Область применения
Настоящее изобретение относится к иммунологии и белковой инженерии и, в частности, к созданию иммуногенных препаратов и вакцин, которые могут быть использованы для профилактики «свиного» гриппа H1N1v-2009.
Актуальность
Грипп является одним из наиболее распространенных вирусных заболеваний человека и животных. Вирусы гриппа типа A различаются по степени патогенности в отношении человека и животных. В последние годы высокопатогенные штаммы H5N1 вызывали локальные вспышки с высокой смертностью в регионах Юго-Восточной Азии. Вирус H1N1 свиного происхождения вызвал пандемию гриппа 2009-10 гг. с неожиданно высоким уровнем смертности среди групп риска и людей среднего возраста. По многим фенотипическим свойствам и филогенетическому происхождению пандемический вирус H1N1v-2009 близок к вирусу, вызвавшему «испанку» в 1918-1920 годах, что подтверждает возможность возврата в циркуляцию среди людей вирусов с высоким потенциалом патогенности. Используемые в настоящее время противогриппозные вакцины основаны на получаемом в куриных эмбрионах вирусе гриппа или его компонентах [Nicholson K., Webster R., Hay A. (1998) Textbook of Influenza // Oxford, Blackwell Science]. Высокая изменчивость поверхностных белков вируса, гемаглютинина и нейраминидазы приводит к возникновению нового эпидемического штамма каждые 1-2 года [Webster et al., (1992), Microbiol. Rev.56. P.152-179], с такой же частотой требуется изготовление «стандартных» штамм-специфических вакцин.
Одним из потенциальных источников антигенной изменчивости вируса гриппа человека является его рекомбинация с вирусами гриппа животных, которая может привести к возникновению нового высокопатогенного рекомбинантного вируса, «неизвестного» иммунной системе человека и потому способного вызвать пандемию. В то же время, создание традиционной вакцины от нового штамма требует длительного времени (от 6 до 9 месяцев), в течение которого появление нового пандемического штамма гриппа может привести к гибели миллионов людей. Таким новым высокопатогенным штаммом является вызвавший пандемию 2009 г. вирус так называемого «свиного» гриппа, относящийся, по последовательностям его гемалютинина и нейраминидазы, к типу H1N1.
Одним из перспективных путей решения проблемы получения «пандемической» вакцины является создание «универсальных» вакцин, основанных на использовании консервативных иммунодоминантных пептидов, например внеклеточного домена M2 белка вируса гриппа, последовательность которого практически инвариантна у всех человеческих штаммов. Однако у вируса «свиного» гриппа H1N1v-2009 эта последовательность заметно отличается от встречающихся в «человеческих» штаммах.
Таким образом, задача создания вакцин против новых вирусов гриппа, включая H1N1v-2009, продолжает оставаться актуальной. Такая вакцина может обеспечить защиту человека от нового пандемического вируса H1N1v-2009, а также от его производных, которые могут появиться в результате рекомбинации между этим штаммом и другими птичьими и человеческим вирусами.
Уровень техники.
1) Современные противогриппозные вакцины.
В таблице 1 представлен перечень и характеристики противогриппозных вакцин, зарегистрированных и использующихся в России.
Все вакцины могут быть разделены на следующие типы:
1. Живые
2. Инактивированные
- цельновирионные
- расщепленные
- субъединичные
Из этого перечня видно, что для таких актуальных вакцин, как противогриппозные, вообще не разрабатывались рекомбинантные эпиотопные вакцины. Однако преимущество генно-инженерных технологий в решении проблем эффективности и безопасности гриппозных вакцин очевидны и их можно перечислить в следующем порядке по степени важности:
1. Гарантированно полное отсутствие яичного белка (овальбумина) - основного аллергена в современных вакцинах.
2. Индивидуальность избранных эпиотопов - гарантии отсутствия аутоиммунных реакций и осложнений. Эта опасность особенно велика для живых и цельновирионных вакцин и не исключается для субъединичных.
3. Относительная простота производства, отсутствие зависимости от природного сырья, например куриных эмбрионов, и полное соответствие требованиям безопасности (отсутствие вирусных компонентов). Хорошие экономические показатели производства.
4. Возможность быстрого развертывания масштабного производства на случай роста потребностей в вакцине в предпандемический и пандемический периоды.
2) Биологически-активные наноструктуры на основе капсидов вирусов животных
Способность биологических макромолекул к самосборке и самоорганизации является одной из отличительных черт живых систем и предоставляет поистине неисчерпаемые возможности для использования биомолекул в качестве «строительных» блоков «молекулярного конструктора» для направленного создания новых наноархитектур с заданными пространственными и функциональными свойствами. Одним из наиболее ярких примеров таких структур, обладающих четкой симметрией, обширными возможностями направленной модификации современными методами генной инженерии, являются вирусные частицы.
Высокая стабильность рекомбинантных вирусноподобных частиц (VLP), их симметричная пространственная организация, легкость получения и очистки VLP, их безопасность, возможности направленной модификации структурных белков вирусов методами генной инженерии делают эти частицы идеальными векторами-носителями для представления чужеродных функционально-значимых пептидов и целых белков. Представление антигенных детерминант-эпитопов на поверхности VLP обеспечивает их высокую иммуногенность за счет использования вириона в качестве адъюванта в сочетании с высоким выходом, легкостью очистки и стабильностью при хранении. Эффективность такого подхода иллюстрируется многочисленными примерами создания VLP для борьбы с такими инфекционными заболеваниями, как малярия, бешенство, вирусные гепатиты B, C и E, папилломавирусная инфекция (рак шейки матки), СПИД и т.д.
Один из наиболее эффективных носителей антигенных детерминант является нуклеокапсидный белок вируса гепатита B человека, а также близкородственных вирусов животных, в дальнейшем коллективно именуемых ВГБ. Мономеры этого белка, состоящие из 183-185 аминокислот, в инфицированных клетках собираются в стабильные агрегаты (вирусоподобные частицы), называемые HBc-частицы. Образуются два типа частиц - частицы диаметром около 30 нм, состоящие из 180 мономеров, и в большем количестве частицы диаметром 34 нм, состоящие из 240 субъединиц.
