Настоящее изобретение относится к иммуностимулирующим олигодезоксинуклеотидам (ODN), векторам и вакцинам, включающим такие олигодезоксинуклеотиды, к их применению в качестве лекарственного средства, к их применению для предупреждения инфекционного заболевания или борьбы с ним и к способам детектирования таких олигодезоксинуклеотидов.
На протяжении последних двух десятилетий в иммунологии выяснялось, что иммунная система позвоночных обладает механизмами для обнаружения микробной инфекции и запуска быстрой активации иммунной системы через посредство опосредуемого рецепторами распознавания уникальных характеристик патогенов, так называемых связанных с патогенами молекулярных структур (PAMP), взаимодействующих с когнатными, распознающими патогены рецепторами хозяина (PRR) (Iwasaki A, Medzhitov R. 2001; Science 327, 291-295. Medzhitov R., 2009. Immunity 30, 766-775).
В настоящее время очевидно, что некоторые формы дезоксирибонуклеиновой кислоты (ДНК) патогенов входят в число этих PAMP. В 1995 были представлены данные, что неметилированные CpG-мотивы в бактериальной ДНК запускают активацию B-клеток мыши (Krieg et al. 1995). Это исследование впервые сформировало связь между специфическим распознаванием бактериальной, иммуностимулирующей, содержащей неметилированные CpG ДНК и уже признанной супрессией CpG, а также широко распространенным метилированием CpG в ДНК млекопитающих. Было обнаружено, что самый эффективный, стимулирующий B-клетки, содержащий неметилированные CpG олигодезоксинуклеотид (CpG-ODN), обладает элементом последовательности GACGTT.
Следующая значительная статья в этой области была опубликована лабораторией Shizuo Akira в Осаке/Японии (Hemmi et al. 2000). С использованием клонирования гена и способа нацеленного выключения гена у мышей смогли определенно установить, что клеточную реакцию у мышей на CpG-ODN опосредует Toll-подобный рецептор 9 (TLR9). Впоследствии было обнаружено, что CpG-ODN являются агонистами в отношении передачи сигнала от TLR9 преимущественно через посредство пути ядерного фактора NF каппа-B (Medzhitov 2001). В последнее десятилетие было опубликовано немало исследований по темам основных исследований и по основным возможным иммунотерапевтическим применениям (обзоры которых приведены, например, в Krieg 2002, 2003, 2006; Klinman 2004, Vollmer 2005, Wilson et al. 2006, Kindrachuk et al. 2008, Dorn and Kippenberger 2008, Vollmer and Krieg 2009, Wilson et al. 2009). Ряд обзорных статей фокусируется на антиинфекционных применениях CpG-ODN (Krieg 2007), применении агонистов TLR9 при лечении рака (Krieg 2007, Weiner 2009), активации TLR9 для лечения астмы и аллергии (Kline 2007, Kline and Krieg 2008, Fonseca and Kline 2009) и в качестве адъювантов для вакцин (Klinman et al. 2004, Klinman 2006, Daubenberger 2007, Wagner 2009, Mutwiri et al. 2009, Klinman et al. 2009).
CpG-ODN также описаны и обсуждены в качестве иммуностимулирующих агентов и адъювантов для вакцин в применениях в ветеринарии, в частности, для жвачных животных, свиней, овец, собак, кур и рыб (Babiuk et al. 2003, Carrington and Secombes 2006, Griebel et al. 2005, Mutwiri et al. 2003, Singh and O′Hagan 2003, Werling and Jungi 2003).
В области применений в ветеринарии для кур описано использование содержащих CpG олигодезоксинуклеотидов, например, в вакцинах для защиты кур от пневмоэнцефалита (Linghua 2007). Недавно было обнаружено, что у кур TLR21 действует как функциональный гомолог TLR9 млекопитающих при распознавании содержащих CpG олигодезоксинуклеотидов (Brownlie et al., 2009).
Конструирование специфических CpG-ODN в качестве иммуномодуляторов до сих пор было довольно произвольным. Это особенно справедливо в отношении CpG-ODN не млекопитающих. Причина этого является многофакторной; прежде всего не известно о корреляции между иммуномодулирующими CpG-мотивами для TLR у человека и для TLR у не человека, не говоря уже о не являющихся млекопитающими видах. Во-вторых, нет в распоряжении клеточных систем с достаточно низким уровнем фона/шума, чтобы селективно исследовать эффекты очень низких концентраций CpG-ODN. Кроме того, нет в распоряжении способов с высокой пропускной способностью, и даже если бы они были, нет четкой корреляции между in vivo и in vitro эффективностями CpG-ODN в качестве иммуномодуляторов у не являющихся млекопитающими видов.
Таким образом, несомненно, существует потребность в новых CpG-ODN, которые оказывают большой иммуномодулирующих эффект и, следовательно, являются эффективными в низких дозах. И существует потребность в селективных и чувствительных системах отбора CpG-ODN для целей ветеринарии, которые демонстрируют корреляцию между in vitro и in vivo CpG-активностями.
Одной из задач настоящего изобретения является обеспечение таких новых CpG-ODN.
В связи с этим, один вариант осуществления настоящего изобретения относится к иммуностимулирующему неметилированному олигодезоксинуклеотиду, имеющему общую формулу:
5′[G]x{TCGTCG}nTCG[G]z 3′,
где n равен 2-100, x равен 3-20, и z равен 0-10,
или его фармацевтически приемлемой соли.
"Иммуностимулирующий неметилированный олигодезоксинуклеотид" относится к олигодезоксинуклеотиду, который содержит неметилированную динуклеотидную последовательность цитидин-фосфат-гуанозин, которая стимулирует инициацию каскадов передачи сигналов, приводящих к активации факторов транскрипции, таких как NF-κB или регулирующий интерфероны фактор 3 (IRF3). Именно эта активация в свою очередь приводит к экспрессии вовлеченных в воспалительный процесс цитокинов и к другим событиям активации клеток. Сайты связывания NF-κB и экспрессия генов, на которую влияет NF-κB, описаны, между прочим, Schindler and Baichwal (1994).
Термин "олигодезоксинуклеотид" означает короткий полимер из дезоксинуклеотидов; т.е. молекулу, включающую множество дезоксирибоз, связанных с фосфатной группой и сменяемым органическим основанием. Таким органическим основанием является замещенный пиримидин или замещенный пурин. Примерами являются цитозин и тимин относительно аденина и гуанина. Олигонуклеотиды согласно настоящему изобретению могут включать модификации. Примерами таких модификаций являются, например, модификации в фосфодиэфирном межнуклеозидном мостике, находящемся на 3′ и/или 5′ конце нуклеозида. Такие модификации относятся, между прочим, к замене фосфодиэфира, например, фосфоротиоатом или фосфородитиоатом.
Другими модификациями являются, например, замены фосфодиэфирного мостика дефосфо-мостиком. Примерами дефосфо-мостиков являются метилгидроксиламинная, формацетальная и диметиленсульфоновая группы.
Тем не менее, другими модификациями являются модификации, которые имеют отношение к замене природного нуклеозидного основания неприродным нуклеозидным основанием, таким как 5-фторцитоцин, 7-дезаза-7-замещенный гуанин, 7-дезаза-8-амещенный гуанин, 2-тиоурацил, дигидроурацил, 5-бромцитозин, 6-амещенные цитозины, N4-амещенные цитозины.
Кроме того, другими модификациями являются модификации, которые имеют отношение к замене сахарной единицы; сахара в виде β-рибозы или сахарной единицы в виде β-D-2′-рибозы модифицированной сахарной единицей, такой как, например, L-2′-дезоксирибоза или 2′-L-арабиноза.
Учебником, дающим дополнительное понимание олигонуклеотидов, является, например, "PCR Primer: A Laboratory Manual", Second Edition, 2003, Edited By Carl W. Dieffenbach, National Institute of Allergy and Infectious Diseases; Gabriela S. Dreksler, Uniformed Services University of the Health Sciences, Cold Spring Harbor Laboratory Press ISBN 978-087969654-2.
Структура 5′{TCGTCG}n 3′ содержащая CpG-мотив, представляет собой активную иммуностимулирующую составляющую ODN согласно настоящему изобретению. Следовательно, настоящим изобретением обеспечиваются иммуностимулирующие олигодезоксинуклеотиды, которые включают этот так называемый "остов".
