Область техники
Настоящее изобретение относится к области биотехнологии, вирусологии, медицины. Оно касается рекомбинантных векторов, которые могут быть использованы в фармацевтической промышленности для производства противогриппозных вакцин.
Предшествующий уровень развития
Грипп - высококонтагиозное инфекционное заболевание, широко распространенное по всему миру. Согласно ВОЗ, основным, традиционным и самым эффективным способом профилактики гриппа или его тяжелых последствий является вакцинация. В настоящее время применяют в практике живые аттенуированные, инактивированные (цельновирионные и сплит-вакцины) и субъединичные вакцины (Есмагамбетов И.Б., Алексеева С.В., Саядян Х.С., Шмаров М.М. Современные подходы к созданию универсальной вакцины против вируса гриппа («Инфекция и иммунитет», 2016, Т. 6, №2, С. 117-132). Большая доля вакцин предназначены для профилактики сезонного гриппа и содержат в качестве антигена три или четыре типа гемагглютинина, циркулирующих на данный период вирусов гриппа.
Сложность заключается в том, что вакцинация против гриппа наиболее эффективна в случаях, когда циркулирующие вирусы в значительной мере соответствуют вирусам, содержащимся в вакцине. Однако вирус гриппа постоянно изменяется и при совместной циркуляции может происходить обмен генетическим материалом, в результате чего возможно появление новых вирусов, отличающихся от всех циркулирующих вариантов и обладающих пандемическим потенциалом, против которых сезонные вакцины могут быть неэффективны.
Для таких измененных вариантов вируса гриппа разрабатывают пандемические вакцины, которые на сегодняшний день являются моновалентными, а процесс производства первых серий может занимать примерно 5-6 месяцев после выявления и изоляции нового штамма [Киселев О.И., Цыбалова Л.М., Покровский В.И. Грипп. Эпидемиология, диагностика, лечение, профилактика. Москва: МИА, 2012, С. 443-466). Индуцированный ими иммунитет, является штаммоспецифическим, т.е. протективным по отношению к идентичным или близкородственным вирусам гриппа.
В связи с этим актуальным является получение вакцинных штаммов, способных индуцировать иммунный ответ широкого спектра против различных вирусов гриппа человека и вирусов гриппа птиц, также способных вызвать заболевания у людей.
В настоящее время наиболее перспективным является направление по разработке новых вакцин на основе консервативных антигенов вируса, которые обладали бы максимальной иммуногенностью (Букринская, А.Г. Вирусология / А.Г. Букринская. - М.: Медицина, 1986. - 336 с. Коротяев, А. И. Медицинская микробиология, иммунология и вирусология / А.И. Коротяев, С.А. Бабичев. - СПб.: СпецЛит, 2002. - 591 с. Одним из подходов для решения такой задачи является поиск и характеристика консервативных антигенных детерминант в молекуле гемагглютинина (НА) вируса гриппа (Кареткина Г.Н. Грипп: новое в лечении и профилактике, 2009, №1, http://www.Ivrach.ru/2009/01/5898073/). При этом, протективный эффект таких универсальных вакцин должен обеспечиваться индукцией комплексного иммунного ответа, базирующегося на выработке кросс-реактивных антител и Т-клеток. (Есмагамбетов И.Б., Алексеева С.В., Саядян Х.С, Шмаров М.М. Современные подходы к созданию универсальной вакцины против вируса гриппа, Инфекция и иммунитет, 2016, Т. 6, №2, С. 117-132).
В качестве примера вакцинного антигена с широким спектром противогриппозного действия можно рассмотреть следующий патент. Известны «Оптимизированные с помощью вычислительных средств антигены с широким спектром реактивности для вирусов гриппа H5N1 и H1N1» (патент РФ 2639551). Предложен способ их получения. Указанные в патенте пептиды способны вызывать иммунный ответ с широким спектром реактивности против вируса гриппа H5N1 и H1N1. Оптимизированные полипептиды НА были разработаны путем проведения серии выравниваний последовательностей белков НА и последующего получения консенсусных последовательностей на основании строения определенных изолятов вирусов гриппа H5N1 и H1N1. Оптимизированные последовательности нуклеиновых кислот могут быть вставлены в соответствующий вектор экспрессии, такой как вектор экспрессии pTR600. Данный патент можно рассматривать в качестве прототипа.
Недостатком указанного выше прототипа является то, что указанные полипептиды не достаточно широко специфичны, т.е. защищают только от вирусов гриппа субтипов Н5 и Н1. Таким образом, их применение в качестве вакцинного антигена может защитить от небольшого и строго определенного количества вирусов гриппа, а, следовательно, в случае появления пандемического варианта других субтипов люди могут заболеть гриппом.
Раскрытие изобретения
Техническая задача заявляемого изобретения направлена на создание штамма рекомбинантной псевдоаденовирусной частицы для создания противогриппозных иммуногенных препаратов, способных вызвать протективный иммунитет против всех существующих и вновь появляющихся вирусов гриппа А субтипов Н1, Н2 и Н5.
Поставленная задача решена за счет того, что получен штамм рекомбинантной псевдоаденовирусной частицы на основе генома аденовируса человека 5 серотипа Ad5-tetOFF-E3-HA125, несущей ген консенсусной последовательности гемагглютинина вируса гриппа А субтипов Н1, Н2, Н5, обогащенный В и Т-клеточными эпитопами вируса гриппа с нуклеотидной последовательностью SEQ ID NO: 2, при этом ген консенсусной последовательности гемагглютинина вируса гриппа А кодирует аминокислотную последовательность SEQ ID NO: 1.
Способ получения гена консенсусной последовательности гемагглютинина, осуществляемый за счет слияния консервативных участков гемагглютининов вируса гриппа А субтипов Н1, Н2 и Н5, при этом общая длина рекомбинантного белка равна нативному гемагглютинину вируса гриппа А - 1701 п.о., содержит Т-клеточные эпитопы и конформационный В-клеточный эпитоп в стебельной части, заключается в следующих последовательных действиях:
- из общедоступной базы генетических данных отбирают аминокислотные последовательности гемагглютининов вирусов гриппа А трех субтипов Н1, Н2 и Н5, выделенные от человека, выравнивают друг относительно друга с помощью общедоступной компьютерной программы, при этом сохраняя конформационный В-клеточный эпитоп в стебельной части, и, таким образом, получают «консенсусные» (усредненные) последовательности для каждого субтипа отдельно;
- с помощью общедоступной базы данных по распространенности аллельных частот выявляют наиболее часто встречающиеся аллели молекул HLA I-класса и HLA II-класса у жителей РФ и оставляют эпитопы с высокой афинностью, сокращая одинаковые последовательности эпитопов, которые распознаются разными аллелями;
- с помощью доступных онлайн сервисов определяют наличие эпитопов в консенсусных последовательностях Н1, Н2 и Н5 для отобранных ранее аллелей HLA I-класса и HLA II-класса.
- накладывают последовательности выявленных эпитопов аллелей HLA I-класса и HLA II-класса на каждую из трех консенсусных последовательностей вируса гриппа субтипов Н1, Н2 и Н5 и определяют наличие соответствующих эпитопов и отмечают их местоположение;
- с помощью общедоступной базы данных иммунологических эпитопов из отобранных ранее Т-клеточных эпитопов HLA отбираются все известные эпитопы с экспериментально подтвержденной активностью и также на каждую из трех консенсусных последовательностей вируса гриппа субтипов Н1, Н2 и Н5 и определяют наличие соответствующих эпитопов и отмечают их местоположение.
- три консенсусные последовательности гемагглютининов вирусов гриппа А трех субтипов Н1, Н2 и Н5 выравнивают относительно друг друга;
- собирают консенсусную последовательность гемагглютинина вируса гриппа А путем слияния того участка гена, на котором отмечено наибольшее количество Т-клеточных эпитопов, выбранного каждый раз из одной из трех консенсусных последовательностей гемагглютининов вирусов гриппа А субтипов Н1, Н2 и Н5, и последовательно включаемого в собираемую консервативную последовательность;
- лученную консервативную аминокислотную последовательность гемагглютинина вируса гриппа А, содержащую участки гемагглютининов вирусов гриппа субтипов Н1, Н2 и Н5 и максимально насыщенную консервативными В- и Т-клеточными эпитопами, переводят в соответствующую ей нуклеотидную последовательность, в которой оптимизируют кодоны общеизвестным способом для более эффективной экспрессии в клетках человека.
Иммуногенный препарат, содержащий штамм рекомбинантной псевдоаденовирусной частицы на основе генома аденовируса человека 5 серотипа по п. 1. для профилактики гриппа А, вызванного субтипами Н1, Н2 и Н5 содержи на дозу:
- рекомбинантная псевдоаденовирусная частица Ad5-tetOFF-Е3-НА125 - 107-109 акт.ед.;
- фармацевтически приемлемый буферный раствор-до 0,5 мл.
Описание фигур
На фигуре 1
представлено схематическое изображение фрагмента гена консенсусной последовательности гемагглютинина вируса гриппа А субтипов Н1, Н2, Н5, обогащенного В и Т-клеточными эпитопами вируса гриппа.
Использованы следующие обозначения:
 - выбранный фрагмент гемагглютинина вируса гриппа субтипа Н1;
- выбранный фрагмент гемагглютинина вируса гриппа субтипа Н1;
 - выбранный фрагмент гемагглютинина вируса гриппа субтипа Н2;
- выбранный фрагмент гемагглютинина вируса гриппа субтипа Н2;
 - выбранный фрагмент гемагглютинина вируса гриппа субтипа Н5.
- выбранный фрагмент гемагглютинина вируса гриппа субтипа Н5.
На фигуре 2
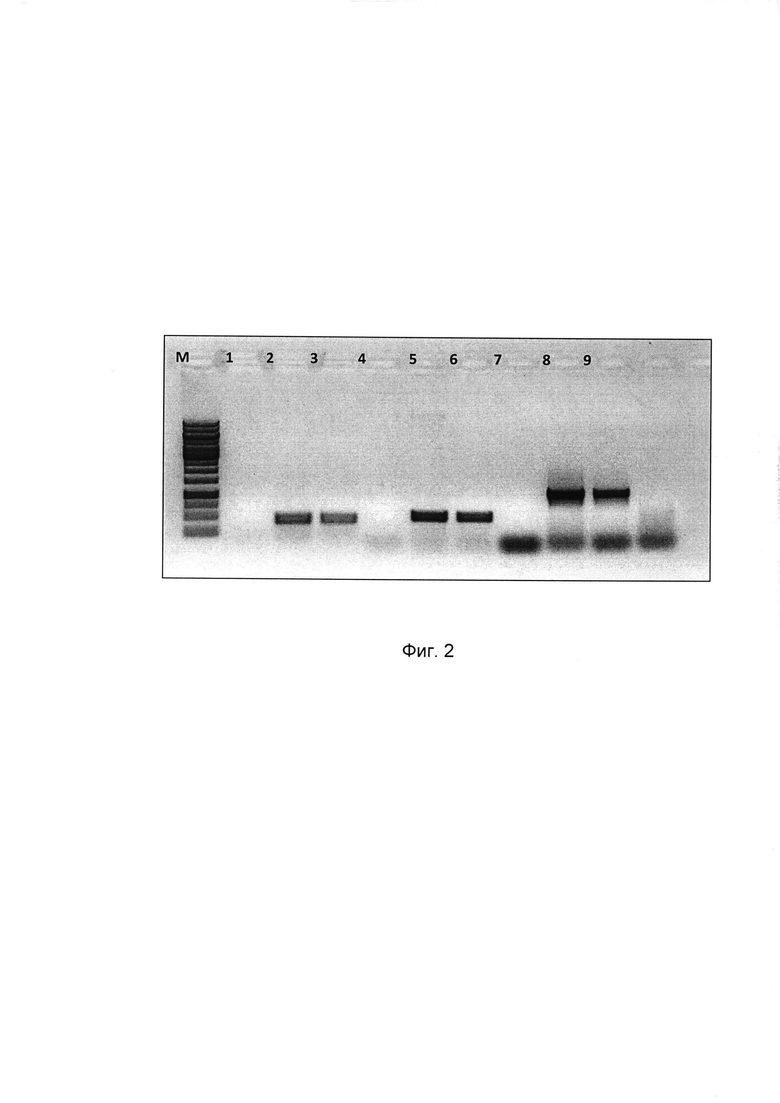
представлена электрофореграмма ПЦР-анализа.
Результаты ПЦР подтверждают подлинность созданной по изобретению рекомбинантной псевдоаденовирусной частицы, так как показывает наличие аденовирусного вектора, отсутствие реплицирующихся частиц аденовируса (по отсутствию Е1 области генома), наличие целевого трансгена.
Обозначения:
М - маркер молекулярного веса Thermo Scientific Gene Ruler 1 kb DNA Ladder;
Треки 1, 2, 3 - на трансген (НА 125), где:
1 - Контроль отрицательный;
2 - Контроль положительный (плазмида);
3 - гексон (440 п.о.)
Треки 4, 5, 6 - на аденовирусный геном (наличие гексона),
где:
4 - Контроль отрицательный;
5 - Контроль положительный (плазмида);
6 - НА 125 (504 п.о.).
