Перекрестные ссылки на родственные заявки
По настоящей заявке испрашивается приоритет предварительной патентной заявки США №61/781,005, поданной 14 марта 2013, и патентной заявки США №14/106,330, поданной 13 декабря 2013, описания которых полностью включены в настоящее описание посредством ссылки.
Область техники, к которой относится настоящее изобретение
Настоящее изобретение относится главным образом к выделению, поддержанию и применению клеточных культур. В частности, оно относится к клеточным композициям и способам дифференцировки плюрипотентных стволовых клеток in vitro в панкреатические эндодермальные клеточные (ПЭК) культуры, обогащенные не-эндокринными мультипотентными панкреатическими прогениторными субпопуляциями (CHGA-), но относительно бедные клетками, коммитированными в эндокринную линию (CHGA+). Настоящее описание также раскрывает, в первую очередь, композиции клеток и способы дифференцировки плюрипотентных стволовых клеток in vitro в эндокринные клетки, которые после имплантации млекопитающему продуцируют инсулин в ответ на стимуляцию глюкозой in vivo. Настоящее описание также раскрывает композиции клеток и способы дифференцировки плюрипотентных стволовых клеток in vitro в эндокринные клетки, продуцирующие инсулин in vitro.
Предшествующий уровень техники
Разработка расширенных популяций человеческих панкреатических β-клеток, которые могут применяться для клеточной трансплантации, является главной задачей исследований диабета. В последние годы внимание было сфокусировано на применении плюрипотентных стволовых клеток в качестве потенциального обновляемого источника β-клеток. Из-за сложной природы панкреатической эндокринной дифференцировки, которая до сих пор не выяснена полностью, дифференцировка плюрипотентных стволовых клеток в зрелые и функциональные β-клетки не была эффективно достигнута in vitro. На сегодняшний день только прогениторные клетки, такие как панкреатические эндодермальные клетки (ПЭК), были трансплантированы грызунам и спустя 8-12 недель продемонстрировали функцию β-клеток, специфичную для человека. Популяции ПЭК клеток включают по меньшей мере две популяции клеток: субпопуляцию не-эндокринных мультипотентных панкреатических прогениторов (CHGA-) и клетки, коммитированные в эндокринную линию (CHGA+). Установлено, что при имплантации ПЭК субпопуляция не-эндокринных мультипотентных панкреатических прогениторов (CHGA-) созревает и образует β-клетки in vivo. Трансплантация популяций ПЭК клеток требует длительного времени созревания in vivo (8-12 недель), которое можно сократить или устранить, если: (a) in vitro ПЭК популяции содержат больше клеток того типа, который созревает в β-клетки in vivo (т.е. не-эндокринных мультипотентных панкреатических прогениторных клеток (CHGA-)), (b) терминальная (β-клеточная) дифференцировка достигается in vitro, или (с) используется популяция прогениторов in vitro, которые более продвинуты по пути дифференцировки, чем те, которые идентифицированы к настоящему времени, т.е. продвинутая в развитии клеточная популяция.
Хотя настоящая заявка не ограничивается какой-либо теорией, возможным объяснением того, почему плюрипотентные стволовые клетки не созревают в полностью функциональные β-клетки in vitro, является то, что клетки, полученные in vitro, не имеют такой экспрессии маркеров в определенные моменты времени, как при развитии панкреатических клеток млекопитающих in vivo. Например, экспрессия NGN3 при дифференцировке in vitro происходит раньше, чем при развитии панкреатических клеток млекопитающих in vivo. Действительно, наблюдалось, что в традиционных протоколах дифференцировки на 4 стадии, как описано автором настоящей заявки во многих публикациях и патентах, включенных посредством ссылки во всей полноте, ПЭК популяции экспрессируют NGN3 и NKX2.2. Таким образом, супрессия или ингибирование экспрессии NGN3 после экспрессии в PDX1 / дифференцировки NKX6.1 со-позитивной субпопуляции не-эндокринных мультипотентных панкреатических прогениторов является важным этапом достижения in vitro дифференцировки плюрипотентных стволовых клеток в зрелые и функциональные β-клетки, или дифференцировки ПЭК популяций, содержащих больше не-эндокринных мультипотентных панкреатических прогениторных клеток (CHGA-), по сравнению с клетками, коммитированными в эндокринную линию (CHGA+).
Идентификация протокола, замедляющего экспрессию NGN3 и NKX2.2, является неочевидной, поскольку протокол дифференцировки не должен прерываться, и предпочтительно повышает формирование субпопуляции не-эндокринных мультипотентных панкреатических прогениторов (CHGA-), и должен минимизировать потерю клеток; или по меньшей мере поддерживать адекватную клеточную массу и выход клеток (т.е. микрограммы, граммы, килограммы клеток).
Кроме того, поскольку экспрессия NGN3 требуется для инициации дифференцировки эндокринных клеток, стимуляция созревания островковых клеток и поддержка функции островков, стимуляция или индукция экспрессии NGN3 в клетках, коммитированных в эндокринную линию (CHGA+), является важным этапом достижения дифференцировки in vitro плюрипотентных стволовых клеток в зрелые и функциональные β-клетки.
Краткое описание настоящего изобретения
В одном аспекте обеспечивается способ получения in vitro клеточной популяции, включающей PDX1-позитивные панкреатические эндодермальные клетки. Способ включает обеспечение контакта клеточной популяции, включающей линии клеток дефинитивной энтодермы, с фактором роста суперсемейства TGFβ и членом семейства Wnt, с генерацией, таким образом, клеточной популяции, включающей PDX1-позитивные панкреатические эндодермальные клетки. В вариантах осуществления фактор роста суперсемейства TGFβ выбран из группы, состоящей из Nodal, активина А, активина В, ВМР2, ВМР4, GDF8, GDF-10, GDF-11 и GDF-15. В вариантах осуществления способ дополнительно включает обеспечение контакта клеточной популяции, включающей линию дефинитивных эндодермальных клеток, с агентом, активирующим киназу ERBB рецептора. В вариантах осуществления фактор роста суперсемейства TGFβ является активином А. В других вариантах осуществления членом семейства Wnt является Wnt3a. В вариантах осуществления агентом, активирующим киназу рецептора ERBB, является херегулин.
В одних вариантах осуществления клеточная популяция, включающая PDX1-позитивные панкреатические эндодермальные клетки, контактирует по меньшей мере с 25 нг/мл фактора роста суперсемейства TGFβ. В других вариантах осуществления клеточная популяция, включающая PDX1-позитивные панкреатические эндодермальные клетки, контактирует по меньшей мере с 50 нг/мл фактора роста суперсемейства TGFβ. В других вариантах осуществления клеточная популяция, включающая PDX1-позитивные панкреатические эндодермальные клетки, контактирует по меньшей мере с 75 нг/мл фактора роста суперсемейства TGFβ.
В одних вариантах осуществления клеточная популяция, включающая PDX1-позитивные панкреатические эндодермальные клетки, контактирует по меньшей мере с 25 нг/мл члена семейства Wnt. В других вариантах осуществления клеточная популяция, включающая PDX1-позитивные панкреатические эндодермальные клетки, контактирует по меньшей мере с 50 нг/мл члена семейства Wnt. В других вариантах осуществления клеточная популяция, включающая PDX1-позитивные панкреатические эндодермальные клетки, контактирует по меньшей мере с 75 нг/мл члена семейства Wnt.
В одних вариантах осуществления клеточная популяция контактирует с количеством агента, активирующего киназу рецептора ERBB, в 15-50 раз меньшим, чем количество каждого из фактора роста суперсемейства TGFβ и члена семейства Wnt. В одних вариантах осуществления по меньшей мере примерно 50% указанной клеточной популяции, включающей PDX1-позитивные панкреатические эндодермальные клетки, является PDX1-позитивными панкреатическими эндодермальными клетками. В других вариантах осуществления по меньшей мере примерно 60% указанной клеточной популяции, включающей PDX1-позитивные панкреатические эндодермальные клетки, является PDX1-позитивными панкреатическими эндодермальными клетками. В других вариантах осуществления по меньшей мере примерно 70% указанной клеточной популяции, включающей PDX1-позитивные панкреатические эндодермальные клетки, является PDX1-позитивными панкреатическими эндодермальными клетками. В других вариантах осуществления по меньшей мере примерно 80% указанной клеточной популяции, включающей PDX1-позитивные панкреатические эндодермальные клетки, является PDX1-позитивными панкреатическими эндодермальными клетками. В других вариантах осуществления по меньшей мере примерно 90% указанной клеточной популяции, включающей PDX1-позитивные панкреатические эндодермальные клетки, является PDX1-позитивными панкреатическими эндодермальными клетками.
В одних вариантах осуществления PDX1-позитивные панкреатические эндодермальные клетки дополнительно экспрессируют NKX6.1 и по существу не экспрессируют NGN3. В других вариантах осуществления PDX1-позитивные панкреатические эндодермальные клетки не экспрессируют CHGA. В других вариантах осуществления PDX1-позитивные панкреатические эндодермальные клетки дополнительно экспрессируют NKX6.1 и PFT1A, и по существу не экспрессируют NGN3. В других вариантах осуществления PDX1-позитивные панкреатические эндодермальные клетки являются мультипотентными клетками, которые могут дифференцироваться в унипотентные незрелые бета-клетки.
В одном аспекте обеспечивается культура человеческих панкреатических эндодермальных клеток, включающая дефинитивную эндодермальную клеточную линию и фактор роста суперсемейства TGFβ и член семейства Wnt. В одних вариантах осуществления фактор роста суперсемейства TGFβ выбран из группы, состоящей из Nodal, активина А, активина В, ВМР2, ВМР4, GDF8, GDF-10, GDF-11 и GDF-15. В других вариантах осуществления способ дополнительно включает обеспечение контакта клеточной популяции, включающей клетки дефинитивной эндодермальной линии, с агентом, активирующим киназу рецептора ERBB. В одних вариантах осуществления фактором роста суперсемейства TGFβ является активин А. В других вариантах осуществления членом семейства Wnt является Wnt3a. В других вариантах осуществления агентом, активирующим киназу рецептора ERBB, является херегулин.
В другом аспекте обеспечиваются унипотентные человеческие незрелые бета-клетки in vitro. В одних вариантах осуществления унипотентные человеческие незрелые бета-клетки экспрессируют INS и NKX6.1, и по существу не экспрессируют NGN3. В других вариантах осуществления унипотентные человеческие незрелые бета-клетки способны к дифференцировке в зрелые бета-клетки. В одних вариантах осуществления дифференцировка осуществляется in vivo. В других вариантах осуществления унипотентные человеческие незрелые бета-клетки образуют часть клеточной популяции. В других вариантах осуществления по меньшей мере 10% клеточной популяции является незрелыми бета-клетками. В других вариантах осуществления по меньшей мере 20% клеточной популяции является незрелыми бета-клетками. В других вариантах осуществления по меньшей мере 30% клеточной популяции является незрелыми бета-клетками. В других вариантах осуществления по меньшей мере 40% клеточной популяции является незрелыми бета-клетками. В других вариантах осуществления по меньшей мере 50% клеточной популяции является незрелыми бета-клетками. В других вариантах осуществления по меньшей мере 60% клеточной популяции является незрелыми бета-клетками. В других вариантах осуществления по меньшей мере 80% клеточной популяции является незрелыми бета-клетками. В других вариантах осуществления по меньшей мере 90% клеточной популяции является незрелыми бета-клетками. В других вариантах осуществления по меньшей мере 98% клеточной популяции является незрелыми бета-клетками. В других вариантах осуществления человеческие незрелые бета-клетки являются моногормональными. В других вариантах осуществления унипотентные человеческие клетки экспрессируют MAFB.
В одном аспекте обеспечивается способ получения унипотентных человеческих незрелых бета-клеток. Способ включает обеспечение контакта клеток человеческой дефинитивной эндодермальной линии in vitro членом суперсемейства TGFβ и членом семейства Wnt, с генерацией незрелых бета-клеток. В одних вариантах осуществления унипотентные человеческие незрелые бета-клетки экспрессируют INS, NKX6.1, и по существу не экспрессируют NGN3. В других вариантах осуществления член суперсемейства TGFβ выбран из группы, состоящей из Nodal, активина А, активина В, ВМР2, ВМР4, GDF8, GDF-10, GDF-11 и GDF-15. В одних вариантах осуществления способ дополнительно включает обеспечение контакта клеток человеческой дефинитивной эндодермальной линии in vitro с агентом, активирующим киназу рецептора ERBB. В других вариантах осуществления фактор роста суперсемейства TGFβ является активином А. В других вариантах осуществления членом семейства Wnt является Wnt3a. В других вариантах осуществления агент, активирующий киназу рецептора ERBB, является херегулином. В других вариантах осуществления фактором роста суперсемейства TGFβ является BMP.
В одних вариантах осуществления способ дополнительно включает обеспечение контакта клеток человеческой дефинитивной эндодермальной линии in vitro с никотинамидом, ретиноевой кислотой или аналогом ретиноевой кислоты, ингибитором rho-киназы или ингибитором гамма-секретазы. В одних вариантах осуществления ретиноевая кислота выбрана из группы, состоящей из полностью транс-ретиноевой кислоты (RA), 13-цис-ретиноевой кислоты (13-cis-RA), и аротиноидной кислоты (TTNPB). В других вариантах осуществления ретиноевая кислота является полностью транс-ретиноевой кислотой (RA). В других вариантах осуществления ретиноевая кислота является 13-цис-ретиноевой кислотой (13-cis-RA). В других вариантах осуществления ретиноевая кислота является аротиноидной кислотой (TTNPB). В других вариантах осуществления ингибитор rho-киназы выбран из группы, состоящей из Y-27632, Fasudil, Н-1152Р, Wf-536 и Y-30141. В других вариантах осуществления ингибитор rho-киназы является Y-27632.
В одних вариантах осуществления ингибитор гамма-секретазы выбран из группы, состоящей из ингибитора гамма-секретазы I (GSI I), ингибитора гамма-секретазы II (GSI II), ингибитора гамма-секретазы III (GSI III), ингибитора гамма-секретазы IV (GSI IV), ингибитора гамма-секретазы V (GSI V), ингибитора гамма-секретазы VI (GSI VI), ингибитора гамма-секретазы VII (GSI VII), ингибитора гамма-секретазы IX (GSI IX), (DAPT), ингибитора гамма-секретазы XI (GSI XI), ингибитора гамма-секретазы XII (GSI XII), ингибитора гамма-секретазы XIII (GSI XIII), ингибитора гамма-секретазы XIV (GSI XIV), ингибитора гамма-секретазы XVI (GSI XVI), ингибитора гамма-секретазы XVII (GSI XVII), ингибитора гамма-секретазы XIX (GSI XIX), ингибитора гамма-секретазы XX (GSI XX), ингибитора гамма-секретазы XXI (GSI XXI), ингибитора гамма40-секретазы I, ингибитора гамма40-секретазы II и RO4929097. В одних вариантах осуществления ингибитором гамма-секретазы является RO4929097. В других вариантах осуществления ингибитором гамма-секретазы является GSI IV.
В другом аспекте обеспечивается способ получения зрелых бета-клеток in vivo. Способ включает: (а) обеспечение клеток человеческой дефинитивной эндодермальной линии in vitro с членом суперсемейства TGFβ и членом семейства Wnt, с генерацией незрелых бета-клеток; (b) трансплантацию незрелых бета-клеток с этапа (а) млекопитающему субъекту; и обеспечение дифференцировки незрелых бета-клеток in vivo с получением популяции клеток, включающей зрелые бета-клетки. В одних вариантах осуществления способ дополнительно включает обеспечение продукции инсулина зрелыми бета-клетками в ответ на стимуляцию глюкозой. В одних вариантах осуществления фактор роста суперсемейства TGFβ выбран из группы, состоящей из Nodal, активина А и активина В, GDF-8, GDF-11 и GDF-15. В других вариантах осуществления способ дополнительно включает обеспечение контакта клеток человеческой дефинитивной эндодермальной линии in vitro агентом, активирующим киназу рецептора ERBB. В других вариантах осуществления фактором роста суперсемейства TGFβ является активин А. В других вариантах осуществления членом семейства Wnt является Wnt3a. В других вариантах осуществления агентом, активирующим киназу рецептора ERBB, является херегулин.
В одних вариантах осуществления клетки человеческой дефинитивной эндодермальной линии контактируют по меньшей мере с 25 нг/мл фактора роста суперсемейства TGFβ. В других вариантах осуществления клетки человеческой дефинитивной эндодермальной линии контактируют по меньшей мере с 50 нг/мл фактора роста суперсемейства TGFβ. В других вариантах осуществления клетки человеческой дефинитивной эндодермальной линии контактируют по меньшей мере с 75 нг/мл фактора роста суперсемейства TGFβ.
В одних вариантах осуществления клетки человеческой дефинитивной эндодермальной линии контактируют по меньшей мере с 25 нг/мл члена семейства Wnt. В других вариантах осуществления клетки человеческой дефинитивной эндодермальной линии контактируют по меньшей мере с 50 нг/мл члена семейства Wnt. В других вариантах осуществления клетки человеческой дефинитивной эндодермальной линии контактируют по меньшей мере с 75 нг/мл члена семейства Wnt.
В одних вариантах осуществления клетки человеческой дефинитивной эндодермальной линии контактируют с количеством агента, активирующего киназу рецептора ERBB, которое в 5-15 раз меньше количества фактора роста суперсемейства TGFβ и члена семейства Wnt. В других вариантах осуществления по меньшей мере 10% клеточной популяции является незрелыми бета-клетками. В других вариантах осуществления по меньшей мере 20% клеточной популяции является незрелыми бета-клетками В других вариантах осуществления по меньшей мере 30% клеточной популяции является незрелыми бета-клетками. В других вариантах осуществления по меньшей мере 40% клеточной популяции является незрелыми бета-клетками. В других вариантах осуществления по меньшей мере 50% клеточной популяции является незрелыми бета-клетками. В других вариантах осуществления по меньшей мере 60% клеточной популяции является незрелыми бета-клетками. В других вариантах осуществления по меньшей мере 70% клеточной популяции является незрелыми бета-клетками. В других вариантах осуществления по меньшей мере 80% клеточной популяции является незрелыми бета-клетками. В других вариантах осуществления по меньшей мере 90% клеточной популяции является незрелыми бета-клетками. В других вариантах осуществления незрелые бета-клетки экспрессируют INS и NKX6.1, и по существу не экспрессируют NGN3. В других вариантах осуществления незрелые бета-клетки экспрессируют INS, NKX6.1 и MAFB, и по существу не экспрессируют NGN3.
В одном аспекте обеспечиваются унипотентные человеческие незрелые бета-клетки in vitro. В одних вариантах осуществления унипотентные человеческие незрелые бета-клетки экспрессируют INS и NKX6.1, и по существу не экспрессируют NGN3. В одних вариантах осуществления унипотентные человеческие незрелые бета-клетки способны к созреванию в зрелые бета-клетки.
В одном аспекте обеспечивается способ получения зрелых бета-клеток in vivo. Способ включает: (а) обеспечение контакта клеток человеческой дефинитивной эндодермальной линии in vitro с членом суперсемейства TGFβ и членом семейства Wnt, с генерацией незрелых бета-клеток; (b) трансплантацию незрелых бета-клеток с этапа (а) млекопитающему субъекту; и (с) обеспечение созревания незрелых бета-клеток in vivo с получением популяции клеток, включающей инсулин-секретирующие бета-клетки.
В одном аспекте обеспечиваются унипотентные человеческие незрелые бета-клетки, экспрессирующие INS и NKX6.1, и по существу не экспрессирующие NGN3. В одних вариантах осуществления унипотентные человеческие незрелые бета-клетки способны созревать в зрелые бета-клетки. В других вариантах осуществления унипотентные человеческие незрелые бета-клетки дополнительно экспрессируют MAFB.
В одном аспекте обеспечиваются in vitro унипотентные человеческие незрелые бета-клетки, экспрессирующие INS и NKX6.1, и по существу не экспрессирующие NGN3. В одних вариантах осуществления унипотентные человеческие незрелые бета-клетки способны созревать в зрелые бета-клетки. В других вариантах осуществления унипотентные человеческие незрелые бета-клетки дополнительно экспрессируют MAFB.В одном аспекте обеспечивается способ получения унипотентных человеческих незрелых бета-клеток. Способ включает обеспечение контакта клеток человеческой дефинитивной эндодермальной линии in vitro с членом суперсемейства TGFβ и членом семейства Wnt, с получением унипотентных человеческих незрелых бета-клеток. В одних вариантах осуществления унипотентные человеческие незрелые бета-клетки экспрессируют INS, NKX6.1, и по существу не экспрессируют NGN3. В других вариантах осуществления способ дополнительно включает обеспечение контакта клеток человеческой дефинитивной эндодермальной линии in vitro с агентом, активирующим киназу рецептора ERBB. В других вариантах осуществления фактором роста суперсемейства TGFβ является активин А. В других вариантах осуществления членом семейства Wnt является Wnt3a. В других вариантах осуществления агентом, активирующим киназу рецептора ERBB, является херегулин.
В одном аспекте обеспечивается способ получения зрелых бета-клеток in vivo. Способ включает: (а) обеспечение контакта клеток человеческой дефинитивной эндодермальной линии in vitro с членом суперсемейства TGFβ и членом семейства Wnt, с генерацией незрелых бета-клеток; (b) трансплантацию незрелых бета-клеток с этапа (а) млекопитающему субъекту; и (с) обеспечение созревания незрелых бета-клеток in vivo с получением популяции клеток, включающей инсулин-секретирующие бета-клетки. В одних вариантах осуществления способ дополнительно включает обеспечение контакта клеток человеческой дефинитивной эндодермальной линии in vitro с агентом, активирующим киназу рецептора ERBB. В других вариантах осуществления фактором роста суперсемейства TGFβ является активин А. В других вариантах осуществления членом семейства Wnt является Wnt3a. В других вариантах осуществления агентом, активирующим киназу рецептора ERBB, является херегулин. В других вариантах осуществления клетки человеческой дефинитивной эндодермальной линии контактируют по меньшей мере с 50 нг/мл фактора роста суперсемейства TGFβ. В других вариантах осуществления клетки человеческой дефинитивной эндодермальной линии контактируют по меньшей мере с 25 нг/мл члена семейства Wnt. В других вариантах осуществления клетки человеческой дефинитивной эндодермальной линии контактируют с количеством агента, активирующего киназу рецептора ERBB, которое в 5-15 раз меньше количества фактора роста суперсемейства TGFβ и члена семейства Wnt. В других вариантах осуществления незрелые бета-клетки экспрессируют INS и NKX6.1, и по существу не экспрессируют NGN3. В других вариантах осуществления незрелые бета-клетки экспрессируют INS, NKX6.1 и MAFB, и по существу не экспрессируют NGN3.
Один вариант осуществления обеспечивает композицию и способ дифференцировки плюрипотентных стволовых клеток или клеток-предшественников, полученных из плюрипотентных стволовых клеток, в эндокринные и не-эндокринные популяции. В одном аспекте эндокринные клетки экспрессируют по меньшей мере CHGA (или CHGA+), а не эндокринные клетки не экспрессируют CHGA (или CHGA-). В одном аспекте эти клеточные популяции обозначают как эндокринные (CHGA+) и неэндокринные (CHGA-) субпопуляции, или просто CHGA+ или CHGA- субпопуляции, или просто эндокринные и не-эндокринные субпопуляции. В другом аспекте эти эндокринные и не-эндокринные субпопуляции могут быть субпопуляциями мультипотентных прогениторов/предшественников, такими как субпопуляции не-эндокринных мультипотентных панкреатических прогениторов или субпопуляции эндокринных мультипотентных панкреатических прогениторов; или они могут быть унипотентными субпопуляциями, такими как незрелые эндокринные клетки, предпочтительно, незрелые бета-клетки, незрелые глюкагоновые клетки, и тому подобные.
Один вариант осуществления обеспечивает композицию и способ дифференцировки плюрипотентных стволовых клеток in vitro в панкреатические эндодермальные клеточные (ПЭК) культуры, включающие как эндокринные, так и эндокринные субпопуляции, где подавляется экспрессия NGN3. В одном аспекте экспрессия NKX2.2 также подавлена в ПЭК культуре. В одном аспекте экспрессия CHGA также подавлена в ПЭК культуре. В одном аспекте экспрессия PDX1 и NKX6.1 совместно проявляется на одной клетке в ПЭК культуре. В одном аспекте менее 50%, предпочтительно меньше 40%, 30%, более предпочтительно меньше 20%, 105, 5%, 2%, 1% клеток в ПЭК культуре экспрессируют NGN3, NKX2.2 или CHGA. В одном аспекте ПЭК культура обогащена субпопуляцией неэндокринных мультипотентных панкреатических прогениторов (CHGA-). В одном аспекте ПЭК культура экспрессирует маркеры (PDX1 и/или NKX6.1) для субпопуляции неэндокринных мультипотентных панкреатических прогениторов (CHGA-). В одном аспекте ПЭК культуру с обильной субпопуляцией не-эндокринных мультипотентных панкреатических прогениторов (CHGA-) имплантируют млекопитающему хозяину, и трансплантат созревает и продуцирует инсулин in vivo в ответ на глюкозу. В одном аспекте ПЭК культура с обильной субпопуляцией не-эндокринных мультипотентных панкреатических прогениторов (CHGA-) созревает in vivo быстрее, чем ПЭК культуры без обильной субпопуляции не-эндокринных мультипотентных панкреатических прогениторов (CHGA-). В одном аспекте ПЭК культура с обильной субпопуляцией неэндокринных мультипотентных панкреатических прогениторов (CHGA-) имеет по меньшей мере в 2 раза улучшенную функцию in vivo, оцениваемую путем измерения продукции С-пептида (инсулина) в ответ на стимуляцию глюкозой в течение примерно 10 недель или примерно 12 недель после имплантации, чем у имплантированных ПЭК культур без обильной субпопуляции не-эндокринньк мультипотентных панкреатических прогениторов (CHGA-).
Один вариант осуществления обеспечивает композицию и способ дифференцировки плюрипотентных человеческих стволовых клеток in vitro, где экспрессия генов, типичных для эндокринной дифференцировки и развития, таких как NGN3 и NKX2.2, подавлена до последующей продукции не-эндокринных мультипотентных панкреатических прогениторов (CHGA- и PDX1 / NKX6.1 совместная экспрессия).
Один вариант осуществления обеспечивает способ дифференцировки плюрипотентных стволовых клеток in vitro в ПЭК культурах, включающий этапы: (а) получения PDX1-негативной популяции клеток энтодермы передней части пищеварительного тракта; (b) обеспечения контакта клеточной популяции с этапа (а) с агентом, активирующим член семейства рецептора TGFβ, с генерацией клеточной культуры, содержащей ПЭК, где ПЭК по существу не экспрессируют маркер, выбранный из группы, включающей CHGA, NGN3 или NKX2.2. В одном аспекте ПЭК включают субпопуляцию клеток, совместно экспрессирующих PDX1 и NKX6.1. В одном аспекте ПЭК включают субпопуляцию не-эндокринных мультипотентных панкреатических прогениторных (CHGA-) клеток. В одном аспекте агент, активирующий член семейства рецептора TGFβ, выбран из группы, состоящей из Nodal, активина А, активина В, ВМР2, ВМР4, GDF8, GDF-10, GDF-11, GDF-15 и их смеси. В одном аспекте активин А добавляют в количестве 100 нг/мл, 75 нг/мл, 50 нг/мл, 25 нг/мл или 10 нг/мл на стадии 3; и 15 нг/мл, 10 нг/мл или 5 нг/мл на стадии 4. В одном аспекте ПЭК культура, образованная путем добавления активина А к популяции PDX1-негативных клеток энтодермы передней части пищеварительного тракта, имеет по меньшей мере 2-кратное улучшение функции in vivo, оцениваемой по продукцию С-пептида (инсулина) в ответ на стимуляцию глюкозой, по сравнению с ПЭК культурами без обильной субпопуляции не-эндокринных мультипотентных прогениторов (CHGA-).
В одном аспекте ПЭК культура с субпопуляцией не-эндокринных мультипотентных прогениторов (CHGA-) имеет по меньшей мере 2-кратное улучшение функции in vivo, оцениваемой по продукцию С-пептида (инсулина) в ответ на стимуляцию глюкозой, по сравнению с ПЭК клеточными культурами без обильной субпопуляции не-эндокринных мультипотентных прогениторов (CHGA-).
Один вариант осуществления обеспечивает способ дифференцировки плюрипотентных стволовых клеток in vitro в клеточную популяцию, содержащую ПЭК, включающий этапы: (а) получения PDX1-негативной популяции клеток энтодермы передней части пищеварительного тракта; (b) обеспечения контакта клеточной популяции с этапа (а) с агентом, активирующим член семейства рецептора TGFβ, с генерацией клеточной культуры, содержащей ПЭК, где ПЭК по существу не экспрессируют маркер, выбранный из группы, включающей CHGA, NGN3 или NKX2.2. В одном аспекте ПЭК популяция включает клетки, совместно экспрессирующие PDX1 и NKX6.1. В одном аспекте популяция ПЭК включает субпопуляцию не-эндокринных мультипотентных панкреатических прогениторных (CHGA-) клеток. В одном аспекте агент, активирующий член семейства рецептора TGFβ, выбран из группы, состоящей из Nodal, активина А, активина В, ВМР2, ВМР4, GDF8, GDF-10, GDF-11, GDF-15 и их смеси. В одном аспекте агент, активирующий член семейства рецептора TGFβ, является активином А. В одном аспекте активин А добавляют в количестве 100 нг/мл, 75 нг/мл, 50 нг/мл, 25 нг/мл или 10 нг/мл на стадии 3; и 15 нг/мл, 10 нг/мл или 5 нг/мл на стадии 4. В одном аспекте лиганд семейства EGF выбран из группы, включающий херегулин (также известный как нейрегулин-1 (NRG1)), нейрегулин-2 (NRG2), нейрегулин-3 (NRG3), нейрегулин-4 (NRG4), EGF (эпидермальный фактор роста) и бета-целлюлин. В одном аспекте лигандом семейства EGF является херегулин. В одном аспекте херегулин добавляют в количестве 2 нг/мл, 5 нг/мл, 10 нг/мл, 20 нг/мл, 15 нг/мл, 30 нг/мл, 40 нг/мл, 50 нг/мл, 60 нг/мл, 70 нг/мл или 80 нг/мл. В одном аспекте ПЭК культура, образованная путем добавления активина А и херегулина к популяции PDX1-негативных клеток энтодермы передней части пищеварительного тракта, имеет по меньшей мере 2-кратное улучшение функции in vivo, измеряемой по продукции С-пептида (инсулина) в ответ на стимуляцию глюкозой, по сравнению с ПЭК культурами без обильных субпопуляций не-эндокринных мультипотентных прогениторов (CHGA-).
Один вариант осуществления обеспечивает способ дифференцировки плюрипотентных стволовых клеток in vitro в клеточную популяцию, содержащую ПЭК, включающий этапы: (а) получения PDX1-негативной популяции клеток энтодермы передней части пищеварительного тракта; (b) обеспечения контакта клеточной популяции с этапа (а) по меньшей мере с агентом, активирующим член семейства рецептора TGFβ, с лигандом семейства EGF или агентом, активирующим путь передачи сигналов Wnt, по отдельности или в комбинации, с генерацией клеточной культуры, содержащей ПЭК, где ПЭК по существу не экспрессируют маркер, выбранный из группы, включающей CHGA, NGN3 или NKX2.2. В одном аспекте ПЭК популяция включает клетки, совместно экспрессирующие PDX1 и NKX6.1. В одном аспекте ПЭК клеточная культура включает субпопуляцию не-эндокринных мультипотентных панкреатических прогениторных (CHGA-) клеток. В одном аспекте агент, активирующий член семейства рецептора TGFβ, выбран из группы, состоящей из Nodal, активина А, активина В, ВМР2, ВМР4, GDF8, GDF-10, GDF-11, GDF-15 и их смеси. В одном аспекте агент, активирующий член семейства рецептора TGFβ, является активином А. В одном аспекте агент, активирующий путь передачи сигнала Wnt, является членом семейства Wnt, выбранным из группы, включающей WNT1, WNT2, WNT2B, WNT3, WNT3A, WNT4, WNT5A, WNT5B, WNT6, WNT7A, WNT7B, WNT8A, WNT8B, WNT9A, WNT9B, WNT10A, WNT10B, WNT11, WNT16 и Wnt3A. В другом аспекте активин А добавляют в количестве 100 нг/мл, 75 нг/мл, 50 нг/мл, 25 нг/мл или 10 нг/мл на стадии 3; и 15 нг/мл, 10 нг/мл или 5 нг/мл на стадии 4. В одном аспекте херегулин добавляют в количестве 2 нг/мл, 5 нг/мл, 10 нг/мл, 20 нг/мл, 15 нг/мл, 30 нг/мл, 40 нг/мл, 50 нг/мл, 60 нг/мл, 70 нг/мл или 80 нг/мл. В одном аспекте WNT добавляют в количестве от 5 нг/мл, 25 нг/мл, 50 нг/мл до 75 нг/мл, более предпочтительно, 50 нг/мл.
В одном аспекте активин А добавляют по отдельности или в комбинации с WNT на стадии 3. В другом аспекте активин А добавляют с WNT, или WNT и херегулином на стадии 3. В еще одном аспекте активин А добавляют по отдельности или в комбинации с херегулином на стадии 4.
В одном аспекте более 10%, предпочтительно, больше 20%, 30%, 40%, и более предпочтительно, больше 50%, 60%, 70%, 80%, 90%, 95%, 98% или 100% клеток в клеточной популяции являются субпопуляцией не-эндокринных мультипотентных прогениторов (CHGA-). В одном аспекте культура ПЭК клеток, образованная путем добавления активина А, херегулина и WNT, к популяции PDX1-негативных клеток энтодермы передней части пищеварительного тракта, имеет по меньшей мере 2-кратное улучшение функции in vivo, измеряемой по продукции инсулина в ответ на стимуляцию глюкозой, по сравнению с ПЭК клеточными культурами без обильных субпопуляций неэндокринных мультипотентных прогениторов (CHGA-).
Один вариант осуществления обеспечивает композицию и способ дифференцировки плюрипотентных стволовых клеток in vitro в ПЭК культуры, включающие минимум клеток, коммитированных в эндокринную линию (CHGA+). В одном аспекте клетки, коммитированные в эндокринную линию, характеризуются экспрессией CHGA. В одном аспекте меньше 50%, предпочтительно меньше 40%, 30%, более предпочтительно, меньше 20%, 10%, 5%, 2%, 1% клеток из клеток в ПЭК культуре являются клетками, коммитированными в эндокринную линию (CHGA+).
Один вариант осуществления обеспечивает композицию и способ дифференцировки плюрипотентных стволовых клеток in vitro по существу до культур ПЭК, и далее дифференцировки культуры ПЭК в эндокринные клетки in vitro. В одном аспекте эндокринные клетки могут продуцировать инсулин in vitro. В одном аспекте эндокринные клетки, секретирующие инсулин in vitro, могут продуцировать инсулин в ответ на стимуляцию глюкозой. В одном аспекте больше 10%, предпочтительно, больше 20%, 30%, 40%, и более предпочтительно, больше 50%, 60%, 70%, 80%, 90%, 95%, 98% или 100% клеток в клеточной популяции являются эндокринными клетками.
Один вариант осуществления обеспечивает способ дифференцировки плюрипотентных стволовых клеток in vitro в эндокринные клетки, включающий этапы: (а) получения клеточной популяции, включающей ПЭК; (b) обеспечение контакта клеточной популяции с этапа (а) с факторами роста, выбранными из ингибитора хэджхог, тромбоцитарного фактора роста (PDGF), члена семейства факторов роста фибробластов, ретиноида, инсулиноподобного фактора роста 1 и 2 (IGF1 и IGF2) и никотинамида, с получением эндокринных клеток. В одном аспекте эндокринные клетки экспрессируют CHGA. В одном аспекте эндокринные клетки являются незрелыми инсулин-продуцирующими бета-клетками in vitro. В одном аспекте эндокринные клетки являются загружаемыми в имплантируемое полупроницаемое инкапсулирующее устройство, зрелыми и способными к продукции инсулина в ответ на стимуляцию глюкозой in vivo. В одном аспекте больше чем 10%, предпочтительно больше чем 20%, 30%, 40%, и более предпочтительно, больше чем 50%, 60%, 70%, 80%, 90%, 95%, 98% или 100% клеток в клеточной популяции являются эндокринными клетками. В одном аспекте SSH добавляют в количестве от 10 нг/мл до 100 нг/мл. В одном аспекте PDGF добавляют в количестве от 10 нг/мл до 100 нг/мл. В одном аспекте BMP добавляют в количестве от 10 нг/мл до 20 нг/мл. В одном аспекте FGF2 добавляют в количестве от 2 нг/мл до 20 нг/мл. В одном аспекте IGF1 или IGF2 добавляют в количестве от 25 нг/мл до 100 нг/мл. В одном аспекте вышеуказанные соединения добавляют в комбинации друг с другом. В одном аспекте членом семейства факторов роста фибробластов является FGF2. В одном аспекте ретиноид является ретиноевой кислотой и/или аналогами ретиноевой кислоты, такими как TTNPB или ТТ3, инсулиноподобный фактор роста I (IGF-I или IGF1) и -II (IGF-II или IGF2). В одном аспекте костным морфогенетическим белком является ВМР4. В одном аспекте ингибитором хэджхог является sonic Hedgehog, KAAD-циклопамин, аналоги KAAD-циклопамина, иервин, аналоги иервина; антитела, блокирующие путь хэджхог; и любые другие ингибиторы функции пути хэджхог, известные рядовому специалисту в данной области техники.
Один вариант осуществления обеспечивает способ дифференцировки плюрипотентных стволовых клеток in vitro в эндокринные клетки, включающий этапы: (а) получения клеточной популяции, содержащей ПЭК; (b) обеспечения контакта клеточной популяции с этапа (а) с ингибитором гамма-секретазы, с получением эндокринных клеток in vitro. В одном аспекте эндокринные клетки экспрессируют CHGA. В одном аспекте эндокринными клетками являются незрелые инсулин-продуцирующие бета-клетки in vitro. В одном аспекте эндокринными клетками являются инсулин-секретирующие клетки, которые продуцируют инсулин в ответ на стимуляцию глюкозой in vitro. В одном аспекте эндокринные клетки являются загружаемыми в имплантируемое полупроницаемое инкапсулирующее устройство, зрелыми и способными к продукции инсулина в ответ на стимуляцию глюкозой in vivo. В одном аспекте ингибитор гамма-секретазы выбран из группы, включающей N-[N-(3,5-дифторфенацетил-L-аланил)]-S-фенилглицин t-бутиловый эфир (DAPT), RO44929097, 1-(S)-эндо-N-(1,3,3)-триметилбицикло[2.2.1]гепт-2-ил)-4-фторфенил сульфонамид, WPE-III31C, S-3-[N'-(3,5-дифторфенил-альфа-гидроксиацетил)-L-аланилил]амино-2,3-дигидро-1-метил-5-фенил-1Н-1,4-бензодиазепин-2-он, (N)-[(S)-2-гидрокси-3-метил-бутирил]-1-(L-аланинил)-(S)-1-амино-3-метил-4,5,6,7-тетрагидро-2Н-3-бензазепин-2-он, BMS-708163 (Авагацестат), BMS-708163, Семагацестат (LY450139), Семагацестат (LY450139), MK-0752, MK-0752, YO-01027, YO-01027 (Дибензазепин, DBZ), LY-411575, LY-411575, или LY2811376. В некоторых вариантах осуществления ингибитор гамма-секретазы присутствует в клеточной культуре или клеточной популяции в концентрации в диапазоне примерно от 0,01 мкМ до 1000 мкМ. В предпочтительных вариантах осуществления ингибитор гамма-секретазы присутствует в клеточной культуре или клеточной популяции в концентрации в диапазоне примерно от 0,1 мкМ до 100 мкМ. В одном аспекте ингибитор гамма-секретазы удаляют после добавления.
Варианты осуществления, описанные в настоящей заявке, обеспечивают композиции и способы дифференцировки плюрипотентных человеческих стволовых клеток in vitro в эндокринные клетки. В одном аспекте эндокринные клетки экспрессируют CHGA. В одном аспекте эндокринные клетки могут продуцировать инсулин in vitro. В одном аспекте эндокринные клетки являются незрелыми эндокринными клетками, такими как незрелые бета-клетки. В одном аспекте клетки, продуцирующие инсулин in vitro, могут продуцировать инсулин с ответ на стимуляцию глюкозой.
Один вариант осуществления обеспечивает способ получения инсулина in vivo у млекопитающего, включающий: (а) загрузку популяции эндокринных клеток в имплантируемое полупроницаемое устройство; (b) имплантацию устройства с эндокринными клетками млекопитающему хозяину; и (с) созревание популяции эндокринных клеток в указанном устройстве in vivo, где по меньшей мере некоторые из эндокринных клеток являются инсулин-секретирующими клетками, продуцирующими инсулин в ответ на стимуляцию глюкозой in vivo, с продукцией инсулина in vivo у млекопитающего. В одном аспекте эндокринная клетка получена из клеточной композиции, содержащей ПЭК с более высокой субпопуляцией не-эндокринных мультипотентных панкреатических прогениторов (CHGA-). В другом аспекте эндокринная клетка получена из клеточной композиции, содержащей ПЭК с уменьшенной эндокринной субпопуляцией (CHGA+). В другом аспекте эндокринная клетка является незрелой эндокринной клеткой, предпочтительно, незрелой бета-клеткой.
В одном аспекте эндокринные клетки, полученные in vitro из плюрипотентных стволовых клеток, экспрессируют больше PDX1 и NKX6.1, по сравнению с PDX-1-позитивными панкреатическими эндодермальными популяциями, или не-эндокринными (CHGA-) субпопуляциями, которые являются PDX1/NKX6.1 позитивными. В одном аспекте эндокринные клетки, полученные in vitro из плюрипотентных стволовых клеток, экспрессируют PDX1 и NKX6.1 относительно больше, чем субпопуляция ПЭК неэндокринных мультипотентных панкреатических прогениторов (CHGA-). В одном аспекте костный морфогенетический белок (BMP) и аналог ретиноевой кислоты (RA) по отдельности или в комбинации добавляют в клеточную культуру для получения эндокринных клеток с повышенной экспрессией PDX1 и NKX6.1, по сравнению с субпопуляцией ПЭК не-эндокринных мультипотентных панкреатических прогениторов (CHGA-). В одном аспекте BMP выбраны из группы, включающей ВМР2, ВМР5, ВМР6, ВМР7, ВМР8 и ВМР4, и более предпочтительно, ВМР4. В одном аспекте аналог ретиноевой кислоты выбран из группы, включающей полностью транс-ретиноевую кислоту и TTNPB (4-[(Е)-2-(5,6,7,8-тетрагидро-5,5,8,8-тетраметил-2-нафталенил)-1-пропенил]бензойную кислоту, аротиноидную кислоту), или 0,1-10 мкМ АМ-580 (4-[(5,6,7,8-тетрагидро-5,5,8,8-тетраметил-2-нафталенил)карбоксамидо]бензойную кислоту), и более предпочтительно, TTNPB.
В одном варианте осуществления обеспечивается способ дифференцировки плюрипотентных стволовых клеток in vitro в эндокринные и незрелые эндокринные клетки, предпочтительно, незрелые бета-клетки, включающий диссоциацию и ре-ассоциацию агрегатов. В одном аспекте диссоциацию и ре-ассоциацию осуществляют на стадии 1, стадии 2, стадии 3, стадии 4, стадии 5, стадии 6 или стадии 7, или их комбинации. В одном аспекте клетки дефинитивной энтодермы, PDX1-негативной энтодермы передней части пищеварительного тракта, PDX1-позитивной энтодермы передней части пищеварительного тракта, ПЭК и/или эндокринные клетки и эндокринные прогениторы/предшественники являются диссоциированными и ре-ассоциированными. В одном аспекте на стадии 7 диссоциированные и ре-ассоциированные клеточные агрегаты состоят из меньшего числа не-эндокринных (CHGA-) субпопуляций, по сравнению с эндокринными (CHGA+) субпопуляциями. В одном аспекте более 10%, предпочтительно, больше 20%, 30%, 40%, и более предпочтительно, больше 50%, 60%, 70%, 80%, 90%, 95%, 98% или 100% клеток в клеточной популяции являются эндокринными (CHGA+) клетками.
В одном варианте осуществления обеспечивается способ дифференцировки плюрипотентных стволовых клеток in vitro в эндокринные клетки путем удаления эндокринных клеток, полученных во время стадии 4 продукции ПЭК, с обогащением, таким образом, субпопуляции не-эндокринных мультипотентных панкреатических прогениторов (CHGA-), которые являются PDX1+ и NKX6.1+.
В одном варианте осуществления обеспечивается способ дифференцировки плюрипотентных стволовых клеток in vitro в эндокринные клетки, где клетки со стадии 6 и стадии 7, как показано в Таблице 17, культивируют с низкими уровнями матригеля примерно от 0,02% до 2%, предпочтительно, примерно от 0,02% до 0,05%, предпочтительно, примерно от 0,05% до 1%, предпочтительно 0,05%.
В одном варианте осуществления обеспечивается способ улучшения клеточной адгезии ПЭК и агрегатов эндокринных клеток путем обработки клеточных культур матригелем и ингибитором rho-киназы или ROCK. В одном варианте осуществления ингибитором rho-киназы или ROCK является Y-27632.
В одном варианте осуществления культуры ПЭК, обогащенные субпопуляцией неэндокринных мультипотентных прогениторов (CHGA-), получают, не добавляя член семейства ноггина на стадии 3 и/или стадии 4. В одном варианте осуществления культуры ПЭК, относительно бедные клетками, коммитированными в эндокринную линию (CHGA+), получают, не добавляя член семейства ноггина на стадии 3 и/или стадии 4. В одном аспекте член семейства ноггина является соединением, выбранным из группы, включающей ноггин, хордин, фоллистатин, фоллистатин-подобные белки, Cerberus, Coco, Dan, гремлин, склеростин, PRDC (белок, связанный с Dan и Cerberus).
В одном варианте осуществления обеспечен способ поддержки эндокринных клеток в культуре, путем их культивирования в средах, содержащих высокие уровни экзогенной глюкозы, где экзогенную глюкозу добавляют в количестве примерно от 1 мМ до 25 мМ, примерно от 1 мМ до 20 мМ, примерно от 5 мМ до 15 мМ, примерно от 5 мМ до 10 мМ, примерно от 5 мМ до 8 мМ. В одном аспекте средой является DMEM, CMRL или среда на основе RPMI.
В одном варианте осуществления обеспечен способ дифференцировки плюрипотентных стволовых клеток in vitro в эндокринные клетки с диссоциацией и ре-ассоциацией клеточных агрегатов, и без них. В одном аспекте недиссоциированные или диссоциированные и ре-ассоциированные клеточные агрегаты криоконсервируют или замораживают на стадии 6 и/или стадии 7 без влияния на функцию in vivo эндокринных клеток. В одном аспекте криоконсервированные культуры эндокринных клеток оттаивают, культивируют, и при трансплантации функционируют in vivo.
В другом варианте осуществления обеспечивают систему культивирования для дифференцировки плюрипотентных стволовых клеток в эндокринные клетки, включающую по меньшей мере агент, способный к супрессии или ингибированию экспрессии эндокринных генов во время ранних стадий дифференцировки, и агент, способный к индукции экспрессии эндокринных генов во время поздних стадий дифференцировки. В одном аспекте агент, способный к супрессии или ингибированию экспрессии эндокринных генов, добавляют в систему культивирования, состоящую из панкреатических PDX1-негативных клеток передней части пищеварительного тракта. В одном аспекте агент, способный к индукции экспрессии эндокринных генов, добавляют в систему культивирования, состоящую из PDX1-позитивных панкреатических эндодермальных прогениторов или ПЭК. В одном аспекте агент, способный к супрессии или ингибированию экспрессии эндокринных генов, является агентом, активирующим семейство рецепторов TGFβ, предпочтительно, он является активином, предпочтительно, он является активином с высокими уровнями, а затем активином с низкими уровнями. В одном аспекте агент, способный к индукции экспрессии эндокринных генов, является ингибитором гамма-секретазы, выбранным из группы, состоящей из N-[N-(3,5-дифторфенацетил-L-аланил)]-S-фенилглицин t-бутилового эфира (DAPT), RO44929097, DAPT (N-[N-(3,5-дифторфенацетил-L-аланил)]-S-фенилглицин t-бутилового эфира), 1-(S)-эндо-N-(1,3,3)-триметилбицикло[2.2.1]гепт-2-ил)-4-фторфенил сульфонамида, WPE-III31C, S-3-[N'-(3,5-дифторфенил-альфа-гидроксиацетил)-L-аланилил]амино-2,3-дигидро-1-метил-5-фенил-1Н-1,4-бензодиазепин-2-она, (N)-[(S)-2-гидрокси-3-метил-бутирил]-1-(L-аланилил)-(S)-1-амино-3метил-4,5,6,7-тетрагидро-2Н-3-бензазепин-2-она, BMS-708163 (Авагацестата), BMS-708163, Семагацестата (LY450139), Семагацестата (LY450139), MK-0752, MK-0752, YO-01027, YO-01027 (дибензазепина, DBZ), LY-411575, LY-411575, или LY2811376. В одном аспекте высокие уровни активина означают уровни выше 40 нг/мл, 50 нг/мл, и 75 нг/мл. В одном аспекте высокие уровни активина применяют во время стадии 3 или перед продукцией клеток панкреатической энтодермы передней части пищеварительного тракта. В одном аспекте низкие уровни активина означают меньше 30 нг/мл, 20 нг/мл, 10 нг/мл и 5 нг/мл. В одном аспекте низкие уровни активина применяют во время стадии 4 или для получения ПЭК. В одном аспекте ингибируемый или индуцируемый эндокринный ген является NGN3. В другом варианте осуществления активин А и Wnt3A применяют по отдельности или в комбинации для ингибирования эндокринной экспрессии, предпочтительно, для ингибирования NGN3 экспрессии перед продукцией клеток панкреатической энтодермы передней части пищеварительного тракта, или предпочтительно во время стадии 3. В одном аспекте ингибитор гамма-секретазы, предпочтительно RO44929097 или DAPT, применяют в системе культивирования для индукции экспрессии эндокринных генов после продукции ПЭК, или предпочтительно, во время стадий 5, 6 и/или 7.
Культура клеток in vitro, содержащая эндокринные клетки, в которых по меньшей мере 5% указанных человеческих клеток экспрессируют эндокринный маркер, выбранный из группы, состоящей из инсулина (INS), NK6 гомеобокса 1(NKX6.1), панкреатического и дуоденального гомеобокса 1 (PDX1); фактора транскрипции, связанного с локусом 2 (NKX2.2); спаренного бокса 4 (РАХ4), нейрогенной дифференцировки 1 (NEUROD), forkhead box Al (FOXA1), forkhead box A2 (FOXA2), семейства snail «цинковых пальцев» 2 (SNAIL2), и семейства А и В онкогена мышечно-апоневротической фибросаркомы (MAFA и MAFB), и по существу не экспрессируют маркер, выбранный из группы, состоящей из нейрогенина 3 (NGN3), островков 1 (ISL1), гепатоцитарного ядерного фактора 6 (HNF6), GATA-связывающего белка 4 (GATA4), GATA-связывающего белка 6 (GATA6), панкреато-специфического фактора транскрипции 1a (PTF1A) и SRY (полоопределяющего участка Y)-9 (SOX9), где указанные эндокринные клетки являются унипотентными и могут созревать в панкреатические бета-клетки.
Эндокринные клетки предпочтительно являются незрелыми бета-клетками.
Предпочтительно, по меньшей мере 10% указанных человеческих клеток являются незрелыми бета-клетками.
Предпочтительно, по меньшей мере 20% указанных человеческих клеток являются незрелыми бета-клетками.
Предпочтительно, по меньшей мере 50% указанных человеческих клеток являются незрелыми бета-клетками.
Эти клетки предпочтительно являются не-рекомбинантными.
Предпочтительно, чтобы клетки были получены из человеческих плюрипотентных стволовых клеток, выбранных из группы, состоящей из человеческих эмбриональных стволовых клеток (hESC), индуцированных плюрипотентных стволовых клеток (iPSC), партенотных клеток, клеток эмбриональной карциномы (ЕС), мезоэндодермальных клеток, клеток дефинитивной энтодермы, PDX1-1 негативных клеток панкреатической энтодермы передней части пищеварительного тракта, PDX1-позитивных клеток панкреатической энтодермы передней части пищеварительного тракта, и панкреатических эндодермальных клеток (ПЭК).
Обеспечивается способ получения незрелых бета-клеток, включающий этапы: (а) обеспечения контакта популяции PDX1-негативных клеток энтодермы передней части пищеварительного тракта с фактором, активирующим член семейства рецептора TGFβ, с получением популяции PDX1-позитивных клеток панкреатической энтодермы; и (b) обеспечения контакта популяции PDX-позитивных клеток панкреатической энтодермы с ингибитором гамма-секретазы, с получением незрелых бета-клеток, экспрессирующих INS и NKX6.1.
Предпочтительно, чтобы популяция PDX1-негативных клеток энтодермы передней части пищеварительного тракта контактировала с членом семейства Wnt и херегулина.
Член семейства рецептора TGFβ предпочтительно выбран из группы, состоящей из активина А, активина В, Nodal, GDF-8, GDF-10, GDF-11 и GDF15.
Член семейства рецептора TGFβ предпочтительно является активином A, GDF-8 и GDF-11.
Член семейства рецептора TGFβ предпочтительно является активином А.
Член семейства рецептора Wnt предпочтительно выбран из группы, состоящей из WNT1, WNT2, WNT2B, WNT3, WNT3A, WNT4, WNT5A, WNT5B, WNT6, WNT7A, WNT7B, WNT8A, WNT8B, WNT9A, WNT9B, WNT10A, WNT10B, WNT11, WNT16 и WNT3A.
Член семейства рецептора Wnt предпочтительно является Wnt3a.
Член семейства херегулина (HRG) предпочтительно является HRG1, HRG2, HRG3 и HRG4.
Член семейства HRG предпочтительно является HRG1 и HRG2.
Обеспечивается способ получения инсулина in vivo, включающий этапы: (а) обеспечения контакта популяции PDX1-негативных клеток энтодермы передней части пищеварительного тракта с фактором, активирующим член семейства рецептора TGFβ, с получением популяции PDX1-позитивных клеток панкреатической энтодермы; и (b) обеспечения контакта популяции PDX-позитивных клеток панкреатической энтодермы с ингибитором гамма-секретазы, с получением незрелых бета-клеток, экспрессирующих INS и NKX6.1; (с) имплантации незрелых бета-клеток с этапа (b); и (d) созревания клеток в зрелые бета-клетки, секретирующие инсулин in vivo в ответ на стимуляцию глюкозой.
Предпочтительные характеристики и аспекты изобретения являются следующими.
На стадии 7 агенты могут также модулировать экспрессию генов. Предпочтительно, член суперсемейства FGF добавляют на стадии 7. Предпочтительно, FGF2 добавляют на стадии 7. FGF2 может повышать экспрессию SST и/или подавлять экспрессию INS, GCG или GHRL. Предпочтительно, BMP добавляют на стадии 7. BMP может повышать экспрессию INS, PDX1 или IDI.
Популяция незрелых бета-клеток дает начало функции in vivo. Предпочтительно, популяция незрелых бета-клеток является CHGA+. Предпочтительно, популяция незрелых бета-клеток является способной к функционированию in vivo, т.е. при трансплантации секретирует инсулин в ответ на глюкозу в крови. Предпочтительно, популяция эндокринных клеток при трансплантации может развиваться и созревать в функциональные клетки панкреатических островков. Предпочтительно, популяция незрелых бета-клеток обогащена эндокринными клетками (или обеднена неэндокринными клетками). В предпочтительном варианте осуществления более примерно 50% в популяции незрелых бета-клеток являются CHGA+. В более предпочтительном варианте осуществления больше примерно 60%, или 70%, или 80%, или 90%, или 95%, или 98%, или 100% клеток в популяции незрелых бета-клеток являются CHGA+. В предпочтительном варианте осуществления менее примерно 50% клеток в популяции незрелых бета-клеток являются CHGA-. В более предпочтительном варианте осуществления меньше примерно 15% клеток в популяции незрелых бета клеток являются CHGA-. В более предпочтительном варианте осуществления меньше примерно 10%, или 5%, или 3%, или 2%, или 1%, или 0,5%, или 0% клеток в популяции незрелых бета-клеток являются CHGA-. Далее, экспрессия некоторых маркеров может быть супрессирована при продукции незрелых бета-клеток, такая как экспрессия NGN3 во время стадии 3. Популяция незрелых бета-клеток предпочтительно является моногормональной (т.е. только INS). Популяция незрелых бета-клеток предпочтительно одновременно экспрессирует другие клеточные маркеры, включая NKX6.1 и PDX1. Популяция незрелых бета-клеток предпочтительно является моногормональной и одновременно экспрессирует другие клеточные маркеры, включая NKX6.1 и PDX1. Популяция незрелых бета-клеток предпочтительно имеет больше INS клеток, экспрессирующих один гормон, в процентном отношении от всей INS популяции. Популяция незрелых бета-клеток предпочтительно имеет по меньшей мере 50% INS клеток, экспрессирующих один гормон, в процентном отношении от всей INS популяции. Популяция незрелых бета-клеток предпочтительно является CHGA+/INS+/NKX6.1+ (тройной позитивной). В предпочтительном варианте осуществления более примерно 25% клеток в популяции незрелых бета-клеток являются CHGA+/INS+/NKX6.1+ (тройными позитивными). В предпочтительном варианте осуществления более примерно 30%, или 40%, или 50%, или 60%, или 70%, или 80%, или 90%, или 95%, или 100% клеток в популяции незрелых бета-клеток являются CHGA+/INS+/NKX6.1+ (тройными позитивными).
Обогащенная популяция незрелых бета-клеток получена путем диссоциации и реагрегации клеточных агрегатов. Обогащенная популяция незрелых бета-клеток предпочтительно диссоциирована и реагрегирована на стадии 7. Обогащенная популяция незрелых бета-клеток предпочтительно диссоциирована и реагрегирована примерно на 25 день, 26 день, или 27 день, или 28 день, или 29 день, или 30 день, или позже.
Комплексная среда является важной для поддержки эндокринных клеток. Предпочтительно, комплексная среда является CMRL или RPMI. Предпочтительно, комплексную среду применяют на стадии 6 или стадии 7, или и на стадии 6, и на стадии 7. CMRL повышает экспрессию эндокринных маркеров. RPMI повышает экспрессию эндокринных маркеров.
Криоконсервация не снижает популяции незрелых бета-клеток на стадии 7. Криоконсервация не снижает экспрессию CHGA+ на стадии 7.
Клетки на стадии 7 являются NKX6.1-позитивными. Клетки на стадии 7 являются С-пептид-позитивными. Клетки на стадии 7 являются NKX6.1- и С-пептид-позитивными. Клетки на стадии 7 являются моногормональными. Клетки на стадии 7 экспрессируют только INS и GCG.
Диссоциация и реагрегация клеток на стадии 7 не влияет на функцию in vivo.
Криоконсервация клеток на стадии 7 не влияет на функцию in vivo.
Клетки на стадии 7 могут быть криоконсервированы, и сохранять свою функцию in vivo.
Повышение концентрации глюкозы повышает экспрессию INS, GCG или SST. Предпочтительно, концентрация глюкозы увеличивается на стадии 6, или на стадии 7, или на обеих стадиях. Предпочтительно, экспрессия GHRL снижается с повышением концентрации глюкозы.
Незрелые бета-клетки in vitro, которые являются моногормональными (только INS).
Незрелые бета-клетки in vitro, которые совместно экспрессируют NKX6.1 и PDX1.
Незрелые бета-клетки in vitro, которые являются моногормональными (только INS) и совместно экспрессируют NKX6.1 и PDX1.
Незрелые бета-клетки in vitro, которые могут созревать in vivo в клетки, секретирующие инсулин в ответ на глюкозу в крови.
Незрелые бета-клетки in vitro, которые являются CHGA+, INS+ и NKX6.1+.
Предпочтительно, больше 10% от всей клеточной популяции in vitro включают незрелые бета-клетки, которые являются CHGA+, INS+ и NKX6.1+. Незрелые бета-клетки, составляющие больше 50% от всей клеточной популяции in vitro, включают незрелые бета-клетки, которые являются CHGA+, INS+ и NKX6.1+. Незрелые бета-клетки, составляющие больше 80% от всей клеточной популяции in vitro, включают незрелые бета-клетки, которые являются CHGA+, INS+ и NKX6.1+.
Очищенные незрелые бета-клетки. Очищенные незрелые бета-клеточные предшественники. Предпочтительно, незрелые бета-клетки ли предшественники бета-клеток являются очищенными из популяций клеток со стадии 7 с применением сенсора цинка. Предпочтительно, слабая флюоресценция из-за присутствия Ру 1 сенсора, связанного с цинком, усилена для бета-клеток. Предпочтительно, яркая флюоресценция из-за присутствия Ру 1 сенсора, связанного с цинком, усилена для альфа-клеток. Способ очистки бета-клеток с применением сенсора цинка. Способ очистки альфа-клеток с применением сенсора цинка. Очищенные альфа-клетки. Очищенные предшественники альфа-клеток.
Обогащенная клеточная популяция, которая экспрессирует маркеры бета-клеточной линии, такие как INS, IAPP, PDX1, NKX6.1, РАХ4, PCSK1, G6PC2, GCK, или SLC30A8.
Обогащенная клеточная популяция, которая экспрессирует маркеры альфа-клеточной линии, такие как GCG, ARX, или SLC30A8.
Обогащенная клеточная популяция, в которой более 50% клеток являются INS-позитивными. Предпочтительно, чтобы клеточная популяция содержала больше 90% INS-позитивных клеток. В предпочтительных вариантах осуществления 50%-99% клеток являются lNS-позитивными клетками.
Способ получения инсулина in vivo, включающий: (а) получение популяции CHGA+ клеток in vitro; и (b) трансплантацию CHGA+ клеток с продукцией инсулина in vivo.
