[0001] Данная заявка испрашивает приоритет предварительной заявки на патент США №61/429409, поданной 3 января 2011 г.; предварительной заявки на патент США №61/431376, поданной 10 января 2011 г. Все предшествующие заявки полностью включены в данное описание путем ссылки.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
[0002] В настоящее время существует много рекомбинантных полипептидов и белков, используемых в терапевтическом лечении ряда заболеваний. Все эти рекомбинантные полипептиды и белки производят для реализации с использованием первичных постоянных линий нечеловеческих или человеческих диплоидных клеток. Например, некоторые из них получают с использованием бактерий, таких как Е. coli, a другие получают с использованием дрожжей или различных линий клеток яичников животного происхождения. Бактерии непригодны для получения некоторых полипептидов и белков, в частности, если профили гликозилирования и другие белковые модификации являются важными для активности связывания с биологическими рецепторами, биологической активности, биораспределения или фармакокинетики биопрепарата или для иммунологического распознавания реципиентом. Поскольку гликозилирование является критическим фактором, то клетка яичника китайского хомячка в настоящее время является одной из наиболее широко используемых клеточных линий для биологического производства.
[0003] К сожалению, необходимость использовать рекомбинантный полипептид или белок в течение длительного периода времени или для продолжительной терапии может привести к выработке у пациента нейтрализующих антител к продукту, что делает пациента менее восприимчивым или невосприимчивым к данному лекарственному средству. В некоторых случаях пациент может перейти на другое лекарственное средство того же класса, такого как анти-ФНО биопрепараты, такие как Энбрел, также известный как Этанерцепт; Ремикаид, также известный как Инфликсимаб; Цетолизумаб и Хумира, также известный как D2E7, которые используют для стандартного лечения заболеваний, таких как ревматоидный артрит, ювениальный ревматоидный артрит, псориаз, псориатический артрит, анкилозирующий спондилоартрит, язвенный колит и болезнь Крона. Однако, выработка нейтрализующих антител к одному конкретному анти-ФНО биопрепарату обычно приводит к тому, что у пациента в конечном случае вырабатываются нейтрализующие антитела к другому анти-ФНО биологическому продукту. В ряде случаев не существует альтернативного лечения пациента, и поэтому образование этих нейтрализующих антител оставляет пациента без вариантов лечения. Даже в том случае, когда существуют подходящие варианты лечения, предрасположенность к выработке нейтрализующих антител к другим лекарственным средствам того же класса в конечном счете означает, что пациент может остаться без вариантов лечения.
[0004] Другие биологические продукты, которые могут нейтрализоваться антителами человека, включают: Натализумаб или Тисабри, представляющий инновационное средство лечения рассеянного склероза, и Денозумаб - полностью человеческое моноклональное антитело к рецептору-активатору лиганда ядерного фактора каппа-В (RANKL), разрешенное для лечения остеопороза и индуцированных химиотерапией переломов костей с потенциалом использования для лечения рака молочной железы, вызванного ГЗТ и гормональными контрацептивами. Абатецепт является гибридным белком CTLA-4, разрешенным для использования при ревматоидном артрите у пациентов, у которых лечение анти-ФНО средствами перестало давать хорошие результаты. Хотя его использование начали слишком недавно, чтобы уже были сведения об индуцированных Абатецептом нейтрализующих антителах, также существует возможность того, что он может индуцировать ответ нейтрализующих антител человека.
[0005] Было показано, что другие полипептиды и белки, помимо лекарственных средств на основе антител и гибридных белков, также вызывают иммунный ответ при длительном лечении. Например, рекомбинантный человеческий эритропоэтин вызывает образование нейтрализующих антител, которые снижают его эффективность и могут привести к редкому синдрому аплазии. Дополнительно, частота выработки антител к средствам на основе факторов свертывания крови у больных гемофилией составляет до 25-30% пациентов. Это является основной проблемой факторов свертывания крови. Терапия рака и гепатита B с использованием рекомбинантного интерферона альфа 2a также усложняется генерированием нейтрализующих антител к лечению. Другой проблемой является невосприимчивость детей с малым ростом к длительной терапии гормонами роста. Известно об образовании нейтрализующих антител к некоторым инсулиновым продуктам.
[0006] Другие биологические лекарственные средства, которые обладают способностью вызывать образование нейтрализующих антител, включают цельную кровь, сыворотку, пулы плазмы или другие первичные исходные клетки биологических веществ, например, человеческий альбумин, ингибитор альфа 1-протеиназы человека, комплекс антигемофильного фактора и фактора Виллебранда человека, внутривенный противоботулинический иммуноглобулин BabyBig, ингибитор эстеразы C1, фибриносилант, фибриноген, иммуноглобулин внутривенный, иммуноглобулин подкожный, концентрат белка C, иммуноглобулин внутривенный Rho(D), тромбин, комплекс фактор Виллебранда/фактор VIII свертывания крови.
[0007] Рекомбинантные полипептиды и белки вызывают иммунные ответы и выработку нейтрализующих антител, что обусловлено рядом характеристик, включающих: срок биологического лечения, интервал повторного введения, аминокислотный состав биопрепарата и модификации биологического препарата, такие как гликозилирование, метилирование, нитросилация, сиалирование, фосфорилирование, сульфатирование, пренилирование, селенирование, убиквитинирование, витамин-зависимые модификации, ассоциации связывания с белком, ацилирование, гликирование, трехмерные конфигурации и сверхспирализацию. Таким образом, существует потребность в способах получения полипептидный и белковых продуктов, обладающих пониженными уровнями антигенности у животных, которых лечат данным биологическим продуктом.
[0008] Содержание всех источников, цитируемых в данном документе, полностью включено в данный документ посредством ссылок.
ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0009] Настоящее изобретение удовлетворяет указанную потребность путем обеспечения способов получения биопрепаратов, таких как полипептиды или белки, нуклеиновые кислоты, вирусы и вакцины, путем трансфецирования или трансформирования полученных искусственным путем плюрипотентных стволовых клеток (spPSC) или эндогенных плюрипотентных стволовых клеток (ePSC). Эти клетки получают из видов, которые лечат и трансфецируют векторами, которые экспрессируют целевой биопрепарат, и индуцируют экспрессию биологического продукта трансфецированной или трансформированной spPSC или ePSC.
[0010] Также настоящее изобретение обеспечивает способ получение рекомбинантного полипептида или белка, включающий получение spPSC из клеток взрослого организма или выделение ePSC животного и трансфецирование или трансформирование указанных spPSC или ePSC нуклеиновой кислотой, кодирующей указанный полипептид или белок, в условиях, при которых полипептид или белок экспрессируются указанной стволовой клеткой.
[0011] В альтернативном варианте осуществления настоящего изобретения spPSC получают или ePSC выделяют из клеток той же этнической группы, что и пациент, которому вводят рекомбинантный полипептид или белок. Разные этнические группы могут иметь отличающиеся профили гликозилирования и полиморфизмы, различные в разных группах. Этнические группы - это группы, которые имеют одинаковые типы крови и тканей. Таким образом, согласно настоящему изобретению рекомбинантные полипептиды и белки получают из spPSC или ePSC, причем ePSC или spPSC получают из клеток, выделенных из той же этнической группы, что и пациент, которому вводят рекомбинантный полипептид или белок. Пациенту будут вводить рекомбинантный полипептид или белок, продуцируемый spPSC или ePSC, причем spPSC получены или ePSC выделены из клеток, принадлежащих этнической группе, к которой принадлежит указанный пациент.
[0012] Настоящее изобретение также обеспечивает способ введения полипептида или белка пациенту-животному, включающий получение spPSC, таких как индуцированные плюрипотентные стволовые клетки (iPSC), из указанных клеток указанного животного или выделение ePSC и трансфецирование или трансформирование указанных spPSC или ePSC нуклеиновой кислотой, кодирующей указанный полипептид или белок, в условиях, при которых полипептид или белок экспрессируются указанной плюрипотентной стволовой клеткой, выделение указанного полипептида или белка из указанной индуцированной плюрипотентной стволовой клетки и введение указанного выделенного полипептида или белка указанному пациенту.
[0013] Настоящее изобретение обеспечивает способ персонализированного получения терапевтических полипептидов или белков, делающий тем самым spPSC или ePSC коммерчески доступными и полезными. Настоящее изобретение относится к способам, позволяющим создавать пациент-специфические spPSC или ePSC для получения пациент-специфических полипептидов или белков, чтобы устранить проблемы образования нейтрализующих антител, которые обычно связаны с полипептидами или белками для длительного и постоянного использования. Пациентом может быть любое животное, предпочтительно млекопитающее и, более предпочтительно, человек. Настоящее изобретение также обеспечивает способ получения нуклеиновых кислот или вирусов, включающий трансфецирование или трансформирование spPSC или ePSC вектором в условиях, при которых продуцируются желательная нуклеиновая кислота или вирус.
[0014] Кроме того, в настоящем изобретении предложены способы получения пациент-специфической и орган- или клеточно-специфической клеточной линии для продуцирования тщательно подобранных посттранскрипционно модифицированных биопрепаратов для терапевтического использования. Пациент-специфические клеточные линии могут быть получены, используя SCNT, индуцированное перепрограммирование, партеногенез или способы ANT-OAR перепрограммирования, или они могут быть выделены из пациента-мишени. Таким образом, плюрипотентные стволовые клетки, полученные или выделенные, могут генетически модифицироваться, используя стандартные процедуры молекулярной биотехнологии для экспрессирования соответствующих терапевтических средств, например, используя инсерционные или эписомальные векторы экспрессии или способы гомологичной рекомбинации. Генетически модифицированная клеточная линия может быть размножена в культуре и сохранена для периодического биологического массового производства, планируемого на основе срока хранения изготавливаемого биопрепарата (Пример 2). Альтернативно, полученные пациент-специфические клеточные линии могут быть дифференцированы до клеточного типа, который нормально экспрессирует повышенные уровни желательного терапевтического белка, и затем использованы для биологического производства. Дифференцирование может проводиться для каждого массового производства или может быть осуществлено в крупных размерах, а дифференцированные пациент-специфические клеточные линии сохраняться для последующих массовых производств, основанных на сроке хранения изготавливаемого терапевтического средства (Пример 3).
[0015] Дополнительно, поскольку известно, что профили гликозилирования и другие посттрансляционные модификации различаются у разных типах тканей и клеток, линии пациент-специфических стволовых клеток могут быть получены из зрелых или соматических клеток, выделенных из органа или из типов клеток, эндогенно экспрессирующих указанный биопрепарат. Как, например, затем быть использованы методы SCNT, PGA, ANT-OAR или методы перепрограммирования для получения линии плюрипотентных клеток для получения биологических веществ. Таким образом, плюрипотентные стволовые клетки, полученные или выделенные, могут быть генетически модифицированы с использованием стандартных процедур молекулярной биотехнологии для экспрессирования целевого терапевтического средства. Генетически модифицированная клеточная линия может быть размножена в культуре и сохранена для периодического биологического массового производства, планируемого на основе срока хранения изготавливаемого биопрепарата (смотри Пример 4). Развивая преимущество свойств «памяти» перепрограммированных клеток (iPS-клетки), пациент- и ткане- или клеточно-специфические iPS-клетки могут быть индуцированы для дифференцировки обратно до своего типа клеток происхождения, чтобы максимально полно создать клеточную линию, поддающуюся эндогенной посттрансляционной модификации. Клетки iPS могут быть генетически модифицированы для экспрессирования целевого терапевтического средства перед редифференцировкой в первоначального выделенный тип клеток или после редифференцировки в первоначальный тип клеток (смотри Пример 5).
