Область техники, к которой относится изобретение
Настоящее изобретение относится к области биотехнологии и медицины и, в частности к иммуностимулирующим олигодезоксинуклеотидам (ODN) и фармацевтическим композициям, содержащим такие ODN.
Уровень техники
Вакцины могут спасти больше жизней (и ресурсов), чем любое другое медицинское вмешательство (Nossal, 1998). Благодаря всемирным программам вакцинации частота заболеваний многими смертельными болезнями резко сократилась. Хотя такое представление действительно для целого ряда болезней, например, туберкулеза, дифтерии, коклюша, кори и столбняка, не существует эффективных вакцин для многих инфекционных болезней, в том числе большинства вирусных инфекций, таких как СПИД. Также не существует эффективных вакцин для других болезней, инфекционных или неинфекционных, посягающих на миллионы жизней миллионов пациентов в год, включая малярию или рак. Кроме того, быстрое появление бактерий и микроорганизмов, устойчивых к антибиотикам, вызывает к жизни альтернативные способы лечения вакцинами, что является логичным выбором. Наконец, большая потребность в вакцинах также иллюстрируется тем фактом, что инфекционные болезни, а не сердечно-сосудистые нарушения или рак или травмы, остаются самой основной причиной смерти и потери трудоспособности в мире (Bloom and Widdus, 1998).
С иммунологической точки зрения, одна из основных проблем в области вакцин на сегодняшний день состоит в том, что традиционные вакцины (и/или иммуномодулирующие соединения, содержащиеся в таких препаратах) создаются для индукции высокого уровня антител (Harrow and Lane, 1988). Однако свои собственные антитела неэффективны при предупреждении большого ряда заболеваний, включая большинство болезней, вызываемых вирусами, внутриклеточными бактериями, некоторыми паразитами, и рак. Примерами таких болезней являются, но не ограничиваются перечисленным, болезни, вызванные вышеуказанным вирусом ВИЧ или видом Plasmodium в случае малярии. На многих экспериментальных системах показано, что в случае таких показаний важна клеточная ветвь иммунной системы, включая Т-клетки, а не гуморальная ветвь. Следовательно, требуются новые передовые технологии для преодоления ограничений традиционных вакцин. Необходимо сосредоточиться на технологиях, которые надежно индуцируют клеточную иммунную систему, включая антигенспецифические Т-клетки, узнающие молекулы, экспрессированные в клетках, зараженных патогенными микроорганизмами. В идеале, создаются вакцины, индуцирующие как Т-клетки, отличающие заболевшие и/или инфицированные клетки от здоровых клеток, так и одновременно антитела, секретированные В-клетками, узнающими патогенные микроорганизмы во внеклеточных компартментах.
Некоторые созданные вакцины состоят из живых аттенюированных организмов, где существует опасность реверсии к вирулентному штамму дикого типа. В частности, у хозяев с ослабленным иммунитетом это обстоятельство может стать сценарием с угрозой для жизни. С другой стороны, вакцины вводят в виде сочетания антигенов, происходящих от патогенного микроорганизма, и соединений, индуцирующих или усиливающих иммунные реакции против таких антигенов (такие соединения обычно называют адъювантами), так как субъединичные вакцины сами по себе, как правило, неэффективны.
Хотя нет сомнения в том, что вышеописанные вакцины важны для консервативного лечения, существует такой недостаток, что из-за своей сложности они могут вызывать серьезное побочное действие, например, на антигены, содержащиеся в вакцинах, что отражает перекрестную активность в отношении молекул, экспрессированных клетками вакцинированных индивидуумов. Кроме того, трудно удовлетворить существующие требования регулирующих органов, например, Всемирной организации здравоохранения (ВОЗ), Управления по санитарному надзору за пищевыми продуктами и медикаментами (FDA) и аналогичных европейских учреждений, в отношении строгих технических условий на состав вакцины и механизмы индукции иммунитета.
Антигенпрезентирующие клетки принадлежат к врожденной иммунной системе, развивающейся как первая линия защиты хозяина, которая своевременно ограничивает инфекцию после воздействия микроорганизмов (Hoffman et al., 1999). Клетки врожденной иммунной системы узнают конфигурации или относительно неспецифические структуры, экспрессированные на их клетках-мишенях, вероятнее, чем более сложные специфические структуры, которые узнает адаптивная иммунная система (Hoffman et al., 1999). Примерами клеток врожденной иммунной системы являются макрофаги и дендритные клетки, а также гранулоциты (например нейтрофилы), природные киллерные клетки и другие клетки. Напротив, клетки адаптивной иммунной системы узнают специфические антигенные структуры, включая пептиды, в случае Т-клеток, и пептиды, а также трехмерные структуры в случае В-клеток. Адаптивная иммунная система значительно специфичнее и сложнее, чем врожденная иммунная система, и улучшается после повторного воздействия данного патогенного микроорганизма/антигена. Филогенетически врожденная иммунная система значительно старше и может быть обнаружена уже в очень примитивных организмах. Тем не менее врожденная иммунная система является критическим фактором во время начальной фазы антигенного воздействия, так как кроме ограничения патогенных микроорганизмов, клетки врожденной иммунной системы, т.е. АПК (АРС), примируют клетки адаптивной иммунной системы и таким образом стимулируют специфические иммунные реакции, приводящие к клиренсу "незваных гостей". В итоге клетки врожденной иммунной системы, и в частности АПК, играют критическую роль во время фазы индукции иммунных реакций путем а) сдерживания заражения с помощью системы узнавания примитивных структур и b) примирования клеток адаптивной иммунной системы, приводящего к специфическим иммунным реакциям и запоминанию, результатом которых является клиренс вторгающихся патогенных микроорганизмов или других клеток-мишеней (Roitt et al., 1998). Указанные механизмы также могут быть важными для выведения или сдерживания опухолевых клеток.
Как указывалось выше, клетки врожденной иммунной системы узнают структуры, экспрессированные на их соответствующих мишенях. Примерами являются липополисахариды (ЛПС) в случае грамотрицательных бактерий, микобактериальные гликолипиды, липотейхоевые кислоты грамположительных бактерий, маннаны дрожжей и двухцепочечные РНК вирусов (Hoffman et al., 1999). Кроме того, они могут узнавать такие структуры, как с измененными гликозилированиями белков на опухолевых клетках.
Полученные в последнее время данные описывают ДНК простейших или низших эукариотов как дополнительную структуру, узнаваемую врожденной (но возможно, также адаптивной) иммунной системой млекопитающих (и, вероятно, большинства, если не всех, позвоночных) (Kreig, 1996; Lipford et al., 1998).
Иммунная система узнает низшие организмы, включая бактерии, из-за обычных структурных различий и различий в последовательностях между патогенным микроорганизмом и ДНК хозяина. В частности, мишенями являются короткие нити ДНК, образующиеся у беспозвоночных, или в форме коротких синтетических ODN, содержащих неметилированные динуклеотиды цитозин-гуанин (CpG) в контексте определенных оснований (Kreig et al., 1995). Мотивы CpG обнаруживаются с ожидаемой частотой в бактериальной ДНК, но со значительно меньшей частотой в ДНК позвоночных (Lipford et al., 1998; Pisetsky, 1999). Кроме того, мотивы CpG беспозвоночных (т.е. бактериальные) являются неметилированными, в то время как последовательности CpG позвоночных метилированы. Такие различия между бактериальной ДНК и ДНК позвоночных позволяют позвоночным узнавать ДНК беспозвоночного как сигнал опасности.
Природные ODN, содержащие CpG ДНК, а также тиофосфатзамещенные (с заменой фосфатных остатков на тиофосфатные) ODN, содержащие мотивы CpG (CpG-ODN), являются не только сильными активаторами пролиферации иммунных клеток и гуморальных иммунных реакций (Krieg et al., 1995), но также стимулируют сильные клеточные иммунные реакции (обзор в Lipford et al., 1998). ДНК/ODN, содержащие неметилированные мотивы CpG, могут непосредственно активировать моноциты (дендритные клетки, макрофаги) и В-клетки. Подобным образом, природные киллерные (NK) клетки непосредственно не активируются, но реагируют на EI-12 (интерлейкин 12), образованный моноцитами, с заметным повышением продуцирования ими IFN-γ (Chace et al., 1997). В результате индукция моноцитов и NK-клеток CpG-ДНК промотирует индукцию ответов типа Th-1 и рост цитотоксических Т-клеток.
Известно, что рибонуклеиновая кислота на основе инозина и цитозина, подобно полиинозиновой-полицитидиловой кислоте (поли-I:С), промотирует Th-1-специфические иммунные реакции. Известно, что она стимулирует продукцию цитокинов в макрофагах, таких как IL-1α и IL-12 (Manetti et al., 1995), она также известна как сильный индуктор интерферона типа 1 (Manetti et al., 1995) и как сильный стимулятор NK-клеток (Cavanaugh et al., 1996).
Однако такое действие строго ограничено рибонуклеиновой кислотой, содержащей инозиновые и цитидиновые остатки (WO 98/16247).
Раскрытие изобретения
Исследования авторов настоящего изобретения показали, что ODN, содержащие неметилированные мотивы CpG, хотя и эффективны при стимулировании иммунной системы, имеют существенные недостатки, особенно в отношении специфичности (высокий фон) и индукции побочного действия, такого как высокое системное образование TNF-α. Известно, что значительное системное высвобождение TNF-α вызывает токсический шок, который может вызвать смерть больного.
Поэтому целью настоящего изобретения являются подходящие новые ODN, не оказывающие такого сильного побочного действия как ODN на основе последовательностей CpG. Другой целью является снижение побочного действия фармацевтических композиций, содержащих известные ODN, и безопасные и эффективные хорошо переносимые фармацевтические композиции с эффективными иммуностимулирующими свойствами, подходящие для вакцинации животных, в частности млекопитающих, включая людей.
