Область техники, к которой относится изобретение
Изобретение относится к области молекулярной биологии, вирусологии, медицине и ветеринарии и может быть использовано для идентификации вирусов гриппа А и В, типирования и субтипирования вирусов гриппа А с одновременным определением генетических маркеров патогенности и устойчивости к противогриппозным препаратам. Изобретение также включает биологический микрочип (биочип) для осуществления способа и набор олигонуклеотидных зондов для иммобилизации в элементах биочипа.
Уровень техники
Вирусы гриппа (типы А, В и С) относятся к семейству Ortomyxoviridae. Они циркулируют в природных резервуарах, вызывая инфекцию у человека, птиц, многих видов млекопитающих; и являются причиной сезонных эпидемий и периодических пандемий, которые уносят миллионы жизней. Наибольшую эпидемиологическую опасность представляет вирус гриппа типа А (далее по тексту - ВГА). Различные генетические варианты вирусов гриппа (штаммы) различаются так называемыми сайтами патогенности - нуклеотидными последовательностями в составе РНК, кодирующими синтез факторов, обуславливающих тяжесть заболевания, контагиозность, восприимчивость к лекарственным противовирусным средствам. Высокая изменчивость вируса приводит к появлению с периодичностью в 2-3 года новых патогенных для человека генетических вариантов и мутаций, изменяющих устойчивость к противогриппозным препаратам.
Для борьбы с гриппом существуют две стратегии: вакцинация и лекарственная терапия. Эффективность вакцинации ограничена изменениями, происходящими в геноме вируса от сезона к сезону, а действие лекарственных средств - возникновением устойчивых штаммов. Основные препараты, применяемые для терапии вируса, действуют на разные мишени: белок М2, образующий ионный канал (препараты группы адамантанов, например, амантадин, ремантадин) (Jefferson T., Demicheli V., et al., Amantadine and rimantadine for influenza A in adults. Cochrane Database of Syst. Rev., 2006. 2, CD001169); нейраминидазу, отщепляющую остаток N-ацетилнейраминовой кислоты (препараты группы ингибиторов нейраминидазы, например, ланинамивир, озельтамивир, перамивир, занамивир) (Ikematsu H., Kawai N. Laninamivir octanoate: a new long-acting neuraminidase inhibitor for the treatment of influenza. Expert Review of Anti-infective Therapy, 2011. v. 9 (10), p. 851-857).
Появление штаммов, обладающих выраженным пандемическим потенциалом, а также не снижающийся уровень заболеваемости гриппом обусловливают необходимость поиска новых высокочувствительных и надежных способов определения вирусного подтипа для прогнозирования эпидемий, осуществления эпидемиологического контроля, своевременного создания противогриппозных вакцин, учитывающих генетические особенности возбудителя грядущей эпидемии.
Известен ряд методов для определения подтипов вируса гриппа, дифференциации подтипов гемагглютинина и нейраминидазы, определения чувствительности к противогриппозным препаратам.
(1) Выделение вируса гриппа на развивающихся куриных эмбрионах или клеточных линиях с последующим анализом. Наиболее распространенными методами идентификации подтипов ВГА являются реакция торможения гемагглютинации (РТГА) с антисыворотками к эталонным штаммам вируса гриппа для определения подтипа гемагглютинина и реакция ингибирования нейраминидазной активности (РИНА) - для определения подтипа нейраминидазы (WHO Manual on animal influenza diagnosis and surveillance. WHO, Geneva, 2002, document WHO/CDS/CSR/NCS/2002.5 Rev.1, http://www.who.int). Метод явдяется "золотым стандартом" при определении вирусного подтипа (Stamboulian D., Bonvehi Р. Ε., Nacinovich F. M., Сох N. Influenza Infect. Dis. Clin. North. Am., 2000, v. 14., p. 141-166), и также используется для определения патогенности и устойчивости вирусов гриппа к противовирусным препаратам (Bouvier N.M., Lowen А.С. Animal models for influenza virus pathogenesis and transmission. Viruses, 2010, v. 2 (8), p. 1530-1563).
Преимуществом культивирования вируса является то, что вирус может быть использован для дальнейших исследований, молекулярно-генетической характеризации и др. Недостатком является повышенная биологическая опасность, трудоемкость и длительное время, требуемое для культивирования (от 3 до 7 дней).
(2) Методы, основанные на использовании поли- или моноклональных антител, иммуноферментный и иммунофлуоресцентный анализ (Belshe R.B., Smith M.H., Hall C.B., Betts R. Hay A.J. Genetic basis of resistance to rimantadine emerging during treatment of influenza virus infection. J. Virol., 1988, v. 62, p. 1508-1512; Tkaceova M., Varenckova E., Baker I.C., Love J.M., Ziegler T. Evaluation of monoclonal antibodies for subtyping of currently circulating human type A Influenza viruses. J. Clin. Microbiol, 1997, p. 1196-1198). Методы с использованием антител требуют наличия набора специфичных антител для каждого подтипа ВГА, которые дорогостоящи и трудны в получении, и применяются для ограниченного количества подтипов.
(3) Методы определения чувствительности вирусов к различным лекарственным препаратам, включающие:
- метод подавления бляшкообразования (Matrosovich M., Matrosovich Т., Garten W., Klenk H.-D. New low-viscosity overlay medium for viral plaque assays. Virology Journal, 2006, v. 3, p.63). Применяется преимущественно для определения устойчивости вирусов гриппа к препаратам группы адамантанов и не подходит для определения устойчивости к ингибиторам нейраминидазы;
- МТТ колориметрический тест: оценка клеточной и лекарственной цитотоксичности, основанная на способности митохондриальных дегидрогеназ конвертировать водорастворимый 3-(4,5-диметилтиазол-2-ил)-2,5-дифенил-2Н-тетразолиум бромид (МТТ) в формазан, который кристаллизуется внутри клетки (Watanabea W., Konnoa K., Ijichi K., Inouea H., et al. МТТ colorimetric assay system for the screening of anti- orthomyxo-and anti-paramyxoviral agents. J. Virol. Methods, 1994, v. 48, p. 257-265). Метод требует наличия специальной аппаратуры и не обладает достаточной чувствительностью;
- анализ активности нейраминидазы хемилюминесцентным (Buxton R.C., Edwards В., Juo R. R., et al. Development of a sensitive chemiluminescent neuraminidase assay for the determination of influenza virus susceptibility to zanamivir. Anal. Biochem., 2000, v. 280, p. 291-300) и флюорометрическим методом (Potier M., Mameli L., Belisle M., Dallaire L., Melançon S.B. Fluorometric assay of neuraminidase with a sodium (4-methylumbelliferyl-α-D-N-acetylneruaminate) substrate. Anal. Biochem., 1979, v. 94, p. 287-296). Методы не применимы для анализа резистентности к препаратам группы адамантанов;
- проточная цитометрия инфицированных клеток (McSharry, J.J. McDonough А.С, Olson B.A., Drusano G.L. Phenotypic drug susceptibility assay for influenza virus neuraminidase inhibitors. Clin. Diagn. Lab. Immunol., 2004, v. 11, p. 21-28). Требуется дорогостоящее специальное оборудование (проточный цитофлюориметр).
В последнее время все более широкое распространение получают молекулярно-генетические методы анализа, основанных на анализе последовательностей вирусного генома, включая последовательности генов НА и NA и сайты патогенности (Wang R., Taubenberger J.K. Methods for molecular surveillance of influenza. Expert. Rev. Anti Infect. Ther., 2010, v. 8(5), p. 517-527). Проводится амплификация соответствующих участков генома с последующим анализом результатов различными методами.
(4) Полное или частичное секвенирование вирусных генов. Описана амплификация любых возможных подтипов ВГА с помощью универсальной праймерной системы с последующим секвенированием (Hoffmann Ε., Stech J., Guan Y., Webster R.G., Perez D.R. Universal primer set for the full-length amplification of all influenza A viruses. Arch. Virol., 2001, v. 146, p. 2275-2289). Для исследования резистентности вируса к препаратам группы адамантанов использовали метод прямого секвенирования сегментов MP и NA (Laplante J.M., Marshall S.A., Shudt M., Van T.T., et al. Influenza antiviral resistance testing in New York and Wisconsin, 2006-2008: methodology and surveillance data. J. Clin. Microbiol., 2009, v. 47, p.1372-1378; Bright R.A., Shay D.K., Shu В., Cox N.J., Klimov A.I. Adamantane resistance among influenza A viruses isolated early during the 2005-2006 influenza season in the United States. JAMA, 2006, v. 295, p. 891-894, а также метод пиросеквенирования этих же сегментов (Deyde V.M., Tung Ν., Bright R.A., et al. Detection of molecular markers of antiviral resistance in influenza A (H5N1) viruses using a pyrosequencing method. Antimicrob. Agents Chemother., 2009, v. 53 (3), p. 1039-1047).
К недостаткам методов, основанных на секвенировании, относится их трудоемкость, дороговизна, необходимость дорогостоящего оборудования (например, автоматический секвенатор), а также значительные затраты времени и реагентов на дополнительную очистку продуктов амплификации перед проведением секвенирования.
(5) Проведение полимеразной цепной реакции (ПЦР) и последующий электрофоретический анализ продуктов (Poddar S.K. Influenza virus types and subtypes detection by single step single tube multiplex reverse transcription-polymerase chain reaction (RT-PCR) and agarose gel electrophoresis, J. Virol. Methods, 2002 v. 99 (1-2), p. 63-70).
(6) Проведение ПЦР и рестрикции с последующим анализом полиморфизма длин рестрикционных фрагментов (L.A. Cooper, K. Subbarao. A simple restriction fragment length polymorphism-based strategy that can distinguish the internal genes of human H1N1, H3N2, and H5N1 influenza A viruses. J. Clin. Microbiol., 2000, p. 2579-2583), Suzuki H., Saito R., Masuda H., Oshitani H., Sato M., Sato I. Emergence of amantadine-resistant influenza A viruses: epidemiological study. J. Infect. Chemother., 2003, v. 9, p.195-200).
(7) Изотермическая амплификация (Poon L.L., Leung C.S., Chan K.H. et al. Detection of human influenza A viruses by loop-mediated isothermal amplification. J. Clin. Microbiol., 2005, v. 43, p. 427-430).
(8) ПЦР-гетеродуплексный анализ (Ellis J.S., Zambon M.C. Combined PCR-heteroduplex mobility assay for detection and differentiation of influenza A viruses from different animal species. J. Clin. Microbiol, 2001, p. 4097-4102).
(9) Методы ПЦР с детекцией в режиме реального времени. Используются как для типирования ВГА (Stone В., Burrows J., Schepetiuk S., et al. Rapid detection and simultaneous subtype differentiation of influenza A viruses by real time PCR. J. Virol. Methods, 2004, v. 117, p. 103-112), так и для определения мутаций, приводящих к лекарственной устойчивости ВГА (Hindiyeh M., Ram D., Mandelboim M., Meningher T., et al. Rapid detection of influenza A pandemic (H1N1) 2009 virus neuraminidase resistance mutation H275Y by real-time reverse transcriptase PCR. J Clin. Microbiol., 2010; v. 48 (5), p. 1884-1887; Carr M.J., Sayre N., Duffy M., Connell J., Hall W.W. Rapid molecular detection of the H275Y oseltamivir resistance gene mutation in circulating influenza A (H1N1) viruses. J. Virol. Methods, 2008, v. 153 (2), p. 257-262).
Для ПЦР в реальном времени описаны также методики с количественным определением вируса (Lee C.W., Suarez D.L. Application of real-time RT-PCR for the quantitation and competitive replication study of H5 and H7 subtype avian influenza virus. J. Virol. Methods, 2004, v. 119, p. 151-158).
К недостаткам вышеуказанных ПЦР-методов (5)-(9) относится их трудоемкость, а также ограниченность в числе определяемых подтипов вируса. Метод изучения полиморфизма и ПЦР-гетеродуплексный анализ чрезвычайно усложняются при увеличении количества дифференцируемых подтипов. Метод ПЦР в реальном времени требует постановки отдельных реакций для определения каждой мутации и при этом весьма дорог для рутинного анализа.
(10) Многопараметрический анализ генетического материала с использованием ДНК-микроматриц (микрочипов), представляющих собой массив иммобилизованных на твердой фазе олигонуклеотидов, способных специфично связываться с детектируемыми последовательностями. Для идентификации и типирования вируса гриппа описаны микрочипы двух типов: ДНК-микроматрицы высокой плотности и специализированные олигонуклеотидные микрочипы.
Описано несколько вариантов секвенирующего микрочипа высокой плотности для анализа вируса гриппа и других респираторных патогенов (Lin В., Malanoski А.Р., Wang Ζ., Blaney K.M., et al. Universal detection and identification of avian influenza virus by use of resequencing microarrays. J. Clin. Microbiol., 2009, v. 47 (4), p.988-993; Leclercq I., Berthet N., Batéjat С, Rousseaux С. Use of consensus sequences for the design of high density resequencing microarrays: the influenza virus paradigm, BMC Genomics, 2010, v. 11, p. 586-590). Известны микроматрицы на основе технологии Agilent для общего скрининга вирусов «Virochip», разрабатываемые компанией Agilent technologies, США, содержащие ~36000 зондов, полученные на основе базы данных 1500 вирусов GenBank (Chen Е.С., Miller S.A., DeRisi J.L., Chiu C.Y. Using a pan-viral microarray assay (Virochip) to screen clinical samples for viral pathogens, J. Vis. Exp., 2011 Apr 27; (50), 2536).
Компанией CombiMatrix Corp., США запатентован микрочип для идентификации ряда патогенов, включающих аденовирусы, энтеровирусы, вирус герпеса, SARS, вирус Эпштейна-Барр, вирусы гриппа А и В и др., а также субтипирования ВГА (Microarray for pathogen identification, заявка на патент США №20070092871). Этой же компанией представлен полупроводниковый микрочип высокой плотности для идентификации 15 вариантов гемагглютинина и 9 вариантов нейраминидазы (Lodes M.J., Suciu D., Elliott M., Stover A.G., et al. Use of semiconductor-based oligonucleotide microarrays for influenza a virus subtype identification and sequencing. J. Clin. Microbiol., 2006, v. 44, p. 1209-1218). Около 9000 олигонуклеотидных зондов синтезировались непосредственно на микрочипе, результаты регистрировали с помощью специализированного оптического сканера.