[0016] Например, гормон роста обычно интенсивнее продуцируется соматотропными клетками в передней доле гипофиза, а также экспрессируется на высоком уровне в клетках плаценты (тромбобласты), и языка и кожи вульвы или ануса. Экспрессия белка фактора VIII высока в клетках почечных канальцев, но он умеренно экспрессируется рядом тканей и типов клеток согласно Атласу белков человека. Антитела обычно продуцируются B-клетками, которые созревают в зародышевых центрах селезенки и других лимфоидных органах. Терапевтическое продуцирование антител с высокими уровнями антителозависимой клеточной цитотоксичности (ADCC) определяют по уровню GDP-D-маннозо-4,6-дегидратазы (GMD), которая может помещать N-ацетилглюкозамин (GlcNac) в положении раздвоения антител подтипа IgG1, присутствующей в производящей клеточной линии (J Biol Chem., Vol.273, pp.14582-14587, 1998 and BMC Biotechnol., 7: 84-97 (2007). Продуцирование антител с высокой активностью АЗКЦ не всегда является оптимальным при использовании производственной линии клеток CHO (J Biol Chem. 278: 3466-3473, 2003). В настоящем изобретении предложены новые клеточные линии млекопитающих для оптимальной активности АЗКЦ продуцируемых антител, гибридных белков и цитотоксических биопрепаратов.
[0017] Известно, что факторы транскрипции, ассоциированные с высокими уровнями биологического продуцирования, могут быть котрасфектированы совместно с целевым геном для оптимизации уровней экспрессии в пациент-специфической клеточной линии. Например, высокие уровни экспрессии Pit-1 могут вызывать интенсивную экспрессию пролактина в некотором клеточном типе, одновременно блокируя или предотвращая экспрессию гормона роста (Genes Dev, 3: 946-958 1989).
[0018] Продуцирование моноклональных антител может быть увеличено путем оптимизации генного кодона с использованием таких систем, как разработанные Sino Biological Inc, Peoples Republic of China, морфогенетические агенты (Proc Natl Acad Sci, 103: 3557-3562, 2006) или другие стандартные биотехнологические методы.
Получение искусственно полученных плюрипотентных стволовых клеток
[0019] Любой тип искусственно полученной плюрипотентной стволовой клетки может быть использован для получения персонализированных биопрепаратов настоящего изобретения. Двумя основными категориями являются индуцированные или перепрограммированные плюрипотентные стволовые клетки (iPSC) и стволовые клетки, полученные путем переноса ядра (SCNT), ANT-OAR и партеногенеза.
[0020] Перенос ядер соматических клеток (SCNT) является методикой, в которой в яйцеклетку с удаленным ядром инъецируют ядра зрелых соматических клеток и имплантируют в подготовленную матку реципиента с получением живорождений, которые дают полные генетические ядерные клоны. Дополнительно, плюрипотентные стволовые клетки получают в культуре после применения методов SCNT (Cell Reprogram. 12: 105-113, 2010 and Genome Res., 19: 2193-2201, 2009).
[0021] Перенос измененных ядер. перепрограммирование с использованием яйцеклетки (ANT-OAR) является методикой, близкой SCNT, однако донорские ядра генетически изменяют перед введением в яйцеклетку-реципиент, тем самым предотвращая дифференцировку ANT-ооцита и образование целого организма (Genome Res. 19: 2193-2201, 2009).
[0022] Партеногенез (PGA) также используют для получения плюрипотентных стволовых клеток, применяя такие методики как перенос ядер, освобожденных от вителлинового слоя, активация партеногенеза; и методики клонирования, такие как SCNT, также партеногенез (PGA) используют для получения перепрограммированных плюрипотентных стволовых клеток (Cell Reprogram., 12: 105-113, 2010 and Nature, 450: 497-502 2007).
[0023] Эти плюрипотентные стволовые клетки могут поддерживаться в культуре на протяжении довольно длительных неопределенных периодов, что делает их потенциальным источником для биологического производства, например, рекомбинантных белков, ДНК и вирусов.
Индуцированные или перепрограммированные spPSC
[0024] Индуцированные плюрипотентные стволовые клетки подобны природным плюрипотентным стволовым клеткам, таким как эмбриональные стволовые клетки (ES), во многих отношениях, таких как экспрессирование генов указанных стволовых клеток и белков, профили метилирования хроматина, время удвоения, образование эмбриоидных телец, образование тератомы, образование жизнеспособной химеры, потенция и дифференцируемость.
[0025] Индуцированные плюрипотентные стволовые клетки, обычно сокращенно называемые iPS-клетки или iPSC, являются типом плюрипотентных стволовых клеток, искусственно полученных из неплюрипотентных клеток, обычно зрелой соматической клетки, путем индуцирования "принудительной" (форсированной) экспрессии специфических генов (Nature Reports Stem Cells. 2007).
[0026] Индуцированные плюрипотентные стволовые клетки (iPSC) получают путем трасфекции некоторых ассоциированных со стволовыми клетками генов в неплюрипотентные клетки. Индуцированные плюрипотентные стволовые клетки обычно получают путем трансфекции указанных ассоциированных со стволовыми клетками генов в такие неплюрипотентные клетки, как зрелые фибробласты. Как правило, трансфекцию осуществляют посредством вирусных векторов, таких как ретровирусы, или ретротранспозонов. Трансфецированные гены включают исходные регуляторы транскрипции Oct-3/4 (Pou5f1) и Sox2, хотя предполагается, что другие гены повышают эффективность индукции. Через 3-4 недели небольшие количества трансфецированных клеток начинают становиться морфологически и биохимически похожими на плюрипотентные стволовые клетки, обычно их выделяют посредством морфологической селекции, по времени удвоения или с использованием гена-репортера и антибиотической селекции.
[0027] Эмбриональные стволовые клетки, полученные из фибробластов, зрелых фибробластов и других клеток, перепрограммируют до плюрипотентного состояния путем слияния с эмбриональными стволовыми клетками (Cell. 126: 652-655, 2006 and Stem Cell Rev, 2: 331-340, 2006), путем добавления 4 генов, используя методы ретровирусной трансфекции (Cell. 126: 663-676, 2006), с помощью одинарной кассеты или методов бицистронной лентивирусной трансфекции [Stem Cells; 27: 543-549, 2009, and Stem Cells., 27: 1042-1049, 2009), и путем эндогенной стимуляции плюрипотентности соответствующих факторов транскрипции (Stem Cells., 27: 3053-3062, 2009). Индукция плюрипотентности также может быть достигнута путем модификации режима метилирования или полиаденелирования генома (PLoS One., 4: е8419, 2009), с помощью микроРНК (Dev Biol., 344: 16-15, 2010), низкомолекулярных активаторов соответствующих факторов транскрипции, эпигенетического перепрограммирования (Regen Med., 2: 795-816, 2007), с помощью белкового перепрограммирования (Blood 116: 386-395, 2010), добавлением супернатанта культуры клеток или клеточных экстрактов из плюрипотентных клеток в культуре, с использованием химических, радиационных или других средств генной мутации для реактивации плюрипотентных генов, и путем добавления факторов роста или цитокинов, или агентов клеточной сигнализации, которые индуцируют или поддерживают эндогенные плюрипотентные состояния.
[0028] При использовании ретровирусов для перепрограммирования клеток до плюрипотентных состояний существует опасность, аналогичная связанной с исследованиями генной терапии иммунодефицита. Для удаления ретровирусов после завершения перепрограммирования используют такие эксцизионные методы, как Cre-lox, а методы с использованием транспозона piggyBac полностью исключают потребность в ретровирусах (Curr Opin Biotechnol., 20: 516-521, 2009).
[0029] Человеческие iPSC получают путем трансформирования человеческих фибробластов в плюрипотентные стволовые клетки, с использованием четырех базовых генов: Oct3/4, Sox2, Klf4 и c-Myc с ретровирусной системой. Человеческие iPSC также получают с использованием OCT4, SOX2, NANOG и другой ген LIN28, применяя лентивирусную систему. Для переноса необходимых четырех генов в ДНК клеток кожи и печени также используют аденовирус, что дает клетки, идентичные эмбриональным стволовым клеткам (Science 322 (5903): 945-949, 2008). Перепрограммирование зрелой клетки в iPSC также можно осуществить с использованием плазмиды без какой-либо вирусной трансфекционной системы вообще (Science 322 (5903): 949-953, 2008). IPSC получают с использованием транспозонной системы piggyBac, минициклической технологии, перепрограммирования, стимулированного белком или кондиционированной средой.
[0030] Образование iPS-клеток в значительной степени зависит от генов, используемых для индукции. Было определено, что Oct-3/4 и указанные представители семейства генов Sox (Sox1, Sox2, Sox3 и Sox15) являются ключевыми регуляторами транскрипции, играющими активную роль в процессе индукции, и отсутствие их делает индукцию невозможной. Кроме того, было определено, что дополнительные гены, включая указанные представители семейства Klf (Klf1, Klf2, Klf4 и Klf5), семейства Myc (C-myc, L-myc и N-myc), Nanog и LIN28, повышают эффективность индукции.
- Oct-3/4 (Pou5f1) (кДНК, доступная от Bioclone, San Diego CA) (Nucleic Acids Res. 20 (17): 4613-20, 1992): Oct-3/4 - одно из семейств октамерных факторов транскрипции ("Oct") и играет ключевую роль в поддержании плюрипотентности. Отсутствие Oct-3/4 в клетках Oct-3/4+, таких как бластомеры и эмбриональные стволовые клетки, приводит к спонтанной трофобластной дифференцировке, а присутствие Oct-3/4, соответственно, приводит к возникновению плюрипотентности и способности эмбриональных стволовых клеток к дифференцировке. Ряд других генов семейства "Oct", включая близких родственников Oct-3/4 - Oct1 и Oct6, не в состоянии вызвать индукцию, что демонстрирует эксклюзивность Oct-3/4 для процесса индукции.
- семейство Sox: семейство генов Sox связано с поддережанием плюрипотентности подобно Oct-3/4, хотя оно связано с мультипотентными и унипотентными стволовыми клетками, в отличие от Oct-3/4, который экспрессируется исключительно в плюрипотентных стволовых клетках (Dev Biol. 227 (2): 239-55, 2000). Наряду с тем, что Sox2 (кДНК можно приобрести в Bioclone, Сан-Диего, Калифорния, США) был исходным геном, применяемым для индукции (Mamm. Genome 5 (10): 640-642, 1995), было обнаружено, что другие гены семейства Sox так же работают в индукционном процессе. Sox1 (кДНК можно приобрести в Bioclone, Сан-Диего, Калифорния, США) обеспечивает получение iPS-клеток с той же эффективностью, что и Sox2, а гены Sox3 (человеческая кДНК, можно приобрести в Bioclone, Сан-Диего, Калифорния, США), Sox15 и Sox18 также генерируют iPS-клетки, хотя и с более низкой эффективностью.