Такая цель достигается с помощью иммуностимулирующих олигодезоксинуклеиновых кислот (ODN), имеющих строение, согласно формуле (I)
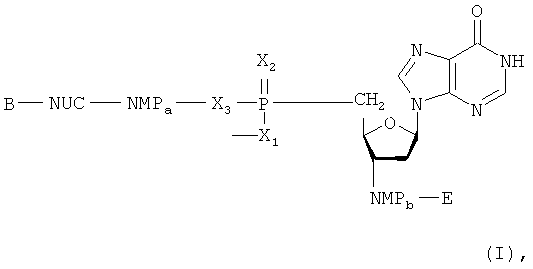
где любой Х представляет собой О или S,
и где
любой NMP представляет собой монофосфат или монотиофосфат дезоксинуклеозида, выбранный из группы, состоящей из дезоксиаденозин-, дезоксигуанозин-, дезоксиинозин-, дезоксицитозин-, дезоксиуридин-, дезокситимидин-, 2-метилдезоксиинозин-, 5-метилдезоксицитозин-, дезоксипсевдоуридин-, дезоксирибозопурин-, 2-аминодезоксирибозопурин-, 6-S-дезоксигуанин-, 2-диметилдезоксигуанозин- или N-изопентенилдезоксиаденозин-монофосфата или -монотиофосфата,
NUC представляет собой 2'-дезоксинуклеозид, выбранный из группы, состоящей из дезоксиаденозин-, дезоксигуанозин-, дезоксиинозин-, дезоксицитозин-, дезоксиуридин-, дезокситимидин-, 2-метилдезоксиинозин-, 5-метилдезоксицитозин-, дезоксипсевдоуридин-, дезоксирибозопурин-, 2-аминодезоксирибозопурин-, 6-S-дезоксигуанин-, 2-диметилдезоксигуанозин- или N-изопентенил-дезоксиаденозина,
а и b являются целыми числами от 0 до 100, при условии, что а+b равно величине от 4 до 150,
В и Е являются группами, обычными для 5'- или 3'-концов молекул нуклеиновых кислот.
Неожиданно оказалось, что ODN, содержащие дезоксиинозиновые остатки (I-ODN), показывают иммуностимулирующее действие, сравнимое с действием ODN, содержащих мотивы CpG, или во многих случаях даже его превосходящее. Кроме того, ODN, по настоящему изобретению, вызывают более специфические иммунные реакции на данный антиген или фрагмент антигена, чем CpG-ODN. Кроме того, ODN по настоящему изобретению, снижают индукцию вредных побочных реакций, в частности, индукцию системного TNF-α или IL-6.
В то время как описано определенное иммуностимулирующее действие инозинсодержащих молекул РНК, таких как поли-IC, или молекул, указанных в WO 98/16247, неожиданно оказалось, что хорошими иммуностимулирующими ODN могут быть молекулы дезоксинуклеиновых кислот, содержащие дезоксиинозиновые остатки.
Кроме того, I-ODN по настоящему изобретению, в отличие от ODN на основе специфического мотива CpG, не зависят от специфического мотива или палиндромной последовательности, как это описывается для CpG-олигонуклеотидов (см., например, ЕР 0468520А2, WO 96/02555, WO 98/18810, WO 98/37919, WO 98/52581, WO 99/51259 и WO 99/56755, включенные в данное описание в качестве ссылок). Следовательно, одна группа I-ODN, по настоящему изобретению, может, предпочтительно, содержать мотив CI (и, следовательно, ODN, описанные в указанных ссылках, где один или несколько остатков гуанозина заменены остатками дезоксиинозина, являются предпочтительными вариантами ODN настоящего изобретения). Это не является необходимостью для основного свойства иммуностимулирования, так как проявление иммуностимулирующих свойств I-ODN с инозином также не определяется связью с местоположением CI или IC.
Следовательно, I-ODN, по настоящему изобретению, представляет собой молекулу ДНК, содержащую остаток дезоксиинозина, находящийся, предпочтительно, в одноцепочечной форме.
I-ODN, по настоящему изобретению, можно выделить с помощью рекомбинантных методов или синтезировать химически. В последнем случае I-ODN, по настоящему изобретению, также может содержать модифицированные олигонуклеотиды, которые можно синтезировать с использованием стандартных химических трансформаций, такие как метилфосфонаты или другие модифицированные по фосфору олигонуклеотиды, такие как фосфотриэфиры, фосфоамидаты и фосфородитиораты. Также можно использовать другие модифицированные не по фосфору олигонуклеотиды (Stirchak et al., MAR, 17 (1989), 6129-6141), однако, предпочтительным из монофосфатов или монотиофосфатов для применения в настоящем изобретения является монофосфат 2'-дезоксинуклеозида.
NMP I-ODN, по настоящему изобретению, выбирают, предпочтительно, из группы, состоящей из дезоксиаденозин-, дезоксигуанозин-, дезоксиинозин-, дезоксицитозин-, дезоксиуридин-, дезокситимидин-, 2-метилдезоксиинозин-, 5-метилдезоксицитозин-монофосфата или -монотиофосфата (как правило, фосфатная или тиофосфатная группа находится в 5' дезоксирибозы). В то время как для ODN на основе мотива CpG существенно, что указанный мотив не метилирован, неожиданно, это несущественно в случае ODN, по настоящему изобретению, где, например, 2-метилдезоксиинозиновый или 5-метилдезоксицитозиновый остатки не оказывают отрицательного действия на иммуностимулирующие свойства ODN по настоящему изобретению. С другой стороны, в позиции 2 рибозогруппы вместо 2-дезоксиформ NMP могут присутствовать также другие, в особенности, инертные, группы, такие как, например, -F, -NH2, -СН3, в особенности, -СН3. Конечно, для ODN, по настоящему изобретению, исключается наличие групп -ОН и SH в позиции 2 рибозы, в особенности, для остатка рибозы в случае NMP, содержащего инозин.
Длина ODN, по настоящему изобретению, находится в интервале стандартных ODN, используемых согласно существующему уровню техники. Поэтому молекулы с общей длиной до 4 и свыше 150 показывают постепенное падение иммуностимулирующего потенциала. Предпочтительные ODN содержат от 10 до 60, в частности от 15 до 40, оснований (нуклеозидов), причем предполагается, что в таких предпочтительных вариантах a+b в формуле I составляет от 10 до 60, предпочтительно - от 15 до 40.
В то время как молекулы рибонуклеиновой кислоты, содержащие инозин и цитидин, описанные в уровне техники как иммуностимуляторы, являются крупными и относительно неидентифицированными полинуклеиновыми кислотами с молекулярной массой значительно более 200000 (коммерчески доступная полиинозиновая-полицитидиловая кислота от Sigma Chemicals имеет молекулярную массу, колеблющуюся от 220000 до 460000 (по меньшей мере 500-1000 остатков C+I), молекулы по настоящему изобретению являются молекулами ДНК значительно меньшей длины и установленной длины и состава, причем хорошо воспроизводятся в продуктах.
Также предпочтительно, когда содержащий дезоксиинозин NMP I-ODN, соответствующего формуле I, является монофосфатной группой с одним-четырьмя атомами серы, и когда также другие NMP, в частности - все другие NMP, присутствуют в виде монотиофосфатов нуклеозидов, поскольку такие ODN проявляют более высокую устойчивость к нуклеазам (в случае настоящего изобретения ясно, что "моно" в "монотиофосфат" относится к фосфату, т.е., что в каждом NMP присутствует одна фосфатная группа (один атом фосфора)). Предпочтительно, чтобы в NMP, согласно настоящему изобретению, по меньшей мере один из X1 и X2 представлял собой S, и по меньшей мере один из Х3 и Х4 представлял собой О. Предпочтительно, когда Х3 и Х4 представляют собой О. (Х3 может быть образован (из-за синтеза NMP) от фосфатной группы или от 3'-группы NMP-рибозы.)
Предпочтительно, ODN, по настоящему изобретению, содержат последовательность
hhh wdi dhh h
nhh hhh wdi nhh hhh hhh wn,
nhh wdi din hhh hdi ndi nh,
nhh hhh wdi dhh hhh hhh wn или
nhh wdi did hhh hdi ddi dh,
где
любой n представляет собой монофосфат или монотиофосфат 2'-дезоксинуклеозида, выбранный из группы, состоящей из дезоксиаденозин-, дезоксигуанозин-, дезоксицитозин- или дезокситимидин-монофосфата или монотиофосфата,
любой h представляет собой монофосфат или монотиофосфат 2'-дезоксинуклеозида, выбранный из группы, состоящей из дезоксиаденозин-, дезоксицитозин- или дезокситимидин-монофосфата или -монотиофосфата,
i представляет собой дезоксиинозинмонофосфат или -монотиофосфат,
любой w представляет собой монофосфат или монотиофосфат 2'-дезоксинуклеозида, выбранный из группы, состоящей из дезоксиаденозин- или дезокситимидин-монофосфата или -монотиофосфата, и
любой d представляет собой монофосфат или монотиофосфат 2'-дезоксинуклеозида, выбранный из группы, состоящей из дезоксиаденозин-, дезоксигуанозин- или дезокситимидин-монофосфата или -монотиофосфата.
Как описано выше, специфический мотив (такой как CpG или палиндром) не является необходимым для I-ODN по настоящему изобретению. Однако ODN, содержащие мотив CI, предпочтительны, так что в предпочтительном варианте воплощения изобретения ODN, соответствующий формуле I, содержит по меньше мере один 2'-дезоксицитозин-монофосфат или -монотиофосфат, 3'-соседний с 2'-дезоксиинозин-монофосфатом или -монотиофосфатом, для образования такого 5'-СI 3'-мотива.
Предпочтительные ODN, по настоящему изобретению, содержат одну или несколько последовательностей
gacitt,
iacitt,
gaictt,
iaictt,
где
а представляет собой дезоксиаденозин-монофосфат или -монотиофосфат,
g представляет собой дезоксигуанозин-монофосфат или -монотиофосфат,
i представляет собой дезоксиинозин-монофосфат или -монотиофосфат,
с представляет собой дезоксицитозин-монофосфат или -монотиофосфат, и
t представляет собой дезокситимидин-монофосфат или -монотиофосфат.
ODN, по настоящему изобретению, являются особенно подходящими для применения в области фармацевтики, например, для применения в качестве лекарства для животного или для человека. Они специфически адаптированы для того, чтобы действовать как иммуностимулирующее средство, в частности, вместе с вакцинами.
Следовательно, настоящее изобретение также относится к фармацевтической композиции, содержащей ODN по настоящему изобретению.
Так как предпочтительной фармацевтической композицией по настоящему изобретению является вакцина, такая композиция, кроме ODN, по настоящему изобретению, должна содержать антиген. Потенциал такого антигена для возникновения защитной/иммунной реакции у вакцинированного индивидуума сильно возрастает за счет сочетания его с ODN, по настоящему изобретению, в частности, из-за их иммуностимулирующей активности.