Известно, что HBc-частицы могут быть использованы в качестве высокоиммуногенного носителя чужеродных пептидов, стимулирующего T-клеточный иммунный ответ. Два района HBc могут быть использованы для презентации чужеродных пептидов на поверхности HBc-частицы, - N-конец белка и так называемая иммунодоминантная петля, расположенная между 75 и 85 аминокислотными остатками белка. Было показано, что C-концевая аргинин-богатая область белка (150-183 аминокислотные остатки) может быть удалена без нарушения самосборки частиц, более того, в отличие от образованных полноразмерным белком, такие частицы при сборке не включают нуклеиновые кислоты клетки-хозяина.
Химерные HBc частицы часто имеют менее упорядоченную структуру, чем частицы, не содержащие эпитопов [Schodel et al., (1994) J. Exp. Med., 180:1037-1046]. Bo многих случаях вставка эпитопа вызывает настолько сильную дестабилизацию, что гибридные частицы либо вообще не могут быть получены, либо оказываются настолько нестабильными, что не могут быть использованы в качестве компонента вакцин [Schodel et al. (1994) Infect. Immunol., 62:1669-1676]. Поэтому показанная ниже упорядоченная структура и стабильность созданных в настоящем изобретении рекомбинантных HBc-частиц, содержащих эпитопы вирусов гриппа птиц, не являются заранее ожидаемыми.
Использование рекомбинантных HBc-частиц для получения противогриппозных вакцин описано в работе [Neirynck et al., (1999) Nature Med., 5(10):1157-1163] и последующих работах этой группы. Их авторами получена серия рекомбинантных HBc-частиц, содержащих внеклеточный домен M2 (M2E) белка вируса гриппа человека и показана их высокая иммуногенность и защита от инфекции человеческими вирусами в опытах на лабораторных животных. В этих работах была использована M2E последовательность SLLTEVETPIRNEWGCRCNDSSD, присутствующая в практически неизменном виде во всех человеческих штаммах вируса, начиная с изолированного в 1933 г. штамма A/WSN/33.
Белок M2 вирусов гриппа типа A принципиально важен для конструирования универсальных вакцин в связи с рядом недооцененных его свойств:
1. локализация и экспонирование на поверхности вириона,
2. критическая важность в процессах фьюжина и почкования (ранняя и поздняя стадии инфекции),
3. высокий консерватизм среди всех подтипов вируса вируса гриппа A.
Выбор высококонсервативного антигена предусматривал индукцию антител, способных к подавлению репродукции к большинству подтипов A вирусов гриппа. Такая вакцина получила название «универсальной» - предполагающей защиту фактически от всех вирусов гриппа типа A.
Поэтому созданная этими авторами вакцина оказалась «универсальной» в отношении человеческого гриппа. Однако у вирусов гриппа животных последовательность M2E отличается от приведенной выше. Например, M2e вируса «свиного» гриппа A/California/04/2009(H1N1) имеет последовательность SLLTEVET PTRSEWECRCSDSSD (подчеркнуты отличия от консенсусной "человеческой" последовательности (Фиг.1). Опубликованные данные об эффективности вакцин, созданных на основе человеческого М2Е, в отношении «свиного» гриппа H1N1v-2009 отсутствуют. Более того, полученные в ходе реализации настоящего изобретения результаты свидетельствуют о том, что M2E-HBc частицы, содержащие «свиной» M2E, эффективны против «свиного» гриппа A/California/04/2009(H1N1), но не против человеческого штамма A/PR/8/34. Также не обладала протективностью в отношении штамма A/PR/8/34 ранее созданная нами с использованием аналогичных подходов кандидатная вакцина против вируса гриппа птиц (Равин Н.В., Киселев О.И., Скрябин К.Г. 2009. Универсальная вакцина против вируса гриппа птиц. Патент РФ №2358981).
Таким образом, существующие в настоящее время методы создания противогриппозных вакцин, имеющие в своей основе конструирование рекомбинантных M2E-HBc частиц, содержащих «человеческий» M2E, не решают задачу создания вакцины против вируса «свиного» гриппа H1N1v-2009. В связи с этим целью изобретения являлась разработка подходов к созданию «универсальных» вакцин и, в частности, вакцины, обеспечивающей эффективную защиту против эволюционирующих вирусов «свиного» гриппа типа H1N1, представляющих особую опасность для людей в пандемический период.
Раскрытие изобретения
В настоящем изобретении ставилась задача создания рекомбинантной кандидатной вакцины против вируса «свиного» гриппа (H1N1v-2009). В основу решения поставленной задачи было положено конструирование рекомбинантных вирусоподобных частиц на основе ядерного антигена вируса гепатита B (HBc), представляющих на своей поверхности полипептиды внеклеточного домена M2 белка вируса «свиного» гриппа A/California/04/2009(H1N1).
Фактически задача была решена путем:
а) дизайна конструкции для экспрессии функционально важного и доступного эпитопа наиболее консервативного белка вирусов гриппа типа A,
б) синтеза гена, кодирующего гибридный белок, включающий М2Е и лишенный C-концевой области HBc антиген вируса гепатита В,
в) создания штамма Е. coli - продуцента гибридного белка M2Es-HBc,
г) разработки протокола выделения и очистки рекомбинантных M2Es-HBc частиц,
д) проведения испытаний полученных препаратов на лабораторных животных, результаты которых свидетельствуют о высокой иммуногенности вирусоподобных M2Es-HBc частиц и 100% защите животных от летальной гриппозной инфекции.