Треки 7, 8, 9 - реплицирующиеся аденовирусные частицы (по наличию области Е1 генома аденовируса должны отсутствовать), где:
7 - Контроль отрицательный;
8 - Контроль положительный (Ad5-wt);
9 - Е1 (размер фрагмента 1000 п.о.).
Таким образом, результаты проведенного анализа подтверждают создание рекомбинантных псевдоаденовирусных частиц на основе генома аденовируса человека 5 серотипа, несущих ген консенсусной последовательности гемагглютинина вируса гриппа А субтипов Н1, Н2, Н5, обогащенной В- и Т-клеточными эпитопами вируса гриппа, так как на представленном форезе присутствует целевая полоса на гексон аденовируса в опыте и положительном контроле, равная 440 п.о., полоса на НА125 в опыте и положительном контроле, равная 504 п.о., а также отсутствует область Е1 генома аденовируса, что говорит о создании рекомбинантной псевдоаденовирусной частицы по изобретению.
На фигуре 3.
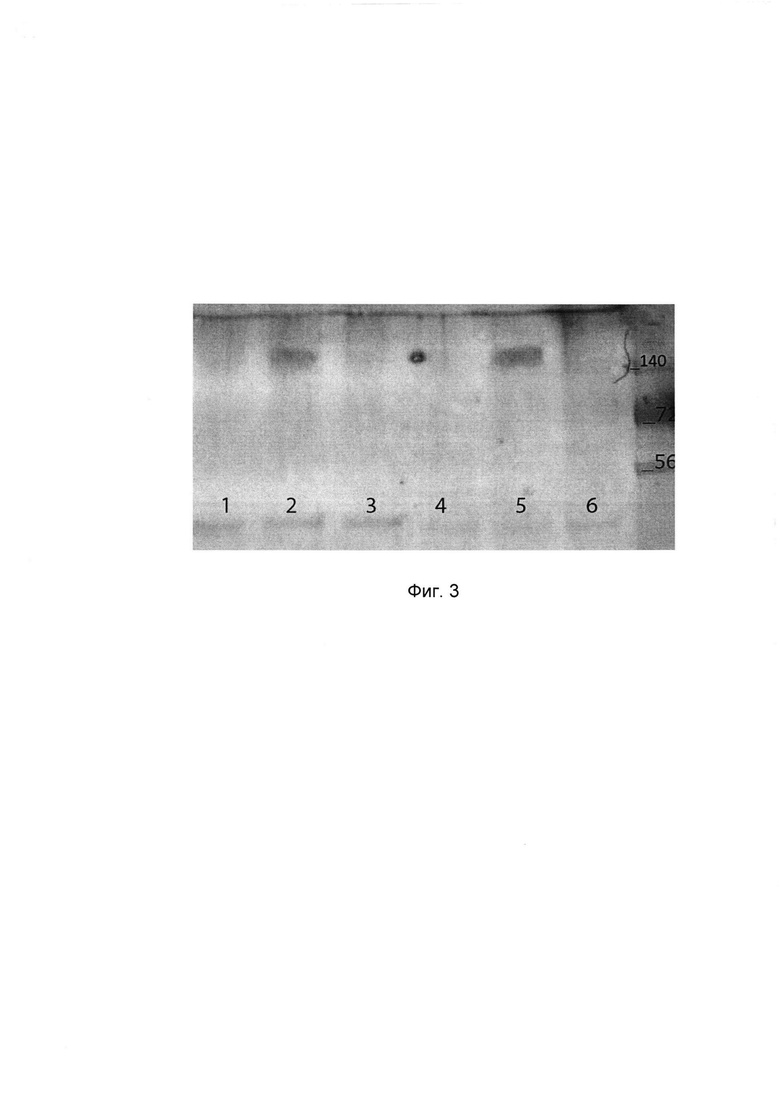
представлены результаты иммуноблоттинга при определении экспрессии целевых белков рекомбинантных псевдоаденовирусных частиц в дыхательных путях мыши через 72 часа после однократного интраназального введения в дозе 4,0×107 акт. ед.
Обозначения:
1 - экстракт лизированной ткани носовой перегородки интактной мыши;
2 - экстракт лизированной ткани носовой перегородки мыши, получавшей препарат;
3 - экстракт лизированной ткани носовой перегородки мыши, получавшей Ad5-null;
4 - экстракт лизированной ткани легких интактной мыши;
5 - экстракт лизированной ткани легких мыши, получавшей препарат;
6 - экстракт лизированной ткани легких мыши, получавшей Ad5-null;
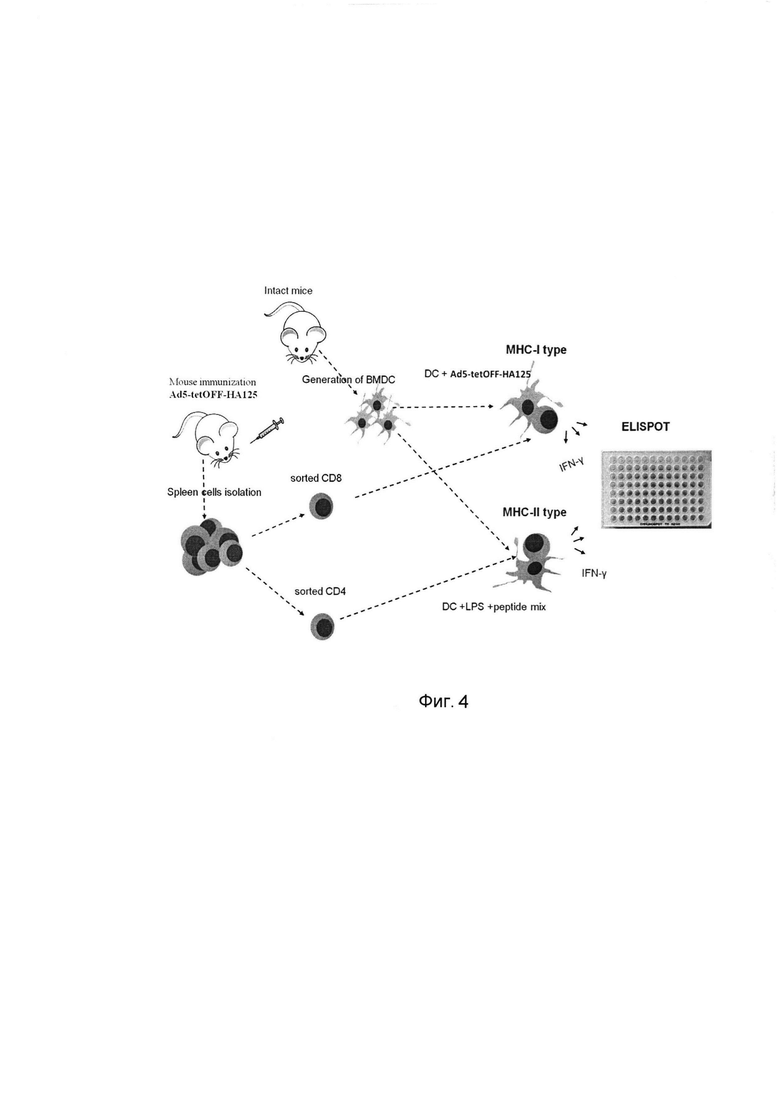
7 - маркер молекулярного веса, кДА. На фигуре 4
представлена схема изучения специфического Т-клеточного ответа у мышей, иммунизированных Ad5-tetOFF-HA125, к целевому антигену, представленному в комплексе с молекулами МНС I и МНС II.
Из костного мозга аллогенных интактных мышей, с помощью культивирования in vitro в присутствии GM-CSF, получают дендритные клетки. Дендритные клетки, трансдуцированные Ad5-tetOFF-HA125, превращаются в антиген-презентирующие клетки, экспрессирующие эпитопы целевого антигена в комплексе с молекулами МНС I. Дендритные клетки, активированные LPS и нагруженные смесью пептидов целевого антигена, превращаются в антиген-презентирующие клетки, экспрессирующие эпитопы целевого антигена в комплексе с молекулами МНС II.
Из селезенки иммунизированных и контрольных мышей методом клеточной сортировки на проточном цитофлуориметре-сортировщике получают высокоочищенные (чистота 95-97%) популяции CD4 и CD8 Т-клеток. При сокультивировании CD8 Т-клеток с дендритными клетками, экспрессирующими эпитопы целевого антигена в комплексе с молекулами МНС I, и CD4 Т-клеток с дендритными клетками, экспрессирующими эпитопы целевого антигена в комплексе с молекулами МНС II, Т-клетки начинают секретировать IFN-γ. Детекция секреции IFN-γ от единичных клеток производится с помощью набора ELISPOT.
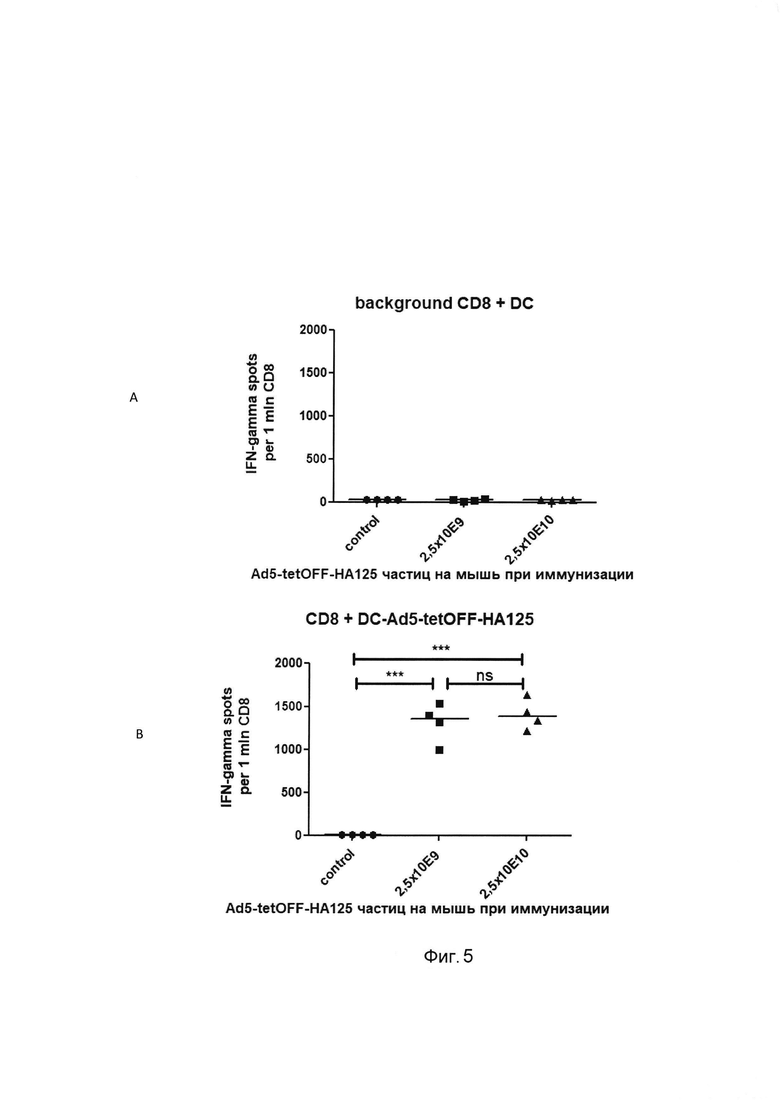
На фигуре 5
показаны результаты определения методом ELISPOT количества CD8+IFN-γ секретирующих клеток, специфичных к эпитопам целевого антигена в комплексе с молекулами МНС I, в селезенке мышей, иммунизированных Ad5-tetOFF-HA125.
Ось абсцисс - экспериментальные группы, ось ординат - число IFN-γ секретирующих клеток в пересчете на 1 млн CD8+ Т-клеток.
А - фоновый уровень секреции IFN-γ при сокультивировании сортированных CD8+ Т-клеток и аллогенных дендритных клеток. В - стимуляция выработки IFN-γ сортированными CD8 Т-клетками при трансдукции дендритных клеток Ad5-tetOFF-HA125 (за вычетом фоновых значений).
В лунки 96-луночного планшета для ELISPOT помещали по 50 тыс ВМ DC или 50 тыс ВМ DC, трансдуцированных 100 БОЕ Ad5-tetOFF- НА125 на клетку, и по 50 тыс сортированных CD8 Т-клеток от индивидуальных мышей тестируемых групп. Планшет инкубировали при 37°С, 5% CO2 в течение 20 часов и обрабатывали в соответствии с протоколом фирмы производителя. Данные приведены для индивидуальных мышей. Достоверность отличий между группами в тесте one-way ANOVA/Bonferroni обозначена *** р<.001.
