Перекрестная ссылка на родственные заявки
Данная заявка заявляет приоритет предварительной заявки на патент США 61/367809, поданной 26 июля 2010 г., содержание которой включено в виде ссылки в настоящую заявку.
Область техники, к которой относится изобретение
Данное изобретение относится к трансгенным позвоночным животным и конкретнее к трансгенным позвоночным животным для разработки лекарственных препаратов для человека.
Уровень техники
В последующем обсуждении будут описаны некоторые статьи и способы для представления предшествующего уровня техники и введения. Ничто, содержащееся в данном документе, не следует истолковывать в качестве «допущения» предшествующего уровня техники. Заявитель точно сохраняет свое право продемонстрировать, где это является подходящим, что цитированные статьи и способы не составляют предшествующий уровень техники согласно применимым законодательным положениям.
Гены, кодирующие иммуноглобулины человека и мыши, хорошо охарактеризованы. Berman et al., 1988, EMBO J., 7:727-738 описывают локус VH IgG человека, содержащий генные сегменты VH.D и JH. Sakano et al., 1981, Nature, 280:562-565 описывают дополнительный сегмент генов тяжелых цепей иммуноглобулинов. Blankenstein and Kruwinkel, 1987, Eur. J. Immunol., 17:1351-1357 описывают мышиную вариабельную область тяжелой цепи. Получение трансгенных животных, таких как мыши, имеющих различные иммуноглобулиновые локусы, позволило использовать таких трансгенных животных в научных и опытно-практических применениях, например, для разработки лекарственных препаратов и в фундаментальных исследованиях в различных биологических системах. Получение трансгенных мышей, несущих гены иммуноглобулинов человека, описано в международных заявках WO 90/10077 и WO 90/04036. В WO 90/04036 описана трансгенная мышь с интегрированным «мини» локусом иммуноглобулина человека. В WO 90/10077 описан вектор, содержащий доминантную регуляторную область иммуноглобулина для применения в получении трансгенных животных.
Разработаны многочисленные способы для замещения эндогенных областей мышиных иммуноглобулинов последовательностями человеческих иммуноглобулинов для получения частично или полностью человеческих антител в целях разработки лекарственных препаратов. Примеры таких мышей включают животных, описанных, например, в патентах США № 7145056; 7064244; 7041871; 6673986; 6596541; 6570061; 6162963; 6130364; 6091001; 6023010; 5593598; 5877397; 5874299; 5814318; 5789650; 5661016; 5612205 и 5591669. Во многих случаях у мышей с полностью гуманизированными иммуноглобулинами продукция антител находится ниже обычных уровней за счет менее эффективной V(D)J-рекомбинации и ограниченной продукции антител в результате частичного добавления гена. В других случаях получение мышиной кодирующей последовательности, которая «обменена» на человеческие последовательности, является длительным и дорогостоящим процессом за счет подхода замещения отдельных мышиных экзонов на синтетические человеческие аналоги.
Основываясь на приведенном выше анализе, становится очевидным, что существует необходимость в эффективных и экономичных способах эффективного получения человеческих антител. Конкретнее, в данной области имеется необходимость в позвоночных животных, отличных от человека, содержащих области иммуноглобулинов человека, и в трансгенных животных, обладающих способностью правильно отвечать на антиген.
В соответствие с вышеизложенной целью, изобретение относится к трансгенным животным, отличным от человека, которые способны продуцировать антитело с V-областями человека.
Сущность изобретения
Раздел «Сущность изобретения» приводится для обоснования выбора концепций в упрощенной форме, которые ниже подробно описаны в разделе «Подробное описание изобретения». Данный раздел «Сущность изобретения» не предназначен для указания ключевых или основных признаков заявленной сущности изобретения или для ограничения объема заявленной сущности изобретения. Другие признаки, детали, применения и преимущества заявленной сущности изобретения станут понятными из последующего раздела «Подробное описание изобретения», включая аспекты, показанные на прилагающихся фигурах и определенные в прилагаемой формуле изобретения.
Настоящее изобретение относится к клеткам позвоночных животных, отличных от человека, и позвоночным животным, отличным от человека, имеющим геном, содержащий введенную частично человеческую область иммуноглобулинов, где указанная введенная область содержит кодирующие последовательности локуса вариабельной области иммуноглобулина человека и некодирующие последовательности, основанные на эндогенном локусе вариабельной области иммуноглобулина позвоночного животного, отличного от человека. Предпочтительно трансгенные клетки и животные по изобретению имеют геномы, в которых частично или полностью удалена эндогенная иммуноглобулиновая область.
Для продукции человеческих моноклональных антител у позвоночных животных, отличных от человека, требуется как минимум, чтобы хозяин имел, по меньшей мере, один локус, который будет экспрессировать белки тяжелой цепи иммуноглобулина человека, и один локус, который будет экспрессировать белки легкой цепи иммуноглобулина человека.
В некоторых аспектах частично человеческий локус вариабельной области иммуноглобулина содержит человеческие кодирующие последовательности VH и некодирующие последовательности VH, основанные на эндогенной области VH позвоночного животного, отличного от человека. В данных аспектах частично человеческий локус вариабельной области иммуноглобулина дополнительно содержит кодирующие последовательности D- и J-генов человека и некодирующие последовательности D- и J-генов, основанные на эндогенном геноме позвоночного животного-хозяина, отличного от человека.
В других аспектах иммуноглобулиновая область содержит введенную область, включающую человеческие кодирующие последовательности VL и некодирующие последовательности VL, основанные на эндогенной области VL позвоночного животного, отличного от человека. Более предпочтительно введенная частично человеческая область иммуноглобулина, содержащая человеческие кодирующие последовательности VL, дополнительно содержит кодирующие последовательности J-гена человека и некодирующие последовательности J-гена, основанные на эндогенном геноме позвоночного животного-хозяина, отличного от человека.
В некоторых аспектах позвоночное животное представляет собой млекопитающее, и предпочтительно млекопитающее является грызуном, например мышью или крысой. В других аспектах позвоночное животное является птицей, например цыпленком.
В одном конкретном аспекте изобретение относится к способу получения клетки позвоночного животного, отличного от человека, содержащей частично человеческую область иммуноглобулина, где указанный способ включает: а) введение двух или более нацеленных на рекомбиназу сайтов в клетку позвоночного животного, отличного от человека, и интеграцию, по меньшей мере, одного сайта в клеточный геном выше и, по меньшей мере, одного сайта ниже геномной области, содержащей эндогенный локус вариабельной области иммуноглобулина; и b) введение частично человеческого локуса вариабельной области иммуноглобулина, содержащего человеческие кодирующие последовательности вариабельной области иммуноглобулина и некодирующие последовательности, основанные на эндогенной вариабельной области иммуноглобулина позвоночного животного-хозяина, отличного от человека, в клетку позвоночного животного-хозяина, отличного от человека посредством обмена, опосредованного рекомбиназой.
В конкретном аспекте данного способа введенная частично человеческая область иммуноглобулина содержит человеческие кодирующие области гена VH и дополнительно содержит i) кодирующие последовательности D- и J-генов человека и ii) некодирующие последовательности D- и J-генов, и пре-DJ-области, основанные на эндогенном геноме позвоночного животного-хозяина, отличного от человека. Частично человеческие иммуноглобулиновые области предпочтительно вводят в клетку-хозяин с использованием нацеленных на рекомбиназу сайтов, которые вводятся выше эндогенных генов VH иммуноглобулина и ниже эндогенной области D- и J-генов.
В других аспектах кодирующие области гена VH происходят (по меньшей мере, частично) из других источников - например, они могут представлять рационально или иначе сконструированные последовательности, последовательности, которые представляют собой комбинацию человеческой и другой сконструированной последовательностей, или последовательности от других видов, таких как приматы, отличные от человека.
В еще одном конкретном аспекте введенная частично человеческая область иммуноглобулина содержит человеческие кодирующие области гена VL и дополнительно содержит i) кодирующие последовательности J-гена человека и ii) некодирующие последовательности J-гена, основанные на эндогенном геноме позвоночного животного-хозяина, отличного от человека. Частично человеческие иммуноглобулиновые области предпочтительно вводят в клетку-хозяин с использованием нацеленных на рекомбиназу сайтов, которые вводятся выше эндогенных генов VL иммуноглобулина и ниже эндогенной J-области.
Предпочтительно частично человеческая область иммуноглобулина синтезируется в виде одной нуклеиновой кислоты и вводится в клетку позвоночного животного-хозяина, отличного от человека, в виде одной нуклеиновокислотной области. Частично человеческую область иммуноглобулина также можно синтезировать в виде двух или более смежных сегментов и вводить в клетку позвоночного животного-хозяина в этих двух отдельных сегментах. Частично человеческую нуклеиновую кислоту также можно получить с использованием рекомбинантных способов и выделить до введения нуклеиновой кислоты в клетку позвоночного животного-хозяина, отличного от человека.
В еще одном предпочтительном аспекте способ дополнительно обеспечивает делецию геномной области, фланкированной двумя введенными сайтами рекомбиназы, до проведения стадии b).
В еще одном аспекте изобретение относится к способам получения клетки позвоночного животного, отличного от человека, содержащей частично человеческую область иммуноглобулина, где указанный способ включает: а) введение двух или более сайтов сайт-специфической рекомбинации, которые не способны рекомбинировать друг с другом, в геном клетки позвоночного животного-хозяина, отличного от человека, где, по меньшей мере, один сайт рекомбинации вводится выше эндогенного локуса вариабельной области иммуноглобулина и, по меньшей мере, один сайт рекомбинации вводится ниже эндогенного локуса вариабельной области иммуноглобулина; b) обеспечение вектора, содержащего частично человеческую область иммуноглобулина, имеющую i) человеческие кодирующие последовательности вариабельной области иммуноглобулина и ii) некодирующие последовательности, основанные на эндогенной вариабельной области иммуноглобулина клетки-хозяина, где частично человеческая область фланкирована теми же двумя сайтами сайт-специфической рекомбинации, которые фланкируют эндогенную вариабельную область иммуноглобулина клетки-хозяина со стадии а); с) введение вектора со стадии b) и сайт-специфической рекомбиназы, способной узнавать два сайта рекомбиназы, в клетку; d) обеспечение того, что происходит рекомбинантное событие между геномом клетки со стадии а) и частично человеческой областью иммуноглобулина, приводящее к замене эндогенного локуса вариабельной области иммуноглобулина частично человеческим локусом области иммуноглобулина. В конкретном аспекте данного способа частично человеческая область иммуноглобулина содержит кодирующие последовательности гена VH иммуноглобулина и дополнительно включает i) кодирующие последовательности D- и J-генов человека и ii) некодирующие последовательности D- и J-генов и пре-DJ-области, основанные на эндогенном геноме позвоночного животного-хозяина, отличного от человека. Нацеленные на рекомбиназу сайты вводятся выше эндогенных генов VH и ниже эндогенных последовательностей D- и J-генов.
В еще одном конкретном аспекте данного способа, способ дополнительно обеспечивает делецию геномной области, фланкированной двумя веденными сайтами рекомбиназы до проведения стадии с).
Изобретение относится к еще одному способу получения клетки трансгенного позвоночного животного, отличного от человека, где указанный способ включает: а) обеспечение клетки позвоночного животного, отличного от человека, имеющей геном, который содержит две группы сайтов сайт-специфической рекомбинации, которые не способны рекомбинировать друг с другом, и которые фланкируют участок эндогенной иммуноглобулиновой области генома хозяина; b) делецию участка эндогенного локуса вариабельной области иммуноглобулина генома введением рекомбиназы, которая узнает первую группу сайтов сайт-специфической рекомбинации, где такая делеция в геноме сохраняет вторую группу сайтов сайт-специфической рекомбинации; с) обеспечение вектора, содержащего частично человеческий локус вариабельной области иммуноглобулина человека, включающий человеческие кодирующие последовательности и некодирующие последовательности, основанные на эндогенной вариабельной области иммуноглобулина, фланкированной второй группой сайтов сайт-специфической рекомбинации; d) введение вектора со стадии с) и сайт-специфической рекомбиназы, способной узнавать вторую группу сайтов рекомбиназы, в клетку; и е) обеспечение того, что происходит рекомбинантное событие между геномом клетки и частично человеческим локусом вариабельной области иммуноглобулина, приводящее к замене эндогенного локуса вариабельной области иммуноглобулина частично человеческим локусом вариабельной области иммуноглобулина.
Предпочтительно клетка млекопитающего, отличного от человека, для применения в каждом из указанных выше способов представляет клетку млекопитающего, и более предпочтительно эмбриональную стволовую клетку млекопитающего (ES-клетку). В других аспектах клетка может представлять клетку птицы и предпочтительно птичью примордиальную зародышевую клетку.
После того как клетки подверглись замене эндогенного локуса вариабельной области иммуноглобулина, отбирают клетки, содержащие введенную частично человеческую вариабельную область иммуноглобулина, и предпочтительно их выделяют. В предпочтительном аспекте изобретения клетки являются эмбриональными стволовыми клетками млекопитающего (ES-клетками), отличного от человека, и затем выделенную ES-клетку используют для получения трансгенного млекопитающего, отличного от человека, экспрессирующего частично человеческий локус вариабельной области иммуноглобулина. В других аспектах клетки являются примордиальными зародышевыми клетками, и затем выделенную зародышевую клетку используют для получения трансгенной птицы, экспрессирующей частично человеческую вариабельную область иммуноглобулина.
В конкретном аспекте изобретение относится к способу получения клетки млекопитающего, отличного от человека, содержащей частично человеческую область иммуноглобулина, где способ включает: а) обеспечение эмбриональной стволовой клетки (ES-клетки) млекопитающего, имеющей геном, который содержит два сайта сайт-специфической рекомбинации, которые не способны рекомбинировать друг с другом и которые фланкируют участок иммуноглобулиновой области; b) обеспечение вектора, содержащего частично человеческую область иммуноглобулина, включающую человеческие кодирующие последовательности вариабельной области иммуноглобулина и некодирующие последовательности, основанные на эндогенной вариабельной области иммуноглобулина, где указанная частично человеческая область фланкирована теми же двумя сайтами сайт-специфической рекомбинации, которые фланкируют участок иммуноглобулиновой области в ES-клетке; с) приведение в контакт ES-клетки и указанного вектора с сайт-специфической рекомбиназой, способной узнавать два сайта рекомбиназы в соответствующих условиях для обеспечения рекомбинантного события, приводящего к замене эндогенного участка иммуноглобулиновой области частично человеческой областью иммуноглобулина в ES-клетке.
В еще одном аспекте изобретение относится к способу получения трансгенного млекопитающего, отличного от человека, содержащего частично человеческую область иммуноглобулина, где указанный способ включает: а) введение одного или более сайтов сайт-специфической рекомбинации, которые не способны рекомбинировать другом с другом, в геном клетки позвоночного животного, отличного от человека; b) обеспечение вектора, содержащего частично человеческую область иммуноглобулина, имеющую i) человеческие кодирующие последовательности вариабельной области, и ii) некодирующие последовательности, основанные на эндогенной вариабельной области, где частично человеческая область фланкирована теми же сайтами сайт-специфической рекомбинации, как и введенные в геном клетки-хозяина со стадии а); с) введение вектора со стадии b) и сайт-специфической рекомбиназы, способной узнавать одну группу сайтов рекомбиназы, в клетку; d) обеспечение того, что происходит рекомбинантное событие между геномом клетки со стадии а) и частично человеческой областью иммуноглобулина, приводящее к замене эндогенной вариабельной области иммуноглобулина частично человеческой областью иммуноглобулина; е) отбор клетки, которая содержит частично человеческую область иммуноглобулина; и f) применение клетки для получения трансгенного животного, содержащего частично человеческую область иммуноглобулина.
В конкретном аспекте частично человеческая область иммуноглобулина содержит человеческие кодирующие области VH, кодирующие последовательности D- и J-генов человека и некодирующие последовательности D- и J-генов, и пре-DJ-области, основанные на эндогенном геноме позвоночного животного-хозяина, отличного от человека. Затем вводятся сайты сайт-специфической рекомбинации выше эндогенных генов VH иммуноглобулина и ниже эндогенных областей D- и J-генов.