Первый аспект настоящего изобретения связан с получением рекомбинантной нуклеиновой кислоты (синтетического гена), кодирующей рекомбинантную белковую молекулу (рекомбинантный белок) M2Es-HBc, аминокислотная последовательность которого, начиная с N-конца, состоит из остатка метионина, последовательности SLLТEVETPTRSEWECRCSDSSD внеклеточного домена M2 белка вируса «свиного» гриппа A/California/04/2009(H1N1) от 2-ой до 24-ой аминокислоты (M2Es), изображенной на фиг.1, и последовательности ядерного антигена вируса гепатита B (HBc) от 4-й до 149-й аминокислоты, относительно первого метионина в нативном HBc, за которой, при необходимости, может следовать один цистеин.
Вторым аспектом изобретения является создание рекомбинантного вектора экспрессии и штамма Е. coli - продуцента рекомбинантного белка M2Es-HBc. С этой целью рекомбинантную нуклеиновую кислоту, кодирующую M2Es-HBc, клонировали в экспрессионном векторе pQE60 под контролем РТ5-lас промотора, репрессируемого LacI и индуцируемого IPTG, в результате чего был создан рекомбинантный вектор экспрессии.
Соответственно, третий аспект изобретения относится к оптимизации условий продукции рекомбинантного белка M2Es-HBc в клетках Е. coli и разработке методов выделения и очистки образуемых им вирусоподобных M2Es-HBc частиц.
Четвертый аспект изобретения связан с проведением испытаний полученных препаратов вирусоподобных M2Es-HBc частиц на лабораторных животных. При этом было показано, что (1) препараты M2Es-HBc частиц не токсичны для лабораторных животных (мышей), (2) иммунизация мышей приводит к образованию в их крови антител, специфически распознающих М2Е пептид в составе M2Es-HBc частиц, (3) иммунизация мышей препаратом M2Es-HBc частиц обеспечивает их 100% защиту от летальной гриппозной инфекции. Таким образом, вирусоподобные M2Es-HBc частицы могут быть использованы в качестве основы вакцины против "свиного" гриппа H1N1v-2009.
Краткое описание чертежей
Фиг.1 - Сравнение аминокислотных последовательностей внеклеточных доменов М2 белков различных штаммов вируса гриппа типа А человека и животных. Отличия от консенсусной последовательности М2е «человеческого» вируса гриппа выделены серым.
Фиг.2 - Аминокислотная последовательность рекомбинантной белковой молекулы M2Es-НВс. Последовательность аминокислот из M2 белка подчеркнута.
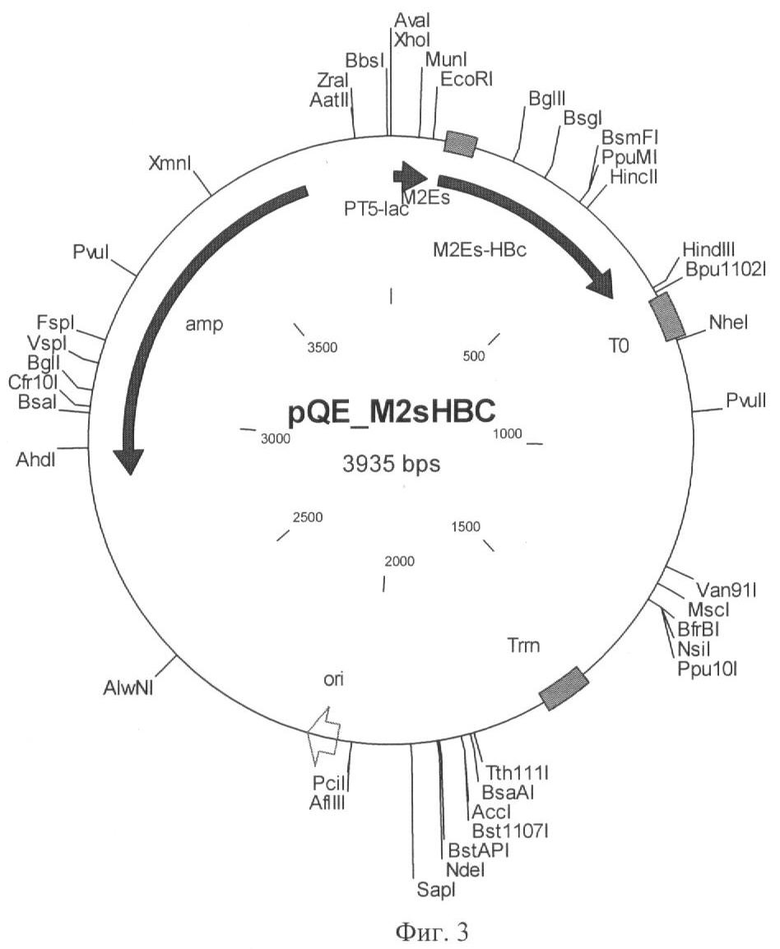
Фиг.3 - Структура рекомбинантного вектора экспрессии pQE-M2EsHBc.
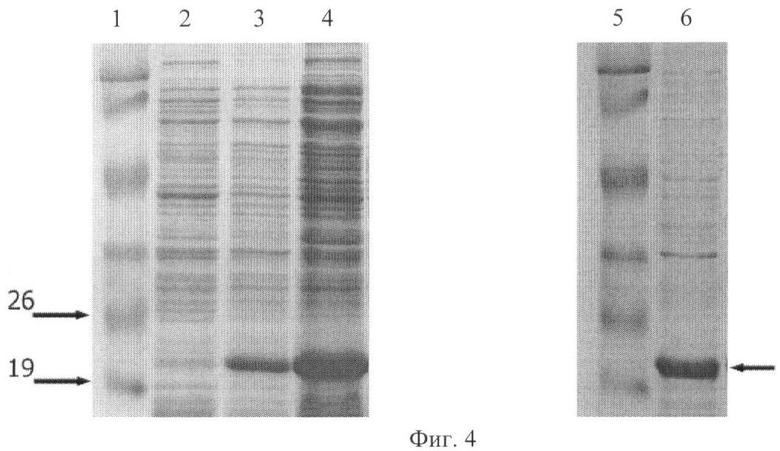
Фиг.4 - Экспрессия и очистка рекомбинантного белка M2Es-HBc.
