Область техники
Заявленное изобретение относится к области медицины и здравоохранения, в частности профилактики гриппа. Предложена вакцина для профилактики гриппа А, вызываемого различными вирусами птичьего гриппа А.ё
Предшествующий уровень техники
Грипп - высококонтагиозное вирусное заболевание млекопитающих (в т.ч. человека) и птиц. Хотя большинство вирусов птичьего гриппа не вызывают болезнь у людей, некоторые из них представляют отдельный интерес для медицинского и научного сообществ, так как способны инфицировать людей и имеют пандемический потенциал. По данным Всемирной Организации Здравоохранения к таким вирусам можно отнести подтип вирусов птичьего гриппа H5N1, циркулирующий в настоящее время среди домашней птицы и вызвавший с 1997 г. случаи заболевания и смерти людей. Другие подтипы вирусов птичьего гриппа, включая H7N7 и H9N2, также инфицировали людей, в том числе со смертельным исходом.
Поэтому проблема создания высокоиммуногенных и безопасных гриппозных вакцин, обеспечивающих иммунитет, способный защитить человека сразу от различных вариантов вируса птичьего гриппа А, является актуальной, с развитием современных биотехнологий возобновился интерес к созданию новых вакцин, которые могут индуцировать широкий защитный иммунитет.
Иммунизация современными гриппозными вакцинами является научно обоснованным эффективным способом массовой профилактики гриппа. Современные вакцины против вируса гриппа А вызывают образование, главным образом, штамм-специфичных антител к поверхностным гликопротеинам вируса гриппа: гемагглютинину и нейраминидазе. Однако вирусы гриппа А способны постоянно модифицировать свою структуру в результате генетических изменений. Антигенные вариации являются результатом молекулярных изменений в поверхностных белках - гемагглютинине и/или нейраминидазе. Подобная генетическая вариабельность способствует изменению антигенной структуры настолько существенно, что специфический иммунитет, выработанный на полученную вакцину, может быть не эффективен. Таким образом, вакцины, основанные на одном из поверхностных антигенов вируса гриппа, характеризуются узкой специфичностью действия, что является существенным недостатком.
Одним из наиболее перспективных антигенов вируса гриппа А для создания вакцин широкого спектра действия является белок М2, формирующий ионный канал. М2 - один из трех белков вируса гриппа А, экспрессируемых на поверхности вириона, и, в отличие от гемагглютинина и нейраминидазы, он является высоко консервативным. Это делает белок М2е хорошим кандидатом для создания вакцин широкого спектра действия. Эктодомен белка М2 вируса гриппа А является небольшим (24 а.к.) белком, причем филогенетический анализ показывает, что близкими по аминокислотному составу являются белки вирусов гриппа А различных субтипов, но циркулирующих в человеческой популяции или среди птиц. Таким образом, возможно создание консенсусных («усредненных») последовательностей М2е белка, позволяющих «перекрыть» или большинство штаммов вирусов гриппа человека, или вирусов гриппа птиц.
Известно, что применение различных консервативных антигенов вируса гриппа для создания современных генетических вакцин так же является перспективным. Такими антигенами могут быть М1 и М2 белки вируса гриппа А, нуклеопротеин NP вируса гриппа А и даже полимеразный белок РВ2. Так как изменчивость этих белков от штамма к штамму значительно ниже, чем у гликопротеинов вируса гриппа, возможно создание на их основе вакцин широкого спектра действия, защищающих от различных субтипов вируса гриппа А. В настоящее время активно ведутся разработки новых рекомбинантных вакцин на основе эктодомена белка М2 вируса гриппа.
Недостатком консервативных антигенов вируса гриппа является их низкая иммуногенность. Для повышения иммуногенности требуется применение адъювантов, в том числе и молекулярных адъювантов или индукторов врожденного иммунного ответа.
Большинство решений известных из уровня техники основано на применении экспрессионных систем на основе E. coli.
Одним из аналогов предлагаемого изобретения является вакцина против инфекции, вызываемой вирусом гриппа птиц, включающая в качестве активного агента вирусоподобные частицы на основе ядерного антигена вируса гепатита В, несущие на своей поверхности полипептиды эктодомена М2 белка вируса гриппа птиц и экспрессируемые в E. coli с дальнейшей очисткой. Вакцина продемонстрировала высокую иммуногенность, а также протективность после заражения гомологичным по М2е пептиду штаммом вируса гриппа птиц на лабораторных животных, что позволяет рассматривать ее в качестве кандидата на универсальную вакцину против вируса гриппа птиц типа А. (Патент РФ №2358981). Недостатком подобной конструкции рекомбинантной белковой молекулы можно считать наличие одной копии эктодомена М2 белка вируса гриппа птиц, что занижает иммуногенные возможности препарата, кроме того, вакцина не предназначена для профилактики у человека птичьего гриппа и защищает только от гомологичного по М2е пептиду штаммом вируса гриппа птиц (A/Duck/Potsdam 1402-6/1986 (H5N2)) после двукратной или трехкратной иммунизации.
Также известно исследование по внутримышечной иммунизации мышей рекомбинантным фъюжн-белком 4xM2e.HSP70c, состоящим из последовательных повторов белка М2е и белка HSP70 Mycobacterium tuberculosis, полученным в Е. coli, при заражении животных летальной дозой вирусов гриппа A H1N1, H3N2 или H9N2 приводит к значительному снижению падения веса, уменьшению титра вируса в легких и менее выраженному проявлению симптомов заболевания (Ebrahimi S., et. al. In contrast to conventional inactivated influenza vaccines, 4xM2e.HSP70c fusion protein fully protected mice against lethal dose of H1, H3 and H9 influenza A isolates circulating in Iran, Virology 430 (2012) 63-72). У фъюжн-белка, включающего белок HSP70 Mycobacterium tuberculosis, существенным недостатком может явиться сенсибилизация организма с последующей невозможностью проведения диагностических аллергических исследований на туберкулез из-за получения ложноположительных результатов.
Таким образом, существует необходимость в подборе более подходящих адъювантов для включения в состав фъюжн-белка. Известны исследования по повышению иммуногенности кандидатной субъединичной вакцины на основе М2 белка вируса гриппа А с помощью флагеллина - лиганда для Толл-подобного рецептора 5 (TLR5). TLR активируют систему врожденного иммунитета и во многом определяют развитие адаптивного иммунитета, использование этого механизма является современным подходом к созданию вакцин. Слитные (фъюжн) белки, полученные в результате продукции E. coli, состоящие из флагеллина и антигенов вируса гриппа, при иммунизации были эффективнее, чем просто антигены вируса гриппа [Huleatt J.W., et.al., Potent immunogenicity and efficacy of a universal influenza vaccine candidate comprising a recombinant fusion protein linking influenza M2e to the TLR5 ligand flagellin, Vaccine, 2008].
Наиболее близким решением к изобретению является композиция, содержащая в своем составе рекомбинантный фъюжн-белок из 4-х М2е и лиганда TLR5 - флагеллина, выбранного из группы: S. typhimurium fljB/STF2, Е. coli flagellin fliC, S. muenchen flagellin fliC, предназначенная для стимуляции иммунного ответа у субъекта. Данное решение принято за прототип (Заявка WO 2006/069262). Представленный в данной заявке фъюжн-белок в сущности своей является рекомбинантным, экспрессированным in vitro в культуре клеток (Drosophila Dmel-2 cells трансдуцируется плазмидой, несущей соответствующий ген фъюжн-белка). Для получения различных фармацевтических композиций наработанные таким образом рекомбинантные фъюжн-белки должны быть извлечены из культуральной жидкости и тщательно очищены многоступенчатым путем. В итоге субъекту вводят препарат, действующим веществом которого является готовый рекомбинантный фъюжн-белок. Однако у прототипа существует ряд недостатков. Получение рекомбинантных белков в условиях in vitro в больших количествах является значимой биотехнологической проблемой - сложно масштабировать процесс культивации клеточных культур и последующую многостадийную очистку наработанных целевых белков. В результате процесс получения рекомбинантных белков получается длительным, трудоемким и очень дорогостоящим. Кроме того, фармацевтические композиции, содержащие в своем составе готовые белки, обладают известными недостатками - имеют короткие сроки выведения, в качестве вакцин слабо иммуногенны и требуют двух-трехкратных введений, а также обладают повышенным аллергогенным потенциалом.
Следовательно, актуальным является поиск новых промышленно применимых технологических решений по созданию более дешевых, эффективных и безопасных противогриппозных вакцин, основанных на консервативных белках вируса птичьего гриппа А.
Раскрытие и изобретения
Технической задачей изобретения являлось создание противогриппозной вакцины широкого спектра действия на основе рекомбинантных псевдоаденовирусных частиц, экспрессирующих гены эктодоменов белка М2 вируса гриппа А, с молекулярным адъювантом, способной эффективно защищать человека от заражения вирусами птичьего гриппа типа А.
Задача решается за счет того, что создана противогриппозная вакцина широкого спектра действия против птичьего гриппа А на основе эктодомена белка М2, при этом эктодомен белка М2 экспрессируется рекомбинантными псевдоаденовирусными частицами непосредственно в организме субъекта в виде фъюжн-белка, состоящего из последовательно расположенных четырех эктодоменов белка М2 вируса гриппа А птиц и активатора TLR5 - флагеллина Salmonella enterica serovar dublin, кроме того, вакцина дополнительно содержит молекулярный адъювант и буферный раствор, взятые в эффективных количествах.
Заявленная противогриппозная вакцина содержит на дозу:
- рекомбинантные псевдоаденовирусные частицы, экспрессирующие ген фъюжн-белка -106-109 акт.ед.;
- молекулярный адъювант - 20-100 Ед;
- фармацевтически приемлемый буфер - до 0,5 мл.
В заявленной противогриппозной вакцине нуклеотидная последовательность гена фъюжн-белка, который состоит из последовательно расположенных четырех эктодоменов белка М2 и активатора TLR5 - флагеллина Salmonella enterica serovar Dublin, представлена SEQ ID NO:1. При этом нуклеотидная последователльность гена фъюжн-белка SEQ ID NO:1 кодирует аминокислотную последовательность фъюжн-белка SEQ ID NO:2.