Изобретение относится к еще одному способу получения трансгенного животного, отличного от человека, содержащего частично человеческую область иммуноглобулина, где указанный способ включает: а) обеспечение клетки позвоночного животного, отличного от человека, имеющей геном, который содержит две группы сайтов сайт-специфической рекомбинации, которые не способны рекомбинировать друг с другом, и которые фланкируют участок эндогенного локуса вариабельной области иммуноглобулина генома хозяина; b) делецию участка эндогенной иммуноглобулиновой области генома хозяина введением рекомбиназы, которая узнает первую группу сайтов сайт-специфической рекомбинации, где такая делеция в геноме сохраняет вторую группу сайтов сайт-специфической рекомбинации; с) обеспечение вектора, содержащего частично человеческий локус вариабельной области иммуноглобулина, включающий человеческие кодирующие последовательности и некодирующие последовательности, основанные на эндогенном локусе вариабельной области иммуноглобулина, фланкированным второй группой сайтов сайт-специфической рекомбинации; d) введение вектора со стадии с) и сайт-специфической рекомбиназы, способной узнавать вторую группу сайтов сайт-специфической рекомбинации, в клетку и е) обеспечение того, что происходит рекомбинантное событие между геномом клетки и частично человеческой вариабельной областью иммуноглобулина, приводящее к замене эндогенной иммуноглобулиновой области частично человеческой вариабельной областью иммуноглобулина; f) отбор клетки, которая содержит частично человеческую вариабельную область иммуноглобулина; и g) применение клетки для получения трансгенного животного, содержащего частично человеческую вариабельную область иммуноглобулина.
Изобретение относится к еще одному способу получения трансгенного млекопитающего, отличного от человека, содержащего частично человеческую область иммуноглобулина, где указанный способ включает: а) обеспечение эмбриональной стволовой клетки (ES-клетки) млекопитающего, отличного от человека, имеющей геном, который содержит два сайта сайт-специфической рекомбинации, которые не способны рекомбинировать друг с другом, и которые фланкируют участок иммуноглобулиновой области; b) обеспечение вектора, содержащего частично человеческую область иммуноглобулина, включающую человеческие кодирующие последовательности вариабельной области и некодирующие последовательности, основанные на эндогенном гене вариабельной области, где частично человеческая область фланкирована теми же двумя сайтами сайт-специфической рекомбинации, которые фланкируют участок иммуноглобулиновой области в ES-клетке; с) приведение в контакт указанной ES-клетки и указанного вектора с сайт-специфической рекомбиназой, способной узнавать два сайта рекомбиназы в соответствующих условиях для обеспечения рекомбинантного события, приводящего к замене эндогенного участка иммуноглобулиновой области частично человеческой областью иммуноглобулина в ES-клетке; d) отбор ES-клетки, которая содержит замещенный участок нуклеиновой кислоты и применение указанной эмбриональной стволовой клетки; и е) применение клетки для получения трансгенного животного, содержащего частично человеческий локус вариабельной области иммуноглобулина для получения гетерозиготного «частично человеческого» животного.
В конкретном аспекте изобретения трансгенные позвоночные животные, отличные от человека, представляют собой млекопитающих, и предпочтительно млекопитающие являются грызунами, например мышью или крысой. В еще одних аспектах трансгенные позвоночные животные, отличные от человека, представляют птицу, например цыпленка.
Целью изобретения является обеспечение клеток позвоночного животного, отличного от человека, и трансгенных млекопитающих, отличных от человека, экспрессирующих введенный локус вариабельной области иммуноглобулина, содержащий человеческие кодирующие последовательности вариабельной области иммуноглобулина и некодирующие последовательности, основанные на эндогенном геноме хозяина.
Кроме того, целью является получение В-клеток от трансгенных животных, которые способны экспрессировать частично человеческие антитела, содержащие последовательности человеческой VH, где такие клетки иммортализованы для обеспечения источника моноклонального антитела, специфического для конкретного антигена.
Еще одной целью является обеспечение человеческих вариабельных областей, клонированных из В-клеток, для применения в продукции и/или оптимизации антител для диагностических и терапевтических применений.
Еще одной целью изобретения является обеспечение гибридомных клеток, которые способны продуцировать частично человеческие моноклональные антитела, содержащие человеческие последовательности вариабельной области.
Данные и другие аспекты, цели и признаки изобретения более подробно описаны ниже.
Краткое описание фигур
На фигуре 1 показана схема проведения примерного способа из предпочтительного варианта осуществления изобретения.
На фигуре 2 приведена схематичная диаграмма, показывающая введение первой группы сайтов сайт-специфической рекомбинации в геном клетки млекопитающего, отличного от человека, посредством нацеленного на гомологичные участки вектора.
На фигуре 3 приведена другая схематичная диаграмма, показывающая введение первой группы сайтов сайт-специфической рекомбинации в геном клетки млекопитающего, отличного от человека, посредством нацеленного на гомологичные участки вектора.
На фигуре 4 приведена схематичная диаграмма, показывающая введение второй группы сайтов сайт-специфической рекомбинации в геном клетки млекопитающего, отличного от человека, посредством нацеленного на гомологичные участки вектора.
На фигуре 5 приведена схематичная диаграмма, показывающая делецию эндогенной иммуноглобулиновой области клетки-хозяина.
На фигуре 6 приведена схематичная диаграмма, показывающая введение частично человеческой области иммуноглобулина посредством сайт-специфического нацеленного на гомологичные участки вектора.
На фигуре 7 приведена схематичная диаграмма, показывающая введение частично человеческой области иммуноглобулина, содержащей дополнительные мышиные последовательности с использованием сайт-специфического нацеленного вектора.
На фигуре 8 приведена схематичная диаграмма, показывающая введение частично человеческой области иммуноглобулина, содержащей дополнительные мышиные последовательности, в мышиную область тяжелой цепи.
На фигуре 9 приведена схематичная диаграмма, показывающая введение частично человеческой области иммуноглобулина, содержащей дополнительные мышиные последовательности, в мышиную область каппа-цепи.
На фигуре 10 приведена схематичная диаграмма, показывающая введение частично человеческой области иммуноглобулина, содержащей дополнительные мышиные последовательности, в мышиную область лямбда-цепи.
На фигуре 11 приведена схематичная диаграмма, показывающая введение частично человеческой области иммуноглобулина, содержащей миниген человеческой VH посредством сайт-специфического нацеленного вектора.
Определения
Термины, использованные в данном документе, предназначены иметь ясные и общепринятые значения, которые понимаются специалистами в данной области. Следующие определения приводятся для оказания помощи читателю в понимании настоящего изобретения, но не предназначаются для варьирования или ограничения иначе значений таких терминов, если не указано иначе.
В том смысле, в котором в данном документе используется термин «частично человеческая», он относится к нуклеиновой кислоте, содержащей одновременно последовательности человека и млекопитающего или животного, отличного от человека, включая нуклеиновую кислоту, содержащую одновременно последовательности человека и млекопитающего, отличного от человека. В контексте термина «частично человеческие последовательности» по изобретению частично человеческие нуклеиновые кислоты содержат последовательности кодирующих областей человеческого иммуноглобулина и некодирующие последовательности, основанные на эндогенной иммуноглобулиновой области млекопитающего, отличного от человека. Термин «основанные на», использованный по отношению к эндогенным некодирующим последовательностям млекопитающего, отличного от человека, относится к последовательностям, которые соответствуют некодирующей последовательности и имеют относительно высокую степень гомологии с некодирующими последовательностями эндогенных локусов млекопитающего-хозяина, например млекопитающего, от которого получена ES-клетка. Предпочтительно некодирующие последовательности имеют, по меньшей мере, 80%, более предпочтительно 90% гомологию с соответствующими некодирующими последовательностями, найденными в эндогенных локусах клетки позвоночного животного-хозяина, отличного от человека, в которую вводится частично человеческая молекула, содержащая некодирующие последовательности.
В том смысле, в котором в данном документе используется термин «нацеленный на гомологичные участки вектор», он относится к вектору, содержащему нуклеиновую кислоту, кодирующую нацеленную последовательность, сайт сайт-специфической рекомбинации, и необязательно селективный маркерный ген, которые используются для модификации иммуноглобулиновой области опосредованной гомологией рекомбинацией в клетке-хозяине. Например, нацеленный на гомологичные участки вектор можно использовать в настоящем изобретении для введения сайта сайт-специфической рекомбинации в определенную область генома клетки-хозяина.
В том смысле, в котором в данном документе используется термин «вариабельная область иммуноглобулина», он относится к нуклеотидной последовательности, которая кодирует всю или участок вариабельной области молекулы антитела, или ко всей или фрагменту регуляторной нуклеотидной последовательности, которая регулирует экспрессию молекулы антитела. Иммуноглобулиновые области для тяжелых цепей могут включать, не ограничиваясь этим, все или часть V-, D-, J- и свич-областей, включая интроны. Иммуноглобулиновая область для легких цепей может включать, не ограничиваясь этим, V- и J-области, их выше фланкирующие последовательности, интроны, ассоциированные с или смежными с геном константной области легкой цепи.
В том смысле, в котором в данном документе используется термин «сайт-специфическая рекомбинация», он относится к процессу рекомбинации между двумя совместимыми сайтами рекомбинации, включающему следующие три события: а) делецию заранее выбранной нуклеиновой кислоты, фланкированной сайтами рекомбинации; b) инверсию нуклеотидной последовательности заранее определенной нуклеиновой кислоты, фланкированной сайтами рекомбинации, и с) двухсторонний обмен нуклеиновокислотных областей, приближающихся к сайтам рекомбинации, расположенных в различных нуклеиновокислотных молекулах. Следует понимать, что такой двухсторонний обмен сегментами нуклеиновой кислоты приводит к интеграционному событию, если одна или обе нуклеиновокислотные молекулы являются кольцевыми.
В том смысле, в котором в данном документе используется термин «нацеленная последовательность», он относится к последовательности, гомологичной ДНК-последовательностям в геноме клетки, которые фланкируют или являются смежными к участкам иммуноглобулиновой области, предназначенной для модификации. Фланкирующая или смежная последовательность может находиться внутри самого локуса или выше или ниже кодирующих последовательностей в геноме клетки-хозяина. Нацеленные последовательности вставляются в рекомбинантные ДНК-векторы, которые используются для трансфекции таким образом, чтобы последовательности, предназначенные для вставки в геном клетки, такие как последовательность сайта рекомбинации, фланкировались нацеленными последовательностями вектора.
В том смысле, в котором в данном документе используется термин «сайт-специфический нацеленный вектор», он относится к вектору, содержащему нуклеиновую кислоту, кодирующую сайт сайт-специфической рекомбинации, частично человеческую нуклеиновую кислоту и необязательно селектируемый маркерный ген, которые используют для модификации эндогенной области иммуноглобулина у хозяина с использованием опосредованной рекомбиназой сайт-специфической рекомбинации. Сайт рекомбинации нацеленного вектора подходит для сайт-специфической рекомбинации с другим соответствующим сайтом рекомбинации, который был вставлен в геномную последовательность клетки-хозяина (например, посредством нацеленного на гомологичные участки вектора), смежную к иммуноглобулиновой области, которая предназначена для модификации. Интеграция частично человеческой последовательности в сайт рекомбинации в иммуноглобулиновой области приводит к замене эндогенной области введенной частично человеческой областью.
В том смысле, в котором в данном документе используется термин «трансген», он используется для описания генетического материала, который был вставлен или предназначен для искусственной вставки в геном клетки и, в частности, клетки позвоночного животного-хозяина. В том смысле, в котором в данном документе используется термин «трансген», он относится к частично человеческой нуклеиновой кислоте, например частично человеческой нуклеиновой кислоте в форме экспрессионной конструкции и/или нацеленного вектора.
Под выражением «трансгенное животное» понимается животное, отличное от человека, обычно млекопитающее, содержащее экзогенную нуклеиновокислотную последовательность, находящуюся в виде экстрахромосомного элемента в части его клеток, или она стабильно интегрирована в ДНК его зародышевой линии (т.е. геномной последовательности большинства или всех его клеток). В настоящем изобретении частично человеческую нуклеиновую кислоту вводят в зародышевую линию таких трансгенных животных генетической манипуляцией, например, с эмбрионами или эмбриональными стволовыми клетками животного-хозяина согласно способам, хорошо известным в данной области.
Термин «вектор» включает плазмиды и вирусы, и любую молекулу ДНК или РНК, независимо от того, являются они самореплицирующимися или нет, которые можно использовать для трансформации и трансфекции клетки.
Подробное описание изобретения
В практике способов, описанных в настоящем документе, можно использовать, если не указано иначе, обычные методы и протоколы органической химии, полимерных технологий, молекулярной биологии (включая рекомбинантные методы), клеточной биологии, биохимии и технологии секвенирования, которые известны специалистам в данной области. Такие обычные способы включают синтез полимерных микрочипов, гибридизацию и лигирование полинуклеотидов, и детектирование гибридизации с использованием метки. Конкретные иллюстрации подходящих методов можно найти при обращении к примерам, представленным в данном документе. Однако, конечно, также можно использовать другие равноценные обычные методы. Такие широко известные методы и протоколы можно найти в обычных лабораторных руководствах, таких как Green et al., Eds., 1999, Genome Analysis: A Laboratory Manual Series (Vols. I-IV); Weiner, Gabriel, Stephens, Eds., 2007, Genetic Variation: A Laboratory Manual; Dieffenbach, Dveksler, Eds., 2003, PCR Primer: A Laboratory Manual; Bowtell and Sambrook, 2003, DNA Microarrays: A Molecular Cloning Manual; Mount, 2004, Bioinformatics: Sequence and Genome Analysis; Sambrook and Russell, 2006, Condensed Protocols from Molecular Cloning: A Laboratory Manual; и Sambrook and Russell, 2002, Molecular Cloning: A Laboratory Manual (все изданы Cold Spring Harbor Laboratory Press); Stryer L., 1995, Biochemistry (4th Ed.) W.H.Freeman, New York N.Y.; Gait «Oligonucleotide Synthesis: A Practical Approach», 1984, IRL Press, London; Nelson and Cox, 2000, Lehninger, Principles of Biochemistry 3rd Ed., W.H. Freeman Pub., New York N.Y.; и Berg et al., 2002, Biochemistry (5th Ed.) W.H.Freeman, New York N.Y.; все источники в полном объеме включены в данный документ для сведении для всех целей.
Следует обратить внимание, что в данном документе и в прилагаемой формуле изобретения, формы единственного числа «a», «an» и «the» включают множественные формы, если по контексту четко не указано иначе. Таким образом, например, обращение к «локусу» относится к одному или более локусам, и обращение к «способу» включает обращение к эквивалентным стадиям и способам, известным специалистам в данной области и т.д.
Если не указано иначе, то все технические и научные термины, использованные в данном документе, имеют то же значение, которое обычно понимается специалистами в области, к которой относится изобретение. Все публикации, упомянутые в данном документе, включены в данный документ для сведения в целях описания и раскрытия устройств, формуляций и методологий, которые можно использовать по отношению к описанному изобретению.
В тех случаях, когда приводятся пределы значений, то понятно, что каждое промежуточное значение, находящееся между верхней и нижней границей такого предела и любое установленное или промежуточное значение в указанных пределах, включается в изобретение. Верхняя и нижняя границы данных более узких пределов могут независимо включаться в более узкие пределы, и они также включаются в объем изобретения, в зависимости от любой специально исключенной границы в указанных пределах. В тех случаях, когда указанные пределы включают одну или обе границы, то они также включаются в объем изобретения.
В последующем описании приводятся многочисленные конкретные детали для обеспечения полного понимания настоящего изобретения. Однако специалистам в данной области, очевидно, понятно, что настоящее изобретение может практиковаться без одной или более таких конкретных деталей. В других случаях хорошо известные признаки и способы, хорошо известные специалистам в данной области, не были описаны во избежание неопределенностей в описании изобретения.