Показаны результаты анализа белковых препаратов с помощью SDS-PAGE.
1 и 5, - маркер молекулярной массы (показаны положения маркеров 19 и 26 кДа)
2, - белковый препарат из штамма DLT1270/pQE-M2EsHBc-l до индукции синтеза продукта
3, - то же, но через 16 часов после индукции синтеза продукта
4, - фракция растворимых белков, выделенная из штамма-продуцента после индукции синтеза продукта.
5, - очищенный препарат M2EsHBc частиц
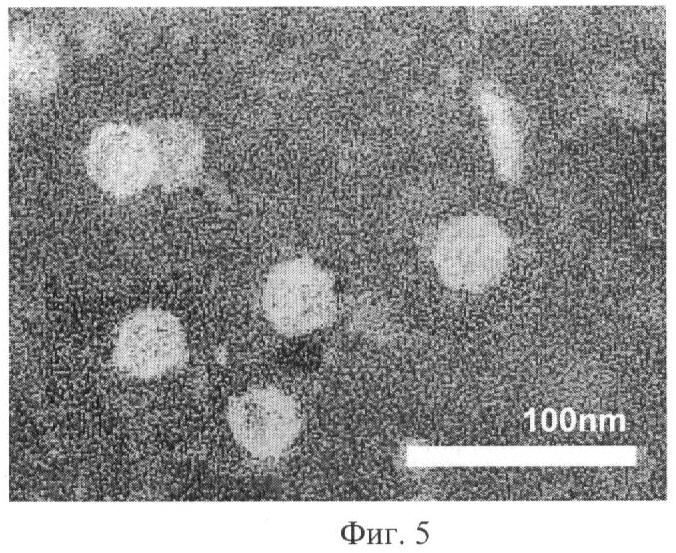
Фиг.5 - Анализ структуры вирусоподобных M2Es-HBc частиц с помощью электронной микроскопии. Шкала соответствует 100 нм.
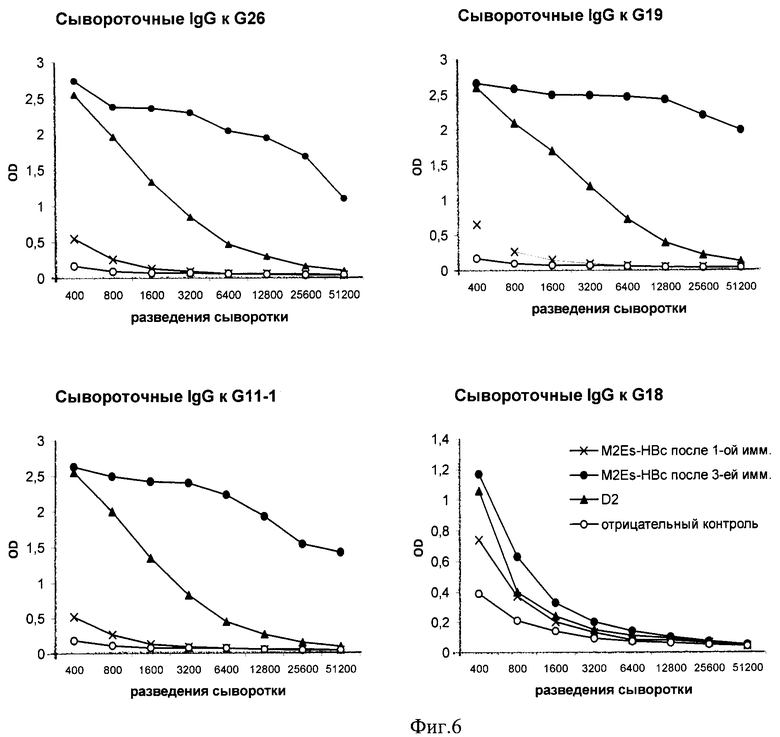
Фиг.6 - Сывороточные IgG антитела иммунизированных вирусоподобными M2Es-HBc частицами мышей, связывающиеся с синтетическими M2e пептидами. (A), - сывороточные IgG к G26; (B), - сывороточные IgG к G18; (C), - сывороточные IgG к G11-1; (D), - сывороточные IgG к G19.
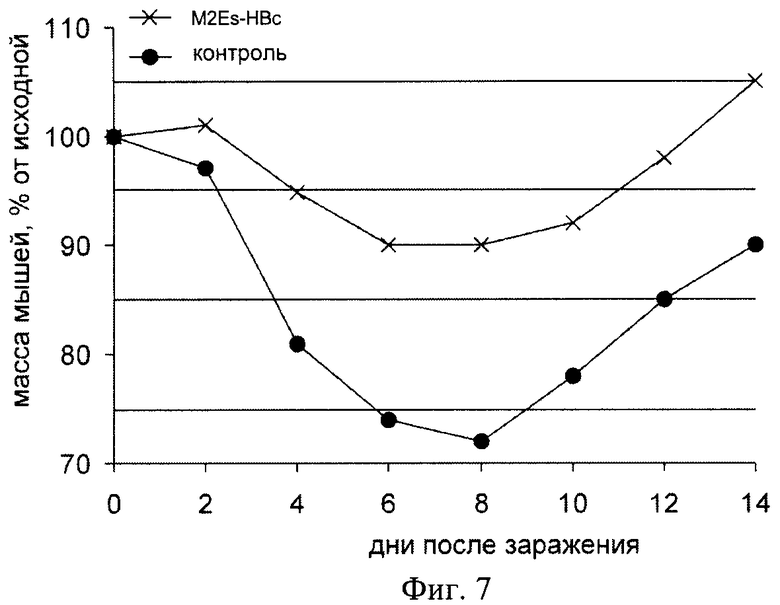
Фиг.7 - Динамика массы мышей, иммунизированных вирусоподобными M2EsHBc частицами, после заражения вирусом гриппа A/California/04/2009(H1N1).