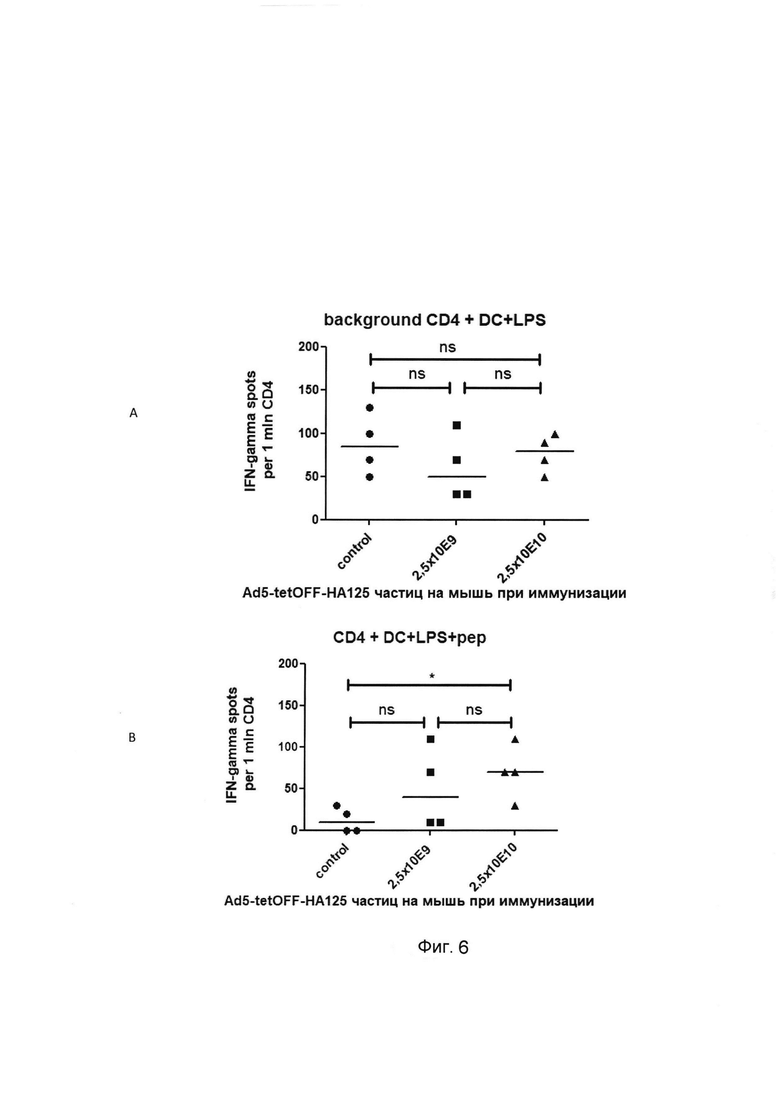
На фигуре 6
показаны результаты определения методом ELISPOT количества CD4+IFN-γ секретирующих клеток Т-клеток, специфичных к эпитопам целевого антигена в комплексе с молекулами МНС II, в селезенке мышей, иммунизированных Ad5-tetOFF-HA125.
Ось абсцисс - экспериментальные группы, ось ординат - число IFN-γ секретирующих клеток в пересчете на 1 млн CD4+ Т-клеток. А - фоновый уровень секреции IFN-γ при сокультивировании сортированных CD4 Т-клеток и аллогенных дендритных клеток, активированных LPS. В - стимуляция выработки IFN-γ сортированными CD4+ Т-клетками при добавлении к активированным дендритным клеткам смеси пептидов из целевого антигена (за вычетом фоновых значений).
В лунки 96-луночного планшета для ELISPOT помещали по 50 тыс ВМ DC, активированных LPS 100 нг/мл, и по 100 тыс сортированных CD4+ Т-клеток от индивидуальных мышей тестируемых групп. В ряд лунок добавляли смесь пептидов целевого антигена. Планшет инкубировали при 37°С, 5% CO2 в течение 20 часов и обрабатывали в соответствии с протоколом фирмы производителя. Данные приведены для индивидуальных мышей. Достоверность отличий между группами в тесте one-way ANOVA/Bonferroni обозначена * р<0.05.
На фигуре 7 изображены:
А - результаты белкового электрофореза;
Б - результат вестерн-блотта.
Обозначения: 1 - отрицательный контроль (клетки трансдуцированные рекомбинантным вирусом гриппа без экспрессионной кассеты), 2 - белок НА125 от клеток трансдуцированных рекомбинантным аденовирусом Ad-HA125 в денатурирующем буфере, 3 - белок НА125 от клеток трансдуцированных рекомбинантным аденовирусом Ad-HA125 в не денатурирующем буфере, М - маркер молекулярного веса Amersham (кат. № RPN756E), 4 - белок НА от штамма A/California/04/2009 H1N1 в денатурирующем буфере, 5 - белок НА от штамма A/California/04/2009 H1N1 в не денатурирующем буфере, 6 - белок НА от штамма A/Japan/305/1957 H2N2 в денатурирующем буфере, 7 - белок НА от штамма A/Japan/305/1957 H2N2 в не денатурирующем буфере, 8 - белок НА от штамма A/Egypt/N05056/2009 H5N1 в денатурирующем буфере, 9 - белок НА от штамма A/Egypt/N05056/2009 H5N1 в не денатурирующем буфере.
В треке №3 видна полоса в районе 130-250 кДа (расчетная масса белка 190 кДа), она соответствует полосам из положительных контролей - треки 5, 7 и 9. В треках 2, 4, 6 и 8 нет полосы, так как эпитоп к антителу CR9114 является конформационным и в денатурирующих условиях четвертичная конформация разрушается. Разрушение конформации белка видно по результатам белкового электрофореза - масса мономера в среднем составляет 63,4 кДа (треки 4, 6 и 8).
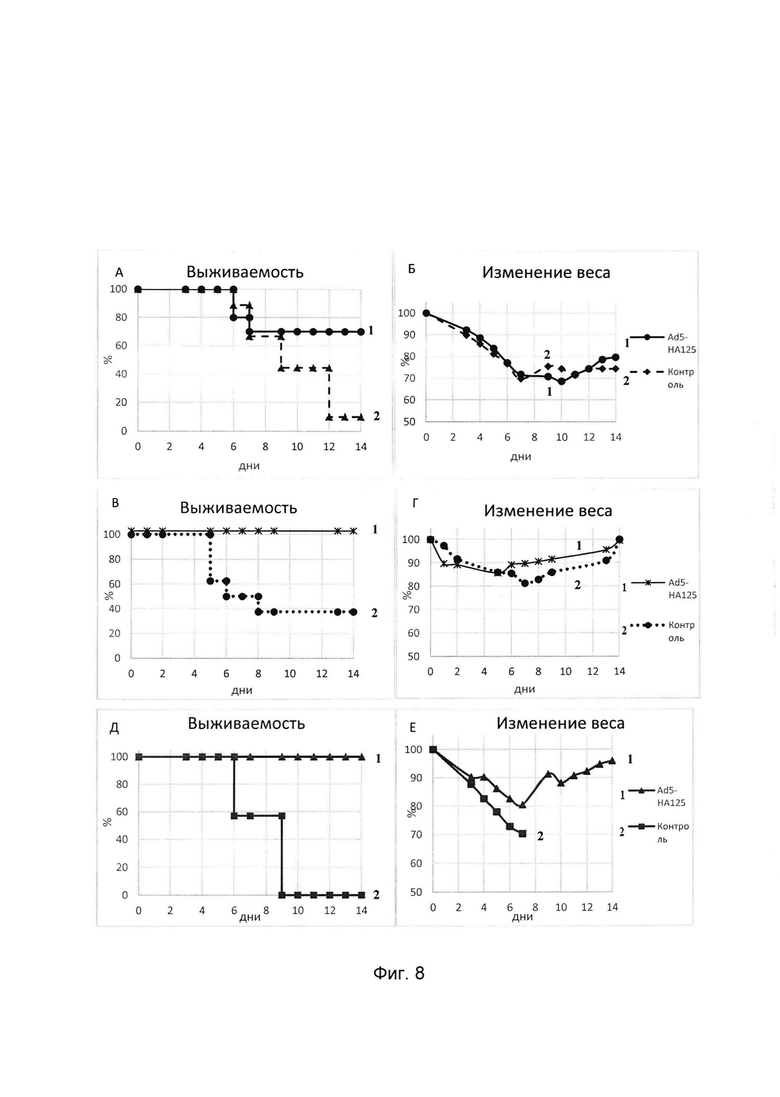
На фигуре 8
представлены результаты оценки иммуногенных свойств иммуногенного препарата, содержащего штамм рекомбинантной псевдоаденовирусной частицы на основе генома аденовируса человека 5 серотипа, несущей ген консенсусной последовательности гемагглютинина вируса гриппа А субтипов Н1, Н2, Н5, обогащенный В и Т-клеточными эпитопами вируса гриппа:
А - кривые выживаемости мышей, иммунизированных Ad5-НА125, после заражения 15ЛД50 вируса гриппа А /California/20/09H1N1, (1  - мыши, иммунизированные Ad5-НА125, 2
- мыши, иммунизированные Ad5-НА125, 2  - контрольные мыши, получавшие буфер для хранения аденовирусных препаратов);
- контрольные мыши, получавшие буфер для хранения аденовирусных препаратов);
Б - кривые изменения веса мышей, иммунизированных Ad5-НА125, после заражения 15ЛД50 вируса гриппа A /California/20/09 H1N1 (1  - мыши, иммунизированные Ad5-HA125, 2
- мыши, иммунизированные Ad5-HA125, 2 - контрольные мыши, получавшие буфер для хранения аденовирусных препаратов);
- контрольные мыши, получавшие буфер для хранения аденовирусных препаратов);
В - кривые выживаемости мышей, иммунизированных Ad5-НА125, после заражения 15ЛД50 вируса гриппа A/BlackDuck/NewJersey/1580/78(H2N3), (1  - мыши, иммунизированные Ad5-HA125, 2
- мыши, иммунизированные Ad5-HA125, 2  - контрольные мыши, получавшие буфер для хранения аденовирусных препаратов);
- контрольные мыши, получавшие буфер для хранения аденовирусных препаратов);
Г. - кривые изменения веса мышей после заражения мышей вирусом гриппа A/BlackDuck/NewJersey/1580/78(H2N3) (1  - мыши, иммунизированные Ad5-HA125, 2
- мыши, иммунизированные Ad5-HA125, 2  - контрольные мыши, получавшие фармацевтически приемлемый буфер;
- контрольные мыши, получавшие фармацевтически приемлемый буфер;
Д - кривые выживаемости мышей после заражения вирусом гриппа A/Duck(Mallard)/Pensylvania/10218/84 H5N2 (1  - мыши, иммунизированные Ad5-HA125, 2
- мыши, иммунизированные Ad5-HA125, 2  - контрольные мыши, получавшие фармацевтически приемлемый буфер;
- контрольные мыши, получавшие фармацевтически приемлемый буфер;
Е - кривые изменения веса мышей после заражения мышей вирусом гриппа A/Duck(Mallard)/Pensylvania/10218/84 H5N2 (1  -мыши, иммунизированные Ad5-HA125, 2
-мыши, иммунизированные Ad5-HA125, 2  - контрольные мыши, получавшие фармацевтически приемлемый буфер.
- контрольные мыши, получавшие фармацевтически приемлемый буфер.
Реализация изобретения
Использование вирусных векторов является перспективным направлением при разработке вакцин. Наиболее хорошо изученными являются векторы на основе рекомбинантных аденовирусов. По данным базы данных Gene Therapy Clinical Trials World (provided by the Journal of Gene Medicine) http://www.abedia.com/wiley/vectors.php в 20% протоколов клинических испытаний в качестве вектора использовали аденовирусы.
В качестве рекомбинантного вектора по изобретению использовали аденовирусный вектор на основе аденовируса человека 5 серотипа - рекомбинантые псевдоаденовирусные частицы. Рекомбинантные аденовирусы обладают рядом преимуществ, благодаря которым они используются в качестве векторов, экспрессирующих целевые антигены, в частности белки вируса гриппа для получения кандидатных гриппозных вакцин. Геном большинства аденовирусов хорошо охарактеризован и прост для манипуляций, ДНК аденовируса остается в экстрахромосомной форме. Векторы способны трансдуцировать, как делящиеся, так и постмитотические клетки. Рекомбинантные аденовирусы могут быть получены в количестве более 1010 вирусных (физических) частиц/мл, что позволяет легко масштабировать технологический процесс их получения, также они не способны к размножению, кроме как в особых условиях, возможных только в специальных линиях клеток in vitro; обеспечивают высокий уровень экспрессии целевого гена в клетке-мишени. При иммунизации они обеспечивает индукцию как клеточного, так и гуморального иммунного ответа против вируса гриппа и выводятся из организма в течение 4-5 недель. Еще одним преимуществом является то, что процесс получения нового рекомбинантного аденовируса занимает несколько недель, что может позволить быстро реагировать на меняющуюся эпидемиологическую обстановку в максимально сжатые сроки, чего нельзя сделать при получении классических вакцинных штаммов.
В качестве трансгена в рекомбинантные псевдоаденовирусные частицы встраивали ген консенсусной последовательности гемагглютинина вируса гриппа А субтипов Н1, Н2, Н5, предварительно обогащенный В и Т-клеточными эпитопами вируса гриппа. Обогащение В и Т-клеточными эпитопами гена консенсусной последовательности гемагглютинина вируса гриппа А субтипов Н1, Н2, Н5 позволило индуцировать иммунитет широкого спектра действия ко всем вирусам гриппа внутри каждого из субтипов Н1, Н2, Н5, а также вновь появляющимся в будущем новым вирусам гриппа указанных субтипов.