Описаны проточные микрочипы для анализа ВГА на основе микроканалов (Kessler Ν., Ferraris О., Palmer K., Marsh W., Steel A. Use of the DNA flowthruchip, a three-dimensional biochip, for typing and subtyping of influenza viruses. J. Clin. Microbiol., 2004, v. 42, p. 2173-2185). Олигонуклеотидные зонды выбирали на основе анализа консервативных участков геномов 7 вирусов.
Гибридизация на микроматрицах высокой плотности требует сложного компьютерного обсчета полученных результатов и дорогостоящих расходных материалов. Чрезвычайно высокая стоимость микрочипов не позволяет использовать их для рутинной медицинской практики.
В работах различных авторов описаны гибридизационные микрочипы низкой плотности, которые отличаются методами получения, типом используемых олигонуклеотидных зондов, типом детекции. Большинство микрочипов изготавливаются путем нанесения растворов олигонуклеотидов на стеклянную или пластиковую подложку, т.е. олигонуклеотидные зонды находятся на поверхности подложки, однако имеются примеры и микрочипов других типов.
Микрочипы, представляющие собой подложки с иммобилизованными фрагментами ДНК, описаны, например, в работах Li J., Chen S., and Evans D. H. Typing and subtyping influenza virus using DNA microarrays and multiplex reverse transcriptase PCR. J. Clin. Microbiol., 2001, v. 39, p. 696-704; Sengupta S., Onodera K., Lai Α., Melcher U. Molecular detection and identification of influenza viruses by oligonucleotide microarray hybridization. J. Clin. Microbiol., 2003, v. 41, p. 4542-4550. Имеется патентная заявка США по методам и реагентам для детекции, типирования и субтипирования вируса гриппа на основе микрочипов (DNA array analysis as a diagnostic for current and emerging strains of influenza, патентная заявка WO 2008/054830, заявка на патент США №20090124512), а также заявка по детекции устойчивости вируса гриппа к различным антивирусным агентам (Detection of antiviral resistance in influenza a using DNA microarray, заявка WO/2008/079450).
Описан микрочип для идентификации подтипов вируса гриппа по последовательности M гена вируса (Dawson E.D., Moore C.L., Smagala J.A. MChip: a tool for influenza surveillance. Anal. Chem, 2006, v. 78, p. 7610-7615), a также микрочип для анализа мутаций сегмента MP, ведущих к устойчивости к адамантанам (Townsend M., Smagala J., Dawson Ε., Deyde V., et al. Detection of adamantane-resistant influenza on a microarray. J. Clin. Virol, 2008, v. 42, p. 117-123).
Имеется несколько работ российских авторов, в которых описаны микрочипы для анализа вируса гриппа, представляющие собой подложки с нанесенными на них олигонуклеотидными зондами («двумерные микрочипы»). Микрочипы для субтипирования ВГА, разработанные в Институте химической биологии и фундаментальной медицины СО РАН, получают путем нанесения растворов на стеклянный слайд (Рябинин В.А., Костина Е.В., Неверов А.А., Максакова Г.А., Синяков А.Н. Типирование гемагглютинина вируса гриппа А с использованием гибридизационного микрочипа. Биоорг. хим., 2010, т. 36 (6), с. 688-699; Ryabinin V.A., Kostina Ε.V., Maksakova G.A., Neverov A.A., Chumakov K.M., Sinyakov A.N. Universal oligonucleotide microarray for sub-typing of Influenza A virus. PLoS One, 2011, v. 6 (4), e17529). Микрочип содержит зонды для определения H1-, Н2-, Н3-, Н5-, Н7-, Н9-субтипов гемагглютинина и N1-, N2-, N7-субтипов нейраминидазы и, а также зонды, выявляющие ген матриксного белка ВГА для выявления субтипов, патогенных для человека. Получен патент РФ №2470076, дата публикации 20.12.2012, касающийся способа выбора ДНК-зондов для микрочиповой диагностики, биочипа и способа типирования гена нейраминидазы и гемагглютинина ВГА. Способ включает проведение двухэтапной ПЦР кДНК вируса с последующей гибридизацией полученных флуоресцентно меченных ампликонов с ДНК-микрочипом. Теми же авторами опубликована работа по определению субтипов ВГА H1N1, H5N1, H2N2, H3N2, H9N2, H7N9 методом ПЦР на микрочипе: Костина Е.В., Рябинин В.А., Терновой В.А., Синяков А.Н., ПЦР на микрочипе для определения субтипов вируса гриппа А, циркулирующих в человеческой популяции, Биорг. хим., 2015, т. 41 (1). с. 124-128. Микрочипы позволяют анализировать лишь ограниченное число субтипов ВГА. Кроме того, не описано использование данных микрочипов для анализа клинических образцов.
Описан диагностический олигонуклеотидный микрочип для субтипирования вирусов гриппа А (Васин А.В., Сандыбаев Н.Т., Плотникова М.А, et al. Универсальный диагностический олигонуклеотидный микрочип для определения и субтипирования вирусов гриппа А человека и животных. Вопр. вирусол., 2013, т. 58 (5), с. 32-37). Микрочип также получен методом нанесения олигонуклеотидные зондов, содержащих амино-линкер, на подложку с последующей ковалентной пришивкой под действием УФ-излучения.
Получен патент РФ №2538168, дата публикации 10.01.2015 (ЗАО «ИмДи»), касающийся наборов праймеров и зондов для идентификации и типирования вирусов гриппа А и В, биологического микрочипа и тест-системы на основе биочипов. Микрочип и тест-система предназначены только для идентификации и типирования вируса. Субтипирования и анализ мутаций не проводится.
Микрочипы, полученные нанесением на альдегидные стекла описаны в работе китайских авторов Tian M., Tian Y., Li Y., Lu H., Li X., et al., Microarray multiplex assay for the simultaneous detection and discrimination of influenza A and influenza В viruses, Indian J. Microbiol., 2014, v. 54 (2), p. 211-217 (детекция вирусов гриппа А и В, субтипирование субтипов H1 и Н3).
Имеется несколько заявок на получение патентов Китая: Flu/human avian influenza virus detection gene chip and production method and use», заявка CN 101392302; Detection of influenza A virus epidemic isolates, typing gene chip and using method», заявка CN 101487061; Gene chip for detecting influenza A virus and preparation method and application thereof», заявка CN 101701266.
К недостаткам вышеописанных микрочипов низкой плотности относится их неуниверсальность: описаны микрочипы либо только для идентификации и типирования вируса гриппа, либо субтипирования вируса гриппа А, либо для анализа устойчивости к ограниченному числу антивирусных препаратов. Как правило, микрочипы позволяют идентифицировать лишь небольшое количество типов и субтипов вируса.
Микрочипы для молекулярного типирования ВГА на основе трехмерных гидрогелевых элементов разработаны в ИМБ РАН (Москва) (Fesenko Е.Е., Kireyev D.E., Gryadunov D.A., Mikhailovich V.M., et al. Oligonucleotide microchip for subtyping of influenza A virus. Influenza Other Resp. Viruses, 2007, v. 1 (3), p. 121-129; Гейдаров P.H., Фесенко E.E., Шаскольский Б.Л., et al. Определение генетических детерминант устойчивости вируса гриппа А к адамантанам и ингибиторам нейраминидазы на биологическом микрочипе. Докл. Акад. Наук, 2015, т. 460, №1, с. 102-106). Процедура анализа включает двухстадийную амплификацию фрагментов генов НА и NA с последующей гибридизацией на биочипе и флуоресцентной детекцией. Сконструирован микрочип для идентификации 15 подтипов НА и подтипов N1 и N2 нейраминидазы, а также микрочип для выявления мутаций в генах белков М2 (S31N, V27A, L26F и А30Т) и NA (H274Y), приводящих к возникновению устойчивости вируса к адамантанам и ингибиторам нейраминидазы. К недостаткам данных микрочипов относится ограниченное число анализируемых субтипов ВГА, что не позволяет определять все возможные варианты вируса, циркулирующие в данном эпидемиологическом сезоне.
Компанией GENOMICA S.A.U., Испания выпускается тест-система CLART® PneumoVir для детекции и генотипирования наиболее распространенных вирусов, вызывающих респираторные инфекции у человека (аденовирусы, некоторые субтипы ВГА, VSR-A, VSR-B и др., всего 19 вирусов) на основе микроматриц низкой плотности, элементы которых располагают на дне пробирки объемом 1,5 мл (Array-Tube-AT) или колодца стандартного иммунологического стрипа (CL ART Strips-CS). Детекция осуществляется после проведения ОТ-ПЦР с образованием продуктов, меченных биотином, и основана на образовании нерастворимого осадка: Kouni S., Karakitsos P., Chranioti Α., et al. Evaluation of viral co-infections in hospitalized and non-hospitalized children with respiratory infections using microarrays, Clin. Microbiol. Infect., 2013, v. 19 (8), p. 772-777).
Другим примером коммерчески доступных микроматриц для анализа вируса гриппа являются микрочипы FluType производства Alere Technologies GmbH, Германия. Чипы представляют собой микротитровальные стрипы, содержащие иммобилизованные ДНК-зонды для генотипирования ВГА. Возможен одновременный анализ 96 образцов. Тест-система, помимо стрипов (чипов), включает специфические ПЦР-праймеры для типирования, реагенты для детекции на основе фермента пероксидазы: Gall Α., Hoffmann В., Harder T., Grund С, et al. Design and validation of a microarray for detection, hemagglutinin subtyping, and pathotyping of avian influenza viruses. J. Clin. Microbiol, 2009, v. 47, p. 327-334; Bandt D., Monecke S., Scott C, Gall Α., Hoffmann В., Ehricht R. Economic high-throughput-identification of influenza A subtypes from clinical specimens with a DNA-oligonucleotide microarray in an outbreak situation, Mol. Cell. Probes, 2012, v. 26 (1), p. 6-10. Недостатком данных систем также является ограниченное число анализируемых типов вируса.
Многопараметрический анализ достигается также при использовании микрочипов, представляющих собой суспензию флуоресцентно маркированных микросфер (мультиплексная технология хМАР, детекция с использованием технологии Luminex). Окрашивание микросфер смешанными в различных соотношениях флуорофорами позволяет получить множество типов микросфер с различными спектральными характеристиками. Олигонуклеотидные зонды расположены на поверхности микросфер. Описано применение микросфер для субтипирования ВГА и детекции ряда респираторных патогенов (Multiplex detection of respiratory pathogens, заявка WO/2008/143450, заявка на патент США №20090088331; Munro S.B., Kuypers J., Jerome K.R. Comparison of a Multiplex Real-Time PCR Assay with a Multiplex Luminex Assay for Influenza Virus Detection, J. Clin. Microbiol., 2013, v. 51(4), p, 1124-1129).
Имеется также несколько заявок на получение патентов Китая, касающихся метода детекции и типирования вирусов гриппа с использованием технологии хМАР (в данных заявках они называются «жидкими» чипами ("liquid chips")): Method for detecting flu and H5N1 avian influenza virus by using liquid chip, заявка CN 101392298; Method for detecting influenza virus by liquid phase chip, заявка CN 101717830). Предложенные варианты чипа позволяют идентифицировать лишь ограниченное число подтипов ВГА.
Таким образом, многопараметрический анализ образца достигается при использовании микрочипов высокой и низкой плотности. Микроматрицы высокой плотности чрезвычайно дорогостоящи и сложны в изготовлении; в настоящее время они не получили широкого применения и используются только в научных лабораториях. Основным недостатком всех описанных вариантов гибридизационных микрочипов низкой плотности для анализа вируса гриппа является их неуниверсальность: микрочипы позволяют проводить либо только идентификацию вируса, либо только типирование и/или субтипирование ВГА, причем число определяемых подтипов ограничено, либо только анализ чувствительности к некоторым противовирусным препаратам. Все эти недостатки преодолены настоящим изобретением.
Раскрытие изобретения
Предлагаемое изобретение обеспечивает способ идентификации РНК вирусов гриппа А и В с одновременным определением вариантов гемагглютинина и нейраминидазы вируса гриппа А, идентификацией генетических маркеров патогенности и устойчивости к противогриппозным препаратам на биологических микрочипах (биочипах), представляющих собой подложку с гелевыми элементами, содержащими иммобилизованные специфичные олигонуклеотидные зонды. Изобретением также обеспечиваются биочип, используемый в способе идентификации РНК вирусов гриппа, и набор олигонуклеотидных зондов, используемый для получения биочипа. Способ выгодно отличается от способов, известных из уровня техники, тем, что предоставляет возможность одновременно (в одном эксперименте) осуществить идентификацию РНК вируса гриппа и определить его типовую принадлежность (тип А либо тип В), выполнить субтипирование вируса гриппа А с определением молекулярных вариантов генов гемагглютинина и нейраминидазы, молекулярного анализа генетических маркеров, характеризующих степень патогенности вируса гриппа А и его устойчивость к основным противовирусным препаратам. Способ не требует дорогостоящего оборудования и высококвалифицированного персонала.
Анализ наличия в геноме вируса гриппа генетических маркеров патогенности важен для оценки степени патогенности возбудителя для человека и его восприимчивости к основным противовирусным препаратам. Способ может использоваться для эпидемиологического мониторинга заболевания гриппом и разработки стратегии вакцинации и лечения в каждый конкретный эпидемический сезон.
Процедура определения в предлагаемом способе основана на проведении реакции обратной транскрипции для получения кДНК с использованием вирусной РНК в качестве матрицы, далее проведение полимеразной цепной реакции (ПНР) с последующей гибридизацией полученного одноцепочечного флуоресцентно-меченного ПЦР-продукта на биологическом микрочипе. Биологический микрочип представляет собой подложку с упорядоченно расположенными гидрогелевыми элементами, содержащими ковалентно иммобилизованные олигонуклеотидные зонды, которые обеспечивают гибридизацию со специфическими участками генома вируса гриппа, определяющими его тип, субтип и генетические маркеры, включая детерминанты устойчивости к противовирусным препаратам.