Было обнаружено, что остов олигодезоксинуклеотида согласно настоящему изобретению, структура 5′{TCGTCG}n 3′, должен быть представлен по меньшей мере два, предпочтительно три раза. Следовательно, n должно быть равно по меньшей мере двум. Было также обнаружено, что активность олигодезоксинуклеотидов увеличивается при увеличении n. Этот эффект нивелируется, когда n увеличивается. По существу, число n остовной структуры должно быть, следовательно, равно по меньшей мере 2. Предпочтительно диапазоном n является 3≤n≤100, просто из-за того факта, что чем длиннее синтетическая последовательность, тем труднее ее создать. На практике диапазон n предпочтительно составляет 3≤n≤18. Более предпочтительно, диапазон n составляет 4≤n≤18, даже более предпочтительно диапазон n составляет 5≤n≤18, даже еще предпочтительнее диапазон n составляет 6≤n≤18.
Идентификация CpG-ODN согласно настоящему изобретению была обеспечена, между прочим, в результате использования более селективной системы детектирования, чем системы, используемые в настоящее время для обнаружения активации NF-κB. Brownlie и др. (2009), описывают репортерную систему на основе люциферазы для NF-κB. Другие системы основаны, например, на измерении транскрипта IL-8 или секреции цитокинов, или детектировании секреции NO.
В отличие от этого, в настоящем изобретении используется система детектирования на основе секретируемой щелочной фосфатазы (SEAP). SEAP является репортерным ферментом в системах млекопитающих (Yang et al., 1997). Эта система, как оказалось, является неожиданно чувствительной и, кроме того, неожиданно обеспечивает близкую корреляцию между in vitro и in vivo активностями проверенных CpG-ODN. Система на основе SEAP использовалась вместе с пара-нитрофенилфосфатом (pNPP) в качестве субстрата.
Другим усовершенствованием по сравнению с существующими системами было введение и прочное сохранение в клетках плазмиды, несущей ген SEAP. До сих пор во всех системах детектирования использовалась транзиторная трансфекция клеток геном-репортером. Именно благодаря введению и прочному сохранению в клетках гена-репортера теперь впервые удалось построить кривую зависимости ответной реакции от дозы. Такая кривая важна, если должно быть осуществлено надежное сравнение между активностями различных CpG-ODN.
Следовательно, способы и линии клеток, подробно описанные в разделе "Примеры" настоящего изобретения, впервые создают возможность для проведения надежного одновременного сравнения между различными CpG-ODN. Дополнительные детали использованной системы приведены в разделе "Примеры".
Было обнаружено, что увеличение числа G′ на 5′-конце структуры 5′[G]x{TCGTCG}nTCG[G]z 3′ приводит к увеличению активности CpG-ODN. Значение x должно быть равно по меньшей мере 3, но увеличение численности G вплоть до 20 G приводит к увеличению активности CpG-ODN. Поэтому более предпочтительно x равно 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20 в этом порядке возрастания предпочтения.
Было также обнаружено, что увеличение числа G′ на 3′-конце структуры 5′[G]x{TCGTCG}nTCG[G]z 3′ приводит к (небольшому) уменьшению активности CpG-ODN. Значение z должно быть равно 10 или меньше 10, но уменьшение числа G вплоть до 0 G приводит к увеличению активности CpG-ODN. Следовательно, более предпочтительно, когда z равно 9, 8, 7, 6, 5, 4, 3, 2, 1 или 0, в этом порядке возрастания предпочтения.
Как указано выше, возможны некоторые виды модификаций в фосфодиэфирном межнуклеозидном мостике, находящемся на 3′ и/или 5′ конце нуклеозида. Но по существу, в зависимости от пути синтеза, обычными распространенными типами связей между двумя нуклеотидами являются фосфодиэфирные (PDE) связи и фосфоротиоатные (PTO) связи. Для увеличения стабильности и иммуностимулирующего эффекта CpG-ODN структурные элементы синтетических олигодезоксинуклеотидов обычно обеспечены фосфоротиоатомами, так что они образуют PTO связи. Однако неожиданно было обнаружено, что когда только нуклеотиды 5′[G]x и 3′[G]z связаны PTO связями, а другие нуклеотиды связаны PDE связями, эффективность олигодезоксинуклеотида согласно настоящему изобретению дополнительно увеличивается. (В таких случаях связь 5′[G]x с {TCGTCG}nTCG[G]z 3′ является PTO связью, в то время как связь 5′[G]x{TCGTCG}nTCG с [G]z 3′ является PDE связью.)
Следовательно, другая предпочтительная форма этого варианта осуществления относится к олигодезоксинуклеотидам согласно настоящему изобретению, где нуклеотиды 5′[G]x и 3 [G]z имеют фосфоротиоатное связывание, а другие нуклеотиды имеют фосфодиэфирное связывание.
Как правило, содержащие CpG олигодезоксинуклеотиды активны в наномолярных количествах, как в in vitro системе исследования, так и in vivo. Однако неожиданно содержащие CpG олигодезоксинуклеотиды согласно настоящему изобретению даже активны в пикомолярных (субнаномолярных) количествах; их EC50 меньше 1 нМ. Полумаксимальной эффективной концентрацией (EC50) олигодезоксинуклеотида является количество олигодезоксинуклеотида, которое необходимо для индукции количества репортерного фермента SEAP (который создает окрашенный продукт с абсорбционной способностью при 405 нм) в репортерных клетках (HEK293-pNifty2-TLR21 кур или HD11-pNifTy2Hyg), которое дает полумаксимальную скорость ферментативной реакции. Если EC50 олигодезоксинуклеотида меньше 1 нМ в этих клетках, он считается активным в пикомолярных (субнаномолярных) количествах.
Вполне возможно соединить олигодезоксинуклеотид согласно настоящему изобретению с носителем или гаптеном, через реакционно-способную химическую группу. Такое соединение увеличивает иммуностимулирующий эффект комбинированных молекул. Простыми примерами таких компонентов являются, например, дигоксигенин, аминогексил-, техасский красный и биотин. Предпочтительные носители или гаптены представляют собой 3′- и 5′-меченый техасский красный и 5′-меченый дигоксигенин. Соединение олигодезоксинуклеотидов с гаптенами/носителями хорошо известно в данной области техники.
Другой вариант осуществления настоящего изобретения относится к вектору, включающему иммуностимулирующий неметилированный олигодезоксинуклеотид согласно настоящему изобретению. Таким вектором может быть молекула нуклеиновой кислоты, такая как плазмида, вирус, бактериофаг или любой другой вектор, используемый в молекулярной биологии. Просто в качестве примера: вектором, включающим иммуностимулирующий неметилированный олигодезоксинуклеотид, может быть, например, молекула ДНК, такая как плазмида, которая может увеличиваться в числе в бактериях, в которую был клонирован иммуностимулирующий неметилированный олигодезоксинуклеотид согласно настоящему изобретению. Предпочтительно такая плазмида имеет активное начало репликации, приводящее к присутствию большой численности плазмиды в хозяине. Выращивание таких бактерий в большом масштабе с последующим выделением плазмид обеспечивает альтернативу для синтетического получения иммуностимулирующего неметилированного олигодезоксинуклеотида согласно настоящему изобретению.
Одной из задач настоящего изобретения является обеспечение новых CpG-ODN, которые могут использоваться в качестве успешных иммуностимулирующих компонентов в вакцинах, которые предупреждают инфекционное заболевание или борются с ним, вместе с антигенным компонентом или генетической информацией, кодирующей антигенный компонент, и фармацевтически приемлемым носителем.
Как правило, термин "антигенный компонент" относится к составу материала, который включает по меньшей мере один эпитоп, который может индуцировать, стимулировать или усиливать иммунный ответ после введения человеку или животному.
Антигенный компонент может быть любым видом антигенного компонента, но предпочтительно происходит из микроорганизма или вируса, который в своей форме дикого типа является патогенным для людей или животных.
Антигенным компонентом может быть цельный патоген, предпочтительно в инактивированной или аттенуированной форме, экстракт из патогена или иммуногенный белок патогена. Если антигенным компонентом является иммуногенный белок патогена, то иммуногенный белок предпочтительно экпрессирован в in vitro культивируемых клетках и выделен из них.
Следовательно, другой вариант осуществления относится к вакцине для предупреждения инфекционного заболевания или борьбы с ним, отличающейся тем, что указанная вакцина включает иммуностимулирующее количество олигодезоксинуклеотида согласно настоящему изобретению и/или вектор согласно настоящему изобретению, иммуногенное количество антигенного компонента или генетической информации, кодирующей антигенный компонент, и фармацевтически приемлемый носитель.
Конечно, иммуностимулирующее количество олигодезоксинуклеотида и иммуногенное количество антигенного компонента сильно взаимосвязаны. Одним из преимуществ настоящего изобретения является то, что присутствие олигодезоксинуклеотида согласно настоящему изобретению может уменьшить количество антигенного компонента, которое необходимо для предупреждения инфекционного заболевания или борьбы с ним. Количество антигенного компонента, которое необходимо для предупреждения инфекционного заболевания или борьбы с ним, называют иммуногенным количеством антигенного компонента. Иммуностимулирующим количеством олигодезоксинуклеотида является количество, которое способно к уменьшению иммуногенного количества антигенного компонента, т.е. количества антигенного компонента, которое необходимо для предупреждения инфекционного заболевания или борьбы с ним. То есть, по существу, формулировки "иммуностимулирующее количество олигодезоксинуклеотида" и "иммуногенное количество" должны рассматриваться в зависимости друг от друга.