Для решения поставленной технической задачи были проведены следующие работы:
1. Получен ген консенсусной последовательности гемагглютинина вируса гриппа А субтипов Н1, Н2, Н5, обогащенный В и Т-клеточными эпитопами вируса гриппа.
2. Сконструирован штамм рекомбинантной псевдоаденовирусной частицы на основе генома аденовируса человека 5 серотипа, несущий в своем геноме экспрессионную кассету гена консенсусной последовательности гемагглютинина вируса гриппа А субтипов Н1, Н2, Н5, обогащенного В и Т-клеточными эпитопами вируса гриппа.
3. Получен иммуногенный препарат, содержащий штамм рекомбинантной псевдоаденовирусной частицы на основе генома аденовируса человека 5 серотипа, несущей ген консенсусной последовательности гемагглютинина вируса гриппа А субтипов Н1, Н2, Н5, обогащенный В и Т-клеточными эпитопами вируса гриппа.
4. Определено наличие экспрессии целевого белка методом иммуноблоттинга.
5. Оценен гуморальный ответ на введение иммуногенного препарата, содержащего штамм рекомбинантной псевдоаденовирусной частицы на основе генома аденовируса человека 5 серотипа, несущей ген консенсусной последовательности гемагглютинина вируса гриппа А субтипов Н1, Н2, Н5, обогащенный В и Т-клеточными эпитопами вируса гриппа, методом ИФА.
6. Оценена индукция Т- и В- клеточного иммунного ответа у мышей, иммунизированных иммуногенным препаратом, содержащим штамм рекомбинантных псевдоаденовирусных частиц по изобретению.
7. Оценены протективные свойства иммуногенного препарата, содержащего рекомбинантные псевдоаденовирусные частицы по изобретению.
Пример 1.
Получение гена консенсусной последовательности гемагглютинина вируса гриппа А субтипов Н1, Н2, Н5, обогащенного В и Т-клеточными эпитопами вируса гриппа
В данном примере рассмотрен ход работ по получению аминокислотной и нуклеотидной последовательностей трансгена (гена консенсусной последовательности гемагглютинина вируса гриппа А субтипов Н1, Н2, Н5, обогащенного В и Т-клеточными эпитопами вируса) гриппа который в следующем примере будет встроен в рекомбинантную псевдоаденовирусную частицу.
Получение консенсусной последовательности гена гемагглютинина происходит путем слияния консервативных участков гемагглютининов вируса гриппа А субтипов Н1, Н2 и Н5, при этом общая длина рекомбинантного белка равна нативному гемагглютинину вируса гриппа А (1701 п.о.), содержит Т-клеточные эпитопы и конформационный В-клеточный эпитоп в стебельной части.
Процесс получения заключается в следующих последовательных действиях:
1) Из общедоступной базы генетических данных Influenza virus Resource (https://www.ncbi.nlm.nih.gov/genomes/FLU/Database/nph-select.cgi?go=database) отбирали аминокислотные последовательности гемагглютининов вирусов гриппа А трех субтипов Н1, Н2 и Н5, выделенные от человека, выравнивали друг относительно друга с помощью общедоступной компьютерной программы, при этом сохраняя известный конформационный В-клеточный эпитоп в стебельной части, и, таким образом, получили «консенсусные» (усредненные) последовательности для каждого субтипа отдельно;
Всего количество включенных последовательностей составляло:
Н1: 5991 белковых сиквенсов;
Н5: 218 белковых сиквенсов;
Н2: 80 белковых сиквенсов.
2) С помощью общедоступной базы данных по распространенности аллельных частот - The Allele Frequency Net Database (http://www.allelefrequencies.net/), выявляли наиболее часто встречающиеся аллели молекул HLA I-класса и HLA II-класса у жителей РФ и оставляли эпитопы с высокой афинностью, сокращая одинаковые последовательности эпитопов, которые распознаются разными аллелями;
Согласно этой базе данных у жителей России наиболее часто встречаются следующие серотипы молекул HLA:
HLA I-класса: А*02, А*24, С*03, С*04, С*07;
HLA II-класса: DQA1*01, DQB1*06; DRB1*01, DRB1*04, DRB1*07, DRB1*11, DRB1*15.
3) Далее с помощью доступных онлайн сервисов определяли наличие эпитопов в консенсусных последовательностях Н1, Н2 и Н5 для отобранных ранее аллелей HLA I-класса (http://www.cbs.dtu.dk/services/NetMHCcons/) и HLA II-класса (http://www.cbs.dtu.dk/services/NetMHCII/). Таким образом было определено наличие следующих эпитопов: HLA 1-класса:
HLA-A0201, HLA-A0202, HLA-A0203, HLA-A0205, HLA-A0206, HLA-A0207, HLA-A0211, HLA-A0212, HLA-A0216, HLA-A0217, HLA-A0219, HLA-A0250, HLA-A2402, HLA-A2403, HLA-C0303, HLA-C0401, HLA-C0701, HLA-C0702
HLA II-класса:
HLA-DQA10101-DQB10501, HLA-DQA10102-DQB10602, HLA-DRB10101, HLA-DRB10401, HLA-DRB10404, HLA-DRB10405, HLA-DRB10701, HLA-DRB11101, HLA-DRB11501.
4) В общеизвестной компьютерной программе (например, Geneious) накладывали последовательности выявленных эпитопов аллелей HLA I-класса и HLA II-класса, на каждую из трех консенсусных последовательностей вируса гриппа субтипов Н1, Н2 и Н5, определяли наличие соответствующих эпитопов и отмечали их местоположение;
Для выполнения данной работы создавали базу данных эпитопов от трех сиквенсов, принадлежащие одному HLA-классу, переносили на одну страницу Xlsx-файла и сохраняли только эпитопы с высокой аффинностью. Повторяющиеся значения удаляли, и конечные данные переносили в общеизвестную компьютерную программу (например, Geneious) (т.е. удаляли одинаковые сиквенсы эпитопов к одному и тому же аллелю, например RMNYYWTLV HLA-A0202 и RMNYYWTLV HLA-A0202). В компьютерной программе сохраняли только уникальные последовательности: то есть сокращали одинаковые последовательности эпитопов, которые распознаются разными аллелями (например, последовательность RMNYYWTLV распознают такие аллели как HLA-A0201, HLA-A0202, HLA-A0203, HLA-A0206, HLA-A0207, HLA-A0211, HLA-A0212, HLA-A0216, HLA-А0250, HLA-C0401). Таким образом, были получены две базы данных эпитопов: к HLA I-классу и HLA II-классу.
5) С помощью общедоступной базы данных иммунологических эпитопов из отобранных ранее Т-клеточных эпитопов HLA отбираются все известные эпитопы с экспериментально подтвержденной активностью и также на каждую из трех консенсусных последовательностей вируса гриппа субтипов Н1, Н2 и Н5 и определяют наличие соответствующих эпитопов и отмечают их местоположение;
В процессе отбирались все известные Т-клеточные эпитопы, дающие положительный результат (связывания Т-клетками HLA-тетрамеров, продукция различных воспалительных цитокинов и т.д.) в исследованиях.
6) Три консенсусные последовательности гемагглютининов вирусов гриппа А трех субтипов Н1, Н2 и Н5 выравнивали относительно друг друга;
7) Собирали консенсусную последовательность гемагглютинина вируса гриппа А путем слияния того участка гена, на котором отмечено наибольшее количество Т-клеточных эпитопов, выбранного каждый раз из одной из трех консенсусных последовательностей гемагглютининов вирусов гриппа А субтипов Н1, Н2 и Н5, и последовательно включали в собираемую консервативную последовательность;
Были использованы следующие обозначения, например HLA-DRB10101 (3): HLA-DRB10101 - это название эпитопа; (3) -количество аллелей, которое распознает данный эпитоп.
В качестве примера представлен процесс получения фрагмента консенсусной последовательности из:
1) Н1 консенсуса, определены следующие:
а) HLA I-класса: HLA-A0211 (3), HLA-C0303 (2), HLA-C0401, HLA-A0206 (4);
б) HLA II-класса: HLA-DRB10101 (3), HLA-DRB10701, HLA-DRB10401 (4), HLA-DRB-10101 (3), HLA-DRB10701, HLA-DRB10101, HLA-DRB10701, HLA-DRB10701;
2) Н2 консенсуса определены следующие:
а) HLA I-класса: HLA-C0303, HLA-A0201 (11);
б) HLA II-класса: HLA-DRB10101 (3), HLA-DRB10101 (2), HLA-DRB10101, HLA-DRB10701;
3) Н5 консенсуса определены следующие:
а) HLA I-класса: HLA-A0211 (5), HLA-A0201 (10), HLA-A0201 (5);
б) HLA II-класса: HLA-DRB10101 (2), HLA-DRB10701, HLA-DRB10404(2), HLA-DRB10101, HLA-DRB10701.
Таким образом, при сравнительном анализе трех ранее составленных консенсусных последовательностей генов гемагглютинина субтипов в состав первого фрагмента консенсусной последовательности был включен участок Н1 консенсуса:
а) HLA I-класса: HLA-A0211 (3), HLA-C0303 (2), HLA-C0401, HLA-A0206 (4);
б) HLA II-класса: HLA-DRB10101 (3), HLA-DRB10701, HLA-DRB10401 (4), HLA-DRB-10101 (3), HLA-DRB10701, HLA-DRB10101, HLA-DRB10701, HLA-DRB10701.
По такому же принципу был составлен второй участок оптимизированной последовательности:
1) из Н1 консенсуса определены следующие:
а) HLA I-класса: HLA-C0401; HLA-A0250;
б) HLA II-класса: HLA-DRB10101, HLA-DRB10101 (2), HLA-DRB10101, HLA-DRB10401 (2);
2) из Н2 консенсуса определены следующие:
а) HLA I-класса: HLA-C0401, HLA-C0701; HLA-C0701 (2);
б) HLA II-класса: HLA-DRB10701, HLA-DRB10101, HLA-DRB10101 (2), HLA-DRB10101;
3) из Н5 консенсуса определены следующие:
а) HLA I-класса: HLA-C0401, HLA-A0207 (2), HLA-A0250, HLA-А0211 (2), HLA-A0205 (6), HLA-C0702, HLA-A0206 (2).
б) HLA II-класса: HLA-DRB10701, HLA-DRB10701, HLA-DRB10101 (2), HLA-DRB10101.
На данном этапе после сравнения трех консенснусных последовательностей был выбран участок Н5 консенсуса для включения в оптимизированную последовательность:
а) HLA I-класса: HLA-C0401, HLA-A0207 (2), HLA-A0250, HLA-А0211 (2), HLA-A0205 (6), HLA-C0702, HLA-A0206 (2);
б) HLA II-класса: HLA-DRB10701, HLA-DRB10701, HLA-DRB10101 (2), HLA-DRB10101.
По такому же принципу были определены остальные участки оптимизированной последовательности.
8) Полученную консервативную аминокислотную последовательность гемагглютинина вируса гриппа А, содержащую участки гемагглютининов вирусов гриппа субтипов Н1, Н2 и Н5 и максимально насыщенную консервативными (В- и Т-клеточными) эпитопами, переводят в соответствующую ей нуклеотидную последовательность, в которой оптимизируют кодоны общеизвестным способом для более эффективной экспрессии в клетках человека.
Таким образом, в результате работ была получена сначала аминокислотная (SEQ ID NO: 1), а затем и нуклеотидная (SEQ ID NO: 2) последовательности гемагглютинина вируса гриппа, содержащего части гемагглютининов вирусов гриппа субтипов Н1, Н2 и Н5 и максимально насыщенного консервативными (В- и Т-клеточными) эпитопами вируса гриппа.
Для теоретической оценки наличия иммуногенных свойств у полученного гена далее была проведена оценка встречаемости среди населения РФ Т-клеточных эпитопов, входящих в сконструированную консервативную последовательность гемагглютинина НА125 с помощью онлайн ресурса располагающегося в базе данных иммунологических эпитопов (http://tools.iedb.org/tools/population/iedb_input). Установлено, что доля жителей России, распознающих хотя бы один эпитоп, составляет - 94,69%, среднее число распознаваемых человеком эпитопов - 15,70%, а также минимальное число эпитопов, распознаваемых 90% людей - 2,94%. Полученные результаты говорят о высоких иммуногенных свойствах разработанного гена консенсусной последовательности гемагглютинина вируса гриппа А субтипов Н1, Н2, Н5, обогащенной В и Т-клеточными эпитопами вируса гриппа.
На фиг. 1
представлено схематическое изображение фрагмента полученной консенсусной последовательности гена гемагглютинина вируса гриппа, состоящего из различных участков гемагглютининов субтипов Н1, Н2 и Н5 (НА 125).
Пример 2.
Конструирование штамма рекомбинантной
псевдоаденовирусной частицы на основе генома аденовируса человека 5 серотипа, несущей в своем геноме экспрессионную кассету гена консенсусной последовательности гемагглютинина вируса гриппа А субтипов Н1, Н2, Н5, обогащенного В и Т-клеточными эпитопами вируса гриппа.
В данном примере рассмотрен процесс встраивания полученного гена консенсусной последовательности гемагглютинина вируса гриппа А субтипов Н1, Н2, Н5, обогащенного В и Т-клеточными эпитопами вируса гриппа в рекомбинантную псевдоаденовирусную частицу на основе генома аденовируса человека 5 серотипа.