Визуализация результатов гибридизации проводится флуоресцентным методом с помощью возбуждения флуоресценции после реакции меченого ПЦР-продукта с иммобилизованными на микрочипе зондами. Интерпретация результатов осуществляется в зависимости от того, с какими из иммобилизованных зондов прошла реакция гибридизации.
Предлагаемый способ включает:
а) обеспечение биочипа, представляющего собой подложку с гелевыми элементами, в каждом из которых иммобилизован набор олигонуклеотидных зондов, относящийся к следующим группам:
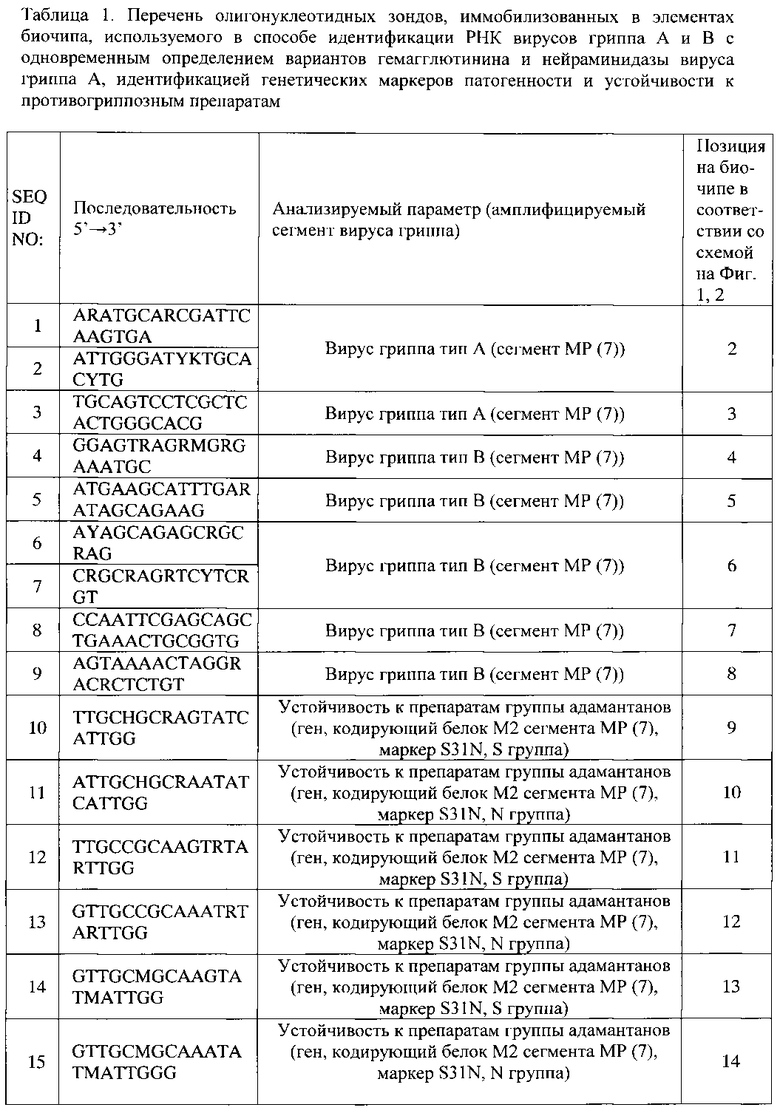
- группа зондов, последовательности которых представлены SEQ ID NO: 1-9 (Таблица 1), соответствующая фрагментам вирусных сегментов MP (7), для определения наличия РНК вируса гриппа и определения типа вируса (тип А или тип В);
- группа зондов, последовательности которых представлены SEQ ID NO: 112-178 (Таблица 1), соответствующая фрагментам вируса гриппа А, для определения молекулярного варианта гемагглютинина (Н1-Н16);
- группа зондов, последовательности которых представлены SEQ ID NO: 73-111 (Таблица 1), соответствующая фрагментам вируса гриппа А, для определения молекулярного варианта нейраминидазы (N1-N9);
- группа зондов, последовательности которых представлены SEQ ID NO: 9-25 (Таблица 1), для идентификации генетических маркеров S31N, V27A, А30Т, L26F в гене М2 вирусного сегмента MP вируса гриппа А, ассоциированных с устойчивостью вируса к лекарственным препаратам группы адамантанов;
- группа зондов, последовательности которых представлены SEQ ID NO: 26-37 (Таблица 1), для идентификации генетического маркера H274Y в гене нейраминидазы вируса гриппа А субтипа N1, ассоциированных с устойчивостью вируса к лекарственным препаратам группы ингибиторов нейраминидазы;
- группа зондов, последовательности которых представлены SEQ ID NO: 66-72 (Таблица 1), для определения состояния сайта протеолитического расщепления гемагглютинина вируса гриппа А, ассоциированного с повышенной патогенностью вируса (для субтипов Н5 и Н7);
- группа зондов, последовательности которых представлены SEQ ID NO: 38-51 (Таблица 1), для анализа генетических маркеров в рамке считывания вирусного белка PB1-F2, который является одним из ключевых факторов подавления вирусом гриппа иммунного ответа хозяина, включающих последовательности для идентификации мутации N66S и стоп-кодонов в 12 и 58 положении;
- группа зондов, последовательности которых представлены SEQ ID NO: 52-65 (Таблица 1), для анализа генетических маркеров в структуре PDZ-связывающего домена в гене NS1, детерминирующего активацию сигнальных систем клетки, обеспечивающих контроль над противовирусным ответом при гриппозной инфекции, включающих последовательности для анализа длины белка NS1 (LVT-0, LVT-11, LVT-13), последовательности аминокислот RSKV (низкопатогенный вариант вируса), RSEV (низкопатогенный вариант вируса), KSEV (высокопатогенный вариант вируса), ESEV (высокопатогенный вариант), ESKV (высокопатогенный вариант вируса), EPEV (высокопатогенный вариант вируса);
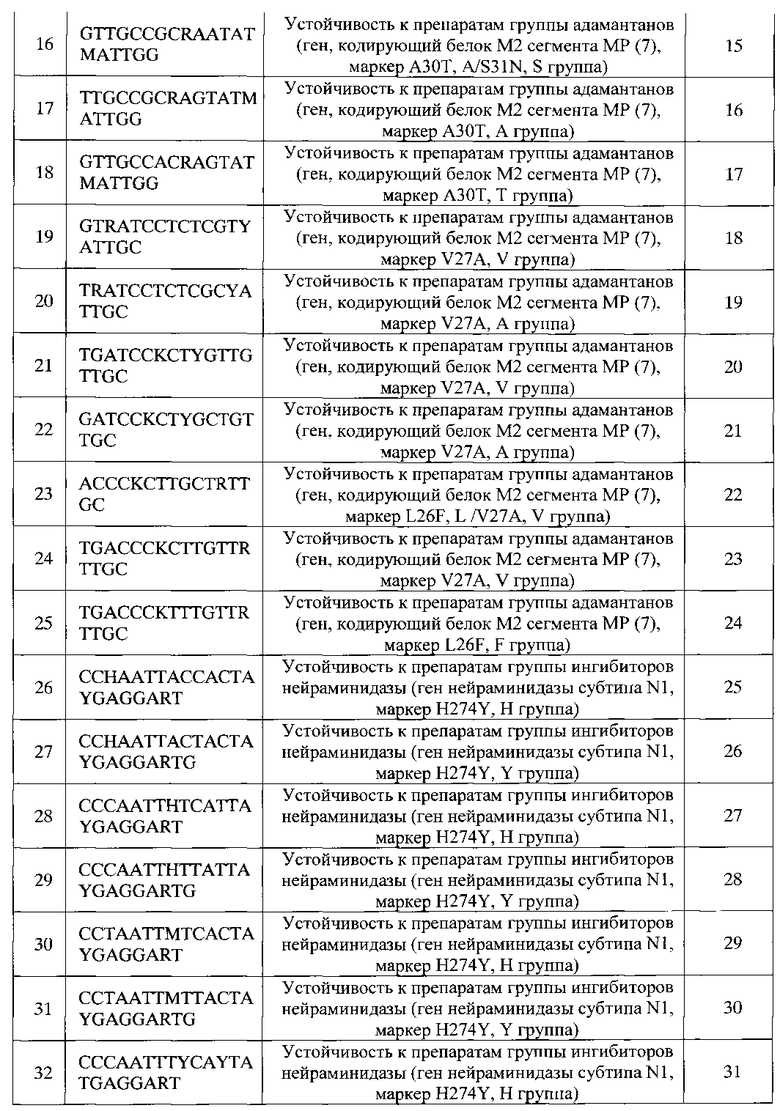
б) проведение реакции обратной транскрипции для получения кДНК с использованием вирусной РНК в качестве матрицы и далее ПЦР с использованием олигонуклеотидных праймеров, комплементарных фрагментам генома вируса гриппа, в буферном растворе, включающем смесь четырех дезоксинуклеозидтрифосфатов, один из которых содержит флуоресцентную метку, с получением продуктов амплификации в виде преимущественно одноцепочечных флуоресцентно-меченных фрагментов ДНК;
в) гибридизацию амплифицированных флуоресцентно-меченных продуктов, полученных на стадии (б), на биочипе, полученном на стадии (а);
г) регистрацию и интерпретацию результатов гибридизации на биочипе, проведенной на стадии (в), путем сравнения интенсивности флуоресценции сигналов в пределах каждой из групп ячеек, в которых образовались совершенные и несовершенные гибридизационные дуплексы.
Для анализа по данному способу используется РНК, выделенная из биологического материала, полученного от человека (мазки из полости носа и ротоглотки, бронхоальвеолярный лаваж, промывные воды бронхов, секционный материал) и от птиц (клоакальные мазки, фрагменты внутренних органов) или культуры вирусов. ПЦР на стадии (б), в результате которой осуществляется получение одноцепочечных фрагментов вирусного генома и их флуоресцентное маркирование за счет встраивания флуоресцентного красителя в продукт.
Регистрацию результатов на стадии (г) проводят с помощью прибора-анализатора флуоресценции, оснащенного специализированным программным обеспечением, что позволяет использовать программную автоматическую обработку интенсивностей сигналов с последующей интерпретацией результатов.
Изобретение также обеспечивает биочип, используемый в способе идентификации РНК вирусов гриппа А и В с одновременным определением вариантов гемагглютинина и нейраминидазы вируса гриппа А, идентификацией генетических маркеров патогенности и устойчивости к противогриппозным препаратам, представляющий собой подложку с гелевыми элементами, в каждом из которых иммобилизован набор зондов, причем последовательности зондов представлены SEQ ID NO: 1-178 (Таблица 1).
Изобретением также предлагается набор олигонуклеотидных зондов, используемый для получения биочипа, используемого в способе идентификации РНК вирусов гриппа А и В с одновременным определением вариантов гемагглютинина и нейраминидазы вируса гриппа А, идентификацией генетических маркеров патогенности и устойчивости к противогриппозным препаратам, причем зонды имеют последовательности SEQ ID NO: 1-178 (Таблица 1).
Далее изобретение будет раскрыто подробнее со ссылками на фигуры и примеры, которые приводятся исключительно с целью иллюстрации и пояснения сущности заявленного изобретения, но которые не предназначены для ограничения объема притязаний.
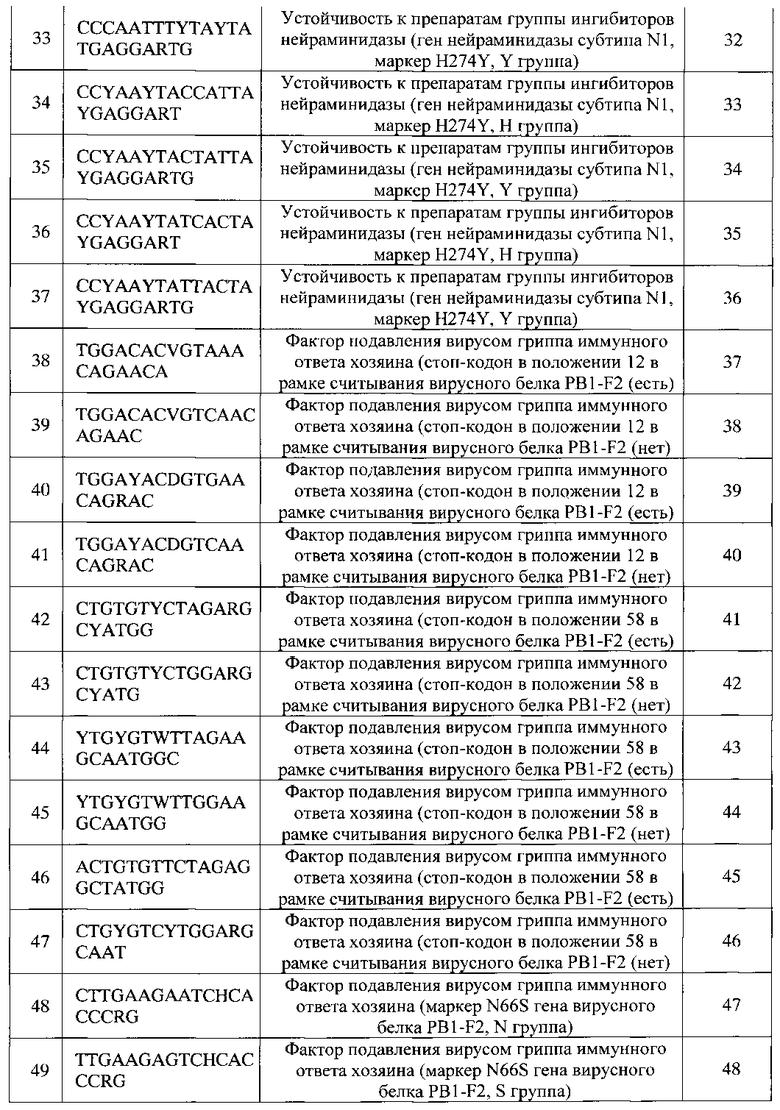


Краткое описание фигур
Фиг. 1 представляет схему размещения олигонуклеотидов на биочипе для идентификации РНК вирусов гриппа А и В с одновременным определением вариантов гемагглютинина и нейраминидазы вируса гриппа А, идентификацией генетических маркеров патогенности и устойчивости к противогриппозным препаратам.