CHGA+ клетки предпочтительно диссоциируют и реагрегируют на стадии 7.
CHGA+ клетки предпочтительно экспрессируют PDX1 и NKX6.1.
CHGA+ клетки предпочтительно экспрессируют INS и NKX6.1.
CHGA+ клетки предпочтительно культивируют в CMRL или RPMI.
CHGA+ клетки предпочтительно криоконсервируют и оттаивают перед трансплантацией.
CHGA+ клетки предпочтительно культивируют в среде, содержащей высокую концентрацию глюкозы.
Эндокринные клетки in vitro, которые дают начало физиологически функциональной клетке, секретирующей инсулин in vivo.
Незрелые бета-клетки, адекватно специализированные in vitro, которые дают начало физиологически функциональной клетке, секретирующей инсулин in vivo.
Адекватно специализированные незрелые бета-клетки предпочтительно диссоциируют и реагрегируют на стадии 7.
Адекватно специализированные незрелые бета-клетки предпочтительно экспрессируют PDX1 HNKX6.1.
Адекватно специализированные незрелые бета-клетки предпочтительно экспрессируют INS и NKX6.1.
Адекватно специализированные незрелые бета-клетки предпочтительно культивируют в CMRL или RPMI.
Адекватно специализированные незрелые бета-клетки предпочтительно криоконсервируют и оттаивают перед трансплантацией.
Адекватно специализированные незрелые бета-клетки предпочтительно культивируют в среде, содержащей высокую концентрацию глюкозы.
Способ получения обогащенной популяции незрелых бета-клеток in vitro, включающий: получение популяции CHGA+ клеток и сортировку. Способ получения обогащенной клеточной популяции, экспрессирующей INS, IAPP, PDX1, NKX6.1 или РАХ4 in vitro, включающий: получение популяции CHGA+ клеток и сортировку.
Способ получения обогащенной популяции альфа-клеток in vitro, включающий: получение популяции CHGA+ клеток и сортировку.
Способ получения обогащенной популяции клеток, экспрессирующих GCG или ARX in vitro, включающий: получение популяции CHGA+ клеток и сортировку.
Обогащенные популяции альфа- или бета-клеток предпочтительно сортируют с применением Ру 1 сенсора цинка.
В одних вариантах осуществления незрелые клетки экспрессируют INS и NKX6.1, и по существу не экспрессируют NGN3. В других вариантах осуществления незрелые клетки экспрессируют INS, NKX6.1 и MAFB, и по существу не экспрессируют NGN3.
Краткое описание чертежей
Фигура 1 является фотографическим изображением агрегатной суспензионной культуры дедифференцированных перепрограммированных клеток или также упомянутых в настоящем описании как iPS клетки. См. Пример 1.
Фигуры 2A-L являются гистограммами, демонстрирующими относительные уровни экспрессии генов ОСТ4 (Фиг. 2А), BRACHYURY (Фиг. 2В), CER1 (Фиг. 2С), GSC (Фиг. 2D), FOXA2 (Фиг. 2Е), FOXA1 (Фиг. 2F), HNF6 (Фиг. 2G), PDX1 (Фиг. 2Н), PTF1A (Фиг. 2I), NKX6.1 (Фиг. 2J), NGN3 (Фиг. 2K) и INS (Фиг. 2L). Уровни экспрессии нормализованы до средних уровней экспрессии конститутивных генов, экспрессии циклофилина G и ТАТА-связывающего белка (ТВР). Графики изображают кратность положительной регуляции против наименьшей точки данных в наборе данных. См. Пример 1.
Фигуры 3 являются микрофотографиями иммуноцитохимии (ИЦХ) культур человеческих iPS клеток (Е2021 с применением G4 hIPS клеточной линии) с 4 стадии дифференцировки (клетки PDX1-позитивной панкреатической энтодермы) с применением антител, специфичных (панель А) к PDX1; (панель В) к NKX6.1; (панель С) к PTF1A; и (панель D) к Dapi. См. Пример 2.
Фигуры 4 являются изображениями иммуноцитохимии (ИЦХ) культур iPS клеток (Е2021 с применением G4 hIPS клеточной линии) с 5 стадии дифференцировки с применением лигандов, специфичных (панель А) к глюкагону; (панель В) к инсулину; (панель С) к соматостатину; и (панель D) к Dapi. См. Пример 2.
Фигуры 5 являются схемой расположения матрицы и ключа матрицы, обеспеченных в Proteome Profiler™ биочипах человеческих фосфо-RTK антител от R&D Systems. Схема расположения на Фигуре 5А показывает координаты или расположение RTK-антител. Идентичность или наименование RTK-семейства или антител описаны в ключе, Фигура 5В. Позитивные сигналы, наблюдаемые на проявленной пленке, таким образом, можно идентифицировать путем наложения слайда, как на Фигуре 5А, и идентификации сигналов путем ссылки на координаты на накладке (Фигура 5А) с наименованием RTK на Фигуре 5В. См. Пример 4.
Фигуры 6 являются анализом RTK матрицы полученных из iPS клеток панкреатических эндодермальных клеток (ПЭК) в различных условиях (панели А, В, С и D, как описано в Примере 5). Фосфорилирование тирозина определенных RTK наблюдали путем идентификации сигналов от высокой к низкой интенсивности. Идентифицированы или заключены в рамку члены семейств IGF1R/IR и ERBB (EGFR). См. Пример 5.
Фигуры 7А-С являются графиками, демонстрирующими концентрации С-пептида человека и инсулина в сыворотке крови при имплантации мышам для экспериментов Е2314, Е2356 и Е2380 (Фиг. 7А), Е2347 (Фиг. 7В), и Е2354 (Фиг. 7С). Фиг. 7А: Обработка PDX1-экспрессирующих клеток херегулином in vitro улучшает стимулированную глюкозой секрецию инсулина при трансплантации и созревание in vivo. Фиг. 7В: Стимулированные глюкозой уровни С-пептида на 23 неделе после трансплантации, за 3 недели до индукции STZ диабета на мышиной модели. У мышей, которым имплантировали ПЭК, анализировали в указанные моменты времени после трансплантации сывороточные уровни человеческого С-пептида натощак, и спустя 30 минут и 60 минут после интраперитонеального введения глюкозы. На Фиг. 7С ПЭК инкапсулировали с устройством для инкапсулирования клеток (Encaptra® EN20, или EN20, ViaCyte, Сан-Диего, Калифорния), и в некоторых случаях устройства имели микроперфорации (pEN20, ViaCyte, Сан-Диего, Калифорния). Такие устройства описаны в патенте США №8,278,106, описание которого полностью включено в настоящее описание посредством ссылки. См. Пример 7.
Фигуры 8А и 8В являются графиками, демонстрирующими результаты анализа глюкозы крови мышей, леченных STZ, в Эксперименте №2347. Показаны чувствительные к глюкозе инсулин-секретирующие бета-клетки, полученные из имплантированных ПЭК (иПЭК), контролирующие уровень глюкозы на модели STZ-индуцированного диабета. На фигуре 8А показана глюкоза крови для каждой из 13 мышей (базовая линия с херегулином и без него). На фигуре 8В показаны комбинированные средние значения для каждого лечения (базовая линия с херегулином и без него). Результаты анализа произвольных уровней глюкозы в крови не-натощак показаны для 13 мышей, которым имплантировали иПЭК, до 14 суток перед лечением STZ (день 0), и для тех же самых мышей после лечения STZ и после удаления трансплантатов. Животные, леченные STZ, получали STZ примерно 26 недель после трансплантации (день 0). На 28 неделе после трансплантации, примерно 2 недели после начала лечения STZ, иПЭК трансплантаты эксплантировали (удаляли). Образец для анализа глюкозы крови не-натощак собирали на протяжении времени для каждого из животных. См. Пример 7.
Фиг. 9А-Е являются гистограммами, демонстрирующими относительные уровни экспрессии PDX1 (Фиг. 9А), NKX6.1 (Фиг. 9В), PTF1A (Фиг. 9С), NKX2.2 (Фиг. 9D), и NGN3 (Фиг. 9Е). См. Пример 8.
Фиг. 10 является фотографическим изображением агрегатной суспензионной культуры, где PDX1-негативные клетки энтодермы передней части пищеварительного тракта (стадия 2) дифференцировались в PDX1-позитивные клетки энтодермы передней части пищеварительного тракта (стадия 3) с добавлением 50 нг/мл активина А. См. Пример 8.
Фиг. 11 является фотографическим изображением агрегатных суспензионных культур с конце стадии 3 (день 8, верхняя панель) и 4 (день 12, нижняя панель) с применением 5 нг/мл АСТ5) или 10 нг/мл (ACT10) активина А на стадии 3, или 5 нг/мл (АСТ5) активина на стадиях 3 и 4. См. Пример 8.
Фиг. 12 являются фотографическими изображениями агрегатных суспензионных культур на стадии 4 (А-Е, верхняя панель) или в конце стадии 4 (А-Е, нижняя панель). На верхней панели показано 10 нг/мл активина и 2 нг/мл херегулина (ACT 10 HGN 2), или 20 нг/мл активина А и 2 нг/мл херегулина (ACT 20 HGN 2), или 20 нг/мл активина А и 10 нг/мл херегулина (ACT 20 HGN 10) или 10 нг/мл активина А, 2 нг/мл херегулина и 50 нг/мл WNT (ACT 10 HGN 2 WNT 50) при добавлении на стадии 3. На нижней панели показано 25 нг/мл активина A (ACT 25), 50 нг/мл активина A (ACT 50), 75 нг/мл активина A (ACT 75) или 100 нг/мл активина A (ACT 100) при добавлении на стадии 3. Во всех случаях были добавлены низкие уровни активина и херегулина на стадии 4. См. Пример 9.
Фиг. 13А-С являются гистограммами, демонстрирующими относительные уровни экспрессии генов NGN3 (Фиг. 13А), NKX2.2 (Фиг. 13В) и NKX6.1 (Фиг. 13С) при добавлении активина, херегулина и WNT (AHW) на стадии 3, и добавлении активина и херегулина (АН) на стадии 4, как описано в Примере 9.
Фиг. 14А-В являются гистограммами, демонстрирующими относительные уровни экспрессии генов NKX6.1 (Фиг. 14А) и NGN3 (Фиг. 14В) при добавлении активина, херегулина и WNT (AHW) на стадии 3, и добавлении активина и херегулина (АН) на стадии 4, как описано в Примере 10.
Фиг. 15 является графиком, демонстрирующим концентрации человеческого С-пептида в сыворотке мышей, которым имплантировали инкапсулированные ПЭК (контроль) и инкапсулированные модифицированные ПЭК (полученные с применением активина, херегулина и WNT (AHW) на стадии 3, и активина и херегулина (АН) на стадии 4). Уровни экспрессии анализировали спустя 11 недель после трансплантации натощак, и спустя 30 минут и 60 минут после интраперитонеального введения глюкозы. См. Пример 10.
Фиг. 16 является графиком, демонстрирующим концентрации человеческого С-пептида в сыворотке мышей, которым имплантировали инкапсулированные ПЭК (контроль) и инкапсулированные модифицированные ПЭК (полученные с применением активина, херегулина и WNT (AHW) на стадии 3, и активина и херегулина (АН) на стадии 4). Уровни экспрессии анализировали спустя 14 недель после трансплантации натощак, и спустя 30 минут и 60 минут после интраперитонеального введения глюкозы. См. Пример 10.
Фиг. 17 является гистограммой, демонстрирующей относительные уровни экспрессии гена нейрогенина 3 на 13, 15 и 17 день после обработки ингибитором гамма-секретазы на стадии 5, как показано в Примере 11.
Фиг. 18A-D являются гистограммами, демонстрирующими относительные уровни экспрессии генов PDX1 (Фиг. 18А), NKX6.1 (Фиг. 18В), SOX9 (Фиг. 18С) и PTF1A (Фиг. 18D) после дифференцировки в соответствии с Таблицей 17 для стадий 6 и 7, с использованием только FBS, матригеля и ингибитора rho-киназы в дополнение к основной DMEM среде, как описано в Примере 12.
Фиг. 19A-D являются гистограммами, демонстрирующими относительные уровни экспрессии генов INS (Фиг. 19А), GCG (Фиг. 19В), GHRL (Фиг. 19С) и SST (Фиг. 19D) в тех же самых условиях дифференцировки, как на Фиг. 18 и как описано в Примере 12.
Фиг. 20 является графиком, демонстрирующим концентрации человеческого С-пептида в сыворотке мышей с имплантированными инкапсулированными эндокринными клетками, полученными так, как описано в Примере 12 (Е2395, Rag 2, неделя 10 GSIS). Уровни экспрессии анализировали спустя 10 недель после трансплантации натощак и спустя 60 минут после интраперитонеального введения глюкозы. См. Пример 12.
Фиг. 21 является графиком, демонстрирующим концентрации человеческого С-пептида в сыворотке мышей с имплантированными инкапсулированными эндокринными клетками, полученными так, как описано в Примере 12 (Е2395, Rag 2, неделя 15 GSIS). Уровни экспрессии анализировали спустя 15 недель после трансплантации натощак и спустя 60 минут после интраперитонеального введения глюкозы. См. Пример 12.
Фиг. 22A-D являются микрофотографиями, демонстрирующими агрегаты эндокринных клеток, дифференцированные и созревшие in vivo, как описано в Примере 12. Область клеток окрашена DAPI (Фиг. 22А) или антителами к NKX6.1 (Фиг. 22В и Фиг. 22С, окрашено ядро), и хроматину А (Фиг. 22В и Фиг. 22D, цитоплазма). Одна и та же область показана на всех изображениях. См. Пример 12.
Фиг. 23А-С являются микрофотографиями, демонстрирующими агрегаты эндокринных клеток, как описано в Примере 12. На Фиг. 23А показана область клеток, окрашенная на INS (цитоплазматическая) и NKX6.1 (ядерная). На Фиг. 23В показана область клеток, окрашенная на INS (цитоплазматическая) и PDX1 (ядерная). На Фиг. 23С показана та же самая область клеток, окрашенная с DAPI. См. Пример 12.
Фиг. 24A-D являются гистограммами, демонстрирующими данные Nanostring мРНК по относительным уровням экспрессии генов INS (Фиг. 24А), NKX6.1 (Фиг. 24В), PDX1 (Фиг. 24С) и ID1 (Фиг. 24D) в условиях дифференцировки, описанных в Примере 13.
Фиг. 25А-С являются микрофотографиями, демонстрирующими агрегаты эндокринных клеток, как описано в Примере 13. Фиг. 25А является контролем без добавления ТТ или BMP, Фиг. 25В показывает добавление ТТ на стадии 7, Фиг. 25С показывает добавление BMP на стадии 7. Фиг. 25А-С демонстрируют окрашивание на С-пептид (цитоплазматическое) и PDX1 (ядерное).
Фиг. 26А-В являются микрофотографиями, демонстрирующими агрегаты эндокринных клеток, как описано в Примере 13. Фиг. 26А является добавлением ТТ или BMP на стадиях 6 и 7, Фиг. 25В показывает добавление ТТ на стадии 7. Фиг. 25А-В демонстрируют окрашивание на С-пептид (цитоплазматическое) и PDX1 (ядерное).
Фиг. 27А-С являются микрофотографиями, демонстрирующими агрегаты эндокринных клеток, как описано в Примере 13, и изображают те же самые области, как на Фиг. 25. Фиг. 27А является контролем без добавления ТТ или BMP, Фиг. 27В показывает добавление ТТ на стадии 7, Фиг. 27С показывает добавление BMP на стадии 7. Фит. 27А-С демонстрируют окрашивание на С-пептид (цитоплазматическое) и NKX6.1 (ядерное).
Фиг. 28А-В являются микрофотографиями, демонстрирующими агрегаты эндокринных клеток, как описано в Примере 13, и изображают те же самые области, как на Фиг. 26. Фиг. 28А является обработкой ТТ и BMP на стадиях 6 и 7, а Фиг. 28В показывает добавление ТТ и BMP на стадии 7. Фиг. 28А и В демонстрируют окрашивание на С-пептид (цитоплазматическое) и NKX6.1 (ядерное).
Фиг. 29А-Е являются гистограммами, демонстрирующими относительные уровни экспрессии генов эндокринных агрегатов, обработанных с никотинамидом (правый столбец, d21 gi- d13-d15) или без него (левый столбец; d21 gi- d13-d15+NIC d15) на стадии 6, и анализированных на 21 день, как описано в Примере 13.
Фиг. 30 демонстрирует фотографические изображения (дни 21, 22 и 23) суспензионных культур агрегатов эндокринных клеток, диссоциированных и реагрегированных в начале стадии 7, с последующим добавлением FBS по отдельности, FBS (панель А) и 0,05% матригеля (MG; панель В), FBS и 10 мкМ Y-27632 (Y; панель С), или FBS, 0,05% матригеля (MG) и 10 мкМ Y-27632 (Y; панель D), как описано в Примере 14.
Фиг. 31A-D являются гистограммами, демонстрирующими относительные уровни экспрессии генов NKX6.1 (Фиг. 31A), NKX2.2 (Фиг. 31В), PDX1 (Фиг. 31С) и INS (Фиг. 31D), сравнивающими условия дифференцировки с иоггином или без него на 1, 2 и 3 дни. См. пример 15.
Фиг. 32А-С являются гистограммами, демонстрирующими данные Nanostring мРНК по относительным уровням экспрессии генов INS и GCG (Фиг. 32А), GHRL и SST (Фиг. 32В), и PDX1 и ID1 (Фиг. 32С), описывающими эффект экспрессии генов указанными факторами. См. Пример 16.
Фиг. 33А-С являются гистограммами, демонстрирующими данные Nanostring мРНК по относительным уровням экспрессии генов PDX1, NKX6.1, PTF1A, SOX9 (Фиг. 33А); INS, GCG, PPY, GHRL (Фиг. 33В) и PCSK1, GCK, SLC30A8, G6PC2 (Фиг. 33С), описывающие эндокринные (CHGA+) субпопуляции и не-эндокринные (CHGA-) субпопуляции в реагрегированных культурах, по сравнению с не реагрегированными культурами. См. Пример 17.
Фиг. 34 является графиком, демонстрирующим концентрации человеческого С-пептида в сыворотке мышей, которым имплантировали инкапсулированные эндокринные клетки, полученные, как описано в Примере 18, где эндокринные (CHGA+) клеточные типы были обогащены путем диссоциации и реагрегации в начале стадии 7 (образец re-agg), и их сравнивали с клетками, дифференцированными таким же образом, но не реагрегированными (контрольный образец). Уровни экспрессии анализировали спустя 21 неделю после трансплантации натощак, спустя 30 минут, и спустя 60 минут после интраперитонеального введения глюкозы. См. Пример 18.
Фиг. 35А-С являются гистограммами, показывающими данные Nanostring мРНК относительных уровней экспрессии генов CHGA, INS, GCG, GHRL, PPY, SST (Фиг. 35A); GCK, G6CP2, UCN3, PCSK1 (Фиг. 35B); HNF1a, HNF4A, SLC30A8, CADH1 (Фиг. 35C), описывающими эффект сред на основе DMEM и CMRL во время стадии 7. См. Пример 19.
Фиг. 36А-С являются микрофотографиями, показывающими агрегаты эндокринных клеток на стадии 7, как описано в Примере 20. Агрегаты не реагрегировали на стадии 7. Одна и та же область на Фиг. 36А и Фиг. 36В окрашена на INS (цитоплазматический) и GCG (цитоплазматический), соответственно; а на Фиг. 36С окрашена на NKX6.1 (ядерный) и С-пептид (цитоплазматический).
Фиг. 37А-С являются микрофотографиями, демонстрирующими агрегаты эндокринных клеток на стадии 7, как описано в Примере 20. Агрегаты реагрегировали на стадии 7. Одна и та же область на Фиг. 37А и Фиг. 37В окрашена на INS (цитоплазматический) и GCG (цитоплазматический), соответственно; а на Фиг. 37С окрашена на NKX6.1 (ядерный) и С-пептид (цитоплазматический).
Фиг. 38 является графиком, показывающим концентрации человеческого С-пептида в сыворотке мышей, которым имплантировали инкапсулированные эндокринные клетки, полученные, как описано в Примере 20, где клеточные культуры были замороженными (криоконсервированными) и оттаявшими (F/T), и не диссоциированными и реагрегированными (левая сторона), или они были не замороженными (свежими) и диссоциированными и реагрегированными в начале стадии 7 (правая сторона). Уровни экспрессии анализировали спустя 12 недель после трансплантации натощак, спустя 30 минут, и спустя 60 минут после интраперитонеального введения глюкозы. См. Пример 21.
Фиг. 39А-В являются гистограммами, показывающими данные Nanostring мРНК относительных уровней экспрессии генов INS, GCG (Фиг. 39А) и SST, GHRL (Фиг. 39В), описывающими влияние экзогенного высокого уровня глюкозы на стадии 6 на экспрессию эндокринных гормонов. См. Пример 21.
Фиг. 40А-В являются микрофотографией, демонстрирующей агрегаты эндокринных клеток, как описано в Примере 21, с высоким уровнем экзогенной глюкозы во время стадии 7. На Фиг. 40А показано окрашивание на INS (цитоплазматический), а на Фиг. 40В на GCG (цитоплазматический).
Фиг. 41. График проточной цитометрии, показывающий обогащение незрелых эндокринных бета-клеток со стадии 7, вначале обработанных Pyl, цинк-связывающим агентом, и сортированными с применением флюоПЭКценции. См. Пример 24.
Фиг. 42А-С являются гистограммами, показывающими данные Nanostring мРНК относительных уровней экспрессии генов INS, GCG, GHRL, IAPP, SST и SST (Фиг. 42A); PAX4, PDX1, ARX, NKX6.1 (Фиг. 42В); и PCSK1, GCK, G6PC2 и SLC30A8 (Фиг. 42C), характеризующими и идентифицирующими клетки, обогащенные при сортировке с применением цинка. См. Пример 24.
Фиг. 43 является схематической диаграммой стадий 1-4 продукции панкреатических эндодермальных клеток (ПЭК) in vitro и развития и созревания инсулин-секретирующих бета-клеток in vivo. Диаграмма также изображает две основных субпопуляции ПЭК, эндокринных (CHGA+) и не-эндокринных (CHGA-) клеток. Эмбриональные стволовые клетки (ESC), мезоэнтодерма (ME), дефинитивная энтодерма (DE), энтодерма передней части пищеварительного тракта (FG), энтодерма задней части пищеварительного тракта (pFG), и панкреатический эпителий или панкреатическая энтодерма (РЕ).
Фиг. 44 является схематической диаграммой стадий 1-7 получения эндокринных клеток in vitro. Диаграмма также показывает, что высокие уровни активина (стадия 3) с последующими низкими уровнями активина и удалением WNT (стадия 4) подавляют или ингибируют экспрессию NGN3, который позже экспрессируется во время стадии 5 с добавлением ингибитора гамма-секретазы. Эмбриональные стволовые клетки (ESC), мезоэнтодерма (ME), дефинитивная энтодерма (DE), энтодерма передней части пищеварительного тракта (FG), энтодерма задней части пищеварительного тракта (pFG), и панкреатический эпителий или панкреатическая энтодерма (РЕ).
Фиг. 45 является схематической диаграммой стадий 1-7 получения эндокринных клеток in vitro. Диаграмма также показывает стадии, факторы роста, типы клеток и обозначения маркеров каждого типа клеток: эмбриональные стволовые клетки (ESC), мезоэнтодерма (ME), дефинитивная энтодерма (DE), энтодерма передней части пищеварительного тракта (FG), энтодерма задней части пищеварительного тракта (pFG), и панкреатический эпителий или панкреатическая энтодерма (РЕ), пре-бета клетки (pre-В; или незрелые бета-клетки) и бета-клетки (β-клетки).
Подробное описание настоящего изобретения
Настоящее изобретение станет лучше понятным со ссылкой на следующее подробное описание предпочтительных вариантов осуществления изобретения и примеров, включенных него. Однако перед раскрытием и описанием представленных соединений, композиций и способов, необходимо понять, что настоящее изобретение не ограничивается специфическими типами клеток, специфическими слоями фидерных клеток, специфическими условиями, или специфическими способами, и т.д., и как таковое, может варьировать. Многочисленные модификации и варианты станут понятными для специалистов в данной области техники. Необходимо понимать, что терминология, используемая в настоящей заявке, предназначена только для описания специфических вариантов осуществления, и не предназначена для ограничения. Ссылки на все патентные и не-патентные публикации включены посредством ссылки во всей полноте.
Различные клеточные композиции, полученные из плюрипотентных стволовых клеток, описаны в настоящей заявке, и могут быть найдены в поданных автором настоящей заявки заявках на патент США №№: 10/486,408, озаглавленной «Способы культивирования HESC на фидерных клетках, поданной 6 августа 2002; 11/021,618, озаглавленной «Дефинитивная энтодерма», поданной 23 декабря 2004; 11/115,868, озаглавленной «PDX1-экспрессирующая энтодерма», поданной 26 апреля 2005; 11/165,305, озаглавленной «Способы идентификации факторов для дифференцировки дефинитивной энтодермы», поданной 23 июня, 2005; 11/573,662, озаглавленной «Способы повышения дефинитивной эндодермальной дифференцировки плюрипотентных человеческих эмбриональных стволовых клеток с ингибиторами PI-3 киназы», поданной 15 августа 2005; 12/729,084, озаглавленной PDX1-экспрессирующей дорзальной и вентральной энтодермы передней части пищеварительного тракта», поданной 27 октября 2005; 12/093,590, озаглавленной «Маркеры дефинитивной энтодермы», поданной 14 ноября 2005; 11/993,399, озаглавленной «Композиции для культивирования эмбриональных стволовых клеток и способы их применения», поданной 20 июня 2006; 11/588,693, озаглавленной «PDX1-экспрессирующая дорзальная и вентральная энтодерма передней части пищеварительного тракта», поданной 27 октября 2006; 11/681,687, озаглавленной «Эндокринные прогениторы/клетки-предшественники, панкреатические гормон-экспрессирующие клетки и способы их получения», поданной 2 марта 2007; 11/807,223, озаглавленной «Способы культивирования и получения одноклеточных популяций из HESC», поданной 24 мая 2007; 11/773,944, озаглавленной «Способы продукции панкреатических гормонов», поданной 5 июля, 2007; 11/860,494, озаглавленной «Способы увеличения продукции дефинитивной энтодермы», поданной 24 сентября 2007; 12/099,759, озаглавленной «Способы продукции панкреатических гормонов», поданной 8 апреля 2008; 12/107,020, озаглавленной «Способы очистки эндодермальных и панкреатических эндодермальных клеток, полученных из человеческих эмбриональных стволовых клеток», поданной 21 апреля 2008; 12/618,659, озаглавленной «Инкапсулирование панкреатической клеточной линии, полученной из человеческих плюрипотентных стволовых клеток», поданной 13 ноября 2009; 12/765,714 и 13/761,078, озаглавленных «Клеточные композиции, полученные из дедифференцированных перепрограммированных клеток», поданных 22 апреля 2010 и 6 февраля 2013; 11/838,054, озаглавленной «Композиции и способы, пригодные для культивирования дифференцируемых клеток», поданной 13 августа 2007; 12/264,760, озаглавленной «Композиции агрегатных суспензий стволовых клеток, и способы их дифференцировки», поданной 4 ноября 2008; 13/259,15, озаглавленной «Малые молекулы, поддерживающие рост плюрипотентных клеток», поданной 27 апреля 2010; PCT/US11/25628, озаглавленной «Система загрузки для инкапсулирующего устройства», поданной 21 февраля 2011; 13/992,931, озаглавленной «Агенты и способы для ингибирования плюрипотентных стволовых клеток», поданной 28 декабря 2010; и заявках на патент США №№: 29/408,366, поданной 12 декабря 2011; 29/408,368, поданной 12 декабря 2011; 29/423,365, поданной 31 мая 2012; и 29/447,944, поданной 13 марта 2013; и предварительных заявках на патент США №№: 61/774,443, озаглавленной «Полупроницаемые макро-имплантируемые клеточные инкапсулирующие устройства», поданной 7 марта 2013; 61/775,480, озаглавленной «Криоконсервация, гибернация и хранение при комнатной температуре инкапсулированных агрегатов панкреатических эндодермальных клеток», поданной 8 марта 2013; и 61/781,005, озаглавленной «Дифференцировка in vitro плюрипотентных стволовых клеток в панкреатические эндодермальные клетки (ПЭК) и эндокринные клетки», поданной 14 марта 2013.
Определения
Необходимо понимать, что все численные диапазоны, выраженные в настоящей заявке, включают конечные точки, установленные далее, и описывают все целые числа между конечными точками в установленном численном диапазоне.
Если не указано иное, термины, применяемые в настоящей заявке, необходимо понимать в соответствии с их обычным применением рядовым специалистом в соответствующей области техники. Кроме того, для целей настоящего описания и формулы изобретения, если не указано иное, все числа, выражающие количества ингредиентов, проценты или пропорции материалов, условия реакции, и другие численные величины, используемые в данном описании и формуле изобретения, необходимо понимать как модифицированные во всех случаях термином «примерно». Соответственно, если не указано противоположное, численные параметры, установленные в следующем описании и формуле изобретения, являются приблизительными, и могут варьировать, в зависимости от необходимых свойств, ожидаемых при осуществлении настоящего изобретения. В крайнем случае, и не с целью ограничения заявки доктриной эквивалентов в объеме формулы изобретения, каждый численный параметр должен быть истолкован по меньшей мере в свете ряда сообщенных значимых чисел и путем применения обычных методик округления.
При осуществлении вариантов, описанных в настоящей заявке, если не указано иное, применяются обычные методики клеточной биологии, молекулярной биологии, генетики, химии, микробиологии, рекомбинантной ДНК, и иммунологии.
Термин «клетка», использующийся в настоящей заявке, также относится к индивидуальным клеткам, клеточным линиям, или культурам, полученным из таких клеток. «Культура» означает композицию, включающую изолированные клетки того же самого или отличающегося типа. «Культура», «популяция», или «клеточная популяция», использующиеся в настоящей заявке, могут быть и применяются взаимозаменяемо, и их значения ясно определяются контекстом. Например, термин «популяция» может означать клеточную культуру из более чем одной клетки, имеющую одну и ту же идентификационную характеристику, или может означать культуру из более чем одного типа клеток, имеющих различные идентификационные характеристики, например, популяция в одном контексте может быть субпопуляцией в другом контексте. Термин «субпопуляция» означает субкомплекс из клеточной культуры или популяции, при использовании для описания некоторых клеточных типов в пределах клеточной культуры или клеточной популяции.
Использующийся в настоящей заявке термин «тотипотентные стволовые клетки» относится к клеткам, обладающим способностью к дифференцировке во все клетки, составляющие организм, такие как клетки, полученные при слиянии яйцеклетки и сперматозоида. Клетки, полученные путем первых нескольких делений оплодотворенной яйцеклетки, также могут быть тотипотентными. Эти клетки могут дифференцироваться в эмбриональные и экстраэмбриональные типы клеток. Плюрипотентные стволовые клетки, например, такие как ES клетки, могут давать начало любому типу клеток плода или взрослого организма. Однако по отдельности они не могут развиться в плод или взрослое животное, поскольку не обладают потенциалом к развитию экстраэмбриональной ткани. Экстраэмбриональная ткань является, например, полученной из экстраэмбриональной энтодермы, и может дополнительно классифицироваться на париетальную энтодерму (мембрану Рейхерта) и висцеральную энтодерму (образует часть желточного мешка). Как париетальная, так и висцеральная энтодерма поддерживают развитие эмбриона, но сами не образую эмбриональных структур. Также существуют другие экстраэмбриональные ткани, включая экстраэмбриональную мезодерму и экстраэмбриональную эктодерму.
В некоторых вариантах осуществления «плюрипотентную клетку» применяют в качестве исходного материала для дифференцировки в эндодермальную линию, или более конкретно, в клеточные типы панкреатической энтодермы. Использующиеся в настоящей заявке термины «плюрипотентность» или «плюрипотентные клетки» или их эквиваленты означают клетки, которые способны как к пролиферации в клеточной культуре, так и к дифференцировке в множество линиеспецифических клеточных популяций, которые проявляют мультипотентные свойства; например, и плюрипотентные ES клетки, и индуцированные плюрипотентные стволовые (iPS) клетки могут давать начало каждой из трех эмбриональных клеточных линий. Однако плюрипотентные клетки не могут быть способными к образованию всего организма. То есть, плюрипотентные клетки не являются тотипотентными.
В некоторых вариантах осуществления плюрипотентные клетки, применяемые в качестве исходного материала, являются стволовыми клетками, включая hES клетки, hEG клетки, iPS клетки, даже партеногенные клетки и тому подобные. Использующийся в настоящем изобретении термин «эмбриональные» означает ряд стадий развития организма, начиная от единственной зиготы, и заканчивая многоклеточной структурой, которая больше не содержит плюрипотентных или тотипотентных клеток, иных чем развитые половые клетки. В дополнение к эмбриону, полученному при слиянии гамет, термин «эмбриональный» относится к эмбриону, полученному путем переноса ядра соматической клетки. В еще одном варианте осуществления плюрипотентные клетки не получены, или не получены непосредственно из эмбриона, например, iPS клетки получены из не-плюрипотентной клетки, например, мультипотентной клетки или окончательно дифференцированной клетки. Человеческие линии плюрипотентных стволовых клеток, используемые в настоящей заявке, включают hESC и iPSC, например, СуТ49, СуТ25, СуТ203, СуТ212, BG01, BG02, BG03, или любые из тех, которые перечислены в Таблицах 4 и 5, или любые, известные на сегодняшний день или публично доступные или полученные позже.
Человеческие плюрипотентные стволовые клетки также могут быть определены или охарактеризованы по наличию некоторых факторов транскрипции и белков клеточной поверхности, включая факторы транскрипции Oct-4, Nanog и Sox-2, которые формируют коровый регуляторный комплекс, обеспечивающий супрессию генов, приводящих к дифференцировке и поддержанию плюрипотентности; и антигены клеточной поверхности, такие как гликолипиды SSEA3, SSEA4 и кератансульфатные антигены, Tra-1-60 and Tra-1-81, и щелочная фосфатаза.
Использующийся в настоящей заявке термин «индуцированные плюрипотентные стволовые клетки» или «iPS клетки» или «iPSC» означает тип плюрипотентных стволовых клеток, искусственно полученных из не-плюрипотентной клетки, как правило, взрослой соматической клетки, или окончательно дифференцированной клетки, такой как фибробласт, гемопоэтическая клетка, миоцит, нейрон, эпидермальная клетка, или тому подобной, путем вставки некоторых генов или продуктов генов, обозначаемых как факторы перепрограммирования. См. Takahashi и др., Cell 131:861-872 (2007); Wernig и др., Nature 448:318-324 (2007); Park и др., Nature 451:141-146 (2008). Эти и последующие известные способы получения iPS хорошо известны, и то, как выделены или получены iPS, не ограничивается настоящим изобретением. Индуцированные плюрипотентные стволовые клетки по существу подобны натуральным человеческим плюрипотентным стволовым клеткам, таким как hES клетки, во многих отношениях, включая экспрессию некоторых генов и белков стволовых клеток, характер метилирования хроматина, время удвоения, формирование эмбриоидных телец, формирование тератомы, формирование жизнеспособной химеры, и потенцию и способность к дифференцировке. Человеческие iPS клетки обеспечивают источник плюрипотентных стволовых клеток без связанного применения эмбрионов.
Использующийся в настоящей заявке термин «перепрограммирование», «перепрограммированная» или их эквиваленты относятся к процессу, который обеспечивает для клетки измеримо повышенную способность к образованию потомства по меньшей мере одного нового клеточного типа, в культуре или in vivo, по сравнению с теми же самыми условиями без перепрограммирования. В некоторых вариантах осуществления, описанных в настоящей заявке, соматические клетки «перепрограммированы» в плюрипотентные клетки. В некоторых аспектах соматические клетки перепрограммированы так, чтобы после достаточной пролиферации измеряемая часть клеток in vivo или в клеточной культуре in vitro проявляла фенотипические характеристики нового плюрипотентного типа клеток. Без перепрограммирования такие соматические клетки не дают начало потомству, проявляющему фенотипические характеристики нового плюрипотентного типа клеток. Если даже без перепрограммирования соматические клетки могут давать начало потомству, проявляющему фенотипические характеристики нового плюрипотентного типа клеток, доля потомства этих соматических клеток, проявляющего фенотипические характеристики нового плюрипотентного типа клеток, измеримо увеличивается по сравнению с тем, что было до перепрограммирования.
Использующееся в настоящей заявке выражение «перепрограммирование дифференцировки» относится к процессу, изменяющему клетку до образования потомства по меньшей мере одного нового типа клеток с новым статусом дифференцировки, либо в культуре, либо in vivo, по сравнению с теми же самыми условиями без перепрограммирования дифференцировки. Этот процесс включает дифференцировку, дедифференцировку и трансдифференцировку. Следовательно, использующееся в настоящей заявке выражение «дифференцировка» означает процесс, при котором менее специализированные клетки становятся более специализированным типом клеток. Напротив, выражение «дедифференцировка» означает клеточный процесс, при котором частично или окончательно дифференцированные клетки обращаются на раннюю стадию развития, такую как клетки, обладающие плюрипотентностью или мультипотентностью. Далее, выражение «трансдифференцировка» означает процесс трансформации одного дифференцированного клеточного типа в другой дифференцированный клеточный тип.
Использующийся в настоящей заявке термин «мультипотентность» или «мультипотентная клетка» или их эквиваленты означают клеточный тип, который может давать начало ограниченному количеству других конкретных типов клеток. То есть, мультипотентные клетки являются коммитированными для одного или нескольких эмбриональных клеточных зачатков, и таким образом, в отличие от плюрипотентных клеток, не могут давать начало каждой из трех эмбриональных клеточных линий, а также экстраэмбриональных клеток. Мультипотентные соматические клетки являются более дифференцированными, по сравнению с плюрипотентными клетками, но не являются окончательно дифференцированными. Плюрипотентные клетки, таким образом, обладают более высокой потенцией, чем мультипотентные клетки. Факторы, определяющие потенцию, которые могут перепрограммировать соматические клетки или использоваться для генерации iPS клеток, включают такие факторы, как Oct-4, Sox2, FoxD3, UTF1, Stella, Rex1, ZNF206, Sox15, Myb12, Lin28, Nanog, DPPA2, ESG1, Otx2 или их комбинации, но не ограничиваются ими.
Использующийся в настоящей заявке термин «унипотентная», «унипотентность» или «унипотентная клетка», или их эквиваленты означают тип клеток, который может давать начало только одной другой клеточной линии. Например, незрелые бета-клетки, как описано в настоящей заявке, обладают способностью к дифференцировке только в инсулиновые бета-клетки, и не обладают потенциалом, например, к дифференцировке в глюкагоновые (альфа) клетки, соматостатиновые (дельта) клетки и секретирующие панкреатический полипептид (гамма) клетки. Напротив, эндокринные клетки-предшественники, PDX1-позитивные панкреатические эндодермальные клетки, клетки дефинитивной энтодермы, или клетки мезоэнтодеромы, и hES клетки все являются мультипотентными или плюрипотентными (hESC), которые могут давать начало каждой из панкреатических альфа-, бета-, дельта- и гамма-островковых клеток. Подобным образом, панкреатические бета-, дельта- и гамма- островковые клетки являются клеточными линиями из эндокринных клеток-предшественников, PDX1-позитивных панкреатических эндодермальных клеток, клеток дефинитивной энтодермы, или клеток мезоэнтодермы, и hES клеток.
Использующийся в настоящей заявке термин «моногормональные» и «полигормональные» клетки означают клетки, экспрессирующие только один гормон (например, незрелые бета-клетки и бета-клетки экспрессируют только белок инсулин, а не белки глюкагон или соматостатин), или экспрессируют более чем один или множество гормонов (например, эндокринные предшественники или прогениторные клетки содержат субпопуляции клеток, экспрессирующие 2, 3 или 4 или больше гормонов в одной и той же клетке). Использующийся в настоящей заявке термин «агент, активирующий тирозинкиназу ERBB рецептора» включает по меньшей мере 16 различных лигандов семейства EGF, связывающихся с ERBB рецепторами: EGF (эпидермальный фактор роста), AG или AREG (амфирегулин), и TGF-α (трансформирующий фактор роста-альфа), Btc (бетацеллюлин), HBEGF (гепарин-связывающий EGF), и Ereg (эпирегулин), нейрегулины (или херегулины), такие как нейрегулин-1, -2, -3 и -4 (или херегулин-1, -2, -3 и -4), но не ограничивается ими. Однако настоящее изобретение подразумевает любой лиганд, способный к связыванию с любым одним из четырех ERBB рецепторов или их комбинацией для индукции образования гомо- и гетеродимерных рецепторных комплексов, приводя к активации мембранного киназного домена и последующему фосфорилированию. См. также Таблицу 13.
Термин «клеточная линия» означает все стадии развития клеточного типа, от самой ранней клетки-предшественника до полностью созревшей клетки (т.е. специализированной клетки). Например, «дефинитивная эндодермальная клеточная линия» или «PDX1-негативная эндодермальная клеточная линия» или «PDX1-позитивная панкреатическая эндодермальная клеточная линия», «линия эндокринных клеток-предшественников», или «эндокринная клеточная линия», или «линия незрелых бета-клеток» и тому подобное означает клетки, полученные или дифференцированные из клеток дефинитивной энтодермы, клеток PDX1-негативной энтодермы, клеток PDX1-позитивной панкреатической энтодермы, и тому подобное. Клетка дефинитивной энтодермы является линией мезонтодермальных клеток, одной из ее предшественников. Клетка PDX1-позитивной панкреатической энтодермы является линией клетки дефинитивной энтодермы, одной из ее предшественников. Эндокринный предшественник является линией PDX1-позитивной панкреатической клетки, клетки дефинитивной энтодермы и клетки мезоэнтодермы, все из которых являются ее предшественниками. Незрелая бета-клетка является линией эндокринной клетки-предшественника, PDX1-позитивной панкреатической клетки, клетки дефинитивной энтодермы и клетки мезоэнтодермы, все из которых являются ее предшественниками. Бета-клетка является единственной линией, например, незрелой бета-клетки. Далее, все эндодермальные клеточные линии, описанные в настоящей заявке, являются hES клеточными линиями.
«Клетка-предшественник» или «прогениторная клетка», использующаяся в настоящей заявке, может быть любой клеткой на пути клеточной дифференцировки, способной к дифференцировке в более зрелую клетку. Как таковая, клетка-предшественник может быть плюрипотентной клеткой, или она может быть частично дифференцированной мультипотентной клеткой, или обратимо дифференцированной клеткой. Термин «популяция клеток-предшественников» означает группу клеток, способных к развитию в более зрелый или дифференцированный клеточный тип. Популяция клеток-предшественников может содержать клетки, являющиеся плюрипотентными, клетки, которые являются линиеспецифическими стволовым клетками (т.е. клетками, способными развиваться в клетки не всех эктодермальных линий, или например, клетками только нейрональной линии), и клетки, которые являются обратимо линиеспецифическими стволовыми клетками. Таким образом, термин «прогениторная клетка» или «клетка-предшественник» может означать «плюрипотентную клетку» или «мультипотентную клетку».
«Эндокринный прогенитор/клетка-предшественник», использующийся в настоящей заявке, означает мультипотентную клетку из дефинитивной эндодермальной линии, экспрессирующей по меньшей мере один маркер, выбранный из перечня, состоящего из нейрогенина 3 (NEUROG3), PDX1, PTF1A, SOX9, NKX6.1, HNF1b, GATA4, HNF6, FOXA1, FOXA2, GATA6, MYT1, ISLET1, NEUROD, SNAIL2, MNX1, IA1, RFX6, PAX4, РАХ6, NKX2.2, MAFA и MAFB, которая может далее дифференцироваться в клетки эндокринной системы, включая гормон-экспрессирующие клетки панкреатических островков, но не ограничиваясь ими. Эндокринные прогениторы/клетки-предшественники не могут дифференцироваться во многие другие типы клеток, тканей и/или органов, по сравнению с менее специфически дифференцированными клетками дефинитивной эндодермальной линии, такими как PDX1-позитивные панкреатические эндодермальные клетки или дефинитивные эндодермальные клетки, или мезоэндодермальные клетки. Эндокринные прогениторы/клетки-предшественники описаны подробно по меньшей мере в патенте США №8,129,182 от автора настоящей заявки.
Использующиеся в настоящей заявке термины «развиваться из плюрипотентных клеток», «дифференцироваться из плюрипотентных клеток», «созревать из плюрипотентных клеток», или «продуцированный из плюрипотентных клеток», «полученный из плюрипотентных клеток», «дифференцированный из плюрипотентных клеток» и эквивалентные выражения относятся к продукции дифференцированного типа клеток из плюрипотентных клеток in vitro или in vivo, например, в случае эндокринных клеток, созревающих из трансплантированных PDX1 панкреатических эндодермальных клеток in vivo, как описано в Международной патентной заявке №PCT/US2007/015536, озаглавленной «Способы получения панкреатических гормонов», описание которой полностью включено в настоящее описание посредством ссылки. Все такие термины относятся к развитию клетки от стадии с потенциалом дифференцировки по меньшей мере в две различных клеточных линии до специализированной и окончательно дифференцированной клетки. Такие термины могут применяться взаимозаменяемо для целей настоящей заявки. Варианты осуществления, описанные в настоящей заявке, подразумевают условия культивирования, позволяющие такой дифференцировке быть обратимой, так чтобы плюрипотентность или по меньшей мере способность к дифференцировке в более чем одну клеточную линию могла быть избирательно восстановлена.
Термин «фидерная клетка» означает культуру клеток, выращенную in vitro и секретирующую по меньшей мере один фактор в культуральную среду, и которую можно применять для поддержки роста другой необходимой клетки в культуре. Использующийся в настоящей заявке термин «слой фидерных клеток» можно применять взаимозаменяемо с термином «фидерная клетка». Фидерная клетка может включать монослой, где фидерные клетки покрывают поверхность культуральной чашки полным слоем перед ростом сверху других клеток, или может включать кластеры клеток. В предпочтительном варианте осуществления фидерная клетка включает адгезивный монослой.
Подобным образом, варианты осуществления, в которых плюрипотентные клеточные культуры или агрегатные плюрипотентные суспензионные культуры растут в определенных условиях без применения фидерных клеток, являются «бесфидерными». Бесфидерные способы культивирования повышают масштабируемость и воспроизводимость плюрипотентной клеточной культуры и снижают риск контаминации, например, инфекционными агентами из фидерных клеток или других компонентов культур животного происхождения. Бесфидерные способы также описаны в патенте США №6,800,480, Bodnar и др. (принадлежит Geron Corporation, Менло-Парк, Калифорния). Однако, и в отличие от патента США №6,800,480, варианты осуществления, описанные в настоящей заявке, являющиеся плюрипотентными, мультипотентными или дифференцированными клеточными культурами, являются бесфидерными и не содержат дополнительно эндогенного или экзогенного экстрацеллюлярного матрикса, т.е. культуры, описанные в настоящей заявке, свободны от экстрацеллюлярного матрикса, а также являются бесфидерными. Например, в патенте США №6,800,480 экстрацеллюлярный матрикс готовят путем культивирования фибробластов, лизиса фибробластов in situ, с последующим отмыванием того, что осталось после лизиса. Альтернативно, в патенте США №6,800,480 экстрацеллюлярный матрикс может также быть приготовлен из изолированного компонента матрикса или комбинации компонентов, выбранных из коллагена, плацентарного матрикса, фибронектина, ламинина, мерозина, тенасцина, гепаринсульфата, хондроитинсульфата, дерматансульфата, аггрекана, бигликана, тромбоспондина, витронектина и декорина. Варианты осуществления, описанные в настоящей заявке, не продуцируют экстрацеллюлярный матрикс путем роста фидерного или фибробластного слоя и лизиса клеток для получения экстрацеллюлярного матрикса; и не требуют первоначального покрывания сосуда для культивирования ткани компонентом экстрацеллюлярного матрикса или комбинацией компонентов экстрацеллюлярного матрикса, выбранных из коллагена, плацентарного матрикса, фибронектина, ламинина, мерозина, тенасцина, гепаринсульфата, хондроитинсульфата, дерматансульфата, аггрекана, бигликана, тромбоспондина, витронектина и декорина. Следовательно, агрегатные суспензионные культуры, описанные в настоящей заявке для плюрипотентных, мультипотентных и дифференцированных клеток, не требуют фидерного слоя, лизированных фидерных или фибробластных клеток для получения покрытия из экстрацеллюлярного матрикса, экзогенно добавленного экстрацеллюлярного матрикса или компонента матрикса; скорее применение растворимого компонента человеческой сыворотки, как описано в Международной заявке PCT/US2008/080516, озаглавленной «Способы и композиции для бесфидерной среды для плюрипотентных стволовых клеток, содержащей человеческую сыворотку», включенной посредством ссылки во всей полноте, преодолевает потребность в фидерных клетках или фидерном монослое, а также преодолевает потребность в эндогенном экстрацеллюлярном матриксе из фидерной или фибробластной клетки или из экзогенно добавленных компонентов экстрацеллюлярного матрикса.
Использующиеся в настоящей заявке термины «кластер» и «скопление» или «агрегат» могут применяться взаимозаменяемо, и обычно означают группу клеток, которая не диссоциирована на отдельные клетки, а затем агрегирована до формирования кластеров, или имеет тесные контакты между клетками. Термин «реагрегирована», использующийся в настоящей заявке, означает, что кластеры, скопления и/или агрегаты диссоциируют на более мелкие кластеры, скопления и/или агрегаты из отдельных клеток, а затем формируют новые межклеточные контакты путем повторной агрегации с образованием кластеров, скоплений и/или агрегатов. Эта диссоциация обычно по природе осуществляется вручную (например, с применением пастеровской пипетки), но подразумеваются и другие средства диссоциации. Суспензия агрегатов плюрипотентных или мультипотентных клеточных культур является по существу такой, как описано в Международных публикациях PCT/US2007/062755, озаглавленной «Композиции и способы культивирования дифференцированных клеток» и PCT/US2008/082356, озаглавленной «Суспензионные композиции агрегатов стволовых клеток и способы их дифференцировки».
Термин «суспензия из отдельных клеток» или ее эквиваленты означает суспензию плюрипотентных, мультипотентных или окончательно дифференцированных отдельных клеток, или суспензию отдельных клеток, полученных их плюрипотентной или мультипотентных клеток, любыми механическими или химическими средствами. Существует несколько способов диссоциации клеточных кластеров до получения суспензий отдельных клеток из первичных тканей, прикрепленных клеток в культуре, и агрегатов, например, физические усилия (механическая диссоциация, такая как с помощью скребка для клеток, измельчение с помощью пипетки с узким поперечным сечением, тонкоигольная аспирация, дизагрегация с помощью вортекса и принудительная фильтрация через мелкую сетку из нейлона или нержавеющей стали), ферменты (ферментативная диссоциация таким ферментами, как трипсин, коллагеназа, Accutase™ и тому подобное), или комбинация того и другого. Далее, способы и условия культуральных сред, пригодных для поддержки диссоциации на отдельные клетки плюрипотентных, мультипотентных или дифференцированных клеток, применяются для экспансии, сортировки клеток, и установленного высева для анализа на многолуночных планшетах и обеспечения автоматизации процедур культивирования и клональной экспансии.
В предпочтительных вариантах осуществления способы культивирования не включают продуктов животного происхождения. В другом предпочтительном варианте осуществления способы культивирования не содержат ксеногенных продуктов. В еще более предпочтительных вариантах осуществления одно или несколько условий или требований для коммерческого производства лечебных средств на основе клеток человека удовлетворяются или соответствуют лучшим показателям с помощью способов культивирования, описанных в настоящей заявке.
Популяция плюрипотентных клеток может быть далее культивирована в присутствии некоторых дополнительных факторов роста для получения популяции клеток, которые являются или будут развиваться в различные клеточные линии, или могут быть избирательно обратимыми, чтобы быть способными к развитию в различные клеточные линии. Термин «дополнительный фактор роста» используется в самом широком контексте, и означает вещество, эффективное для активации роста плюрипотентной клетки, поддержки выживания клетки, стимуляции дифференцировки клетки и/или стимуляции обращения дифференцировки клетки. Далее, дополнительный фактор роста может быть веществом, секретируемым фидерной клеткой в среду. Такие вещества включают цитокины, хемокины, малые молекулы, нейтрализующие антитела, и белки, но не ограничиваются ими. Факторы роста могут также включать межклеточные сигнальные полипептиды, которые контролируют развитие и поддержку клетки, а также форму и функции тканей. В предпочтительных вариантах осуществления дополнительный фактор роста выбран из группы, состоящей из фактора стволовых клеток (SCF), онкостатина М (OSM), цилиарного нейротрофического фактора (CNTF), интерлейкина-6 (IL-6) в комбинации с растворимым рецептором интерлейкина-6 (IL-6R), фактора роста фибробластов (FGF), костного морфогенетического белка (BMP), фактора некроза опухоли (TNF), и гранулоцитарно-макрофагального колониестимулирующего фактора (GM-CSF).
В некоторых способах продукции клеток, как описано в настоящей заявке, факторы роста удаляют из клеточной культуры или популяции клеток после их добавления. Например, фактор роста, такой как активин А, активин В, GDF-8 или GDF-11, можно добавить и удалить в пределах примерно одного дня, примерно двух дней, примерно трех дней, примерно четырех дней, примерно пяти дней, примерно шести дней, примерно семи дней, примерно восьми дней, примерно девяти дней или примерно десяти дней после их добавления. В некоторых вариантах осуществления факторы дифференцировки не удаляют из клеточной культуры как таковые, но их устранение из клеточной среды означает удаление, например, замену среды для культивирования клеток, содержащей активин, на среду, не содержащую активин.
Поскольку эффективность процесса дифференцировки можно регулировать путем модификации определенных параметров, которые включают условия роста клеток, концентрации факторов роста и продолжительность этапов культивирования, но не ограничиваются ими, процедуры дифференцировки, описанные в настоящей заявке, могут приводить примерно к 5%, примерно к 10%, примерно к 15%, примерно к 20%, примерно к 25%, примерно к 30%, примерно к 35%, примерно к 40%, примерно к 45%, примерно к 50%, примерно к 55%, примерно к 60%, примерно к 65%, примерно к 70%, примерно к 75%, примерно к 80%, примерно к 85%, примерно к 90%, примерно к 95%, или к более чем примерно 95% превращению плюрипотентных клеток, которые включают индуцированные плюрипотентные клетки, в мультипотентные или дифференцированные клетки, например, окончательной энтодермы, энтодермы передней части пищеварительного тракта, PDX1-позитивной энтодермы передней части пищеварительного тракта, PDX1-позитивной панкреатической энтодермы или PDX1/NKX6.1 со-позитивной панкреатической энтодермы, эндокринного предшественника или NGN3/NKX2.2 со-позитивного эндокринного предшественника, и гормон-секретирующим эндокринным клеткам или INS, GCG, GHRL, SST, PP единично-позитивные эндокринные клетки. В способах, в которых применяют выделение клеток препервичной полоски или мезоэндодермальных клеток, можно выделить по существу чистую популяцию клеток препервичной полоски или мезоэнтодермы.
Общие способы получения клеток эндодермальной линии из hES клеток описаны в родственных заявках США, как указано выше, и D'Amour и др. 2005 Nat Biotechnol. 23:1534-41, опубликованной он-лайн 28 октября 2005; D'Amour и др. 2006 Nat Biotechnol. 24(11):1392-401, опубликованной он-лайн 19 октября 2006; Kroon и др. (2008) Nat Biotechnol. 26 (4):443-452, опубликованной он-лайн 20 февраля 2008; Kelly и др. (2011) Nat. Biotechnol. 29(8):750-6, опубликованной он-лайн 31 июля 2011; и Schulz и др. (2012) PLosOne, 7(5): е37004, опубликованной он-лайн 18 мая 2012. D'Amour и др. описывают 5-этапный протокол дифференцировки: этап 1 (приводит в основном к продукции дефинитивной энтодермы); этап 2 (приводит в основном к продукции PDX1-негативной энтодермы передней части пищеварительного тракта); этап 3 (приводит в основном к продукции PDX1-позитивной энтодермы передней части пищеварительного тракта); этап 4 (приводит в основном к продукции панкреатической энтодермы или панкреатического эндокринного предшественника) и этап 5 (приводит в основном к продукции гормон-экспрессирующих эндокринных клеток).
Термин «трофоэктодерма» относится к мультипотентным клеткам с относительно высокой экспрессией маркеров, выбранных из группы, состоящей из HAND1, Eomes, MASH2, ESXL1, HCG, KRT18, PSG3, SFXN5, DLX3, PSX1, ETS2, и ERRB генов, по сравнению с уровнями экспрессии HAND1, Eomes, MASH2, ESXL1, HCG, KRT18, PSG3, SFXN5, DLX3, PSX1, ETS2, и ERRB в не-трофоэктодермальных клетках или клеточных популяциях.
«Экстраэмбриональная энтодерма» означает мультипотентную клетку с относительно высокими уровнями экспрессии маркеров, выбранных из группы, состоящей из SOX7, SOX17, THBD, SPARC, DAB1 или AFP генов, по сравнению с уровнями экспрессии SOX7, SOX17, THBD, SPARC, DAB1 или AFP в не-экстраэмбриональных эндодермальных клетках или клеточных популяциях.
Термин «клетки препервичной полоски» относится к мультипотентным клеткам с относительно высокими уровнями экспрессии генов маркеров FGF8 и/или NODAL, по сравнению с клетками с низкой экспрессией BRACHURY, FGF4, SNAI1, SOX17, FOXA2, SOX7 и SOX1.
Термин «мезоэндодермальная клетка» относится к мультипотентной клетке с относительно высокими уровнями экспрессии генов маркеров BRACHURY, FGF4, SNAI1 MLXL1 и/или WNT3, по сравнению с клетками с низкой экспрессией SOX17, CXCR4, FOXA2, SOX7 и SOX1.
Термин «дефинитивная энтодерма (DE)» означает мультипотентную эндодермальную клеточную линию, которая может дифференцироваться в клетки кишечной трубки или органы, полученные из кишечной трубки. В соответствии с некоторыми вариантами осуществления, клетки дефинитивной энтодермы являются клетками млекопитающих, а в предпочтительном варианте осуществления клетки дефинитивной энтодермы являются человеческими клетками. В некоторых вариантах осуществления настоящего изобретения клетки дефинитивной энтодермы экспрессируют или незначительно экспрессируют некоторые маркеры. В некоторых вариантах осуществления один или несколько маркеров, выбранных из SOX17, CXCR4, MIXL1, GATA4, HNF3β, GSC, FGF17, VWF, CALCR, FOXQ1, CMKOR1 и CRIP1, экспрессируются в клетках дефинитивной энтодермы. В других вариантах осуществления один или несколько маркеров, выбранных из ОСТ4, альфа-фетопротеина (AFP), тромбомодулина (ТМ), SPARC, SOX7 и HNF4-альфа, не экспрессируются или незначительно экспрессируются клетками дефинитивной энтодермы. Популяции клеток дефинитивной энтодермы и способы их получения также описаны в заявке США №11/021,618, озаглавленной «Дефинитивная энтодерма», поданной 23 декабря 2004.
Термин «PDX1-негативные эндодермальные клетки передней части пищеварительного тракта» или «эндодермальные клетки передней части пищеварительного тракта», или их эквиваленты означают клетки, экспрессирующие SOX17, HNF1β (HNF1B), HNF4-альфа (HNF4A) и FOXA1 маркеры, но по существу не экспрессирующие PDX1, AFP, SOX7, или SOX1. PDX1-негативные популяции эндодермальных клеток передней части пищеварительного тракта и способы их получения также описаны в заявке США №11/588,693, озаглавленной «PDX1-экспрессирующая дорзальная и вентральная энтодерма передней части пищеварительного тракта», поданной 27 октября 2006.
Термин «PDX1-позитивные дорсальные эндодермальные клетки передней части пищеварительного тракта» (дорсальные PDX1-позитивные эндодермальные клетки передней части пищеварительного тракта» или просто «PDX1-позитивная энтодерма», или их эквиваленты означают означает клетки, экспрессирующие один или несколько маркеров, выбранных из Таблицы 1 и/или один или несколько маркеров, выбранных из Таблицы 2, также описанных в родственной заявке США №11/588,693, озаглавленной «PDX1-экспрессирующая дорзальная и вентральная энтодерма передней части пищеварительного тракта», поданной 27 октября 2006, а также заявке США №11/115,868, озаглавленной «PDX1-экспрессирующая энтодерма», поданной 26 апреля 2005.