- семейство Klf: Klf4 из семейства генов Klf является фактором образования мышиных iPS-клеток. Klf2 (кДНК можно приобрести в Bioclone, Сан-Диего, Калифорния, США) и Klf4 (кДНК можно приобрести в Bioclone, Сан-Диего, Калифорния, США) являются факторами, способными обеспечивать получение iPS-клеток, а родственные гены Klf1 (кДНК можно приобрести в Bioclone, Сан-Диего, Калифорния, США) и Klf5 (кДНК можно приобрести в Bioclone, Сан-Диего, Калифорния, США) действуют аналогичным образом, хотя и с более низкой эффективностью.
- семейство Myc: гены семейства Myc являются протоонкогенами, связанными с раком. C-myc (кДНК можно приобрести в Bioclone, Сан-Диего, Калифорния, США) является фактором, участвующим в образовании мышиных iPS-клеток. Однако, c-myc возможно не являются необходимыми для образования человеческих iPS-клеток. Использование семейства генов "myc" в индуцировании iPS-клеток является проблемой для возможности клинической терапии iPS-клетками, так как у 25% мышей, которым трансплантируют индуцированные c-myc клетки iPS, развились летальные тератомы. N-myc (кДНК можно приобрести в Bioclone, Сан-Диего, Калифорния, США) и L-myc были идентифицированы для индуцирования вместо c-myc с близкой эффективностью.
- Nanog: (кДНК можно приобрести в Bioclone, Сан-Диего, Калифорния, США) Nanog, наряду с Oct-3/4 и Sox2, является необходимым для активации плюрипотентности в эмбриональных стволовых клетках (Cell 113 (5): 643-55, 2003).
- LIN28: (кДНК можно приобрести в Bioclone, Сан-Диего, Калифорния, США) LIN28 является связывающим мРНК белком, экспрессируемым в эмбриональных стволовых клетках и в эмбриональных раковых клетках, связанных с дифференцировкой и пролиферацией (Dev Biol 258 (2): 432-42,2003).
Идентичность искусственно полученных плюрипотентных стволовых клеток
[0031] Полученные spPSC в значительной степени похожи на выделенные в природных условиях плюрипотентные стволовые клетки (такие как мышиные или человеческие эмбриональные стволовые клетки (ESC), mESC и hESC, соответственно) в следующих отношениях, тем самым подтверждая идентичность, аутентичность и плюрипотентность spPSC выделенным в природных условиях плюрипотентным стволовым клеткам:
Биологические свойства клеток:
- Морфология: iPSC морфологически близки к ESC. Каждая клетка имеет округлую форму, большое ядрышко и небольшое количество цитоплазмы. Колонии iPSC также подобны колониям ESC. Человеческие iPSC образуют заостренные, плоские, плотно упакованные колонии, подобные hESC, а мышиные iPSC образуют колонии, подобные mESC, менее плоские и более агрегированные колонии, чем колонии hESC.
- Ростовые свойства: в связи с тем, что стволовые клетки по определению должны самообновляться, время удвоения и митотическая активность существенны для ESC. iPSC митотически активны, активно самообновляются, пролиферируют и делятся со скоростью, равной ESC.
- Маркеры стволовых клеток: iPSC экспрессируют те же антигенные маркеры клеточной поверхности, что и ESC. Человеческие iPSC экспрессируют маркеры, специфические для hESC, включая SSEA-3, SSEA-4, TRA-1-60, TRA-1-81, TRA-2-49/6Е и Nanog. Мышиные iPSC экспрессируют SSEA-1, но не SSEA-3 или SSEA-4, подобно mESC.
- Гены стволовых клеток: iPSC экспрессируют гены, экспрессируемые в недифференцированных ESC, включая Oct-3/4, Sox2, Nanog, GDF3, REX1, FGF4, ESG1, DPPA2, DPPA4 и hTERT.
- Теломеразная активность: теломеразы необходимы для поддержания клеточного деления, неограниченного пределом Хейфлика, равным ~50 клеточных делений. hESC демонстрируют высокую теломеразную активность для поддержания самообновления и пролиферации, iPSC также демонстрируют высокую теломеразную активность и экспрессируют hTERT (обратная транскриптаза теломеразы человека), необходимый компонент теломеразо-белкового комплекса.
[0032] Плюрипотентность: iPSC способны к дифференцировке аналогично ESC, в полностью дифференцированные ткани:
- Нейрональная дифференцировка: iPSC могут дифференцировать в нейроны, экспрессирующие βIII-тубулин, тирозингидроксилазу, AADC, DAT, ChAT, LMX1B и MAP2. Присутствие катехоламин-ассоциированных ферментов может указывать на то, что iPSC, подобно hESC, могут дифференцировать в допаминэргические нейроны. После дифференцировки связанные со стволовыми клетками гены подавляются.
- Кардиальная дифференцировка: iPSC могут дифференцировать в кардиомиоциты, которые начинают спонтанно пульсировать. Кардиомиоциты экспрессировали TnTc, MEF2C, MYL2A, MYHCβ и NKX2.5. После дифференцировки связанные со стволовыми клетками гены подавлялись.
- Образование тератомы: iPSC, инъецированные в иммунодефицитную мышь, через девять недель спонтанно образуют тератомы. Тератомы - это опухоли множественного происхождения, содержащие ткани, производные от трех зародышевых листков - эндодермы, мезодермы и эктодермы, это не похоже на другие опухоли, которые обычно представлены только одним типом клеток. Образование тератомы - это основной тест на плюрипотентность.
- Эмбриоидные тельца: в культуре hESC спонтанно образуют сфероподобные эмбриоподобные структуры, названные "эмбриоидными тельцами", которые состоят из кора митотически активных и дифференцирующих hESC и периферии - полностью дифференцированных клеток всех трех зародышевых слоев. iPSC также образуют эмбриоидные тельца и имеют периферийные дифференцированные клетки.
- Тетраплоидная комплементация: iPS образуют мышиные фетальные фибробласты, инъецируемые в тетраплоидные бластоциты (которые сами могут образовывать только экстраэмбриональные ткани), из них могут развиваться целые, не химерные, фертильные мыши, хотя и с низкой степенью успеха. Метод тетраплоидной комплементации - это биологический метод, при котором клетки двух эмбрионов млекопитающих объединяют с образованием нового эмбриона. Его используют для конструирования генетически модифицированных организмов для изучения последствий некоторых мутаций на эмбриональное развитие и изучении плюрипотентных стволовых клеток.
[0033] Индуцированные плюрипотентные стволовые клетки (iPS) получали из клеток брыжейки кишечника (Cell Reprogram., 12: 237-247, 2010), пигментированных эпителиальных клеток сетчатки (Stem Cells., 28: 1981-1991, 2010), амниотических клеток (Differentiation., 80: 123-129, 2010), фибробластов (J Vis Exp., 8: 1553, 2009), зрелых нейральных клеток (Nature., Vol.454, pp.646-650, 2008), пульпы зуба (J Dent Res, Vol.89, pp.773-778, 2010), жировых клеток (Cell Transplant., 19: 525-536, 2010), клеток яичника (J Reprod Dev., 56: 481-494, 2010) и многих других клеток эмбриональных, фетальных и зрелых источников. Теоретически, iPS-клетки могут быть получены из любого типа клеток, хотя пока еще не все 220 типов клеток тела систематически изучены. Несколько недавних исследований продемонстрировали, что iPS-клетки сохраняют ′память′ о типе клеток своего происхождения. Это выражается в том, что iPS-клетки редифференцируют в культуре, иногда спонтанно, предпочтительно в клетки своего типа происхождения.
Выделение эндогенных стволовых клеток
[0034] Стволовые клетки, включая эндогенные плюрипотентные стволовые клетки (ePSC), могут быть охарактеризованы и выделены с помощью специфических антигенов, экспрессируемых на их поверхности. Плюрипотентные стволовые клетки могут быть охарактеризованы по экспрессии стадиеспецифического эмбрионального антигена (SSEA), факторов транскрипции Oct4 и Nanog и других маркеров наряду с другими способами. Основной тип эндогенных плюрипотентых стволовых клеток, который был выделен на сегодняшний день, представлен очень маленькими эмбрионоподобными стволовыми клетками (VSEL).
[0035] VSEL представляют собой маленькие (у мышей 3-5 микрон в диаметре, у людей 3-7 микрон в диаметре) с высоким ядерно-цитоплазматическим отношением. VSEL положительны по SSEA1, Oct4, Nanog, Rex1 и другим маркерам плюрипотентных стволовых клеток, и по CD133, CD34, AP, cMet, LIF-R и CXCR4 (J Am Coll Cardiol 53(1): 10-20, 2009; Stem Cell Rev 4: 89-99, 2008). Они отрицательны по CD45. VSEL меньше по размеру, чем HSC (3-6 против 6-8 мкм), и имеют более высокое ядерно-цитоплазматическое отношение. Ядро VSEL большое, содержит хроматин открытого типа и окружено узким кольцом цитоплазмы с многочисленными митохондриями. Следовательно, их морфология согласуется с примитивными PSC.
[0036] Абсолютное количество циркулирующих VSEL в периферической крови крайне низкое (от 1 до 2 клеток в 1 мл крови при стационарных условиях) и поэтому для их идентификации и выделения применяют специальные проточно-цитометрические протоколы. Фенотипические маркеры, используемые для идентификации VSEL, включают отрицательную экспрессию CD45 (мыши и человек), положительную экспрессию Sca-1 (мыши), CXCR4, CD133 и CD34 (мыши и человек), положительные для стволовых клеток - предшественников маркеры (другими словами Oct-4, Nanog и SSEA) и экспрессию ряда маркеров, характеризующих стволовые клетки эпибластной/зародышевой линии.
[0037] С использованием только клеточного сортера с активацией флуоресценции, сортирующий все VSEL, присутствующие в 100 мл пуповинной крови, выделение VSEL может быть завершено за 4 рабочих дня. Более эффективный и экономичный протокол трехступенчатого выделения позволяет получить 60% выход исходного количества Lin-/CD45-/CD133þ UCB-VSEL. Протокол включает лизис эритроцитов в гипотоническом растворе хлорида аммония, селекцию клеток CD133þ с помощью иммуномагнитных шариков и сортировку Lin-/CD45-/CD133þ клеток с помощью FACS с шариками маркеров размера для контроля. Выделенные клетки сильно обогащены Oct-4þ и SSEA-4þ популяцией маленьких, в высокой степени примитивных Lin-/CD45-/CD133þ клеток.
[0038] Другие методы сортировки VSEL основаны на присутствии нескольких поверхностных маркеров и диаметре клеток. Вкратце, начальный шаг - это лизис эритроцитов, позволяющий получить фракцию ядросодержащих клеток. Буфер для лизиса эритроцитов используют вместо центрифугирования с фиколом, потому что второй прогон может истощить популяцию очень маленьких клеток. Затем клетки окрашивают с антителами к Sca-1 (мышиные VSEL) или CD 133 (человеческие VSEL), пангемопоэтическому антигену (CD45), маркерам гематопоэтических линий (lin) и CXCR4 и сортируют, используя мультипараметрические системы сортировки живых стерильных клеток (MoFlo, Beckman Coulter; FACSAria, Beckton Dickinson). Этот метод использует "растянутые лимфоцитарные ворота", чтобы охватить объекты диаметром 2-10 мкм, включая приблизительно 95% VSEL.