Само собой разумеется, что, если вакцина включает генетическую информацию, кодирующую антигенный компонент, количество антигенного компонента, экспрессируемого этой генетической информацией, должно быть достаточным для предупреждения инфекционного заболевания или борьбы с ним, т.е. оно должно быть иммуногенным количеством.
Тот факт, что неметилированные олигодезоксинуклеотиды согласно настоящему изобретению являются иммуностимулирующими, означает, что они увеличивают иммунологическую эффективность антигенных компонентов в вакцинах. По этой причине, вакцины согласно настоящему изобретению будут во многих случаях включать меньше антигенного компонента или генетической информации, кодирующей антигенный компонент, чем в случае, если олигодезоксинуклеотиды согласно настоящему изобретению не будут присутствовать. В некоторых случаях антигенный компонент как таковой, без добавления иммуностимулирующих олигонуклеотидов, может обладать такими низкими иммуногенными свойствами, что большие количества должны вводиться в любом случае, хотя и без достижения желаемого уровня иммуногенности. В таких случаях антигенный компонент может вводиться в обычной высокой концентрации, однако теперь вместе с олигодезоксинуклеотидом согласно настоящему изобретению, чтобы подобным образом достичь желаемого уровня иммуногенности.
Таким образом, количество антигенного компонента или генетической информации, кодирующей антигенный компонент, вводимого с олигонуклеотидом согласно настоящему изобретению, будет, как показывает опыт, равно или меньше количества, вводимого в отсутствие олигонуклеотида. Специалисту в данной области, участвующему в производстве конкретных вакцин, будет известно это количество для данной конкретной вакцины. Также в разделе "Примеры" дается, например, вполне достаточное указание в отношении количества антигенных компонентов, используемых, например, в трех различных вакцинах на основе инактивированных вирусов: вакцине против вируса пневмоэнцефалита птиц, вакцине против вируса инфекционного бронхита и вакцине против ринотрахеита индеек.
Количество олигодезоксинуклеотида согласно настоящему изобретению, которое необходимо вводить вместе с антигенным компонентом или генетической информацией, кодирующей антигенный компонент, зависит как от выбранного олигодезоксинуклеотида, так и от антигенного компонента. Очень подходящее количество олигодезоксинуклеотида согласно настоящему изобретению будет обычно варьировать от 1 до 100 наномолей. Очень хорошие in vivo результаты были получены, например, при использовании 1-10 мкг олигодезоксинуклеотидов согласно настоящему изобретению со средней длиной, составляющей 30 дезоксинуклеотидов, которые, как было обнаружено, являются активными в vitro исследованиях в наномолярном диапазоне. Если олигодезоксинуклеотид выбран из группы олигодезоксинуклеотидов, которые активны в пикомолярном диапазоне, специалисту в данной области будет понятно, что до проверки наномолярных количеств следует проверить количества, которые меньше, возможно гораздо меньше, 1 наномоля, т.е. пикомолярные количества.
Вакцины согласно настоящему изобретению включают фармацевтически приемлемый носитель. Природа этого носителя зависит, между прочим, от пути введения. Если введение осуществляют пероральным или интраназальным путем, носитель может быть настолько простым, как стерильная вода, физиологический солевой раствор или буфер. Если предпочтительным путем является инъекция, носитель должен быть предпочтительно изотоническим и удовлетворять особым требованиям в отношении pH, что делает его подходящим для инъекции. Однако такие носители широко известны в данной области техники.
Вакцины согласно настоящему изобретению могут, помимо антигенного компонента или генетической информации, кодирующей антигенный компонент, и олигодезоксинуклеотида согласно настоящему изобретению, включать адъювант. Как правило, адъювантами являются вещества, которые усиливают иммунный ответ хозяина неспецифическим образом. В данной области техники известно множество адъювантов, которые являются подходящими, такие как полный и неполный адъювант Фрейнда, витамин E, неионные блоксополимеры и полианионы, такие как декстран сульфата, карбопол и пиран, гидроксид алюминия. Также часто используемыми являются фосфат алюминия, сапонины, растительные масла, такие как токоферол и минеральные масла. Очень эффективными адъювантами являются эмульсии типа "масло-в-воде" и особенно эмульсии типа "вода-в-масле", далее также называемые адъювантами типа "масло-в-воде" и адъювантами типа "вода-в-масле". Такие эмульсии хорошо известны в данной области техники. Таким образом, предпочтительно вакцина включает адъювант типа "вода-в-масле".
Предпочтительно антигенный компонент представляет собой вирус или микроорганизм, который в своей форме дикого типа является патогенным для домашней птицы, или происходит из него.
Более предпочтительно, когда указанный вирус или микроорганизм выбирают из группы, состоящей из вируса инфекционного бронхита, вируса пневмоэнцефалита птиц, вируса инфекционного бурсита (болезни Гамборо), агента, вызывающего анемию у кур, реовируса птиц, Mycoplasma gallisepticum, вируса ринотрахеита индеек, Haemophilus paragallinarum (Coryza), поксвируса кур, вируса энцефаломиелита птиц, вируса синдрома снижения несучести, вируса инфекционного ларинготрахеита, вируса герпеса индеек, Eimeria spp., Ornithobacterium rhinotracheale, Pasteurella multocida, Mycoplasma synoviae, Salmonella spp. и Escherichia coli.
Кроме того, другой вариант осуществления настоящего изобретения относится к иммуностимулирующему неметилированному олигодезоксинуклеотиду согласно настоящему изобретению для применения в качестве лекарственного средства.
Кроме того, другой вариант осуществления настоящего изобретения относится к иммуностимулирующему неметилированному олигодезоксинуклеотиду согласно настоящему изобретению для применения в предупреждении инфекционного заболевания у домашней птицы или борьбы с ним.
До сих пор во всех системах детектирования использовалась транзиторная трансфекция клеток геном-репортером. Такие транзиторные системы не создают возможность для надежного одновременного сравнения эффективности CpG-ODN. Как сказано выше, основным усовершенствованием по сравнению с существующими системами было введение и прочное сохранение в клетках плазмиды, содержащей ген-репортер. Прочное означает, что плазмида остается находящейся в клетке после нескольких циклов деления клетки.
Часто прочного сохранения плазмиды достигают посредством выращивания клеток под давлением одного или более селективных агентов, таких как антибиотики, ген устойчивости к которым присутствует на плазмиде. Утрата плазмиды будет в таком случае приводить к гибели клетки, которая утратила плазмиду. Остающиеся жизнеспособные клетки будут все еще содержать плазмиду. Другим способом прочного сохранения будет трансфекция линеаризованной плазмидой. Такие плазмиды обычно интегрируются в геном клетки и поэтому прочно сохраняются. Таким образом, еще один вариант осуществления настоящего изобретения относится к клетке, включающей TLR21-рецептор и плазмиду, кодирующую ген-репортер для NF-κB, которая прочно сохраняется в клетке. Такие клетки очень подходят для применения при отборе содержащих CpG молекул, более конкретно отбора содержащих CpG молекул согласно настоящему изобретению. В разделе "Примеры" дается вполне достаточное указание в отношении того, как получить такую клетку, включающую плазмиду, кодирующую ген-репортер, которая может прочно сохраняться в клетке.
Как также отмечено выше, системы детектирования, основанные на секретируемой щелочной фосфатазе (SEAP), как обнаружено, очень подходят для используемой системы детектирования. Таким образом, предпочтительно геном-репортером является ген, кодирующий секретируемую щелочную фосфатазу.
По существу, любая клетка или линия клеток, содержащая и экспрессирующая TLR21, которая позволяет вводить и предпочтительно прочно сохранять плазмиду, несущую ген-репортер для NF-κB, предпочтительно ген SEAP, описанный выше, подходит для проверки TLR21-специфических CpG-ODN. Предпочтительным примером такой линии клеток, подходящей для проверки TLR21-специфических CpG-ODN, является линия клеток кур HD11. Следовательно, предпочтительно линией клеток для использования в системе детектирования является линия клеток HD11, включающая стабильную плазмиду, кодирующую ген-репортер.