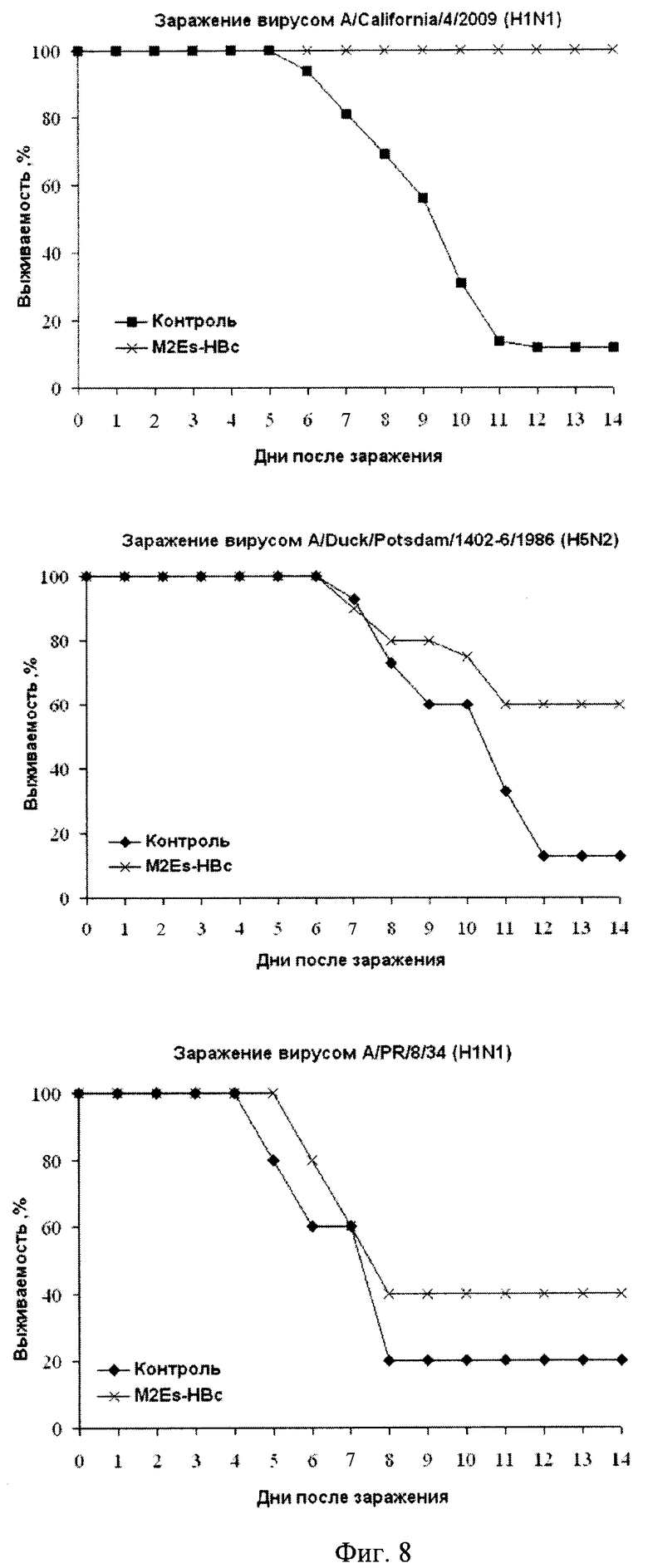
Фиг.8 - Динамика гибели мышей, иммунизированных вирусоподобными M2EsHBc частицами, после заражения различными штаммами вируса гриппа.
Осуществление изобретения.
Пример 1. Рекомбинантная белковая молекула M2Es-HBc и кодирующая ее рекомбинантная нуклеиновая кислота.
Для получения иммуногенных вирусоподобных частиц использовали рекомбинантный белок, включающий внеклеточный домен М2 белка вируса гриппа (М2Е домен) и фрагмент HBc антигена ВГБ. Рекомбинантная белковая молекула, M2Es-HBcl включала последовательно следующие элементы: N-концевой метионин, последовательность аминокислот SLLTEVETPTRSEWECRCSDSSD, соответствующую M2E домену из штамма A/California/04/2009(H1N1) вируса гриппа, аминокислотную последовательность HBc [Bichko, V., Pushko, P., Dreilina, D., Pumpen, P. and Gren, E. (1985) FEBS Lett. 185 (1), 208-212], начиная с аспарагина в положении +4 до валина в положении +149 (относительно первого метионина в нативном HBc), и цистеин на С- конце (Фиг.2). Введение C-концевого цистеина имело целью повысить стабильность рекомбинантных M2E-HBc частиц, но не обязательно для получения высокоактивных вирусоподобных частиц.
Для получения описанной выше рекомбинантной белковой молекулы была синтезирована кодирующая ее рекомбинантная нуклеиновая кислота (ген). На первом этапе часть последовательности этого гена была получена в результате проведения ПЦР с использованием праймеров M2sF1 (GAA АСС CCG ACC CGT AGC GAA TGG GAA TGC CGT TGC AGC) и HBC-R (AAG АТС ТСА GCA AAC AAC AGT AGT СТС CGG AAG) и ДНК ранее полученной нами плазмиды pQE-M2HBc-2 [Равин Н.В., Киселев О.И., Скрябин К.Г. 2009. Универсальная вакцина против вируса гриппа птиц. Патент РФ №2358981] в качестве матрицы. ПЦР проводили при следующих условиях: (1) 94°C - 30 сек, (2) 50°С - 60 сек, (3) 70°С - 90 сек, шаги 1-3 повторяли 30 раз. На следующем этапе полученный фрагмент ДНК был использован в качестве матрицы для проведения ПЦР-амплификации с праймерами M2sF2 (CTC ATG AGC CTG CTG АСС GAA GTG GAA ACC CCG ACC CGT AGC) и HBC-R. ПЦР проводили при следующих условиях: (1) 94°C - 30 сек, (2) 50°С - 60 сек, (3) 70°С - 90 сек, шаги 1-3 повторяли 30 раз.
