Настоящая заявка является частичным продолжением заявки США рег. N 07/983946, поданной 12/1/92, которая во всех случаях и во всей полноте вводится в настоящее описание посредством ссылки.
Изобретение относится к комбинированным технологиям рекомбинантных ДНК и моноклональных антител, используемых для разработки новых биологических материалов, а в частности, для продуцирования неиммуногенных (для человека) иммуноглобулинов, специфичных для белка L-селектина, а также к использованию указанных иммуноглобулинов in vitro и in vivo.
Способность клеток к взаимной адгезии играет важную роль в созревании, нормальной физиологии и в патологических процессах. Указанная способность опосредуется факторами адгезии, в основном, гликопротеинами, экспрессированными на клеточных мембранах. Во многих случаях, фактор адгезии на одном типе клетки связывается с другим фактором адгезии, экспрессированным на другом типе клеток, образуя пару "контррецептор-рецептор". Существует три основных класса факторов адгезии, а именно интегрины, селектины и члены суперсемейства иммуноглобулинов (Ig) (см. работы Springer, Nature 346:425 (1990); Osborn, Cell 62:3 (1880); Hynes, Cell 69:11 (1992), которые во всей своей полноте вводятся в настоящее описание посредством ссылки). Указанные молекулы имеют особенно важное значение для взаимодействия лейкоцитов и тромбоцитов друг с другом, с внеклеточным матриксом и васкулярным эндотелием.
Интегрины представляют собой гетеродимерные трансмембранные гликопротеины, состоящие из α-цепи (120-180 кДа) и β-цепи (90-110 кДа), имеющих, в основном, короткие цитоплазматические домены. Все α- субъединицы имеют гомологичную последовательность и содержат общие элементы, также как и β-субъединицы. Три известных интегрина, содержащих β-субъединицу, обозначаемую β2, играют важную роль в функции Т-клеток, нейтрофилов и моноцитов. LFA-I (αLβ2) широко распространен на лимфоцитах, гранулоцитах и моноцитах. Его контррецептор представляет собой молекулу ICAM-1 (и, вероятно, молекулу меньшего значения ICAM-2), принадлежащую к семейству Ig, которая экспрессируется на многих клетках, включая лейкоциты, и активируется на васкулярном эндотелии цитокинами, такими как TNF и IL-1. Блокирование LFA-1 на T-клетках антителами к α- или β-субъединице в значительной степени ингибирует адгезивно-зависимые функции, такие как CTL-опосредованный лизис клеток-мишеней. Mac-1 (αMβ2) располагается на нейтрофилах и моноцитах, а его контррецептором также является ICAM-1 (а возможно ICAM-2). Кроме того, Mac-1 представляет собой рецептор активированного комплемента 3 (CR3) и связывается с C3bi-фрагментом. Третий β2-интегрин, P15095 (αxβ2), также находится на нейтрофилах и моноцитах, но, по всей вероятности, он играет менее значительную роль. α-Субъединицы, LFA-1, Mac-1 и P150.95 имеют также соответствующие CD - обозначения CD11a, CD11b и CD11c, а β2 обозначается также CD18, так что LFA-1 представляет собой CD11a/CD18, а Mac-1 представляет собой CD11b/CD18.
Существует три известных селектина, которые ранее обозначались LECCAM, а в настоящее время обозначаются как L-селектин (именуемый также LECCAM-1, Меl-14 или LAM-1), E-селектин (именуемый также ELAM-1) и P-селектин (именуемый также GMP140 или PAD GEM). Все указанные селектины были секвенированы на уровне кДНК и имеют гомологичные последовательности и идентичные фрагменты, включая лектин-подобный домен. L-селектин имеет двойную функцию: с одной стороны, он является "хоминг"-рецептором на T-клетках для наружных эндотелиальных венул периферических лимфатических узлов; а с другой стороны, он является фактором адгезии на нейтрофилах для эндотелия (Hallmann et al., Biochem. Biophys. Res. Commun. 174:236 (1991); эта работа во всей своей полноте вводится в настоящее описание посредством ссылки). E-селектин и P-селектин индуцируются на эндотелии цитокинами, но с различной кинетикой. L-селектин на нейтрофилах является контррецептором E-селектина и P-селектина (Picker и др., Cell 66:921 (1991); эта работа во всей своей полноте вводится в настоящее описание посредством ссылки), хотя, возможно, что все три указанные рецепторы имеют также другие контррецепторы. В частности, E-селектин связывается с углеводной группой сиалил-Льюиса x (SLex)(Lowe et al., Cell 63:475, (1990); эта работа во всей своей полноте вводится в настоящее описание посредством ссылки), тогда как этот углевод, как известно, присутствует на L-селектине (Picker et al., Cell 66:921 (1991)) и может, очевидно, присутствовать на других белках. E-селектин экспрессируется, в основном, в воспалительных участках кожи, а также служит в качестве фактора адгезии для "хоминга" Т-клеток кожи, которые могут участвовать в воспалительных процессах (Picker et al. , Nature 349:796 (1991); эта работа вводится во всей своей полноте посредством ссылки).
Как показывают различные анализы, все антитела к CD11a, CD11b, CD18, L-селектину и E-селектину в большей или меньшей степени блокируют связывание нейтрофилов с активированными эндотелиальными клетками, но наиболее полное ингибирование достигается, в основном, комбинацией антитела к CD18 и антитела к L- или E-селектину (см., например, Luscinskas, J. Immunol., 142:2257 (1989); эта работа во всей своей полноте вводится в настоящее описание посредством ссылки). Лишь совсем недавно получила широкое признание модель, которая объясняет вышеуказанные факты трехстадийным процессом адгезии (Butcher, Cell 67:1033(1991); эта работа целиком вводится в настоящее описание посредством ссылки). В первой стадии нейтрофилы обратимо связываются с воспаленным васкулярным эндотелием посредством селектинов, которые очень хорошо связываются в условиях кровотока, заставляя нейтрофилы буквально "катиться" вдоль стенок сосудов. Затем нейтрофилы активируются рядом стимуляторов, находящихся вокруг или высвобождаемых эндотелием, включая IL-8, PAF и C5a. Активированные нейтрофилы выделяют L-селектин и активируют Mac-1. В конечной стадии связывание Mac-1 с ICAM-1 и вероятно с другими контррецепторами на клетках эндотелия способствует стабильной адгезии и транссудации через эндотелий.
В принципе, антитела или другие антагонисты таких факторов адгезии, как интегрин и селектин, могут остановить этот процесс, предотвращая связывание нейтрофилов с эндотелием и их транссудацию в ткани. Следовательно, указанные антитела могут быть использованы для лечения множества патологических состояний, в основе которых лежит воспалительный процесс.
Например, в живых моделях антител против CD 18, которые связываются с LFA-1 и Мac-1, были использованы для снижения повреждений при ишемической реперфузии (см. , например, Vedder et sl., J. Clin. Invest., 81:939 (1988); Vedder et al. , Proc. Natl. Acad. Sci. USA 87:2643 (1990); патент США N 4797277). Эти антитела также способствуют снижению нейтрофил-опосредованных повреждений в легких, обусловленных различными инсультами (Doerschuk et al., J. Immunol 144:2327 (1990) и Mulligan et al., J.Immunol. 148:1847 (1992)), включая сепсис, вызванный грамотрицательными бактериями (Walsh et al., Surgery 110:205 (1991)). В экспериментах с кроликами антитела против CD 18 также защищали от летального исхода, обусловленного менингитом (Tuomanen et al. , J. Exp. Med., 170:959 (1990)). Эти антитела могут быть также использованы для предупреждения или лечения отторжения трансплантата, поскольку они блокируют функцию T-клеток.
Например, инъекция антител против L-селектина или E-селектина грызунам способствует подавлению аккумуляции нейтрофилов внутри воспаленных очагов брюшины (Jutila et al., J. Immunol., 143:3318 (1989) и Milligan et al., J. Clin. Invest. 88:1396 (1991)). С помощью видеомикроскопа для витальных исследований было обнаружено, что антитело против L-селектина в высокой степени ингибирует "прокатывание" лейкоцитов вдоль эндотелия сосудистой стенки мезентериальных венул кролика, временно выведенных на поверхность тела (Von Andrian et al., Proc. Natl. Acad. Sci. USA. 88:7538 (1991)). Антитело против E-селектина способствует значительному снижению васкулярных нарушений, индуцированных отложением иммунных комплексов в коже или легких крыс, а также значительному снижению аккумуляции нейтрофилов на этих участках (Mulligen et al. , J. Clin. Invest. 88:1396 (1991)). Кроме того, в модели приобретенной бронхиальной астмы у приматов антитело против E-селектина способствует значительному снижению притока нейтрофилов в легкие, и ассоциированную с ним позднюю обструкцию дыхательных путей после ингаляции антигена (Gundel et al. , J. Clin. Invest. 88:1407 (1991)).
Было получено несколько антител, включая мышиное DREG-55, мышиное DREG-56 и мышиное DREG-200, которые связываются с L-селектином человека (Kishimoto et al. , Proc. Natl.Acad. Sci. USA 87:2244 (1990); эта работа целиком вводится в настоящее описание посредством ссылки). Эти антитела частично или полностью блокируют связывание лимфоцитов человека с наружными эндотелиальными венулами периферических лимфатических узлов, а также связывание нейтрофилов человека со стимулированными клетками пупочной вены человека (Kishimoto et al., Blood 78:805, (1991); эта работа целиком вводится в настоящее описание посредством ссылки). Способность указанных антител блокировать связывание нейтрофилов с эндотелиальными клетками свидетельствует о том, что антиген, с которым они связываются, а именно L-селектин, может быть подходящей мишенью для потенциальных терапевтических агентов.
К сожалению, использование моноклональных антител, не являющихся человеческими, например, таких как мышиное антитело DREG-200, для лечения человека имеет некоторые недостатки, особенно в режимах повторной терапии, как поясняется ниже. Например, мышиные моноклональные антитела имеют относительно короткое время полужизни в кровотоке, а также не обладают другими важными иммуноглобулиновыми функциональными свойствами, необходимыми для использования в лечении человека.
И вероятно, что более важно, нечеловеческие моноклональные антитела содержат значительные фрагменты аминокислотных последовательностей, которые могут быть иммуногенными при их введении человеку. Многочисленные исследования показали, что после инъецирования пациенту чужеродного антитела иммунный ответ, индуцированный в организме пациента против этого антитела, может быть настолько сильным, что сведет на нет терапевтический эффект антитела уже в самом начале лечения. Более того, поскольку в настоящее время постоянно разрабатываются новые мышиные или другие антигенные (по отношению к человеку) моноклональные антитела для лечения самых различных заболеваний, то может оказаться, что после первого или нескольких введений человеку чужеродных антител последующее лечение даже с использованием совершенно другой терапии будет неэффективным или опасным из-за перекрестной реактивности. Определенный успех был получен при продуцировании так называемых "химерных антител" (в которых, например, мышиные вариабельные области соединены с человеческими константными областями), но несмотря на это, проблема, связанная со значительной иммуногенностью, все же не была решена.
Для решения проблемы иммуногенности чужеродных антител было предпринято несколько попыток получения "очеловеченных" (гуманизированных) антител. Переход от мышиного к "очеловеченному" антителу предусматривает определенный компромисс в отношении некоторых конкурирующих факторов, и выбор того или иного решения зависит от конкретно используемых антител. Для минимизации иммуногенности в иммуноглобулине должна сохраниться по возможности большая часть человеческой последовательности акцептора. Однако, для сохранения аутентичной способности к связыванию иммуноглобулиновый остов должен содержать достаточное количество замещений в последовательности акцептора человека в целях обеспечения трехмерной конформации CDR-областей, по возможности более близкой к конформации мышиного донорного иммуноглобулина. В результате этого многие "очеловеченные" антитела, продуцированные до настоящего времени, обнаруживают значительную потерю аффинности связывания по сравнению с соответствующими мышиными антителами. См., например, Jones et al., Nature 321: 522-525 (1986); Shearman et al., J. Immunol. 147:4366-4373 (1991); Kettleborough Protein Engineering 4: 773-783 (1991); Gorman et al., Proc. Natl. Acad. Sci. USA 88:4181-4185 (1991); Tempest et al., Boitechnology 9:266-271 (1991); Riechmann et al., Nature 332:323 (1988) и публикация EPO N 0239400 (каждая из указанных работ вводится во всей своей полноте посредством ссылки).
В соответствии с вышеуказанным очевидно, что необходимо разработать улучшенные формы очеловеченных иммуноглобулинов, специфичных к антигену L-селектину, которые являются не только, в основном, неиммуногенными для человека, но и могли бы быть легко и без больших экономических затрат продуцированы способом, приемлемым для изготовления лекарственных средств, и для использования в других целях. Эти и другие проблемы были решены с помощью настоящего изобретения.
Изобретение относится к новым композициям, которые могут быть использованы, например, для лечения воспалительных заболеваний у человека и которые содержат "очеловеченные" иммуноглобулины, способные к специфическому связыванию L-селектином. Указанные иммуноглобулины могут обладать двумя парами комплексов "легкая цепь/тяжелая цепь", где по крайней мере одна цепь содержит одну или несколько гипервариабельных областей, функционально соединенных с сегментами каркасных областей иммуноглобулина человека. Например, мышиные гипервариабельные области с дополнительными натуральными аминокислотными остатками мыши или без них могут быть введены в каркасные области иммуноглобулина человека в целях продуцирования "очеловеченных" иммуноглобулинов, способных к связыванию с L-селектином при уровнях аффинности, составляющих более чем 107 М-1. Указанные "очеловеченные" иммуноглобулины также обладают способностью блокировать связывание CD R-донорного мышиного моноклонального антитела с L-селектином.
Иммуноглобулины настоящего изобретения, включая связывающие фрагменты и другие их производные, могут быть легко продуцированы с использованием техники рекомбинантных ДНК, где конечную экспрессию в трансфецированных клетках осуществляют предпочтительно в иммортализованных эукариотических клетках, таких как клетки миеломы или гибридомы. Полинуклеотиды, содержащие первую последовательность, кодирующую каркасные области "очеловеченного" иммуноглобулина, и вторую серию последовательностей, кодирующих нужные гипервариабельные области иммуноглобулина, могут быть продуцированы синтетически либо путем объединения соответствующих кДНК и геномных ДНК-сегментов.