Линии клеток кур, такие как линия клеток HD11, демонстрируют полный набор TLR кур. В определенных условиях это может порождать некоторую фоновую активность. По этой причине более предпочтительными линиями клеток являются линии клеток не домашней птицы, такие как линии клеток млекопитающих. Примером такой линии клеток млекопитающих является клетка HEK293, в которую был клонирован TLR21. Такая линия клеток является более специфически селективной в отношении TLR21-активирующих сигналов. По этой причине более предпочтительно, когда линией клеток для использования в системе детектирования является линия клеток млекопитающих HEK293, которая включает прочно сохраняемый ген-репортер и в которую был клонирован TLR21.
Еще один вариант осуществления настоящего изобретения относится к способу детектирования иммуностимулирующих олигодезоксинуклеотидов согласно настоящему изобретению, который включает стадии: a) приведения олигодезоксинуклеотида в контакт с клеткой согласно настоящему изобретению, b) определения уровня продукта гена-репортера.
В предпочтительной форме этого способа продуктом гена-репортера является SEAP.
Более предпочтительная форма данного варианта осуществления относится к способу детектирования иммуностимулирующих олигодезоксинуклеотидов согласно настоящему изобретению, в котором клеткой является клетка линии клеток кур HD11 или линии клеток HEK293, в которую был клонирован TLR21 кур.
Примеры
Пример 1
Клонирование гена и гетерологичная экспрессия TLR21 кур
Недавние успехи в поиске TLR у кур говорят о том, что TLR21 является функциональным гомологом TLR9 млекопитающих у видов птиц (Keestra 2008, Brownlie et al. 2009).
Основные положения клонирования гена TLR21
На основе последовательности NM_001030558 в базе данных Genbank была синтезирована пара праймеров для амплификации с помощью полимеразной цепной реакции (ПЦР) гена TLR21 кур:
Ga-TLR21-for1 (прямой праймер):
GAAGCTTACCATGATGGAGACAGCGGAGAAGGC
Ga-TLR21-rev1 (обратный праймер):
GGCGGCCGCTACATCTGTTTGTCTCCTTCCCTG
Праймеры были разработаны так, чтобы они обеспечивали рестрикционые сайты для клонирования (с подчеркиванием) и последовательность Козака (курсивный шрифт), фланкирующие инициирующий кодон и стоп-кодон (жирный шрифт). Используя эти праймеры и тотальную РНК кур в качестве матрицы, выполняли ОП-ПЦР. Продукт ПЦР ожидаемого размера (~3000 п.о.) клонировали в pCR2.1-Topo и секвенировали 5 независимых плазмидных клонов (P1, P2, P12, P13, P14).
Последовательность ДНК TLR21 кур, которая использовалась:
AAGCTTACCATGATGGAGACAGCGGAGAAGGCATGGCCCAGCACCAGGATGTGCCCCTCCCACTGCTGTCCACTCTGGCTGCTGCTGCTGGTGACAGTGACACTGATGCCGATGGTGCACCCGTATGGCTTTCGCAACTGCATTGAGGATGTCAAGGCACCTTTGTACTTCCGCTGCATCCAGCGCTTCCTGCAGTCGCCGGCCCTGGCAGTGTCTGACCTGCCACCACATGCCATCGCGCTCAATCTGTCATACAACAAAATGCGCTGCCTGCAGCCCTCTGCCTTTGCCCACCTGACACAGCTGCATACCCTGGACCTGACCTACAACCTCCTGGAGACCCTCTCCCCTGGTGCCTTCAATGGGCTGGGTGTGCTGGTGGTGCTGGACCTGTCTCACAACAAGCTGACCACACTTGCTGAAGGGGTGTTCAACAGCTTGGGCAACCTGTCCTCGCTGCAGGTACAACATAACCCCCTCAGCACGGTGTCACCAAGTGCTCTGCTACCCCTGGTCAACCTGCGCCGCCTGTCTCTACGGGGCGGGCGGCTGAATGGGTTGGGGGCAGTGGCAGTGGCAGTGCAGGGCTTGGCACAGCTGGAGCTGTTGGACCTATGTGAAAACAACCTGACAACGCTGGGGCCAGGCCCACCGCTACCCGCCTCGCTGCTCACCCTGCAGCTGTGCAACAACTCGCTGAGGGAGTTAGCGGGGGGCAGCCCGGAGATGCTATGGCACGTGAAGATACTCGACCTCTCCTACAACAGTATCTCACAGGCGGAGGTCTTCACCCAGCTCCACCTGCGCAACATCAGCCTGCTCCACCTGATCGGCAACCCCTTGGATGTCTTCCACCTGTTGGACATCTCTGACATCCAACCTCGCAGCCTGGATTTCTCTGGGTTGGTGCTGGGGGCTCAGGGGCTGGATAAGGTGTGCCTGAGGCTGCAGGGTCCCCAGGCCTTGCGGCGGCTGCAGCTACAACGCAACGGGCTGAAGGTGCTGCATTGTAATGCACTGCAGTTGTGTCCTGTGCTGAGAGAGCTGGACCTGTCCTGGAACCGGCTACAGCACGTGGGCTGTGCCGGCCGGCTGCTGGGCAAGAAGCAGCGGGAGAAGCTGGAAGTGCTGACAGTGGAACACAACCTGCTGAAGAAACTGCCGTCTTGCCTGGGGGCCCAGGTGCTGCCTCGGCTGTACAACATTTCCTTCCGCTTTAACCGCATCCTGACTGTTGGGCCCCAAGCCTTTGCCTACGCCCCGGCCCTGCAGGTGTTGTGGCTCAATATTAACAGCCTGGTGTGGCTGGACAGGCAGGCACTGTGGAGGCTGCACAACCTGACAGAGCTGCGCCTGGACAACAACCTGCTGACCGACCTCTATCACAACTCCTTCATTGACCTCCACAGACTGCGCACCCTCAACCTGCGCAACAACCGTGTCTCCGTCCTCTTCTCTGGTGTCTTCCAGGGGCTGGCTGAGCTGCAGACGCTGGATTTAGGGGGCAACAACTTGCGCCACCTGACTGCACAGTCACTGCAGGGGCTGCCCAAACTGCGCAGGCTGTACCTGGACCGCAACAGATTGCTGGAGGTGAGCAGCACTGTGTTCGCCCCAGTGCAGGCTACCCTGGGGGTGCTGGACCTGCGGGCCAACAACCTGCAGTACATCTCACAGTGGCTGCGCAAGCCGCCACCCTTCCGCAACCTGAGCAGCCTGTACGACCTGAAGCTGCAGGCGCAGCAGCCCTATGGACTGAAGATGCTGCCTCACTACTTCTTCCAGGGCTTGGTGAGGCTGCAGCAGCTGTCGCTGTCACAGAACATGCTGCGGTCCATCCCACCGGATGTCTTCGAGGACTTGGGCCAGCTGCGCTCCCTGGCATTGGCTGACAGCAGCAATGGGCTGCATGACCTGCCTGACGGCATCTTCAGAAACCTGGGCAACCTGCGGTTCCTGGACCTGGAGAATGCAGGGCTGCACTCGCTCACTCTGGAAGTCTTCGGCAATCTCAGCCGGCTGCAGGTGCTGCACTTGGCCAGAAACGAGCTGAAGACCTTCAATGACAGCGTTGCCAGCCGGCTGTCCTCCTTGCGCTACCTGGACCTGCGCAAGTGTCCGCTCAGCTGCACCTGTGACAACATGTGGCTGCAGGGCTGGCTGAACAACAGCCGTGTGCAGGTTGTCTACCCCTACAACTACACCTGTGGCTCACAGCACAATGCCTACATCCACAGCTTTGACACACACGTCTGCTTCCTGGACCTGGGGCTCTATCTCTTTGCTGGGACTGCACCGGCAGTGCTGCTGCTGCTGGTGGTGCCGGTGGTGTACCACCGCGCCTACTGGAGGCTGAAGTACCACTGGTACCTTCTGCGGTGCTGGGTCAACCAGCGGTGGCGGCGGGAGGAAAAGTGCTACCTCTATGACAGCTTTGTGTCCTACAATTCAGCTGATGAAAGTTGGGTGTTGCAGAAGCTGGTGCCTGAGCTGGAGCACGGTGCCTTCCGCCTCTGCTTGCACCACCGCGACTTCCAGCCGGGCCGCAGCATCATTGACAACATTGTGGATGCTGTCTACAACAGCCGGAAGACGGTGTGCGTGGTGAGCCGCAGCTACCTGCGCAGCGAGTGGTGCTCTCTAGAGGTGCAGTTGGCCAGCTACCGGCTGTTGGATGAGCGGCGTGACATCCTGGTACTGGTGCTGCTGGAGGACGTGGGTGATGCTGAGCTGTCTGCCTACCACCGCATGCGGCGGGTGCTGCTGCGGCGCACCTACCTGCGCTGGCCTCTTGACCCCGCAGCTCAGCCGCTCTTTTGGGCACGGCTGAAGAGGGCACTGAGGTGGGGAGAGGGAGGAGAGGAGGAGGAAGAAGAAGGTTTGGGTGGAGGGACGGGAAGGCCCAGGGAAGGAGACAAACAGATGTAGCGGCCGC
Трансфекция HEK293-pNifTy2-Zeo (клональной клеточной линии) плазмидой pcDNA3.1(+)-Neo-TLR21 кур
Клетки эмбриональной почки человека (HEK) 293 были созданы в 70-годах ХХ столетия в результате вирусной трансформации (Graham et al., 1977) и в настоящее время имеются в распоряжении научного сообщества благодаря депозитариям линий клеток, таким как ATCC.
