Изобретение относится к области биотехнологии, а именно генной инженерии, и касается рекомбинантной вакцины для профилактики папилломавирусной инфекции, содержащей в своем составе в качестве активного компонента рекомбинантный капсидный белок L1 вируса папилломы человека типа 16, 18, 6, 11, полученный путем культивирования рекомбинантных штаммов дрожжей Hansenula polymorpha, а также способа получения вакцины.
Вирус папилломы человека (HPV - Human Papilloma Virus), поражая эпителиальные клетки, вызывает различные заболевания у человека. На основании отличий в последовательности ДНК выделяют более 200 типов вируса папилломы. По данным ВОЗ более 630 миллионов человек являются носителями папилломавирусной инфекции, из них у 190 миллионов наблюдаются клинические проявления. Вирус папилломы человека типа 16 и 18 вызывает эпителиальную дисплазию слизистой половых органов, именно эти типы вируса папилломы ассоциированы с большей частью преинвазивных и инвазивных карцином аногенитальной сферы (рак шейки матки, влагалища, вульвы и анального канала), из которых рак шейки матки - один из самых распространенных и опасных видов новообразований у женщин. Ежегодно раком шейки матки заболевают около 500 тысяч женщин во всем мире, причем более половины этих случаев заканчиваются летальным исходом. Вирус папилломы человека типов 6 и 11 не относятся к онкогенным: папилломавирусная инфекция в этом случае проявляется в виде образования доброкачественных бородавок, незлокачественных кондилом или папиллом на слизистой половых органов или дыхательных путей. В мире ежегодно регистрируется 30 миллионов новых случаев генитальных бородавок, связанных с папилломавирусной инфекцией. Клинические проявления инфекции не обнаруживаются длительное время, в то же время ранняя диагностика папилломавирусной инфекции довольно сложна. Именно поэтому в борьбе с распространением такого тяжелого заболевания предпочтение отдается профилактике папилломавирусной инфекции, в частности, путем индуцирования иммунного ответа у человека при вакцинации.
Вирус папилломы относится к семейству пaповирусов (Papoviridae ), имеет диаметр 40-50 нм. Капсид вируса образован из 72 белковых капсомеров, состоящих из основного белка L1 и минорного белка L2. Геном вируса представлен в виде кольцевой двуцепочечной ДНК, которая содержит 8 ранних генов (Е1-Е7) и два поздних гена, кодирующих белки капсида L1 и L2. Главный белок L1 (55-60 кДа) способен сам собираться в вирусоподобные частицы (VLP, virus-like particles), сходные по строению с вирионами, но не содержащие вирусную ДНК. Белок L1 обладает высокой иммуногенностью и индуцирует образование антител, нейтрализующих инфекционный вирус.
В настоящее время рекомбинантные белки L1 HPV представляют собой перспективные иммуногенные компоненты для создания профилактических вакцин, направленных против опасных типов папилломавируса. Такие рекомбинантные вакцины безопасны, поскольку не содержат потенциально онкогенного вирусного генома. Экспрессируемый в эукариотической клетке рекомбинантный белок L1 способен к самосборке в капсидоподобную структуру, в основном отражающую VLP, морфологически и антигенно эквивалентную аутентичным вирионам. Иммунизация подобными VLP способствует продукции антител, нейтрализующих вирус, что приводит к защите против HPV инфекции.
Затруднения, возникающие при создании эффективных папилломавирусных вакцин, прежде всего связаны с разработкой оптимальных экспрессионных систем для продукции рекомбинантных белков HPV, обладающих иммуногенными свойствами природных белков HPV L1, способных к образованию конформационно правильных VLP.
Известно получение белков L1 и L2 HPV типа 16 путем культивирования клеток СV-1 млекопитающих, инфицированных рекомбинантным вектором вируса коровьей оспы [Zhou J et al., Virology v. 185, p. 251-257, 1991]. Известна экспрессия папилломавирусного белка L1, в том числе типов 6 и 11, способного спонтанно собираться в вирусоподобные частицы и обладающего иммуногенностью, в эукариотической системе на основе культивируемых клеток насекомых (бакуловирусная система) [Kirnbauer R. et al., Proc.Natl.Acad.Sci USA, v. 89, p. 12180-12184, 1992], US 8, 062, 642, US 9, 050, 287, US 9, 745, 351, US 6,165,471. В научной и патентной литературе представлены примеры использования для получения рекомбинантных белков HPV системы экспрессии на основе дрожжей Saccharomyces cerevisiae, которым отдается предпочтение благодаря международно признанной безопасности дрожжей, способности к наработке больших количеств белка в нативной конформации. Так, в патенте RU 2206608, 2003, С 12 N 7/ 00 описывается создание дрожжевого продуцента белка L1 НРV или L2 НРV разных типов, в том числе HPV 6 и 11, на примере Saccharomyces cerevisiae. Клонирование генов L1 и L2 HPV осуществляли с использованием геномной ДНК, экстрагированной из клеток Caski (АТСC № СRL 1550). Конструировали экспрессионный плазмидный дрожжевой вектор, содержащий последовательность ДНК, кодирующую соответствующий полипептид, под контролем промотора GAL10 и трансформировали им клетки S. cerevisiae. Способность рекомбинантных клеток S. cerevisiae экспрессировать белки L1 или L2 НРV анализировали иммуноблотингом.
Известно использование метилотрофных дрожжей Pichia pastoris для получения HPV16-L1 [Bazan S.B. et al., Arch. Virol. 2009, 154(10), 1609-17]. В геном P. pastoris был введен ген, кодирующий HPV16-L1, под контролем регулируемого метанолом промотора. Этот ген имел оптимизированный кодоновый состав. Очистку белка L1 выполняли с использованием гепарин-сефарозной хроматографии с последующей стадией сборки вирусоподобных частиц. Образование биологически активных VLP подтверждали методами иммуноэлектронной микроскопии и гемагглютинации.