"Очеловеченные" иммуноглобулины могут быть использованы отдельно, т.е., в основном, в чистом виде либо в сочетании с химиотерапевтическим средством, таким как нестероидное противовоспалительное средство (например, аспирин); кортикостероид или иммунодепрессант. Все указанные соединения особенно эффективны при лечении воспалительных заболеваний. "Очеловеченные" иммуноглобулины или их комплексы могут быть получены в виде фармацевтически приемлемых лекарственных препаратов, форма которых может варьироваться в зависимости от способа введения.
На фиг.1 представлены последовательности кДНК и транслированные аминокислоты последовательности вариабельных областей легкой цепи (A) и тяжелой цепи (B) мышиного антитела DREG-200. Зрелая тяжелая цепь начинается с аминокислоты 20 E, а зрелая легкая цепь начинается с аминокислоты 21 D, которым предшествуют сигнальные последовательности.
На фиг. 2 - аминокислотные последовательности вариабельных областей зрелой легкой цепи (A) и тяжелой цепи (B) мышиного антитела DREG-200 (верхние строки) и "очеловеченного" антитела DREG-200 (нижние строки). Три CDR в каждой цепи подчеркнуты. Аминокислотные остатки в каркасных областях, которые были заменены мышиными аминокислотами, или типичные аминокислоты человека в "очеловеченном" антителе подчеркнуты двойной чертой.
На фиг. 3 - нуклеотидные последовательности генов, кодирующих вариабельные области легкой цепи (A) и тяжелой цепи (B) "очеловеченного" антитела DREG-200 и начинающихся, и кончающихся XbaI-сайтами; и транслированные аминокислотные последовательности, включая сигнальные последовательности.
На фиг. 4 - конкурентное связывание мышиного и "очеловеченного IgG1 и IqG4 DREG-200. Клетками-мишенями являются клетки 2-1, линия мышиных пре-B-клеток, которые были трансфецированы геном человеческого L-селектина, а поэтому экспрессировали человеческий L-селектин (Berg et al., Biochem. Biophys. Res. Comm. 184: 1048 (1992)). 5•105 клеток инкубировали с 3 нг 1251-меченного мышиного антитела (2 мкКи/мкг) вместе с возрастающими количествами мышиного или "очеловеченного" антитела-конкурента в 0,2 мл буфера для связывания (PBS + 2%FBs +0,1% азида) в течение 1 часа при 4oC. Клетки промывали и осаждали, после чего измеряли их связанную радиоактивность. Затем вычисляли концентрации связанного и свободного меченного антитела.
На фиг. 5 - связывание человеческих нейтрофилов с IL-I-стимулированными эндотелиальными клетками пупочной вены человека (HU V EC). Нейтрофилы сначала обрабатывали иррелевантным контрольным антителом, мышиным антителом DREG-200 или очеловеченным антителом IgG1 DREG-200, или вообще не обрабатывали антителом (слева).
На фиг. 6 - защита ишемическо-реперфузированной ткани сердца с помощью "очеловеченного" антитела DREG-200. На рис. проиллюстрирован эксперимент, проведенный на кошках, обработанных "очеловеченным" антителом DREG-200 или контрольным антителом; слева направо: область под угрозой/полная вентрикулярная область; область некротической ткани/область под угрозой и область некротической ткани/полная область левого желудочка. Величины в скобках означают +/- среднеквадратичное отклонение для шести кошек; высота отрезков означает среднее значение.
Определения
Термин "в основном идентичный" или "в основном гомологичный" означает, что две пептидные последовательности при оптимальном сравнении их первичной структуры, например, с помощью программ GAP или BESTFIT с использованием весов предполагаемых брешей имеют идентичность по крайней мере 65%, а предпочтительно, по крайней мере 80-90%, а более предпочтительно, по крайней мере 95% или более (например, 99%). При этом, предпочтительно, если положения остатков, которые не являются идентичными, отличаются друг от друга консервативными аминокислотными замещениями.
Для классификации консервативных или неконсервативных аминокислотных замещений аминокислоты могут быть разделены на следующие группы: Группа I (гидрофобные боковые цепи): норлейцин, met, ala, val, leu, ile; Группа II (нейтральные гидрофильные боковые цепи): сys, ser, thr; Группа III (кислотные боковые цепи): asp, glu; Группа IV (основные боковые цепи): asn, gln, his, lys, arg; Группа V (остатки, влияющие на ориентацию цепи): gly, pro и Группа VI (ароматические боковые цепи): trp, tyr, phe. Консервативными замещениями являются замещения между аминокислотами одного и того же класса. Неконсервативные замещения представляют собой замену аминокислоты одного класса на аминокислоту другого класса.
Аминокислоты из вариабельных областей зрелых тяжелой и легкой цепей иммуноглобулинов обозначаются Hx и Lx соответственно, где x представляет собой число, обозначающее положение аминокислот в соответствии со схемой Kobat в "Sequences of Proteins of Immunological Interest (National Institutes of Health, Bethesda, MD, 1987 и 1991). В этой работе Kabat перечисляется много аминокислотных последовательностей антител для каждого подкласса, а также перечисляются наиболее часто встречающиеся аминокислоты для каждого положения остатка в данном подклассе. Kabat использовал метод, в котором каждой аминокислоте в перечисленных последовательностях приписывался номер, и этот метод нумерации аминокислотных остатков был принят специалистами за стандарт. Схема Kabat может быть использована и в отношении других антител, не включенных в список Kabat, путем сравнения первичной структуры исследуемого антитела с одной из консенсусных последовательностей в списке Kabat. Использование системы, нумерации Kabat, позволяет легко идентифицировать аминокислоты в эквивалентных положениях различных антител. Например, аминокислота в положении L 50 человеческого антитела занимает эквивалентное положение по отношению к положению аминокислоты L 50 мышиного антитела.
Начиная от N-конца и кончая C-концом, легкая и тяжелая цепи включают в себя домены FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4. Приписывание аминокислот к каждому домену проводится в соответствии с определениями Kabat (1987) и (1991), (см. выше) или Chothia & Lesk, J. Mol. Biol. 196:901-917 (1987); Chothia et al., Nature 342:878-883 (1989).
Подробное описание изобретения
В основном, классификация, используемая ниже, а также при описании лабораторных процедур; методы молекулярной генетики, химии нуклеиновых кислот и гибридизации, описанные ниже, являются хорошо известными и широко используются специалистами в этой области. В методах рекомбинантных ДНК, синтеза полинуклеотидов, культивировании клеток и трансгенного введения генетического материала (например, электропорация, микроинъекции, липофекция) используется традиционная техника. В основном, ферментные реакции, синтез олигонуклеотидов и стадии очистки осуществляют в соответствии с инструкциями производителей. Способы и процедуры, в основном, осуществляют в соответствии со стандартной техникой, обычно применяемой в данной области, а также в соответствии с описаниями, имеющимися в литературе, ссылки на которую приводятся в настоящей заявке. Для удобства читателя в настоящей заявке также приводится описание хорошо известных процедур. Вся нужная информация вводится в настоящее описание посредством ссылки.
"Очеловеченные" антитела против L-селектина
Настоящее изобретение относится к получению "очеловеченных" иммуноглобулинов, специфически связывающихся с эпитопами L-селектина. Указанные иммуноглобулины, в основном, имеют высокую аффинность связывания с L-селектином, составляющую по крайней мере около 107 М-1, а предпочтительно от 108 М-1 до 109 М-1 или более, и обладают способностью связываться, например, с нейтрофилами. "Очеловеченные" иммуноглобулины имеют каркас человеческого иммуноглобулина, и одну или несколько областей, определяющих комплементарность (CDR-Complementarity determining regions), происходящих от иммуноглобулина, обычно мышиного иммуноглобулина, способного к специфической реакции с L-селектином. В предпочтительном варианте настоящего изобретения одна или несколько CDR-областей происходят от мышиного антитела DREG-200, а "очеловеченный" иммуноглобулин, в целом, имеет изотип IgG1 или IqG4. Таким образом, иммуноглобулины настоящего изобретения, которые могут быть получены в больших количествах экономически эффективным способом, предназначены для использования в лечении воспалительных заболеваний у человека с применением различной техники.
Известно, что основная структурная единица антитела состоит из тетрамера. Каждый тетрамер состоит из двух идентичных пар полипептидных цепей, при этом пара имеет одну "легкую" (около 25 кДа) и одну "тяжелую" цепь (около 50-70 кДа). NG2-конец каждой цепи начинается с вариабельной области, состоящей приблизительно из 100-110 или более аминокислот, которые ответственны, главным образом, за узнавание антигена. COOH-часть каждой цепи определяет константная область, которая ответственна, главным образом, за эффекторную функцию.
Легкие цепи подразделяются на каппа- и лямбда-цепи. Тяжелые цепи классифицируются как гамма-, мю-, альфа-, дельта- или эпсилон-цепи, которые определяют изотип антитела, а именно IgG, IgM, IgA, IgD и IgE, соответственно. В легких и тяжелых цепях вариабельные и константные области соединены "J"-областью, состоящей примерно из 12 аминокислот или более; причем в тяжелой цепи, кроме того, имеется область "D", состоящая примерно из 10 или более аминокислот (См. например, Fundamental Immunology, Paul, W., Ed., Chapter 7 pp. 131-166, Raven Press, N.Y. (1984); эта работа целиком вводится в настоящее описание посредством ссылки).
Вариабельные области каждой пары легкой/тяжелой цепей образуют антигенсвязывающий центр. Консервативные каркасные области всех цепей имеют, в основном, одинаковое строение и соединяются посредством трех гипервариабельных областей, называемых также участками, определяющими комплементарность (CDR) (см., например, "Sequences of Proteins of Immunological Interest", Kabat E. и др., U.S. Department of Health and Human Services (1987) и Chothia & Lesk, J. Mol. Biol., 196:901-917 (1987); эти работы целиком вводятся в настоящее описание посредством ссылки). CDR двух цепей каждой пары перемещался с каркасными участками, обладающими способностью связываться со специфическим эпитопом.
Используемый в настоящем описании термин "иммуноглобулин" относится к белку, состоящему из одного или нескольких полипептидов, в основном, кодируемых генами иммуноглобулина. Такими генами являются каппа-, лямбда-, альфа-, гамма-, дельта-, эпсилон- и мю-гены константных областей, а также множество генов вариабельной области иммуноглобулина. Иммуноглобулины могут существовать, помимо антител, в виде других форм, например, таких как FV, Fab, и (Fab')2, а также в виде гетеровалентных антител (например, Lanzavechia и др. , Eur. J. Immunol. 17:105 (1987)) и одиночных цепей (например, Huston et al. , Proc. Natl. Acad. Sci. USA 85:5879-5883 (1988) и Bird et al., Science 242: 423-426 (1988); указанные работы во всей полноте вводятся в настоящее описание посредством ссылки). (В общих чертах, см. Hood et al., Immunology (Benjamin N.Y., 2nd ed., 1984), Harlow & Lane, Antibodies: A Laboratory Manual (Cold Spring Harbor Laboratory, 1988) и Hunkapiller & Hood, Nature, 323: 15-16 (1986); все указанные работы вводятся в настоящее описание во всей своей полноте посредством ссылки).
Химерные антитела представляют собой такие антитела, у которых гены легкой и тяжелой цепей были сконструированы обычно методами генной инженерии из сегментов иммуноглобулиновых генов, принадлежащих к различным видам. Например, вариабельные (V) сегменты генов от моноклонального мышиного антитела могут быть соединены с константными (C) сегментами иммуноглобулина человека, таким как V1 и V4. Таким образом, типичное терапевтическое химерное антитело представляет собой гибридный белок, состоящий из V или антигенсвязывающего домена, происходящего от мышиного антитела, и C или эжекторного домена, происходящего от антитела человека, хотя могут быть использованы антитела и других видов млекопитающих.
Используемый в настоящем описании термин "каркасная область" относится к таким участкам вариабельных областей легкой и тяжелой цепей иммуноглобулина, которые являются относительно консервативными (т.е. менее вариабельными, чем CDR) у иммуноглобулинов одного вида (как было определено Kabat и др., см. выше). Используемый в настоящем описании термин "каркасная область человека" означает каркасную область, которая, в основном, идентична (например, на 85% или более) каркасной области натурального антитела человека или консенсусной последовательности нескольких таких антител.
Используемый в настоящем описании термин "очеловеченный иммуноглобулин" относится к иммуноглобулину, который включает в себя каркас иммуноглобулина человека; и по крайней мере одну гипервариабельную область (CDR), происходящую от антитела, не являющегося человеческим и в котором любая присутствующая константная область, в основном, идентична константной области человеческого иммуноглобулина по крайней мере примерно на 85-90%, а предпочтительно, по крайней мере на 95 %. Следовательно, все части "очеловеченного" иммуноглобулина, за исключением, возможно, CDR, являются, в основном, идентичными соответствующим частям одного или нескольких последовательностей нативного иммуноглобулина человека. Так, например, к "очеловеченному" иммуноглобулину не относится химерное антитело, имеющее мышиную вариабельную область и человеческую константную область.
По сравнению с антителами, а в некоторых случаях и с химерными антителами "очеловеченные" антитела имеют по крайней мере три потенциальных преимущества при их использовании для лечения человека, а именно:
(1) поскольку эффекторная часть "очеловеченного" антитела происходит от человеческого иммуноглобулина, то это антитело лучше взаимодействует с другими частями иммунной системы человека (например, с большей эффективностью разрушает клетки мишени благодаря комплементзависимой цитотоксичности (CDC) или антителозависимой клеточно-опосредованной цитотоксичности (ADCC)).
(2) Человеческая иммунная система не должна распознавать каркасную или C-область "очеловеченного" антитела как чужеродную, а поэтому антительный ответ организма против такого инъецированного антитела должен быть меньше, чем ответ против целиком чужеродного мышиного антитела, либо частично чужеродного химерного антитела.