Вакцина может содержать самые разные антигены. Примерами таких антигенов являются полностью убитые организмы, такие как инактивированные вирусы или бактерии, грибы, простейшие или даже раковые клетки. Антигены также могут состоять из субфракций таких организмов/тканей, белков или, в их самой простой форме, из пептидов. Антигены также могут распознаваться иммунной системой в форме гликозилированных белков или пептидов, и также могут представлять собой или содержать полисахариды или липиды. Можно использовать короткие пептиды, так как, например, Т-цитотоксичные клетки (CTL) узнают антигены в форме коротких пептидов, как правило, длиной в 8-11 аминокислот, в сочетании с главным комплексом гистосовместимости (ГКГС, МНС) (Rammensee et al., Immunogenetics, 41 (1995), 178-228). В-Клетки узнают более длинные пептиды, длиной начиная примерно с 15 аминокислот (Harrow et al., Cold Spring Harbor: Cold Spring Harbor Laboratory (1988)). В отличие от Т-клеточных эпитопов, трехмерная структура В-клеточных антигенов также может быть важна для узнавания антителами. Для того, чтобы получить длительные антигенспецифические иммунные реакции, полезны адъюванты для запуска каскадов иммунных реакций, в которые, по необходимости, вовлекаются все клетки иммунной системы. В первую очередь адъюванты действуют, но не ограничиваются по способу такого своего действия, на так называемые антигенпрезентирующие клетки (АПК). Такие клетки, как правило, сначала сталкиваются с антигеном(-ами) с последующим представлением процессированного или немодифицированного антигена иммунному эффектору. Также могут участвовать промежуточные типы клеток. Только эффекторные клетки с соответствующей специфичностью активируются при мощной иммунной реакции. Адъювант также может локально удерживать антигены и другие привнесенные совместно факторы. Кроме того, адъювант может действовать как хемоатрактант для других иммунных клеток, или может действовать локально и/или системно как стимулятор для иммунной системы.
Согласно предпочтительному варианту воплощения изобретения, в качестве антигенов используют Т-клеточные эпитопы. С другой стороны, также предпочтительным может быть сочетание Т-клеточных эпитопов и В-клеточных эпитопов.
Антигены, используемые в настоящем изобретении, не являются критичным фактором. Конечно, согласно настоящему изобретению, также возможно использование смесей различных антигенов. Предпочтительно, в качестве таких антигенов используют белки или пептиды, полученные от вирусных или бактериальных патогенных микроорганизмов, или от грибов или паразитов (включая дериватизированные антигены или гликозилированные или липидированные антигены, или полисахариды или липиды). Другим предпочтительным источником антигенов являются опухолевые антигены. Предпочтительные патогенные микроорганизмы выбирают из числа вируса иммунодефицита человека (ВИЧ), вирусов гепатита А и В, вируса гепатита С (HCV), вируса саркомы Рауса (RSV), вируса Эпштейна-Барр (EBV), вируса гриппа, ротавируса, Staphylococcus aureus, Chlamydia pneumonias, Chlamydia trachomatis, Mycobacterium tuberculosis, Streptococcus pneumonias, Bacillus anthracis, Vibrio cholerae, вида Plasmodium (Pl. falciparum, Pl. vivax, и т.п.), вида Aspergillus или Candida albicans. Антигены также могут представлять собой молекулы, экспрессированные раковыми клетками (опухолевые антигены). Способ дериватизации может включать выделение белков в чистом виде из патогенных микроорганизмов/раковых клеток, инактивацию патогенного микроорганизма, а также протеолитическую или химическую дериватизацию или стабилизацию такого белка. На тех же условиях в фармацевтической композиции, по настоящему изобретению, также можно использовать опухолевые антигены (вакцины против рака) или аутоиммунные антигены. С такими композициями можно осуществлять вакцинацию против опухолей или лечение аутоиммунных заболеваний.
В случае пептидных антигенов в настоящее изобретение включают применение пептидных мимитопов/агонистов/ сверхагонистов/антагонистов или пептидов, измененных в некоторых позициях без воздействия на иммунологические свойства, или непептидных мимитопов/агонистов/ сверхагонистов/антагонистов (обзор в Sparbier and Walden, 1999). Пептидные антигены также могут содержать элонгации или по карбокси- или по аминоконцу пептидного антигена, облегчающие взаимодействие с поликатионным(и) соединением(ями) или иммуностимулирующим(и) соединением(ями). Для лечения аутоиммунных заболеваний можно применять пептидные антагонисты.
Антигены также можно дериватизировать с целью включения молекул, усиливающих презентацию антигена и направление антигена к антигенпрезентирующим клеткам.
В одном из вариантов воплощения изобретения фармацевтическая композиция служит для придания толерантности к белкам или белковым фрагментам и пептидам, которые принимают участие в аутоиммунных заболеваниях. Антигены, используемые в таких вариантах, служат для того, чтобы вызвать толерантность иммунной системы или иммунные реакции по типу отрицательной связи против эпитопов, принимающих участие в аутоиммунных заболеваниях.
Предпочтительно, фармацевтическая композиция по настоящему изобретению, в частности, в форме вакцины, также содержит поликатионный полимер, предпочтительно, поликатионный пептид, в частности, полиаргинин, полилизин, или антимикробный пептид.
Поликатионное(ые) соединение(я) для применения, согласно настоящему изобретению, могут представлять собой любое поликатионное соединение, демонстрирующее характерное действие согласно WO 97/30721. Предпочтительные поликатионные соединения выбирают из числа основных полипептидов, органических поликатионных соединений, основных полиаминокислот или их смесей. Такие полиаминокислоты должны иметь цепь длиной по меньшей мере в 4 аминокислотных остатка (см., тафтсин, как описывается в Goldman et al. (1983)). Особенно предпочтительными являются вещества, содержащие пептидные связи, подобные полилизину, полиаргинину, и полипептиды, содержащие свыше 20%, в частности, свыше 50%, основных аминокислот в ряду из более 8, в частности, более 20, аминокислотных остатков или их смеси. Другие предпочтительные поликатионные соединения и содержащие их фармацевтические композиции описываются в WO 97/30721 (например, полиэтиленимин) и WO 99/38528. Предпочтительно, такие полипептиды содержат от 20 до 50 аминокислотных остатков, в частности, от 30 до 200 остатков.
Такие поликатионные соединения можно получить химически или рекомбинантно, или их можно выделить из природных источников.
Катионные (поли)пептиды также могут представлять собой антибактериальные(микробные) пептиды со свойствами, рассматриваемыми в Ganz and Lehrer, 1999; Hancock, 1999. Такие (поли)пептиды могут быть прокариотического, животного или растительного происхождения, или могут быть получены химическими или рекомбинантными методами (Andreu and Rivas, 1998; Ganz and Lehrer, 1999; Simmaco et al., 1998). Пептиды также могут принадлежать к классу дефинзинов (Ganz, 1999; Ganz and Lehrer, 1999). Последовательности таких пептидов можно найти, например, в базе данных антимикробных последовательностей по адресу в Интернете http://www.bbcm.univ.trieste.it/~tossi/pagl.html.
Такие защитные пептиды хозяина или дефензивы также являются предпочтительной формой поликатионного полимера, согласно настоящему изобретению. Как правило, в качестве поликатионного полимера используют соединение, допускающее в качестве конечного продукта активацию (или регуляцию по типу отрицательной связи) адаптивной иммунной системы, предпочтительно, опосредуемую АПК (включая дендритные клетки).
Особенно предпочтительными для применения в качестве поликатионных веществ в настоящем изобретении являются антимикробные пептиды, образованные кателицидином, или их производные (А 1416/2000, включен в данное описание в качестве ссылки), в частности, антимикробные пептиды, образованные от кателицидина млекопитающих, предпочтительно - человека, коровы или мыши, или нейроактивные соединения, такие как гормон роста (человека).
Поликатионные соединения, полученные из природных источников, включают ВИЧ-REV или ВИЧ-ТАТ (дериватизированные катионные пептиды, пептиды antennapedia, хитозан или другие производные хитина) или другие пептиды, полученные из таких пептидов или белков биохимическими или рекомбинантными способами. Другими предпочтительными поликатионными соединениями являются кателин или родственные или образованные из кателина вещества. Например, мышиный кателин представляет собой пептид с аминокислотной последовательностью NH2-RLAGLLRKGGEKIGEKLKKIGOKIKNFFQKLVPQPE-СООН. Родственные кателину вещества или его производные содержат всю последовательность кателина или ее части с, по меньшей мере, 15-20 аминокислотными остатками. Дериватизация может включать замещение или модификацию природных аминокислот аминокислотами, которые не входят в число стандартных аминокислот. Кроме того, в такие молекулы кателина можно ввести дополнительные катионные остатки. Такие молекулы кателина предпочтительны для сочетания с антигеном и иммуномодулирующим ODN по настоящему изобретению. Однако неожиданно такие молекулы кателина оказались также эффективными в качестве адъюванта для антигена без добавления других адъювантов. Поэтому возможно применение таких молекул кателина как эффективных адъювантов в композициях вакцин без других иммуноактивирующих веществ или с ними.
Другим предпочтительным поликатионным веществом для применения, согласно настоящему изобретению, является синтетический пептид, содержащий по меньшей мере 2 мотива KLK, разделенных линкером из 3-7 гидрофобных аминокислот (А 1789/2000, включен в данное описание в качестве ссылки).
Стало неожиданностью, что иммуностимулирующее действие фармацевтической композиции, по настоящему изобретению, оказалось существенно выше, чем можно было ожидать от сложения действия каждого из отдельных компонентов или даже сложения действия ODN или поликатионного соединения с действием антигена.
В и Е в формуле I представляют собой группы, обычные для 5'- и/или 3'-концов молекул нуклеиновых кислот. Примеры таких групп хорошо известны специалистам в данной области техники (см., например, "Oligonucleotides and Analogues - A Practical Approach" (1991), ed. Eckstein, Oxford University Press). В случае I-ODN по настоящему изобретению В и/или Е выбирают, предпочтительно, из числа -Н, -СН3, -СОСН3, -ОН, -СНО, фосфатной, тиофосфатной, сульфатной или тиосульфатной группы, или фосфоалкильной группы, в частности, с алкилом C1-C6, и/или концевой аминогруппы (аминогруппу можно использовать, например, для дополнительного мечения I-ODN по настоящему изобретению, например, -PO4-(CH2)n-NH2 или -РO4-(СН2)n-NН-метка). Особенно предпочтительно, когда В являются нуклеозидами, в частности, 2'-дезоксинуклеотидами, указанными выше (т.е. без фосфатной или тиофосфатной группы). С другой стороны, такие группы также могут содержать линкерные группы для других молекул, в частности молекул носителя или меток. В таких формах ODN, где ODN связываются с твердыми поверхностями или частицами или метками, такие поверхности, частицы или метки, и т.п., затем также являются частью групп В и/или Е.
