Изобретение относится к области биотехнологии растений, использующей методы генной инженерии, и может быть применено для получения генетически модифицированных древесных растений, экспрессирующих различные гетерологичные гены.
Древесные растения, относящиеся как к сельскохозяйственным культурам, так и к лесным породам, имеют очень большое экономическое значение. Сбор плодов в России исчисляется миллионами тонн, а объемы лесозаготовок - десятками миллионов кубометров. Появление генетически модифицированных деревьев с новыми признаками, которые невозможно придать методами традиционной селекции, может значительно повысить эффективность как сельского, так и лесного хозяйства. Однако в силу биологических особенностей древесных растений эффективность их генетической трансформации значительно ниже, чем у травянистых растений.
Первая трансгенная лесная порода, гибрид тополя с осиной, была получена в 1987 году (Fillatti J.J., Sellmer J., McCown В., Haissig В., Comai L. Agrobacterium mediated transformation and regeneration of Populus. Mol. Gen. Genet. 1987. 206:192-199), а первая трансгенная плодовая культура, яблоня - в 1989 году (James D.J., Passey A.J., Barbara D.J. Genetic transformation of apple (Malus pumilu Mill.) using a disarmed Ti-binary vector. Plant Cell Rep. 1989. 7:658-661), то есть всего через несколько лет после получения первых трансгенных растений в 1983 году. Однако в настоящее время древесные трансгенные растения распространены несоизмеримо меньше, чем травянистые. В 2012 году трансгенные растения занимали в мире 170 млн. га (James С. 2012. Global Status of Commercialized Biotech/GM Crops: 2012. ISAAA Brief No. 44. ISAAA: Ithaca. NY) и из них на долю трансгенных деревьев, тополя и папайи, приходились, в лучшем случае, только десятки тысяч гектаров. Не в последнюю очередь такое положение дел связано с низкой эффективностью трансформации древесных растений.
Эффективность (частота) трансформации рассчитывается как отношение количества первичных трансгенных растений (для которых подтверждено наличие вставки в геноме) к количеству эксплантов, используемых для трансформации, и представляется в процентах. Трансформацию древесных растений (за исключением хвойных) осуществляют агробактериальным способом и на ее эффективность может влиять большое количество факторов, связанных как с растением и его культивированием, так и с агробактериями. Эти факторы включают генотип растения, тип экспланта, состав питательной среды, бактериальный штамм и вектор, повреждение растительной ткани, а также подавление и элиминацию агробактериальной инфекции после кокультивации.
Концентрация агробактерий на стадии кокультивации, во время которой осуществляется перенос Т-ДНК в геном растений, оказывает сильное влияние на частоту трансформации. Очень высокая концентрация агробактерий может или вызывать повреждение некоторых типов трансформируемых тканей, что приводит к снижению образования каллуса или регенерантов, или приводить к такой избыточной инфицированности эксплантов, с которыми будет невозможно справиться бактериостатическими антибиотиками, используемыми для элиминации агробактерий, или потребуются такие высокие концентрации антибиотиков, которые также могут вызвать гибель растительных тканей. С другой стороны, очень низкая концентрация агробактерий может стать причиной невысокой частоты трансформации. Например, при агробактериальной трансформации эмбриогенного каллуса риса при периоде кокультивации 1-3 дня было получено всего несколько трансгенных линий из-за недостаточного времени для завершения или достижения максимального уровня переноса Т-ДНК агробактерий в растительный геном, а период кокультивации в 5-6 дней также значительно снизил трансформацию из-за повреждений эксплантов, вызванных избыточным ростом агробактерий (Rahman Z.A., Seman Z.A., Basirun N., Julkifle A.L., Zainal Z., Subramaniam
S. Preliminary investigations of Agrobacterium-mediated transformation in indica rice MR219 embryogenic callus using gusA gene. African Journal of Biotechnology. 2011. 10: 7805-7813).
По этой причине очень важно выбрать правильную продолжительность кокультивации: не очень длительную, чтобы агробактерий не размножились слишком сильно, и не очень короткую, когда они трансформируют растительные клетки с низкой частотой. Это особенно важно для эксплантов древесных растений: из-за низкой частоты трансформации для них обычно увеличивают продолжительность кокультивации, а из-за низкой частоты регенерации их обычно выдерживают на среде для регенерации длительное время. В обоих случаях возникает риск неконтролируемого развития агробактерий.
Продолжительность кокультивации зависит от очень многих факторов: использующегося штамма агробактерий, типа и стадии растительной ткани, температуры, изначальной концентрации агробактерий, состава среды для культуры ткани и т.д. Все эти факторы могут варьировать в широких пределах и определить оптимальное время кокультивации довольно сложно.