[0039] Эндогенные стволовые клетки могут содержаться в мононуклеарной клеточной фракции из костного мозга, цельной крови, пуповинной крови, жировой ткани или других источников или могут быть очищены путем селекции по CD34, CD133, CD105, CD117, SSEA1-4, вытеснению красителя или другим специфическим антигенам стволовых клеток. Стволовые клетки могут быть выделены из цельной крови, костного мозга, пуповинной крови, жировой ткани, тканевых соскобов нейроэпителия и других источников стволовых клеток, которые могут быть разделены на суспензии отдельных клеток, например, ткань пуповины, центрифугированием в градиенте плотности, с использованием фиколл-гипак или другие коммерчески доступных градиентов. Стволовые клетки могут быть выделены из мононуклеарной клеточной фракции в результате таких процедур. В альтернативном варианте стволовые клетки могут быть обнаружены в других фракциях после центрифугирования в градиенте плотности (Stem Cells Dev. 2011 [Epub ahead of print]). Например, пуповинная кровь может быть разведена 1:1 в ФБР, осторожно разлита в Histopaque 1077 (Sigma) и отцентрифугирована при 1500 оборотов за минуту при комнатной температуре в течение 30 минут. Полученные слои, как изображено на Фигуре, могут быть далее обработаны для выделения стволовых клеток. Слой 1 - это слой тромбоцитов, слой 2 - лейкоцитарная пленка, содержащая мононуклеарные клетки, слой 3 - слой фиколла, а слой 4 - осадок эритроцитов, который также содержит VSEL. Слои 1, 2 и 3 собирают, разбавляют подходящей средой, такой как DMEM F12 с или без ФБР, и центрифугируют снова с получением клеточного осадка. Слой 4 разбавляют подходящей средой, такой как DMEM F12, и центрифугируют при 800 оборотов за минуту на протяжении 15 минут при комнатной температуре на стандартной настольной центрифуге. Стволовые клетки преимущественно выделяют из слоя 2 (лейкоцитарная пленка) и слоя 4 (осадок эритроцитов), с использованием проточно-цитометрических способов, указанных выше. На Фигуре 1 показан обычный вид слоев, полученных из цельной крови после центрифугирования в градиенте. 1 показывает тромбоциты; 2 - лейкоцитарную пленку с мононуклеарами и стволовыми клетками; 3 - фиколл, а 4 - осадок эритроцитов и стволовых клеток.
[0040] Дополнительно, поскольку профили гликозилирования и другие посттрансляционные модификации могут различаться в разных типах клеток и тканей, линии пациент-специфических стволовых клеток готовят из зрелых или соматических клеток, выделенных из органа или из клеток, эндогенно экспрессирующих биопрепарат. Смотри Rajpert-De Meyts E, et al. "Changes in the profile of simple mucin-type O-glycans and polypeptide GalNAc-transferases in human testis and testicular neoplasms are associated with germ cell maturation and tumour differentiation"., Virchows Arch, Vol.451: 805-814 (2007). Смотри Pevalova M., et al. "Post-translational modifications of tau protein". Bratisi Lek Listy, 107: 346-353 (2006).
ПОДРОБНОЕ ОПИСАНИЕ ПРЕДПОЧТИТЕЛЬНОГО ВАРИАНТА ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Продуцирование иммортализованных spPSC и ePSC
[0041] В предпочтительном варианте осуществления настоящего изобретения spPSC и ePSC являются иммортализоваными, преимущественно путем невирусной индукции большого T-антигена, обычно с использованием полиомавируса обезьяний вирус 40 (SV40). Смотри Rose, M.R. et al., (1983). "Expression of the Large T Protein of Polyoma Virus Promotes the Establishment in Culture of "Normal" Rodent Fibroblast Cell Lines". PNAS 80: 4354-4358 (1983); и Hofmann, M.C. et al. "Immortalization of зародышевые клетки and somatic testicular cells using the SV40 large T antigen" Experimental Cell Research, 201: 417-435 (1992).
Обзор варианта осуществления с использованием искусственно полученных, предпочтительно индуцированных, плюрипотентных стволовых клеток или, более предпочтительно, выделенных эндогенных плюрипотентных стволовых клеток
[0042] В предпочтительном варианте осуществления изобретения:
1. Выделяют эндогенные плюрипотентные стволовые клетки;
2. ePSC иммортализуют;
3. Иммортализованные ePSC трансфецируют целевым геном, вирусом или нуклеиновой кислотой с использованием невирусной методики;
4. Затем трансфецированные, иммортализованные ePSC индуцируют для дифференцировки в зародышевую линию клеток, предпочтительно клеток яичника, что обеспечивает возможность более эффективной экспрессии нуклеиновокислотного продукта;
5. Теперь дифференцированную клетку можно индуцировать для экспрессии соответствующего нуклеиновокислотного продукта с ранее трансфецированного вектора, содержащего целевые нуклеиновые кислоты.
[0043] В другом предпочтительном варианте осуществления изобретения:
1. Выделяют соматические клетки;
2. Соматические клетки трансформируют в индуцированные плюрипотентные стволовые клетки (iPS-клетки);
3. iPS-клетки иммортализуют;
4. Иммортализованные iPS-клетки трансфецируют целевым геном, вирусом или нуклеиновой кислотой;
5. Затем трансфецированные, иммортализованные iPS-клетки индуцируют для редифференцировки в соматические клетки, что обеспечивает более эффективную экспрессию белка;
6. Теперь редифференцированная клетка может быть индуцирована для экспрессии целевого белка с ранее трансфецированного вектора, содержащего соответствующий ген.
[0044] В еще более предпочтительном варианте осуществления изобретения:
1. Выделяют эндогенные плюрипотентные стволовые клетки;
2. Иммортализованные ePSC трансфецируют целевым геном, вирусом или нуклеиновой кислотой, используя невирусную технологию;
3. Затем трансфецированные, иммортализованные ePSC индуцируют для дифференцировки в зародышевую линию клеток, предпочтительно клеток яичника, что обеспечивает более эффективную экспрессию продукта - нуклеиновой кислоты;
4. Теперь дифференцированная клетка может быть индуцирована для экспрессии целевого продукта - нуклеиновой кислоты с ранее трансфецированного вектора, содержащего целевые нуклеиновые кислоты.
[0045] Способы реализации предпочтительных вариантов осуществления этого изобретения хорошо известны специалистам в этой области техники.
[0046] В альтернативных вариантах осуществления изобретения ePSC или spPSC возможно трансфецируют вектором, содержащим целевые нуклеиновые кислоты, перед иммортализацией. См. Du С. et al. "Generation of Variable and Fixed Length siRNA from a novel siRNA Expression Vector", Biomed. & Biophys. Res. Comm. 345: 99-105 (2006); York Zhu, патентная заявка США №12/313,554, поданная 21 ноября 2008 г. Либо ePSC, либо spPSC индуцируют для редифференцировки, затем клетки могут быть иммортализованы, и иммортализованные редифференцированные клетки трансфецируют вектором, содержащим целевые нуклеиновые кислоты. Другая возможность состоит в том, что клетки ePSC или spPSC индуцируют для редифференцировки, редифференцированные клетки трансфецируют соответствующими нуклеиновыми кислотами, и редифференцированные трансфецированные клетки возможно иммортализуют.
[0047] В предпочтительном варианте ePSC- или spPSC- клетки размножают в культуре перед редифференцировкой, предпочтительно в среде для культивирования клеток, содержащей аутологичную сыворотку человека и фактор стволовых клеток или фактор ингибирования лейкемии.
[0048] Продуцируемый полипептид или белок может быть любым полипептидом или белком. Особый интерес представляют полипептиды или белки, выбранные из группы, состоящей из эритропоэтина, фактора VIII, фактора IX, тромбина, антитела или фрагмента антитела, альфа-интерферона, альфа-интерферона 2A и 2B (смотри патенты США №№4810645 и 4874702), бета-интерферона (смотри патент США №4738931), консенсусного интерферона (смотри патент США №5661009), гормона роста, антигемофильного фактора, Г-КСФ, ГМ-КСФ, растворимого рецептора, гибридных белков, таких как гибридный или растворимый рецептор к константной области иммуноглобулина (Ig) (см. патент США №5155027), ТФР-β, костных морфогенетических белков (BMP), ТФРα, интерлейкина-2, β-глюкоцереброзидазы или ее аналогов, ингибитора альфа 1-протеиназы, фибрина, фибриногена, фактора Виллебранда, имиглюцеразы, агалзидазы-бета, ларонидазы, алглюкозидазы-альфа, альфа-тиротропина и альфа-тимозина.
[0049] Любое антитело или фрагмент антитела могут быть получены согласно процессу настоящего изобретения. Особый интерес представляют те антитела или фрагменты антител, которые связывают мишень, где указанную мишень выбырают из группы, состоящей из молекулы фактора некроза опухоли (TNF), рецептора фактора роста, молекулы фактора роста эндотелия сосудов (VEGF), интерлейкина 1, интерлейкина 4, интерлейкина 6, интерлейкина 11, интерлейкина 12, гамма-интерферона, активатора рецептора лиганда ядерного фактора каппа-В (RANKL) и Blys.
Индукция дифференцировки стволовых клеток
[0050] Для оптимизации продуцирования целевого белка трансфецированные ePSC- или spPSC-клетки следует индуцировать для дифференцировки в соматическую клетку.
[0051] В альтернативном варианте осуществления изобретения популяцию ePSC- или spPSC-клеток размножают, индуцируют дифференцировку, и дифференцированные клетки затем могут быть трансфецированы целевой нуклеиновой кислотой. Стволовые клетки индуцируют для дифференцировки по направлению к соматическим типам клеток в культуре путем добавления в нее различных факторов роста (Blood., 85: 2414-2421, 1995), изменения питательных веществ в культуральной среде, манипуляций с условиями культивирования, такими как давление кислорода (ВМС Cell Biol., 11: 94, 2010) или температура, или путем культивирования стволовых клеток на разных внеклеточных матриксах, наряду с другими способами, известными специалисту в области клеточной биологии и клеточной дифференцировки. Например, ретиноевая кислота, ТФР-β, костные морфогенетические белки (BMP), аскорбиновая кислота и бета-глицерофосфат обеспечивают получение остеобластов; индометацин, IBMX (3-изобутил-1-метилксантин), инсулин и трийодотиронин (Т3) обеспечивают получение адипоцитов; αFGF, βFGF, витамин D3, ФНО-β и ретиноевая кислота обеспечивают получение миоцитов (CARDIAC-DERIVED STEM CELLS. (WO/1999/049015) март 1998). Зародышевые клетки получают из плюрипотентных стволовых клеток, применяя монослойную культуру, образование эмбриоидных телец (EB), коагрегацию с BMP4-продуцирующими клетками и использование кондиционированной клетками яичек или яичников среды или образование EB с рекомбинантными костными морфогенетическими белками человека (BMP) (PLoS One. 2009; 4(4): е5338). Маркерные гены зародышевых клеток включают содержащий домен PR 1, с доменом ZNF (PRDM1, также известный как BLIMP1), содержащий домен PR 14 (PRDM14), белкаргининметилпротеазу 5 (PRMT5), DPPA3, IFITM3, GDF3, c-KIT, хемокиновый (C-X-C мотив) рецептор 4 (CXCR4), NANOS1-3, DAZL, VASA, семейство генов PIWI (PIWIL1 и PIWIL2, известные как HIWI и HILI у людей, соответственно), Mut-L гомолог-1 (MLH1), белок 1 синаптонемного комплекса 1 (SCP1) и SCP3. Полученные зародышевые линии клеток модифицируют для экспрессии соответствующих генных или белковых продуктов, аналогично использованию клеток яичника китайского хомячка (CHO) и других используемых в настоящее время производственных клеточных линий. Стратегии дифференцировки для получения различных линий соматических клеток из стволовых клеток на разных стадиях хорошо известны специалистам в области биологии стволовых клеток.