После электрофоретического анализа выделяли ПЦР-фрагмент размером 0,52 т.п.н., который лигировали с ДНК вектора pUC19, предварительно обработанной рестриктазой SmaI. Полученную плазмиду, которая была отобрана по результатам рестрикционного анализа, обозначали pUC-M2EsHBc и использовали для получения фрагмента, содержащего рекомбинантную нуклеиновую кислоту, кодирующую белок M2Es-HBC. Для этого ее обрабатывали рестриктазами PagI и Bg1II, выделяли фрагмент размером 0,52 т.п.н., представляющий собой искомую рекомбинантную нуклеиновую кислоту.
Пример 2. Создание рекомбинантного вектора экспрессии и штамма E. coli - продуцента рекомбинантного белка M2Es-HBc.
Для создания рекомбинантного вектора экспрессии рекомбинантную нуклеиновую кислоту, представляющую собой фрагмент ДНК размером 0,52 т.п.н., вырезанный из плазмиды pUC-M2EsHBc с помощью рестриктаз PagI и Bg1II, клонировали в экспрессионном векторе pQE60 по сайтам NcoI и Bg1II. Полученный рекомбинантный вектор экспрессии был обозначен как pQE-M2sHBc (Фиг.3) и использован в дальнейшей работе. В этом векторе рекомбинантная нуклеиновая кислота, кодирующая M2Es-HBc, оперативно связана с промотором PT5-lac и терминатором транскрипции Т0 (Фиг.3), что обеспечивает ее экспрессию в клетке бактерии Escherichia coli (Фиг.4). Отсутствие обусловленных ПЦР мутаций в синтезированном гене было подтверждено с помощью ДНК-секвенирования.
Для получения штамма-продуцента рекомбинантного белка M2Es-HBc рекомбинантный вектор экспрессии pQE-M2sHBc вводили в клетки Е. coli штамма DLT1270 с помощью трансформации. Штамм DLT1270, являющийся производным штамма DH10B [Grant et al., Proc.Natl. Acad. Sci. USA. 1990. v.87(12), p.4645-4649], содержит ген репрессора лактозного оперона lacI, интегрированный в хромосому. Культуру трансформированных клеток DLT1270/ pQE-M2sHBc выращивали в стандартных условиях, подходящих для экспрессии рекомбинантного белка, например, в LB-бульоне до середины логарифмической фазы роста (OD600=0,5) при 37°C, добавляли IPTG до концентрации 1mM (достаточной для полной индукции промотора) и выращивали клетки еще в течение 16 часов при 37°C. Пробы для определения белка отбирали в начальный момент времени, а также через 16 часов после индукции. Определение уровня экспрессии рекомбинантного белка M2Es-HBc проводили путем анализа суммарных белковых препаратов, выделенных из бактериальных культур с помощью SDS-PAGE. Максимальный уровень продукции рекомбинантного белка M2Es-НВс составлял около 30-40% общего клеточного белка (Фиг.4).
Пример 3. Выделение и очистка вирусоподобных частиц, образуемых рекомбинантным белком M2Es-HBc
Клетки E.coli штамма-продуцента рекомбинантного белка M2Es-HBc после индукции (см. выше) осаждали центрифугированием 3000 об/мин 30 минут и ресуспендировали в 50 мМ Tris-HCl буфере pH 8,0, содержащем 0,5 M NaCl, 15 мМ ЭДТА и 20% сахарозы из расчета 1 мл буфера на 50 мл культуральной жидкости. Клеточную суспензию обрабатывали лизоцимом (1 мг/мл) в течение 15 минут при +4°C, затем клетки разрушали ультразвуком. К полученному лизату добавляли 1/20 объема раствора полиэтиленгликоля (50% вес/объем) и инкубировали 30 минут при +4°C. Затем проводили центрифугирование в течение 10 минут при 13000 об/мин. К супернатанту добавляли 1/5 объема насыщенного раствора сульфата аммония, перемешивали и оставляли на 30 минут при +4°C. Образовавшийся после центрифугирования осадок белков растворяли в 1 мл того же буфера и повторно осаждали сульфатом аммония в тех же условиях. Полученный осадок растворяли в 1 мл физиологически-приемлемого носителя, - 50 мМ Tris-HCl буфера с pH 8,0, содержащего 0,15 M NaCl, 1 мМ ЭДТА и 20% сахарозы. Полученный препарат рекомбинантного белка M2Es-HBc содержал по данным SDS-PAGE около 90% белка M2Es-HBc в концентрации примерно 1,5 мг/мл.
Указанные условия выделения и очистки подбирались экспериментальным путем и могут варьироваться в известных среднему специалисту в этой области значениях.
Факт образования рекомбинантным белком M2Es-HBc вирусоподобных частиц был показан с помощью электронной микроскопии выделенного препарата, в качестве контроля был использован препарат HBc-частиц, не содержащих М2Е полипептида, а в остальном эквивалентный настоящим частицам. Электронно-микроскопический анализ (Фиг.5) подтвердил факт самосборки рекомбинантных белковых молекул M2Es-HBc в вирусоподобные частицы размером 30-40 нанометров, сходные с частицами, образуемыми HBc антигеном, не содержащим M2E полипептида.
Пример 4. Иммуногенность кандидатной вакцины на основе M2Es-HBc частиц.
Препарат вирусоподобных M2Es-HBc частиц, растворенный в физиологически-приемлемом носителе, тестировали в качестве кандидатной противогриппозной вакцины. В качестве физиологически-приемлемого носителя могут быть использованы различные водные растворы, используемые для растворения медицинских препаратов, известные среднему специалисту в этой области.
Для характеристики иммуногенности и протективного действия был проведен эксперимент по иммунизации мышей очищенным препаратом вирусоподобных M2Es-HBc частиц. Опытную группу из 60 животных иммунизировали подкожно с применением для первого введения вакцины адъюванта TiterMax Gold Adjuvant (Sigma), а для двух последующих иммунизации - неполного адъюванта Фрейнда (Sigma). Схема иммунизации приведена в таблице 2.