(3) Сообщалось (Shaw D., et al., J. Immunol., 138:4534-4538 (1987)), что инъецированные мышиные антитела имеют более короткое время полужизни в кровотоке человека, чем нормальные антитела. Инъецированные "очеловеченные" антитела, по всей вероятности, имеют такое же время полужизни, что и натуральные антитела человека, если учесть, что эти антитела попадают в кровоток в меньших и не в столь частых дозах.
В одном из своих вариантов настоящее изобретение относится к сегментнам рекомбинантных ДНК, кодирующим CDR-области тяжелой и/или легкой цепи иммуноглобулина, способного связываться с нужным эпитопом L-селектина, например, такого как мышиные моноклональные антитела DREG-200, DREG-55 или DREG-56 (Kishimoto et al. (1990), см.выше; эта работа во всей своей полноте вводится в настоящее описание посредством ссылки). ДНК-сегменты, кодирующие указанные области, сшивают с ДНК-сегментами, кодирующими соответствующие каркасные области, происходящие от иммуноглобулина человека. Характерные ДНК-последовательности, которые после экспрессии кодируют полипептидные цепи, содержащие CDR-области тяжелой и легкой цепей моноклонального мышиного антитела DREG-200, показаны на фиг. 1. Благодаря вырождению кодона и некритическим аминокислотным замещениям, эти последовательности могут быть легко заменены другими ДНК-последовательностями, как, например, описано ниже. Подробное описание конструирования и продуцирования "очеловеченных" иммуноглобулинов приводится в общих чертах в переуступленных заявках per. N 07/290975 и 07/310252, поданных 28 декабря 1988 и 13 февраля 1989 соответственно, которые во всей своей полноте вводятся в настоящее описание посредством ссылки.
Кроме того, ДНК-сегменты могут содержать ДНК-последовательность, регулирующую экспрессию и соответствующим образом присоединенную к последовательностям, кодирующим "очеловеченный" иммуноглобулин, например, натуральные или гетерологичные промоторные области. Предпочтительными регуляторными последовательностями являются эукариотические промоторные системы, присутствующие в векторах, способных к трансформации или трансфекции эукариотических клеток-хозяев, хотя могут быть также использованы и регуляторные последовательности для прокариотических клеток. После введения вектора в соответствующую клетку-хозяина эту клетку выдерживают в условиях, подходящих для высокого уровня экспрессии нуклеотидных последовательностей, а затем, если это необходимо, осуществляют сбор и очистку легких цепей, тяжелых цепей, димеров легких/тяжелых цепей или интактных антител, связывающихся фрагментов или других форм иммуноглобулинов.
Таким образом, нуклеиновокислотные последовательности настоящего изобретения, обладающие способностью к продуцированию нужных "очеловеченных" антител, могут быть сконструированы из различных полинуклеотидов (таких, как геномная или кДНК, РНК, синтетические олигонуклеотиды и т.п.) и компонентов (таких, как V-, J-, D- и C-области) с использованием различной техники. Наиболее распространенным методом такого конструирования является соединение соответствующих геномных и синтетических последовательностей, хотя могут быть также использованы и кДНК-последовательности (см.публикацию Европатента N 0239400 и Riechmann, L. et al., Nature 332: 323-327 (1988); обе эти работы во всей своей полноте вводятся в настоящее описание посредством ссылки).
ДНК-последовательности константных областей человека могут быть выделены с использованием известных процедур из различных клеток человека, а предпочтительно изиммортализованных B-клеток (Kabat см.выше, и WP87/02671). CDR, используемые для получения иммуноглобулинов настоящего изобретения, могут быть получены от моноклональных антител, способных связываться с L-селектином, и продуцированы в любом подходящем источнике, взятом от млекопитающего, например, такого, как мышь, крыса, кролик или другое позвоночное животное, способное к продуцированию антител, с использованием методов, хорошо известных специалистам. Подходящие клетки-источники для ДНК-последовательностей, а также клетки-хозяева для экспрессии и секреции иммуноглобулинов могут быть получены из ряда источников, например, таких как Американская коллекция типовых культур (Catalogue of Cell Lines and Hybridomas, 5-th ed. (1985) Rockville, MD; эта работа во всей своей полноте вводится в настоящее описание посредством ссылки). В предпочтительных вариантах настоящего изобретения CDR-последовательности соответствуют CDR-последовательностям антитела DREG-200, антитела мыши DREG-55 или антитела DREG-56 и могут содержать вырожденные нуклеотидные последовательности, кодирующие соответствующие аминокислотные последовательности CDR мышиного антитела DREG-200, DREG-55 или DREG-56.
Помимо "очеловеченных" иммуноглобулинов, подробно описанных в настоящей заявке, могут быть легко сконструированы и продуцированы другие "в основном, гомологичные" модифицированные иммуноглобулины с использованием хорошо известной техники рекомбинантных ДНК. В качестве источника каркасной последовательности могут быть использованы и другие антитела человека, отличающиеся от антител Eu, обсуждаемых в Примере 2. Эти каркасные последовательности должны быть в высокой степени идентичными вариабельным каркасным доменам мышиного антитела DREG-200, от которых происходят области CDR. Вариабельные каркасные области тяжелой и легкой цепей могут происходить от последовательностей одного и того же антитела либо от последовательностей разных антител человека. Так, например, каждая из каркасных областей тяжелой и легкой цепей может происходить от нескольких антител человека. Такими последовательностями могут быть последовательности природных антител человека, либо консенсусные последовательности нескольких антител человека (см. Carter и др. WO 92/22653 (1992)).
Непосредственное и неестественное соседство мышиных CDR-областей с вариабельной каркасной областью человека может приводить к конформационным затруднениям, которые, если их не скорректировать посредством замещения некоторых аминокислотных остатков, могут привести к потере аффинности связывания. Выбор аминокислотных остатков для замещения осуществляется частично с помощью компьютерного моделирования. Для этих целей широко используется компьютерная аппаратура и программное обеспечение для проецирования трехмерного изобретения молекул иммуноглобулина. В общих чертах, молекулярные модели продуцируют исходя из известных структур иммуноглобулиновых цепей или их доменов. Моделируемые цепи сравнивают с цепями или доменами известных трехмерных структур для анализа сходства их аминокислотных последовательностей и, если указанные цепи или домены обнаруживают высокую степень сходства, то их используют в качестве отправной точки для конструирования молекулярной модели. Выбранные сходные структуры модифицируют с учетом различия между тактически существующими аминокислотами в моделируемых цепях или доменах иммуноглобулина и аминокислотами, имеющимися в исходной структуре. Затем модифицированные структуры собирают с получением модели составного иммуноглобулина. И, наконец, полученную модель совершенствуют путем энергетической минимизации и контроля за тем, чтобы все атомы находились на соответствующем расстоянии друг от друга и чтобы длина связей и углы находились в пределах химически допустимых диапазонов. Стадии продуцированной трехмерной компьютерной модели для вариабельных областей мышиного антитела DREG-200 подробно обсуждаются в Примере 2. Эта модель, в свою очередь, может быть отправной точкой предсказания трехмерной структуры антитела, содержащего гипервариабельные области мышиного DREG-200, замещенные в каркасных чел. структурах. Кроме того, могут быть сконструированы модели, представляющие структуру в том случае, если вводятся дополнительные аминокислотные замещения, обсуждаемые ниже.
Вообще говоря, замещение человеческих аминокислотных остатков мышиными остатками должно быть минимизировано, так как введение мышиных остатков увеличивает риск индуцирования у человека HAMA-ответ. Аминокислоты для замещения выбирают исходя из их возможного влияния на CDR-конформацию и/или связывание с антигеном. Исследование такого возможного влияния проводят путем моделирования, оценки характеристик аминокислот в конкретных положениях или эмпирического наблюдения влияния замещения или мутагенеза конкретных аминокислот.
Если аминокислота в вариабельной каркасной области мышиного антитела DREG-200 отличается от аминокислоты в эквивалентной вариабельной каркасной области человека, то человеческая аминокислота должна быть заменена эквивалентной мышиной аминокислотой в случае, если имеются основания ожидать, что данная аминокислота:
/1/ нековалентно контактирует непосредственно с антигеном, или
/2/ находится в непосредственной близости или как-нибудь иначе взаимодействует с CDR-областью /например, находится на расстоянии примерно  от CDR-области/.
от CDR-области/.
Другими кандидатами на замещение являются аминокислоты акцепторного каркаса иммуноглобулина человека, которые несвойственны для иммуноглобулина человека в этом положении /например, аминокислота H113 человеческого антитела Eu/. Эти аминокислоты могут быть замещены аминокислотами из эквивалентного полонения более типичных иммуноглобулинов человека. Альтернативно, аминокислоты из эквивалентных положений мышиного антитела DREG-200 могут быть введены в каркасные области человека, если такие аминокислоты являются типичными для иммуноглобулина человека в эквивалентном положении.
Вообще говоря, замещение всех или большинства аминокислот, удовлетворяющее вышеуказанным критериям, является желательным. Однако, иногда, имеется некоторая неопределенность относительно того, какая именно аминокислота отвечает вышеуказанному критерию, и в этом случае продуцируют альтернативный вариант иммуноглобулинов, один из которых содержит данное замещение, а другой - нет. "Очеловеченные" антитела настоящего изобретения обычно имеют замещение в каркасной области легкой цепи соответствующим остатком мышиного антитела DREG-200 по крайней мере в 1, 2, 3, 4 и обычно 5 положениях, выбранных из следующих положений: L87, L54, L66, L76 и L93. "Очеловеченные" антитела также обычно содержат замещение остатком каркасной области мышиной тяжелой цепи по крайней мере в 1, 3, 5, 7, 9, 10, 11, а обычно 12 положениях, выбранных из следующих положений: H93, H95, H98, H111, H112, H115, H30, H98, H111, H27, H48 и H72. В предпочтительном варианте настоящего изобретения, если акцепторным иммуноглобулином с тяжелой цепью человека является антитело Eu, то тяжелая цепь также имеет замещение в H113. Это положение обычно замещается аминокислотой из эквивалентного положения иммуноглобулина человека, имеющего более типичные аминокислотные остатки.
Обычно, CDR-области в "очеловеченных" антителах являются, в основном и предпочтительно, идентичными соответствующим CDR-областям мышиного антитела DREG-200. Однако, в некоторых случаях может оказаться предпочтительным заменить один остаток в CDR-области, например, для создания сходства с местом связывания лиганда L-селектина. Иногда можно, хотя и нежелательно, сделать одно или несколько консервативных аминокислотных замещений CDR-остатков, если это, конечно, заметно не повлияет на аффинность связывания полученного "очеловеченного" иммуноглобулина.
За исключением конкретных аминокислотных замещений, обсуждаемых выше, каркасные области "очеловеченных" иммуноглобулинов, в основном и в большинстве случаев, идентичны каркасным областям антител человека, от которых они происходят. Однако, в некоторых вариантах настоящего изобретения каркасные области могут отличаться от нативных последовательностей по своей первичной структуре, например, несколькими аминокислотными замещениями, концевыми или промежуточными вставками и делециями, и т.п. Подходящими компонентами для полипептидов настоящего изобретения могут также служить стереоизомеры /например, D-аминокислоты/ двадцати главных аминокислот, ненатуральные аминокислоты, такие как α,α-дизамещенные аминокислоты, N-алкил-аминокислоты, молочная кислота и другие второстепенные аминокислоты. Очевидно, что многие из аминокислот каркасной области могут вносить свой небольшой или косвенный вклад в специфичность или аффинность антитела. Поэтому многие отдельные консервативные замещения остатков каркасной области являются вполне допустимыми, если только они не оказывают неблагоприятного воздействия на специфичность или аффинность полученного "очеловеченного" иммуноглобулина. Однако, в основном, такие замещения являются нежелательными. Модификации генов могут быть легко осуществлены с использованием хорошо известной техники, такой как сайт-специфический мутагенез /см. Gillman & Smith, Gene 8:81-97 /1989/ и Robers et al. , Nature 328:731-734 /1987/; обе из указанных работ во всей своей полноте вводятся в настоящее описание посредством ссылки/.
Альтернативно могут быть продуцированы полипептидные фрагменты, содержащие лишь часть первичной структуры антитела и обладающие одной или несколькими активностями иммуноглобулина /например, активностью связывания/. Эти полипептидные фрагменты могут быть продуцированы путем протеолитического расщепления интактных антител известными методами либо путем инсерции стоп-кодонов в нужных положениях в векторы pVk и pVg1- dhfr с использованием сайт-специфического мутагенеза, например, так, чтобы после CH1 продуцировались Fab-фрагменты или после шарнирной области продуцировались /Fab'/2-фрагменты. Одноцепочечные антитела могут быть продуцированы путем соединения VL и VH с ДНК-линкером /см. Huston et al., см. выше, и Bird et al. , см.выше/. Например, FV - или Fab-фрагменты могут быть продуцированы в E.coli в соответствии с методами, описанными Buchner & Rudolph, Bio/Technology 9: 157-162 /1991/ и Skerra et al., Bid Technology 9:273-277 /1991/ /эти работы во всей своей полноте вводятся в настоящее описание посредством ссылки/. FV- и Fab-фрагменты могут быть также продуцированы путем экспрессии кодирующих полинуклеотидов в эукариотических клетках, предпочтительно в клетках млекопитающего. Кроме того, поскольку, подобно многим генам, гены, ассоциированные с иммуноглобулинами, содержат отдельные функциональные области, каждая из которых обладает одной или несколькими характерными биологическими активностями, то эти гены могут быть лигированы с функциональными областями от других генов /например, ферментов, см. переуступленную заявку США рег. N 132387, поданную 15 дек.1987, которая вводится во всей полноте в настоящее описание посредством ссылки/, в результате чего могут быть получены гибридные белки /например, иммунотоксины/, обладающие новыми свойствами.