Позиция 0 - элементы, не содержащие иммобилизованные соединения (фоновые ячейки);
позиция 1 - элементы, содержащие флуоресцентный маркер;
позиции 2-177 - элементы, содержащие иммобилизованные олигонуклеотидные зонды (последовательности зондов приведены в Таблице 1).
Фиг. 2 представляет структуру биочипа в соответствии со схемой на Фиг. 1 с указанием элементов для определения анализируемых параметров/мутаций.
А, В - ячейки для определения типа вируса гриппа: тип А или тип В;
N1-N9 - ячейки для определение субтипа нейраминидазы вируса гриппа А;
H1-Н16 - ячейки для определение субтипа гемагглютинина вируса гриппа А;
S31, А30, V27, L26, Н274, N66 - ячейки для определения генетических маркеров вируса гриппа А, связанных с патогенностью и устойчивостью к противовирусным препаратам;
Блок PB1-F2 - ячейки для определения стоп-кодонов в рамке считывания вирусного белка PB1-F2;
Блок NS1 - ячейки для определения стоп-кодонов и структуры вирусного белка NS1;
Блок HPAI - ячейки для анализа структуры сайта протеолитического расщепления гемагглютинина вируса гриппа A (HPAI).
Фиг. 3 представляет флуоресцентное изображение (гибридизационную картину) биочипа и отчет программы после проведения анализа биологического образца (мазок из полости носа и ротоглотки): обнаружен вирус гриппа, тип А, субтип H1N1; имеется мутация в белке М2 сегмента MP (7) S31N, приводящая к устойчивости к препаратам группы адамантанов; функциональный вирусный белок PB1-F2 не синтезируется, имеется стоп кодон по положению 12; обнаружен вариант белка NS1, характерный для низкопатогенных штаммов, С-концевое укорачивание на 11 аминокислот.
Фиг. 4 представляет флуоресцентное изображение (гибридизационную картину) биочипа и отчет программы после проведения анализа биологического образца (мазок из полости носа и ротоглотки): обнаружен вирус гриппа, тип В.
Фиг. 5 представляет флуоресцентное изображение (гибридизационную картину) биочипа и отчет программы после проведения анализа биологического образца (мазок из полости носа и ротоглотки) здорового донора: вирус гриппа не обнаружен.
Фиг. 6 представляет флуоресцентное изображение (гибридизационную картину) биочипа и отчет программы после проведения анализа образца культуры вируса, штамм вируса гриппа A/Coot/Chany/204/2005, субтип H2N2, детерминанты патогенности: умеренно патогенный функциональный вирусный белок PB1-F2, вариант белка NS 1, характерный для высокопатогенных штаммов (ESEV).
Фиг. 7 представляет флуоресцентное изображение (гибридизационную картину) биочипа и отчет программы после проведения анализа биологического образца (промывные воды бронхов): обнаружен вирус гриппа, тип А, субтип H3N2, имеется мутация S31N в белке М2 сегмента MP (7), приводящая к устойчивости к адамантанам, обнаружен умеренно патогенный функциональный вирусный белок PB1-F2.
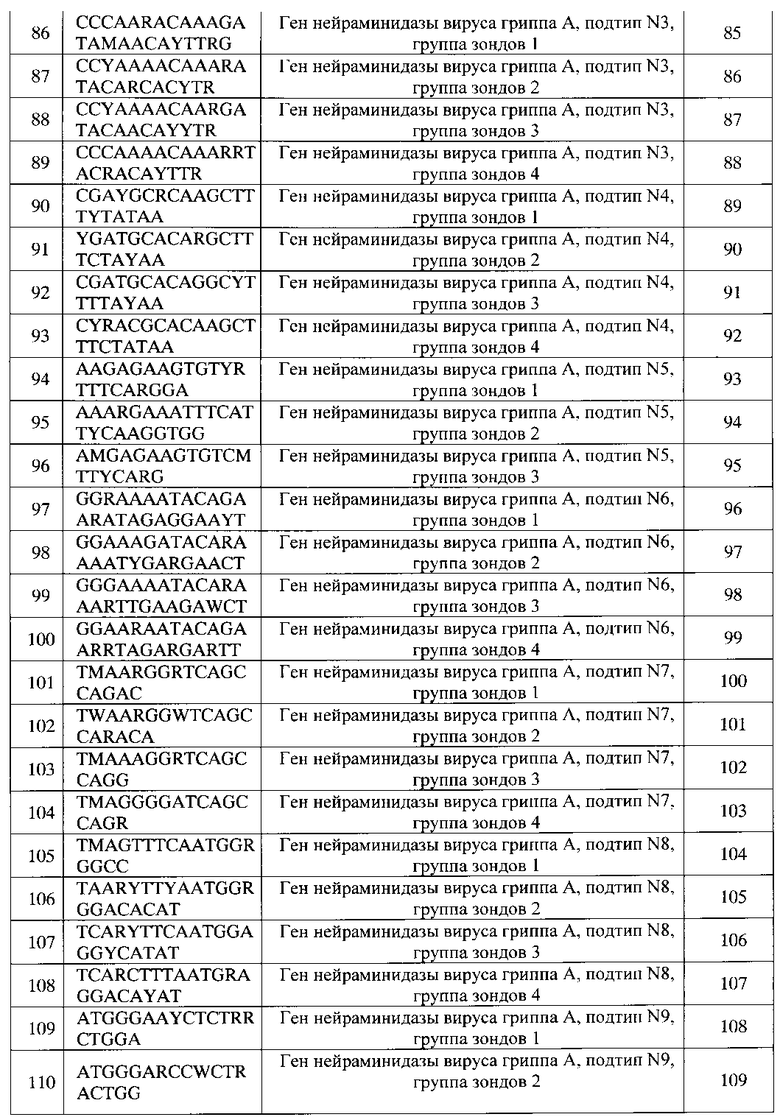
Фиг. 8 представляет флуоресцентное изображение (гибридизационную картину) биочипа и отчет программы после проведения анализа образца культуры вируса, штамм вируса гриппа A/Mallard/Altai/846/2011, субтип H4N6, детерминанты патогенности: высокопатогенный функциональный вирусный белок PB1-F2 (мутация N66S), вариант белка NS1, характерный для высокопатогенных штаммов (ESEV).
Фиг. 9 представляет флуоресцентное изображение (гибридизационную картину) биочипа и отчет программы после проведения анализа биологического образца (секционный материал, суспензия внутренних органов домашних птиц): обнаружен вирус гриппа, тип А, субтип H5N1 ("птичий грипп"), имеется мутация N66S в белке PB1-F2, приводящая к повышенной патогенности вируса, а также вариант белка NS1, характерный для высокопатогенных штаммов (ESEV).
Фиг. 10 представляет флуоресцентное изображение (гибридизационную картину) биочипа и отчет программы после проведения анализа образца культуры вируса, штамм вируса гриппа A/Teal/Amurskaya oblast/928/2009, субтип H6N6, обнаружен умеренно патогенный функциональный вирусный белок PB1-F2.
Фиг. 11 представляет флуоресцентное изображение (гибридизационную картину) биочипа и отчет программы после проведения анализа образца культуры вируса, штамм вируса гриппа A/Mallard/NT/00, субтип H7N3, имеется вариант белка NS1, характерный для высокопатогенных штаммов (ESEV).
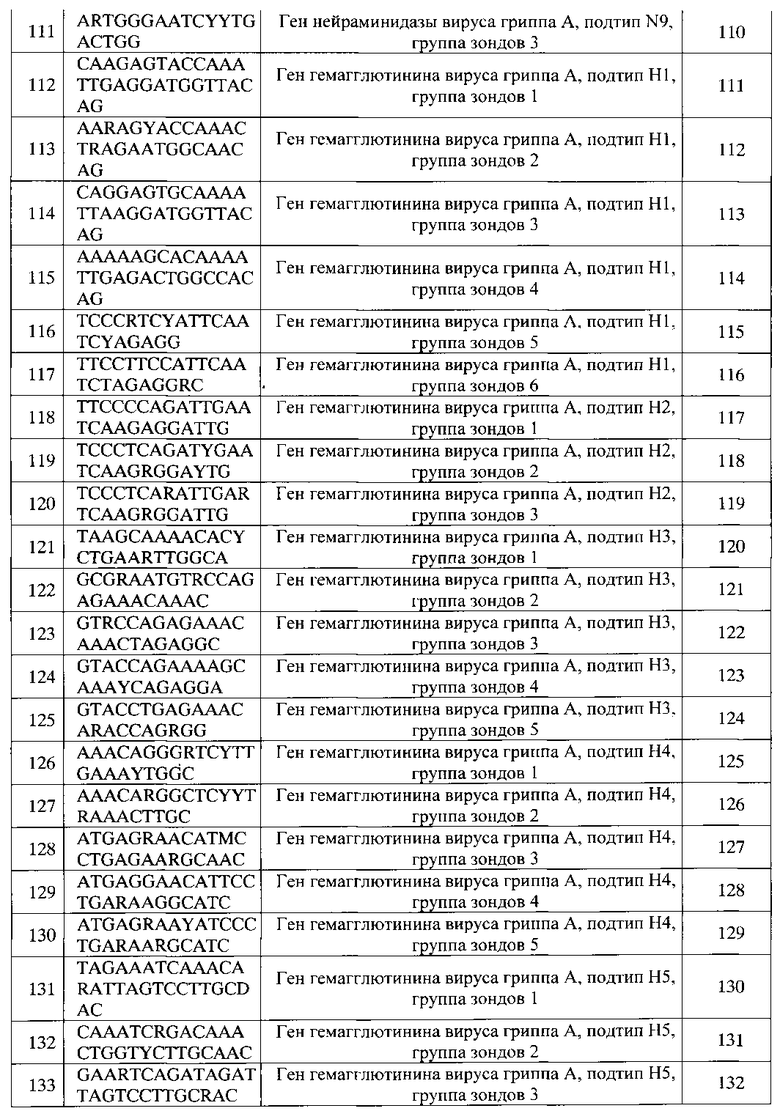
Фиг. 12 представляет флуоресцентное изображение (гибридизационную картину) биочипа и отчет программы после проведения анализа образца культуры вируса, штамм вируса гриппа A/Shoveler/Novosibirsk/246/2011, субтип H8N4, имеется высокопатогенный функциональный вирусный белок PB1-F2 (мутация N66S) и вариант белка NS1, характерный для высокопатогенных штаммов (ESEV).
Фиг. 13 представляет флуоресцентное изображение (гибридизационную картину) биочипа и отчет программы после проведения анализа препарата плазмидной ДНК, содержащей синтетический фрагмент последовательности гена НА 9 субтипа и гена NA 9 субтипа.
Фиг. 14 представляет флуоресцентное изображение (гибридизационную картину) биочипа и отчет программы после проведения анализа образца культуры вируса, штамм вируса гриппа A/Pochard/Siberia/249/2008, субтип H10N7, имеется умеренно патогенный функциональный вирусный белок PB1-F2, и вариант белка NS1, характерный для высокопатогенных штаммов (ESEV).
Фиг. 15 представляет флуоресцентное изображение (гибридизационную картину) биочипа и отчет программы после проведения анализа образца культуры вируса, штамм вируса гриппа A/Mew Gull/Kamchatka/131/2008, субтип H11N8, имеется умеренно патогенный функциональный вирусный белок PB1-F2, и вариант белка NS1, характерный для высокопатогенных штаммов (ESEV).
Фиг. 16 представляет флуоресцентное изображение (гибридизационную картину) биочипа и отчет программы после проведения анализа образца культуры вируса, штамм вируса гриппа A/Mallard/Alberta/234/2007, субтип H12N5, детерминанты патогенности: высокопатогенный функциональный вирусный белок PB1-F2 (мутация N66S), вариант белка NS1, характерный для высокопатогенных штаммов (ESEV).
Фиг. 17 представляет флуоресцентное изображение (гибридизационную картину) биочипа и отчет программы после проведения анализа образца культуры вируса, штамм вируса гриппа A/Common gull/Altai/811/2011, субтип H13N2, детерминанты патогенности: умеренно патогенный функциональный вирусный белок PB1-F2, вариант белка NS1, характерный для высокопатогенных штаммов (ESEV).
Фиг. 18 представляет флуоресцентное изображение (гибридизационную картину) биочипа и отчет программы после проведения анализа образца культуры вируса, штамм вируса гриппа A/Mallard/Gurjev/263/1982, субтип H14N5, детерминанты патогенности: умеренно патогенный функциональный вирусный белок PB1-F2, вариант белка NS1, характерный для высокопатогенных штаммов (ESEV).
Фиг. 19 представляет флуоресцентное изображение (гибридизационную картину) биочипа и отчет программы после проведения анализа образца культуры вируса, штамм вируса гриппа A/Teal/Chany/7119/2008, субтип H15N4, детерминанты патогенности: умеренно патогенный функциональный вирусный белок PB1-F2, вариант белка NS1, характерный для высокопатогенных штаммов (ESEV).
Фиг. 20 представляет флуоресцентное изображение (гибридизационную картину) биочипа и отчет программы после проведения анализа образца культуры вируса, штамм вируса гриппа A /Common gull/Altai/805/2011, субтип H16N3, имеется умеренно патогенный функциональный вирусный белок PB 1-F2.
Фиг. 21 представляет флуоресцентное изображение (гибридизационную картину) биочипа и отчет программы после проведения анализа биологического образца (промывные воды бронхов): обнаружен вирус гриппа, тип А, субтип H1N1, обнаружена мутация нераминидазы H274Y, приводящая к устойчивости к ингибиторам нейпамини дазы.
Фиг. 22 представляет флуоресцентное изображение (гибридизационную картину) биочипа и отчет программы после проведения анализа препарата плазмидной ДНК, содержащей синтетический фрагмент гена М2 с мутациями, приводящими к заменам в белке L26F, V27A, А30Т.
Фиг. 23 представляет флуоресцентное изображение (гибридизационную картину) биочипа и отчет программы после проведения анализа образца культуры вируса, штамм вируса гриппа A A/Common gull/Chany/P/2006, субтип H5N1, детерминанты патогенности: сайт протеолиза, характерный для высокопатогенных штаммов (HPAI-сайт), умеренно патогенный функциональный вирусный белок PB1-F2, вариант белка NS1, характерный для высокопатогенных штаммов (ESKV).