Конечно, любая ионизованная форма (соль) или таутомерные формы молекул формулы I охватываются данной формулой I.
Фармацевтическая композиция, по настоящему изобретению, также может содержать другие активные ингредиенты (фармацевтически активные вещества), в частности, вещества, полезные в связи с вакциной. Предпочтительными вариантами таких других активных ингредиентов являются цитокины, противовоспалительные вещества, антимикробные вещества или их сочетания.
Конечно, фармацевтическая композиция, по настоящему изобретению, также может содержать вспомогательные вещества, в частности, фармацевтически приемлемый носитель, буферные вещества, стабилизаторы или их сочетания.
Относительные количества ингредиентов в фармацевтической композиции по настоящему изобретению весьма зависят от потребностей в отдельном антигене и от животного/человека, к которому должна применяться данная композиция. Поэтому фармацевтическая композиция, по настоящему изобретению, содержит, предпочтительно, один или несколько ODN предпочтительно, в количестве от 1 пг до 10 г, предпочтительно - от 1 нг до 1 г, предпочтительнее - от 100 нг до 10 мг, в частности - от 10 мкг до 1 мг. Антиген, как и поликатионный полимер, можно применять в подобных дозах, предпочтителен интервал от 1 до 10000 мг антигена и от 0,1 до 1000 мг поликатионного соединения на вакцинацию.
Композиции настоящего изобретения можно применять к пациенту, например, кандидату на вакцинацию, в эффективных количествах, например, с интервалами в одну неделю, в две недели или в месяц. Пациенты, которых лечат композициями настоящего изобретения, также могут быть вакцинированы повторно или только один раз. Предпочтительным применением настоящего изобретения является активная иммунизация, в частности, людей или животных, без защиты от специфического антигена.
Способ введения композиции настоящего изобретения не является критичным фактором, например, подходящим является подкожное, внутримышечное, интрадермальное или трансдермальное введение, а также оральное введение.
Композицию настоящего изобретения также можно применять раздельно, например, инъецируя иммуностимулирующее вещество отдельно от антигенной/поликатионной композиции. Следовательно, настоящее изобретение также относится к набору, содержащему в качестве одного компонента композицию, содержащую антиген и поликатионный полимер, и в качестве второго компонента - композицию, содержащую иммуностимулятор или хемотактическое вещество.
Компоненты можно применять в одном и том же месте или в одно и то же время, однако, также возможно применение в разных местах или в разное время, или в течение разного периода времени. Также возможны варианты системного или местного применения композиций или компонентов, соответственно.
Детали настоящего изобретения описываются с помощью приведенных далее примеров и фигур, но изобретение, естественно, данными примерами не ограничивается.
Перечень фигур
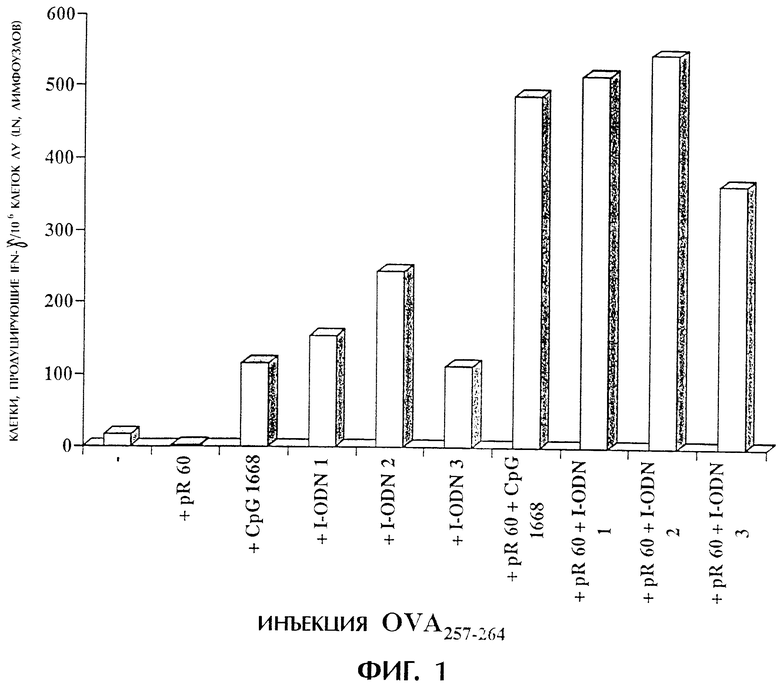
Фиг.1 показывает иммунную реакцию против полученного из овальбумина пептида OVA257-264 после инъекции OVA257-264, поли-L-аргинина (pR 60) и содержащих дезоксиинозин I олигодезоксинуклеотидов (I-ODN) или CpG 1668. Мышам делают инъекции в подушечки задних лап указанными смесями. Через четыре дня дренирование клеток лимфоузлов стимулируют OVA257-264 ex vivo. Через 24 часа с использованием анализа методом ELISPOT определяют число клеток, продуцирующих IFN-γ. Результаты выражают в числе пятен/1×106 клеток лимфоузлов.
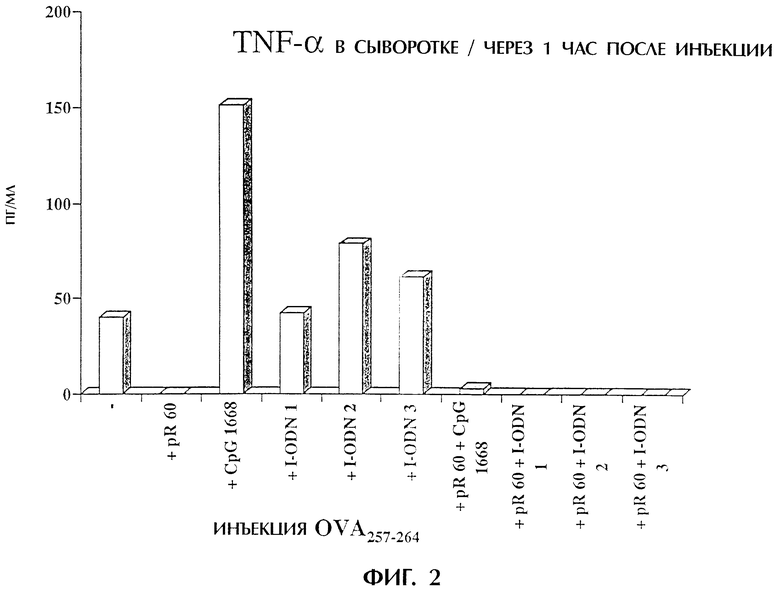
Фиг.2 показывает индукцию системного продуцирования TNF-α после инъекции OVA257-264, поли-L-аргинина (pR 60) и I-содержащих олигодезоксинуклеотидов (I-ODN) или CpG 1668. Мышам делают инъекции в подушечки задних лап указанными смесями. Через час после инъекции берут кровь из хвостовой вены и получают сыворотку. Концентрацию TNF-α в сыворотке определяют с использованием метода ELISA.
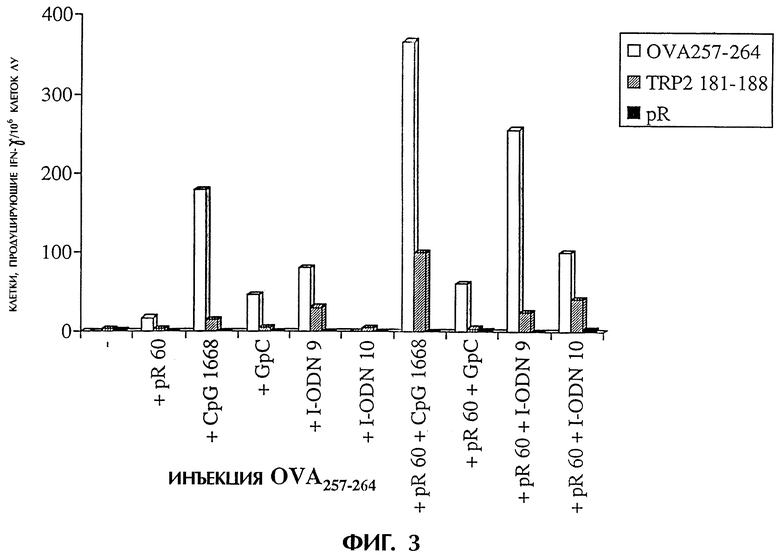
Фиг.3 показывает иммунную реакцию против полученного из овальбумина пептида OVA257-264 после инъекции OVA257-264, поли-L-аргинина (pR 60) и содержащих дезоксиинозин олигодезоксинуклеотидов (I-ODN), CpG 1668 или GpC. Мышам делают инъекции в подушечки задних лап указанными смесями. Через четыре дня стимулируют ex vivo дренирование клеток лимфоузлов OVA257-264, несоответствующим пептидом mTRP2181-188 (родственный мышиной тирозиназе протеин-2, VYDFFVWL) или pR 60. Через 24 часа с использованием анализа методом ELISPOT определяют число клеток, продуцирующих IFN-γ. Результаты выражают в числе пятен/1×106 клеток лимфоузлов со стандартным отклонением при трехкратном повторе.
Фиг.4 показывает индукцию системного продуцирования TNF-(после инъекции OVA257-264, поли-L-аргинина (pR 60) и I-содержащих олигодезоксинуклеотидов (I-ODN), GpC или CpG 1668. Мышам делают инъекции в подушечки задних лап указанными смесями. Через час после инъекции берут кровь из хвостовой вены и получают сыворотку. С использованием цитокинспецифического ELISA в сыворотке определяют концентрацию TNF-α и IL-6.