Экспрессия последовательностей "очеловеченного" иммуноглобулина в бактериальных хозяевах может быть использована для благоприятного выбора последовательностей "очеловеченного" иммуноглобулина с более высокой аффинностью посредством мутагенеза CDR-областей и продуцирования фаговых дисплейных библиотек, которые могут быть скринированы на наличие CDR-вариантов "очеловеченного" иммуноглобулина, обладающих высокой аффинностью и/или высокой специфичностью связывания с L-селектином. Одним из потенциальных преимуществ такого усовершенствованного подбора аффинности является генерирование CDR-вариантов "очеловеченного" иммуноглобулина, имеющих более высокую аффинность связывания и/или более низкую перекрестную реактивность по отношению к молекулам, не являющимся L-селектинами. Методы продуцирования фаговых дисплейных библиотек, содержащих последовательности вариабельных областей иммуноглобулина, известны специалистам /см., например, Cesareni, FEBS Lett 307: 66-70 /1992/; Swimmer et al., Proc. Natl. Acad. Sci. USA 89:3756-60 /1992/; Gram et al., Proc. Natl. Acad. Sci. USA 89-3576-80 /1992/;
Clackson et al. , Nature 352:624-8 /1991/; Scott & Smith, Science 249: 386-90 /1990/; Garrard et al., Bid Techniques 9:1373-1377 /1991/; все указанные работы во всей своей полноте вводятся в настоящее описание посредством ссылки/. Полученные в результате последовательности CDR-вариантов "очеловеченного" иммуноглобулина затем экспрессируют в подходящем хозяине, обеспечивающем эффективную экспрессию.
Как указывалось ранее, ДНК-последовательности могут быть экспрессированы в соответствующих клетках-хозяевах после того, как эти последовательности будут надлежащим образом присоединены /т.е. таким образом, чтобы обеспечивалось их функционирование/ к последовательности, регулирующей экспрессию. Для этих целей могут быть использованы экспрессирующие векторы, которые способны к репликации в хозяйских организмах либо как эписомы, либо как интегральная часть хромосомной ДНК хозяина. Обычно векторы экспрессии содержат селективные маркеры, например, резистентность к тетрациклину /tetR/, в G418-резистентность /neoR/, резистентность к микофенольной кислоте /gpt/, HSV-tk, которые позволяют обнаруживать клетки, трансформированные нужными ДНК-последовательностями /см. например, патент США 4704362, который полностью вводится в настоящее описание посредством ссылки/.
Одним из прокариотических хозяев, конкретно используемых для клонирования ДНК-последовательностей настоящего изобретения, является E.coli. Другими подходящими микробными хозяевами являются бациллы, такие как Bacillus Subtilis; другие энтеробактерии, такие как Salmonella, Serratia, и различные виды Pseudomonas. В этих прокариотических хозяевах могут быть также экспрессированы векторы, которые обычно содержат регуляторные последовательности, совместимые с данной клеткой-хозяином /например, сайт-инициации репликации/. Кроме того, могут также присутствовать любые другие хорошо известные промоторы, такие как промоторная система лактозы, промоторная система триптофана /trp/, промоторная система бета-лактамазы или промоторная система фага-лямбда. Эти промоторы обычно контролируют экспрессию, необязательно, посредством операторной последовательности и имеют последовательности сайта связывания с рибосомой и т.п. для инициации и терминации транскрипции и трансляции.
Для экспрессии могут быть также использованы и другие микробы, такие как дрожжи. Предпочтительным хозяином является Saccharomyces, а подходящими векторами являются векторы, имеющие последовательности, контролирующие экспрессию, такие как промоторы, например, промоторы 3-фосфоглицераткиназы или других гликолитических ферментов, а также сайт инициации репликации, терминирующие последовательности и т.п., если это необходимо.
Для экспрессии "очеловеченного" иммуноглобулина настоящего изобретения могут быть использованы растения и культуры растительных клеток /Larrick & Fry, Hum. Antibodies Hybridomas 2/4/:172-89 /1991/; Benvenuto et al., Plant Mol. Biol. 17/4/:865-74 /1991/; Durin et al., Plant Mol. Biol. 15/2/:281-93 /1990/; Hiatt et al., Nature 342:76-8 /1989/; все указанные работы вводятся в настоящее описание посредством ссылки/. Предпочтительными растительными хозяевами являются, например, Arabidopsis, Nicotiana tabacum, Nicotiana rustica и Solanum tuberosum. Предпочтительным полигенным экспрессирующим кластером, подходящим для экспрессии полинуклеотидных последовательностей, кодирующих "очеловеченные" антитела настоящего изобретения, направленные против L-селектина, является плазмида pMOG18, в которой инсертированная полинуклеотидная последовательность, кодирующая цепь "очеловеченного" иммуноглобулина, является соответственно сшитой с CAMV35S-промотором, связанным с дуплицированным энхансером; причем указанная плазмида используется в соответствии с методом, описанным Sijmons et al., Bio/Technology, 8:217-221 /1990/; эта работа вводится в настоящее описание посредством ссылки/. Альтернативно, экспрессию "очеловеченных иммуноглобулинов в растениях осуществляют предпочтительно по методу Hiatt и др. /см.выше/, за исключением того, что иммуноглобулиновые последовательности, используемые Hiatt и др. /см. выше/, заменяют полинуклеотидными последовательностями, кодирующими "очеловеченные" антитела настоящего изобретения против L-селектина. Для экспрессии последовательностей "очеловеченного" иммуноглобулина могут быть также использованы векторы на основе Т-ДНК Agrobacterium tumifaciens, а предпочтительно такие векторы, которые содержат маркерный ген, кодирующий резистентность к спектиномицину, или другой маркерный ген.
Для продуцирования "очеловеченных" иммуноглобулинов настоящего изобретения могут быть также использованы культуры клеток насекомых, но, в основном, используется экспрессирующая система на основе бакуловируса. "Очеловеченные" иммуноглобулины могут быть продуцированы путем экспрессии полинуклеотидных последовательностей, кодирующих "очеловеченные" иммуноглобулины, с использованием методики Putlitz и др. /Bio/Technology, 8:651-654 /1990/, эта работа полностью вводится в настоящее описание посредством ссылки/. Метод Putlitz и др. может быть модифицирован, например, вместо кДНК-последовательностей тяжелой и легкой цепи мышиного моноклонального антитела Ab6A4, используемых Putlitz и др. , могут быть введены полинуклеотидные последовательности, кодирующие "очеловеченные" антитела против L-селектина.
Для экспрессии и продуцирования полипептидов настоящего изобретения, помимо микроорганизмов и растений, могут быть использованы клеточные культуры млекопитающих /см. Winnacker, From Genes to Clones (VCH Publisher, NY, 1987; эта работа полностью вводится в настоящее описание посредством ссылки/. При этом, предпочтительными клетки млекопитающих, поскольку специалистами получены подходящие линии клеток-хозяев, способные секретировать интактные иммуноглобулины, например линии клеток CHO, различные линии клеток COS, HEIa-клетки, предпочтительно клеточные линии миеломы, и т.п. либо трансформированные B-клетки или гибридомы. Экспрессирующие векторы, используемые для этих клеток, могут содержать регуляторные последовательности, такие как сайт инициации репликации, промотор, энхансер /Queen и др., Immunol. Rev. 89:49-68 /1986/; эта работа полностью вводится в настоящее описание посредством ссылки/, а также необходимые сайты, регулирующие процессинг, например, такие как сайты связывания с рибосомой, сайты сплайсинга РНК, сайты полиаденилирования, и последовательности терминации транскрипции. Предпочтительными регуляторными последовательностями являются промоторы, происходящие от генов иммуноглобулина, SV 40, аденовируса, бычьего вируса папилломы, цитомегаловируса и т.п. Обычно экспрессирующий вектор содержит маркерный ген, такой как neoR.
Трансгены, кодирующие "очеловеченный" иммуноглобулин настоящего изобретения, могут быть использованы для генерирования трансгенных животных, не относящихся к человеку, которые экспрессируют "очеловеченный" иммуноглобулин, обычно, в выделяемых жидкостях организма, таких как молоко или сыворотка. Указанные трансгены содержат полинуклеотидную последовательность, кодирующую "очеловеченные" иммуноглобулины и надлежащим образом соединенную с промотором, обычно с присоединенным энхансером, таким как иммуноглобулин грызунов или промотор/энхансер казеинового гена /Buhler et al., Bio/Technology 8: 140-143, /1990/; Meade et al., Bio/Technology 8:443-446 /1990/; эти работы вводятся целиком в настоящее описание посредством ссылки/. Трансгены могут быть введены в клетки и эмбрионы в соответствии с известной методикой, в результате чего /см.ниже/ могут быть получены гомологичные рекомбинантные конструкции. Предпочтительными животными, не относящимися к человеку, являются мыши, крысы, овцы, коровы и козы; при этом предпочтительно, если для экспрессии используется коровье молоко. См. WO 91/08216 /1991/ /эта работа вводится в настоящее описание посредством ссылки/. Очистку "очеловеченных" антител настоящего изобретения осуществляют в соответствии с известными методами очистки, обычно применяемыми для очистки иммуноглобулинов.
Векторы, содержащие ДНК-сегменты /например, последовательности, кодирующие тяжелую и легкую цепь, и регуляторные последовательности/, могут быть перенесены в клетки-хозяева известными методами, выбор которых зависит от типа клетки-хозяина. Например, для прокариотических клеток обычно применяют трансфекцию с использованием хлорида кальция, тогда как для других клеток-хозяев могут быть использованы такие методы, как обработка фосфатом кальция, липофекция, биобаллистика, трансдукция с помощью вируса или электропорация. Для клеток растений и тканей предпочтительно использовать метод баллистического трансгенеза с помощью вольфрамовых частиц /В общих чертах см. Maniatis et al. , Molecular Cloning: A Laboratory Manual /Cold Spring Harbor Press, 1982/; эта работа вводится в настоящее описание посредством ссылки/.
После экспрессии интактные антитела, их димеры, отдельные легкие и тяжелые цепи или другие формы иммуноглобулина настоящего изобретения могут быть очищены в соответствии со стандартными процедурами, такими как, например, осаждение сульфатом аммония, аффинная хроматография, колоночная хроматография, гель-электрофорез/в общих чертах, см. Scopes, R., Protein Purificatin (Springer-Verlag, NY, 1982; эта работа целиком вводится в настоящее описание посредством ссылки/. В основном, очищенные иммуноглобулины, имеющие, предпочтительно, гомогенность по крайней мере около 90-95%, а наиболее предпочтительно 98-99%, могут быть использованы в фармацевтических целях. Затем очищенные /частично или до гомогенности/ полипептиды могут быть использованы в терапевтических целях/например, экстракорпорально/ либо в исследовательских и аналитических процедурах, в иммунофлуоресцентном окрашивании и т. п. /в общих чертах, см. Immunological Methods, vols. I и II /Lefkovits & Pernis, eds. Academic Press, NY, 1979 and 1981/.
В предпочтительном варианте настоящего изобретения продуцируются "очеловеченные" иммуноглобулины, которые связываются с L-селектином с аффинностью связывания, составляющей по крайней мере 1•107 М-1 при стандартных условиях связывания /например, фосфатно-буферный физиологический раствор, содержащий 2% плодной телячьей сыворотки; при 25oC/. Одним из примеров таких "очеловеченных" иммуноглобулинов является "очеловеченное" антитело DREG-200, имеющее аминокислотную последовательность, показанную на фиг. 2. /Далее, в некоторых случаях, "очеловеченное" антитело DREG-200 обозначаться "чел. DREG-200"/. Очеловеченные иммуноглобулины, содержащие области CDR, происходящие от мышиного антитела DREG-55 или от мышиного антитела DREG-56, также могут связываться с L-селектином с аффинностью, составляющей по крайней мере 1•107 М-1.
Очеловеченные антитела настоящего изобретения связываются (в стандартных условиях связывания) с L-селектином предпочтительно с аффинностью, составляющей по крайней мере 1• 108 M-1, более предпочтительно с аффинностью, составляющей по крайней мере 1•108 M-1; а наиболее предпочтительно с аффинностью, составляющей по крайней мере, 1•1010 М-1 или выше. Обычно, аффинность связывания "очеловеченного" иммуноглобулина примерно в три раза превышает аффинность связывания мышиного иммуноглобулина, от которого он происходит. Так, например, аффинность мышиного антитела DREG-200 составляет около 108 М-1.
Компьютеры
В другом варианте своего осуществления настоящее изобретение относится к компьютерам для построения трехмерного изображения антител на мониторе. Например, для целей настоящего изобретения могут быть использованы автоматизированная рабочая установка Silicon Graphics IRIS 4D, работающая в операционной системе UNIX, и пакет программ для молекулярного моделирования QUANTA (Polygen Corp. USA). Компьютеры могут быть использованы для генерирования вариантов "очеловеченных" антител. Вообще говоря, антитела настоящего изобретения уже обладают достаточной аффинностью связывания. Однако, очевидно, что антитела даже с более сильной аффинностью связывания могут быть идентифицированы путем последующей вариации некоторых аминокислотных остатков. С помощью трехмерного изображения можно также идентифицировать множество некритических аминокислот, которые могут быть объектами консервативных замещений, не оказывающих заметного влияния на аффинность связывания данного антитела. Однако, даже консервативные замещения, взятые вместе, могут оказывать значительное влияние на свойства иммуноглобулина. Но многие отдельные консервативные замещения, по всей вероятности, не оказывают неблагоприятного воздействия на свойства иммуноглобулинов.
Человеческие антитела против L-селектина
В другом своем варианте, настоящее изобретение относится к человеческим антителам против L-селектина. Эти антитела продуцируют с использованием различной техники, описанной ниже. Некоторые человеческие антитела выбирают путем экспериментов на конкурентное связывание либо иначе путем определения, имеют ли эти антитела такую же специфичность антигенной детерминанты, что и конкретное мышиное антитело, например, такое как мышиное антитело DREG-200 или его "очеловеченный" вариант. Эти антитела имеют, по всей вероятности, общие ценные терапевтические свойства, продемонстрированные для "очеловеченного" антитела DREG-200.