Фиг. 24 представляет флуоресцентное изображение (гибридизационную картину) биочипа и отчет программы после проведения анализа препарата плазмидной ДНК, содержащей синтетический фрагмент гена NS1, кодирующий последовательность EPEV (высокопатогенный вариант вируса гриппа).
Фиг. 25 представляет флуоресцентное изображение (гибридизационную картину) биочипа и отчет программы после проведения анализа препарата плазмидной ДНК, содержащей синтетический фрагмент гена NS1, кодирующий последовательность KSEV (высокопатогенный вариант вируса гриппа).
Фиг. 26 представляет флуоресцентное изображение (гибридизационную картину) биочипа и отчет программы после проведения анализа препарата плазмидной ДНК, содержащей синтетический фрагмент: последовательность, кодирующая белок NS1 без удлинения и укорачивания.
Фиг. 27 представляет флуоресцентное изображение (гибридизационную картину) биочипа и отчет программы после проведения анализа препарата плазмидной ДНК, содержащей синтетический фрагмент: последовательность белка NS1 с С-концевым укорачиванием на 13 аминокислот.
Осуществление изобретения
Целью изобретения является создание способа многопараметрического анализа для идентификации РНК вируса гриппа, определения типа вируса (тип А или тип В), определения вариантов гемагглютинина и нейраминидазы вируса гриппа А, идентификации генетических маркеров патогенности и устойчивости к противогриппозным препаратам.
Идентификация генетических маркеров патогенности и устойчивости к противогриппозным препаратам включает:
- идентификацию генетических маркеров, ассоциированных с лекарственной устойчивостью вируса гриппа к препаратам группы адамантанов (например, амантадин, ремантадин);
- идентификацию генетических маркеров, ассоциированных с лекарственной устойчивостью к препаратам группы ингибиторов нейраминидазы (например, ланинамивир, озельтамивир, перамивир, занамивир).
- идентификацию генетических маркеров, определяющих состояние сайта протеолитического расщепления гемагглютинина, ассоциированного с повышенной патогенностью вируса гриппа А (для субтипов Н5 и Н7);
- идентификацию генетических маркеров в рамке считывания вирусного белка PB1-F2, который является одним из ключевых факторов подавления вирусом гриппа иммунного ответа хозяина;
- идентификацию генетических маркеров в структуре PDZ-связывающего домена в сегменте NS1, детерминирующего активацию сигнальных систем клетки, обеспечивающих контроль над противовирусным ответом.
Одновременный многопараметрический анализ ДНК достигается при использовании биологических микрочипов - массивов элементов, содержащих олигонуклеотидные зонды. Заявленный способ заключается в обеспечении биочипа с соответствующими иммобилизованными специфичными олигонуклеотидными зондами, проведение реакции обратной транскрипции для получения кДНК с использованием вирусной РНК в качестве матрицы, проведении ПЦР с использованием олигонуклеотидных праймеров, комплементарных фрагментам генома вируса гриппа, с получением преимущественно одноцепочечных флуоресцентно-меченных фрагментов ДНК, гибридизацию флуоресцентно-меченных продуктов на биочипе, регистрацию и интерпретацию результатов гибридизации.
Биологический микрочип, используемый в предлагаемом способе, представляет собой массив трехмерных гидрогелевых элементов, расположенных на подложке из стекла или пластика и содержащих ковалентно иммобилизованные олигонуклеотидные зонды. Элементы массива - микрокапли заданного объема, как правило, диаметром от 80 до 300 мкм, расстояние между каплями составляет, как правило, 150-500 мкм. Микрочипы изготавливают методом сополимеризационной иммобилизации по технологии, разработанной в ИМБ РАН (публикация международной заявки WO/2003/033539 "Composition for polymerising immobilisation of biological molecules and method for producing said composition"; Rubina A.Y., Pan′kov S.V. et al., Hydrogel drop microchips with immobilized DNA: properties and methods for large-scale production, Anal. Biochem. 2004, v. 325, p. 92-106).
При изготовлении биочипа полимеризационную смесь, содержащую гелеобразующие мономеры и подлежащие иммобилизации олигонуклеотидные зонды, наносят на подложку в виде массива микрокапель с помощью автоматического микродозатора (робота). Полимеризация геля с одновременной ковалентной иммобилизацией зондов происходит под действием УФ-излучения. Изготовление биочипа на основе гидрогелей для идентификации и анализа РНК вируса гриппа описано в Примере 1.
Для определения наличия РНК вируса гриппа, а также типа вируса гриппа (грипп А или грипп В) используются олигонуклеотидные зонды, соответствующие фрагментам относительно консервативной последовательности сегмента MP (7) вируса гриппа (SEQ ID NO: 1-9).
Поверхностные гликопротеины - гемагглютинин (НА) и нейраминидаза (NA) являются существенными факторами, характеризующими вирус гриппа А. По предлагаемому способу определяются 16 субтипов гемагглютинина и 9 субтипов нейраминидазы. Используются группы олигонуклеотидных зондов, соответствующие участкам гена НА (подтипы Н1-Н16) (SEQ ID NO: 112-178) и гена NA (N1-N9) (SEQ ID NO: 73-111).
Лекарственные препараты группы адамантанов, например, амантадин и ремантадин, являются блокаторами трансмембранного белка М2 вируса гриппа А. В предлагаемом способе для анализа мутаций, приводящих к устойчивости к препаратам группы адамантанов, используются олигонуклеотидные зонды, соответствующие участкам гена, кодирующего белок М2 сегмента MP (7) вируса с последовательностями SEQ ID NO: 10-25), позволяющими выявить мутации S31N, А30Т, V27A, L26F. Данные мутации характерны для подавляющего большинства резистентных к адамантанам штаммов вируса гриппа А и являются достоверными маркерами устойчивости вируса к лечению этими препаратами.
Нейраминидаза является важной мишенью противовирусной терапии. Препараты, относящиеся к группе селективных ингибиторов нейраминидазы (озельтамивир, занамивир, перамивир и др.) являются средством лечения и профилактики адамантан-устойчивых штаммов гриппа. Причиной резистентности вируса гриппа А к ингибиторам нейраминидазы в большинстве случаев является мутация H274Y в гене NA, характерная для распространенного у человека подтипа H1N1. В предлагаемом способе используется группа олигонуклеотидных зондов с последовательностями SEQ ID NO: 26-37, позволяющие идентифицировать мутацию H274Y в гене нейраминидазы субтипа N1.
Наличие сайта протеолиза гемагглютинина является важным маркером патогенности вируса гриппа. Гемагглютинин - белок, обеспечивающий узнавание клеточных рецепторов и проникновение вируса в клетку. Для выполнения этой функции НА должен пройти созревание, заключающееся в специфическом расщеплении клеточными протеазами молекулы НА в сайте протеолиза. Последовательность сайта протеолиза различна для различных подтипов гемагглютинина. Для высокопатогенных штаммов (подтипы Н5 и Н7) характерно наличие полиосновного сайта протеолиза, в то время как у низкопатогенных штаммов в этом сайте присутствует лишь одна основная аминокислота. Полиосновный сайт легко расщепляется протеазами, экспрессирующимися практически во всех тканях организма хозяина, в то время как сайт протеолиза, характерный для низкопатогенных штаммов, может расщепляться только тканеспецифичными протеазами. Поэтому, наличие полиосновного сайта протеолиза может приводить к системной инфекции и множественному поражению тканей и органов. Группа зондов для определения состояния сайта протеолитического расщепления гемагглютинина, ассоциированного с повышенной патогенностью вируса гриппа А включает последовательности, представленные SEQ ID NO: 66-72.
Вирусный белок PB1-F2, участвующий в индукции апопоптоза в макрофагах, является одним из ключевых факторов подавления вирусом гриппа иммунитета хозяина. Домен, ответственный за индукцию, находится на С-конце аминокислотной последовательности белка PB1-F2. Наличие мутации N66S в гене вирусного белка РВ1-F2 является маркером повышенной патогенности вируса гриппа. Кроме того, наличие стоп ко донов в положениях 12 и 58, приводит к синтезу в инфицированных клетках "дефектных" белков PB1-F2, не способных индуцировать апоптоз. Присутствие либо отсутствие стоп-кодонов по этим положения позволяет сделать вывод о степени патогенности конкретного штамма вируса гриппа А. Группа зондов для анализа генетических маркеров в рамке считывания вирусного белка PB1-F2 включает последовательности для идентификации мутации N66S и стоп-кодонов в 12 и 58 положении, представленные SEQ ID NO: 38-51.
Белок NS1 вируса гриппа А играет первостепенную роль в регуляции первичного иммунного ответа клетки на вирусную инфекцию. Помимо противостояния интерфероновому ответу, NS1 может влиять на вирулентность за счет связывания и нарушения работы клеточных сигнальных белков. Длина С-концевого фрагмента белка вариабельна, и наиболее частыми вариантами являются укорачивание на 11, 13 аминокислот и удлинение на 7 аминокислот. Для выяснения вклада NS1 в степень патогенности вируса гриппа, необходимо: (1) определять наличие укорачивания белка NS1 на 11 и/или 13 аминокислот; (2) определять наличие либо отсутствие удлинения на 7 аминокислот; (3) дифференцировать С-концевые последовательности являющиеся лигандами PDZ-домена и характерные для высокопатогенных штаммов (ESEV, EPEV, ESKV, KSEV) от последовательностей таковыми не являющимися (RSKV, RSEV). Группа зондов для определения генетических маркеров в структуре PDZ-связывающего домена включает последовательности для анализа длины белка NSI (LVT-0, LVT-11, LVT-13), последовательности аминокислот RSKV (низкопатогенный вариант вируса), RSEV (низкопатогенный вариант вируса), KSEV (высокопатогенный вариант вируса), ESEV (высокопатогенный вариант), ESKV (высокопатогенный вариант вируса), EPEV (высокопатогенный вариант вируса), представленные SEQ ID NO: 52-65.
На Фиг. 1 и 2 представлена схема биочипа, гидрогелевые элементы которого содержат иммобилизованные олигонуклеотидные зонды с последовательностями SEQ ID NO: 1-178 (позиции 2-177). Кроме того, биочип содержит элементы, не содержащие иммобилизованные соединения для измерения фоновых сигналов (позиция 0), а также элементы, содержащие флуоресцентный маркер (позиция 1), для правильного позиционирования биочипа при получении флуоресцентного изображения и захвата изображения программным обеспечением биочип-анализатора. Из-за высокой вариабельности вирусов гриппа для идентификации каждого генетического маркера использовано различное число элементов биочипа (в зависимости от степени вариабельности нуклеотидной последовательности конкретного генетического маркера). В каждом элементе иммобилизована группа зондов относительно близкой последовательности с целью определения максимального числа отличающихся друг от друга по нуклеотидной последовательности штаммов вируса. При гибридизации флуоресцентно-меченного продукта с иммобилизованными на чипе олигонуклетидными зондами образуется стабильный комплекс с наиболее комплементарным зондом, что приводит к появлению высокого флуоресцентного сигнала в соответствующем элементе биочипа. Анализ флуоресцентных сигналов позволяет сделать вывод о наличии/отсутствии генетического маркера.
Для проведения анализа по данному способу используется РНК, выделенная из биологического материала, полученного от человека (мазки из полости носа и ротоглотки, бронхоальвеолярный лаваж, промывные воды бронхов, секционный материал) и птиц (клоакальные мазки, фрагменты внутренних органов) или культуры вирусов. Выделение РНК осуществляют любым из известных в данной области способов или с использованием коммерчески доступных наборов реагентов коммерческих наборов и автоматических роботизированных станций, например, набор реагентов «Проба НК» ООО «ДНК-Технология», Россия, QIAamp DSP Virus Kit, Qiagen, Германия, MagMAX™AI/ND Viral RNA Isolation Kits, Ambion, США.
На следующем этапе проведения анализа по данному способу полученную вирусную РНК используют в качестве матрицы для получения кДНК по реакции обратной транкрипции и далее ПЦР с использованием олигонуклеотидных праймеров, комплементарных соответствующим фрагментам генома вируса гриппа, и смеси четырех дезоксинуклеозидтрифосфатов, один из которых содержит флуоресцентную метку, с получением продуктов амплификации в виде преимущественно одноцепочечных флуоресцентно-меченных фрагментов ДНК. ПЦР может осуществляться в одну или несколько стадий. Реакция обратной транскрипции может проводиться одновременно с ПЦР.
Смесь дезоксинуклеозидтрифосфатов содержит дАТФ, дГТФ, дЦТФ и дТТФ; вместо дТТФ можно использовать дУТФ или смесь дТТФ с дУТФ в любом соотношении. Флуоресцентную метку может нести любой из указанных дезоксинуклеозидтрифосфатов.
В качестве флуоресцентного красителя может быть использован любой флуоресцентный краситель, который может быть включен в молекулу дезоксинуклеозидтрифосфата таким образом, чтобы не препятствовать проведению ПЦР и последующей гибридизации. Например, флуоресцентный краситель может быть присоединен к 5′-концу аминоаллильного производного дУТФ. Примеры красителей включают красители флуоресцеинового (TAMRAR, ROXR, JOER), родаминового (Texas RedR), полиметинового (Cy3R, Cy5R, Cy5.5R, Cy7R) рядов, но не ограничиваются ими.
После проведения ПЦР полученные продукты гибридизуют на биочипе, содержащем иммобилизованные олигонуклеотидные зонды, комплементарные исследуемым последовательностям. Гибридизацию проводят в растворе, содержащем буферный компонент для поддержания рН, соль для создания ионной силы и хаотропный (дестабилизирующий водородные связи) агент, в герметичной гибридизационной камере.
В качестве хаотропного могут быть использованы, например, гуанидин тиоцианат, мочевина, формамид и т.п. Иммобилизованные на биочипе дискриминирующие олигонуклеотиды, заявленные в настоящем изобретении, имеют температуры плавления в интервале от 42 до 44°С, что позволяет проводить гибридизацию при 37°С с использованием хаотропного агента.