pNifty2 является плазмидой, которая позволяет детектировать активацию транскрипционного фактора NFκВ, которая является признаком многих иммуностимулирующих действий, активаций Toll-подобных рецепторов среди них. Геном-репортером в pNifTy2, транскрипция/трансляция которого зависит от активации NFκВ, является секретируемая щелочная фосфатаза (SEAP). Детали описаны в спецификации от компании, продающей эту плазмиду: Invivogen. События трансформации/трансфекции с использованием pNifty2 отбирают как в бактериях, так и в клетках млекопитающих посредством добавления зеоцина в среды для роста.
Клетки HEK293 трансфицировали pNifTy2 с использованием стандартных способов (липофекции), отбирали стабильную линию клеток, функциональность осевой линии NFκВ/SEAP устанавливали посредством стимуляции с помощью фактора α некроза опухолей человека (Sigma). Секретируемую SEAP в супернатанте культуры стимулированных клеток определяли с помощью колориметрического исследования в титрационном микропланшете, в котором используется хромогенный субстрат пара-нитрофенилфосфат (pNPP, 5 мМ) в щелочном буфере (50 мМ NaHCO3, pH 9,6, 2 мМ MgCl2). Развитие цвета (λ=405 нм) контролировали считыванием на титрационном микропланшет-ридере. Это считывание данных также использовали для отбора клональных линий (методом предельного разведения) с высокими отношениями сигнала к шуму. Один из таких отобранных клонов (дублированный клон 11) затем использовали для дальнейших исследований с TLR21 кур.
pcDNA3.1(+)-neo представляет собой стандартный вектор для экспрессии в клетках млекопитающих, купленный у Invitrogen. Субклонирование гена TLR21 кур в этот вектор было осуществлено через фланкирующие рестрикционные сайты для Hind III (инициирующий кодон) и Not I (стоп-кодон), которые были введены с помощью ПЦР. (См. фиг.1).
Эту плазмиду затем трансфицировали (с помощью липофекции) в клональную линию HEK293-pNifty2-zeo, и рекомбинантные клетки отбирали посредством добавления и зеоцина, и G418 в среду для роста. Функциональность результирующей поликлональной рекомбинантной клеточной линии оценивали посредством стимуляции культуры с помощью ODN-X4 и ODN-HEK1-PTO и детектирования SEAP. Лучшие клональные линии затем идентифицировали методом предельного разведения с последующей стимуляцией и детектированием SEAP.
SEAP является репортерным ферментом в системах млекопитающих (Yang et al., 1997). SEAP является секретируемой формой щелочной фосфатазы эмбриона человека. Ее основными преимуществами являются высокая стабильность и очень высокая специфическая активность, которые обеспечивают чувствительность и надежность детектирования. Для обнаружения SEAP описано несколько субстратов, но был выбран экономичный и устойчивый pNPP, поскольку продукт его реакции пара-нитрофенолят детектируется с высокой чувствительностью (ε405=18500 M-1см-1). В своих схемах проверки авторы настоящего изобретения выполняют кинетические исследования, поскольку они обеспечивают более широкий динамический диапазон квантификации.
Клетки HEK293-pNifTy2-Zeo трансфицировали pcDNA3.1(+)-Neo-TLR21 кур (линеаризованной с помощью Pvu I), и поликлональную клеточную линию отбирали посредством пополнения среды 350 мкг/мл зеоцина и 600 мкг/мл G418. Проверку на функциональность проводили посредством стимуляции клеток с помощью ODN-X4 (PDE) и ODN-HEK1 (PTO). Секретируемую щелочную фосфатазу (SEAP) продуцировали отобранные клетки, но не родительская клеточная линия HEK293-pNifTy2-Zeo. Выполняли клонирование одной клетки, и отдельные клоны анализировали в отношении их способности реагировать на ODN-X4 (PDE) (GGGGGGTTCGTTTTCGTTTTCGTTGGGGG) и ODN-HEKl (PTO) (TCGTCGTTTTGTCGTTTGTCGTT).
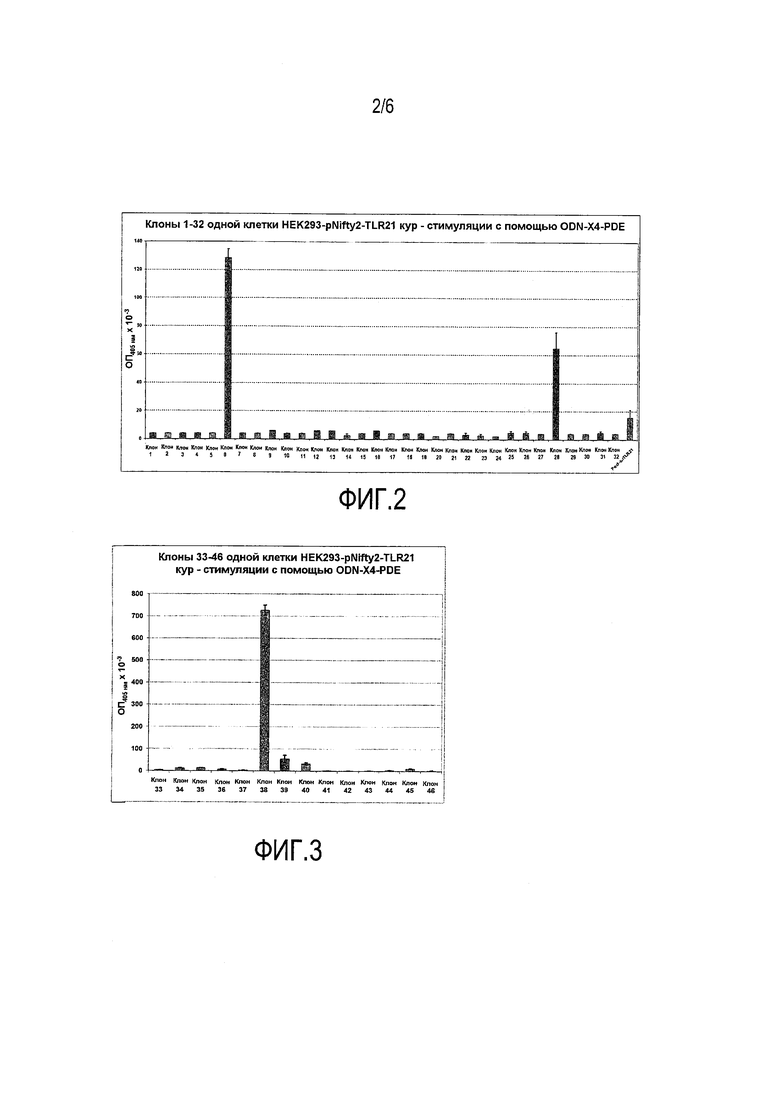
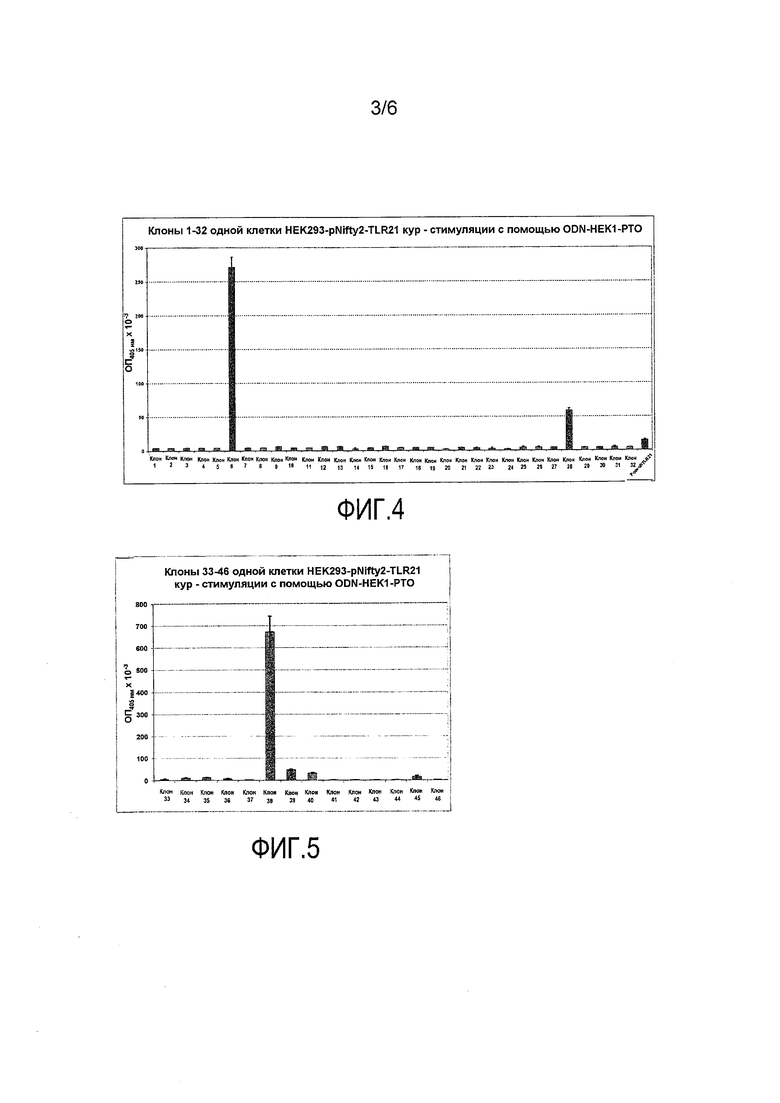
Из 46 клональных линий клеток, устойчивых как к зеоцину, так и к G418, только 3, несомненно, реагировали на ODN стимулы, в то время как 3-4 дополнительные клеточные линии продемонстрировали более слабые сигналы. 85% отобранных клонов были, следовательно, нефункциональными.