Известна вакцина для создания иммунного ответа против папилломавируса человека, включающая вирусоподобные частицы HPV16, HPV18 и, по меньшей мере, одного из типов HPV31, HPV45, HPV52 (RU 2420313, 2010). VLP получали путем экспрессии в клетках Trichoplusia ni, инфицированных рекомбинантным бакуловирусом, кодирующим соответствующий ген L1 HPV. После очистки каждый тип VLP независимо адсорбировали на гидроокиси алюминия с получением адсорбированных концентрированных моновалентов, которые затем смешивали в нужном соотношении. Дозировка компонентов вакцины варьирует в зависимости от состояния, пола, возраста, массы индивидуума, пути введения и может составлять 1-100 мкг каждой VLP.
В патенте US 7,709,010, 04.05.2010 описано создание поливалентной вакцины против папилломавирусной инфекции, включающей VLP L1 разных типов HPV, в том числе и типов 6, 11. VLP получены путем экспрессии капсидного белка L1 в рекомбинантных дрожжевых клетках S.cerevisiae. Выделенные и очищенные VLP адсорбировали на алюминиевом адъюванте и адъюванте ISCOM-типа. Преимущество системы экспрессии на основе дрожжей S.cerevisiae заключается в относительно низкой себестоимости и легкой адаптации к крупномасштабному росту в ферментерах.
Известна рекомбинантная вакцина для профилактики папилломавирусной инфекции, содержащая в качестве антигена L1 VLP HPV типа 16 и 18, и/или 56 (RU 2546213, 13.02.2015). VLP HPV получали путем культивирования рекомбинантных клеток дрожжей H. Polymorpha.
Большая часть известных из уровня техники вакцин для профилактики папилломавирусной инфекции еще не прошла необходимые клинические испытания. В настоящее время существуют две вакцины, разрешенные к применению: квадривалентная профилактическая вакцина Гардасил производства компании Merck Sharp & Dohme B.V. и бивалентная вакцина Церварикс производства компании Glaxo SmithKline Biologicals S.A.
Гардасил (ЛС-002293, 24.11.06) - рекомбинантная квадривалентная вакцина, состоящая из вирусоподобных частиц HPV 6, 11, 16, 18 типов. VLP получены путем экспрессии вирусных капсидных белков L1 в дрожжевых клетках Saccharomyces cerevisiae, очищены и адсорбированы на алюминиевом адъюванте (аморфный гидроксифосфатсульфат алюминия). Каждая доза вакцины (0,5 мл) содержит рекомбинантные антигены: тип 6 - 20 мкг, тип 11 - 40 мкг, тип 16 - 40 мкг, тип 18 - 20 мкг, а также 225 мкг алюминиевого адъюванта и буферный раствор. Рекомендуемый курс вакцинации состоит из трех доз (0 -2 - 6).
Церварикс (ЛСР - 006423/08, 11.08.08) - рекомбинантная бивалентная вакцина, состоящая из вирусоподобных частиц HPV 16 и 18 типов. VLP получены с использованием рекомбинантных бакуловирусов на культуре клеток Trichoplusia ni. Каждая доза вакцины (0,5 мл) содержит рекомбинантные антигены: тип 16-20 мкг, тип 18 - 20 мкг, МФЛ(3-О-дезацил-4-монофосфорил липид А) - 50 мкг, алюминия гидроксид - 0,5 мг.
Несмотря на то, что указанные вакцины уже используются, остается еще много неизученных вопросов, связанных с их применением. Так, не исследованы отдаленные результаты вакцинации, отсутствуют данные о корреляции образующихся антител с защитным эффектом. Отмечается также, что при вакцинации продуцируется большое число специфичных антител, однако не все антитела способны нейтрализовать вирус папилломы. К недостаткам вакцин можно отнести реактогенность, побочные явления, ограничения в применении, дороговизну.
Ближайшим аналогом является патент RU 2206608, 2003, С 12 N 7/ 00, в котором описывается способ получения вирусоподобных частиц белка L1 НРV или L2 НРV разных типов. Способ включает конструирование экспрессионного плазмидного дрожжевого вектора, содержащего последовательность ДНК, кодирующую соответствующий полипептид, под контролем промотора GAL10 и трансформирование им клетки Saccharomyces cerevisiae, культивирование трансформированной клетки, сбор VLP из трансформированной клетки и очистку VLP с помощью хроматографии. Способность рекомбинантных клеток S. cerevisiae экспрессировать белки L1 или L2 НРV анализировали иммуноблотингом. Для создания вакцины против папилломавирусной инфекции очищенные VLP смешивали с фармацевтически приемлемым носителем, стабилизатором или адъювантом. Вакцина содержит 0,1-100 мкг, предпочтительно 1-20 мкг активного антигена. К недостаткам использования системы экспрессии на основе Saccharomyces cerevisiae можно отнести нестабильность рекомбинантных клеток, связанную с относительно высокой частотой потерь автономной плазмиды при делении клеток. При культивировании штамма в неселективных для плазмиды условиях это приводит к накоплению клеток, не содержащих плазмиды и неспособных к синтезу целевого белка. Это существенно осложняет культивирование рекомбинантного штамма и увеличивает себестоимость целевого продукта. Для индукции экспрессии рекомбинантных белков используется достаточно дорогой реагент - галактоза, что увеличивает себестоимость целевого продукта.
Задачей изобретения является создание высокоиммуногенной, нетоксичной, не обладающей побочными эффектами рекомбинантной вакцины для профилактики папилломавирусной инфекции, содержащей в качестве антигена L1 VLP HPV типа 16 , 18, 6, 11, а также способ ее получения. Задача решена за счет создания рекомбинантных штаммов дрожжей H. Polymorpha, стабильно и эффективно продуцирующих главный капсидный белок вируса папилломы человека типа 11, 6, соответственно, обладающий необходимыми биологическими свойствами (правильная конформация, иммуногенность, чистота) и низкой себестоимостью. VLP L1 типа 16 и 18 получали согласно патенту RU 2546213. Дрожжевой продуцент должен обеспечивать возможность контролировать отсутствие мутаций в экспрессируемом чужеродном гене, иметь контролируемый уровень экспрессии рекомбинантного гена, чтобы обеспечить высокую эффективность укладки синтезируемого полипептида с образованием природной конформации получаемого продукта, иметь достаточно высокий выход целевого белка, обеспечивающий экономическую целесообразность использования продуцента.