[0052] Известно, что факторы транскрипции, ассоциированные с высокими уровнями биологической продукции, могут быть котрансфецированы с целевым геном для оптимизации уровней экспрессии в пациент-специфической клеточной линии. Например, высокие уровни экспрессии Pit-1 могут обеспечивать экспрессию пролактина на высоком уровне в клеточном типе, одновременно блокируя или предотвращая экспрессию гормона роста (Genes Dev, 3: 946-958, 1989).
[0053] Продуцирование моноклональных антител повышают путем оптимизации кодонов гена, используя системы, такие как разработанные Sino Biological Inc, морфогенетические или другие стандартные биотехнологические методы.
[0054] Согласно настоящему изобретению рекомбинантные полипептиды и белки продуцируются в ePSC и spPSC, где spPSC и ePSC получены или выделены из клеток специфической породы или этнической группы вследствие того, что некоторые породы или этнические группы конкретных видов животных имеют разные профили гликозилирования полипептидов или белков, продуцируемых конкретной породой или этнической группой. Согласно настоящему изобретению этническая группа - это группа, представители которой имеют наследуемые обычным путем признаки, часто состоящим из общих предков или эндогамии (практика заключать браки в пределах специфической группы, например, евреи ашкенази). Как правило, это в высокой степени биологически самоподдерживающаяся группа. Примеры этнических групп, которые могут иметь различные профили гликозилирования в полипептидах и белках, продемонстрированы в Levinson, David (1998), Ethnic Groups Worldwide: A Ready Reference Handbook, Greenwood Publishing Group.
СПОСОБЫ И СИСТЕМЫ ДЛЯ ТЕРАПИИ СТВОЛОВЫМИ КЛЕТКАМИ
[0055] Настоящее изобретение также включает способ стимулирующей терапии стволовыми клетками без образования тератом. Настоящее изобретение позволяет применять наблюдаемую ′память′ репрограммированных соматических клеток, называемых в настоящей заявке искусственно полученными плюрипотентными стволовыми клетками (spPSC) для обеспечения более благоприятного терапевтического эффекта. Память spPSC, которая сообщает предпочтение редифференцировать в направлении типа клеток происхождения перед перепрограммированием, обеспечивает средство для повышения безопасности и терапевтической пользы spPSC для регенеративной медицины.
Общие шаги
[0056] Настоящее изобретение включает:
1. Выделение соответствующих соматических клеток, в частности, соматических клеток, которые желательно регенерировать;
2. Превращение (трансформацию) указанных соматических клеток в искусственно полученные плюрипотентные стволовые клетки (spPSC), в частности, индуцированные плюрипотентные стволовые клетки (iPSC), описанные выше;
3. Размножение популяции spPSC, где spPSC сохраняют присущую им эпигенетическую память о соматических клетках;
4. Редифференцировку spPSC в культуре до редифференцированных соматических клеток, имеющих оригинальный тип соматических клеток; и
5. Введение или доставку редифференцированных соматических клеток в участок тела, где желателен этот клеточный тип.
[0057] Ряд недавних исследований продемонстрировал, что spPSC-клетки сохраняют ′память′ типе клеток, из которого они произошли (Stem Cells. 28: 1981-1991, 2010), (Nature, 467 (7313): 285-90, 2010), (Nat Biotechnol, 28,: 848-855, 2010) and (Mol Hum Reprod., 16: 880-885, 2010). Это выражается в том, что spPSC-клетки в культуре предпочтительно, иногда спонтанно, редифференцируются в клетки типа, из которого они произошли. Ученые и клиницисты решительно сосредоточили внимание на получении плюрипотентных стволовых клеток, которые могут быть полезны в клинической терапии, поиске новых лекарств, моделировании болезней или скрининге токсичности (Curr Opin Biotechnol., 20: 516-521, 2009), упуская терапевтическое преимущество, заключающееся в том, что аспект памяти плюрипотентных стволовых клеток обеспечивает безопасную терапию человека. Более того, ученые, опубликовавшие данные об аспектах ′памяти′ перепрограммированных плюрипотентных стволовых клеток, сосредоточили внимание на ограничениях, которые это свойство spPSC клеток создает для терапии, и упустили важность этого свойства для обеспечения терапевтической полезности этих клеток.
[0058] Аспект памяти spPSC наблюдается для соматических клеток различного происхождения. Например, первичные фетальные эпителиальные клетки сетчатки были перепрограммированы в iPSC, с использованием лентивирусной экспрессии OCT4, SOX2, LIN28 и Nanog (Stem Cells. 28: 1981-1991, 2010), и прошли стандартные тесты на плюрипотентность; они образовывали тератомы и экспрессировали маркеры плюрипотентных стволовых клеток. После удаления основного ФРФ (фактора роста фибробластов) из питательной среды несколько линий spPSC сетчатки спонтанно редифференцировали обратно до эпителиальной клеточной линии сетчатки дифференцировки. Приблизительно 60% клеток, которые спонтанно дифференцировали от фетальных ретинальных эпителиальных spPSC клеток человека, по сравнению с 5-16% ретинальных эпителиальных клеток из spPSC из фетальных легочных фибробластов человека или ESC клеток человека. Однако, одной из трех spPSC-клеток из фетальных эпителиальных клеток сетчатки человека вообще не удалось дифференцировать до ретинальных эпителиальных клеток. Kim et al. перепрограммировали клетки-предшественники костного мозга и фибробласты кожи от старых мышей, используя ретровирусное введение Oct4, Sox2, Klf4 и Myc (Nature, 467 (7313): 285-90, 2010). Все полученные ими линии стволовых клеток демонстрировали плюрипотентность на основании критериев, обычно применяемых для человеческих образцов. Последующая дифференцировка перепрограммированных им плюрипотентных стволовых клеток продемонстрировала, что гематопоэтические исходные клетки редифференцируют до гематопоэтических линий охотнее, чем фибробластные источники, и, аналогично, фибробластные исходные клетки редифференцируют до мезенхимальных линий более охотно, чем гематопоэтические источники. Авторы также показали, что эта склонность редифференцировать предпочтительно в соматическую линию, из которой они произошли, может быть частично преодолена путем дифференцировки в гематопоэтическую линию с последующим новым раундом плюрипотентного перепрограммирования, а затем дополнительной дифференцировки. Например, перепрограммированные клетки, полученные из нервных клеток-предшественников, были дифференцированы до гематопоэтических линий, затем перепрограммированы до плюрипотентности и показали более высокое образование гематопоэтических колоний, чем предшественники нервных клеток, которые были дифференцированы до гематопоэтических клеток.
[0059] Подобные исследования предпочтения перепрограммированных клеток редифференцировать в типы клеток, из которых они получены, были выполнены с использованием невирусного перепрограммирования фетальных нейральных клеток-предшественников человека (PLoS One., 4: е7076-е7088, 2009). Полученные перепрограммированные клетки экспрессировали ряд маркеров плюрипотентности, маркеры всех трех зародышевых листков, но однако были способны образовывать эмбриоидные тельца в культуре и тератомы; используя GeneChip-анализ, авторы продемонстрировали, что перепрограммированные нервные клетки-предшественники сохранили некоторую экспрессию генов нервных стволовых клеток. Polo et al. (Nat Biotechnol, 28,: 848-855, 2010), полученные плюрипотентные перепрогамированные клетки из кончика хвоста мыши давали фибробласты, В-клетки селезенки, гранулоциты костного мозга и прекурсоры скелетных мышц. Автономные исследования дифференцировки показали, что перепрограммированные из В-клетки селезенки и гранулоцита костного мозга spPSC-клетки образуют гематопоэтические предшественники более эффективно, чем происходящие из фибробласта или скелетной мышцы spPSC-клетки. Интересно, что серийное пассирование этих разных клеточных линий spPSC привело к исчезновению генетических различий и различий в метилировании к 16 пассажу, в отличие от 4 пассажа согласно более ранним результатам. Примечательно, что этот феномен неодинакового потенциала дифференцировки не ограничивался перепрограммированными соматическими клетками, а также наблюдался для линий эмбриональных стволовых клеток, которые обнаружили отличающиеся генетические сигнатуры и спонтанное предпочтение к дифференцировке в определенные клеточные линии (Nat Biotechnol., 26: 313-315, 2008) (Hum Reprod Update., 13: 103-120, 2007) (Dev Biol., 307: 446-459, 2007) (BMC Cell Biol, 10: 44, 2009).
[0060] Стволовые клетки могут быть охарактеризованы и выделены при помощи специфических антигенов, экспрессируемых на их поверхности. Плюрипотентные стволовые клетки могут быть охарактеризованы экспрессией стадиеспецифического эмбрионального антигена (SSEA), факторов транскрипции Oct4 и Nanog и других маркеров, а также другими методами. Эмбриональную стволовую клетку, полученную из фибробластов и зрелых фибробластов и других клеток, перепрограммируют до состояния плюрипотентности путем слияния с эмбриональными стволовыми клетками (Cell 126: 652-655, 2006 and Stem Cell Rev, 2: 331-340, 2006), путем добавления 4 генов, используя методы ретровирусной трансфекции (Cell 126: 663-676, 2006), с помощью одинарной кассеты или способов бицистронной лентивирусной трансфекции (Stem Cells 27: 543-549, 2009 and Stem Cells 27: 1042-1049, 2009), и путем эндогенной стимуляции необходимых для плюрипотентности факторов транскрипции Stem Cells 27: 3053-3062, 2009 Индукция плюрипотентности также может быть достигнута путем модификации режима метилирования или полиаденелирования генома (PLoS One 4: е8419, 2009), с помощью микроРНК (Dev Biol. 344: 16-25, 2010), низкомолекулярных активаторов соответствующих факторов транскрипции, эпигенетического перепрограммирования (Regen Med. 2: 795-816, 2007 с помощью белкового перепрограммирования (Blood 116: 386-395, 2010), добавлением супернатанта культуры клеток или клеточных экстрактов из плюрипотентных клеток при культивировании, с помощью химических, радиационных или других средств генной мутации для реактивации генов плюрипотентности, и путем добавления факторов роста или цитокинов, или агентов клеточной сигнализации, которые индуцируют или поддерживают эндогенные состояния плюрипотентности. При использовании ретровирусов для перепрограммирования клеток до плюрипотентных состояний существуют опасности, которые напоминают исследования генной терапии иммунодефицита. Эксцизионные методы, такие как Cre-lox, или способы с использованием транспозона piggyBac используют для удаления ретровирусов после завершения перепрограммирования (Curr Opin Biotechnol. 20: 516-521, 2009). Способы клонирования, такие как SCNT, и партеногенез (PGA) (Cell Reprogram. 12: 105-113, 2010) также используют для получении\ перепрограммированных плюрипотентных стволовых клеток (Nature 450: 497-502, 2007).