Антитела, имеющие требуемую специфичность антигенной детерминанты, могут быть также идентифицированы путем их скрининга на способность блокировать взаимодействие нейтрофилов и эндотелиальных клеток. Простой визуальный анализ на обнаружение такого взаимодействия был описан Kishimoto и др. (1991) (см. выше). Вкратце, этот способ заключается в том, что монослои клеток, происходящих из пупочной вены человека, стимулируют интерлейкином IL -1. Затем к этим слоям при определенных условиях добавляют нейтрофилы, необработанные или заранее обработанные испытуемым антителом, после чего с помощью микроскопа определяют число прилипающих нейтрофилов. В одном из методов, нейтрофилы получают от пациентов с дефицитом прилипания лейкоцитов. См. Anderson и др. Ann. Rev. Med. 38:175 (1987). Нейтрофилы от таких пациентов не имеют рецепторов интегрина, что очень удобно, поскольку связывание этих рецепторов с нейтрофилами могло бы затемнить эффекты блокирования связывания с L-селектином.
а. Методология с использованием триомы
Существо этого метода и подходящий партнер для получения клеточных гибридов (SPAZ-4), используемый в этом методе, описаны Oestberg и др. (Hybridoma 2:361-367 (1983); Oestberg (патент США 4634664) и Engleman и др. (патент США N 4634666) (каждая из этих работ во всей своей полноте вводится в настоящее описание посредством ссылки). Клеточные линии, продуцирующие антитело и полученные указанным способом, были названы триомами, поскольку они происходят из трех клеток - из двух человеческих и одной мышиной. Сначала линию клеток мышиной миеломы гибридизировали с B-лимфоцитом человека, в результате чего получали ксеногенную гибридную клетку, не продуцирующую антитело, такую как клеточная линия SPAZ-4, описанную Oestberg (см. выше). Затем полученную ксеногенную клетку гибридизировали с иммунизированным B-лимфоцитом человека, в результате чего получали клеточную линию триомы, продуцирующую антитело. Было установлено, что триомы продуцируют антитело более стабильно, чем обыкновенные гибридомы, полученные из клеток человека.
Иммунизированные B-лимфоциты получали из крови, селезенки, лимфатических узлов или костного мозга человека-донора. In vivo-иммунизация живых людей L-селектином нежелательна из-за риска продуцирования неблагоприятного ответа. Поэтому B-лимфоциты, обычно, иммунизируют in vitro с использованием полипептида L-селектина, его антигенного фрагмента, или клетки, несущей указанный полипептид или его фрагмент. Если необходимо получить антитела против конкретного антигена или эпитопа, то для in vitro-иммунизации предпочтительно использовать этот антиген или эпитоп. B-лимфоциты, обычно, экспонируют антигеном в течение 7-14 дней в такой среде, как RPMI-1640 (см. Engleman, см. выше), содержащей 10% плазму человека.
Иммунизированные B-лимфоциты гибридизируют с ксеногенными гибридными клетками, такими как SPAZ-4, хорошо известными способами. Например, клетки обрабатывают 40-50% полиэтиленгликолем (MW 1000-4000) примерно при 37oC в течение около 5-10 минут. Затем клетки выделяют из гибридизационной смеси и культивируют в селективной среде для получения нужных гибридов (например, HAT или AH). Клоны, секретирующие антитела с требуемой специфичностью связывания, идентифицируют путем анализа среды с триомной культурой на способность к связыванию с L-селектином или его фрагментом. Затем триомы, продуцирующие антитела с требуемой специфичностью, субклонируют с использованием методологии серийного разведения и культивируют in vitro в культуральной среде. После этого полученные клетки триомы анализируют на способность к связыванию с L-селектином или его фрагментом.
Хотя триомы являются генетически стабильными, они не продуцируют очень высоких уровней антител. Уровни экспрессии могут быть увеличены путем клонирования генов антител, полученных из триомы, в одном или нескольких экспрессирующих векторах с последующей трансформацией вектора в клеточную линию, например, такую как обсуждаемые клеточные линии, используемые для экспрессии рекомбинантных или "очеловеченных" иммуноглобулинов.
b. Трансгенные млекопитающие, не относящиеся к человеку
Человеческие антитела против L-селектина могут быть также продуцированы от трансгенных млекопитающих, не являющихся человеком, и содержащие трансгены, которые кодируют по крайней мере сегмент локуса иммуноглобулина человека. Обычно эндогенный локус иммуноглобулина таких трансгенных млекопитающих функционально инактивируют. Предпочтительно, если сегмент локуса иммуноглобулина человека включает в себя нереаранжированные последовательности компонентов тяжелой и легкой цепи. Инактивация генов эндогенного иммуноглобулина и введение генов экзогенного иммуноглобулина могут быть осуществлены путем направленной гомологичной рекомбинации или путем введения УАС-хромосом. Полученные в результате трансгенные млекопитающие обладают способностью к функциональной реаранжировке последовательностей компонентов иммуноглобулина, а также к экспрессии сывороточного спектра антител различных изотипов, кодируемых генами иммуноглобулина человека, но при этом эти млекопитающие не обладают способностью экспрессировать гены эндогенного иммуноглобулина. Продуцирование таких млекопитающих и их свойства подробно описаны Lonberg и др., 093/12227 (1993); Kucherlapati WO 91/10741 (1991) (каждая из указанных работ полностью вводится в настоящее описание посредством ссылки). Особенно подходящими являются трансгенные мыши. Антитела против L-селектина могут быть получены путем иммунизации трансгенных млекопитающих, не относящихся к человеку (например, описанных Lonberg или Kucherlapati (см. выше)), с использованием L-селектина или его фрагмента. Моноклональные антитела могут быть получены, например, путем гибридизации B-клеток, полученных от вышеуказанных млекопитающих, с соответствующими линиями клеток миеломы с использованием известной технологии Kohler-Milstein.
c. Методы фаговой индикации
Другой способ получения чел. антител против L-селектина заключается в скрининге ДНК-библиотеки из B-клеток человека в соответствии с общим протоколом Huse и др. Science 246:1275- 1281 (1989). Для этого выбирают антитела, связывающиеся с L-селектином, или его фрагментов. Затем клонируют и амплифицируют последовательности, кодирующие указанные антитела (или связывающиеся фрагменты). Протокол, разработанный Huse, может быть использован с большей эффективностью в сочетании с методикой фаговой индикации. См., например, Dawer и др. WO 91/17271 и Mc Cafferty и др. WO 92/01047 (каждая из указанных работ во всей полноте вводится в настоящее описание посредством ссылки). В этой методике продуцируются библиотеки фага, члены которой отображают на своей поверхности различные антитела. Эти антитела обычно представляют собой FV- или Fao-фрагменты. Выбор фага, указывающего на антитело с нужною специфичностью, осуществляют по наиболее высокой аффинности в отношении полипептида L-селектина или его фрагмента.
В одном из своих вариантов метод фаговой индикации заключается в том, что могут быть продуцированы человеческие антитела, имеющие специфичность связывания выбранного мышиного антитела. См. Winter, WO 92/20791. В этом методе в качестве исходного материала используют вариабельную область либо тяжелой, либо легкой цепи выбранного мышиного антитела /например, мышиного антитела DREG-200/. Если, например, в качестве исходного материала выбирают вариабельную область легкой цепи, то фаговую библиотеку конструируют для тех членов, которые обнаруживают одинаковую вариабельную область легкой цепи (т. е. мышиный исходный материал) и различные вариабельные области тяжелой цепи. Вариабельные области тяжелой цепи получают из библиотеки реаранжированных вариабельных областей тяжелой цепи иммуноглобулина человека. Затем выбирают фаг, обнаруживающий сильное специфическое связывание с L-селектином (например, по крайней мере 108 М-1, а предпочтительно, по крайней мере 109 М-1). Вариабельная область тяжелой цепи человека, полученная от этого фага, служит впоследствии в качестве исходного материала для конструирования дополнительной фаговой библиотеки. В этой библиотеке каждый фаг указывает на одинаковую вариабельную область тяжелой цепи (т.е. область, идентифицированную из первой фаговой библиотеки) и на разные вариабельные области легкой цепи. Вариабельные области легкой цепи получают из библиотеки перегруппированных вариабельных областей легкой цепи человека. И аналогично выбирают фаг, обнаруживающий сильное специфическое связывание с L-селектином. Этот фаг указывает на вариабельные области, принадлежащие целиком человеческому антителу против L-селектина. Указанные антитела обычно имеют идентичную или схожую специфичность антигенной детерминанты, что и исходный мышиный материал (например, мышиное антитело DREG-200).
Методы использования
Антитела настоящего изобретения могут быть использованы для лечения заболеваний, связанных с воспалительными процессами, в частности, опосредованными нейтрофилами или Т-клетками. Предпочтительным применением антител настоящего изобретения является лечение или профилактика таких состояний, как ишемическо-реперфузионные повреждения, вызванные инфарктом миокарда, инсультом (например, "ударом"), почечным, печеночным или селезеночным инфарктом, операцией на головном мозге, операцией на сердце (например, артериокоронарного шунтирования), пластической операцией на сосудах и т.п. Другими предпочтительными применениями антител настоящего изобретения является лечение сепсиса, респираторного дистресс-синдрома у взрослых и недостаточности различных органов. Антитела настоящего изобретения могут быть использованы для лечения различных повреждений, вызванных травмами, ожогами, отморожениями, или повреждений спинного мозга. Кроме того, антитела настоящего изобретения могут быть также использовали для лечения аутоиммуных заболеваний, например, таких как ревматоидный артрит, системная красная волчанка, рассеянный склероз, диабет I-типа и увеит; для лечения воспалительных заболеваний кожи, таких как псориаз, и для лечения менингита и энцефалита. Другими типичными применениями указанных антител являются предупреждение и лечение отторжения трансплантированного органа и ревущий "трансплантат против хозяина".
Любой иммуноглобулин настоящего изобретения может быть использован в сочетании с другими антителами, в частности, с человеческими или "очеловеченными" антителами, связывающимися с другими адгезивными молекулами. Например, подходящими для этой цели являются иммуноглобулины, обладающие специфичностью к CD11A, CD11b, CD18, E-селектину, P-селектину, и ICAM-1. Другими подходящими антителами являются антитела, специфичные к лимфокинам, таким как IL-1, IL-2 и IFN- γ , и к другим рецепторам.
Антитела настоящего изобретения могут быть также использованы в виде отдельно вводимых композиций, применяемых в сочетании с химиотерапевтическими агентами. Такими агентами могут быть нестероидные противовоспалительные лекарственные средства и кортикостероиды либо другие хорошо известные лекарственные средства (например, циклоспорин). В основном, иммуноглобулины настоящего изобретения предназначены для использования в комбинации с другими лекарственными средствами, обычно используемыми в современной медицине для лечения конкретных заболеваний.
В некоторых терапевтических методах антитела против L-селектина могут быть использованы в комбинации с тромболитическими средствами. В ранее применяемых методах, лечение пациентов с острым инфарктом миокарда часто проводилось путем открытия закупоренной коронарной артерии. Повторное открытие закупоренной коронарной артерии может быть достигнуто путем введения тромболитических агентов, растворяющих сгустки, вызванные обструкцией артерии, и тем самым восстанавливающих коронарный кровоток. Реперфузия сосудов может быть также достигнута путем экстренной чрезкожной катетерной коронарной ангиопластики (PTCA), осуществляемой с помощью баллонной дилатации закупоренной артерии и суженного сегмента коронарной артерии. Однако, при использовании указанных методов восстановление коронарного кровотока приводит к ишемическо- реперфузионному повреждению сосудов.
В методах настоящего изобретения ишемическо-реперфузионное повреждение сосудов можно уменьшить или предупредить с использованием комбинации тромболитического средства или PTCA и человеческого или "очеловеченного" антитела против L-селектина. Антитела, используемые для профилактических целей, обычно вводят перед введением или одновременно с введением тромболитического агента или инициацией PTCA. Часто после или во время введения тромболитического агента или проведения ангиопластического лечения вводят дополнительные дозы антитела. Интервал между профилактическим введением антитела и началом тромболитического или ангиопластического лечения, в основном, составляет 5-30 мин, предпочтительно 5-20 мин, а более предпочтительно 5-10 мин. Антитела настоящего изобретения вводят парентерально, предпочтительно путем внутривенной инъекции, в дозах, составляющих 0,1 - 10 мг/кг веса тела, предпочтительно 0,14 - 5 мг/кг, а наиболее предпочтительно 0,3-3 мг/кг. Указанные антитела могут быть введены в виде внутривенной инъекции ударной дозы, например, в течение 1 - 5 мин в виде повторных инъекций с меньшими дозами либо в виде внутривенного вливания. Инъекция ударной дозы обычно используется для профилактических доз или в критических случаях. Дополнительные дозы антител могут быть повторно введены (например, каждые 4-6 ч), во время или после тромболитического или ангиопластического лечения острого инфаркта миокарда в тех же самых количествах, что указаны выше, в целях достижения оптимальных уровней антитела в плазме.
Тромболитические агенты представляют собой лекарственные средства, обладающие способностью непосредственно или косвенно стимулировать растворение тромбов in vivo. Такими тромболитическими средствами являются тканевый активатор плазминогена (см. EP-B0 093619), активаза, альтеплаза, дутеплаза, силтеплаза, стрептокиназа, анистреплаза, урокикиназа, гепарин, варфарин и кумарин. Дополнительными тромоболитическими средствами являются саруплаза и активатор плазминогена вампировых. См. Harris, Protejn Engineering, 6:449-458 (1987); PCT-EP 90/00194; патент США 4 970 159). Тромболитические средства вводят пациенту в количестве, достаточном для частичного рассасывания, или для предупреждения образования тромбов и их осложнений. Количество, адекватное для достижения этой цели, определяется как "терапевтически эффективная доза" или "эффективная доза". Количества, эффективные для использования в вышеуказанных целях, зависят от тяжести состояния, общего состояния пациента, способа введения и используемой комбинации с другими лекарственными средствами. Часто терапевтически эффективные дозы тромболитических агентов и схемы их введения утверждаются Управлением по контролю за качеством пищевых продуктов и медикаментов (FDA) для независимого использования, например, для альтеразы эта доза составляет 100 мг, а для стрептокиназы 1,5 млн.ME.