Для всех дальнейших исследований использовали клональную клеточную линию 38, которая порождала, вне всяких сомнений, самый высокий сигнал считывания SEAP в ответ на стимуляцию с помощью ODN-X4 (PDE) и ODN-HEKl (PTO).
Фиг.2-5 дают общее представление о SEAP активности различных клональных клеточных линий, устойчивых как к зеоцину, так и к G418.
Пример 2
В этом ряде гомополимеров, основанном на мотиве 5′[G]x{TCGTCG}nTCG[G]z 3′, 3′-dG отрезок укорочен до одного G в случае всех проверенных ODN.
Например: пример X23N-5 представляет собой 5′[G]x{TCGTCG}nTCG[G]z 3′, где x=7, z=0 и n=2: G7{TCGTCG}2TCG.
Например: пример X23N-7 представляет собой 5′[G]x{TCGTCG}nTCG[G]z 3′, где x=7, z=0 и n=3: G7{TCGTCG}3TCG.
X23N-5 GGGGGGGTCGTCGTCGTCGTCG
X23N-7 GGGGGGGTCGTCGTCGTCGTCGTCGTCG
X23N-9 GGGGGGGTCGTCGTCGTCGTCGTCGTCGTCGTCG
X23N-11 GGGGGGGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCG
X23N-13 GGGGGGGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCG
X23N-15 GGGGGGGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCG
X23N-17 GGGGGGGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCG
X23N-19 GGGGGGGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCG
X23N-21 GGGGGGGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCG
X23N-23 GGGGGGGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCG
X23N-25 GGGGGGGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCG
X23N-27 GGGGGGGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCGTCG
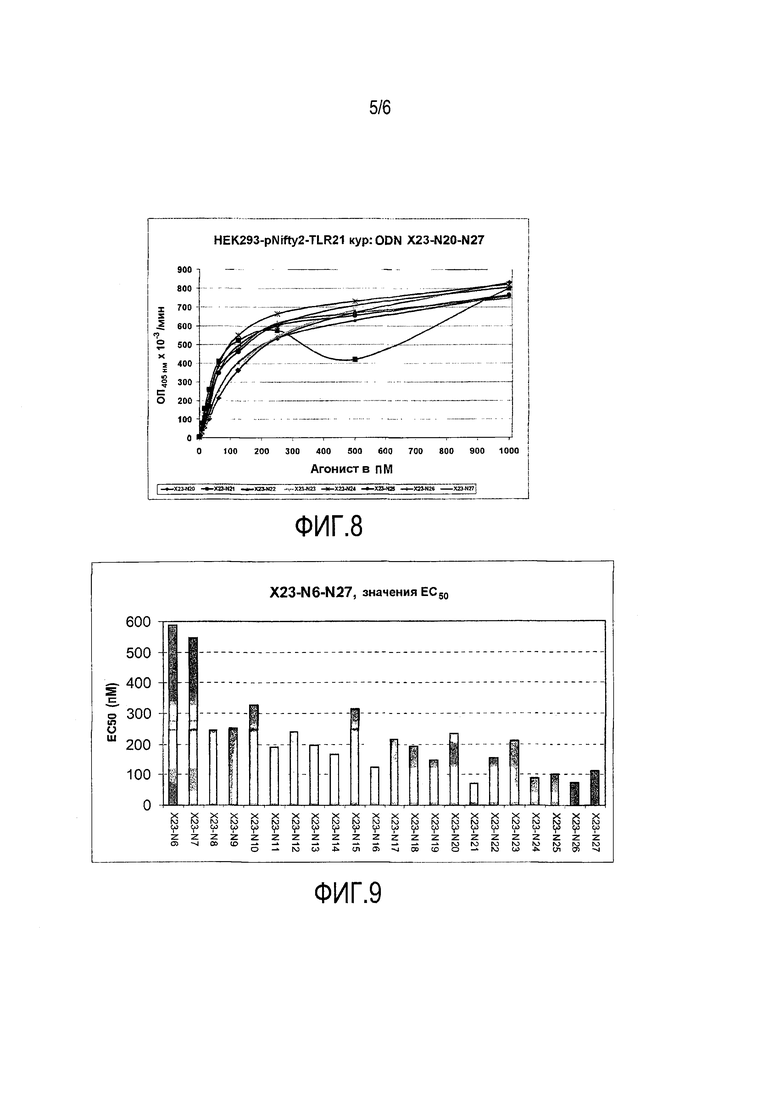
На фиг.6-8 продемонстрирован эффект X23-4-X23N-27 на индукцию количества окрашенного продукта с абсорбционной способностью при 405 нм репортерного фермента SEAP в репортерных клетках HEK293-pNifty2-TLR21 кур.
Как ни удивительно, на основе фиг.6 можно сделать вывод, уже в случае n=3, иммуностимулирующий эффект является неожиданно высоким. Этот эффект четко выражен количественно в неожиданно маленьком значении EC50 на фиг.9 (см. ниже).
Кроме того, на основе фиг.6-8 можно сделать вывод, что увеличение с n=3 до n=13 дает даже дальнейшее увеличение активности (= уменьшение EC50). Это увеличение демонстрирует довольно быстрое нивелирование после достижения значения n, равного 5.
На фиг.9 представлено значение EC50 для X23-4-X23N-27 в пикомолях.
Как можно видеть на фиг.9, уже при значении n=4, EC50 только немного выше 200 пМ, тогда как, начиная с n=10, значение EC50 достигает плато немного меньше 100 пМ.
Пример 3:
В этом ряде гомополимеров, основанном на мотиве 5′[G]x{TCGTCG}nTCG[G]z 3′, число 3′-G изменяли, в то время как n=4.