Способом получения штамма рекомбинантной псевдоаденовирусной частицы на основе генома аденовируса человека 5 серотипа, несущей ген консенсусной последовательности гемагглютинина вируса гриппа А субтипов Н1, Н2, Н5, обогащенного В и Т-клеточными эпитопами вируса гриппа была гомологичная рекомбинация между плазмидой, несущей кассету с системой Tet-off, позволяющей блокировать экспрессию подконтрольных генов путем добавления доксициклина и ген НА125 ДНК аденовируса 5 серотипа с делецией Е1 области в клетках E.coli штамма BJ5183, сборка рекомбинантных псевдоаденовирусных частиц в клетках линии HEK293 (Human embryonic kidney 293) после трансфекции рекомбинантной ДНК Ad5-tetOFF-E3-HA125.
Процесс получения штамма рекомбинантной псевдоаденовирусной частицы на основе генома аденовируса человека 5 серотипа, несущей ген консенсусной последовательности гемагглютинина вируса гриппа А субтипов Н1, Н2, Н5, обогащенного В и Т-клеточными эпитопами вируса гриппа состоял из нескольких основных этапов, суть которых известна специалисту данной области, основывался на общеизвестных методиках (например, Sambrook J. et al., Molecular cloning: a laboratory manual, 3rd ed., Russell, 2001, том 1, 2, 3 - Молекулярное клонирование - лабораторное руководство) и поэтому в данном примере подробно не рассматривается, указывая лишь на самые основные этапы:
1) Постановка трансфекции пермессивной культуры эукариотических клеток плазмидой, несущей ген консенсусной последовательности гемагглютинина вируса гриппа А, обогащенный В и Т-клеточными эпитопами;
2) Получение рекомбинантной псевдоаденовирусной частицы на основе генома аденовируса человека 5 серотипа с кассетой гена-регулятора ТТА промотора TETOFF в ЕЗ области аденовирусного генома;
3) Получение штамма рекомбинантной псевдоаденовирусной частицы на основе генома аденовируса человека 5 серотипа с кассетой целевого гена в Е1 области аденовирусного генома;
В результате проведенных генно-инженерных манипуляций были получены клоны плазмиды pAdTetHA125_E3tTA, содержащей ген гемагглютинина по изобретению, под контролем TetOff промотора и ген-регулятор tTA промотора TetOff в делетированной Е3 области аденовирусного генома.
Далее эти плазмиды использовались для трансфекции пермиссивной клеточной культуры клеток HEK293 с последующим получением определенного титра штамма рекомбинантной псевдоаденовирусной частицы, который применялся для дальнейшего наращивания.
4) Наращивание для депонирования полученного штамма рекомбинантной псевдоаденовирусной частицы на основе генома аденовируса человека 5 серотипа, несущей ген консенсусной последовательности гемагглютинина вируса гриппа А субтипов Н1, Н2, Н5, обогащенный В и Т-клеточными эпитопами вируса гриппа.
Штамм рекомбинантных псевдоаденовирусных частиц наращивали в пермиссивной культуре клеток почки человека HEK293 в присутствии доксициклина после трансдукции клеток линии HEK293. Для роста клеток линии HEK293 в адгезионной культуре использовали среду DMEM (Invitrogen, №52100-047, США), содержащую 25 мМ глюкозы, 4 мМ L-глутамина и 10% эмбриональной бычьей сыворотки, специальный инкубатор с поддержанием температуры +37°С и 5% CO2.
Полученный штамм рекомбинантных псевдоаденовирусных частиц может храниться при температуре не выше -70°С в среде для культивирования DMEM (Invitrogen, кат.№52100-047, США) с 10% эмбриональной бычьей сыворотки (Ну Clone, кат. №SV30160.03, Великобритания), собранной вместе с клетками HEK293 через 40-46 часов после трансдукции клеток линии HEK293.
Клеточную линию 293-HEK (клетки почки эмбриона человека, human embryo kidney) использовали для определения титра рекомбинантных псевдоаденовирусных частиц методом бляшкообразования.
По окончании наращивания активность (продуктивность) полученного штамма рекомбинантных псевдоаденовирусных частиц на основе генома аденовируса человека 5 серотипа, несущих ген консенсусной последовательности гемагглютинина вируса гриппа А субтипов Н1, Н2, Н5, обогащенный В и Т-клеточными эпитопами вируса гриппа, составляла от 5×107- до 108 акт. ед./мл (бляшкообразующих единиц на 1 мл культуры).
Получение штамма рекомбинантной псевдоаденовирусной частицы на основе генома аденовируса человека 5 серотипа, несущей ген консенсусной последовательности гемагглютинина вируса гриппа А субтипов Н1, Н2, Н5, обогащенный В и Т-клеточными эпитопами вируса гриппа, было подтверждено результатами ПЦР со специфическими праймерами (представлен на фигуре 2).
Штамм рекомбинантной псевдоаденовирусной частицы на основе генома аденовируса человека 5 серотипа, несущей ген консенсусной последовательности гемагглютинина вируса гриппа А субтипов Н1, Н2, Н5, обогащенный В и Т-клеточными эпитопами вируса гриппа (Ad5-tetOFF-E3-HA125) зарегистрирован в Государственной коллекции микроорганизмов при ФГБУ «НИЦ эпидемиологии и микробиологии имени Н.Ф. Гамалеи» (Музей генно-инженерно-модифицированных микроорганизмов) под номером 21. Продукт, синтезируемый штаммом - консенсусный гемагглютинин вируса гриппа А, обогащенный В и Т-клеточными эпитопами (НА125).
Пример 3.
Получение иммуногенного препарата, содержащего штамм рекомбинантной псевдоаденовирусной частицы на основе генома аденовируса человека 5 серотипа, несущей ген консенсусной последовательности гемагглютинина вируса гриппа А субтипов Н1, Н2, Н5, обогащенный В и Т-клеточными эпитопами вируса гриппа.
Получение иммуногенного препарата, содержащего штамм рекомбинантных псевдоаденовирусных частиц на основе генома аденовируса человека 5 серотипа, несущих ген консенсусной последовательности гемагглютинина вируса гриппа А субтипов Н1, Н2, Н5, обогащенный В и Т-клеточными эпитопами вируса гриппа необходимо для изучения его иммуногенных и протективных свойств, в качестве обоснования возможности применения в противогриппозных вакцинах.
Для получения иммуногенного препарата, содержащего штамм рекомбинантных псевдоаденовирусных частиц использовали культуру клеток почки эмбриона человека HEK 293, адаптированную для роста в суспензии. Клеточную суспензию, полученную в примере 2, содержащую штамм рекомбинантных псевдоаденовирусных частиц Ad5-tetOFF-E3-HA125 с титром 5×107-108 акт. ед./мл, использовали для наращивания с целью приготовления иммуногенного препарата с эффективным количеством рекомбинантных псевдоаденовирусных частиц.
Для наработки необходимых титров рекомбинантных псевдоаденовирусных частиц волновой биореактор с 4500,0 мл суспензии пермиссивной клеточной культуры 293HEK засевали клеточной суспензией объемом 500,0 мл, содержащей рекомбинантные псевдоаденовирусные частицы с титром 5×107-108 акт. ед./мл. Для наращивания рекомбинантных псевдоаденовирусных частиц Ad5-tetOFF-E3-HA125 и достижения титра 2×108 акт. ед./мл, их культивировали в течение 48 часов в присутствие доксициклина в концентрации 5,0 мкг/мл. Затем клеточную массу очищали с помощью общеизвестных методов ультрафильтрации и высокоэффективной жидкостной хроматографии в несколько этапов:
1) Осаждение клеточной массы центрифугированием;
2) Перемораживание осадка;
3) Обработка бензоназой;
4) Центрифугирование ля удаления клеточного дебриса;
5) Ультрафильтрация супернатанта;
6) Анионобменная хроматография;
7) Эксклюзионная хроматогафия;
8) Нормальная фильтрация.
Для стерилизации полученный препарат фильтровали через систему фильтров с размером пор 0,22 мкМ и разбавляли стерильным фармацевтически приемлемым буферным раствором, например: 10 mМ TrisHCl, 75 mM NaCl, 1 mM MgCl2, 5% сахароза, 0,05% полисорбат 80, 0,5% этанол, 100 мкм ЭДТА, рН 8.0, с получением препарата с заданной активностью 107-109 акт. ед. рекомбинантных псевдоаденовирусных частиц Ad5-tetOFF-Е3-НА125 на дозу 0,5 мл.
Таким образом, представленная технология позволяет выпускать иммуногенный препарат, содержащий на дозу 0,5 мл:
- рекомбинантные псевдоаденовирусные частицы Ad5-tetOFF-E3-HA125 - 107-109 акт. ед.;
- фармацевтически приемлемый буферный раствор - до 0,5 мл.
В конкретном примере произведенный иммуногенный препарат содержал 4,0×108 акт. ед./дозу 0,5 мл рекомбинантных псевдоаденовирусных частиц на основе генома аденовируса человека 5 серотипа, несущих ген консенсусной последовательности гемагглютинина вируса гриппа А субтипов Н1, Н2, Н5, обогащенный В и Т-клеточными эпитопами вируса гриппа. Данный иммуногенный препарат был использован для дальнейших исследований экспрессии целевого гена, иммуногенности и протективных свойств.
Пример 4.
Определение наличия экспрессии целевого белка методом иммуноблоттинга
В примере показано определение экспрессии целевого белка в месте введения иммуногенного препарата, содержащего штамм рекомбинантной псевдоаденовирусной частицы на основе генома аденовируса человека 5 серотипа, несущей ген консенсусной последовательности гемагглютинина вируса гриппа А субтипов Н1, Н2, Н5, обогащенный В и Т-клеточными эпитопами вируса гриппа методом иммуноблоттинга. Для выявления и идентификации в тканях экспрессии целевого антигена вируса гриппа - консенсусного гемагглютинина опытным мышам однократно интраназально вводили иммуногенный препарат, содержащий штамм рекомбинантных псевдоаденовирусных частиц в дозе 4,0×107 акт. ед. в объеме 50 мкл на животное. Для контроля использовали мышей, иммунизированных вирусным вектором, не несущим трансгена Ad5-null и интактные мыши.
Проводили анализ методом электрофореза с последующим иммуноблоттингом. Для этого через 72 часа после интраназальной иммунизации мыши были бескровно умерщвлены и из них были извлечены ткани носовой перегородки и легких.
Ткани помещали в лизирующий буфер CCLR (Е1531 Promega Cell Culture Lysis 5X Reagent), содержащий ингибиторы протеиназ (S8820 SIGMA SIGMAFAST™ Protease Inhibitor Tablets) и гомогенизировали с бусами (19-644 Omni International Garnet Bead Material 0.7mm) двукратно (4 м/с, no 20 секунд). После гомогенизации пробирки инкубировали 10 минут на льду, дебрис осаждали центрифугированием (12000g, 1 мин) и надосадочную жидкость отбирали для дальнейшего анализа. Определение содержания общего белка в образцах проводили по стандартному методу Брэдфорд, на спектрофотометре BioMate 3S (ThermoScientific). Фракционирование белков проводили методом электрофореза в градиентном 10-20% ПААГ-ДСН, в буферной системе Лэммли, с использованием вертикального прибора для минигелей Mini-Protean 3 Cello (BIO-RAD) и готовых пластин геля Mimi-Protein TGX Stain-free Precast Gel S (BIO-RAD). Для анализа экспрессии рекомбинантного белка НА125, несущего антигенные детерминанты конформационного типа, в пробы добавили равный объем 2-кратного диссоциирующего буфера без меркаптоэтанола.
По данным иммуноблоттинга (Фигура 3) в реакции с рекомбинантными антителами в образцах экстрактов тканей дыхательных путей (носовая перегородка и легкие) иммунизированных мышей идентифицирован рекомбинантный белок НА125 (темная полоса на треках 2 и 5, молекулярная масса более 140 кДа, что соответствует гемагглютинину вируса гриппа А). В дыхательных путях контрольных животных экспрессия целевых антигенов отсутствует.
Как видно из представленных результатов, через 72 часа после введения иммуногенного препарата, содержащего штамм рекомбинантных псевдоаденовирусных частиц по изобретению в месте введения (назальная перегородка) и легких иммунизированных мышей наблюдается экспрессия целевых антигенов - гена консенсусной последовательности гемагглютинина вируса гриппа А субтипов Н1, Н2, Н5, обогащенного В и Т-клеточными эпитопами вируса гриппа, что соответствует поставленной цели изобретения.
Пример 5.
Оценка гуморального ответа на введение иммуногенного препарата, содержащего штамм рекомбинантной псевдоаденовирусной частицы на основе генома аденовируса человека 5 серотипа, несущей ген консенсусной последовательности гемагглютинина вируса гриппа А субтипов Н1, Н2, Н5, обогащенного В и Т-клеточными эпитопами вируса гриппа, методом ИФА
Иммуногенные свойства созданного штамма Ad5-tetOFF-E3-НА125 изучались методом ИФА.