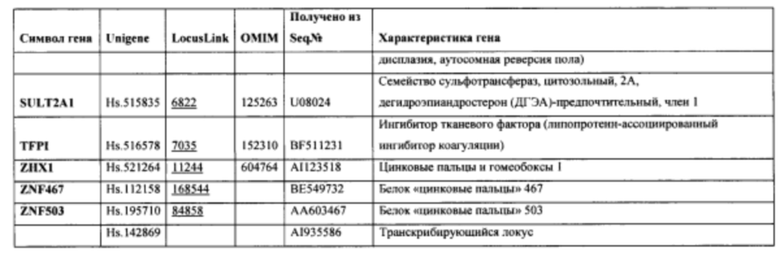

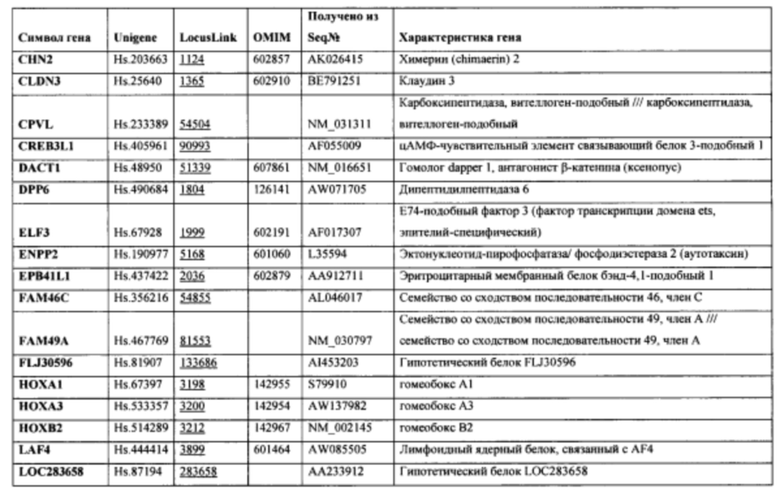


Термины «панкреатическая энтодерма», «панкреатический эпителиальный», «панкреатический эпителий» (сокращенно «ПЭ»), «панкреатический прогенитор», «PDX-1 позитивная панкреатическая энтодерма» или их эквиваленты, такие как панкреатические эндодермальные клетки («ПЭК»), все являются предшественниками или прогениторными панкреатическими клетками. ПЭК, как описано в настоящей заявке, является популяцией прогениторных клеток после 4 стадии дифференцировки (примерно 12-14 день) и включает по меньшей мере две основных различных популяции: (i) панкреатические прогениторные клетки, которые экспрессируют PDX1 и NKX6.1, но не экспрессируют CHGA (или CHGA-негативные, CHGA-); или «не-эндокринные мультипотентные прогениторные субпопуляции (CHGA-)», или «не-эндокринные (CHGA-) субпопуляции», или «не-эндокринные (CHGA-) клетки», или их эквиваленты, и (ii) полигормональные эндокринные клетки, которые экспрессируют CHGA (CHGA-позитивные, CHGA+), или «эндокринные мультипотентные прогениторные субпопуляции (CHGA+)», или «эндокринные (CHGA+) субпопуляции», или «эндокринные (CHGA+) клетки», или их эквиваленты. ПЭК панкреатические прогениторные субпопуляции, которые экспрессируют PDX1 и NKX6.1, но не CHGA, также обозначаются как «не-эндокринная мультипотентная панкреатическая прогениторная субпопуляции (CHGA-)» или «неэндокринная прогениторная субпопуляция», «не-эндокринная (CHGA-) субпопуляция», «мультипотентная прогениторная субпопуляция», и тому подобное. ПЭК полигормональная эндокринная клеточная субпопуляция, которая экспрессирует CHGA, также обозначается как «клетки, коммитированные в эндокринную клеточную линию (CHGA+)», или «эндокринные клетки», или «CHGA+ клетки», и тому подобное. Не углубляясь в теорию, полагают, что клеточная популяция, которая экспрессирует NKX6.1, но не CHGA, является более активным или терапевтическим компонентом ПЭК, в то время как популяция CHGA-позитивных полигормональных эндокринных клеток далее дифференцирует и созревает in vivo в глюкагон-экспрессирующие островковые клетки. См. Kelly и др. (2011) «Cell-surface markers for the isolation of pancreatic cell types derived from human embryonic stem cells», Nat Biotechnol. 29(8):750-756 («Маркеры клеточной поверхности для выделения типов панкреатических клеток, полученных из эмбриональных стволовых клеток человека»), опубликовано он-лайн 31 июля 2011, и Schulz и др. (2012), «А Scalable System for Production of Functional Pancreatic Progenitors from Human Embryonic Stem Cells», PLosOne 7(5): 1-17, e37004 («Масштабируемые системы для продукции функциональных панкреатических прогениторов из эмбриональных стволовых клеток человека»).
До сих пор, иногда панкреатические эндодермальные клетки применяют без ссылки на ПЭК, как описано выше, но обозначают в целом как типы клеток по меньшей мере на 3 и 4 стадии дифференцировки. Применение и значение станет ясным из контекста. Панкреатическая энтодерма, полученная из плюрипотентных стволовых клеток, и по меньшей мере hES и hIPS клеток, таким образом, отличается от других типов эндодермальных клеточных линий, на основе отличающихся или высоких уровней экспрессии маркеров, выбранных из PDX1, NKX6.1, PTF1A, СРА1, cMYC, NGN3, РАХ4, ARX и NKX2.2 маркеров, но по существу не экспрессирующих гены, являющиеся признаком панкреатических эндокринных клеток, например, CHGA, INS, GCG, GHRL, SST, MAFA, PCSK1 и GLUT1. Кроме того, некоторые «эндокринные прогениторные клетки», экспрессирующие NGN3, могут дифференцироваться в другие непанкреатические структуры (например, двенадцатиперстную кишку). Популяции панкреатических эндодермальных или эндокринных прогениторных клеток и способы для них также описаны, например, а патентной заявке США №11/773,944, озаглавленной «Способы продукции панкреатических гормонов», поданной 5 июля 2007, и патентной заявке США №12/107,020, озаглавленной «Способы очистки энтодермы и панкреатических эндодермальных клеток, полученных из эмбриональных стволовых клеток человека», поданной 21 апреля 2008, описания которых полностью включены в настоящее описание посредством ссылки.
Термин «эндокринная клетка» или «панкреатическая островковая гормон-экспрессирующая клетка», «панкреатическая эндокринная клетка», «панкреатическая островковая клетка», «панкреатические островки» или их эквиваленты означают клетку, которая может быть полигормональной или моногормональной. Таким образом, клетки могут экспрессировать один или несколько панкреатических гормонов, которые имеют по меньшей мере некоторые из функций человеческой клетки панкреатического островка. Гормон-экспрессирующие клетки панкреатических островков могут быть зрелыми или незрелыми, и дополнительно дифференцированными или дальше коммитированными по пути дифференцировки, чем клеточные типы эндокринных прогениторов/предшественников, из которых они получены.
Использующеейся в настоящей заявке выражение «адекватно специализированные эндокринные клетки» или «культуры 7 стадии», или «незрелые эндокринные клетки», включая «незрелые бета-клетки», означают популяции эндокринных клеток, полученные in vitro, которые способны к функционированию in vivo, например, незрелые бета-клетки, которые при трансплантации секретируют инсулин в ответ на глюкозу крови. Адекватно специализированные эндокринные клетки или культуры 7 стадии могут иметь дополнительные характеристики, включая следующее: при трансплантации адекватно специализированные эндокринные клетки могут развиваться и созревать в функциональные клетки панкреатических островков. Адекватно специализированные эндокринные клетки могут быть обогащены эндокринными клетками (или обеднены неэндокринными клетками). В предпочтительном варианте осуществления более примерно 50% клеток в популяции адекватно специализированных эндокринных клеток являются CHGA+. В более предпочтительном варианте осуществления больше примерно 60% или 70% или 80% или 90% или 95% или 98% или 100% клеток в популяции адекватно специализированных эндокринных клеток являются CHGA+. В предпочтительном варианте осуществления меньше примерно 50% клеток в популяции адекватно специализированных эндокринных клеток являются CHGA-. В более предпочтительном варианте осуществления меньше примерно 15% клеток в популяции адекватно специализированных эндокринных клеток являются CHGA-. В более предпочтительном варианте осуществления меньше примерно 10% или 5% или 3% или 2% или 1% или 0,5%, или 0% клеток в популяции адекватно специализированных эндокринных клеток являются CHGA-. Далее, экспрессия определенных маркеров может быть супрессирована в адекватно специализированных эндокринных клетках, такая как экспрессия NGN3 во время стадии 3. Адекватно специализированные эндокринные клетки могут иметь повышенную экспрессию NGN3 на стадии 5. Адекватно специализированные эндокринные клетки могут быть моногормональными (например, только INS, только GCG или только SST). Адекватно специализированные эндокринные клетки могут совместно экспрессировать другие маркеры незрелых эндокринных клеток, включая NKX6.1 и PDX1. Адекватно специализированные эндокринные клетки могут моногормональными и совместно экспрессировать другие маркеры незрелых эндокринных клеток, включая NKX6.1 и PDX1. Адекватно специализированные эндокринные клетки могут содержать больше INS клеток, экспрессирующих единственный гормон, в качестве процентного содержания от общей популяции INS. В предпочтительном варианте осуществления адекватно специализированные эндокринные клетки содержат по меньшей мере 50% 1NS клеток, экспрессирующих единственный гормон, в качестве процентного содержания от общей популяции INS. Адекватно специализированные эндокринные клетки могут быть CHGA+/INS+/NKX6.1+ (тройными позитивными). В предпочтительном варианте осуществления более примерно 25% клеток в клеточной популяции являются CHGA+/INS+/NKX6.1+ (тройными позитивными). В предпочтительном варианте осуществления более примерно 30% или 40% или 50% или 60% или 70% или 80% или 90% или 95%, или 100% клеток в клеточной популяции являются CHGA+/INS+/NKX6.1+ (тройными позитивными).
Термин «незрелая эндокринная клетка», в частности, «незрелая бета-клетка», или его эквиваленты, означают клетку, полученную из любой другой эндокринной клетки-предшественника, включая эндокринный прогенитор/клетку-предшественник, панкреатическую эндодермальную (РЕ) клетку, панкреатическую клетку передней части пищеварительного тракта, клетку дефинитивной энтодермы, мезоэндодермальную клетку, или любую другую ранее выделенную клетку, описанную позже, которая экспрессирует по меньшей мере маркер, выбранный из группы, состоящей из INS, NKX6.1, PDX1, NEUROD, MNX1, NKX2.2, MAFA, РАХ4, SNAIL2, FOXA1 или FOXA2. Предпочтительно, незрелая бета-клетка, описанная в настоящей заявке, экспрессирует INS, NKX6.1 и PDX1, и более предпочтительно, совместно экспрессирует INS и NKX6.1. Термины «незрелая эндокринная клетка», «незрелая панкреатическая гормон-экспрессирующая клетка», или «незрелый панкреатический островок», или их эквиваленты означают, например, по меньшей мере унипотентную незрелую бета-клетку, или пре-бета клетку, как описано на Фиг. 45, и не включают другие незрелые клетки; например, термины не включают незрелые альфа (глюкагоновую) клетку, или незрелую дельта (соматостатиновую) клетку, или незрелую эпсилон (грелиновую) клетку, или незрелую клетку, секретирующую панкреатический полипептид (РР).
Условия среды для культивирования или роста многих стволовых клеток указаны в вариантах осуществления, описанных в настоящей заявке, включая среды с определенным составом, кондиционированные среды, бесфидерные среды, бессывороточные среды и тому подобное. Как применятся в настоящей заявке, термин «среда для выращивания» или «коктейль», или его эквиваленты означают среду, в которой недифференцированные или дифференцированные стволовые клетки (например, эмбриональные стволовые клетки приматов) пролиферируют in vitro. Характеристики среды включают среду, в которой культивируют клетки, и поддерживающую структуру (такую, как субстрат на твердой поверхности), если она присутствует. Способы культивирования или поддержки плюрипотентных клеток и/или дифференцировки плюрипотентных клеток также описаны в PCT/US2007/062755, озаглавленной «Композиции и способы, пригодные для культивирования дифференцируемых клеток», поданной 23 февраля 2007; заявке США №11/993,399, озаглавленной «Композиции для культивирования эмбриональных стволовых клеток и способы их применения», поданной 20 декабря 2007; и заявке США №11/875,057, озаглавленной «Способы и композиции для бесфидерных сред для плюрипотентных стволовых клеток, содержащих человеческую сыворотку», поданной 19 октября 2007.
Термин «по существу» или «в основном» означает минимальное или сниженное количество компонента или клетки, присутствующих в любой клеточной популяции или культуре, например, культуры незрелых бета-клеток, описанные в настоящей заявке, являются «по существу или в основном однородными», «по существу или в основном незрелыми бета-клетками, экспрессирующими INS, NKX6.1 и PDX1, и по существу или полностью не экспрессирующими NGN3». Другие примеры включают «по существу или полностью hES клетки», «по существу или полностью клетки дефинитивной энтодермы», «по существу или полностью клетки энтодермы передней части пищеварительного тракта», «по существу или полностью PDX1-негативные клетки энтодермы передней части пищеварительного тракта», «по существу или полностью PDX1-позитивные панкреатические эндодермальные клетки», «по существу или полностью панкреатические эндокринные клетки-предшественники», «по существу или полностью панкреатические эндокринные клетки», и тому подобное, но не ограничиваются ими.
Что касается клеток в клеточных культурах или клеточных популяциях, термин «по существу не содержит» означает, что указанный тип клеток, который не содержит клеточная культура или клеточная популяция, присутствует в количестве менее примерно 10%, менее примерно 9%, менее примерно 8%, менее примерно 7%, менее примерно 6%, менее примерно 5%, менее примерно 4%, менее примерно 3%, менее примерно 2% или менее примерно 1% от общего числа клеток, присутствующих в клеточной культуре или клеточной популяции.
Термин «со сниженным содержанием сыворотки» или «бессывороточная», или их эквиваленты означают клеточные культуры, выращенные в среде, содержащей сниженное количество сыворотки или по существу свободной от сыворотки, или без сыворотки. В определенных условиях культивирования концентрации сыворотки могут находиться в диапазоне примерно от 0 об. % до 10 об. %. Например, в некоторых способах дифференцировки концентрация сыворотки в среде может составлять меньше примерно 0,05% (о/о), меньше примерно 0,1% (о/о), меньше примерно 0,2% (о/о), меньше примерно 0,3% (о/о), меньше примерно 0,4% (о/о), меньше примерно 0,5% (о/о), меньше примерно 0,6% (о/о), меньше примерно 0,7% (о/о), меньше примерно 0,8% (о/о), меньше примерно 0,9% (о/о), меньше примерно 1% (о/о), меньше примерно 2% (о/о), меньше примерно 3% (о/о), меньше примерно 4% (о/о), меньше примерно 5% (о/о), меньше примерно 6% (о/о), меньше примерно 7% (о/о), меньше примерно 8% (о/о), меньше примерно 9% (о/о) или меньше примерно 10% (о/о). В некоторых способах клетки препервичной полоски выращивают без сыворотки или без замены сыворотки. В еще одних способах клетки препервичной полоски выращивают в присутствии В27. В таких способах концентрация В27 добавки может быть в диапазоне примерно от 0,1% (о/о) до 20% (о/о).
В других способах клеточные культуры выращивают в присутствии В27. В таких способах концентрация добавки В27 может быть в диапазоне примерно от 0,1% (о/о) до 20% (о/о), или в концентрациях выше примерно 20% (о/о). В некоторых способах концентрация В27 в среде составляет примерно 0,1% (о/о), примерно 0,2% (о/о), примерно 0,3% (о/о), примерно 0,4% (о/о), примерно 0,5% (о/о), примерно 0,6% (о/о), примерно 0,7% (о/о), примерно 0,8% (о/о), примерно 0,9% (о/о), примерно 1% (о/о), примерно 2% (о/о), примерно 3% (о/о), примерно 4% (о/о), примерно 5% (о/о), примерно 6% (о/о), примерно 7% (о/о), примерно 8% (о/о), примерно 9% (о/о), примерно 10% (о/о), примерно 15% (о/о) или примерно 20% (о/о). Альтернативно, концентрация внесенной добавки В27 может быть измерена в виде кратности крепости коммерческого маточного раствора В27. Например, В27 поставляется Invitrogen (Карлсбад, Калифорния) в виде 50Х маточного раствора. Добавление достаточного количества такого маточного раствора к достаточному объему среды для выращивания обеспечивает среду с добавлением необходимого количества В27. Например, добавление 10 мл 50Х маточного раствора В27 к 90 мл среды для выращивания обеспечивает среду для выращивания с 5Х В27. Концентрация добавки В27 может составлять примерно 0,1Х, примерно 0,2Х, примерно 0,3Х, примерно 0,4Х, примерно 0,5Х, примерно 0,6Х, примерно 0,7Х, примерно 0,8Х, примерно 0,9Х, примерно 1X, примерно 1,1Х, примерно 1,2Х, примерно 1,3Х, примерно 1,4Х, примерно 1,5Х, примерно 1,6Х, примерно 1,7Х, примерно 1,8Х, примерно 1,9Х, примерно 2Х, примерно 2,5Х, примерно 3Х, примерно 3,5Х, примерно 4Х, примерно 4,5Х, примерно 5Х, примерно 6Х, примерно 7Х, примерно 8Х, примерно 9Х, примерно 10Х, примерно 11Х, примерно 12Х, примерно 13Х, примерно 14Х, примерно 15Х, примерно 16Х, примерно 17Х, примерно 18Х, примерно 19Х, примерно 20Х и более чем примерно 20Х.
В еще одном аспекте, где требуются низкие уровни инсулина для дифференцировки, В27 не применяют, поскольку В27 содержит высокие уровни инсулина. Например, автор настоящей заявки определил уровни инсулина в 100х маточных растворах В27 и ITS и 1х рабочих маточных растворах в каждой из RPMI. Уровни инсулина определяли с применением сверхчувствительного набора для ИФА инсулина Mercodia в соответствии с инструкциями производителя. Анализы проводили на не открытых 100х и 50х маточных растворах В27 и ITS, соответственно, получая их от Life Technologies (Карлсбад, Калифорния). Результаты анализа показаны ниже в Таблице 3.

В Таблице 3 показано, что примерно 3 мкг/мл и 10 мкг/мл инсулина присутствует в 1x В27 и 1x ITS, соответственно. Далее, концентрация инсулина в 100х маточном растворе ITS соответствует концентрации инсулина, указанной производителем. Life Technologies не указывает концентрацию инсулина для 50х В27, так что нельзя сравнить с уровнями инсулина, указанными производителем, однако, вышеуказанная концентрация инсулина около 160 мкг/мл точно основана на измерении 100Х маточного раствора ITS, которое проводили в то же самое время и на основе McLean и др. (2007) выше, где показано, что инсулин и инсулиноподобный фактор роста (IGF) оба являются хорошо известными агонистами PI3-киназа зависимой передачи сигнала, позволяя предположить, что ингибитор PI3-киназы (например, LY 294002) будет стимулировать формирование дефинитивной энтодермы путем ингибирования эффекторов инсулина/ IGF. См. Фиг. 6А и В, в частности, экспрессия SOX17 снижается в 4 раза с 1 мкг/мл инсулина. Инсулин и/или IGF присутствуют на биологически активных уровнях в различных добавках для сред, таких как KSR, FCS (эмбриональная телячья сыворотка), FBS (фетальная бычья сыворотка), В27 и StemPro34 могут ингибировать дифференцировку дефинитивной энтодермы в таких условиях культивирования, см. Jiang и др. (2007) выше.
Использующиеся в настоящей заявке «экзогенные» или «экзогенно добавленные» соединения, такие как агенты, компоненты, факторы роста, факторы дифференцировки, и тому подобные, в контексте культур или кондиционированных сред, означают соединения, которые добавляют в культуры или среды для дополнения любыми соединениями или факторами роста, которые уже могут присутствовать или не присутствовать в культуре или среде. Например, в некоторых вариантах осуществления клеточные культуры и/или клеточные популяции не включают экзогенно добавленного ретиноида.
Использующийся в настоящей заявке термин «ретиноид» означает ретинол, ретиналь или ретиноевую кислоту, а также производные любого из этих соединений. В предпочтительном варианте осуществления ретиноид является ретиноевой кислотой.
Термин «фактор роста семейства FGF», «фактор роста фибробластов» или «член семейства фактора роста фибробластов» означает FGF, выбранный из группы, состоящей из FGF1, FGF2, FGF3, FGF4, FGF5, FGF6, FGF7, FGF8, FGF9, FGF10, FGF11, FGF12, FGF13, FGF14, FGF15, FGF16, FGF17, FGF18, FGF19, FGF20, FGF21, FGF22 и FGF23. В некоторых вариантах осуществления «фактор роста семейства FGF», «фактор роста фибробластов» или «член семейства фактора роста фибробластов» означает любой фактор роста, обладающий гомологией и/или функцией, подобной известному члену семейства фактора роста фибробластов.
Термин «фактор роста суперсемейства TGFβ» или «лиганд суперсемейства TGFβ» или «активатор сигнального пути TGFβ» или его эквиваленты означают более 30 структурно связанных белков, включая субсемейства, включающие TGFβ1, TGFβ2, TF-β3, GDF-15, GDF-9, ВМР-15, ВМР-16, ВМР-3, GDF-10, ВМР-9, ВМР-10, GDF-6, GDF-5, GDF-7, ВМР-5, ВМР-6, ВМР-7, ВМР-8, ВМР-2, ВМР-4, GDF-3, GDF-1, GDF 11, GDF8, активины βC, βЕ, βА и βВ, ВМР-14, GDF-14, MIS, ингибин-альфа, Lefty1, Lefty2, GDNF, нейротурин, персефин и артемин. См. Chang и др. (2002) Endocrine Rev. 23(6):787-823. Эти лиганды, как правило, синтезируются в виде пропептидов примерно из 400-500 аминокислот (АК), и «фактор роста суперсемейства TGFβ» или «лиганд суперсемейства TGFβ» или «активатор сигнального пути TGFβ» или его эквиваленты могут быть непроцессированным белком или его протеолитическим пептидом.
Термин «лиганд ERBB» означает лиганд, связывающийся с любым из ErbB рецепторов, которые в свою очередь могут димеризоваться с другим ErbB рецептором, или функционировать как мономер, таким образом активируя тирозинкиназную активность ErbB части мономерного или димерного или гетеродимерного рецептора. Не ограничивающие примеры ErbB лигандов включают нейрегулин-1; сплайс-варианты и изоформы нейрегулина-1; включая HRG-β, HRG-α, Neu фактор дифференцировки (NDF), ацетилхолиновый рецептор - индуцирующую активность (ARIA), глиальный фактор роста 2 (GGF2), и фактор, полученный из сенсорных и моторных нейронов (SMDF); нейрегулин-2; сплайс-варианты и изоформы нейрегулина-2, включая NRG2-β, но не ограничиваясь им; эпирегулин и бирегулин, но не ограничиваясь ими.
Использующийся в настоящей заявке термин «экспрессия» означает продукцию материала или вещества, а также уровень или количество продукции материала или вещества. Таким образом, определение экспрессии специфического маркера означает детекцию относительного или абсолютного количества экспрессируемого маркера, или просто детекцию присутствия или отсутствия маркера.
Использующийся в настоящей заявке термин «маркер» означает любую молекулу, которую можно наблюдать или выявлять. Например, маркер может включать нуклеиновую кислоту, такую как транскрипт специфического гена, полипептидный продукт гена, полипептид - не-генный продукт, гликопротеин, углевод, гликолипид, липид, липопротеин, или малую молекулу (например, молекулы с молекулярной массой меньшей 10,000 а.е.м.), но не ограничивается ими.
Для большинства маркеров, описанных в настоящей заявке, обеспечивается официальный символ гена Международной организации по изучению генома человека (HUGO). Такие символы, которые разработаны Комитетом по номенклатуре генов HUGO, обеспечивают уникальные аббревиатуры для каждого из названных генов человека и продуктов генов. Эти символы генов легко распознаются и могут легко быть ассоциированы с соответствующим уникальным геном человека и/или белковой последовательностью рядовым специалистом в данной области техники.
В соответствии с обозначениями HUGO, определены следующие символы генов: GHRL - грелин; IAPP - островковый амилоидный полипептид; INS - инсулин; GCG - глюкагон; ISL1 - фактор транскрипции ISL1; РАХ6 - ген спаренных боксов 6; РАХ4 - ген спаренных боксов 4; NEUROG3 - нейрогенин 3 (NGN3); NKX2-2 - NKX2-фактор-транскрипции-зависимый, локус 2 (NKX2.2); NKX6-1 - NKX6-фактор транскрипции-зависимый, локус 1 (NKX6.1); IPF1 - промоторный инсулиновый фактор 1 (PDX1); ONECUT1 - one cut домен, член семейства 1 (HNF6); HLXB9 - гомеобокс В9 (НВ9); TCF2 - фактор транскрипции 2, печеночный (HNF1b); FOXA1 - forkhead box Al; HGF - фактор роста гепатоцитов; IGF1 - инсулиноподобный фактор роста 1; POU5F1 - POU домен, класс 5, фактор транскрипции 1 (ОСТ4); NANOG - Nanog гомеобокс; SOX2 - SRY (полоопределяющий участок Y)-бокс 2; CDH1 - кадгерин 1, тип 1, Е-кадгерин (ECAD); Т - brachyury гомолог (BRACH); FGF4 - фактор роста фибробластов 4; WNT3 - бескрылого типа MMTV семейства сайта интеграции, член 3; SOX17 - SRY (полоопределяющий участок Y)-бокс 17; GSC - гусекоид; CER1 - (cerberus 1, cysteine-knot суперсемейства, гомолог (CER); CXCR4 - хемокина (С-Х-С мотив) рецептор 4; FGF17 - фактор роста фибробластов 17; FOXA2 - forkhead бокс А2; SOX7 - SRY (полоопределяющий участок Y)-бокс 7; SOX1 - SRY (полоопределяющий участок Y)-бокс 1; AFP - альфа-фетопротеин; SPARC - секретируемый белок, кислый, богатый цистеином (остеонектин); и THBD - тромбомодулин (ТМ), NCAM - молекула адгезии нервных клеток; SYP - синаптопсин; ZIC1 - Zic семейства член 1; NEF3 - нейрофиламент 3 (NFM); SST - соматостатин; MAFA - v-maf мышечно-апоневротической фибросаркомы онкогена гомолог A; MAFB - v-maf мышечно-апоневротической фибросаркомы онкогена гомолог В; SYP - синаптопсин; CHGA - хромогранин А (паратиреоидный секреторный белок 1); NGN3 - нейрогенин 3, NKX2.2 - NK2 гомеобокс 2; NKX6.1 - NK6 гомеобокс 1; ID1 - ингибитор связывания ДНК 1, GHRL - грелин/обестатин препропептид, GSK - киназа гликоген-синтазы, G6PC2-глюкозо-6-фосфатаза, UCN3 - урокортин 3, PCSK1 - пропротеин-конвертаза субтилизин/кексин тип 1, SLC30A8 - семейства носителей растворенных веществ 30 (транспортера цинка) член 8, и CADH1 - Е-кадгерин. Термины «фактор роста фибробластов 7» (FGF7) и «фактор роста кератиноцитов» (KGF) являются синонимами.
Развитие плюрипотентных клеток в различные мультипотентные и/или дифференцированные клетки можно контролировать путем определения относительной экспрессии генов, или маркеров генов, характерных для специфических клеток, по сравнению с экспрессией второго или контрольного гена, например, конститутивных генов. В некоторых способах экспрессию определенных маркеров оценивают путем детекции присутствия или отсутствия маркера. Альтернативно, экспрессию некоторых маркеров можно определить путем измерения уровня, на котором маркер присутствует в клетке из клеточной культуры или клеточной популяции. В таких способах измерение экспрессии маркеров может быть качественным или количественным. Одним способом количественного определения экспрессии маркеров, продуцируемых генами маркеров, является применение количественной ПЦР (Q-PCR). Способы выполнения Q-PCR хорошо известны в данной области техники. Другие способы, известные в данной области техники, также можно применять для количественного определения экспрессии гена маркера. Например, экспрессию продукта маркерного гена можно определить с применением антител, специфичных для интересующего продукта гена маркера.
Кроме того, многочисленные способы и/или методологии доступны для анализа многих генов с высокой чувствительностью и эффективностью. Например, по меньшей мере с nCounter System, поставляемой Nanostring (Сиэттл, Вайтоминг, США), потребители в настоящее время могут анализировать уровни экспрессии до 800 генов одновременно, с чувствительностью, сопоставимой с системами количественной ПЦР и менее чем за 15 минут времени операций на реакцию. Следовательно, общую РНК во всех образцах (например, на стадии 4 ПЭК и стадии 7 культур эндокринных прогениторов/предшественников и эндокринных клеток, соответственно) можно выделить (например, в трех повторностях) с применением 6100 экстрактора нуклеиновых кислот (Applied Biosystems; Фостер-Сити, Калифорния, США); и примерно 100 нг требуется для реакции количественного определения экспрессии гена с применением Nanostring nCounter System. Вместо аналоговой детекции, nCounter System применяет цифровую детекцию, где транскрипт каждого гена выявляется зондом, связанным с сегментом ДНК, прикрепленным к уникальной нити окрашенных флюорофоров (молекулярному штрих-коду). Таким образом, идентификация каждого транскрипта зависит только от порядка флюоресцентов на нити, а не от интенсивности флюоресцентов. Во-вторых, число общих транскриптов в образце определяют путем подсчета общего числа выявленных конкретных нитей флюоресцентов (штрих-кода). Далее, этот способ не требует амплификации целевой мРНК, так, диапазоном является биологический диапазон экспрессии, как правило, с трехкратной амплитудой. В настоящее время система позволяет измерять всего лишь 1,2-кратные изменения единственного транскрипта в виде 20 копий на клетку (10 фМ) со статистической достоверностью (р<0,05). Для генов, экспрессируемых на уровнях от 0,5 до 20 копий на клетку, 1,5-кратные различия уровней экспрессии можно выявить с тем же самым уровнем достоверности.
В некоторых способах наивысшая экспрессия следующих генов, по сравнению с более низкой экспрессией других генов, является показателем для некоторых популяций клеток, например, SOX17, SOX7, AFP или THBD являются показателями экстраэмбриональной энтодермы; NODAL и/или FGF8 являются показателями препервичной полоски; brachyury, FGF4, SNAI1 и/или WNT3 являются показателями мезоэнтодермы; CER, GSC, CXCR4, SOX17 и FOXA2 являются показателями дефинитивных эндодермальных клеток; SOX17, FOXA2, FOXA1, HNF1B и HNF4A являются показателями энтодермы передней части пищеварительного тракта (или PDX1-негативной энтодермы); PDX1, HNF6, SOX9 и PROX1 являются показателями PDX1-позитивной энтодермы; PDX1, NKX6.1, PTFA1, CPA и cMYC являются показателями панкреатического эпителия (РЕ или панкреатического прогенитора); NGN3, РАХ4, ARX и NKX2.2 являются показателями эндокринных прогениторов/клеток-предшественников; а INS, GCG, GHRL, SST и PP являются показателями различных эндокринных клеток; относительно высокая экспрессия генов MAFA - MAFB является показателем инсулин-секретирующих эндокринных клеток; а относительно высокая экспрессия генов MAFB - MAFA является показателем глюкагон-секретирующих эндокринных клеток. На некоторых фигурах и чертежах показаны только те «характерные» или «ключевые» маркеры, которые типичны для конкретного типа клеток конкретной линии. Рядовому специалисту в данной области техники понятно, что другие маркеры или гены экспрессированы, но не показаны и не описаны, например, гены, которые конститутивно экспрессированы, или экспрессированы в каждой или во множестве, или в большинстве клеточных типов определенной линии или всех линий.
Способы продукции индуцированных плюрипотентных стволовых (iPS) клеток
Варианты осуществления, описанные в настоящей заявке, не ограничиваются каким-либо одним типом iPS клеток или каким-либо одним способом получения iPS клеток. Варианты осуществления не ограничиваются и не зависят от уровней эффективности продукции iPS клеток, поскольку существуют различные способы. Варианты осуществления, описанные в настоящей заявке, применяются к дифференцировке iPS клеток в эндодермальные клеточные линии, и к их применению.
Исследования с применением определенных ядерных перепрограммирующих факторов обеспечивают получения плюрипотентных стволовых клеток или подобных плюрипотентным стволовых клеток из собственных соматических клеток пациента. Эти клетки также называются индуцированными плюрипотентными (iPS) стволовыми клетками. Настоящее изобретение описывает различные линии iPS клеток, предоставленные Shinya Yamanaka, Университет Киото. Однако, другие линии iPS клеток, например, описанные James Thomson и др., включены в настоящее изобретение. См. публикацию США 20090047263, Международную публикацию WO2005/80598, публикацию США 20080233610 и Международную публикацию WO2008/11882. Таким образом, использующийся в настоящей заявке термин «индуцированные плюрипотентные стволовые (iPS) клетки» означает клетки, обладающие свойствами, подобными свойствам других плюрипотентных стволовых клеток, например, hES клетки, hEG клетки, pPS клетки (плюрипотентные стволовые клетки приматов), партеногенные клетки, и тому подобное.
Ядерные программирующие факторы описаны в публикации США 20090047263, Международной публикации WO2005/80598, публикации США 20080233610 и Международной публикации WO2008/11882, и применяются для индукции перепрограммировании дифференцированных клеток без применения яйцеклеток, эмбрионов или ES клеток. Способы приготовления индуцированных iPS клеток из соматических клеток путем применения ядерных перепрограммирующих факторов подобны тем, которые описаны в настоящем изобретении, и не ограничиваются конкретно. В предпочтительных вариантах осуществления ядерный перепрограммирующий фактор контактирует с соматическими клетками в среде, в которой соматические клетки и индуцированные плюрипотентные стволовые клетки могут пролиферировать. Преимуществом некоторых вариантов осуществления, описанных в настоящей заявке, является то, что индуцированные плюрипотентные стволовые клетки можно приготовить путем контакта ядерного перепрограммирующего фактора с соматическими клетками при отсутствии яйцеклеток, эмбрионов или эмбриональных стволовых (ES) клеток. С применением ядерного перепрограммирующего фактора ядро соматической клетки можно перепрограммировать для получения iPS клетки или «ES-подобной клетки».
Плюрипотентные стволовые клетки, описанные в настоящей заявке, являющиеся hES клетками или iPS клетками, могут экспрессировать любое количество маркеров плюрипотентных клеток, включая щелочную фосфатазу (АР); ABCG2; стадиеспецифический эмбриональный антиген-1 (SSEA-1); SSEA-3; SSEA-4; TRA-1-60; TRA-1-81; Tra-2-49/6E; ERas/ECAT5, Е-кадгерин; βIII-тубулин; альфа-гладкомышечный актин (альфа-SMA); фактор роста фибробластов 4 (Fgf4), Cripto, Dax1; белок «цинковые пальцы» 296 (Zfp296); N-ацетилтрансферазу-1 (Nat1); ассоциированный с ES клетками транскрипт 1 (ЕСАТ1); ESG1/DPPA5/ECAT2; ЕСАТ3; ЕСАТ6; ЕСАТ7; ЕСАТ8; ЕСАТ9; ECAT10; ЕСАТ15-1; ЕСАТ15-2; Fthl17; Sal14; фактор транскрипции недифференцированных эмбриональных клеток (Utf1); Rex1; р53; G3PDH; теломеразу, включая TERT; молчащие гены X хромосомы; Dnmt3a; Dnmt3b; TRIM28; F-бокс содержащий белок 15 (Fbx15); Nanog/ECAT4; Oct3/4; Sox2; Klf4; c-Myc; Esrrb; TDGF1; GABRB3; Zfp42, FoxD3; GDF3; CYP25A1; дифференцировочный ассоциированный с плюрипотентностью 2 (DPPA2); Т-клеточной лимфомы точка разрыва 1 (Tcl1); DPPA3/Stella; DPPA4 и тому подобные, но не ограничиваясь ими. Необходимо понять, что настоящее изобретение не ограничивается маркерами, перечисленными в настоящей заявке, и охватывает такие маркеры, как маркеры клеточной поверхности, антигены, и другие продукты генов, включая EST, РНК (включая микроРНК и антисмысловые РНК), ДНК (включая гены и кДНК), и их части.
В одном варианте осуществления линии iPS клеток, применяемые в настоящей заявке, содержат следующие гены ядерных перепрограммирующих факторов: семейство генов Oct, семейство генов Klf, и семейство генов Sox. В одной линии iPS клеток обеспечен каждый из трех видов генов: Oct3/4, Klf4, и Sox2. Применялись другие продукты генов линии iPS из каждого из следующих трех видов: ген семейства Oct, ген семейства Klf, и ген семейства Мус, например, Oct3/4, Klf4 и с-Мус. Соответственно, необходимо понять, что ядерный перепрограммирующий фактор может быть с геном семейства Мус или без него.
Ядерные перепрограммирующие факторы, описанные в настоящей заявке и также известные в данной области техники, можно применять для генерации iPS клеток из дифференцированных взрослых соматических клеток, и они не ограничиваются типом перепрограммируемых соматических клеток, т.е. любой вид соматических клеток можно перепрограммировать или дедифференцировать. Поскольку перепрограммирование соматических клеток не требует яйцеклетки и/или эмбриона, iPS клетка может быть клеткой млекопитающего, таким образом, обеспечивая возможность генерации пациент-или болезнь-специфических плюрипотентных стволовых клеток.
Описаны вирусные, невирусные и неинтегрирующие вирусные способы генерации индуцированных плюрипотентных стволовых клеток (iPSC) с применением аденовирусов, плазмид или вырезания перепрограммирующих факторов с применением Cre-loxP3, или piggyback транскрипции. См. Stadtfeld, М., и др., Science 322, 945-949 (2008); Okita, K. и др., Science 322, 949-953 (2008); Kaji, K. и др. Nature 458, 771-775 (2009); Soldner, F. и др. Cell 136, 964-977 (2009); и Woltjen, K. и др. Nature 458, 766-770 (2009), которые включены посредством ссылки во всей полноте. См. также патентную заявку США №20100003757 от Mack, А. и др. (опубликовано 7 января 2010) и №PCT/US2009/037429 от Shi и др. Эти способы, однако, обладают низкой перепрограммирующей эффективностью (<0,003%), и могут оставлять остаточные векторные последовательности, несмотря на вырезание, что ограничивает терапевтическое применение. Например, вирусная интеграция в геном хозяина и гиперэкспрессия вышеуказанных факторов транскрипции является потенциально характеристикой, которая отличает ES клетки и iPS клетки. См. Solder, F. и др., Cell 136:964-977 (2009); Foster и др., Oncogene 24:1491-1500 (2005); и Hochedlinger, K. и др., Cell 121:465-477 (2005).
В других вариантах осуществления настоящего изобретения способы генерации iPSC включают эписомальные векторы, полученные из вируса Эпштейн-Барра. См. Yu, J. и др. Science 324, 797-801 (2009) и заявку США №20100003757 от Mack, А. и др. опубликованную 7 января 2010. Эти способы требуют трех отдельных плазмид, несущих комбинацию семи факторов, включая онкоген SV40.
В другом варианте осуществления изобретения способы генерации iPSC включают iPSC на белковой основе от мышиных и человеческих фетальных и неонатальных клеток. См. Zhou, Н. и др. Cell Stem Cell 4, 381-384 (2009); и Kim, D. и др. Cell Stem Cell 4, 472-476 (2009). Эти методологии осуществляют с применением химической обработки (например, вальпроевой кислотой в случае Zhou и др. 2009, выше) или многих циклов обработки (Kim и др. 2009, выше).
В другом варианте осуществления настоящего изобретения можно применять миникольцевые векторы или плазмиды, которые являются молекулами сверхспиральной ДНК, утратившей бактериальное происхождение репликации и генов устойчивости к антибиотикам. См. See Chen, Z.-Y. и др., Mol. Ther. 8, 495-500 (2003); Chen, Z.-Y. и др., Hum. Gene Ther. 16, 126-131 (2005); и Jia, F. и др., Nature Methods Advance Publication Online, 7 февраля 2010. Эти методологии генерируют iPSC с более высокой эффективностью трансфекции и более длительной эктопической экспрессией, поскольку они имеют меньшую активацию экзогенных механизмов сайленсинга.
В другом варианте осуществления настоящего изобретения iPS клетки могут быть генерированы от людей - пациентов, включая больных диабетом, боковым амиотрофическим склерозом, спинальной мышечной дистрофией и болезнью Паркинсона. См. Maehr и др. PNAS USA 106(37): 15768-73 (2009); Dimos и др., Science, 321:1218-21 (2008); Ebert и др. Nature 457:277-80 (2009); Park и др. Cell 134:877-886 (2008); и Soldner и др., Cell 136:964-977. По меньшей мере одним преимуществом получения hIPS клеток от пациентов со специфическими заболеваниями является то, что полученные клетки могут содержать генотип и иметь клеточный ответ для заболевания человека. Также, см. Таблицу 4, перечисляющую по меньшей мере некоторые существующие человеческие iPS клеточные линии. Эта информация была получена из литературы и доступна в базах данных, включая например, Реестр стволовых клеток Национального института здравоохранения (NIH), Реестр эмбриональных стволовых клеток человека и Международный реестр стволовых клеток, расположенный в Медицинской школе Массачусетского университета, Ворчестер, Массачусетс, США. Эти базы данных периодически обновляются, когда становятся доступными клеточные линии и получена регистрация.
Варианты осуществления композиций и способов, описанных в настоящей заявке, подразумевают применение различных дифференцируемых плюрипотентных стволовых клеток приматов, включая человеческие плюрипотентные стволовые клетки, такие как hESC, включая СуТ49, СуТ212, СуТ203, СуТ25, (коммерчески доступные по меньшей мере ко времени подачи настоящей заявки от ViaCyte Inc., расположенном в 3550 General Atpmics Court, Сан-Диего, Калифорния, 92121) BGOl, BG02 и MEL1, и индуцированные плюрипотентные стволовые (iPS) клетки, такие как iPSC-482c7 и iPSC-603 (Cellular Dynamics International, Inc., Мэдисон, Висконсин) и iPSC-G4 (далее «G4») и iPSC-B7 (далее «В7») (Shinya Yamanaka, Центр исследования iPS клеток, Университет Киото), но не ограничиваясь ими; исследования с применением этих и других человеческих iPS клеток подробно описаны в заявках на патент США №№12/765,714 и 13/761,078, обе из них озаглавлены «Клеточные композиции из дедифференцированных перепрограммированных клеток», поданных 22 апреля 2010 и 6 февраля 2013, соответственно. Некоторые из этих человеческих плюрипотентных стволовых клеток зарегистрированы в национальных реестрах, таких как Реестр Национального института здравоохранения (NIH), и перечислены в Реестре стволовых клеток человека NIH (например, СуТ49, регистрационный номер 0041). Информацию по СуТ49, другим доступным клеточным линиям также можно найти в Интернете на stemcells.nih.gov/research/registry. Другие клеточные линии, например, BG01 и BG0lv, поставляются и распространяются третьими лицами от WiCell®, филиалом Висконсинского международного банка стволовых клеток (WISC) (номер по каталогу BG01) и АТСС (номер по каталогу SCRC-2002), соответственно. В то время как другие клеточные линии, описанные в настоящей заявке, могут быть не зарегистрированы и не распределяться биологическим репозиторием, таким как WiCell® или АТСС, такие клеточные линии доступны напрямую или опосредованно от основных исследователей, лабораторий и/или институтов. Публичный запрос на клеточные линии и реагенты, например, является обычным для рядовых специалистов в данной области техники. Как правило, передача этих клеток или материалов осуществляется путем стандартного соглашения по передаче материалов между собственником клеточной линии или материала, и потребителем. Передача этих типов материалов осуществляется частным образом в исследовательской среде, в частности, в области науки о жизни. Фактически, автор заявки рутинно передавал клетки со времени их получения и характеристики, включая СуТ49 (2006), СуТ203 (2005), Cyt212 (2009), СуТ25 (2002), BG01 (2001), BG02 (2001), BG03 (2001) и BG01v (2004), посредством таких соглашений с коммерческими и некоммерческими промышленными партнерами и соавторами. Год в скобках после каждой клеточной линии в предыдущем списке указывает год, когда клеточные линии или материалы становятся публично доступными и бессмертными (например, из клеточных банков, где они изготовлены), и таким образом, разрушение другого эмбриона не проводится и не требуется, благодаря получению этих клеточных линий для создания композиций и осуществления способов, описанных в настоящей заявке.
Таблицы 4 и 5 являются не ограничивающими перечнями некоторых iPSC и hESC, соответственно, которые доступны во всем мире для исследовательских и/или коммерческих целей, и пригодны для применения в способах и композициях из настоящего изобретения. Информация из Таблиц 4 и 5 получена из литературы и публичных баз, включая, например Реестр стволовых клеток Национального института здравоохранения (NIH), Реестр эмбриональных стволовых клеток человека и Международный реестр стволовых клеток медицинской школы Массачусетс кого университета, США. Эти базы данных периодически обновляются, когда клеточные линии становятся доступными и зарегистрированными.
Человеческие iPSC, описанные в настоящей заявке (по меньшей мере iPSC-603 и iPSC-482-c7), получали от Cellular Dynamics International, Inc. (Мэдисон, Висконсин, США).








Другим преимуществом применения hIPS клеток является то, что hIPS клетки будут соответствовать иммунологически аутологичной клеточной популяции; и пациент-специфические клетки можно получать для изучения происхождения и прогрессирования заболевания. Таким образом, можно понять первопричины заболевания, что позволит создать представление, ведущее к разработке профилактического и терапевтического лечения заболевания.
Плюрипотентные человеческие эмбриональные стволовые (hES) клетки
Некоторые варианты осуществления направлены на способы получения дефинитивных эндодермальных клеток и наконец, любого типа клеток, полученных из эндодермальной линии, включая энтодерму передней части пищеварительного тракта, панкреатическую энтодерму, эндокринные клетки-предшественники и/или гормон-экспрессирующие клетки панкреатических островков, но не ограничиваясь ими, с применением человеческих эмбриональных стволовых (hES) клеток в качестве исходного материала. Эти hES клетки могут быть клетками, происходящими из морулы, внутренней клеточной массы эмбриона или клетками, полученными из эмбриональных половых гребней. Человеческие эмбриональные стволовые клетки могут поддерживаться в культуре в плюрипотентном состоянии без существенной дифференцировки с применением способов, известных в данной области техники. Такие способы описаны, например, в патентах США №№5,453,357; 5,670,372; 5,690,926; 5,843,780; 6,200,806 и 6,251,671.
В некоторых способах плюрипотентные стволовые клетки, например, hES клетки, поддерживаются на фидерном слое. В таких способах может применяться любой фидерный слой, позволяющий плюрипотентной клетке сохраняться в плюрипотентном состоянии. Одним обычно используемым фидерным слоем для культивирования человеческих эмбриональных стволовых клеток является слой мышиных фибробластов. Недавно были разработаны фидерные слои человеческих фибробластов для применения при культивировании плюрипотентных клеток (см. патентную заявку США №2002/0072117). Альтернативные способы позволяют поддерживать плюрипотентные клетки без применения фидерного слоя.
Плюрипотентные клетки, раскрытые в настоящей заявке, можно поддерживать в культуре с сывороткой или без сыворотки, с экстрацеллюлярным матриксом или без него, с FGF или без него. В некоторых процедурах поддержки плюрипотентных клеток применяют замену сыворотки. Эти и другие способы культивирования и дифференцировки плюрипотентных или мультипотентных клеток, соответственно, описаны в PCT/US2007/062755, поданной 23 февраля 2007, и озаглавленной «Композиции и способы культивирования дифференциальный клеток», и PCT/US2008/080516, поданной 20 октября 2008, и озаглавленной «Способы и композиции для бесфидерных сред для плюрипотентных стволовых клеток, содержащих человеческую сыворотку».
Изобретение, раскрытое в настоящей заявке, пригодно для всех hES клеточных линий, и по меньшей мере тех, которые перечислены в Таблице 5, которые являются коммерчески доступными от указанной компании, или могут быть приобретены от WiCell и указаны на веб-сайте wicell.org/home/stem-cell-lines/order-stem-cell-lines/obtain-stem-cell-lines.cmsx. Эта информация получена из литературы и публичных баз данных, включая, например, Реестр стволовых клеток Национального института здравоохранения (NIH), Реестр эмбриональных стволовых клеток человека и Международный реестр стволовых клеток, расположенный в Медицинской школе Массачусетского университета, Ворчестер, Массачусетс, США. Эти базы данных периодически обновляются, когда становятся доступными клеточные линии и получена регистрация. Имеется по меньшей мере 254 коммерческих линий iPSC, перечисленных в Международной реестре стволовых клеток, и 1211 коммерческих линий hESC. В таблице 5 внизу не включены все перечисленные hESC и iPSC, но скорее приведены примеры потенциально доступных плюрипотентных стволовых клеток.