Были проверены следующие конструкции:
X23-six 5′-GGGGGGTCGTCGTCGTCGTCGTCGGGGG-3′
(=X23, стандарт 1)
X4-pent 5′-GGGGGGTTCGTTTTCGTTTTCGTTTTCGTTTTCGTTGGGGG-3′
(=стандарт 2)
X23-nine-3455 5′-GGGGGTCGTCGTCGTCGTCGTCGTCGTCGTCGGGGG-3′
(→ n=4, x=5, z=4)
X23-nine-3451 5′-GGGGGTCGTCGTCGTCGTCGTCGTCGTCGTCG-3′
(→ n=4, x=5, z=0)
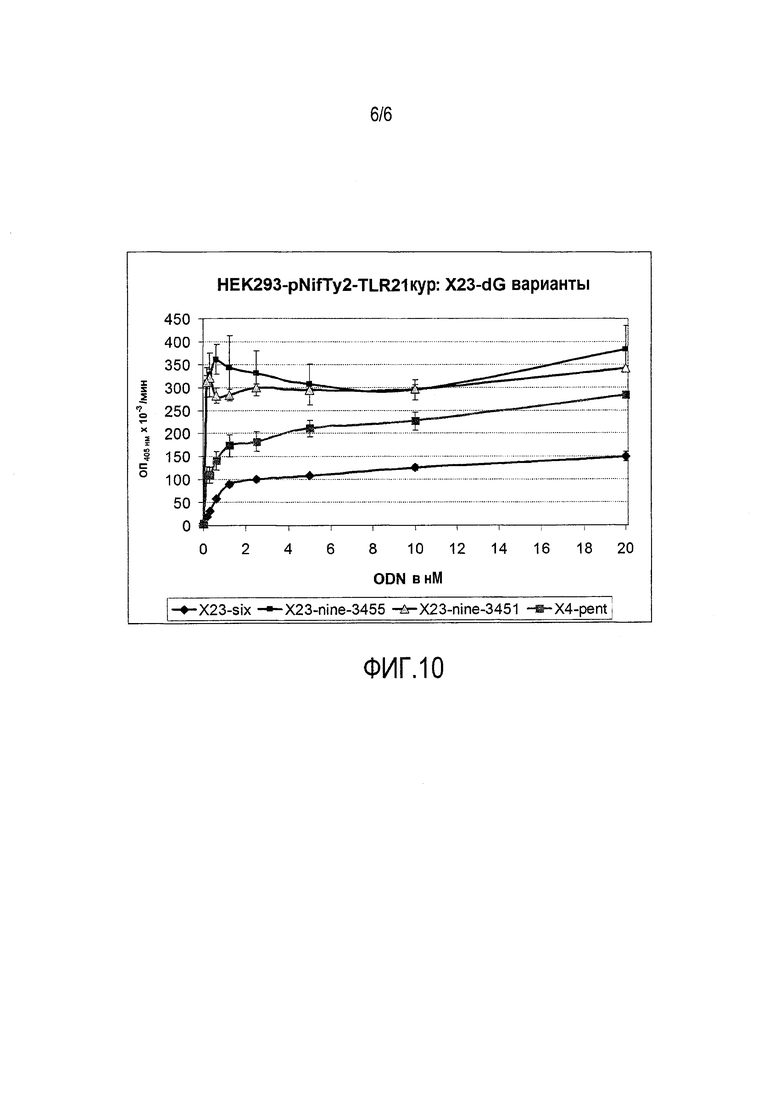
На фиг.10 продемонстрирован эффект изменения числа 3′-G на индукцию количества окрашенного продукта с абсорбционной способностью при 405 нм репортерного фермента SEAP в репортерных клетках HEK293-pNifty2-TLR21 кур.
Значения EC50 для этих CpG-мотивов являются следующими:
X4-pent (стандартный контроль): 340 пМ
X23-nine-3455 (n=4, x=5, z=4): <<100 пМ
X23-nine-3451 (n=4, x=5, z=0) <<100 пМ
На основе данной фигуры можно сделать вывод, что число 3′-G не является особенно важным, при условии, если z=4 или меньше 4, что касается EC50.
Подписи к фигурам:
Фиг.1: Карта плазмиды pcDNA3.1(+)-chiTLR21
Фиг.2-5: общее представление о SEAP активности различных клональных клеточных линий, устойчивых как к зеоцину, так и к G418.
Фиг.6-8: На этих фигурах представлен эффект X23-4-X23N-27 на индукцию количества окрашенного продукта с абсорбционной способностью при 405 нм репортерного фермента SEAP в репортерных клетках (HEK293-pNifty2-TLR21 кур).
Фиг.9: На этой фигуре представлены значения EC50 для X23-4-X23N-27 в пикомолях.
Фиг.10: На этой фигуре представлены значения EC50 для X4-pent (стандартного контроля), X23-six, X23-nine-3455 и X23-nine-3451.
Библиография:
Babiuk L.A., Gomis S., Hecker R., 2003. Molecular approaches to disease control. Poult. Sci. 82, 870-875.
Brownlie, R., Zhu J., Allan B., Mutwiri G.K., Babiuk L.A., Potter A., Griebel P., 2009. Chicken TLR21 acts as a functional homologue to mammalian TLR9 in the recognition of CpG oligodeoxynucleotides. Mol. Immunol. 46, 3163-3170.
Carrington A.C., Secombes C.J., 2006. A review of CpGs and their relevance to aquaculture. Vet. Immunol. Immunopathol. 112, 87-101.
Daubenberger C.A., 2007. TLR9 agonists as adjuvants for prophylactic and therapeutic vaccines. Curr. Opin. Mol. Ther. 9, 45-52.
Dorn A., Kippenberger S., 2008. Clinical application of CpG-, non-CpG-, and antisense oligodeoxynucleotides as immunomodulators. Curr. Opin. Mol. Ther. 10, 10-20.
Fonseca D.E., Kline J.N., 2009.Use of CpG oligodeoxynucleotides in treatment of asthma and allergic disease. Adv. Drug Deliv. Rev. 61, 256-262.
Graham, F.L., Smiley, J., Russell, W.C., Nairn, R., 1977. Characteristics of a human cell line transformed by DNA from human adenovirus type 5. J. Gen. Virol. 36, 59-74.
Griebel P.J., Brownlie R., Manuja A., Nichani A., Mookherjee N., Popowych Y., Mutwiri G., Hecker R., Babiuk L.A., 2005. Bovine toll-like receptor 9: a comparative analysis of molecular structure, function and expression. Vet. Immunol. Immunopathol. 108, 11-16.
Hemmi H., Takeuchi O., Kawai T., Kaisho T., Sato S., Sanjo H., Matsumoto M., Hoshino K., Wagner H., Takeda K., Akira S., 2000. A Toll-like receptor recognizes bacterial DNA. Nature 408, 740-745.
Iwasaki A, Medzhitov R. Regulation of adaptive immunity by the innate immune system. 2010. Science 327, 291-295.
Keestra A.M., 2008. Molecular dissection of the chicken Toll-like receptor repertoire. PhD thesis (Proefschrift), University of Utrecht, The Netherlands Kline J.N., 2007. Immunotherapy of asthma using CpG oligodeoxynucleotides. Immunol. Res. 39, 279-286.
Kline J.N., Krieg A.M., 2008. Toll-like receptor 9 activation with CpG oligodeoxynucleotides for asthma therapy. Drug News Perspect. 21, 434-439.
Klinman D.M., 2004. Immunotherapeutic uses of CpG oligodeoxynucleotides. Nat. Rev. Immunol. 4, 249-258.
Klinman D.M, Currie D., Gursel I., Verthelyi D., 2004. Use of CpG oligodeoxynucleotides as immune adjuvants. Immunol. Rev. 199, 201-216.
Klinman D.M., 2006. Adjuvant activity of CpG oligodeoxynucleotides. Int. Rev. Immunol. 25, 135-154.
Klinman D.M., Klaschik S., Sato T., Tross D., 2009. CpG oligodeoxynucleotides as adjuvants for vaccines targeting infectious diseases. Adv. Drug Deliv. Rev. 61, 248-255.
Kindrachuk J., Potter J., Wilson H.L., Griebel P., Babiuk L.A., Napper S., 2008.
Activation and regulation of toll-like receptor 9: CpGs and beyond. Mini Rev. Med. Chem. 8, 590-600.
Krieg A.M., Yi A,K., Matson S, Waldschmidt T,J., Bishop G.A., Teasdale R., Koretzky G.A., Klinman D.M., 1995. CpG motifs in bacterial DNA trigger direct B-cell activation. Nature 374, 546-549.
Krieg A.M., 2002. CpG motifs in bacterial DNA and their immune effects. Annu. Rev. Immunol. 20,709-760.
Krieg A.M., 2003. CpG motifs: the active ingredient in bacterial extracts? Nat. Med. 9, 831-835.
Krieg A.M., 2006. Therapeutic potential of Toll-like receptor 9 activation. Nat. Rev. Drug Discov. 5, 471-484.
Krieg A.M., 2007a. Anti-infective applications of toll-like receptor 9 agonists. Proc. Am. Thorac. Soc. 4, 289-294.
Krieg A.M., 2007b. Development of TLR9 agonists for cancer therapy. J. Clin. Invest. 117, 1184-1194.
Linghua Zhang et al., 2007. Vaccination with Newcatle disease vaccine and CpG oligodeoxynucleotides induces specific immunity and protection against Newcastle disease virus in SPF chicken. Vet. Immun. And Immunopath. 115, 216-222.
Medzhitov R., 2001. CpG DNA: security code for host defense. Nat. Immunol. 2, 15-16.
Medzhitov R., Approaching the asymptote: 20 years later. 2009. Immunity 30, 766-775).
Mutwiri G., van Drunen Littel-van den Hurk S., Babiuk L.A., 2009. Approaches to enhancing immune responses stimulated by CpG oligodeoxynucleotides. Adv. Drug Deliv. Rev. 61, 226-232.