В качестве молекулярного адъюванта заявленная вакцина содержит активатор TLR-4, а именно, кислый пептидогликан растительного происхождения с молекулярной массой от 1000 до 40000 кДа.
Одним из наиболее эффективных на сегодняшний день подходов для создания безопасных и эффективных вакцин нового поколения является использование генетических вакцин, в том числе базирующихся на рекомбинантных аденовирусных векторах. При введении в организм таких вакцин происходит попадание генетического материала в клетки организма и экспрессия в них генов целевых белков патогена. В результате антигены соответствующих патогенов распознаются иммунной системой, что приводит к индукции как гуморального, так и клеточного иммунного ответа. На сегодняшний момент наиболее перспективными и часто используемыми для создания генетических вакцин являются рекомбинантные псевдоаденовирусные частицы (РПАН), созданные на основе аденовируса человека пятого серотипа. Вакцины на основе РПАН имеют ряд преимуществ:
- РПАН являются репликативно-дефектными и не способны вызывать заболевания. Безопасность аденовирусов человека пятого серотипа с делетированными Е1 и Е3 областями генома подтверждается целым рядом проведенных клинических испытаний различных вакцинных и терапевтических препаратов на их основе [Van Kampen K.R. et.al., Vaccine. 2005];
- не требуют многократных дополнительных введений, так как экспрессия целевого антигена (фъюжн-белка) происходит непосредственно in vivo в организме иммунизированного субъекта;
- на сегодняшний момент разработаны быстрые и гибкие технологии получения РПАН, позволяющие реализацию масштабного производства различных кандидатных вакцин на их основе на одной технологической линии, без ее переоборудования и изменения регламента;
- РПАН позволяют проводить интраназальную иммунизацию и, как следствие, индуцируют образование мукозального иммунного ответа;
- по сравнению с рекомбинантными белками, получаемыми в культуре эукариотов, более дешевы и высокоиммуногенны, так как небольшая доза РПАН позволяет продуцировать в организме значительные количества белка.
Вышеперечисленные свойства делают РПАН хорошей технологической платформой для создания широкого спектра вакцин против различных патогенов.
В вакцине по изобретению реализуется еще один способ повышения иммуногенности вакцины - введение в ее состав еще одного активатора врожденного иммунного ответа - кислого пептидогликана с молекулярной массой 1000-40000 кДа. Это биологически активный сахарид, выделенный из растительного сырья и являющийся эффективным адъювантом. Действие препарата заключается в активации Толл-подобного рецептора 4 (TLR4), значительном повышении эффективности антиген-специфических и неспецифических реакций иммунной системы, в усилении иммунных механизмов защиты от инфекций. Исходя из современного уровня знаний, стимуляция врожденного иммунитета инициирует специфический иммунный ответ. Изучение TLR выявило связь между врожденным и приобретенным иммунитетом. Взаимосвязь врожденного и приобретенного иммунитета осуществляется посредством дендритных клеток, специализированных фагоцитов, сконцентрированных в селезенке, лимфоузлах и коже. Дендритные клетки, являясь антиген-презентующими клетками, ответственны за стимуляцию иммуннокомпетентных клеток. Они экспрессируют высокий уровень ко-стимуляторных молекул, необходимых для активации Т-лимфоцитов, что является началом специфического иммунитета.
Краткое описание фигур
На фиг. 1 представлена нуклеотидная последовательность созданного гена фъюжн-белка (SEQ ID NO:1).
На фигуре 2 представлена аминокислотная последовательность (SEQ ID NO:2) созданного фъюжн-белка, кодируемая геном SEQ ID NO:1.
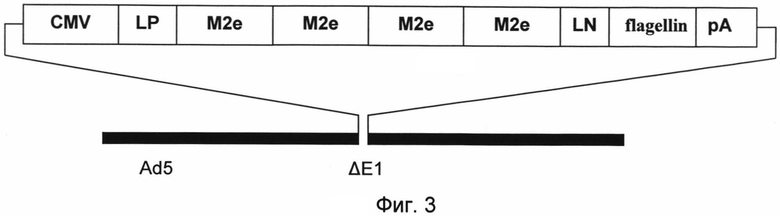
На фигуре 3 представлена схема генома РПАН, несущих генетическую конструкцию, экспрессирующую ген фъюжн-белка, содержащий гены эктодоменов М2 белка и флагеллина (SEQ ID NO:1).
Изображены составные части конструкции:
CMV - промотор цитомегаловируса человека,
LP - ген лидерного пептида секретируемой щелочной фосфатазы SEAP,
М2е - консенсусная нуклеотидная последовательность эктодомена М2 белка вируса гриппа А (в четырех последовательных повторах),
LN - линкер,
flagellin - консервативные N и С-концевые домены белка флагеллина Salmonella enterica serovar Dublin,
pA - сигнал полиаденилирования SV40 (Simian vacuolating virus 40),
Ad5 - геномная часть аденовируса человека 5 серотипа,
ΔЕ1 - делеция Е1 области аденовирусного генома.
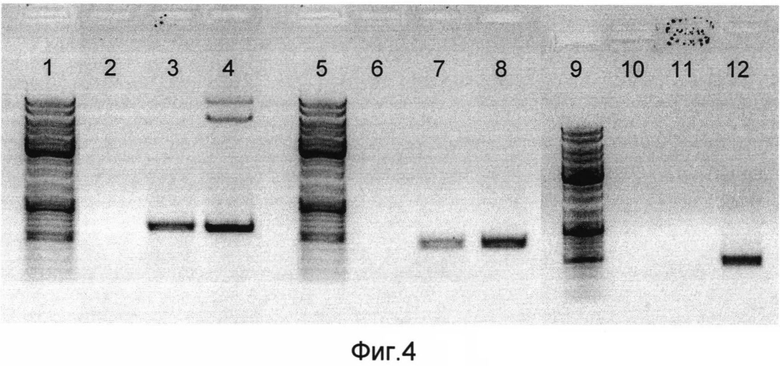
На фигуре 4 представлены результаты ПЦР-анализа ДНК рекомбинантной псевдоаденовирусной частицы на основе аденовируса человека 5 серотипа (РПАН) со вставкой гена фъюжн-белка (SEQ ID NO:1).
Положительным результатом является темная полоса на дорожке геля.
На дорожках в зависимости от номера отображено:
1, 5, 9 - маркер молекулярного веса (10000 п.н.);
2, 6, 10 - отрицательный контроль;
3 - ДНК РПАН, ПЦР с праймерми, комплементарными гену эктодомена М2 белка вируса гриппа и гену флагеллина;
4 - положительный контроль, ПЦР с праймерми, комплементарными гену эктодомена М2 белка вируса гриппа и гену флагеллина (pShuttle-CMV-M4-FI);
7 - ДНК РПАН, ПЦР с праймерми, комплементарными гену гексона аденовируса человека пятого серотипа;
8 - положительный контроль, ПЦР с праймерми, комплементарными гену гексона аденовируса человека пятого серотипа (pAd-Easy);
11 - ДНК РПАН, ПЦР с праймерми, комплементарными Е1 области генома аденовируса человека пятого серотипа;
12 - положительный контроль, ПЦР с праймерми, комплементарными Е1 области генома аденовируса человека пятого серотипа (геном аденовируса «дикого типа»).
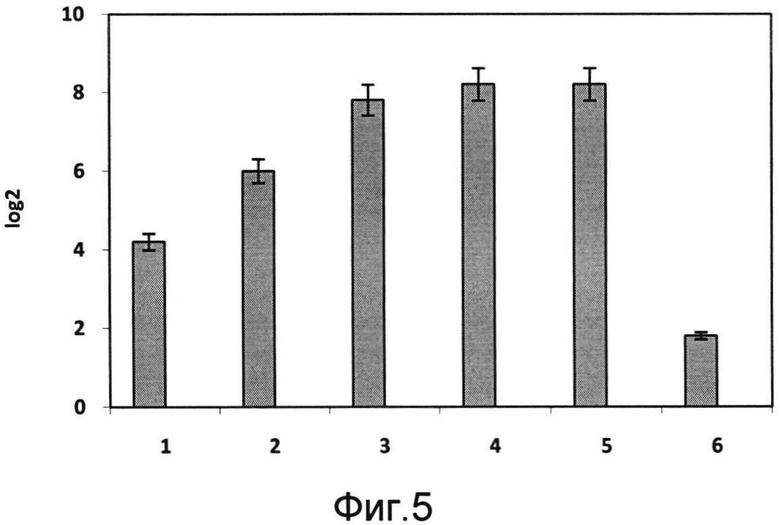
На фигуре 5 показаны уровни специфических антител к М2е вируса гриппа A H5N2 в сыворотках крови мышей, интраназально иммунизированных:
1 группа - РПАН в дозе 108 акт.ед. без адъюванта;
2 группа - РПАН в дозе 108 акт.ед.+10 ЕД адъюванта;
3 группа - РПАН в дозе 108 акт.ед.+20 ЕД адъюванта;
4 группа - РПАН в дозе 108 акт.ед.+100 ЕД адъюванта;
5 группа - РПАН в дозе 108 акт.ед.+110 ЕД адъюванта;
6 группа - отрицательный контроль, 0,9% NaCl.
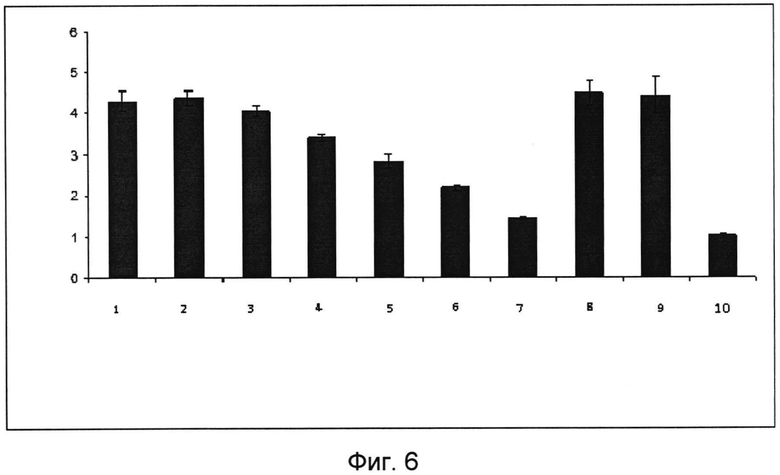
На фигуре 6 изображены диаграммы оптической плотности растворов лизированных клеток, экспрессирующих Толл-подобный рецептор 4 (HEK293-hTLR4) после добавления к ним кислого пептидогликана (КПГ) в различных концентрациях, а также положительные и отрицательные контроли.