Альтернативные способы получения плюрипотентных стволовых клеток
Существуют способы получения плюрипотентных стволовых клеток, таких как ES клетки млекопитающих, без разрушения эмбриона. Вкратце, Advanced Cell Technology (Ворчестер, Массачусетс, США) опубликовано 3 научных статьи в журналах, описывающих получение мышиных и человеческих ES клеток из единичных бластомеров, когда эмбрион оставался интактным, и таким образом, не происходило его разрушения. Позднее в 2005 Chung и др. вначале описали способы получения мышиных ES клеток из единственного бластомера. См. Chung и др. (2006) Nature 439: 216-219, опубликовано онлайн 16 октября 2005. Chung и др. (2006) описывают взятие биоптатов из эмбриона с применением методик микроманипуляции, подобных методикам, используемым для преимплантационной генетической диагностики (ПГД); см. страницу 217. К этому времени Chung и др. (2006) совместно культивировали бластомерные клеточные линии с другими эмбриональными стволовыми клетками, но позже были разработаны способы, где это не требовалось. См. Chung и др. (2008) «Human Embryonic Stem Cell Lines Generated without Embryo Destruction», Cell Stem Cell 2: 113-117 («Линии человеческих эмбриональных стволовых клеток, генерированные без деструкции эмбрионов»).
Преимплантационную генетическую диагностику применяют для анализа генетических аномалий эмбрионов перед имплантацией. Способ выполняют на ранних стадиях у нормально развивающихся эмбрионов (например, на стадии восьми клеток), где можно изучить генетический вклад обоих родителей, путем создания отверстия в вителлиновом слое и аспирации или экструзии или диссекции одного или двух бластомеров через отверстие. Генетический анализ с применением флюоцентной гибридизации in situ (FISH) и полимеразной цепной реакции (ПЦР) обычно применяют в ПГД для анализа хромосомных аномалий и моногенных нарушений, соответственно. Затем, если нет генетических аномалий, оставшийся интактным эмбрион имплантируют женщине-пациентке на нормальный период беременности. Методики преимплантационной генетической диагностики были описаны уже в 2004 различными группами, включая Staessen, С. и др. (2004), «Comparison of blastocyst transfer with or without preimplantation genetic diagnosis for aneuploidy screening in couples with advanced maternal age: a prospective randomized controlled trial», Hum. Reprod. 19: 2849-2858 («Сравнение переноса бластоцисты с проведением или без проведения преимплантационной генетической диагностики для скрининга анеуплоидии, связанной с большим возрастом матери: проспективное рандомизированное контролируемое испытание») и Monni и др. (2004) «Preimplantation genetic diagnosis for beta-thalassaemia: the Sardinian experience». Prenat Diagn 24: 949-954 («Преимплантационная генетическая диагностика бета-талассемии: Сардинский опыт»). Так, Chung и др. (2006) просто описывает способы, в которых применяется экстракция бластомеров от эмбрионов на ранних стадиях без деструкции эмбрионов»).
Далее, в августе 2006, в публикации от Klimanskaya и др., второго автора в Chung и др. (2006) и также от Advanced Cell Technology, описана процедура, которая хотя и не была эффективной (только 2% из выделенных бластомеров генерировали hES клеточную линию), показала, что подобные ПГД методики делают возможным получение человеческих ES клеточных линий из единственного бластомера; это незначительно отличалось от того, что было вначале описано Chung и др. (2006) выше. См. Klimanskanya и др. (2006) «Human embryonic stem cell lines derived from single blastomeres», Nature 444, 481-485 («Линии человеческих эмбриональных стволовых клеток, полученные из единственного бластомера»). Klimanskaya и др. совместно культивировали новые полученные hES клеточные линии с другими ES клетками, которые, как установил Chung и др. (2006), могут быть критическими. Chung и др. (2006) установил, что «неясно, действительно ли успех системы совместного культивирования ES в (его) исследовании связан с веществами, секретируемыми ES клетками, или необходим контакт между клетками». См. Chung и др. (2006) выше, стр. 218, правая колонка. Но позже в исследовании от 2008 того же самого Chung, и др., выше, было показано, что hES клеточные линии вовсе не требуют совместного культивирования с ES клетками, поскольку культивирование изолированных бластомеров в среде с ламинином повышало их способность давать начало hESC. См. Chung и др. (2008), «Human Embryonic Stem Cell Lines Generated without Embryo Destruction» («Линии человеческих эмбриональных стволовых клеток, генерированные без деструкции эмбриона»), Cell Stem Cell (2): 113-117, p. 116, опубликовано он-лайн 10 января 2008. Далее, hES клетки, полученные таким способом, имеют те же самые характеристики, как и другие человеческие плюрипотентные стволовые клетки, включая hES клетки, включая способность к сохранению недифференцированного состояния в течение более 6 месяцев, и демонстрируют нормальный кариотип и экспрессию маркеров плюрипотентности, включая Oct-4, SSEA-3, SSEA-4, TRA-1-60, TRA-1-81, Nanog и щелочную фосфатазу; и могут дифференцироваться и вызывать образование всех трех (3) эмбриональных зародышевых слоев in vitro и формирование тератом in vivo.
Таким образом, совместное культивирование отдельных бластомеров с другими ES клетками не является критическим, как вначале было постулировано Chung и др. (2006), и их культивируют на одном слое, который является общим компонентом многих экстрацеллюлярных матриксов (коммерческих или приготовленных, например, путем лизиса фидерных клеток) и обычно применяется и известен для выращивания клеток млекопитающих, и это было известно и доступно ко времени первой демонстрации получения мышиных ES клеточных линий из единственного бластомера из публикации Chung и др. (2006). Таким образом, вполне возможно, что рядовой специалист в данной области техники с учетом методологий Chung и др. (2006) выше и с применением знаний, доступных, как ранее описано Straessen и др. (2004) выше, может получить hES клеточную линию из единственного бластомера при сохранении эмбриона или предотвращении его деструкции.
Суспензия агрегатов плюрипотентных стволовых клеток, и клетки, полученные из плюрипотентной стволовой клетки
В отличие от ранее известных способов тканевой инженерии, которые основаны на высеве индивидуальных клеток на полимерные каркасы, матрицы и/или гели, варианты осуществления, описанные в настоящей заявке, могут применять суспензии агрегатов клеток, полученные из плюрипотентной стволовой клетки, суспензий отдельных клеток или суспензий дифференцированных отдельных клеток, полученных из них. Способы обработки и/или производства суспензий агрегатов стволовых клеток и дифференцировки клеток из них описаны в PCT/US2007/062755, поданной 23 февраля 2007, и озаглавленной «Композиции и способы для культивирования дифференцированных клеток», и теперь в патентах США №8,211,699 и №8,415,158; и в PCT/US2008/080516, поданной 20 октября 2008, и озаглавленной «Способы и композиции для бесфидерных сред для плюрипотентных стволовых клеток, содержащих человеческую сыворотку», и теперь патенте США №8,334,138, и в Schulz Т. и др. (2012) выше.
Варианты осуществления, описанные в настоящей заявке, относятся к способам генерации агрегата плюрипотентных клеток в суспензии из плюрипотентной адгезивной культуры, путем культивирования плюрипотентной клетки в условиях адгезивной ростовой культуры, обеспечивающих экспансию в недифференцированном состоянии; диссоциации адгезивной культуры плюрипотентных клеток в суспензионную культуру отдельных клеток; контактирования суспензионной культуры отдельных клеток при первых дифференцирующих условиях культивирования, обеспечивающих образование клеточных агрегатов из hES в суспензии путем перемешивания суспензионной культуры из отдельных клеток в течение такого периода времени, когда суспензионная культура из отдельных клеток образует в суспензии агрегаты клеток, полученные из плюрипотентных; и таким образом, генерации клеточных агрегатов в суспензии, полученных из плюрипотентных клеток. В предпочтительных вариантах осуществления перемешивание суспензионной культуры отдельных клеток выполняют путем вращения со скоростью 80-160 об./мин. В некоторых других вариантах осуществления, описанных в настоящей заявке, ингибитор rho-киназы применяют для облегчения агрегации, роста, пролиферации, экспансии и/или клеточной массы плюрипотентных стволовых клеток.
Выражение «по существу однородный» или «по существу однородный по размеру и форме», или их эквиваленты, относится к разбросу по однородности агрегатов, и составляет не более примерно 20%. В другом варианте осуществления разброс по однородности агрегатов составляет не более примерно 15%, 10% или 5%.
В еще одном варианте осуществления суспензии агрегатов hES клеток культивировали в среде, по существу свободной от сыворотки, и кроме того, не содержащей экзогенно добавленного фактора роста фибробластов (FGF). Это отличается от патента США №7,005,252 от Thomson, J., в котором было необходимо культивирование hES клеток в среде без сыворотки, но содержащей экзогенно добавленные ростовые факторы, включая FGF. В некоторых вариантах осуществления суспензии агрегатов iPS клеток культивировали в среде, по существу свободной от сыворотки и/или дополнительно при отсутствии экзогенно добавленного фактора роста фибробластов (FGF).
Хотя точное число клеток на агрегат не является критическим, специалистам в данной области техники понятно, что размер каждого агрегата (и таким образом, число клеток на агрегат) ограничивается способностью кислорода и нутриентов к диффузии в центральные клетки, и что это число может также варьировать в зависимости от типа клеток и питательных потребностей этого типа клеток. Клеточные агрегаты могут содержать минимальное число клеток (например, две или три клетки) на агрегат, или могут содержать многие сотни или тысячи клеток на агрегат. Как правило, клеточные агрегаты содержать сотни или тысячи клеток на агрегат. Для целей настоящего изобретения клеточные агрегаты, как правило, имеют размер примерно от 50 микрон до 600 микрон, хотя, в зависимости от типа клеток, размер может быть меньше или больше этого диапазона. В одном варианте осуществления клеточные агрегаты имеют размер примерно от 50 микрон до 250 микрон, или примерно от 75 до 200 микрон, и предпочтительно, примерно от 100 до 150 микрон.
Другие способы описывают получение эмбриоидных телец (ЕВ). Использующиеся в настоящей заявке термины «эмбриоидные тельца», «агрегатные тельца» или их эквиваленты означают агрегаты из дифференцированных или недифференцированных клеток, которые появляются, когда ES клетки перерастают в однослойных культурах, или поддерживаются в суспензионных культурах в не-дифференцирующей среде, или дифференцируются с помощью ненаправленных протоколов в ткани из множества зародышевых листков. То есть, ЕВ не образуются из суспензии из отдельных клеток или плюрипотентных стволовых клеток, как описано в настоящей заявке, и не образуются из адгезивных культур или мультипотентных клеток, полученных из плюрипотентных. Эти характеристики по отдельности явно отличают настоящее изобретение от эмбриоидных телец.
В отличие от эмбриоидных телец, которые являются смесью дифференцированных и недифференцированных клеток, и как правило, состоят из клеток из нескольких зародышевых листков и проходят через произвольную дифференцировку, клеточные агрегаты, описанные в настоящей заявке, по существу или в основном являются гомо-клеточными, существуя в виде агрегатов из плюрипотентных, мультипотентных, бипотентных или унипотентных типов клеток, например, эмбриональных клеток, дефинитивной энтодермы, энтодермы передней части пищеварительного тракта, PDX1 позитивной панкреатической энтодермы, панкреатических эндокринных клеток и тому подобного.
Способы, описанные в настоящей заявке, никоим образом не требуют предварительного покрытия сосудов для культивирования экстрацеллюлярным матриксом, например, как описано в патенте США №6,800,480, Bodnar и др., принадлежащем Geron Corporation. В некоторых вариантах осуществления, описанных в настоящей заявке, iPS клетки можно культивировать таким же способом, как и другие плюрипотентные стволовые клетки, например, hES и iPS клетки, которые культивируют с применением растворенной человеческой сыворотки, как подробно описано в патенте США от авторов настоящей заявки №8,334,138; и в Schulz Т. и др. (2012) выше.
Способы, описанные в настоящей заявке, никоим образом не требуют экзогенно добавленного фактора роста фибробластов (FGF) из иного источника, чем просто слой фидерных фибробластов, как описано в патенте США №7,005,252, Thomson, J., принадлежащем Wisconsin Alumni Research Foundation (WARF).
Композиции мультипотентных и дифференцированных клеток
Клеточные композиции, полученные посредством способов из настоящей заявки, включают клеточные культуры, содержащие плюрипотентные стволовые клетки, клетки препервичной полоски, мезоэнтодермы, дефинитивной энтодермы, энтодермы передней части пищеварительного тракта, PDX1-позитивной энтодермы передней части пищеварительного тракта, PDX1-позитивной панкреатической энтодермы или PDX1/NKX6.1-со-позитивной панкреатической энтодермы, эндокринного прогенитора/предшественника или NGN3/NKX2.2-co-позитивного эндокринного прогенитора/предшественника, и гормон-секретирующие эндокринные клетки или INS, GCG, GHRL, SST, PP одинарно-позитивные эндокринные клетки, где по меньшей мере примерно 5-90% клеток в культуре являются полученными клетками препервичной полоски, мезоэнтодермы, дефинитивной энтодермы, энтодермы передней части пищеварительного тракта, PDX1-позитивной энтодермы передней части пищеварительного тракта, PDX1-позитивной панкреатической энтодермы или PDX1/NKX6.1-со-позитивной панкреатической энтодермы, эндокринного прогенитора/предшественника или NGN3/NKX2.2-co-позитивного эндокринного прогенитора/предшественника, и гормон-секретирующими эндокринными клетками или INS, GCG, GHRL, SST, РР одинарно-позитивными эндокринными клетками.
Некоторые варианты осуществления, описанные в настоящей заявке, относятся к композициям, таким как клеточные популяции и клеточные культуры, содержащие как плюрипотентные клетки, так и стволовые клетки, такие как стволовые клетки и iPS клетки, и мультипотентные клетки, такие как клетки препервичной полоски, мезоэнтодермы или дефинитивной энтодермы, а также более дифференцированные, но все еще потенциально мультипотентные клетки, такие как клетки PDX1-позитивной энтодермы передней части пищеварительного тракта, PDX1-позитивной панкреатической энтодермы или PDX1/NKX6.1-со-позитивной панкреатической энтодермы, эндокринного прогенитора/предшественника или NGN3/NKX2.2-co-позитивного эндокринного прогенитора/предшественника, и гормон-секретирующие эндокринные клетки или INS, GCG, GHRL, SST, PP одинарно-позитивные эндокринные клетки. Например, с применением способов, описанных в настоящей заявке, можно получить композиции, содержащие смеси плюрипотентных стволовых клеток и других мультипотентных или дифференцированных клеток. В некоторых вариантах осуществления получают композиции, содержащие по меньшей мере примерно 5 мультипотентных или дифференцированных клеток на каждые 95 плюрипотентных клеток. В других вариантах осуществления получают композиции, содержащие по меньшей мере примерно 95 мультипотентных или дифференцированных клеток на каждые 5 плюрипотентных клеток. Дополнительно подразумеваются композиции, содержащие другие отношения мультипотентных или дифференцированных клеток к плюрипотентным клеткам. Например, подразумеваются композиции, содержащие по меньшей мере 1 мультипотентную или дифференцированную клетку примерно на каждые 1,000,000 плюрипотентных клеток, по меньшей мере 1 мультипотентную или дифференцированную клетку примерно на каждые 100,000 плюрипотентных клеток, по меньшей мере 1 мультипотентную или дифференцированную клетку примерно на каждые 10,000 плюрипотентных клеток, по меньшей мере 1 мультипотентную или дифференцированную клетку примерно на каждые 1000 плюрипотентных клеток, по меньшей мере 1 мультипотентную или дифференцированную клетку примерно на каждые 500 плюрипотентных клеток, по меньшей мере 1 мультипотентную или дифференцированную клетку примерно на каждые 100 плюрипотентных клеток, по меньшей мере 1 мультипотентную или дифференцированную клетку примерно на каждые 10 плюрипотентных клеток, по меньшей мере 1 мультипотентную или дифференцированную клетку примерно на каждые 5 плюрипотентных клеток, и примерно на каждую 1 плюрипотентную клетку, и по меньшей мере примерно 1,000,000 мультипотентных или дифференцированных клеток примерно на каждую 1 плюрипотентную клетку.
Некоторые варианты осуществления, описанные в настоящей заявке, относятся к клеточным культурам или клеточным популяциям, содержащим по меньшей мере примерно от 5% мультипотентных или дифференцированных клеток до 99% мультипотентных или дифференцированных клеток. В некоторых вариантах осуществления клеточные культуры или клеточные популяции содержат клетки млекопитающих. В предпочтительных вариантах осуществления клеточные культуры или клеточные популяции содержат человеческие клетки. Например, некоторые специфические варианты осуществления относятся к клеточным культурам, содержащим человеческие клетки, где по меньшей мере примерно от 5% до 99% от человеческих клеток являются мультипотентными или дифференцированными клетками. Другие варианты осуществления относятся к клеточным культурам, содержащим человеческие клетки, где по меньшей мере примерно 5%, по меньшей мере примерно 10%, по меньшей мере примерно 15%, по меньшей мере примерно 20%, по меньшей мере примерно 25%, по меньшей мере примерно 30%, по меньшей мере примерно 35%, по меньшей мере примерно 40%, по меньшей мере примерно 45%, по меньшей мере примерно 50%, по меньшей мере примерно 55%, по меньшей мере примерно 60%, по меньшей мере примерно 65%, по меньшей мере примерно 70%, по меньшей мере примерно 75%, по меньшей мере примерно 80%, по меньшей мере примерно 85%, по меньшей мере примерно 90%, по меньшей мере примерно 95%, по меньшей мере примерно 98%, по меньшей мере примерно 99%, или больше 99% человеческих клеток являются мультипотентными или дифференцированными клетками. В вариантах осуществления, где клеточные культуры или клеточные популяции содержат человеческие фидерные клетки, вышеуказанные проценты рассчитывают по отношению к человеческим фидерным клеткам в клеточных культурах.
Композиции и способы, описанные в настоящей заявке, имеют некоторые полезные характеристики. Например, клеточные культуры и клеточные популяции, включающие мультипотентные клетки, например, клетки препервичной полоски и/или клетки мезоэнтодермы, а также способы получения клеточных культур и клеточных популяций, пригодны для моделирования ранних этапов человеческого развития. Далее, композиции и способы, описанные в настоящей заявке, можно также использовать для терапевтического вмешательства в патологические состояния, такие как сахарный диабет. Например, поскольку клетки препервичной полоски и/или мезоэнтодермальные клетки служат в качестве источника только ограниченного числа тканей, они могут служить для получения чистых тканей или клеточных типов. В некоторых способов получения клеток препервичной полоски плюрипотентные клетки, используемые в качестве исходного материала, являются плюрипотентными стволовыми клетками, например, hES, hEG или iPS клетками.
Трофоэктодермальные клетки
С помощью способов, описанных в настоящей заявке, можно получить композиции, содержащие трофоэктодермальные клетки, по существу свободные от других типов клеток. В некоторых вариантах осуществления, описанных в настоящей заявке, трофоэктодермальные клеточные популяции или клеточные культуры, полученные способами, описанными в настоящей заявке, по существу имеют высокую экспрессию маркеров, выбранных из группы, состоящей из HAND1, Eomes, MASH2, ESXL1, HCG, KRT18, PSG3, SFXN5, DLX3, PSX1, ETS2 и ERRB генов, по сравнению с уровнями экспрессии HAND1, Eomes, MASH2, ESXL1, HCG, KRT18, PSG3, SFXN5, DLX3, PSX1, ETS2 и ERRB в не-трофоэктодермальных клетках или клеточных популяциях.
Клетки препервичной полоски
С применением способов, описанных в настоящей заявке, можно получить композиции, содержащие клетки препервичной полоски, по существу свободные от других типов клеток. В некоторых вариантах осуществления, описанных в настоящей заявке, популяции или культуры клеток препервичной полоски, полученные способами, описанным в настоящей заявке, по существу экспрессируют FGF8 и/или NODAL маркерные гены, по сравнению с BRACHURYlow, FGF4 low, SNAI1 low, SOX17 low, FOXA2 low, SOX7 low и SOX1 low. Клетки препервичной полоски и способы получения клеток препервичной полоски описаны в патенте США от автора настоящей заявки №7,958,585, озаглавленном «Клетки препервичной полоски и мезоэнтодермы», изданном 26 июля 2011.
Экстраэмбриональные клетки
С применением способов, описанных в настоящей заявке, можно получить композиции, содержащие экстраэмбриональные клетки, по существу свободные от других типов клеток. Клетки примитивной, висцеральной и париетальной энтодермы являются экстраэмбриональными клетками. Клетки примитивной энтодермы дают начало клеткам висцеральной и париетальной энтодермы. Клетки висцеральной энтодермы являются эндодермальными клетками, которые образуют часть желточного мешка. Функцией клеток висцеральной энтодермы является захват и транспорт нутриентов. Клетки париетальной энтодермы соприкасаются с экстраэмбриональной тканью, известной как мембрана Рейхерта. Одной из ролей клеток париетальной энтодермы является продукция компонентов базальной мембраны. Вместе клетки висцеральной энтодермы и клетки париетальной энтодермы образуют то, что часто обозначают как экстраэмбриональная энтодерма. Как предполагает наименование, клетки экстраэмбриональной энтодермы не дают начало эмбриональным структурами, образуемым при развитии. Напротив, клетки дефинитивной энтодермы и другие клетки эндодермальных линий или панкреатических линий, описанные в настоящей заявке, являются эмбриональными или полученными из эмбриональных клеток, и дают начало тканям, которые получены из кишечной трубки, образуемой при развитии эмбриона. В некоторых вариантах осуществления, описанных в настоящей заявке, популяции или культуры экстраэмбриональных клеток, полученных способами, описанными в настоящей заявке, по существу имеют высокую экспрессию маркеров, выбранных из группы, состоящей из генов SOX7, SOX17, THBD, SPARC, DAB1, HNF4alpha или AFP, по сравнению с уровнями экспрессии по меньшей мере SOX7, SOX17, THBD, SPARC, DAB1 или AFP, которые на экспрессируются в других типах клеток или клеточных популяций, например, дефинитивной энтодерме.
Мезоэндодермальные клетки
С применением способов, описанных в настоящей заявке, можно получить композиции, содержащие мезоэндодермальные клетки, по существу свободные от других типов клеток. В некоторых вариантах осуществления, описанных в настоящей заявке, популяции или культуры мезоэндодермальных клеток, полученные способами, описанным в настоящей заявке, по существу имеют высокую экспрессию маркеров, выбранных из группы, состоящей из FGF4, SNAI1 MIXL1 и/или WNT3 маркерных генов, по сравнению с SOX17 low, CXCR4 low, FOXA2 low, SOX7 low и SOX1 low. Мезоэндодермальные клетки и способы получения мезоэндодермальных клеток подробно описаны в патенте США, принадлежащему автору настоящей заявки, №7,958,585, озаглавленном «Клетки препервичной полоски и мезоэнтодермы», изданном 26 июля 2011.
Способы скрининга
В некоторых вариантах осуществления применяются способы скрининга для получения определенных клеточных популяций, включающих плюрипотентные, мультипотентные и/или дифференцированные клетки, такие как человеческие плюрипотентные стволовые клетки, индуцированные плюрипотентные стволовые клетки, клетки препервичной полоски, мезоэндодермальные клетки, клетки дефинитивной энтодермы, клетки энтодермы передней части пищеварительного тракта или PDX1-негативные клетки энтодермы передней части пищеварительного тракта, PDX1-позитивные клетки энтодермы передней части пищеварительного тракта или PDX1-позитивные панкреатические эндодермальные клетки или панкреатические прогениторные клетки, эндокринные клетки-предшественники, и/или эндокринные клетки. Популяция клеток затем обеспечивается потенциальным фактором дифференцировки. В первый момент времени, который отмечается до или примерно в то же время, что и обеспечение потенциального фактора дифференцировки, определяют экспрессию маркера. Альтернативно, экспрессию маркера можно определить после обеспечения потенциального фактора дифференцировки. Во второй момент времени, который отмечается после первого момента времени и после этапа обеспечения потенциального фактора дифференцировки клеточной популяции, вновь определяют экспрессию того же самого маркера. Определяют, способен ли потенциальный фактор дифференцировки к стимуляции дифференцировки панкреатических клеток-предшественников, путем сравнения экспрессии маркера в первый момент времени с экспрессией маркера во второй момент времени. Если экспрессия маркера во второй момент времени повышается или снижается, по сравнению с экспрессией маркера в первый момент времени, то потенциальный фактор дифференцировки способен к стимуляции дифференцировки панкреатических прогениторных клеток.
Некоторые варианты осуществления способов скрининга, описанных в настоящей заявке, используют популяции или культуры клеток, содержащие клетки человеческой дефинитивной энтодермы, PDX-1 негативные клетки энтодермы передней части пищеварительного тракта, PDX-1 позитивные клетки энтодермы передней части пищеварительного тракта, PDX-1 позитивные панкреатические эндодермальные клетки, или панкреатические прогениторные клетки или эндокринные прогениторы/клетки-предшественники. Например, клеточная популяция может быть по существу очищенной популяцией PDX-1 позитивных панкреатических эндодермальных клеток или панкреатических прогениторных клеток. Например, клеточная популяция может быть обогащенной популяцией человеческих панкреатических прогениторных клеток, где по меньшей мере 50%-97% человеческих клеток в клеточной популяции являются человеческими панкреатическими прогениторными клетками, остаток включает эндокринные прогениторы/предшественники или эндокринные клетки и другие типы клеток. Обогащение популяций панкреатических прогениторов подробно описано в принадлежащих автору настоящей заявки патентной заявке США №12/107,020, озаглавленной «Способы очистки энтодермы и панкреатических эндодермальных клеток, полученных из человеческих эмбриональных стволовых клеток», поданной 21 апреля 2008, теперь патенте США №8,338,170, и соответствующей публикации Kelly и др. (2011) выше.
В вариантах осуществления способов скрининга, описанных в настоящей заявке, клеточная популяция контактирует или иным образом обеспечена потенциальным (тестируемым) фактором дифференцировки. Потенциальный фактор дифференцировки может содержать любую молекулу, которая может обладать потенциалом стимуляции дифференцировки любой из вышеупомянутых клеток, например, человеческих панкреатических прогениторных клеток. В некоторых вариантах осуществления, описанных в настоящей заявке, потенциальный фактор дифференцировки включает молекулу, которая известна как фактор дифференцировки для одного или нескольких типов клеток. В альтернативных вариантах осуществления потенциальный фактор дифференцировки включает молекулу, которая неизвестна как стимулятор клеточной дифференцировки. В предпочтительных вариантах осуществления потенциальный фактор дифференцировки включает молекулу, которая неизвестна как стимулятор дифференцировки человеческих панкреатических прогениторных клеток. Скрининг факторов, дифференцирующих дефинитивную энтодерму, например, подробно описан в принадлежащей автору настоящей заявки патентной заявке США №12/093,590, озаглавленной «Маркеры дефинитивной энтодермы», поданной 21 июля 2008.
В дополнение к определению экспрессии по меньшей мере одного маркера в первый момент времени, некоторые варианты осуществления способов скрининга, обеспеченные в настоящей заявке, подразумевают определение экспрессии по меньшей мере одного маркера во второй момент времени, после второго момента времени, и который отмечается после обеспечения клеточной популяции потенциальным фактором дифференцировки. В таких вариантах осуществления экспрессию того же самого маркера определяют и в первый, и во второй момент времени. В некоторых вариантах осуществления экспрессию множества маркеров определяют и в первый, и во второй момент времени. В таких вариантах осуществления экспрессию одного и того же множества маркеров и в первый, и во второй момент времени. В некоторых вариантах осуществления экспрессию маркеров определяют во множестве моментов времени, каждый из которых отмечается после первого момента времени, и каждый из которых отмечается после обеспечения клеточной популяции потенциальным фактором дифференцировки. В некоторых вариантах осуществления экспрессию маркеров определяют путем Q-PCR. В других вариантах осуществления экспрессию маркеров определяют с помощью иммуноцитохимии.
В некоторых вариантах осуществления способов скрининга, описанных в настоящей заявке, маркер, чью экспрессию определяют в первый и второй моменты времени, является маркером, связанным с дифференцировкой панкреатических прогениторных клеток в клетки, являющиеся предшественниками окончательно дифференцированных клеток, которые образуют ткани панкреатических островков. Такие клетки могут включать незрелые гормон-экспрессирующие клетки панкреатических островков.
В некоторых вариантах осуществления способов скрининга, обеспеченных в настоящей заявке, обеспечивается достаточное время между обеспечением клеточной популяции потенциальным фактором дифференцировки и определением экспрессии маркера во второй момент времени. Достаточное время между обеспечением клеточной популяции потенциальным фактором дифференцировки и определением экспрессии маркера во второй момент времени может составлять от всего лишь примерно 1 час вплоть до примерно 10 дней. В некоторых вариантах осуществления экспрессию по меньшей мере одного маркера определяют много раз после обеспечения клеточной популяции потенциальным фактором дифференцировки. В некоторых вариантах осуществления достаточное время составляет по меньшей мере примерно 1 час, по меньшей мере примерно 6 часов, по меньшей мере примерно 12 часов, по меньшей мере примерно 16 часов, по меньшей мере примерно 1 сутки, по меньшей мере примерно 2 суток, по меньшей мере примерно 3 суток, по меньшей мере примерно 4 суток, по меньшей мере примерно 5 суток, по меньшей мере примерно 6 суток, по меньшей мере примерно 7 суток, по меньшей мере примерно 8 суток, по меньшей мере примерно 9 суток, по меньшей мере примерно 10 суток, по меньшей мере примерно 11 суток, по меньшей мере примерно 12 суток, по меньшей мере примерно 13 суток, по меньшей мере примерно 14 суток, по меньшей мере примерно 1 неделю, по меньшей мере примерно 2 недели, по меньшей мере примерно 3 недели, по меньшей мере примерно 4 недели, по меньшей мере примерно 5 недель, по меньшей мере примерно 6 недель, по меньшей мере примерно 7 недель, или по меньшей мере примерно 8 недель.
В некоторых вариантах осуществления способов, описанных в настоящей заявке, дополнительно определяют, повышается или снижается ли экспрессия маркера во второй момент времени по сравнению с экспрессией этого маркера в первый момент времени. Повышение или снижение экспрессии по меньшей мере одного маркера указывает, что потенциальный фактор дифференцировки способен к стимуляции дифференцировки эндокринных прогениторов/клеток-предшественников. Подобным образом, если определяют экспрессию множества маркеров, дополнительно определяют, повышается или снижается ли экспрессия множества маркеров во второй момент времени по сравнению с экспрессией этого множества маркеров в первый момент времени. Повышение или снижение множества маркеров можно определить путем измерения или иной оценки количества, уровня или активности маркера в клеточной популяции в первый и второй моменты времени. Такое определение может быть относительным по другим маркерам, например, экспрессии конститутивных генов, или абсолютным. В некоторых вариантах осуществления, где экспрессия маркера повышается во второй момент времени, по сравнению с первым моментом времени, повышение является по меньшей мере примерно 2-кратным, по меньшей мере примерно 5-кратным, по меньшей мере примерно 10-кратным, по меньшей мере примерно 20-кратным, по меньшей мере примерно 30-кратным, по меньшей мере примерно 40-кратным, по меньшей мере примерно 50-кратным, по меньшей мере примерно 60-кратным, по меньшей мере примерно 70-кратным, по меньшей мере примерно 80-кратным, по меньшей мере примерно 90-кратным, по меньшей мере примерно 100-кратным, или более чем по меньшей мере примерно 100-кратным. В некоторых вариантах осуществления повышение является менее чем 2-кратным. В вариантах осуществления, где экспрессия маркеров снижается во второй момент времени, по сравнению с первым моментом времени, снижение является по меньшей мере примерно 2-кратным, по меньшей мере примерно 5-кратным, по меньшей мере примерно 10-кратным, по меньшей мере примерно 20-кратным, по меньшей мере примерно 30-кратным, по меньшей мере примерно 40-кратным, по меньшей мере примерно 50-кратным, по меньшей мере примерно 60-кратным, по меньшей мере примерно 70-кратным, по меньшей мере примерно 80-кратным, по меньшей мере примерно 90-кратным, по меньшей мере примерно 100-кратным, или более чем по меньшей мере примерно 100-кратным. В некоторых вариантах осуществления снижение является менее чем 2-кратным.
Мониторинг продукции мультипотентных или дифференцированных клеток
Развитие плюрипотентных клеток в мультипотентные клетки, другие мультипотентные клетки или дифференцированные клетки, такие как панкреатические прогениторы или эндокринные гормон-секретирующие клетки, можно контролировать путем определения экспрессии маркеров, характерных для специфических клеток, включая генетические маркеры и фенотипические маркеры, такой как экспрессия островковых гормонов и процессинг проинсулина в инсулин и С-пептид в эндокринных клетках. В некоторых способах экспрессию некоторых маркеров определяют путем детекции присутствия и отсутствия маркера. Альтернативно, экспрессию некоторых маркеров можно определить путем измерения уровня, в котором маркер присутствует в клетках или клеточной культуре или клеточной популяции. Например, в некоторых способах определяют экспрессию маркеров, характерных для незрелых гормон-экспрессирующих клеток панкреатических островков, а также отсутствие существенной экспрессии маркеров, характерных для плюрипотентных клеток, клеток дефинитивной энтодермы, энтодермы передней части пищеварительного тракта, PDX1-позитивной энтодермы передней части пищеварительного тракта, эндокринных прогениторов/предшественников, клеток экстраэмбриональной энтодермы, мезодермы, эктодермы, зрелых гормон-экспрессирующих клеток панкреатических островков и/или других типов клеток.
Как описано в связи с мониторингом продукции других менее дифференцированных типов клеток линии дефинитивной энтодермы, можно применять качественные или полуколичественные методики, такие как способы блоттинга и иммунохимии, для измерения экспрессии маркера. Альтернативно, можно провести точное количественное определение экспрессии маркера с помощью такой методики, как Q-PCR или nCounter от Nanostring, или родственных мультиплексных технологий с высокой пропускной способностью, позволяющих анализировать и определять сотни или больше маркеров одновременно. Кроме того, необходимо понять, что на полипептидном уровне многие из маркеров гормон-экспрессирующих клеток панкреатических островков являются секретируемыми белками. Таким образом, можно применять методики для измерения содержания экстрацеллюлярного маркера, такие как ИФА.
В других вариантах осуществления иммуноцитохимию применяют для детекции белков, экспрессируемых вышеупомянутыми генами. В других вариантах осуществления Q-PCR может применяться в сочетании с иммуногистохимическими методиками или методиками проточной цитометрии для эффективной и точной характеристики и идентификации количества и относительных пропорций таких маркеров в исследуемом типе клеток. В одном варианте осуществления Q-PCR позволяет количественно определить уровень экспрессии РНК в клеточной культуре, содержащей смешанную популяцию клеток. Однако Q-PCR не позволяет предположить или оценить, имеется ли совместная экспрессия маркеров или белков в одной и той же клетке. В другом варианте осуществления Q-PCR применяют в сочетании со способами проточной цитометрии для характеристики и идентификации типов клеток. Таким образом, с применением комбинации способов, описанных в настоящей заявке, и таких как те, которые описаны выше, можно осуществить и продемонстрировать полную характеристику и идентификацию различных типов клеток, включая типы клеток эндодермальной линии.
Например, в одном предпочтительном варианте осуществления панкреатические прогениторы или клетки панкреатической энтодермы или PDX-1-позитивные клетки панкреатической энтодермы, экспрессирующие по меньшей мере PDX1, Nkx6.1, PTF1A, CPA и/или cMYC, как показано Q-PCR и/или ICC, по меньшей мере совместно экспрессирующие PDX1 и Nkx6.1 как показано ICC, и не экспрессирующие другие маркеры, включая SOX17, CXCR4 или CER, идентифицируются как PDX1-позитивные экспрессирующие клетки. Подобным образом, для надлежащей идентификации зрелых гормон-секретирующих панкреатических клеток, in vitro или in vivo, например, показано, что С-пептид (продукт надлежащего процессинга проинсулина в зрелой и функционирующей β клетке) и инсулин совместно экспрессируются, посредством ICC, в инсулин-секретирующей клетке.
Другие способы, известные в данной области техники, можно также применять для количественного определения экспрессии маркерного гена. Например, можно выявить экспрессию продукта маркерного гена с применением антител, специфичных для интересующего продукта маркерного гена (например, посредством вестерн-блоттинга, проточной цитометрии, и тому подобного). В некоторых способах определяют экспрессию маркерных генов, характерных для клеток, полученных из hES, а также отсутствие существенной экспрессии маркерных генов, характерных для клеток, полученных из hES. Другие способы характеристики и идентификации типов клеток, полученных из hES, описаны в родственных заявках, как указано выше.
Обобщение продукции PDX1-позитивной панкреатической энтодермы (Стадии 1-4) и продукции инсулина in vivo
Способы продукции определенных клеток эндодермальной линии и панкреатической эндодермальной линии обеспечены в настоящей заявке, и обсуждаются далее в других родственных заявках, таких как патенты США №№7,534,608; 7,695,965; и 7,993,920, озаглавленные «Способы получения панкреатических гормонов»; в патенте США №8,129,182, озаглавленном «Эндокринные прогениторы/клетки-предшественники, панкреатические гормон-экспрессирующие клетки и способы получения». См. также Таблицу 17 и Фигуры 42, 43 и 44.
Вкратце, способы направленной дифференцировки для плюрипотентных стволовых клеток, например, hES и iPS, могут быть описаны по меньшей мере четырьмя или пятью, или шестью или семью стадиями, в зависимости от необходимой культуры клеток в терминальной стадии (ПЭК или эндокринных клеток). Стадия 1 является продукцией дефинитивной энтодермы из плюрипотентных стволовых клеток, и занимает от 2 до 5 суток, предпочтительно от 2 до 3 суток. Плюрипотентные стволовые клетки суспендируют в средах, содержащих RPMI, фактор роста, являющийся членом суперсемейства TGFβ, такой как активин А, активин В, GDF-8 или GDF-11 (100 нг/мл), член семейства Wnt или активатор Wnt пути, такой Wnt3a (25 нг/мл), и альтернативно, ингибитор rho-киназы или ROCK, такой как Y-27632 (10 мкМ) для усиления роста и/или выживания и/или пролиферации и/или адгезии клеток. Спустя примерно 24 часа среды заменяют на среды, содержащие RPMI с сывороткой, такой как 0,2% FBS, и фактор роста, являющийся членом суперсемейства TGFβ, такой как активин А, активин В, GDF-8 или GDF-11 (100 нг/мл), и альтернативно, ингибитор rho-киназы или ROCK еще на 24 (день 1) или 48 часов (день 2). Альтернативно, спустя примерно 24 часа в среде, содержащей активин/ Wnt3a, клетки культивируют еще 24 часа в среде, содержащей один активин (т.е. в среде, не содержащей Wnt3a). Важно, что продукция дефинитивной энтодермы требует условий культивирования клеток с низким содержанием сыворотки, и таким образом, с низким содержанием инсулина или инсулиноподобного фактора роста. См. McLean и др. (2007) Stem Cells 25: 29-38. McLean и др. также показали, что обеспечение контакта hES клеток с инсулином в концентрациях всего 0,2 мкг/мл на Стадии 1 может быть вредным для продукции дефинитивной энтодермы. Другие специалисты в данной области техники модифицировали Стадию 1 дифференцировки плюрипотентных клеток в дефинитивную энтодерму по существу так, как описано в настоящей заявке и в D'Amour и др. (2005), например, по меньшей мере, в Agarwal и др., «Efficient Differentiation of Functional Hepatocytes from Human Embryonic Stem Cells», Stem Cells (2008) 26:1117-1127 («Эффективная дифференцировка функциональных гепатоцитов из человеческих эмбриональных стволовых клеток»); Borowiak и др., «Small Molecules Efficiently Direct Endodermal Differentiation of Mouse and Human Embryonic Stem Cells», (2009) Cell Stem Cell 4:348-358 («Малые молекулы эффективно направляют эндодермальную дифференцировку мышиных и человеческих эмбриональных стволовых клеток»); и Brunner и др., «Distinct DNA methylation patterns characterize differentiated human embryonic stem cells and developing human fetal liver», (2009) Genome Res. 19:1044-1056 («Отличающийся характер метилирования ДНК характеризует дифференцированные человеческие эмбриональные стволовые клетки и развивающуюся человеческую фетальную печень»). Надлежащая дифференцировка, спецификация, характеристика и идентификация определителя необходимы для получения других клеток эндодермальной линии. Клетки дефинитивной энтодермы на этой стадии совместно экспрессируют SOX17 и HNF3β (FOXA2), и не экспрессируют ощутимо по меньшей мере HNF4alpha, HNF6, PDX1, SOX6, PROX1, PTF1A, CPA, cMYC, NKX6.1, NGN3, РАХ3, ARX, NKX2.2, INS, GSC, GHRL, SST или PP. Отсутствие экспрессии HNF4α в дефинитивной энтодерме поддерживается и подробно описывается по меньшей мере в Duncan и др. (1994), «Expression of transcription factor HNF-4 in the extraembryonic endoderm, gut, and nephrogenic tissue of the developing mouse embryo: HNF-4 is a marker for primary endoderm in the implanting blastocyst, Proc. Natl. Acad. Sci, 91:7598-7602 («Экспрессия фактора транскрипции HNF-4 в экстраэмбриональной энтодерме, кишечнике и нефрогенной ткани развивающегося мышиного эмбриона: HNF-4 является маркером первичной энтодермы и имплантированной бластоцисте») и Si-Tayeb и др. (2010), «Highly Efficient Generation of Human Hepatocyte-Like cells from Induced Pluripotent Stem Cells», Hepatology 51:297-305 («Высокоэффективная генерация человеческих гепатоцит-подобных клеток из индуцированных плюрипотентных стволовых клеток»).
Стадия 2 принимает культуру клеток дефинитивной энтодермы со стадии 1 и продуцирует клетки энтодермы передней части пищеварительного тракта или PDX1-негативной энтодермы передней части пищеварительного тракта путем инкубации суспензионных культур с RPMI с низкими уровнями сыворотки, такими как 0,2% FBS, в разведении ITS 1:1000, 25 нг KGF (или FGF7), и альтернативно ROCK ингибитором в течение 24 часов (день 2 - день 3). Спустя 24 часа (день 3 - день 4), среду заменяют на ту же самую среду минус ингибитор TGFβ, но альтернативно с ROCK ингибитором для усиления роста, выживания и пролиферации клеток, еще на 24 часа (день 4 - день 5) или 48 часов (день 6). Критическим этапом для надлежащей спецификации энтодермы передней части пищеварительного тракта является удаление факторов роста семейства TGFβ. Таким образом, ингибитор TGFβ можно добавить на Стадии 2 клеточных культур, такой как 2,5 мкМ ингибитор TGFβ №4, или 5 мкМ SB431542, специфический ингибитор киназы, подобной рецептору активина (ALK), который является рецептором TGFβ I типа. Клетки энтодермы передней части пищеварительного тракта или PDX1-негативной энтодермы передней части пищеварительного тракта, полученные со Стадии 2, совместно экспрессируют SOX17, HNF1β и HNF4alpha и не проявляют выраженной совместной экспрессии по меньшей мере ни SOX17 и HNF3β (FOXA2), ни HNF6, PDX1, SOX6, PROX1, PTF1A, CPA, cMYC, NKX6.1, NGN3, РАХ3, ARX, NKX2.2, INS, GSC, GHRL, SST, или PP, которые являются отличительной чертой дефинитивной энтодермы, PDX1-позитивной панкреатической энтодермы или панкреатических прогениторных клеток или эндокринных прогениторов/предшественников, а также типичных клеток полигормонального типа.
Стадия 3 (дни 5-8) получения ПЭК принимает культуру клеток энтодермы передней части пищеварительного тракта со Стадии 2 и продуцирует клетки PDX1-позитивной энтодермы передней части пищеварительного тракта с помощью DMEM или RPMI в 1% В27, 0,25 мкМ KAAD циклопамина, ретиноида, такого как 0,2 мкМ ретиноевой кислоты (RA) или аналога ретиноевой кислоты, такого как 3 нМ TTNPB (или СТТ3, который является комбинацией KAAD циклопамина и TTNPB), и 50 нг/мл Noggin в течение примерно 24 (день 7) - 48 часов (день 8). В частности, авторы заявки применяли DMEM с высоким содержанием глюкозы примерно с 2003, и все патентные и не-патентные источники, раскрытые к настоящему времени, применяли DMEM с высоким содержанием глюкозы, даже если не упоминалось «DMEM с высоким содержанием глюкозы» и тому подобное. Это отчасти обусловлено тем, что производители, такие как Gibco, не называют их среду DMEM как таковую, например DMEM (Кат.№11960) и Knockout DMEM (Кат. №10829). Это заслуживает внимания, поскольку на момент подачи настоящей заявки Gibco поставляет большую часть DMEM продуктов, но еще не указывает «высокое содержание глюкозы» в некоторых из своих DMEM продуктов, содержащих высокое количество глюкозы, например, Knockout DMEM (Кат. №10829-018). Таким образом, можно предположить, что в каждом случае, когда описывается DMEM, это означает DMEM с высоким содержанием глюкозы, и это понятно другим специалистам, проводящим исследования и разработки в данной области техники. Более подробное описание применения экзогенного высокого количества глюкозы приведено в Примере 21. Вновь, ROCK ингибитор или ингибитор rho-киназы, такой как Y-27632, можно применять для усиления роста, выживания, пролиферации и стимуляции клеточной адгезии. PDX1-позитивные клетки передней части пищеварительного тракта, полученные со Стадии 3, совместно экспрессируют PDX1 и HNF6, а также SOX9 и PROX, и не проявляют заметной экспрессии маркеров, характерных для клеток дефинитивной энтодермы или энтодермы передней части пищеварительного тракта (PDX1-негативной энтодермы передней части пищеварительного тракта) или PDX1-позитивной энтодермы передней части пищеварительного тракта, как описано на Стадиях 1 и 2.
Вышеуказанная стадия 3 способа является одной из четырех стадий получения ПЭК. Для получения эндокринных прогениторов/предшественников и эндокринных клеток, как подробно описано в Примерах 9-24 ниже, в дополнение к ноггину, KAAD-циклопамину и ретиноиду применяют активин, Wnt и херегулин, по отдельности и/или в комбинации, для супрессии NGN3 при сохранении хорошей массы клеточных агрегатов. См. Примеры 8, 9 и 10.
Стадия 4 (дни 8-14) получения ПЭК принимает среды от Стадии 3 и заменяет их на среды, содержащие DMEM с 1% о/о В27 добавки, плюс 50 нг/мл KGF и 50 нг/мл EGF и иногда также 50 нг/мл ноггина и ингибитора ROCK, и дополнительно включает активин по отдельности или в комбинации с херегулином. Эти новые способы дают начало панкреатическим прогениторным клеткам, совместно экспрессирующим по меньшей мере PDX1 и Nkx6.1, а также PTF1A. Эти клетки не проявляют существенной экспрессии маркеров, характерных для клеток дефинитивной энтодермы или энтодермы передней части пищеварительного тракта (PDX1-негативной энтодермы передней части пищеварительного тракта), как описано выше для Стадий 1, 2 и 3. См. Фиг. 44.
Альтернативно, клетки со Стадии 4 можно дополнительно дифференцировать на Стадии 5 для продукции эндокринных прогениторов/предшественников или прогениторных типов клеток и/или моно- или полигормональных панкреатических эндокринных типов клеток в среде, содержащей DMEM в 1% о/о В27 в течение примерно 1-6 суток (предпочтительно, примерно 2 суток, т.е. дней 13-15). Эндокринные прогениторы/предшественники, полученные со Стадии 5, совместно экспрессируют по меньшей мере CHGA, NGN3 и Nkx2.2, но не проявляют заметной экспрессии маркеров, характерных для клеток дефинитивной энтодермы или энтодермы передней части пищеварительного тракта (PDX1-негативной энтодермы передней части пищеварительного тракта) или PDX1-позитивной энтодермы передней части пищеварительного тракта или прогениторных клеток, как описано выше на Стадиях 1, 2, 3 и 4 для продукции ПЭК. См.Фиг.44.
Для продукции ПЭК PDX1-позитивную панкреатическую энтодерму, полученную со Стадии 4, загружают и полностью содержат в устройстве для макро-инкапсулирования и трансплантируют пациенту, и клетки PDX1-позитивной панкреатической энтодермы созревают в панкреатические гормон-секретирующие клетки, или панкреатические островки, например, инсулин-секретирующие клетки, in vivo (также обозначенные как «функционирующие in v/vo». Инкапсулирование клеток PDX1-позитивной панкреатической энтодермы и продукция инсулина in vivo подробно описаны в заявке США №12/618,659 («заявке '659»), озаглавленной «Инкапсулирование панкреатической клеточной линии, полученной из человеческих плюрипотентных стволовых клеток», поданной 13 ноября 2009. Заявка '659 испрашивает приоритет предварительной патентной заявки 61/114,857, озаглавленной «Инкапсулирование панкреатических прогениторов, полученных из HES клеток», поданной 14 ноября 2008; и предварительной патентной заявки США №61/121,084, озаглавленной «Инкапсулирование панкреатических эндодермальных клеток», поданной 9 декабря 2008, и теперь патентов США №№8,278,106 и 8,424,928. Способы, композиции и устройства, раскрытые в настоящей заявке, представляют предпочтительные варианты осуществления, и являются примерными, и не предназначены для ограничения объема изобретения. Изменения и другие виды применения очевидны для специалистов в данной области техники и охватываются сущностью изобретения и определены объемом описания. Соответственно, для специалиста в данной области техники понятно, что различные замены и модификации могут быть выполнены в изобретении, раскрытом в настоящей заявке, не выходя за рамки и сущность настоящего изобретения.
Например, активин А, член суперсемейства факторов роста TGFβ или сигнальных белков, используют для получения дефинитивной энтодермы из плюрипотентных клеток, например, hES клеток и iPS клеток, однако, можно применять другие члены суперсемейства TGFβ, например, GDF-8 и GDF-11, для получения дефинитивной энтодермы, такие как те, которые описаны в международной заявке PCT/US2008/065686, озаглавленной «Факторы роста для получения дефинитивной энтодермы», поданной 3 июня 2008.
В другом контексте, активин по отдельности или в комбинации с Wnt и херегулином может подавлять и ингибировать экспрессию NGN3 на высоких и низких уровнях; на низких уровнях по отдельности или в комбинации с херегулином, или альтернативно на высоких уровнях в комбинации с WNT и херегулином, можно поддерживать клеточную массу, и таким образом, выход. См. Примеры 8, 9 и 10.
Для получения ПЭК ретиноевую кислоту (RA) применяют для дифференцировки PDX1-негативных клеток энтодермы передней части пищеварительного тракта на Стадии 2 в PDX1-позитивные клетки энтодермы передней части пищеварительного тракта на Стадии 3. Однако, можно применять другие ретиноиды или аналоги ретиноевой кислоты, такие как 4-[(Е)-2-(5,6,7,8-тетрагидро-5,5,8,8-тетраметил-2-нафталенил)-1-пропенил]бензойная кислота (или TTNPB) и подобные аналоги (например, 4-HBTTNPB). Для получения эндокринных прогениторов/клеток-предшественников ретиноевую кислоту или любые ее аналоги можно добавлять во время стадий 6 и/или 7 для индукции экспрессии генов гормонов. См. Примеры 13 и 16.
Ноггин (noggin) является белком, например, который инактивирует сигнальные белки - члены суперсемейства TGFβ, такие как костный морфогенетический белок-4 (ВМР4). Однако другие BMP; ингибиторы, такие как хордин и белок Twisted Gastrulation (скрученной гаструляции) (Tsg) или анти-ВМР нейтрализующие антитела могут предотвратить связывание BMP с рецепторами клеточной поверхности, таким образом, эффективно ингибируя передачу сигнала BMP. Альтернативно, ген человеческого ноггина был клонирован и секвенирован. См. патент США №6,075,007. Анализ последовательности ноггина показал карбокси-концевой участок, имеющий гомологию с ингибитором протеаз типа Куница, что указывает на возможность наличия у ингибиторов протеаз типа Куница схожего эффекта в ингибировании BMP. В Примере 15 описано применение ноггина для повышения продукции не-эндокринных (CHGA-) субпопуляций.
Наконец, устройства для макро-инкасулирования, описанные в настоящей заявке и в заявке США №12/618,659, заявках на патент США №№29/447,944; 29/408,366; 29/408,368; 29/423,365; и 61/774,443, озаглавленной «Полупроницаемые макро-имплантируемые устройства для инкапсулирования клеток», поданной 7 марта 2013, являются лишь примерными, и не предназначены для ограничения объема настоящего изобретения. В частности, изменения конструкции устройства, такие как изменения размера устройства, множества камер или субкомпартментов в устройстве, или множестве портов, или механизмах загрузки и извлечения устройства, охвачены сущностью настоящего изобретения. Таким образом, для специалиста в данной области техники очевидно, что различные замены и модификации не только описанных способов дифференцировки, но и инкапсулирующего устройства, также могут быть выполнены в раскрытом изобретении, не выходя за рамки и сущность настоящего изобретения. См. Примеры 10, 12 и 18.
Что касается некоторых способов дифференцировки плюрипотентных клеток в клетки дефинитивной энтодермы, вышеупомянутые факторы роста обеспечивают для клеток так, чтобы факторы роста присутствовали в культурах в концентрациях, достаточных для индукции дифференцировки по меньшей мере части плюрипотентных клеток в клетки дефинитивной энтодермы и клетки панкреатической линии. В некоторых способах вышеупомянутые факторы роста присутствуют в клеточной культуре в концентрации по меньшей мере примерно 5 нг/мл, по меньшей мере примерно 10 нг/мл, по меньшей мере примерно 25 нг/мл, по меньшей мере примерно 50 нг/мл, по меньшей мере примерно 75 нг/мл, по меньшей мере примерно 100 нг/мл, по меньшей мере примерно 200 нг/мл, по меньшей мере примерно 300 нг/мл, по меньшей мере примерно 400 нг/мл, по меньшей мере примерно 500 нг/мл, по меньшей мере примерно 1000 нг/мл, по меньшей мере примерно 2000 нг/мл, по меньшей мере примерно 3000 нг/мл по меньшей мере примерно 4000 нг/мл, по меньшей мере примерно 5000 нг/мл или больше примерно 5000 нг/мл. Другие агенты или факторы роста присутствуют в клеточных культурах в концентрации по меньшей мере 0,1 мМ или 10 мМ, или больше.
В некоторых способах агенты для дифференцировки плюрипотентных стволовых клеток и/или факторы роста удаляют из клеточной культуры после их добавления. Например, факторы роста можно удалить в течение примерно одних суток, примерно двух суток, примерно трех суток, примерно четырех суток, примерно пяти суток, примерно шести суток, примерно семи суток, примерно восьми суток, примерно девяти суток или примерно десяти суток после их добавления. В предпочтительных способах факторы роста удаляют спустя примерно 4 суток после их добавления. Удаление агента и/или фактора роста может сопровождаться заменой среды при отсутствии агента и/или фактора роста, или применением другого агента, который ингибирует функцию этого агента и/или фактора роста.
В одном предпочтительном варианте осуществления культуры клеток на стадии 3 включают агенты, способные к подавлению, супрессии, ингибированию клеток, коммитированных в эндокринные линии (CHGA+), и тому подобному, но не ограничиваются ими. Такие агенты также включают активин, херегулин и WNT, и комбинации этих трех агентов в количествах, эффективных для стимуляции дифференцировки и/или индукции экспрессии маркеров, показательных для субпопуляции не-эндокринных мультипотентных панкреатических прогениторов (CHGA-), для подавления или минимизации экспрессии маркеров клеток, коммитированных в эндокринную линию (CHGA+). Маркеры, показательные для клеток, коммитированных в эндокринную линию (CHGA+), включают NGN3, NKX2.2 и другие, но не ограничиваются ими.
В другом предпочтительном варианте осуществления культуры клеток на стадии 4 включают агенты, способные к подавлению, супрессии, ингибированию, клеток, коммитированных в эндокринные линии (CHGA+), и тому подобному, но не ограничиваются ими. Такие агенты также включают активин и херегулин, и их комбинации, в количествах, эффективных для стимуляции дифференцировки и/или индукции экспрессии маркеров, показательных для субпопуляции не-эндокринных мультипотентных панкреатических прогениторов (CHGA-), и для подавления или минимизации экспрессии маркеров клеток, коммитированных в эндокринную линию (CHGA+). Маркеры, показательные для клеток, коммитированных в эндокринную линию (CHGA+), включают NGN3, NKX2.2 и другие, но не ограничиваются ими. Средства супрессии по меньшей мере NGN3 во время стадий 3 и 4 описаны подробно в Примерах 8-21 ниже.
В одном варианте осуществления культуры клеток на стадии 4 дополнительно дифференцируются на стадии 5 по меньшей мере с ноггином, KGF, EGF, и ингибитором передачи сигналов или пути Notch. Маркеры, показательные для эндокринной дифференцировки, включают NGN3, NKX2.2 и другие, но не ограничиваются ими. В предпочтительном варианте осуществления на стадиях 1-7 применяют агенты, которые симулируют или эффективно имитируют in vitro то, что наблюдалось в исследованиях развития in vivo. Например, на стадиях 3 и 4 из настоящего описания и в соответствии с Таблицей 17, применение агентов, способных к замедлению, репрессии, супрессии и/или ингибированию эндокринной дифференцировки и/или замедлению, репрессии, супрессии и/или ингибированию маркеров, показательных для эндокринной дифференцировки, включает NGN3 and NKX2.2, но не ограничивается ими. На стадиях 5,6 и/или 7, клеточные культуры обрабатывают агентами, способными к индукции, повышению и/или стимуляции эндокринной дифференцировки, например, путем использования ингибитора пути Notch, такого как ингибитор гамма-секретазы. Вкратце, условия культивирования клеток на стадиях 3 и 4 подавляют эндокринные фенотипы, в то время как условия культивирования клеток на стадиях 5, 6 и/или 7 постепенно индуцируют эндокринные фенотипы, включая инсулин (INS), глюкагон (GCG), соматостатин (SST), панкреатический полипептид (РР), грелин (GHRL), семейства носителей растворенных веществ 30 член 8 (SLC30A8), глюкозо-6-фосфатазу 2 (G6PC2), прогормона конвертазу 1 (PCSK1) и глюкозокиназу (GCK), но не ограничиваясь ими.
В одном варианте осуществления PDX1-позитивные панкреатические эндодермальные клетки дифференцированы в эндокринные клетки и эндокринные прогениторы/предшественники путем продолжения инкубации PDX1-позитивных панкреатических эндодермальных клеток в присутствии ингибитора передачи сигналов Notch, например, ингибитора гамма-секретазы, такого как R04929097, используемого по отдельности или в комбинации с ретиноидами, такими как ретиноевая кислота. Присутствие ингибитора передачи сигналов Notch индуцирует экспрессию NGN3 во время стадии 5. В других вариантах осуществления ингибитор гамма-секретазы обеспечивают в начале способа дифференцировки, например, плюрипотентной стадии, и оставляют в клеточной культуре на протяжении дифференцировки гормон-экспрессирующих клеток панкреатических островков. В других вариантах осуществления ингибитор гамма-секретазы добавляют после начала дифференцировки, но перед дифференцировкой до стадии PDX1-позитивной энтодермы передней части пищеварительного тракта. В предпочтительных вариантах осуществления ингибитор гамма-секретазы обеспечивают в клеточной культуре или клеточной популяции примерно в то же самое время, что и обеспечение факторов дифференцировки, индуцирующих превращение дефинитивной энтодермы в PDX1-позитивную энтодерму. В других предпочтительных вариантах осуществления ингибитор гамма-секретазы обеспечивают для клеточной культуры или клеточной популяции после дифференцировки существенной части клеток в клеточной культуре или клеточной популяции в клетки PDX1-позитивной энтодермы передней части пищеварительного тракта.