Изобретение в целом
В гуморальной иммунной системе продуцируется репертуар различных антител посредством комбинаторного и соединительного разнообразия локусов генов цепей IgH (Igh) и IgG (Igl) в процессе, названном V(D)J-рекомбинацией. В развивающейся В-клетке первое рекомбинационное событие, которое должно произойти, находится между сегментами одного D-гена и одного J-гена локуса тяжелой цепи, и ДНК между этими двумя генами делецируется. После такой D-J-рекомбинации происходит объединение одного V-гена из области выше вновь образовавшегося DJ-комплекса, с формированием реарранжированного V(D)J-гена. Все другие гены между сегментами V и D нового V(D)J-гена уже на этот момент делецированы из генома отдельной В-клетки. Такой реарранжированный ген в конечном счете экспрессируется на поверхности В-клетки в виде IgH-полипептида, который связывается с IgL с образованием рецептора В-клетки. Мышиные и человеческие локусы Ig являются очень сложными, простираясь по области размером примерно 2 млн.п.н., включая несколько сегментов гена константной области, сегменты J-гена, сегменты D-гена и большее число вариабельных генов.
Настоящее изобретение относится к клеткам позвоночных животных, отличных от человека, содержащим введенную частично человеческую нуклеиновую кислоту, содержащую кодирующие области человеческих вариабельных областей и некодирующие последовательности из генома позвоночного животного-хозяина, например мышиные геномные некодирующие последовательности, когда млекопитающее-хозяин является мышью. Такая частично человеческая нуклеиновая кислота позволяет трансгенному животному продуцировать репертуар тяжелых цепей, содержащих человеческие VH области, в то время оставляя регуляторные последовательности и другие элементы, которые можно найти в промежуточных последовательностях в определенном геноме хозяина, которые помогают осуществлять эффективную продукцию антител и узнавание антигена. Настоящее изобретение относится к применению синтетической или рекомбинантно продуцированной частично человеческой области, содержащей человеческие кодирующие последовательности и «нечеловеческие» некодирующие последовательности из локуса VH.
Поскольку способы по изобретению могут иметь преимущество обеспечения двух или более групп сайтов сайт-специфической рекомбинации в сконструированном геноме, то стадия рекомбинация позволяет провести многочисленные вставки в частично человеческий локус.
В предпочтительных аспектах изобретения такая частично человеческая область, предназначенная для введения в клетку позвоночного животного-хозяина, содержит все или достаточное число известных генов VH человека. Однако в некоторых случаях может быть желательным использовать подгруппу таких генов VH, и в конкретных случаях можно использовать даже одну человеческую кодирующую последовательность VH, в клетках и животных по изобретению.
Предпочтительные аспекты изобретения включают млекопитающих и клетки млекопитающих, отличных от человека, содержащих частично человеческий локус иммуноглобулина, который включает гены VH человека и дополнительно включает кодирующие области D- и J-генов и пре-DJ-области, основанные на эндогенном геноме млекопитающего-хозяина, отличного от человека. В некоторых аспектах введенная частично человеческая область может содержать один или более полностью рекомбинированных V(D)J-сегментов.
В конкретном аспекте изобретения трансгенное млекопитающее, отличное от человека, содержит введенную нуклеиновую кислоту, включающую многочисленные гены VH человека с промежуточными последовательностями, основанными на промежуточных последовательностях, находящихся в локусах млекопитающего-хозяина, отличного от человека, и кодирующие последовательности D- и J-генов человека. В особенно предпочтительном аспекте частично человеческая нуклеиновая кислота содержит гены VH человека, пре-D-область, основанную на геноме млекопитающего-хозяина, отличного от человека, например геноме мыши, и экзоны D и J человека.
В примерном варианте осуществления, как более подробно описано в разделе «Примеры», полный эндогенный локус VH иммуноглобулина мыши (включая локус J558) делецирован, и экзоны VH локуса J558 области VH мыши замещаются нуклеиновой кислотой, содержащей 44 из генов VH человека, которые в результате перемежаются с некодирующими областями, которые соответствуют некодирующим последовательностям мыши. Полная введенная область VH иммуноглобулина дополнительно содержит человеческие D- и J-экзоны, а также гены VH. В данном аспекте пре-D-область размером 10 т.п.н. содержит мышиные последовательности, в то время как D- и J-области включают человеческие кодирующие последовательности. Предпочтительно D- и J-области обеспечиваются в виде человеческой кодирующей DJ-области, содержащей D-гены человека и J-гены человека.
В способах по изобретению используется комбинация гомологичной рекомбинации и сайт-специфической рекомбинации для получения клеток и животных по изобретению. Нацеленный на гомологичные участки вектор вначале используется для введения сайтов сайт-специфической рекомбинации в геном млекопитающего-хозяина в требуемом положении в эндогенных локусах иммуноглобулина. Инсерция сайта сайт-специфической рекомбинации в геномную последовательность посредством гомологичной рекомбинации ассоциированной нацеленной последовательности с геномной ДНК в условиях in vivo предпочтительно не модифицирует аминокислотную последовательность молекулы антитела, которая экспрессируется трансфектированной клеткой. При таком подходе сохраняется правильная транскрипция и трансляция иммуноглобулиновых генов, которые продуцируют требуемое антитело после вставки сайтов рекомбинации и необязательно любой дополнительной последовательности, такой как селектируемый маркерный ген. Однако в некоторых случаях возможно вставить сайт рекомбиназы и другие последовательности в последовательность локуса иммуноглобулина таким образом, что аминокислотная последовательность молекулы антитела изменяется в результате инсерции, но антитело по-прежнему сохраняет достаточные функциональные свойства для выполнения требуемой цели, и изобретение также включает такие инсерции.
Приведенные в качестве примера методологии гомологичной рекомбинации описаны в патентах США № 6689610; 6204061; 5631153; 5627059; 5487992 и 5464764, каждый из которых в полном объеме включен в данный документ для сведения.
В конкретных аспектах изобретения нацеленный на гомологичные участки вектор можно использовать для замещения некоторых последовательностей в эндогенном геноме, а также для введения сайтов сайт-специфической рекомбинации и селектируемых маркеров. Например, нацеленный на гомологичные участки вектор, используемый для введения 3'-элементов области VH гена, можно использовать для замещения мышиных пре-D- и DJ-последовательностей человеческими эквивалентными последовательностями.
Сайт-специфическая рекомбинация
Сайт-специфическая рекомбинация отличается от общей гомологичной рекомбинации в том отношении, что короткие специфические ДНК-последовательности, которые необходимы для узнавания рекомбиназы, являются единственными сайтами, в которых происходит рекомбинация. Для сайт-специфической рекомбинации необходимы специфические рекомбиназы для узнавания сайтов и катализа рекомбинации в этих сайтах. Было показано, что ряд бактериофаговых и дрожжевых систем сайт-специфической рекомбинации, каждая из которых содержит рекомбиназу и специфические узнаваемые сайты, функционирует в эукариотических клетках для интеграции ДНК и, следовательно, они применимы для использования в настоящем изобретении, и они включают P1 Cre/lox бактериофага, дрожжевую системы FLP-FRT и систему Dre тирозинового семейства сайт-специфических рекомбиназ. Такие системы и способы описаны, например, в патентах США № 7422889; 7112715; 6956146; 6774249; 5677177; 5885836; 5654182 и 4959317, которые включены в данный документ для сведения в отношении способов применения таких рекомбиназ. Способ обмена экспрессионными кассетами, опосредованного рекомбиназой (RMCE), упрощается при использовании комбинации сайта дикого типа и мутантного сайта loxP (или FRT и т.д.) вместе с негативным отбором. Однако он имеет место, и когда используются только немутантные сайты и/или в отсутствии отбора. Но эффективность в этом случае является очень низкой, поскольку в большей степени имеют место реакции вырезания, а не инсерции, и (без включения позитивного отбора) не будет происходить обогащения соответствующим образом мутированными клетками.
Известно, что другие системы на основе тирозинового семейства, такие как интеграза Int бактериофага лямбда, интеграза HK2022, а также системы, относящиеся к отдельному сериновому семейству рекомбиназ, такие как бактериофаговые интегразы phiC31, R4Tp901, функционируют в клетках млекопитающих с использованием их соответствующих сайтов рекомбинации (Tronche F. et al., 2002), и они применимы для использования в настоящем изобретении.
В способах по изобретению предпочтительно используются сайты сайт-специфической рекомбинации, в которых функционирует одна и та же рекомбиназа, но которая не способствует рекомбинации между сайтами. Например, сайт Lox P и мутированный сайт Lox P можно интегрировать в геном хозяина, но введение Cre хозяину не будет способствовать рекомбинации двух сайтов; в большей степени сайт Lox P будет рекомбинировать с другим сайтом Lox P, а мутированный сайт будет рекомбинировать только с другим мутированным аналогичным образом сайтом LoxP. Примеры таких мутированных сайтов рекомбинации включают сайты, которые содержат комбинацию инвертированных повторов, или сайты, которые содержат сайты рекомбинации, имеющие мутантные спейсерные последовательности. Например, имеются две группы вариантных сайтов рекомбиназы для получения стабильной интегративной рекомбинации Cre-loxP. Оба содержат мутации последовательностей в последовательности узнавания Cre либо в спейсерной области размером 8 п.н. или инвертированных повторах размером 13 п.н. Мутанты по спейсорной последовательности, такие как lox511 (Hoess R.H. et al., Nucleic Acids Res., 1986, 14:2287-2300), lox5171 и lox2272 (Lee G. and Saito I., Gene, 1998, 216:55-65), m2, m3, m7 и m11 (Langer S.J. et al., Nucleic Acids Res., 2002, 30:3067-3077), легко рекомбинируют друг с другом, но обладают существенно пониженной степенью рекомбинации с сайтом дикого типа. Данная группа мутантов использовалась для инсерции ДНК посредством обмена экспрессионными кассетами, опосредованного рекомбиназой (RMCE), с использованием невзаимодействующих сайтов рекомбинации Cre-Lox и невзаимодействующих сайтов рекомбинации FLP (Baer A. and Bode J., Curr. Opin. Biotechnol., 2001, 12:473-480; Albert H. et al., Plant J., 1995, 7:649-659; Seiber J. and Bode J., Biochemistry, 1997, 36:1740-1747; Schlake T. and Bode J., Boichemistry, 1994, 33:12746-12751).
Мутанты на основе инвертированных повторов представляют вторую группу вариантных сайтов рекомбиназы. Например, сайты LoxP могут содержать измененные основания в левом инвертированном повторе (мутант LE) или правом инвертированном повторе (мутант RE). Мутант LE, lox71, содержит участок размером 5 п.н. в 5'-конце левого инвертированного повтора, чья последовательность дикого типа заменена на TACCG (Araki et al., Nucleic Acids Res., 1997, 25:868-872). Аналогично мутант RE, lox66, содержит пять 3' оснований, замененных на CGGTA. Мутанты на основе инвертированных повторов используются для интеграции плазмидных инсертов в хромосомную ДНК с мутантом LE, сконструированным в виде «целевого» хромосомного сайта loxP, в который рекомбинирует мутантный RE. После рекомбинации сайты loxP располагаются в цис-положении, фланкируя вставленный сегмент. Механизм рекомбинации является таковым, что после рекомбинации один сайт loxP является двойным мутантом (содержащим обе мутации на основе инвертированных повторов LE и RE) и другой остается сайтом дикого типа (Lee L. and Sadowski P.D., Prog. Nuleic Acid Res. Mol. Biol., 2005, 80:1-42; Lee L. and Sadowski P.D., J. Mol. Biol., 2003, 326:397-412). Двойной мутант в достаточной степени отличается от сайта дикого типа, который не распознается Cre-рекомбиназой, и вставленный сегмент не вырезается.
В некоторых аспектах сайты сайт-специфической рекомбинации можно ввести в интроны, а не в кодирующие области нуклеиновой кислоты или регуляторные последовательности. При этом можно избежать случайного разрыва каких-либо регуляторных последовательностей или кодирующих областей, необходимых для правильной экспрессии антител после вставки сайтов сайт-специфической рекомбинации в геном животной клетки.
Введение сайтов сайт-специфической рекомбинации можно достичь обычными методами гомологичной рекомбинации. Такие методы описаны в следующих источниках, таких как Sambrook and Russell, 2001 (Molecular Cloning: A Laboratory Manual 3rd Ed. (Cold Spring Harbor, N.Y.: Cold Spring Laboratory Press) и Nagy A., 2003 (Manipulating the mouse embryo: A Laboratory Manual, 3rd Ed. (Cold Spring Harbor, N.Y.: Cold Spring Laboratory Press). Genetic Recombination: Nucleic acid, Homology (Biology), Homologous Recombination Non-homologous End Joining, DNA Repair, Bacteria, Eukaryote, Meiosis, Adaptive Immune System, V(D)J Recombination by Frederic P. Miller, Agnes F. Vandome and John McBrewster (Paperback - Dec. 23, 2009).
Специфическую рекомбинацию в геноме можно обеспечить с использованием векторов, сконструированных для позитивного или отрицательного отбора, как это известно в данной области. Для упрощения идентификации клеток, которые подверглись реакции замещения, можно использовать соответствующую систему гена-маркера, и клетки отобрать, например, при использовании селекционной среды. Однако для гарантии того, что геномная последовательность по существу не содержит ненужных нуклеиновокислотных последовательностей в двух концевых точках участка замещения или смежных с ними областях, желательно, чтобы маркерную систему/ген можно было удалить после отбора клеток, содержащих замещенную нуклеиновую кислоту.
В одном предпочтительном аспекте способов по настоящему изобретению клетки, в которых имело место замещение всего или части эндогенного иммуноглобулина, подвергаются негативному отбору при воздействии токсина или лекарственного препарата. Например, клетки, которые сохраняют экспрессию HSV-TK, можно отобрать применением аналогов нуклеозидов, таких как ганцикловир. В еще одном аспекте изобретения клетки, содержащие делецию эндогенной иммуноглобулиновой области, можно позитивно отобрать при использовании маркерного гена, который можно необязательно удалить из клеток после или в результате рекомбинантного события. Система позитивного отбора, которую можно использовать, основана на применении двух нефункциональных фрагментов гена-маркера, такого как HPRT, которые сводятся вместе посредством рекомбинантного события. Такие два фрагмента приводятся в функциональную ассоциацию при проведении успешной реакции замещения, и функционально восстановленный маркерный ген фланкируется с каждой стороны дополнительными сайтами сайт-специфической рекомбинации (которые являются другими по сравнению с сайтами сайт-специфической рекомбинации, использованным в реакции замещения), так, чтобы в итоге маркерный ген можно было вырезать из генома с использованием соответствующей сайт-специфической рекомбиназы.
Рекомбиназа может находиться в виде чистого белка или конструкции, временно экспрессированной внутри клетки, для обеспечения рекомбиназной активности. Альтернативно клетку можно использовать для получения трансгенного животного, которое можно скрестить с животным, которое экспрессирует указанную рекомбиназу, для получения потомства, у которого отсутствует маркерный ген и ассоциированные сайты рекомбинации.
Получение трансгенных животных
В конкретных аспектах изобретение относится к способам получения трансгенных животных, содержащих введенную частично человеческую область иммуноглобулина.
В одном аспекте клетка-хозяин, использованная для замещения эндогенных генов иммуноглобулина, представляет собой эмбриональную стволовую клетку (ES-клетку), которую затем можно использовать для получения трансгенного млекопитающего. Таким образом, согласно одному аспекту способы по изобретению дополнительно включают: выделение эмбриональной стволовой клетки, которая содержит введенную частично человеческую область иммуноглобулина и применение указанной ES-клетки для получения трансгенного животного, которое содержит замещенный частично человеческий локус иммуноглобулина.