Общепринятым является эмпирический способ определения продолжительности кокультивации и обычно он составляет от 2 до 6 дней. Например, известен способ агробактериальной трансформации одуванчика, который состоит из стадии преинкубирования эксплантов, стадии контактирования и кокультивации эксплантов с агробактериальными клетками и стадии культивирования эксплантов на селективной среде, причем период кокультивации продолжается около 2 дней (US Patent 7,026,529. Due et al. Methods for Agrobacterium-mediated transformation of dandelion. April 11, 2006). Известен также способ трансформации однодольных растений, который состоит из стадии инфицирования эксплантов агробактериями, стадии кокультивации на среде для кокультивации, стадию культивирования на среде покоя и стадию регенерации на среде для регенерации, причем стадия кокультивации продолжается от 1 до 7 дней (US Patent 8,404,930. Wu et al. Methods for improving monocot transformation. March 26,2013).
Недостатком этих способов является то, что продолжительность кокультивации определяется эмпирически и обычно остается неизменным при трансформации растений одного вида или даже рода и семейства, тогда как размножение агробактерий на эксплантах сильно зависит от ряда факторов - состояния эксплантов, исходной концентрации бактериальной суспензии, продолжительности инкубации эксплантов, температуры, состава питательной среды и т.д.
Задачей изобретения является повышение частоты трансформации путем использования индикатора размножения агробактерий во время кокультивации, показывающего предел, после которого ее необходимо завершить.
Поставленная задача решается благодаря тому, что экспланты помещаются на один слой фильтровальной бумаги с определенными характеристиками (размер пор 5-8 мкм) и появление на питательной среде под слоем фильтровальной бумаги колоний агробактерий в определенном количестве (под проекцией 10-20% эксплантов) служит сигналом прекращения стадии кокультивации.
Для трансформации используют растения, выбираемые из группы, включающей абрикос, айву, алычу, вишню, грушу, персик, сливу, черешню, яблоню, березу, бук, вяз, иву, каштан, клен, липу, осину, тополь, ясень.
Примеры в детальном описании приводятся для подвоя груши ГП 217, генотипа осины Pt и генотипа березы бп3ф1.
Однако перечень растений для использования в данном изобретении не ограничивается указанными растениями.
В качестве эксплантов для трансформации, в частности, используют листья груши и березы in vitro и междоузлия осины in vitro.
Примеры в детальном описании приводятся для маркерного гена gus, генов ксилоглюканазы Xeg, лакказы Lac, глутаминсинтетазы GS.
Возможность осуществления предлагаемого способа подтверждается представленными примерами, но не ограничивается ими.
Пример 1. Подготовка штаммов бактерий A.tumefaciens CBE21 (pCBE21, pBI121), A.tumefaciens CBE21 (pCBE21, pBI-Xeg), A.tumefaciens CBE21 (pCBE21, pBI-Lac) и A.tumefaciens CBE21 (pCBE21, pGS) для трансформации растений.
Для трансформации растений используют ночную культуру бактерий A.tumefaciens. Для этого 100 мкл суспензии клеток бактерий A.tumefaciens CBE21 (pCBE21, pBI121), A.tumefaciens CBE21 (pCBE21, pBI-Xeg), A.tumefaciens CBE21 (pCBE21, pBI-Lac) и A.tumefaciens CBE21 (pCBE21, pGS) добавляют к 50 мл жидкой среды LB, содержащей 50 мг/л канамицина, и инкубируют в течение ночи на термостатируемом орбитальном шейкере при 28°C и 120-150 об/мин, после чего центрифугируют полученную суспензию 5 минут при 4000 об/мин, осадок промывают жидкой средой MS и повторяют центрифугирование и промывание. После осаждения клеток их заливают 50 мл жидкой среды MS и ресуспендируют.
Пример 2. Трансформация растений груши клетками бактерий A.tumefaciens CBE21 (pCBE21, pBI121).
А) Подготовка растительного материала.
Для трансформации растений груши используют листовые экспланты с укорененных растений in vitro. Размножение культуры груши проводят на питательной среде, содержащей минеральные соли QL (Quoirin М., Lepoivre P. Improved medium for in vitro culture of Prunus sp. Acta Hort. 1977. 78: 437-442), витамины JS (Jacobini A., Standardi A. La moltiplicazione in vitro del melo cv. Wellspur. Rivisia della Ortoflorofrutticoltura Italiana. 1982. 66: 217-229), 1,5 мг/л БАП, 0,2 мг/л ИМК, 0,3 мг/л ГК, 100 мг/л мезоинозита, 20 г/л сахарозы и 7 г/л агара. Укоренение побегов груши проводят на питательной среде, содержащей минеральные соли QL с уменьшенным вдвое содержанием NH4NO3, витамины MS (Murashige Т., Skoog F. A revised medium for rapid growth and bioassays with tobacco tissue culture. Physiol. Plantarum. 1962. 15: 473-497), 0,5 мг/л ИМК, 10 г/л сахарозы и 7 г/л агара. Растения выращивают при фотопериоде 16/8 часов, температуре 22-24°C и освещенности 3000-3500 люкс.