Изображены столбцы:
1 - кислый пептидогликан в концентрации 15 мкг/мл
2 - кислый пептидогликан в концентрации 5 мкг/мл
3 - кислый пептидогликан в концентрации 1,67 мкг/мл
4 - кислый пептидогликан в концентрации 566 нг/мл
5 - кислый пептидогликан в концентрации 185 нг/мл
6 - кислый пептидогликан в концентрации 61,7 нг/мл
7 - кислый пептидогликан в концентрации 20,6 нг/мл
8 - липополисахарид (положительный контроль) 10 мкг/мл
9 - липополисахарид (положительный контроль) 1 мкг/мл
10 - Отрицательный контроль (К-) - HEK293-hTLRnull.
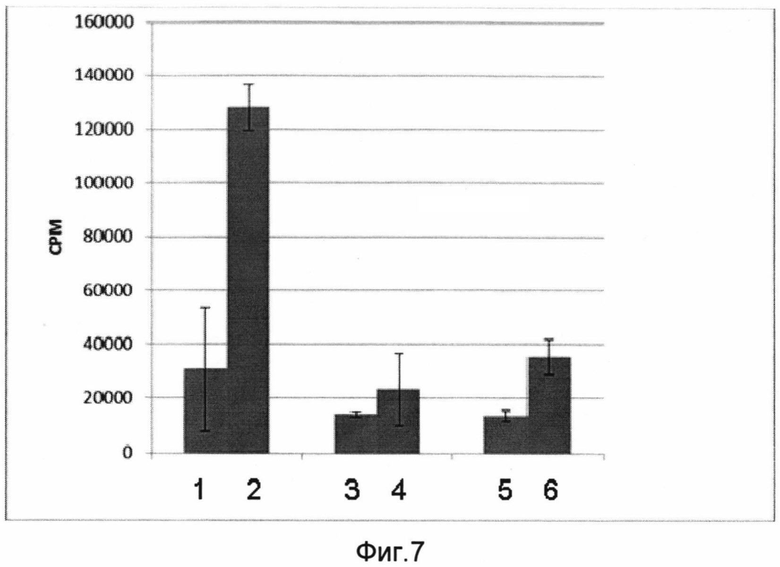
На фигуре 7 показаны результаты определения уровня лимфопролиферации в культуре клеток лимфоузлов иммунизированных мышей в ответ на добавление антигена.
Столбцы:
1 - уровень лимфопролиферации в группе, получавшей вакцину без последующей стимуляции культуры клеток антигеном;
2 - уровень лимфопролиферации в группе, получавшей вакцину с последующей стимуляцией культуры клеток антигеном;
3 - уровень лимфопролиферации в группе, получавшей фосфатный буфер без последующей стимуляции культуры клеток антигеном (отрицательный контроль);
4 - уровень лимфопролиферации в группе, получавшей фосфатный буфер с последующей стимуляцией культуры клеток антигеном (отрицательный контроль);
5 - уровень лимфопролиферации в группе, получавшей РПАН без трансгена, без последующей стимуляции культуры клеток антигеном (отрицательный контроль);
6 - уровень лимфопролиферации в группе, получавшей РПАН без трансгена, с последующей стимуляцией культуры клеток антигеном (отрицательный контроль);
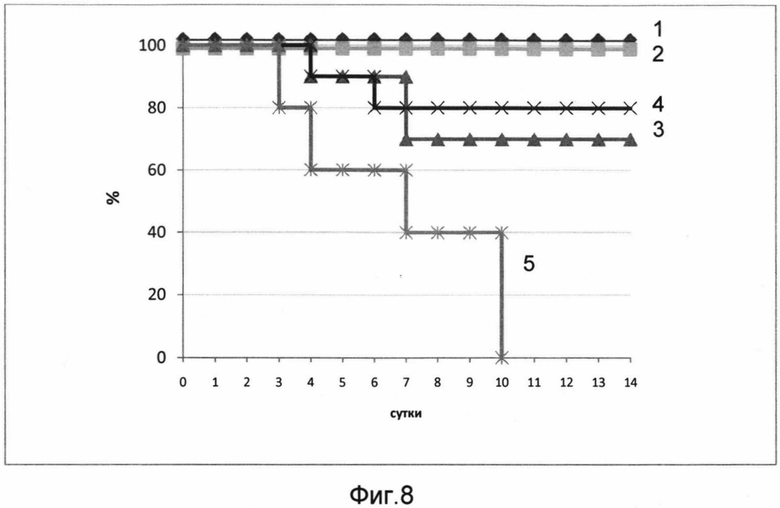
На фигуре 8 представлены кривые выживаемости мышей, получавших вакцину и контрольное вещество, зараженных 50 ЛД50 вируса гриппа A/Mallard duck/Pennsylvania/10218/84 (H5N2).
Кривые представляют данные по группам, которым вводили:
1 - 109 акт.ед. РПАН+100 ЕД адъюванта
2 - 109 акт.ед. РПАН+20 ЕД адъюванта
3 - 106 акт.ед. РПАН+20 ЕД адъюванта
4 - 106 акт.ед. РПАН+100 ЕД адъюванта
5 - контрольное вещество (буфер)
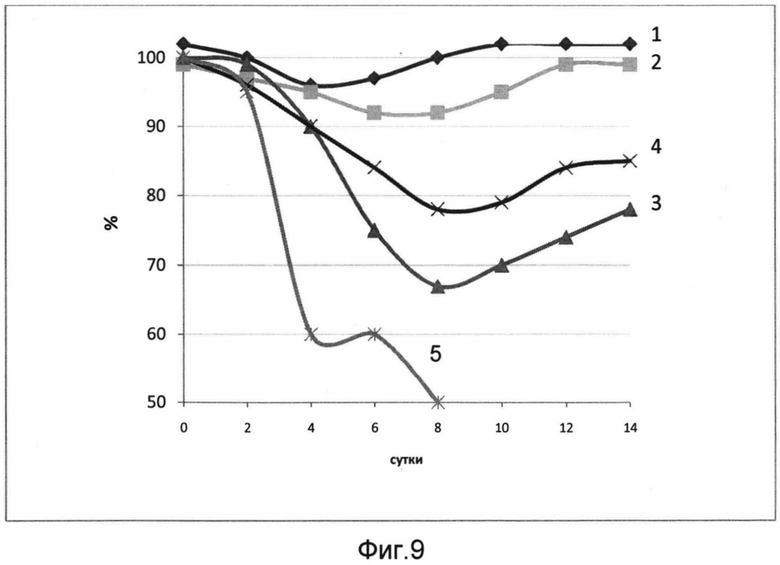
На фигуре 9 изображены кривые падения веса мышей, получавших вакцину и контрольное вещество, зараженных 50 ЛД50 вируса гриппа A/Mallard duck/Pennsylvania/10218/84 (H5N2).
Кривые представляют данные по группам, которым вводили:
1 - 109 акт.ед. РПАН+100 ЕД адъюванта
2 - 109 акт.ед. РПАН+20 ЕД адъюванта
3 - 106 акт.ед. РПАН+20 ЕД адъюванта
4 - 106 акт.ед. РПАН+100 ЕД адъюванта
5 - контрольное вещество (буфер)
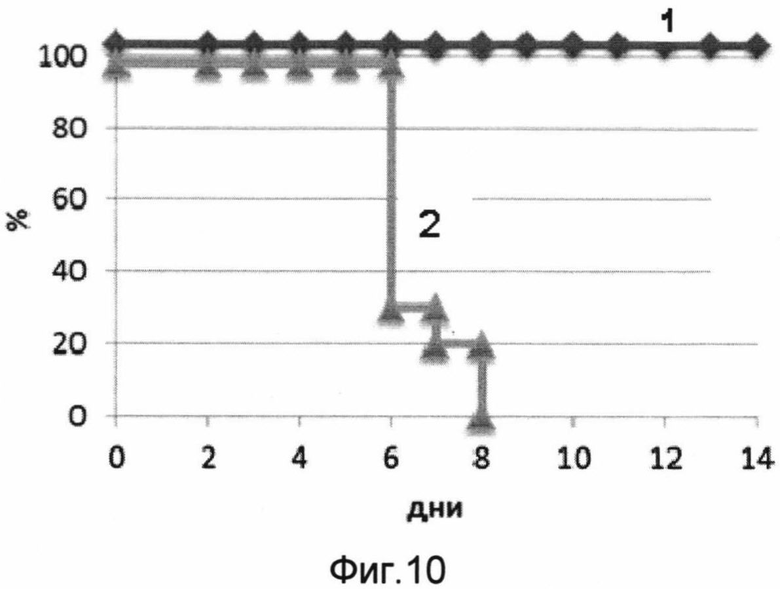
На фигуре 10 изображены кривые выживаемости мышей в течение срока наблюдения 14 дней после заражения 50 ЛД50 вируса гриппа A H5N2.
1 - кривая выживаемости мышей, получивших вакцину перед заражением.
2 - кривая выживаемости мышей, получавших буферный раствор (отрицательный контроль).