В другом примере трансгенное животное является птицей, и животное получают с использованием примордиальных зародышевых клеток. Таким образом, согласно другому аспекту способы по изобретению дополнительно включают: выделение примордиальной зародышевой клетки, которая содержит введенную частично человеческую область иммуноглобулина и применение указанной зародышевой клетки для получения трансгенного животного, которое содержит замещенный частично человеческий локус иммуноглобулина. Способы получения таких трансгенных птиц раскрыты, например, в патентах США № 7323618 и 7145057, которые включены в данный документ для сведения.
Примеры
Последующие примеры приводятся для обеспечения специалистов в данной области полным раскрытием и описанием того, как осуществить и использовать настоящее изобретение, и они не предназначаются для ограничения объема того, что рассматривают заявители в качестве их изобретения, также они не предназначаются для того, чтобы представить или подразумевать, что примеры, приведенные ниже являются всеми или только проведенными экспериментами. Специалистам в данной области, очевидно, понятно, что можно сделать многочисленные изменения и/или модификации по отношению к изобретению, описанному в конкретных вариантах осуществления, не отступая от сущности или объема изобретения, как широко описано. Следовательно, настоящие варианты осуществления во всех аспектах следует рассматривать в качестве иллюстративных и не ограничивающих.
Были предприняты усилия для гарантированного обеспечения точности по отношению к использованным терминам и цифрам (например, к векторам, количествам, температуре и т.д.), но следует учитывать некоторые экспериментальные ошибки и отклонения. Если не указано иначе, то части являются частями по массе, молекулярная масса является средней молекулярной массой, температура выражена в градусах Цельсия и давление является атмосферным или близким к нему.
Примеры иллюстрируют нацеливание обоими 5'-вектором и 3'-вектором, которые фланкируют сайт рекомбинации и введения синтетической ДНК. Специалистам в данной области при чтении заявки, очевидно, понятно, что первым может быть нацеливание 5'-вектора, а за ним следует 3'-вектор, или вначале идет нацеливание 3'-вектора, а за ним следует 5'-вектор. В некоторых обстоятельствах нацеливание можно проводить одновременно с двойным механизмом детектирования.
Пример 1: введение частично человеческой области иммуноглобулина в локус гена VH генома мыши
Приведенный в качестве примера способ замещения участка генома млекопитающего частично человеческой областью иммуноглобулина показан на фигурах 1-6. На фигуре 1 приведена схема, показывающая различные стадии данного примерного аспекта способов по изобретению. Данный способ обеспечивает введение первого сайта сайт-специфической рекомбинации в геном млекопитающего, который можно ввести в 5'- или 3'-участок эндогенной области VH генома млекопитающего. Затем следует введение 102 второго сайта сайт-специфической рекомбинации в геном млекопитающего, который в сочетании с первым сайтом сайт-специфической рекомбинации фланкирует эндогенную иммуноглобулиновую область. Фланкированная эндогенная область делецируется 104 и вводится синтетическая нуклеиновая кислота, содержащая человеческие и «нечеловеческие» последовательности 106 посредством опосредованного рекомбиназой обмена.
Приведенный в качестве примера способ, показывающий введение частично мышиной-человеческой области иммуноглобулина в геномный локус мышиной ES-клетки, более подробно приведен на фигурах 2-6. На фигуре 2 показан нацеленный на гомологичные участки вектор 201, содержащий слитый белок пуромицин фосфотрансфераза-тимидинкиназа (puroΔTK) 203, фланкированный двумя различными сайтами узнавания рекомбиназы, например FRT 207 и loxP 205, для Flp и Cre, и модифицированными сайтами, например, для FRT 209 и loxP 211, которые не могут рекомбинировать соответственно с немодифицированными сайтами 207 и 205. Нацеленный вектор содержит кДНК человеческого рецептора дифтерийного токсина (hDTR) 217 для применения в негативном отборе клеток, экспрессирующих введенную конструкцию, на последующих стадиях. Нацеленный вектор также необязательно содержит визуальный маркер, такой как флуоресцентный зеленый белок (GFP) (не показан). Области 213 и 215 являются гомологичными соответственно к 5'- и 3'-участкам смежной области 223 в эндогенном мышином локусе, который является 5'-участком геномной области 219, содержащей мышиные эндогенные гены VH. Нацеленный на гомологичные участки вектор 201 вводится 202 в мышиную ES-клетку, которая имеет иммуноглобулиновую область 229, содержащую эндогенные гены VH 219, пре-D-область 221, область J-гена 225 и область константного гена 227 иммуноглобулиновой области. Сайты сайт-специфической рекомбинации и кДНК hDTR 217 нацеленного на гомологичные участки вектора 201 интегрируются 204 в 5' мышиный геном мышиной эндогенной области гена VH.
На фигуре 3 показана эффективность того же подхода, который приведен на фигуре 2, за исключением того, что добавляется дополнительная группа сайтов сайт-специфической рекомбинации, например сайт Rox 331 и модифицированный сайт Rox 333 для применения с Dre-рекомбиназой. На фигуре 3 показан нацеленный на гомологичные участки вектор 301, содержащий слитый белок пуромицин фосфотрансфераза-тимидинкиназа 303, фланкированный сайтами узнавания рекомбиназы FRT 307, loxP 305 и Rox 331 и модифицированными сайтами FRT 309, loxP 311 и Rox 333, которые не способны рекомбинировать соответственно с немодифицированными сайтами 307, 305 и 331. Нацеленный вектор также содержит кДНК человеческого рецептора дифтерийного токсина (hDTR) 317. Области 313 и 315 являются гомологичными соответственно для 5'- и 3'-участков смежной области 323 в эндогенном мышином локусе, который является 5'-участком геномной области 319, содержащей мышиные эндогенные гены VH. Нацеленный на гомологичные участки вектор 301 вводится 302 в мышиную иммуноглобулиновую область 329, которая содержит эндогенные гены VH 319, пре-D-область 321, область J-гена 325 и область константного гена 327 иммуноглобулиновой области. Сайты сайт-специфической рекомбинации и кДНК hDTR 317 нацеленного на гомологичные участки вектора 301 интегрируются 304 в 5' мышиный геном эндогенной области мышиного гена VH.
Как показано на фигуре 4, обеспечивается второй нацеленный на гомологичные участки вектор 401, содержащий миниген гипоксантинфосфорибозилтрансферазы (HPRT) 435 и ген резистентности к неомицину 437, и сайты узнавания рекомбиназы FRT 407, loxP 405 для Flp и Cre, которые обладают способностью рекомбинировать с сайтами FRT 407 и loxP 405, интегрированными с помощью первого нацеленного на гомологичные участки вектора. Области 431 и 433 являются гомологичными соответственно для 5'- и 3'-участков смежной области 441 в эндогенном мышином локусе, который является 3'-участком геномной области, содержащей мышиные эндогенные гены VH, D и J, и 5'-участком области константного гена. Нацеленный на гомологичные участки вектор 401 вводится 402 в модифицированную мышиную иммуноглобулиновую область, которая содержит эндогенные гены VH 419, пре-D-область 421, область J-гена 425 и область константного гена 427. Сайты сайт-специфической рекомбинации и миниген HPRT 435 и ген резистентности к неомицину 437 нацеленного на гомологичные участки вектора 401 интегрируются 404 в 5'-участок мышиного генома мышиной эндогенной области гена VH.
После того как сайты рекомбинации введены в геном млекопитающего-хозяина, эндогенная область иммуноглобулинового домена затем подвергается рекомбинации введением одной из рекомбиназ, соответствующих сайтам сайт-специфической рекомбинации в геноме, в данном примере рекомбиназ FLP или Cre. Как показано на фигуре 5, когда вводится FLP 502, то область, содержащая сайты сайт-специфической рекомбинации (509, 511, 507 и 505), и ген puroΔTK 503 сохраняются с дополнительным сайтом рекомбинации FLP 507 на данный момент присутствующих 3' других двух сайтов рекомбинации 507 и 505. 3'-область сайтов рекомбинации - включая hDTR 517, эндогенный иммуноглобулиновый домен (519, 521, 525) и гены HPRT 527 и Neo 529, введенные с использованием нацеленного на гомологичные участки вектора, делецируются. В том случае, когда используется Cre-рекомбиназа для опосредованной рекомбиназой делеции 504, то область делеции остается такой же, но только один сайт сайт-специфической рекомбинации 507 остается непосредственно в 3'-области гена puroΔTK. Способ зависит от второй нацеленности, которая скорее будет иметь место в той же хромосоме, чем в ее гомологе (т.е. в цис-положении, а не в транс-положении). Если нацеленность имеет место в транс-положении, то клетки не будут чувствительными к негативному отбору после Cre-рекомбинации.
Первичный скрининг на делецию эндогенной области иммуноглобулина можно провести саузерн-блоттингом, или первичными скринингами на основе полимеразной цепной реакции (ПЦР) с подтверждением результатов вторичными скринингами на основе саузерн-блоттинга и/или потери нативного аллеля колПЦР. HPRT позволит провести негативный отбор (6-тиогуанин-зависимый) в HPRT-дефицитных ES-клетках. ES-клетки с делецированной иммуноглобулиновой областью можно отобрать негативной селекцией с использованием гена hDTR.
На фигуре 6 показано введение частично человеческой последовательности в модифицированный мышиный геном. Сайт-специфический нацеленный вектор 629, содержащий частично человеческую область иммуноглобулина 610, предназначенную для введения в геном млекопитающего-хозяина, вводится 602 в геномную область 601 с делецированной эндогенной иммуноглобулиновой областью, содержащей сайты сайт-специфической рекомбинации (609, 611, 607 и 605) и ген puroΔTK 603. Сайт-специфический нацеленный вектор, состоящий из частично человеческой области иммуноглобулина, содержит i) область VH 619, содержащую 44 человеческих кодирующих областей VH и промежуточные последовательности, основанные на мышиных геномных эндогенных последовательностях; ii) пре-DJ-область размером 10 т.п.н. 621, содержащую мышиную последовательность и iii) DJ-область 625, содержащую кодирующие области D- и J-генов человека и промежуточные последовательности, основанные на эндогенных последовательностях мышиного генома. Частично человеческая область иммуноглобулина фланкирована сайтами рекомбинации (609, 611, 605 и 607), что позволит происходить рекомбинации с модифицированным эндогенным локусом. После введения соответствующей рекомбиназы 604 частично человеческая область иммуноглобулина интегрируется в геном выше области константного гена 627.
Первичный скрининг на введение частично человеческой иммуноглобулиновой области можно провести саузерн-блоттингом, или первичными скринингами на основе ПЦР с подтверждением результатов вторичными скринингами на основе саузерн-блоттинга и/или потери нативного аллеля колПЦР. Делеция гена HPRT 605 в качестве части рекомбинантного события позволит идентифицировать клетки, которые не подверглись рекомбинантному событию с использованием (6-тиогуанин-зависимого) негативного отбора.
Пример 2: введение частично человеческой области иммуноглобулина в геном мыши
В некоторых аспектах частично человеческая область иммуноглобулина будет содержать элементы, описанные в примере 1, но с наличием дополнительных последовательностей, например последовательностей, добавленных в стратегических целях для введения дополнительных регуляторных последовательностей, для гарантии обеспечения требуемого пространственного распределения во введенной иммуноглобулиновой области, для того чтобы некоторые кодирующие последовательности находились в адекватном соседнем расположении с другими последовательностями, смежными с замещенной областью иммуноглобулина и тому подобное. На фигуре 7 показано введение второй примерной частично человеческой последовательности в модифицированный геном мыши, полученный, как показано на фигурах 2-5 и описано в примере 1 выше.
Сайт-специфический нацеленный вектор, содержащий частично человеческую область иммуноглобулина 710, предназначенную для введения в геном млекопитающего-хозяина, вводится 702 в геномную область 701 с делецированной эндогенной иммуноглобулиновой областью, содержащей сайты сайт-специфической рекомбинации (709, 711, 707 и 705) и ген puroΔTK 703. Сайт-специфический нацеленный вектор, содержащий частично человеческую область иммуноглобулина, включает i) область VH 719, содержащую 1-43 человеческих кодирующих областей VH и промежуточные последовательности, основанные на эндогенных последовательностях мышиного генома; ii) пре-DJ-область размером 10 т.п.н. 721, содержащую мышиную последовательность; iii) DJ-область 725, содержащую кодирующие области D- и J-генов человека и промежуточные последовательности, основанные на эндогенных последовательностях мышиного генома и iv) нефункциональную область JH-гена мыши. Частично человеческая область иммуноглобулина фланкирована сайтами рекомбинации 709, 711, 705 и 707, что позволяет происходить рекомбинации с модифицированным эндогенным локусом. После введения соответствующей рекомбиназы 704 частично человеческая область иммуноглобулина интегрирует в геном выше области константного гена 727.
Как описано в примере 1, первичный скрининг на введение частично человеческой иммуноглобулиновой области можно провести саузерн-блоттингом, или первичными скринингами на основе ПЦР с подтверждением результатов вторичными скринингами на основе саузерн-блоттинга и/или потери нативного аллеля колПЦР. Делеция гена HPRT 705 в качестве части рекомбинантного события позволит идентифицировать клетки, которые не подверглись рекомбинантному событию с использованием (6-тиогуанин-зависимого) негативного отбора.
Пример 3: введение частично человеческой области иммуноглобулина в локус гена тяжелой цепи иммуноглобулина мышиного генома
Способ замещения участка генома млекопитающего частично человеческой областью иммуноглобулина показан на фигуре 8. В данном способе использовали введение первого сайта сайт-специфической рекомбинации в геном млекопитающего с последующим введением второго сайта сайт-специфической рекомбинации в геном млекопитающего. Два сайта фланкировали полный кластер сегментов генов VH-, DH- и JH-областей. Фланкированную эндогенную область делецировали с использованием соответствующей сайт-специфической рекомбиназы, как описано в данном документе.
Нацеленные векторы 803, 805, использованные для введения сайтов сайт-специфической рекомбиназы, с каждой стороны кластера сегментов генов VH-, DH- и JH-областей, в области мышиного иммуноглобулина дикого типа 801, содержали дополнительный сайт сайт-специфической рекомбиназы, который был модифицирован таким образом, что он по-прежнему эффективно распознавался рекомбиназой, но не рекомбинировал с немодифицированными сайтами. Данный сайт располагался в нацеленном векторе таким образом, что после делеции кластера сегментов генов VH-, DH- и JH-областей он не мог использоваться для второго сайт-специфического рекомбинационного события, в котором ненативный фрагмент ДНК продвигался в модифицированный локус VH. Способ продвижения ДНК в локус с использованием сайт-специфической рекомбиназы относится в данном документе к «обмену экспрессионными кассетами, опосредованному рекомбиназой». В данном примере ненативная ДНК представляла собой синтетическую нуклеиновую кислоту, содержащую человеческие и «нечеловеческие» последовательности.
Конструировали два ген-нацеленных вектора для осуществления только что описанного способа. Один из векторов 803 содержал мышиную геномную ДНК, отобранную с 5'-конца локуса выше наиболее дистального сегмента гена вариабельной области. Другой вектор 805 содержал мышиную геномную ДНК, отобранную внутри локуса вблизи сегментов гена J-области.
Ключевыми признаками 5'-вектора 803 в направлении от 5' к 3' были следующие: ген, кодирующий субъединицу дифтерийного токсина А (DTA) под транскрипционным контролем модифицированного промотора гена тимидинкиназы вируса герпеса простого типа I, сопряженного с двумя мутантными энхансерами транскрипции из полиомавируса; участок мышиной геномной ДНК размером 4,5 т.п.н., картированный выше наиболее дистального сегмента гена вариабельной области в локусе тяжелой цепи; генный сегмент J-области (нефункциональный); последовательность узнавания FRT для рекомбиназы Flp; участок геномной ДНК, содержащий промотор мышиного гена Polr2a; последовательность инициации транскрипции (метиониновый кодон, вставленный в консенсусную последовательность «Козак»); мутированная последовательность узнавания loxP (известная как сайт lox5171) для Cre-рекомбиназы; последовательность терминации транскрипции/полиаденилирования; последовательность узнавания loxP для Cre-рекомбиназы; ген, кодирующий слитый белок, состоящий из белка, придающего резистентность к пуромицину, слитого с усеченной формой тимидинкиназы (pu-TK) под транскрипционным контролем промотора из мышиного гена фосфоглицераткиназы 1, и участок мышиной геномной ДНК размером 3 т.п.н., картированный вблизи к последовательности размером 4,5 т.п.н. в векторе, и расположенный в нативной относительной ориентации.