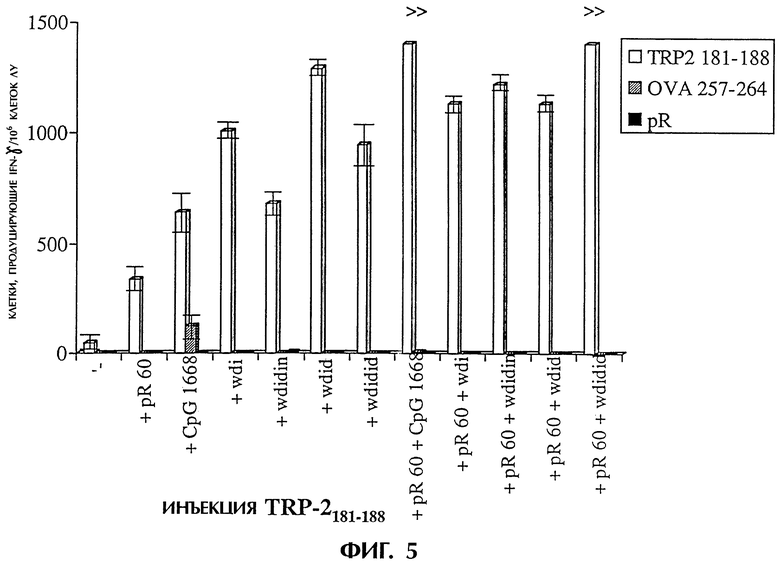
Фиг.5 показывает иммунную реакцию против полученного из овальбумина пептида OVA257-264 после инъекции TRP-2, поли-L-аргинина, CpG 1668 или случайных 20-мерных последовательностей, содержащих дезоксиинозин. Мышам делают инъекции в подушечки задних лап указанными смесями. Через четыре дня стимулируют ех vivo дренирование клеток лимфоузлов TRP-2, OVA257-264 или pR 60. Через 24 часа с использованием анализа методом ELISPOT определяют число клеток, продуцирующих IFN-γ. Результаты выражают в числе пятен/1×106 клеток лимфоузлов со стандартным отклонением при трехкратном повторе.
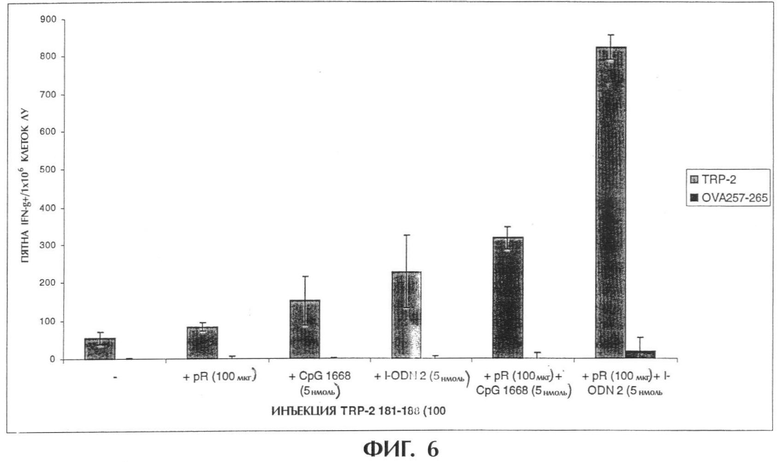
Фиг.6 показывает комбинированную инъекцию I-ODN и поли-L-аргинина (pR 60) вместе с пептидом, полученным из меланомы.
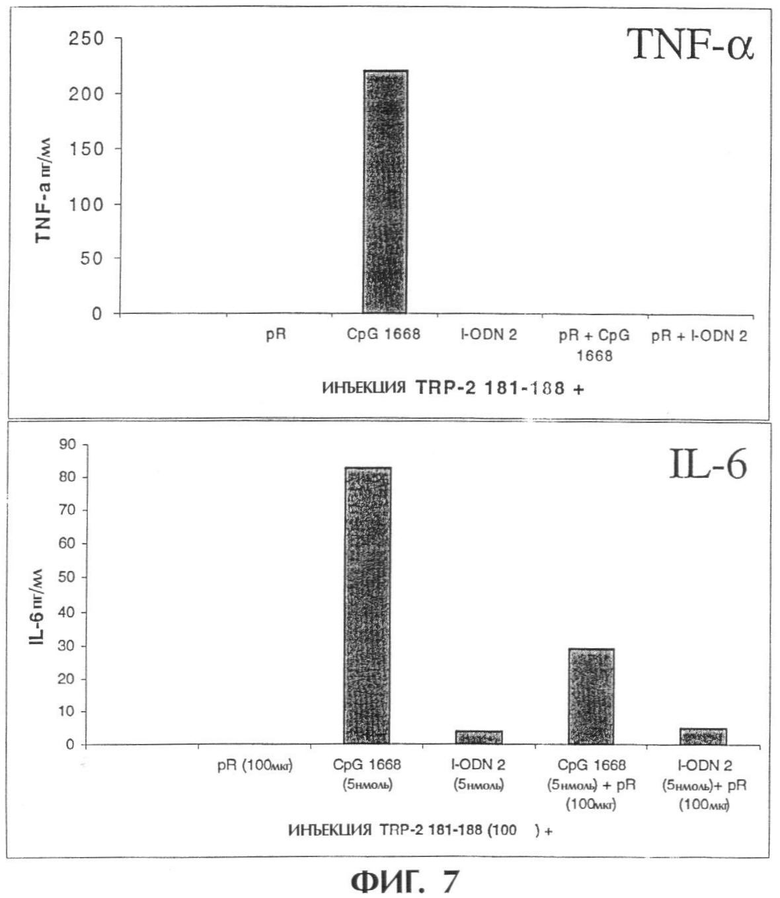
Фиг.7 показывает, что комбинированная инъекция I-ODN и pR 60 вместе с пептидом, полученным из меланомы, снижает индукцию системного продуцирования TNF-α и IL-6.
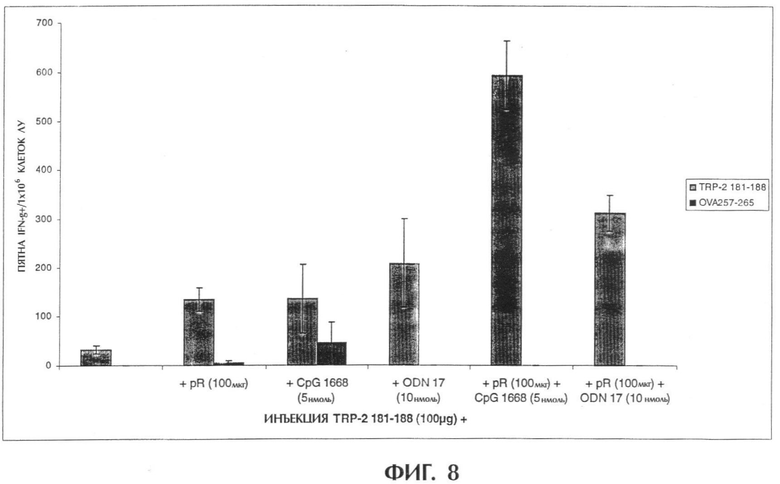
Фиг.8 показывает комбинированную инъекцию случайного 10-мерного I-ODN и pR 60 вместе с пептидом, полученным из меланомы.
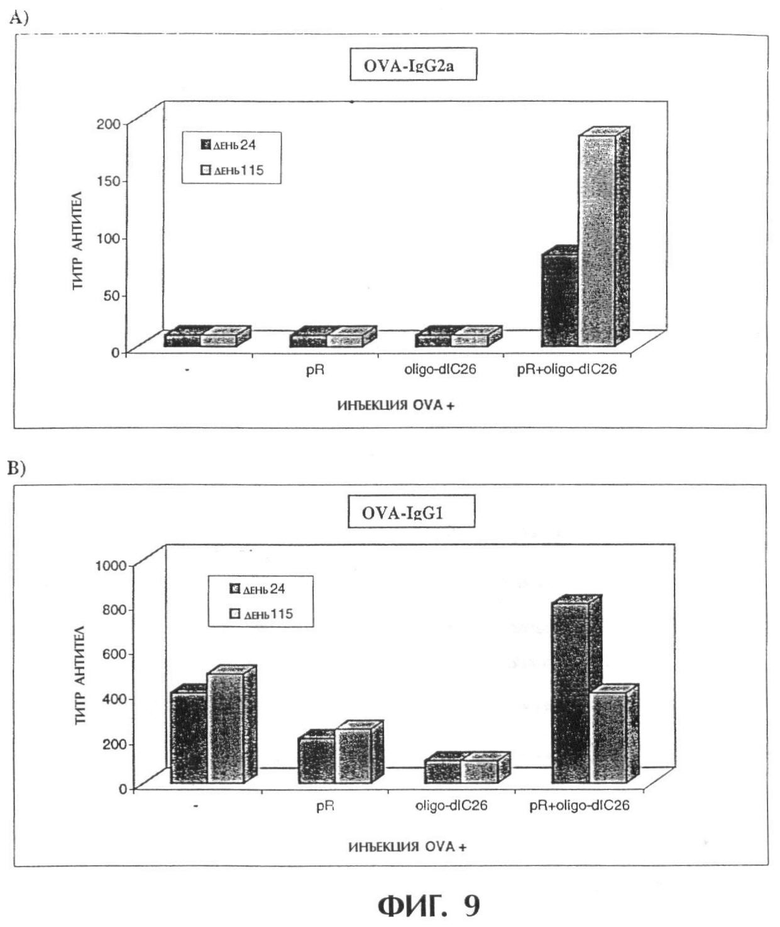
Фиг.9 показывает, что комбинированное применение овальбумина (OVA) с олиго-dIC26-мер и pR усиливает продуцирование OVA-специфических IgG-антител. Мышам подкожно в подушечки лап делают инъекции указанными смесями. В дни 24 и 115 после инъекции собирают сыворотки и подвергают скринингу методом ELISA на OVA-специфические антитела IgG2a (А) и IgG1 (В). Результаты приводятся в виде титра антител.
Осуществление изобретения
Во всех экспериментах используют тиофосфатзамещенные ODN (с заменой фосфатных остатков тиофосфатными остатками, называемые далее "тиофосфатзамещенными олигодезоксинуклеотидами"), так как такие ODN показывают более высокую устойчивость к нуклеазам (Ballas et al., 1996; Krieg et al., 1995; Parronchi et al., 1999).
Пример 1
Комбинированная инъекция различных I-ODN и поли-L-аргинина (pR 60) синергично усиливает иммунную реакцию против пептида, полученного из овальбумина
tcc atg acg ttc ctg atg ct, синтезированы NAPS GmbH, G ttingen.