Для изучения иммуногенности мышам линии BALB/c (самки весом 18-20 грамм) однократно интраназально вводили препарат Ad5-tetOFF-E3-HA125 в дозе 2×108 акт. ед./мышь. В каждой группе было 6 животных. В качестве контрольной группы использовали мышей, получавших интраназально буферный раствор для хранения аденовирусных препаратов. Через 3 недели после иммунизации у мышей отбирали кровь, получали сыворотки.
Определение титра IgG в сыворотках крови мышей проводили методом непрямого ИФА, широко известного специалисту. В качестве антигенов использовали рекомбинантные гемагглютинины вирусов гриппа A H1N1, H5N2 и H2N2 (A/Egypt/N05056/2009 H5N1 (Sino biological Inc., Китай, кат №1702-V08H), A/California/04/2009 H1N1 (Sino biological Inc., Китай, кат №11055-V08H2), A/Japan/305/1957 H2N2 (Sino biological Inc., Китай, кат №11088-V08H), A/New Caledonia/20/1999 H1N1 (Sino biological Inc., Китай, кат №11683-V08H)).
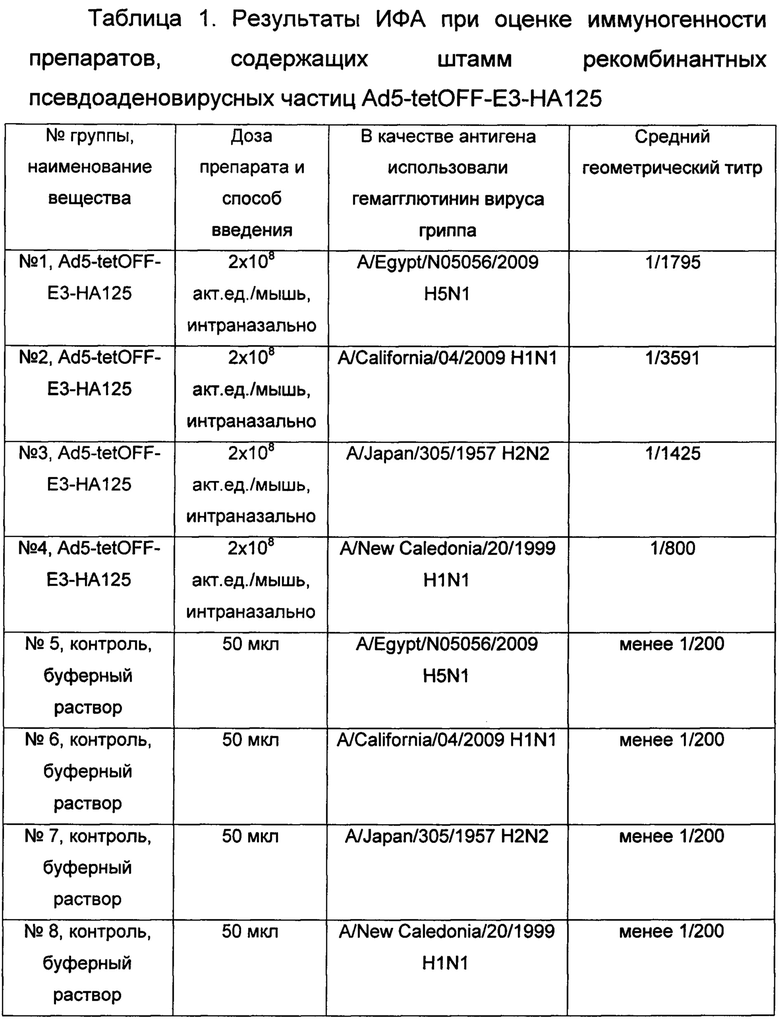
Таким образом, из представленных в таблице результатов следует, что иммуногенный препарат, содержащий штамм рекомбинантных псевдоаденовирусных частиц на основе генома аденовируса человека 5 серотипа, несущих ген консенсусной последовательности гемагглютинина вируса гриппа А субтипов Н1, Н2, Н5, обогащенный В и Т-клеточными эпитопами вируса гриппа вызывает в организме иммунизированных животных индукцию высоких титров специфических антител против различных вирусов гриппа А субтипов Н1, Н5 и Н2.
Пример 6.
Оценка Т- и В- клеточного иммунного ответа у мышей, иммунизированных иммуногенным препаратом, содержащим штамм рекомбинантных псевдоаденовирусных частиц по изобретению
Известно, что наличие специфического клеточного иммунного ответа в отношении различных субтипов вируса гриппа позволяет говорить об индукции иммунитета, позволяющего защищать от множества штаммов вируса гриппа А, в нашем случае ко всем вирусам гриппа А субтипов Н1, Н2 и Н5, что является современным требованием к противогриппозным вакцинам широкого спектра действия.
Для изучения Т- клеточного иммунного ответа у мышей был использован полученный в примере 5 иммуногенный препарат, содержащий Ad5-tetOFF-E3-HA125. Для этой цели применяли метод ELISPOT, позволяющий выявлять Т-лимфоциты, специфичные к белку НА125, по их способности секретировать IFN-γ в ответ на реактивацию дендритными клетками, презентирующими белок НА125. Дендритные клетки получали из костного мозга интактных мышей и культивировали в присутствии гранулоцитарно-макрофагального колониестимулирующего фактора.
Для проведения опыта мышей (Balb/c, самки) иммунизировали однократно интраназально препаратом, содержащим штамм рекомбинантных псевдоаденовирусных частиц, экспрессирующих ген консенсусной последовательности гемагглютинина вируса гриппа А субтипов Н1, Н2, Н5, обогащенный В и Т-клеточными эпитопами вируса гриппа в дозах 4,0×106 акт. ед. и 4,0×107 акт. ед. в объеме 50 мкл. В качестве контрольной группы использованы животные, получившие рекомбинантный аденовирусный вектор, не несущий целевого трансгена и фосфатный буфер. Количество мышей в опытных и контрольной группе составляло по 4 особи, срок после иммунизации составлял 30 дней, после чего мышей эвтаназировали бескровным методом и в асептических условиях извлекали селезенку.
Затем спленоциты реактивировали in vitro дендритными клетками, презентирующими эпитопы антигена НА125 и определяли количество спленоцитов, отвечающих секрецией IFN-γ в ответ на реактивацию. Для реактивации спленоцитов in vitro использовали 2 варианта дендритных клеток. В первом варианте, дендритные клетки предварительно активировали LPS для индукции экспрессии молекул МНС класса II и ко-стимулирующих молекул CD80/CD86. Затем предварительно активированные дендритные клетки инкубировали в присутствии антигенных пептидов:
Пептиды производства фирмы ООО "Алма-Бион"


Пептидные антигены разводили в дистиллированной воде в концентрации 0,3 мг/мл и фильтровали через 0,2 мкм. Аликвоты хранили замороженные при -80°С.
В качестве контроля реактивации использовали дендритные клетки, активированные LPS, без нагрузки антигеном. При втором варианте постановки эксперимента дендритные клетки трансдуцировали рекомбинантным вирусным вектором, экспрессирующим ген НА125 в количестве 100 акт. ед./клетку и по истечению двух суток после трансдукции, использовали их для реактивации спленоцитов (схема постановки эксперимента представлена на фигуре 4). Методом ELISPOT, проводимым в соответствии с представленной на фигуре последовательностью определяли количество CD4 и CD8 Т-клеток, секретирующих IFN-γ.
В результате проведенных исследований установлено, что однократная интраназальная иммунизация препаратом, содержащим штамм рекомбинантных псевдоаденовирусных частиц Ad5-tetOFF-HA125 в дозах 4,0×106 и 4,0×107 акт. ед на мышь вызывает через 30 суток массовое появление в селезенке CD8+ Т-клеток, которые специфически вырабатывающих IFN-γ в ответ на презентацию эпитопов целевого антигена в комплексе с молекулами МНС I. Число IFN-γ секретирующих клеток составляет - около 1500 на 1 млн спленоцитов (0.15% IFN-γ от всех спленоцитов) при иммунизации Ad5-tetOFF-HA125 в дозе 2,5×109 частиц и около 2800 на 1 млн спленоцитов (0.28% IFN-γ от всех спленоцитов) при дозе иммунизации 4,0×107 акт. ед. на мышь. Количество CD4+IFN-γ секретирующих клеток, специфичных к эпитопам целевого антигена в комплексе с молекулами МНС II, было незначительным (Фигуры 5 и 6).
Таким образом, было заключено, что развитие иммунного ответа на антиген НА125, экспрессируемый рекомбинантыми псевдоаденовирусными частицами по изобретению, происходит преимущественно в направлении цитотоксических Т-клеток (CD8+), и протективность, соответственно, обеспечивается преимущественно цитотоксическим действием, что свидетельствует о возникновении иммунитета широкого спектра действия к вирусам гриппа А субтипов Н1, Н2, Н5.
Наличие В-клеточных эпитопов было доказано экспериментальным взаимодействием антигена НА125 с универсальными антителами к стебельной части гемагглютинина - CR9114 по экспрессии рекомбинантного белка НА125 методом вестерн-блотта.
Для получения этого антитела, был получен рекомбинантный аденовирус, несущий ген антитела CR9114. Клетки А549 были трансдуцированны этим рекомбинантным аденовирусом и через 12 часов среду DMEM меняли на новую с 2% содержание сыворотки. Через 5 дней среда была собрана и использована в качестве первичных антител в вестерн-блотте.
В качестве положительного контроля были использованы следующие белки производства Альмабион:
Influenza A H1N1 (A/California/04/2009) Hemagglutinin Protein, SB кат. №11055-M08P2 100 мкг.
Influenza A H2N2 (A/Japan/305/1957) Hemagglutinin Protein, SB кат. №11088-V08H 100 мкг.
Influenza A H5N1 (A/Egypt/N05056/2009) Hemagglutinin Protein, SB кат. №11702-V08H 100 мкг.
В качестве вторичных антител были использованы anti-Human антитела компании Sigma-Aldrich (кат.№I1886-2ML).
Рекомбинантным аденовирусом Ad-HA125 трансдуцировали клетки линии А549, через двое суток клетки собирали, лизировали и ставили вестерн-блотт. Результаты показаны на фигуре 7.
Таким образом, по результатам эксперимента было показано, что, во-первых, универсальное антитело CR9114 распознает эпитоп только в нативной конформации, во-вторых, нативная конформация по крайней мере стебельной части сохраняется в рекомбинантном белке НА125 и, в-третьих, CR9114 способен распознавать конформационный эпитоп в гемагглютинине НА125. Было заключено, что Ad5-tetOFF-E3-НА125 содержит в составе гена гемагглютинина В-клеточный эпитоп.
Пример 7.
Оценка протективных свойств иммуногенной композиции, содержащей штамм рекомбинантных псевдоаденовирусных частиц по изобретению.
Оценку протективных свойств препарата на основе штамма Ad5-HA125 проводили на мышах линии BALB/c (самках весом 18-20 грамм). Мыши были иммунизированы однократно интраназально под легким эфирным наркозом в дозе 2×108 акт. ед./мышь. В каждой группе было 10 животных. В качестве контрольной группы использовали мышей, получавших интраназально буферный раствор препарата. Через 28 дней после иммунизации мышей интраназально под легким эфирным наркозом заражали вирусами гриппа А в дозе 15ЛД50 на животное. Для приготовления нужных разведений вирусов гриппа использовали среду Хенкса с гидролизатом лактальбумина и антибиотиком гентомицином (ПанЭко, Россия). Оценку выживаемости и изменения веса животных проводили в течение 14 дней после заражения. Все работы выполняли в соответствии с «Правилами проведения работ с использованием экспериментальных животных».
Для пробоя мышей были использованы следующие штаммы вирусов гриппа:
A/California/20/09 H1N1
A/BlackDuck/NewJersey/1580/78(H2N3)
A/Duck(Mallard)/Pensylvania/10218/84 H5N2
В ходе опыта иммуногенный препарат, содержащий штамм рекомбинантных псевдоаденовирусных частиц на основе генома аденовируса человека 5 серотипа, несущих ген консенсусной последовательности гемагглютинина вируса гриппа А субтипов Н1, Н2, Н5, обогащенный В и Т-клеточными эпитопами вируса гриппа, продемонстрировал 100% протекцию иммунизированных мышей от заражения вирусом гриппа А субтипа Н5, 100% протекцию при заражении вирусом гриппа А субтипа Н2 и 70% протекцию при заражении вирусом гриппа А субтипа Н1 (Фигура 9).
Полученные данные свидетельствуют об индукции защитного иммунитета в отношении вируса гриппа А субтипов Н1, Н2, Н5.
Иммуногенный препарат на основе штамма рекомбинантной псевдоаденовирусной частицы на основе генома аденовируса человека 5 серотипа Ad5-tetOFF-E3-HA125, несущей ген консенсусной последовательности гемагглютинина вируса гриппа А субтипов Н1, Н2, Н5 может применяться, как самостоятельно в качестве противогриппозной вакцины, так и в качестве одного из действующих веществ многокомпонентной по составу противогриппозной вакцины.