В другом варианте осуществления клеточные культуры со стадии 5 дополнительно дифференцируются на стадии 6 и 7. Во время этих стадий применяют агенты, способные к надлежащей специализации эндокринных прогениторов/предшественников или прогениторных клеток для дифференцировки в более продвинутые в развитии и/или зрелые эндокринные клетки in vivo. Такие агенты включают ретиноид или ретиноевую кислоту, BMP, никотинамид, IGF, белки хэджхог и тому подобные, но не ограничиваются ими. В предпочтительном варианте осуществления TTNPB (4-[Е-2-(5,6,7,8-тетрагидро-5,5,8,8-тетраметил-2-нафталенил)-1-пропенил] бензойная кислота или аротиноидная кислота) является селективным и высоко активным аналогом ретиноевой кислоты с аффинностью к рецепторам ретиноевой кислоты (RAR) α, β, и γ, которые являются нуклеарными факторами транскрипции. TTNPB и другая ретиноевая кислота или аналоги ретиноевой кислоты вызывают лиганд-активированную транскрипцию генов, обладающих элементами, чувствительными к ретиноевой кислоте. Следовательно, другие агенты или лиганды, способные к активации элементов, чувствительных к ретиноевой кислоте или связыванию любого из RAR, потенциально пригодны и применимы в настоящем изобретении для стимуляции дифференцировки эндокринных клеток.
В другом варианте осуществления культуры клеток со стадии 5 или стадии 6 или стадии 7 могут быть дополнительно обработаны матригелем по отдельности или в комбинации с ингибиторам rho-киназы для улучшения клеточной адгезии. Альтернативно, ингибитор rho-киназы может быть добавлен на любой стадии из стадий 5, 6 или 7 в концентрациях, достаточных для стимуляции выживания, массы и выхода клеток. Альтернативно, клеточные культуры могут быть диссоциированы и ре-диссоциированы во время или между стадиями 6 и 7, для удаления или уменьшения нежелательных клеточных типов, включая субпопуляции не-эндокринных панкреатических прогениторов (CHGA-), но не ограничиваясь ими. См. Пример 14.
В другом варианте осуществления любые из стадий 1-7 могут быть продлены на 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или больше дней, или сокращены на 1, 2, 3, 4, 5, 6, 7 или больше дней. Например, в одном предпочтительном варианте осуществления и в соответствии с Таблицей 17, стадия 6 занимает больше примерно 6 дней, а стадия 7 больше примерно 9 дней. Однако обе эти стадии могут быть сокращены для приспособления к продукции эндокринных прогениторов/предшественников, например, продлены и увеличены до 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40 или больше дней для получения более продвинутых на пути развития эндокринных клеток.
В другом варианте осуществления обеспечиваются способы получения более продвинутых на пути развития ПЭК или панкреатических эндокринных прогениторов/предшественников или панкреатических эндокринных клеток. В одном аспекте настоящего изобретения обеспечивается эндокринная клетка, экспрессирующая единственный панкреатический гормон, являющаяся со-позитивной по PDX1 и NKX6.1, например, незрелая бета-клетка, одновременно экспрессирующая INS, PDX1 и NKX6.1. В другом аспекте продвинутая по пути развития клетка может быть ПЭК культурой, которая состоит главным образом из не-эндокринных панкреатических прогениторных клеток, экспрессирующих по меньшей мере PDX и NKX6.1, но не состоит минимально из клеток, коммитированных в эндокринную линию (CHGA+), экспрессирующую по меньшей мере CHGA+, NGN3 и/или NKX2.2. Эти ПЭК культуры продвинуты по пути развития, поскольку они считаются более родственными тем, которые были показаны in vivo в исследованиях развития при панкреатической дифференцировке. См. Jorgensen и др. (2007), «An Illustrated Review of Early Pancreas Development in the Mouse», Endocrine Reviews 28(6):685-705 («Иллюстрированный обзор раннего развития поджелудочной железы у мышей») и Rukstalis and Habener (2009), «Neurogenin3: A master regulator of pancreatic islet differentiation and regeneration», Islets 1(3): 177-184 («Нейрогенин 3: главный регулятор дифференцировки и регенерации панкреатических островков»). Другим примером продвинутой по пути развития клетки или клеточной культуры является пример, в котором клетки диссоциированы и реагрегированы или реассоциированы, так что реагрегированная клеточная культура состоит из более однородной популяции клеток. Эта более однородная популяция клеток в одном аспекте изобретения не содержит клеточных типов на более ранних стадиях, например, панкреатических эндокринных прогениторов/предшественников или эндокринных клеток, которые преимущественно состоят из клеток, экспрессирующих единственный гормон, являющихся со-позитивными по PDX1 и NKX6.1, но не содержат клеток или субпопуляций со стадий 3 или 4, или ПЭК или субпопуляций ПЭК, включающих не-эндокринные мультипотентные панкреатические прогениторные (CHGA-) клетки, или остаточные/тройные негативные клетки (CHGA-/PDX1-/NKX6.1-) и тому подобные, или мульти-гормон-экспрессирующие клетки (т.е. клетки, экспрессирующие более одного гормона в любом одном типе клеток), но не ограничивающихся ими.
В другом варианте осуществления клеточные культуры, обеспеченные в настоящей заявке, предпочтительно являются адекватно специализированными, что имеет специфическое и определенное значение в области биологии развития. В контексте биологии развития, существует механизм, посредством которого клетки получают определенное направление или являются специализированными или адекватно специализированными. Поскольку дифференцировка клеточной культуры in vitro не имеет временной организации и меток, типичных для исследований развития in vivo, применение и надежность клеточных маркеров (поверхностных или внутренних маркеров, таких факторов транскрипции), в частности, сигнатурные или отличительные маркеры, характерные для специфического типа клеток, являются существенными. Таким образом, ссылка на адекватно специализированные клетки, клеточные культуры, субпопуляции и популяции означает, что эти клетки имеют различные направления, и эти направления являются более определенными и детерминированными на основе их экспрессии маркеров, профиля экспрессируемых маркеров, и что важно, маркеров, которые они не экспрессируют. Надлежащая специализация клеточных культур in vitro зависит от исследований развития подобных клеток in vivo, таким образом, в одном предпочтительном варианте осуществления исследования развития in vivo являются руководством для дифференцировки in vitro и характеристики адекватно специализированных клеток.
В некоторых вариантах осуществления изобретения, описанных в настоящей заявке, эксендин 4 обеспечивают для дифференцировки клеточной культуры или клеточной популяции примерно в то же самое время, что и ингибитор гамма-секретазы. В некоторых вариантах осуществления эксендин 4 обеспечивают так, чтобы он присутствовал в клеточной культуре или клеточной популяции в концентрации по меньшей мере примерно от 0,1 нг/мл до 1000 нг/мл.
Необходимо понять, что экспрессия маркеров NGN3, NKX2.2 и/или РАХ4 индуцируется в диапазоне различных уровней в эндокринных прогениторах/клетках-предшественниках, в зависимости от условий дифференцировки. Как таковая, в некоторых вариантах осуществления, описанных в настоящей заявке, экспрессия маркеров NGN3, NKX2.2 и/или РАХ4 в эндокринных прогениторах/клетках-предшественниках или клеточных популяциях превышает по меньшей мере в 2 раза - 10000 раз экспрессию маркеров NGN3, NKX2.2 и/или РАХ4 в не-эндокринных прогениторах/клетках-предшественниках или клеточных популяциях, например, плюрипотентных стволовых клетках, клетках дефинитивной энтодермы, клетках PDX1-позитивной энтодермы передней части пищеварительного тракта, незрелых гормон-экспрессирующих клетках панкреатических островков, зрелых гормон-экспрессирующих клетках панкреатических островков, клетках экстраэмбриональной энтодермы, клетках мезодермы и/или клетках эктодермы. В других вариантах осуществления экспрессия маркеров NGN3, NKX2.2 и/или РАХ4 в эндокринных прогениторах/клетках-предшественниках или клеточных популяциях превышает по меньшей мере примерно в 4 раза, по меньшей мере в 6 раз - в 10000 раз экспрессию маркеров NGN3, NKX2.2 и/или РАХ4 в не-эндокринных прогениторах/клетках-предшественниках или клеточных популяциях, например, плюрипотентных стволовых клетках, клетках дефинитивной энтодермы, клетках PDX1-позитивной энтодермы передней части пищеварительного тракта, незрелых гормон-экспрессирующих клетках панкреатических островков, зрелых гормон-экспрессирующих клетках панкреатических островков, клетках экстраэмбриональной энтодермы, клетках мезодермы и/или клетках эктодермы. В некоторых вариантах осуществления экспрессия маркеров NGN3, NKX2.2 и/или РАХ4 в эндокринных прогениторах/клетках-предшественниках или клеточных популяциях неограниченно превышает экспрессию маркеров NGN3, NKX2.2 и/или РАХ4 в неэндокринных прогениторах/клетках-предшественниках или клеточных популяциях, например, плюрипотентных стволовых клетках, таких как iPS клетки и hES клетки, клетках дефинитивной энтодермы, клетках PDX1-позитивной энтодермы передней части пищеварительного тракта, незрелых гормон-экспрессирующих клетках панкреатических островков, зрелых гормон-экспрессирующих клетках панкреатических островков, клетках экстраэмбриональной энтодермы, клетках мезодермы и/или клетках эктодермы.
Другие варианты осуществления настоящего изобретения относятся к композициям, таким как клеточные культуры или клеточные популяции, содержащим человеческие клетки, включая человеческие эндокринные прогениторы/клетки-предшественники, где экспрессия маркера NGN3 превышает экспрессию маркера AFP, SOX7, SOX1, ZIC1, NFM, MAFA, SYP, CHGA, INS, GCG, SST, GHRL и/или PAX6 по меньшей мере примерно на 2% от человеческих клеток. В других вариантах осуществления экспрессия маркера NGN3 превышает экспрессию маркера AFP, SOX7, SOX1, ZIC1, NFM, MAFA, SYP, CHGA, INS, GCG, SST, GHRL и/или PAX6 по меньшей мере примерно на 5%-98% от человеческих клеток. В некоторых вариантах осуществления процент человеческих клеток в клеточных культурах или популяциях, где экспрессия маркера NGN3 превышает экспрессию маркера AFP, SOX7, SOX1, ZIC1, NFM, MAFA, SYP, CHGA, INS, GCG, SST, GHRL и/или PAX6, рассчитывают независимо от фидерных клеток.
Необходимо понять, что некоторые варианты осуществления настоящего изобретения относятся к композициям, таким как клеточные культуры или клеточные популяции, содержащим человеческие эндокринные прогениторы/клетки-предшественники, где экспрессия маркера NKX2.2 и/или РАХ4 превышает экспрессию маркера AFP, SOX7, SOX1, ZIC1, NFM, MAFA, SYP, CHGA, INS, GCG, SST, GHRL и/или PAX6 по меньшей мере от примерно 2% до более чем примерно 98% человеческих клеток. В некоторых вариантах осуществления экспрессия маркера NKX2.2 и/или РАХ4 превышает экспрессию маркера AFP, SOX7, SOX1, ZIC1, NFM, MAFA, SYP, CHGA, INS, GCG, SST, GHRL и/или PAX6 по меньшей мере от 5% до 98% человеческих клеток. В некоторых вариантах осуществления процент человеческих клеток в клеточных культурах или популяциях, где экспрессия маркера NKX2.2 и/или РАХ4 превышает экспрессию маркера AFP, SOX7, SOX1, ZIC1, NFM, MAFA, SYP, CHGA, INS, GCG, SST, GHRL и/или PAX6, рассчитывают независимо от фидерных клеток.
С применением способов, описанных в настоящей заявке, можно получить композиции, содержащие эндокринные прогениторы/клетки-предшественники, по существу свободные от других типов клеток. В некоторых вариантах осуществления настоящего изобретения клеточные популяции или клеточные культуры эндокринных прогениторов/клеток-предшественников, полученные посредством способов, описанных в настоящей заявке, по существу свободны от клеток, проявляющих значительную экспрессию маркеров AFP, SOX7, SOX1, ZIC1 и/или NFM. В некоторых вариантах осуществления популяции или культуры эндокринных клеток-предшественников, полученные способами, описанными в настоящей заявке, по существу свободны от клеток, проявляющих значительную экспрессию маркеров AFP, SOX7, SOX1, ZIC1, NFM, MAFA, SYP, CHGA, INS, GCG, SST, GHRL и/или PAX6.
В других способах получения незрелых гормон-экспрессирующих клеток панкреатических островков, описанных в настоящей заявке, HGF обеспечивают для клеток так, чтобы он присутствовал в клеточной культуре или клеточной популяции в концентрациях, достаточных для стимуляции дифференцировки по меньшей мере части эндокринных прогениторов/клеток-предшественников в незрелые гормон-экспрессирующие клетки панкреатических островков. В некоторых вариантах осуществления HGF присутствует в клеточной культуре или клеточной популяции в концентрации по меньшей мере примерно 1 нг/мл, по меньшей мере примерно от 5 нг/мл до 1000 нг/мл.
В других способах получения незрелых гормон-экспрессирующих клеток панкреатических островков, описанных в настоящей заявке, IGF1 обеспечивают для клеток так, чтобы он присутствовал в клеточной культуре или клеточной популяции в концентрациях, достаточных для стимуляции дифференцировки по меньшей мере части эндокринных прогениторов/клеток-предшественников в незрелые гормон-экспрессирующие клетки панкреатических островков. В некоторых вариантах осуществления IGF1 присутствует в клеточной культуре или клеточной популяции в концентрации по меньшей мере примерно от 1 нг/мл до 1000 нг/мл.
В некоторых вариантах осуществления способов получения незрелых гормон-экспрессирующих клеток панкреатических островков, как описано в настоящей заявке, один или несколько из никотинамида, эксендина 4, HGF и IGF1 обеспечивают после удаления из клеточных культур одного или нескольких ранее обеспеченных факторов дифференцировки. В других вариантах осуществления один или несколько из никотинамида, эксендина 4, HGF и IGF1 обеспечивают для клеточной культуры или клеточной популяции, содержащей один или несколько факторов дифференцировки, которые обеспечены раньше, или обеспечены примерно в то же самое время, как один или несколько из никотинамида, эксендина 4, HGF и IGF1. В предпочтительных вариантах осуществления факторы дифференцировки, которые обеспечены раньше, или обеспечены примерно в то же самое время, как один или несколько из никотинамида, эксендина 4, HGF и IGF1, включают DAPT, FGF-10, KAAD-циклопамин, активин А, активин В, ВМР4 и/или RA, но не ограничиваются ими.
Другие варианты осуществления настоящего изобретения относятся к композициям, таким как клеточные культуры или клеточные популяции, содержащие человеческие клетки, включая человеческие незрелые гормон-экспрессирующие клетки панкреатических островков, где экспрессия маркера MAFB, SYP, CHGA, NKX2.2, ISL1, PAX6, NEUROD, PDX1, HB9, GHRL, IAPP, INS GCG, SST, PP и/или С-пептида превышает экспрессию маркера NGN3, MAFA, МОХ1, CER, POU5F1, AFP, SOX7, SOX1, ZIC1 и/или NFM по меньшей мере примерно в 2% человеческих клеток. В других вариантах осуществления экспрессия маркера MAFB, SYP, CHGA, NKX2.2, ISL1, PAX6, NEUROD, PDX1, HB9, GHRL, IAPP INS GCG, SST, PP и/или С-пептида превышает экспрессию маркера NGN3, MAFA, МОХ1, CER, POU5F1, AFP, SOX7, SOX1, ZIC1 и/или NFM по меньшей мере примерно в 5% человеческих клеток, по меньшей мере примерно от 10% человеческих клеток до 95% человеческих клеток, или по меньшей мере примерно в 98% человеческих клеток.
В некоторых вариантах осуществления настоящего изобретения клеточные культуры и/или клеточные популяции, содержащие незрелые гормон-экспрессирующие клетки панкреатических островков, также включают среду, содержащую один или несколько секретируемых гормонов, выбранных из грелина, инсулина, соматостатина и/или глюкагона. В других вариантах осуществления среда содержит С-пептид. В предпочтительном варианте осуществления концентрация одного или нескольких секретируемых гормонов или С-пептида в среде находится в диапазоне от по меньшей мере примерно 1 пмоль грелина, инсулина, соматостатина, глюкагона или С-пептида на мкг клеточной ДНК до по меньшей мере примерно 1000 пикомоль (пмоль) грелина, инсулина, соматостатина, глюкагона или С-пептида на мкг клеточной ДНК.
Способ получения инсулина in vivo
В некоторых вариантах осуществления полученные in vitro панкреатические клетки-прогениторы или типы клеток PDX-1-позитивной панкреатической энтодермы, или их эквиваленты, описанные выше, трансплантируют млекопитающему субъекту. Эти способы подробно описаны в Международной заявке PCT/US2007/015536, озаглавленной «Способы получения панкреатических гормонов», и Kroon и др. (2008) выше. В предпочтительном варианте осуществления млекопитающим субъектом является человеческий субъект. Особо предпочтительными субъектами являются те, кто признан имеющим состояние, ограничивающее способность субъекта к продукции достаточных уровней инсулина в ответ на физиологически высокие концентрации глюкозы в крови. Диапазон уровней глюкозы в крови, представляющий физиологически высокий уровень глюкозы крови для какого-либо конкретного вида млекопитающих, может легко определить рядовой специалист в данной области техники. Любой стойкий уровень глюкозы в крови, приводящий к выраженному заболеванию или состоянию, считается физиологически высоким уровнем глюкозы в крови.
Дополнительные варианты осуществления настоящего изобретения относятся к клетке, секретирующей инсулин in vivo, полученной из in vitro плюрипотентной стволовой клетки или ее потомства, например, к мультипотентным клеткам, таким как клетка PDX-1 позитивной энтодермы передней части пищеварительного тракта, клетка PDX-1-позитивной панкреатической энтодермы или панкреатическая прогениторная клетка, эндокринный прогенитор/предшественник, такой как NGN3-позитивный эндокринный прогенитор/предшественник, или функциональная дифференцированная гормон-секретирующая клетка, такая как инсулин-, глюкагон-, соматостатин-, грелин- или панкреатический полипептид-секретирующая клетка. Любые из вышеописанных окончательно дифференцированных или мультипотентных клеток могут быть трансплантированы хозяину, или млекопитающему, и созревать в физиологически функциональные гормон-секретирующие клетки, такие как инсулин-секретирующие клетки, в ответ на высокие уровни глюкозы в крови хозяина. В предпочтительных вариантах осуществления клетки не образуют тератомы in vivo, и если она формируется, остаются локализованными в области трансплантата и могут легко быть отделены или удалены. В особо предпочтительных вариантах осуществления клетки не содержат какой-либо кариотипической аномалии во время процесса дифференцировки in vitro, или при трансплантации млекопитающему in vivo, или при созревании и развитии в функциональные островки in vivo.
Далее, хотя варианты осуществления, описанные в настоящей заявке, относятся к сконструированной или генетически рекомбинантной плюрипотентной клетке, мультипотентной или дифференцированной клетке, полученной из плюрипотентной клетки, такой как человеческая iPS клетка, на основе описания, обеспеченного в настоящей заявке, предполагается, что поскольку iPS клетки демонстрируют схожую физиологию и профили экспрессии маркерных генов с hES клетками и полученными из hES клетками, то они должны иметь схожие физиологические характеристики in vivo.
Способ инкапсулирования панкреатических прогениторов
В некоторых вариантах осуществления композиция плюрипотентных, мультипотентных и дифференцированных клеток, описанная в настоящей заявке, может быть инкапсулирована в биологическом и/или не-биологическом механическом устройстве, где инкапсулирующее устройство отделяет и/или изолирует клеточные композиции от хозяина.
Способы инкапсулирования подробно описаны в заявке США №61/114,857, поданной 14 ноября 2008, озаглавленной «Инкапсулирование панкреатических прогениторов, полученных из HES клеток», и заявке США №61/121,086, поданной 12 декабря 2008, озаглавленной «Инкапсулирование клеток панкреатической энтодермы».
В одном варианте осуществления устройство для инкапсулирования содержит клетки, полученные из плюрипотентных, например, клетки PDX-1-позитивной энтодермы передней части пищеварительного тракта, клетки PDX-1-позитивной панкреатической энтодермы или прогениторные клетки, эндокринный прогенитор/предшественник, такой как NGN3-позитивный эндокринный прогенитор/предшественник, или функционально дифференцированные гормон-секретирующие клетки, такие как инсулин-, глюкагон-, соматостатин-, грелин-, или панкреатический полипептид-секретирующие клетки, в полупроницаемой мембране, предотвращающей проникновение трансплантированной клеточной популяции, хранящейся в устройстве, и в то же самое время обеспечивающий проход некоторых секретируемых полипептидов, например, инсулина, глюкагона, соматостатина, грелина, панкреатического полипептида и тому подобного. Альтернативно, устройство имеет множество мембран, включая васкуляризованную мембрану.
Применение агентов для усиления и индукции роста, выживания, пролиферации и клеточной адгезии человеческих плюрипотентных стволовых клеток, например. hES клеток и iPS клеток
На клеточную регуляцию можно влиять путем трансдукции экстрацеллюлярных сигналов через мембрану, что в свою очередь модулирует биохимические пути в клетке. Фосфорилирование белков представляет один вариант, при котором внутриклеточные сигналы передаются от молекулы к молекуле, в итоге приводя к клеточному ответу. Эти каскады трансдукции сигнала являются высоко регулируемыми, и часто перекрываются, о чем свидетельствует существование многих киназ, а также фосфатаз. Отмечалось, что у человека белковые тирозинкиназы, как известно, играют существенную роль в развитии многих патологических состояний, включая диабет, рак, а также связаны с широким рядом врожденных синдромов. Серин-треонин-киназы, например, Rho киназы, являются классом ферментов, ингибирование которых может быть пригодным для лечения заболеваний человека, включая диабет, рак и множество сердечнососудистых заболеваний и СПИД. Большинство ингибиторов, идентифицированных на сегодняшний день, действуют на участок связывания АТФ. Такие конкурирующие с АТФ ингибиторы продемонстрировали селективность благодаря своей способности к нацеливанию на менее консервативные области участка связывания АТФ.
Семейство Rho киназы малых ГТФ-связывающих белков содержит по меньшей мере 10 членов, включая Rho А-Е и G, Rac 1 и 2, Cdc42, и ТС10. Ингибиторы часто обозначаются как ROK или ROCK ингибиторы, или ингибиторы Rho-киназы, и термины применяются взаимозаменяемо в настоящей заявке. Эффекторные домены RhoA, RhoB, и RhoC имеют одну и ту же аминокислотную последовательность, и по-видимому, имеют схожие внутриклеточные мишени. Rho киназа действует в качестве первичного медиатора, регулирующего последующие звенья каскада Rho, и существует в двух изоформах: α (ROCK2) и β (ROCK1). Белки семейства Rho киназы имеют каталитический (киназный) домен в их N-концевом домене, суперспиральный домен в средней части, и гипотетический плекстрин-гомологичный (РН) домен в С-концевом участке. Rho-связывающий домен ROCK локализован в С-концевой части суперспирального домена, и связывание ГТФ-связанной формы Rho приводит к повышению киназной активности. Rho/Rho-киназа-опосредованный путь играет важную роль в передаче сигнала, инициированной многими агонистами, включая ангиотензин II, серотонин, тромбин, эндотелин-1, норадреналин, тромбоцитарный фактор роста, АТФ/АДФ и экстрацеллюлярные нуклеотиды, и уротензин II. Через модуляцию своих целевых эффекторов/субстратов Rho киназа играет важную роль в различных клеточных функциях, включая сокращение гладких мышц, организацию актинового цитоскелета, клеточную адгезию и подвижность, и экспрессию генов. Благодаря роли, которую Rho киназные белки играют в опосредовании ряда клеточных функций, считающихся связанными с патогенезом атеросклероза, ингибиторы этих киназ могут также быть пригодными для лечения или профилактики различных атеросклеротических сердечнососудистых заболеваний, и вовлечены в сокращение эндотелия.
В некоторых вариантах осуществления агенты, которые стимулируют и/или поддерживают клеточный рост, выживание, пролиферацию и клеточную адгезию, добавляют в различные среды для культивирования клеток, включая ингибиторы Rho-киназы Y-27632, Fasudil (также обозначаемый как НА1077), Н-1152Р и ITS (инсулин/трансферрин/селен; Gibco), Wf-536, Y-30141 (описан в патенте США №5,478,838, включенном посредством ссылки во всей полноте) и их производные, и антисмысловую нуклеиновую кислоту для ROCK; нуклеиновую кислоту, индуцирующую интерференцию РНК (например, миРНК), конкурирующие пептиды, пептиды-антагонисты, ингибирующие антитела, ScFV-фрагменты антител, доминантно-негативные варианты, и их векторы экспрессии, но не ограничиваясь ими. Далее, поскольку другие низкомолекулярные соединения известны как ингибиторы ROCK, такие соединения или их производные также можно применять в настоящем изобретении (например, см. патентные заявки США №№20050209261, 20050192304, 20040014755, 20040002508, 20040002507, 20030125344 и 20030087919, и Международные патентные публикации №№2003/062227, 2003/059913, 2003/062225, 2002/076976 и 2004/039796).
В настоящем изобретении можно также применять комбинацию одного или двух или более ингибиторов ROCK. Эти агенты функционируют, в свою очередь, путем стимуляции ре-ассоциации диссоциированных hES клеток, iPS или дифференцированных клеточных культур, например, дефинитивной энтодермы, энтодермы передней части пищеварительного тракта, панкреатической энтодермы, панкреатического эпителия, популяций панкреатических прогениторов, эндокринных прогениторов и популяций, и тому подобного. Подобным образом, агенты могут функционировать, когда клеточная диссоциация не происходит. Увеличение роста, выживания, пролиферации и клеточной адгезии человеческих плюрипотентных стволовых клеток достигалось независимо от того, были ли клетки получены из клеточных агрегатов в суспензии или из адгезионных культур (с компонентами экстрацеллюлярного матрикса или без низ, с сывороткой или без нее, с фидерными фибробластами или без них, с FGF или без него, с активином или без него). Повышение выживания этих клеточных популяций облегчает и улучшает системы очистки с применением клеточного сортера, таким образом, повышает выход клеток. Применение ингибиторов Rho киназы, таких как Y27632, может обеспечивать экспансию дифференцированных типов клеток, а также стимуляцию их выживания во время пассажей диссоциированных единичных клеток или криоконсервирования. Хотя ингибиторы Rho киназы, такие как Y27632, тестировали на человеческих плюрипотентных стволовых клетках (например, hES и iPS клетках), и клетках, дифференцированных из них, ингибиторы Rho киназы можно применять для других типов клеток, например, в целом, эпителиальных клеток, включая клетки тонкой кишки, легких, тимуса, почки, но не ограничиваясь ими, а также нервных типов клеток, таких как пигментный эпителий сетчатки.
Концентрация ингибитора ROCK в среде для культивирования клеток не ограничивается той, что описана в примерах ниже, с условием, что концентрация позволяет достичь необходимых эффектов, таких как усиление, повышение и/или стимуляция роста, выживания, пролиферации и адгезии клеток. Специалисту в данной области техники понятно, что может быть необходима оптимизация различных ингибиторов ROCK в различных условиях. Например, при использовании Y-27632 предпочтительная концентрация может быть в диапазоне примерно от 0,01 до 1000 мкМ, более предпочтительно примерно от 0,1 до 100 мкм, и еще более предпочтительно примерно от 1,0 до 50 мкМ, и наиболее предпочтительно примерно от 5 до 20 мкМ. Когда применяют Fasudil/HA1077, его можно использовать в концентрации, превышающей примерно в два или три раза концентрацию вышеупомянутого Y-27632. Когда применяют Н-1152, его можно применять в концентрации, составляющей, например, около 1/10, 1/20, 1/30, 1/40, 1/50 или 1/60 от концентрации вышеупомянутого Y-27632. Концентрация используемого ингибитора ROCK зависит отчасти от биологической активности и мощности ингибитора, и условий, в которых его применяют.
Время и период лечения ингибитором ROCK может быть ограниченным или не ограниченным, в зависимости от необходимых эффектов, таких как усиление, повышение и/или стимуляция роста, выживания, пролиферации (клеточной массы) и клеточной адгезии. Однако добавление ингибитора ROCK может также влиять на дифференцировку необычным образом, как лучше описано в Примере 7. Примеры внизу описывают культуры человеческих плюрипотентных стволовых клеток и/или культуры дифференцированных клеток, обработанных в течение примерно 12 часов, 24 часов, 48 часов или больше.
Плотность культур человеческих плюрипотентных стволовых клеток, обработанных ингибитором ROCK, также не ограничена, если при этой плотности достигаются необходимые эффекты, такие как усиление, повышение и/или стимуляция роста, выживания, пролиферации и адгезии клеток. Клеточную плотность засеянных клеток можно регулировать в зависимости от множества факторов, включая применение адгезионных или суспензионных культур, специфическую рецептуру используемой среды для культивирования, условия роста и предполагаемое применение культивируемых клеток, но не ограничиваясь ими. Примеры плотности клеточных культур включают 0,01 х 105 клеток/мл, 0,05 х 105 клеток/мл, 0,1 х 105 клеток/мл, 0,5 х 105 клеток/мл, 1,0 х 105 клеток/мл, 1,2 х 105 клеток/мл, 1,4 х 105 клеток/мл, 1,6 х 105 клеток/мл, 1,8 х 105 клеток/мл, 2,0 х 105 клеток/мл, 3,0 х 105 клеток/мл, 4,0 х 105 клеток/мл, 5,0 х 105 клеток/мл, 6,0 х 105 клеток/мл, 7,0 х 105 клеток/мл, 8,0 х 105 клеток/мл, 9,0 х 105 клеток/мл, или 10,0 х 105 клеток/мл, или больше, например, до 5 х 107 клеток/мл, или любое значение между ними, но не ограничиваясь ими, при культивировании с хорошим клеточным ростом, выживанием, пролиферацией и адгезией.
Применение агентов, активирующих члены семейства рецептора TGFβ
В другом варианте осуществления агенты, активирующие член семейства рецептора TGFβ, включают члены TGFβ суперсемейства факторов роста, как описано в настоящей заявке. Использующийся в настоящей заявке термин «член суперсемейства TGFβ» или его эквиваленты означает более 30 структурно связанных белков, включая субсемейства, включающие TGFβ1, TGFβ2, TF-β3, GDF-15, GDF-9, BMP-15, BMP-16, ВМР-3, GDF-10, ВМР-9, ВМР-10, GDF-6, GDF-5, GDF-7, ВМР-5, ВМР-6, ВМР-7, ВМР-8, ВМР-2, ВМР-4, GDF-3, GDF-1, GDF 11, GDF8, активины βС, βЕ, βА и βВ, ВМР-14, GDF-14, MIS, ингибин-альфа, Lefty1, Lefty2, GDNF, нейротурин, персефин и артемин. См. Chang и др. (2002) Endocrine Rev. 23(6):787-823.
Член семейства TGFβ может быть замещен, или использован в сочетании с активатором пути передачи сигналов TGFβ, который является соединением, стимулирующим один или несколько полипептидов или взаимодействий, участвующих в трансдукции или ином осуществлении изменений свойств клетки в ответ на член семейства TGFβ. Путь передачи сигнала TGFβ включает сами члены семейства TGFβ. Члены суперсемейства TGFβ передают сигналы к ядру путем передачи сигнала через рецепторы серин-треонин-киназы II и I типа и внутриклеточные эффекторы, известные как Smad. Эти рецепторы относятся к двум субсемействам, известным как рецепторы I и II типа, которые совместно действуют для связывания лиганда и передачи сигнала (Attisano и др., Mol Cell Biol 16 (3), 1066-1073 (1996)). Лиганды связывают рецепторы I и II типа на клеточной поверхности, стимулируя активацию рецептора I типа через фосфорилирование. Этот активированный комплекс, в свою очередь, активирует внутриклеточные Smad, которые собирают мульти-субъединичные комплексы, регулирующие транскрипцию. Члены суперсемейства TGFβ разделяются на две подгруппы передачи сигнала: те, чья функция связана с TGFβ/активином, и те, чья функция связана с субсемейством BMP/GDF. Считают, что большинство лигандов TGFβ связываются вначале с рецептором II типа, и этот комплекс лиганд/рецептор II типа затем рекрутирует или фосфорилирует рецептор I типа (Mathews, L S, Endocr Rev 15:310-325 (1994); Massague, Nature Rev: Mol Cell Biol. 1, 169-178 (2000)). Киназа рецептора II типа фосфорилирует и активирует киназу рецептора I типа, которая затем фосфорилирует и активирует белки Smad. Лиганды TGFβ/активин связываются с рецепторами II типа TGFβ и активина, и могут активировать Smad-2 и -3, Nodal и Lefty сигнал посредством этого пути активинового типа. BMP/GDF лиганды связываются рецепторами BMP II типа, и могут активировать Smad 1, 5, и 8. См. Derynck, R и др. Cell 95, 737-740 (1998)). При стимуляции лигандом Smad перемещаются к ядру и функционируют в качестве компонентов комплексов транскрипции.
Передача сигнала TGFβ регулируется позитивно и негативно через различные механизмы. Позитивная регуляция усиливает сигнал до уровня, достаточного для биологической активности. Лиганды суперсемейства TGFβ связываются с рецептором II типа, который привлекает и фосфорилирует рецептор I типа. Рецептор I типа затем фосфорилирует SMAD, регулируемые рецептором (R-SMAD, например, SMAD1, SMAD2, SMAD3, SMAD5, и SMAD8) которые связывают общий медиатор Smad или coSMAD. Комплексы R-SMAD/coSMAD аккумулируются в ядре, где они действуют, как факторы транскрипции, и участвуют в регуляции экспрессии целевого гена. Например, факторы роста и дифференцировки включают 1, 2, 3, 5, 6, 7, 8, 9, 10, 11 и 15. В одном предпочтительном варианте осуществления GDF8 и GDF11 являются членами семейства TGFβ, которые также являются активаторами пути передачи сигнала TGFβ (позитивной регуляции), и действуют путем связывания с экстрацеллюлярной частью лиганд-связывающего домена ActRII рецептора, а затем образуют комплекс с ActRI, приводя к ингибированию Smad7 негативного регулятора и фосфорилированию комплекса Smad2/Smad3. Комплекс Smad2/Smad3 ассоциируется с Smad4 для регуляции экспрессии некоторых генов.
Как с применением любого агента, концентрация любого члена суперсемейства TGFβ в среде для культивирования клеток не ограничивается описанной в примерах ниже, если концентрация позволяет достичь необходимых эффектов, например, чтобы активировать член семейства рецептора TGFβ. Например, при использовании активинов, например, активина А и/или В, или GDF8 and GDF-11, предпочтительная концентрация может быть в диапазоне примерно от 10 до 300 нМ, более предпочтительно, примерно от 50 до 200 нМ, и еще более предпочтительно примерно от 75 до 150 нМ, и наиболее предпочтительно примерно от 100 до 125 нМ. Рядовой специалист в данной области техники сможет легко протестировать любую концентрацию, и с применением стандартных методик в данной области техники определить эффективность такой концентрации, например, оценить дифференцировку путем определения экспрессии и отсутствии экспрессии панели генетических маркеров для любого типа клеток.
Применение агентов для продукции эндокринных клеток
В одном варианте осуществления настоящее изобретение обеспечивает способы получения популяций и/или субпопуляций типа панкреатической эндодермальной линии, в частности, ПЭК и/или клеток панкреатической эндодермальной линии, с применением ингибитора пути Notch, включая ингибитор гамма-секретазы, но не ограничиваясь им, в количестве, эффективном для стимуляции эндокринной дифференцировки и/или для индукции экспрессии некоторых маркеров, которые являются критерием эндокринных клеток и показателем эндокринной дифференцировки. Описаны многочисленные ингибиторы гамма-секретазы. См., например, патенты США №№6,756,511; 6,890,956; 6,984,626; 7,049,296; 7,101,895; 7,138,400; 7,144,910; и 7,183,303. Ингибиторы гамма-секретазы легко доступны, например, от Calbiochem (Ла-Холья, Калифорния); ингибитор гамма-секретазы I, (GSI I), Z-Leu-Leu-норлейцин-СНО; ингибитор гамма-секретазы II; ингибитор гамма-секретазы III, (GSI III), N-бензилоксикарбонил-Leu-лейцинал; ингибитор гамма-секретазы IV, (GSI IV), N-(2-нафтоил)-Вал-фенилаланинал; ингибитор гамма-секретазы V, (GSI V), N-бензилоксикарбонил-Leu-фенилаланинал; ингибитор гамма-секретазы VI, (GSI VI), 1-(S)-эндо-N-(1,3,3)-триметилбицикло[2.2.1]гепт-2-ил)-4-фторфенил сульфонамид; ингибитор гамма-секретазы VII, (GSI VII), ментилоксикарбонил-LL-CHO; ингибитор гамма-секретазы IX, (GSI IX), (DAPT), N-[N-(3,5-дифторфенацетил-L-аланил)]-S-фенилглицин t-бутиловый эфир; ингибитор гамма-секретазы X, (GSI X), {1S-бензил-4R-[1-(1S-карбамоил-2-фенэтилкарбамоил)-1S-3-метилбутилкарбамоил]-2R-гидрокси-5-фенилпентил}карбаминовой кислоты трет-бутиловый эфир; ингибитор гамма-секретазы XI, (GSI XI), 7-амино-4-хлоро-3-метоксиизокумарин; ингибитор гамма-секретазы XII, (GSI XII), Z-Ile-Leu-CHO; ингибитор гамма-секретазы XIII, (GSI XIII), Z-Tyr-Ile-Leu-CHO; ингибитор гамма-секретазы XIV, (GSI XIV), Z-Цис(t-Бу)-Ile-Leu-CHO; ингибитор гамма-секретазы XVI, (GSI XVI), N-[N-3,5-дифторфенацетил]-L-аланил-S-фенилглицин метиловый эфир; ингибитор гамма-секретазы XVII, (GSI XVII), WPE-III-31C); ингибитор гамма-секретазы XIX, (GSI XIX), (2S,3R)-3-(3,4-дифторфенил)-2-(4-фторфенил)-4-гидрокси-N-((3S)-2-оксо- -5-фенил-2,3-дигидро-1Н-бензо[е][1,4]диазепин-3-ил)-бутирамид; ингибитор гамма-секретазы XX, (GSI XX), (дибензазепин (DBZ)), (S,S)-2-[2-(3,5-дифторфенил)ацетиламино]-N-(5-метил-6-оксо-6,7-дигидро-5Н-дибензо[b,d]азепин-7-ил)пропионамид, ингибитор гамма-секретазы XXI, (GSI XXI), (S,S)-2-[2-(3,5-дифторфенил)-ацетиламино]-N-(1-метил-2-оксо-5-фенил-2,3-дигидро-1Н-бензо[е][1,4]диазепин-3-ил)-пропионамид; ингибитор гамма40-секретазы I, N-транс-3,5-диметоксициннамоил-Ile-лейцинал; ингибитор гамма40-секретазы II, N-трет-бутилоксикарбонил-Gly-Val-валинал; и изовалерил-V-V-Sta-А-Sta-ОС3.
В одном аспекте ингибитор гамма-секретазы выбран из группы, состоящей из одного или нескольких из: Z-Leu-Leu-норлейцин-СНО, N-бензилоксикарбонил-Leu-лейцинала, N-(2-нафтоил)-Val-фенилаланинала, N-бензилоксикарбонил-Leu-фенилаланинал, 1-(S)-эндо-N-(1,3,3) триметилбицикло[2.2.1]гепт-2-ил)-4-фторфенилсульфонамида, метилоксикарбонил-LL-CHO, N-[N-(3,5-дифторфенацетил-L-аланил)]-S-фенилглицин t-бутилового эфира, {1S-бензил-4R-[1-(1S-карбамоил-2-фенэтилкарбамоил)-1S-3-метилбутилкарбамоил]-2R-гидрокси-5-фенилпентил}карбаминовой кислоты трет-бутилового эфира, 7-амино-4-хлоро-3-метоксиизокумарина, Z-Ile-Leu-CHO, Z-Tyr-Ile-Leu-СНО, Z-Cys(t-Bu)-Ile-Leu-CHO, N-[N-3,5-дифторфенацетил]-L-аланил-S-фенилглицин метилового эфира, (2S,3R)-3-(3,4-дифторфенил)-2-(4-фторфенил)-4-гидрокси-N-((3S)-2-оксо-5-фенил-2,3-дигидро-1Н-бензо[е][1,4]диазепин-3-ил)-бутирамида, (S,S)-2-[2-(3,5-дифторфенил)-ацетиламино]-N-(1-метил-2-оксо-5-фенил-2-,3-дигидро-1Н-бензо[е][1,4]диазепин-3-ил)-пропионамида, N-транс-3,5-диметоксициннамоил-Ile-лейцинала, N-трет-бутиоксикарбонил-Gly-Val-валинала, изовалерил-V-V-Sta-A-Sta-OCH.sub.3; и где почечное заболевание является гломерулярным поражением почек или тубулярным поражением почек. В одном аспекте настоящего заболевания используют примерно от 0,5 до 10 мМ, от 0,5 до 5 мМ, от 0,5 до 3 мМ, предпочтительно от 0,5 до 1 мМ ингибитора гамма-секретазы на любой или на всех стадиях 1-7, описанных в настоящей заявке, и в соответствии с Таблицами 8 и 17.
В одном варианте осуществления инсулиноподобный фактор роста (IGF) является частью сложной системы, которую клетки применяют для сообщения с физиологической средой, и таким образом, может быть использован для дифференцировки PSC в ПЭК и/или панкреатические клетки эндокринной линии. Эта сложная система (часто обозначаемая как IGF «ось») состоит из двух клеточно-поверхностных рецепторов (IGF1R и IGF2R), двух лигандов (инсулиноподобного фактора 1 (IGF-1) и инсулиноподобного фактора 2 (IGF-2)), семейства из шести высокоаффинных IGF-связывающих белков (IGFBP-1 - IGFBP-6), а также связанных IGFBP разрушающих ферментов, вместе обозначаемых как протеазы. Известно, что IGF связывается с IGF1R, рецептором инсулина, рецептором IGF-2, инсулин-связанным рецептором, и возможно, другими рецепторами. Так, в одном аспекте настоящего изобретения факторы роста, которые связываются с этими рецепторами, включают IGF1 и IGF2, но не ограничиваются ими, поскольку любой фактор роста, агент или морфоген, который связывается с любым или с комбинацией этих рецепторов, потенциально оказывает такой же самый физиологический эффект, как тот, который описан и предусмотрен в настоящей заявке. В одном аспекте настоящего изобретения примерно от 5 до 200 нг/мл, от 10 до 100 нг/мл, от 10 до 75 нг/мл, предпочтительно от 10 до 50 нг/мл, предпочтительно от 20 до 50 нг/мл IGF применяют на любой или на всех стадиях 1-7, описанных в настоящей заявке, и в соответствии с Таблицами 8 и 17.
В одном варианте осуществления тромбоцитарный фактор роста (PDGF) используют для дифференцировки PSC в ПЭК и/или клетки, коммитированные в эндокринную линию (CHGA+). PDGF является главным белковым фактором роста, который широко описан как белковый митоген многочисленных видов клеток. PDGF принадлежит к семейству димерных изоформ полипептидных цепей, А, В, С и D, которые действуют через различные рецепторы тирозинкиназы: PDGFR-α и PDGFR-β. В одном аспекте изобретения лиганд является PDGF или его функциональным эквивалентом, включая PDGFaa, PDGFab или PDGFbb, но не ограничиваясь ими, который связывается с этими двумя типами рецепторов. В одном аспекте настоящего изобретения примерно от 5 до 100 нг/мл, от 10 до 75 нг/мл, от 10 to 50 нг/мл, предпочтительно от 10 до 20 нг/мл PDGF применяют на любой или на всех стадиях 1-7, описанных в настоящей заявке, и в соответствии с Таблицами 8 и 17.
В одном варианте осуществления никотинамид, также известный как ниацинамид и никотиновой кислоты амид, являющийся амидом никотиновой кислоты (витамина В3/ ниацина) используют для дифференцировки PSC в ПЭК и/или клетки панкреатической эндокринной линии. Никотиновая кислота, также известная как ниацин, превращается в никотинамид in vivo, и хотя ниацин и никотинамид являются идентичными по своим витаминным функциям, никотинамид не обладает фармакологическим и токсическим эффектами ниацина. Таким образом, другой витамин В или производные витамина В с функцией, подобной никотинамиду, применяются в настоящем изобретении. В одном аспекте настоящего изобретения примерно от 5 до 100 нг/мл, от 10 до 75 нг/мл, от 10 до 50 нг/мл, предпочтительно от 10 до 20 нг/мл никотинамида применяют на любой или на всех стадиях 1-7, описанных в настоящей заявке, и в соответствии с Таблицами 8 и 17.
В другом варианте осуществления семейство белков хэджхог применяют для дифференцировки PSC в ПЭК и/или клетки панкреатической эндокринной линии. Семейство хэджхог внутриклеточных сигнальных молекул позвоночных, упомянутое в настоящем изобретении, состоит по меньшей мере из четырех членов, включая Desert hedgehog (Dhh), Sonic hedgehog (Shh), Indian hedgehog (Ihh) и Moonrat hedgehog (Mhh), но не ограничиваясь ими. В одном аспекте примерно от 5 до 200 нг/мл, от 10 до 200 нг/мл, от 10 до 150 нг/мл, предпочтительно от 10 до 100 нг/мл белка хэджхог применяют на любой или на всех стадиях 1-7, описанных в настоящей заявке, и в соответствии с Таблицами 8 и 17.
После общего описания настоящего изобретения можно получить дополнительное разъяснение со ссылкой на определенные конкретные примеры, которые приведены в настоящей заявке с целью иллюстрации, и не предназначены для ограничения.
Примеры
Необходимо также понимать, что вышеизложенное относится к предпочтительным вариантам осуществления настоящего изобретения, и многочисленные изменения могут быть выполнены, не выходя за рамки настоящего изобретения. Изобретение далее иллюстрировано следующими примерами, которые не должны рассматриваться как ограничивающие объем настоящего изобретения. Напротив, необходимо ясно понимать, что изобретение может выполняться в различных других вариантах осуществления, модификациях и эквивалентах, которые после прочтения описания могут быть предложены специалистом в данной области техники без изменения сущности настоящего изобретения и/или не выходя за рамки формулы изобретения.
Пример 1. Дифференцировка человеческих iPS клеток в панкреатические прогениторные и эндокринные клетки через дефинитивную энтодерму и эндодермальные промежуточные формы
Человеческие индуцированные плюрипотентные стволовые (iPS) клетки дифференцировали в суспензионных агрегатах с применением четырех (4) этапной процедуры в течение примерно 2 недель (или 14 дней) для генерации популяции панкреатических клеточных типов, включая панкреатические прогениторы, эндокринные прогениторы и гормон-экспрессирующие эндокринные клетки. Человеческие линии iPS клеток, применяемые в настоящей заявке, были предоставлены S. Yamanaka, Университет Киото, Япония, и Cellular Dynamics International, Inc. (GDI).
Эти iPS клетки, описанные в настоящей заявке, вначале получали от Shinja Yamanaka, а позднее от CDI. Недифференцированные iPS клетки выращивали на митотически инактивированных мышиных эмбриональных фибробластах, или предпочтительно без фидеров (без фидерного слоя фибробластов) в DMEM/F12, содержащей 20% замены нокаутной сыворотки. Дифференцировку инициировали путем диссоциации недифференцированных iPS клеток на отдельные клетки с применением аккутазы, образцы клеток отбирали на выделение и анализ РНК. Клетки ресуспендировали по 1-2 миллиона клеток на миллилитр в RPMI+0,2% о/о FBS, содержащей 1:5000 разведение инсулина-трансферрина-селена (ITS), активин А (100 нг/мл), wnt3a (50 нг/мл), и ингибитор rho-киназы или ROCK ингибитор, Y-27632 в 10 мкМ, помещая в 6-луночный планшет с ультранизким прикреплением, помещая на ротационную платформу и вращая примерно при 100 об./мин. Культуры вращали при 100 об./мин в оставшейся части процесса дифференцировки с ежедневной заменой среды. Рост, пассирование и пролиферация iPSC были по существу такими, как описано в патентах США №№7,961,402 и 8,211,699.
Способы, описанные в настоящей заявке для получения агрегатных суспензионных культур плюрипотентных клеток, например, hES или iPS клеток, и клеток, полученных из плюрипотентных клеток, являются по существу такими, как описано в PCT/US2007/062755, поданной 23 февраля 2007 и озаглавленной «Композиции и способы для культивирования различных клеток» и PCT/US2008/080516, поданной 20 октября 2008 и озаглавленной «Способы и композиции для бесфидерных сред для плюрипотентных стволовых клеток, содержащих человеческую сыворотку».
Способы, описанные в настоящей заявке, можно упростить, прежде всего, покрытием сосуды для культивирования экстрацеллюлярным матриксом, например, как описано в патенте США 6,800,480 от Bodnar и др., принадлежащем Geron Corporation. Эти способы, а также другие способы культивирования других плюрипотентных стволовых клеток, например, hES и iPS клеток, можно осуществить с применением растворимой человеческой сыворотки, по существу, как описано в заявке США PCT/US2008/080516, поданной 20 октября 2008 и озаглавленной «Способы и композиции для бесфидерных сред для плюрипотентных стволовых клеток, содержащих человеческую сыворотку».
Способы, описанные в настоящей заявке, можно упростить с помощью экзогенно добавленного фактора роста фибробластов (FGF), полученного из иного источника, чем просто фидерный слой фибробластов, как описано в патенте США №7,005,252, Thomson, J., принадлежащем Wisconsin Alumni Research Foundation (WARF).
В течение примерно первых 24 часов вращения, отдельные клетки, прилипающие друг к другу, образуют клеточные агрегаты, и достаточные клеточные образцы были взяты для изоляции и анализа РНК. Клеточные агрегаты находились в диапазоне примерно от 60 микрон до 120 микрон в диаметре. Спустя примерно 1 день (24 часа) после помещения образцов iPS клеток на ротационную платформу, в культуры подавали RPMI+0,2% о/о FBS, содержащей 1:5000 разведение ITS, активин А (100-200 нг/мл), и Wnt3a (50-100 нг/мл), или примерно на один день (время 0 до 1 дня) и дополнительный день, или в ту же самую среду, но без Wnt3a (день 1 - день 2). Ежедневно образцы клеток отбирали для выделения и анализа РНК. После двух дней дифференцировки в культуры вносили RPMI+0,2% о/о FBS, содержащую 1:1000 разведение ITS, KGF (или FGF7, 25 нг/мл), и ингибитор TGFβ №4 (2,5 мкМ) на один день (или 24 часа, день 2 - день 3). На следующие два дня (день 3 - день 5) в агрегатные суспензии iPS клеток вносили те же самые коктейли из среды и факторов роста, за тем исключением, что ингибитор TGFβ удаляли из среды для культивирования. Вновь, образцы клеток отбирали для выделения РНК в конце этой стадии (стадия 2, или день 5). Для стадии 3 (день 5 - день 8) среду для культивирования клеток заменяли на DMEM+1% о/о В27 добавки, содержащей TTNPB [4-[Е-2-(5,6,7,8-тетрагидро-5,5,8,8-тетраметил-2-нафталенил)-1-пропенил] бензойную кислоту] (3 нМ), KAAD-циклопамин (0,25 мкМ) и ноггин (50 нг/мл). Вновь, клеточные образцы отбирали для выделения и анализа РНК в конце этой стадии (стадия 3, день 8). Для стадии 4 (день 8 - день 14) среды меняли на DMEM+1% о/о В27 добавку, содержащую ноггин (50 нг/мл), KGF (50 нг/мл) и EGF (50 нг/мл). Вновь, клеточные образцы отбирали для выделения и анализа РНК в конце стадии 4 (или дня 14).
ПЦР в режиме реального времени проводили для определения экспрессии гена для различных маркерных генов в течение курса дифференцировки. Экспрессия генов специфических маркеров вначале нормализовалась до среднего уровня экспрессии конститутивных генов, экспрессии циклофилина G и ТАТА-связывающего белка (ТВР). Нормализованную относительную экспрессию генов затем отображали на диаграммах относительно уровня экспрессии в недифференцированных iPS клетках, и показывали кратность возрастания различных маркеров дифференцировки. Для ОСТ4 экспрессия генов была нормализована для установки самого низкого образца в наборе данных (день 14), и таким образом, представляющего кратность снижения в ходе дифференцировки.
Фигуры 2A-L являются диаграммами, демонстрирующими относительную экспрессию идентифицированного гена (например, Oct4, Brachyury, Cer1, GSC, FOXA2, FOXA1, HNF6, PDX1, PTF1A, NKX6.1, NGN3 и INS) относительно уровня экспрессии того же самого гена в недифференцированных iPS клетках. Уровень экспрессии генов был нормализован для установки конститутивных генов (контроль), а сравнение уровня экспрессии генов в два различных момента времени показало, имеется ли повышающая или понижающая регуляция гена или маркера экспрессии. Для ОСТ4 (Фиг. 2А) экспрессия гена была нормализована, и образец с самым низким уровнем экспрессии был установлен на 1 (день 14). Таким образом, как показано на Фиг. 2, относительные уровни экспрессии ОСТ4 представляют кратность понижающей регуляции (ось Y) в течение курса дифференцировки (ось X, стадия 0-4, или день 0 - день 14).
Как показано на фигуре 2А, ОСТ4 (POU5F1) экспрессируется на высоких уровнях в недифференцированных iPS клетках, и проявляет последующую понижающую регуляцию в ходе дифференцировки (день 0 - день 14). В день 14 уровни экспрессии ОСТ4 более чем в 3000 раз снижаются по сравнению с уровнями экспрессии, наблюдавшимися в недифференцированных клетках. Напротив, имеется временная повышающая регуляция экспрессии гена brachyury (BRACHYURY, Фиг. 2В) в течение первых 2 дней (день 1 и день 2). Временная повышающая регуляция гена brachyury была результатом направленной дифференцировки плюрипотентных/ iPS клеток в мезоэнтодермы путем применения активина А и wnt3a. Мезоэнтодерма была дополнительно дифференцирована в дефинитивную энтодерму во время дней 2-3 путем длительного воздействия активина А, на что указывала повышающая регуляция CER1, GSC и FOXA2 в конце стадии 1 в день 3 (Фиг. 2С-Е). Во время стадии 2 дефинитивную энтодерму дальше подвергали направленной дифференцировке в энтодерму кишечной трубки, как показано повышающей регуляцией FOXA1, сохранением экспрессии FOXA2 и понижающей регуляцией CER1 и GSC на день 6 дифференцировки (Фиг. 2С-Р). Во время стадии 3, при воздействии ретиноида, циклопамина и ноггина, энтодерму кишечной трубки далее подвергали направленной дифференцировке в энтодерму передней части пищеварительного тракта/ PDX1-экспрессирующую энтодерму, на что указывала повышающая регуляция HNF6 и PDX1 в день 8 (Фиг. 2G-H). Во время стадии 4, при воздействии KGF и EGF, энтодерма передней части пищеварительного тракта/ PDX1-экспрессирующая энтодерма дополнительно направлялась к дифференцировке в панкреатические прогениторы, эндокринные прогениторы, и гормон-экспрессирующие эндокринные клетки, на что указывала повышающая регуляция PTF1A, NKX6-1, NGN3 и INS в день 14 (Фигура 2I-L).
Пример 2. Ингибиторы Rho-киназы стимулируют рост, выживание, пролиферацию и адгезию iPS клеток
Способы дифференцировки различных hES и iPS клеточных линий были по существу такими же, как описано в настоящей заявке и в Примере 1. В дополнение к условиям культивирования, как описано для стадий 1, 2, 3, 4 и 5, ингибитор апоптоза и/или ингибитор Rho-киназы или ROCK добавляли в среды для культивирования для повышения и активизации роста, выживания, пролиферации и адгезии клеток во время дифференцировки. Как правило, примерно 10 мкМ ингибитора Rho-киназы, например, Y-27632, добавляли в клеточные культуры на каждой из стадий. Альтернативно, ингибитор Rho-киназы добавляли по меньшей мере на стадиях 1 и 2 и стадиях 4 и 5, или любой их комбинации. Морфология и профили экспрессии генетических маркеров суспензионных (агрегатных) культур дифференцированных iPS клеток были по существу схожими с суспензионными клеточными культурами, полученными из hES клеток.
На Фигурах 3 и 4 показаны результаты иммуноцитохимии (ICC) iPS клеточных культур со стадий 4 и 5, соответственно. На Фигуре 3 показаны клеточные агрегаты со Стадии 4, экспрессирующие типичные генетические маркера, характерные для PDX1-позитивной панкреатической энтодермы (также обозначаемой как панкреатический эпителий или панкреатические прогениторы), включая PDX1/ NKX6.1 со-позитивные клетки. Хотя это не показано на Фиг. 3, на Стадии 4 клетки не экспрессируют гормон-секретирующих белковых или генетических маркеров, более типичных для клеток Стадии 5, таких как инсулин (INS), глюкагон (GCG), соматостатин (SST) и панкреатический полипептид (РР). На Фигуре 4 показаны клеточные агрегаты гормон-экспрессирующих клеток со Стадии 5. Результаты ICC были дополнительно подтверждены с применением QPCR. Однако поскольку QPCR является общим популяционным исследованием общего уровня РНК, экспрессируемой в образце или клеточной культуре, нельзя определенно показать, что любая клетка экспрессирует множество маркеров.
Пример 3. Инкапсулирование панкреатических прогениторов, полученных из IPS
На сегодняшний день описаны способы продукции IPS клеток и источники для продукции IPS клеток. Однако нет достаточного описания дифференцировки каких-либо IPS клеток в какие-либо дифференцированные клетки для потенциального применения в клеточной терапии для лечения конкретного заболевания, например, диабета.
Чтобы определить, действительно ли культуры PDX1-позитвной панкреатической энтодермы или культуры панкреатических прогениторных клеток со Стадии 4, полученные из человеческих iPS клеток, полностью способны к развитию и созреванию in vivo в чувствительные к глюкозе инсулин-секретирующие клетки, популяции панкреатических прогениторов, по существу, как описано в Примерах 1 и 2, загружали в макро-инкапсулирующие устройства, по существу подобные тем, которые описаны в заявке США №12/618,659, озаглавленной «Инкапсулирование панкреатических клеточных линий, полученных из человеческих плюрипотентных стволовых клеток», поданной 13 ноября 2009; и патентах США №№7,534,608 и 7,695,965, озаглавленных «Способы получения панкреатических гормонов». Вкратце, примерно 5-10-20 мкл осажденных под действием гравитации агрегатов клеточных суспензий загружали в каждое устройство, содержащее по существу примерно 3 х 106 клеток.
Инкапсулированные клетки в устройстве затем готовили к имплантации млекопитающему, например, мыши с иммунной недостаточностью SCID/Bg, но могли имплантировать более крупным животным, включая крыс, свиней, обезьян или людей. Способы имплантации инкапсулированных клеток и устройства являются по существу такими, как описано в патентной заявке США №12/618,659, патентах США №№7,534,608 и 7,695,965, включая панкреатические прогениторные клетки, имплантированные на GELFOAM матрикс и введенные под эпидидимальную жировую ткань (EFP).
Для данных исследований не была нужна иммуносупрессия, однако иммуносупрессия может требоваться для некоторых млекопитающих в начальный переходный период, пока прогениторы внутри устройства не созреют полностью и не станут отвечать на глюкозу. У некоторых млекопитающих режим иммуносупрессии может занимать примерно 1, 2, 3, 4, 5, 6 или больше недель, и будет зависеть от млекопитающего.
Трансплантированным клеткам позволяли дифференцироваться и дополнительно созревать in vivo. Чтобы определить, действительно ли трансплантированные клетки обладают нормальной физиологической функцией, например, как натуральные бета-клетки, нужно определить уровни человеческого инсулина путем тестирования уровней человеческого С-пептида. Человеческий С-пептид отщепляется или процессируется из человеческого проинсулина, таким образом, специфическое определение человеческого С-пептида, а не эндогенного С-пептида указывает на секрецию инсулина из пересаженных (экзогенных) клеток. У животных с имплантатами определяли уровни человеческого С-пептида примерно каждые две, три или четыре недели путем инъекции болюса аргинина или глюкозы, предпочтительно глюкозы. Созревшие бета-клетки (полученные из дифференцированных плюрипотентных iPS клеток) должны быть физиологически функциональными и чувствительными к глюкозе, подобно натуральным или эндогенным бета-клеткам. Как правило, количества человеческого С-пептида выше 50 пМ или среднего базового (порогового) уровня, являются индикатором функции бета-клеток из трансплантированных прогениторов.
Подобно описанным в Kroon и др. (2008) выше, заявке США №12/618,659, патентах США №№7,534,608; 7,695,965 и 7,993,920, инкапсулированные панкреатические прогениторы, полученные из hIPS клеток, как ожидается, созревают в функционирующие кластеры панкреатических островков, содержащие эндокринные, ацинарные и дуктальные клетки, подобно натуральным островкам. Также предполагается, что очищенные или обогащенные панкреатические прогениторы, полученные из hIPS клеток перед трансплантацией, также созревают и развиваются в функциональные панкреатические островки и продуцируют инсулин in vivo. Некоторые варианты осуществления для очистки и обогащения различных дифференцированных клеточных популяций описаны более подробно в заявке США №12/107,020, озаглавленной «Способы очистки эндодермальных и панкреатических эндодермальных клеток, полученных из hES клеток», поданной 8 апреля 2008, теперь патенте США 8,338,170. Также предполагается, что панкреатические прогениторы, полученные из hIPS клеток, которые были подвергнуты криоконсервации, могут оттаивать и адаптировать в культуре перед трансплантацией, и созревать и продуцировать инсулин in vivo, соответственно. Гипогликемия может быть облегчена у животных с индуцированным диабетом, которым трансплантировали панкреатические прогениторы, полученные из hIPS клеток.
В итоге, полностью инкапсулированные панкреатические прогениторные клетки, полученные из hIPS клеток в макро-инкапсулирующем устройстве, созревают в физиологически функциональные панкреатические островки, и как ожидается, продуцируют инсулин в ответ на глюкозу in vivo.
Пример 4. Композиции панкреатических прогениторных и гормон-секретирующих клеток
Популяции дифференцированных hIPS клеток анализировали с применением проточной цитометрии для определения содержания клеток PDX1-позитивной панкреатической энтодермы или панкреатических прогениторных клеток (на стадии 4); и эндокринных клеток или эндокринных клеток-предшественников (на стадии 5), как показано в Таблицах 6а, 6b и 7, соответственно. В таблице 6b представлены те же данные, что и в Таблице 6а, но представлены подобно данным из Таблицы 10 для сравнения. Популяции из таблицы 6а перекрываются, например, общее число клеток превышает 100%, поскольку общее число PDX1+ и NKX6.1+ перекрывается с NKX6.1+/PDX1+/CHGA- клеточными популяциями (5-я колонка из Таблицы 6а). Таблица 6b включает только данные PDX1+ и тройные негативные, что не показано в Таблице 6а. Некоторые из этих иПЭК трансплантатов, а также другие, с применением по существу схожих рецептур, имплантировали животным для определения функции in vivo, однако, уровни человеческого сывороточного С-пептида были недостаточно стойкими для какой-либо потенциальной терапевтической цели (данные не показаны). Показанные значения являются процентами от общих клеток, принадлежащих к данной популяции. Ряд панкреатических прогениторов (NKX6.1(+) /PDX1(+) /Хромогранин А(-)) и очень маленькая популяция NKX6.1+/PDX1-/CHGA- находятся в агрегатах клеточных суспензий, что согласуется с тем, что наблюдалось в агрегатах суспензий панкреатических прогениторных клеток, полученных из hES клеток и агрегировавших на стадии ESC, как описано в заявке США №12/264,760, озаглавленной «Композиции агрегатных суспензий стволовых клеток, и способы их дифференцировки», поданной 4 ноября 2008. Уровни эндокринных клеток и/или эндокринных клеток-предшественников также по существу согласовались с полученными в клеточных культурах, полученных из hES клеток из заявки США 12/107,020, озаглавленной «Способы очистки клеток энтодермы и клеток панкреатической энтодермы, полученных из hES клеток», поданной 8 апреля 2008. Подобно агрегатам суспензий, полученных из hES клеток, изменение концентраций различных факторов роста в среде для культивирования на определенных стадиях дифференцировки (например, стадии 4), должно повышать и/или снижать определенные популяции клеток панкреатической энтодермы, эндокринных клеток, клеток PDX1-позитивной энтодермы или не-панкреатических типов клеток.



