.7 г/л смеси солей и витаминов "Yeast Nitrogen Base" (Difco), 20 г/л D-глюкозы и 20 г/л агара. Один из полученных трансформантов был обозначен КБТ11/pPV-182. У этого трансформанта было подтверждено наличие интактного гена DAK-HPV18 (см. пример 5) и интактного гена MOX-HPV18 (см. пример 6).
Пример 5. Определение наличия интактного гена DAK-HPV18 в геноме трансформантов Н. polymorpha.
Из анализируемых клонов получали препараты геномной ДНК и использовали в качестве матрицы для проведения полимеразной цепной реакции (ПЦР) с олигонуклеотидами dakFwl (SEQ ID NO:5) и AMI20 (SEQ ID NO:6) в качестве праймеров. ПЦР проводили с использованием полимеразы Pfu (Fermentas). При наличии в геноме гена DAK-HPV18 образовывался продукт ПЦР длиной 1739 п.н. Для подтверждения интактности этого гена определяли последовательность нуклеотидов полученного продукта ПЦР.
Пример 6. Определение наличия интактного гена MOX-HPV18 в геноме трансформантов Н. polymorpha.
Из анализируемых клонов получали препараты геномной ДНК и использовали в качестве матрицы для проведения полимеразной цепной реакции (ПЦР) с олигонуклеотидами МОХ20 (SEQ ID NO:3) и MOX3′TRP3 (SEQ ID NO:4) в качестве праймеров. ПЦР проводили с использованием полимеразы Pfu (Fermentas). При наличии в геноме гена MOX-HPV18 образовывался продукт ПЦР длиной 1717 п.н. Для подтверждения интактности этого гена определяли последовательность нуклеотидов полученного продукта ПЦР.
Пример 7. Культивирование штамма Н. polymorpha KBTll/pPV-182, содержащего две экспрессионные кассеты HPV18-L1, одна из которых с промотором МОХ, а другая с промотором DAK.
Ферментацию штамма-продуцента дрожжей Н. polymorpha осуществляли в два этапа. На первом этапе в режиме фед-бэтч при температуре 30°С наращивали биомассу в культуральной среде, содержащей 4% дрожжевого экстракта, 2% бакто-пептона и 4% глицерина. Когда плотность сырой биомассы достигала 150 г/л (примерно через 28-32 часа ферментации), добавляли индуктор до концентрации 0.5-0.8% и поддерживали на этом уровне в течение 48-72 часов.
Пример 8. Выделение и очистка рекомбинантного белка HPV18-L1 из клеток Н. polymorpha.
После ферментации штамма-продуцента белок HPV18-L1 (аминокислотная последовательность - SEQ ID NO:7) выделяли согласно опубликованной методике [Kim H.J. et al., Protein Expression and Purification, v.70, p.68-74, 2010] с небольшими изменениями. Клетки из культуральной жидкости осаждали центрифугированием при 4000g в течение 15 минут при 4°С. Осажденную биомассу ресуспендировали до концентрации 380 г влажных клеток на литр суспензии в буфере для экстракции: PBS (рН 7.2) с добавлением 1.7 mM EDTA, 2 mM PMSF, 0.01% Tween-80. Далее клетки разрушали в мельнице Dyno-Mill типа KDL, используя стеклянные шары диаметром 0.5-0.7 мм. Для удаления основной массы нецелевых белков полученный экстракт насыщали сульфатом аммония до 45% от насыщения и осажденные белки отделяли центрифугированием при 12000 g в течение 30 минут. Осадок ресуспендировали в 9 объемах буфера PBS (рН 7.2)+0.01% Tween-80 для достижения 5% насыщения сульфатом аммония и инкубировали в течение 24 часов при комнатной температуре для полного растворения целевого белка и осаждения части примесных белков, которые затем отделяли центрифугированием при 12000 g в течение 30 минут. Полученный супернатант диализовали в буфер для связывания: PBS (рН 7.0), содержащий 0.2 М NaCl и 0.01% Tween-80 с последующей адсорбцией на сорбент Heparin Sepharose CL-6B (GE Healthcare, USA). После промывки сорбента тем же буфером для связывания белок HPV18-L1 элюировали линейным градиентом от 0.2 М до 1.0 М NaCl. Объединенные фракции, содержащие антиген (по иммуноблоту и SDS-PAGE), концентрировали ультрафильтрацией в тангенциальном потоке на мембране 500 KDa и разделяли при помощи зонального ультрацентрифугирования (Beckman Coulter, USA) в градиенте глицерин/сахароза. Фракции, содержащие целевой белок, объединяли, а затем очищали от глицерина и сахарозы гельфильтрацией на сорбенте Toyopearl HW-65 (ToyoSoda Corp., Japan). Чистоту полученного таким образом рекомбинантного белка HPV18-L1 определяли методом SDS-PAGE электрофореза. Если чистота белка была недостаточна (менее 95%) производили дополнительную очистку при помощи катионобменной хроматографии на сорбенте Toyopearl SP-650M.