Для получения рекомбинантного штамма-продуцента главного капсидного белка L1 HPV11 в геном реципиентного штамма DLT2 H. polymorpha последовательно интегрируют две экспрессионные кассеты, содержащие по одной копии нужного гена. Первая кассета содержит фрагмент ДНК с рекомбинантным геном, кодирующим HPV11-L1, под контролем промотора гена MOX (метанолоксидаза) и селективный маркер (ген TRP3 H. polymorpha). Вторая экспрессионная кассета содержит аналогичный ген HPV11-L1, но под контролем промотора гена DAK (ген кодирует дигидроксиацетон киназу) дрожжей H. polymorpha и селективный маркер (ген LEU2 S. cerevisiae). В результате трансформации получают рекомбинантный штамм, содержащий одну копию гена HPV11-L1 под контролем промотора MOX и одну копию гена HPV11-L1 под контролем промотора DAK. При этом искусственные гены интегрированы непосредственно в геном штамма-продуцента, т.е. находятся в составе хромосом дрожжей H. polymorpha. Это обеспечивает высокую митотическую стабильность штамма, давая возможность оптимизировать условия его культивирования без учета риска накопления клеток, потерявших способность синтезировать чужеродный белок. Кроме того, наличие только одной копии рекомбинантного гена в каждой экспрессионной кассете с уникальной промоторной и терминальной областью позволяет контролировать отсутствие мутаций этого гена в процессе его введения в геном и при культивировании штамма. Контроль за отсутствием мутаций осуществляют путем анализа нуклеотидной последовательности продуктов полимеразной цепной реакции с использованием праймеров, уникальных к специфическим последовательностям в кодирующей области каждой экспрессионной кассеты.
Преимуществом предложенного изобретения также является возможность получения стабильно высокого уровня синтеза целевого белка без нарушения эффективности его укладки. Использование экспрессионных кассет с двумя разными промоторами (MOX и DAK), максимальная активность которых достигается при разных условиях культивирования, обеспечивает большую равномерность синтеза продукта в процессе культивирования штамма. При этом не превышается уровень синтеза белка, который может привести к возможному нарушению эффективности укладки и, как следствие, снижению продуктивности штамма.
В результате отбора клонов, способных синтезировать белок HPV11-L1, был выделен трансформант, наиболее отвечающий требуемым свойствам. Полученный рекомбинантный штамм позволяет добиваться более высоких уровней синтеза рекомбинантного белка при более простых способах получения культур высокой плотности, а также выделять белок HPV11-L1 в виде VLP с правильной конформацией, обладающий необходимыми антигенными и иммуногенными свойствами, с достаточно высоким выходом. Образование VLP подтверждали с использованием просвечивающей электронной микроскопии.
Штамм депонирован 21.11.2017 в коллекции ЗАО НПК «Комбиотех» под номером КБТ-17/рPV-112. Штамм является новым и ни в патентной, ни в научно-технической литературе не описан.
Аналогичным образом был получен рекомбинантный штамм дрожжей H. polymorpha, продуцирующий VLP HPV6-L1. Штамм депонирован 21.11.2017 в коллекции ЗАО НПК «Комбиотех» под номером КБТ-17/рPV-062. Штамм является новым и ни в патентной, ни в научно-технической литературе не описан.
Полученные рекомбинантные дрожжевые штаммы, продуцирующие главный капсидный белок HPV типа 6,11, соответственно, культивировали раздельно в условиях, подходящих для экспрессии главного капсидного белка HPV. После выделения и очистки рекомбинантные антигены использовали в составе вакцины для профилактики папилломавирусной инфекции человека. Рекомбинантная вакцина содержит в виде VLP белок L1 типа 16 и 18, 6, 11 в эффективном количестве, адьювант, физиологически приемлемый разбавитель и может содержать мертиолят в качестве консерванта, а также L- гистидин и Твин-80 в качестве стабилизатора. VLP L1 типа 16 и 18 получали согласно RU 2546213. Выделенные и очищенные VLP L1 типа 16 и 18 не имели мутаций, имели чистоту не менее 95%, обладали антигенной активностью и иммунной специфичностью. Очищенные VLP L1 4-х типов адсорбировали на адъюванте (гидроокись алюминия) и смешивали в нужном соотношении, или очищенные VLP первоначально смешивали в нужном соотношении, а затем адсорбировали на адъюванте. Под эффективным количеством понимается такое количество активного начала настоящего изобретения, которое достаточно для защиты от инфекции HPV. Точное количество будет зависеть от конкретных обстоятельств и может быть оценено специалистом в данной области с использованием известных методик.
Технический результат предложенного изобретения заключается в повышении иммунногенности вакцины (улучшении биологических свойств: антигенность, иммуногенность, правильная сборка VLP), снижении себестоимости, а также в создании вакцины, направленной против распространенных на территории РФ типов HPV 16,18,6,11.
Изобретение может быть проиллюстрировано следующими примерами.
В приводимых примерах все генно-инженерные операции производили согласно стандартным методикам и инструкциям компаний производителей ферментов и наборов для манипуляций с ДНК in vitro. Трансформацию клеток Escherichia coli и Hansenula polymorpha осуществляли согласно ранее описанным методам (Inoue et al, 1990, Gene, 96:23-28. и Bogdanova et al., 1995, Yeast 11:343-353, соответственно). Синтез фрагментов ДНК, а также определение последовательности нуклеотидов производились ЗАО “Евроген” г. Москва. Были использованы синтетические фрагменты ДНК, нуклеотидные последовательности которых приведены в Списке последовательностей.
Пример 1. Получение плазмидного вектора p11HPV3, содержащего рекомбинантный ген DAK-HPV11-L1 и селективный маркер ген LEU2 S. cerevisiae.
Фрагмент ДНК DAK-HPV11 (SEQ ID NO: 1) и препарат плазмиды AMIpSL1 (Agaphonov et al., 1999, Yeast 15:541-551), содержащей ген LEU2 S. cerevisiae в качестве дрожжевого селективного маркера, был гидролизован рестриктазами BsrGI и HindIII. Полученные препараты фрагмента ДНК с рекомбинантным геном и линеаризованной плазмиды AMIpSL1 внесли в реакционную смесь для проведения лигирования в концентрации 30 нг/мкл. После инкубации с Т4 ДНК лигазой в течение 2 часов, 2 мкл реакционной смеси использовали для трансформации штамма E. coli DH5α. Из нескольких выросших трансформантов выделили плазмидную ДНК. На основании рестрикционного анализа была отобрана плазмида, содержащая фрагмент DAK-HPV11. Определение последовательности нуклеотидов этой плазмиды подтвердило, что она содержит ген, кодирующий HPV11-L1 под контролем промотора DAK H. polymorpha. Полученную плазмиду обозначили p11HPV3.