Пример 5. Тирозинкиназы ПЭК рецептора
Вышеописанные примеры по существу подобны тем, что описаны в Таблице 7 ниже, адаптированным из Schulz и др., (2012), выше. Эти и другие способы, описанные в настоящей заявке, можно найти во многих патентных и не-патентных публикациях заявителя, включая патенты США №№7,964,402; 8,211,699; 8,334,138; 8,008,07; и 8,153,429.

hESC Agg.: агрегаты hESC; XF НА: DMEM/F12, содержащая GlutaMAX, с добавлением 10% о/о Xeno-free KnockOut заменителя сыворотки, 1% о/о не-незаменимых аминокислот, 0,1 мМ 2-меркаптоэтанола, 1% о/о пенициллина/стрептомицина (все от Life Technologies), 10 нг/мл херегулина-1β (Peprotech) и 10 нг/мл активина A (R&D Systems); SP: StemPro® hESC SFM (Life Technologies); r0.2FBS: RPMI 1640 (Mediatech); 0,2% FBS (HyClone), 1x GlutaMAX-1 (Life Technologies), 1% о/о пенициллина/стрептомицина; ITS: Инсулин-трансферрин-селен (Life Technologies), разбавленный 1:5000 или 1:1000; A100: 100 нг/мл рекомбинантного человеческого активина A (R&D Systems); W50: 50 нг/мл рекомбинантного мышиного Wnt3A (R&D Systems); K25: 25 нг/мл рекомбинантного человеческого KGF (R&D Systems); IV: 2,5 мкМ TGF-β RI киназы ингибитора IV (EMD Bioscience); db: DMEM HI глюкоза (HyClone) с добавлением 0,5х В-27 добавки (Life Technologies), 1x GlutaMAX, и 1% о/о пенициллина/стрептомицина; СТТ3: 0,25 мкМ KAAD-циклопамина (Toronto Research Chemicals) и 3 нМ TTNPB (Sigma-Aldrich); N50: 50 нг/мл рекомбинантного человеческого ноггина (R&D Systems); K50: 50 нг/мл рекомбинантного человеческого KGF (R&D Systems); Е50: 50 нг/мл рекомбинантного человеческого EGF (R&D Systems); без подачи: указано, что в клетки не вносили повторно среду в указанный день; db, DMEM (с высоким содержанием глюкозы).
Когда вышеуказанные способы применяли к iPS клеткам и панкреатическим прогениторам, трансплантированным животным, автор заявки не смог впоследствии получть ту же самую устойчивую функцию in vivo, по сравнению с тем, когда те же самые способы применяли к hESC и панкреатическим прогениторам, полученным из hES. Это было удивительно, поскольку iPS клетки являются человеческими плюрипотентными стволовыми клетками, которые имеют морфологию и характеристики экспрессии генов hESC, и могут формировать как эмбриоидные тельца in vitro, так и тератомы in vivo, что указывает, что они могут формировать клетки из всех трех зародышевых слоев. См., по меньшей мере, например, Yu и др. (2007); патентную публикацию США №2009/0047263, Международную патентную публикацию №WO2005/80598; патентную публикацию США №2008/0233610; и Международную патентную публикацию №WO2008/11882, выше. Эти ссылки описывают, что iPS клетки удовлетворяют критериям определения для ESC. Таким образом, ожидается, что iPS клетки могут заменять ESC в протоколе дифференцировки in vitro для получения панкреатических прогениторных клеток, полученных из hES, которые дальше созревают и развиваются в полностью функциональные чувствительные к глюкозе клетки in vivo. Однако, с учетом не согласующихся данных по функциональности in vivo с применением вышеуказанных способов, автор заявки предложил использовать рецептуру сред для дифференцировки, уникальную для панкреатических прогениторов и/или панкреатических эндодермальных клеток (ПЭК), т.е. клеток, полученных на стадии 4 из hiPSC (или «иПЭК»), которые способны к обеспечению по существу схожих устойчивых уровней функции in vivo, которая впоследствии наблюдалась для ПЭК, полученных из hESC.
Авторы настоящей заявки ранее сообщали, что эндокринные (CHGA+) клетки, присутствующие в ПЭК, являются полигормональными эндокринными клетками, и не являются субпопуляцией клеток ПЭК, которые дают начало островкам, содержащим чувствительные к глюкозе инсулин-секретирующие клетки in vivo. См. Kelly et. al. (2011) выше. Скорее, они являются популяцией не-эндокринных клеток (CHGA- клеток), особенно тех, которые совместно экспрессируют NKX6.1 и PDX-1, которые считаются ПЭК, которые действительно дают начало функциональным островкам in vivo. Таким образом, авторы заявки исследовали, действительно ли модуляция, изменение или сдвиг взаимных соотношений эндокринных и не-эндокринных субпопуляций может влиять впоследствии на функцию in vivo.
Предыдущие попытки расшифровки передачи сигналов рецептор-лиганд в hESC привели к успешной идентификации факторов роста, стимулирующих самообновление и обеспечивающих разработку определенных условий среды для культивирования. См. Wang et al (2007) выше. Wang и др. идентифицировали херегулин-1β в качестве лиганда, связывающегося с ERBB3 и индуцирующего димеризацию с ERBB2 для влияния на самообновление hESC в этом контексте. ERBB является рецепторной тирозинкиназой (RTK), a RTK являются широко экспрессируемыми мембранными белками, которые действуют в качестве рецепторов для факторов роста и других экстраклеточных сигнальных молекул. При связывании лиганда они подвергаются фосфорилированию тирозина в специфических участках в цитоплазматическом хвосте и выделяют сигнальный каскад для связывания других белковых субстратов, вовлеченных в RTK-опосредованную трансдукцию сигнала. Функция RTK в некоторых процессах развития, включая регуляцию выживания клеток, пролиферацию и подвижность, и ее роль в образовании рака хорошо документирована. Также известно, что рецепторы ERBB тирозинкиназы экспрессируются на протяжении развития фетальной человеческой поджелудочной железы, хотя специфическая роль некоторых ERBB рецепторов и их лигандов неизвестна. См. Mari-Anne Huotari и др. (2002) «ERRB Signaling Regulates Lineage Determination of Developing Pancreatic Islet Cells in Embryonic Organ Culture», Endocrinology 143(11): 4437-4446 («ERRB сигналинг регулирует дифференцировку линий при развитии клеток панкреатических островков в эмбриональной органной культуре»).
Благодаря роли ERBB RTK сигналинга в самообновлении плюрипотентных стволовых клеток и их экспрессии в фетальной человеческой поджелудочной железе, как показано Wang и др. (2007) выше, и ERBB RTK экспрессии в человеческой фетальной поджелудочной железе, авторы настоящей заявки стали исследовать потенциальную активацию RTK in vitro в клетках панкреатической энтодермы (ПЭК), полученных из hES, в попытке идентифицировать рецепторы и лиганды, которые могут улучшать специализацию ПЭК во время дифференцировки, или экспансию путем стимуляции самообновления, или каких-либо других неизвестных механизмов, которые стимулируют созревание до физиологически функциональных островковых гормон-секретирующих клеток in vivo. ПЭК генерировали в суспензии в дифференцированных агрегатах, по существу как описано в Таблице 8, за исключением следующих модификаций.
Четыре образца ПЭК генерировали для RTK блоттинг-анализа. «Равновесный» образец ПЭК агрегатов в db-N50 K50 Е50 собирали в конце стадии 4 (или d13). «Обедненный» образец представляли d12 ПЭК агрегаты с добавлением db (DMEM с высоким содержанием глюкозы или DMEM с высоким содержанием глюкозы с добавлением 0,5х В-27 добавки (Life Technologies)) среды по отдельности (без факторов роста), и собирали в d13. Два «активированных» образца питали и культивировали в средах db на d12, затем на d13 питали либо db-K50 Е50 средами, либо db средами, содержащими 2% FBS, в течение 15 минут до сбора. Такие условия были предназначены для выявления RTK, которые были активными в условиях стадии 4, и чей ответ мог быть выявлен с импульсом KGF, EGF и инсулина (присутствовавшего в добавке В27), или сыворотке. Активация сывороткой предназначалась в качестве стимула факторов роста широкого спектра действия, потенциально идентифицирующего RTK, которые присутствовали на ПЭК и могли быть активированы, но не стимулированы представленными условиями стадии 4.
Анализ RTK проводили по существу так, как описано ранее в Wang et al, (2007) выше. Вкратце, Proteome Profiler™ человеческие фосфо-RTK матрицы антител (R&D Systems) применяли в соответствии с инструкциями производителя. Белковые лизаты готовили в 1% NP-40, 20 мМ Трис-HCl (рН 8,0), 137 мМ NaCl, 10% глицерине, 2,0 мМ ЭДТА, 1,0 мМ натрия ортованадата, 10 мкг/мл апротинина, и 10 мкг/мл лейпептина. 50 мкг свежих белковых лизатов инкубировали в течение ночи с нитроцеллюлозными мембранами, на которые наносили двойные пятна с 42 анти-RTK антителами и 5 негативными контрольными антителами, а также 8 анти-фосфотирозин-позитивных контрольных пятен (Фигура 5А). Матричные антитела улавливали экстрацеллюлярные домены фосфорилированных и нефосфорилированных RTK, а связанные фосфо-RTK выявляли с универсальным анти-фосфо-тирозиновым антителом, конъюгированным с пероксидазой хрена (HRP) с применением хемилюминесценции. См. фигуру 5 для размещения RTK матрицы, а также Таблицу 9 внизу для перечисления RTK в матрице.
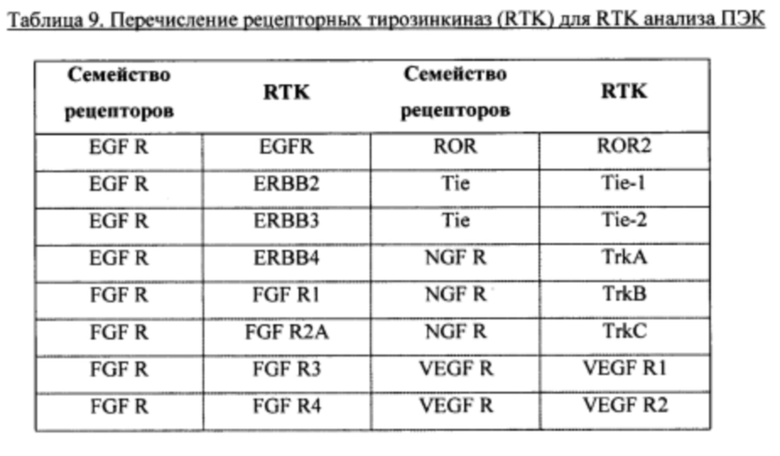

Анализ пятен RTK (Фигура 6А) показал, что инсулин- и IGF1-рецепторы (IR, IGF1R, соответственно) были фосфорилированы и активированы во всех условиях, подобно тому, что наблюдалось раньше с hESC. См. Wang et al (2007) выше. EGF рецептор (EGFR, также известный как ERBB1) был фосфорилирован в равновесных состояниях, которые ожидались с учетом присутствия EGF в среде на стадии 4. Действительно, низкоуровневое фосфорилирование ERBB2 наблюдалось как в равновесном состоянии, так и в обедненных условиях. Фосфорилирование и EGFR, и ERBB2 было повышено в каждом из активированных условий, что подтверждало способность анализа к выявлению активации в ответ на импульс лиганда. Фосфорилированный VEGFR3 был также выявлен во всех условиях, и был повышен в активированных образцах. Это позволило предположить, что ПЭК продуцируют эндогенный VEGFR3 лиганд, возможными кандидатами являются VEGF-C и D. Сывороточный стимул, как оказалось, активировал дополнительные рецепторы, включая низкие уровни ERBB3 фосфорилирования. Обнаружение фосфорилированного ERBB2/3 показывает, что херегулин-подобный член семейства EGF может активировать сигналинг в ПЭК. TIE-2 является одним из двух рецепторов ангиопоэтина, и как кажется, фосфорилируется в низких уровнях в ответ на сыворотку. Ангиопоэтин 1 и ангиопоэтин 4, как известно, являются активирующими лигандами TIE-2, в то время как ангиопоэтин 2 и ангиопоэтин 3 функционируют как контекстно-зависимые конкурентные антагонисты. HGF-рецептор (HGFR) также фосфорилировался в ответ на сывороточный стимул, что позволило предположить, что фактор роста гепатоцитов может также вызывать сигналинг в ПЭК. Наконец, в то время как был выявлен низкий уровень фосфорилирования эфрин В 2 RTK (ЕРНВ2), эфрин/Eph сигналинг является мембрано-связанной системой сигналинга между клетками, и вряд ли легко используется в дифференцировке ПЭК. ИнтеПЭКно, что ERBB4 не был фосфорилирован. Таким образом, анализ RTK выделил некоторые рецепторы, которые фосфорилировались в ПЭК, или могли быть фосфорилированы в ответ на различные условия, например, сыворотку. Эти результаты позволили предположить, что некоторые растворимые лиганды могут проявлять RTK сигналинг в ПЭК и возможно, воздействовать на клеточную пролиферацию, дифференцировку и/или специализацию, и таким образом, возможно, влиять на последующее созревание в функционирующие панкреатические островки in vivo.
Пример 6. Херегулин и FGF2 факторы роста влияют на ПЭК, полученные из композиций hESC
С учетом анализов RTK, которые показали, что некоторые RTK были активированы (или фосфорилированы) в определенных условиях, как описано выше в Примере 5, и поскольку было выявлено, что по меньшей мере ERBB2 и ERBB3 были активированы в ПЭК (спустя 13 дней дифференцировки из стадий 1-4), автор настоящей заявки решил определить эффект херегулина при применении для клеток на стадии 3 и 4.
Предварительные исследования проводили с применением херегулина и FGF. В некоторых из этих исследований включали ингибитор Rho-киназы, Y-27632. Эти предварительные исследования показали, что обработка плюрипотентных стволовых клеток в течение одного дня на стадии 1 с 10 нг/мл херегулина-1β (той же самой концентрацией и изомером херегулина, как описано в Wang и др. (2007)) повышала размер клеточных агрегатов, полученных их hES клеток в суспензионной культуре, по сравнению с размером агрегатов, полученных их hES клеток в суспензионной культуре без херегулина-1β (Hrg1β). Повышение размера клеточных агрегатов является предпочтительным в том, что приводит к более высокой клеточной массе для последующей имплантации и анализа функций у животных. Кроме того, размер диска агрегатов повышался при увеличении Hrg1β от 10 нг/мл до 50 нг/мл на стадии 3. Этот результат также наблюдался, когда 50 нг/мл другого фактора роста, FGF2, использовали на стадии 3, по сравнению с культурами при отсутствии FGF2. Повышение размера клеточных агрегатов также наблюдалось, когда культуры на стадии 3 подвергали дополнительным дням воздействия FGF2, например, 3 дня с 50 нг/мл FGF2, по сравнению с двумя днями.
В Таблице 10 приведено краткое изложение анализа проточной цитометрией ПЭК клеток, обработанных Hrg1β и FGF2 на стадии 3. Эндокринные клетки обозначались как CHGA позитивные (или CHGA+) клетки, а не-эндокринные клетки обозначались как CHGA негативные (или CHGA-) клетки. Эндокринные (CHGA+) и не-эндокринные клетки (CHGA-) могли позитивно окрашиваться на другие маркеры, например, позитивные на PDX1 и/или NKX6.1. Клетки, которые не окрашивались на какие-либо из тестируемых маркеров, обозначались как тройные негативные клетки или остальные клетки (CHGA-/NKX6.1-/PDX1).

Hg, Херегулин-β; FGF2, Фактор роста фибробластов 2; Hrg10, 10 нг/мл херегулина-1β; Hrg50, 50 нг/мл херегулина-1β; 2d FGF-50, 50 нг/мл FGF2 в течение 2 дней на стадии 3; 3d FGF2-50, 50 нг/мл FGF2 в течение 3 дней на стадии 3.
Чтобы определить, действительно ли повышение размера клеточных агрегатов влияет на субпопуляции ПЭК, композиции из ПЭК популяций анализировали проточной цитометрией. По сравнению с контрольными культурами, где не применяли ни Hrg1β, ни FGF2 для дифференцировки клеток, не-эндокринная субпопуляция ПЭК (CHGA-) повышалась от 54,01% до 61,2% при добавлении 10 нг/мл Hrg1β на стадии 3, и повышалась от 54,01% до 64,2% при добавлении 50 нг/мл Hrg1β на стадии 3. На эндокринную субпопуляцию (CHGA+) не оказывала существенного влияния обработка 10 нг/мл Hrg1β, тем более с 50 нг/мл. При этом относительные уровни остальных клеток снижались, и тем более с 50 нг/мл Hrg1β. Таким образом, повышение размера клеточных агрегатов при обработке Hrg1β связано главным образом с повышением не-эндокринных субпопуляций, относительно эндокринных и остальных субпопуляций.
Влияние FGF2 на культуры стадии 3 было схожим, но более выраженным, чем для Hrg1β. Например, не-эндокринная субпопуляция ПЭК (CHGA-) повышалась, как с Hrg1β. Основным эффектом FGF2 в этих культурах было существенное снижение эндокринной субпопуляции. В некоторых случаях эти клетки почти не выявлялись за 3 дня обработки (от 32,9% до 0,33%). Таким образом, повышение размера клеточных агрегатов для культур, обработанных FGF2, было главным образом связано с повышением неэндокринных, а в некоторых случаях остальных субпопуляций (от 13,1%) до 22,7% в течение 3 дней на стадии 3).
Таким образом, херегулин и/или FGF2, как кажется, играют роль в специализации клеток в популяциях ПЭК. Это было неожиданным с учетом того, что Wang et al (2007) выше сообщали, что херегулин по отдельности играет роль в обновлении клеток при использовании в среде с плюрипотентными стволовыми клетками.
Пример 7. Способы улучшения функции трансплантата из ПЭК in vivo путем обработки клеточных культур, полученных из iPSC, херегулином
Поскольку способы в соответствии с Таблицей 8 при применении к iPSC для получения иПЭК не обеспечивали устойчивой функции in vivo у животных, авторы настоящей заявки разработали другие способы для получения иПЭК. Изменения стандартного способа, указанного в Таблице 8, включают: оптимизацию числа пассажей iPSC; модуляцию уровней BMP сигналинга; модуляцию параметров агрегации суспензии iPSC во время экспансии и дифференцировки (например, скорости сдвига, скорости вращения, и тому подобное); оптимизацию концентраций, времени применения и продолжительности применения факторов роста, таких как Wnt, активин и ингибиторы rho-киназы; и обработку другими факторами роста на различных стадиях 1-4 протокола дифференцировки в качестве кандидатов для улучшения клеточной массы, пролиферации, дифференцировки, выживания и тому подобного (например, ERBB лиганды), но не ограничиваются ими. Многие повторные эксперименты были проведены по отдельности, или в комбинации, для определения того, какие способы дифференцировки для iPSC на стадиях 1-4 могут быть оптимизированы. Такие оптимизированные способы дифференцировки, продуцирующие иПЭК популяции, которые затем трансплантировали, привели к устойчивым чувствительным к глюкозе инсулин-секретирующим клеткам in vivo, подобным тем. что наблюдалось и сообщалось для hESC. В Таблице 11 ниже описаны базовые условия, с херегулином и без него, которые, как было показано, дифференцируют iPSC в иПЭК, которые позднее созревают в чувствительные к глюкозе островковые клетки in vivo. Базовые условия были подобны тем, которые описаны в Примерах 1, 2 и 5, а также в Таблице 8, за тем исключением, что херегулин добавляли на стадиях 3 и 4. Хотя использовали 30 нг/мл Hrg1β, пригодны концентрации в диапазоне от 10 нг/мл до 50 нг/мл, или даже больше 50 нг/мл. Кроме того, добавление ингибитора rho-киназы, Y-27632, поддерживали в дифференцирующих культурах, как описано в Примере 2.

iPSC Aggs: агрегаты iPSC; KSR: нокаутная сыворотка (Life Technologies); F10: 10 нг/мл bFGF (R&D Systems); A10: 10 нг/мл активина A (R&D Systems); A100: 100 нг/мл активина A; r0.2FBS: RPMI 1640 (Mediatech); 0,2% FBS (HyClone), 1x GlutaMAX-1 (Life Technologies), 1% о/о пенициллина/стрептомицина; ITS: Инсулин-трансферрин-селен (Life Technologies), разбавленный 1:5000 или 1:1000; A100: 100 нг/мл рекомбинантного человеческого активина A (R&D Systems); K25: 25 нг/мл рекомбинантного человеческого KGF (R&D Systems); СТТ3: 0,25 мкМ KAAD-циклопамина (Toronto Research Chemicals) и 3 нМ TTNPB (Sigma-Aldrich); N50: 50 нг/мл рекомбинантного человеческого ноггина (R&D Systems); K50: 50 нг/мл рекомбинантного человеческого KGF (R&D Systems); Е50: 50 нг/мл рекомбинантного человеческого EGF (R&D Systems); Y10: 10 мкМ Y-27632 (маточный раствор 20 мМ, 2000Х; Н30: 30 нг/мл херегулина (маточный раствор 100 мкг/мл); db, DMEM (с высоким содержанием глюкозы)
Для определения эффекта добавления херегулина, или херегулина и ингибитора rho-киназы в клеточные субпопуляции на стадии 3 и 4, иПЭК популяции анализировали проточной цитометрией. В Таблице 12 приведено краткое изложение анализа посредством проточной цитометрии различных популяций иПЭК с применением набора рецептур, приведенных в Таблице 11, а также таких рецептур, модифицированных путем повышения концентрации активина до 200 нг/мл. Кроме того, в Таблице 12 показаны общие условия, установленные для каждого набора экспериментов (базовые условия с херегулином или без него), и относительные проценты типов клеток в популяции иПЭК (эндокринных, не-эндокринных, только PDX1 и тройных негативных или остальных клеточных субпопуляций). В Таблице 12 также описаны данные, касающиеся функции in vivo клеток, полученных в каждом эксперименте.


BL, базовые условия; hIPSC, человеческие индуцированные плюрипотентные стволовые клетки; Hg30, 30 нг/мл херегулина-1l St 3+4, Стадии 3 и 4; hESC, человеческие эмбриональные стволовые клетки; CHGA, хромогранин А.
При определенных условиях отношение субпопуляций клеток в ПЭК (hESC, Е2354) и иПЭК (Е2314, Е2344, Е2347) популяциях изменялось. Например, иногда процент эндокринных (CHGA+) клеток снижался, а процент не-эндокринных клеток (CHGA-/NKX6.1+/PDX1+) повышался, по сравнению с базовыми условиями (без херегулина). Хотя кажется, что херегулин был ответственным за изменение пропорций эндокринных клеток относительно не-эндокринных клеток в этих ПЭК и иПЭК популяциях, в эксперименте #2380 (Е2380), уровень эндокринных (CHGA+) скорее повышался, чем снижался с добавлением херегулина.
Чтобы определить, действительно ли изменение в составе ПЭК и иПЭК популяций влияет на функцию in vivo, ПЭК и иПЭК трансплантаты из большинства экспериментов, описанных в Таблице 12, трансплантировали мышам подкожно, как описано ранее в другом патенте автора настоящей заявки и не-патентных публикациях, включая Schulz и др. (2012) и Kroon et al (2008), выше, и патенты США №№7,534,608; 7,695,965; 7,993,920 и 8,278,106, выше. Вкратце, ПЭК и иПЭК полностью инкапсулировали биодеградируемым полупроницаемым устройством для инкапсулирования клеток, некоторая часть которого включала микроперфорации. Устройства были произведены автором настоящей заявки и описаны подробно в патенте США №8,278,106, озаглавленном «Инкапсулирование панкреатических клеток из человеческих плюрипотентных стволовых клеток», поданной 13 ноября 2009. Анализы стимулированной глюкозой секреции инсулина (GSIS) проводили, начиная примерно 56 дней после имплантации. Кровь собирали до (натощак) и спустя 30 и/или 60 минут после применения глюкозы. Функцию трансплантата оценивали путем измерения концентраций С-пептида в сыворотке в ответ на применение глюкозы.
Количество человеческого С-пептида, высвобождаемого в сыворотку, является показателем количества высвобождаемого инсулина. С-пептид является коротким пептидом из 31 аминокислоты, соединяющим или связывающим А и В-цепи проинсулина и препроинсулина, который секретируется функциональными бета- или инсулин-секретирующими клетками. Как обсуждалось раньше Kroon и др. (2008) выше и другими, анализ человеческого С-пептида пригоден для оценки высвобождения инсулина, генерируемого de novo имплантированными клетками. Таким образом, уровни человеческого С-пептида в сыворотке этих животных являются мерой функции in vivo зрелых ПЭК и иПЭК трансплантатов. Человеческий С-пептид определялся в сыворотке по меньшей мере спустя 8 недель после имплантации. С дополнительными неделями после имплантации и натощак, уровни стимулированного глюкозой С-пептида повышались со сдвигом пиковых уровней С-пептида от 60 минут до 30 минут после применения глюкозы, что является показателем более быстрого ответа на глюкозную нагрузку, когда инсулиновые клетки созревают. У некоторых мышей не проявлялась функция, или их умерщвляли из-за плохого состояния здоровья; однако, эти мыши относились к группам, которые в остальном были животными с высоким проявлением функции, что, таким образом, скорее указывает на недостаточность приживления, чем на неспособность имплантированных клеток к дифференцировке и функционированию.
Фигуры 7А-С демонстрируют уровни человеческого С-пептида в сыворотке после применения глюкозы для всех экспериментов, указанных в таблице 12, за исключением Е2344. Фигуры 7А-С показывают, что по сравнению с контролями базового уровня, эти трансплантаты, полученные при обработке херегулином, в целом обеспечивали более высокие уровни сывороточного человеческого С-пептида. Например, на Фигуре 7А в эксперименте 2380 отмечалось примерно 5-кратное повышение (933 пМ: 200 пМ спустя 60 минут после применения глюкозы) для трансплантатов, полученных при обработке херегулином, по сравнению с трансплантатами, полученными без херегулина (базовый уровень). Херегулин, как кажется, оказывает меньшее влияние на ПЭК, полученные из hESC, поскольку эксперимент 2354 (Фиг. 7С) не показал более высоких уровней сывороточного С-пептида в трансплантатах, полученных при обработке херегулином, по сравнению с контролями базового уровня. Далее, при сравнении ПЭК, полученных из iPSC, иПЭК трансплантаты имели сопоставимую функцию in vivo с ПЭК трансплантатами (например, сравните Фиг. 7А и Фиг. 7В (iPSC трансплантаты) с Фиг. 7С (СуТ203 hESC). По существу, иПЭК трансплантаты являются такими же устойчивыми, как ПЭК трансплантаты. Кроме того, взаимные отношения эндокринных и не-эндокринных клеток, которые, как кажется, влияют на некоторые популяции иПЭК (например, Е2314, Е2347 и Е2354), не влияют на функцию in vivo, поскольку иПЭК из Е2380, которые не имеют того же самого сдвига в эндокринной и не-эндокринной субпопуляции, также показали хорошую функцию (см. Фиг. 7А-С).
В дополнение к анализу стимулированной глюкозой секреции инсулина, трансплантаты зрелых иПЭК тестировали, чтобы определить, действительно ли они по отдельности способны поддержать эугликемию, подобно эугликемии, поддерживаемой ПЭК, полученными из hESC, если бета-клетки животного-хозяина разрушены. Это включало разрушение бета-клеток мыши, которой проводили имплантацию, с применением бета-клеточного токсина, стрептозотоцина (STZ), который проявляет большую цитотоксичность против мышиных бета-клеток, по сравнению с человеческими бета-клетками. Произвольные измерения глюкозы крови не-натощак проводили для каждой мыши до и после обработки STZ. При удалении иПЭК трансплантата на 13 день после обработки STZ гипергликемия восстанавливалась (отмечался подъем уровня глюкозы), что демонстрировало контроль гликемии иПЭК трансплантатом, а не собственной поджелудочной железой мыши (см. Фиг. 8А и Фиг. 8В).
Кроме того, выражен синергетический эффект, когда херегулин и ингибитор rho-киназы были обеспечены во время 1-4 стадий дифференцировки (см. Таблицу 11). Например, iPSC, обработанные херегулином на стадиях 3 и 4 без ингибитора rho-киназы, приводили к ощутимо меньшей клеточной массе, что делало имплантацию невозможной. Дополнительное доказательство синергизма херегулина и ингибитора rho-киназы было приведено в некоторых экспериментах, например, Е2356, Е2380, в то время как базовые условия с ингибитором rho-киназы по отдельности не обеспечивали устойчивой функции, как у трансплантата с ингибитором rho-киназы и херегулином (см. Фигуры 7А и В). Как кажется, обработка херегулином и ингибитором rho-киназы не была дополнительной, поскольку добавление херегулина по отдельности обеспечивало недостаточную клеточную массу для трансплантации, а добавление ингибитора rho-киназы по отдельности (базовые условия) вызывало плохую функцию in vivo. Таким образом, обеспечение херегулина по отдельности или ингибитора rho-киназы по отдельности по существу не является схожим с суммой эффектов двух комбинированных веществ. Таким образом, по отдельности ни один из них не обеспечивает устойчивого ответа на глюкозу in vivo, но при объединении они обеспечивают ответ на глюкозу, схожий с ответом клеток, полученных из hES клеток. Соответственно, представляется, что обеспечение и херегулина, и ингибитора rho-киназы является синергетическим, поскольку их комбинированный эффект превышает сумму этих эффектов каждого из них по отдельности. Таким образом, иПЭК, обработанные ингибитором rho-киназы и херегулином, созревают in vivo, проявляя стимулированную глюкозой секрецию инсулина, и являясь способными к сохранению эугликемии на мышиной модели диабета (см. Фиг. 7А-В и Фиг. 8А-В).
Функция ERBB требует связывания лиганда, димеризации рецепторов, и миграции рецептора. Вариабельность каждого процесса может привести к дифференциальной регуляции рецепторов и нисходящих сигналов, которые они контролируют.Например, различные ERBB лиганды связываются с ERBB рецепторами с различной аффинностью, таким образом, изменяя характеристики и динамику образования ERBB димеров. В Таблице 13 показано множество различных комбинаций лигандов и комплексов, связывающих рецепторы. Обзоры, касающиеся сложности этой системы, обеспечены Oda, и др. (2005) «А comprehensive pathway map of epidermal growth factor receptor signaling», Mol. Syst. Biol., 1 (2005) («Исчерпывающая карта пути сигналинга рецептора эпидермального фактора роста») и Lazzara и др. (2009) «Quantitative modeling perspectives on the ERBB system of cell regulatory processes», Experimental Cell Research 315(4):717-725 («Перспективы количественного модулирования на системе ERBB регуляторных клеточных процессов»).

ERRB рецепторные тирозинкиназы: ErbBl (также обозначаемый как Her1, или рецептор эпидермального фактора роста, EGFR); ErbB2 (также обозначаемый как рецептор человеческого эпидермального фактора роста, или Her2; или Neu); ErbB3 (также обозначаемый как Her3), ErbB4 (также обозначаемый как Her4), ERBB лиганды: EGF, эпидермальный фактор роста; TGFα, трансформирующий фактор роста α; HB-EGF, гепарин-связывающий EGF-подобный фактор роста; EPR, эпирегулин; EPG, эпиген; AR, амфирегулин, Hrg1, херегулин-1 или нейрегулин-1; Hrg2, херегулин-2 или нейрегулин-2; Hrg3, херегулин-3 или нейрегулин-3; Hrg4, херегулин-4 или нейрегулин-4; херегулин применяется взаимозаменяемо с нейрегулином.
Huotari и др. предположил, что нейрегулин-4 может модулировать относительные уровни субпопуляций эндокринных клеток путем повышения числа соматостатиновых (дельта) клеток при расходе глюкагоновых (альфа) клеток, и что нейрегулин-4 не влияет на отношение экзокринных (например, секретирующих амилазу) и эндокринных (например, секретирующих β-инсулин, α-глюкагон, δ-соматостатин, РР-панкреатический полипептид) клеток. Однако эти исследования выполняли путем инкубации нейрегулина 4 на тотальных органных культурах тканей, полученных от мышей в день Е12.5. Эти клеточные популяции после эксплантации у мышей были дифференцированы дальше клеточных популяций со стадии 3 (например, PDX1 негативной энтодермы передней части пищеварительного тракта) и/или стадии 4 (PDX1 позитивной энтодермы передней части пищеварительного тракта, описанных в настоящей заявке. Нейрегулин-4 связывает только ERBB4 RTK, так что только эндокринная субпопуляция тотальной органной культуры мыши может быть модулирована нейрегулином-4 в этом контексте. Таким образом, не ожидается, что обработка на стадии 3 (PDX1 негативной энтодермы передней части пищеварительного тракта) и/или стадии 4 (PDX1 позитивной энтодермы передней части пищеварительного тракта) клеток, как описано в настоящей заявке с лигандом, иным чем ERBB, например, Hrg1, будет модулировать относительные эндокринные субпопуляции как в Huotari, поскольку уже было показано, что Hrg1 связывается с ERBB3 и индуцирует димеризацию ERBB2/3. Однако, из-за низкоуровневого экспрессии ERBB2 и 3 в ПЭК, как показано на Фиг. 6, неясно, будут ли типы клеток на стадии 3 и 4 экспрессировать низкие или высокие уровни ERBB2 и 3 для связывания с Hrg1.
Далее, в другом контексте, автор настоящей заявки описали, что Hrg1 связывается ERRB 2/3 и стимулирует самообновление плюрипотентных стволовых клеток (см. Wang et al (2007)). Хотя возможно, что Hrg1 может действовать с той же самой емкостью в контексте стадии 3 и 4, автор настоящей заявки ранее описывал, что большая часть клеточной экспансии для продукции ПЭК осуществляется на стадии плюрипотентных стволовых клеток (стадия 0). Во время стадии 0 hESC выращивали, пассировали и подвергали экспансии в течение примерно двух (2) недель. Таким образом, большая часть клеточной экспансии или самообновления для обеспечения клеточной экспансии не происходит во время стадий 1-4. См. Schulz и др. (2012) выше. Также, если предположить, что ERRB2/3 присутствует во время стадий 3 и 4, можно ожидать, что херегулин будет проявлять тот же самый эффект, как с плюрипотентными стволовыми клетками (самообновление), в отличие от влияния на направленную дифференцировку. Различие в функции, как кажется, зависит от контекста, т.е. плюрипотентные стволовые клетки против клеточного типа энтодермы или панкреатической линии.
В целом, обеспечивая херегулин, или херегулин и ингибитор rho-киназы in vitro для клеток энтодермы передней части пищеварительного тракта (стадия 3) и PDX1 экспрессирующих клеток панкреатической энтодермы (конец стадии 3 и стадия 4), получают популяции ПЭК и иПЭК, которые после трансплантации созревают и развиваются в чувствительные к глюкозе инсулин-секретирующие клетки in vivo (см. Фигуры 7 и 8). Такое применение херегулина, или херегулина и ингибитора rho-киназы указано нами в первый раз. Такое применение и эффект не отличаются от ранее описанных в патентной или не-патентной литературе.
Пример 8. Активин подавляет экспрессию NGN3 и продукцию клеток, коммитированных в эндокринную линию, в культурах панкреатических эндодермальных клеток (ПЭК).
Панкреатические эндодермальные клетки, или «ПЭК» являются панкреатической популяцией клеток, включающей главным образом две различных субпопуляции: (i) субпопуляцию не-эндокринных мультипотентных панкреатических прогениторов (CHGA-) (далее «не-эндокринных (CHGA-)»), которые развиваются и созревают, давая начало чувствительным к глюкозе инсулин-секретирующим бета-клеткам in vivo; и (ii) субпопуляцию клеток, коммитированных в эндокринную линию (CHGA+) (далее «эндокринные (CHGA+)»), которые могут давать начало другим не-инсулин-секретирующим или островковым поддерживающим клеткам при созревании in vivo. Неэндокринная (CHGA-) субпопуляция является CHGA негативной, и преимущественно PDX1 и NKX6.1 позитивной, в то время как субпопуляция клеток, коммитированных в эндокринную линию, является CHGA позитивной и в основном PDX1 и NKX6.1 негативной. Для не-эндокринной (CHGA-) субпопуляции требуется примерно 8-12 недель или больше для развития и созревания в чувствительные к глюкозе инсулин-секретирующие бета-клетки в количествах, позволяющих проводить системное определение инсулинового ответа in vivo (с помощью ИФА на С-пептид). Однако требуется меньше 8 недель чувствительным к глюкозе клеткам для развития и созревания in vivo во время нормального эмбрионального развития. Таким образом, остается необходимым (1) получение популяции ПЭК с повышенной не-эндокринной (CHGA-) субпопуляцией и/или (2) истинной эндокринной (CHGA+) клеточной популяции, чувствительной к глюкозе in vitro и/или продвинутой в развитии популяции (например, адекватно специализированной и имеющей определенные сигнатурные маркеры, соответствующие тем, которые описаны для подобных панкреатических популяций in vivo) и/или прогениторной клеточной популяции, способной к сокращению и/или снижению продолжительности периода развития и созревания in vivo.
Что касается первой задачи, необходимо получить популяцию ПЭК, где неэндокринная (CHGA-) субпопуляция остается интактной, и в то же самое время дифференцировка в клетки, коммитированные в эндокринную линию (CHGA+) подавляется, подобно тому, что естественно происходит in vivo. В Примерах ниже описаны способы получения таких популяций ПЭК.
Считается, что при нормальном развитии in vivo нейрогенин-3 (NGN3), фактор транскрипции, является главным регулятором развития эндокринных клеток, и что экспрессия NGN3 не возникает после повышающей регуляции PDX1 и NKX6.1, например, в клетках панкреатической эндодермальной линии in vivo. См. Rukstalis and Habener (2009), «Neurogenin3: A master regulator of pancreatic islet differentiation and regeneration», Islets 1(3): 177-184 («Нейрогенин-3: главный регулятор дифференцировки и регенерации панкреатических островков»). В соответствии с Таблицей 8, на стадии 3 ретиноевую кислоту (RA) или аналог ретиноида, 4-[(E)-2-(5,6,7,8-тетрагидро-5,5,8,8-тетраметил-2-нафталенил)-1-пропенил]бензойную кислоту или «TTNPB» (или «ТТ3» в Таблице 8) применяют для индукции экспрессии PDX1. Однако в то же самое время ретиноевая кислота также преждевременно индуцирует экспрессию NGN3, которая инициирует эндокринную специализацию. Таким образом, супрессия, подавление или ингибирование NGN3 во время стадии 3 и/или стадии 4 может обеспечить один подход для минимизации клеток, коммитированных в субпопуляцию эндокринной линии (CHGA+) в ПЭК и/или получения эндокринных клеток на более поздних стадиях развития in vitro и/или in vivo. Важно, что любой супрессор, репрессор или ингибитор NGN3 должен в то же самое время не супрессировать, не репрессировать и не ингибировать не-эндокринные (CHGA-) клетки, и предпочтительно, повышать эту субпопуляцию во время продукции ПЭК in vitro.
Исследовали способность различных факторов роста к супрессии генов, таких как NGN3, на стадиях 3 и 4. Одним вероятным фактором роста является активин, который выполняет разнообразные функции при экспансии и дифференцировке стволовых клеток. Например, при низких уровнях (например, 10 нг/мл) активин способствует поддержанию плюрипотентности плюрипотентных стволовых клеток и пролиферации стволовых клеток. При более высоких уровнях (например, 100 нг/мл) активин действует в качестве фактора роста и дифференцировки, ответственного за продукцию дефинитивной энтодермы на стадии 1 (см., например, Таблицу 8). Для определения его влияния на репрессию клеток, коммитированных в эндокринную линию (CHGA+) при формировании ПЭК, автор заявки вначале тестировал активин в средней концентрации примерно 50 нг/мл при стандартной адгезивной дифференцировке. На этом уровне активин был способен к супрессии экспрессии NGN3, без супрессии экспрессии генов PDX1 и NKX6.1 в ПЭК (сравни Фиг. 9D-Е с Фиг. 9А-В). Однако когда тот же самый уровень активина использовали с более масштабируемой суспензионной культурой клеточных агрегатов во время стадии 3, общая клеточная масса резко снижалась, и в некоторых случаях клеточные агрегаты исчезали (сравни контроль с агрегатами клеток, обработанными активином, на Фиг. 10). Подобные уменьшения клеточных агрегатов наблюдались для активина при 25 нг/мл с 50 нг/мл (данные не показаны).
С учетом этих результатов, более низкие концентрации активина (например, 5 нг/мл и 10 нг/мл) тестировали на стадиях 3 и 4. Как показано на Фиг. 11, при более низких концентрациях активина клеточные агрегаты были способными сохранять клеточную массу и выход, по сравнению с контролем (сравни Фиг. 11В-С на 8 день и Фиг. 11B-D на 12 день с Фиг. 11А, контролем, на 8 и 12 день). Таким образом, уровни активина менее примерно 25 нг/мл пригодны для сохранения клеточной массы и выхода. Автор заявки затем провел анализ проточной цитометрией этих клеточных культур, чтобы определить, действительно ли активин при низких уровнях на стадиях 1 и 4 оказывает влияние на композицию клеточных агрегатов; см. Таблицу 14. В Таблице 14 показано, что низкие уровни активина не снижают продукцию не-эндокринной (CHGA-) субпопуляции, а когда обработку проводят на обеих стадиях 3 и 4, это может фактически увеличивать эту субпопуляцию (см. Таблицу 14, А5, результаты для стадий 3 и 4). Однако кажется, что дифференцировка в клетки, коммитированные в эндокринную линию (CHGA+) все еще индуцируется в значительной степени с очень схожими процентами эндокринных (CHGA+) клеток, с активином и без него (см. Таблицу 13, колонку по эндокринным клеткам). Таким образом, хотя активин может супрессировать или ингибировать экспрессию NGN3 и NKX2.2, как наблюдалось при Q-PCR, при более низких уровнях (позволяющих поддерживать клеточную массу), он не снижает субпопуляцию, коммитированную в эндокринную линию (CHGA+) при анализе в конце стадии 4. Предпочтительно, снижаются только PDX1 и тройные негативные субпопуляции, по сравнению с контролем, в частности, когда активин применяют на стадиях 3 и 4.

Пример 9. Активин, херегулин и WNT подавляют продукцию клеток, коммитированных в эндокринную линию в культурах ПЭК
Пример 8 показывает, что умеренные уровни активина (например, 50 нг/мл) во время стадий 3 и 4 были способны к супрессии или репрессии экспрессии NGN3 (Фиг. 9), но те же самые уровни не поддерживали клеточную массу. Более низкие уровни активина (например, 5 и 10 нг/мл) были способны к поддержанию клеточной массы (Фиг. 11), но были недостаточными для продолжения репрессии эндокринной дифференцировки во время стадии 4, поскольку процент клеток эндокринной линии (CHGA+) оставался по существу схожим с контролем (Таблица 14, колонка CHGA+) и с тем, что описано в предыдущих Примерах. Таким образом, кажется, что подавление дифференцировки в клетки, коммитированные в эндокринную линию (CHGA+), в первую очередь во время стадии 3 на этих низких уровнях является недостаточным, поскольку предполагается, что продолжение супрессии дифференцировки этой субпопуляции во время стадии 4 все еще необходимо. См. Rukstalis и др. (2009) выше. Таким образом, были исследованы дополнительные факторы роста, способные к предотвращению потери клеток и выхода (т.е. противостоящие эффекту средних и высоких уровней активина) при одновременном отсутствии влияния на продукцию субпопуляций не-эндокринных мультипотентных панкреатических прогениторов и супрессию дифференцировки клеток, коммитированных в эндокринную линию (CHGA+).
С учетом вышеуказанных примеров, описывающих херегулин в дозе 30 нг/мл во время стадий 3 и 4, часто меняющих относительные уровни эндокринной (CHGA+) и неэндокринной (CHGA-) субпопуляций ПЭК, автор заявки проанализировал, какие эффекты, снижающие концентрации херегулина, возможны при объединении с активином, в отношении продукции ПЭК.
Плюрипотентные стволовые клетки дифференцировали с применением стандартного протокола в соответствии с Таблицей 8, за исключением стадии 3, активин включали в количестве 10 нг/мл или 20 нг/мл, в комбинации с херегулином в количестве 2 нг/мл или 10 нг/мл; а во время стадии 4 активин снижали до 5 нг/мл с херегулином до 1 нг/мл в течение двух дней, затем с 10 нг/мл. Как показано на Фиг. 12 (день 10; изображения на верхней панели), активин в количестве 10 нг/мл и херегулин 2 нг/мл показали некоторое снижение клеток, по сравнению с контролем (сравни Фиг. 12А и В, верхняя панель). Повышение активина до 20 нг/мл с херегулином, остающимся на уровне 2 нг/мл, усиливало потерю клеток (сравни Фиг. 12В и С, верхняя панель). Однако было возможно предотвращение большей потери клеток при более высоких концентрациях активина (20 нг/мл) путем повышения херегулина до 10 нг/мл при уровне активина 20 нг/мл (сравни Фиг. 12D и С, верхняя панель). Таким образом, когда уровни херегулина являются слишком низкими, наблюдается значительная потеря клеток, но когда уровни херегулина повышаются, например, по меньшей мере до 20 нг/мл, имеется изменение в процентом содержании не-эндокринных клеток (CHGA-), по сравнению с субпопуляциями эндокринной линии (CHGA+)
Количественный ПЦР анализ проводили для определения относительных уровней экспрессии генов в дни 8, 10 и 12 дифференцировки с вышеуказанными комбинациями активина и херегулина (Фиг. 13). Подобно тому, что показано в Примере 8 (Фиг. 9), активин на уровне 10 и 20 нг/мл был способен к репрессии NGN3 и NKX2.2 (другого маркера эндокринной дифференцировки) без уменьшения экспрессии NKX6.1 (сравни Фиг. 13А и В с С). Тот факт, что экспрессия NKX6.1 остается высокой, указывает, что продукция не-эндокринной (CHGA-) субпопуляции остается неизменной, или на нее не оказывается влияние. В частности, однако, при 20 нг/мл активина и 10 нг/мл херегулина, или при 10 нг/мл активина и 2 нг/мл херегулина, активин-зависимая потеря клеток, которая наблюдалась раньше, когда активин применяли на уровне 25 нг/мл и 50 нг/мл (Фиг. 10), была снижена (Фиг. 12D и Е, верхняя панель). Таким образом, херегулин, как кажется, эффективно противодействует потере клеток, исходно наблюдаемой при использовании активина на стадии 3. И хотя остается некоторая индукция NGN3 и NKX2.2 (т.е. они уже не так высоко репрессируются, как это наблюдалось при использовании 50 нг/мл активина на стадиях 3 и 4 при адгезивной дифференцировке в Примере 8, Фиг. 9), она существенно снижалась, по сравнению со стандартными условиями (сравни контрольные условия и условия А20Н10 с Фиг. 13А и В).
Хотя на уровне выше 20 нг/мл активин может дальше подавлять маркеры NGN3 и NKX2.2, и таким образом, еще больше подавлять эндокринную дифференцировку, более высокие уровни херегулина затем потребуются для поддержки клеточной массы для смещения эффекта дозы активина при высоких концентрациях. Еще более высокие уровни херегулина, как наблюдалось в вышеуказанных примерах, могут изменять относительную продукцию эндокринной (CHGA+) и не-эндокринной (CHGA-) субпопуляций РЕС. Тем не менее, влияние более высоких уровней активина на репрессию экспрессии NGN3 еще необходимо. Таким образом, авторы заявки изучили дополнительные факторы роста или морфогены, которые могут влиять на образование передней или задней энтодермы, и контролировать другие аспекты развития, включая WNT, FGF, PDGF, ретиноевую кислоту (RA или TTNPB), BMP, хэджхог, EGF, IGF и тому подобные, но не ограничиваясь ими.
В частности, влияние на каноничные пути передачи сигналов Wnt исследовали путем добавления Wnt3A в среду дифференцировки с активином и херегулином на стадиях 3 и/или 4. Вновь, какая-либо комбинация требовалась для супрессии дифференцировки клеток, коммитированных в эндокринную линию (CHGA+), как наблюдалось по репрессии NGN3 и NKX2.2 экспрессии и сохранению хорошей клеточной массы (т.е. ограниченной потере клеточной массы);в то же самое время не наблюдалось влияния на не-эндокринную субпопуляцию мультипотентных панкреатических прогениторов. Одной такой исследованной смесью было 50 нг/мл Wnt вместе с 10 нг/мл активина и 2 нг/мл херегулина на стадии 3; и как указано выше, на стадии 4, 5 нг/мл активина и 1 нг/мл херегулина в течение двух (2) дней, впоследствии 5 нг/мл активина и 10 нг/мл херегулина. Это привело к увеличению клеточной массы (ПЭК), по сравнению с применением активина и херегулина в тех же самых концентрациях по отдельности (сравни Фиг. 12D и Е, верхняя панель), и еще большей клеточной массе, по сравнению с контролем (сравни Фиг. 12А и Е, верхняя панель). QPCR анализ клеточных агрегатов в этих условиях (например, при комбинации активина, херегулина и Wnt на стадии 3, и активина и херегулина на стадии 4) показал, что отмечалась значительная супрессия клеток, коммитированных для дифференцировки в эндокринную линию (CHGA+), как наблюдалось по снижению экспрессии NGN3 и NKX2.2 на стадиях 3 и 4, в частности, в дни 8, 10 и 12 (Фиг. 13А и В). Далее, экспрессия NKX6.1, исходно репрессированная на 8 день, была существенно повышена на 10 и 12 дни (стадия 4) (Фиг. 13С). Таким образом, развитие не-эндокринной (CHGA-) субпопуляции не нарушалось с добавлением Wnt, и дифференцировка клеток, коммитированных в эндокринную линию (CHGA+), как кажется, замедлялась или подавлялась, и не было явного снижения клеточной массы.
Для определения клеточного состава ПЭК популяции с комбинацией активина, херегулина и Wnt во время стадии 3 проводили проточную цитометрию после стадии 4, и клеточный состав ПЭК показан в таблице 14. Как показано, комбинация факторов роста (активина, херегулина и Wnt, или «AHW») эффективно снижала дифференцировку клеток, коммитированных в эндокринную линию (CHGA+), как наблюдалось по снижению процента этой субпопуляции (15,6 против 58,8) при повышении не-эндокринной (CHGA-) субпопуляции (58,1 против 23,4), по сравнению с контрольными клеточными культурами при стандартном протоколе (Таблица 8). Данные проточной цитометрии поддерживают и согласуются с данными Q-PCR о том, что комбинация трех факторов (AHW) в целом замедляет или подавляет дифференцировку клеток, коммитированных в эндокринную линию (CHGA+) в ПЭК, без влияния на продукцию не-эндокринной (CHGA-) субпопуляции. Замедленная экспрессия этих генов in vitro согласуется с установлением (или замедлением) развития поджелудочной железы in vivo, в то время как экспрессия NGN3 в эндокринной дифференцировке происходит после установления экспрессии PDX1 и NKX6.1 в не-эндокринной (CHGA-) субпопуляции. Кроме того, не нарушалась масса и выход ПЭК.
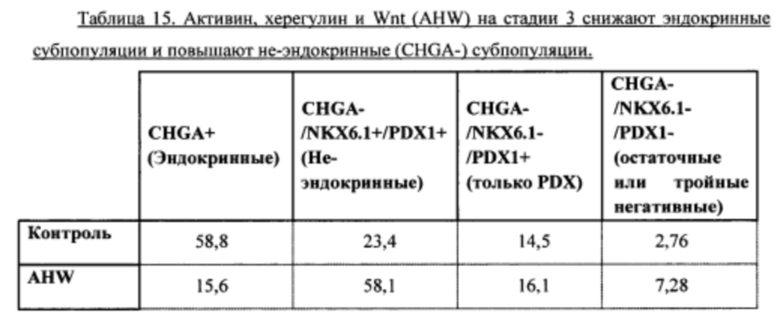
С учетом результатов с применением комбинации активина, херегулина и Wnt, уровни активина вновь титровали на стадии 3 для установления диапазона концентраций активина, которые можно применять. Тестируемые концентрации активина были в диапазоне примерно от 0, 25, 50, 75 до 100 нг/мл с 50 нг/мл Wnt и 5 нг/мл херегулина на стадии 3. Активин уменьшали до 5 нг/мл, херегулин до 5 нг/мл, a Wnt не включали на стадии 4. Получали изображения клеток в конце стадии 4 (день 13), показанные на Фиг. 12 (нижняя панель). Потеря клеток вновь была явной при более высоких концентрациях активина, например, от 75 до 100 нг/мл (Фиг. 12D и Е, нижняя панель). Важно, что включение Wnt на стадии 3 позволяло применять гораздо более высокие концентрации активина на стадии 3, которые в ином случае были невозможными из-за потери клеток.
Для анализа эффекта продолжения воздействия активина и херегулина на стадии 4, в контексте активина, херегулина и Wnt на стадии 3, человеческие ESC дифференцировали с применением трех условий: (i) стандартный протокол (см. таблицу 8; или Протокол №1 в Таблице 17); (ii) комбинация активина (75 нг/мл), херегулина (5 нг/мл) и Wnt (50 нг/мл на стадии 3, с последующими стандартными факторами на стадии 4; и (iii) комбинация активина (75 нг/мл), херегулина (5 нг/мл) и Wnt (50 нг/мл) на стадии 3, с последующим дополнительным активином (5 нг/мл) и херегулином (5 нг/мл) на стадии 4, см. Таблицу 17. Проточную цитометрию проводили после стадии 4 для всех трех условий, и клеточный состав ПЭК показан в Таблице 17. AHW на стадии 3 снижали клетки, коммитированные в эндокринную линию, и повышали содержание не-эндокринной (CHGA-) субпопуляции, по сравнению с контролем (21,8 против 42,2 для клеток, коммитированных в субпопуляцию эндокринной линии; и 40,1 против 27,6 для неэндокринной (CHGA-) субпопуляции), а дополнительная обработка АН на стадии 4 дополнительно уменьшала клетки, коммитированные в эндокринную линию, и дополнительно повышала содержание не-эндокринной (CHGA-) субпопуляции (6,46 против 21,8 для клеток, коммитированных в эндокринную линию; и 72,7 против 40,1 для субпопуляции не-эндокринных (CHGA-) прогениторов).

Пример 10. Трансплантаты ПЭК, полученные с применением комбинации активина, херегулина и Wnt, обладают улучшенной функцией чувствительности к глюкозе in vivo
В Примере 9 показано, что на основе морфологии (например, микроскопических изображений малого увеличения для дифференцированных клеточных агрегатов), данных Q-PCR и проточной цитометрии, можно предположить, что комбинация активина, херегулина и Wnt («AHW») позволяет эффективно подавлять клетки, коммитированные для дифференцировки в эндокринную линию (CHGA+), как наблюдалось по репрессии экспрессии NGN3 и NKX2.2, поддерживать или повышать не-эндокринные (CHGA-) субпопуляции, в то же время не оказывая отрицательного влияния, или улучшая клеточную массу или выход. Чтобы определить, действительно или эти изменения оказывают влияние на окончательное развитие и созревание чувствительных к глюкозе инсулин-секретирующих клеток, агрегаты ПЭК, полученные с AHW, имплантировали животным, в сочетании с ПЭК, полученными по стандартному протоколу в соответствии с Таблицей 8 или Протоколу №1 из Таблицы 17.
В частности, человеческие плюрипотентные стволовые клетки дифференцировали с применением комбинации этих трех факторов (AHW) на стадии 3, а затем активина и херегулина по отдельности на стадии 4. Контрольные ПЭК готовили по существу в соответствии с Таблицей 8. Клеточные агрегаты анализировали посредством QPCR в дни 8, 10, 12 и 14, как показано на Фиг. 14. В ПЭК, полученных с применением контрольных и модифицированных AHW протоколов, наблюдалась экспрессия NKX6.1 (Фиг. 14А), хотя отмечалась наиболее стойкая экспрессия NKX6.1 с применением AHW на 14 день (после стадии 4). В то же самое время, дифференцировка клеток, коммитированных в эндокринную линию (CHGA+), существенно подавлялась с протоколом AHW, как наблюдалось по репрессии экспрессии NGN3, особенно на день 8, по сравнению с контрольным протоколом (Фиг. 14В).
Агрегаты ПЭК от обоих протоколов загружали в запатентованные автором заявки имплантируемые полупроницаемые инкапсулирующие устройства на 14 день дифференцировки, и имплантировали SCID-beige мышам, по существу, как описано раньше (n=5 животных для каждого из AHW и контрольного способов, всего n=10). На 11 неделе после имплантации ПЭК, полученные с применением AHW протокола, продуцировали в среднем около 900 пМ С-пептида спустя один час (60') после нагрузки глюкозой, в то время как контрольные ПЭК вырабатывали в среднем около 400 пМ С-пептида. См. Фиг. 15, сравнивающую AHW животных с контрольными. В частности, на 11 неделе после имплантации у 3 из 5 мышей с AHW имплантатами отмечено в среднем более 1200 пМ человеческого С-пептида (Фиг. 15, животные №№3514, 3516 и 3518), в то время как у контрольных животных уровни С-пептида были значительно ниже. Действительно, только у 1 из 5 контрольных животных отмечены уровни, близкие к 600 пМ (Фиг. 15, животное №3524). Таким образом, на основе уровней С-пептида, на 11 неделе после имплантации протокол AHW был способен к продукции ПЭК популяции по меньшей мере с двукратным улучшением функции in vivo, по результатам измерения уровня человеческого С-пептида после стимуляции глюкозой. Это, возможно, коррелирует с более быстрым временем созревания.
На 14 неделе после имплантации AHW ПЭК имплантаты продолжали превосходить контрольные имплантаты, с более быстрым ответом секреции инсулина, поскольку значения С-пептида спустя 30 минут после глюкозной нагрузки для AHW ПЭК имплантатов были выше, чем значения спустя 60 минут после нагрузки для контрольных имплантатов (Фиг. 16). В частности, у 3 из 4 животных с AHW ПЭК имплантатами отмечалось в среднем более 3500 пМ С-пептида спустя 30 минут после глюкозной нагрузки, по сравнению с 3 контрольными животными, у которых отмечалось в среднем примерно 2300 пМ, но спустя 60 минут после глюкозной нагрузки (Фиг. 16). Эти данные показывают, что дифференцировка с применением комбинации этих трех факторов роста (AHW) позволяет получить популяцию ПЭК, которая лучше функционирует (т.е. повышает сывороточные уровни С-пептида спустя 30 минут после глюкозной нагрузки, что указывает на более быструю чувствительность к глюкозе для имплантированных ПЭК, полученных из AHW) или потенциально является более активной, чем ПЭК, полученные по стандартному протоколу, в те же самые изучаемые моменты времени. Не ограничивая настоящую заявку какой-либо теорией, считается, что повышенная активность AHW ПЭК имплантатов связана с тем фактом, что комбинация AHW (1) замедляет или подавляет дифференцировку клеток, коммитированных в эндокринную линию (CHGA+) в ПЭК, (2) увеличивает субпопуляцию не-эндокринных мультипотентных панкреатических прогениторов в ПЭК, и (3) поддерживает хорошую клеточную массу.
Пример 11. Культуры панкреатических эндокринных клеток in vitro
В примерах 9 и 10 описано спланированное замедление, репрессия, супрессия или ингибирование дифференцировки клеток, коммитированных в эндокринную линию (CHGA+), на стадиях 3 и 4. В попытке получить продвинутые в развитии эндокринные клетки-предшественники или эндокринные клетки in vitro, авторы заявки тестировали различные способы на стадиях 5, 6 и/или 7. Например, авторы заявки анализировали, какие факторы роста, по отдельности или в комбинации, способны к стимуляции или индукции эндокринной дифференцировки, т.е. индукции экспрессии NGN3.
В Таблице 17 внизу обобщены различные способы продукции эндокринных клеток вместе со стандартным протоколом получения ПЭК (Протокол №1) и протоколом, используемым для получения ПЭК (Протокол №2). В таблице 17, Протокол №3 указано число факторов роста в специфические дни и стадии, однако, авторы заявки тестировали, и специалисту в данной области техники понятно, что не все факторы роста, по отдельности или в комбинации, нужно использовать в указанный определенный день/стадию. Таким образом, в Таблице 17 показано, что многие циклы (например, факторов роста, продолжительности и тому подобного) протестированы авторами настоящей заявки.