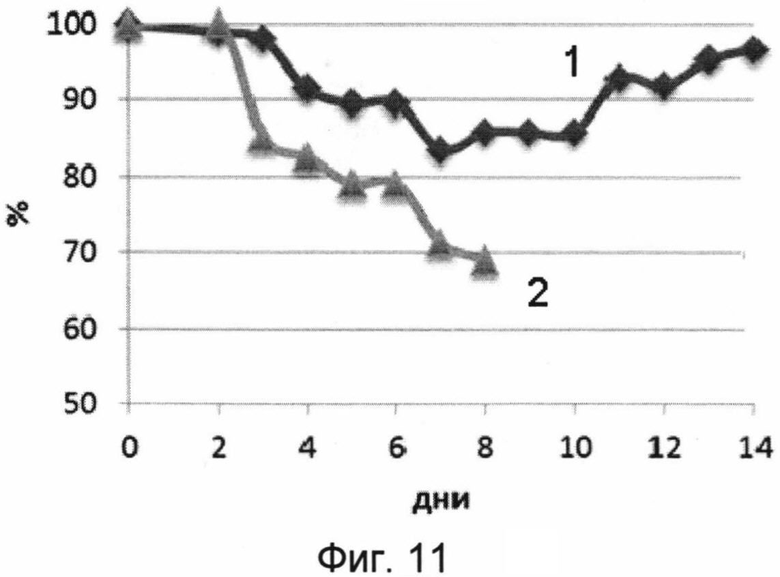
На фигуре 11 изображены кривые падения веса (суммарного в %) мышей в течение срока наблюдения 14 дней после заражения 50 ЛД50 вируса гриппа A H5N2.
1 - кривая падения веса мышей, получивших вакцину перед заражением.
2 - кривая падения веса мышей, получавших буферный раствор (отрицательный контроль).
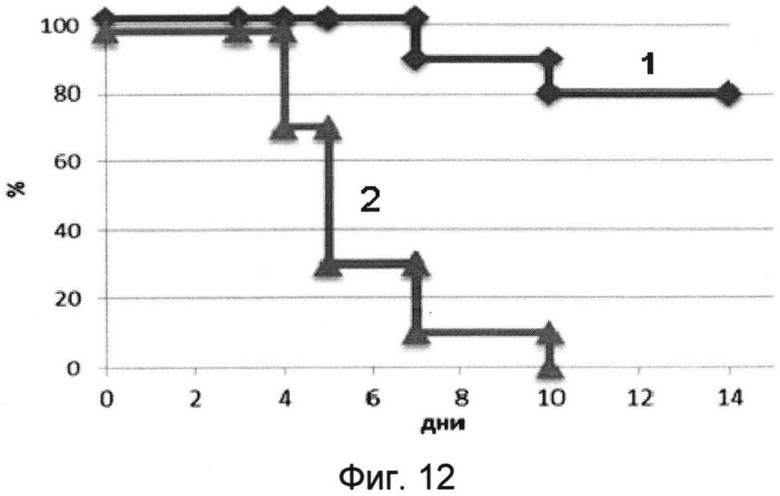
На фигуре 12 изображены кривые выживаемости мышей в течение срока наблюдения 14 дней после заражения 50 ЛД50 вируса гриппа A H2N3.
1 - кривая выживаемости мышей, получивших вакцину перед заражением.
2 - кривая выживаемости мышей, получавших буферный раствор (отрицательный контроль).
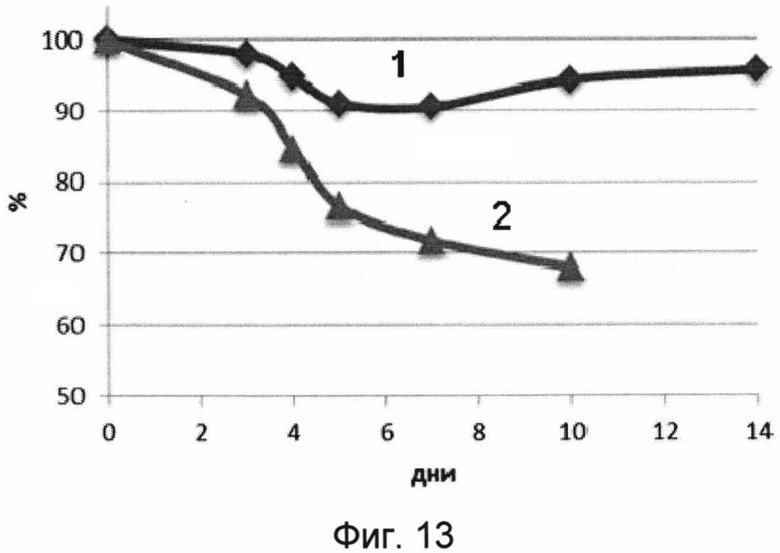
На фигуре 13 изображены кривые падения веса (суммарного в %) мышей в течение срока наблюдения 14 дней после заражения 50 ЛД50 вируса гриппа A H2N3.
1 - кривая падения веса мышей, получивших вакцину перед заражением.
2 - кривая падения веса мышей, получавших буферный раствор (отрицательный контроль).
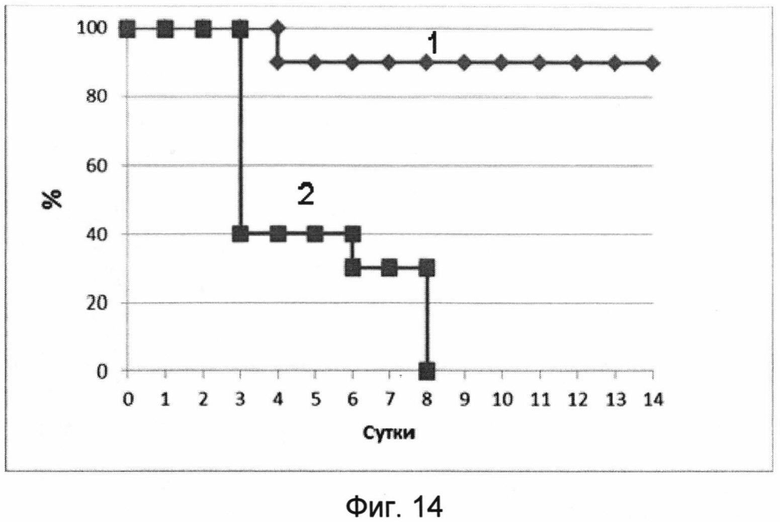
На фигуре 14 изображены кривые выживаемости мышей в течение срока наблюдения 14 дней после заражения 50 ЛД50 вируса гриппа A H1N1.
1 - кривая выживаемости мышей, получивших вакцину перед заражением.
2 - кривая выживаемости мышей, получавших буферный раствор (отрицательный контроль).
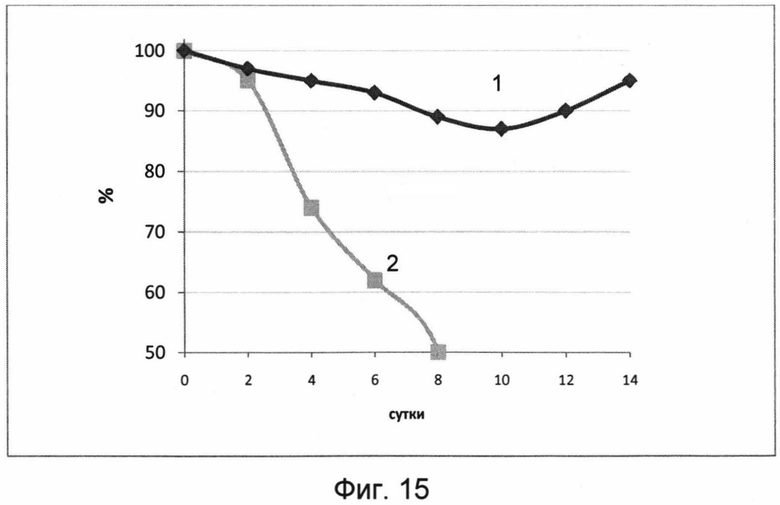
На фигуре 15 изображены кривые падения веса (суммарного в %) мышей в течение срока наблюдения 14 дней после заражения 50 ЛД50 вируса гриппа A H1N1.
1 - кривая падения веса мышей, получивших вакцину перед заражением.
2 - кривая падения веса мышей, получавших буферный раствор (отрицательный контроль).
Примеры осуществления заявленного изобретения
Пример 1. Создание нуклеотидной последовательности, кодирующей фъюжн-белок, состоящий из четырех эктодоменов белка М2 и лиганда для Толл-подобного рецептора 5 - флагеллина.
Пример 2. Конструирование рекомбинантной псевдоаденовирусной частицы, экспрессирующей ген разработанного фъюжн-белка, состоящего из четырех эктодоменов белка М2 и лиганда для Толл-подобного рецептора 5 - флагеллина.
Пример 3. Подбор эффективного молекулярного адъюванта, способного активировать врожденный иммунитет.
Пример 4. Подбор эффективного состава вакцины
Пример 5. Получение вакцины
Пример 6. Оценка Т-клеточного ответа на вакцину
Пример 7. Оценка протективности вакцины
Пример 8. Исследование широты действия противогриппозной вакцины
Пример 9. Доклинические исследования безопасности вакцины
Пример 1
В данном примере приведен способ создания нуклеотидной последовательности фъюжн-белка, состоящего из четырех эктодоменов белка М2 и лиганда для Толл-подобного рецептора 5 - флагеллина, необходимой для дальнейшего встраивания в рекомбинантные псевдоаденовирусные частицы.
Исходные нуклеотидные и аминокислотные последовательности для создания целевого гена были взяты из официального общедоступного источника NCBI, GenBank, США (www.ncbi.nlm.nih.gov). Работа с последовательностями осуществлялась с помощью компьютерной программы AliBee - Multiple Alignment.
Сначала была создана уникальная последовательность белка М2е, для этого были отобраны и выровнены относительно друг друга нуклеотидные последовательности гена М2е белка нескольких десятков штаммов вирусов гриппа А птичьего происхождения и составлен консенсусный («усредненный») вариант. Далее аминокислота цистеин в положениях 17 и 19 гена М2е была заменена на серин для того, чтобы лишить белок возможности собираться в тетрамер и иметь способность секретироваться.
Для повышения иммуногенности белка М2е вируса гриппа птиц был выбран лиганд Толл-подобного рецептора 5 флагеллин и была создана последовательность гена флагеллина на основе Salmonella enterica serovar dublin (GenBank №AAA27081), которая представлена консервативными N и С-концевыми доменами, центральный гипервариабельный регион был исключен, а в положении 176-402 аминокислот был встроен линкер, связывающей N и С-концевые домены.