Экспериментальные группы (5 мышей на группу)
1. OVA257-264
2. OVA257-264+pR 60
3. OVA257-264+CpG 1668
4. OVA257-264+I-ODN 1
5. OVA257-264+I-ODN 2
6. OVA257-264+I-ODN 3
7. OVA257-264+CpG 1668+pR 60
8. OVA257-264+I-ODN 1+pR 60
9. OVA257-264+I-ODN 2+pR 60
10. OVA257-264+I-ODN 3+pR 60
В день 0 мышам в каждую подушечку задних лап делают инъекцию 100 мл (общий объем, 50 мл на подушечку) композиций, содержащих вышеуказанные соединения. Животных умерщвляют через 4 дня после инъекции и собирают подколенные лимфатические узлы. Лимфатические узлы пропускают через 70-мм клеточный фильтр и дважды промывают средой DMEM (GEBCO BRL), содержащей 5% фетальной телячьей сыворотки (FCS, SIGMA chemicals). Клетки разводят средой DMEM/5% FCS до 3×106 клеток/мл. IFN-γ определяют методом ELISPOT при трехкратном повторе, как описано ранее (Miyahira et al., 1995). Такой способ является широко используемой процедурой, позволяющей количественно определить антигенспецифические Т-клетки. Лимфоциты стимулируют ex vivo контрольной фоновой средой, пептидом OVA257-264 или конканавалином A (Con А). Подсчитывают пятна, представляющие отдельные продуцирующие IFN-γ Т-клетки, и из результатов для всех образцов вычитают фоновые пятна. Большое число пятен, обнаруживаемых после стимуляции Con А (данные не приводятся), указывает на хорошее состояние используемых лимфоцитов. Для каждой экспериментальной группы мышей на фигуре 1 приводится число пятен/1×106 клеток.
Через один час после инъекции берут кровь из хвостовой вены и получают сыворотку для определения с помощью ELISA индукции системного продуцирования TNF-α (фигура 2).
Пример 2
Замена гуанозина на дезоксиинозин превращает неиммуногенную GpC-последовательность в высокоиммуногенную последовательность, особенно при сочетании с поли-L-аргинином (pR 60)
Экспериментальные группы (5 мышей на группу)
OVA257-264
OVA257-264+PR 60
OVA257-264+CpG 1668
OVA257-264+GpC
OVA257-264+I-ODN 9
OVA257-264+I-ODN 10
OVA257-264+CpG 1668+pR 60
OVA257-264+GpC+pR 60
OVA257-264+I-ODN 9+pR 60
OVA257-264+I-ODN 10+pR 60
В день 0 мышам в каждую подушечку задних лап делают инъекцию 100 мкл (общий объем, 50 мкл на подушечку) композиций, содержащих вышеуказанные соединения. Животных умерщвляют через 4 дня после инъекции и собирают подколенные лимфатические узлы. Лимфатические узлы пропускают через 70-мм клеточный фильтр и дважды промывают средой DMEM (GIBCO BRL), содержащей 5% фетальной телячьей сыворотки (FCS, SIGMA chemicals). Клетки разводят средой DMEM/5% FCS до 3×106 клеток/мл. IFN-γ определяют методом ELISPOT при трехкратном повторе, как описано ранее (Miyahira et al., 1995). Такой способ является широко используемой процедурой, позволяющей количественно определить антигенспецифические Т-клетки. Лимфоциты стимулируют ex vivo при трехкратном повторе средой (фон), пептидом OVA257-264, несоответствующим пептидом mTRP-2181-188 (родственный мышиной тирозиназе протеин-2, VYDFFVWL), pR 60 и конканавалином A (Con А). Подсчитывают пятна, представляющие отдельные продуцирующие IFN-γ Т-клетки, и из результатов для всех образцов вычитают фоновые пятна. Большое число пятен, обнаруживаемых после стимуляции Con А (данные не приводятся), указывает на хорошее состояние используемых лимфоцитов. Для каждой экспериментальной группы мышей на фигуре 3 приводится число пятен/1×106 клеток и стандартное отклонение при трехкратном повторе стимуляции ex vivo. Через один час после инъекции берут кровь из хвостовой вены, и получают сыворотку для определения с использованием цитокин-специфического ELISA индукции системного продуцирования TNF-α и EL-6 (фигура 4).
Пример 3
Комбинированная инъекция случайных 20-мерных последовательностей, содержащих дезоксиинозин, и пептида, полученного из меланомы, вызывает сильную иммунную реакцию против пептида, которая может быть даже усилена совместным применением поли-L-аргинина (pR 60)
Экспериментальные группы (5 мышей на группу)
1. TPR-2
2. TPR-2+pR 60
3. TPR-2+CpG 1668
4. TPR-2+wdi
5. TPR-2+wdidin
6. TPR-2+wdid
7. TPR-2+wdidid
8. TPR-2+CpG 1668+pR 60
9. TPR-2+wdi+pR 60
10. TPR-2+wdidin+pR 60
11. TPR-2+wdid+pR 60
12. TPR-2+wdidid+pR 60
В день 0 мышам в каждую подушечку задних лап делают инъекцию 100 мкл (общий объем, 50 мкл на подушечку) композиций, содержащих вышеуказанные соединения. Животных умерщвляют через 4 дня после инъекции и собирают подколенные лимфатические узлы. Лимфатические узлы пропускают через 70-мм клеточный фильтр и дважды промывают средой DMEM (GIBCO BRL), содержащей 5% фетальной телячьей сыворотки (FCS, SIGMA chemicals). Клетки разводят средой DMEM/5% FCS до 3×106 клеток/мл. IFN-γ определяют методом ELISPOT при трехкратном повторе, как описано ранее (Miyahira et al., 1995). Такой способ является широко используемой процедурой, позволяющей количественно определить антигенспецифические Т-клетки. Лимфоциты стимулируют ex vivo при трехкратном повторе средой (фон), пептидом TPR-2, несоответствующим пептидом OVA257-264, pR 60 и конканавалином A (Con А). Подсчитывают пятна, представляющие отдельные продуцирующие IFN-γ Т-клетки, и из результатов для всех образцов вычитают фоновые пятна. Большое число пятен, обнаруживаемых после стимуляции Con А (данные не приводятся), указывает на хорошее состояние используемых лимфоцитов. Для каждой экспериментальной группы мышей на фигуре 5 приводится число пятен/1×106 клеток, приводится стандартное отклонение при трехкратном повторе стимуляции ех vivo.
Пример 4
Комбинированная инъекция I-ODN и поли-L-аргинина (pR 60) синергично усиливает иммунную реакцию против пептида, полученного из меланомы
Экспериментальные группы (5 мышей на группу)
1. TPR-2181-188
2. TPR-2181-188+pR 60
3. TPR-2181-188+CpG 1668
4. TPR-2181-188+I-ODN 2
5. TPR-2181-188+CpG 1668+pR 60
6. TPR-2181-188+I-ODN 2+pR 60
В день 0 мышам в каждую подушечку задних лап делают инъекцию 100 мкл (общий объем, 50 мкл на подушечку) композиций, содержащих вышеуказанные соединения. Животных умерщвляют через 4 дня после инъекции, и собирают подколенные лимфатические узлы. Лимфатические узлы пропускают через 70-мм клеточный фильтр и дважды промывают средой DMEM (GIBCO BRL), содержащей 5% фетальной телячьей сыворотки (FCS, SIGMA chemicals). Клетки разводят средой DMEM/5% FCS до 3×106 клеток/мл. IFN-γ определяют методом ELISPOT при трехкратном повторе, как описано ранее (Miyahira et al., 1995). Такой способ является широко используемой процедурой, позволяющей количественно определить антигенспецифические Т-клетки. Лимфоциты стимулируют ex vivo при трехкратном повторе контрольной фоновой средой, пептидом TPR-2181-186, несоответствующим пептидом OVA257-264 и конканавалином A (Con А). Подсчитывают пятна, представляющие отдельные продуцирующие IFN-γ Т-клетки и из результатов для всех образцов вычитают фоновые пятна. Большое число пятен, обнаруживаемых после стимуляции Con А (данные не приводятся), указывает на хорошее состояние используемых лимфоцитов. Для каждой экспериментальной группы мышей на фигуре 6 приводится число пятен/1×106 клеток, приводится стандартное отклонение при трехкратном повторе стимуляции ех vivo.
Через один час после инъекции берут кровь из хвостовой вены и получают сыворотку для определения с использованием специфического ELISA индукции системного продуцирования TNF-α и IL-6 (фигура 7).
Пример 5
Комбинированная инъекция случайного 10-мерного I-ODN и поли-L-аргинина (pR 60) синергично усиливает иммунную реакцию против пептида, полученного из меланомы.
Экспериментальные группы (5 мышей на группу)
1. TPR-2181-188
2. TPR-2181-188+pR60
3. TPR-2181-188+CpG 1668
4. TPR-2181-188+ODN 17
5. TPR-2181-188+CpG 1668+pR 60
6. TPR-2181-188+ODN 17+pR 60
В день 0 мышам в каждую подушечку задних лап делают инъекцию 100 мкл (общий объем, 50 мкл на подушечку) композиций, содержащих вышеуказанные соединения. Животных умерщвляют через 4 дня после инъекции и собирают подколенные лимфатические узлы. Лимфатические узлы пропускают через 70-мм клеточный фильтр и дважды промывают средой DMEM (GIBCO BRL), содержащей 5% фетальной телячьей сыворотки (FCS, SIGMA chemicals). Клетки разводят средой DMEM/5% FCS до 3×106 клеток/мл. IFN-γ определяют методом ELISPOT при трехкратном повторе, как описано ранее (Miyahira et al., 1995). Такой способ является широко используемой процедурой, позволяющей количественно определить антигенспецифические Т-клетки. Лимфоциты стимулируют ex vivo при трехкратном повторе контрольной фоновой средой, пептидом TPR-2181-188, несоответствующим пептидом OVA257-264 и конканавалином A (Con А). Подсчитывают пятна, представляющие отдельные продуцирующие IFN-γ Т-клетки, и из результатов для всех образцов вычитают фоновые пятна. Большое число пятен, обнаруживаемых после стимуляции Con А (данные не приводятся), указывает на хорошее состояние используемых лимфоцитов. Для каждой экспериментальной группы мышей на фигуре 8 приводится число пятен/1×106 клеток и стандартное отклонение при трехкратном повторе стимуляции ех vivo.