Б) Генетическая трансформация эксплантов растений груши.
Для трансформации используют листья с укорененных растений, которые находятся на среде для укоренения в течение 1,5-2 месяцев. У листьев удаляют черешки и верхушки (у крупных листьев - также и боковые стороны) и наносят несколько надрезов перпендикулярно центральной жилке, не доводя их до краев листа. Подготовленные таким образом экспланты помещают на 40-50 минут в суспензию агробактерий, после чего осушают стерильными фильтрами и размещают на фильтрах, расположенных в чашках Петри на поверхности среды для кокультивации, содержащей минеральные соли NN (Nitsch J.P., Nitsch С. Haploid plants from pollen grains. Science. 1969. 163: 85-87), витамины JS, 3 мг/л ТДЗ, 0,5 мг/л НУК, 0,1 мг/л ГК, 100 мг/л мезоинозита, 100 мг/л гидролизата казеина, 30 г/л сахарозы и 7 г/л агара. В каждую чашку помещают по 10-15 эксплантов. Начиная с двух суток. периодически проверяют наличие колоний агробактерий на поверхности среды под проекцией эксплантов. Кокультивацию эксплантов с агробактериями проводят в темноте при температуре 23°C в течение 2 суток и более с 12-часовым интервалом при условии отсутствия на поверхности среды колоний агробактерий и при обнаружении колоний агробактерий под проекций 10-20, 50-60 и 90-100% эксплантов. После периода кокультивации экспланты переносят на среду для регенерации и селекции трансформантов того же состава, содержащую дополнительно 25 мг/л канамицина и 500 мг/л цефотаксима. На этой среде экспланты выдерживают в течении 4-6 субкультиваций по четыре недели в темноте при температуре 23°C. Регенерированные побеги пересаживают на среду для размножения, содержащую 25 мг/л канамицина и 250 мг/л цефотаксима.
Пример 3. Трансформация растений осины клетками бактерий A.tumefaciens CBE21 (pCBE21, pBI-Xeg).
А) Подготовка растительного материала.
Для трансформации растений осины используют междоузлия растений in vitro. Размножение культуры осины проводят на безгормональной питательной среде WPM (Lloyd G., McCown В.Н. Commercially feasible micropropagation of mountain laurel, (Kalmia latifolia) by use of shoot tip culture. Combined Proceedings of International Plant Propagators' Society. 1980. 30: 421-427), содержащей 1 мг/л витаминов MS, 30 г/л сахарозы и 7 г/л агара. Растения выращивают при фотопериоде 16/8 часов, температуре 22-24°C и освещенности 3000-3500 люкс.
Б) Генетическая трансформация эксплантов растений осины.
Для трансформации используют междоузлия растений возрастом 3-4 недели, нарезанные на сегменты 5-10 мм. Междоузлия помещают в 9 мл жидкой среды MSm и добавляют 1 мл агробактериальной суспензии. Инокуляцию проводят в течении 30 мин при слабом помешивании, после чего экспланты помещают на стерильную фильтровальную бумагу, немного подсушивают и размещают на фильтрах, расположенных в чашках Петри на поверхности среды для кокультивации, содержащей минеральные соли MSm, витамины MS, 0,5 мг/л 2,4-Д, 30 г/л сахарозы и 7 г/л агара. В каждую чашку помещают по 20 эксплантов. Начиная с двух суток, периодически проверяют наличие колоний агробактерий на поверхности средуыпод проекцией эксплантов. Кокультивацию эксплантов с агробактериями проводят в темноте при температуре 23°C в течение 2 суток и более с 12-часовым интервалом при условии отсутствия на поверхности среды колоний агробактерий и при обнаружении колоний агробактерий под проекций 10-20, 50-60 и 90-100% эксплантов. После периода кокультивации экспланты промывают в дистиллированной воде с добавлением 1 г/л цефотаксима в течение 20-30 минут и затем дважды в воде без цефотаксима. Отмытые экспланты подсушивают на фильтрах и переносят на среду для регенерации и селекции трансформантов того же состава, содержащую вместо 0.5 мг/л 2,4-Д, 0,5 мг/л зеатина и дополнительно 30 мг/л канамицина и 500 мг/л цефотаксима. На этой среде экспланты выдерживают в условиях 16-часового светового дня при 22-23°C с пересадкой каждые 4 недели. Регенерированные побеги пересаживают на среду для размножения, содержащую 30 мг/л канамицина и 250 мг/л цефотаксима.