Предпочтительная композиция настоящего изобретения предусматривает использование иммуноглобулина настоящего изобретения, включенного в иммунотоксины, которые нейтрализуют клетки, экспрессирующие L-селектин. Иммунотоксины характеризуются тем, что они состоят из двух компонентов и используются для лизиса отобранных клеток in vitro или in vivo. Один компонент иммунотоксина представляет собой цитотоксичный агент, оказывающий фатальное действие на клетку при его присоединении или абсорбции. Другой компонент, известный как "направленный носитель", является средством доставки токсичного агента к клеткам конкретного типа, например к клеткам, экспрессирующим антигенную детерминанту L-селектина. Эти два компонента обычно связывают друг с другом химическим путем с использованием хорошо известных процедур. Например, если цитотоксичный агент является белком, а второй компонент представляет собой интактный иммуноглобулин, то эти компоненты могут быть связаны с помощью перекрестно-сшивающих агентов (кросс-линкеров), например SPDP, карбодиимида, глутаральдегида или т.п. Продуцирование различных иммунотоксинов хорошо известно специалистам, см., например, работу "Monoclonal Antibody-Toxin Conjugates: Aiming the Magic Bullet" Thorpe et al., Monoclonal Antibodies in Clinical Medicine, Academic Press, pp. 168-190 (1982), которая вводится в настоящее описание посредством ссылки. Указанные компоненты могут быть также связаны генетически (см. Chaudhary et al., Nature 339: 394 (1989); эта работа вводится в настоящее описание посредством ссылки).
Для получения иммунотоксинов могут быть использованы различные цитотоксичные агенты. Такими цитотоксичными агентами могут быть радионуклиды, такие как йод-131 или другие изотопы йода, иттрий-90, рений-188 и висмут-212 или другие альфа-излучатели; различные химиотерапевтические средства, такие, как виндезин, метотрексат, адриамицин и цисплатин; а цитотоксичные белки, такие как белки, инактивирующие рибосому, например противовирусный белок фитоллаки американской, экзотоксин A Pseudomonas, рицин, дифтерийный токсин, цепь рицина A и т.п., либо агент, активный на клеточной поверхности, такой как фосфолипаза (например, фосфолипаза C). (См. в общих чертах, переуступленная заявка на патент США рег. N 07/290968; "Chimeric Toxins', Olsnas & Phil, Pharmac. There., 25:355-381 (1982) и Monoclonal Antibodies for Cancer Detection and Therapy (изд. Baldwin & Byers, Academic Press, 1985), pp. 159-179, 22-266; все указанные работы вводятся в настоящее описание посредством ссылки).
В качестве компонента, используемого для доставки цитотоксичного агента, могут быть использованы иммуноглобулины настоящего изобретения. Предпочтительными являются интактные иммуноглобулины или их связывающие фрагменты, такие как Fab или Fv. Обычно, антитела, используемые в иммунотоксинах, являются человеческими иммуноглобулинами изотипа IgM или IqG, но, если необходимо, то могут быть также использованы константные области иммуноглобулинов других млекопитающих.
Антитела и фармацевтические композиции настоящего изобретения используются предпочтительно для парентерального введения, например подкожного, внутримышечного или внутривенного введения. Антитела настоящего изобретения могут быть также использованы для местного применения, для чреззондового питания или лаважа, для внутрибрюшинных инъекций, для изготовления глазных мазей и мазей для наружного применения, для внутричерепных инъекций (обычно, в желудочек головного мозга), для интраперикардиальных инъекций или для интрасиновиальных инъекций. Композиции для парентерального введения, обычно, содержат раствор иммуноглобулина или его смесь, растворенную в приемлемом носителе, а предпочтительно в водном носителе. В этих целях могут быть использованы различные водные носители, например вода, забуферная вода, фосфатно-буферный раствор (PBS); 0,4% физиологический раствор, 0,3% глицин, раствор альбумина человека и т.п. Эти растворы являются стерильными и, в основном, не содержат каких-либо частиц. Композиции настоящего изобретения могут быть стерилизованы стандартными способами, хорошо известными специалистам. Эти композиции могут содержать фармацевтически приемлемые добавки, необходимые для создания физиологических условий, такие как pH-корректирующие и забуферивающие агенты, агенты, регулирующие токсичность, и т.п., например, ацетат натрия, хлорид натрия, хлорид калия, хлорид кальция и лактат натрия. Концентрация антитела в этих композициях может широко варьироваться, например, примерно от менее чем 0,005%, а обычно по крайней мере от около 1% до не менее чем 15 или 20 мас.%; и выбор концентрации антитела осуществляют, главным образом, исходя из объема жидкости, вязкости и т.п., а также в зависимости от выбранного способа введения.
Так, например, типичная фармацевтическая композиция для инъекций может содержать 1 мл стерильной забуференной воды и 1-70 мг иммуноглобулина. Типичная композиция для внутривенных вливаний может содержать 250 мл стерильного раствора Рингера и 150 мг антитела. Методы получения парентеральных композиций хорошо известны специалистам и подробно описаны в литературе (см. , например Remington's Pharmaceutical Science (15-ое изд., Mack Publishing Company, Easton, Pennsylvania, 1980; эта работа вводится в настоящее описание посредством ссылки). Композиции, подходящие для лаважа или для других способов введения, могут быть изготовлены в соответствующих формах в зависимости от целей их использования. Некоторые фармацевтическое композиции могут содержать как антитело против L-селектина, так и тромболитическое средство.
Для хранения антитела настоящего изобретения могут быть заморожены или лиофилизованы с последующим их восстановлением в соответствующем носителе непосредственно перед использованием. Было установлено, что при использовании обычных иммуноглобулинов может быть с успехом применена традиционная техника лиофилизации и восстановления. Однако, следует отметить, что лиофилизация и последующее восстановление могут привести к различной степени потери активности антител (например, в случае обычных иммуноглобулинов антитела IgM обнаруживают большую тенденцию к потере активности, чем антитела IgG), а поэтому может оказаться необходимым скорректировать используемые уровни для компенсации возможных потерь.
Композиции настоящего изобретения, содержащие антитела или их смесь, могут быть введены в профилактических и/или терапевтических целях. Для терапевтических целей композиции вводят пациенту, страдающему воспалительными заболеваниями, в количестве, достаточном для излечения или по крайней мере для частичного подавления данного заболевания и его осложнений. Количество, адекватное для достижения этой цели, определяется как "терапевтически эффективная доза". Эффективная доза зависит от тяжести заболевания, общего состояния иммунной системы пациента и составляет, в основном, от около 1 до около 200 мг антитела на дозу, а обычно от 5 до 70 мг на пациента. Схема введения лекарственного средства может варьироваться в зависимости от конкретного заболевания и состояния пациента и обычно колеблется от одной ударной дозы или продолжительного вливания в день до многократного введения в течение 1 дня (например, каждые 4-5 часов) в соответствии с назначением лечащего врача исходя из состояния пациента. При этом следует иметь в виду, что материалы настоящего изобретения могут быть использованы при серьезных заболеваниях в опасных для жизни или потенциально опасных для жизни ситуациях. В таких случаях для минимизации присутствия инородных веществ и снижения вероятности отторжения "чужеродного материала", которое достигается благодаря применению иммуноглобулинов настоящего изобретения, может оказаться желательным введение избыточного количества этих антител.
В профилактическом применении композиции, содержание антитела настоящего изобретения или их смесь, вводят пациенту, еще не страдающему конкретной болезнью, в целях усиления его сопротивляемости. Количество антитела, вводимого в этих целях, определяют как "профилактически эффективную дозу". В таких случаях точные количества антитела также зависят от общего состояния здоровья пациента и общего уровня его иммунитета и, в основном, составляют от 1 до 70 мг на дозу. Предпочтительными профилактическими целями использования композиций настоящего изобретения являются предупреждение респираторного дистресс-синдрома у взрослых пациентов, страдающих травмами или сепсисом; предупреждение отторжения трансплантированного органа и предупреждение реперфузионного повреждения у пациентов, страдающих ишемией. Для серьезно больных пациентов могут быть использованы дозы, составляющие от около 50 до 150 мг "очеловеченного" или человеческого иммуноглобулина на дозу либо более высокие дозы.
Выбор доз и схемы введения (т.е. разовое или многократное введение) композиций настоящего изобретения обычно осуществляет лечащий врач. Но в любом случае фармацевтические композиции должны включать в себя определенное количество антител, достаточное для эффективного лечения пациента.
Кроме того, антитела настоящего изобретения могут найти широкое применение в in vitro-исследованяях. Например, эти антитела могут быть использованы для детекции антигенов L-селектина в целях выделения специфических лейкоцитов или т.п.
Другими примерами такого применения являются, но не ограничиваются ими, "очеловеченный" иммуноглобулин DREG-200, который может быть иммобилизован и подвергнут взаимодействию с кровью, взятой у пациента, для удаления клеток крови, несущих антигены L-селектина, после чего оставшаяся кровь с удаленными L-селектин-несущими клетками может быть снова введена пациенту. Любые остаточные "очеловеченные" антитела, присутствующие в L-селектин-обедненной и вновь введенной пациенту крови (например, в результате отщепления от иммобилизирующего носителя), будут иметь очень небольшую или совсем незначительную антигенность по сравнению с антителом.
При использовании в целях диагностики антитела могут быть меченными или немеченными. Немеченные антитела могут быть использованы в комбинации с другими меченными антителами ("вторые" антитела), которые являются реактивными по отношению к "очеловеченным" или человеческим антителам, например антитела, специфичные к константным областям иммуноглобулина человека. Альтернативно, антитела могут быть подвергнуты прямому мечению. Для этого могут быть использованы различные метки широкого ряда, например, такие как радионуклиды, флуоресцентные вещества, ферменты, ферментные субстраты, конферменты, ингибиторы ферментов, лиганды (в частности, гаптены) и т.п. В этих целях могут быть использованы различные типы иммуноанализов, хорошо известных специалистах.
Нижеприведенные примеры иллюстрируют настоящее изобретение и не должны рассматриваться как некое ограничение объема изобретения. При этом следует отметить, что хотя приведенные ниже примеры относятся к использованию мышиного антитела DREG-200 для продуцирования "очеловеченного" антитела против L-селектина с высокой аффинностью связывания, однако, для продуцирования "очеловеченного" антитела могут быть также использованы гипервариабельные области (CDR), происходящие от мышиного антитела DREG-55, мышиного антитела DREG-56 или другого моноклонального антитела, способного связываться с эпитопом L-селектина.
Пример 1: Клонирование кДНК тяжелой и легкой цепи
кДНК для генов вариабельных доменов тяжелой и легкой цепи мышиного антитела DREG-200 клонировали с использованием полимеразно-цепных реакций (описанных Co и др. в. J. Immu. Vol. 148:1149 (1992) и в переуступленной заявке США рег. N 07/634278), а также 3'-праймеров, которые гибридизируются с константными областями и содержат HindIII-сайты, и 5'-праймеров, которые гибридизируются с IgG-концами и содержат EcoRI-сайты. PCR-амплифицированные фрагменты переваривали ферментами EcoRI и HindIII и клонировали в вектор pUC18 для секвенирования. Для мышиного антитела DREG-200 были секвенированы по крайней мере два гамма-1-специфических клона и два каппа-специфических клона. Гамма-1-клоны и каппа-клоны имели идентичную последовательность. кДНК-последовательности вариабельного домена и введенные аминокислотные последовательности представлены на фиг. 1.
Пример 2: Компьютерное моделирование "очеловеченных" антител
Для сохранения высокой аффинности связывания в "очеловеченных" антителах были проведены нижеследующие процедуры, описанные Queen и др. (см. Queen и др. Proc. Natl. Acad. Sci. USA 86:10029 (1989) и WO 90/07861; эти работы полностью вводятся в настоящее описание посредством ссылки). Для исходного донорного мышиного антитела необходимо выбрать более гомологичное акцепторное человеческое антитело, поскольку в этом случае при объединении мышиных CDR-областей с чел. каркасными областями меньше вероятности внести искажения в CDR, которые могут привести к снижению аффинности связывания антитела. Предпочтительно, если гомология (т.е., процент идентичности последовательности) между каркасной вариабельной областью тяжелой цепи "очеловеченного" иммуноглобулина и каркасной вариабельной областью тяжелой цепи донорного иммуноглобулина составляет по крайней мере 65%. Обычно, для получения каркасных последовательностей тяжелую и легкую цепь выбирают от одного и того же антитела человека для того, чтобы по возможности снизить несовместимость при сборке этих двух цепей. В результате поиска гомологичных последовательностей исходя из базы данных белковых последовательностей (осуществляемого с использованием программного обеспечения Micro Genie Sequence Analysis Soft Ware (Beckman)) было выбрано антитело Eu, которое было затем использовано для получения каркасных последовательностей в целях "очеловечивания" мышиного антитела DREG-200.
Для конструирования модели вариабельной области мышиного DREG-200 использовали компьютерную программу ENCAD(Levitt, J. Mol. Biol. 168:595 (1983); эта работа вводится во всей полноте посредством ссылки). Полученную модель использовали для определения аминокислотных последовательностей в каркасной области DREG-200 мыши, которая была достаточно близкой к CDR-областям, чтобы иметь возможность взаимодействовать с этими областями (категория 4, см. ниже). Для конструирования вариабельных областей легкой и тяжелой цепей "очеловеченного" DREG-200 в каждом положении аминокислоты выбирали так, чтобы они совпадали с аминокислотами в антителе Eu, если только это положение не подпадает под одну или более из нижеследующих 5 категорий:
(1) Это положение находится внутри области CDR;
(2) аминокислота в Eu является не характерной для антител человека в этом положении, а аминокислота мышиного DREG-200 является типичной для человеческих антител в этом положении;
(3) Это положение находится в непосредственной близости от CDR;
(4) В соответствии с описанной выше моделью имеется предположение, что данная аминокислота может находиться в непосредственной близости к антигенсвязывающей области (CDR).