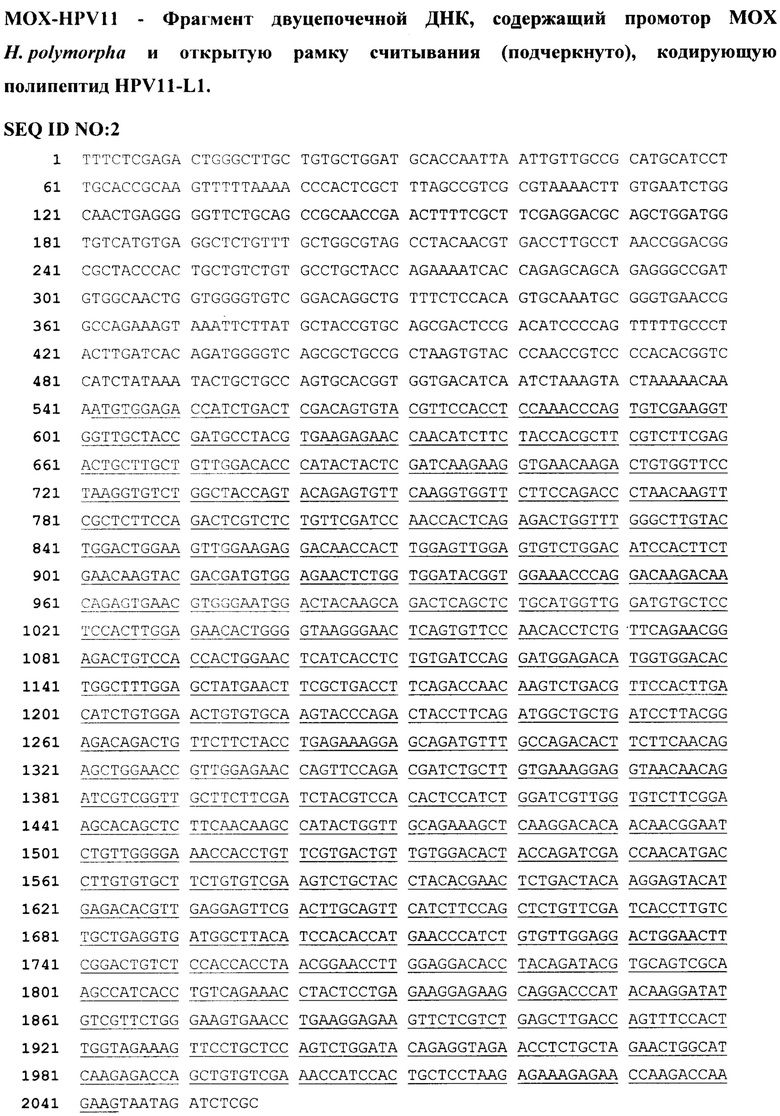
Пример 2. Получение плазмидного вектора p11HPV2, содержащего рекомбинантный ген MOX-HPV11-L1 и неполный ген TRP3 H. polymorpha в качестве селективного маркера.
Фрагмент ДНК MOX-HPV11 (SEQ ID NO: 2) и препарат плазмиды pTZ-MOX (Agaphonov et al., 1995, Yeast, 11:1241-1247), содержащей неполный ген TRP3 H. polymorpha в качестве дрожжевого селективного маркера, были гидролизованы рестриктазами BglII и XhoI. Продукт гидролиза фрагмента MOX-HPV11 и продукт гидролиза плазмиды pTZ-MOX длиной 4.3 т.п.н. были выделены после электрофоретического разделения в агарозном геле. Полученные препараты ДНК внесли в реакционную смесь для проведения лигирования в концентрации 30 нг/мкл. После инкубации с ДНК лигазой Т4 в течение 2 часов, 2 мкл реакционной смеси использовали для трансформации штамма E. coli DH5α. Из нескольких выросших трансформантов выделили плазмидную ДНК. На основании рестрикционного анализа была отобрана плазмида, содержащая фрагмент MOX-HPV11. Определение последовательности нуклеотидов этой плазмиды подтвердило, что она содержит ген, кодирующий HPV11-L1 под контролем промотора MOX H. polymorpha. Полученную плазмиду обозначили p11HPV2.
Пример 3. Получение штамма H. polymorpha, содержащего экспрессионную кассету HPV11-L1 с промотором МОХ.
Штамм H. polymorpha DLT2 [Agaphonov et al., 1994, Yeast v10, pp.509-513; коллекция штаммов лаборатории молекулярной генетики РКНПК МЗ РФ] трансформировали плазмидой p11HPV2, гидролизованной рестриктазами Ecl136II и XhoI. Трансформантов отбирали на среде, содержащей 6.7 г/л смеси солей и витаминов "Yeast Nitrogen Base" (Difco), 20 г/л D-глюкозы, 20 г/л агара и 60 мг/л лейцина. Среди полученных трансформантов были отобраны клоны, способные продуцировать HPV11-L1. Один из таких трансформантов был обозначен DLT2/p11HPV2.
Пример 4. Получение штамма H. polymorpha, содержащего две экспрессионные кассеты HPV11-L1, одна из которых с промотором МОХ, а другая с промотором DAK.
Штамм DLT2/p11HPV2 (пример 3) трансформировали плазмидой p11HPV3, гидролизованной рестриктазой BsrGI. Трансформантов отбирали на среде, содержащей 6.7 г/л смеси солей и витаминов "Yeast Nitrogen Base" (Difco), 20 г/л D-глюкозы и 20 г/л агара. Один из полученных трансформантов был обозначен КБТ17/pPV-112. У этого трансформанта было подтверждено наличие интактного гена MOX-HPV11 (см. пример 5) и интактного гена DAK-HPV11 (см. пример 6).
Пример 5. Определение наличия интактного гена МОХ-HPV11 в геноме трансформантов H. polymorpha.