Для создания конечной генетической конструкции, кодирующей слитный (фъюжн-белок), (SEQ ID NO:1) были последовательно расположены гены четырех эктодоменов белка М2 (М2е) с серинами в положениях 17 и 19, линкер, и ген флагеллина Salmonella enterica serovar dublin. Для эффективной внеклеточной экспрессии к N-концу фъюжн-белка был присоединен лидерный пептид (LP) секретируемой щелочной фосфатазы (SEAP). Нуклеотидная последовательность гена фъюжн-белка SEQ ID NO:1 кодировала аминокислотную последовательность SEQ ID NO:2.
В таблице 1 представлены нуклеотидные и аминокислотные последовательности компонентов экспрессируемой конструкции гена фъюжн-белка, содержащей лидерный пептид, четыре М2е белка и флагеллин, связанный с ними линкером.


Полученную уникальную нуклеотидную последовательность гена фъюжн-белка синтезировали общеизвестным химическим методом и лигировали в общеизвестную вспомогательную плазмиду для дальнейшего переклонирования.
Пример 2
В данном примере показано получение рекомбинантных псевдоаденовирусных частиц, экспрессирующих фъюжн-белок, состоящий из четырех эктодоменов белка М2 и лиганда для Толл-подобного рецептора 5 - флагеллина.
На данном этапе была получена рекомбинантная псевдоаденовирусная частица на основе генома аденовируса человека 5-го серотипа, содержащая вставку гена фъюжн-белка SEQ ID NO:1, созданного в примере 1.
Все нижеописанные работы по клонированию проводили с использованием общеизвестных стандартных методик, описанных в руководствах для специалистов данной области.
Получение конструкции рекомбинантной псевдоаденовирусной частицы на основе генома аденовируса человека пятого серотипа (размером 70-80 нм) проводили методом гомологичной рекомбинации между общеизвестной плазмидой pJM17 (McGrory W.J. et al., A simple technique for the rescue of early regionl mutations into infectious human adenovirus type 5, Virology, V.163, №2, 1988 - Простая техника для удаления раннего региона 1 в инфекционном аденовирусе человека 5 типа), содержащей геномную часть аденовируса человека 5 серотипа с нарушенной Е1 областью, и вспомогательной плазмидой pACCMVpLpA (Roth M.G., Methods in cell biology, №43, c. 175 - Методы в клеточной биологии.), (Go′ mez-Foix A. et al., Adenovirus-mediated transfer of the muscle glycogen phosphorylase gene into hepatocytes confers altered regulation of glycogen metabolism, J. Biol. Chem., 1992, №267 (35), 15, c. 25129-25134 - Обеспечиваемый аденовирусом перенос гена фосфорилазы мышечного гликогена в гепатоциты меняет регуляцию метаболизма гликогена). Предварительно в шаттл-плазмиду pACCMVpLpA переклонировали SEQ ID NO:1 из вспомогательной плазмиды, в результате получили pACCMVpLpA-M4FL с экспрессирующей кассетой с целевым трансгеном (геном фъюжн-белка), фланкированной участками генома аденовируса, которые участвуют в дальнейшей рекомбинации. Гомологичную рекомбинацию проводили в клетках линии НЕК293 после котрансфекции ее плазмидами pJM17 и pACCMVpLpA-M4FL. Так как плазмида pJM17 содержала бактериальный сайт инициации репликации (Ori) и ген устойчивости к ампициллину внутри области Е1 геномной части аденовируса, то эта плазмида не могла быть упакована в аденовирусные вирионы и, таким образом, предотвращалось размножение аденовирусов «дикого типа». После рекомбинации, Ori и ген устойчивости к ампициллину исчезали, замещаясь кассетой с целевым трансгеном. Рекомбинантные ДНК упаковывались в капсид вирионов, в результате чего образовались рекомбинантные псевдоаденовирусные частицы, не способные размножаться в непермиссивных культурах клеток, в том числе, в обычных клетках человека, из-за отсутствия Е1 в геноме рекомбинантных псевдоаденовирусных частиц.
Таким образом, после проведенных рекомбинаций, была получена РПАН с экспрессирующей кассетой в области делеции Е1 генома, содержащей промотор цитомегаловируса человека (CMV), ген лидерного пептида секретируемой щелочной фосфатазы SEAP (LP), гена фъюжн-белка (включающего четыре последовательных повтора уникальной последовательности М2е, линкер (LN), ген флагеллина, кодирующего консервативные N и С-концевые домены белка флагеллина Salmonella enterica serovar dublin), сигнала полиаденилирования SV40 (Simian vacuolating virus 40) - pA.
Схема экспрессирующей кассеты, включенной в геном полученного РПАН, представлена на фигуре 3.
Препарат, содержащий рекомбинантные псевдоаденовирусные частицы, получали следующим способом. Через несколько дней после трансфекции, на культуре клеток НЕК293 образовывались бляшки, их отбирали пастеровской пипеткой, полученный материал размножали на клетках НЕК293 до получения титра 108 акт.ед./мл. Далее осуществляли хроматографическую очистку, в результате получили препарат с титром 2×109 акт.ед./мл.
Активность препаратов РПАН здесь и далее оценивали стандартным методом титрования на культуре чувствительных клеток НЕК293 в реакциях бляшкообразования.
Для подтверждения создания конструкции рекомбинантной псевдоаденовирусной частицы по изобретению на основе аденовируса человека 5 серотипа (РПАН) со вставкой гена фъюжн-белка (SEQ ID NO:1) проводили ПЦР (полимеразная цепная реакция) по известной стандартной методике. Для постановки ПЦР использовали пары праймеров, комплементарных: гену гексона аденовируса человека 5 серотипа; на область Е1 (которая должна отсутствовать у созданных рекомбинантных псевдоаденовирусных частиц); на ген фъюжн-белка. По результатам ПЦР было заключено, что последовательность ДНК рекомбинантной псевдоаденовирусной частицы на основе аденовируса человека 5 серотипа - обнаружена, ген фъюжн-белка (SEQ ID NO:1) - обнаружен, Е1 область, которая должна быть делетирована у рекомбинантных псевдоаденовирусных частиц, - не обнаружена (смотри фиг. 4).
Таким образом, нами была получена рекомбинантная псевдоаденовирусная (отсутствует Е1 область аденовируса) частица на основе аденовируса человека 5 серотипа (есть гексон аденовируса человека 5 серотипа) со вставкой гена фъюжн-белка, состоящего из четырех эктодоменов белка М2 и лиганда для Толл-подобного рецептора 5 - флагеллина (присутствует ген фъюжн-белка - SEQ ID NO:1), что говорит о соответствии данной конструкции заявляемым по изобретению требованиям.
Далее была показана способность сконструированных рекомбинантных псевдоаденовирусных частиц экспрессировать ген фъюжн-белка и иммуногенность, т.е. их функциональную активность.
В исследовании были использованы мыши линии Balb/c (самки), 7-8 недель (массой 16-18 г), в каждой экспериментальной группе было 10 животных.
Эксперимент проводили на мышах после интраназального введения 50 мкл препарата РПАН, полученного ранее во 2 примере (всего 108 акт.ед.). Биологический материал (кровь) забирали у животных до введения, а также после введения кандидатной вакцины на 28 день. В сыворотке крови определяли титры специфических антител изотипа IgG к М2е белку, консенсусному для вирусов гриппа птиц, в ИФА.
Для постановки ИФА использовали искусственно синтезированный пептид М2е, консенсусный для вирусов гриппа птиц, конъюгированный с носителем KLH (гемоцианин лимфы улитки).
Иммуноферментный анализ проводили общепринятым методом. Пептид М2е иммобилизировали на 96-луночные планшеты с высокой сорбционной способностью (Greiner, Германия) в концентрации 3 мкг/мл (в карбонатном буфере, pH 9,5-9,6), выдерживали ночь при 4°C. Планшеты обрабатывали блокирующим буфером (0,01 М ФБР, pH 7,2-7,4 с 5% эмбриональной телячьей сывороткой) в течение 1 часа при комнатной температуре, отмывали 3 раза ФБР с твином.
В лунки планшет добавляли по 100 мкл 2-х кратных разведений сывороток в блокирующем буфере, инкубировали 1 час при комнатной температуре. Для определения специфических антител изотипа IgG в качестве конъюгата использовали кроличьи моноклональные антимышиные IgG (Invitrogen, USA) в разведении 1:1000, меченные пероксидазой хрена. В качестве субстрата использовали тетраметил бензидин (ТМБ) (Invitrogen, USA). Учет реакции проводили при длине волны 450 нм. За титр принимали наибольшее разведение сыворотки, которое дает оптическую плотность по крайней мере в 2 раза больше, чем сыворотка, взятая до иммунизации, в том же разведении.
На 28 день после иммунизации средние титры сывороточных IgG у мышей, иммунизированных препаратом РПАН со вставкой фъюжн-белка, были существенно выше, у контрольных групп. Средний геометрический титр составил соответственно 8444,851 (р<0.05). У животных, получавших фосфатный буфер и контрольные РПАН, титры были менее чем 100. В таблице 2 представлены результаты определения иммуногенности препарата РПАН со вставкой фъюжн-белка

Таким образом, полученные результаты говорят о способности РПАН экспрессировать фъюжн-белок и показывают его иммуногенность.
Заключено, что сконструированная рекомбинантная псевдоаденовирусная частица на основе аденовируса человека 5 серотипа, содержащая вставку гена фъюжн-белка, состоящего из четырех эктодоменов белка М2 и лиганда для Толл-подобного рецептора 5 - флагеллина, продуцирует фъюжн-белок в организме лабораторных животных. Наличие специфической активности - иммуногенности у препарата РПАН означает возможность использования в качестве действующего вещества противогриппозной вакцины.
Пример 3
Подбор эффективного молекулярного адъюванта, способного активировать врожденный иммунитет.
В данном примере рассмотрен вопрос подбора молекулярного адъюванта, способного усиливать иммуногенность РПАН со вставкой фъюжн-белка по изобретению и сохраняющего свою активность в составе препарата.