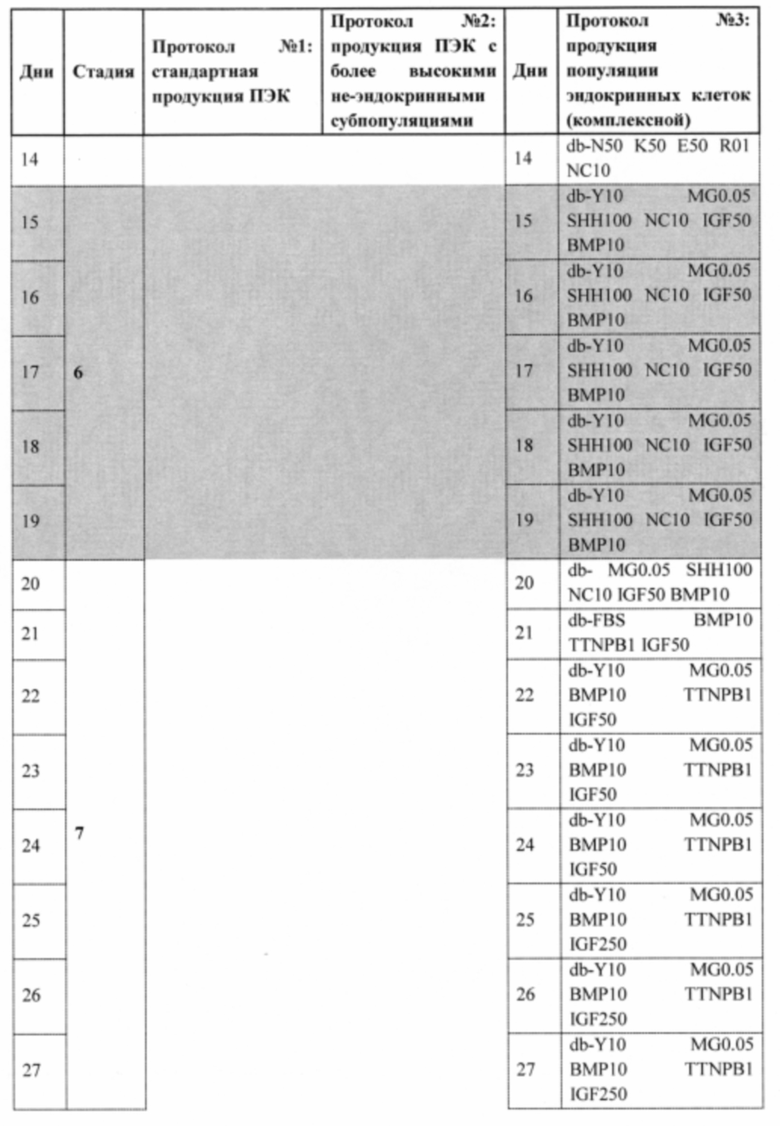

hESC Agg.: hESC агрегаты; XF HA: DMEM/F12 содержащий GlutaMAX, с добавлением 10 об. % Xeno-free KnockOut заменителя сыворотки, 1 об. % не незаменимых аминокислот, 0,1 мМ 2-меркаптоэтанол, 1 об. % пенициллин/стрептомицин (все от Life Technologies), 10 нг/мл херегулина-1b (Peprotech) и 10 нг/мл активина A (R&D Systems); SP: StemPro® hESC SFM (Life Technologies); r0.2FBS: RPMI 1640 (Mediatech); 0.2% FBS (HyClone), 1x GlutaMAX-1 (Life Technologies), 1 об. % v/v пенициллин/стрептомицин; cb: CMRL: CMRL 1066, 1x Glutamax, 1 об. % пенициллин/стрептомицин, 2% B-27; db: DMEM Hi глюкоза (HyClone) с добавлением 0,5x B-27 добавки (Life Technologies), 1x GlutaMAX, и 1 об. % пенициллин/стрептомицин; A100, A50, A5: 100 нг/мл, 50 нг/мл, 5 нг/мл рекомбинантного человеческого активина A (R&D Systems); ВМР20, ВМР10: 20 нг/мл, 10 нг/мл ВМР4 (Peprotech); СТТ3: 0,25 мМ KAAD-циклопамин (Toronto Research Chemicals) и 3 нМ TTNPB (Sigma-Aldrich); Е50: 50 нг/мл рекомбинантного человеческого EGF (R&D Systems); Н10, Н5: 10 нг/мл, 5 нг/мл херегулина 1b; IGF25: 25 нг/мл IGF2 (Peprotech); ITS: инсулин-трансферрин-селен (Life Technologies), разбавленный 1:5000 или 1:1000; IV: 2,5 мМ TGF-b RI ингибитора киназы IV (EMD Bioscience); K50, K25: 50 нг/мл, 25 нг/мл рекомбинантный человеческий KGF (R&D Systems, или Peprotech); MG0.05: 0,05% MATRIGEL (BD Biosciences); N50: 50 нг/мл рекомбинантный человеческий Noggin (R&D Systems); NC10: 10 мМ никотинамид; PDGF10: 10 нг/мл тромбоцитарный фактор роста (PDGF, R&D Systems); RO1: ингибитор гамма-секретазы, RO4929097, 1 мМ; SHH100: 100 нг/мл sonic hedgehog; TTNPB1: 1 нМ TTNPB (Sigma-Aldrich); W50: 50 нг/мл рекомбинантного мышиного Wnt3A (R&D Systems); Y10: 10 мМ Y-27632 (Tocris Bioscience).
Ранее авторы настоящей заявки раскрыли применение ингибитора гамма-секретазы для получения эндокринных прогениторов/предшественников в патенте США №8,129,182, «Эндокринные прогениторы/клетки-предшественники, панкреатические гормон-экспрессирующие клетки и способы их получения», изданном 6 марта 2012. Как применяется и заявлено в патенте '182, для дальнейшей индукции дифференцировки PDX1-позитивных панкреатических эндодермальных клеток по направлению к эндокринной линии, были описаны факторы роста или агенты, ингибирующие сигнальный путь Notch. Это достигалось путем применения ингибитора гамма-секретазы (например, DAPT или N-[N-(3,5-дифторфенацетил)-L-аланил]-S-фенилглицин t-бутилового эфира или родственных аналогов). Ингибиторы гамма-секретазы блокируют интрамембранное расщепление молекулы Notch, таким образом, предотвращая высвобождение активированного внутриклеточного домена Notch. Применение на 2-4 день ингибитора гамма-секретазы DAPT, либо на последние дни добавления ретиноевой кислоты, либо немедленно после удаления ретиноевой кислоты, использовали для индукции экспрессии NGN3. Здесь использовали другой ингибитор гамма-секретазы RO44929097 во время стадии 5. См. Таблицу 17.
Как указано в Таблице 17, для направления дифференцировки, и/или созревания эндокринной линии во время стадий 6 и 7 применяли комбинацию факторов роста, включая ВМР4 («ВМР»), Sonic Hedgehog («SHH»), тромбоцитарного фактора роста («PDGF»), FGF2 («FGF»), ретиноевой кислоты и/или аналогов ретиноевой кислоты, таких как TTNPB («TTNPB»), инсулиноподобного фактора роста I (IGF-I или «IGF1») и II (IGF-II или «IGF2»), производного витамина В3, никотинамида («NC»), или комбинацию одного или нескольких из этих факторов роста, но не ограничиваясь ими. Одна такая комбинация описана в Таблице 17, Протокол №3.
Например, ПЭК, полученные из протокола AHW, по существу, как описано выше в Примере 10, дополнительно обрабатывали на стадии 5 (дни 13 и 14) ингибитором гамма-секретазы RO4929097 («R01»), как правило, в течение 2 дней при 1 мкМ, в соответствии с Таблицей 17, Протокол №3. Эта обработка на стадии 5 приводила к индукции экспрессии NGN3, а затем к эндокринной дифференцировке. Кроме того, никотинамид включали во время стадий 5 и 6 также для индукции экспрессии NGN3 или эндокринной дифференцировки. Экспрессию гена NGN3 анализировали посредством Nanostring анализа мРНК после стадии 4 (13 день культивирования), после стадии 5 (15 день культивирования), и спустя 2 дня на стадии 6. Как показано на Фиг. 17, ингибитор гамма-секретазы сильно индуцирует экспрессию NGN3 после стадии 5 на 15 день, по сравнению с низким уровнем экспрессии NGN3 в конце стадии 4 (день 13). После удаления ингибитора гамма-секретазы на 15 день (первый день стадии 6), экспрессия NGN3 снижалась, как наблюдалось на 17 день (Фиг. 17). Это временное повышение экспрессии NGN3 во время стадии 5 согласуется с временной экспрессией, наблюдаемой при развитии бета-клеток in vivo. См. Jorgensen и др. (2007) см. выше.
Отмечаем, что общее число дней для любой из стадий может варьировать и варьирует, например, стадия 5, 6 и/или 7 может быть сокращена или продлена по количеству дней. Это также применяется к более ранним стадиям 1, 2, 3 и 4.
Кроме того, ингибитор rho-киназы может быть добавлен в среду для дифференцировки непрерывно или периодически во время любой из стадий 1, 2, 3, 4, 5, 6 и 7 для стимуляции выживания клеток и клеточной массы и выхода. Ингибитор rho-киназы на стадиях 1, 2 и 4, например, как кажется, стимулирует индуцированную дифференцировку плюрипотентных стволовых клеток, например, путем поддержки клеточной массы и выхода.
Также, примерно на 20 день (т.е. на стыке стадий 6 и 7 из Протокола №3 из таблицы 17), клеточные агрегаты могут диссоциировать и реагрегировать. Эта диссоциация и реагрегация удаляет некоторые субпопуляции клеток, таким образом, оставляя преимущественно популяцию эндокринных клеток и/или эндокринных прогениторов/предшественников (CHGA+) клеток. Эта физическая манипуляция также облегчает анализ эндокринных популяций, и составляет этап очистки, если необходимо. См. Пример 14 для подробностей.
Далее, начиная со стадии 6 и во время стадии 7, если необходимо, можно использовать очень низкую концентрацию, например, около 0,05% матригеля, чтобы способствовать сохранению клеточных агрегатов и/или предотвращению миграции эндокринных клеток из клеточных агрегатов. Применение матригеля или любых других комплексных клеточных матриксов может зависеть от самих специфических клеточных агрегатов (например, агрегатов ПЭК, агрегатов эндокринных прогениторов/клеток-предшественников, как описано ниже, и тому подобного), и возможно, от PSC линии, из которой получены клеточные агрегаты. Можно применять другие комплексные матриксы или определенные матриксы, например, сыворотку, ламинин, фибронектин. коллаген и другие белки экстрацеллюлярного матрикса, для выполнения по существу схожих функций. См. подробности в Примере 14.
Добавление инсулиноподобного фактора роста (IGF) 1 и/или 2 (IGF1 или IGF2) можно осуществить на стадии 6 или 7, или во время стадии 6, например, начиная на день 16, 17 или 18. Плюрипотентные стволовые клетки дифференцировали по существу так, как описано выше, в соответствии с Таблицей 17, за исключением IGF1 в дозе 50 или 200 нг/мл, или IGF2 в дозе 25 или 100 нг/мл, добавляемых в клеточные культуры на стадии 6, в частности, на день 17 примерно на 3 дня. В клеточных агрегатах анализировали мРНК с применением Nanostring после стадии 6 (день 20). Эти предварительные исследования показали, что IGF2 был более способным к индукции экспрессии INS, чем IGF1 (данные не показаны), однако, более длительная обработка могла вызвать более устойчивый эффект для IGF2 по отдельности, или для IGF1 в комбинации с IGF2.
В таблице 17 описано применение различных факторов роста в определенных концентрациях однако, изобретение не ограничено специфическими концентрациями, описанными в настоящей заявке, поскольку специалисту в данной области техники понятно, что изменения концентрации факторов роста являются стандартными в данной области техники, и фактически, описаны в вышеуказанных примерах, по меньшей мере с изменениями уровней активина и херегулина и их влияния на дифференцировку и состав клеток. Модификации в настоящей заявке включают 5, 10, 20, 50 - 100 нг/мл активина; 5, 25, 50 - 75 нг/мл Wnt; 2, 5, 10, 30 нг/мл херегулина; 10 - 200 нг/мл sonic hedgehog (SHH); 10 - 100 нг/мл PDGF; 10 - 20 нг/мл BMP; 2 - 20 нг/мл FGF2; и 25 - 200 нг/мл IGF1 и/или IGF2, но не ограничиваются ими. Кроме того, можно применять любые комбинации этих факторов, и стадия или период, когда их применяют, могут варьировать, даже значительно, не выходя за рамки раскрытого изобретения.
Пример 12. Функция панкреатических эндокринных клеток in vivo
На сегодняшний день обогащенные популяции эндокринных клеток, полученных in vitro из PSC, не дают начало чувствительным к глюкозе инсулин-секретирущим бета-клеткам in vivo. Таким образом, авторы настоящей заявки проверяли, действительно ли вышеуказанные способы в соответствии с таблицей 17 и описанные в Примере 11 выше, позволяют получить первые эндокринные клетки in vitro, чувствительные к глюкозе in vivo.
Человеческие плюрипотентные стволовые клетки дифференцировали в соответствии с Протоколом №3 из Таблицы 17 для стадий 1-5, однако, на стадиях 6 и 7 применяли только FBS, матригель и ингибитор rho-киназы, в дополнение к среде на основе DMEM с В-27 добавкой. Экспрессию РНК анализировали посредством Nanostring, с моментами времени в начале стадий 5 (д14) и 6 (д15), и в конце стадий 6 (д20) и 7 (д26). На Фигурах 18A-D и 19A-D показаны относительные уровни экспрессии мРНК для маркеров субпопуляции не-эндокринных мультипотентных панкреатических прогениторов, включая PDX1, NKX6.1, SOX9, и PTF1A, и маркеров эндокринных клеток, включая инсулин (INS), глюкагон (GCG), соматостатин (SST) и грелин (GHRL). В конце стадии 4 (день 13) отмечалась высокая экспрессия PDX1, SOX9, NKX6.1 и PTF1A. Все эти маркеры снижались во время стадии 5, когда осуществлялась эндокринная дифференцировка или индукция (день 15). В конце стадии 6 (день 20) или начале стадии 7 уровни мРНК PDX1, NKX6.1 и PTF1A снижались, при этом в то же самое время уровни мРНК INS, GCG, GHRL и SST повышались (сравните FIG.18A-D и F1G.19A-D на 20 день). В частности, снижение экспрессии PDX1 и PFT1A и резкое сопутствующее повышение относительной экспрессии мРНК INS, GCG, GHRL и SST на стадиях 5 и 6 обусловлено комбинацией активина, херегулина и Wnt (AHW), и в частности, активина на стадиях 3 и 4. То есть, включение активина на стадиях 3 и 4 подавляет эндокринную дифференцировку на стадиях 3 и 4, в то время как гамма-секретаза на стадии 5 индуцирует эндокринную дифференцировку на стадиях 5, 6 и 7. Схожие уровни экспрессии NKX6.1 на 20 и 26 дни (стадии 6 и 7), по сравнению с более ранними моментами времени, согласуются с экспрессией этого маркера в панкреатических эндокринных клетках, а также субпопуляциях не-эндокринных мультипотентных панкреатических прогениторов. Таким образом, на более поздних стадиях (например, стадиях 5, 6 и 7) клетки были направлены на эндокринную дифференцировку, в то время как они были супрессированы от этого на более ранних стадиях, стадиях 3 и 4.
После примерно стадии 6, клеточные агрегаты были диссоциированы и реагрегированы (более подробно описано ниже в Примере 14), что удаляло некоторые клеточные типы, полученные во время стадий 3 и 4, включая некоторые субпопуляции ПЭК, например, не-эндокринные (CHGA-) клетки. Удаление не-эндокринных (CHGA-) субпопуляций и/или экзокринных клеток и/или экзокринных прогениторов и/или протоковых клеток и/или протоковых прогениторов можно наблюдать по резкому снижению SOX9 и PTF1A после реагрегации (Фиг. 18, сравните день 20 и день 26). После стадии 7 эндокринные клетки загружали в устройства на день 26 и трансплантировали RAG2 мыши на 27 день, по существу как описано ранее для трансплантации (или имплантации) мышам SCID-Beige. На Фиг. 20 показаны сывороточные уровни человеческого С-пептида натощак и спустя один час (60 минут) после применения или нагрузки глюкозой. Спустя 10 недель после имплантации, у всех пяти животных продуцировался инсулин, как наблюдалось по человеческому сывороточному С-пептиду. Спустя 15 недель после имплантации (Фиг. 21) у всех животных проявлялись повышенные уровни продукции инсулина, по сравнению с уровнем спустя 10 недель после имплантации, и у 2 из 5 животных в среднем составили 1500 пМ С-пептида, что является доказательством устойчивой чувствительности к глюкозе in vivo.
Перед загрузкой эндокринных клеток в устройства на 26 день, отбирали образцы, чтобы убедиться, что функциональные бета-клетки развивались, созревали и появлялись из трансплантированных эндокринных клеток со стадий 6 и 7, а не из, например, субпопуляции остальных не-эндокринных мультипотентных панкреатических прогениторов. Проводили иммунохимических анализ образцов с применением эндокринных и не-эндокринных маркеров. Замороженные срезы образцов на 26 день совместно окрашивали антителами к DAPI (Фиг. 22А), NKX6.1 и хромогранину А (Фиг. 22В). Ясно, что подавляющее большинство NKX6.1 позитивных (красных) клеток также позитивно окрашивались на хромогранин (CHGA+), эндокринный маркер. Таким образом, кажется, что не-эндокринные мультипотентные панкреатические прогениторные клетки, как показано по отсутствию CHGA-/NKX6.1+ клеток, не присутствуют в каком-либо значительном количестве в этих препаратах эндокринных клеток.
Образцы этих агрегатов эндокринных клеток также совместно окрашены на NKX6.1 и PDX1 (Фиг. 23А-В). На Фиг. 23А-В показано, что большинство инсулин-секретирующих клеток также экспрессируют NKX6.1 и PDX1. Это неожиданный результат, учитывая, что в ранее опубликованных отчетах полученные in vitro инсулин-экспрессирующие клетки из PSC не проявляли совместной экспрессии NKX6.1 и/или PDX1. См. патент США 7,033,831 и родственные заявки от Geron Corporation and Jiang и др. (2007), «Generation of insulin-producing islet-like clusters from human embryonic stem cells», Stem Cells 2007 Aug; 25(8): 1940-53, опубл. 17 мая 2007 («Генерация инсулин-продуцирующих подобных островкам кластеров из человеческих эмбриональных стволовых клеток»).
Взятые вместе, эти данные обеспечивают неопровержимое доказательство того, что, прежде всего, полученная из PSC in vitro культура эндокринных клеток может (1) совместно экспрессировать INS, PDX1 и NKX6.1 in vitro и вероятно, является чувствительной к глюкозе in vitro; и (2) могут давать начало чувствительным к глюкозе инсулин-секретирующим клеткам in vivo.
Пример 13. Ретиноевая кислота, никотинамид и BMP повышают экспрессию PDX1 и NKX6.1 в эндокринных клетках
Признаком зрелых бета-клеток является повышающая регуляция NKX6.1 и PDX1, на уровне отдельных клеток, относительно уровней, наблюдаемых в не-эндокринных (CHGA-) субпопуляциях ПЭК. Примеры 11 и 12 описывают продукцию эндокринных клеток, где большинство клеток имеют уровни экспрессии NKX6.1 и PDX1, подобные уровням, наблюдаемым в не-эндокринной (CHGA-) субпопуляции ПЭК. Таким образом, необходимо идентифицировать факторы роста, которые специфически усиливают экспрессию по меньшей мере двух маркеров в инсулин-позитивных эндокринных клетках.
Два потенциальных фактора включают TTNPB, аналог ретиноевой кислоты или ретиноида, и ВМР4, член семейства костных морфогенетических белков, которое является частью суперсемейства трансформирующего фактора роста-бета. TTNPB и ВМР4 тестировали в различных комбинациях на стадиях 6 и 7 и в различных концентрациях, но типично в количестве 0,5 нМ и 10 нг/мл, соответственно. Как показано анализом мРНК Nanostring на Фиг. 24, BMP на стадиях 6 и 7, на стадии 7 отдельно, или в комбинации с TTNPB, может одновременно повышать экспрессию инсулина и PDX1 (Фиг. 24А и С). Напротив, TTNPB по отдельности на тех же самых стадиях (стадиях 6 и 7 и стадии 7 по отдельности), как кажется, не повышают экспрессию PDX1, даже слегка понижает экспрессию PDX1, по сравнению с BMP по отдельности (сравните Фиг. 24С, колонки 4 и 5, с колонками 2 и 3). Эффекты BMP в отношении экспрессии NKX6.1 являются менее явными, поскольку при всех условиях (BMP по отдельности, TTNPB по отдельности, BMP и TTNPB в комбинации, обработка на каждой из стадий 6 и 7, или на стадии 7 по отдельности), экспрессия NKX6.1 была относительно стабильной. В качестве дополнительного индикатора специфичности BMP, для BMP также наблюдалось повышающая регуляция экспрессии генов ингибиторов дифференцировки (ID) (например, ID1), который, как известно, регулируется членами суперсемейства TGF-β, включая ВМР4 (Фиг. 24D). Таким образом, не ограничивая эту заявку какой-либо теорией, кажется, что BMP является фактором, избирательно индуцирующим повышающую регуляцию PDX1, и ID1.
Чтобы определить, действительно ли некоторые их этих маркеров совместно экспрессируются в какой-либо одной клетке, агрегаты эндокринных клеток фиксировали на 27 день (после стадии 7) дифференцировки, и замороженные срезы готовили и окрашивали посредством иммуноцитохимических (ИЦХ) методик на С-пептид, NKX6.1 и PDX1. См. Фиг. 25-28. Здесь, вместо окрашивания с INS, клетки окрашивали с антителами против С-пептида, центрального фрагмента про-инсулина, и таким образом, это окрашивание также является показателем инсулин-экспрессирующих клеток (Фиг. 25). Результаты ИЦХ согласуются с данными Nanostring выше, т.е. что BMP по отдельности, или в комбинации с TTNPB, на стадиях 6 и 7 или только на стадии 7, был способен к индукции экспрессии PDX1 в инсулин- (С-пептид) -секретирующих клетках (Фиг. 25 и 26).
Несмотря на тот факт, что данные по мРНК позволили предположить, что BMP и TTNPB по отдельности или в комбинации оказывают слабое влияние на экспрессию NKX6.1, исследования ИЦХ показали, что на клеточном уровне совместная экспрессия NKX6.1 и С-пептид- (или инсулин-) экспрессирующих клеток является достаточно стойкой (Фиг. 27 и 28), т.е. NKX6.1 окраска является яркой, и также часто ярче, чем наблюдаемая в субпопуляции не-эндокринных мультипотентных панкреатических прогениторов. Таким образом, экспрессия NK.X6.1, как кажется, преимущественно совместно экспрессируется с С-пептид- (или инсулин-) экспрессирующими клетками. Объединенные данные мРНК (Nanostring) и белка (ИЦХ) также показали, что в определенном контексте может быть предел применения только одной методологии для исходной характеристики типа клеток. Таким образом, данные ИЦХ показали, что BMP по отдельности или в комбинации с TTNPB на стадиях 6 и 7 или стадии 7 отдельно был способен к повышению уровней PDX1 и NKX6.1 в С-пептид- (или инсулин-) экспрессирующих клетках (Фиг. 27 и 28). Наиболее сильный эффект наблюдался, когда применяли два фактора в комбинации, но BMP по отдельности также способен к этому. TTNPB может также независимо регулировать PDX1 и NKX6.1. Например, обработка TTNPB в течение нескольких дней, например, 2, 3 или 4 дней во время стадии 7 была способна к индукции экспрессии NKX6.1 без присутствия BMP (данные не показаны). Таким образом, применение этих факторов роста, в частности, TTNPB, может зависеть от времени.
Обращаясь специально к никотинамиду, на Фиг. 29 показано, что никотинамид в условиях культивирования на стадии 6 повышает экспрессию INS и снижает экспрессию GCG. приводя к 3-кратному повышению отношения экспрессии INS/GCG. Функция никотинамида является неясной, но отмечалось, что никотинамид предотвращает потерю бета-клеток у грызунов, связанную с применением стрептозотоцина, путем ингибирования PARP-1, поли(аденозин-трифосфат [АДФ]-рибозы) полимеразы-1, и путем предотвращения истощения НАД+. См. Masiello, Р. и др. (1998) «Experimental NIDDM Development of a New Model in Adult Rats Administered Streptozotocin and Nicotinamide», Diabetes, 47(2):224-9 («Экспериментальное развитие ИНСД на новой модели взрослых крыс с применением стрептозотоцина и никотинамида»). Никотинамид может также повышать эндокринную дифференцировку и содержание инсулина в островках плода человека. См. Otonkoski Т. и др. (1993), «Nicotinamide is a potent inducer of endocrine differentiation in cultured human fetal pancreatic cells». J Clin Invest 92:1459-1466 («Инсулин является мощным индуктором эндокринной дифференцировки в культивируемых панкреатических клетках плода человека»).
Вновь, зрелые бета-клетки in vivo экспрессируют высокие уровни PDX1 и NKX6.1, и высокие уровни PDX1 и NKX6.1 необходимы для надлежащей функции. Таким образом, BMP, и BMP в комбинации с TTNPB, путем индукции высокой экспрессии обоих маркеров в инсулин-позитивных клетках указывают, что впервые возможно получить эндокринные клетки, чувствительные к глюкозе, не только in vivo, но также in vitro.
Как упоминалось ранее, сроки для стадий 6 и 7 не являются жесткими, и могут быть сокращены, или даже продлены, и специалисту в данной области техники понятно, что 5 и 10 дней для стадии 6 и 7, соответственно, можно легко изменить; в частности, если применять по существу те же самые факторы роста, как описано в настоящей заявке и в соответствии с Таблицей 17.
Пример 14. Матригель по отдельности и в комбинации с ингибитором rho-киназы улучшает клеточную адгезию ПЭК и агрегатов эндокринных клеток
В Примере 11 выше клеточные агрегаты были диссоциированы и реагрегированы и реассоциированы между стадиями 6 и 7. Диссоциацию осуществляли с применением аккутазы в день 20 или около того, и реагрегировали в ротационной культуре, по существу, как описано в Schulz и др. выше. При реагрегации удалялись некоторые субпопуляции ПЭК, включая любые оставшиеся не-эндокринные (CHGA-) субпопуляции, а также другие не-эндокринные клетки, как описано более подробно в Kelly и др. (2011), выше. Эндокринные клетки обладают аффинностью друг к другу, однако это взаимодействие in vitro является относительно слабым, даже реагрегированные клетки на стадиях 6/7 были лишь свободно ассоциированными. Таким образом, проводили исследования для тестирования различных факторов и агентов на их способность к повышению взаимодействия между клетками в агрегатах эндокринных клеток.
Было установлено, что добавление Y-27632, ингибитора rho-киназы, и разбавленного матригеля (0,05 об. %) позволяет стимулировать более тесные ассоциации реагрегированных агрегатов эндокринных клеток. Комбинация Y-27632 и матригеля повышала, например, межклеточное взаимодействие в клеточных агрегатах, по сравнению с каждым из компонентов по отдельности.
Дифференцированные культуры получали по существу так, как описано в Примере 12, на 20 день диссоциировали аккутазой и реагрегировали в db с 5% FBS (более низкие концентрации FBS, т.е. 2% или 1%, также являются эффективными), и ДНК-азой. Y-27632 не включали в среду для реагрегации, поскольку его добавление приводило к включению не-эндокринной (CHGA-) субпопуляции, а также других не-эндокринных клеток во вновь сформированные агрегаты. На следующий день и каждый день после этого, клеточные агрегаты обрабатывали Y-27632 или матригелем, или обоими компонентами. Стереомикроскопические изображения получали спустя 1, 2 и 3 дня после агрегации, как показано на Фиг. 30. Фиг. 30 демонстрирует, что комбинация матригеля, Y-27632 и db-FBS была наиболее эффективной. Эта комбинация была более эффективной, чем применение db-FBS по отдельности, или db-FBS и матригеля, или db-FBS и Y-27632 (сравните Фиг. 30А-D). Клеточные агрегаты при использовании комбинированных агентов выглядели морфологически более плотными, чем все другие образцы (Фиг. 30D). Вероятно, что агенты улучшают контакт между клетками и взаимодействия между клетками и матриксом; хотя более поздние эксперименты при отсутствии FBS и только с матригелем и Y-27632 дали те же самые результаты. Эти более плотные клеточные агрегаты выглядят более морфологически схожими с тканью органа. Это предпочтительно, поскольку клеточные агрегаты могут лучше выдерживать стресс ротационной культуры, загрузку в устройства, обработку для трансплантации, имеют усовершенствованное развитие и созревание in vitro и in vivo, и так далее.
Пример 15. Отсутствие ноггина приводит к получению ПЭК, обогащенных субпопуляцией не-эндокринных мультипотентных прогениторов с ограниченным содержанием эндокринных клеток и без потребности во фракционировании или очистке популяции ПЭК.
Как обсуждалось выше, необходима продукция популяции ПЭК (стадия 4), высоко обогащенной субпопуляцией не-эндокринных мультипотентных панкреатических прогениторов (CHGA-), но относительно бедной клетками, коммитированными в эндокринную линию (CHGA+). Такие клеточные популяции могут, например, быть пригодными для скрининга молекул или условий, которые специфически индуцируют их дифференцировку в эндокринную клеточную линию. Соединения с такой активностью могут быть пригодными для приложений в регенеративной медицине для генерации эндокринных клеток, включая панкреатические бета-клетки in vivo или ex vivo. Поскольку известно, что фракционирование и очистка специфических клеточных популяций из более сложных клеточных смесей вызывает некоторую потерю эффективности из-за клеточной гибели и по другим причинам, полезно иметь возможность получения высоко обогащенных клеточных популяций с необходимым фенотипом посредством только эффективной направленной дифференцировки, не требующей этапов очистки.
Недифференцированные человеческие ESC размножали и дифференцировали по существу так, как описано выше в Таблице 17 Протокол №1 для этапов 1-4 и в Schulz и др. (2012) выше, за тем исключением, что во время трех (3) дней стадии 3, 4 сравнивали различные условия: (i) 0 ноггина («без ноггина») в течение всех 3 дней; (ii) 1 день с ноггином 50 нг/мл, и 2 дня без дополнительного ноггина; (iii) 2 дня с ноггином при 50 нг/мл и 1 день без дополнительного ноггина; и (iv) 3 дня с ноггином при 50 нг/мл. После стадии 3 для каждого из этих условий (i)-(iv) добавляли стандартный коктейль для среды дифференцировки для стадии 4, в соответствии с Таблицей 8 и Таблицей 17, но без дополнительного ноггина. Анализ проточной цитометрией после стадии 4 показал, что при отсутствии ноггина получается более 90% субпопуляции не-эндокринных мультипотентных панкреатических прогениторов и менее 5% клеток, коммитированных в эндокринную линию (CHGA+). См. таблицу 17. Далее, с каждым дополнительным днем обработки ноггином получалась меньшая субпопуляция не-эндокринных мультипотентных панкреатических прогениторов (сравните 90,6% без ноггина и 64,6% с 3 днями обработки ноггином). Напротив, каждый день обработки ноггином повышает уровни продукции клеток, коммитированных в эндокринную линию (CHGA+).

Q-PCR анализ клеточных культур после стадии 4 (день 12) показал, что каждый дополнительный день обработки ноггином приводил к повышению уровней экспрессии маркеров эндокринной линии, включая NKX2.2 и INS (Фиг. 31B и D), что соответствовало данным проточной цитометрии в Таблице 18. Напротив, уровни экспрессии маркерных генов для субпопуляции не-эндокринных мультипотентных панкреатических прогениторов, включая NKX6.1 (и PDX1), были относительно обильными в любой период времени обработки ноггином.
Пример 16. Агенты, модулирующие экспрессию генов эндокринной линии на стадии 7
Затем мы определяли, действительно ли имеются какие-либо дополнительные модуляторы экспрессии генов панкреатической линии. Способы дифференцировки были по существу схожими с теми, которые были описаны в Примерах 13 и 14 (т.е. AHW на стадии 3, АН на стадии 4, ингибиторы гамма-секретазы и rho-киназы на стадии 5, матригель и ингибитор rho-киназы на стадиях 6 и 7). Кроме того, другие агенты тестировали по их способности модулировать экспрессию генов, включая BMP, ноггин, FGF1, FGF2, FGF7, EGF, Hrg, HGF, SCF, или TTNPB в дни 30-35, во время стадии 7. Экспрессию РНК анализировали посредством Nanostring в конце продленной стадии 7 (день 35). На Фиг. 32А-С показано, что BMP повышает экспрессию INS, PDX1 и ID1, что согласуется с данными, показанными на Фиг. 24 и подробно описанными в Примере 13. BMP также снижали экспрессию GHRL. FGF1 (кислый FGF) осуществлял повышающую регуляцию GCG и SST. Неожиданно, FGF2 на дни 30-35 стадии 7 приводил к существенному повышению экспрессии SST при сопутствующем ингибировании или супрессии генов других гормонов (например, INS, GCG, GHRL; FIG.32B, SST панель).
Эти исследования показали, что некоторые агенты могут еще модулировать и влиять на экспрессию генов, и таким образом, дифференцировку клеточных культур и полученные типы клеток из этих более дифференцированных/коммитированных клеток.
Пример 17. Диссоциация и реагрегация клеточных культур снижает неэндокринные (CHGA-) субпопуляции и повышает адекватно специализированные эндокринные (CHGA+) субпопуляции.
Ранее отмечалось, что диссоциация и реагрегация клеточных культур может эффективно истощать или уменьшать присутствие определенных субпопуляций реагрегированных клеток, и следовательно, реагрегацию можно применять для обогащения некоторых субпопуляции клеток. Исследования были проведены для определения эффектов, если они имеются, реагрегированных клеточных культур с их обогащенными субпопуляциями, например, эндокринными (CHGA+) субпопуляциями. (См. Пример 14).
Способы дифференцировки были по существу подобными тем, которые описаны в Примере 14, включая диссоциацию и реагрегацию клеточных агрегатов, полученных из hES, перед стадией 7 (день 20). На Фигурах 33А-С показан анализ РНК после стадии 7 (примерно 29 день), демонстрирующий, что не-эндокринные (CHGA-) субпопуляции были существенно уменьшены в не-реагрегированных клеточных культурах, как было показано посредством Nanostring анализа с применением комбинации маркеров, включая PTF1A, SOX9, PDX1 и NKX6.1 (Фиг. 33А). Фигура 33В также показывает, что реагрегация приводит к повышению маркеров эндокринных клеток и клеток панкреатических островков, включая INS, GCG, панкреатический полипептид (PPY), GHRL, семейства носителей растворенных веществ 30 член 8 (SLC30A8), глюкозо-6-фосфатазу 2 (G6PC2), прогормон-конвертазу 1 (PCSK1) и глюкокиназу (GCK).
По отдельности, эти маркеры наблюдались в других типах клеток, например, PTF1A также экспрессируется в экзокринных клетках, SOX9 в протоковых клетках, PDX1 и NKX6.1 в бета-клетках. Так, авторы заявки получили данные по различным ПЭ, эндокринным и островковым маркерам в качестве эталона для определения значения экспрессии любого маркера. И хотя были определены четыре ПЭ и не-эндокринных (CHGA-) маркера (PTF1A, SOX9, PDX1 и NKX6.1), только PTF1A и SOX9 субпопуляции были истощены в реагрегированных клеточных культурах в конце стадии 7 (примерно день 25, день 26, день 27, день 28, день 29, день 30 и больше). Напротив, экспрессия PDX1 и NKX6.1, которая также наблюдалась в бета-клетках и панкреатических эндокринных клетках, оставалась высокой после стадии 7. См. Фиг. 33А.
Подобным образом, другие эндокринные маркеры (например, бета- и альфа-клеток), включая SLC30A8, G6PC2, PCSK1 и GCK, наблюдались при высоких уровнях экспрессии на 20 день в реагрегированных клеточных культурах, по сравнению с теми культурами, которые были не диссоциированными и реагрегированными (или «не-реагг.»). См. Фиг. 33С. Например, SLC30A8 является транспортером цинка, связанным с секрецией инсулина, и некоторые аллели SLC30A8 могут повышать риск развития диабета 2 типа; глюкозо-6-фосфатаза 2 (G6PC2) является ферментом, находящимся в панкреатических островках; прогормон-конвертаза 1 (PCSK1), вместе с PCSK2, перерабатывает проинсулин в инсулин в панкреатических островках; и наконец, глюкокиназа (GCK) облегчает фосфорилирование глюкозы в глюкозо-6-фосфат и экспрессируется в клетках печени, поджелудочной железы, кишечника, и головного мозга, где она регулирует углеводный метаболизм, действуя в качестве датчика глюкозы, запуская сдвиг метаболизма или функции клеток в ответ на подъем или падение уровней глюкозы, такой как после приема пищи или натощак. Мутации GCK могут вызывать необычные формы диабета или гипогликемию. Повышение экспрессии этих маркеров на стадии 7 указывает, что диссоциация и реагрегация клеточных культур истощает или снижает не-эндокринные (CHGA-) субпопуляции, и повышает эндокринные (CHGA+) субпопуляции. Таким образом, авторы заявки дополнительно улучшили способ получения адекватно специализированных эндокринных клеток in vitro для функции in vivo.
Анализ проточной цитометрией клеточных культур на стадиях 1-7 также подтвердил, что диссоциация и реагрегация клеточных культур, как указано выше, повышает общие эндокринные субпопуляции (обогащение CHGA+ клеток) с одновременным снижением не-эндокринных (CHGA-) субпопуляций или ПЭ субпопуляции клеток. См. Таблицу 19 ниже.

На сегодняшний день функциональные бета-клетки могут только быть дифференцированными через этап трансплантации in vivo, и подлинно функциональные бета-клетки полностью in vitro еще не описаны. См. Zhou, и др. (2008), Nature 455, 627-632. Так, вначале авторы настоящей заявки показали, что более 50% популяции эндокринных клеток (CHGA+) in vitro содержат меньше 10% не-эндокринных клеток (CHIGA-) и являются адекватно специализированными. И такие клетки способны к развитию и созреванию в подлинно функциональные бета-клетки и островки in vivo; см. Пример 18 ниже.
Пример 18. Продукция инсулина in vivo эндокринными (CHGA+) субпопуляциями
В качестве последующего исследования для Примера 17, были выполнены исследования для определения эффектов in vivo, если они имеются, реагрегированных клеточных культур с их обогащенными эндокринными (CHGA+) субпопуляциями.
Культуры человеческих плюрипотентных стволовых клеток размножали и дифференцировали подобно тому, как описано выше в Примерах 12-12 и 16 и в Schulz и др. (2012), выше, за тем исключением, что примерно в начале стадии 7 (день 20) культуры собирали и разделяли на 2 образца: (1) клеточные агрегаты, которые были диссоциированы и реагрегированы («реагрегированная клеточная культура»); и (2) клеточные агрегаты, которые были не диссоциированы и реагрегированы (контроль). Далее, оба образца дополнительно включали BMP, RA или аналог RA, TTNPB, и ингибитор rho-киназы и 0,05% матригель во время стадии 7 (дни 20-28 клеточных культур). Перед трансплантацией образцов отбирали пробы для анализа Nanostring и проточной цитометрии (Таблица 19 выше). В Таблице 19 показано, что реагрегированные клеточные культуры примерно на 29 день содержали примерно на 11% больше эндокринных (CHGA+) субпопуляций, чем контроль. Важно, что реагрегированный образец содержал на 9,25% меньше не-эндокринных (CHGA-) или ПЭ типов клеток (только 1,45%), чем образцы, которые были не реагрегированы.
Два образца по отдельности загружали и инкапсулировали в полупроницаемые биосовместимые устройства для доставки лекарств ENCAPTRA® EN20™ от авторов настоящей заявки (исследовательское устройство, предназначенное для животных) и трансплантировали подкожно, по существу, как описано в Примере 10 выше, всем из 10 мышей SCID-Beige (5 мышей получали реагрегированную клеточную культуру; 5 мышей получали контрольные клеточные культуры).
На Фигуре 34 показаны, спустя 21 неделю после трансплантации или при тестировании на живых организмах, сывороточные уровни человеческого С-пептида натощак (первый из трех столбцов слева для каждого животного), спустя 30 минут (30'; второй столбец) и один час (60'; третий столбец) после применения или нагрузки глюкозой. На 21 неделе после имплантации у всех кроме двух животных (животные №4317 и 4323; по одной из каждой группы) получены устойчивые уровни инсулина, как наблюдалось по более чем 932-2691 пМ человеческого сывороточного С-пептида. Однако когда показатели при стимуляции сравнивали между двумя группами (разница между уровнями сывороточного С-пептида 30' или 60' и уровнем сывороточного С-пептида натощак) не отмечалось существенной разницы функции in vivo между группами с реагрегированной клеточной культурой и контролем. Так, трансплантированные культуры или трансплантаты, состоящие примерно из 98% эндокринных/ CHGA+ реагрегированных клеточных культур или примерно 87% эндокринных/ CHGA+ (контроль), проявляли функцию in vivo. Так, авторы настоящей заявки впервые показали, эндокринная (CHGA+) клеточная популяция in vitro (адекватно специализированные эндокринные клетки) может давать начало физиологически функциональным инсулин-секретирующим клеткам при имплантации.
Таким образом, авторы настоящей заявки впервые продемонстрировали, что in vitro популяция адекватно специализированных эндокринных клеток при трансплантации развивается и созревает в функциональные панкреатические островковые клетки, которые секретируют инсулин в ответ на глюкозу крови. Далее функция in vivo подобна той, которая ранее была описана автором настоящей заявки для ПЭК и показана в Schulz и др. (2012), выше. Далее, кажется, что поскольку адекватно специализированные эндокринные клетки были получены посредством способов, описанных в настоящей заявке на стадиях 1-7, дополнительное обогащение эндокринных клеток (или истощение не-эндокринных клеток) через диссоциацию и реагрегацию является необязательным и не требуется для функции in vivo. Эти аспекты изобретения не были показаны до сегодняшнего дня. К настоящему времени авторы настоящей заявки и другие показали только функцию in vivo с применением клеточной популяции панкреатических прогениторов, а не эндокринной клеточной популяции. Действительно, как описано в Kelly и др. (2011) выше, показано, что эндокринные (CHGA+) субпопуляции ПЭК или препараты панкреатических прогениторов не обеспечивают функцию in vivo.
Пример 19. Комплексные основные среды для культивирования эндокринных клеток
Комплексные основные среды, такие как CMRL и RMPI, широко применяются для культивирования островковых клеток и сохранения клеточной массы островковых клеток. Среда CMRL, как правило, является более сложной (больше компонентов или ингредиентов), чем RPMI. И CMRL, и RMPI являются более сложными, чем DMEM, обычно применяемая и менее сложная среда для клеточной культуры. Так, авторы настоящей заявки проверяли, являются ли такие сложные основные среды благоприятными для поддержки эндокринных клеток на последних стадиях дифференцировки, например, на стадиях 6 и 7.
Трансплантация островков требует культивирования и пересадки островков с адекватной клеточной массой островков и минимальной токсичность. Кратковременная культура островков была связана с быстро деградацией и потерей жизнеспособности и контроля глюкозы. См. S. Matsumoto, и др. (2003), «А comparative evaluation of culture conditions for short-term maintenance (<24 hr.) of human islets isolated using the Edmonton protocol», Cell Tissue Banking, 4, p. 85. («Сравнительная оценка условий культивирования для кратковременной поддержки (менее 24 часов) человеческих островков (поджелудочной железы) с применением протокола Эдмонтона»); и N.J.M. London, и др. (1998), «Isolation, Culture and Functional Evaluation of Islets of Langerhans», Diabetes & Metabolism, 23, 200-207 («Выделение, культивирование и функциональная оценка островков Лангерганса»). В 1978 в исследовании Andersson и др. (1978) сравнивалась эффективность ТСМ, RPMI, CMRL, DMEM и Hams F10 для культивирования мышиных панкреатических островков. См. Andersson А. (1978) «Isolated mouse pancreatic islets in culture: Effects of serum and different culture media on the insulin production of the islets», Diabetelogia, 14, 397-404 («Изолированные мышиные панкреатические островки в культуре: влияние сыворотки и различных культуральных сред на продукцию инсулина в островках»). Результаты Andersson показали, что Ham's F10 обеспечивает островковые культуры с наибольшим содержанием инсулина, но RMPI обеспечивает наилучшую скорость биосинтеза инсулина в культурах; и что это, возможно, обусловлено высоким содержанием глюкозы и никотинамида в среде. Другие исследователи, такие как Davalli и др. (1992), показали, что ТСМ с аденозин-фосфатом и ксантином были лучшими для культивирования свиных панкреатических островков, чем CMRL и RMPI. См. Davalli, А.М. и др. (1992), «In vitro function of adult pig islets: Effect of culture in different media», Transplantation, 60, 854-860 («Функция панкреатических островков взрослых свиней in vitro: эффект культуры в различных средах»). Кроме того, Davalii и др. предположили, что высокое содержание глюкозы в Ham's F10 и RMPI было, возможно, токсичным для панкреатических островков свиней. Таким образом, островки от различных видов могут иметь различные требования к культуре, и нет единой рецептуры, пригодной для всех эндокринных и островковых клеток. См. Andersson (1978), р. 4 выше.
В таблице 20 ниже описаны только те компоненты в средах CMRL, RPMI и DMEM, которые варьируют между разными типами сред, т.е. Таблица 20 не перечисляет компонентов, которые присутствуют во всех 3 типах основных сред, но лишь перечисляет компоненты, содержащиеся в CMRL, но не содержащиеся в RPMI и/или DMEM. В Таблице 20 показано, что CMRL является наиболее сложной средой (наибольшее число компонентов), a DMEM является наименее сложной средой (наименьшее число компонентов). «Е» указывает наличие компонента в основной среде, а «Н» указывает отсутствие компонента в основной среде. Вкратце, в DMEM отсутствует примерно 7 компонентов (Н, затененные ячейки), которые присутствуют в CMRL и RPMI.


Способы получения культур эндокринных клеток были по существу подобными тем, которые описаны выше, за исключением стадии 6 (день 15), или середины стадии 7 (день 23), или середины стадии 7 (день 26), или стадий 6 и 7 (дни 15-25), где использовали основную среду DMEM или CMRL. Кроме того, в некоторые культуры добавляли инсулиноподобный фактор роста (IGF), никотинамид (NC), глюкозу (Glc), и/или ингибитор rho-киназы (Y-27632).
В одном эксперименте клетки на стадии 7, начиная примерно со дня 23, культивировали в основной среде CMRL или DMEM с добавкой В27, матригелем, BMP, IGF и Y-27632. Nanostring анализ образцов на 26 день (стадия 7) показал, что как правило, клетки на стадии 7, культивируемые в CMRL, имеют повышенную неспецифическую экспрессию генов и гормональных, и панкреатических эндокринных маркеров, по сравнению с клетками на той же стадии, культивированными в DMEM. См. Фиг. 35А-С. Таким образом, CMRL может быть важной для поддержки эндокринных клеток.
Соответствующие исследования также проводили с применением RPMI, сравниваемой со средами на основе DMEM и CMRL (данные не показаны). CMRL и RMPI среды оказывали схожее влияние на клеточные культуры, и обе были лучше, чем DMEM; таким образом, компоненты, не присутствующие в DMEM и присутствующие и в CMRL, и в RPMI, могут быть важными для поддержки эндокринных клеток во время стадии 6 и/или 7, включая восстановленный глутатион, аминокислоты, гидрокси-L-пролин, L-пролин, L-аспарагиновую кислоту, L-глутаминовую кислоту и витамины, биотин и пара-аминобензойную кислоту.
Пример 20. Криоконсервация не снижает число эндокринных клеток
Коммерчески пригодная клеточная терапия требует масштабного производства клеточного продукта, и средств хранения продукта в течение длительного продукта для применения по запросу пациента. Таким образом, любой клеточный продукт, клетки по отдельности или инкапсулированные клетки, такие как устройство для доставки лекарств ENCAPTRA®, могут требовать криоконсервации или других средств долговременного хранения без вредного или токсического влияния на клетки, и без влияния на терапевтический эффект клеток, или в данном случае на продукцию инсулина in vivo в ответ на глюкозу крови. Таким образом, авторы настоящей заявки изучали, действительно ли эндокринные клетки, криоконсервированные на стадии 7, сохраняют свою способность к развитию и функционированию in vivo, подобно тому, что наблюдалось для свежих (не-криоконсервированных) и криоконсервированных ПЭК, как подробно описано в патентах от авторов настоящей заявки U.S. №№8,278,106 и 8,425,928, оба озаглавлены «Инкапсулирование панкреатических клеток, полученных из человеческих плюрипотентных стволовых клеток», изданных 2 октября 2012 и 23 апреля 2013, соответственно, и заявке на патент США №61/775,480, озаглавленной «Криоконсервация, гибернация и хранение при комнатной температуре инкапсулированных панкреатических клеточных агрегатов», поданной 7 марта 2013, и Kroon et al (2008) выше.
Способы получения культур эндокринных клеток были по существу подобными тем, которые описаны выше, за тем исключением, что клеточные культуры все были в среде на основе CMRL во время стадии 7 и разделялись на следующие образцы: (1) не диссоциированные или реагрегированные, но криоконсервированные примерно на 23 день в течение нескольких часов, а потом размороженные и культивированные в течение нескольких дней в среде на основе CMRL с добавкой В27, BMP, TTNPB, и ингибитором rho-киназы (Не реаг./Криоконсервированные); (2) не диссоциированные или реагрегированные и не криоконсервированные (Не реаг./Не криоконсервированные); и (3) диссоциированные и реагрегированные и не криоконсервированные (Реаг./Не криоконсервированные). Все 3 образца культивировали в одних и тех же условиях среды стадии 7. Образцы анализировали проточной цитометрией на 27 день. См. Таблицу 21 ниже.

На основе анализа проточной цитометрии в Таблице 1, для тех культур, которые не были диссоциированы или реагрегированы (образцы 1 и 2), криоконсервация клеток (образец 2) не снижала общего числа эндокринных (CHGA+) клеток, по сравнению со свежими клетками (образец 1) (88,57 против 89,97). Как ожидалось, общая эндокринная (CHGA+) популяция была выше в культурах, которые были диссоциированы и реагригированы, и не криоконсервированы (92,85). Таким образом, диссоциация и реагрегация влияет на число эндокринных (CHGA+) клеток, как описано выше в Примере 17, но криоконсервация культур на стадии 7, как кажется, не влияет на изменение клеточного состава.
Далее образцы анализировали с применением клеточной иммуногистохимии с окрашиванием на NKX6.1, С-пептид, INS, и GCG. См. Фиг. 36А-В. Эндокринные (CHGA+) клеточные агрегаты со стадии 7 (Не-реаг./Не криоконсервированные; образец 2) показали совместную экспрессию или совместное окрашивание NKX6.1 (ядерное) и С-пептида (цитоплазматическое) (Фиг. 36А-С). Совместное окрашивание не наблюдалось в подобном анализе ПЭК со стадии 4 протокола дифференцировки. Клеточные культуры со стадии 7 были также первично моногормональными, как можно видеть на Фиг. 36А-С, что было продемонстрировано отдельным окрашиванием INS (36А; цитоплазма) и GCG (36В; цитоплазма) клеток, т.е. большинство индивидуальных клеток не проявляют совместной экспрессии INS и GCG, как можно видеть в эндокринных (CHGA+) субпопуляциях ПЭК со стадии 4. Фиг. 37 демонстрирует микрофотографии со схожим характером окрашивания, но от клеточных культур, которые были диссоциированы и реагрегированы в начале стадии 7.
Фигура 37 сравнивает функцию трансплантата из клеточных культур 1 и 3 из Таблицы 20 выше. Анализ функции in vivo трансплантатов на стадии 7, пересаженных обеим группам животных на 12 неделе, показал, что культуры, которые были криоконсервированы, но не реагрегированы (Образец 1), обеспечивали схожие сывороточные уровни С-пептида спустя 30 и 60 минут после глюкозной нагрузки, по сравнению с теми культурами, которые не были криоконсервированы, но были реагрегированы (Образец 3). Эти уровни С-пептида были сопоставимы с тем, которые наблюдались на 10 и 15 неделе на Фиг. 20 и 21 (например, на 15 неделе средний уровень С-пептида составил 995 пМ спустя 60 минут после применения глюкозы). Так, ожидается и предполагается, что последующий анализ покажет, что уровни С-пептида будут повышаться до более 1000 пМ спустя 30 и/или 60 минут, у большинства животных, после стимуляции глюкозой, подобно тому, что описано в Примере 18 и наблюдается на Фиг. 34. Фактически, у животного №4490 трансплантат образца 1 обеспечивал сывороточные уровни С-пептида на 12 неделе, которые были сопоставимы с уровнями у животного №4320 в Примере 18 (Фиг. 34) спустя 21 неделю после имплантации.
Таким образом, криоконсервация эндокринных клеток на стадии 7, как кажется, не влияет на их функцию in vivo.
Пример 21. Повышенные концентрации глюкозы влияют на экспрессию гормонов
Уровни глюкозы в рецептурах для клеточных культур находятся в диапазоне от 1 г/л (5,5 мМ) до 10 г/л (55 мМ), где некоторые среды содержат примерно 5,5 мМ глюкозы, что приближено к нормальным уровням сахара в крови in vivo. Уровни глюкозы, приближающиеся к 10 мМ, являются пред-диабетическими уровнями, а уровни выше 10 мМ аналогичны диабетическому состоянию. Иначе говоря, концентрация глюкозы выше 10 мМ имитирует гипергликемические условия in vivo. Высокий уровень глюкозы может вызвать, например, пост-трансплантационные вторичные модификации, включая гликацию, глиоксилирование и карбонильный стресс. Поскольку отчеты об эффектах высокой концентрации глюкозы в отношении различных типов клеток in vitro варьируют, авторы настоящей заявки предложили изучить влияние высокой концентрации глюкозы (т.е. 4,5 г/л или 24,98 мМ глюкозы) на эндокринные клетки стадии 6 и/или 7.
Глюкоза является растворимым гексозным сахаром, добавляемым в среды для культивирования клеток, включая среду Эймса; основную среду Игла (ВМЕ); BGJb среду в модификации Фиттона-Джексона; среду Клика; среду CMRL-1066; модифицированную Дульбекко среду Игла (DMEM); DMEM/Ham смесь нутриентов F-12 (50:50); F-12 в модификации Куна; среду Фишера; среду H-Y (Hybri-Max®); среду Дульбекко, модифицированную по способу Исков (IMDM); модифицированную среду Мак-Коя 5А; среды MCDB; среду 199; минимальную эссенциальную среду Игла (ЕМЕМ); среду NCTC; смесь нутриентов, Ham's F-10; смесь нутриентов, Ham's F-12; смесь нутриентов Ham's F-12 в модификации Kaighn (F12K); RPMI-1640; бессывороточную/безбелковую среду для гибридомы; среду Waymouth MB; среду Williams Е, и различные патентованные среды. L-15 среда содержит галактозу вместо глюкозы. См. справочник по средам Sigma-Aldrich, доступный на веб-сайте: sigmaaldrich.com/life-science/cell-culture/learning-center/media-expert/glucose.html.
Среды, включающие уровни добавленной глюкозы более 10 мМ, включают по меньшей мере DMEM/Ham смесь нутриентов F-12 (50:50), содержащую 17,5 мМ глюкозы; DMEM (Hi Glucose), GMEM и IMDM, содержащие 25 мМ глюкозы; и H-Y среду (Hybri-Max®) и бессывороточную/безбелковую среду для гибридомы, содержащие 22,6 и 28,9 мМ глюкозы, соответственно. См. справочник по средам Sigma-Aldrich доступный на вебсайте: sigmaaldrich.com/life-science/cell-culture/learning-center/media-expert/glucose.html. Поскольку различные культуральные среды содержат широко варьирующие уровни глюкозы, и что эффекты глюкозы, как правило, меняются от одной системы культивирования клеток к другой, изучали влияние высокой концентрации глюкозы на эндокринные клетки.
Вновь, способы получения культур эндокринных клеток были по существу подобны тем, которые описаны выше, за тем исключением, что во время стадий 6 и/или 7 культуры обрабатывали средой на основе CMRL с высоким содержанием экзогенной глюкозы (общая концентрация глюкозы 4,5 г/л, или 25 мМ), или без нее. Так, это составило 3,5 г/л в дополнение к 1000 мг/л или 1 г/л (5,5 мМ) глюкозы, уже присутствующей в среде CMRL. При обоих условиях с экзогенной глюкозой и без нее добавляли те же самые факторы роста, включая никотинамид, BMP, RA (или TTNPB), и альтернативно, ингибитор rho-киназы и матригель. Анализ Nanostring показал, что когда высокое количество глюкозы добавляли в начале стадии 6, то в конце стадии 6 отмечалась повышенная экспрессия INS, GCG и SST и сниженная экспрессия GHRL (Фиг. 39А и В). Таким образом, повышение экзогенной глюкозы на стадиях 6 и 7, за исключением грелина, повышает экспрессию гормонов.
Пример 22. Получение незрелых бета-клеток
Примеры 8-21 описывают различные итеративные способы получения эндокринных клеток in vitro, включая применение высокого уровня активина, по отдельности или в комбинации с Wnt и/или херегулином на стадии 3; низким уровнем активина на стадии 4, по отдельности или в комбинации с херегулином; ноггина и ингибитора гамма-секретазы с KGF, EGF и ингибитором rho-киназы на стадии 5 или без них; и одного или более из никотинамида, ретиноевой кислоты, BMP и матригеля на стадиях 6-7. Популяции эндокринных клеток, полученные такими способами, были не только моногормональными (например, только INS, только GCG или только SST; см. Фиг. 39), но также одновременно экспрессировали другие маркеры незрелых эндокринных клеток, включая NKX6.1 и PDX1.
Анализ проточной цитометрией проводили на двух различных культурах стадии 7 с применением окраски на гормоны INS, SST и GCG (набор данных А и набор данных В). Один набор данных (А) получали после анализа многих повторений стадий 1-7, а другой набор данных (В) получали раньше по времени, например, при отсутствии никотинамида, BMP, CMRL и тому подобного. См. Таблицу 22 ниже. В таблице 22 показано общее процентное содержание позитивного окрашивания на INS, SST и GCG (см. 5 колонок слева), негативного окрашивания на INS, SST и GCG (см. 5 колонок справа), и моногормонального окрашивания (см. среднюю колонку). Окрашивание на гормоны для этих двух наборов данных (А и В) сравнивали с тем, что было получено в культурах ПЭК на стадии 4, которые поддерживали в культуре примерно до 26 дня.

В целом, общая субпопуляция INS-позитивных клеток (сумма одинарно INS-позитивных плюс INS/SST со-позитивных плюс INS/GCG со-позитивных плюс INS/SST/GCG со-позитивных) является в 3 раза больше на стадии 7 эндокринных культур, чем на стадии 4 ПЭК культур (49,3 против 22,7 против 15,8). Соответственно, общая субпопуляция только с INS (без одновременной экспрессией с SST или GCG) также является более высокой в культурах на стадии 7, чем в культурах ПЭК на стадии 4 (22,7 против 12,9 против 3,7). Важно, что в культурах на стадии 7 отмечается больше клеток, экспрессирующих единственный гормон INS в процентном содержании от общей INS популяции (22,7/49,3=46% в А и 12,7/22,7= ~57% в В), по сравнению с культурами ПЭК на стадии 4 (3,7/15,8=23%). Таким образом, видно, что культуры эндокринных клеток на стадии 7 состоят из большего числа клеток, экспрессирующих один гормон, в частности, клеток, экспрессирующих только INS, чем в культурах ПЭК на стадии 4. И по меньшей мере в Примерах 18 и 20 показано, что эндокринные клетки на стадии 7, хотя и все еще незрелые, развиваются и созревают в физиологически функциональные панкреатические островки, способные секретировать инсулин в ответ на глюкозу крови при трансплантации; в то время как эндокринные (CHGA+) субпопуляции культур ПЭК на стадии 4 не способны к этому in vivo. См. Фиг. 44 и 45 и Примеры выше для более подробного описания.
Пример 23. Эндокринные (CHGA+) клетки со стадии 7 и стадии 4 (ПЭК) отличаются друг от друга.
Хотя анализ культур со стадии 7 (адекватно специализированных эндокринных) и со стадии 4 (ПЭК) использует те же самые антитела (например, CHGA, INS, NKX6.1) для окрашивания и анализа клеток, субпопуляции адекватно специализированных эндокринных (CHGA+) клеток со стадий 7 и 4 (ПЭК) не являются одними и теми же.
Ранее авторы настоящей заявки сообщили, что происхождение функциональных бета-клеток, наблюдаемых после трансплантации культур ПЭК на 4 стадии, связано с панкреатическими прогениторами или не-эндокринными (CHGA-) клетками, а не с первичными эндокринными (CHGA+) клетками. Kelly и др. (2011), выше, показали, что когда культуры ПЭК были обогащенными (очищенными) эндокринными (CHGA+) субпопуляциями и трансплантированными, они не давали начала функциональными панкреатическими островками или бета-клетками, по сравнению с необогащенными культурами ПЭК. См. Kelly et al (2011), рр. 3-4, Фиг. 4 и дополнительная Таблица 1; Schulz и др. (2012) выше; и патенты США №№7,534,608; 7,695,965; 7993920; и 8,278,106. Примеры 17 и 18 и Таблица 19 описывает новую чистую (98,2%) эндокринную (CHGA+) клеточную культуру со стадии 7, которая при трансплантации приводит устойчивому функциональному трансплантату in vivo (Фиг. 34). С учетом Примеров 17 и 18 и других результатов, описанных в настоящей заявке, функциональная популяция культур со стадии 7 является эндокринными (CHGA+) клетками со стадии 7, и с незначительным процентом не-эндокринных (CHGA-) клеток (2,08%). Это отличается от культур ПЭК 4 стадии, которые описаны подробно автором настоящей заявки в предыдущих документах, где функция in vivo связана с не-эндокринными (CHGA-) субпопуляциями. Таким образом, в целом, эндокринные субпопуляции со стадий 7 и 4 не являются сопоставимыми или одними и теми же.
В Таблице 23 показаны данные по проточной цитометрии, сравнивающие общие эндокринные (CHGA+) и не-эндокринные (CHGA-) субпопуляции со стадии 7 (адекватно специализированные эндокринные) и стадии 4 (ПЭК), и демонстрирующие, что субпопуляции не являются эквивалентными и отличаются друг от друга. Например, общий процент CHGA+ субпопуляции был существенно выше для стадии 7, по сравнению с культурами со стадии 4 (85,2 против 39,4). Соответственно, общий процент CHGA-субпопуляции был существенно ниже для стадии 7, по сравнению с культурами со стадии 4 (14,8 против 60,1). Далее, истинные эндокринные клетки, например, бета-клетки, не только экспрессируют CHGA+, на также одновременно экспрессируют по меньшей мере INS и NKX6.1. Таким образом, истинные эндокринные клетки экспрессируют CHGA+/INS+/NKX6.1+ (тройные позитивные) и также способны к функционированию in vivo. В Таблице 23 показано, что культуры на стадии 7 содержат почти в 5 раз больше CHGA+/INS+/NKX6.1+ (тройных позитивных клеток) клеток, чем культуры на стадии 4 (37,8 против 7,8); и даже хотя на стадии 4 CHGA+/INS+/NKX6.1+ субпопуляции могут быть адекватно специализированными, как обсуждалось выше, эти эндокринные субпопуляции со стадии 4 не функционируют in vivo при трансплантации, в то время как не-эндокринные (CHGA-) субпопуляции со стадии 4 не созревают и не функционируют in vivo. Таким образом, авторы настоящей заявки обозначают эндокринные клетки на стадии 7 как «незрелые эндокринные клетки» или «незрелые бета-клетки», а не эндокринные прогениторы/клетки-предшественники, поскольку эти клетки в своем развитии коммитированы к превращению в зрелые бета-клетки in vivo.