В результате этих операций удается выделить около 20% белка L1, присутствующего в осветленном клеточном лизате. Выход очищенного HPV18-L1 составлял не менее 50 мг/л культуральной жидкости.
Пример 9. Анализ рекомбинантного HPV18-L1.
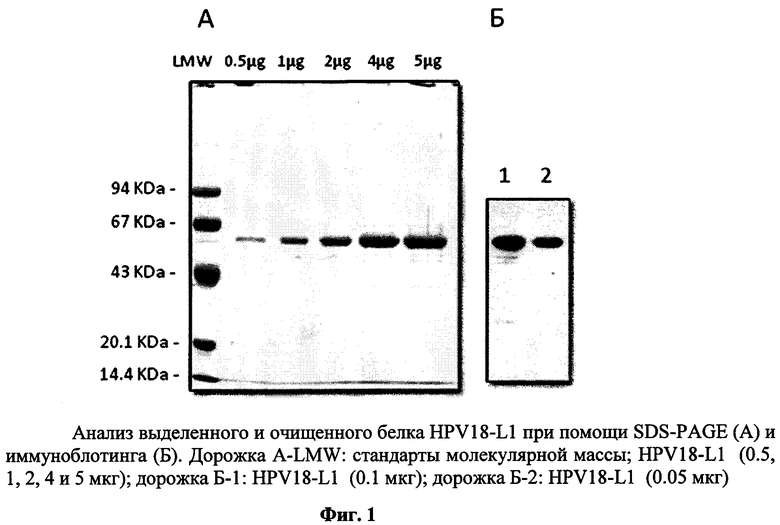
1) Чистота рекомбинантного целевого белка определяется электрофорезом в полиакриламидном геле в восстанавливающих условиях (SDS-PAGE) путем окрашивания с Coomassie Brilliant Blue R-250 и анализа интенсивности полос отсканированных гелей посредством компьютерной программы NIH Image. Чистота выделенного рекомбинантного белка HPV18-L1 составляет не менее 95% (фиг.1А).
2) Иммуноспецифичность рекомбинантного HPV18-L1 определяют методом иммуноблотинга. Первичными моноклональными антителами к белку HPV-L1 (Camvir-1, AbCam, UK) окрашивают иммуноблот с последующей визуализацией при помощи специфических антител к иммуноглобулинам мыши, коньюгированными с пероксидазой хрена, по методике улучшенной хемилюминисценции ECL (Amersham, UK). Рекомбинантный антиген HPV18-L1 представлен в виде полосы мономера белка размером 55-60 кДа (фиг.1Б).