Ключевыми признаками 3'-вектора 805 в направлении от 5' к 3' были следующие: ген, кодирующий субъединицу дифтерийного токсина А (DTA) под транскрипционным контролем модифицированного промотора гена тимидинкиназы вируса герпеса простого типа I, сопряженного с двумя мутантными энхансерами транскрипции из полиомавируса; участок мышиной геномной ДНК размером 3,7 т.п.н., содержащий сегменты мышиного гена J-области, ориентированные таким образом, что конец области, который картирован ближе всего к сегментам гена вариабельной области тяжелой цепи, наиболее близко находился к гену DTA в векторе; миниген, кодирующий гипоксантингуанинфосфорибозилтрансферазу человека (HPRT) под транскрипционным контролем промотора мышиного гена Polr2a; ген резистентности к неомицину под контролем промотора мышиного гена фосфоглицераткиназы 1; последовательность узнавания loxP для Cre-рекомбиназы, и участок мышиной геномной ДНК размером 2,1 т.п.н., который картирован в геноме непосредственно ниже фрагмента размером 3,7 т.п.н. с двумя фрагментами, ориентированными в той же конфигурации, что и в геноме мыши.
Мышиные эмбриональные стволовые клетки (ES-клетки) (полученные от мышей C57B1/6NTac) трансфектировали электропорацией 3'-вектором 805, следуя широко используемым протоколам. До электропорации ДНК-вектор линеаризовали рестриктазой NotI. Трансфектированные клетки высевали и через ≥24 ч их помещали в условия отбора по резистентности к препарату с использованием аналога неомицина G418. Колонии резистентных к препарату ES-клеток физически извлекали с чашек после того, как они становились видимыми невооруженным глазом через неделю. Собранные колонии разделяли, повторно высевали в микролуночные планшеты и культивировали в течение нескольких суток. Затем каждый из клеточных клонов разделяли таким образом, чтобы часть клеток можно было заморозить в качестве архивного материала, а остальные использовали для выделения ДНК для проведения аналитических исследований.
ДНК из клонов ES-клеток подвергали скринингу ПЦР с использованием широко применяемого ген-нацеленного анализа. Использовали четыре теста, и в каждом случае одну из последовательностей олигонуклеотидных ПЦР-праймеров картировали вне области идентичности между 3'-вектором 805 и геномной ДНК, в то время как другие картировали в новой ДНК между двумя плечами геномной идентичности в векторе (т.е. в HPRT или элементах гена neo). Согласно стандартному дизайну данные тесты предназначены для детектирования участков ДНК, которые будут находиться только в клонах клеток, полученных из трансфектированных клеток, которые подверглись полностью правильной гомологичной рекомбинации между направленным на тяжелую цепь 3'-вектором и геномом.
Проводили две отдельные трансфекции с 3'-вектором 805. Первая из них давала в целом два позитивных клона примерно из 300 клонов, подвергшихся скринингу, с использованием четырех анализов ПЦР. Вторая давала в целом 6 позитивных клонов, также примерно из 300 клонов, подвергшихся скринингу. В целом из двух трансфекций было отобрано шесть ПЦР-позитивных клонов для размножения с последующим дополнительным анализом с использованием саузерн-блоттинга.
Саузерн-блоттинг проводили согласно широко используемым протоколам с использованием трех зондов и геномной ДНК, расщепленной многочисленными рестриктазами, выбранными таким образом, чтобы комбинация зондов и гидролизатов позволяла идентифицировать структуру нацеленного локуса в клонах как правильно модифицированную в результате гомологичной рекомбинации. Один из зондов картирован к ДНК-последовательности, фланкирующей одну сторону области идентичности между нацеленным на 3'-тяжелую цепь вектором и геномной ДНК; второй зонд картирован вне области идентичности, но с другой ее стороны; и третий зонд картирован внутри новой ДНК между двумя плечами геномной идентичности в векторе (т.е. в HPRT или элементах гена neo).
Шесть ПЦР-позитивных клонов ES-клеток анализировали кариотипически с использованием флуоресцентной гибридизации in situ, предназначенной для обнаружения наиболее часто появляющихся хромосомных аберраций, которые возникают в мышиных ES-клетках. Клоны с такими аберрациями исключались из дальнейшего использования. Клоны ES-клеток, которые принимались за клоны, имеющие предполагаемую правильную геномную структуру, основываясь на данных саузерн-блоттинга, и в которых также отсутствовали детектируемые хромосомные аберрации, что основывалось на кариотипическом анализе, отбирали для дальнейшего использования.
Приемлемые клоны модифицировали 5'-вектором 803 с использованием методов и скрининговых тестов, которые в основном аналогичны по дизайну таковым с использованием 3'-вектора 805, за исключением того, что использовали отбор по пуромицину вместо отбора по G418/неомицину. ПЦР, зонды и гидролизаты также были приспособлены для соответствия геномной области, модифицированной 5'-вектором 805.
Клоны ES-клеток, которые были мутированы предполагаемым образом обоими 3'-тяжелая цепь и 5'-тяжелая цепь векторами, т.е. выделяют двойные-нацеленные клетки, несущие обе сконструированные мутации, после векторного нацеливания. Клоны должны подвергнуться генному нацеливанию в одной хромосоме, а не в гомологичных хромосомах (т.е. сконструированные мутации, созданные нацеленными векторами, должны быть в большей степени в цис-положении в одной цепи ДНК, чем в транс-положении гомологичных цепей ДНК). Клоны с цис-положением мутаций отличимы от таковых с транс-положением при использовании таких аналитических методов, как флуоресцентная гибридизация in situ метафазных пластинок с использованием зондов, которые гибридизуются с новой ДНК, находящейся в двух ген-нацеленных векторах между их плечами геномной идентичности. Два типа клонов также можно отличить один от другого трансфекцией их вектором, экспрессирующим Cre-рекомбиназу, и затем сравнением числа колоний, которые выдерживают отбор по ганцикловиру против гена тимидинкиназы, введенного 5'-вектором 803, и анализом фенотипа резистентности к препарату выживших клонов с помощью скринингового теста «родственный отбор», в котором некоторые клетки из клона тестируются на резистентность к пуромицину или G418/неомицину. Предполагается, что клетки с цис-положением мутаций дают примерно в 103 раз больше резистентных к ганцикловиру клонов, чем клетки с транс-положением в эксперименте такого типа. Большинство полученных цис-производных клонов, резистентных к ганцикловиру, также являются чувствительными к пуромицину и G418/неомицину в противоположность транс-производным клонам, резистентным к ганцикловиру, которые будут сохранять резистентность к обоим препаратам. Двойные-нацеленные клоны клеток с цис-положением сконструированных мутаций в локусе тяжелой цепи отбирают для дальнейшего использования.
Двойные-нацеленные клоны клеток трансфектируют вектором, экспрессирующим Cre-рекомбиназу, и трансфектированные клетки затем помещают в условия отбора по ганцикловиру, как в аналитическом протоколе, описанном выше. Выделяют резистентные к ганцикловиру клоны клеток и анализируют ПЦР и саузерн-блоттингом на присутствие предполагаемой делеции между двумя сконструированными мутациями, полученными с помощью 5'-нацеленных на тяжелую цепь и 3'-нацеленных на тяжелую цепь векторов. В данных клонах Cre-рекомбиназа приводит к рекомбинации 802 между сайтами loxP, введенными в локус тяжелой цепи двумя векторами, с получением конструкции, показанной в 807. Поскольку сайты loxP располагаются в одинаковых относительных ориентациях в двух векторах, то рекомбинация приводит к вырезанию круга ДНК, содержащего полный геномный фрагмент между двумя сайтами loxP. Круг не содержит ориджина репликации и, таким образом, не будет реплицироваться во время митоза и, следовательно, будет потерян из клонов клеток по мере того, как они подвергаются размножению. Полученные клоны несут делецию ДНК, которая первоначально находилась между двумя сайтами loxP.
Клоны ES-клеток, несущие делецию последовательности в одной из двух гомологичных копий их локуса тяжелой цепи иммуноглобулина, подвергают повторной трансфекции 804 с помощью экспрессионного вектора с Cre-рекомбиназой вместе с участком ДНК 809, содержащим частично человеческий локус тяжелой цепи иммуноглоблина, включающий сегменты генов V-, D- J-областей. Ключевыми признаками данного фрагмента ДНК 809 являются следующие: сайт lox5171; открытая рамка считывания гена резистентности к неомицину (без инициатора метионинового кодона, но в рамке считывания и в смежном положении с непрерывной открытой рамкой считывания в сайте lox5171); последовательность терминации транскрипции/полиаденилирования; сайт FRT; совокупность 44 сегментов гена вариабельной области тяжелой цепи, каждый состоящий из человеческих кодирующих последовательностей, вставленных в мышиные некодирующие последовательности; участок геномной ДНК размером 7,5 т.п.н. непосредственно выше кластера сегментов гена D-области в локусе мышиной тяжелой цепи; участок геномной ДНК размером 58 т.п.н., содержащий сегменты генов D- и J-областей; сайт loxP в противоположной относительной ориентации к сайту lox5171.
Трансфектированные клоны помещают в условия отбора по G418, которые приводят к обогащению клонами клеток, подвергшихся обмену экспрессионными кассетами, опосредованному рекомбиназой, в которых частично человеческая донорная ДНК 809 полностью интегрируется в делецированный локус тяжелой цепи иммуноглобулина между сайтами loxP и lox5171 с получением области ДНК, показанной в 811. Остальные элементы из 5'-вектора 803 удаляются посредством FLP-опосредованной рекомбинации 806, приводя к конечному гуманизированному локусу, показанному в 813.
Клоны резистентных к G418 ES-клеток анализируют ПЦР и саузерн-блотингом для определения того, насколько они прошли предполагаемый обмен экспрессионными кассетами, опосредованный рекомбиназой, без нежелательных реарранжировок или делеций. Клоны с наличием предполагаемой геномной структуры выбирают для дальнейшего применения.
Клоны ES-клеток, несущие частично человеческую ДНК тяжелой цепи иммуноглобулина 813 в мышином локусе тяжелой цепи подвергают микроинъекции в мышиные бластоцисты из штамма DBA/2 с получением частично химерной мыши, полученной из ES-клетки, согласно стандартным методам. Отбирают химерных мышей-самцов с наиболее высокими уровнями вклада ES-клеток в их шерстном покрове для спаривания с мышами-самками. Мыши-самки выбора будут представлять штамм C57Bl/6NTac и также будут нести трансген, кодирующий рекомбиназу Flp, который экспрессируется в их зародышевой линии. Потомство, полученное в результате этих спариваний, анализируют на присутствие частично человеческого локуса тяжелой цепи иммуноглобулина и на потерю FRT-фланкированного гена резистентности к неомицину, который был создан на стадии обмена экспрессионными кассетами, опосредованного рекомбиназой. Мышей, которые несут частично человеческий локус, будут использовать для создания колонии мышей.
Пример 4: введение частично человеческой области иммуноглобулина в локус гена иммуноглобулиновой каппа-цепи мышиного генома
Другой способ замещения участка генома млекопитающего частично человеческой областью иммуноглобулина показан на фигуре 9. Данный способ обеспечивает введение первого сайта сайт-специфической рекомбинации в геном млекопитающего, который можно ввести 5'- или 3'-область основного кластера сегментов генов VK- и JK-областей генома млекопитающих с последующим введением второго сайта сайт-специфической рекомбинации в геном млекопитающего, который в сочетании с первым сайтом сайт-специфической рекомбинации фланкирует полный кластер сегментов генов VK- и JK-областей. Затем фланкированная эндогенная область делецируется и замещается с использованием соответствующей сайт-специфической рекомбиназы.
Нацеленные векторы, использованные для введения сайтов сайт-специфической рекомбинации с каждой стороны кластера сегментов генов VK- и JK-областей, также включают дополнительный сайт сайт-специфической рекомбинации, который модифицирован таким образом, что он по-прежнему эффективно распознается рекомбиназой, но не рекомбинирует с немодифицированными сайтами. Данный сайт находится в нацеленном векторе так, что после делеции кластера сегментов генов VK- и JK-областей, он может использоваться для второго сайт-специфического рекомбинантного события, в котором ненативный фрагмент ДНК продвигается в модифицированный локус VK посредством обмена экспрессионными кассетами, опосредованного рекомбиназой. В данном примере ненативная ДНК представляет собой синтетическую нуклеиновую кислоту, содержащую как человеческие, так и «нечеловеческие» последовательности.
Конструировали два ген-нацеленных вектора для осуществления только что описанного способа. Один из векторов 903 содержал мышиную геномную ДНК, взятую с 5'-конца локуса выше самого дистального сегмента гена вариабельной области. Другой вектор 905 содержал мышиную геномную ДНК, взятую внутри локуса вблизи сегментов гена J-области.
Ключевыми признаками 5'-вектора 903 были следующие: ген, кодирующий субъединицу дифтерийного токсина А (DTA) под транскрипционным контролем модифицированного промотора гена тимидинкиназы вируса герпеса простого типа I, сопряженного с двумя мутантными энхансерами транскрипции из полиомавируса; участок мышиной геномной ДНК размером 6 т.п.н., картированный выше самого дистального сегмента гена вариабельной области в локусе каппа-цепи; последовательность узнавания FRT для рекомбиназы Flp; участок геномной ДНК, содержащий промотор мышиного гена Polr2a; последовательность инициации транскрипции (метиониновый кодон, вставленный в консенсусную последовательность «Козак»); мутированная последовательность узнавания loxP (известная как сайт lox5171) для Cre-рекомбиназы; последовательность терминации транскрипции/полиаденилирования; последовательность узнавания loxP для Cre-рекомбиназы; ген, кодирующий слитый белок, состоящий из белка, придающего резистентность к пуромицину, слитого с усеченной формой тимидинкиназы (pu-TK), под транскрипционным контролем промотора из мышиного гена фосфоглицераткиназы 1, и участок мышиной геномной ДНК размером 2,5 т.п.н., картированный близко к последовательности размером 6 т.п.н. в векторе и расположенный в нативной относительной ориентации.
Ключевыми признаками 3'-вектора 905 были следующие: ген, кодирующий субъединицу дифтерийного токсина А (DTA) под транскрипционным контролем модифицированного промотора гена тимидинкиназы вируса герпеса простого типа I, сопряженного с двумя мутантными энхансерами транскрипции из полиомавируса; участок мышиной геномной ДНК размером 6 т.п.н., взятый вблизи сегментов гена J-области каппа-цепи, ориентированный таким образом, что конец фрагмента, который картирован ближе всего к сегментам гена вариабельной области каппа-цепи, наиболее близко находился к гену DTA в векторе; миниген, кодирующий гипоксантингуанинфосфорибозилтрансферазу человека (HPRT) под транскрипционным контролем промотора мышиного гена Polr2a; ген резистентности к неомицину под контролем промотора мышиного гена фосфоглицераткиназы 1; последовательность узнавания loxP для Cre-рекомбиназы, и участок мышиной геномной ДНК размером 3,6 т.п.н., который картирован в геноме непосредственно ниже фрагмента размером 6 т.п.н., также включенного в вектор, с двумя фрагментами, ориентированными в той же относительной конфигурации, что и в геноме мыши.