Из анализируемых клонов получали препараты геномной ДНК и использовали в качестве матрицы для проведения полимеразной цепной реакции (ПЦР) с олигонуклеотидами MOX20 (SEQ ID NO:3) и MOX3’TRP3 (SEQ ID NO:4) в качестве праймеров. ПЦР проводили с использованием полимеразы Pfu (Fermentas). При наличии в геноме гена МОХ-HPV11 образовывался продукт ПЦР длиной 1696 п.н. Для подтверждения интактности этого гена, определяли последовательность нуклеотидов полученного продукта ПЦР.
Пример 6. Определение наличия интактного гена DAK-HPV11 в геноме трансформантов H. polymorpha.
Из анализируемых клонов получали препараты геномной ДНК и использовали в качестве матрицы для проведения полимеразной цепной реакции (ПЦР) с олигонуклеотидами dakFw1 (SEQ ID NO:5) и AMI20 (SEQ ID NO:6) в качестве праймеров. ПЦР проводили с использованием полимеразы Pfu (Fermentas). При наличии в геноме гена DAK-HPV11 образовывался продукт ПЦР длиной 1718 п.н. Для подтверждения интактности этого гена, определяли последовательность нуклеотидов полученного продукта ПЦР.
Пример 7. Культивирование штамма H. polymorpha КБТ17/pPV-112, содержащего две экспрессионные кассеты HPV11-L1, одна из которых с промотором МОХ, а другая с промотором DAK.
Ферментацию штамма-продуцента дрожжей H. polymorpha осуществляли в два этапа. На первом этапе в режиме фед-бэтч при температуре 30°С наращивали биомассу в культуральной среде, содержащей 4% дрожжевого экстракта, 2% бакто-пептона и 4% глицерина. Когда плотность сырой биомассы достигала 150 г/л (примерно через 28-32 часа ферментации), добавляли индуктор до концентрации 0.5-0.8 % и поддерживали на этом уровне в течение 48-72 часов.
Пример 8. Выделение и очистка рекомбинантного белка HPV11-L1 из клеток H. polymorpha.
После ферментации штамма-продуцента белок НРV11-L1 (аминокислотная последовательность - SEQ ID NO:7) выделяли согласно опубликованной методике [Kim H.J. et al., Protein Expression and Purification, v.70, p. 68-74, 2010] с небольшими изменениями. Клетки из культуральной жидкости осаждали центрифугированием при 4000g в течение 15 минут при 4°С. Осажденную биомассу ресуспендировали до концентрации 300 г влажных клеток на литр суспензии в буфере для экстракции: PВS (рН 7.2) с добавлением 1.7mM EDTA, 2mM PMSF, 0.01% Tween-80. Далее клетки разрушали в мельнице Dyno-Mill типа KDL, используя стеклянные шары диаметром 0.5-0.7 мм. Для удаления основной массы нецелевых белков полученный клеточный лизат насыщали сульфатом аммония до 35% от насыщения и осажденные белки отделяли центрифугированием при 12000g в течение 30 минут. Осадок ресуспендировали в 9 объемах буфера PBS (pH 7.2) + 0.01% Tween-80 для достижения 5% насыщения сульфатом аммония и инкубировали в течение 24 часов при комнатной температуре для полного растворения целевого белка и осаждения части примесных белков, которые затем отделяли центрифугированием при 12000g в течение 30 минут. Полученный супернатант диализовали в буфер для связывания: РВS (рН 6.2), содержащий 0.2 M NaCl и 0.01% Tween-80 с последующей адсорбцией на сорбент SP Sepharose FF (GE Healthcare, USA). После промывки сорбента тем же буфером для связывания, белок HPV11-L1 элюировали линейным градиентом от 0.2 M до 1.5 M NaCl. Объединенные фракции, содержащие антиген (по иммуноблоту и SDS-PAGE), концентрировали ультрафильтрацией в тангенциальном потоке на мембране Pellicon Biomax 300 KDa (Millipore, USA) и разделяли при помощи зонального ультрацентрифугирования (Beckman Coulter, USA) в градиенте глицерин / сахароза. Фракции, содержащие целевой белок, объединяли, а затем очищали от глицерина и сахарозы гельфильтрацией на сорбенте Тoyopearl HW-65F (ToyoSoda Corp., Japan). Чистоту полученного таким образом рекомбинантного белка НРV11-L1 определяли методом SDS-PAGE электрофореза. Если чистота белка была недостаточна (менее 95%) производили дополнительную очистку при помощи анионообменной хроматографии на сорбенте Тoyopearl DEAE-650M (ToyoSoda Corp., Japan).
В результате этих операций удается выделить около 15-20% белка L1, присутствующего в клеточном лизате (таблица 1). Выход очищенного HPV11-L1 составлял не менее 50 мг / кг влажной биомассы дрожжей.
Таблица 1. Типичный баланс выделения и очистки HPV11-L1. Концентрация общего белка на первых стадиях определялась по Биуретовому методу. На остальных стадиях - по методу Лоури. Содержание и чистота HPV11-L1 определялась при помощи иммуноблотинга и SDS-PAGE электрофореза.