Пример 4. Трансформация растений осины клетками бактерий A.tumefaciens CBE21 (pCBE21, pBI-Lac).
Трансформацию проводят аналогично примеру 3.
Пример 5. Трансформация растений березы клетками бактерий A.tumefaciens CBE21 (pCBE21, pGS).
А) Подготовка растительного материала.
Для трансформации растений березы используют листовые экспланты с растений in vitro. Размножение культуры березы проводят на питательной среде WPM, содержащей 0,6 мг/л БАП, 0,1 мг/л ИМК, 20 г/л сахарозы и 7 г/л агара. Растения выращивают при фотопериоде 16/8 часов, температуре 22-24°C и освещенности 3000-3500 люкс.
Б) Генетическая трансформация эксплантов растений березы.
Для трансформации используют листья с растений in vitro возрастом 1 месяц. У листьев удаляют черешки и верхушки (у крупных листьев - также и боковые стороны) и наносят несколько надрезов перпендикулярно центральной жилке, не доводя их до краев листа. Подготовленные таким образом экспланты помещают на 40-50 минут в суспензию агробактерий, после чего осушают стерильными фильтрами и размещают на фильтрах, расположенных в чашках Петри на поверхности среды для кокультивации, содержащей минеральные соли MS, 5 мг/л зеатина, 5 мг/л БАП, 0,2 мг/л ИМК, 30 г/л сахарозы и 7 г/л агара. В каждую чашку помещают по 10-15 эксплантов. Начиная с двух суток, периодически проверяют наличие колоний агробактерий на поверхности среды под проекцией эксплантов. Кокультивацию эксплантов с агробактериями проводят в темноте при температуре 23°C в течение 2 суток и более с 12-часовым интервалом при условии отсутствия на поверхности среды колоний агробактерий и при обнаружении колоний агробактерий под проекций 10-20, 50-60 и 90-100% эксплантов. После периода кокультивации экспланты промывают в дистиллированной воде с добавлением 1 г/л цефотаксима в течение 20-30 минут и затем дважды в воде без цефотаксима. Отмытые экспланты подсушивают на фильтрах и переносят на среду для регенерации и селекции трансформантов того же состава, содержащую дополнительно 50 мг/л канамицина и 500 мг/л цефотаксима. На этой среде экспланты выдерживают в условиях 16-часового светового дня при 22-23°C с пересадкой каждые 4 недели. Регенерированные побеги пересаживают на среду для размножения, содержащую 50 мг/л канамицина и 250 мг/л цефотаксима.
Пример 6. Полимеразная цепная реакция (ПЦР) на присутствие фрагмента последовательности гена gus.
Присутствие гена gus в трансгенных растениях груши подтверждают методом ПЦР с праймерами, специфичными для последовательности промотора CaMV 35S и внутренней части маркерного гена:
1) 5′-CTGCCGACAGTGGTCCCAAAGATGGACCC-3′
2) 5′-GAATCCTTTGCCACGCAAGTCCGCATCTT-3′.
Геномную ДНК из растений груши выделяют по Rogers and Bendich (Rogers S.O., Bendich A.J. Extraction of total cellular DNA from plants, algae and fungi. In: Plant Molecular Biology Manual. Gelvin S.B., Schilperoort R.A. (Eds.). Kluwer Academic Publisher, 1994. D1: 1-8). Для выделения используют листья растений in vitro (около 100 мг). Полученную растительную ДНК используют в качестве матрицы в ПЦР-анализах. Реакционная смесь содержит 16 мМ (NH4)2SO4, 0,01% BSA, 200 мкМ каждого дНТФ, 0,4 мкМ каждого олигонуклеотида, 0,05 единиц/мкл Taq полимеразы, 1-5 нг/мкл геномной ДНК. Реакцию проводят в объеме 50 мкл при следующих условиях: 96°C - 3 мин; 30 циклов: 94°C - 1 мин, 60°C - 1 мин, 72°C - 1 мин, затем 72°C - 5 мин.