Mutwiri G., Pontarollo R., Babiuk S., Griebel P., van Drunen Littel-van den Hurk S., Mena A., Tsang C., Alcon V., Nichani A., Ioannou X., Gomis S., Townsend H., Hecker R., Potter A., Babiuk L.A., 2003. Biological activity of immunostimulatory CpG DNA motifs in domestic animals. Vet. Immunol. Immunopathol. 91, 89-103.
Schindler, U., and Baichwal, V.R., 1994. Moll. Cell. Biol. 14: 5820-5831.
Singh M., O′Hagan D.T., 2003. Recent advances in veterinary vaccine adjuvants. Int. J. Parasitol. 33, 469-478.
Vollmer J., 2005. Progress in drug development of immunostimulatory CpG oligodeoxynucleotide ligands for TLR9. Expert Opin. Biol. Ther. 5, 673-682.
Vollmer J., Krieg A.M., 2009. Immunotherapeutic applications of CpG oligodeoxynucleotide TLR9 agonists. Adv. Drug Deliv. Rev. 61, 195-204.
Wagner H., 2009. The immunogenicity of CpG-antigen conjugates. Adv. Drug. Deliv. Rev. 61, 243-247.
Weiner G.J., 2009. CpG oligodeoxynucleotide-based therapy of lymphoid malignancies. Adv. Drug Deliv. Rev. 61, 263-267.
Werling D., Jungi T.W., 2003. TOLL-like receptors linking innate and adaptive immune response. Vet. Immunol. Immunopathol. 91, 1-12.
Wilson H.L., Dar A., Napper S.K., Marianela Lopez A., Babiuk L.A., Mutwiri G.K., 2006. Immune mechanisms and therapeutic potential of CpG oligodeoxynucleotides. Int. Rev. Immunol. 25, 183-213.
Wilson K.D., de Jong S.D., Tam Y.K., 2009. Lipid-based delivery of CpG oligodeoxynucleotides enhances immunotherapeutic efficacy. Adv. Drug Deliv. Rev. 61, 233-242.
Yang, T.T., Sinai, P., Kitts, P.A., Kain, S.R., 1997. Quantification of gene expression with a secreted alkaline phosphatase reporter system. Biotechniques 23, 1110-1114.
| название | год | авторы | номер документа |
|---|---|---|---|
| ИММУНОСТИМУЛИРУЮЩИЕ ОЛИГОДЕЗОКСИНУКЛЕОТИДЫ | 2012 |
|
RU2587633C2 |
| ИММУНОСТИМУЛИРУЮЩИЕ ОЛИГОДЕЗОКСИНУКЛЕОТИДЫ | 2011 |
|
RU2615457C2 |
| УСИЛЕНИЕ ИММУННОГО ОТВЕТА ПТИЦ, ИНДУЦИРОВАННОГО ВИРУСНЫМ ВЕКТОРОМ | 2014 |
|
RU2701805C2 |
| ВАКЦИННАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ ИММУНОСТИМУЛИРУЮЩИЕ ОЛИГОДЕЗОКСИНУКЛЕОТИДЫ | 2001 |
|
RU2413520C2 |
| КОМПЛЕКС, СОДЕРЖАЩИЙ ОЛИГОНУКЛЕОТИД, ОБЛАДАЮЩИЙ ИММУНОСТИМУЛИРУЮЩЕЙ АКТИВНОСТЬЮ, И ЕГО ПРИМЕНЕНИЕ | 2014 |
|
RU2723943C2 |
| ИММУНОСТИМУЛИРУЮЩИЕ ОЛИГОДЕЗОКСИНУКЛЕОТИДЫ | 2001 |
|
RU2293573C2 |
| ИММУНОСТИМУЛИРУЮЩИЕ ОЛИГОНУКЛЕОТИДЫ | 2009 |
|
RU2477753C2 |
| ИММУНОСТИМУЛИРУЮЩИЕ НУКЛЕИНОВЫЕ КИСЛОТЫ | 2000 |
|
RU2245149C2 |
| Иммуностимулирующие олигонуклеотиды | 2012 |
|
RU2610690C2 |
| CPG-ОЛИГОНУКЛЕОТИДНЫЕ АНАЛОГИ, СОДЕРЖАЩИЕ ГИДРОФОБНЫЕ Т-АНАЛОГИ С УСИЛЕННОЙ ИММУНОСТИМУЛИРУЮЩЕЙ АКТИВНОСТЬЮ | 2007 |
|
RU2477315C2 |







Настоящее изобретение относится к иммуностимулирующим олигодезоксинуклеотидам общей формулы:
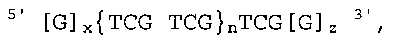
где х = 3-20, z = 0-10, n = 2-100,
и может быть использовано в медицине, конкретно в ветеринарии. Полученные иммуностимулирующие олигодезоксинуклеотиды или включающие их векторы и лекарственные средства могут быть использованы для предупреждения инфекционного заболевания у домашней птицы. Настоящее изобретение позволяет стимулировать активацию факторов транскрипции NF-kB и IRF3, а также повысить экспрессию вовлеченных в воспалительный процесс цитокинов, что способствует формированию активного иммунитета к различным инфекциям у домашней птицы. 5 н. и 4 з.п. ф-лы, 10 ил., 3 пр.
1. Иммуностимулирующий неметилированный олигодезоксинуклеотид, имеющий общую формулу
5' [G]x {TCG TCG}n TCG [G]z
3' ,
где
х = 3-20,
z = 0-10,
n = 2-100,
или его фармацевтически приемлемая соль,
который, необязательно, соединен с носителем или гаптеном.
2. Иммуностимулирующий неметилированный олигодезоксинуклеотид по п. 1, характеризующийся тем, что
х = 3-10;
z <5;
n = 3-18.
3. Олигодезоксинуклеотид по п. 1, в котором нуклеотиды 5′[G]x и 3′[G]z связаны с помощью фосфоротиоатной связи, а другие нуклеотиды связаны с помощью фосфодиэфирной связи.
4. Олигодезоксинуклеотид по любому из пп. 1-3, который соединен с носителем или гаптеном.
5. Вектор, включающий олигодезоксинуклеотид по любому из пп. 1-3, где указанный вектор способен экспрессировать указанный олигодезоксинуклеотид.
6. Вакцина для предупреждения инфекционного заболевания у домашней птицы, характеризующаяся тем, что указанная вакцина включает иммуностимулирующее количество олигодезоксинуклеотида по любому из пп. 1-4 и/или иммуностимулирующее количество вектора по п. 5, иммуногенное количество убитого или аттенуированного антигенного компонента или нуклеиновой кислоты, кодирующей антигенный компонент, и фармацевтически приемлемый носитель, где
указанный антигенный компонент представляет собой вирус или микроорганизм, который в своей форме дикого типа является патогенным для домашней птицы, или происходит из него.
7. Вакцина по п. 6, характеризующаяся тем, что указанный вирус или микроорганизм выбирают из группы, состоящей из вируса инфекционного бронхита, вируса пневмоэнцефалита птиц, вируса инфекционного бурсита (болезни Гамборо), агента, вызывающего анемию у кур, реовируса птиц, Mycoplasma gallisepticum, вируса ринотрахеита индеек, Haemophilus paragallinarum (Coryza), поксвируса кур, вируса энцефаломиелита птиц, вируса синдрома снижения несучести, вируса инфекционного ларинготрахеита, вируса герпеса индеек, Eimeria spp., Ornithobacterium rhinotracheale, Pasteurella multocida, Mycoplasma synoviae, Salmonella spp.и E.coli.
8. Применение иммуностимулирующего неметилированного олигодезоксинуклеотида по любому из пп. 1-4 в комбинации с иммунологическим количеством убитого или аттенуированного антигенного компонента или нуклеиновой кислоты, кодирующей антигенный компонент, который представляет собой вирус или микроорганизм, который в своей форме дикого типа является патогенным для домашней птицы, или происходит из него, для предупреждения инфекционного заболевания у домашней птицы.
9. Применение вектора по п. 5 в комбинации с иммунологическим количеством убитого или аттенуированного антигенного компонента или нуклеиновой кислоты, кодирующей антигенный компонент, который представляет собой вирус или микроорганизм, который в своей форме дикого типа является патогенным для домашней птицы, или происходит из него, для предупреждения инфекционного заболевания у домашней птицы.
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| Способ приготовления мыла | 1923 |
|
SU2004A1 |
| ВАКЦИННАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ ИММУНОСТИМУЛИРУЮЩИЕ ОЛИГОДЕЗОКСИНУКЛЕОТИДЫ | 2001 |
|
RU2413520C2 |