(мг/мл)
(мг)
(г)
%
%
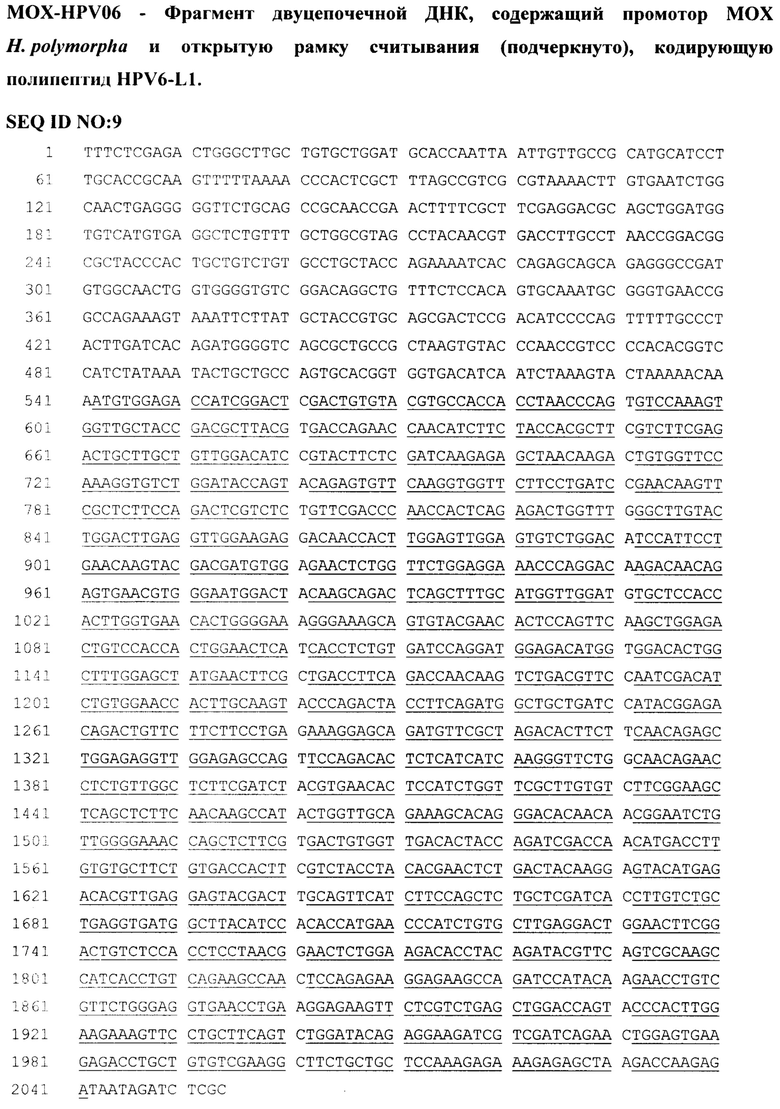
Пример 9. Получение рекомбинантного штамма дрожжей H. polymorpha КБТ-17/рPV-062, продуцирующего VLP HPV6-L1 (SEQ ID NO:10), осуществляли аналогично примерам 1-4 с использованием фрагментов ДНК DAK-HPV6 (SEQ ID NO:8) и MOX-HPV6 (SEQ ID NO:9). Наличие интактного гена DAK-HPV6 в полученных клетках подтверждали согласно примеру 6 образованием ПЦР продукта длиной 1715 п.н. Наличие интактного гена MOX-HPV6 в полученных клетках подтверждали согласно примеру 5 образованием ПЦР продукта длиной 1693 п.н. Для подтверждения интактности этого гена определяли последовательность нуклеотидов полученного продукта.
Культивирование штамма H. polymorpha КБТ-17/рPV-062, продуцирующего VLP HPV6-L1, осуществляли аналогично примеру 7. Выделение и очистку VLP HPV6 осуществляли аналогично примеру 8 (таблица 2). Выход очищенного HPV6-L1 составлял не менее 50 мг / кг влажной биомассы дрожжей.
Таблица 2. Баланс выделения и очистки HPV6-L1. Концентрация общего белка на первых стадиях определялась по Биуретовому методу. На остальных стадиях - по методу Лоури. Содержание и чистота HPV6-L1 определялась при помощи иммуноблотинга и SDS-PAGE электрофореза.
(мг/мл)
(мг)
(мг)
%
%
Пример 10. Анализ рекомбинантных антигенов HPV11-L1, HPV6-L1
1) Чистота рекомбинантного целевого белка определяется электрофорезом в полиакриламидном геле в восстанавливающих условиях (SDS-PAGE) путем окрашивания c Coomassie Brilliant Blue R-250 и анализа интенсивности полос отсканированных гелей посредством компьютерной программы NIH Image. Чистота выделенных рекомбинантных белков НРV-L1 составляет не менее 95% (фиг. 1А НРV11-L1, фиг. 2А НРV6-L1).
2) Иммуноспецифичность рекомбинантных антигенов определяют методом иммуноблотинга. Первичными антителами к белку НРV6-L1 (cat# CABT-B8785, Creative Diagnostics, USA) или белку НРV11-L1 (cat# CABT-B8787, Creative Diagnostics, USA) окрашивают иммуноблот с последующей визуализацией при помощи специфических антител к иммуноглобулинам мыши, коньюгированными с пероксидазой хрена, по методике улучшенной хемилюминисценции ЕCL (Аmersham, UK). Рекомбинантный антиген HPV-L1 представлен в виде полосы мономера белка в районе 55 кДа (фиг. 1Б НРV11-L1, фиг. 2Б НРV6-L1).
3) Образование VLP рекомбинантных антигенов НРV11-L1 и НРV6-L1 подтверждали методом просвечивающей электронной микроскопии (фиг. 3, 4). Представленные образцы были исследованы методом негативного контраста (в качестве контрастера использовали водный раствор фосфорновольфрамовой кислотой) с отмывкой дистиллированной водой, в трансмиссионном электронном микроскопе JEOL 100B (Japan) при ускоряющем напряжении 80 kV, съемка производилась при инструментальных увеличениях от 15000 до 59000. Средний размер вирусоподобных частиц составил ~50нм.
4) Иммуногенность рекомбинантных НРV-L1 определяли в тесте на мышах линии Balb/c с массой 12-14 г после однократной вакцинации антигеном, сорбированном на гидроокиси алюминия. Значение ED/50 (ED/50 - доза антигена, вызывающая сероконверсию у 50% мышей) составляет менее 100 нг, что является хорошим показателем иммуногенных свойств рекомбинантного белка.
Культурально-морфологические особенности штаммов: клетки округлой формы, небольшие по размеру, на агаризованной среде YPD образуют крупные круглые колонии с выраженной выпуклой серединой.
Хранение - при -70°С в виде суспензии клеток в стерильном 30-50%-ном растворе глицерина.
Генетические особенности: Штаммы не являются зоопатогенными или фитопатогенными.
Способ, условия и состав сред для размножения штаммов: инкубирование прокачиванием при 30°С в питательной среде состава: 2% пептона, 1% дрожжевого экстракта, 2% глюкозы.
Условия и состав среды для ферментации: прокачивание при 30°С и рН 5.0-5.5 в питательной среде, содержащей до 4 % глицерина.
Активность штамма определяли в осветленном гомогенизате клеток методом иммуноблотинга или иммуноферментного анализа. Активность штаммов - не менее 15 мг/л культуральной жидкости.