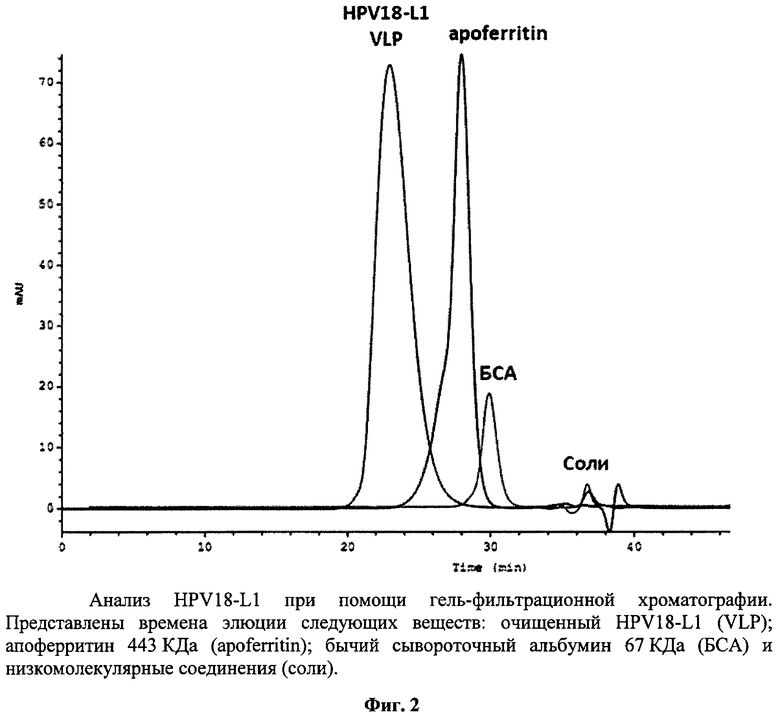
3) Образование VLP рекомбинантного белка HPV18-L1 подтверждали методом гель-фильтрационной хроматографии на полимерной колонке фирмы Tosoh Bioscience (Japan) TSK G5000PW диаметром 7.5 мм и длиной 60 см, соединенной с предколонкой PrePW диаметром 7.5 мм и длиной 7.5 см. Размер пор сорбента составляет 1000 Å, что позволяет отделить правильно собранные VLP от мономеров белка L1, а также агрегатов. Разделение проводили в буфере 10 mM KH2PO4, (рН 7.2), 0.03% Tween-20 при скорости потока 0.6 мл/мин. Мониторинг процесса осуществляли по поглощению на 280 нм. Время элюции белка L1 соответствует времени элюции вирусоподобных частиц (рис.2).
4) Иммуногенность рекомбинантного HPV-L1 определяли в тесте на мышах линии Balb/c с массой 12-14 г после однократной вакцинации белком HPV18-L1, сорбированном на гидроокиси алюминия. Значение ED/50 (доза антигена, вызывающая сероконверсию у 50% мышей) составляет не более 100 нг, что является хорошим показателем иммуногенных свойств рекомбинантного белка. Относительная потенция (отношение дозы, выраженной в ED/50, аналога к ED/50 испытуемого образца) HPV18-L1 по изобретению в сравнении с HPV18-L1, полученным в соответствии с ближайшим аналогом, составляет более 1.0 (1.0-1.25).
Культурально-морфологические особенности штамма: клетки округлой формы, небольшие по размеру, на агаризованной среде YPD образуют крупные круглые колонии с выраженной выпуклой серединой.
Хранение - при -70°С в виде суспензии клеток в стерильном 30-50%-ном растворе глицерина.
Генетические особенности: штамм не является зоопатогенным или фитопатогенным.
Способ, условия и состав сред для размножения штамма: инкубирование прокачиванием при 30°С в питательной среде состава: 2% пептона, 1% дрожжевого экстракта, 2% глюкозы.
Условия и состав среды для ферментации: прокачивание при 30°С и рН 5.0-5.5 в питательной среде, содержащей до 4% глицерина.
Активность штамма определяли в осветленном гомогенизате клеток методом иммуноблотинга или иммуноферментного анализа. Активность штамма - не менее 50 мг/л культуральной жидкости.