Важными компонентами, необходимыми для повышения иммуногенности вакцины, являются адъюванты. Способность адъювантов повышать иммуногенность вакцинного антигена при совместном их применении реализуется через взаимодействие с паттерн-распознающими рецепторами, активация которых приводит к дополнительной стимуляции иммунных реакций.
В наших экспериментах был обнаружен такой молекулярный адъювант - кислый пептидогликан, усиливающий иммуногенность РПАН со вставкой трансгена по изобретению.
Для подтверждения способности кислого пептидогликана усиливать иммуногенность РПАН со вставкой фъюжн-белка был проведен эксперимент с возрастающими дозировками адъюванта. Всего было сформировано 6 групп по 10 мышей. Мышей однократно интраназально иммунизировали 50 мкл препарата с содержанием:
1 группа - РПАН в дозе 108 акт.ед. без адъюванта;
2 группа - РПАН в дозе 108 акт.ед.+10 ЕД адъюванта;
3 группа - РПАН в дозе 108 акт.ед.+20 ЕД адъюванта;
4 группа - РПАН в дозе 108 акт.ед.+100 ЕД адъюванта;
5 группа - РПАН в дозе 108 акт.ед.+110 ЕД адъюванта;
6 группа - контроль, 0,9% NaCl.
Через три недели после иммунизации у животных брали кровь для определения в сыворотке крови уровня антител к вирусу гриппа птиц A/Mallard duck/PA/10218/84 (H5N2). Уровень специфических антител к вирусу гриппа А птиц H5N2 определяли с помощью ИФА, по методике, описанной во 2 примере (Фиг. 5).
Титр антител к М2е вируса гриппа птиц в сыворотках крови мышей, получавших только РПАН в дозе 108 акт.ед. без адъюванта, составил 4,2 log2; при добавлении 10 ЕД адъюванта - увеличился до 6,0 log2, при добавлении 20 ЕД адъюванта - увеличился до 7,8 log2, при добавлении 100 ЕД адъюванта - увеличился до 8,2 log2, при добавлении 110 ЕД адъюванта - остался 8,2 log2. В контроле - отрицательный результат.
Полученные результаты свидетельствовали о значительном повышении иммуногенности РПАН со вставкой фъюжн-белка при одновременном интраназальном введении с заявляемым молекулярным адъювантом, что позволяет совместно использовать их для создания высокоиммуногенных композиций.
Было проведено определение способности к взаимодействию кислого пептидогликана с Toll-подобными рецепторами, другими словами, к способности активировать врожденный иммунитет. Способность молекулярного адъюванта специфически связываться с Толл-подобными (TLR) рецепторами человека была оценена в условиях in vitro, широко известным специалистам методом, используя набор эукариотических клеточных линий на основе линии клеток эмбрионального почечного эпителия человека (НЕК293), экспрессирующих различные типы Толл-подобных рецепторов. В геном данных клеток включен ген бета-галактозидазы под NF-kB зависимым промотором. Таким образом, активация транскрипционного фактора NF-kB в результате взаимодействия лиганда со специфичным к нему Толл-подобным рецептором приводит к экспрессии клетками бета-галактозидазы. Активность бета-галактозидазы после добавления к клеткам молекулярного адъюванта определяли путем добавления субстрата для этого фермента - о-нитрофенил-β-Д-галактопиранозид. Уровень активности β-галактозидазы определяли спектрофотометрически (414 нм) по конверсии о-нитрофенил-β-Д-галактопиранозида.
Была изучена способность молекулярного адъюванта к активации следующих Толл-рецепторов: TLR2, TLR3, TLR4, TLR5, TLR7, TLR8, TLR9. Для эксперимента использовались клетки линии hTLR2/CD14, HEK293-hTLR4/CD14-MD2, HEK293-hTLR5, НЕК293-hTLR7, HEK293-hTLR8, HEK293-hTLR9, отрицательным контролем служили клетки, не несущие Толл-подобных рецепторов (НЕК293-hTLRnull).
Из всех перечисленных Толл-рецепторов молекулярный адъювант активировал Толл-подобный рецептор 4 (TLR4) (Фигура 6).
Основываясь на полученных результатах, был сделан вывод, что исследованный кислый пептидогликан из всех вышеперечисленных рецепторов, способен вызывать активацию Толл-подобного рецептора 4 (TLR4) врожденного иммунитета в условиях in vitro в концентрации от 61,7 нг/мл до 15 мкг/мл.
Таким образом, в результате проведенных исследований было установлено, что кислый пептидогликан из всех вышеперечисленных рецепторов способен вызывать активацию TLR4, что позволяет рассматривать его в качестве молекулярного адъюванта, активирующего врожденный иммунитет, и использовать его для повышения протективности вакцинных препаратов РПАН.
Для оценки стабильности молекулярного адъюванта в составе вакцины, ее хранили в течение 6 месяцев (методом ускоренного старения при 25°C) и каждые 3 месяца проверяли активность адъюванта с помощью клеток, экспрессирующих Толл-подобный рецептор 4 и несущих в своем геноме ген бета-галактозидазы под NF-kB зависимым промотором.
Состав вакцины был доведен буфером до объема дозы 0,5 мл. Буферный раствор может быть любой, поддерживающий жизнеспособность рекомбинантных псевдоаденовирусных частиц и не влияющий на стабильность молекулярного адъюванта (например, 10 mM TrisHCl, 75 mM NaCl, 1 mM MgCl2, 5% сахароза, 0,05% полисорбат 80, 0,5% Этанол, 100 мкм ЭДТА, pH 8.0). В таблице 3 представлены результаты оценки стабильности молекулярного адъюванта в вакцине.

Кроме того, была показана стабильность pH, цветности, прозрачности вакцины при хранении методом ускоренного старения.
Данным экспериментом показано отсутствие влияния РПАН и молекулярного адъюванта при совместном формулировании на активность друг друга, что говорит о стабильности разработанной вакцины.
Таким образом, было заключено, что кислый пептидогликан обнаружил свойства молекулярного адъюванта в отношении РПАН со вставкой фъюжн-белка по изобретению, способного активировать TLR4, а значит врожденный иммунитет. Адъювант стабилен в составе вакцины и не влияет на стабильность РПАН.
Таким образом, задача по разработке эффективного состава вакцины выполнена за счет добавления к рекомбинантным псевдоаденовирусным частицам молекулярного адъюванта и буферного раствора.
Пример 4
Подбор эффективного состава вакцины
Осуществлялся по результатам доклинического исследования иммуногенности различных композиций вакцины.
Проводили однократную иммунизацию вакциной в объеме 50 мкл. Опытные и контрольные группы были по 10 мышей линии Balb/c. На 42 сутки после однократной иммунизации у мышей была взята кровь, из которой была получена сыворотка. Оценка уровня антител к М2е белку вируса гриппа A H5N2 была проведена с помощью иммуно-ферментного анализа (ИФА). В качестве антигена был использован химически синтезированный пептид М2е вируса гриппа птиц. Для проведения анализа 96-луночный высокосвязывающий, плоскодонный планшет сенсибилизировали антигеном в концентрации 50 мкг/мл в 20 мМ калиево-фосфатном буфере pH 8,0. Сенсибилизацию проводили на +4°C в течение 12 часов. По окончании сенсибилизации планшет отмывали от не связавшегося антигена рабочим буфером (20 мМ калиево-фосфатный буфер pH 8,0 с 0,05% Твин 20). Далее в лунки планшета вносили двукратные разведения исследуемых сывороток крови лабораторных мышей. По окончании инкубации (1 час при+37°C на шейкере) планшет отмывали и вносили антитела к мышиному IgG, конъюгированные с пероксидазой, в разведении 1:10000 в рабочем буфере. После окончания инкубации планшет отмывали и для проявления реакции использовали ТМВ-индикаторную смесь. Далее реакцию останавливали 4М H2SO4 и оптическую плотность окрашенного продукта определяли при длине волны 450 нм.
Результаты проведенного анализа представлены в таблице 4. Данные представлены в виде среднегеометрического титра (СГТ) для каждой группы мышей.
В таблице 4 представлены результаты иммуногенности различных составов вакцины
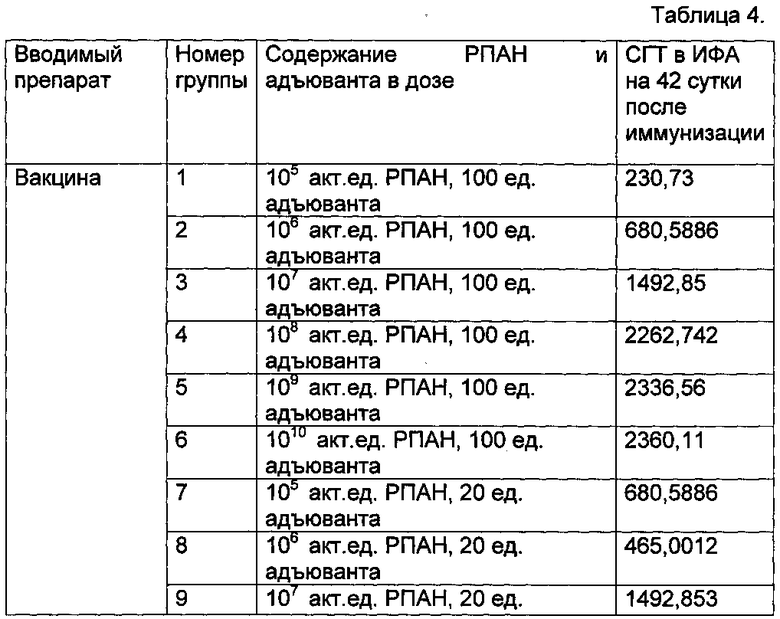

Таким образом, была показана иммуногенность полученной вакцины в эффективных дозах 106-109 акт.ед., как в сочетании с 20 ед. адъюванта, так и в сочетании со 100 ед. адъюванта, при однократной схеме иммунизации. Дозы с содержанием 105 акт.ед. РПАН не показали высокой иммуногенности, а с 1010 акт.ед. РПАН - показали иммуногенность, достоверно сравнимую с 109 акт.ед. РПАН, что делает их применение не экономичным. Наиболее оптимальным по сочетанию иммуногенность и экономическая выгода был признан состав, содержащий: 108 акт.ед. РПАН в сочетании с 20 ЕД адъюванта.