[0061] Значительное количество ресурсов - экономических, интеллектуальных и трудовых - было вложено в обнаружение различных источников плюрипотентных стволовых клеток, в первую очередь в надежде на то, что эти стволовые клетки будут пригодны для регенеративной медицинской терапии человека. К сожалению, за исключением зрелых VSEL (Stem Cell Rev 4: 89-99, 2008), все плюрипотентные стволовые клетки, выделенные на сегодняшний день, являются не удачными из-за проблем с образованием тератом, опухолей и даже неопластических свойств. Поэтому требуется необходимо как-то применять эти плюрипотентные клетки для того, чтобы экономические, интеллектуальные и трудовые инвестициями, сделанные в течение последних 15 лет или около того, были оправданы.
Стволовые клетки могут быть индуцированы к дифференцировке по направлению к типам соматических клеток в культуре путем добавления в нее различных факторов роста (Blood 85: 2414-2421, 1995), изменения питательных веществ в культуральной среде, манипуляций с условиями культивирования, такими как давление кислорода (BMC Cell Biol. 11: 94, 2010 или температура, или путем культивирования стволовых клеток на разных внеклеточных матриксах, наряду с другими способами, известными специалисту в области клеточной биологии и клеточной дифференцировки. Например, ретиноевая кислота, ТФР-β, костные морфогенетические белки (BMP), аскорбиновая кислота и бета-глицерофосфат обеспечивают получение остеобластов; индометацин, IBMX (3-изобутил-1-метилксантин), инсулин и трийодотиронин (Т3) обеспечивают получение адипоцитов; αFGF, βFGF, витамин D3, TNF-β и ретиноевая кислота обеспечивают получение микоцитов (WO/1999/049015, март 1998). Стратегии дифференцировки для получения различных линий соматических клеток из стволовых клеток на разных стадиях хорошо известны специалистам в области биологии стволовых клеток.
[0062] Терапия стволовыми клетками исследуется и совершенствуется для лечения многих заболеваний человека. Информация о клинических испытаниях, содержащаяся на веб-сайте Национальных институтов здравоохранения www.clinicaltrials.gov, насчитывает свыше 3000 исследований стволовых клеток. Апробируемые заболевания включают: злокачественные опухоли крови, лейкемии, лимфомы, рак, остеопороз, апластическую анемию и цитопении, серповидно-клеточную болезнь и талассемию, дефицит лимбальных стволовых клеток, рак молочной железы, острый инфаркт миокарда, болезнь коронарных артерий, болезнь периферических сосудов, сердечную недостаточность, сахарный диабет 1 типа, сахарный диабет 2 типа, инсульт, повреждение спинного мозга, нейробластому, рассеянный склероз, системный склероз, красную волчанку, хроническое заживление ран, ожоги, заживление переломов, восстановление хрящей, опухоли ЦНС, остеоартрит, почечную недостаточность, болезнь Паркинсона, миеломы, диабетическую стопу, цирроз печени и билиарный цирроз печени, дилатационную кардиомиопатию, анемию, пигментную дистрофию сетчатки, болезнь Крона, диабетическую нейропатию, мастоцитоз, рак яичников, эпилепсию, миастению гравис, аутоиммунные заболевания, гранулематозную болезнь, остеонекроз, печеночную недостаточность, тазовую мышечную дисфункцию, липодистрофию, демиелинизирующее заболевание, дефекты хрящей, заболевание сетчатки глаза, волчаночный нефрит, болезнь Альцгеймера, травматическое поражение мозга, саркому, миозиты, гипергликемию, макулярную дистрофию, язвенный колит, мышечную дегенерацию и другие.
[0063] Стволовые клетки могут быть выделены, с использованием различных маркеров, известных специалистам в данной области. Например, перечень общеизвестных маркеров стволовых клеток можно найти на http://stemcells.nih.gov/info/scireport/appendixe.asp#eii. Нервные стволовые клетки можно выделить с помощью CD133; мезенхимальные стволовые и клетки-предшественники - с использованием рецептора костного морфогенетического белка (BMPR); гематопоэтические клетки - с помощью CD34, мезенхимальные стволовые клетки - с помощью комбинации маркеров CD34+Scal+Lin; гематопоэтические и мезенхимальные стволовые клетки - с помощью ckit, Stro1 или Thy1; нервные и панкреатические прогениторы - с помощью нестина; эктодермные, нервные и панкреатические прогениторы - с помощью виментина; и других маркеров.
[0064] Настоящее изобретение дополнительно обеспечивает способы безопасного перепрограммирования и редифференцировки соматических клеток для регенеративной медицины. Полезные соматические клетки могут включать полностью дифференцированные соматические клетки, клетки-предшественники или более примитивные стволовые клетки. Зависимо от органа, нуждающегося в регенеративной терапии, более примитивные стволовые клетки могут быть более или менее доступными путем пункции органа, биопсии, соскобов или хирургического доступа. Использование более примитивных стволовых клеток является предпочтительным, когда возможен доступ к этим клеткам. Менее предпочтительным является использование прогениторных клеток, но более предпочтительным, чем использование полностью дифференцированных соматических клеток.
[0065] Выделение соматических клеток из конкретного органа, намеченного для лечения, перепрограммирование этих соматических клеток, кратковременное размножение в культуре, гарантирующее сохранение врожденной ′памяти′ spPSC, последующая редифференцировка в культуре до типа клеток, из которого они получены, с последующим терапевтическим применением обеспечивают возможность использования способов на основе плюрипотентных стволовых клеток для лечения пациентов, обеспечивая пониженную опасность образования опухоли или тератомы.
[0066] Перепрограммированные соматические клетки используют между пассажами 1 и 12, более предпочтительно на пассаже 4. Перепрограммированные соматические клетки могут быть дифференцированы до клеточного типа для желательной регенерации согласно стандартным способам клеточной биологии и дифференцировки. Полученную терапевтическую клетку можно вводить внутривенно, внутриартериально, внутримышечно или другими инъекционными способами, используя стандартные инъекционные средства, которые могут включать катетеры, такие как NOGAStar или MyoStar инъекционные катетеры, или другое пригодное катетерное инъекционное устройство. В альтернативном варианте, терапевтические клетки можно вводить в ткань-мишень минимально инвазивными или менее инвазивными хирургическими способами. Терапевтические клетки можно вводить в буферной композиции, содержащей 0-15% аутологичного сывороточного альбумина человека, более предпочтительно 5%. Для лечения заболевания центральной нервной системы предпочтительно забуферить терапевтические клетки, используя аутологичную спинномозговую жидкость. Терапевтические клетки также можно вводить с использованием подложки, такой как коллаген, фибриноген, или другого экстрацелюлярного матрикса или комбинации экстрацелюлярных матриксов, или с помощью нити, например, созданной с помощью альгината, или других стандартных способов для удержания клеток на месте и сохранения контакта с органом, нуждающимся в регенеративном восстановлении.
[0067] В предпочтительном варианте вводят минимум 500 терапевтических клеток, обычно в клеточной терапии используют от 15 миллионов до 500 миллионов клеток. Более предпочтительно, от 15 миллионов до 100 миллионов клеток будут вводить для обеспечения максимальной пользы.
[0068] Для примера, для клеточной терапии неврологических заболеваний, таких как болезни Альцгеймера, Паркинсона, инсульт, болезнь Хантингтона, рассеянный склероз, паралич и другие заболевания центральной нервной системы (ЦНС), предпочтительно выделять нейроэпителий, используя жесткие эндоскопы, как известно из опубликованных источников (J Spinal Cord Med. 29: 191-203, 2006). Нервные стволовые клетки и клетки-предшественники или обкладочные нейроэпителиальные клетки (elfactory enseathing cells) выделяют из нейроэпителия способами, хорошо известными специалистам в области регенеративной медицины. В альтернативном варианте стволовые клетки нервного валика могут быть выделены из волосяных фолликулов человека (Folia Biol. 56: 149-157, 2010). В еще более предпочтительном варианте нервные стволовые клетки перепрограммируют путем добавления определенных факторов, размножают и пассируют на протяжении 4 пассажей, редифференцируют до конкретных типов клеток центральной нервной системы путем добавления определенных факторов, а затем вводят пациенту для регенеративной терапии.
[0069] Изобретение обеспечивает способы безопасного перепрограммирования и редифференцировки соматических клеток для регенеративной медицины. Применимые соматические клетки могут включать полностью дифференцированные соматические клетки, клетки-предшественники или более примитивные стволовые клетки. В зависимости от органа, нуждающегося в регенеративной терапии, более примитивные стволовые клетки могут быть более или менее доступными для получения путем пункции органа, биопсии, соскобов или хирургического доступа. Использование более примитивных стволовых клеток является предпочтительным, если возможен доступ к этим клеткам. Использование клеток-предшественников является менее предпочтительным, но более предпочтительным, чем использование полностью дифференцированных соматических клеток.
[0070] Для восстановления спинальных повреждений нейроэпителий выделяют с помощью жесткого эндоскопа, нервные стволовые клетки выделяют, используя нейросферный анализ, впервые описанный в 1992 авторами Reynolds и Weiss (Science, 255: 1707-1710, 1992), эпидермальный фактор роста (EGF) и основной фактор роста фибробластов (bFGF) добавляют к клеткам, которые несколько дней поддерживают в культуре нейроэпителия, для стимулирования роста нейросфер, нервные стволовые клетки способны возобновляться в культуре в течении 7-10 дней. Полученные нервные стволовые клетки перепрограммируют путем добавления эписомальных векторов для доставки OCT4 и NANOG с гигромициновой селекцией на протяжении 5-7 дней. Клетки пассируют и размножают до пассажа 4 с получением неинтегрированных колоний, а затем дифференцируют до обкладочных нейроэпителиальных и стволоподобных нервных клеток-предшественников, используя аутологичную спинномозговую жидкость или определенные факторы. Доступ к поврежденному спинному мозгу получают в ходе хирургической операции, используя стандартный срединный разрез и заднюю срединную миелотомию, рубцовую ткань удаляют насколько возможно, терапевтические клетки забуферивают аутологичной спинномозговой жидкостью, высеивают в биоактивную подложку и наносят непосредственно на поврежденный спинной мозг.
ПРИМЕРЫ
Пример 1
Генетическая модификация пациент-специфических искусственно полученных плюрипотентных стволовых клеток для продуцирования рекомбинантных белков
[0071] Искусственно полученные плюрипотентные стволовые клетки (spPSC), такие как SCNT или PGA, или ANT-OAR или iPSC, брали от пациентов для генетической модификации, чтобы индуцировать биологическое продуцирование. SCNT производные стволовых клеток готовили путем переноса ядер клеток пациента в подготовленную безъядерную яйцеклетку. ANT-OAR производные стволовых клеток готовили путем генетической модификации ядерных ДНК пациента перед перенесением модифицированных ядер в подготовленную безъядерную яйцеклетку. iPSC производные стволовых клеток готовили путем перепрограммирования клеток пациента, используя генетическую модификацию, активаторы плюрипотентного фактора транскрипции, эпигенетическую модификацию или другие способы, известные из уровня техники, как указано выше. Полученную пациент-специфическую синтетически произведенную линию стволовых клеток «сохраняли» как главный клеточный банк и рабочий банк клеток для последующей генетической модификации для биологического продуцирования.