Для определения иммуногенности кандидатной вакцины были исследованы сыворотки мышей после их иммунизации M2Es-HBc. Сыворотки собирали через 2 недели после 1-ой и после 3-ей иммунизации, титры антител определяли в пуле сывороток мышей каждой группы (3-5 мышей). Отрицательный контроль - сыворотка не иммунизированных мышей, положительный контроль - моноклинальные антитела к M2e Потсдам (D2).
ИФА для определения титра специфических антител проводили общепринятым методом. 96-луночные планшеты с высокой сорбционной способностью (Greiner, Германия) покрывали синтетическими пептидами G-11-1 (М2е вируса гриппа птиц штамма A/Chicken/Kurgan/05/2005 (H5N1)), G19 (M2e вируса гриппа птиц штамма А/Duck/ Potsdam 1402-6/1986 (H5N2)), G26 (M2e вируса «свиного» гриппа штамма A/California/7/2009 (H1N1)), G18 (M2e вируса гриппа человека штамма PR/8/34) в концентрации 5 мкг/мл и 3M2eHBc в концентрации 3 мкг/мл (в карбонатном буфере, pH 9,5-9,6), выдерживали ночь при 4°C. Планшеты обрабатывали блокирующим буфером (0,01 М ФСБ, рН=7,2-7,4 с 5% ЭТС) в течение 1 часа при комнатной температуре, отмывали 3 раза ФСБ с твином. Пул сывороток мышей каждой группы исследовали в дубликатах. В лунки планшет добавляли 100 мкл 2-кратных разведений сывороток (начиная с 1:400) в блокирующем буфере, инкубировали 1 час при комнатной температуре. В качестве конъюгата использовали кроличьи поликлональные антимышиные IgG (Abcam, UK) в разведении 1:8000 меченые пероксидазой хрена. В качестве субстрата использовали ТМБ. Учет реакции проводили при длине волны 450 нм. За титр принимали наибольшее разведение сыворотки, которое дает оптическую плотность, по крайней мере, в 2 раза больше, чем сыворотка не иммунизированных мышей в том же разведении.
Данные ИФА представлены на Фиг.6 и суммированы в Таблице 3.
Таким образом, после иммунизации мышей кандидатной вакциной на основе вирусоподобных M2Es-HBc частиц образуются в высоких титрах сывороточные антитела изотипа IgG, связывающиеся как с синтетическим пептидом G-26 «свиного» гриппа A/California/7/2009, так и с синтетическими пептидами, последовательности которых соответствуют «птичьим» (A/Chicken/Kurgan/05/2005 и А/Duck/ Potsdam 1402-6/1986) и "человеческим" (PR/8/34) вирусам гриппа. Наивысший титр антител наблюдали после 3-кратной иммунизации.
Пример 5. Протективное действие кандидатной вакцины на основе M2Es-HBc частиц на модели летальной гриппозной инфекции.
Для заражения животных, иммунизированных кандидатными вакцинами были использованы следующие вирусы гриппа, адаптированные к мышам: A/Duck/Potsdam/1402-6/1986 (H5N2), A/California/04/2009 (H1N1) и A/PR/8/34 (H1N1). Вирусы вводили интраназально в дозе 5 LD/50 по 50 мкл/мышь под легким эфирным наркозом (1 LD/50, - доза, соответствующая 50% летальности).
После заражения проводили ежедневное наблюдение за животными. Протективное действие кандидатной вакцины оценивали с помощью двух параметров:
1. Определения динамики падения массы тела мышей после заражения;
2. Выживаемости мышей после заражения.
На Фиг.7 показана динамика падения массы тела животных после заражения, что может являться показателем тяжести протекания заболевания. После инфекции вес иммунизированных животных снижался (до 90% от начального), но в значительно меньшей степени, чем у мышей контрольной группы (до 70% от начального веса). Эти результаты показывают, что иммунизация кандидатной вакциной не предотвращает гриппозную инфекцию, но облегчает ее протекание.
На Фиг.8 показаны результаты основного показателя, по которому оценивается эффективность протективного действия M2Es-HBc частиц - динамика гибели мышей после заражения 5 LD/50 вируса «свиного» гриппа A/California/4/2009 (H1N1). Полученные данные однозначно свидетельствуют о 100% протективном действии кандидатной вакцины на основе вирусоподобных частиц M2Es-HBc после трехкратной иммунизации. За весь период наблюдения все животные в этой группе остались живыми. В контрольной группе мышей при этих условиях заражения осталось в живых лишь 12% животных. Частичная защита от инфекции наблюдалась и в отношении штаммов гриппа, у которых аминокислотная последовательность M2e пептида отличается от таковой в использованном для иммунизации препарате. Так, при инфицировании мышей вирусом «птичьего» штамма A/Duck/Potsdam/1 402-6/1986 (H5N2) наблюдали 60% выживаемость иммунизированных животных против 13% в контрольной группе (статистическая значимость P<0,006, тест Фишера). При инфицировании «человеческим» гриппом A/PR/8/34 выживаемость животных составила 40% в опытной группе и 20% - в контрольной. Эти результаты показывают функциональную значимость различий последовательностей М2е пептидов вирусов гриппа человека и животных, что необходимо учитывать при разработке «универсальных» противогриппозных вакцин на основе M2e.