Для положений, подпадающих под эти категории, использовались аминокислоты от мышиного антитела DREG-200.
Кроме того, конкретное положение могло подпадать под пятую категорию, а именно:
(5) Аминокислота Eu является в высокой степени нехарактерной для человеческих антител в этом положении, аминокислота DREG-200, хотя и отличается от аминокислоты Eu, но также является нехарактерной. В этих случаях для данного положения использовали аминокислоту, типичную для человеческого антитела.
Аминокислоты каждой категории показаны в Таблице 1. Некоторые аминокислоты могут подпадать под более чем одну категорию. Конечные последовательности вариабельных доменов легкой и тяжелой цепи "очеловеченного" антитела DREG-200 показаны на фиг. 2 в сравнении с последовательностями мышиного DREG-200.
Для конструирования генов, кодирующих "очеловеченные" антитела, были выбраны (в основном, с использованием кодонов, обнаруживаемых в мышиной последовательности) нуклеотидные последовательности, кодирующие белковые последовательности тяжелых и легких цепей "очеловеченного" антитела, включая сигнальные пептиды. Для создания сайтов рестрикции или для удаления нежелательных сайтов некоторые вырожденные кодоны были заменены. Нуклеотидные последовательности генов также включают в себя донорные сигналы сплайсинга и XbaI-сайт на каждом конце. Нуклеотидные последовательности и кодированные вариабельные домены легкой и тяжелой цепи "очеловеченного" иммуноглобулина показаны на фиг. 3. Каждый ген был сконструирован из четырех перекрывающихся синтетических олигонуклеотидов, описанных в литературе (см. Co и др., J. Immunol. 148:1149 (1992) и переуступленная заявка США рег. N 07/634278, указанные работы целиком вводятся в настоящее описание посредством ссылки). Затем гены, кодирующие вариабельные области тяжелой и легкой цепи, лигировали в XbaI- сайты экспрессирующих векторов pVg1-dhfr или pVk (см. переуступленную заявку США рег. N 07/634278) в соответствующей ориентации в целях продуцирования генов полной тяжелой и легкой цепи. Реакции осуществляли в стандартных условиях, хорошо известных специалистам (Maniatis и др., см. выше).
Плазмиды для тяжелой и легкой цепей были введены в клетки мышиной миеломы Sp2/O путем электропорации, и клетки отбирали для экспрессии gpt. Клоны скринировали путем оценки продуцирования человеческого антитела в супернатанте культуры с помощью ELISA и из клонов о наилучшим продуцированием выделяли антитело. Затем "очеловеченное" IgGI-антитело DREG-200 очищали путем пропускания супернатанта тканевой культуры через колонку с A-Сефарозой стафилококкового белка CL-4B (Phafmacia). Связанное антитело элюировали 0,2 М Глицином-HCl (pH 3,0) и нейтрализовали 1 М Трисом (pH 8,0). Затем буфер заменяли на PBS путем пропускания через PD10-колонку (Pharmacia) или путем диализа. Для получения клеток, продуцирующих наиболее высокие уровни антитела, трансфецированные клоны могут быть культивированы в возрастающих концентрациях метотрексата.
Для продуцирования "очеловеченного" антитела DREG-200 изотипа IgG4 сначала был сконструирован другой вектор pVg4-dnfr. Для этого XbaI-BamH1-фрагмент вектора pVg1-dnfr, содержащего константную γ 1-область, заменяли фрагментом (приблизительно в 2000 п.о.) гена константной γ 4-области иммуноглобулина человека (Ellison и Hood, Proc. Natl. Acad. Sci. USA 79:1984 (1982)), который простирался от Hind111-сайта, находящегося перед CH1-экзоном γ 4-гена до 270 п. о. после Nsi1-сайта, расположенного за CH4-экзоном этого гена, с использованием известной техники, включая полимеразно-цепную реакцию. Затем ген, кодирующий вариабельную область тяжелой цепи "очеловеченного" антитела DREG-200, клонировали в Xba1-сайт вектора pVg4-dnfr. Полученную плазмиду, несущую ген тяжелой цепи, вместе с вышеописанной плазмидой, несущей ген легкой цепи, вводили в клетки Sp2/O, клоны отбирали,
а "очеловеченное" антитело DREG-200 изотипа IgG4 очищали как описано выше для антитела IgG1.
Пример 3. Свойства "очеловеченных" антител
Аффинность "очеловеченных антител DREG-200 против L-селектина определяли путем конкурентного связывания с мышиным антителом DREG-200, меченным радиоактивным йодом (фиг. 4). Аффинность связывания вычисляли по методу Berzofsky (J. A. Berzofsky и L.J. Berkower, в"Fundamental Immunology (ed. W.E. Paul), Raven Press (New York), 595 (1984), эта работа целиком вводится в настоящее описание посредством ссылки). Аффинность "очеловеченных" антител DREG-200 примерно в 2 раза превышает аффинность мышиного антитела DREG-200. Аналогичные результаты могут быть получены, если аффинность антитела против L-селектина измерять на нейтрофилах человека.
Способность мышиного и "очеловеченного" антитела DREG-200 блокировать прилипание нейтрофилов к эндотелиальным клеткам определяли посредством модификации аналитического метода Hallmann et al., Bio Chem. Biophys. Res. Comm. 174:236 (1991). В частности, эндотелиальные клетки пуповины (HUVEC, от Clonetics, San Diego) культивировали до состояния сплошности в EGM-среде (Clonetics) на предметных стеклах с лунками Lab-Tek 8 (Nunc, Naperville, IL). Перед использованием HUVEC-клетки стимулировали в течение 4 часов с использованием 20 нг/мл IL - 1β (R&D Systems, Minneapolis, MN). Посредством центрифугирования в градиенте плотности нейтрофилы выделяли из светлых слоев кровяного сгустка, который был очищен от эритроцитов путем осаждения декстраном, после чего их количество доводили до 107/мл. Нейтрофилы (100 мкл) предварительно инкубировали в течение 20 минут на льду с различными концентрациями антитела (в 100 мкл RPMI). Предметные стекла с HUVEC-клетками промывали при отсутствии 1L - 1β и помещали в роторный шейкер (100 об./мин) при 4oC. Затем в лунки добавляли необработанные или обработанные антителом нейтрофилы и предметные стекла инкубировали на шейкере в течение 30 минут при 4oC. Затем предметные стекла промывали путем погружения 10 раз в сосуд со RPMI-средой, фиксировали в 1% глутаральдегиде в RPMI и оставляли на воздухе для осушки. Количественную оценку прилипания нейтрофилов проводили путем подсчета нейтрофилов, связанных с определенными участками монослоя эндотелиальных клеток, при помощи микроскопа. Как показано на фиг. 5, "очеловеченное" lgG1 и мышиное антитела DREG-200 эффективно блокируют связывание нейтрофилов с HUVEC, тогда как иррелевантное контрольное антитело к такому блокированию неспособно. "Очеловеченное" IgG4-антитело DREG-200 аналогичным образом блокирует связывание нейтрофилов с эндотелиальными клетками.
Пример 4.
Влияние антитела чел. DREG-200 на повреждение мио- после реперфузии.
Исследовали степень реального защитного действия "очеловеченного" антитела DREG-200 изотипа IgG4 (очел. DREG-200) на ишемическую ткань миокарда после реперфузии. Самцов взрослых кошек (2,8-4,2 кг) анестезировали натрия пентобарбиталом (30 мг/кг, i. v.). В срединный разрез вводили трахеотомическую канюлю, после чего кошек помещали в условия искусственной вентиляции легких под переменным положительным давлением (небольшой респиратор для животных, Harvard, Dover, МА). В правую внешнюю яремную вену вводили полиэтиленовый катетер для дополнительного введения пентобарбитала для поддержания нужного уровня наркоза при операции и для введения антител. Другой полиэтиленовый катетер вставляли через левую бедренную артерию и помещали в абдоминальной аорте для измерения среднего артериального давления (МАВР) с помощью датчика давления (Cobe Instruments, Lakewood, CO). После грудинной торакотомии вскрывали передний перикард и наносили шов шелковой нитью (3-0) вокруг левой передней нисходящей (LAD) коронарной артерии на расстоянии 8-10 мм от ее начала. Затем через апикальную ямку в левый жердочек вводили высокоточный датчик давления с наконечником катетера (Model MPC 500 с блоком управления - Model TCB 500, Milla Instruments Inc., Houston, TX). Катетер устанавливали посредством наблюдения за LV-давлением и формой dP/dt-волны, а затем закрепляли на месте шелковым швом. Для определения частоты пульса (HR) и элевации ST-сегмента использовали стандартное отведение II скалярной электрокардиограммы (ECG). Элевацию ST-сегментов определяли путем анализа записи ECG при 50 мм/с каждые 2O минут. ECG, MABP, LVP и dP/dt постоянно контролировали на мониторе с блоком Hewlett - Packard 78304 A(Hewlett - Packard, Palo Alto, CA) и записывали на осцилографическом рекордере (Gould Inc., Cleveland, OH) каждые 20 минут. PRI-индекс, приблизительная потребность миокарда в кислороде, вычисляли как отношение MABP и HP/1000.
После завершения всех хирургических процедур кошек оставляли на 30 минут для стабилизации и в течение этого времени записывали базовые показания ECG, MABP, LVP и dP/dt. Ишемию миокарда (MI) индуцировали путем стягивания обратимой лигатуры, первоначально помещенной вокруг LAD, в целях достижения полной закупорки сосудов. Эту ситуацию определяли как нулевой момент времени. Через 80 минут после коронарной окклюзии внутривенно вводили в качестве ударной дозы 2 мг/кг веса тела очел. DREG-200 (изотип IgG4) или контрольное антитело Mab hu ABL - 364 (т.е. "очеловеченное" контрольное Mab, соответствующее изотипу IgG4) (за 10 минут до реперфузии, R). Затем через 10 минут (т.е. в целом через 90 минут после ишемической окклюзии, I) лигатуру вокруг LAD развязывали и ишемический миокард подвергали реперфузии в течение 4,5 часов.
Животных произвольно разделяли на три основные группы. Группе из шести кошек с ложной MI + R вводили очел. DREG-200 (2 мг/кг), группе из 6 кошек MI + R вводили контрольное антитело Mab hu ABL - 364 (2 мг/кг) и группе из 6 кошек MI + R вводили очел. DREG-200 (2 мг/кг). Группу кошек с ложной MI + R подвергали аналогичным хирургическим процедурам, за исключением того, что LAD коронарная артерия не подвергалась окклюзии.
По окончании 4,5-часового периода реперфузии лигатуру вокруг LAD снова затягивали. После этого в левый желудочек быстро инъецировали 20 мл 0,5% голубого Эванс в целях окрашивания зоны миокарда, которая подвергалась перфузии посредством открытых коронарных артерий. Эта зона "под угрозой" определялась путем негативного окрашивания. Сразу после этой инъекции сердце быстро вырезали и помещали в подогретый, насыщенный кислородом K-H-раствор. Левую огибающую (LCX) и LAD-коронарную артерии выделяли и удаляли для последующего исследования вазоактивности коронарного кольца и прилипания PMN. Правый желудочек, крупные сосуды и жировую ткань осторожно удаляли, а левый желудочек разрезали на секции толщиной 3 мм параллельно атриовентрикулярной борозде. Неокрашенную часть миокарда (т.е. полная зона риска или ишемическая зона) отделяли от части миокарда, окрашенного голубым Эванса (т.е. безопасная зона или неишемическая зона). Зону под угрозой (или зону риска) разрезали на небольшие кубики и инкубировали в 0,1% нитросинем тетразолии в фосфатном растворе (pH 7,4) при 37oC в течение 15 минут. Тетразолиевый краситель образует синий формазановый комплекс в присутствии клеток миокарда, содержащий активные дигидрогеназы и их кофакторы. Необратимо поврежденную или некротическую часть миокарда зоны риска, которая не окрашивалась, отделяли от окрашенной части миокарда (т.е. ишемической, но не некротической зоны). После этого все три части миокарда (т.е. неишемическая, ишемическая, но не некротическая, и ишемическая некротическая ткань) взвешивали. Полученные результаты выражали в виде площади некротической сердечной ткани, вычисленной как % либо от зоны риска, либо от полной массы левого желудочка.
В соответствии с этими критериями повреждение сердечной ткани было значительно уменьшено (p < 0,001) у кошек, обработанных очел. DREG-200. Тогда, как у группы, обработанной контрольным антителом, около 30% миокарда зоны риска развивалось в некротическую ткань, причем у очел. DREG-200-oбpaботанной группы количество некротической ткани составляло менее 15% (p < 0,01), т.е. уменьшилось на 50-60% (см. фиг. 6). Значительного различия между мокрыми массами зон риска, выраженными как процент от полной массы левого желудочка, для двух ишемических групп не наблюдалось, что указывает на то, что ишемии миокарда подвержены сравнительно одинаковые области у этих двух групп. Таким образом, очеловеченное антитело DREG-200 обеспечивает значительную защиту от повреждений после реперфузии.
Заметная способность очел. DREG-200 защищать ишемическую ткань была также проиллюстрирована благодаря измерениям активности креатинкиназы в плазме, биохимического маркера повреждения миокарда. Пробы артериальной крови (2 мл) брали сразу после наложения лигатуры, а затем через каждый час. Эту кровь собирали в полиэтиленовые пробирки, содержащие 200 ME натриевой соли гепарина. Полученные пробы центрифугировали при 2000 x г при 4oC в течение 20 минут, после чего плазму декантировали для биохимического анализа. Концентрацию белка в плазме определяли с использованием биурета по методу Gornall и др., J. Biol. Chem. 177:751-766 (1949). Активность креатинкиназы (СK) в плазме измеряли по методу Rosalki, J. Lab. Clin. Med. 69:696-705 (1967) и выражала как ME/мкг белка.