[0072] Зародышевые клетки получали из плюрипотентных стволовых клеток, применяя монослойную культуру, образование эмбриоидных телец (EB), коагрегацию с BMP4-продуцирующими клетками, использование кондиционированной тестикулярными или овариальными клетками среды, или образование EB с использованием рекомбинантных человеческих костных морфогенетических белков (BMP). Зародышевые клетки индентифицировали по экспрессии маркерных генов, которые могут включать содержащий домен PR 1, с доменом ZNF (PRDM1, также известный как BLIMP 1), содержащий домен PR 14 (PRDM14), белкаргининметилпротеазу 5 (PRMT5), DPPA3, IFITM3, GDF3, c-KIT, хемокиновый (C-X-C мотив) рецептор 4 (CXCR4), NANOS1-3, DAZL, VASA, PIWI семейство генов (PIWIL1 и PIWIL2, известное как HIWI и HILI у людей, соответственно), Mut-L гомолог-1 (MLH1), белок 1 синаптонемного комплекса 1 (SCP1) и SCP3. Полученные зародышевые клетки трансфецировали соответствующим геном, таким как Фактор VIII, способами, обычно используемыми для продуцирования рекомбинантного Фактора VIII.
Пример 2
Генетическая модификация пациент-специфических искусственно полученных плюрипотентных стволовых клеток для продуцирования рекомбинантного инсулина.
[0073] Искусственно полученные плюрипотентные стволовые клетки (spPSC), такие как стволовые клетки, полученные методом SCNT или PGA, или ANT-OAR, или iPSC, от пациентов использовали для генетической модификации, чтобы индуцировать биологическое продуцирование. полученные методом SCNT стволовые клетки готовили путем переноса ядер клеток пациента в подготовленную безъядерную яйцеклетку. Полученные методом ANT-OAR стволовые клетки готовили путем генетической модификации ядерных ДНК пациента перед переносом модифицированных ядер в подготовленную безъядерную яйцеклетку. Полученные методом iPSC стволовые клетки готовили путем перепрограммирования клеток пациента, используя генетическую модификацию, активаторы плюрипотентного фактора транскрипции, эпигенетическую модификацию или другие способы, известные из уровня техники, как указано выше. Полученную пациент-специфическую синтетически произведенную линию стволовых клеток «сохраняли» как главный клеточный банк и рабочий банк клеток для последующей генетической модификации для биологического продуцирования.
[0074] Экспрессию предшественников инсулина в пациент-специфических стволовых клетках осуществляли согласно способам, обычно используемым для продуцирования инсулина в S. cerevisiae: [Kjeldsen T., et al., "Engineering-enhanced protein secretory expression in yeast with application to insulin". 21, May 2002, J Biol Chem., 277: 18245-18248 (May 2002); Zhang B., et al., "Intracellular retention of newly synthesized insulin in yeast is caused by endoproteolytic processing in the Golgi complex"., J Cell Biol., 153: 1187-1198 (June 2001); and Kristensen C. et al., "Alanine scanning mutagenesis of insulin"., J Biol Chem., 272: 12978-12983 (May 1997)], или E. Coli [Son YJ., et al. "Effects of beta-mercaptoethanol and hydrogen peroxide on enzymatic conversion of human proinsulin to insulin"., J. Microbiol Biotechnol., 18: 983-989 (May 2008)], потом обрабатывали и очищали согласно стандартным способам. Экспрессирующую предшественники инсулина пациент-специфическую клеточную линию «сохраняли» как главный клеточный банк и рабочий банк клеток для последующего продуцирования инсулина.
Пример 3
Получение бета-клеток для продуцирования инсулина с использованием генетической модификации пациент-специфических искусственно полученных плюрипотентных стволовых клеток.
[0075] Для продуцирования инсулина от пациентов брали соматические клетки для генетической модификации с получением spPSC, которые использовали для индукции биологической продукции. spPSC-производные стволовые клетки готовили путем перепрограммирования клеток пациента с использованием генетической модификации, активаторов плюрипотентного фактора транскрипции, эпигенетической модификации или других способов, известных из уровня техники. Полученную пациент-специфическую линию стволовых клеток «сохраняли» как главный клеточный банк и рабочий банк клеток для последующей генетической модификации для биологического продуцирования. Или эндогенные плюрипотентные стволовые клетки (ePSC) выделяли согласно методам, описанным выше и сохраняли.
[0076] Экспрессию предшественников инсулина в пациент-специфических стволовых клетках осуществляли согласно способам, обычно используемым для продукции инсулина в S. cerevisiae или E. coli, как описано выше. После генной трансфекции подходящими кодирующими инсулин генными конструктами клетки дифференцировали в направлении бета-клеточной линии согласно стандартным протоколам, описанным в [Shi, Y., et. Al. "Inducing embryonic stem cells to differentiate into pancreatic beta cells by a novel three-step approach with activin A and all-trans retinoic acid". Stem Cells., 23: 656-662 (2005); или Tateishi, K., et. Al. "Generation of insulin-secreting islet-like clusters from human skin fibroblasts"., J Biol Chem., 283: 31601-31607 (2008)] of the Beta Cell Biology Consortium, http://www.protocolonline.org/prot/Cell_Biology/Stem_Cells/Differentiation_of_Stem_Cell/index.html. Protocol Online. [Online] [Cited: Dec 19, 2010.]
[0077] Полученный экспрессированный биологический продукт затем обрабатывали и очищали согласно стандартным способам. Полученную пациент-специфическую, экспрессирующую предшественники инсулина, линию стволовых клеток «сохраняли» как главный клеточный банк и рабочий банк клеток для последующей генетической модификации для биологического продуцирования.
[0078] В альтернативном варианте полученные пациент-специфические стволовые клетки дифференцировали в направлении бета-клеточной линии согласно стандартным протоколам, описанным в Shi et. Al. id. или Tateishi et. Al. id of the Beta Cell Biology Consortium, см. выше. Экспрессию предшественников инсулина в дифференцированных пациент-специфических стволовых клетках осуществляли согласно способам, обычно используемым для продуцирования инсулина в S. cerevisiae: или Е. coli, как описано выше. Полученный экспрессированный биологический продукт затем обрабатывали и очищали согласно стандартным способам.
Пример 4
Выделение зрелых (соматических) клеток, продуцирующих антитела, для перепрограммирования и трансфекции для продукции терапевтических средств на основе антител.
[0079] В-клетки, продуцирующие антитела, выделяли из периферической крови, костного мозга и других легкодоступных источников гематопоэтических клеток для получения пациент-специфических продуцирующих клеточных линий для получения биопрепаратов на основе терапевтических антител. В-клетки выделяли, используя доступные наборы для определения CD19 экспрессии (StemCell Technologies). Для отбора клеток, продуцирующих максимальные уровни иммуноглобулина (Ig), применяли методы серийных разведении или клеточной сортировки. После непродолжительного размножения клональную клетку, продуцирующую высокие уровни Ig, перепрограммировали до состояния плюрипотентной клетки или клетки-предшественника, используя стандартные методы перепрограммирования. Полученные пациент-специфические стволовые клетки трансфецировали кодирующими целевое антитело генными конструктами, используя стандартные способы и методы молекулярной биологии. Полученное экспрессированное терапевтическое антитело обрабатывали и очищали согласно биотехнологическим способам на самом высоком современном уровне, общедоступным или хранимым в режиме конфиденциальности владельцами композиции для терапевтического средства на основе антитела. Способы получения желательного очищенного содержащего антитело продукта включают ионообменную хроматографию.
Пример 5
Выделение зрелых (соматических) клеток, продуцирующих антитела, для перепрограммирования и трансфекции для производства биологических терапевтических средств на основе антител с редифференцировкой до клеток, продуцирующих соматические антитела.
[0080] B-клетки, продуцирующие антитела, выделяли из периферической крови, костного мозга и других легкодоступных источников гематопоэтических клеток с целью получения пациент-специфических продуцирующих клеточных линий для продуцирования биопрепаратов на основе терапевтических антител. Для отбора клеток, продуцирующих максимальные уровни иммуноглобулина (Ig), применяли методы серийных разведении или клеточной сортировки. После непродолжительного размножения клональную клетку, продуцирующую высокие уровни Ig, перепрограммировали до состояния плюрипотентной или прогениторной клетки, используя стандартные методы перепрограммирования. Полученные пациент-специфические стволовые клетки трансфектировали желательными кодирующими антитело генными конструктами, используя стандартные способы и методы. После генной модификации линию плюрипотентных пациент-специфических клеток дифференцировали до зрелых B-клеток, продуцирующих антитело, путем культивирования клеток в присутствии CD40L, BAFF, активации Toll-подобного рецептора (TLR) [Hayashi E.A., et al. "TLR4 promotes B cell maturation: independence and cooperation with B lymphocyte-activating factor"., J Immunol., 184: 4662-4672 (2010), или других факторов созревания B-клеток, известных из уровня техники, таких как активация рецептора B-клеток (BCR) и активация семейства лигандов Notch-рецептора [Palanichamy A. et al. "Novel human transitional B cell populations revealed by B cell depletion therapy". 10, May 2009, J Immunol., Vol.182, pp.5982-5993 (2009); Thomas M.D. et al., "Regulation of peripheral B cell maturation"., Cell Immunol., 239: 92-102 (2006).
[0081] Титры и аффинность терапевтического антитела могут быть улучшены путем использования таких способов, как морфогенетические методы, описанные Li J., et al., "Human antibodies for immunotherapy development generated via a human B cell hybridoma technology"., Proc Natl Acad Sci, 103: 3557-3562 (2006). Полученное экспрессированное терапевтическое антитело обрабатывали и очищали согласно биотехнологическим способам на самом высоком современном уровне, общедоступным или хранимым в режиме конфиденциальности владельцами композиции для терапевтического средства на основе антитела.
[0082] В альтернативном варианте линию плюрипотентных пациент-специфических клеток дифференцировали до зрелых B-клеток, продуцирующих антитело, путем культивирования клеток в присутствии CD40L, BAFF, активации Toll-подобного рецептора (TLR) (смотри Hayashi, et al. id), или других факторов созревания B-клеток, известных из уровня техники, таких как активация рецептора B-клеток (BCR) и активация семейства лигандов Notch-рецептора (смотри Palanichamy A. et al. id., and Thomas, M.D. et al. id.). Титры и аффинность терапевтического антитела могут быть улучшены путем использования таких способов, как морфогенетические методы, как описано (смотри Li, et al. id.).
[0083] Полученные пациент-специфические клетки, продуцирующие антитело, трансфецировали кодирующими целевое антитело генными конструктами, используя стандартные способы и методы. Полученное экспрессированное терапевтическое антитело обрабатывали и очищали согласно биотехнологическим способам на самом высоком современном уровне, общедоступным или хранимым в режиме конфиденциальности владельцами композиции для терапевтического средства на основе антитела.
Пример 6
Получение пациент-специфических клеточных линий для продуцирования высокоактивных АЗКЦ антител.