Пример 24. Очистка эндокринных клеток из клеточных популяций со стадии 7 с применением датчика цинка
Секреторные везикулы бета-клеток содержат высокие концентрации цинка. Цинк накапливается в везикулах по меньшей мере под действием транспортера цинка SLC30A8 (или ZnT8). Связывающие цинк флюоресцентные зонды используют для визуализации цинка в клетках путем абсорбции и повышения эмиссии на специфических длинах волн при связывании с цинком и без цинка. В публикациях показано, что большая часть зондов может не локализоваться надлежащим образом для детекции цинка в везикулах бета-клеток. Интересно, что PyDPy1 (или Py1; Chemical Communications, 2011, 47:7107-9) может поступать в везикулы, но не применялся с бета-клетками до сегодняшнего дня. Py1 повышает интенсивность флюоресценции в 50-80 раз при связывании с Zn2+. Впервые авторы настоящей заявки продемонстрировали получение популяции эндокринных клеток, или незрелых бета-клеток, способной к функционированию in vivo. Таким образом, предпочтительно иметь средства для дополнительного обогащения этой популяции для дополнительных анализов in vitro, для применения в качестве инструмента скрининга, или для обеспечения обогащенной популяции незрелых бета-клеток для трансплантации.
Эндокринные клетки на стадии 7, или незрелые бета-клетки, обрабатывали Py1, и сортировали с помощью флюоресценции, отбирая клетки с высоким содержанием цинка. Датчик цинка Py1 (синтезированный по заказу ChemoGenics Biopharma, Research Triangle Park, Северная Каролина) ресуспендировали при 10 мМ в ДМСО и разбавляли в DPBS (-/-)/ 0,25% BSA (Буферный раствор А) до 5 микромолярной итоговой концентрации (окрашивающий раствор). Дифференцированные клеточные агрегаты на 7 стадии дважды промывали DPBS(-/-) и диссоциировали Accumax. Accumax гасили добавлением основной среды, содержащей В-27. Полученную клеточную суспензию фильтровали через сетку с отверстиями 40 микрон, центрифугировали, промывали буферным раствором А, и ресуспендировали в окрашивающем растворе. Окрашивание проводили для 15 минут, клетки центрифугировали, промывали буферным раствором А и ресуспендировали буферным раствором А для сортировки. Клетки сортировали проточной цитометрией в CMRL/50% FBS при 4°С. После сортировки клетки центрифугировали и ресуспендировали в буферном растворе для выделения РНК.
В результатах, показанных на Фиг. 40, клетки, охваченные или заключенные в многоугольник, являются живыми клетками, у которых наблюдалась повышенная флюоресценция из-за присутствия Py1 датчика, связанного с цинком. Эту клеточную популяцию разделяли примерно на два равных потока, и сортировали в две пробирки, обозначенный «Яркий» и «Тусклый», в соответствии с интенсивностью их флюоресценции. Из клеток готовили РНК и подвергали анализу Nanostring.
Как видно на Фиг. 42А-С, тусклая популяция обогащена маркерами бета-клеточной линии, такими как INS, IAPP, PDX1, NKX6.1, РАХ4, PCSK1, G6PC2, GCK и SLC30A8. Это показывает, что бета-клетки или незрелые бета-клетки могут быть очищены из клеточной популяции со стадии 7 с применением датчика цинка. В предыдущих экспериментах было определено, что 630000 INS мРНК единиц при Nanostring соответствуют примерно 49% INS-позитивных клеток путем проточной цитометрии (данные не показаны). Таким образом, очищенная тусклая популяция имела значение INS при Nanostring 1,200,000, что соответствовало примерно 93% INS-позитивных клеток (1,200,000/630,000 х 49%).
Напротив, отсортированная яркая популяция была обогащена маркерами альфа-клеточной линии, такими как GCG, ARX и SLC30A8 (Фиг. 41, 42, 43). Подобно бета-клеткам, глюкагоновые клетки, как известно, поглощают цинка, поэтому датчик Py1 можно применять для связывания цинка в бета- и глюкагоновых клетках, но разделяя или сортируя по различным уровням флюоресценции.
Обобщение способов получения культур ПЭК (стадии 1-4) эндокринных клеток (стадии 1-7).
В итоге, изобретения, описанные в настоящей заявке, направлены по меньшей мере на ПЭК и незрелые эндокринные клетки и на способы получения таких клеток, включающие по меньшей мере стадии 1-4 получения ПЭК, и стадии 1-7 для получения эндокринных клеток. Фигуры 43, 44 и 45 являются диаграммами, обобщающими некоторые аспекты клеточных композиций авторов настоящей заявки, и способов получения, описанных в настоящей заявке.
Авторы настоящей заявки ранее сообщали, что эндокринные (CHGA+) после стадии 4 протокола дифференцировки не развиваются в зрелые и функционирующие клетки панкреатических островков при трансплантации in vivo. См. клеточный тип «EN» после стадии 4 на Фиг. 43 и 44. Эти эндокринные (CHGA+) субпопуляции имеют раннюю или незрелую экспрессию NGN3 (экспрессию NGN3 перед совместной экспрессией PDX1 и NKX6.1). См. также Rukstalis и др. (2009), выше. Напротив, не-эндокринные (CHGA-) субпопуляции ПЭК, которые не экспрессировали NGN3, развивались и созревали в эндокринные клетки in vivo. Эта замедленная экспрессия NGN3 до последующего созревания in vivo не-эндокринных (CHGA-) субпопуляций ПЭК показана в Примере 10 (Фиг. 15-16), где комбинация активина, Wnt и херегулина эффективно подавляла NGN3, по сравнению с контролем, а трансплантация этих ПЭК, по сравнению с контролем, давала начало улучшенной функции in vivo. См. Фиг. 15-16.
Авторы настоящей заявки затем попытались получить культуру адекватно специализированных эндокринных клеток, способных развиваться и созревать в панкреатические островки in vivo и вырабатывать инсулин в ответ на уровни глюкозы крови, подобно тому, что наблюдалось во всех случаях с ПЭК, в частности, неэндокринной (CHGA-) субпопуляции ПЭК. Для этого авторы настоящей заявки выполнили много повторных экспериментов для подавления или ингибирования экспрессии NGN3 на стадиях 3 и 4. В Примерах 8-11 подробно описано применение авторами настоящей заявки активина по отдельности или в комбинации с другими агентами, такими как Wnt и херегулин, и в различных концентрациях, для влияния на экспрессию или супрессию NGN3. На Фиг. 44 также обобщено влияние активина на стадиях 3 и 4.
Поскольку было продемонстрировано, что экспрессия NGN3 может быть замедлена, и это не влияет на функцию in vivo, авторы настоящей заявки разработали способы индукции экспрессии эндокринных маркеров на стадиях после формирования ПЭК (стадии 4). Примеры 11-14 и 16-22 описывают многие повторения и способы, применяемые для оптимизации культуральных условий для получения адекватно специализированных эндокринных популяций, которые могут давать начало функции in vivo. В частности, авторы настоящей заявки использовали ингибитор гамма-секретазы на стадии 5 для индукции экспрессии NGN3 и эндокринной дифференцировки. Авторы настоящей заявки использовали такие реагенты, как CMRL на стадиях 6 и 7 для повышения экспрессии эндокринных маркеров; BMP для повышения INS и PDX1. На Фигурах 44 и 45 обобщены и описаны эти попытки.
Необходимо понимать, что исходно требуется применение множества методологий для характеристики и идентификации клеток (например, Q-ПЦР, ИЦХ, проточной цитометрии, количественного определения С-пептида и тому подобного). После полной характеристики и идентификации таких клеток в определенных условиях культивирования для дифференцировки клеток, часто применяют Q-ПЦР и Nanostring multiplex RNA в качестве единственных способов анализа, чтобы определить, действительно ли получен такой тип клеток.
Способы, композиции и устройства, описанные в настоящей заявке, представляют предпочтительные варианты осуществления, и являются примерными и не предназначены для ограничения объема изобретения. Изменения и другие виды применения очевидны для специалистов в данной области техники и охватываются сущностью настоящего изобретения и определяются объемом описания. Соответственно, для специалиста в данной области техники очевидно, что различные замены и модификации могут быть выполнены в раскрытом изобретени, не выходя за рамки и сущность настоящего изобретения.
Использующееся в формуле изобретения и в описании, выражение «по существу состоящий из» означает включение любых элементов, перечисленных после выражения, и ограничено другими элементами, которые не мешают или способствуют активности или действию, указанному в описании для перечисленных элементов. Таким образом, выражение «по существу состоящий из» указывает, что перечисленные элементы являются необходимыми или обязательными, но что другие элементы являются необязательными и могут присутствовать или не присутствовать, в зависимости от того, влияют или не влияют они на активность или действие перечисленных элементов. Далее, подразумевается, что в вариантах осуществления, где приведены численные величины, такие как количества, концентрации, проценты, пропорции или диапазоны, обозначенная величина может быть «по меньшей мере примерно» численным значением, «примерно» численным значением или «по меньшей мере» численным значением.
Варианты осуществления настоящего изобретения
Вариант осуществления 1. Унипотентная человеческая незрелая бета-клетка in vitro.
Вариант осуществления 2. Унипотентная человеческая незрелая бета-клетка из варианта 1, где указанная унипотентная человеческая незрелая бета-клетка экспрессирует INS и NKX6.1 и по существу не экспрессирует NGN3.
Вариант осуществления 3. Унипотентная человеческая незрелая бета-клетка из варианта 1, где указанная унипотентная человеческая незрелая бета-клетка способна к дифференцировке в зрелую бета-клетку.
Вариант осуществления 4. Унипотентная человеческая незрелая бета-клетка из варианта 3, где указанная дифференцировка осуществляется in vivo.
Вариант осуществления 5. Унипотентная человеческая незрелая бета-клетка из варианта 1, где указанная унипотентная человеческая незрелая бета-клетка образует часть клеточной популяции.
Вариант осуществления 6. Унипотентная человеческая незрелая бета-клетка из варианта 5, где по меньшей мере примерно 10% указанной клеточной популяции является незрелыми бета-клетками.
Вариант осуществления 7. Унипотентная человеческая незрелая бета-клетка из варианта 5, где по меньшей мере примерно 20% указанной клеточной популяции является незрелыми бета-клетками.
Вариант осуществления 8. Унипотентная человеческая незрелая бета-клетка из варианта 5, где по меньшей мере примерно 30% указанной клеточной популяции является незрелыми бета-клетками.
Вариант осуществления 9. Унипотентная человеческая незрелая бета-клетка из варианта 5, где по меньшей мере примерно 40% указанной клеточной популяции является незрелыми бета-клетками.
Вариант осуществления 10. Унипотентная человеческая незрелая бета-клетка из варианта 5, где по меньшей мере примерно 50% указанной клеточной популяции является незрелыми бета-клетками.
Вариант осуществления 11. Унипотентная человеческая незрелая бета-клетка из варианта 5, где по меньшей мере примерно 60% указанной клеточной популяции является незрелыми бета-клетками.
Вариант осуществления 12. Унипотентная человеческая незрелая бета-клетка из варианта 5, где по меньшей мере примерно 80% указанной клеточной популяции является незрелыми бета-клетками.
Вариант осуществления 13. Унипотентная человеческая незрелая бета-клетка из варианта 5, где по меньшей мере примерно 90% указанной клеточной популяции является незрелыми бета-клетками.
Вариант осуществления 14. Унипотентная человеческая незрелая бета-клетка из варианта 5, где по меньшей мере примерно 98% указанной клеточной популяции является незрелыми бета-клетками.
Вариант осуществления 15. Унипотентная человеческая незрелая бета-клетка из варианта 1, где человеческая незрелая бета-клетка является моногормональной.
Вариант осуществления 16. Унипотентная человеческая незрелая бета-клетка из варианта 1, где указанная унипотентная человеческая клетка экспрессирует MAFB.
Вариант осуществления 17. Способ получения унипотентной человеческой незрелой бета-клетки, включающий обеспечение контакта клеток дефинитивной эндодермальной линии in vitro с членом суперсемейства TGFβ и членом семейства Wnt, с генерацией незрелых бета-клеток.
Вариант осуществления 18. Способ из варианта 4, где указанная упипотентная человеческая незрелая бета-клетка экспрессирует INS, NKX6.1 и по существу не экспрессирует NGN3.
Вариант осуществления 19. Способ из варианта 4, где член суперсемейства TGFβ выбран из группы, состоящей из Nodal, активина А, активина В, ВМР2, ВМР4, GDF8, GDF-10, GDF-11 и GDF-15.
Вариант осуществления 20. Способ из варианта 4, дополнительно включающий обеспечение контакта клеток дефинитивной эндодермальной линии in vitro с агентом, активирующим киназу рецептора ERRB.
Вариант осуществления 21. Способ из варианта 4, где фактор роста суперсемейства TGFβ является активином А.
Вариант осуществления 22. Способ из варианта 4, где членом семейства Wnt является Wnt3a.
Вариант осуществления 23. Способ из варианта 6, где агент, активирующий киназу рецептора ERBB, является херегулином.
Вариант осуществления 24. Способ из варианта 4, где фактор роста суперсемейства TGFβ является BMP.
Вариант осуществления 25. Способ из варианта 4, дополнительно включающий обеспечение контакта клеток человеческой дефинитивной эндодермальной линии in vitro с никотинамидом, ретиноевой кислотой или аналогом ретиноевой кислоты, ингибитором rho-киназы или ингибитором гамма-секретазы.
Вариант осуществления 26. Способ из варианта 25, где ретиноевая кислота состоит из полностью транс-ретиноевой кислоты (RA), 13-цис-ретиноевой кислоты (13-cis-RA), и аротиноидной кислоты (TTNPB).
Вариант осуществления 27. Способ из варианта 26, где ретиноевая кислота является полностью транс-ретиноевой кислотой (RA).
Вариант осуществления 28. Способ из варианта 26, где ретиноевая кислота является 13-цис-ретиноевой кислотой (13-cis-RA).
Вариант осуществления 29. Способ из варианта 26, где ретиноевая кислота является аротиноидной кислотой (TTNPB).
Вариант осуществления 30. Способ из варианта 25, где ингибитор rho-киназы выбран из группы, состоящей из Y-27632, Fasudil, Н-1152Р, Wf-536 и Y-30141.
Вариант осуществления 31. Способ из варианта 30, где ингибитор rho-киназы является Y-27632.
Вариант осуществления 32. Способ из варианта 25, где ингибитор гамма-секретазы выбран из группы, состоящей из ингибитора гамма-секретазы I (GSI I), ингибитора гамма-секретазы II (GSI II), ингибитора гамма-секретазы III (GSI III), ингибитора гамма-секретазы IV (GSI IV), ингибитора гамма-секретазы V (GSI V), ингибитора гамма-секретазы VI (GSI VI), ингибитора гамма-секретазы VII (GSI VII), ингибитора гамма-секретазы IX (GSI IX), (DAPT), ингибитора гамма-секретазы XI (GSI XI), ингибитора гамма-секретазы XII, (GSI XII), ингибитора гамма-секретазы XIII (GSI XIII), ингибитора гамма-секретазы XIV (GSI XIV), ингибитора гамма-секретазы XVI (GSI XVI), ингибитора гамма-секретазы XVII (GSI XVII), ингибитора гамма-секретазы XIX (GSI XIX), ингибитора гамма-секретазы XX (GSI XX), ингибитора гамма-секретазы XXI (GSI XXI), ингибитора гамма40-секретазы I, ингибитора гамма40-секретазы II и RO4929097.
Вариант осуществления 33. Способ из варианта 32, где ингибитор гамма-секретазы является RO4929097.
Вариант осуществления 34. Способ из варианта 32, где ингибитор гамма-секретазы является GSI IV.
Вариант осуществления 35. Способ получения зрелых бета-клеток in vivo, включающий: (а) обеспечение контакта клеток человеческой дефинитивной эндодермальной линии in vitro с членом суперсемейства TGFβ и членом семейства Wnt с генерацией незрелых бета-клеток; (b) трансплантацию незрелых бета-клеток с этапа (а) млекопитающему субъекту; и (с) обеспечение дифференцировки незрелых бета-клеток in vivo до получения популяции клеток, включающих зрелые бета-клетки.
Вариант осуществления 36. Способ из варианта 10, дополнительно включающий обеспечение продукции инсулина указанными зрелыми бета-клетками в ответ на стимуляцию глюкозой.
Вариант осуществления 37. Способ из варианта 10, где указанный фактор роста из суперсемейства TGFβ выбран из группы, состоящей из Nodal, активина А и активина В, GDF8, GDF-11 и GDF-15.
Вариант осуществления 38. Способ из варианта 10, дополнительно включающий обеспечение контакта клеток человеческой дефинитивной эндодермальной линии in vitro с агентом, активирующим киназу рецептора ERBB.
Вариант осуществления 39. Способ из варианта 10, где фактор роста суперсемейства TGFβ является активином А.
Вариант осуществления 40. Способ из варианта 10, где член семейства Wnt является Wnt3a.
Вариант осуществления 41. Способ из варианта 37, где агентом, активирующим киназу рецептора ERBB, является херегулин.
Вариант осуществления 42. Способ из варианта 10, где клетки человеческой дефинитивной эндодермальной линии контактируют по меньшей мере с 25 нг/мл фактора роста суперсемейства TGFβ.
Вариант осуществления 43. Способ из варианта 10, где клетки человеческой дефинитивной эндодермальной линии контактируют по меньшей мере с 50 нг/мл фактора роста суперсемейства TGFβ.
Вариант осуществления 44. Способ из варианта 10, где клетки человеческой дефинитивной эндодермальной линии контактируют по меньшей мере с 75 нг/мл фактора роста суперсемейства TGFβ.
Вариант осуществления 45. Способ из варианта 10, где клетки человеческой дефинитивной эндодермальной линии контактируют по меньшей мере с 25 нг/мл члена семейства Wnt.
Вариант осуществления 46. Способ из варианта 10, где клетки человеческой дефинитивной эндодермальной линии контактируют по меньшей мере с 50 нг/мл члена семейства Wnt.
Вариант осуществления 47. Способ из варианта 10, где клетки человеческой дефинитивной эндодермальной линии контактируют по меньшей мере с 75 нг/мл члена семейства Wnt.
Вариант осуществления 48. Способ из варианта 37, где клетки человеческой дефинитивной эндодермальной линии контактируют с количеством агента, активирующего киназу рецептора ERBB, в 5-15 раз меньшим количества фактора роста суперсемейства TGFβ и члена семейства Wnt.
Вариант осуществления 49. Способ из варианта 10, где по меньшей мере 10% указанной клеточной популяции являются незрелыми бета-клетками.
Вариант осуществления 50. Способ из варианта 10, где по меньшей мере 20% указанной клеточной популяции являются незрелыми бета-клетками.
Вариант осуществления 51. Способ из варианта 10, где по меньшей мере 30% указанной клеточной популяции являются незрелыми бета-клетками.
Вариант осуществления 52. Способ из варианта 10, где по меньшей мере 40% указанной клеточной популяции являются незрелыми бета-клетками.
Вариант осуществления 53. Способ из варианта 10, где по меньшей мере 50% указанной клеточной популяции являются незрелыми бета-клетками.
Вариант осуществления 54. Способ из варианта 10, где по меньшей мере 60% указанной клеточной популяции являются незрелыми бета-клетками.
Вариант осуществления 55. Способ из варианта 10, где по меньшей мере 70% указанной клеточной популяции являются незрелыми бета-клетками.
Вариант осуществления 56. Способ из варианта 10, где по меньшей мере 80% указанной клеточной популяции являются незрелыми бета-клетками.
Вариант осуществления 57. Способ из варианта 10, где по меньшей мере 90% указанной клеточной популяции являются незрелыми бета-клетками.
Вариант осуществления 58. Способ из варианта 10, где незрелые бета-клетки экспрессируют INS и NKX6.1, и по существу не экспрессируют NGN3.
Вариант осуществления 59. Способ из варианта 10, где незрелые бета-клетки экспрессируют INS, NKX6.1 и MFAB, и по существу не экспрессируют NGN3.
Вариант осуществления 60. Унипотентные человеческие незрелые бета-клетки in vitro, экспрессирующие INS и NKX6.1, и по существу не экспрессирующие NGN3.
Вариант осуществления 61. Унипотентные человеческие незрелые бета-клетки из варианта 60, где указанные унипотентные человеческие незрелые бета-клетки способны к созреванию в зрелые бета-клетки.
Вариант осуществления 62. Унипотентные человеческие незрелые бета-клетки из варианта 60, где указанные унипотентные человеческие незрелые бета-клетки дополнительно экспрессируют MAFB.
Вариант осуществления 63. Способ получения унипотентных человеческих незрелых бета-клеток, включающий обеспечение контакта клеток человеческой дефинитивной эндодермальной линии in vitro с членом суперсемейства TGFβ и членом семейства Wnt, с получением, таким образом, унипотентных человеческих незрелых бета-клеток.
Вариант осуществления 64. Способ из варианта 63, где указанные унипотентные человеческие незрелые бета-клетки экспрессируют INS, NKX6.1, и по существу не экспрессируют NGN3.
Вариант осуществления 65. Способ из варианта 63, дополнительно включающий обеспечение контакта клеток человеческой дефинитивной эндодермальной линии in vitro с агентом, активирующим киназу рецептора ERBB.
Вариант осуществления 66. Способ из варианта 63, где фактором роста суперсемейства TGFβ является активин А.
Вариант осуществления 67. Способ из варианта 63, где членом семейства Wnt является Wnt3a.
Вариант осуществления 68. Способ из варианта 65, где агентом, активирующим киназу рецептора ERBB, является херегулин.
Вариант осуществления 69. Способ получения зрелых клеток in vivo, включающий: (а) обеспечение контакта клеток человеческой дефинитивной эндодермальной линии in vitro с членом суперсемейства TGFβ и членом семейства Wnt с генерацией незрелых бета-клеток; (b) трансплантацию незрелых бета-клеток с этапа (а) млекопитающему субъекту; и (с) обеспечение созревания незрелых бета-клеток in vivo до получения популяции клеток, включающих инсулин-секретирующие бета-клетки.
Вариант осуществления 70. Способ из варианта 69, дополнительно включающий обеспечение контакта клеток человеческой дефинитивной эндодермальной линии in vitro с агентом, активирующим киназу рецептора ERBB.
Вариант осуществления 71. Способ из варианта 69, где фактором роста суперсемейства TGFβ является активин А.
Вариант осуществления 72. Способ из варианта 69, где членом семейства Wnt является Wnt3a.
Вариант осуществления 73. Способ из варианта 70, где агентом, активирующим киназу рецептора ERBB, является херегулин.
Вариант осуществления 74. Способ из варианта 69, где клетки человеческой дефинитивной эндодермальной линии контактируют по меньшей мере с 50 нг/мл фактора роста суперсемейства TGFβ.
Вариант осуществления 75. Способ из варианта 69, где клетки человеческой дефинитивной эндодермальной линии контактируют по меньшей мере с 25 нг/мл члена семейства Wnt.
Вариант осуществления 76. Способ из варианта 70, где клетки человеческой дефинитивной эндодермальной линии контактируют с количеством агента, активирующего киназу рецептора ERBB, в 5-15 раз меньшим количества фактора роста суперсемейства TGFβ и члена семейства Wnt.
Вариант осуществления 77. Способ из варианта 69, где незрелые бета-клетки экспрессируют INS и NKX6.1, и по существу не экспрессируют NGN3.
Вариант осуществления 78. Способ из варианта 69, где незрелые бета-клетки экспрессируют INS, NKX6.1 и MAFB, и по существу не экспрессируют NGN3.


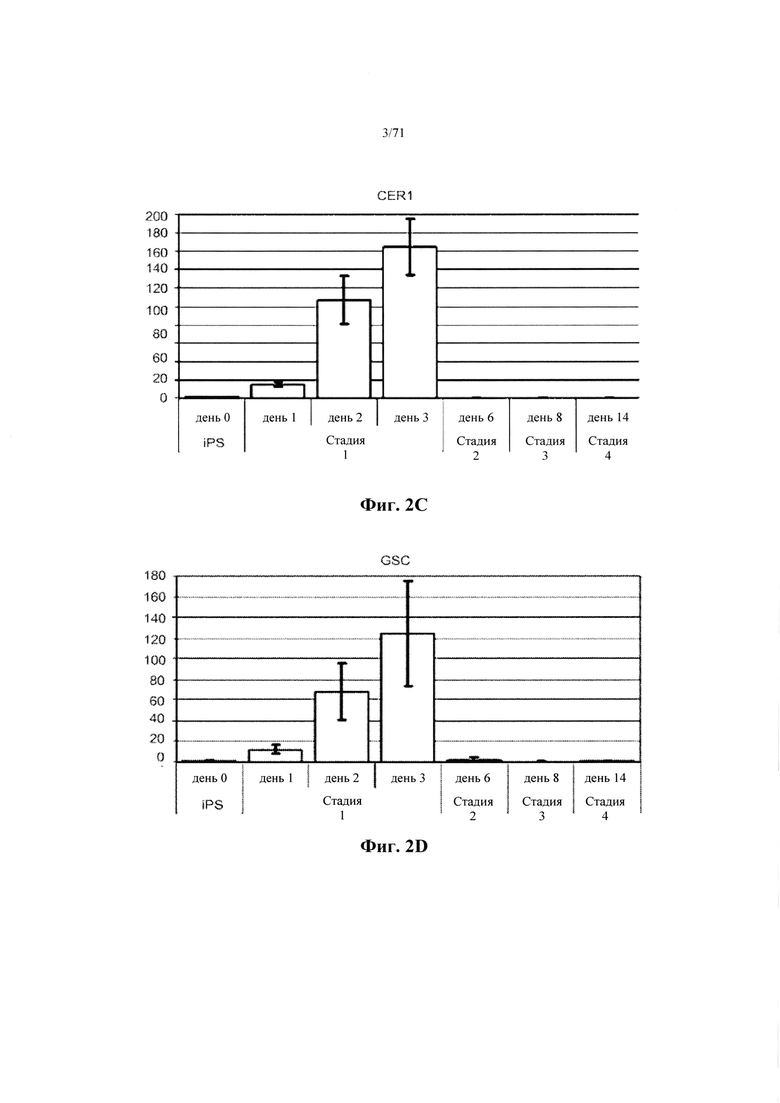


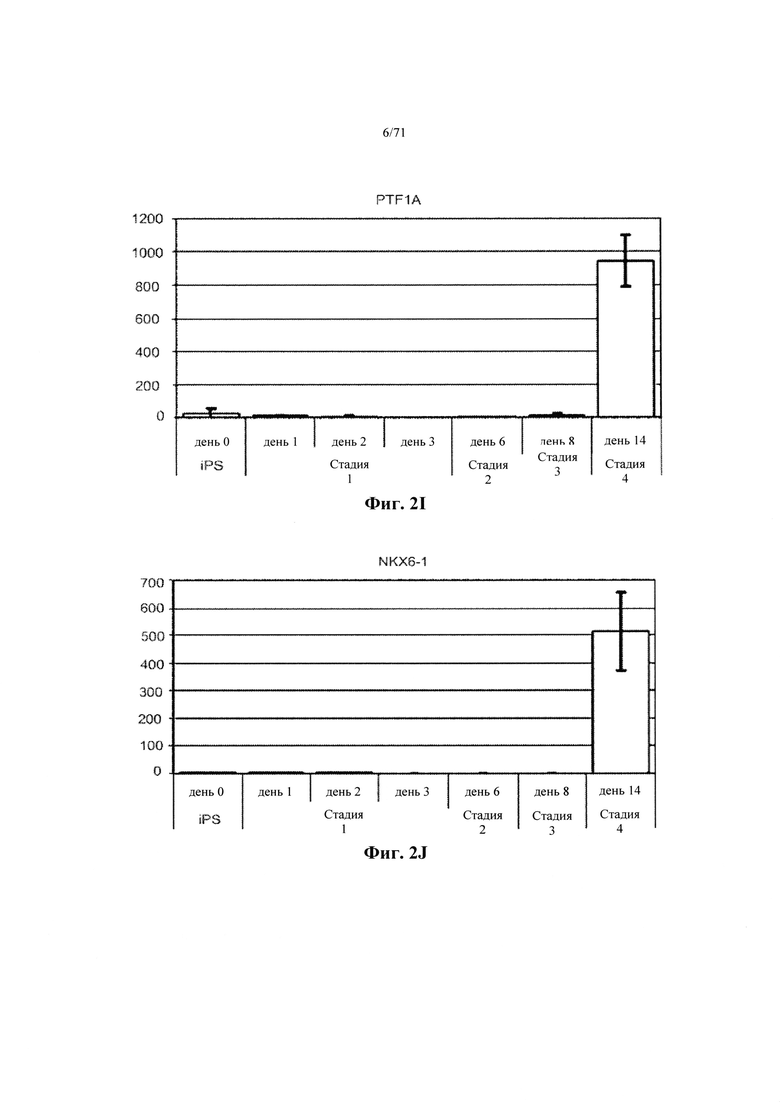























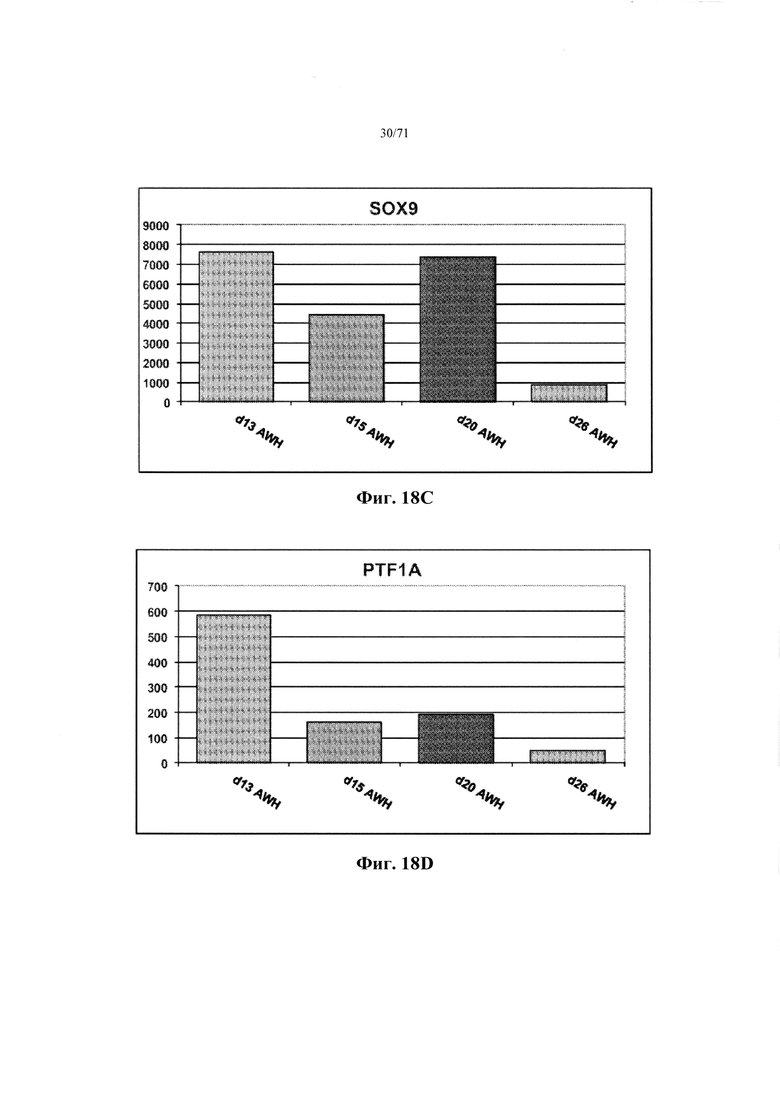

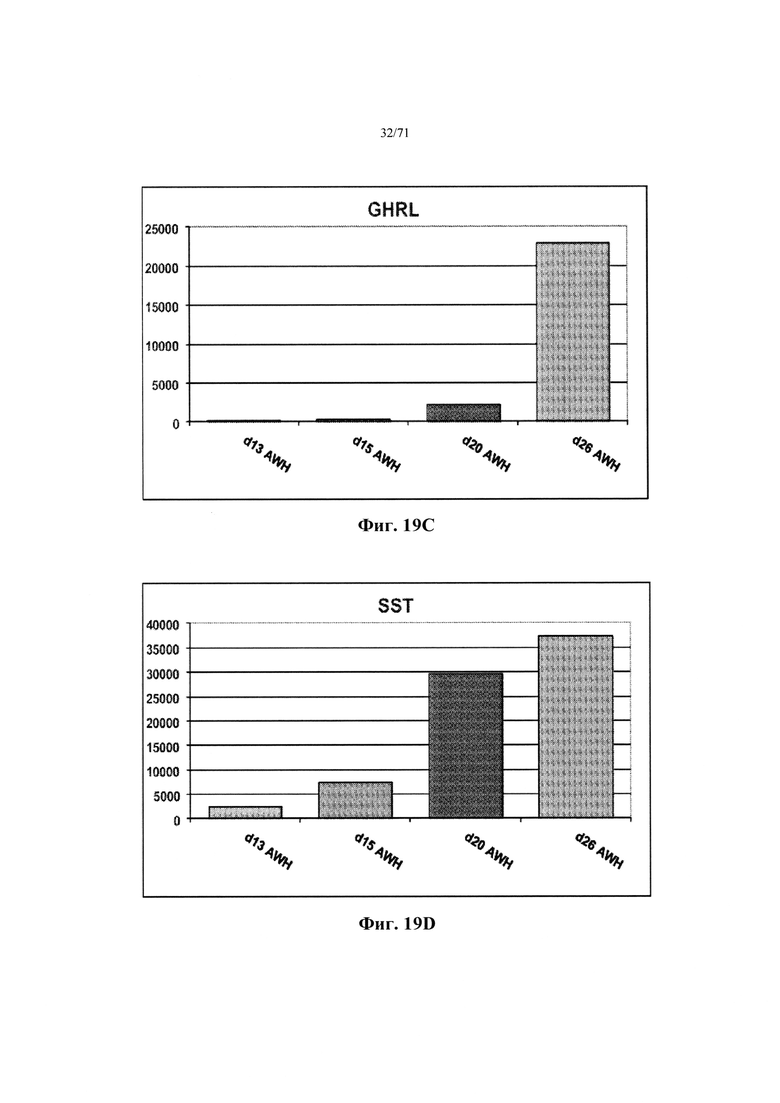
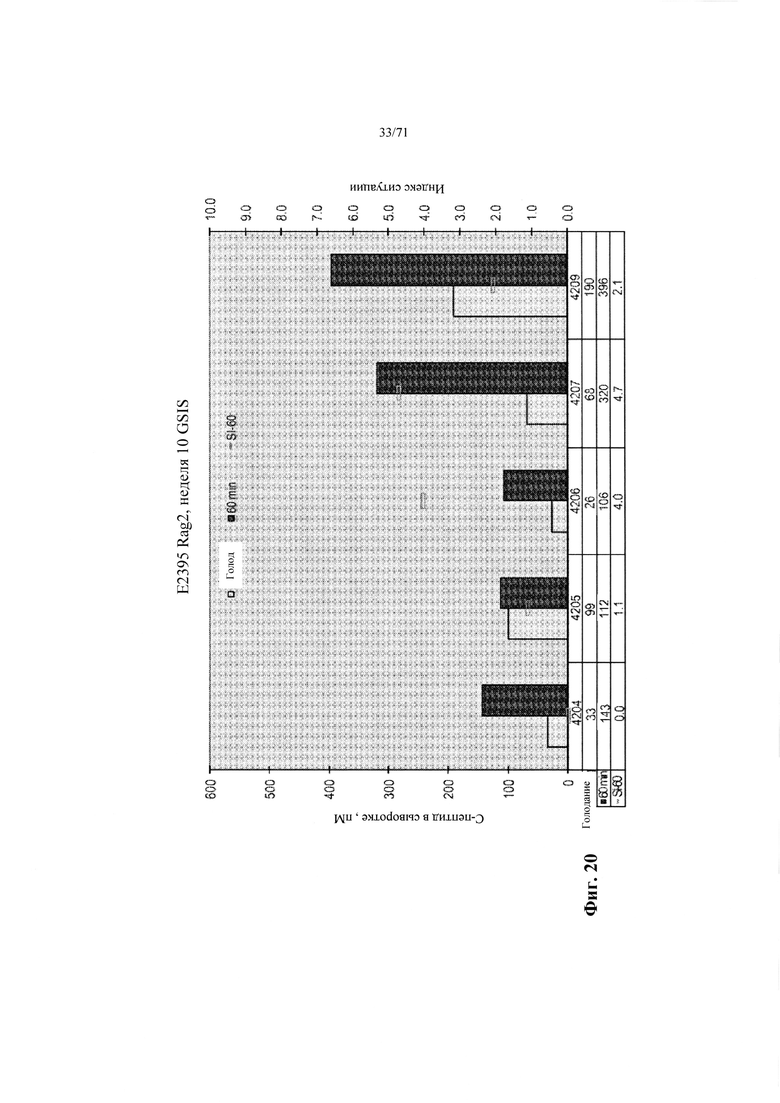

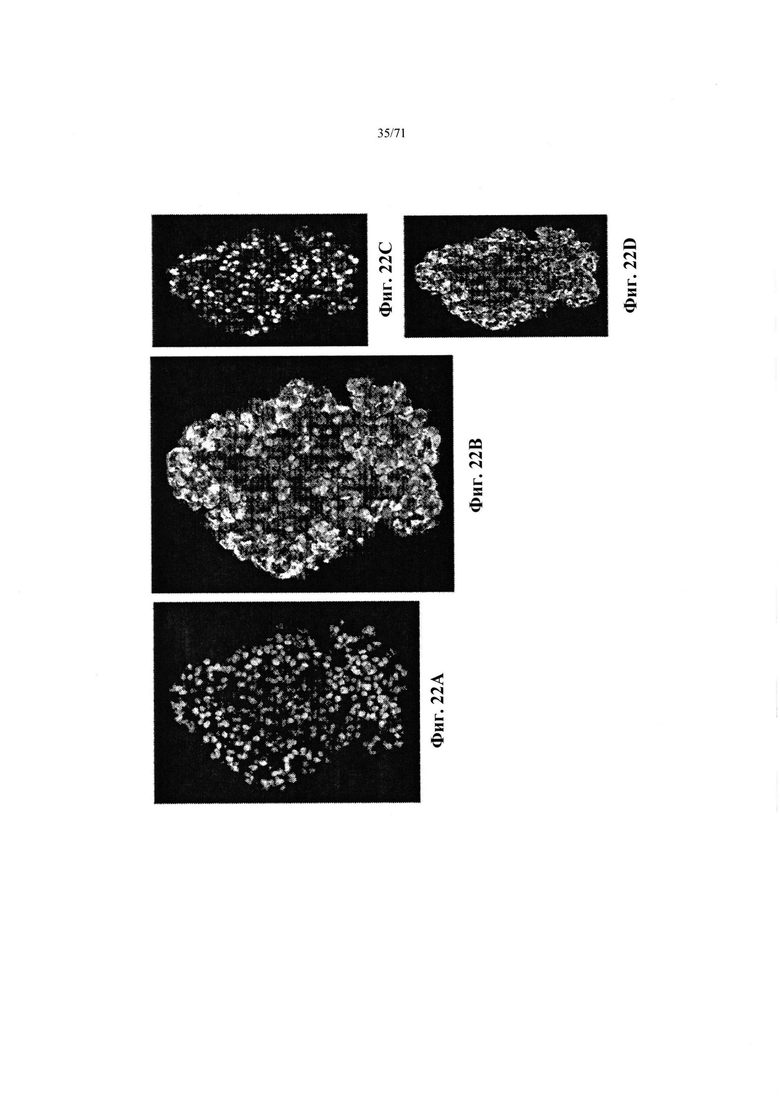













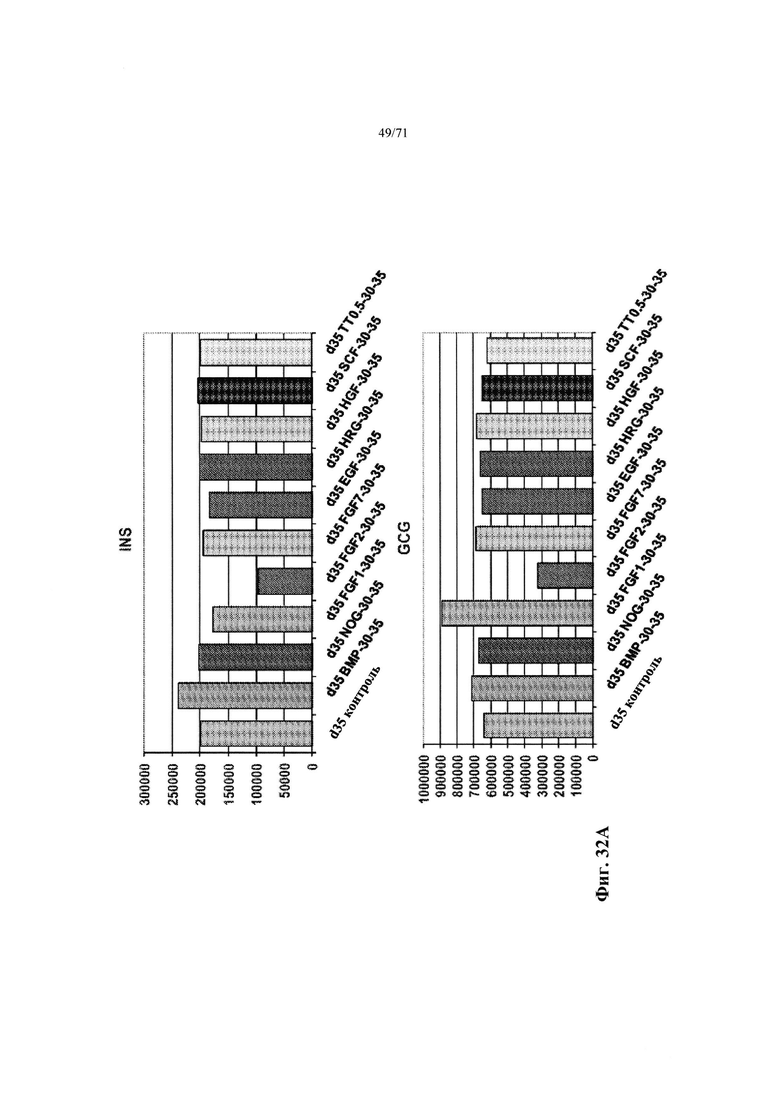





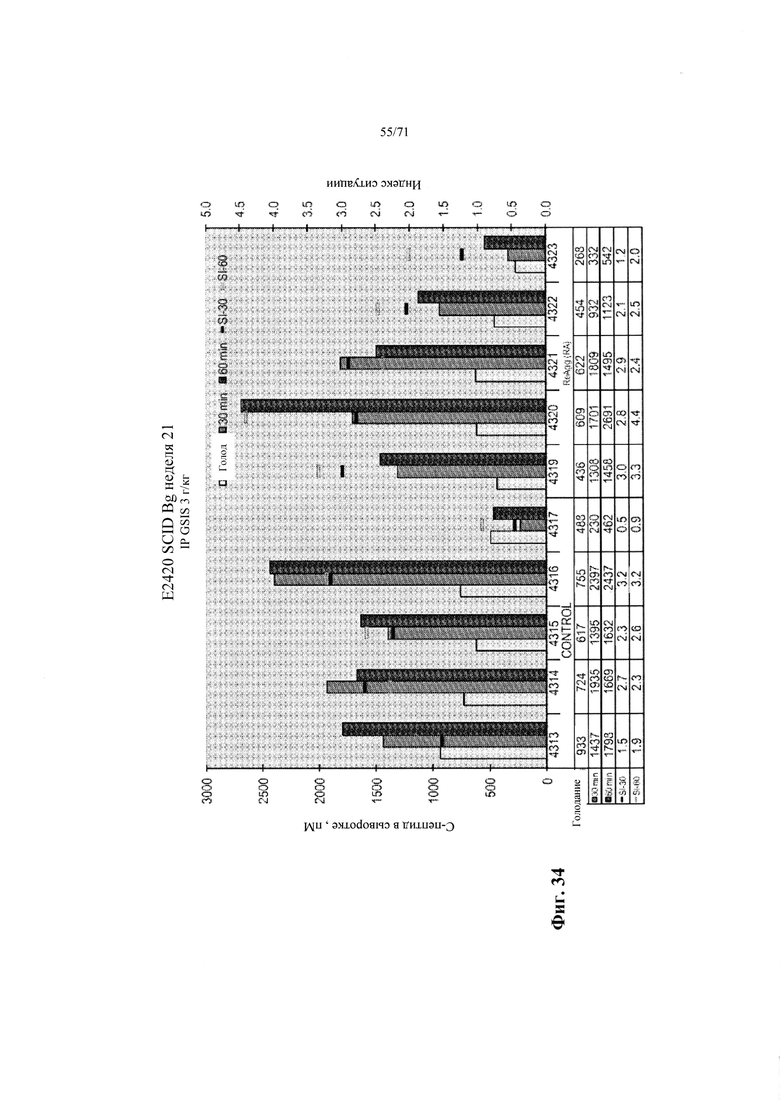





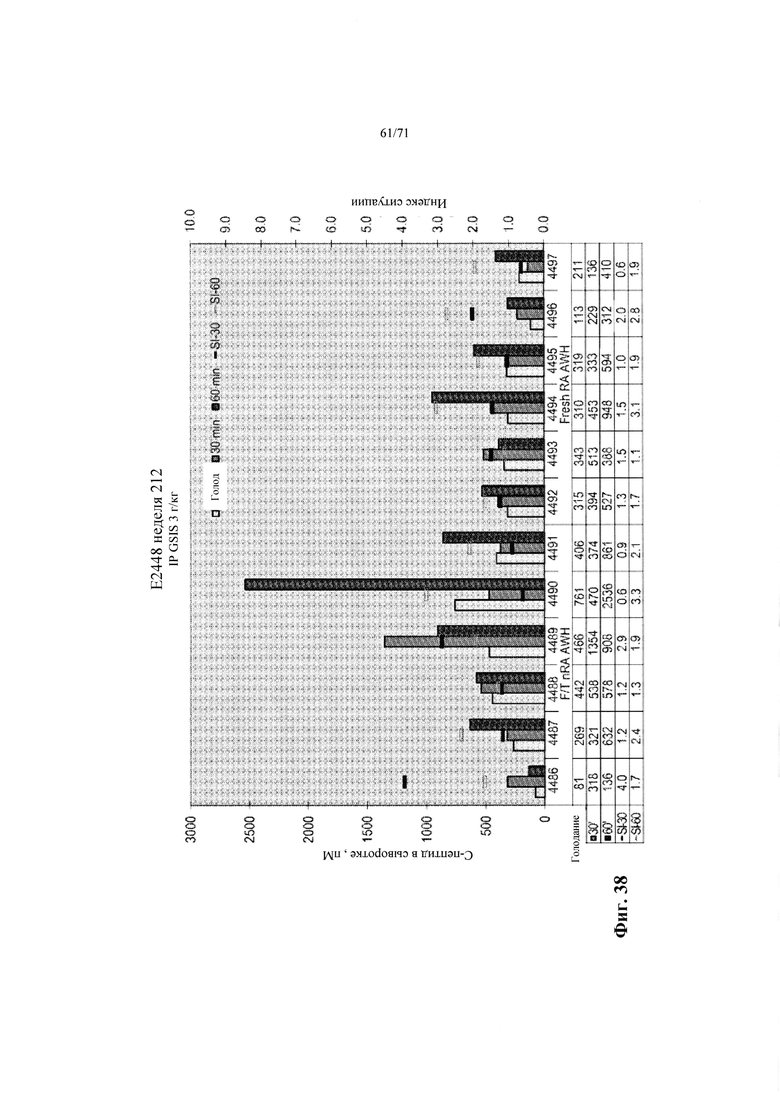


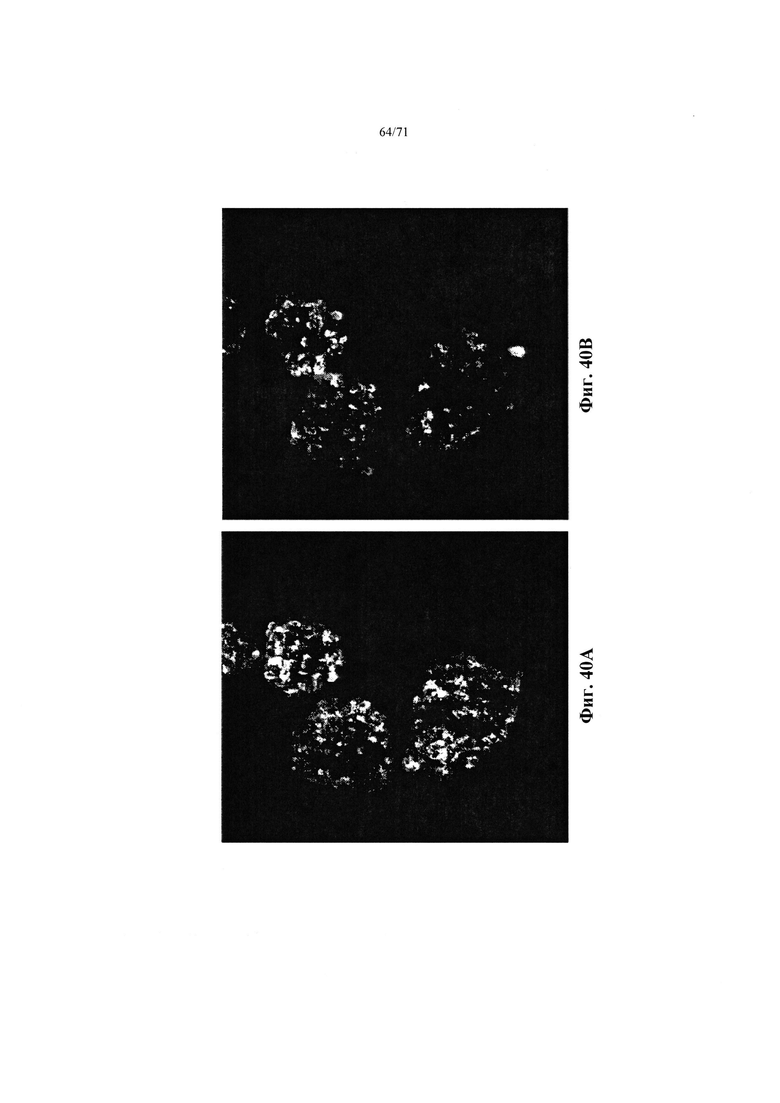
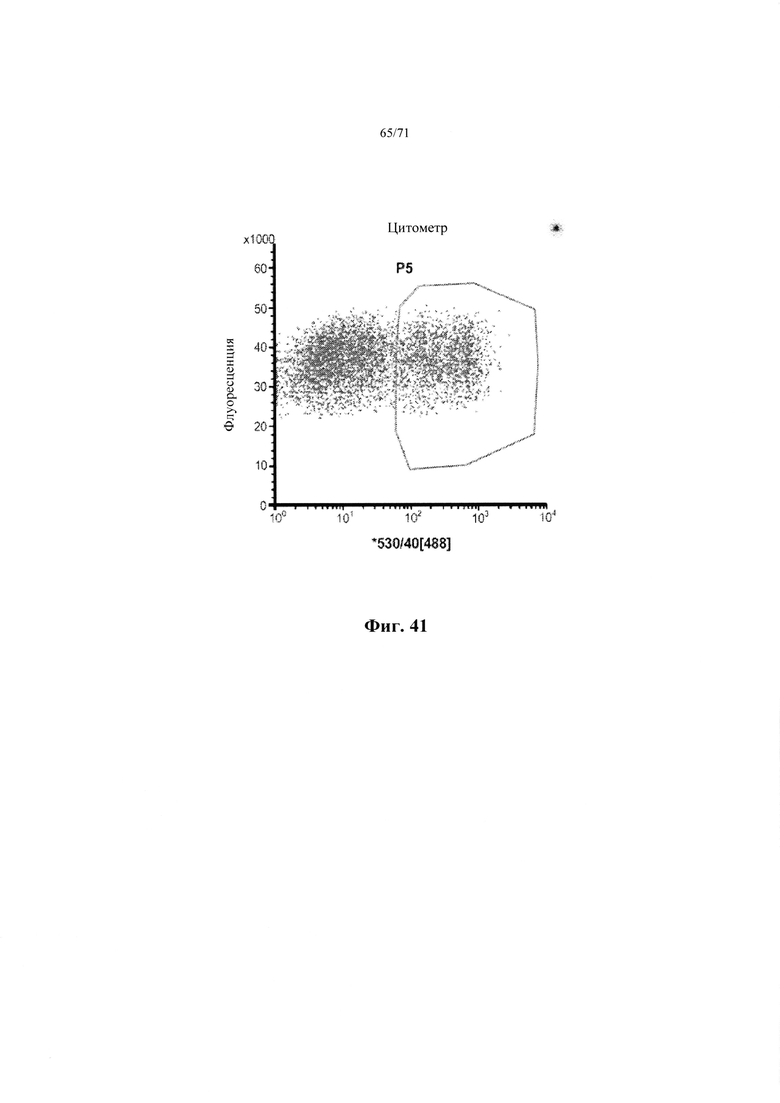





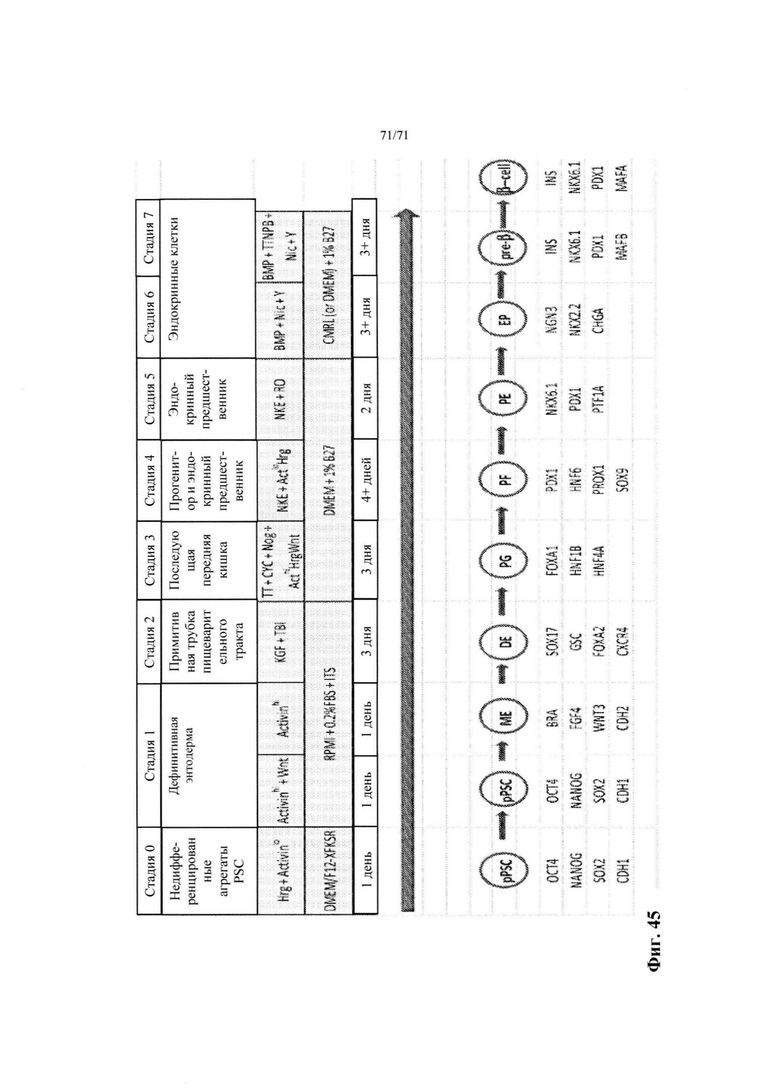
Изобретение относится к области биотехнологии, а именно к композиции клеток человека и криоконсервированным композициям клеток человека in vitro для применения при лечении диабета. Указанные композиции содержат эндокринные клетки или суспензионные агрегаты, содержащие эндокринные клетки, полученные без использования эмбриона человека, в которых по меньшей мере 10% или 50% клеток являются незрелыми бета-клетками, экспрессирующими INS и NKX6.1. Также композиция и криоконсервированная композиция содержат Y-27632 в эффективном количестве, способствующем выживанию указанных клеток в указанной композиции клеток. Изобретение позволяет расширить арсенал технических средств. 4 н. и 14 з. п. ф-лы, 23 табл., 71 ил., 24 пр.
1. Композиция клеток человека in vitro для применения при лечении диабета, содержащая: эндокринные клетки, где по меньшей мере 10% указанных клеток человека являются незрелыми бета-клетками, экспрессирующими INS и NKX6.1; и Y-27632 в эффективном количестве, способствующем выживанию указанных клеток в указанной композиции клеток, при этом из указанных клеток человека исключены клетки, выделенные из стволовых клеток эмбрионов человека, полученных исключительно способом, требующим использование указанного эмбриона.
2. Композиция клеток по п. 1, отличающаяся тем, что указанная композиция клеток находится в полупроницаемом инкапсулирующем устройстве.
3. Композиция клеток по п. 1 или 2, отличающаяся тем, что указанные незрелые бета-клетки человека по существу не экспрессируют NGN3.
4. Криоконсервированная композиция клеток человека in vitro для применения при лечении диабета, содержащая: суспензионные агрегаты, содержащие эндокринные клетки, где по меньшей мере 10% клеток в указанных агрегатах являются унипотентными незрелыми бета-клетками, экспрессирующими INS и NKX6.1; и криоконсервирующий агент, при этом из указанных клеток человека исключены клетки, выделенные из стволовых клеток эмбрионов человека, полученных исключительно способом, требующим использование указанного эмбриона.
5. Композиция по п. 4, отличающаяся тем, что указанные суспензионные агрегаты находятся в полупроницаемом инкапсулирующем устройстве.
6. Композиция по п. 4 или 5, отличающаяся тем, что указанные суспензионные агрегаты имеют размер от 50 до 600 мкн.
7. Криоконсервированная композиция клеток человека in vitro для применения при лечении диабета, содержащая: эндокринные клетки, где по меньшей мере 10% указанных клеток человека являются незрелыми бета-клетками, экспрессирующими INS и NKX6.1; и криоконсервирующий агент, при этом из указанных клеток человека исключены клетки, выделенные из стволовых клеток эмбрионов человека, полученных исключительно способом, требующим использование указанного эмбриона.
8. Композиция клеток по п. 7, отличающаяся тем, что указанные незрелые бета-клетки человека по существу не экспрессируют NGN3.
9. Композиция клеток по п. 7 или 8, отличающаяся тем, что указанные незрелые бета-клетки находятся в полупроницаемом инкапсулирующем устройстве.
10. Композиция клеток по любому из пп. 7-9, отличающаяся тем, что указанные незрелые бета-клетки находятся в перфорированном инкапсулирующем устройстве.
11. Композиция клеток по любому из пп. 7-10, отличающаяся тем, что указанные незрелые бета-клетки человека дополнительно экспрессируют панкеротический диоденальный гомеобоксфактор-1 (PDX-1).
12. Композиция клеток по любому из пп. 7-11, отличающаяся тем, что указанные незрелые бета-клетки человека обладают способностью к созреванию в зрелые бета-клетки.
13. Композиция клеток по любому из пп. 7-12, отличающаяся тем, что указанные незрелые бета-клетки дополнительно экспрессируют MAFB.
14. Композиция клеток по любому из пп. 7-13, дополнительно содержащая хромогранин А(СНGА)-отрицательные клетки.
15. Композиция клеток по любому из пп. 7-14, дополнительно содержащая соединение, выбранное их группы, включающей костный морфогенный белок, белок "sonic hedgehog", фактор роста тромбоцитов, член суперсемейства FGF, ретиноевую кислоту, аналоги ретиноевой кислоты, инсулиноподобный фактор роста, производное витамина В3, и никотинамид.
16. Композиция клеток по любому из пп. 7-15, дополнительно содержащая матрикс.
17. Криоконсервированная композиция клеток человека in vitro для применения при лечении диабета, содержащая: эндокринные клетки, где по меньшей мере 50% указанных клеток человека являются незрелыми бета-клетками, экспрессирующими INS и NKX6.1; и криоконсервирующий агент, при этом из указанных клеток человека исключены клетки, выделенные из стволовых клеток эмбрионов человека, полученных исключительно способом, требующим использование указанного эмбриона.
18. Композиция клеток по п. 17, отличающаяся тем, что указанные незрелые бета-клетки находятся в полупроницаемом инкапсулирующем устройстве.
| DONGHUI Z., et al | |||
| "Highly efficient differentiation of human ES cells and iPS cells into mature pancreatic insulin-producing cells", Cell Research | |||
| Колосоуборка | 1923 |
|
SU2009A1 |
| US5453357A, 26.09.1995 | |||
| US20030087919A1, 08.05.2003 | |||
| ВЕЧКАНОВ Е.М | |||
| и др | |||
| Основы клеточной инженерии: Учебное пособие | |||
| - Ростов-на-Дону, 2012 | |||
| Регулятор для ветряного двигателя в ветроэлектрических установках | 1921 |
|
SU136A1 |
| КОЖУХАРОВА И.В | |||
| "Новые линии | |||