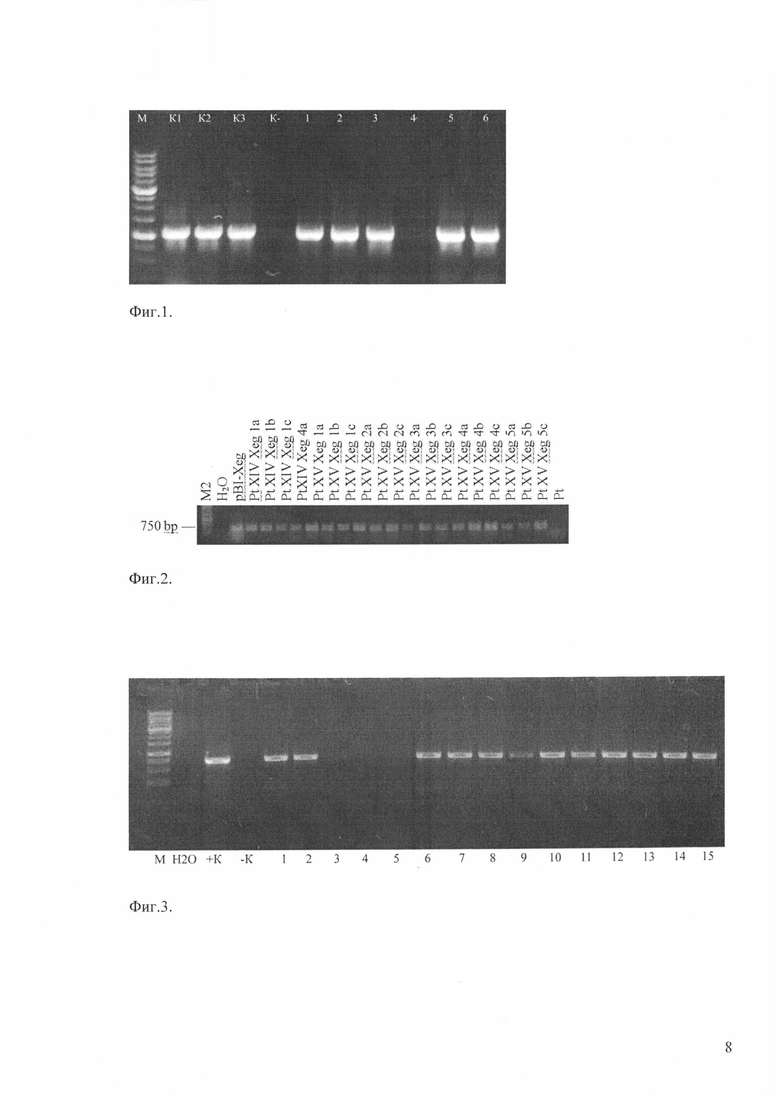
Продукты ПЦР анализируют в 1,8% агарозном геле с добавлением бромистого этидия. Гель фотографируют в ультрафиолете при длине волны 260-280 нм. Появление продукта ПЦР (ДНК размером 1149 н.п.) при использовании указанных праймеров, а также при условии отсутствия его в реакциях, поставленных на контрольной ДНК, свидетельствует о присутствии искомого гена в ДНК исследуемых растений (Фиг. 1).
Пример 7. Полимеразная цепная реакция (ПЦР) на присутствие фрагмента последовательности гена Xeg.
Присутствие гена Xeg в трансгенных растениях осины подтверждают методом ПЦР с праймерами, специфичными для кодирующей области трансгенной конструкции:
1) 5′-GAAATGGCTAATGCCACTACATT-3′
2) 5′-GATTTAGGCAACATCGGCAG-3′.
Геномную ДНК из растений осины выделяют по методу Rogers and Bendich (1994). Для выделения используют листья растений in vitro (около 100 мг). Полученную растительную ДНК используют в качестве матрицы в ПЦР-анализах. Реакционная смесь содержит 16 мМ (NH4)2SO4, 200 мкг/мл бычьего сывороточного альбумина, 200 мкМ каждого дНТФ, 0,8 мкМ каждого олигонуклеотида, 0,15 единиц/мкл Taq полимеразы, 1-5 нг/мкл геномной ДНК. Реакцию проводят в объеме 25 мкл при следующих условиях: 96°C - 3 мин; 30 циклов: 95°C - 45 сек, 62°C - 45 сек, 72°C - 1 мин, затем 72°C - 5 мин.