Фрагменты ДНК образуют совершенные гибридизационные дуплексы только с соответствующими полностью комплементарными олигонуклеотидами. Дискриминацию совершенных и несовершенных дуплексов выполняют путем сравнения интенсивностей флуоресценции ячеек микрочипа, в которых образовались дуплексы. Интенсивность сигнала в ячейках, в которой образовались совершенные гибридизационные дуплексы выше, чем в ячейках, где образовались несовершенные дуплексы. Регистрацию и интерпретацию результатов гибридизации на биочипе проводят путем сравнения интенсивности флуоресценции сигналов в пределах каждой из групп ячеек, в которых образовались совершенные и несовершенные гибридизационные дуплексы.
Регистрация результатов гибридизации на микрочипах может быть осуществлена с помощью любых приборов-анализаторов флуоресценции, оснащенного специализированным программным обеспечением, позволяющим использовать программную автоматическую обработку интенсивностей сигналов с последующей интерпретацией результатов, например, с помощью коммерческих сканирующих устройств GenePix 4000 В (Axon Instruments, США), ′GenePix Pro′, ′Acuity′ (Axon Instruments, США), портативноного биочип-анализатора: "Комплекс универсальный аппаратно-программный для анализа биологических микрочипов" (ООО «БИОЧИП-ИМБ», Россия).
Проведение анализа: идентификации РНК вирусов гриппа А и В с одновременным определением вариантов гемагглютинина и нейраминидазы вируса гриппа А, идентификацией генетических маркеров патогенности и устойчивости к противогриппозным препаратам, по данному способу описано в Примере 2. Результаты анализа представлены на Фиг. 3-27.
Данным изобретением также предлагается биочип для идентификации РНК вирусов гриппа А и В с одновременным определением вариантов гемагглютинина и нейраминидазы вируса гриппа А, идентификацией генетических маркеров патогенности и устойчивости к противогриппозным препаратам, представляющий собой подложку с гидрогелевыми элементами, в каждом из которых иммобилизован набор специфических олигонуклеотидных зондов. Пример биочипа представлен на Фиг. 1 и 2, примеры результатов анализа с его использованием представлены на Фиг. 3-27.
Нуклеотидные последовательности (SEQ ID NO: 1-178) зондов для иммобилизации в гидрогелевых элементах микрочипа, являющегося предметом изобретения, представлены в Таблице 1.
Далее изобретение проиллюстрировано примерами, которые предназначены для обеспечения лучшего понимания сущности изобретения, и которые не должны рассматриваться как ограничивающие данное изобретение.
Пример 1
Изготовление биочипа на основе гидрогелей для идентификации и анализа РНК вируса гриппа
Олигонуклеотидные зонды с последовательностями SEQ ID NO: 1-178 синтезировали на автоматическом синтезаторе 394 DNA/RNA synthesizer (Applied Biosystems, США). Олигонуклетиды для иммобилизации в гидрогелевых ячейках биочипа содержали спейсер со свободной аминогруппой 3′-Amino-Modifier С7 CPG 500 (Glen Research, США). Изготовление биочипов производили в соответствии с процедурой, описанной ранее (Rubina A.Y., PanTsov S.V. et al. Hydrogel drop microchips with immobilized DNA: properties and methods for large-scale production. Anal. Biochem., 2004, v. 325, p. 92-106).
Полимеризационные смеси, содержащие смесь гелеобразующих мономеров (производные метакриламида, метилен-бис-акриламида, стабилизаторы и др.) и иммобилизуемые соединения (олигонуклеотиды или флуоресцентный краситель IMD 504), помещали ячейки 384-луночного планшета. С использованием робота QArray (Genetix, Великобритания) смеси из лунок планшета переносили в виде микрокапель на поверхность пластиковых подложек. Полимеризация геля с одновременной ковалентной иммобилизацией олигонуклеотидов в структуре геля происходила под действием УФ-излучения (УФ-лампа Sylvania GTE F15T8350 BL, Великобритания), 40 мин, 20°С, в потоке азота. После полимеризации подложки с массивами гидрогелевых ячеек (микрокапель) промывали 0,1 M фосфатным буфером, водой и высушивали в беспылевой атмосфере.
Полученные биочипы представляли собой совокупность полусферических ячеек диаметром 100 мкм, расстояние между ячейками - 300 мкм. Структура биочипа представлена на Фиг. 1, 2.
Микрочип содержит 192 гидрогелевых элемента. Элементы 2-177 (Фиг. 1) содержат иммобилизованные олигонуклеотидные зонды с последовательностями SEQ ID NO: 1-178, указанными в Таблице 1;
ячейки 2-8 содержат олигонуклеотидные зонды (SEQ ID NO: 1-9), соответствующие фрагментам последовательности сегмента MP (7) вируса гриппа для определения типа вируса гриппа: тип А или тип В;
ячейки 9 - 24 - зонды (SEQ ID NO: 10-25), соответствующие участкам гена, кодирующего белок М2, для определения генетических маркеров вируса гриппа А, связанных с устойчивостью к противовирусным препаратам группы адамантанов (мутации S31N, А30Т, V27A, L26F);
ячейки 25 - 36 - зонды (SEQ ID NO: 26-37), соответствующие участкам гена нейраминидазы подтипа вируса гриппа H1N1, для идентификации мутации H274Y, приводящей к устойчивости к ингибиторам нейраминидазы;
ячейки 37 - 46 - зонды (SEQ ID NO: 38-47) для анализа генетических маркеров в рамке считывания вирусного белка PB1-F2: стоп-кодоны в 12 и 58 положении;
ячейки 47 - 50 - зонды (SEQ ID NO: 48-51) для анализа генетических маркеров в рамке считывания вирусного белка PB1-F2: идентификация мутации N66S, приводящей к повышенной патогенности вируса;
ячейки 51 - 56 - зонды (SEQ ID NO: 52-57) для анализа С-концевых последовательностей белка NSI: ESEV, EPEV, ESKV, KSEV - последовательности, характерные для высокопатогенных штаммов, RSKV, RSEV последовательности, характерные для низкопатогенных штаммов вируса гриппа А;
ячейки 57 - 64 - зонды (SEQ ID NO: 58-65) для анализа длины С-концевого фрагмента белка NS1: укорачивание на 11 и/или 13 аминокислот, удлинение на 7 аминокислот;
ячейки 65 - 71 - зонды (SEQ ID NO: 66-72) для определения наличия сайта протеолитического расщепления гемагглютинина (HPAI сайт) 5-го и 7-го подтипов вируса гриппа А;
ячейки 72 - 110 - зонды (SEQ ID NO: 73-111), соответствующие участкам гена нейраминидазы, для определения субтипа нейраминидазы (N1-N9);
ячейки 11 - 177 - зонды (SEQ ID NO: 112-178), соответствующие участкам гена гемагглютинина, для определения субтипа гемагглютинина (Н1-Н16).
В состав чипа также входят маркерные точки, содержащие иммобилизованный флуоресцентный краситель IMD 504 (ООО «БИОЧИП», Москва) (элементы 1 на Фиг. 1) для правильного позиционирования биочипа, выполняемого компьютерной программой при анализе флуоресцентного изображения после проведения гибридизации, и ячейки, не содержащие иммобилизованные соединения (элементы 0 на Фиг. 1), необходимые для вычисления фонового значения интенсивности флуоресценции.
Пример 2. Процедура проведения анализа: идентификация РНК вирусов гриппа А и В с одновременным определением вариантов гемагглютинина и нейраминидазы вируса гриппа А, идентификацией генетических маркеров патогенности и устойчивости к противогриппозным препаратам с использованием биологического микрочипа
Проведение анализа включало стадии: подготовки биологических образцов (биологический материал, полученный от человека и птиц или культуры вирусов); выделения РНК; проведения реакции обратной транкрипции (ОТ) для получения кДНК при использовании РНК вируса гриппа в качестве матрицы; ПЦР-амплификации с получением одноцепочечных флуоресцентно-меченных фрагментов ДНК; гибридизации амплифицированных флуоресцентно-меченных продуктов на биочипах; регистрации и интерпретации результатов гибридизации.
а) Подготовка биологических образцов
Мазки из полости носа и ротоглотки. Мазки брали сухими стерильными зондами с вискозными тампонами (ФБУН ЦНИИ Эпидемиологии, кат. №6100/SG/CS) вращательными движениями с поверхности миндалин, небных дужек и задней стенки ротоглотки. После взятия материала рабочую часть зонда с вискозным тампоном помещали в стерильную пробирку с 0,5 мл транспортной среды (ФБУН ЦНИИ Эпидемиологии, кат. №957). Конец зонда отламывали, придерживая крышкой пробирки с расчетом, чтобы он позволил плотно закрыть пробирку. Содержимое закрытой пробирки перемешивали на вортексе и центрифугировали в течение 5 с при 5000 об/мин на микроцентрифуге для удаления капель с внутренней поверхности крышки пробирки. Для выделения РНК отбирали 100 мкл образца.
Бронхоальвеолярный лаваж или промывные воды бронхов. Содержимое закрытой пробирки перемешивали на вортексе и центрифугировали в течение 10 мин при 10000 об/мин. Отбирали надосадочную жидкость, используя наконечник с фильтром, оставляя над осадком 200 мкл жидкости, в которой ресуспендировали осадок. Для выделения РНК отбирали 100 мкл полученной суспензии.
Секционный материал и фрагменты внутренних органов птиц гомогенизировали с использованием стерильных фарфоровых ступок и пестиков, затем готовили 10% суспензию на стерильном физиологическом растворе либо фосфатном буфере. Суспензию переносили в пробирку 1,5 мл и центрифугировали при 10000 об/мин в течение 5 мин. Для выделения РНК отбирали 100 мкл надосадочной жидкости.
Культура вирусов Растворяли 10 мкл культуральной жидкости в 90 мкл деионизированной воды. Для выделения РНК отбирали 100 мкл раствора, б) Выделение РНК Выделение РНК из биологических образцов проводили с использованием набора «Проба НК» ООО «ДНК-Технология», Россия.
в) Подготовка праймеров для проведения обратной транскрипции и ПЦР
При выборе последовательностей праймеров для проведения обратной транскрипции и ПЦР использовали базы данных нуклеотидных последовательностей, известные специалистам в данной области, например, база данных http://www.ncbi.nlm.nih.gov/Genbank/index.html. Специфичность праймеров проверяли, например, с помощью программного обеспечения, использующего поиск в базах нуклеотидных последовательностей по алгоритму BLAST (например, www.ncbi.nlm.nih.gov/BLAST). В частности, избегали таких последовательностей, которые способны гибридизоваться с последовательностями генома человека.
Перечень используемых в данном примере праймеров приведен в Таблице 2. Праймеры синтезировали на автоматическом синтезаторе 394 DNA/RNA synthesizer (Applied Biosystems, США). В качестве флуоресцентной метки использовали краситель Imd 500- dUTP №49, d-UTP-конъюгат (ООО «БИОЧИП-ИМБ»), максимум поглощения при 645 нм, максимум флуоресценции при 670 нм. Введение метки проводили в соответствии с рекомендациями производителя.


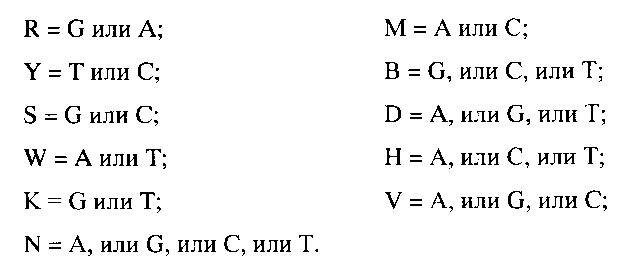
г) Обратная транскрипция (ОТ) для получения кДНК при использовании РНК вируса гриппа в качестве матрицы
К 4 мкл раствора выделенной РНК добавляли 1 мкл (20 пмоль) праймера для ОТ и инкубировали в термостате при +72°С в течении 5 минут. Капли осаждали кратковременным центрифугированием и пробирку переносили в лед.
В пробирку добавляли следующие компоненты:
10-кратный ОТ буфер (50 мМ трис-HCl, рН 8.3, 3 мМ MgCl2, 75 мМ KCl; 10 мМ дитиотреитол - 1 мкл;
смесь дезоксинуклеозидтрифосфатов (дНТФ), 2,5 мМ - 4 мкл;
обратная транскриптаза M-MLV (20 ед./мкл) - 0,5 мкл;
вода, свободная от РНКаз, - 1,5 мкл.
Инкубировали смесь в течении 60 мин при +42°С. Реакцию останавливали прогреванием в течении 10 мин при +95°С. Полученную кДНК использовали для проведения ПЦР.