Заявленный штамм рекомбинантной псевдоаденовирусной частицы по изобретению по сравнению с прототипом имеет ряд существенных преимуществ:
- наличие Т-клеточных эпитопов в составе гемагглютинина вируса гриппа позволяет выработать долговременную иммунологическую память на патоген, что позволит для поддержания иммунитета проводить ревакцинации 1 раз в несколько лет, а не 1 раз в год, как предлагается для существующих вакцин;
- присутствуют конформационные В-клеточные эпитопы на консервативные области гемагглютинина, а именно на его стебельную часть. Известно, что гуморальный иммунитет играет основную роль в защите против вируса гриппа и именно на выработку антител к стебельной части НА направленно большинство исследований вирусологов.
- позволяет de-novo активировать наивные Т-хелперы, что, при естественном заражении, во-первых, вызывает появление антител к экзотическому гемагглютинину, а, во-вторых, более быстрое (в 2-3 дня) появление специфических антител, так как известно, что для индукции гуморального иммунного ответа В-лимфоцитам, необходима помощь в активации со стороны хелперных Т-лимфоцитов. При этом более ранние встречи с другим штаммом вируса гриппа, мешают de-novo активации наивных В-лимфоцитов и, следовательно, сероконверсии после иммунизации. Этот феномен известен в литературе как изначальный «антигенный грех» (антигенный импринтинг или иммунологическое старшинство) и его объяснение лежит в нескольких иммунологических механизмах (маскирование эпитопов, конкуренция за антиген и т.д.). Для того чтобы его преодолеть, в целевой антиген рекомбинантной частицы включены эпитопы гемагглютининов Н1, Н2 и Н5 именно для Т-хелперных лимфоцитов.
Таким образом, поставленная задача, а именно, создание штамма рекомбинантной псевдоаденовирусной частицы, способной вызвать протективный иммунитет против всех существующих и вновь появляющихся вирусов гриппа А субтипов Н1, Н2 и Н5, - выполнена.
Промышленная применимость
Все приведенные примеры также подтверждают промышленную применимость заявленного изобретения.
Перечень сокращений
293-HEK - клетки линии эмбриональной почки человека
Ad5 - аденовирус человека 5 серотипа
CD4+- Т-лимфоциты субпопуляции CD4+
CD8+- Т-лимфоциты субпопуляции CD8+
CMV - цитомегаловирус человека
DMEM - Dulbecco modified Eagle's medium; минимальная среда Игла, модифицированная Дельбекко
НА - гемагглютинин
HLA - человеческий лейкоцитарный антиген
М - маркер молекулярного веса
М2 - белок ионного канала
mCMV - минимальный промотор цитомегаловируса человека
МНС - главный комплекс гистосовместимости NP - нуклеопротеин
NS - неструктурный белок вируса гриппа
RCA - репликативно-компетентный аденовирус
ТМВ - тетраметилбензидин индикаторная смесь
TRE - тетрациклин-зависимый элемент
tTA - тетрациклин контролируемый трансактиватор
ДНК - дезоксирибонуклеиновая кислота
ИФА - иммуноферментный анализ
ЛД50 - 50% летальная доза
п.о. - пар оснований
ПЦР - полимеразная цепная реакция
РНК - рибонуклеиновая кислота
ЦПД - цитопатическое действие.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> федеральное государственное бюджетное учреждение «Национальный исследовательский центр эпидемиологии и микробиологии имени почетного академика Н.Ф. Гамалеи» Министерства здравоохранения Российской Федерации
<120> Штамм рекомбинантной псевдоаденовирусной частицы на основе генома аденовируса человека 5 серотипа Ad5-tetOFF-E3-HA125, несущей ген консенсусной последовательности гемагглютинина вируса гриппа А субтипов H1,Н2,Н5 для создания противогриппозных иммуногенных препаратов, способ получения гена
<160> 2
<170> BISSAP 1.3.6
<210> 1
<211> 4275
<212> DNA
<213> Influenza virus
<400> 1
cgtgaggctc cggtgcccgt cagtgggcag agcgcacatc gcccacagtc cccgagaagt 60
tggggggagg ggtcggcaat tgaaccggtg cctagagaaa gtggcgcggg gtaaactggg 120
aaagtgatgt cgtgtactgg ctccgccttt ttcccgaggg tgggggagaa ccgtatataa 180
gtgcagtagt cgccgtgaac gttctttttc gcaacgggtt tgccgccaga acacaggtaa 240
gtgccgtgtg tggttcccgc gggcctggcc tctttacggg ttatggccct tgcgtgcctt 300
gaattacttc cacgcccctg gctgcagtac gtgattcttg atcccgagct tcgggttgga 360
agtgggtggg agagttcgag gccttgcgct taaggagccc cttcgcctcg tgcttgagtt 420
gaggcctggc ttgggcgctg gggccgccgc gtgcgaatct ggtggcacct tcgcgcctgt 480
ctcgctgctt tcgataagtc tctagccatt taaaattttt gatgacctgc tgcgacgctt 540
tttttctggc aagatagtct tgtaaatgcg ggccaagatc tgcacactgg tatttcggtt 600
tttggggccg cgggcggcga cggggcccgt gcgtcccagc gcacatgttc ggcgaggcgg 660
ggcctgcgag cgcggccacc gagaatcgga cgggggtagt ctcaagctgg ccggcctgct 720
ctggtgcctg gcctcgcgcc gccgtgtatc gccccgccct gggcggcaag gctggcccgg 780
tcggcaccag ttgcgtgagc ggaaagatgg ccgcttcccg gccctgctgc agggagctca 840
aaatggagga cgcggcgctc gggagagcgg gcgggtgagt cacccacaca aaggaaaagg 900
gcctttccgt cctcagccgt cgcttcatgt gactccacgg agtaccgggc gccgtccagg 960
cacctcgatt agttctcgag cttttggagt acgtcgtctt taggttgggg ggaggggttt 1020
tatgcgatgg agtttcccca cactgagtgg gtggagactg aagttaggcc agcttggcac 1080
ttgatgtaat tctccttgga atttgccctt tttgagtttg gatcttggtt cattctcaag 1140
cctcagacag tggttcaaag tttttttctt ccatttcagg tgtcgtgaaa actaccccta 1200
aaagccagga tccaccatgc agtggacctc cctcctgctg ctggcagggc tcttctccct 1260
ctcccaggcc cagtatgaag atgaccctca ttggtggttc cactacctcc gcagccagca 1320
gtccacctac tacgatccct atgaccctta cccgtatgag acctacgagc cttaccccta 1380
tggggtggat gaagggccag cctacaccta cggctctcca tcccctccag atccccgcga 1440
ctgcccccag gagtgcgact gcccacccaa cttccccacg gccatgtact gtgacaatcg 1500
caacctcaag tacctgccct tcgttccctc ccgcatgaag tatgtgtact tccagaacaa 1560
ccagatcacc tccatccagg aaggcgtctt tgacaatgcc acagggctgc tctggattgc 1620
tctccacggc aaccagatca ccagtgataa ggtgggcagg aaggtcttct ccaagctgag 1680
gcacctggag aggctgtacc tggaccacaa caacctgacc cggatgcccg gtcccctgcc 1740
tcgatccctg agagagctcc atctcgacca caaccagatc tcacgggtcc ccaacaatgc 1800
tctggagggg ctggagaacc tcacggcctt gtacctccaa cacaatgaga tccaggaagt 1860
gggcagttcc atgaggggcc tccggtcact gatcttgctg gacctgagtt ataaccacct 1920
tcggaaggtg cctgatgggc tgccctcagc tcttgagcag ctgtacatgg agcacaacaa 1980
tgtctacacc gtccccgata gctacttccg gggggcgccc aagctgctgt atgtgcggct 2040
gtcccacaac agtctaacca acaatggcct ggcctccaac accttcaatt ccagcagcct 2100
ccttgagcta gacctctcct acaaccagct gcagaagatc cccccagtca acaccaacct 2160
ggagaacctc tacctccaag gcaataggat caatgagttc tccatcagca gcttctgcac 2220
cgtggtggac gtcgtgaact tctccaagct gcaggtgctg cgcctggacg ggaacgagat 2280
caagcgcagc gccatgcctg ccgacgcgcc cctctgcctg cgccttgcca gcctcatcga 2340
gatctaagaa ttccctgtga cccctcccca gtgcctctcc tggccctgga agttgccact 2400
ccagtgccca ccagccttgt cctaataaaa ttaagttgca tcattttgtc tgactaggtg 2460
tccttctata atattatggg gtggaggggg gtggtatgga gcaaggggca agttgggaag 2520
acaacctgta gggcctgcgg ggtctattgg gaaccaagct ggagtgcagt ggcacaatct 2580
tggctcactg caatctccgc ctcctgggtt caagcgattc tcctgcctca gcctcccgag 2640
ttgttgggat tccaggcatg catgaccagg ctcagctaat ttttgttttt ttggtagaga 2700
cggggtttca ccatattggc caggctggtc tccaactcct aatctcaggt gatctaccca 2760
ccttggcctc ccaaattgct gggattacag gcgtgaacca ctgctccctt ccctgtcctt 2820
acgcgtagaa ttggtaaaga gagtcgtgta aaatatcgag ttcgcacatc ttgttgtctg 2880
attattgatt tttggcgaaa ccatttgatc atatgacaag atgtgtatct accttaactt 2940
aatgattttg ataaaaatca ttaactagtc catggctgcc tcgcgcgttt cggtgatgac 3000
ggtgaaaacc tctgacacat gcagctcccg gagacggtca cagcttgtct gtaagcggat 3060
gccgggagca gacaagcccg tcagggcgcg tcagcgggtg ttggcgggtg tcggggcgca 3120
gccatgaccc agtcacgtag cgatagcgga gtgtatactg gcttaactat gcggcatcag 3180
agcagattgt actgagagtg caccatatgc ggtgtgaaat accgcacaga tgcgtaagga 3240
gaaaataccg catcaggcgc tcttccgctt cctcgctcac tgactcgctg cgctcggtcg 3300
ttcggctgcg gcgagcggta tcagctcact caaaggcggt aatacggtta tccacagaat 3360
caggggataa cgcaggaaag aacatgtgag caaaaggcca gcaaaaggcc aggaaccgta 3420
aaaaggccgc gttgctggcg tttttccata ggctccgccc ccctgacgag catcacaaaa 3480
atcgacgctc aagtcagagg tggcgaaacc cgacaggact ataaagatac caggcgtttc 3540
cccctggaag ctccctcgtg cgctctcctg ttccgaccct gccgcttacc ggatacctgt 3600
ccgcctttct cccttcggga agcgtggcgc tttctcatag ctcacgctgt aggtatctca 3660
gttcggtgta ggtcgttcgc tccaagctgg gctgtgtgca cgaacccccc gttcagcccg 3720
accgctgcgc cttatccggt aactatcgtc ttgagtccaa cccggtaaga cacgacttat 3780
cgccactggc agcagccact ggtaacagga ttagcagagc gaggtatgta ggcggtgcta 3840
cagagttctt gaagtggtgg cctaactacg gctacactag aagaacagta tttggtatct 3900
gcgctctgct gaagccagtt accttcggaa aaagagttgg tagctcttga tccggcaaac 3960
aaaccaccgc tggtagcggt ggtttttttg tttgcaagca gcagattacg cgcagaaaaa 4020
aaggatctca agaagatcct ttgatctttt ctacggggtc tgacgctcag tggaacgaaa 4080
actcacgtta agggattttg gtcatgagat tatcaaaaag gatcttcacc tagatccttt 4140
taaattaaaa atgaagtttt aaatcaatct aaagtatata tgagtaaact tggtctgaca 4200
gttaccaatg cttaatcagt gaggcaccta tctcagcgat ctgtctattt cgttcatcca 4260
tagttgcctg actcc 4275
<210> 2
<211> 565
<212> PRT
<213> Influenza virus
<400> 2
Met Lys Ala Ile Leu Val Val Leu Leu Tyr Thr Phe Ala Thr Ala Asn
1 5 10 15
Ala Asp Thr Leu Cys Ile Gly Tyr His Ala Asn Asn Ser Thr Asp Thr
20 25 30
Val Asp Thr Val Leu Glu Lys Asn Val Thr Val Thr His Ser Lys Asn
35 40 45
Ile Leu Glu Lys Thr His Asn Gly Lys Leu Cys Asp Leu Asp Gly Val
50 55 60
Lys Pro Leu Ile Leu Arg Asp Cys Ser Val Ala Gly Trp Leu Leu Gly
65 70 75 80
Asn Pro Met Cys Asp Glu Phe Ile Asn Val Pro Glu Trp Ser Tyr Ile
85 90 95
Val Glu Lys Ala Asn Pro Ala Tyr Gly Leu Cys Tyr Pro Gly Ser Phe
100 105 110
Asn Asp Tyr Glu Glu Leu Lys His Leu Leu Ser Ser Val Lys His Phe
115 120 125
Glu Arg Phe Glu Ile Phe Pro Lys Thr Ser Arg Trp Thr Gln His Thr
130 135 140
Thr Thr Gly Gly Ser Arg Ala Cys Ala Val Ser Gly Asn Pro Ser Phe