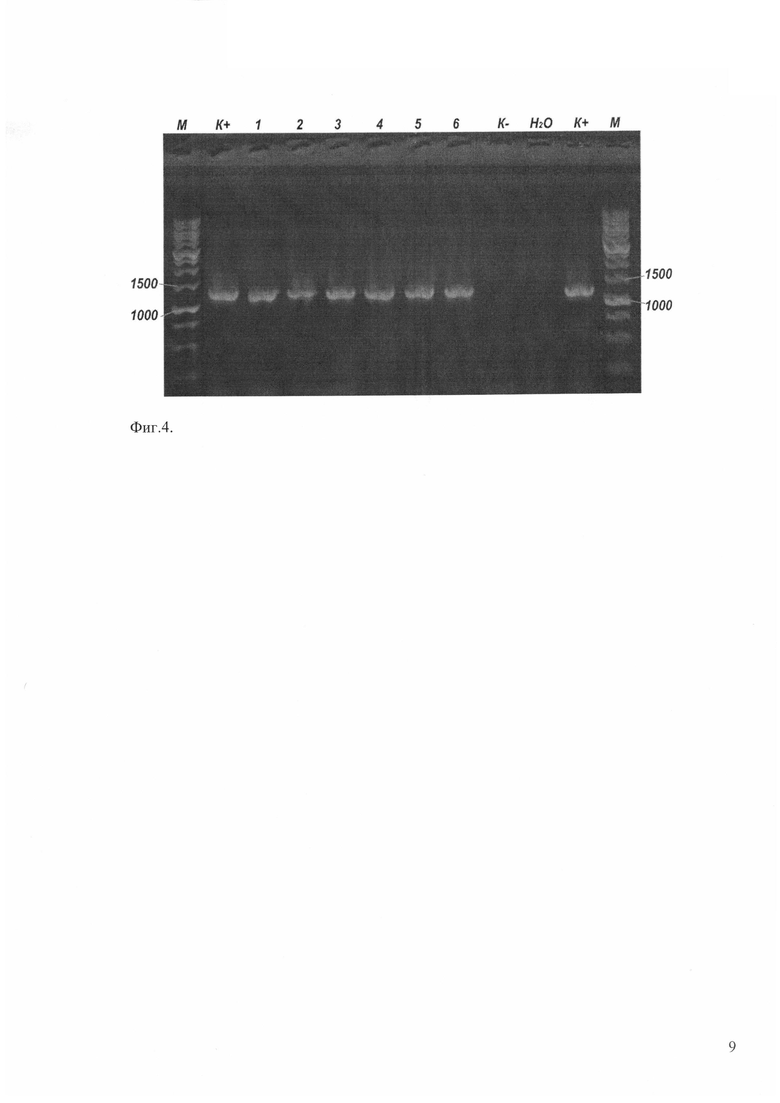
Продукты ПЦР анализируют в 1,8% агарозном геле с добавлением бромистого этидия. Гель фотографируют в ультрафиолете при длине волны 260-280 нм. Появление продукта ПЦР (ДНК размером 761 н.п.) при использовании указанных праймеров, а также при условии отсутствия его в реакциях, поставленных на контрольной ДНК, свидетельствует о присутствии искомого гена в ДНК исследуемых растений (Фиг. 2).
Пример 8. Полимеразная цепная реакция (ПЦР) на присутствие фрагмента последовательности гена Lac.
Присутствие гена Lac в трансгенных растениях осины подтверждают методом ПЦР с праймерами, специфичными для кодирующей области трансгенной конструкции:
1) 5′-TCATTGACAACTTGACGAAC-3′
2) 5′-GCAGGATAGCCGAGTTAATAC-3′
Геномную ДНК из растений осины выделяют по Rogers and Bendich (1994). Для выделения используют листья растений in vitro (около 100 мг). Полученную растительную ДНК используют в качестве матрицы в ПЦР-анализах. Реакцию проводят с использованием смеси ScreenMix (Евроген, Москва) в объеме 11 мкл при следующих условиях: 95°C - 5 мин; 30 циклов: 95°C - 45 сек, 60°C - 30 сек, 72°C - 1 мин, затем 72°C - 5 мин.
Продукты ПЦР анализируют в 1,8% агарозном геле с добавлением бромистого этидия. Гель фотографируют в ультрафиолете при длине волны 260-280 нм. Появление продукта ПЦР (ДНК размером 724 н.п.) при использовании указанных праймеров, а также при условии отсутствия его в реакциях, поставленных на контрольной ДНК, свидетельствует о присутствии искомого гена в ДНК исследуемых растений (Фиг. 3).
Пример 9. Полимеразная цепная реакция (ПЦР) на присутствие фрагмента последовательности гена GS.
Присутствие гена GS в трансгенных растениях березы подтверждают методом ПЦР с праймерами, специфичными для кодирующей области трансгенной конструкции:
1) 5′-TTATGAGCTCTTAGCTTATCAGTGACGCTGG-3′
2) 5′-CTCATCTAGACTGGTTTGTTTTGGAGAGTGGC-3′.