Полученные данные об эффективности кандидатной вакцины против «свиного» гриппа на мышах позволяют экстраполировать достигнутый эффект и на человека, поскольку механизмы функционирования иммунной системы у всех млекопитающих в основном сходны, а лабораторные животные являются стандартной моделью для создания вакцин. Следовательно, полученный препарат может рассматриваться в качестве кандидата на вакцину против «свиного» гриппа H1N1v-2009, предназначенную для человека.
| название | год | авторы | номер документа |
|---|---|---|---|
| КРОСС-РЕАКТИВНАЯ РЕКОМБИНАНТНАЯ ВАКЦИНА ПРОТИВ ВИРУСА ГРИППА А ЧЕЛОВЕКА | 2020 |
|
RU2742336C1 |
| РЕКОМБИНАНТНАЯ ПРОТИВОГРИППОЗНАЯ ВАКЦИНА С ШИРОКИМ СПЕКТРОМ ЗАЩИТЫ И СПОСОБ ЕЕ ПОЛУЧЕНИЯ | 2017 |
|
RU2757013C2 |
| УНИВЕРСАЛЬНАЯ ВАКЦИНА ПРОТИВ ВИРУСА ГРИППА ПТИЦ | 2007 |
|
RU2358981C2 |
| Универсальная противогриппозная вакцина | 2015 |
|
RU2618918C2 |
| РЕКОМБИНАНТНЫЙ ВИРУСНЫЙ ВЕКТОР И СИСТЕМА ЭКСПРЕССИИ В КЛЕТКАХ РАСТЕНИЯ ГИБРИДНОГО БЕЛКА, ВКЛЮЧАЮЩЕГО ВНЕКЛЕТОЧНЫЙ ДОМЕН БЕЛКА М2 ВИРУСА ГРИППА, ПРИСОЕДИНЕННЫЙ К ЯДЕРНОМУ АНТИГЕНУ ВИРУСА ГЕПАТИТА В | 2010 |
|
RU2460796C2 |
| ПРОТИВОГРИППОЗНАЯ ВАКЦИНА ШИРОКОГО СПЕКТРА ДЕЙСТВИЯ ПРОТИВ ПТИЧЬЕГО ГРИППА А НА ОСНОВЕ ЭКТОДОМЕНА БЕЛКА М2 | 2014 |
|
RU2571944C1 |
| Антиген для универсальной рекомбинантной вакцины против вируса гриппа птиц и человека | 2021 |
|
RU2839428C2 |
| Кассета, предназначенная для получения плазмидных векторов, используемых для создания клеток-продуцентов вирусоподобных частиц (ВПЧ) вируса гриппа | 2018 |
|
RU2680703C1 |
| MDCK клетка-продуцент белков вируса гриппа (варианты) | 2018 |
|
RU2681482C1 |
| АТТЕНУИРОВАННЫЙ ГРИППОЗНЫЙ ВЕКТОР И МУКОЗАЛЬНАЯ УНИВЕРСАЛЬНАЯ ГРИППОЗНАЯ ВАКЦИНА НА ЕГО ОСНОВЕ | 2016 |
|
RU2660562C2 |


Изобретение относится к области биотехнологии, генной инженерии и вирусологии. Сконструированная рекомбинантная белковая молекула для получения рекомбинантной вакцины против инфекции, вызванной вирусом «свиного» гриппа (H1N1v-2009). Молекула состоит из остатка метионина, последовательности внеклеточного домена М2 белка от 2 до 24 аминокислоты и последовательности ядерного антигена вируса гепатита В от 4 до 149 аминокислоты. Молекула способна образовывать вирусоподобные частицы. Также раскрыта рекомбинантная нуклеиновая кислота, кодирующая такую молекулу, вектор для ее экспрессии, вирусоподобные частицы, образованные такими молекулами, и вакцина, основанная на полученных вирусоподобных частицах. Полученная вакцина может рассматриваться в качестве кандидата на рекомбинантную вакцину против вируса «свиного» гриппа. 6 н.п. ф-лы, 8 ил., 3 табл., 5 пр.
1. Рекомбинантная белковая молекула для получения рекомбинантной вакцины против инфекции, вызванной вирусом «свиного» гриппа (H1N1v-2009), аминокислотная последовательность которой, начиная с N-конца, состоит из остатка метионина, последовательности внеклеточного домена М2 белка (М2Е) вируса «свиного» гриппа от 2-й до 24-й аминокислоты, представленной на фиг.1, и последовательности ядерного антигена вируса гепатита В (НВс) от 4-й до 149-й аминокислоты, относительно первого метионина в нативном НВс, которая способна образовывать вирусоподобные частицы размером около 30-40 нм.
2. Рекомбинантная нуклеиновая кислота для получения рекомбинантной вакцины против инфекции, вызванной вирусом «свиного» гриппа (H1N1v-2009), кодирующая рекомбинантную белковую молекулу по п.1 и состоящая из последовательно расположенных частей: триплета нуклеотидов, кодирующего метионин, нуклеотидной последовательности, соответствующей аминокислотной последовательности фрагмента М2Е, и нуклеотидной последовательности, соответствующей аминокислотной последовательности фрагмента НВС.
3. Рекомбинантный вектор экспрессии, содержащий нуклеиновую кислоту по п.2, оперативно связанную с регуляторными элементами, обеспечивающими ее экспрессию в клетке бактерии Escherichia coli.
4. Способ получения рекомбинантной вакцины против инфекции, вызываемой вирусом «свиного» гриппа (H1N1v-2009), включающий введение нуклеиновой кислоты по п.2 в клетку бактерии Escherichia coli, продукцию рекомбинантной белковой молекулы в указанной клетке с образованием вирусоподобных частиц, их выделение и смешивание с физиологически приемлемым носителем.
5. Вирусоподобная частица, образованная рекомбинантными белковыми молекулами по п.1, предназначенная для использования в качестве компонента вакцины против инфекции, вызываемой вирусом «свиного» гриппа (H1N1v-2009).
6. Вакцина против инфекции, вызываемой вирусом «свиного» гриппа (H1N1v-2009), включающая вирусоподобные частицы по п.5.
| ZHANG G.G | |||
| et al., "Enhancement of mucosal immune response against the M2eHBc+antigen in mice with the fusion expression products of LTB and M2eHBc+through mucosal immunization route", Vet Res Commun | |||
| Колосоуборка | 1923 |
|
SU2009A1 |
| Колосоуборка | 1923 |
|
SU2009A1 |
| NEIRYNCK S | |||
| et al., "A universal influenza A vaccine based on the extracellular domain of the M2 protein", | |||