Пример 5
Получение вакцины
Разработанная технология должна обеспечивать производство вакцины следующего качества:
- доза 106-109 активных единиц (акт.ед.) РПАН, (рекомбинантных псевдоаденовирусных частиц);
- содержание адъюванта 20-100 Ед;
- объем дозы 0,5 мл (доводится фармацевтически приемлемым буферным раствором).
Получение данной вакцины проходит в несколько этапов.
Для этого полученную в примере №2 клеточную суспензию, содержащую рекомбинантные псевдоаденовирусные частицы с титром 108 ЕД на мл, использовали для дальнейшего наращивания титров рекомбинантных псевдоаденовирусных частиц и приготовления готовой вакцины с заданным содержанием рекомбинантных псевдоаденовирусных частиц.
Таким образом, для наработки необходимых титров рекомбинантных псевдоаденовирусных частиц волновой биореактор с 4500 мл суспензии пермиссивной клеточной культуры 293 засевали клеточной суспензией объемом 500 мл, содержащей рекомбинантные псевдоаденовирусные частицы с титром 108 ЕД активности на мл.
Культивировали для наращивания рекомбинантных псевдоаденовирусных частиц внутри клеток и достижения их активности 2×108 ЕД/мл, ориентировочно в течение 48 часов. По достижении необходимого содержания клеточную массу подавали на очистку, которая состояла из нескольких стадий:
1. Проводили осаждение клеточной массы центрифугированием. Поступающая на очистку суспензия имела не менее 3,3×1011 частиц на 5 л. Центрифугирование проводили при режиме 6000 g в течение 15 мин, при этом жидкий надосадок сливали, а оставшуюся твердую часть, содержащую клетки и рекомбинантных псевдоаденовирусных частиц, подавали на дальнейшие стадии очистки.
2. Извлечение рекомбинантных псевдоаденовирусных частиц из клеточной культуры проводили путем разрушения клеток четырехкратным перемораживанием-оттаиванием. Готовили буферный раствор с pH 8.0: 5 mM Трис HCl, 0.075 MNaCl, 1 mM MgCl2, 5% сахароза, 1% полисорбат 80. Полученный в предыдущую стадию осадок ресуспендировали в 70 мл буфера (коэффициент содержания ×71). Объем раствора составлял 80 мл.
Замораживание проводили в течение 2 часов в жидком азоте, размораживали на водяной бане (при +37°C), не допуская перегрева.
3. Для облегчения дальнейшего удаления геномной клеточной ДНК проводили дополнительную обработку нуклеазой. Для этого добавляли бензоназу до концентрации в растворе 150 U/мл и ставили на мягкое перемешивание с помощью магнитной мешалки на 3 часа при комнатной температуре (21-23°C).
4. Отделение рекомбинантных псевдоаденовирусных частиц от разрушенных клеток осуществляли центрифугированием при 9000 g 10 мин. Отбирали супернатант, содержащий рекомбинантные псевдоаденовирусные частицы.
5. Дальнейшую очистку проводили ультрафильтрацией. Для этого полученный супернатант разводили буфером (50 mM TrisHCl pH 7.5, 1М NaCl, 2 mM MgCl2, 5% сахароза, pH 7,5) до объема не менее 200 мл, перемешивали с помощью магнитной мешалки. В процессе фильтрации объем циркулирующего раствора (ретентата) постоянно доводили до исходного (200 мл).
6. Далее очистку производили путем анионообменной хроматографии.
Ретентат наносили на колонку (AxiChrom 70/300 объемом 400 мл), содержащую анионообменный сорбент Q Sepharose virus licenced. Рекомбинантные псевдоаденовирусные частицы сорбируются на колонке, в то время как примеси не сорбируются, а вымываются буфером А. После удаления примесей рекомбинантные псевдоаденовирусные частицы десорбировали промывкой буфером Б. Условия хроматографирования: поток 193 мл/мин, буфер А (40 mM TrisHCl, 0,27 М NaCl, 2 mM MgCl2, 5% Сахароза, 0,1% Полисорбат 80, pH 7.5), проводимость ~28-30 mS/cm; буфер Б (40 mM TrisHCl, 0.5 М NaCl, 2 mM MgCl2, 5% сахароза, 0.1% полисорбат 80, pH 7.5) проводимость ~50 mS/cm. Элюат в объеме 200 мл отправляли на следующую стадию.
7. Эксклюзионная хроматогафия
Поученный в предыдущей стадии элюат наносили на колонку (AxiChrom 100/300 объемом 800 мл), содержащую сорбент Q Sepharose 4 FastFlow. Высокомолекулярные вещества, не входящие в поры сорбента, элюировали первым пиком (к ним относятся рекомбинантные псевдоаденовирусные частицы), примеси элюировали после выхода пика не реплицирующихся наночастиц. Условия хроматографирования: поток 130 мл/мин, буфер (10 mM TrisHCl, 75 мМ NaCl, 1 mM MgCl2, 5% сахароза, 0,05% полисорбат 80, pH 8.0).
К полученному элюату (80 мл) добавляли этанол до концентрации 0,5% и этилендиаминтетрауксусную кислоту (ЭДТА) до концентрации 100 мкМ, отправляли на следующую стадию.
8. Нормальная фильтрация
Для стерилизации полученного препарата проводили фильтрование через систему фильтров с размером пор 22 мкМ.
После стерилизации полученный объем препарата разбавляют стерильным фармацевтически приемлемым буферным раствором (например, 10 mM TrisHCl, 75 mM NaCl, 1 mM MgCl2, 5% сахароза, 0,05% полисорбат 80, 0,5% Этанол, 100 мкм ЭДТА, pH 8.0) до получения активного компонента с учетом требуемой конечной активности 106-109 акт.ед. рекомбинантных псевдоаденовирусных частиц со вставкой гена фъюжн-белка на дозу, далее добавляют стерильный раствор молекулярного адъюванта с изначальной концентрацией 1000 ЕД/мл, в количестве, необходимом для приготовления требуемой концентрации 20-100 Ед на дозу. Полученный объем препарата разливают по флаконам по 0,5 мл одна доза.
Таким образом, поставленная задача по приготовлению вакцины по изобретению выполнена. Возможно получение вакцины, содержащей:
- рекомбинантных псевдоаденовирусных частиц, экспрессирующих ген фъюжн-белка, который состоит из последовательно расположенных четырех эктодоменов белка М2 (М2е) и активатора TLR 5 флагеллина Salmonella enterica serovar dublin - 106-109 акт.ед.;
- молекулярный адъювант, активатор TLR 4, кислый пептидогликан - 20-100 Ед;
- фармацевтически приемлемый буферный раствор - до 0,5 мл.
Пример 6
Оценка Т - клеточного ответа на вакцину
В данном примере показана способность вакцины стимулировать клеточный иммунитет, т.к. он участвует в регуляции специфического иммунного ответа.
Возможность стимуляции клеточного иммунитета для современной вакцины является необходимым условием. Современные научные данные говорят о необходимости активации Т-лимфоцитов для образования специфического иммунитета. В зависимости от присутствия на их поверхности тех или иных маркеров, они выполняют разные функции, главные из них - индукцию иммунных реакций осуществляют Т-хелперы - CD4.
Для оценки клеточного иммунитета, возникшего в результате интраназальной иммунизации мышей вакциной, проводили лимфопролиферативный анализ иммунокомпетентных клеток иммунизированных мышей после стимуляции антигеном. В качестве контрольных групп использовались мыши, получавшие РПАН без трансгена и фосфатный буфер. Иммунизированные мыши были усыплены эфиром, после чего у них были забраны грудинные и глубокие шейные лимфоузлы, из которых была получена культура иммунокомпетентных клеток. Полученную клеточную культуру стимулировали добавлением антигена (обработанный ультразвуком вирус гриппа птиц A H5N2) в концентрации 10 мкг на 300000 клеток. После инкубации с антигеном определяли уровень пролиферации лимфоцитов по интенсивности включения 3Н-тимидина в ДНК культивируемых клеток, которую определяли в специальном счетчике, сравнивая результаты по числу импульсов в минуту (срт). Результаты анализа показаны на фигуре 7.
В результате Н3-тимидинового анализа было продемонстрировано, что Т-лимфоциты иммунизированных разработанной вакциной мышей после добавления антигена активно включали меченый тимидин, что говорит об их пролиферации в ответ на стимуляцию антигеном. В контрольных группах такой эффект обнаружен не был.
Заключено, что оценка уровня Т-клеточного ответа у иммунизированных вакциной по изобретению животных показала наличие выраженного клеточного ответа, что соответствует современным требованиям к вакцинам.
Пример 7
Оценка протективности вакцины
В эксперименте была показана способность вакцины защищать от летальных доз вируса гриппа.
На 42 сутки после иммунизации мыши были интраназально заражены 50 ЛД50 вируса гриппа A/Mallard duck/Pennsylvania/10218/84 (H5N2), адаптированного для мышей. В опытных группах и контрольной группе было по 10 мышей. В течение 14 суток после заражения за животными вели наблюдение, фиксировали смертность и изменение веса.
Данные эксперимента представлены на фигурах 8 и 9.
В результате проведения эксперимента было показано, что вакцина обладает протективностью, т.е. защищает лабораторных животных от гриппа. Мыши, получавшие вакцину, содержащую диапазон дозировок от минимальной (106 акт.ед. РПАН, 20 ЕД адъюванта) до максимальной (109 акт.ед. РПАН, 100 ЕД адъюванта), были защищены от заражения вирусом гриппа. Мыши, получавшие 109 акт.ед. РПАН, вне зависимости от количества адъюванта, были полностью защищены. Мыши, получавшие 106 акт.ед. РПАН в сочетании как с 20 ЕД, так и со 100 ЕД адъюванта, незначительно теряли в весе, однако смертельных случаев среди этих групп зафиксировано не было. Контрольная группа, получившая буфер, полностью погибла на 10 день.