Пример 6
Комбинированное применение олигодезокси-IC26-мер и поли-L-аргинина (pR) усиливает овальбумин(ОVА)-специфическую гуморальную реакцию
Экспериментальные группы (4 мыши на группу)
1. OVA+олиго-dIC26-мер+PR
2. OVA+олиго-dIC26-мер
3. OVA+pR
4. OVA
В день 0 мышам в каждую подушечку задних лап делают инъекцию 100 мкл (общий объем, 50 мкл на подушечку) композиций, содержащих вышеуказанные соединения. На 24 день после инъекции собирают сыворотку и подвергают скринингу методом ELISA на присутствие OVA-специфических антител. Полученные результаты показывают, что инъекция OVA в сочетании с олиго-dIC и pR усиливает продуцирование OVA-специфических антител IgG по сравнению с инъекцией OVA с каждым одним из указанных веществ (фигура 13А, В). Представляет интерес тот факт, что титры как IgG2a, так и IgG1 повышаются после одной инъекции OVA с олиго-dIC/pR, что предполагает, что вовлекаются как клетки Тh1, так и Th2. Однако через 115 дней в сыворотках мышей, инъецированных OVA и олиго-dIC, все еще обнаруживаются только повышенные уровни IgG2a.
Полученные результаты показывают, что комбинированная инъекция OVA с олиго-dIC и pR усиливает OVA-специфическую гуморальную реакцию. Такая реакция характеризуется продуцированием на ранней фазе как Тh1-, так и Th2-индуцированных изотипов антител, но позднее - только Тh1-индуцированных антител.
Источники информации
Andreu, D., and Rivas, L. (1998). Animal antimicrobial peptides: an overview. Biopolymers 47, 415-433.
Ballas, Z.K., Rasmussen, W.L., and Krieg, A.M. (1996). Induction of NK activity in murine and human cells by CpG motif in oligodeoxynucleotides and bacterial DNA. J Immunol 157, 1840-1845.
Bloom, В.R., and Widdus, R. (1998). Vaccine visions and their global impact. Nat Med 4, 480-484.
Bloom, M.В., Perry-Lalley, D., Robbins, P.F., Li, Y., el-Gamil, M., Rosenberg, S.A., and Yang, J.C. (1997). Identification of tyrosinase-related protein 2 as a tumor rejection antigen for the В 16 melanoma. J Exp Med 185, 453-459.
Buschle, M., Schmidt, W., Berger, M., Schaffner, G., Kurzbauer, R., Killisch, 1., Tiedemarm, J.K., Trska, В., Kirlappos, H., Mechtler, K., Schilcher, F., Gabler, C., and Birntsiel, M.L. (1998). Chemically defined, cell-free cancer vaccines: use of tumor antigen-derived peptides or polyepitope proteins for vaccination. Gene Ther. Mol. Biol. 1, 309-321.
Buschle, M., Schmidt, W., Zauner, W., Mechtler, K., Trska, В., Kirlappos, H., and Birnstiel, M.L. (1997). Transloading of tumor antigen-derived peptides into antigen-presenting cells. Proc. Natl. Acad. Sci. USA 94, 3256-3261.
Cavanaugh, P.F., Jr., Но, Y-К, and Bardos, T.J. (1996). The activation of murine macrophages and natural killer cells by the Partially thiolated double stranded RNA poly (1). mercapto poly(C). Res. Comm. Mol. Pathol. Pharmacol. 91, 131-147.
Chace, J.H., Hooker, N.A., Mildenstein, K.L., Krieg, A.M., and Cowdery, J.S. (1997). Bacterial DNA-induced NK cell IFN-gamma production is dependent on macrophage secretion of IL-12. Clin Immunol Immunopathol 84, 185-193.
Davis, H.L., Weeranta, R., Waldschmidt, T.J., Tygrett, L., Schorr, J., and Krieg, A. M. (1998). CpG DNA is a potent enhancer of specific immunity in mice immunized with recombinant hepatitis В surface antigen. J Immunol 160, 870-876.
Deng, G.M., Nilsson, l.M., Verdrengh, M., Collins, L.V., and Tarkowski, A. (1999). Intra-articularly localized bacterial DNA containing CpG motifs induces arthritis. Nat Med 5, 702-705.
Ganz, Т. (1999). Defensins and host defense [comment]. Science 286, 420-421.
Ganz, Т., and Lehrer, R.l. (1999). Antibiotic peptides from higher eukaryotes: biology and applications. Mol Med Today 5, 292-297.
Hancock, R.E. (1999). Host defence (cationic) peptides: what is their future clinical potential? Drugs 57, 469-473.
Harlow, E., and Lane, D. (1988). Antibodies: a laboratory manual (Cold Spring Harbor: Cold Spring Harbor Laboratory).
Hartmann, G., Weiner, G.J., and Krieg, A.M. (1999). CpG DNA: A potent signal for growth, activation, and maturation of human dendritic cells. Proc Hatl Acad Sci USA 96, 9305-9310.
Hoffmann, J.A., Kafatos, F.C., Janeway, C.A., and Ezekowitz, R.A. (1999). Phylogenetic perspectives in innate immunity. Science 284, 1313-1318.
Klinman, D.M., Yi, A.K., Beaucage, S.L., Conover, J., and Krieg, A.M. (1996). CpG motifs present in bacteria DNA rapidly induce Iymphocytes to secrete interleukin 6, interleukin 12, and interferon gamma. Proc Natl Acad Sci USA 93, 2879-2883.
Krieg, A.M. (1999). CpG DNA: a novel immunomodulator [letter]. Trends Microbiol 7, 64-5.
Krieg, A.M. (1996). An innate immune defense mechanism based on the recognition of CpG motifs in microbial DNA. J Lab Clin Med 128, 128-133.
Krieg, A.M., Yi, A.K., Matson, S., Waldschmidt, T.J., Bishop, G.A., Teasdale, R., Koretzky. G.A., and Klinman, D.M. (1995). CpG motifs in bacterial DNA trigger direct B-cell activation. Nature 374, 546-549.
Krieg. A.M., Yi, A.K., Schorr, J., and Davis, H.L. (1998). The role of CpG dinucleotides in DNA vaccines. Trends Microbiol 6, 23-27.
Lethe, В., van den Eynde, В., van Pel, A., Corradin, G., and Boon, T. (1992). Mouse tumor rejection antigens P815A and P815B: two epitopes carried by a single peptide. Eur J Immunol 22, 2283-2288.
Liljeqvist, S., and Stahl, S. (1999). Production of recombinant subunit vaccines: protein immunogens, live delivery systems and nucleic acid vaccines. J Biotechnol 73, 1-33.
Lipford, G.В., Heeg, K., and Wagner, H. (1998). Bacterial DNA as immune cell activator. Trends Microbiol 6, 496-500.
Manetti, R., Annunziato, F., Tomasevic, L., Gianno, V., Parronchi. P., Romagnani, S. and Maggi, E. (1995). Polyinosinic acid: polycytidylic acid promotes T helper type 1-specific immune responses by stimulating macrophage production of interferon-a and interleukin-12. Eur. J. Immunol. 25, 2656-2660.
Mosmann, T.R., Cherwinski, H., Bond, M.W., Giedlin, M.A., and Coffman, R.L. (1986). Two types of murine helper T cell clone. 1. Definition according to profiles of Iymphokine activities and secreted proteins. J Immunol 136, 2348-2357.
Nossal, G. (1998). Living up to the legacy. Nat Med 4, 475-476.
Oxenius, A., Martinic, M M., Hengartner, H., and Klenerman, P. (1999). CpG-containing oligonucleotides are efficient adjuvants for induction of protective antiviral immune responses with T-cell peptide vaccines. J Virol 73, 4120-4126.
Paillard, F. (1999). CpG: the double-edged sword [comment]. Hum Gene Ther 10, 2089-2090.
Pamer, E.G., Harty, J.Т., and Bevan, M.J. (1991). Precise prediction of a dominant class I MHC-restricted epitope of Listeria monocytogenes. Nature 353, 852-855.
Parronchi, P., Brugnolo, F., Annunziato, F., Manuelli, C., Sampognaro, S., Mavilia, C., Romagnani, S., and Maggi, E. (1999). Phosphorothioate oligodeoxynucleotides promote the in vitro development of human allergen-specific CD4+Т cells into Thl effectors. J Immunol 163. 5946-5953.
Pisetsky, D.S. (1997). Immunostimulatory DNA: a clear and present danger? Nat Med 3, 829-831.
Pisetsky, D.S. (1999). The influence of base sequence on the immunostimulatory properties of DNA. Immunol Res 19, 35-46.
Rammensee, H.G., Friede, Т., Stevanoviic S. (1995), MHC ligands and peptide motifs: first listing. Immunogenetics 41, 178-228.
Rodrigues, M., Nussenzweig, R.S., Romero, P., and Zavala, F. (1992). The in vivo cytotoxic activity of CD8+Т cell clones correlates with their levels of expression of adhesion molecules. J Exp Med 175, 895-905.
Roitt. l., Brostoff, J., and Male, D. (1998). Immunology (London: Mosby International Ltd).
Rotzschke, O., Falk, K., Stevanovic, S., Jung, G., Walden, P., and Rammensee, H.G. (1991). Exact prediction of a natural Т cell epitope. Eur J Immunol 21, 2891-2894.
Schmidt, W., Buschle, M., Zauner, W., Kirlappos, H., Mechtler, K., Trska, В., and Bimstiel, M.L. (1997). Cell-free tumor antigen peptide-based cancer vaccines. Proc. Natl. Acad. Sci. USA 94, 3262-3267.
Schwartz, D.A., Quinn, T.J., Thorne, P.S., Sayeed, S., Yi, A. K., and Krieg, A.M. (1997). CpG motifs in bacterial DNA cause inflammation in the lower respiratory tract, J Clin Invest 100, 68-73.
Shimonkevitz, R., Colon, S., Kappler, J.W., Marrack, P., and Grey, H.M. (1984). Antigen recognition by H2-resctricted Т cells 11. A tryptic ovalbumin peptide that substitutes for processed antigen. J Immunol 133, 2067-2074.
Simmaco, M., Mignogna, G., and Barra, D. (1998). Antimicrobial peptides from amphibian skin; what do they tell us? Biopolymers 47, 435-450.
Sparbier, K., and Walden, P. (1999). Т cell receptor specificity and mimotopes. Curr Opin Immunol 11, 214-218.
Sparwasser, Т., Koch, E.S., Vabulas, R.M., Heeg, K., Lipford, G.В., Ellwart, J.W., and Wagner, H. (1998). Bacterial DNA and Immunostimulatory CpG oligonucleotides trigger maturation and activation of murine dendritic cells. Eur J Immunol 28, 2045-2054.