Мышиные эмбриональные стволовые клетки (ES-клетки), полученные от мышей C57B1/6NTac, трансфектировали электропорацией 3'-вектором 905, следуя широко используемым протоколам. До электропорации ДНК-вектор линеаризовали рестриктазой NotI. Трансфектированные клетки высевали и через ≥24 ч их помещали в условия отбора по резистентности к препарату с использованием аналога неомицина G418. Колонии резистентных к препарату ES-клеток физически извлекали с чашек после того, как они становились видимыми невооруженным глазом через неделю. Собранные колонии разделяли, повторно высевали в микролуночные планшеты и культивировали в течение нескольких суток. Затем каждый из клеточных клонов разделяли таким образом, чтобы часть клеток можно было заморозить в качестве архивного материала, а остальные использовали для выделения ДНК для проведения аналитических исследований.
ДНК из клонов ES-клеток подвергали скринингу ПЦР с использованием широко применяемого ген-нацеленного анализа. Использовали четыре теста, и в каждом случае одну из последовательностей олигонуклеотидных ПЦР-праймеров картировали вне области идентичности между 3'-вектором 905 и геномной ДНК 901, в то время как другие картировали в новой ДНК между двумя плечами геномной идентичности в векторе (т.е. в HPRT или элементах гена neo). Согласно стандартному дизайну данные тесты предназначены для детектирования участков ДНК, которые будут находиться только в клонах клеток, полученных из трансфектированных клеток, которые подверглись полностью правильной гомологичной рекомбинации между нацеленным на тяжелую цепь 3'-вектором 905 и геномом.
Первую трансфекцию проводили 3'-вектором 905, и она давала в целом семнадцать позитивных клонов примерно из 300 клонов, подвергшихся скринингу, с использованием четырех ПЦР.
В целом в результате трансфекции было выделено шесть ПЦР-позитивных клонов для размножения с последующим дополнительным анализом с использованием саузерн-блоттинга. Саузерн-блоттинг проводили согласно широко используемым протоколам; они включали три зонда и геномную ДНК, расщепленную многочисленными рестриктазами, выбранными таким образом, чтобы комбинация зондов и гидролизатов позволяла сделать вывод о структуре нацеленного локуса в клонах и насколько он был правильно модифицирован в результате гомологичной рекомбинации. Один из зондов картирован к ДНК-последовательности, фланкирующей одну сторону области идентичности между нацеленным на 3'-каппа-цепь вектором и геномной ДНК; второй зонд также находился вне области идентичности, но с другой ее стороны; и третий зонд картирован внутри новой ДНК между двумя плечами геномной идентичности в векторе (т.е. в гене neo). С помощью саузерн-блоттинга было установлено присутствие предполагаемого Eco911-продуцированного фрагмента ДНК, соответствующего правильно мутированному фрагменту (в результате гомологичной рекомбинации с нацеленным на каппа-цепь 3'-вектором) локуса каппа-цепи, что детектировали одним из наружных зондов и неомициновым зондом (данные не представлены). Наружный зонд детектирует мутантный фрагмент и также фрагмент дикого типа от немутантной копии локуса каппа-цепи иммуноглобулина в гомологичной хромосоме.
Шесть ПЦР-позитивных клонов ES-клеток анализировали кариотипически с использованием флуоресцентной гибридизации in situ, предназначенной для обнаружения наиболее часто появляющихся хромосомных аберраций, которые возникают в мышиных ES-клетках. Один клон с такими аберрациями исключили из дальнейшего использования. Два кариотипически нормальных клона, которые принимались за клоны, имеющие предполагаемую правильную геномную структуру, основываясь на данных саузерн-блоттинга, отбирали для дальнейшего использования.
Два клона модифицировали 5'-вектором 903 с использованием методов и скрининговых тестов, которые в основном аналогичны по дизайну таковым с использованием 3'-вектора 905, за исключением того, что использовали отбор по пуромицину вместо отбора по G418/неомицину, и протоколы были приспособлены для соответствия геномной области, модифицированной 5'-вектором 903. Целью опытов по трансфекции 5'-вектором 903 является выделение клонов ES-клеток, которые были мутированы предполагаемым образом обоими 3'-вектором 905 и 5'-вектором 903, т.е. выделяли двойные-нацеленные клетки, несущие обе сконструированные мутации. В данных клонах Cre-рекомбиназа вызывает рекомбинацию 902 между сайтами loxP, введенными в локус каппа-цепи двумя векторами, для получения конструкции, представленной в 907.
Затем клоны должны подвергнуться ген-нацеливанию в одной хромосоме, а не в гомологичных хромосомах, т.е. сконструированные мутации с использованием нацеленных векторов должны быть в большей степени в цис-положении в той же цепи ДНК, чем в транс-положении в отдельных гомологичных цепях ДНК. Клоны с цис-положением мутаций отличимы от таковых с транс-положением при использовании таких аналитических методов, как флуоресцентная гибридизация in situ метафазных пластинок с использованием зондов, которые гибридизуются с новой ДНК, находящейся в двух ген-нацеленных векторах между их плечами геномной идентичности. Два типа клонов также можно отличить один от другого их трансфекцией вектором, экспрессирующим Cre-рекомбиназу, и затем сравнением числа колоний, которые выдерживают отбор по ганцикловиру против гена тимидинкиназы, введенного 5'-вектором 903, и анализом фенотипа резистентности к препарату выживших клонов с помощью скринингового теста «родственный отбор», в котором некоторые клетки из клона тестируются на резистентность к пуромицину или G418/неомицину.
Предполагается, что клетки с цис-положением мутаций дают примерно в 103 раз больше резистентных к ганцикловиру клонов, чем клетки с транс-положением в эксперименте такого типа. Большинство полученных цис-производных клонов, резистентных к ганцикловиру, также будут чувствительными к пуромицину и G418/неомицину в противоположность транс-производным клонам, резистентным к ганцикловиру, которые будут сохранять резистентность к обоим препаратам. Клоны клеток с цис-положением сконструированных мутаций в локусе каппа-цепи отбирают для дальнейшего использования.
Двойные-нацеленные клоны клеток трансфектируют вектором, экспрессирующим Cre-рекомбиназу, и трансфектированные клетки затем помещают в условия отбора по ганцикловиру, как в аналитическом протоколе, описанном выше. Выделяют резистентные к ганцикловиру клоны клеток и анализируют ПЦР и саузерн-блоттингом на присутствие предполагаемой делеции между двумя сконструированными мутациями, полученными с помощью 5'-вектора 903 и 3'-вектора 905. В данных клонах Cre-рекомбиназа приводила к рекомбинации между сайтами loxP, введенными в локус каппа-цепи двумя векторами. Поскольку сайты loxP располагаются в одинаковых относительных ориентациях в двух векторах, то рекомбинация приводит к вырезанию круга ДНК, содержащего полный геномный фрагмент между двумя сайтами loxP. Круг не содержит ориджина репликации и, таким образом, не будет реплицироваться во время митоза и, следовательно, будет потерян из клонов клеток по мере того, как они подвергаются размножению. Полученные клоны несут делецию ДНК, которая первоначально находилась между двумя сайтами loxP. Клоны, которые содержат, предполагаемую делецию, будут отобраны для дальнейшего использования.
Клоны ES-клеток, несущие делецию последовательности в одной из двух гомологичных копий их локуса каппа-цепи иммуноглобулина, подвергают повторной трансфекции 904 с помощью экспрессионного вектора с Cre-рекомбиназой вместе с участком ДНК 909, содержащим частично человеческий локус каппа-цепи иммуноглоблина, содержащий сегменты генов V- и J-областей. Ключевыми признаками данного участка ДНК (относящегося к «K-K») являются следующие: сайт lox5171; открытая рамка считывания гена резистентности к неомицину (без инициатора метионинового кодона, но в рамке считывания и в смежном положении с непрерывной открытой рамкой считывания в сайте lox5171); последовательность терминации транскрипции/полиаденилирования; сайт FRT; совокупность 39 сегментов гена вариабельной области каппа-цепи, каждый состоящий из человеческих кодирующих последовательностей, вставленных в мышиные некодирующие последовательности; участок геномной ДНК размером 13,5 т.п.н. непосредственно выше кластера сегментов гена J-области каппа-цепи в локусе мышиной каппа-цепи; участок геномной ДНК размером 2 т.п.н., содержащий 5 сегментов гена J-области человека, вставленных в мышиную некодирующую ДНК; сайт loxP в противоположной относительной ориентации к сайту lox5171.
Во втором независимом опыте альтернативный участок частично человеческой ДНК 909 используют вместо ДНК K-K. Ключевыми признаками данной ДНК (относящейся к «L-K») является следующие: сайт lox5171; открытая рамка считывания гена резистентности к неомицину без инициатора метионинового кодона, но в рамке считывания и в смежном положении с непрерывной открытой рамкой считывания в сайте lox5171; последовательность терминации транскрипции/полиаденилирования; сайт FRT; совокупность 38 сегментов гена вариабельной области лямбда-цепи, каждый состоящий из человеческих кодирующих последовательностей, вставленных в мышиные некодирующие последовательности; участок геномной ДНК размером 13,5 т.п.н. непосредственно выше кластера сегментов гена J-области в локусе мышиной каппа-цепи; участок геномной ДНК размером 2 т.п.н., содержащий 5 сегментов гена J-области лямбда-цепи, вставленных в мышиную некодирующую ДНК; сайт loxP в противоположной относительной ориентации к сайту lox5171.
Трансфектированные клоны из опытов по трансфекции с K-K и L-K помещают в условия отбора по G418, которые приводят к обогащению клонами клеток, подвергшихся обмену экспрессионными кассетами, опосредованному рекомбиназой, в котором частично человеческая донорная ДНК полностью интегрируется в делецированный локус каппа-цепи иммуноглобулина между сайтами loxP и lox5171, которые были помещены там с помощью соответственно 3'-вектора 905 и 5'-вектора 903. Область ДНК, созданная с использованием последовательности K-K, показана в 911 на фигуре 9. Остальные элементы из 5'-вектора 903 удаляются посредством FLP-опосредованной рекомбинации 906, приводя к конечному гуманизированному локусу, показанному в 913.
Клоны резистентных к G418 ES-клеток анализируют ПЦР и саузерн-блотингом для определения того, насколько подверглись предполагаемому обмену экспрессионными кассетами, опосредованному рекомбиназой, без нежелательных реарранжировок или делеций. Оба клона K-K и L-K с наличием предполагаемой геномной структуры отбирают для дальнейшего применения.
Клоны ES-клеток K-K и клоны ES-клеток L-K, каждый несущий частично человеческую ДНК в мышином локусе каппа-цепи, подвергают микроинъекции в мышиные бластоцисты из штамма DBA/2 с получением частично химерной мыши, полученной из ES-клетки, согласно стандартным методам. Отбирают химерных мышей-самцов с наиболее высокими уровнями вклада ES-клеток в их шерстном покрове для спаривания с мышами-самками. Мыши-самки выбора представляют штамм C57Bl/6NTac, и они также должны нести трансген, кодирующий рекомбиназу Flp, который экспрессируется в их зародышевой линии. Потомство, полученное в результате этих спариваний, анализируют на присутствие частично человеческого локуса каппа-цепи иммуноглобулина и на потерю FRT-фланкированного гена резистентности к неомицину, который был создан на стадии обмена экспрессионными кассетами, опосредованного рекомбиназой. Мышей, которые несут частично человеческий локус, будут использовать для создания колоний мышей K-K и L-K.
Мышей, несущих частично человеческий (т.е. гуманизированный) локус тяжелой цепи, полученных, как описано в примере 3, можно спарить с мышами, несущими гуманизированный локус каппа-цепи. В свою очередь, их потомство спаривают по схеме, которая в конечном итоге дает мышей, которые являются гомозиготными для обоих гуманизированных локусов, т.е. гуманизированных в отношении тяжелой цепи и каппа-цепи. Такие мыши продуцируют частично человеческие тяжелые цепи, состоящие из человеческих вариабельных доменов и мышиных константных доменов. Они также продуцируют частично человеческие белки каппа-цепи, состоящие из человеческих вариабельных доменов каппа-цепи и мышиного константного домена каппа-цепи из их каппа локусов. Моноклональные антитела, полученные от этих мышей, состоят из человеческих вариабельных доменов, спаренных с человеческими вариабельными доменами каппа-цепи.
Изменение в схеме размножения включает получение мышей, которые являются гомозиготными для гуманизированного локуса тяжелой цепи, но гетерозиготными в локусе каппа-цепи так, что в одной хромосоме они содержат гуманизированный локус K-K и в другой хромосоме они содержат гуманизированный локус L-K. Такие мыши продуцируют частично человеческие тяжелые цепи, состоящие из человеческих вариабельных доменов и мышиных константных доменов. Они также продуцируют частично человеческие белки каппа-цепи, состоящие из человеческих вариабельных доменов каппа-цепи, и мышиного константного домена каппа-цепи из одного из их каппа локусов. Из другого каппа локуса они будут продуцировать частично человеческие белки лямбда-цепи, состоящие из человеческих вариабельных доменов лямбда-цепи и мышиного константного домена каппа-цепи. Моноклональные антитела, выделенные от этих мышей, состоят из человеческих вариабельных доменов, в некоторых случаях спаренных с человеческими вариабельными доменами каппа-цепи и в других случаях с человеческими вариабельными доменами лямбда-цепи.
Пример 5: введение частично человеческой области иммуноглобулина в локус гена лямбда-цепи иммуноглобулина мышиного генома
Способ замещения участка генома млекопитающих частично человеческой областью иммуноглобулина показан на фигуре 10. Данный способ обеспечивает делецию фрагмента ДНК размером примерно 194 т.п.н. из мышиного локуса лямбда-цепи иммуноглобулина дикого типа 1001 посредством гомологичной рекомбинации, включающей нацеленный вектор 1003, который разделяет идентичность с локусом выше сегмента гена V2 и ниже сегмента гена V1 в непосредственной близости с сегментами J30-, C3-, J1- и С1-генов. Вектор замещает фрагмент ДНК размером 194 т.п.н. элементами, предназначенными обеспечить последующую сайт-специфическую рекомбинацию, в которой ненативный участок ДНК продвигается в модифицированный локус VL посредством обмена экспрессионными кассетами, опосредованного рекомбиназой 1002. В данном примере ненативная ДНК представляет синтетическую нуклеиновую кислоту, содержащую как человеческие, так и «нечеловеческие» последовательности.
Ключевыми признаками ген-нацеленного вектора 1003 для осуществления делеции фрагмента размером 194 т.п.н. были следующие: ген негативного отбора, такой как ген, кодирующий субъединицу дифтерийного токсина А, или ген тимидинкиназы вируса герпеса простого; участок геномной ДНК размером 4 т.п.н. из 5'-области сегмента мышиного гена вариабельной области V2 в лямбда локусе; сайт FRT; участок геномной ДНК, содержащий промотор мышиного гена Polr2a; последовательность инициации трансляции (метиониновый кодон, вставленный в консенсусную последовательность «Козак»); мутированная последовательность узнавания loxP (известная как сайт lox5171) для Cre-рекомбиназы; последовательность терминации транскрипции/полиаденилирования; открытая рамка считывания, кодирующая белок, который придает резистентность к пуромицину; данная открытая рамка считывания должна находиться в антисмысловой цепи относительно промотора Polr2a и последовательности инициации трансляции, следующей за ней; за ней также должна следовать ее собственная последовательность терминации транскрипции/полиаденилирования; последовательность узнавания loxP для Cre-рекомбиназы; последовательность инициации трансляции (метиониновый кодон, вставленный в консенсусную последовательность «Козак») в той же антисмысловой цепи, что и открытая рамка считывания гена резистентности к пуромицину; промотор бета-актина цыпленка и ранний энхансерный элемент цитомегаловируса, ориентированные таким образом, что они регулируют транскрипцию открытой рамки считывания резистентности к пуромицину с инициацией трансляции в кодоне инициации ниже сайта loxP и продолжающийся назад через сайт loxP в открытую рамку считывания пуромицина - все в антисмысловой цепи относительно промотора Polr2a и последовательности инициации трансляции за ней; мутированный сайт узнавания рекомбиназы Flp, известный как сайт «F3»; участок геномной ДНК размером 7,3 т.п.н., содержащей сегменты J3-, C3-, J1- и С1-генов и окружающие последовательности; второй ген негативного отбора, такой как ген, кодирующий субъединицу дифтерийного токсина А, или ген тимидинкиназы вируса герпеса простого.