Пример 8
Исследование широты действия противогриппозной вакцины
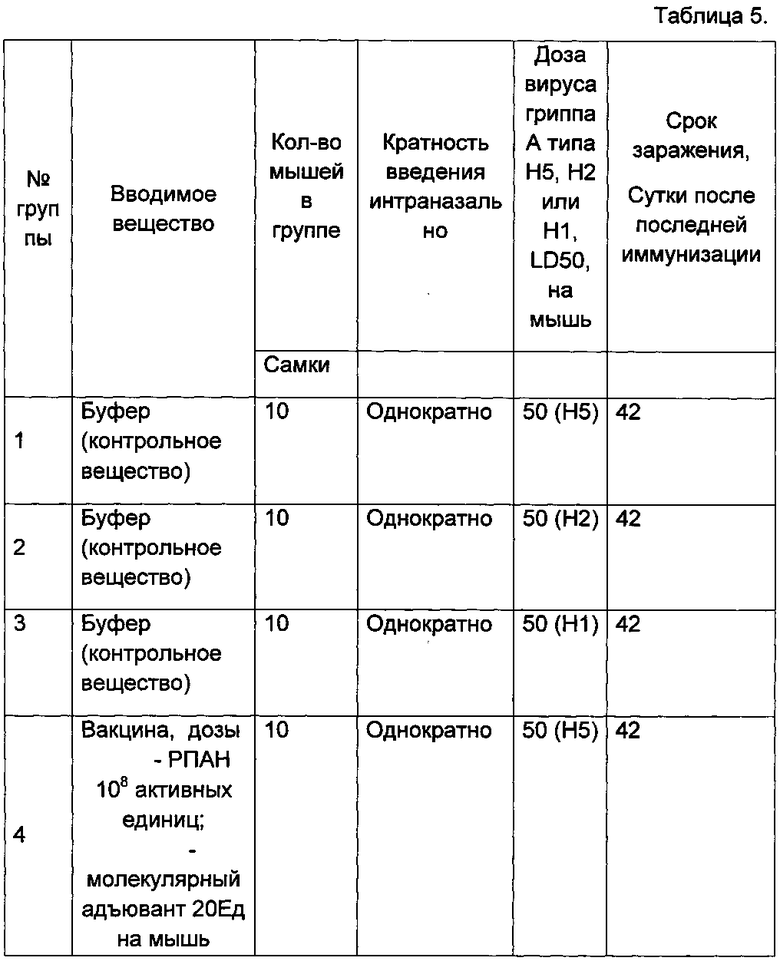
В данном примере показана широта спектра действия разработанной вакцины по протективности в отношении различных штаммов вируса гриппа А субтипов Н5, Н2 и Н1. Для заражения применяли несколько эпидемически актуальных субтипов птичьего вируса гриппа А.
На первом этапе мышам было однократно интраназально введена вакцина. Через 42 дня после иммунизации мыши были интраназально заражены 50ЛД50 вируса гриппа A/Mallard duck/Pennsylvania/10218/84 (H5N2), вируса гриппа A/BlackDuck/NewJersey/1580/78(H2N3) и вируса гриппа A/California/04/09/(H1N1), адаптированных для мышей. Вирусы гриппа A/Mallard duck/Pennsylvania/10218/84 (H5N2) и A/BlackDuck/NewJersey/1580/78(H2N3) являются птичьими, а вирус гриппа A/California/04/09/(H1N1) содержит «птичий» ген М2е. В течение 14 дней после заражения за животными вели наблюдение, фиксировали смертность и изменение веса. Вакцина содержала оптимальный состав: РПАН 108 и адъюванта 20 ЕД в 0,5 мл. Схема эксперимента представлена в таблице 5.
В результате проведения эксперимента было показано, что мыши, получавшие однократно интраназально вакцину, были на 100% защищены от заражения вирусом гриппа птиц A/Mallard duck/Pennsylvania/10218/84 (H5N2). Мыши, получавшие вакцину, незначительно теряли в весе, однако смертельных случаев среди них зафиксировано не было. Контрольная группа полностью погибла на 8 день после заражения. Результаты эксперимента представлены на фигурах 10 и 11.
В результате проведения эксперимента было показано, что мыши, получавшие однократно интраназально лекарственное средство, были на 80% защищены от заражения вирусом гриппа птиц A/BlackDuck/NewJersey/1580/78(H2N3). При этом мыши, получавшие лекарственное средство, незначительно теряли в весе. Контрольная группа полностью погибла на 10 день после заражения. Данные эксперимента представлены на фигурах 12 и 13.
В результате проведения эксперимента было показано, что мыши, получавшие однократно интраназально лекарственное средство, были защищены от заражения вирусом гриппа A/California/04/09/(H1N1). Мыши, получавшие вакцину, показали выживаемость 90%, потеряли до 13% веса. При этом контрольная группа полностью погибла на 8 день после заражения. Данные эксперимента представлены на фигурах 14 и 15.
Таким образом, было показано, что интраназальная однократная иммунизация лекарственным средством, содержащим РПАН в сочетании с молекулярным адъювантом, эффективно (эффективность 80-100%) защищает от летальных доз вирусов гриппа А птиц различных субтипов, что говорит о широком спектре действия разработанной вакцины.
Пример 9
Доклинические исследования безопасности вакцины
Согласно действующей нормативно-методической документации все вакцины, разработанные для клинического применения, должны быть безопасными.
Вакцина прошла требуемый спектр доклинических исследований безопасности, которые включали:
- исследования острой токсичности;
- исследования хронической токсичности;
- исследования репродуктивной токсичности;
- исследования иммунотоксичности и аллергезирующих свойств;
- исследования мутагенной активности.
Доклиническое исследование острой токсичности вакцины не выявило результатов, препятствующих проведению клинических испытаний для медицинского применения, доклинические исследования «хронической» токсичности вакцины не выявили результатов, препятствующих проведению клинических испытаний вакцины у взрослых при ее использовании в заявленной готовой лекарственной форме в дозе, равной 0,5 мл на человека; доклинические исследования репродуктивной токсичности не выявили негативных эффектов, препятствующих проведению клинического исследования вакцины; оценка аллергизирующего действия и иммунотоксичности не выявила отрицательного действия, был установлен стимулирующий эффект препарата на гуморальное звено иммунитета, а вакцина рекомендована для клинических испытаний; при изучении мутагенных свойств в тесте Эймса и тесте учета микроядер, нежелательная мутагенная активность отсутствовала.
Таким образом, разработанная по изобретению вакцина соответствует требованиям безопасности - не обладает токсичностью и реактогенностью, соответствует требованиям 4 класса опасности «вещества малоопасные».
Промышленная применимость
Все приведенные примеры подтверждают промышленную применимость заявленной вакцины.
Разработанная вакцина была признана кандидатным препаратом для клинического применения, как противогриппозная вакцина широкого спектра действия против птичьего гриппа А у человека.


Изобретение относится к области медицины и касается противогриппозной вакцины широкого спектра действия против птичьего гриппа А человека. Охарактеризованная вакцина содержит эктодомен белка М2, молекулярный адъювант и буферный раствор, взятые в эффективных количествах. Эктодомен белка М2 экспрессируется рекомбинантными псевдоаденовирусными частицами на основе генома аденовируса человека пятого серотипа непосредственно в организме субъекта в виде фъюжн-белка, состоящего из последовательно расположенных четырех эктодоменов белка М2 вируса гриппа А птиц и активатора TLR 5 - флагеллина Salmonella enterica serovar Dublin. В качестве молекулярного адъюванта она содержит активатор TLR-4 - кислый пептидогликан растительного происхождения с молекулярной массой от 1000 до 40000 кДа. Представленное изобретение не токсично и может быть использовано для клинического применения у человека. 2 з.п. ф-лы, 15 ил., 5 табл., 9 пр.
1. Противогриппозная вакцина широкого спектра действия против птичьего гриппа А человека на основе эктодомена белка М2, отличающаяся тем, что эктодомен белка М2 экспрессируется рекомбинантными псевдоаденовирусными частицами на основе генома аденовируса человека пятого серотипа непосредственно в организме субъекта в виде фъюжн-белка, состоящего из последовательно расположенных четырех эктодоменов белка М2 вируса гриппа А птиц и активатора TLR 5 - флагеллина Salmonella enterica serovar dublin, кроме того, вакцина дополнительно содержит молекулярный адъювант и буферный раствор, взятые в эффективных количествах, причем нуклеотидная последовательность гена фъюжн-белка, который состоит из последовательно расположенных четырех эктодоменов белка М2 и активатора TLR 5 - флагеллина Salmonella enterica serovar dublin, представлена SEQ ID NO:1, а в качестве молекулярного адъюванта она содержит активатор TLR-4 - кислый пептидогликан растительного происхождения с молекулярной массой от 1000 до 40000 кДа.
2. Противогриппозная вакцина по п. 1, отличающаяся тем, что содержит на дозу:
- рекомбинантных псевдоаденовирусных частиц, экспрессирующих ген фъюжн-белка, - 106-109 акт.ед.,
- молекулярный адъювант - 20-100 ед.,
- фармацевтически приемлемый буфер - до 0,5 мл.
3. Противогриппозная вакцина по п. 3, отличающаяся тем, что нуклеотидная последовательность гена фъюжн-белка SEQ ID NO:1 кодирует аминокислотную последовательность фъюжн-белка SEQ ID NO:2.
| УНИВЕРСАЛЬНАЯ ВАКЦИНА ПРОТИВ ВИРУСА ГРИППА ПТИЦ | 2007 |
|
RU2358981C2 |
| WO 2006069262 A2, 29.06.2006 | |||
| HULEATT JW et al, Potent immunogenicity and efficacy of a universal influenza vaccine candidate comprising a recombinant fusion protein linking influenza M2e to the TLR5 ligand flagellin, Vaccine | |||
| Станок для изготовления деревянных ниточных катушек из цилиндрических, снабженных осевым отверстием, заготовок | 1923 |
|
SU2008A1 |
| EBRAHIMI SM et al., In contrast to conventional | |||