д) 1-я стадия ПЦР для амплификации фрагментов кДНК
Готовили реакционную смесь для проведения 1-го этапа ПЦР, внося в пробирку следующие реагенты (из расчета на 1 реакцию):
- вода деионизованная 21,5 мкл;
- ПЦР-буфер (10 × кратная смесь, содержащая
100 mM трис-HCl, 500 мМ KCl; 15 мМ MgCl2) 3 мкл;
- дНТФ (водный раствор дезоксинуклеозидтрифосфатов,
по 2 ммоль/л дАТФ, дГТФ, дУТФ, дЦТФ) 3 мкл;
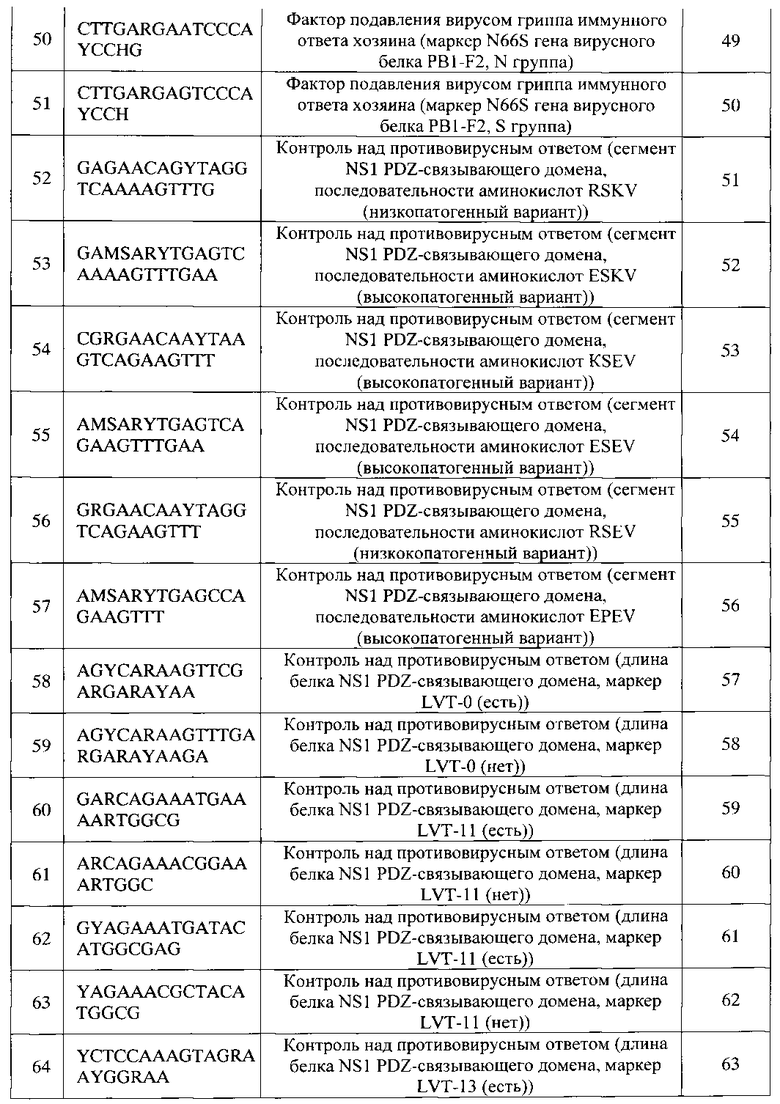
- смесь праймеров для 1-го этапа ПЦР с концентрацией
1 пмоль/мкл каждый 1,0 мкл;
- Taq-полимераза, активность 5 Ед./мкл 0,5 мкл.
К 29 мкл смеси добавляли 1 мкл кДНК, полученной после этапа ОТ. Для отрицательного контрольного образца вместо матрицы вносили деионизованную стерильную воду. Пробирки помещали в ДНК-амплификатор С1000, «Biorad», США и проводили ПЦР по следующей программе: предварительная денатурация ДНК: 94°С, 1 цикл 4 мин; 35 циклов ПЦР: 94°С - 30 с, 63°С - 30 с, 72°С - 1 мин; финальная элонгация при 72°С - 5 мин.
Полученные растворы использовали для проведения 2-го этапа ПЦР.
е) Проведение 2-го этапа ПЦР для получения флуоресцентно-меченных одноцепочечных фрагментов ДНК
Готовили реакционную смесь для проведения 2-го этапа ПЦР, внося в пробирку следующие реагенты (из расчета на 1 реакцию):
- вода деионизованная 21,3 мкл;
- ПЦР-буфер (10 × кратная смесь, содержащая
100 mM трис-HCl, 500 мМ KCl; 15 мМ MgCl2) 3 мкл;
- дНТФ (водный раствор дезоксинуклеозидтрифосфатов,
по 2 ммоль/л дАТФ, дГТФ, дУТФ, дЦТФ) 3 мкл;
- смесь праймеров для 2-го этапа ПЦР с концентрацией
1 пмоль/мкл каждый 1,0 мкл;
- Taq-полимераза, активность 5 Ед./мкл 0,5 мкл.
- флуоресцентно-меченный дУТФ, 1 мкМ 0,2 мкл
В качестве флуоресцентно-меченного субстрата использовали конъюгат дУТФ с флуоресцентным красителем UMD-515 (максимум поглощения при 645 нм, максимум флуоресценции при 670 нм) производства ООО «БИОЧИП-ИМБ».
К 29 мкл смеси добавляли по 1 мкл образца, полученного после 1-го этапа ПЦР. Для отрицательных контрольных образцов вместо матрицы вносили деионизованную стерильную воду или 1 мкл отрицательного контрольного образца, полученного после 1 -ой стадии ПЦР. Пробирки помещали в ДНК-амплификатор С1000, «Biorad», США и проводили ПЦР по следующей программе: предварительная денатурация ДНК: 95°С, 1 цикл 2 мин; 36 циклов ПЦР: 95°С - 20 с, 60°С - 20 с, 72°С - 30 с; финальная элонгация при 72°С - 5 мин.
Полученные растворы использовали для проведения гибридизации на биочипе, полученном, как описано в Примере 1 (Фиг. 1, 2).
ж) Гибридизация флуоресцентно-меченных продуктов на биочипах
К 27 мкл реакционной смеси, полученной после второй стадии ПЦР, добавляли концентрированный раствор гибридизационного буфера так, чтобы конечная концентрация компонентов составляла:
гуанидин тиоцианат - 1М,
HEPES - 50 мМ (рН 7,5),
ЭДТА - 5 мМ.
30 мкл полученной смеси помещали в гибридизационную камеру микрочипа через отверстие в камере. Биочипы помещали в термостат на +37°С и инкубировали в течение 5 ч. По окончании гибридизации удаляли гибридизационные камеры, и биочипы трижды промывали дистиллированной водой, прогретой до 37°С, в течение 30 с и высушивали.
з) Регистрация и интерпретация результатов гибридизации
Результаты гибридизации регистрировали с помощью универсального аппаратно-программного комплекса (УАПК) для анализа изображений диагностических микрочипов с использованием специализированного программного обеспечения "ImageWare" (ООО «БИОЧИП-ИМБ»).
Флуоресцентные изображения биочипов после проведения гибридизации приведены на Фиг. 3-27.
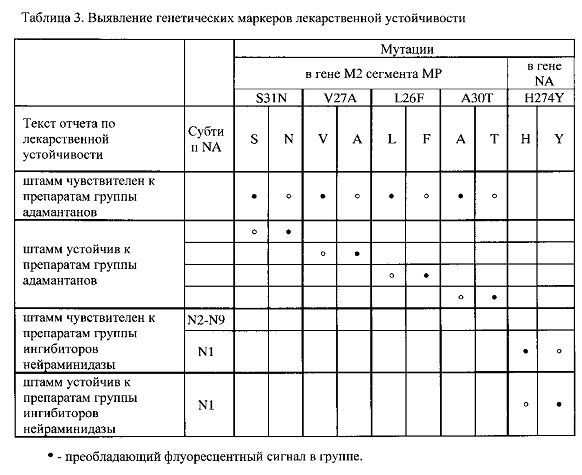
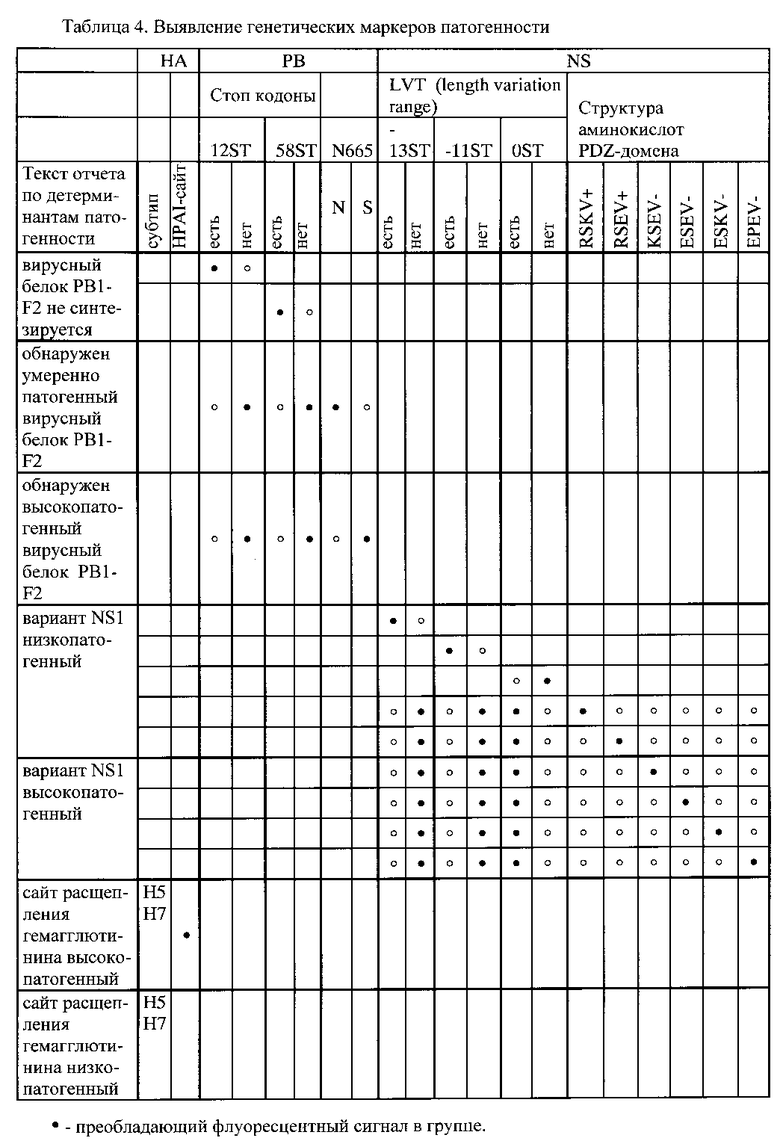
Интерпретация результатов гибридизации на биочипе осуществляется по алгоритму, реализованному в модуле программного обеспечения, путем сравнения интенсивности флуоресценции сигналов в пределах каждой из групп ячеек, в которых образовались совершенные и несовершенные гибридизационные дуплексы. Преобладающие флуоресцентные сигналы в группах для анализа генетических маркеров лекарственной устойчивости указаны в Таблице 3, для анализа генетических маркеров патогенности - в Таблице 4.
После анализа флуоресцентного изображения в отчете программы содержится следующая информация:
«Вирус гриппа не обнаружен» - исследуемый образец отрицательный;
«Вирус гриппа обнаружен» - исследуемый образец положительный;
«Тип вируса гриппа»: А, В;
«Подтип»: гемагглютинин H1-Н16, не определен (Нх); нейраминидаза N1-N9, не определен (Nx);
«Лекарственная устойчивость» (Таблица 3): штамм чувствителен к препаратам группы адамантанов; штамм устойчив к препаратам группы адамантанов (указаны мутации, обуславливающие устойчивость); штамм чувствителен к препаратам группы ингибиторов нейраминидазы; штамм устойчив к препаратам группы ингибиторов нейраминидазы (указана мутация H274Y, обуславливающая устойчивость);
«Детерминанты патогенности» (Таблица 4): Вирусный белок PB1-F2 не синтезируется; обнаружен умеренно патогенный вирусный белок PB1-F2; обнаружен высокопатогенный вирусный белок PB1-F2 (указана мутация N66S, характеризующаяся высокой патогенностью); Состояние вирусного белка PB1-F2 не определено;
Обнаружен вариант NS1, характерный для низкопатогенных штаммов; обнаружен вариант NS1, характерный для высокопатогенных штаммов (указана последовательность аминокислот, характеризующаяся высокой патогенностью);
Вариант NS1 не определен;
Сайт расщепления гемагглютинина высокопатогенный (HPAI). Сайт расщепления гемагглютинина низкопатогенный (LPAI) (для субтипов Н5 и Н7).
Пример 3. Идентификация РНК вирусов гриппа А и В в биологических образцах с В с одновременным определением вариантов гемагглютинина и нейраминидазы вируса гриппа А, идентификацией генетических маркеров патогенности и устойчивости к противогриппозным препаратам
Для проведения анализа использовали биологические микрочипы, полученные, как описано в Примере 1. Процедура анализа описана в Примере 2. Флуоресцентные изображения (гибридизационные картины) и информация, приведенная в отчетах программы, представлены на Фигурах 3-27.
Результаты анализа мазков из полости носа и ротоглотки и промывных вод бронхов представлены на Фигурах 3-5, 7, 21. Образцы биологического материала собирали в период заболеваемости сезонным гриппом на территории России в 2011-1013 гг., и, соответственно, обнаруженные вирусы гриппа А относились к вирусам сезонного гриппа, субтипы H1N1 и H3N2.
Результат анализа секционного материала: суспензии внутренних органов домашних птиц, погибших в период эпизоотии в Новосибирской области в июле 2005 года ("птичий грипп") представлены на Фиг. 9.
Фигуры 6, 10-12, 14-20, 23 представляет результаты анализа культуры вирусов. Референтные штаммы вируса гриппа А были получены из коллекции вирусов ГУ НИИ вирусологии им. Д.И. Ивановского РАМН. Вирусы гриппа предварительно культивировали в 10-дневных куриных эмбрионах. Для выделения РНК использовали коммерческие набора производства НПО «НАРВАК», Россия.
Имеющиеся в наличии образцы биологического материала не содержали всех генетических маркеров патогенности и устойчивости к противогриппозных препаратам, которые могут быть идентифицированы с использованием данного способа, поэтому для анализа некоторых маркеров были получены синтетические фрагменты ДНК, соответствующие участкам генома вируса гриппа, содержащим необходимые мутации, например, синтетический фрагмент последовательности гена НА 9 субтипа и гена NA 9 субтипа ВГА, синтетический фрагмент ДНК, содержащий фрагмент гена NS1, кодирующий последовательность EPEV (высокопатогенный вариант вируса гриппа), и др. Необходимые последовательности генома вируса гриппа были выбраны с помощью базы данных GenBank и синтезированы на синтезаторе ABI 3900 (Applied Biosystems, США). Далее получали модифицированные штаммы Е.coli со специфическими плазмидами. Выделение ДНК проводили по стандартной методике с использованием комплекта «Рибо-сорб» (ООО «Интерлабсервис»). Определение концентрации ДНК проводили с использованием специфических праймеров-зондов в «real-time» ПЦР на приборе для детекции результатов ПЦР в режиме реального времени.
Результаты анализа синтетических фрагментов ДНК показаны на Фигурах 13, 22, 24-27.
Возможность определения наличия РНК вируса гриппа и определения его типа (тип А или тип В) по предлагаемому способу продемонстрирована на Фиг. 3-12, 14-21, 23.
Возможность определения молекулярных вариантов гемагглютинина (Н1-Н16) и нейраминидазы (N1-N9) ВГА по предлагаемому способу продемонстрирована на Фиг. 3, 4, 6-21.