Мышиные эмбриональные стволовые клетки (ES-клетки) (полученные от мышей C57Bl/6NTac) трансфектируют 1002 электропорацией с помощью нацеленного вектора 1003, следуя широко используемым протоколам. Полученная конструкция 1005 будет замещать нативную ДНК последовательностями из нацеленного вектора 1003 в области размером 196 т.п.н.
Перед электропорацией ДНК-вектор линеаризуют редкощепящей рестриктазой, которая разрезает только в прокариотической плазмидной последовательности или полилинкере, ассоциированном с ней. Трансфектированные клетки высевают и через ≥24 ч их помещали в условия отбора по резистентности к препарату с использованием пуромицина. Колонии резистентных к препарату ES-клеток физически извлекают с чашек после того, как они становятся видимыми невооруженным глазом через неделю. Эти собранные колонии разделяют, повторно высевают в микролуночные планшеты и культивируют в течение нескольких суток. Затем каждый из клеточных клонов разделяют таким образом, чтобы часть клеток можно было заморозить в качестве архивного материала, а остальные используют для выделения ДНК для проведения аналитических исследований.
ДНК из клонов ES-клеток подвергают скринингу ПЦР с использованием широко применяемого ген-нацеленного анализа. Использовали четыре теста, и в каждом случае одну из последовательностей олигонуклеотидных ПЦР-праймеров картируют вне области идентичности между нацеленным вектором и геномной ДНК, в то время как другие картируют в новой ДНК между двумя плечами геномной идентичности в векторе (т.е. в гене puro). Согласно стандартному дизайну данные тесты детектируют участки ДНК, которые будут находиться только в клонах клеток, полученных из трансфектированных клеток, которые подверглись полностью правильной гомологичной рекомбинации между нацеленным вектором 1003 и нативной ДНК 1001.
Примерно шесть ПЦР-позитивных клонов из трансфекции 1002 отбирают для размножения с последующим дополнительным анализом с использованием саузерн-блоттинга. Саузерн-блоттинг включает три зонда и геномную ДНК из клонов, которые были расщеплены многочисленными рестриктазами, выбранными таким образом, что комбинация зондов и гидролизатов позволяет идентифицировать, насколько правильно они были модифицированы в результате гомологичной рекомбинации.
Шесть ПЦР-позитивных клонов ES-клеток анализируют кариотипически с использованием флуоресцентной гибридизации in situ, предназначенной для обнаружения наиболее часто появляющихся хромосомных аберраций, которые возникают в мышиных ES-клетках. Клоны с признаками таких аберраций исключаются из дальнейшего использования. Кариотипически нормальные клоны, которые принимались за клоны, имеющие предполагаемую правильную геномную структуру, основываясь на данных саузерн-блоттинга, отбирают для дальнейшего использования.
Клоны ES-клеток, несущие делецию в одной из двух гомологичных копий их локуса лямбда-цепи иммуноглобулина, повторно трансфектируют 1004 с помощью экспрессионного вектора с Cre-рекомбиназой вместе с участком ДНК 1007, содержащим частично человеческий локус лямбда-цепи иммуноглоблина, включающий сегменты генов V-, J- и С-областей. Ключевыми признаками данного участка ДНК 1007 являются следующие: сайт lox5171; открытая рамка считывания гена резистентности к неомицину (без инициатора метионинового кодона, но в рамке считывания и в смежном положении с непрерывной открытой рамкой считывания в сайте lox5171); последовательность терминации транскрипции/полиаденилирования; сайт FRT; совокупность 38 сегментов гена вариабельной области лямбда-цепи, каждый состоящий из человеческих кодирующих последовательностей лямбда-цепи, вставленных в мышиные некодирующие последовательности лямбда-цепи; совокупность J-C-единиц, где каждая единица состоит из сегмента человеческого гена J-области лямбда-цепи и сегмента мышиного гена константного домена лямбда-цепи, вставленных в некодирующие последовательности из мышиного локуса лямбда-цепи (сегменты человеческого гена J-области лямбда-цепи будут представлять сегменты, кодирующие J1, J2, J6 и J7, в то время как сегменты мышиного гена константного домена лямбда-цепи будут представлять С1 и/или С2, и/или С3); мутированный сайт узнавания рекомбиназы Flp, известный как сайт «F3»; открытая рамка считывания, придающая резистентность к гигромицину; открытая рамка считывания располагается в антисмысловой цепи относительно сегмента гена, кодирующего информацию в конструкции; сайт loxP в противоположной относительной ориентации к сайту lox5171.
Трансфектированные клоны помещают в условия отбора по G418 и/или гигромицину, которые приводят к обогащению клонами клеток, подвергшихся обмену экспрессионными кассетами, опосредованному рекомбиназой, в котором частично человеческая донорная ДНК полностью интегрируется в делецированный локус лямбда-цепи иммуноглобулина между сайтами loxP и lox5171, которые были помещены там с помощью ген-нацеленного вектора. Остальные элементы из нацеленного вектора 1003 удаляются посредством FLP-опосредованной рекомбинации 1006, приводя к конечному гуманизированному локусу, показанному в 1011.
Клоны резистентных к G418/гигромицину E-клеток анализируют ПЦР и саузерн-блотингом для определения того, насколько они подверглись предполагаемому обмену экспрессионными кассетами, опосредованному рекомбиназой, без нежелательных реарранжировок или делеций. Клоны с наличием предполагаемой геномной структуры выбирают для дальнейшего применения.
Клоны ES-клеток, несущих частично человеческую ДНК иммуноглобулина 1001 в мышином локусе лямбда-цепи подвергают микроинъекции в мышиные бластоцисты из штамма DBA/2 с получением частично химерной мыши, полученной из ES-клетки, согласно стандартным методам. Отбирают химерных мышей-самцов с наиболее высокими уровнями вклада ES-клеток в их шерстном покрове для спаривания с мышами-самками. Мыши-самки выбора будут представлять штамм C57Bl/6NTac, которые несут трансген, кодирующий рекомбиназу Flp, который экспрессируется в их зародышевой линии. Потомство, полученное в результате этих спариваний, анализируют на присутствие частично человеческого локуса лямбда-цепи иммуноглобулина и на потерю FRT-фланкированного гена резистентности к неомицину и FRT-фланкированного гена резистентности к гигромицину, которые были получены на стадии обмена экспрессионными кассетами, опосредованного рекомбиназой. Мыши, которые несут частично человеческий локус, используются для создания колонии мышей.
В некоторых аспектах мышей, содержащих гуманизированную тяжелую цепь и каппа локус (как описано в примерах 3 и 4), скрещивают с мышами, которые несут гуманизированный локус лямбда-цепи. Мыши, полученные в результате схемы скрещивания такого типа, являются гомозиготными для гуманизированного локуса тяжелой цепи, и они могут быть гомозиготными для гуманизированного локуса K-K или гуманизированного локуса L-K. Альтернативно они могут быть гетерозиготными в каппа локусе, неся локус K-K в одной хромосоме и локус L-K в другой хромосоме. Каждая из этих мышей будет гомозиготной для гуманизированного локуса лямбда-цепи. Моноклональные антитела, полученные от этих мышей, будут состоять из человеческих вариабельных доменов, в некоторых случаях спаренных с человеческими вариабельными доменами каппа-цепи и в других случаях с человеческими вариабельными доменами лямбда-цепи. Вариабельные домены лямбда-цепи будут происходить из гуманизированного локуса L-K или гуманизированного локуса лямбда-цепи.
Пример 6: введение частично человеческого минигена иммуноглобулина в мышиный геном
В некоторых других аспектах частично человеческая область иммуноглобулина будет содержать человеческий миниген вариабельного домена, такой как показан на фигуре 11. В данном случае вместо частично человеческой области иммуноглобулина, содержащей все или по существу все гены VH человека, мышиная иммуноглобулиновая область замещается минигеном 1119, содержащим меньше генов VH человека, например 1-43 человеческих генов VH.
Сайт-специфический нацеленный вектор 1129, содержащий частично человеческую область иммуноглобулина 1100, предназначенную для введения в геном млекопитающего-хозяина, вводится 1102 в геномную область 1101 с делецированной эндогенной областью иммуноглобулина, содержащей сайты сайт-специфической рекомбинации (1109, 1111, 1107 и 1105) и ген puroΔTK 1103. Сайт-специфический нацеленный вектор, содержащий частично человеческую область иммуноглобулина, включает: i) область VH 1119, содержащую все 44 человеческих кодирующих области VH и промежуточные последовательности, основанные на эндогенных последовательностях мышиного генома; ii) пре-DJ-область размером 10 т.п.н. 721, содержащую мышиную последовательность; iii) DJ-область 1125, содержащую кодирующие области D- и J-генов человека и промежуточные последовательности, основанные на эндогенных последовательностях мышиного генома; и iv) мышиную нефункциональную область JH-гена. Частично человеческая область иммуноглобулина фланкируется сайтами рекомбинации (1109, 1111, 1105 и 1107), которые позволят происходить рекомбинации с модифицированным эндогенным локусом. После введения соответствующей рекомбиназы 1104 частично человеческая область иммуноглобулина интегрируется в геном выше области константного гена 1127.
Как описано в примере 1, можно провести первичный скрининг на введение частично человеческого локуса вариабельной области иммуноглобулина саузерн-блоттингом, или первичными скринингами на основе ПЦР с подтверждением результатов вторичными скринингами на основе саузерн-блоттинга и/или потери нативного аллеля колПЦР. Делеция гена HPRT 1105 в качестве части рекомбинантного события позволит идентифицировать клетки, которые не подверглись рекомбинантному событию с использованием (6-тиогуанин-зависимого) негативного отбора.
Приведенное выше описание показывает принципы изобретения. Специалистам в данной области, очевидно, понятно, что можно провести различные переделки, которые не описаны или не показаны подробно в данном документе, которые воплощают принципы изобретения, и включаются, не отступая от сущности и объема. Кроме того, все примеры и условные обозначения, приведенные в данном документе, в основном предназначаются для того, чтобы помочь читателю понять принципы изобретения и идеи, внесенные заявителями в область техники, и их следует истолковывать без ограничения конкретно приведенными примерами и условиями. Кроме того, все положения в данном документе, излагающие принципы, аспекты и варианты осуществления изобретения, а также их конкретные примеры предназначаются для включения их структурных и функциональных эквивалентов. Кроме того, полагается, что такие эквивалентные варианты включают известные в настоящее время эквиваленты и эквиваленты, которые будут разработаны в будущем, т.е. любые разработанные элементы, которые выполняют одинаковую функцию, независимо от структуры. Следовательно, объем настоящего изобретения не ограничивается примерными вариантами осуществления, показанными и описанными в данном документе. В большей степени объем и сущность настоящего изобретения воплощаются в прилагаемой формулой изобретения. В формуле изобретения, которая следует, если только не используется термин «средства», то ни один из признаков или элементов, изложенных в данном документе, не следует рассматривать в качестве ограничений средства-плюс-функция согласно 35 U.S.C. §112, ¶6.