Геномную ДНК из растений березы выделяют по Rogers and Bendich (1994). Для выделения используют листья растений in vitro (около 100 мг). Полученную растительную ДНК используют в качестве матрицы в ПЦР-анализах. Реакционная смесь содержит 67 мМ Tris-HCl, pH 9.0, 16 мМ (NH4)2SO4, 2 мМ MgCl2 0,01% желатина, по 0,2 мМ каждого dNTP, 0,6 мкМ конечной концентрации каждого праймера и 0,2 единицы/мкл Taq полимеразы. Реакцию проводят в объеме 25 мкл при следующих условиях: 92°C - 3 мин; 35 циклов: 92°C - 20 сек, 62°C - 10 сек, 72°C - 1 мин, затем 72°C - 5 мин.
Продукты ПЦР анализируют в 1,8% агарозном геле с добавлением бромистого этидия. Гель фотографируют в ультрафиолете при длине волны 260-280 нм. Появление продукта ПЦР (ДНК размером 1190 н.п.) при использовании указанных праймеров, а также при условии отсутствия его в реакциях, поставленных на контрольной ДНК, свидетельствует о присутствии искомого гена в ДНК исследуемых растений (Фиг. 4).
Пример 10. Сравнение различных периодов кокультивации.
После инокуляции агробактериями экспланты груши, осины и березы находились на стадии кокультивации в течение различного времени: 1) до последней проверки на наличие на поверхности среды колоний агрообактерий, когда они еще не были обнаружены; 2) до момента обнаружения колоний агробактерий на поверхности среды под проекций 10-20% эксплантов; 3) до момента обнаружения колоний агробактерий на поверхности среды под проекций 50-60% эксплантов; 4) до момента обнаружения колоний агробактерий на поверхности среды под проекций 90-100% эксплантов. Регенеранты, полученные с эксплантов соответствующих вариантов, были проверены на наличие последовательностей чужеродных генов, как описано в примерах 6-9. Результаты анализа представлены в таблице 1.


Из представленных данных можно сделать вывод о наличии причинно-следственной связи между продолжительностью периода кокультивации, определяемой по развитию агробактерий под слоем фильтровальной бумаги с определенными характеристиками и эффективностью генетической трансформации древесных видов растений. Максимальная частота трансформации наблюдалась в случае, когда продолжительность стадии кокультивации продолжалась до момента появления на питательной среде колоний агробактерий под проекцией 10-20% эксплантов. Максимальная частота трансформации при указанном индикаторе развития агробактерий наблюдалась для различных видов древесных растений, различных типах эксплантов и различных плазмидных векторах для трансформации, что свидетельствует об универсальности предложенного способа.
Таким образом, изобретение позволяет повысить эффективность генетической трансформации древесных растений путем использования индикатора своевременного прекращения стадии кокультивации с агробактериями, что предотвращает как недостаточную, так и избыточную продолжительность стадии кокультивации.
Краткое описание чертежей
Фиг. 1. ПЦР-анализ трансгенных растений груши, содержащих ген gus. М -молекулярный маркер, К1 - ДНК плазмиды pBI121 (положительный контроль), К2, К3 - ДНК трансгенных растений (положительный контроль), -К - ДНК нетрансформированного растения (отрицательный контроль), 1-6 - ДНК трансформантов груши различных линий.
Фиг. 2. ПЦР-анализ трансгенных растений осины, содержащих ген Xeg. М2 - молекулярный маркер, H2O - отрицательный контроль реакции, pBI-Xeg - плазмидная ДНК (положительный контроль), Pt - ДНК нетрансформированного растения (отрицательный контроль), Pt XI PtXIVXeg…, PtXVXeg… - ДНК трансформантов осины различных линий.
Фиг. 3. ПЦР-анализ трансгенных растений осины, содержащих ген Lac. М - молекулярный маркер, H2O - отрицательный контроль реакции, +K - ДНК плазмиды pBI-Lac (положительный контроль), -К - ДНК нетрансформированного растения (отрицательный контроль), 1-15 - ДНК трансформантов осины различных линий.