Выделенный белок LI HPV типа 18 может быть использован в тест-системах, а также для создания на его основе вакцин для профилактики заболеваний, ассоциированных с вирусом папилломы человека.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНАЯ ВАКЦИНА ДЛЯ ПРОФИЛАКТИКИ ПАПИЛЛОМАВИРУСНОЙ ИНФЕКЦИИ ЧЕЛОВЕКА И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2014 |
|
RU2546243C1 |
| Способ получения рекомбинантной вакцины для профилактики папилломавирусной инфекции человека, рекомбинантная вакцина | 2018 |
|
RU2681174C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ Hansenula polymorpha - ПРОДУЦЕНТ ГЛАВНОГО КАПСИДНОГО БЕЛКА L1 ВИРУСА ПАПИЛЛОМЫ ЧЕЛОВЕКА ТИПА 56 | 2014 |
|
RU2546240C1 |
| Рекомбинантный штамм дрожжей Hansenula polymorpha - продуцент главного капсидного белка L1 вируса папилломы человека типа 11 | 2018 |
|
RU2676160C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ Hansenula polymorpha - ПРОДУЦЕНТ ГЛАВНОГО КАПСИДНОГО БЕЛКА L1 ВИРУСА ПАПИЛЛОМЫ ЧЕЛОВЕКА ТИПА 16 | 2014 |
|
RU2546241C1 |
| Рекомбинантный штамм дрожжей Hansenula polymorpha - продуцент главного капсидного белка L1 вируса папилломы человека типа 6 | 2018 |
|
RU2675471C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ PICHIA ANGUSTA - ПРОДУЦЕНТ КАПСИДНОГО БЕЛКА L1 ВИРУСА ПАПИЛЛОМЫ ЧЕЛОВЕКА ТИПА 18 | 2011 |
|
RU2445358C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ PICHIA ANGUSTA - ПРОДУЦЕНТ КАПСИДНОГО БЕЛКА L1 ВИРУСА ПАПИЛЛОМЫ ЧЕЛОВЕКА ТИПА 16 | 2011 |
|
RU2445357C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ HANSENULA POLYMORPHA - ПРОДУЦЕНТ КАПСИДНОГО БЕЛКА ВИРУСА ГЕПАТИТА Е | 2012 |
|
RU2493249C1 |
| РЕКОМБИНАНТНАЯ ВАКЦИНА ДЛЯ ПРОФИЛАКТИКИ ВИРУСНОГО ГЕПАТИТА Е У ЖИВОТНЫХ | 2012 |
|
RU2501568C1 |


Изобретение относится к биотехнологии, а именно генной инженерии. Предложен штамм дрожжей Hansenula polymorpha - продуцент рекомбинантного белка L1 вируса папилломы человека (HPV) типа 18. Штамм был получен путем введения в геном клетки дрожжей одной копии последовательности ДНК, кодирующей капсидный белок L1 HPV типа 18, под контролем промотора ДАК и одной копии последовательности ДНК, кодирующей капсидный белок L1 HPV типа 18, под контролем промотора МОХ. Штамм обеспечивает высокий выход рекомбинантного белка L1 HPV, обладающего антигенными и иммуногенными свойствами природного антигена. Изобретение может быть использовано для микробиологического получения рекомбинантного белка L1 HPV типа 18. 2 ил., 9 пр.
Рекомбинантный штамм дрожжей Hansenula polymorpha KБT-11/pPV-182, содержащий интегрированный в геном фрагмент ДНК с рекомбинантным геном, кодирующим полипептид L1 HPV типа 18, под контролем промотора гена DAK H. polymorpha (SEQ ID NО:1) и фрагмент ДНК с рекомбинантным геном, кодирующим полипептид L1 HPV типа 18, под контролем промотора гена МОХ Н. polymorpha ( SEQ ID NО:2) - продуцент рекомбинантного капсидного белка L1 вируса папилломы человека (НРV) типа 18.
| ПАПИЛЛОМАВИРУСНЫЕ ВАКЦИНЫ | 1995 |
|
RU2206608C2 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ PICHIA ANGUSTA - ПРОДУЦЕНТ КАПСИДНОГО БЕЛКА L1 ВИРУСА ПАПИЛЛОМЫ ЧЕЛОВЕКА ТИПА 16 | 2011 |
|
RU2445357C1 |
| US 20110086054 A1, 14.04.2011 | |||