| название | год | авторы | номер документа |
|---|---|---|---|
| МЫШИ С ГУМАНИЗИРОВАННОЙ ЛЕГКОЙ ЦЕПЬЮ | 2012 |
|
RU2664181C2 |
| МЫШИ, ЭКСПРЕССИРУЮЩИЕ ГИБРИДНУЮ ЛЕГКУЮ ЦЕПЬ ИММУНОГЛОБУЛИНА | 2011 |
|
RU2601297C2 |
| МЫШИ, ЭКСПРЕССИРУЮЩИЕ ЛЕГКУЮ ЦЕПЬ С ВАРИАБЕЛЬНОЙ ОБЛАСТЬЮ АЛЬФА ЧЕЛОВЕКА И КОНСТАНТНОЙ ОБЛАСТЬЮ МЫШИ | 2011 |
|
RU2590594C2 |
| МЫШИ С ГУМАНИЗИРОВАННОЙ ЛЕГКОЙ ЦЕПЬЮ | 2012 |
|
RU2761639C2 |
| МЫШИ С ГУМАНИЗИРОВАННОЙ УНИВЕРСАЛЬНОЙ ЛЕГКОЙ ЦЕПЬЮ | 2012 |
|
RU2664232C2 |
| ПРОДУЦИРУЮЩИЕ АНТИТЕЛА МЛЕКОПИТАЮЩИЕ, НЕ ЯВЛЯЮЩИЕСЯ ЧЕЛОВЕКОМ | 2020 |
|
RU2820926C2 |
| МЫШИ С ГУМАНИЗИРОВАННОЙ УНИВЕРСАЛЬНОЙ ЛЕГКОЙ ЦЕПЬЮ | 2012 |
|
RU2787781C2 |
| МЫШИ, ЭКСПРЕССИРУЮЩИЕ ЛЕГКУЮ ЦЕПЬ С ВАРИАБЕЛЬНОЙ ОБЛАСТЬЮ λ ЧЕЛОВЕКА И КОНСТАНТНОЙ ОБЛАСТЬЮ МЫШИ | 2011 |
|
RU2724736C2 |
| ЖИВОТНЫЕ, НЕ ЯВЛЯЮЩИЕСЯ ЧЕЛОВЕКОМ, ИМЕЮЩИЕ СКОНСТРУИРОВАННУЮ ЛЕГКУЮ ЦЕПЬ ЛЯМБДА ИММУНОГЛОБУЛИНА, И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2778410C2 |
| ГУМАНИЗИРОВАННЫЕ ГРЫЗУНЫ, КОТОРЫЕ ЭКСПРЕССИРУЮТ ТЯЖЕЛЫЕ ЦЕПИ, СОДЕРЖАЩИЕ ДОМЕНЫ VL | 2013 |
|
RU2664185C2 |
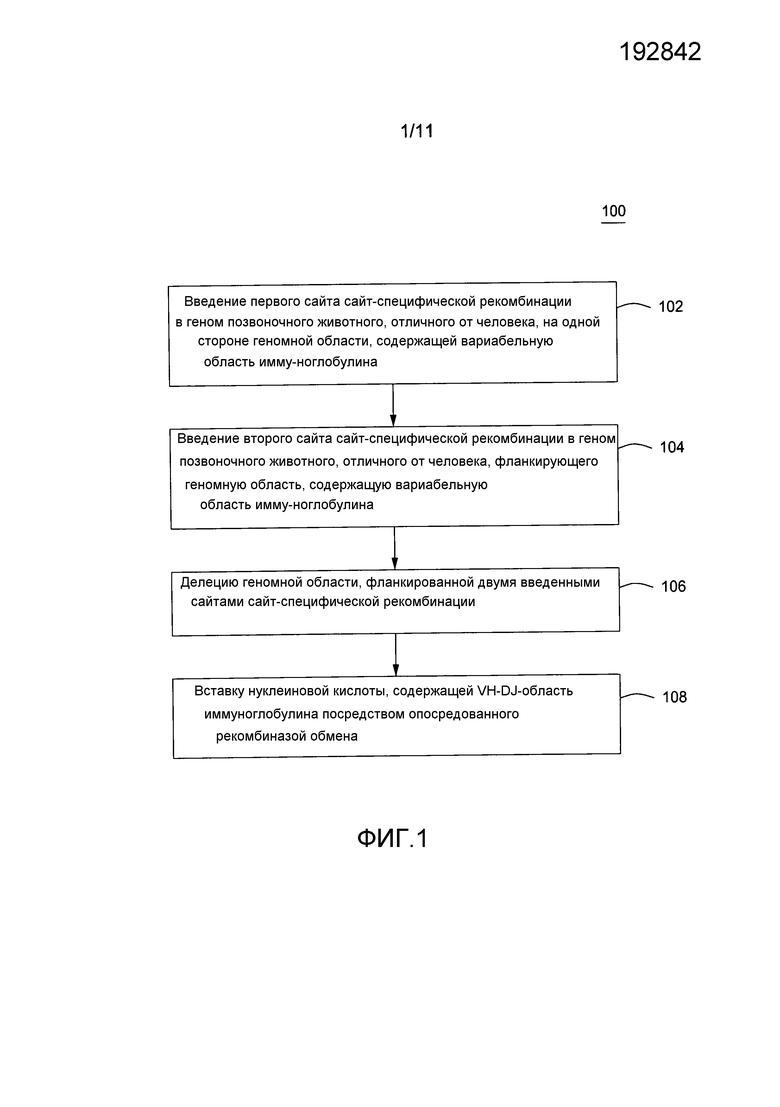

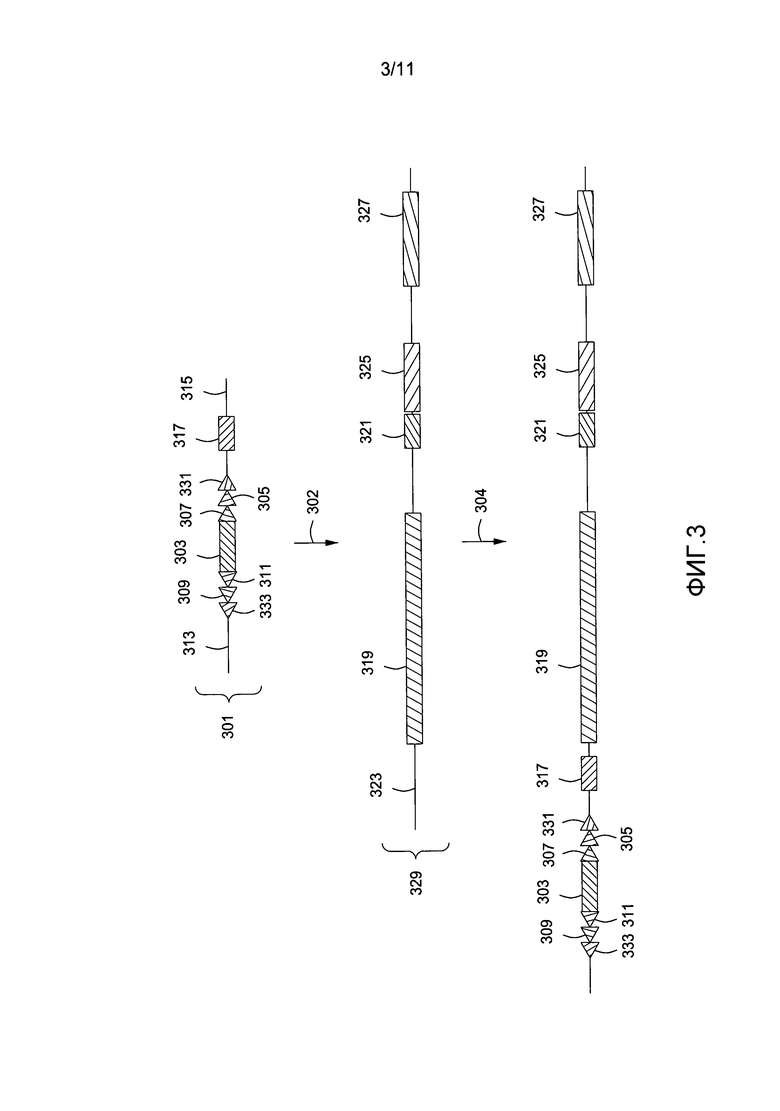

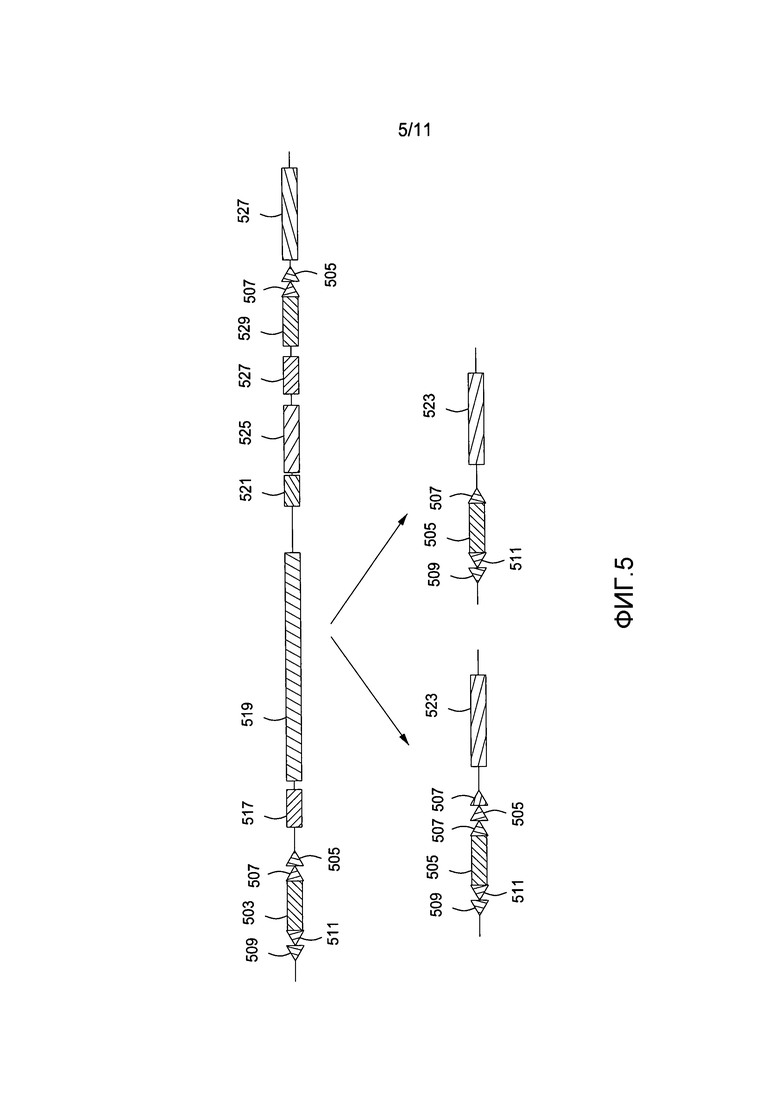




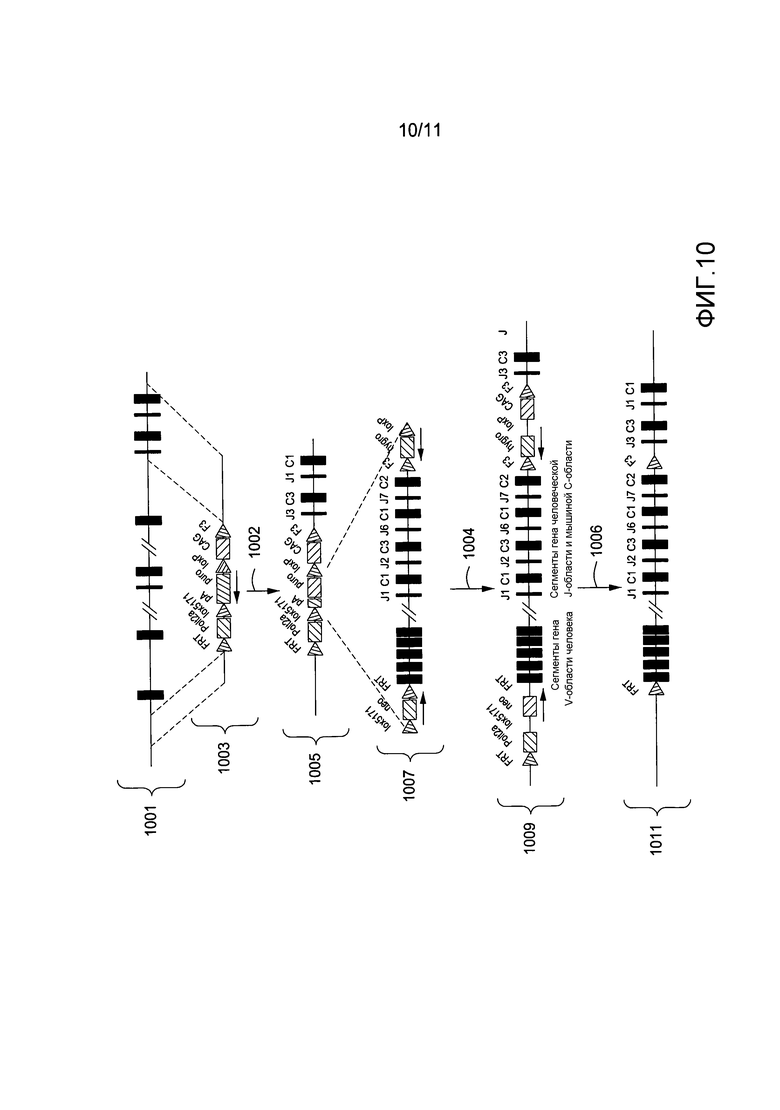
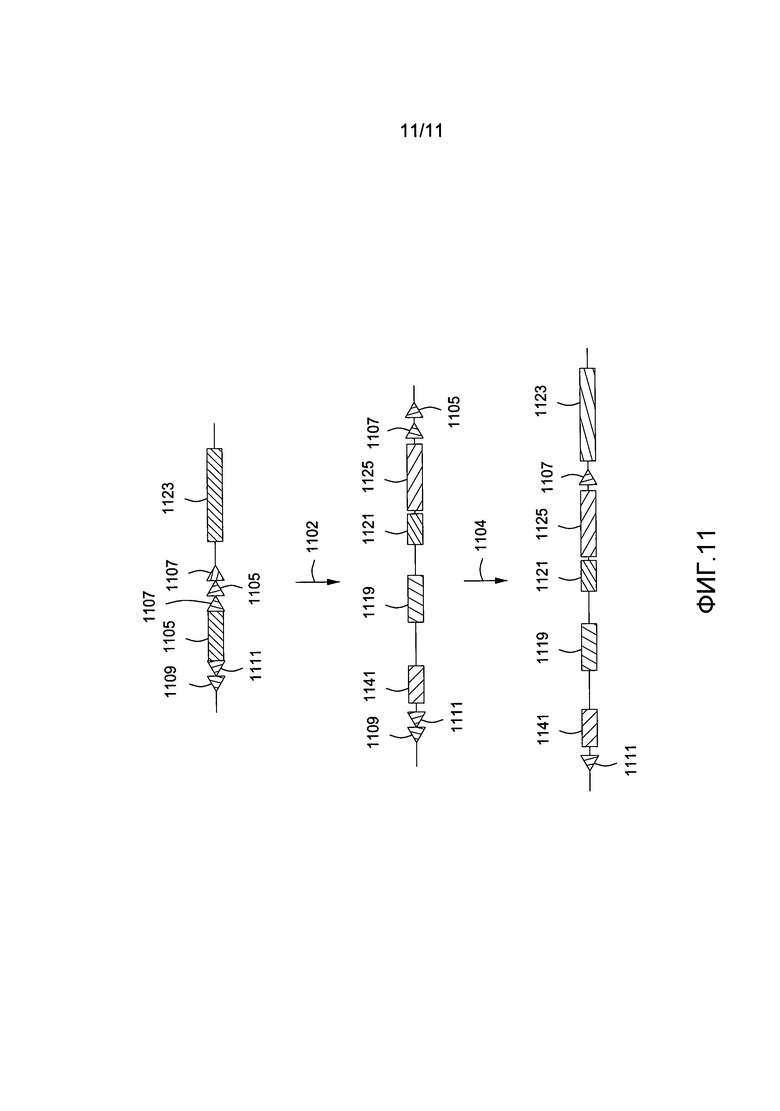
Изобретение относится к области биотехнологии и генной инженерии. Описана трансгенная мышь, имеющая геном, содержащий введенную частично человеческую область иммуноглобулина, где указанная введенная область содержит человеческие кодирующие последовательности VH и некодирующие последовательности VH, основанные на эндогенном геноме мыши. Указанная мышь способна экспрессировать цепи антител с вариабельными областями человека. Также описан способ получения такой мыши. Изобретение может быть использовано в иммунологии. 2 н. и 4 з.п. ф-лы, 11 ил., 6 пр.
1. Трансгенная мышь с геномом, содержащим сконструированный эндогенный локус иммуноглобулина с удаленными его сегментами гена вариабельной области, и в который введена совокупность химерных сегментов гена иммуноглобулина, где каждый из химерных сегментов гена состоит из последовательностей, кодирующих иммуноглобулин человека, и некодирующих последовательностей, основанных на локусе вариабельной области иммуноглобулина мыши, и где присутствие сконструированного локуса придает трансгенной мыши способность экспрессировать цепи антитела с вариабельными областями человека.
2. Трансгенная мышь по п. 1, где введенный локус области иммуноглобулина содержит человеческие кодирующие последовательности VH, DH и JH или кодирующие последовательности VL и JL, где L относится к лямбда- или каппа-цепям.
3. Способ получения трансгенной мыши по п. 1, где указанный способ включает:
а) введение двух или более нацеленных сайтов для одной или более сайт-специфических рекомбиназ в клетку мыши и интеграцию, по меньшей мере, одного сайта в клеточный геном выше и, по меньшей мере, одного сайта ниже геномной области, содержащей сегменты эндогенного гена вариабельной области иммуноглобулина эндогенного локуса иммуноглобулина мыши, где сегменты эндогенного гена вариабельной области иммуноглобулина содержат сегменты V-, D- и J-генов или сегменты V- и J-генов;
b) обеспечение вектора, содержащего частично человеческую вариабельную область иммуноглобулина, содержащую совокупность химерных сегментов гена иммуноглобулина, где каждый из химерных сегментов гена содержит кодирующие последовательности человека и некодирующие последовательности, основанные на локусе вариабельной области иммуноглобулина мыши, и частично человеческую область, фланкированную сайтами сайт-специфической рекомбинации, где сайты рекомбинации способны рекомбинировать с введенными сайтами в клетку мыши на стадии а);
c) введение в клетку вектора со стадии b) и сайт-специфической рекомбиназы, способной узнавать сайты рекомбиназы;
d) обеспечение рекомбинантного события между геномом клетки и частично человеческой вариабельной областью иммуноглобулина, приводящее к замене эндогенного локуса вариабельной области иммуноглобулина частично человеческим локусом вариабельной области иммуноглобулина;
e) отбор клетки, которая содержит частично человеческую иммуноглобулиновую область, полученную на стадии d); и
f) применение клетки для получения трансгенной мыши, содержащей частично человеческую область иммуноглобулина.
4. Способ по п. 3, где клетка является эмбриональной стволовой клеткой (ES-клеткой) мыши.
5. Способ по п. 4, дополнительно включающий стадию делеции участка эндогенной иммуноглобулиновой области генома введением рекомбиназы, которая узнает первую группу сайтов сайт-специфической рекомбинации, где такая делеция в геноме оставляет на месте вторую группу сайтов сайт-специфической рекомбинации, которые не способны рекомбинировать друг с другом после указанной стадии введения и до проведения указанной стадии обеспечения.
6. Способ по п. 5, где клетка является эмбриональной стволовой клеткой (ES-клеткой) мыши.
| СПОСОБ ТРАНССКЛЕРАЛЬНОЙ ГОНИОТОМИИ | 0 |
|
SU212437A1 |
| WO 2008081197 A1, 10.07.2008 | |||
| СПОСОБ ПРОИЗВОДСТВА ИНУЛИНСОДЕРЖАЩЕГО РАСТВОРА ИЗ ТОПИНАМБУРА | 2009 |
|
RU2398784C1 |
| ГУМАНИЗИРОВАННЫЙ ИММУНОГЛОБУЛИН, СПЕЦИФИЧНЫЙ ДЛЯ БЕЛКА L-СЕЛЕКТИНА ЧЕЛОВЕКА | 1993 |
|
RU2151612C1 |