У кошек с ложной MI/R, получавших очел. DREG-200, на протяжении 6 часов наблюдалась слегка повышенная активность СK в плазме, достигая величины 3,8 ± 0,9 ME/мкг белка. Для двух ишемических групп активность CK в плазме слегка возрастала в период ишемии миокарда. У кошек, получавших hu ABL-364, активность CK в крови значительно возрастала в течение первых 30 минут после реперфузии и оставалась повышенной в течение 4 часов периода реперфузии. В противоположность этому, у ишемической группы, обработанной очел. DREG-200, наблюдалась значительно меньшая активность CK в плазме, чем у ишемической группы, обработанной hu ABL-364 (p < 0,05). Этот эффект наблюдался в течение всего периода реперфузии, что свидетельствовало о том, что очел. DREG-200 обладает значительным защитным действием от повреждений миокарда после реперфузии.
Пример 5: Влияние очел. DREG-200 на функцию сердца
Влияние очел. DREG-200 (изотип IgG4) на функцию сердца определяли путем измерения давления в левом желудочке (LVP), первой производной от LVP, dP/dtмакс, показателя сократительной способности миокарда. Все данные были получены с помощью манометра, вводимого посредством наконечника катетера в полость левого желудочка. Все три группы кошек, обсуждаемые в предыдущем примере, показали сравнительно одинаковые начальные величины указанных кардиальных параметров. Для группы с ложной MI на протяжении всего экспериментального периода (6 часов) не наблюдалось значительных изменений в величинах dP/dtмакс. Тогда как в обеих группах MI/R после окклюзии LAD наблюдалось увеличение dP/dtмакс примерно на 65%. У кошек, обработанных hu ABL-364, сократительная способность миокарда не обнаруживала заметного восстановления. Однако, у очел. DREG-200-обработанных MI/R-кошек, dP/dtмакс. восстановилась до контрольных значений через три часа после реперфузии. Таким образом, после 4,5-часовой реперфузии dP/dtмакс. была значительно ниже у hu ABL-364-обработанных кошек, чем у очел. DREG-200-обработанных кошек (p < 0,01). Полученные результаты показали, что очел. DREG-200-антитело не только способствует снижению постреперфузионного некроза миокарда при ишемии, но также способствует улучшению механических функций сердца.
В соответствии с вышеизложенным следует отметить, что иммуноглобулины настоящего изобретения имеют ряд преимуществ по сравнению с другими антителами, специфичными к L-селектину. По сравнению с мышиными моноклональными антителами иммуноглобулины настоящего изобретения могут быть продуцированы с меньшими экономическими затратами и содержат значительно меньше чужеродных аминокислотных последовательностей. Это снижает вероятность продуцирования антигенности после их введения человеку, что значительно повышает терапевтический эффект этих антител.
Все публикации и патентные заявки, цитированные выше, во всех случаях и во всей своей полноте вводятся в настоящее описание посредством ссылки. Хотя настоящее изобретение описано подробно на конкретных примерах его осуществления, однако, следует указать, что эти примеры приводятся лишь в иллюстративных целях, в связи с чем подразумевается, что в настоящее изобретение могут быть внесены различные изменения и модификации, не выходящие за рамки нижеследующей формулы изобретения.
| название | год | авторы | номер документа |
|---|---|---|---|
| РЕКОМБИНАНТНОЕ АНТИТЕЛО, ПОЛИНУКЛЕОТИД (ВАРИАНТЫ), СПОСОБ ПОЛУЧЕНИЯ ТЯЖЕЛЫХ И/ИЛИ ЛЕГКИХ ЦЕПЕЙ РЕКОМБИНАНТНОГО АНТИТЕЛА (ВАРИАНТЫ) | 1989 |
|
RU2126046C1 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА, КОТОРЫЕ РАСПОЗНАЮТ ВЕРОТОКСИН II, И ПРОДУЦИРУЮЩАЯ ИХ ЛИНИЯ КЛЕТОК | 1999 |
|
RU2217166C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА И ИХ ИСПОЛЬЗОВАНИЕ | 1994 |
|
RU2139934C1 |
| АНТИТРОМБОТИЧЕСКОЕ СРЕДСТВО И ГУМАНИЗИРОВАННОЕ МОНОКЛОНАЛЬНОЕ АНТИТЕЛО ПРОТИВ ФАКТОРА ФОН ВИЛЛЕБРАНДА | 1999 |
|
RU2223785C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА К ФАКТОРУ D И ИХ ПРИМЕНЕНИЯ | 2007 |
|
RU2474589C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА К ФАКТОРУ D И ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2678802C2 |
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО К АМИЛОИДУ БЕТА | 2013 |
|
RU2668161C2 |
| ИММУНОГЛОБУЛИНЫ K CD52 ЧЕЛОВЕКА | 2010 |
|
RU2603743C2 |
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО К КОНФОРМАЦИОННОМУ ЭПИТОПУ С3 КОМПОНЕНТА КОМПЛЕМЕНТА ЧЕЛОВЕКА, ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК (ВАРИАНТЫ), ЭКСПРЕССИОННЫЙ ВЕКТОР, СОДЕРЖАЩИЙ ПОСЛЕДОВАТЕЛЬНОСТЬ ДНК (ВАРИАНТЫ), И ШТАММ КЛЕТОК ЯИЧНИКОВ КИТАЙСКОГО ХОМЯЧКА CHO-humC34-ПРОДУЦЕНТ ДАННОГО ГУМАНИЗИРОВАННОГО АНТИТЕЛА | 2016 |
|
RU2630647C1 |
| ХИМЕРНЫЕ И ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА ПРОТИВ ИНТЕГРИНА α5β1, КОТОРЫЕ МОДУЛИРУЮТ АНГИОГЕНЕЗ | 2003 |
|
RU2346701C2 |
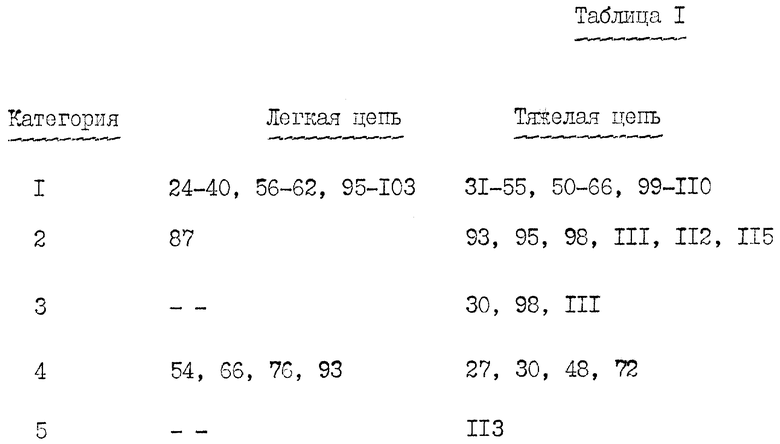



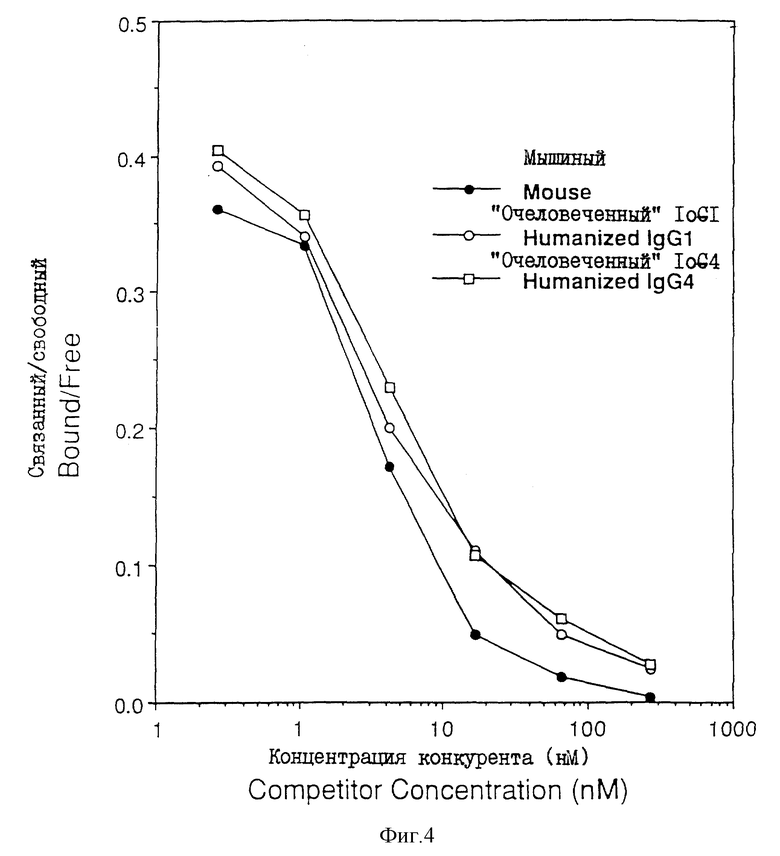
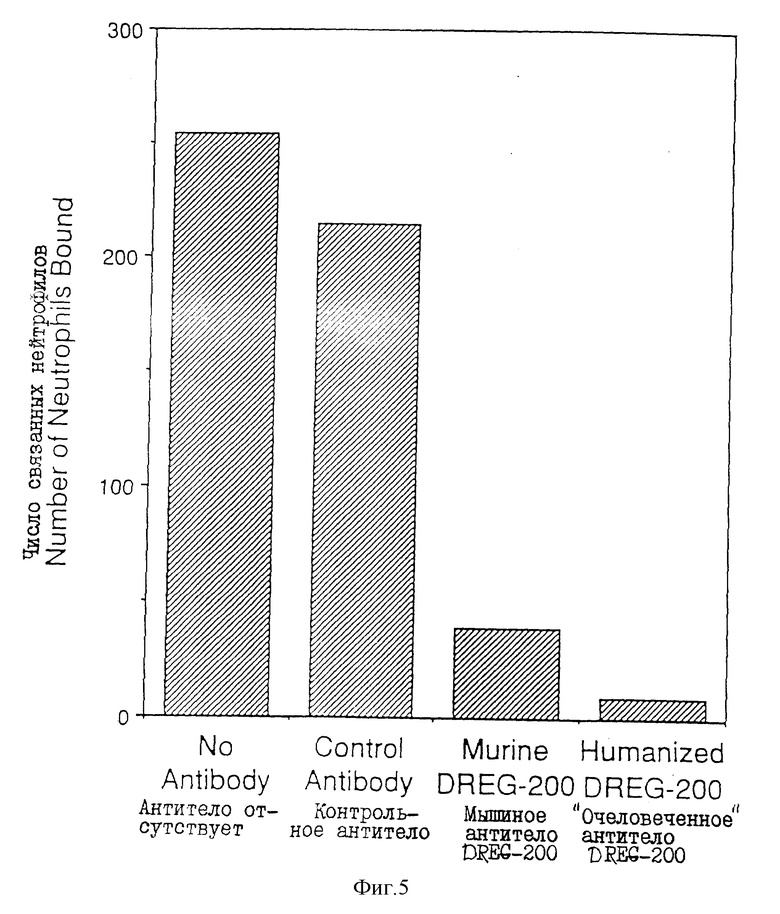
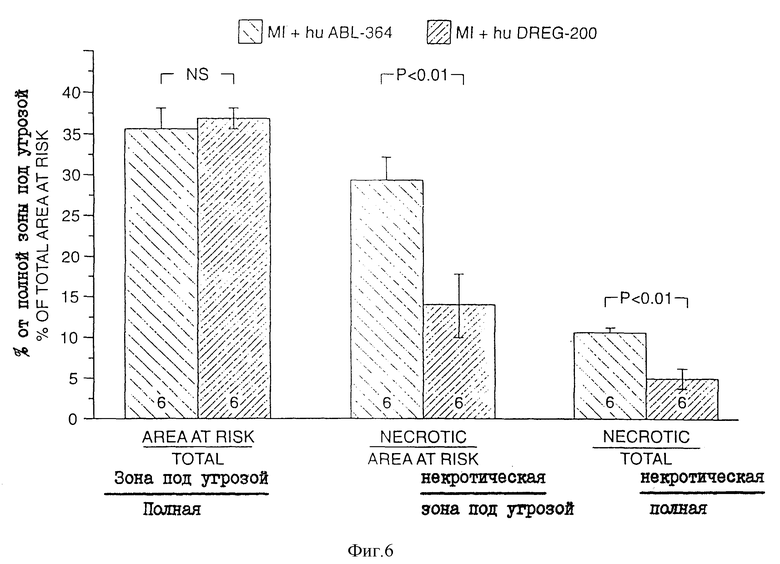
Изобретение относится к биотехнологии и касается гуманизированного иммуноглобулина, специфичного для белка L-селектина человека. Сущность изобретения состоит в том, что гуманизированный иммуноглобулин, специфичный для белка L-селектина человека, представляет собой химерный иммуноглобулин, имеющий области, определяющие комплементарность (CDR), соответствующие CDR-областям донорного мышиного иммуноглобулина, и каркасные области вариабельного района тяжелой и легкой цепи акцепторного иммуноглобулина человека, и специфически связывающийся с L-селектином человека с константой аффинности 107 M-1. Изобретение позволяет получить неиммуногенный для человека иммуноглобулин специфичный для белка L-селектина. 2 с. и 17 з.п. ф-лы, 6 ил. , 1 табл.
 от СDR в указанном гуманизированном иммуноглобулине.
от СDR в указанном гуманизированном иммуноглобулине.
| СПОСОБ ПРОГНОЗИРОВАНИЯ ВОЗНИКНОВЕНИЯ НАРУШЕНИЙ ВЕГЕТАТИВНОЙ РЕГУЛЯЦИИ У ДЕТЕЙ С ЗАДЕРЖКОЙ ВНУТРИУТРОБНОГО РАЗВИТИЯ В АНАМНЕЗЕ | 2003 |
|
RU2257145C2 |
| Автоматический огнетушитель | 0 |
|
SU92A1 |
| Огнетушитель | 0 |
|
SU91A1 |