Выделенные и очищенные VLP HPV16-L1, HPV18-L1, HPV6-L1, HPV11-L1 используют для создания на их основе вакцины для профилактики папилломавирусной инфекции. VLP 4-х типов вируса папилломы смешивают в присутствии адъюванта и физиологически приемлемого разбавителя. Вакцина дополнительно может содержать консервант - мертиолят, а также стабилизаторы L-гистидин и Твин-80.
Конкретные примеры композиций вакцин для профилактики папилломавирусной инфекции:
1. Вакцина для профилактики папилломавирусной инфекции содержит:
20 мкг VLP HPV16-L1, полученного путем культивирования рекомбинантных клеток дрожжей H. polymorpha,
20 мкг VLP HPV18-L1, полученного путем культивирования рекомбинантных клеток дрожжей H. polymorpha,
20 мкг VLP HPV6-L1, полученного путем культивирования штамма КБТ-17/рPV-062,
20 мкг VLP HPV11-L1, полученного путем культивирования штамма КБТ-17/рPV-112
0.5 мг геля гидроокиси алюминия,
до 0.5 мл фосфатно-солевого буфера (ФСБ), представленного 50 mM Na-фосфатного буфера, pH 6.8-7.2 и 0.13 M NaCl.
2. Вакцина для профилактики папилломавирусной инфекции содержит:
20 мкг VLP HPV16-L1, полученного путем культивирования рекомбинантных клеток дрожжей H.polymorpha,
20 мкг VLP HPV18-L1, полученного путем культивирования рекомбинантных клеток дрожжей H.polymorpha,
20 мкг VLP HPV6-L1, полученного путем культивирования штамма КБТ-17/рPV-062,
20 мкг VLP HPV11-L1, полученного путем культивирования штамма КБТ-17/рPV-112
0.5 мг геля гидроокиси алюминия,
60 мкг мертиолята,
до 0.5 мл фосфатно-солевого буфера (ФСБ), представленного 50 mM Na-фосфатного буфера, pH 6.8-7.2 и 0.13 M NaCl.
3. Вакцина для профилактики папилломавирусной инфекции содержит:
40 мкг VLP HPV16-L1, полученного путем культивирования рекомбинантных клеток дрожжей H. polymorpha,
20 мкг VLP HPV18-L1, полученного путем культивирования рекомбинантных клеток дрожжей H.polymorpha,
20 мкг VLP HPV6-L1, полученного путем культивирования штамма КБТ-17/рPV-062,
40 мкг VLP HPV11-L1, полученного путем культивирования штамма КБТ-17/рPV-112
0.5 мг геля гидроокиси алюминия,
780 мкг L-гистидина,
50 мкг Tween-80,
до 0.5 мл фосфатно-солевого буфера (ФСБ), представленного 50 mM Na-фосфатного буфера, pH 6.8-7.2 и 0.13 M NaCl.
Согласно изобретению вакцины для профилактики папилломавирусной инфекции можно получать как с введением в их состав консерванта - мертиолята, так и без него. Это касается и стабилизаторов L-гистидин и Твин-80. Отсутствие в вакцине консерванта и стабилизаторов не сказывается на ее качестве.
Иммуногенность рекомбинантных вакцин, определенная в тесте на мышах линии Balb/c с массой 12-14 г в расчете на ED/50 (доза антигена, вызывающая сероконверсию у 50% мышей), составляет не более 100 нг.
Для сравнения была определена иммуногенность квадривалентной вакцины Гардасил (коммерчески доступная вакцина). Полученные данные показали, что относительная потенция (отношение дозы, выраженной в ED/50, аналога к ED/50 испытуемой вакцины) вакцины по изобретению в сравнении с аналогом составляет не менее 1.0.
Токсичность вакцины определяли путем введения белым мышам 0.5 мл (60 мкг) вакцины внутрибрюшинно и 0.5 мл (60 мкг) вакцины морским свинкам подкожно. Наблюдение в течение 7 суток показало, что вакцина не токсична.




| название | год | авторы | номер документа |
|---|---|---|---|
| Рекомбинантный штамм дрожжей Hansenula polymorpha - продуцент главного капсидного белка L1 вируса папилломы человека типа 6 | 2018 |
|
RU2675471C1 |
| Рекомбинантный штамм дрожжей Hansenula polymorpha - продуцент главного капсидного белка L1 вируса папилломы человека типа 11 | 2018 |
|
RU2676160C1 |
| РЕКОМБИНАНТНАЯ ВАКЦИНА ДЛЯ ПРОФИЛАКТИКИ ПАПИЛЛОМАВИРУСНОЙ ИНФЕКЦИИ ЧЕЛОВЕКА И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2014 |
|
RU2546243C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ Hansenula polymorpha - ПРОДУЦЕНТ ГЛАВНОГО КАПСИДНОГО БЕЛКА L1 ВИРУСА ПАПИЛЛОМЫ ЧЕЛОВЕКА ТИПА 18 | 2014 |
|
RU2546242C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ Hansenula polymorpha - ПРОДУЦЕНТ ГЛАВНОГО КАПСИДНОГО БЕЛКА L1 ВИРУСА ПАПИЛЛОМЫ ЧЕЛОВЕКА ТИПА 56 | 2014 |
|
RU2546240C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ Hansenula polymorpha - ПРОДУЦЕНТ ГЛАВНОГО КАПСИДНОГО БЕЛКА L1 ВИРУСА ПАПИЛЛОМЫ ЧЕЛОВЕКА ТИПА 16 | 2014 |
|
RU2546241C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ PICHIA ANGUSTA - ПРОДУЦЕНТ КАПСИДНОГО БЕЛКА L1 ВИРУСА ПАПИЛЛОМЫ ЧЕЛОВЕКА ТИПА 18 | 2011 |
|
RU2445358C1 |
| РЕКОМБИНАНТНЫЙ ШТАММ ДРОЖЖЕЙ PICHIA ANGUSTA - ПРОДУЦЕНТ КАПСИДНОГО БЕЛКА L1 ВИРУСА ПАПИЛЛОМЫ ЧЕЛОВЕКА ТИПА 16 | 2011 |
|
RU2445357C1 |
| РЕКОМБИНАНТНАЯ ВАКЦИНА ДЛЯ ПРОФИЛАКТИКИ ВИРУСНОГО ГЕПАТИТА Е У ЖИВОТНЫХ | 2012 |
|
RU2501568C1 |
| РЕКОМБИНАНТНАЯ ВАКЦИНА ДЛЯ ПРОФИЛАКТИКИ ВИРУСНОГО ГЕПАТИТА В (ВАРИАНТЫ) | 2015 |
|
RU2603729C2 |



Изобретение относится к биотехнологии, а именно генной инженерии. Предложена рекомбинантная вакцина для профилактики папилломавирусной инфекции человека и способ ее получения. Вакцина создана на основе VLP главного капсидного белка L1 HPV типов 16, 18, 6, 11, полученных путем культивирования рекомбинантных штаммов дрожжей Hansenula polymorphа. Рекомбинантные штаммы, продуцирующие VLP HPV11-L1 и VLP HPV6-L1, были получены путем введения в геном клетки дрожжей одной копии последовательности ДНК, кодирующей капсидный белок L1 НРV типа 11 или 6 под контролем промотора ДАК, и одной копии последовательности ДНК, кодирующей капсидный белок L1 НРV типа 11 или 6 под контролем промотора МОХ. Вакцина содержит эффективное количество VLP HPV11-L1, HPV6-L1, HPV16-L1 и HPV18-L1, адъювант и физиологически приемлемый разбавитель. Предложенный способ позволяет получать высокоиммуногенную, нетоксичную, не обладающую побочными эффектами вакцину. 2 н. и 4 з.п. ф-лы, 4 ил., 2 табл., 3 пр.