[0084] N-ацетилглюкозаминовая (GlcNac) пост-трансляционная модификация иммуноглобулинов важна для антителозависимой клеточно-опосредованной токсичности (АЗКЦ), а нефукозилированные GlcNac остатки имеют более высокую аффинность к Fc гамма рецепторам Mori K., et al., "Non-fucosylated therapeutic antibodies: the next generation of therapeutic antibodies"., Cytotechnology., 55: 109-114 (2007).
[0085] Соответственно, когда целью является терапевтическое антитело с высокими уровнями АЗКЦ, то желательной является пациент-специфическая клеточная линия, способная переносить GlcNac на должном уровне и, желательно, оставлять нефукозилированные GlcNac. Известно, что клетки карциномы экспрессируют высокие уровни GMD, следовательно, поскольку раковые стволовые клетки и плюрипотентные клетки имеют похожие генные сигнатуры, то предполагается, что и плюрипотентные клетки также могут экспрессировать высокие уровни GMD-фермента, ответственного за пост-трансляционное присоединение GlcNac. Среди нормальных тканей, толстая кишка и поджелудочная железа экспрессируют повышенные уровни GMD. Дефицит FUT8, ответственного за фермент, который фукозилирует антитела, будет желательным в линии пациент-специфических стволовых клеток для продуцирования терапевтического антитела, такого как ритуксимаб или герцептин, эффективность которого зависит от активности АЗКЦ. Например, моноклональные антитела, продуцируемые клетками гибридомы YB2/0 крысы, имеют в 50 раз более высокую активность АЗКЦ, чем такие же антитела, продуцируемые с использованием клеток CHO. Полученные из жировой ткани стволовые клетки и зародышевые клеточные линии, также как и лимфома из B-клеток, экспрессируют более высокие чем средний уровни FUT8, тогда как гематопоэтические стволовые клетки (HSC), незрелые B-клетки, нормальные скелетные мышцы экспрессируют более низкие чем средний уровни FUT8.
[0086] Гематопоэтические стволовые клетки выделяли согласно стандартным способам из аспиратов клеточного мозга или подвергнутой аферезу цельной крови, с или без предварительной обработки агентами, мобилизующими стволовые клетки. Затем выделенные HSC перепрограммировали до плюрипотентности, как описано ранее. Полученные пациент-специфические стволовые клетки трансфецировали кодирующими целевое антитело генными конструктами, используя стандартные способы и методы. После трансфекции гена линию плюрипотентных пациент-специфических клеток дифференцировали до зрелых B-клеток, продуцирующих антитело, с помощью культуры клеток в присутствии CD40L, BAFF, активации Toll-подобного рецептора (смотри Hayashi E.A., et al., id.), или других факторов созревания B-клеток, известных из уровня техники, таких как активация рецептора B-клеток (BCR) и активация семейства лигандов Notch-рецептора (смотри Palanichamy A. et al. id., and Thomas, M.D. et al. id.). Титры и аффинность терапевтического антитела могут быть улучшены путем использования таких способов, как морфогенетические методы, как описано (смотри Li, et al. id.). Полученное экспрессированное терапевтическое антитело обрабатывали и очищали согласно биотехнологическим способам на самом высоком современном уровне, общедоступным или хранимым в режиме конфиденциальности владельцами композиции для терапевтического средства на основе антитела.
[0087] В альтернативном варианте, B-клетки, продуцирующие антитела, выделяли из периферической крови, костного мозга и других легкодоступных источников гематопоэтических клеток с целью получения пациент-специфических продуцирующих клеточных линий для продуцирования биопрепаратов на основе терапевтических антител. Для отбора клеток, продуцирующих максимальные уровни иммуноглобулина (Ig), применяли методы серийных разведении или клеточной сортировки. После непродолжительного размножения клональную клетку, продуцирующую высокие уровни Ig, перепрограммировали до состояния плюрипотентной или прогениторной клетки, используя стандартные методы перепрограммирования. Полученные пациент-специфические стволовые клетки трансфектировали желательными кодирующими антитело генными конструктами, используя стандартные способы и методы. После генной модификации линию плюрипотентных пациент-специфических клеток дифференцировали до HSC, незрелых B-клеток или клеток скелетных мышц для продуцирования терапевтических антител, которые лишены или имеют низкое содержание фукозы
[0088] Несмотря на то, что выше предпочтительный вариант осуществления изобретения проиллюстрирован и описан, возможен ряд изменений, не выходящих за рамки сущности и объема изобретения. Таким образом, объем изобретения не ограничивается раскрытием предпочтительного варианта осуществления. Наоборот, изобретение всецело должно определяться следующей формулой изобретения.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0089] Предпочтительные и альтернативные примеры настоящего изобретения подробно описаны ниже со ссылкой на следующие графические материалы:
[0090] На фигуре 1 изображены слои, полученные в результате центрифугирования цельной крови в градиенте.
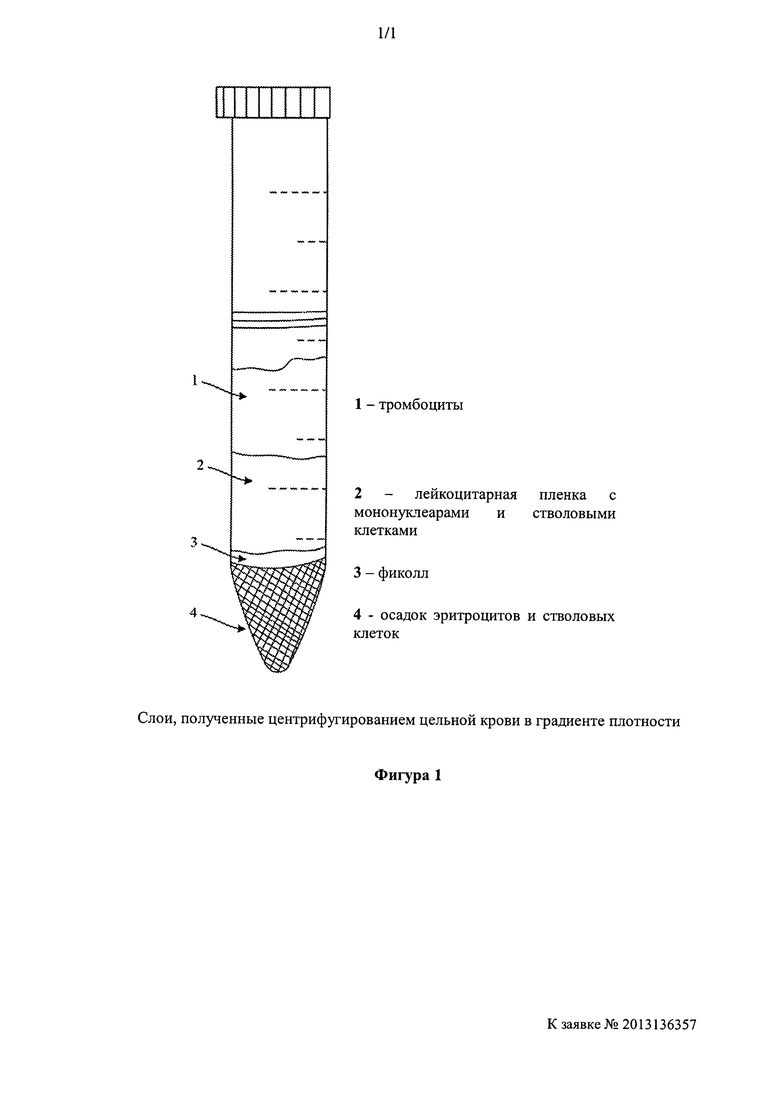
Изобретение относится к области биотехнологии и клеточной инженерии. Предложен способ получения выделенного терапевтического рекомбинантного биологического полипептида или белка для лечения заболевания. Способ предусматривает трансфецирование искусственно полученной плюрипотентной стволовой клетки (spPSC) нуклеиновой кислотой, кодирующей полипептид или белок. Далее осуществляют экспрессию целевого полипептида или белка и их последующее выделение из spPSC. При этом spPSC получена из клетки животного. 7 з.п. ф-лы, 1 ил., 6 пр.
1. Способ получения выделенного терапевтического рекомбинантного биологического полипептида или белка для лечения заболевания, включающий трансфецирование искусственно полученной плюрипотентной стволовой клетки (spPSC) нуклеиновой кислотой, кодирующей указанный терапевтический рекомбинантный биологический полипептид или белок, в условиях, при которых полипептид или белок экспрессируется указанной spPSC, причем полипептид или белок далее выделяют из указанной spPSC, и указанная spPSC получена из клетки животного.
2. Способ по п. 1, в котором терапевтический биологический полипептид или белок выбирают из группы, состоящей из эритропоэтина, фактора VIII, фактора ГХ, тромбина, антитела или фрагмента антитела, альфа-интерферона, альфа-интерферона 2А и 2В, бета-интерферона, гормона роста, антигемофильного фактора, Г-КСФ, ГМ-КСФ, растворимого рецептора, трансформирующего ростового фактора бета (ТФР-β), костных морфогенетических белков (BMP), трансформирующего ростового фактора альфа (ТФР-α), интерлейкина-2, β-глюкоцереброзидазы или ее аналогов, ингибитора альфа 1-протеиназы, фибрина, фибриногена, фактора Виллебранда, имиглюцеразы, агалзидазы-бета, ларонидазы, глюкозидазы-альфа, тиротропина-альфа и тимозина-альфа.
3. Способ по п. 1, дополнительно включающий иммортализацию трансфецированной spPSC.
4. Способ по п. 3, дополнительно включающий индуцирование дифференцировки трансфецированных, иммортализированных spPSC.
5. Способ по п. 1, дополнительно включающий индуцирование дифференцировки трансфецированных spPSC.
6. Способ по п. 5, дополнительно включающий иммортализацию трансфецированных дифференцированных клеток.
7. Способ по п. 1, в котором spPSC выбирают из группы, состоящей из трансфецированных неплюрипотентных стволовых клеток, плюрипотентных стволовых клеток, полученных путем переноса ядра соматической клетки (SCNT плюрипотентные стволовые клетки), плюрипотентных стволовых клеток, полученных путем переноса измененных ядер, перепрограммирования с использованием яйцеклетки (ANT-OAR плюрипотентные стволовые клетки) и плюрипотентных стволовых клеток, полученных путем партеногенеза (PGA плюрипотентных).
8. Способ по п. 1, отличающийся тем, что терапевтический рекомбинантный биологический полипептид или белок выбирают из группы, состоящей из гормонов роста, цитотоксических биопрепаратов и моноклональных антител.
| WO 2006061416 A1, 15.06.2006 | |||
| US 20070155011 A1, 05.07.2007 | |||
| WO 2007081631 A2, 19.07.2007 | |||
| WEN Q | |||
| ET AL., Adenoviral vector-mediated human hepatocyte growth factor gene transfection for its high expression in rabbit bone marrow-derived mesenchymal stem cells, Nan Fang Yi Ke Da Xue Xue Bao, 2007, v | |||
| Прибор с двумя призмами | 1917 |
|
SU27A1 |
| Электрический насос | 1923 |
|
SU1627A1 |
| HIMES B.T | |||
| ET AL., | |||