Sparwasser, Т., Miethke, Т., Lipford, G., Borschert, K., Hacker, H., Heeg, K., and Wagner, H. (1997). Bacterial DNA causes septic shock [letter]. Nature 386, 336-337.
Sparwasser, Т., Miethke, Т., Lipford, G., Erdmann, A., Hacker, H., Heeg, K., and Wagner, H. (1997). Macrophages sense pathogens via DNA mot)&: induction of tumor necrosis factor-alpha-mediated shock. Eur J Immunol 27, 1671-1679.
Weiner, G.J., Liu, H.M., Wooldridge, J.E., Dahle, С.E., and Krieg, A.M. (1997). Immunostimulatory oligodeoxynucleotides containing the CpG motif are effective as immune adjuvants in tumor antigen immunization. Proc Natl Acad Sci USA 94, 10833-10837.
Yew, N.S., Wang, K.X., Przybylska, M., Bagley, R.G., Stedman, M., Marshall, J., Scheule, R.K., and Cheng, S.H. (1999). Contribution of plasmid DNA to inflammation in the lung after administration of cationic lipid: pDNA complexes. Hum Gene Ther 10, 223-234.
| название | год | авторы | номер документа |
|---|---|---|---|
| ИММУНОСТИМУЛИРУЮЩИЕ ОЛИГОДЕЗОКСИНУКЛЕОТИДЫ | 2001 |
|
RU2293573C2 |
| КОМПЛЕКС, СОДЕРЖАЩИЙ ОЛИГОНУКЛЕОТИД, ОБЛАДАЮЩИЙ ИММУНОСТИМУЛИРУЮЩЕЙ АКТИВНОСТЬЮ, И ЕГО ПРИМЕНЕНИЕ | 2014 |
|
RU2723943C2 |
| ВАКЦИННАЯ КОМПОЗИЦИЯ | 2001 |
|
RU2328305C2 |
| АЛЛОГЕННОЕ ПРОТИВООПУХОЛЕВОЕ ТЕРАПЕВТИЧЕСКОЕ СРЕДСТВО | 2004 |
|
RU2354694C2 |
| ВАКЦИНАЦИЯ С ИСПОЛЬЗОВАНИЕМ АЛЬФА3 ДОМЕНА MICA/B ДЛЯ ЛЕЧЕНИЯ РАКА | 2016 |
|
RU2747296C2 |
| МОДИФИЦИРОВАННЫЕ СУПЕРСПИРАЛЬНЫЕ БЕЛКИ С УЛУЧШЕННЫМИ СВОЙСТВАМИ | 2013 |
|
RU2677799C2 |
| Иммуностимулирующие олигонуклеотиды | 2012 |
|
RU2610690C2 |
| СПОСОБЫ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ, АССОЦИИРОВАННЫХ С ВПЧ | 2019 |
|
RU2799784C2 |
| КОМПОЗИЦИЯ ДЛЯ ЛЕЧЕНИЯ РАКА ПРЕДСТАТЕЛЬНОЙ ЖЕЛЕЗЫ | 2008 |
|
RU2583004C2 |
| ИММУНОСТИМУЛИРУЮЩИЕ ОЛИГОНУКЛЕОТИДЫ | 2009 |
|
RU2477753C2 |

Настоящее изобретение относится к области биотехнологии и медицины. Предложена вакцинная композиция, содержащая иммуностимулирующую олигодезоксинуклеиновую кислоту, содержащую дезоксиинозиновые остатки (I-ODN) и антигены. Такие вакцинные композиции вызывают более эффективный иммунный ответ по сравнению с ODN на основе последовательности CpG и снижают индукцию побочных реакций. 12 з.п. ф-лы, 9 ил.
1. Вакцинная композиция, содержащая
иммуностимулируюшую олигодезоксинуклеиновую кислоту (ODN), имеющую строение согласно формуле (I)
где любой Х представляет собой О или S,
и где любой NMP представляет собой монофосфат или монотиофосфат дезоксинуклеозида, выбранный из группы, состоящей из дезоксиаденозин-, дезоксигуанозин-, дезоксиинозин-, дезоксицитозин-, дезоксиуридин-, дезокситимидин-, 2-метилдезоксиинозин-, 5-метилдезоксицитозин-, дезоксипсевдоуридин-, дезоксирибозопурин-, 2-аминодезоксирибозопурин-, 6-S-дезоксигуанин-, 2-диметилдезоксигуанозин- или N-изопентенилдезоксиаденозин-монофосфата или -монотиофосфата, NUC представляет собой 2'-дезоксинуклеозид, выбранный из группы, состоящей из дезоксиаденозин-, дезоксигуанозин-, дезоксиинозин-, дезоксицитозин-, дезоксиуридин-, дезокситимидин-, 2-метилдезоксиинозин-, 5-метилдезоксицитозин-, дезоксипсевдоуридин-, дезоксирибозопурин-, 2-аминодезоксирибозопурин-, 6-S-дезоксигуанин-, 2-диметилдезоксигуанозин- или N-изопентенил-дезоксиаденозина,
а и b являются целыми числами от 0 до 100, при условии, что а+b равно величине от 4 до 150,
В и Е являются группами, обычными для 5'- или 3'-концов молекул нуклеиновых кислот,
и антиген.
2. Вакцинная композиция по п.1, характеризующаяся тем, что в указанном ODN любой NMP выбирают из группы, состоящей из дезоксиаденозин-, дезоксигуанозин-, дезоксиинозин-, дезоксицитозин-, дезоксиуридин-, дезокситимидин-, 2-метилдезоксиинозин-, 5-метилдезоксицитозин-монофосфата или -монотиофосфата.
3. Вакцинная композиция по п.1, характеризующаяся тем, что в указанном ODN a+b равно величине от 10 до 60, предпочтительно от 15 до 40.
4. Вакцинная композиция по п.1, характеризующаяся тем, что в указанном ODN по меньшей мере один из X1 и Х2 представляет собой S, и, по меньшей мере, один из Х3 и Х4 представляет собой О и, предпочтительно, любой NMP представляет собой монотиофосфат нуклеозида.
5. Вакцинная композиция по п.1, характеризующаяся тем, что указанный
ODN содержит последовательность
hhh wdi dhh h
nhh hhh wdi nhh hhh hhh wn,
nhh wdi din hhh hdi ndi nh,
nhh hhh wdi dhh hhh hhh wn или
nhh wdi did hhh hdi ddi dh,
где любой n представляет собой монофосфат или монотиофосфат 2'-дезоксинуклеозида, выбранный из группы, состоящей из дезоксиаденозин-, дезоксигуанозин-, дезоксицитозин- или дезокситимидин-монофосфата или монотиофосфата,
любой h представляет собой монофосфат или монотиофосфат 2'-дезоксинуклеозида, выбранный из группы, состоящей из дезоксиаденозин-, дезоксицитозин- или дезокситимидин-монофосфата или -монотиофосфата, i представляет собой дезоксиинозинмонофосфат или -монотиофосфат, любой w представляет собой монофосфат или монотиофосфат 2'-дезоксинуклеозида, выбранный из группы, состоящей из дезоксиаденозин- или дезокситимидин-монофосфата или -монотиофосфата, и
любой d представляет собой монофосфат или монотиофосфат 2'-дезоксинуклеозида, выбранный из группы, состоящей из дезоксиаденозин-, дезоксигуанозин- или дезокситимидин-монофосфата или монотиофосфата.
6. Вакцинная композиция по п.1, характеризующаяся тем, что указанный ODN содержит, по меньше мере, один 2'-дезоксицитозин-монофосфат или -монотиофосфат, 3'-соседний с 2'-дезоксиинозин-монофосфатом или -монотиофосфатом, в особенности 26-мер дезоксиинозин-дезоксицитозин.
7. Вакцинная композиция по п.1, характеризующаяся тем, что указанный ODN содержит последовательность
gacitt, в особенности tcc atg aci ttc ctg atg ct,
iacitt,
gaictt,
iaictt,
где а представляет собой дезоксиаденозин-монофосфат или -монотиофосфат,
g представляет собой дезоксигуанозин-монофосфат или -монотиофосфат,
i представляет собой дезоксиинозин-монофосфат или -монотиофосфат,
с представляет собой дезоксицитозин-монофосфат или -монотиофосфат, и
t представляет собой дезокситимидин-монофосфат или -монотиофосфат.
8. Вакцинная композиция по п.1, характеризующаяся тем, что указанный ODN содержит последовательность
wdi,
wdid,
wdidin или
wdidid,
где w, d, i и n имеют значения, указанные выше.
9. Вакцинная композиция по п.1, характеризующаяся тем, что в указанном ODN В и Е выбирают независимо из группы, состоящей из -Н, -СН3, -СОН, -СОСН3, -ОН, -СНО, -РO4, -PSO3, -PS2O2, -РS3O, -PS4, -SO3, -PO4-(CH2)1-6-NH2 или -PO4-(CH2)1-6-NH-метка.
10. Вакцинная композиция по п.1, характеризующаяся тем, что она дополнительно содержит поликатионный полимер, предпочтительно, поликатионный пептид, в частности полиаргинин, полилизин, или антимикробный пептид, в особенности антимикробный пептид, происходящий из кателицидина, синтетический пептид, содержащий два мотива KLK, разделенных линкером из 3-7 гидрофобных аминокислот, или гормон роста, в частности гормон роста человека.
11. Вакцинная композиция по п.1, характеризующаяся тем, что она дополнительно содержит другие активные ингредиенты, в частности цитокины, противовоспалительные вещества, антимикробные вещества или их сочетания.
12. Вакцинная композиция по п.1, характеризующаяся тем, что она дополнительно содержит вспомогательные вещества, в частности фармацевтически приемлемый носитель, буферные вещества, стабилизаторы или их сочетания.
13. Вакцинная композиция по любому из предыдущих пунктов, характеризующаяся тем, что она содержит от 1 нг до 1 г, предпочтительно
от 100 нг до 10 мг, в частности - от 10 мкг до 1 мг одного или нескольких ODN.
| WO 9816247 A1, 23.04.1998 | |||
| JINSUK WOO et al., G/C - modified oligodeoxynucleotides with selective complementarity: syntesis and hybridization properties, Nucleic Acids Res., 1996, Vol | |||
| Пишущая машина для тюркско-арабского шрифта | 1922 |
|
SU24A1 |
| ГОЛЬДФАРБ Д.М | |||
| и др | |||
| Иммунология нуклеиновых кислот | |||
| - М.: Наука, с.145-153. | |||