1. Способ получения вакцины для профилактики папилломавирусной инфекции человека, включающий трансформацию дрожжевой клетки молекулой ДНК, кодирующей L1 HPV, получение рекомбинантных штаммов, продуцирующих VLP HPV-L1 разных типов, раздельное культивирование полученных штаммов - продуцентов в подходящих условиях, выделение и очистку VLP HPV-L1, смешивание эффективного количества VLP HPV-L1 разных типов с адъювантом и физиологически приемлемым разбавителем, и получение целевого продукта, отличающийся тем, что дрожжевую клетку Hansenula polymorpha трансформируют последовательно экспрессионной кассетой, содержащей фрагмент ДНК, кодирующий HPV11-L1, под контролем промотора гена МОХ (SEQ ID NO:2) и неполный ген TRP3 H. рolymorpha, и экспрессионной кассетой, содержащей фрагмент ДНК, кодирующий HPV11-L1, под контролем промотора гена DАК (SEQ ID NO:1) и селективный маркер ген LEU2 S. сerevisiae, получают рекомбинантный штамм КБТ17/pPV-112, продуцирующий VLP HPV11-L1, дрожжевую клетку Hansenula polymorpha трансформируют последовательно экспрессионной кассетой, содержащей фрагмент ДНК, кодирующий HPV6-L1, под контролем промотора гена МОХ (SEQ ID NO:9) и неполный ген TRP3 H. рolymorpha, и экспрессионной кассетой, содержащей фрагмент ДНК, кодирующий HPV6-L1, под контролем промотора гена DАК (SEQ ID NO:8) и селективный маркер ген LEU2 S. сerevisiae, получают рекомбинантный штамм КБТ-17/рPV-062, продуцирующий VLP HPV6-L1, выделенные и очищенные VLP HPV11-L1 и VLP HPV6-L1 смешивают с VLP HPV16-L1 и VLP HPV18-L1.
2. Способ по п. 1, отличающийся тем, что дополнительно добавляют мертиолят.
3. Способ по п. 1, отличающийся тем, что дополнительно добавляют L-гистидин и Твин - 80.
4. Рекомбинантная вакцина для профилактики папилломавирусной инфекции человека, содержащая эффективное количество антигена HPV, адъювант и физиологически приемлемый разбавитель, отличающаяся тем, что в качестве антигена HPV рекомбинантная вакцина содержит смесь VLP HPV11-L1, полученной путем культивирования рекомбинантного штамма дрожжей Hansenula polymorpha
КБТ17/pPV-112, содержащего интегрированный в геном фрагмент ДНК, кодирующий HPV11-L1, под контролем промотора гена DАК (SEQ ID NO:1), и фрагмент ДНК, кодирующий HPV11-L1, под контролем промотора МОХ (SEQ ID NO:2), VLP HPV6-L1, полученной путем культивирования рекомбинантного щтамма дрожжей Hansenula polymorpha КБТ-17/рPV-062, содержащего интегрированный в геном фрагмент ДНК, кодирующий HPV6-L1, под контролем промотора гена DАК (SEQ ID NO:8), и фрагмент ДНК, кодирующий HPV6-L1, под контролем промотора МОХ (SEQ ID NO:9), VLP HPV16-L1 и VLP HPV18-L1.
5. Рекомбинантная вакцина для профилактики папилломавирусной инфекции по п. 4, отличающаяся тем, что дополнительно содержит мертиолят.
6. Рекомбинантная вакцина для профилактики папилломавирусной инфекции по п. 4, отличающаяся тем, что дополнительно содержит L-гистидин и Твин - 80.
| Рекомбинантный штамм дрожжей Hansenula polymorpha - продуцент главного капсидного белка L1 вируса папилломы человека типа 11 | 2018 |
|
RU2676160C1 |
| EP 757717 B1, 31.05.2006 | |||
| УСТРОЙСТВО ДЛЯ ПОДВЕСКИ И ТРАНСПОРТИРОВКИ ПЛОТОВ | 1976 |
|
SU595935A1 |
| US 9623098 B2, 18.04.2017 | |||
| US 8039001 B2, 18.10.2011 | |||
| US 20080248062 A1, 09.10.2008 | |||
| ВАКЦИННЫЕ СОСТАВЫ ПРОТИВ ВИРУСА ПАПИЛЛОМЫ ЧЕЛОВЕКА (HPV), СОДЕРЖАЩИЕ АЛЮМИНИЕВЫЙ АДЪЮВАНТ, И СПОСОБЫ ИХ ПОЛУЧЕНИЯ | 2012 |
|
RU2610174C2 |