145 150 155 160
Phe Arg Asn Met Val Trp Leu Thr Lys Glu Gly Ser Ser Tyr Pro Lys
165 170 175
Leu Ser Lys Ser Tyr Ile Asn Thr Asn Gln Glu Asp Leu Leu Val Leu
180 185 190
Trp Gly Ile His His Pro Ile Asp Glu Thr Glu Gln Arg Thr Leu Tyr
195 200 205
Gln Asn Pro Thr Thr Tyr Ile Ser Val Gly Thr Ser Thr Leu Asn Gln
210 215 220
Arg Leu Val Pro Lys Ile Ala Thr Arg Pro Lys Val Asn Gly Gln Gly
225 230 235 240
Gly Arg Met Glu Phe Phe Trp Thr Ile Leu Lys Pro Asn Asp Ala Ile
245 250 255
Thr Phe Glu Ala Thr Gly Asn Leu Val Val Pro Arg Tyr Ala Phe Ala
260 265 270
Met Glu Arg Asn Ala Gly Ser Gly Ile Ile Ile Ser Asp Thr Glu Tyr
275 280 285
Gly Asn Cys Asn Thr Lys Cys Gln Thr Pro Ile Gly Ala Ile Asn Ser
290 295 300
Thr Leu Pro Phe His Asn Ile His Pro Leu Thr Ile Gly Glu Cys Pro
305 310 315 320
Lys Tyr Val Lys Ser Asn Arg Leu Val Leu Ala Thr Gly Leu Arg Asn
325 330 335
Val Pro Ser Ile Gln Ser Arg Gly Leu Phe Gly Ala Ile Ala Gly Phe
340 345 350
Ile Glu Gly Gly Trp Gln Gly Met Val Asp Gly Trp Tyr Gly Tyr His
355 360 365
His Ser Asn Glu Gln Gly Ser Gly Tyr Ala Ala Asp Lys Glu Ser Thr
370 375 380
Gln Lys Ala Phe Asp Gly Ile Thr Asn Lys Val Asn Ser Val Ile Glu
385 390 395 400
Lys Met Asn Thr Gln Phe Glu Ala Val Gly Lys Glu Phe Ser Asn Leu
405 410 415
Glu Arg Arg Leu Glu Asn Leu Asn Lys Lys Met Glu Asp Gly Phe Leu
420 425 430
Asp Val Trp Thr Tyr Asn Ala Glu Leu Leu Val Leu Met Glu Asn Glu
435 440 445
Arg Thr Leu Asp Phe His Asp Ser Asn Val Lys Asn Leu Tyr Asp Lys
450 455 460
Val Arg Met Gln Leu Arg Asp Asn Val Lys Glu Leu Gly Asn Gly Cys
465 470 475 480
Phe Glu Phe Tyr His Lys Cys Asp Asn Thr Cys Met Asn Ser Val Lys
485 490 495
Asn Gly Thr Tyr Asp Tyr Pro Lys Tyr Glu Glu Glu Ser Lys Leu Asn
500 505 510
Arg Asn Glu Ile Lys Gly Val Lys Leu Ser Ser Met Gly Val Tyr Gln
515 520 525
Ile Leu Ala Ile Tyr Ala Thr Val Ala Gly Ser Leu Ser Leu Ala Ile
530 535 540
Met Met Ala Gly Ile Ser Phe Trp Met Cys Ser Asn Gly Ser Leu Gln
545 550 555 560
Cys Arg Ile Cys Ile
565
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Антиген для универсальной рекомбинантной вакцины против вируса гриппа птиц и человека | 2021 |
|
RU2839428C2 |
| РЕКОМБИНАНТНАЯ ПСЕВДОАДЕНОВИРУСНАЯ ЧАСТИЦА НА ОСНОВЕ ГЕНОМА АДЕНОВИРУСА ЧЕЛОВЕКА 5 СЕРОТИПА, ПРОДУЦИРУЮЩАЯ ГЕМАГГЛЮТИНИН ВИРУСА ГРИППА ШТАММА A/Brisbane/59/2007(H1N1) И СПОСОБ ЕЕ ИСПОЛЬЗОВАНИЯ | 2012 |
|
RU2523599C1 |
| Экспрессионный вектор на основе аденовируса человека 5 серотипа, индуцирующий кросс-протективный иммунитет к вирусам гриппа А субтипа Н1, и фармацевтическая композиция на его основе | 2023 |
|
RU2802753C1 |
| ПРОТИВОВИРУСНОЕ ОДНОДОМЕННОЕ МИНИ-АНТИТЕЛО, НУКЛЕОТИДНАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ, ЭКСПРЕССИРУЮЩИЙ РЕКОМБИНАНТНЫЙ ВИРУСНЫЙ ВЕКТОР, ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ И СПОСОБ ПРОФИЛАКТИКИ ИЛИ ТЕРАПИИ ГРИППА ТИПА А | 2013 |
|
RU2536956C1 |
| ХИМЕРНЫЕ ВАКЦИНЫ ПРОТИВ ГРИППА | 2021 |
|
RU2834969C1 |
| РЕКОМБИНАНТНАЯ ПСЕВДОАДЕНОВИРУСНАЯ ЧАСТИЦА НА ОСНОВЕ ГЕНОМА АДЕНОВИРУСА ЧЕЛОВЕКА 5 СЕРОТИПА ДЛЯ ИНДУКЦИИ СПЕЦИФИЧЕСКОГО ИММУНИТЕТА К ВИРУСУ ГРИППА А СУБТИПА Н1N1 И СПОСОБ ЕЕ ИСПОЛЬЗОВАНИЯ В КАЧЕСТВЕ КОМПОНЕНТА ДЛЯ СОЗДАНИЯ ВАКЦИНЫ | 2012 |
|
RU2507257C1 |
| Экспрессионный вектор на основе аденовируса человека 5 серотипа индуцирующий кросс-протективный иммунитет к вирусам гриппа А субтипа H3 и фармацевтическая композиция на его основе. | 2023 |
|
RU2814189C1 |
| КОМПОЗИЦИИ ГЕМАГГЛЮТИНИНА ГРИППА С ГЕТЕРОЛОГИЧНЫМИ ЭПИТОПАМИ И/ИЛИ ИЗМЕНЕННЫМИ САЙТАМИ РАСЩЕПЛЕНИЯ ПРИ СОЗРЕВАНИИ | 2017 |
|
RU2769406C2 |
| Выделенный рекомбинантный вирус на основе вируса гриппа для индукции специфического иммунитета к вирусу гриппа и/или профилактики заболеваний, вызванных вирусом гриппа | 2021 |
|
RU2813150C2 |
| Однодоменное антитело для нейтрализации вирусов и его модификации, и способ их применения для экстренной профилактики заболеваний, вызываемых вирусом гриппа А | 2021 |
|
RU2777073C1 |


Изобретение относится к области биотехнологии, вирусологии, медицины. Описан штамм рекомбинантной псевдоаденовирусной частицы на основе генома аденовируса человека 5 серотипа Ad5-tetOFF-E3-НА125 для создания противогриппозных иммуногенных препаратов, несущей ген консенсусной последовательности гемагглютинина вируса гриппа А субтипов Н1, Н2, Н5, обогащенный В и Т-клеточными эпитопами вируса гриппа с нуклеотидной последовательностью SEQ ID NO: 2, при этом ген консенсусной последовательности гемагглютинина вируса гриппа А кодирует аминокислотную последовательность SEQ ID NO: 1. Так описан способ получения гена консенсусной последовательности гемагглютинина с нуклеотидной последовательностью SEQ ID NO: 2. Кроме того, описан иммуногенный препарат, содержащий штамм рекомбинантной псевдоаденовирусной частицы на основе генома аденовируса человека 5 серотипа по п. 1. для профилактики гриппа А, вызванного субтипами Н1, Н2 и Н5, содержащий на дозу: рекомбинантная псевдоаденовирусная частица Ad5-tetOFF-E3-НА125 - 107-109 акт.ед. и фармацевтически приемлемый буферный раствор - до 0,5 мл. Изобретение расширяет арсенал средств профилактики гриппа А. 3 н.п. ф-лы, 8 ил., 1 табл., 7 пр.
1. Штамм рекомбинантной псевдоаденовирусной частицы на основе генома аденовируса человека 5 серотипа Ad5-tetOFF-E3-НА125 для создания противогриппозных иммуногенных препаратов, несущей ген консенсусной последовательности гемагглютинина вируса гриппа А субтипов Н1, Н2, Н5, обогащенный В и Т-клеточными эпитопами вируса гриппа с нуклеотидной последовательностью SEQ ID NO: 2, при этом ген консенсусной последовательности гемагглютинина вируса гриппа А кодирует аминокислотную последовательность SEQ ID NO: 1.
2. Способ получения гена консенсусной последовательности гемагглютинина с нуклеотидной последовательностью SEQ ID NO: 2, осуществляемый за счет слияния консервативных участков гемагглютининов вируса гриппа А субтипов Н1, Н2 и Н5, при этом общая длина рекомбинантного белка равна нативному гемагглютинину вируса гриппа А - 1701 п. о., содержит Т-клеточные эпитопы и конформационный В-клеточный эпитоп в стебельной части, заключающийся в следующих последовательных действиях:
- из общедоступной базы генетических данных отбирают аминокислотные последовательности гемагглютининов вирусов гриппа А трех субтипов Н1, Н2 и Н5, выделенные от человека, выравнивают друг относительно друга с помощью общедоступной компьютерной программы, при этом сохраняя конформационный В-клеточный эпитоп в стебельной части, и, таким образом, получают «консенсусные» (усредненные) последовательности для каждого субтипа отдельно;
- с помощью общедоступной базы данных по распространенности аллельных частот выявляют наиболее часто встречающиеся аллели молекул HLA I-класса и HLA II-класса у жителей РФ и оставляют эпитопы с высокой афинностью, сокращая одинаковые последовательности эпитопов, которые распознаются разными аллелями;
- с помощью доступных онлайн сервисов определяют наличие эпитопов в консенсусных последовательностях Н1, Н2 и Н5 для отобранных ранее аллелей HLA I-класса и HLA II-класса.
- накладывают последовательности выявленных эпитопов аллелей HLA I-класса и HLA II-класса на каждую из трех консенсусных последовательностей вируса гриппа субтипов Н1, Н2 и Н5 и определяют наличие соответствующих эпитопов и отмечают их местоположение;
- с помощью общедоступной базы данных иммунологических эпитопов из отобранных ранее Т-клеточных эпитопов HLA отбираются все известные эпитопы с экспериментально подтвержденной активностью и также на каждую из трех консенсусных последовательностей вируса гриппа субтипов Н1, Н2 и Н5 и определяют наличие соответствующих эпитопов и отмечают их местоположение;
- три консенсусные последовательности гемагглютининов вирусов гриппа А трех субтипов Н1, Н2 и Н5 выравнивают относительно друг друга;
- собирают консенсусную последовательность гемагглютинина вируса гриппа А путем слияния того участка гена, на котором отмечено наибольшее количество Т-клеточных эпитопов, выбранного каждый раз из одной из трех консенсусных последовательностей гемагглютининов вирусов гриппа А субтипов Н1, Н2 и Н5, и последовательно включаемого в собираемую консервативную последовательность;
- полученную консервативную аминокислотную последовательность гемагглютинина вируса гриппа А, содержащую участки гемагглютининов вирусов гриппа субтипов Н1, Н2 и Н5 и максимально насыщенную консервативными В- и Т-клеточными эпитопами, переводят в соответствующую ей нуклеотидную последовательность, в которой оптимизируют кодоны общеизвестным способом для более эффективной экспрессии в клетках человека.
3. Иммуногенный препарат, содержащий штамм рекомбинантной псевдоаденовирусной частицы на основе генома аденовируса человека 5 серотипа по п. 1. для профилактики гриппа А, вызванного субтипами Н1, Н2 и Н5, содержащий на дозу:
- рекомбинантная псевдоаденовирусная частица Ad5-tetOFF-E3-НА125- 107-109 акт.ед.;
- фармацевтически приемлемый буферный раствор - до 0,5 мл.
| WO 2010058236 A1, 27.05.2010 | |||
| РЕКОМБИНАНТНАЯ ПСЕВДОАДЕНОВИРУСНАЯ ЧАСТИЦА НА ОСНОВЕ ГЕНОМА АДЕНОВИРУСА ЧЕЛОВЕКА 5 СЕРОТИПА ДЛЯ ИНДУКЦИИ СПЕЦИФИЧЕСКОГО ИММУНИТЕТА К ВИРУСУ ГРИППА А СУБТИПА Н1N1 И СПОСОБ ЕЕ ИСПОЛЬЗОВАНИЯ В КАЧЕСТВЕ КОМПОНЕНТА ДЛЯ СОЗДАНИЯ ВАКЦИНЫ | 2012 |
|
RU2507257C1 |
| РЕКОМБИНАНТНАЯ ПСЕВДОАДЕНОВИРУСНАЯ ЧАСТИЦА НА ОСНОВЕ ГЕНОМА АДЕНОВИРУСА ЧЕЛОВЕКА 5 СЕРОТИПА, ПРОДУЦИРУЮЩАЯ ГЕМАГГЛЮТИНИН ВИРУСА ГРИППА ШТАММА A/Brisbane/59/2007(H1N1) И СПОСОБ ЕЕ ИСПОЛЬЗОВАНИЯ | 2012 |
|
RU2523599C1 |
| WO 2013129961 A1, 06.09.2013. | |||