Фиг. 4. ПЦР-анализ трансгенных растений березы, содержащих ген GS. М - молекулярный маркер, +К - ДНК плазмиды pGS (положительный контроль), -К - ДНК нетрансформированного растения (отрицательный контроль), H2O - отрицательный контроль реакции, 1-6 - ДНК трансформантов березы различных линий.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ получения генетически модифицированных древесных растений | 2015 |
|
RU2619175C1 |
| ТРАНСГЕННОЕ РАСТЕНИЕ БЕРЕЗЫ С ПОВЫШЕННОЙ ПРОДУКТИВНОСТЬЮ | 2013 |
|
RU2593721C2 |
| ТРАНСГЕННОЕ РАСТЕНИЕ ОСИНЫ С ПОВЫШЕННЫМ СОДЕРЖАНИЕМ ЦЕЛЛЮЛОЗЫ В ДРЕВЕСИНЕ | 2014 |
|
RU2599445C2 |
| ТРАНСГЕННОЕ РАСТЕНИЕ БЕРЕЗЫ С УСТОЙЧИВОСТЬЮ К ГЕРБИЦИДАМ | 2013 |
|
RU2587623C2 |
| ТРАНСГЕННОЕ РАСТЕНИЕ ОСИНЫ С ПОВЫШЕННОЙ ПРОДУКТИВНОСТЬЮ И МОДИФИЦИРОВАННОЙ ДРЕВЕСИНОЙ | 2013 |
|
RU2593722C2 |
| ТРАНСГЕННОЕ РАСТЕНИЕ ОСИНЫ С ПОНИЖЕННОЙ СКОРОСТЬЮ РАЗЛОЖЕНИЯ ДРЕВЕСИНЫ | 2015 |
|
RU2603081C2 |
| Трансгенное растение березы с ранним цветением | 2015 |
|
RU2619173C1 |
| СПОСОБ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ РАСТЕНИЙ С ПОВЫШЕННОЙ УСТОЙЧИВОСТЬЮ К ФИТОПАТОГЕНАМ | 2002 |
|
RU2261275C2 |
| ТРАНСГЕННОЕ РАСТЕНИЕ ОСИНЫ, УСТОЙЧИВОЕ К ГЕРБИЦИДАМ | 2013 |
|
RU2627189C2 |
| СПОСОБ ПОЛУЧЕНИЯ ФОРМ КАРТОФЕЛЯ in vitro, УСТОЙЧИВЫХ К ВОЗБУДИТЕЛЯМ ФИТОФТОРОЗА И АЛЬТЕРНАРИОЗА | 2013 |
|
RU2524424C1 |
Изобретение относится к области биотехнологии и генной инженерии и представляет собой способ получения генетически модифицированных древесных растений, включающий: а) получение и подготовку к инокуляции растительных эксплантов штамма А. tumefaciens, содержащего предназначенный для введения в растение селективный, маркерный и/или полезный гены; б) получение и подготовку к инокуляции штаммом A. tumefaciens растительных эксплантов; в) инокуляцию эксплантов штаммом A. tumefaciens в жидкой среде; г) кокультивацию эксплантов с агробактериями на среде без селективных агентов и бактериостатических антибиотиков; д) помещение эксплантов на среду, содержащую селективные агенты и бактериостатические антибиотики, с целью получения регенерантов непосредственно из тканей эксплантов или через стадию каллуса; е) отбор полученных на селективной среде регенерантов путем их тестирования на наличие полноразмерной копии целевого гена, где стадию кокультивации завершают при появлении на питательной среде под одним слоем фильтровальной бумаги с размером пор 5-8 мкм колоний агробактерий в количестве 10-20% растительных эксплантов. Изобретение позволяет определить оптимальный период кокультивации и повысить частоту генетической трансформации растений. 1 з.п. ф-лы, 4ил., 1 табл., 10 пр.
1. Способ получения генетически модифицированных древесных растений, включающий: а) получение и подготовку к инокуляции растительных эксплантов штамма А. tumefaciens, содержащего предназначенный для введения в растение селективный, маркерный и/или полезный гены; б) получение и подготовку к инокуляции штаммом A. tumefaciens растительных эксплантов; в) инокуляцию эксплантов штаммом A. tumefaciens в жидкой среде; г) кокультивацию эксплантов с агробактериями на среде без селективных агентов и бактериостатических антибиотиков; д) помещение эксплантов на среду, содержащую селективные агенты и бактериостатические антибиотики, с целью получения регенерантов непосредственно из тканей эксплантов или через стадию каллуса; е) отбор полученных на селективной среде регенерантов путем их тестирования на наличие полноразмерной копии целевого гена, отличающийся тем, что стадию кокультивации завершают при появлении на питательной среде под одним слоем фильтровальной бумаги с размером пор 5-8 мкм колоний агробактерий в количестве 10-20% растительных эксплантов.
2. Способ по п. 1, отличающийся тем, что древесные растения выбраны из группы, включающей абрикос, айву, алычу, вишню, грушу, персик, сливу, черешню, яблоню, березу, бук, вяз, иву, каштан, клен, липу, осину, тополь, ясень.
| ЛЕБЕДЕВ В.Г., генно-инженерные методы в селекции груши обыкновенной (PYRYS COMMUNIS L.) на устойчивость к биотическим и абиотическим факторам, автореферат диссертации, Москва, 2000, с.1-19 | |||
| ЛЕБЕДЕВ В.Г., и др, Генно-инженерные методы в селекции плодовых и ягодных культур, Плодоводство и ягодоводство России (Сборник научных работ), том XII, Москва, |