Возможность идентификации генетических маркеров S31N, V27A, А30Т, L26F в гене М2 вирусного сегмента MP ВГА, ассоциированных с устойчивостью вируса к лекарственным препаратам группы адамантанов, по предлагаемому способу продемонстрирована на Фиг. 3, 7 и 22.
Возможность идентификации генетического маркера H274Y в гене нейраминидазы ВГА субтипа N1, ассоциированного с устойчивостью вируса к лекарственным препаратам группы ингибиторов нейраминидазы, по предлагаемому способу продемонстрирована на Фиг. 21.
Возможность определения состояния сайта протеолитического расщепления гемагглютинина ВГА, ассоциированного с повышенной патогенностью вируса, по предлагаемому способу продемонстрирована на Фиг. 23.
Возможность анализа генетических маркеров в рамке считывания вирусного белка PB1-F2, который является одним из ключевых факторов подавления вирусом гриппа иммунного ответа хозяина: последовательности для идентификации мутации N66S и стоп-кодонов в 12 и 58 положении, по предлагаемому способу продемонстрирована на Фиг. 6-10, 12, 14-20, 23.
Возможность анализа генетических маркеров в структуре PDZ-связывающего домена в гене NS1, детерминирующего активацию сигнальных систем клетки, обеспечивающих контроль над противовирусным ответом при гриппозной инфекции: последовательности для анализа длины белка NSI (LVT-0, LVT-11, LVT-13), последовательности аминокислот RSKV и RSEV (низкопатогенные варианты вируса), KSEV, ESEV, ESKV и EPEV (высокопатогенные варианты вируса), по предлагаемому способу продемонстрирована на Фиг. 3, 4-12, 14-21, 23-27.
Правильность определения наличия РНК вируса гриппа, определения его типа, определения молекулярных вариантов гемагглютинина и нейраминидазы, а также генетических маркеров патогенности и устойчивости к противогриппозным препаратам по предлагаемому способу во всех случаях проверяли независимым методами:
- метод ПЦР с гибридизационно-флуоресцентной детекцией с использованием наборов фирмы АмплиСенс «АмплиСенс® Influenza virus A/B-FL», «АмплиСенс® Influenza virus А-тип-FL»;
- секвенирование нуклеотидных последовательностей генома вируса гриппа, включающее сегменты НА, NA, РВ1, NS, MP генома ВГА и вируса гриппа В. Секвенирование проводилось на автоматическом секвенаторе 3130xL Applied Biosystems (США) с помощью коммерческого набора для проведения секвенирующих реакций «BigDye Terminator v3.1 Ready Reaction Cycle Sequencing Kit», с использованием секвенирующих праймеров М13-прямой (TGTAAAACGACGGCCAGT) и М13-обратный (CAGGAAACAGCTATGACC).
Перечень олигонуклеотидных зондов, иммобилизованных в элементах биочипа, используемого в способе идентификации РНК вирусов гриппа А и В с одновременным определением вариантов гемагглютинина и нейраминидазы вируса гриппа А, идентификацией генетических маркеров патогенности и устойчивости к противогриппозным препаратам
N = А, или G, или C, или Т.
| название | год | авторы | номер документа |
|---|---|---|---|
| БИОЧИП И СПОСОБ ТИПИРОВАНИЯ ГЕНОВ ГЕМАГГЛЮТИНИНА И НЕЙРАМИНИДАЗЫ ВИРУСА ГРИППА А | 2013 |
|
RU2560591C2 |
| НАБОРЫ ОЛИГОНУКЛЕОТИДОВ-ПРАЙМЕРОВ И ЗОНДОВ, БИОЛОГИЧЕСКИЙ МИКРОЧИП И ТЕСТ-СИСТЕМА ДЛЯ ИДЕНТИФИКАЦИИ И ТИПИРОВАНИЯ ВИРУСА ГРИППА А И В С ИХ ИСПОЛЬЗОВАНИЕМ | 2013 |
|
RU2538168C2 |
| СПОСОБ ВЫБОРА ДНК-ЗОНДОВ ДЛЯ МИКРОЧИПОВОЙ ДИАГНОСТИКИ, БИОЧИП И СПОСОБ ТИПИРОВАНИЯ ГЕНА НЕЙРАМИНИДАЗЫ И ГЕМАГГЛЮТИНИНА ВИРУСА ГРИППА А | 2010 |
|
RU2470076C2 |
| СПОСОБ ОПРЕДЕЛЕНИЯ ВИРУЛЕНТНЫХ И ПАТОГЕННЫХ ФОРМ ВИРУСОВ ГРИППА | 2011 |
|
RU2489488C1 |
| Штамм вируса гриппа A/Common Muskrat/Chany Lake/226/05 H2N2-субтипа для использования в диагностике вируса гриппа методами РТГА и ПЦР и исследования эффективности противовирусных препаратов in vitro и in vivo | 2015 |
|
RU2606030C1 |
| ШТАММ ВИРУСА ГРИППА A/BLACK-HEADED GULL/KAMCHATKA/123/2013 H11N8-СУБТИПА ДЛЯ ИСПОЛЬЗОВАНИЯ В ДИАГНОСТИКЕ ВИРУСА ГРИППА МЕТОДАМИ РТГА И ПЦР И ИССЛЕДОВАНИЯ ЭФФЕКТИВНОСТИ ПРОТИВОВИРУСНЫХ ПРЕПАРАТОВ IN VITRO и IN VIVO | 2016 |
|
RU2631932C1 |
| Способ диагностики респираторной вирусной инфекции у детей | 2020 |
|
RU2741508C1 |
| СПОСОБ ФЛУОРЕСЦЕНТНОГО МЕЧЕНИЯ кДНК ВИРУСА ГРИППА ТИПА А | 2011 |
|
RU2480525C2 |
| УСЛОВНО ДЕФЕКТНАЯ ЧАСТИЦА ВИРУСА ГРИППА И СПОСОБЫ ЕЕ ПОЛУЧЕНИЯ (ВАРИАНТЫ) | 2005 |
|
RU2420577C2 |
| ШТАММ ВИРУСА ГРИППА А/common gull/Altai/804/2011/H16N3-СУБТИПА ДЛЯ ПОЛУЧЕНИЯ АНТИГЕНСОДЕРЖАЩЕГО ПРЕПАРАТА, ПОЛИКЛОНАЛЬНОЙ СЫВОРОТКИ И ПРИМЕНЕНИЯ В КАЧЕСТВЕ КОНТРОЛЬНОГО РЕФЕРЕНС-ОБРАЗЦА ПРИ ОЦЕНКЕ СПЕЦИФИЧНОСТИ ТЕСТ-СИСТЕМ НА ОСНОВЕ ПОЛИМЕРАЗНОЙ ЦЕПНОЙ РЕАКЦИИ | 2012 |
|
RU2496872C1 |















Изобретение относится к области молекулярной биологии, вирусологии, ветеринарии и медицине и касается способа идентификации РНК вирусов гриппа А и В с одновременным определением вариантов гемагглютинина и нейраминидазы вируса гриппа А, а также генетических маркеров патогенности и устойчивости к противогриппозным препаратам, на биологических микрочипах. Способ основан на проведении реакции обратной транскрипции для получения кДНК с использованием вирусной РНК в качестве матрицы, полимеразной цепной реакции (ПЦР) и последующей гибридизации полученного одноцепочечного флуоресцентно-меченного ПЦР-продукта на биологическом микрочипе. Биологический микрочип представляет собой подложку с упорядоченно расположенными гидрогелевыми элементами, содержащими ковалентно иммобилизованные олигонуклеотидные зонды, которые обеспечивают гибридизацию со специфическими участками генома вируса гриппа, определяющими его тип, субтип и генетические маркеры, включая детерминанты устойчивости к противовирусным препаратам. 3 н. и 2 з.п. ф-лы, 27 ил., 4 табл., 3 пр.
1. Способ идентификации РНК-вирусов гриппа А и В с одновременным определением вариантов гемагглютинина и нейраминидазы вируса гриппа А, идентификацией генетических маркеров патогенности и устойчивости к противогриппозным препаратам, включающий
а) обеспечение биочипа, представляющего собой подложку с гелевыми элементами, в каждом из которых иммобилизован набор олигонуклеотидных зондов, относящийся к следующим группам:
- группа зондов, последовательности которых представлены SEQ ID NO: 1-9, соответствующая фрагментам вирусных сегментов MP (7), для определения наличия РНК вируса гриппа и определения типа вируса (тип А или тип В);
- группа зондов, последовательности которых представлены SEQ ID NO: 112-178, соответствующая фрагментам вируса гриппа А, для определения молекулярного варианта гемагглютинина (Н1-Н16);
- группа зондов, последовательности которых представлены SEQ ID NO: 73-111, соответствующая фрагментам вируса гриппа А, для определения молекулярного варианта нейраминидазы (N1-N9);
- группа зондов, последовательности которых представлены SEQ ID NO: 10-25, для идентификации генетических маркеров S31N, V27A, А30Т, L26F в гене М2 вирусного сегмента MP вируса гриппа А, ассоциированных с устойчивостью вируса к лекарственным препаратам группы адамантанов;
- группа зондов, последовательности которых представлены SEQ ID NO: 26-37, для идентификации генетического маркера H274Y в гене нейраминидазы вируса гриппа А субтипа N1, ассоциированного с устойчивостью вируса к лекарственным препаратам группы ингибиторов нейраминидазы;
- группа зондов, последовательности которых представлены SEQ ID NO: 66-72, для определения состояния сайта протеолитического расщепления гемагглютинина вируса гриппа А, ассоциированного с повышенной патогенностью вируса (для субтипов Н5 и Н7);
- группа зондов, последовательности которых представлены SEQ ID NO: 38-51, для анализа генетических маркеров в рамке считывания вирусного белка PB1-F2, который является одним из ключевых факторов подавления вирусом гриппа иммунного ответа хозяина, включающих последовательности для идентификации мутации N66S и стоп-кодонов в 12 и 58 положении;
- группа зондов, последовательности которых представлены SEQ ID NO: 52-65, для анализа генетических маркеров в структуре PDZ-связывающего домена в гене NS1, детерминирующего активацию сигнальных систем клетки, обеспечивающих контроль над противовирусным ответом при гриппозной инфекции, включающих последовательности для анализа длины белка NS1 (LVT-0, LVT-11, LVT-13), последовательности аминокислот RSKV (низкопатогенный вариант вируса), RSEV (низкопатогенный вариант вируса), KSEV (высокопатогенный вариант вируса), ESEV (высокопатогенный вариант), ESKV (высокопатогенный вариант вируса), EPEV (высокопатогенный вариант вируса);
б) проведение реакции обратной транскрипции для получения кДНК с использованием вирусной РНК в качестве матрицы и далее ПЦР с использованием олигонуклеотидных праймеров, комплементарных фрагментам генома вируса гриппа, в буферном растворе, включающем смесь четырех дезоксинуклеозидтрифосфатов, один из которых содержит флуоресцентную метку, с получением продуктов амплификации в виде преимущественно одноцепочечных флуоресцентно-меченных фрагментов ДНК;
в) гибридизацию амплифицированных флуоресцентно-меченных продуктов, полученных на стадии (б), на биочипе, полученном на стадии (а);
г) регистрацию и интерпретацию результатов гибридизации на биочипе, проведенной на стадии (в), путем сравнения интенсивности флуоресценции сигналов в пределах каждой из групп ячеек, в которых образовались совершенные и несовершенные гибридизационные дуплексы.
2. Способ по п. 1, характеризующийся тем, что для анализа используется РНК, выделенная из биологического материала, полученного от человека (мазки из полости носа и ротоглотки, бронхоальвеолярный лаваж, промывные воды бронхов, секционный материал) и от птиц (клоакальные мазки, фрагменты внутренних органов) или культуры вирусов.
3. Способ по п. 1, характеризующийся тем, что регистрацию результатов на стадии (г) проводят с помощью прибора-анализатора флуоресценции, оснащенного специализированным программным обеспечением, что позволяет использовать программную автоматическую обработку интенсивностей сигналов с последующей интерпретацией результатов.
4. Биочип, используемый в способе идентификации РНК вирусов гриппа А и В с одновременным определением вариантов гемагглютинина и нейраминидазы вируса гриппа А, идентификацией генетических маркеров патогенности и устойчивости к противогриппозным препаратам, представляющий собой подложку с гелевыми элементами, в каждом из которых иммобилизован набор зондов, причем последовательности зондов представлены SEQ ID NO: 1-178.
5. Набор олигонуклеотидных зондов, используемый для получения биочипа, используемого в способе идентификации РНК вирусов гриппа А и В с одновременным определением вариантов гемагглютинина и нейраминидазы вируса гриппа А, идентификацией генетических маркеров патогенности и устойчивости к противогриппозным препаратам, причем зонды имеют последовательности SEQ ID NO: 1-178.
| НАБОРЫ ОЛИГОНУКЛЕОТИДОВ-ПРАЙМЕРОВ И ЗОНДОВ, БИОЛОГИЧЕСКИЙ МИКРОЧИП И ТЕСТ-СИСТЕМА ДЛЯ ИДЕНТИФИКАЦИИ И ТИПИРОВАНИЯ ВИРУСА ГРИППА А И В С ИХ ИСПОЛЬЗОВАНИЕМ | 2013 |
|
RU2538168C2 |
| СПОСОБ ВЫБОРА ДНК-ЗОНДОВ ДЛЯ МИКРОЧИПОВОЙ ДИАГНОСТИКИ, БИОЧИП И СПОСОБ ТИПИРОВАНИЯ ГЕНА НЕЙРАМИНИДАЗЫ И ГЕМАГГЛЮТИНИНА ВИРУСА ГРИППА А | 2010 |
|
RU2470076C2 |
| СПОСОБ ОБНАРУЖЕНИЯ ВИРУСА ГРИППА А ПОДТИПА H5N1 | 2007 |
|
RU2361924C1 |
| US 2007092871 A1, 26.04.2007 | |||
| RYABININ A.V | |||
| et al., ";Microarray assay for detection and discrimination of Orthopoxvirus species";, J Med Virol | |||
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |

