Родственные заявки
Данная заявка заявляет приоритет 35 USC §119 (е) предварительной заявки США серийный номер US 61/317,261, озаглавленной «Самодоставляющиеся PHKi соединения уменьшенного размера», поданной 24 марта 2010 года, и предварительной заявки США серийный номер US 61/317,597, озаглавленной «Самодоставляющиеся PHKi соединения уменьшенного размера», поданной 25 марта 2010 года, все описания которых включены в данную заявку полностью путем ссылки.
Область техники, к которой относится изобретение
Настоящее изобретение относится к области РНК интерференции (PHKi). Настоящее изобретение более конкретно относится к способам введения молекул нуклеиновой кислоты с улучшенными свойствами доставки in vivo и использования их при эффективном генном сайленсинге.
Уровень техники
Комплементарные олигонуклеотидные последовательности являются перспективными терапевтическими агентами и полезными инструментами исследования для выяснения функций генов. Однако олигонуклеотидные молекулы уровня техники страдают от ряда проблем, которые могут препятствовать их клиническому развитию, и часто затрудняют достижение намеченного эффективного ингибирования экспрессии генов (включая синтеза белка), используя такие композиции in vivo.
Основная проблема заключалась в доставке этих соединений в клетки и ткани. Обычные двухцепочечные PHKi соединения, 19-29 оснований в длину, представляют собой весьма отрицательно заряженные жесткие спирали приблизительно 1,5 на 10-15 нм размером. Молекула данного стержневого типа не может пройти через клеточную мембрану и, как результат, имеет очень ограниченную эффективность как in vitro так и in vivo. В результате, все обычные PHKi соединения требуют своего рода средства доставки, чтобы содействовать их распределению в тканях и клеточному поглощению. Это считается основным ограничением PHKi технологии.
Были предварительные попытки применить химические модификации олигонуклеотидов для улучшения их клеточных свойств поглощения. Одной из таких модификаций было присоединение молекулы холестерина к олигонуклеотиду. Первый доклад о таком подходе был по Letsinger et al., в 1989. Затем, ISIS Pharmaceuticals, Inc. (Carlsbad, CA) сообщил про более продвинутые методы присоединения молекулы холестерина к олигонуклеотиду (Manoharan, 1992).
С открытием siPHK в конце девяностых, подобные типы модификаций были разработаны для этих молекул для повышения их профилей доставки. Молекулы холестерина, сопряженные со слегка модифицированными (Soutschek, 2004) и сильно модифицированными (Wolfram, 2007) siPHK появились в литературе. Yamada et al., 2008 также сообщили об использовании передовых химий линкеров, чтобы еще больше улучшить уровень холестерин-опосредованного поглощения siPHK. Несмотря на все эти усилия, поглощение этих типов соединений по-видимому, ингибируется в присутствии биологических жидкостей, что приводит к весьма ограниченной эффективности генного сайленсинга in vivo, ограничивая применение этих соединений в клинических условиях.
Сущность изобретения
Аспекты настоящего изобретения относятся к модифицированным полинуклеотидам, которые имеют повышенную стабильность и обладают эффективной системной доставки. Полинуклеотиды, связанные с изобретением, включают sd-PHK, в которых по меньшей мере 40% нуклеотидов модифицированы и в которых по меньшей мере два U и/или С включают гидрофобную модификацию, выбранную из группы, состоящей из метильной, октильной, тиофеновой, октин-1-ильной, этинильной, пиридиламидной, изобутильной и фенильной модификации. В некоторых осуществлениях, по меньшей мере 60% нуклеотидов модифицированы.
Модификации могут включать по меньшей мере одну 2'F или 2'O метильную модификацию. В некоторых осуществлениях, множество U и/или С включают гидрофобную модификацию, необязательно выбранную из группы, состоящей из метил октильной, тиофеновой, октин-1-ильной, этинильной, пиридиламидной, изобутильной и фенильной модификации.
sd-PHK может быть присоединена к линкеру. В некоторых осуществлениях, линкер является протонируемым. В определенных осуществлениях, sd-PHK присоединена к множеству линкеров.
sd-PHK может быть связана с липофильной группой, такой, как 3' конец sd-PHK. В некоторых осуществлениях, sd-PHK связана с холестерином. В определенных осуществлениях, sd-PHK присоединена к витамину А или витамину Е.
Ведущая нить sd-PHK может содержать одноцепочечную область, которая содержит по меньшей мере 5 фосфоротиоатных модификаций. В некоторых осуществлениях, sd-PHK содержит по меньшей мере две одноцепочечные области.
В некоторых осуществлениях, sd-PHK разработана для внутривенного, подкожного или интратекального введения. sd-PHK может иметь период полураспада в сыворотке, более чем 12 часов.
Аспекты настоящего изобретения относятся к композициям, содержащим любые из sd-PHK, описанных в данной заявке. В некоторых осуществлениях, композиция содержит две или более различных sd-PHK.
Аспекты настоящего изобретения относятся к двухцепочечным РНК (dsPHK), в которых по меньшей мере 40% нуклеотидов модифицированы и в которых по меньшей мере два U и/или С включают гидрофобную модификацию, выбранную из группы, состоящей из метильной, октильной, тиофеновой, октин-1-ильной, этинильной, пиридиламидной, изобутильной и фенильной модификации. В некоторых осуществлениях, по меньшей мере 60% нуклеотидов модифицированы.
Модификации могут включать по меньшей мере одну 2'F или 2'O метильную модификацию. В некоторых осуществлениях, множество U и/или С включают гидрофобную модификацию, необязательно выбранную из группы, состоящей из метил октильной, тиофеновой, октин-1-ильной, этинильной, пиридил амидной, изобутильной и фенильной модификации.
Аспекты настоящего изобретения относятся к одноцепочечным RISC входящим полинуклеотидам, в которых по меньшей мере 40% нуклеотидов модифицированы и в которых по меньшей мере два U и/или С включают гидрофобную модификацию, выбранную из группы, состоящей из метильной, октильной, тиофеновой, октан-1-ильной, этинильной, пиридил амидной, изобутильной и фенильной модификации. В некоторых осуществлениях, по меньшей мере 60% нуклеотидов модифицированы.
Модификации могут включать по меньшей мере одну 2'F или 2'O метильную модификацию. В некоторых осуществлениях, множество U и/или С включают гидрофобную модификацию, необязательно выбранную из группы, состоящей из метил октильной, тиофеновой, октин-1-ильной, этинильной, пиридил амидной, изобутильной и фенильной модификации. В некоторых осуществлениях, одноцепочечный RISC входящий полинуклеотид в соответствии с любым из пп. 23-26 формулы, где выделенный одноцепочечный RISC входящий полинуклеотид представляет собой miPHK ингибитор.
Аспекты настоящего изобретения относятся к способам доставки нуклеиновой кислоты в отдаленную ткань-мишень у субъекта, который нуждается в этом, включая системное введение субъекту sd-PHK в эффективном количестве для способствования РНК интерференции при помощи sd-PHK в отдаленную ткань-мишень. В некоторых осуществлениях, по меньшей мере 40%, или по меньшей мере 60% нуклеотидов в антисмысловой нити sd-PHK модифицированы.
Модификации могут включать по меньшей мере одну 2'F или 2'O метильную модификацию. В некоторых осуществлениях, по меньшей мере один U или С включает гидрофобную модификацию. В определенных осуществлениях, множество U и/или С включает гидрофобную модификацию. Гидрофобная модификация может быть выбрана из группы, состоящей из октильной, тиофеновой, октин-1-ильной, этинильной, пиридиламидной, изобутильной и фенильной модификации.
sd-PHK может быть присоединена к линкеру. В некоторых осуществлениях, линкер является протонируемым. В определенных осуществлениях, sd-PHK присоединена к множеству линкеров.
sd-PHK может быть связана с липофильной группой, такой, как 3' конец sd-PHK. В некоторых осуществлениях, sd-PHK связана с холестерином, витамином А или витамином Е.
Ведущая нить sd-PHK может содержать одноцепочечную область, которая содержит по меньшей мере 5 фосфоротиоатных модификаций. В некоторых осуществлениях, ведущая нить sd-PHK содержит по меньшей мере две одноцепочечные области.
В некоторых осуществлениях, sd-PHK доставляют в печень, сердце, мозг, легкие или жир. В определенных осуществлениях, sd-PHK доставляют в опухоль. В некоторых осуществлениях, sd-PHK вводят подкожно.
Аспекты настоящего изобретения относятся к способам, включая введение субъекту, имеющему опухоль, sd-PHK в количестве, эффективном для способствования РНК интерференции при помощи sd-PHK в опухоль. В некоторых осуществлениях, sd-PHK вводят в опухоль через внутривенное введение. В других осуществлениях sd-PHK вводят в опухоль через прямую инъекцию в опухоль. В определенных осуществлениях, sd-PHK направлена против гена, кодирующего белок, выбранного из группы, состоящей из VEGF/VEGFR, HER2, PDGF/PDGFR, HDAC, MET, c-kit, CDK, FLT-1, IGF/IGFR, FGF/FGFR, Ras/Raf, Abl, Bcl-2, Src, mTOR, PKC, МАРК, BIRC5, FAS, HIF1A, CDH16, MYC, HRAS и CTNNB1, или их комбинации.
Аспекты настоящего изобретения относятся к способам, включающим введение sd-PHK в центральную нервную систему субъекта, нуждающегося в этом. В некоторых осуществлениях, sd-PHK вводят путем интратекальной доставки.
Описаны в данной заявке способы и композиции для эффективной доставки sd-PHK молекул в отдаленные ткани-мишени. В некоторых осуществлениях, нуклеиновую кислоту доставляют через внутривенное введение. В определенных осуществлениях, внутривенное введение является непрерывной инфузией. В некоторых осуществлениях, нуклеиновую кислоту вводят в течение 2-3 часов. Нуклеиновая кислота может быть введена повторно. В определенных осуществлениях, нуклеиновую кислоту вводят подкожно. В некоторых осуществлениях, нуклеиновую кислоту вводят ежедневно, еженедельно или ежемесячно. В определенных осуществлениях, вторую дозу нуклеиновой кислоты вводят через 1-3 месяца после первой дозы. В некоторых осуществлениях, нуклеиновую кислоту вводят каждые 2-3 месяца.
В некоторых осуществлениях, sd-PHK связана с длинноцепочечным алкилхолестериновым аналогом, витамином А или витамином Е. В некоторых осуществлениях, sd-PHK прикреплена к хлорформиату.
В некоторых осуществлениях, способы, связанные с настоящим изобретением, включают способы лечения легочных состояний, таких как астма, хроническая обструктивная болезнь легких (COPD), рак легких, метастазы легких, фиброз легких, инфекции и инфекционные заболевания, состояниях печени, таких как цирроз и гепатит (например, хронический гепатит, острый гепатит, волчаночный гепатит, аутоиммунный гепатит или вирусный гепатит) или сердечных состояний, таких как ишемическая болезнь сердца, кардиомиопатия, гипертоническая болезнь сердца, пороки сердца, врожденные пороки сердца, миокардит и аритмия.
Аспекты настоящего изобретения относятся к способам доставки нуклеиновой кислоты в отдаленную ткань-мишень, включая подкожное введение субъекту sd-PHK, в количестве, эффективном для способствования РНК интерференции при помощи sd-PHK в отдаленную ткань-мишень. В некоторых осуществлениях, тканью-мишенью являются легкие и sd-PHK молекула доставляется в альвеолярный макрофаг.
В некоторых осуществлениях, тканью-мишенью является сердце, печень или жир. Способы, связанные с настоящим изобретением, могут включать способы лечения или профилактики легочного состояния, состояния сердца или профилактики состояния печени.
Аспекты настоящего изобретения относятся к способам лечения легочного состояния, включая: введение субъекту, имеющему легочное состояние, sd-PHK в количестве, эффективном для способствования РНК интерференции при помощи sd-PHK в альвеолярный макрофаг для лечения легочного состояния. В некоторых осуществлениях, sd-PHK вводят путем инсуффляции или подкожно. В определенных осуществлениях, sd-PHK вводят в композиции сухого порошка и/или с носителем. В некоторых осуществлениях, sd-PHK находится в комплексе с частицей, содержащей глюкан. В определенных осуществлениях, sd-PHK направлена против гена, кодирующего специфический белок альвеолярного макрофага. В некоторых осуществлениях, sd-PHK вводят при помощи интратрахеального спрея.
Дополнительные аспекты настоящего изобретения относятся к композициям, содержащим одну или несколько двухцепочечных молекул нуклеиновой кислоты, выбранной из молекул нуклеиновой кислоты, содержащихся в таблицах 1-5, таким образом что антисмысловая и смысловая нити составляют двухцепочечную молекулу нуклеиновой кислоты.
В некоторых осуществлениях, sd-PHK, связанная с изобретением, направлена против АроВ, МАР4К4, PCSK9, PPIB, KSP, CTGF или VEGF.
Каждое из ограничений в соответствии с настоящим изобретением может включать различные осуществления в соответствии с настоящим изобретением. Таким образом, предполагается, что каждое из ограничений в соответствии с настоящим изобретением с участием одного элемента или комбинаций элементов могут быть включены в каждый аспект в соответствии с настоящим изобретением. Настоящее изобретение не ограничено в применении деталями конструкции и расположением компонентов, изложенных в приведенном ниже описании или показано на чертежах. Изобретение может иметь другие осуществления и практикуется или осуществляется различными способами.
Краткое описание чертежей
Чертежи, которые прилагаются, не предназначены для черчения в масштабе. На чертежах, каждый идентичный или почти идентичный компонент проиллюстрирован на различных фигурах, представленных аналогичными номерами. Для ясности, не каждый компонент может быть помечен на каждом чертеже. На чертежах:
ФИГ. 1 демонстрирует скорость клиренса крови и поглощение печенью sd-PHK после внутривенного введения.
ФИГ. 2 демонстрирует сайленсинг МАР4К4 в печени после ежедневного внутривенного введения sd-PHK.
ФИГ. 3 представляет схематически повторяющийся режим дозирования для введения sd-PHK.
ФИГ. 4 представляет изображения, показывающие поглощение печенью sd-PHK у мышей. Мышам давали дозы 50 мг/кг 1, 2, 3 или 4 раза в течение 24 часов.
ФИГ. 5 представляет изображения, показывающие поглощение sd-PHK в печени, селезенке, сердце, почках и легких после внутривенного введения.
ФИГ. 6 демонстрирует, что повторяющееся дозирование sd-PHK приводит к повышенному поглощению печенью.
ФИГ. 7 представляет сравнение подкожного и внутривенного введения для эффективности в печени. Однократный болюс 80 мг/кг sd-PHK был доставлен для обеих проб.
ФИГ. 8 демонстрирует, что маршрут введения sd-PHK влияет на кинетику клиренса, уровни крови и поглощение тканями. Однократный болюс 80 мг/кг sd-PHK был доставлен.
ФИГ. 9 раскрывает распределение sd-PHK в печени через 24 часа после подкожного или внутривенного введения.
ФИГ. 10 раскрывает распределение sd-PHK в печени через 24 часа после подкожного или внутривенного введения.
ФИГ. 11 раскрывает доставку sd-PHK в кожу после подкожной инъекции.
ФИГ. 12 раскрывает присутствие sd-PHK в клетках после подкожной инъекции.
ФИГ. 13 раскрывает поглощение sd-PHK в легких мышей после введения путем инсуффляции дозировок в диапазоне от 1 мг/кг до 44 мг/кг.
ФИГ. 14 раскрывает поглощение sd-PHK в легких крыс после введения путем инсуффляции дозировок в диапазоне от 1 мг/кг до 10 мг/кг.
ФИГ. 15 раскрывает поглощение sd-PHK в различных клеточных типах после инсуффляции.
ФИГ. 16 раскрывает эффективное поглощение sd-PHK в альвеолярных макрофагах после инсуффляции.
ФИГ. 17 раскрывает клинические наблюдения крыс после введения sd-PHK в легкие. Не наблюдали летаргии или обезвоживания и наблюдали, что легкие нормальные и здоровые.
ФИГ. 18 отображает примеры химических модификаций пассажирской и ведущей нитей последовательностей для МАР4К4 и PCSK9. Символ # относится к модификациям, выбранным из группы, показанной на ФИГ. 19.
ФИГ. 19 отображает примеры 5-уридильных модификаций.
ФИГ. 20 отображает примеры длинноцепочечных алкилхолестериновых аналогов.
ФИГ. 21 отображает пример протонируемого линкера.
ФИГ. 22 отображает примеры 5 метил 2' модифицированных нуклеозидов.
ФИГ. 23 отображает пример хлорформиатного конъюгирования.
ФИГ. 24 отображает конъюгаты витаминов А и Е для ткани-мишени.
ФИГ. 25 отображает пролекарство, имеющее эффективную доставку в печень.
ФИГ. 26 отображает влияние содержания фосфоротиоата и гидрофобного механизма на эффективную доставку sd-PHK in vitro и in vivo.
ФИГ. 27 является схематическим отображением добавления и удлинения множества фосфоротиоатных хвостовых областей на одной или двух нитях sd-PHK молекулы.
ФИГ. 28 отображает добавление множества холестериновых фрагментов в sd-PHK молекулу.
ФИГ. 29 отображает примеры встречающихся в природе рибонуклеотидов.
ФИГ. 30 отображает дополнительные примеры встречающихся в природе рибонуклеотидов.
ФИГ. 31 демонстрирует экспрессию гена АроВ в клетках после введения sd-PHK, мишенью которой является АроВ.
ФИГ. 32 демонстрирует экспрессию гена PCSK9 в клетках после введения sd-PHK, мишенью которой является PCSK9.
ФИГ. 33 демонстрирует экспрессию гена PPIB в клетках после введения sd-PHK, мишенью которой является PPIB.
ФИГ. 34 демонстрирует превращение скрытых PHKi последовательностей для PPIB в sd-PHK последовательности для того же самого гена.
ФИГ. 35 демонстрирует, что sd-PHK, содержащие рибо-линкер, являются эффективными.
ФИГ. 36 демонстрирует, что вариация химии линкеров не влияет на активность сайленсинга sd-PHK in vitro. Оценены две различные химии линкеров, гидроксипролинового линкера и рибо-линкера, на множестве sd-PHK (мишенью которой является Map4k4 или PPIB) в анализах пассивного поглощения для определения линкеров, способствующих самодоставке. HeLa клетки трансфецировали при отсутствии основы доставки (пассивная трансфекция) с sd-PHK при 1 мкМ, 0,1 мкМ или 0,01 мкМ в течение 48 часов. Применение любого линкера приводит к эффективной доставке sd-PHK.
ФИГ. 37 раскрывает сайленсинг в печени после системной доставки sd-PHK.
ФИГ. 38 обеспечивает неограничивающие примеры схем модификации последовательностей.
ФИГ. 39 обеспечивает суммарное влияние sd-PHK химии и структуры на распределение в тканях.
ФИГ. 40 раскрывает, что определенные модели химических модификаций приводят к значительному распределению в сердце и жире.
ФИГ. 41 демонстрирует эффекты октильной модификации sd-PHK, мишенью которой является МАР4К4.
ФИГ. 42 демонстрирует эффекты октильной модификации sd-PHK, мишенью которой является PCSK9.
ФИГ. 43 демонстрирует эффекты октильной модификации sd-PHK, мишенью которой является PCSK9.
ФИГ. 44 демонстрирует эффекты тиофеновой модификации sd-PHK мишенью которой является МАР4К4.
ФИГ. 45 демонстрирует эффекты октин-1-ильной модификации sd-PHK, мишенью которой является МАР4К4.
ФИГ. 46 демонстрирует эффекты октин-1-ильной модификации sd-PHK, мишенью которой является PCSK9.
ФИГ. 47 демонстрирует эффекты этенильной модификации sd-PHK, мишенью которой является МАР4К4.
ФИГ. 48 демонстрирует эффекты этенильной модификации sd-PHK, мишенью которой является PCSK9.
ФИГ. 49 демонстрирует эффекты пиридиламидной модификации sd-PHK, мишенью которой является PCSK9.
ФИГ. 50 демонстрирует эффекты пиридиламидной модификации sd-PHK, мишенью которой является МАР4К4.
ФИГ. 51 демонстрирует эффекты пиридиламидной модификации sd-PHK, мишенью которой является PCSK9.
ФИГ. 52 демонстрирует эффекты пиридиламидной модификации sd-PHK, мишенью которой является PCSK9.
ФИГ. 53 демонстрирует эффекты фенильной модификации sd-PHK, мишенью которой является МАР4К4.
ФИГ. 54 демонстрирует эффекты фенильной модификации sd-PHK, мишенью которой является PCSK9.
ФИГ. 55 обеспечивает сумму 5-уридильных модификаций.
ФИГ. 56 демонстрирует, что модуляция химии в положении 5 уридина приводит к измененному распределению в тканях sd-PHK в печени.
ФИГ. 57 демонстрирует, что модуляция химии в положении 5 уридина приводит к измененному распределению в тканях sd-PHK в сердце и жире.
ФИГ. 58 подытоживает конструкцию синтонов.
ФИГ. 59 обеспечивает неограничивающие примеры синтонов.
ФИГ. 60 обеспечивает дополнительные неограничивающие примеры синтонов.
ФИГ. 61 обеспечивает схематическое изображение линкера, содержащего протонируемые амины, что способствует эндосомальному высвобождению sd-PHK.
ФИГ. 62 обеспечивает схематическую демонстрацию включения протонируемых функциональных групп для способствования высвобождению эндосомов.
ФИГ. 63 обеспечивает дополнительные неограничивающие примеры синтонов.
ФИГ. 64 обеспечивает схематическое изображение GIV «клик» химии, посредством чего олигонуклеотид может быть быстро диверсифицирован.
ФИГ. 65 демонстрирует, что размещение фосфоротиоатного хвоста не изменяет активность сайленсинга.
ФИГ. 66 показывает, что смысловая нить, содержащая дезокси (6) фосфоротиоатный хвост, сохраняет активность сайленсинга.
ФИГ. 67 показывает, что присутствие продолженных дезоксифосфоротиоатных хвостов в смысловой нити или антисмысловой нити может уменьшать эффективность.
ФИГ. 68 показывает, что антисмысловая нить, содержащая 5' дезоксифосфоротиоатный хвост, может иметь уменьшенную эффективность.
ФИГ. 69 раскрывает, что размещение фосфоротиоатного хвоста не изменяет активность сайленсинга.
ФИГ. 70 раскрывает, что sd-PHK с 21-mer ведущей нитью, которая полностью фосфоротиоатно модифицированна, является активной.
ФИГ. 71 показывает, что включение фосфоротиоатного хвоста в смысловую нить может привести к уменьшенной активности сайленсинга.
ФИГ. 72 раскрывает, что стабилизация sd-PHK приводит к повышенной жизнеспособности клеток.
ФИГ. 73 раскрывает, что sd-PHK могут выдерживать изменения в химии линкера.
ФИГ. 74 представляет сумму химической оптимизации sd-PHK, мишенью которой является PPIB.
ФИГ. 75 представляет дополнительные неограничивающие примеры химической оптимизации sd-PHK, мишенью которой является PPIB.
ФИГ. 76 показывает влияние включения 5-метил С и U на активность sd-PHK, мишенью которой является PPIB.
ФИГ. 77 сравнивает нокдаун PPIB, достигнутый при помощи sd-PHK, содержащей 14 mer и 20 mer или содержащей 15 mer и 19 mer.
ФИГ. 78 подытоживает влияние химической оптимизации sd-PHK, мишенью которой является PPIB.
ФИГ. 79 показывает активность оптимизированной sd-PHK, мишенью которой является АроВ.
ФИГ. 80 показывает дополнительные примеры активной химически оптимизированной sd-PHK, мишенью которой является АроВ.
ФИГ. 81 раскрывает, что химически оптимизированная sd-PHK, мишенью которой является АроВ является более эффективной при индукции генного сайленсинга в печени, чем более традиционные соединения.
ФИГ. 82 раскрывает стабильность sd-PHK в сыворотке.
ФИГ. 83 показывает доставку sd-PHK в спинной мозг после инъекции в центральную нервную систему.
ФИГ. 84 раскрывает проникновение в спинной мозг после доставки sd-PHK в центральную нервную систему.
ФИГ. 85 раскрывает симптомы доставки sd-PHK в стволовую часть мозга.
Подробное описание
Аспекты настоящего изобретения относятся к способам и композициям, участвующим в генном сайленсинге. Настоящее изобретение основано, по меньшей мере частично, на удивительном открытии, что sd-PHK молекулы могут быть эффективно доставлены в множество различных тканей. Внутривенное введение, как показано в данной заявке, приводит к эффективному распределению в тканях, таких как печень, селезенка, сердце и легкие, в то время как подкожная доставка показала свою высокую эффективность для доставки в ткани, такие как печень и кожа. Дополнительно, эффективное поглощение и доставка sd-PHK молекул в легкие, включая альвеолярный макрофаг, былы достигнуты за счет инсуффляции. Способы и композиции, описанные в данной заявке, имеют широкое применение для улучшенной in vivo доставки нуклеиновой кислоты.
sd-PHK молекулы
Аспекты настоящего изобретения относятся к sd-PHK молекулам. Как используется в данной заявке, «sd-РНК» или «sd-РНК молекула» относится к самодоставляющейся РНК молекуле, такой, как описано в данной заявке и включено путем ссылки на публикации РСТ номер WO 2010/033247 (заявка № PCT/US 2009/005247), поданной 22 сентября 2009 года под названием «Самодоставляющиеся PHKi соединения уменьшенного размера». Короче говоря, sd-PHK, (также называемая sd-PHKнано) является выделенной асимметричной двухцепочечной молекулой нуклеиновой кислоты, содержащей ведущую нить, с минимальной длиной 16 нуклеотидов, и пассажирскую нить 8-18 нуклеотидов в длину, в которой двухцепочечная молекула нуклеиновой кислоты имеет двухцепочечную область и одноцепочечную область, одноцепочечная область имеет 4-12 нуклеотидов в длину и имеет по меньшей мере три нуклеотидных модификации каркаса. В предпочтительных осуществлениях двухцепочечная молекула нуклеиновой кислоты, имеет один конец, который является тупым или включает один или два нуклеотидных выступа. sd-PHK молекулы могут быть оптимизированы путем химической модификации, а в некоторых случаях путем присоединения гидрофобных конъюгатов.
В некоторых осуществлениях, sd-PHK содержит изолированную двухцепочечную молекулу нуклеиновой кислоты, содержащую ведущую нить и пассажирскую нить, в которой область молекулы, которая является двухцепочечной, состоит из 8-15 нуклеотидов в длину, в которой ведущая нить содержит одноцепочечную область, которая составляет 4-12 нуклеотидов, в которой одноцепочечная область ведущей нити содержит 3, 4, 5, 6, 7, 8, 9, 10, 11 или 12 фосфоротиоатных модификаций, и в которой по меньшей мере 40% нуклеотидов в двухцепочечной нуклеиновой кислоте модифицированы.
Полинуклеотиды в соответствии с настоящим изобретением называют в данной заявке выделенными двухцепочечными или дуплексными нуклеиновыми кислотами, олигонуклеотидами или полинуклеотидами, наномолекулами, нано РНК, sd-PHKнано, sd-PHK или РНК молекулами в соответствии с настоящим изобретением.
sd-PHK гораздо более эффективно поглощаются клетками по сравнению с обычными siPHK. Эти молекулы обладают высокой эффективностью при сайленсинге экспрессии генов-мишеней и имеют значительные преимущества по сравнению с ранее описанными молекулами PHKi, включая высокую активность в присутствии сыворотки, эффективную самодоставку, совместимость с широким спектром линкеров, а также снижение наличия или полное отсутствие химических модификаций, которые связаны с токсичностью.
В отличие от одноцепочечных полинуклеотидов, дуплексные полилинуклеотиды традиционно трудно доставить в клетку, поскольку они имеют жесткую структуру и большое количество отрицательных зарядов, что делает мембранную передачу затрудненной. sd-PHK однако, хотя и частично двухцепочечные, распознаются in vivo, как одноцепочечные и, как таковые, способны эффективно доставляться через клеточные мембраны. В результате полинуклеотиды в соответствии с настоящим изобретением способны во многих случаях самодоставляться. Таким образом, полинуклеотиды в соответствии с настоящим изобретением могут быть сформулированы аналогично обычным PHKi агентам или они могут быть доставлены в клетку или субъекту отдельно (или при помощи носителей не-доставляющего типа), и позволяют самодоставку. В одном осуществлении настоящего изобретения, представлены самодоставляющиеся асимметричные двухцепочечные РНК молекулы, в которых одна часть молекулы напоминает обычный РНК дуплекс и вторая часть молекулы является одноцепочечной.
Олигонуклеотиды в соответствии с настоящим изобретением в некоторых аспектах имеют сочетание асимметричных структур, включая двухцепочечную область и одноцепочечную область из 5 нуклеотидов или более, специфические модели химической модификации и сопряжены с липофильными или гидрофобными молекулами. Этот класс PHKi как соединения имеет высокую эффективность in vitro и in vivo. Считается, что уменьшение размера жесткой области дуплекса в сочетании с фосфоротиоатными модификациями применяемое к одноцепочечной области вносит вклад в наблюдаемую высокую эффективность.
В основе настоящего изобретения лежит, по меньшей мере частично, удивительное открытие, что sd-PHK совместимы с несколькими средствами введения и эффективно распределяются в различных тканях. Как показано в разделе примеров, и, как обсуждается ниже, эффективное системное введение sd-PHK было достигнуто путем внутривенной инъекции, включая использование повторяющегося введения/краткосрочного непрерывного режима инфузии. Удивительно, но эффективное распределение sd-PHK было достигнуто в сердечной ткани с помощью этого метода, хотя сердце традиционно нелегко поразить олигонуклеотидами. Распределение sd-PHK наблюдалось также в тканях, таких как печень, легкие и селезенка путем внутривенного введения.
Другим неожиданным аспектом настоящего изобретения была эффективность с которой sd-PHK была доставлена с использованием подкожного введения. Как показано в разделе примеров, и, как обсуждается далее ниже, в некоторых случаях, метод подкожного введения привел к более высокой концентрации РНК молекул в данной ткани, чем внутривенное введение той же дозы той же РНК молекулы.
Дополнительным неожиданным аспектом настоящего изобретения была эффективная доставка sd-PHK в легкие, и, в частности в альвеолярные клетки макрофагов, после введения путем инсуффлирования. Типичные PHKi подходы ориентированы на эпителиальные клетки, а не альвеолярные клетки макрофагов. Доставка в альвеолярные клетки макрофагов является выгодной для лечения легочных состояний, таких как астма.
Дополнительным неожиданным аспектом настоящего изобретения было открытие, что доставка sd-PHK в ткань-мишень может быть по существу увеличена с помощью стабилизации sd-PHK путем химической модификации. Например, введение стабилизирующей модификации, такой как 2'О метил, 2' дезокси и/или фосфоротиоата может улучшить распределение в тканях. Например, в некоторых случаях, химическая модификация может привести к по меньшей мере 1,5, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100, 105, 110, 115, 120, 125, 130, 135, 140, 145, 150, 155, 160, 165, 170, 175, 180, 185, 190, 195, 200, 225, 250, 275, 300, 325, 350, 375, 400, 425, 450, 475, 500 или более чем 500-кратному увеличению распределения в тканях. В некоторых неорганичивающих примерах, sd-PHK распределяется в печени, сердце, легких, мозге или жире. Как показано в Примерах, повышение уровня стабилизации модификаций было найдено приведет к увеличению генного сайленсинга по сравнению с менее стабильными sd-PHK и повышенному биораспределению в тканях, таких как печень, сердце и жир.
Другим неожиданным аспектом настоящего изобретения была демонстрация того, что введение гидрофобных модификаций sd-PHK усиливает клеточное поглощение и распределение в тканях. Некоторые не ограничивающие примеры гидрофобных модификаций оснований включают: тиофеновую, октильную, октин-1-ильную, этинильную, изобутильную, метильную, пиридил амидную и фенильную. Фигура 57 показывает, что включение гидрофобных модификаций в sd-PHK приводит к изменению распределения в тканях.
В некоторых случаях, по меньшей мере один из С или U остатков включает гидрофобную модификацию. В некоторых случаях, множество С и U содержат гидрофобную модификацию. Например, по меньшей мере 10%, 15%, 20%, 30%, 40%, 50%, 55%, 60% 65%, 70%, 75%, 80%, 85%, 90% или по меньшей мере 95% С и U могут содержать гидрофобную модификацию. В некоторых осуществлениях, все С и U содержат гидрофобную модификацию.
Другим неожиданным аспектом настоящего изобретения является разработка высоко фосфоротиоатных РНК молекул, которые поддерживают возможность ввода RISC. Несколько прошлых групп пытались разработать полностью фосфоротиоатные PHKi соединения, однако, эти соединения не демонстрируют правильный фармакокинетический профиль. Полностью фосфоротиоатные одноцепочечные PHKi соединения также были разработаны ранее, однако, эти соединения не эффективно вводят RISC. Примечательно, что в данной заявке гибридные PHKi соединения были разработаны, которые эффективно вводят RISC и содержат полные фосфоротиоатные каркасы. В некоторых случаях, по меньшей мере 10%, 15%, 20%, 30%, 40%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90% или по меньшей мере 95% нуклеотидов в ведущей нити и/или пассажирской нити фосфоротиоатно модифицированы. В определенных осуществлениях, все нуклеотиды в ведущей нити и/или пассажирской нити фосфоротиоатно модифицированы. В некоторых осуществлениях, РНК молекула является одноцепочечной и по меньшей мере 10%, 15%, 20%, 30%, 40%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90% или по меньшей мере 95% нуклеотидов в одноцепочечной молекуле фосфоротиоатно модифицированы. В определенных осуществлениях, все нуклеотиды в одноцепочечной молекуле фосфоротиоатно модифицированы.
Еще один аспект настоящего изобретения относится к повышению эндосомального высвобождения sd-PHK молекул путем включения протонируемых аминов. В некоторых осуществлениях, протонируемые амины включены в смысловую нить (в часть молекулы, которая удаляется после RISC нагрузки).
В предпочтительном осуществлении, PHKi соединения в соответствии с настоящим изобретением включают асимметричное соединение, содержащее дуплексную область (необходимую для эффективного RISC введения 10-15 оснований в длину) и одноцепочечную область из 4-12 нуклеотидов в длину, с 13 нуклеотидным дуплексом. 6 нуклеотидная одноцепочечная область является предпочтительной в некоторых осуществлениях. Одноцепочечная область новых PHKi соединений также включает 2-12 фосфоротиоатных межнуклеотидных связей (под названием фосфоротиоатные модификации). 6-8 фосфоротиоатных межнуклеотидных связей являются более предпочтительными в некоторых осуществлениях. Дополнительно, PHKi соединения в соответствии с настоящим изобретением также включают уникальную модель химической модификации, которая обеспечивает стабильность и совместима с RISC введением. Комбинация этих элементов привела к неожиданным свойствам, которые весьма полезны для доставки PHKi реагентов in vitro и in vivo.
Модель химической модификации, которая обеспечивает стабильность и совместима с RISC введением, включает модификации смысловой, или пассажирской, нити, а также антисмысловой, или ведущей, нити. Например, пассажирская нить может быть модифицирована любым химическим веществом, которое подтверждает стабильность и не влияет на активность. Такие модификации включают 2' рибо модификации (О-метил, 2'F, 2 дезокси и др.) и модификации каркаса, такие как фосфоротиоатные модификации. Предпочтительная модель химической модификации в пассажирской нити включает О-метильную модификацию С и U нуклеотидов в пассажирской нити или альтернативно пассажирская нить может быть полностью О-метильно модифицирована.
Ведущая нить, например, также может быть модифицирована любой химической модификацией, которая подтверждает стабильность без препятствия RISC введению. Предпочтительная модель химической модификации в ведущей нити включает большинство С и U нуклеотидов, которые 2'F модифицированы и 5' конец фосфорилирован. Другая предпочтительная модель химической модификации в ведущей нити включает 2'O-метильную модификацию положения 1 и C/U в положениях 11-18 и 5' конце химического фосфорилирования. Еще одна предпочтительная модель химической модификации в ведущей нити включает 2'O-метильную модификацию положения 1 и C/U в положениях 11-18 и 5' конце химического фосфорилирования и 2'F модификацию C/U в положениях 2-10. В некоторых осуществлениях пассажирская нить и/или ведущая нить содержит по меньшей мере одну 5-метильную С или U модификацию.
В некоторых осуществлениях, по меньшей мере 30% нуклеотидов в sd-PHK модифицированы. Например, по меньшей мере 30%, 31%, 32%, 33%, 34%, 35%, 36%, 37%, 38%, 39%, 40%, 41%, 42%, 43%, 44%, 45%, 46%, 47%, 48%, 49%, 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59%, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% нуклеотидов в sd-PHK модифицированы. В некоторых осуществлениях, 100% нуклеотидов в sd-PHK модифицированы.
dsPHK, сформулированная в соответствии с настоящим изобретением, также включает PHKori. PHKori относится к классу РНК молекул, описанных и включенных путем ссылки на публикацию РСТ номер WO 2009/102427 (заявка № PCT/US 2009/000852), поданной И февраля 2009 года под названием «МОДИФИЦИРОВАННЫЕ PHKI ПОЛИНУКЛЕОТИДЫ И ИХ ПРИМЕНЕНИЕ».
В некоторых осуществлениях, молекула PHKori содержит двухцепочечный РНК (dsPHK) конструкт из 12-35 нуклеотидов в длину, для ингибирования экспрессии гена-мишени, содержащий: смысловую нить, имеющую 5'-конец и 3'-конец, где смысловая нить сильно модифицирована 2'-модифицированными рибозными сахарами, и где 3-6 нуклеотиды в центральной части смысловой нити не модифицированы 2'-модифицированными рибозными сахарами, и антисмысловую нить, имеющую 5'-конец и 3'-конец, который гибридизуется в смысловую нить и мРНК гена-мишени, где dsPHK ингибирует экспрессию гена-мишени последовательность-зависимым образом.
PHKori может содержать любую из модификаций, описанных в данной заявке. В некоторых осуществлениях, по меньшей мере 30% нуклеотидов в PHKori модифицированы. Например, по меньшей мере 30%, 31%, 32%, 33%, 34%, 35%, 36%, 37%, 38%, 39%, 40%, 41%, 42%, 43%, 44%, 45%, 46%, 47%, 48%, 49%, 50%, 51%, 52%, 53%, 54%, 55%, 56%, 57%, 58%, 59%, 60%, 61%, 62%, 63%, 64%, 65%, 66%, 67%, 68%, 69%, 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% нуклеотидов в PHKori модифицированы. В некоторых осуществлениях, 100% нуклеотидов в sd-PHK модифицированы. В некоторых осуществлениях, только пассажирская нить PHKori содержит модификации.
Описанные выше модели химической модификации олигонуклеотидов в соответствии с настоящим изобретением хорошо переносятся и на самом деле улучшили эффективность асимметричных PHKi соединений. Сочетание модификаций в PHKi, когда их используют вместе в полинуклеотиде, приводит к достижению оптимальной эффективности в пассивном поглощении PHKi. Ликвидация любого из описанных компонентов (стабилизация ведущей нити, фосфоротиоатное растяжение, стабилизация смысловой нити и гидрофобный конъюгат) или увеличение в размерах в некоторых случаях приводит к неоптимальной эффективности и в некоторых случаях к полной потере эффективности. Сочетание элементов приводит к разработке соединения, которое является полностью активным после пассивной доставки в клетки, такие как клетки HeLa.
Данные в примерах, приведенных ниже, демонстрируют высокую эффективность олигонуклеотидов в соответствии с настоящим изобретением in vivo при местном и системном введении. sd-PHK может быть дополнительно улучшена в некоторых случаях за счет улучшения гидрофобности соединений с использованием новых типов химии. Например, одна химия связана с использованием гидрофобных модификаций оснований. Любые основания в любом положении могут быть модифицированы, если модификация приводит к увеличению коэффициента распределения основания. Предпочтительные места для модификации химией представляют собой положения 4 и 5 пиримидинов. Основным преимуществом этих положений являются (а) простота синтеза и (б) отсутствие помех спаривания оснований и формирование А формы спирали, которые необходимы для RISC комплекса нагрузки и распознавания мишеней. Использовали версию sd-PHK соединений, где присутствует множество дезокси уридинов, не мешая общей эффективности соединений. Дополнительно, значительное улучшение в распределении в тканях и клеточного поглощения может быть получено за счет оптимизации структуры гидрофобного конъюгата. В некоторых предпочтительных осуществлениях структура стерина модифицирована для изменения (увеличения/уменьшения) С17 присоединенной цепи. Этот тип модификации приводит к значительному повышению клеточного поглощения и улучшению свойств поглощения в тканях in vivo.
Настоящее изобретение не ограничено в своем применении деталями конструкции и расположением компонентов, изложенными в следующем описании или показанными на чертежах. Настоящее изобретение может иметь другие осуществления и практикуется или осуществляется различными способами. Дополнительно, фразеология и терминология, используемые в данной заявке, приведены с целью описания и не должны рассматриваться как ограничивающие. Использование «включая», «содержащий», или «имеющий», «включающий», «содержащий» и их вариаций в данной заявке предназначено для охвата пунктов, перечисленных ниже в данной заявке, и их эквивалентов, а также дополнительных пунктов.
Таким образом, аспекты настоящего изобретения относятся к изолированным двухцепочечным молекулам нуклеиновой кислоты, содержащим ведущую (антисмысловую) нить и пассажирскую (смысловую) нить. Как используется в данной заявке, термин «двухцепочечный» относится к одной или нескольким молекулам нуклеиновой кислоты, в которых по меньшей мере часть нуклеомономеров являются комплементарными и связанными водородной связью с образованием двухцепочечной области. В некоторых осуществлениях, длина ведущей нити находится в диапазоне 16-29 нуклеотидов. В определенных осуществлениях, ведущая нить содержит 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28 или 29 нуклеотидов в длину. Ведущая нить имеет комплементарность с геном-мишенью. Комплементарность между ведущей нитью и геном-мишенью может существовать в любой части ведущей нити. Комплементарность, как используется в данной заявке, может быть совершенной комплементарностью или менее чем совершенной комплементарностью, до тех пор, пока ведущая нить достаточно комплементарна мишени для обеспечения PHKi. В некоторых осуществлениях комплементарность относится к менее чем 25%, 20%, 15%, 10%, 5%, 4%, 3%, 2% или 1% несоответствию между ведущей нитью и мишенью. Совершенная комплементарность относится к 100% комплементарности. Таким образом, настоящее изобретение имеет то преимущество, что может переносить вариации последовательностей, которые можно было бы ожидать в связи с генетической мутацией, полиморфизмом штаммов или эволюционной дивергенцией. Например, siPHK последовательности с инсерциями, делециями и точечными мутациями по отношению к последовательности-мишени были также признаны эффективными для ингибирования. Дополнительно, не все положения siPHK в равной мере вносят вклад в распознавание мишени. Несоответствия в центре siPHK являются наиболее важными и, по сути, отменяют расщепление РНК-мишени. Несоответствия вверх от центра или вверх от сайта расщепления, которые относятся к антисмысловой нити, переносят, но значительно снижают расщепление РНК-мишени. Несоответствия вниз от центра или сайта расщепления, которые относятся к антисмысловой нити, предпочтительно располагаются рядом с 3' концом антисмысловой нити, например, 1, 2, 3, 4, 5 или 6 нуклеотидов из 3' конца антисмысловой нити, переносятся и снижают расщепления РНК-мишени только незначительно.
Хотя и не желая быть связанным какой-либо конкретной теорией, в некоторых осуществлениях, ведущая нить имеет по меньшей мере 16 нуклеотидов в длину и прикрепляется к Argonaute белку в RISC. В некоторых осуществлениях, когда ведущая нить загружается в RISC, она имеет определенный участок затравки (seed region) и расщепление мРНК-мишени происходит напротив положения 10-11 ведущей нити. В некоторых осуществлениях, 5' конец ведущей нити является фосфорилированным или может быть фосфорилирован. Молекулы нуклеиновой кислоты, описанные в данной заявке, могут называться минимальной запускающей РНК.
В некоторых осуществлениях, длина пассажирской нити находится в диапазоне 8-15 нуклеотидов. В определенных осуществлениях, пассажирская нить содержит 8, 9, 10, 11, 12, 13, 14 или 15 нуклеотидов в длину. Пассажирская нить имеет комплементарность с ведущей нитью. Комплементарность между пассажирской нитью и ведущей нитью может существовать в любой части пассажирской или ведущей нити. В некоторых осуществлениях, существует 100% комплементарность между ведущей и пассажирской нитью в двухцепочечной области молекулы.
Аспекты настоящего изобретения относятся к двухцепочечным молекулам нуклеиновой кислоты с минимальными двухцепочечными областями. В некоторых осуществлениях область молекулы, которая является двухцепочечной, находится в диапазоне 8-15 нуклеотидов в длину. В определенных осуществлениях, область молекулы, которая является двухцепочечной, составляет 8, 9, 10, 11, 12, 13, 14 или 15 нуклеотидов в длину. В определенных осуществлениях двухцепочечная область составляет 13 нуклеотидов в длину. Может существовать 100% комплементарность между ведущей и пассажирской нитью, или может быть одно или несколько несоответствий между ведущей и пассажирской нитью. В некоторых осуществлениях, на одном конце двухцепочечной молекулы, молекула либо имеет тупой конец, или имеет однонуклеотидный выступ. Одноцепочечная область молекулы в некоторых осуществлениях составляет 4-12 нуклеотидов в длину. Например, одноцепочечная область может составлять 4, 5, 6, 7, 8, 9, 10, 11 или 12 нуклеотидов в длину. Однако, в определенных осуществлениях, одноцепочечная область также может быть меньше чем 4 или более чем 12 нуклеотидов в длину. В определенных осуществлениях, одноцепочечная область составляет 6 или 7 нуклеотидов в длину.
PHKi конструкты, связанные с изобретением, могут иметь термодинамическую стабильность (ΔG) менее чем -13 ккал/моль. В некоторых осуществлениях, термодинамическая стабильность (ΔG) составляет менее чем -20 ккал/моль. В некоторых осуществлениях происходит потеря эффективности, когда (ΔG) становится ниже -21 ккал/моль. В некоторых осуществлениях (ΔG) имеет значение выше чем -13 ккал/моль, что совместимо с аспектами настоящего изобретения. Не желая быть связанными теорией, в некоторых осуществлениях молекула с относительно высоким (ΔG) значением может стать активной при относительно высокой концентрации, в то время как молекула с относительно низким (ΔG) значением может стать активной при относительно низкой концентрации. В некоторых осуществлениях, (ΔG) значение может быть выше чем -9 ккал/моль. Эффекты генного сайленсинга опосредованные PHKi конструктами, связанными с настоящим изобретением, содержащие минимальные двухцепочечные области, являются неожиданными, поскольку молекулы имеют практически идентичное строение, но более низкая термодинамическая стабильность была продемонстрирована как неактивная (Rana et al., 2004).
Аспекты настоящего изобретения относятся к химически модифицированным РНК молекулам, которые имеют повышенную стабильность. В некоторых случаях, химически модифицированная sd-PHK молекула имеет период полураспада в сыворотке, превышающий 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или более 24 часов, включая любые промежуточные значения. В определенных осуществлениях, sd-PHK имеет период полураспада в сыворотке, превышающий 12 часов.
Не желая быть связанными теорией, результаты, описанные в данной заявке, предполагают, что участок 8-10 б.п. dsPHK или dsДHK будет структурно распознан белковыми компонентами RISC или ко-факторами RISC. Дополнительно, существует требование свободной энергии для запуска соединения, которое может быть либо воспринято белковыми компонентами и/или достаточно стабильным, чтобы взаимодействовать с такими компонентами, так что оно может быть загружено в Argonaute белок. Если оптимальная термодинамика присутствует и существует двухцепочечная область, которая предпочтительно по меньшей мере имеет 8 нуклеотидов, то дуплекс будут распознан и загружен в PHKi механизм.
В некоторых осуществлениях, термодинамическая стабильность повышается за счет использования оснований LNA. В некоторых осуществлениях, вводятся дополнительные химические модификации. Некоторые не ограничивающие примеры химических модификаций включают: 5' фосфат, 2'-О-метил, 2'-O-этил, 2'-фтор, риботимидин, С-5 пропинил-dC (pdC) и C-5-пропинил-dU (pdU); С-5 пропинил-С (рС) и С-5 пропинил-U (pU); 5-метил С, 5-метил U, 5-метил dC, 5-метил dU метокси, (2,6-диаминопурин), 5'-диметокситритил-N4-этил-2'-дезоксицитидин и MGB (связующий с незначительной бороздкой). Следует иметь в виду, что более одной химической модификации могут быть объединены в одной молекуле.
Молекулы, связанные с изобретением оптимизированы для повышения эффективности и/или снижения токсичности. Например, нуклеотидная длина ведущей и/или пассажирской нити, и/или количество фосфоротиоатных модификаций в ведущей и/или пассажирской нити, может в некоторых аспектах влиять на эффективность РНК молекулы, в то время как замена 2'-фтор (2'F) модификаций на 2'-O-метил (2'ОМе) модификации может в некоторых аспектах влиять на токсичность молекулы. В частности, снижение 2'F содержания в молекуле, по прогнозам, снижает токсичность молекулы. В разделе Примеры представлены молекулы, в которых 2'F модификации были устранены, предлагая преимущества по сравнению с ранее описанными PHKi соединениями из-за прогнозируемого снижения токсичности. Дополнительно, количество фосфоротиоатных модификаций в РНК молекуле может влиять на поглощение молекулы в клетке, например эффективность пассивного поглощения молекулы в клетке. Предпочтительные осуществления молекул, описанных в данной заявке, не имеют 2'F модификации и еще характеризуются равной эффективностью клеточного поглощения и проникновения в ткани. Такие молекулы представляют собой значительное улучшение по сравнению с предыдущим уровнем техники, такие молекулы описаны Accell и Wolfram, которые сильно модифицированы с широким использованием 2'F.
В некоторых осуществлениях, ведущая нить имеет приблизительно 18-19 нуклеотидов в длину и имеет приблизительно 2-14 фосфатных модификаций. Например, ведущая нить может содержать 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или более 14 нуклеотидов, которые фосфатно модифицированы. Ведущая нить может содержать одну или несколько модификаций, которые придают повышенную стабильность без препятствий RISC введению. Фосфатно модифицированные нуклеотиды, такие как фосфоротиоатно модифицированные нуклеотиды, могут находиться на 3' конце, 5' конце или распределены в ведущей нити. В некоторых осуществлениях, 3' концевые 10 нуклеотидов в ведущей нити содержат 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 фосфоротиоатно модифицированных нуклеотидов. Ведущая нить может также содержать 2'F и/или 2'ОМе модификации, которые могут быть расположены по всей молекуле. В некоторых осуществлениях, нуклеотид в положении один ведущей нити (нуклеотид в самом 5' положении ведущей нити) 2'ОМе модифицирован и/или фосфорилирован. С и U нуклеотиды в ведущей нити могут быть 2'F модифицированы. Например, С и U нуклеотиды в положениях 2-10 ведущей нити из 19-и нуклеотидов (или соответствующих положениях в ведущей нити другой длины) могут быть 2'F модифицированы. С и U нуклеотиды в ведущей нити также могут быть 2'ОМе модифицированы. Например, С и U нуклеотиды в положениях 11-18 ведущей нити из 19-и нуклеотидов (или соответствующих положениях в ведущей нити другой длины) могут быть 2'ОМе модифицированы. В некоторых осуществлениях, нуклеотид на самом 3' конце ведущей нити не модифицирован. В определенных осуществлениях, большинство С и U в ведущей нити 2'F модифицированы и 5' конец ведущей нити фосфорилирован. В других осуществлениях, положение 1 и С или U в положениях 11-18 2'ОМе модифицированы и 5' конец ведущей нити фосфорилирован. В других осуществлениях, положение 1 и С или U в положениях 11-18 2'ОМе модифицированы, 5' конец ведущей нити фосфорилирован, и С или U в положениях 2-10 2'F модифицированы.
В некоторых аспектах, оптимальная пассажирская нить составляет приблизительно 11-14 нуклеотидов в длину. Пассажирская нить может содержать модификации, придающие повышенную стабильность. Один или более нуклеотидов в пассажирской нити могут быть 2'ОМе модифицированы. В некоторых осуществлениях, один или более из С и/или U нуклеотидов в пассажирской нити 2'ОМе модифицированы, или все из С и U нуклеотидов в пассажирской нити 2'ОМе модифицированы. В определенных осуществлениях, все нуклеотиды в пассажирской нити 2'ОМе модифицированы. Один или более нуклеотидов в пассажирской нити могут быть также фосфат-модифицированы, например, фосфоротиоат модифицированы. Пассажирская нить может также содержать 2' рибо, 2'F и 2 дезокси модификации или любую из вышеуказанных комбинаций. Как показано в Примерах, модели химической модификации как в ведущей, так и в пассажирской нитях, хорошо переносятся и комбинация химических модификаций показана в данной заявке как приводящая к повышенной эффективности и самодоставке РНК молекул.
Аспекты настоящего изобретения относятся к PHKi конструктам, имеющим продленные одноцепочечные области, относительно двухцепочечных областей, по сравнению с молекулами, которые использовали ранее для PHKi. Одноцепочечная область молекул может быть модифицирована для способствования клеточному поглощению или генному сайленсингу. В некоторых осуществлениях, фосфоротиоатная модификация одноцепочечной области влияет на клеточное поглощение и/или генный сайленсинг. Область ведущей нити, которая фосфоротиоатно модифицирована, может включать нуклеотиды с одноцепочечными и двухцепочечными областями молекулы. В некоторых осуществлениях, одноцепочечная область включает 2-12 фосфоротиоатных модификаций. Например, одноцепочечная область может включать 2, 3, 4, 5, 6, 7, 8, 9, 10, 11 или 12 фосфоротиоатных модификаций. В некоторых случаях, одноцепочечная область содержит 6-8 фосфоротиоатных модификаций. В некоторых осуществлениях, sd-PHK молекула имеет более одной одноцепочечной области.
Молекулы, связанные с настоящим изобретением, также оптимизированы для клеточного поглощения. В РНК молекулах, описанных в данной заявке, ведущая и/или пассажирская нити могут быть присоединены к конъюгату. В определенных осуществлениях конъюгат является гидрофобным. Гидрофобный конъюгат может быть малой молекулой с коэффициентом распределения выше чем 10. Конъюгат может быть молекулой стеринового типа, такой как холестерин, или молекулой с увеличенной длиной полиугольной цепи, присоединенной к С17, и присутствие конъюгата может влиять на способность РНК молекулы поглощаться в клетке с или без липидного трансфекционного реагента. Конъюгат может быть присоединен к пассажирской или ведущей нити через гидрофобный линкер. В некоторых осуществлениях, гидрофобный линкер имеет 5-12С в длину, и/или основан на гидроксипирролидине. В некоторых осуществлениях, гидрофобный конъюгат присоединен к пассажирской нити и CU остатки пассажирской и/или ведущей нити модифицированы. В некоторых осуществлениях, по меньшей мере 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90% или 95% CU остатков в пассажирской нити и/или ведущей нити модифицированы. В некоторых аспектах, молекулы, связанные с настоящим изобретением, являются самодоставляющимися (sd). Как используют в данной заявке, «самодоставка» относится к способности молекулы к доставке в клетку без необходимости в дополнительной основе доставки, такой, как трансфекционный реагент.
Аспекты настоящего изобретения относятся к отбору молекул для использования в PHKi. Исходя их данных, описанных в данной заявке, молекулы, имеющие двухцепочечную область из 8-15 нуклеотидов, могут быть отобраны для использования в PHKi. В некоторых осуществлениях, молекулы отбирают, исходя из их термодинамической стабильности (ΔG). В некоторых осуществлениях, будут отобраны молекулы, имеющие (ΔG) менее чем -13 ккал/моль. Например, значение (ΔG) может составлять -13, -14, -15, -16, -17, -18, -19, -21, -22 или менее чем -22 ккал/моль. В других осуществлениях, значение (ΔG) может превышать -13 ккал/моль. Например, (ΔG) значение может составлять -12, -11, -10, -9, -8, -7 или более чем -7 ккал/моль. Должно быть оценено, что ΔG может быть рассчитано с использованием любого способа, известного в данной области техники. В некоторых осуществлениях ΔG рассчитывают при помощи Mfold, доступного через интернет-сайт Mfold (http://mfold.bioinfo.rpi.edu/cgi-bin/rna-forml.cgi). Способы расчета ΔG описаны в, и включены путем ссылки из таких ссылок: Zuker, М. (2003) Nucleic Acids Res., 31(13): 3406-15; Mathews, D.H., Sabina, J., Zuker, M. and Turner, D.H. (1999) J. Mol. Biol. 288: 911-940; Mathews, D.H., Disney, M.D., Childs, J.L., Schroeder, S.J., Zuker, M., and Turner, D.H. (2004) Proc. Natl. Acad. Sci. 101: 7287-7292; Duan, S., Mathews, D.H., and Turner, D.H. (2006) Biochemistry 45: 9819-9832; Wuchty, S., Fontana, W., Hofacker, I.L., и Schuster, P. (1999) Bjopolymers 49: 145-165.
Аспекты настоящего изобретения относятся к использованию молекул нуклеиновой кислоты, описанных в данной заявке, с минимальными двухцепочечными областями и/или с (ΔG) менее чем -13 ккал/моль, для генного сайленсинга. PHKi молекулы могут быть введены in vivo или in vitro, и эффекты генного сайленсинга могут быть достигнуты in vivo или in vitro.
В определенных осуществлениях, полинуклеотид содержит выступы 5'- и/или 3'-конца. Количество и/или последовательность нуклеотидного выступа на одном конце полинуклеотида могут быть одинаковыми или отличаться от другого конца полинуклеотида. В определенных осуществлениях, один или несколько нуклеотидных выступов могут содержать химическую модификацию(и), такие как фосфоротиоатная или 2'-ОМе модификация.
В определенных осуществлениях, полинуклеотид не модифицирован. В других осуществлениях по меньшей мере один нуклеотид модифицирован. В других осуществлениях, модификация включает 2'-Н или 2'-модифицированный рибозный сахар на 2-м нуклеотиде от 5'-конца ведущей последовательности. «2-й нуклеотид» определяется как второй нуклеотид от 5'-конца полинуклеотида.
Как используется в данной заявке, «2'-модифицированный рибозный сахар» включает рибозные сахара, которые не имеют 2'-ОН группу. «2'-модифицированный рибозный сахар» не включает 2'-дезоксирибозу (находится в немодифицированных канонических ДНК нуклеотидах). Например, 2'-модифицированный рибозный сахар может быть 2'-О-алкил нуклеотидами, 2'-дезокси-2'-фтор нуклеотидами, 2'-дезокси нуклеотидами или их комбинацией.
В определенных осуществлениях, 2'-модифицированные нуклеотиды являются пиримидиновыми нуклеотидами (например, C/U). Примеры 2'-O-алкил нуклеотидов включают 2'-O-метил нуклеотиды, или 2'-O-аллил нуклеотиды.
В определенных осуществлениях, миниРНК полинуклеотид в соответствии с настоящим изобретением с указанной выше 5'-концевой модификацией обладает значительным (например, по меньшей мере приблизительно 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90% или более) менее «вне целевым» генным сайленсингом по сравнению с аналогичными конструктами без указанной 5'-концевой модификации, тем самым значительно улучшая общую специфичность PHKi реагента или терапии.
Как используется в данной заявке, «вне целевой» генный сайленсинг относится к непреднамеренному генному сайленсингу за счет, например, ложной гомологии последовательностей между антисмысловой (ведущей) последовательностью и непреднамеренной целевой мРНК последовательностью.
В соответствии с данным аспектом изобретения, определенные модификации ведущей нити дополнительно повышают нуклеазную стабильность, и/или понижают индукцию интерферона, без значительного снижения PHKi активности (или не снижают PHKi активность вообще). В некоторых осуществлениях, 5'-стволовая последовательность может содержать 2'-модифицированный рибозный сахар, такой, как 2'-О-метил модифицированный нуклеотид, на 2-м нуклеотиде на 5'-конце полинуклеотида и, в некоторых осуществлениях, никаких других модифицированных нуклеотидов. Шпилечная структура, имеющая такую модификацию, может иметь расширенную специфичность к мишени или уменьшенный вне целевой сайленсинг по сравнению с аналогичным конструктом без 2'-O-метил модификации в указанном положении.
Определенные комбинации специфичных модификаций 5'-стволовых последовательностей и 3'-стволовых последовательностей могут привести к дополнительным неожиданным преимуществам, что отчасти проявляется повышенной способностью ингибировать экспрессию генов-мишеней, повышенной стабильностью сыворотки, и/или повышенной специфичностью мишени, и т.д.
В определенных осуществлениях, ведущая нить содержит 2'-O-метил модифицированный нуклеотид, на 2-м нуклеотиде на 5'-конце ведущей нити и никакие другие модифицированные нуклеотиды.
В других аспектах миниРНК структуры в соответствии с настоящим изобретением опосредуют зависимый от последовательностей генный сайленсинг при помощи микроРНК механизма. Как используется в данной заявке, термин «микроРНК» («miPHK»), также известный в уровне техники как «малые временные РНК» («stPHK»), относится к малой (10-50 нуклеотидов) РНК, которая генетически закодирована (например, при помощи вирусных, млекопитающих геномов или геномов растений) и способна направлять или опосредовать РНК сайленсинг. «MiPHK расстройство» относится к болезни или расстройству, характеризующемуся аномальной экспрессией или активностью miPHK.
МикроРНК участвуют в нижней регуляции генов-мишеней в критических маршрутах, таких как развитие и рак у мышей, червей и млекопитающих. Генный сайленсинг с помощью микроРНК механизма достигается за счет специфичного, но пока несовершенного спаривания оснований miPHK и ее мессенжером-мишенью РНК (мРНК). Различные механизмы могут быть использованы в микроРНК-опосредованной нижней регуляции экспрессии мРНК мишени.
miPHK являются некодирующими РНК приблизительно из 22 нуклеотидов, которые могут регулировать экспрессию генов на пост транскрипционном или трансляционном уровне в течение развития растений и животных. Одна общая черта miPHK представляет собой то, что все они иссечены из приблизительно 70 нуклеотидных предшественников РНК ствол-петель, под названием пре-miPHK, вероятно, от Dicer, их РНКзы III-типа фермента, или гомолога. Встречающиеся в природе miPHK экспрессируются эндогенными генами in vivo и обрабатываются от шпилечного или ствол-петля предшественника (пре-miPHK или pri-miPHK) от Dicer или других РНКаз. miPHK могут существовать временно in vivo, как двухцепочечный дуплекс, но только одна нить поглощается комплексом RISC для прямого генного сайленсинга.
В некоторых осуществлениях описана версия sd-PHK соединений, которые являются эффективными при клеточном поглощении и ингибировании активности miPHK. По сути соединения похожи на RISC входящую версию, но большие модели химических модификаций нитей оптимизированы в том, чтобы блокировать расщепление и действовать как эффективный ингибитор активности RISC. Например, соединение может быть полностью или в основном О-метил модифицировано с содержанием PS, описанным выше. Для этих типов соединений 5' фосфорилирование не является необходимым. Наличие двухцепочечной области предпочтительно, так как способствует клеточному поглощению и эффективной RISC загрузке.
Другой маршрут, который использует небольшие РНК как последовательность-специфичные регуляторы, является маршрутом РНК (PHKi) интерференции, который является эволюционно консервативным ответом на присутствие двухцепочечной РНК (dsPHK) в клетке. dsPHK расщепляются в ~20-пар оснований (bp) дуплексов малого вмешательства РНК (siPHK) от Dicer. Эти малые РНК собирают в мультибелковые эффекторные комплексы под названием РНК-индуцированные сайленсинг комплексы (RISC). siPHK затем направляют расщепление мРНК мишени с совершенной комплементарностью.
Некоторые аспекты биогенеза, белковых комплексов, а также функции распределяются между siPHK маршрутом и miPHK маршрутом. Рассматриваемые одноцепочечные полинуклеотиды могут имитировать dsPHK в siPHK механизме или микроРНК в miPHK механизме.
В определенных осуществлениях, модифицированные PHKi конструкты могут иметь улучшенную стабильность в сыворотке и/или спинно-мозговой жидкости по сравнению с немодифицированными PHKi конструктами с той же последовательностью.
В определенных осуществлениях, структура PHKi конструкта не вызывает ответ интерферона в первичных клетках, таких как первичные клетки млекопитающих, включая первичные клетки от человека, мыши и других грызунов, и других нечеловеческих млекопитающих. В определенных осуществлениях, PHKi конструкт также может быть использован для ингибирования экспрессии гена-мишени в организме беспозвоночных.
Для дополнительного повышения стабильности рассматриваемых конструктов in vivo, 3'-конец шпилечной структуры может быть заблокирован защитной группой(ами). Например, защитные группы, такие как инвертированные нуклеотиды, инвертированные абазические фрагменты, или амино-конец модифицированные нуклеотиды, могут быть использованы. Инвертированные нуклеотиды могут содержать инвертированный дезоксинуклеотид. Инвертированные абазические фрагменты могут содержать инвертированный дезоксиабазиновый фрагмент, такой как 3',3'-связанный или 5',5'-связанный дезоксиабазиновый фрагмент.
PHKi конструкты в соответствии с настоящим изобретением способны ингибировать синтез любого белка-мишени, кодируемого геном-мишенью(ями). Изобретение включает способы для ингибирования экспрессии гена-мишени, либо в клетке in vitro или in vivo. Таким образом, PHKi конструкты в соответствии с настоящим изобретением являются полезными для лечения пациентов с заболеванием, которое характеризуется избыточной экспрессией гена-мишени.
Ген-мишень может быть эндогенным или экзогенным (например, вводят в клетку вирусом, или с помощью технологии рекомбинантных ДНК) в клетке. Такие способы могут включать введение РНК в клетку в количестве, достаточном для ингибирования экспрессии гена-мишени. К примеру, такие РНК молекулы могут иметь ведущую нить, комплементарную с нуклеотидной последовательностью гена-мишени, таким образом, что композиция ингибирует экспрессию гена-мишени.
Настоящее изобретение также относится к векторам, экспрессирующим нуклеиновые кислоты в соответствии с настоящим изобретением, и клеткам, содержащим такие векторы или нуклеиновые кислоты. Клетка может быть клеткой млекопитающих in vivo или в культуре, такой как клетка человека.
Настоящее изобретение дополнительно относится к композициям, содержащим рассматриваемые PHKi конструкты, и фармацевтически приемлемый носитель или разбавитель.
Другой аспект настоящего изобретения относится к способу ингибирования экспрессии гена-мишени в клетках млекопитающих, включающему контактирование клетки млекопитающего с любым рассматриваемым PHKi конструктом.
Способ может быть проведен in vitro, ex vivo или in vivo, в, например, клетках млекопитающих в культуре, такой как клетка человека в культуре.
Клетки-мишени (например, клетки млекопитающих) могут контактировать в присутствии реагента доставки, такого как липид (например, катионный липид) или липосома.
Другой аспект настоящего изобретения обеспечивает способ ингибирования экспрессии гена-мишени в клетках млекопитающих, включающий контактирование клетки млекопитающего с вектором, экспрессирующим рассматриваемые PHKi конструкты.
В одном аспекте настоящего изобретения, обеспечивается более длинный дуплексный полинуклеотид, включая первый полинуклеотид, который находится в диапазоне размеров от приблизительно 16 до приблизительно 30 нуклеотидов, второй полинуклеотид, который находится в диапазоне размеров от приблизительно 26 до приблизительно 46 нуклеотидов, причем первый полинуклеотид (антисмысловая нить) комплементарен ко второму полинуклеотиду (смысловая нить) и гену-мишени, и в котором оба полинуклеотида образуют дуплекс и где первый полинуклеотид содержит одноцепочечную область из более чем 6 оснований в длину и модифицированный альтернативной моделью химической модификации, и/или включает конъюгатный фрагмент, который облегчает доставку в клетку. В этом осуществлении от приблизительно 40% до приблизительно 90% нуклеотидов в пассажирской нити, от приблизительно 40% до приблизительно 90% нуклеотидов в ведущей нити, и от приблизительно 40% до приблизительно 90% нуклеотидов в одноцепочечной области первого полинуклеотида являются химически модифицированными нуклеотидами.
В осуществлении, химически модифицированный нуклеотид в полинуклеотидном дуплексе может быть любым химически модифицированным нуклеотидом, известным в данной области техники, таким, как подробно обсуждалось выше. В конкретном осуществлении, химически модифицированный нуклеотид выбран из группы, состоящей из 2'F модифицированных нуклеотидов, 2'-O-метил-модифицированных и 2'дезокси нуклеотидов. В другом конкретном осуществлении, химически модифицированные нуклеотиды получают в результате «гидрофобных модификаций» нуклеотидного основания. В другом конкретном осуществлении, химически модифицированные нуклеотиды являются фосфоротиоатами. В дополнительном конкретном осуществлении, химически модифицированные нуклеотиды являются комбинацией фосфоротиоатов, 2'-О-метил, 2'дезокси, гидрофобных модификаций и фосфоротиоатов. Поскольку такие группы модификаций относятся к модификации рибозного кольца, каркаса и нуклеотида, то возможно, что некоторые модифицированные нуклеотиды будут нести комбинацию всех трех типов модификаций.
В другом осуществлении, химическая модификация не является одинаковой в различных областях дуплекса. В конкретном осуществлении, первый полинуклеотид (пассажирская нить), имеет большое количество разнообразных химических модификаций в различных положениях. Для этого полинуклеотида до 90% нуклеотидов могут быть химически модифицированы и/или имеют несоответствия, которые введены.
В другом осуществлении, химические модификации первого или второго полинуклеотида включают, но не ограничиваясь приведенным, модификацию 5' положения уридина и цитозина (4-пиридил, 2-пиридил, индолил, фенил (С6Н5ОН); триптофанил (C8H6N)CH2CH(NH2)CO), изобутил, бутил, аминобензил; фенил, нафтил и т.д.), где химическая модификация может изменить возможности спаривания оснований нуклеотида. Для ведущей нити важной особенностью этого аспекта изобретения является положение химической модификации относительно 5' конца антисмысловой нити и последовательности. Например, химическое фосфорилирование 5' конца ведущей нити, как правило, полезно для эффективности. О-метильные модификации в участке затравки (seed region) смысловой нити (положение 2-7 относительно 5' конца) обычно плохо переносятся, в то время как 2'F и дезокси хорошо переносятся. Средняя часть ведущей нити и 3' конец ведущей нити являются более обещающими в применяемом типе химических модификаций. Дезокси модификации не переносятся на 3' конце ведущей нити.
Уникальная особенность этого аспекта изобретения включает использование гидрофобной модификации на основаниях. В одном осуществлении, гидрофобные модификации предпочтительно расположены вблизи 5' конца ведущей нити, в других осуществлениях, они локализованы в середине ведущей нити, в другом осуществлении они локализованы на 3' конце ведущей нити, и еще в другом осуществлении они распределены по всей длине полинуклеотида. Тот же тип моделей применяется к пассажирской нити дуплекса.
Другая часть молекулы представляет собой одноцепочечную область. Одноцепочечная область, как ожидают, находится в диапазоне от 6 до 40 нуклеотидов.
В одном осуществлении, одноцепочечная область первого полинуклеотида содержит модификации, выбранные из группы, состоящей из от 40% до 90% гидрофобных модификаций оснований, от 40% до 90% фосфоротиоатов, от 40% до 90% модификаций рибозного фрагмента, а также любой комбинации указанных выше.
Эффективность загрузки ведущей нити (первый полинуклеотид) в RISC комплекс может быть изменена для сильно модифицированных полинуклеотидов, так что в одном осуществлении, дуплексный полинуклеотид включает несоответствие между нуклеотидами 9, 11, 12, 13 или 14 на ведущей нити (первый полинуклеотид) и противоположным нуклеотидом на смысловой нити (второй полинуклеотид) для способствования эффективной загрузке ведущей нити.
Более подробно аспекты настоящего изобретения, описаны в следующих разделах.
Дуплексные характеристики
Двухцепочечные олигонуклеотиды в соответствии с настоящим изобретением могут быть образованы двумя отдельными комплементраными нитями нуклеиновой кислоты. Дуплексное образование может происходить либо внутри, либо снаружи клетки, содержащей ген-мишень.
Как используется в данной заявке, термин «дуплекс» включает область двухцепочечной молекулы нуклеиновой кислоты, которая (которые) связана водородными связями с комплементарной последовательностью. Двухцепочечные олигонуклеотиды в соответствии с настоящим изобретением могут содержать нуклеотидную последовательность, то есть смысловой ген-мишень и комплементарную последовательность, которая является антисмысловым геном-мишенью. Смысловая и антисмысловая нуклеотидные последовательности соответствуют последовательности гена-мишени, например, они идентичны или достаточно идентичны для осуществления ингибирования гена-мишени (например, приблизительно по меньшей мере 98% идентичны, 96% идентичны, 94%, 90% идентичны, 85% идентичны, или 80% идентичны) последовательности гена-мишени.
В определенных осуществлениях, двухцепочечный олигонуклеотид в соответствии с настоящим изобретением является двухцепочечным по всей его длине, например, без выступающей одноцепочечной последовательности на обоих концах молекулы, например, имеет тупые концы. В других осуществлениях отдельные молекулы нуклеиновой кислоты могут быть разной длины. Иными словами, двухцепочечный олигонуклеотид в соответствии с настоящим изобретением не является двухцепочечным по всей его длине. Например, если используются две отдельные молекулы нуклеиновой кислоты, одна из молекул, например, первая молекула, содержащая антисмысловую последовательность, может быть больше, чем вторая молекула, которая гибридизует с ней (оставив часть молекулы одноцепочечной). Аналогичным образом, если используется одна молекула нуклеиновой кислоты, то части молекулы на обоих концах могут остаться одноцепочечными.
В одном осуществлении, двухцепочечный олигонуклеотид в соответствии с настоящим изобретением содержит несоответствия и/или петли или выпуклости, но является двухцепочечным по меньшей мере по приблизительно 70% от длины олигонуклеотида. В другом осуществлении, двухцепочечный олигонуклеотид в соответствии с настоящим изобретением является двухцепочечным по меньшей мере по приблизительно 80% от длины олигонуклеотида. В другом осуществлении, двухцепочечный олигонуклеотид в соответствии с настоящим изобретением является двухцепочечным по меньшей мере по приблизительно 90%-95% от длины олигонуклеотида. В другом осуществлении, двухцепочечный олигонуклеотид в соответствии с настоящим изобретением является двухцепочечным по меньшей мере по приблизительно 96%-98% от длины олигонуклеотида. В определенных осуществлениях, двухцепочечный олигонуклеотид в соответствии с настоящим изобретением содержит по меньшей мере или до 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15 несоответствий.
Модификации
Нуклеотиды в соответствии с настоящим изобретением могут быть модифицированы в различных местах, включая сахарный фрагмент, фосфодиэфирную связь и/или основание.
В некоторых осуществлениях, фрагмент основания нуклеозида может быть модифицирован. Например, пиримидиновое основание может быть модифицировано в 2, 3, 4, 5 и/или 6 положениях пиримидинового цикла. В некоторых осуществлениях, экзоциклический амин цитозина может быть модифицирован. Пуриновое основание также может быть модифицировано. Например, пуриновое основание может быть модифицировано в 1, 2, 3, 6, 7 или 8 положении. В некоторых осуществлениях, экзоциклический амин аденина может быть модифицирован. В некоторых случаях, атом азота в кольце фрагмента основания может быть замещен другим атомом, таким как углерод. Модификация фрагмента основания может быть любой приемлемой модификацией. Примеры модификаций известны специалистам в данной области техники. В некоторых осуществлениях, модификации оснований включают алкилированные пурины или пиримидины, ацилированные пурины или пиримидины или другие гетероциклы.
В некоторых осуществлениях, пиримидин может быть модифицирован в 5 положении. Например, как показано на Фигуре 19, 5 положение пиримидина может быть модифицировано алкильной группой, алкинильной группой, алкенильной группой, ацильной группой, или их замещенными производными. В других примерах, как показано на Фигуре 19, 5 положение пиримидина может быть модифицировано гидроксильной группой или алкоксильной группой или их замещенным производным. Дополнительно, как показано на Фигуре 29, N4 положение пиримидина может быть алкилировано. В других примерах, как показано на Фигуре 30, пиримидина 5-6 связь может быть насыщенной, атом азота в пиримидиновом цикле может быть замещен атомом углерода, и/или O2 и O4 атомы могут быть замещены атомами серы. Следует понимать, что другие модификации также возможны.
В других примерах, как показано на Фигурах 29 и 30, N7 положение и/или N2 и/или N3 положение пурина могут быть модифицированы алкильной группой или их замещенным производным. В дальнейших примерах, как показано на Фигуре 30, третье кольцо может быть гибридизовано с кольцевой системой бициклических пуринов и/или атом азота в системе пуринового кольца может быть замещен атомом углерода. Следует понимать, что другие модификации также возможны.
Неограничивающие примеры пиримидинов, модифицированных в 5 положении, раскрыты в патенте США 5591843, патенте США 7,205,297, патенте США 6,432,963 и патенте США 6,020,483; не ограничивающие примеры пиримидинов, модифицированных в положении N4 раскрыты в патенте США 5,580,731; неограничивающие примеры пуринов, модифицированных в 8 положении, раскрыты в патенте США 6,355,787 и патенте США 5,580,972; не ограничивающие примеры пуринов, модифицированных в положении N6 раскрыты в патенте США 4,853,386, патенте США 5,789,416 и патенте США 7,041,824, и не ограничивающие примеры пуринов, модифицированных в 2 положении, раскрыты в патенте США 4,201,860 и патенте США 5,587,469, все из которых включены в данную заявку путем ссылки.
Неограничивающие примеры модифицированных оснований включают N4,N4-этаноцитозин, 7-деазаксантозин, 7-деазагуанозин, 8-оксо-N6-метиладенин, 4-ацетилцитозин, 5-(карбоксигидроксилметил) урацил, 5-фторурацил, 5-бромурацил, 5-карбоксиметиламинометил-2-тиоурацил, 5-карбоксиметиламинометил урацил, дигидроурацил, инозин, N6-изопентенил-аденин, 1-метиладенин, 1-метилпсевдоурацил, 1-метилгуанин, 1-метилинозин, 2,2-диметилгуанин, 2-метиладенин, 2-метилгуанин, 3-метилцитозин, 5-метилцитозин, N6-метиладенин, 7-метилгуанин, 5-метиламинометил урацил, 5-метокси аминометил-2-тиоурацил, 5-метоксиурацил, 2-метилтио-N6-изопентиладенин, псевдоурацил, 5-метил-2-тиоурацил, 2-тиоурацил, 4-тиоурацил, 5-метилурацил, 2-тиоцитозин и 2,6-диаминопурин. В некоторых осуществлениях, фрагмент основания может быть гетероциклическим основанием, кроме пурина или пиримидина. Гетероциклическое основание может быть необязательно модифицировано и/или замещено.
Сахарные фрагменты включают природные, немодифицированные сахара, например, моносахарид (например, пентозу, например, рибозу, дезоксирибозу), модифицированные сахара и сахарные аналоги. В общем, возможные модификации нуклеомономеров. в частности, сахарного фрагмента, включают, например, замену одной или нескольких гидроксильных групп галогеном, гетероатомом, алифатической группой или функционализацию гидроксильной группы, такой, как эфир, амин, тиол, или тому подобное.
Одной из наиболее полезных групп модифицированных нуклеомономеров являются 2'-O-метил нуклеотиды. Такие 2'-O-метил нуклеотиды могут быть названы «метилированными», и соответствующие нуклеотиды могут быть получены из неметилированных нуклеотидов с последующим алкилированием или непосредственно из метилированных нуклеотидных реагентов. Модифицированные нуклеомономеры могут быть использованы в сочетании с немодифицированными нуклеомономерами. Например, олигонуклеотид в соответствии с настоящим изобретением может содержать как метилированные, так и неметилированные нуклеомономеры.
Некоторые иллюстративные модифицированные нуклеомономеры включают сахар- или каркас-модифицированные рибонуклеотиды. Модифицированные рибонуклеотиды могут содержать не встречающееся в природе основание (вместо встречающегося в природе основания), такое как уридины или цитидины, модифицированные в 5'-положении, например, 5'-(2-амино)пропил уридин и 5'-бром уридин; аденозины и гуанозины, модифицированные в 8-положении, например, 8-бром гуанозин; деаза нуклеотиды, например, 7-деаза-аденозин, а также N-алкилированные нуклеотиды, например, N6-метил аденозин. Также, сахар-модифицированные рибонуклеотиды могут иметь 2'-ОН-группу, замененную Н, алкокси (или OR), R или алкил, галоген, SH, SR, амино (например, NH2, NHR, NR2,), или CN группу, в которой R представляет собой низший алкил, алкенил или алкинил.
Модифицированные рибонуклеотиды также могут иметь фосфодиэфирную группу, присоединенную к прилегающим рибонуклеотидам, замещенным модифицированной группой, например, фосфоротиоатной группой. В целом, различные нуклеотидные модификации могут быть объединены.
Несмотря на то, что антисмысловая (ведущая) нить может быть по существу идентична по меньшей мере части гена-мишени (или генов) по меньшей мере по отношению к свойствам спаривания оснований, последовательность не обязательно должна быть совершенно идентичной, чтобы быть полезной, например, для ингибирования экспрессии фенотипа гена-мишени. Обычно, более высокая гомология может быть использована для компенсации использования более короткого антисмыслового гена. В некоторых случаях, антисмысловая нить в целом будет по существу идентична (хотя в антисмысловой ориентации) гену-мишени.
Использование 2'-O-метил модифицированной РНК также может быть полезно в тех случаях, в которых желательно свести к минимуму клеточные реакции на стресс. РНК имеющая 2'-O-метил нуклеомономеры не могже быть распознана клеточным механизмом, который, как думали, распознает немодифицированную РНК. Использование 2'-O-метилированной или частично 2'-O-метилированной РНК может избежать ответа интерферона на двухцепочечные нуклеиновые кислоты, при сохранении ингибирования РНК мишени. Это может быть полезно, например, для избежания интерферон или другого клеточного ответа на стресс, как в коротких PHKi (например, siPHK) последовательностях, которые вызывают ответ интерферона, так и в более длинных PHKi последовательностях, что может вызвать ответ интерферона.
В целом, модифицированные сахара могут включать D-рибозу, 2'-О-алкил (включая 2'-O-метил и 2'-O-этил), например, 2'-алкокси, 2'-амино, 2'-S-алкил, 2'-гало (включая 2'-фтор), 2'-метоксиэтокси, 2'-аллилокси(-ОСН2СН=СН2), 2'-пропаргил, 2'-пропил, этинил, этенил, пропенил, и циано и тому подобное. В одном осуществлении, сахарный фрагмент может быть гексозой и включен в олигонуклеотид, как описано (Augustyns, K., et al., Nucl. Acids. Res. 18:4711 (1992)). Иллюстративные нуклеомономеры можно найти, например, в патенте США № 5,849,902, включенном в данную заявку путем ссылки.
Определения специфичных функциональных групп и химические термины более подробно описаны ниже. Для целей настоящего изобретения, химические элементы, определены в соответствии с Периодической таблицей элементов, CAS версия, Handbook of Chemistry and Physics, 75th Ed., сведения из первоисточника, и специфические функциональные группы, как правило, определены как описано там. Дополнительно, общие принципы органической химии, а также специфические функциональные фрагменты и реакционная способность, описаны в Organic Chemistry, Thomas Sorrell, University Science Books, Sausalito: 1999, полное содержание которой включено в данную заявку путем ссылки.
Некоторые соединения в соответствии с настоящим изобретением могут существовать в конкретных геометрических или стереоизомерных формах. Настоящее изобретение рассматривает все такие соединения, включая цис- и транс-изомеры, R- и S-энантиомеры, диастереомеры, (D)-изомеры, (L)-изомеры, рацемические смеси, и другие их смеси, как подпадающие под объем настоящего изобретения. Дополнительные асимметричные атомы углерода могут присутствовать в заместителе, таком как алкильная группа. Все такие изомеры, а также их смеси, предназначены для включения в настоящее изобретение.
Изомерные смеси, содержащие любое из различных соотношений изомеров, могут быть использованы в соответствии с настоящим изобретением. Например, если только два изомера объединены, все смеси, содержащие 50:50, 60:40, 70:30, 80:20, 90:10, 95:5, 96:4, 97:3, 98:2, 99:1 или 100:0 соотношения изомеров, предусмотрены настоящим изобретением. Специалист в данной области техники легко поймет, что аналогичные соотношения предусмотрены для более сложных смесей изомеров.
Если, к примеру, конкретный энантиомер соединения в соответствии с настоящим изобретением является целевым, он может быть получен путем асимметрического синтеза, либо путем деривации с хиральным вспомогательным веществом, где в результате диастереомерная смесь разделяется и вспомогательная группа расщепляется, чтобы обеспечить чистые целевые энантиомеры. Альтернативно, если молекула содержит основную функциональную группу, такую как амино или кислотную функциональную группу, такую как карбоксил, диастереомерные соли образуются с соответствующей оптически активной кислотой или основанием, с последующим разрешением диастереомеров, образованных таким образом, путем фракционной кристаллизации или хроматографическими способами, хорошо известными в данной области техники, и последующим восстановлением чистых энантиомеров.
В определенных осуществлениях, олигонуклеотиды в соответствии с настоящим изобретением содержат 3' и 5' концы (за исключением круговых олигонуклеотидов). В одном осуществлении, 3' и 5' концы олигонуклеотида могут быть по существу защищены от нуклеаз, например, путем модификации 3' или 5' связей (например, патент США № 5,849,902 и WO 98/13526). Например, олигонуклеотиды могут быть устойчивыми за счет включения «блокирующей группы». Термин «блокирующая группа», используемый в данной заявке, относится к заместителям (например, кроме ОН групп), которые могут быть присоединены к олигонуклеотидам или нуклеомономерам, в качестве защитных групп или групп сочетания для синтеза (например, FITC, пропил (CH2-CH2-CH3), гликоль (-О-CH2-CH2-О-) фосфат (PO32-), кислый фосфонат или фосфорамидит). «Блокирующие группы» также включают «концевые блокирующие группы» или «экзонуклеазные блокирующие группы», которые защищают 5' и 3' концы олигонуклеотида, включая модифицированные нуклеотиды и не нуклеотидные экзонуклеазные резистентные структуры.
Иллюстративные концевые блокирующие группы включают кэп-структуры (например, 7-метилгуанозин кэп), инвертированные нуклеомономеры, например, с 3'-3' или 5'-5' концевыми инверсиями (см., например, Ortiagao et al., 1992. Antisense Res. Dev. 2:129), метилфосфонат, фосфорамидит, не нуклеотидные группы (например, не-нуклеотидные линкеры, амино линкеры, конъюгаты) и тому подобное. 3' концевой нуклеомономер может содержать модифицированный сахарный фрагмент. 3' концевой нуклеомономер содержит 3'-О, который может быть необязательно замещен блокирующей группой, которая предотвращает 3'-экзонуклеазный распад олигонуклеотида. Например, 3'-гидроксил может быть этерифицирован нуклеотидом через 3'→3' межнуклеотидную связь. Например, алкилокси радикал может быть метокси, этокси или изопропокси, и, предпочтительно, этокси. Необязательно, 3'→3' связанный нуклеотид на 3' конце может быть связан замещающей связью. Чтобы уменьшить нуклеазный распад, 5' наиболее 3'→5' связь может быть модифицированной связью, например, фосфоротиоатной или P-алкилоксифосфотриэфирной связью. Предпочтительно, две 5' наиболее 3'→5' связи являются модифицированными связями. Необязательно, 5' концевой гидрокси фрагмент может быть этерифицирован фосфорсодержащим фрагментом, например, фосфатом, фосфоротиоатом или P-этоксифосфатом.
Специалисту в данной области техники будет понятно, что синтетические способы, описанные в данной заявке, используют различные защитные группы. Под термином «защитная группа», как используется в данной заявке, подразумевают, что определенный функциональный фрагмент, например, О, S или N, временно заблокирован, так что реакция может быть проведена выборочно на другом реактивном сайте в многофункциональном соединении. В определенных осуществлениях, защитная группа реагирует выборочно с хорошим выходом, с получением защищенного субстрата, который устойчив к предполагаемым реакциям; защитная группа должна быть выборочно удаляемой с хорошим выходом при помощи доступных, предпочтительно нетоксичных реагентов, которые не атакуют другие функциональные группы; защитная группа образует легко отделяемые производные (более предпочтительно без генерации новых стереогенных центров), а также защитная группа имеет минимум дополнительных функциональных групп, чтобы избежать дополнительных сайтов реакции. Как указано в данной заявке, кислородные, серные, азотные и углеродные защитные группы могут быть использованы. Гидроксильные защитные группы включают метил, метоксиметил (MOM), метилтиометил (MTM), трет-бутилтиометил, (фенилдиметилсилил)метоксиметил (SMOM), бензилоксиметил (BOM), п-метоксибензилоксиметил (PMBM), (4-метоксифенокси)метил (п-AOM), гуаякольметил (GUM), t-бутоксиметил, 4-пентенилоксиметил (POM), силоксиметил, 2-метоксиэтоксиметил (MEM), 2,2,2-трихлорэтоксиметил, бис(2-хлорэтокси)метил, 2-(триметилсилил)этоксиметил (SEMOR), тетрагидропиранил (THP), 3-бромтетрагидропиранил, тетрагидротиопиранил, 1-метоксициклогексил, 4-метокситетрагидропиранил (MTHP), 4-метокситетрагидротиопиранил, 4-метокситетрагидротиопиранил S,S-диоксид, 1-[(2-хлор-4-метил)фенил]-4-метоксипиперидин-4-ил] (CTMP), 1,4-диоксан-2-ил, тетрагидрофуранил, тетрагидротиофуранил, 2,3,3а,4,5,6,7,7а-октагидро-7,8,8-триметил-4,7-метанобензофуран-2-ил, 1-этоксиэтил, 1-(2-хлорэтокси)этил, 1-метил-1-метоксиэтил, 1-метил-1-бензилоксиэтил, 1-метил-1-бензилокси-2-фторэтил, 2,2,2-трихлорэтил, 2-триметилсилилэтил, 2-(фенилсиленил)этил, трет-бутил, аллил, п-хлорфенил, п-метоксифенил, 2,4-динитрофенил, бензил, п-метоксибензил, 3,4-диметоксибензил, о-нитробензил, п-нитробензил, п-галобензил, 2,6-дихлорбензил, п-цианобензил, п-фенилбензил, 2-пиколил, 4-пиколил, 3-метил-2-пиколил N-оксидо, дифенилметил, п,п'-динитробензилгидрил, 5-дибензосуберил, трифенилметил, α-нафтилдифенилметил, п-метоксифенилдифенилметил, ди(п-метоксифенил)фенилметил, три(п-метоксифенил)метил, 4-(4'-бромфенацилоксифенил)-дифенилметил, 4,4',4''-трис(4,5-дихлорфталимидофенил)метил, 4,4',4''-трис(левулиноилоксифенил)метил, 4,4',4''-трис(бензоилоксифенил)метил, 3-(имидазол-1-ил)бис(4',4''-диметоксифенил)метил, 1,1-бис(4-метоксифенил)-1'-пиренилметил, 9-антрил, 9-(9-фенил)ксантенил, 9-(9-фенил-10-оксо)антрил, 1,3-бензодитиолан-2-ил, бензизотиазолил S,S-диоксидо, триметилсилил (TMS), триэтилсилил (TES), триизопропилсилил (TIPS), диметилизопропилсилил (IPDMS), диэтилизопропилсилил (DEIPS), диметилтексилсилил, t-бутилдиметилсилил (TBDMS), t-бутилдифенилсилил (TBDPS), трибензилсилил, три-п-ксилилсилил, трифенилсилил, дифенилметилсилил (DPMS), трет-бутилметоксифенилсилил (TBMPS), формиат, бензоилформиат, ацетат, хлорацетат, дихлорацетат, трихлорацетат, трифторацетат, метоксиацетат, трифенилметоксиацетат, феноксиацетат, п-хлорфеноксиацетат, 3-фенилпропионат, 4-оксопентаноат (левулинат), 4,4-(этилендитио)пентаноат (левулиноилдитиоацеталь), пивалоат, адамантоат, кротонат, 4-метоксикротонат, бензоат, п-фенилбензоат, 2,4,6-триметилбензоат(мезитоат), алкилметилкарбонат, 9-флуоренилметилкарбонат (Fmoc), алкил этилкарбонат, алкил 2,2,2-трихлорэтилкарбонат (Troc), 2-(триметилсилил)этилкарбонат (TMSEC), 2-(фенилсульфонил)этилкарбонат (Psec), 2-(трифенилфосфино) этилкарбонат (Peoc), алкилизобутилкарбонат, алкилвинилкарбонат, алкилаллилкарбонат, алкил п-нитрофенилкарбонат, алкилбензилкарбонат, алкил п-метоксибензилкарбонат, алкил 3,4-диметоксибензил карбонат, алкил о-нитробензил карбонат, алкил п-нитробензил карбонат, алкил S-бензил тиокарбонат, 4-этокси-1-нафтилкарбонат, метил дитиокарбонат, 2-йодбензоат, 4-азидобутират, 4-нитро-4-метилпентаноат, о-(дибромметил)бензоат, 2-формилбензолсульфонат, 2-(метилтиометокси)этил, 4-(метилтиометокси)бутират, 2-(метилтиометоксиметил) бензоат, 2,6-дихлор-4-метилфеноксиацетат, 2,6-дихлор-4-(1,1,3,3-тетраметилбутил) феноксиацетат, 2,4-бис(1,1-диметилпропил)феноксиацетат, хлордифенилацетат, изобутират, моносукциноат, (E)-2-метил-2-бутеноат, о-(метоксикарбонил)бензоат, α-нафтоат, нитрат, алкил N,N,N',N'-тетраметилфосфордиамидат, алкил N-фенилкарбамат, борат, диметилфосфинотиоил, алкил 2,4-динитрофенилсульфенат, сульфат, метансульфонат (мезилат), бензилсульфонат, и тозилат (Ts). Для защиты 1,2 - или 1,3-диолов, защитные группы включают метилен ацеталь, этилиден ацеталь, 1-трет-бутилэтилиденкеталь, 1-фенилэтилиденкеталь, (4-метоксифенил)этилиден ацеталь, 2,2,2-трихлорэтилиден ацеталь, ацетонид, циклопентилиден кеталь, циклогексилиден кеталь, циклогептилиден кеталь, бензилиден ацеталь, п-метоксибензилиден ацеталь, 2,4-диметоксибензилиден кеталь, 3,4-диметоксибензилиден ацеталь, 2-нитробензилиден ацеталь, метоксиметилен ацеталь, этоксиметилен ацеталь, диметоксиметилен орто эфир, 1-метоксиэтилиден орто эфир, 1-этоксиэтилидин орто эфир, 1,2-диметоксиэтилиден орто эфир, α-метоксибензилиден орто эфир, 1-(N,N-диметиламино)этилиден производное, α-(N,N'-диметиламино)бензилиден производное, 2-оксациклопентилиден орто эфир, ди-трет-бутилсилилен группа (DTBS), 1,3-(1,1,3,3-тетраизопропилдисилоксанилиден) производное (TIPDS), тетра-трет-бутоксидисилоксан-1,3-диилиден производное (TBDS), циклические карбонаты, циклические боронаты, этилборонат и фенилборонат. Амино-защитные группы включают метилкарбамат, этилкарбамат, 9-флуоренилметилкарбамат (Fmoc), 9-(2-сульфо)флуоренилметилкарбамат, 9-(2,7-дибром)флуоренилметилкарбамат, 2,7-ди-трет-бутил-[9-(10,10-диоксо-10,10,10,10-тетрагидротиоксантил)]метилкарбамат (DBD-Tmoc), 4-метоксифенацилкарбамат (Phenoc), 2,2,2-трихлорэтилкарбамат (Troc), 2-триметилсилилэтилкарбамат (Теос), 2-фенилэтилкарбамат (hZ), 1-(1-адамантил)-1-метилэтилкарбамат (Adpoc), 1,1-диметил-2-галоэтилкарбамат, 1,1-диметил-2,2-дибромэтилкарбамат (DB-трет-BOC), 1,1-диметил-2,2,2-трихлорэтилкарбамат (TCBOC), 1-метил-1-(4-бифенилил)этилкарбамат (Врос), 1-(3,5-ди-трет-бутилфенил)-1-метилэтилкарбамат (t-Bumeoc), 2-(2'- и 4'-пиридил)этилкарбамат (Pyoc), 2-(N,N-дициклогексилкарбокамидо)этилкарбамат, трет-бутилкарбамат (ВОС), 1-адамантилкарбамат (Adoc), винилкарбамат (Voc), аллилкарбамат (Alloc), 1-изопропилаллилкарбамат (Ipaoc), коричный карбамат (Coc), 4-нитроциннамилкарбамат (Noc), 8-хинолилкарбамат, N-гидроксипиперидинилкарбамат, алкилдитиокарбамат, бензилкарбамат (Cbz), п-метоксибензилкарбамат (Moz), п-нитобезилкарбамат, п-бромбензилкарбамат, п-хлорбензилкарбамат, 2,4-дихлорбензилкарбамат, 4-метилсульфинилбензилкарбамат (Msz), 9-антрилметилкарбамат, дифенилметилкарбамат, 2-метилтиоэтилкарбамат, 2-метилсульфонилэтилкарбамат, 2-(п-толуолсульфонил)этилкарбамат, [2-(1,3-дитианил)]метилкарбамат (Dmoc), 4-метилтиофенилкарбамат (Mtpc), 2,4-диметилтиофенилкарбамат (Bmpc), 2-фосфониоэтилкарбамат (Peoc), 2-трифенилфосфониоизопропилкарбамат (Ppoc), 1,1-диметил-2-цианоэтилкарбамат, м-хлор-п-ацилоксибензилкарбамат, п-(дигидроксиборил)бензилкарбамат, 5-бензизоксазолилметилкарбамат, 2-(трифторметил)-6-хромонилметилкарбамат (Tcroc), м-нитрофенилкарбамат, 3,5-диметоксибензилкарбамат, о-нитробензилкарбамат, 3,4-диметокси-6-нитробензилкарбамат, фенил(о-нитрофенил)метилкарбамат, фенотиазинил-(10)-карбонилпроизводное, N'-п-толуолсульфониламинокарбонилпроизводное, N'-фениламинотиокарбонилпроизводное, трет-амилкарбамат, S-бензилтиокарбамат, п-цианобензилкарбамат, циклобутилкарбамат, циклогексилкарбамат, циклопентилкарбамат, циклопропилметилкарбамат, п-децилоксибензилкарбамат, 2,2-диметоксикарбонилвинилкарбамат, о-(N,N-диметилкарбоксамидо)бензилкарбамат, 1,1-диметил-3-(N,N-диметилкарбоксамидо)пропилкарбамат, 1,1-диметилпропинилкарбамат, ди(2-пиридил)метилкарбамат, 2-фуранилметилкарбамат, 2-йодэтилкарбамат, изоборнилкарбамат, изобутилкарбамат, изоникотинилкарбамат, п-(п'-метоксифенилазо)бензилкарбамат, 1-метилциклобутилкарбамат, 1-метилциклогексилкарбамат, 1-метил-1-циклопропилметилкарбамат, 1-метил-1-(3,5-диметоксифенил)этилкарбамат, 1-метил-1-(п-фенилазофенил)этилкарбамат, 1-метил-1-фенилэтилкарбамат, 1-метил-1-(4-пиридил)этилкарбамат, фенилкарбамат, п-(фенилазо)бензилкарбамат, 2,4,6-три-трет-бутилфенилкарбамат, 4-(триметиламмоний) бензилкарбамат, 2,4,6-триметилбензилкарбамат, формамид, ацетамид, хлорацетамид, трихлорацетамид, трифторацетамид, фенилацетамид, 3-фенилпропанамид, пиколинамид, 3-пиридилкарбоксамид, N-бензоилфенилаланин производное, бензамид, п-фенилбензамид, о-нитрофенилацетамид, о-нитрофеноксиацетамид, ацетоацетамид, (N'-дитиобензилоксикарбониламино)ацетамид, 3-(п-гидроксифенил)пропанамид, 3-(о-нитрофенил)пропанамид, 2-метил-2-(о-нитрофенокси)пропанамид, 2-метил-2-(о-фенилазофенокси)пропанамид, 4-хлорбутанамид, 3-метил-3-нитробутанамид, о-нитроциннамид, N-ацетилметионин производное, о-нитробензамид, о-(бензоилоксиметил) бензамид, 4,5-дифенил-3-оксазолин-2-он, N-фталимид, N-дитиасукцинамид (Dts), N-2,3-дифенилмалеимид, N-2,5-диметилпиррол, N-1,1,4,4-тетраметилдисилилазациклопентановый аддукт (STABASE), 5-замещенный 1,3-диметил-1,3,5-триазациклогексан-2-он, 5-замещенный 1,3-дибензил-1,3,5-триазациклогексан-2-он, 1-замещенный 3,5-динитро-4-пиридон, N-метиламин, N-аллиламин, N[2-(триметилсилил)этокси] метиламин (SEM), N-3-ацетоксипропиламин, N-(1-изопропил-4-нитро-2-оксо-3-пирролин-3-ил) амин, соли четвертичного аммония, N-бензиламин, N-ди(4-метоксифенил)метиламин, N-5-дибензосубериламин, N-трифенилметиламин (Tr), N-[(4-метоксифенил)дифенилметил]амин (MMTr), N-9-фенилфлуорениламин (PhF), N-2,7-дихлор-9-флуоренилметиленамин, N-ферроценилметиламино (Fcm), N-2-пиколиламино N'-оксид, N-1,1-диметилтиометиленамин, N-бензилиденамин, N-п-метоксибензилиденамин, N-дифенилметиленамин, N-[(2-пиридил)мезитил] метиленамин, N-(N',N'-диметиламинометилен)амин, N,N'-изопропилидендиамин, N-п-нитробензилиденамин, N-салицилиденамин, N-5-хлорсалицилиденамин, N-(5-хлор-2-гидроксифенил) фенилметиленамин, N-циклогексилиденамин, N-(5,5-диметил-3-оксо-1-циклогексенил) амин, N-боран производное, N-дифенилбориновой кислоты производное, N-[фенил (пентакарбонилхрома или вольфрама)карбонил]амин, N-медь хелат, N-цинк хелат, N-нитроамин, N-нитрозоамин, амин N-оксид, дифенилфосфинамид (Dpp), диметилтиофосфинамид (Mpt), дифенилтиофосфинамид (Ppt), диалкилфосфорамидаты, дибензил фосфорамидат, дифенил фосфорамидат, бензолсульфенамид, о-нитробезолсульфенамид (Nps), 2,4-динитробензолсульфенамид, пентахлорбензолсульфенамид, 2-нитро-4-метоксибензолсульфенамид, трифенилметилсульфенамид, 3-нитропиридинсульфенамид (Npys), п-толуолсульфонамид (Ts), бензолсульфонамид, 2,3,6,-триметил-4-метоксибензолсульфонамид (Mtr), 2,4,6-триметоксибензолсульфонамид (Mtb), 2,6-диметил-4-метоксибензолсульфонамид (Pme), 2,3,5,6-тетраметил-4-метоксибензолсульфонамид (Mte), 4-метоксибензолсульфонамид (Mbs), 2,4,6-триметилбензолсульфонамид (Mts), 2,6-диметокси-4-метилбензолсульфонамид (iMds), 2,2,5,7,8-пентаметилхроман-6-сульфонамид (Pmc), метансульфонамид (Ms), β-триметилсилилэтансульфонамид (SES), 9-антраценсульфонамид, 4-(4',8'-диметоксинафтилметил)бензолсульфонамид (DNMBS), бензилсульфонамид, трифторметилсульфонамид, и фенацилсульфонамид. Иллюстративные защитные группы подробно описаны в данной заявке. Однако, следует понимать, что настоящее изобретение не ограничивается этими защитными группами, а, скорее, множество дополнительных эквивалентных защитных групп может быть легко идентифицировано с использованием вышеуказанных критериев и использованы в способе в соответствии с настоящим изобретением. Дополнительно, множество защитных групп описано в Protective Groups in Organic Synthesis, Third Ed. Greene, T.W. and Wuts, P.G., Eds., John Wiley & Sons, New York: 1999, полное содержание которой включено в данную заявку путем ссылки.
Следует иметь в виду, что соединения, описанные в данной заявке, могут быть замещены любым количеством заместителей или функциональных фрагментов. В общем, термин «замещенный», предшествует ли ему термин «необязательно» или нет, и заместители, содержащиеся в формуле настоящего изобретения, относятся к замене водородных радикалов в данной структуре на радикал указанного заместителя. Когда более чем одно положение в любой данной структуре может быть замещено более чем одним заместителем, выбранным из указанной группы, заместитель может быть либо одинаковым, или разным в каждом положении. Как используется в данной заявке, термин «замещенный» предполагается для включения всех допустимых заместителей органических соединений. В широком аспекте, допустимые заместители включают ациклические и циклические, разветвленные и неразветвленные, карбоциклические и гетероциклические, ароматические и неароматические заместители органических соединений. Гетероатомы, такие, как азот, могут иметь водородные заместители и/или любые допустимые заместители органических соединений, описанные в данной заявке, которые удовлетворяют валентности гетероатомов. Дополнительно, настоящее изобретение не должно ограничиваться каким-либо образом допустимыми заместителями органических соединений. Комбинации заместителей и переменные, предусмотренные настоящим изобретением, предпочтительно являются теми, которые приводят к образованию стабильных соединений, полезных для лечения, например, инфекционных заболеваний или пролиферативных расстройств. Термин «стабильный», используемый в данной заявке, предпочтительно относится к соединениям, которые обладают стабильностью, достаточной для получения, и которые поддерживают целостность соединения в течение достаточного периода времени, чтобы быть обнаруженными и предпочтительно в течение достаточного периода времени, чтобы быть полезными для целей, изложенных в данной заявке.
Термин «алифатический», как используется в данной заявке, включает как насыщенные, так и ненасыщенные, неразветвленные, разветвленные, ациклические, циклические или полициклические алифатические углеводороды, которые необязательно замещены одной или несколькими функциональными группами. Как будет понятно специалисту в данной области техники, «алифатический» предназначен в данной заявке для включения, но не ограничиваясь приведенным, алкильных, алкенильных, алкинильных, циклоалкильных, циклоалкенильных и циклоалкинильных фрагментов. Таким образом, как используется в данной заявке, термин «алкил» включает неразветвленные, разветвленные и циклические алкильные группы. Аналогичное соглашение относится и к другим общим терминам, таким как «алкенил», «алкинил», и тому подобное. Дополнительно, как используется в данной заявке, термины «алкил», «алкенил», «алкинил» и т.п. охватывают как замещенные, так и незамещенные группы. В определенных осуществлениях, как используется в данной заявке, «низший алкил» используется для обозначения алкильных групп (циклических, ациклических, замещенных, незамещенных, разветвленных или неразветвленных), имеющих 1-6 атомов углерода.
В определенных осуществлениях, алкильные, алкенильные, и алкинильные группы, задействованные в настоящем изобретении, содержат 1-20 алифатических атомов углерода. В некоторых других осуществлениях алкильные, алкенильные и алкинильные группы, задействованные в настоящем изобретении, содержат 1-10 алифатических атомов углерода. В других осуществлениях алкильные, алкенильные и алкинильные группы, задействованные в настоящем изобретении, содержат 1-8 алифатических атомов углерода. В других осуществлениях алкильные, алкенильные и алкинильные группы, задействованные в настоящем изобретении, содержат 1-6 алифатических атомов углерода. В других осуществлениях алкильные, алкенильные и алкинильные группы, задействованные в настоящем изобретении, содержат 1-4 атомов углерода. Примеры алифатических групп, таким образом, включают, но не ограничиваясь приведенным, например, метил, этил, п-пропил, изопропил, циклопропил, -СН2-циклопропил, винил, аллил, н-бутил, втор-бутил, изобутил, трет-бутил, циклобутил, -СН2-циклобутил, н-пентил, втор-пентил, изопентил, трет-пентил, циклопентил, -СН2-циклопентил, н-гексил, втор-гексил, циклогексил, -СН2-циклогексил фрагменты и т.п., которые опять таки, могут нести один или нескольких заместителей. Алкенильные группы включают, но не ограничиваясь приведенным, например, этенил, пропенил, бутенил, 1-метил-2-бутен-1-ил и тому подобное. Иллюстративные алкинильные группы включают, но не ограничиваясь приведенным, этинил, 2-пропинил (пропаргил), 1-пропинил и тому подобное.
Некоторые примеры заместителей описанных выше алифатических (и других) фрагментов соединений согласно настоящему изобретению, включают, но не ограничиваясь приведенным, алифатические; гетероалифатические; арил; гетероарил; арилалкил, гетероарилалкил; алкокси, арилокси; гетероалкокси; гетероарилокси; алкилтио; арилтио, гетероалкилтио; гетероарилтио; -F; -Cl; -Br; -I; -ОН; -NO2; -CN; -CF3; -CH2CF3; -CHCl2; -СН2ОН; -СН2СН2ОН; -CH2NH2; -CH2SO2CH3; -C(O)Rx; -CO2(Rx); -CON(Rx)2, -OC(O)Rx; -OCO2Rx; -OCON(Rx)2, -N(Rx)2, -S(O)2Rx; -NRx(CO)Rx, где каждый независимо включает, но не ограничиваясь приведенным, алифатический, гетероалифатический, арил, гетероарил, арилалкил или гетероарилалкил, где любой из алифатических, гетероалифатических, арилалкильных или гетероарилалкильных заместителей, описанных выше в данной заявке, может быть замещенным или незамещенным, разветвленным или неразветвленным, циклическим или ациклическим, и где любой из арильных или гетероарильных заместителей, описанных выше в данной заявке, может быть замещенным или незамещенным. Дополнительные примеры общеприменимых заместителей проиллюстрированы конкретными осуществлениями, описанными в данной заявке.
Термин «гетероалифатический», используемый в данной заявке, относится к алифатическим фрагментам, которые содержат один или более атомов кислорода, серы, азота, фосфора или атомов кремния, например, вместо атомов углерода. Гетероалифатические фрагменты могут быть разветвленными, неразветвленными, циклическими или ациклическими и включают насыщенные и ненасыщенные гетероциклы, такие как морфолино, пирролидинил и т.д. В определенных осуществлениях, гетероалифатические фрагменты замещены путем независимой замены одного или нескольких атомов водорода одним или несколькими фрагментами, включая, но не ограничиваясь приведенным, алифатический; гетероалифатический; арил; гетероарил; арилалкил; гетероарилалкил; алкокси; арилокси; гетероалкокси; гетероарилокси; алкилтио; арилтио; гетероалкилтио; гетероарилтио, -F; -Cl; -Br; -I; -ОН; -NO2; -CN; -CF3; -CH2CF3; -CHCl2; -СН2ОН; -СН2СН2ОН; -CH2NH2; -CH2SO2CH3; -C(O)Rx; -CO2(Rx); -CON(Rx)2, -OC(O)Rx; -OCO2Rx; -OCON(Rx)2, -N(Rx)2, -S(O)2Rx; -NRx(CO)Rx, где каждый независимо включает, но не ограничиваясь приведенным, алифатический, гетероалифатический, арил, гетероарил, арилалкил или гетероарилалкил, где любой из алифатических, гетероалифатических, арилалкильных или гетероарилалкильных заместителей, описанных выше в данной заявке, может быть замещенным или незамещенным, разветвленным или неразветвленным, циклическим или ациклическим, и где любой из арильных или гетероарильных заместителей, описанных выше в данной заявке, может быть замещенным или незамещенным. Дополнительные примеры общеприменимых заместителей проиллюстрированы конкретными осуществлениями, описанными в данной заявке.
Термины «гало» и «галоген», используемые в данной заявке, относятся к атому, выбранному из фтора, хлора, брома и йода.
Термин «алкил» включает насыщенные алифатические группы, включая неразветвленные алкильные группы (например, метил, этил, пропил, бутил, пентил, гексил, гептил, октил, нонил, децил и т.д.), разветвленные алкильные группы (изопропил, трет-бутил, изобутил и т.д.), циклоалкильные (алициклические) группы (циклопропил, циклопентил, циклогексил, циклогептил, циклооктил), алкил замещенные циклоалкильные группы и циклоалкил замещенные алкильные группы. В определенных осуществлениях, неразветвленная или разветвленная алкильная цепь имеет 6 или менее атомов углерода в каркасе (например, C1-С6 для неразветвленной цепи, С3-С6 для разветвленной цепи), и более предпочтительно 4 или менее. Дополнительно, предпочтительные циклоалкилы имеют 3-8 атомов углерода в кольцевой структуре, а более предпочтительно имеют 5 или 6 атомов углерода в кольцевой структуре. Термин C1-С6 включает алкильные группы, содержащие от 1 до 6 атомов углерода.
Дополнительно, если не указано иное, термин алкил включает «незамещенные алкилы» и «замещенные алкилы», последние из которых относятся к алкильным фрагментам, имеющим независимо выбранные заместители, замещающие водород одним или более атомами углерода в углеводородном каркасе. Такие заместители могут включать, например, алкенил, алкинил, галоген, гидроксил, алкилкарбонилокси, арилкарбонилокси, алкоксикарбонилокси, арилоксикарбонилокси, карбоксилат, алкилкарбонил, арилкарбонил, алкоксикарбонил, аминокарбонил, алкиламинокарбонил, диалкиламинокарбонил, алкилтиокарбонил, алкоксил, фосфат, фосфонато, фосфинато, циано, амино (включая алкиламино, диалкиламино, ариламино, диариламино и алкилариламино), ациламино (включая алкилкарбониламино, арилкарбониламино, карбамоил и уреидо), амидино, имино, сульфгидрил, алкилтио, арилтио, тиокарбоксилат, сульфаты, алкилсульфинил, сульфонато, сульфамоил, сульфонамидо, нитро, трифторметил, циано, азидо, гетероциклил, алкиларил или ароматический или гетероароматический фрагмент. Циклоалкилы могут быть замещены, например, заместителями, описанными выше. «Алкиларильный» или «арилалкильный» фрагмент представляет алкил, замещенный арилом (например, фенилметил (бензил)). Термин «алкил» включает также боковые цепи природных и неприродных аминокислот. Термин «n-алкил» означает незамещенную алкильную группу с неразветвленной цепью.
Термин «алкенил» включает ненасыщенные алифатические группы, аналогичные по длине и возможно замещенные на алкилы, описанные выше, но которые содержат по меньшей мере одну двойную связь. Например, термин «алкенил» включает неразветвленные алкенильные группы (например, этиленил, пропенил, бутенил, пентенил, гексенил, гептенил, октенил, ноненил, деценил и т.д.), разветвленные алкенильные группы, циклоалкенильные (алициклические) группы (циклопропенил, циклопентенил, циклогексенил, циклогептенил, циклооктенил), алкильные или алкенильные замещенные циклоалкенильные группы и циклоалкильные или циклоалкенильные замещенные алкенильные группы. В определенных осуществлениях, неразветвленная или разветвленная алкенильная группа имеет 6 или менее атомов углерода в каркасе (например, С2-C6 для неразветвленной цепи, С3-C6 для разветвленной цепи). Дополнительно, циклоалкенильные группы могут иметь 3-8 атомов углерода в кольцевой структуре, а более предпочтительно имеют 5 или 6 атомов углерода в кольцевой структуре. Термин С2-С6 включает алкенильные группы, содержащие от 2 до 6 атомов углерода.
Дополнительно, если не указано иное, термин алкенил включает «незамещенные алкенилы» и «замещенные алкенилы», последний из которых относится к алкенильным фрагментам, имеющим независимо выбранные заместители, замещающие водород на один или более атомов углерода в углеводородном каркасе. Такие заместители могут включать, например, алкильные группы, алкинильные группы, галогены, гидроксил, алкилкарбонилокси, арилкарбонилокси, алкоксикарбонилокси, арилоксикарбонилокси, карбоксилат, алкилкарбонил, арилкарбонил, алкоксикарбонил, аминокарбонил, алкиламинокарбонил, диалкиламинокарбонил, алкилтиокарбонил, алкоксил, фосфат, фосфонато, фосфинато, циано, амино (включая алкиламино, диалкиламино, ариламино, диариламино и алкилариламино), ациламино (включая алкилкарбониламино, арилкарбониламино, карбамоил и уреидо), амидино, имино, сульфгидрил, алкилтио, арилтио, тиокарбоксилат, сульфаты, алкилсульфинил, сульфонато, сульфамоил, сульфонамидо, нитро, трифторметил, циано, азидо, гетероциклил, алкиларил или ароматический или гетероароматический фрагмент.
Термин «алкинил» включает ненасыщенные алифатические группы, аналогичные по длине и возможно замещенные на алкилы, описанные выше, но которые содержат по меньшей мере одну тройную связь. Например, термин «алкинил» включает неразветвленные алкинильные группы (например, этинил, пропинил, бутинил, пентинил, гексинил, гептинил, октинил, нонинил, деценил и т.д.), разветвленные алкинильные группы, и циклоалкильные или циклоалкенильные замещенные алкинильные группы. В определенных осуществлениях, неразветвленная или разветвленная алкинильная группа имеет 6 или менее атомов углерода в каркасе (например, С2-С6 для неразветвленной цепи, С3-С6 для разветвленной цепи). Термин С2-С6 включает алкинильные группы, содержащие от 2 до 6 атомов углерода.
Дополнительно, если не указано иное, термин алкинил включает «незамещенные алкинилы» и «замещенные алкинилы», последний из которых относится к алкинильным фрагментам, имеющим независимо выбранные заместители, замещающие водород на один или более атомов углерода в углеводородном каркасе. Такие заместители могут включать, например, алкильные группы, алкинильные группы, галогены, гидроксил, алкилкарбонилокси, арилкарбонилокси, алкоксикарбонилокси, арилоксикарбонилокси, карбоксилат, алкилкарбонил, арилкарбонил, алкоксикарбонил, аминокарбонил, алкиламинокарбонил, диалкиламинокарбонил, алкилтиокарбонил, алкоксил, фосфат, фосфонато, фосфинато, циано, амино (включая алкиламино, диалкиламино, ариламино, диариламино и алкилариамино), ациламино (включая алкилкарбониламино, арилкарбониламино, карбамоил и уреидо), амидино, имино, сульфгидрил, алкилтио, арилтио, тиокарбоксилат, сульфаты, алкилсульфинил, сульфонато, сульфамоил, сульфонамидо, нитро, трифторметил, циано, азидо, гетероциклил, алкиларил или ароматический или гетероароматический фрагмент.
Если количество атомов углерода не указано иным образом, «низший алкил», используемый в данной заявке, означает алкильную группу, как определено выше, но имеющую от одного до пяти атомов углерода в ее каркасной структуре. «Низший алкенил» и «низший алкинил» имеют цепь длиной, например, 2-5 атомов углерода.
Термин «алкокси» включает замещенные и незамещенные алкильные, алкенильные и алкинильные группы, ковалентно связанные с атомом кислорода. Примеры алкоксигрупп включают метокси, этокси, изопропилокси, пропокси, бутокси и пентокси группы. Примеры замещенных алкоксигрупп включают галогенированные алкоксигруппы. Алкоксигруппы могут быть замещены независимо выбранными группами, такими, как алкенил, алкинил, галоген, гидроксил, алкилкарбонилокси, арилкарбонилокси, алкоксикарбонилокси, арилоксикарбонилокси, карбоксилат, алкилкарбонил, арилкарбонил, алкоксикарбонил, аминокарбонил, алкиламинокарбонил, диалкиламинокарбонил, алкилтиокарбонил, алкоксил, фосфат, фосфонато, фосфинато, циано, амино (включая алкиламино, диалкиламино, ариламино, диариламино и алкилариламино), ациламино (включая алкилкарбониламино, арилкарбониламино, карбамоил и уреидо), амидино, имино, сульфгидрил, алкилтио, арилтио, тиокарбоксилат, сульфаты, алкилсульфинил, сульфонато, сульфамоил, сульфонамидо, нитро, трифторметил, циано, азидо, гетероциклил, алкиларил или ароматические или гетероароматические фрагменты. Примеры галогенных замещенных алкокси групп включают, но не ограничиваясь приведенным, фторметокси, дифторметокси, трифторметокси, хлорметокси, дихлорметокси, трихлорметокси и т.д.
Термин «гетероатом» включает атомы любого элемента, кроме углерода и водорода. Предпочтительными гетероатомами являются азот, кислород, сера и фосфор.
Термин «гидрокси» или «гидроксил» включает группы с -OH или –O– (с соответствующим противоионом).
Термин «галоген» включает фтор, бром, хлор, йод и др. Термин «пергалогенированный» обычно относится к фрагменту, в котором все атомы водорода заменены на атомы галогенов.
Термин «замещенный» включает независимо выбранные заместители, которые могут быть размещены во фрагменте и которые позволяют молекуле выполнять свои функции. Примеры заместителей включают алкильную, алкенильную, алкинильную, арильную, (CR'R'')0-3NR'R'', (CR'R'')0-3CN, NO2, галоген, (CR'R'')0-3C(галоген)3, (CR'R'')0-3CH(галоген)2, (CR'R'')0-3CH2(галоген), (CR'R'')0-3CONR'R'', (CR'R''0-3S(O)1-2NR'R'', (CR'R'')0-3CHO, (CR'R'')0-3О(CR'R'')0-3H, (CR'R'')0-3S(О)0-2R', (CR'R'')0-3О(CR'R'')0-3H, (CR'R'')0-3COR', (CR'R'')0-3CO2R', или (CR'R'')0-3OR' группы, где каждый R' и R'' независимо представляет собой водород, С1-С5 алкильную, C2-C5 алкенильную, C2-C5 алкинильную или арильную группу, или R 'и R'' вместе взятые, являются бензилиденовой группой или -(CH2)2O(CH2)2- группой.
Термин «амин» или «амино» включает соединения, или фрагменты, в которых атом азота ковалентно связан с по меньшей мере одним атомом углерода или гетероатомом. Термин «алкиламино» включает группы и соединения, в которых азот связан с по меньшей мере одной дополнительной алкильной группой. Термин «диалкиламино» включает группы, в которых атом азота связан с по меньшей мере двумя дополнительными алкильными группами.
Термин «простой эфир» включает соединения, или фрагменты, которые содержат кислород, связанный с двумя различными атомами углерода или гетероатомами. Например, термин включает «алкоксиалкил», который относится к алкильной, алкенильной или алкинильной группе, ковалентно связанной с атомом кислорода, который ковалентно связан с другой алкильной группой.
Термины «полинуклеотид», «нуклеотидная последовательность», «нуклеиновая кислота», «молекула нуклеиновой кислоты», «последовательность нуклеиновой кислоты» и «олигонуклеотид» относятся к полимеру из двух или более нуклеотидов. Полинуклеотидами могут быть ДНК, РНК, или производные или их модифицированные версии. Полинуклеотид может быть одноцепочечным или двухцепочечным. Полинуклеотид может быть модифицирован во фрагменте основания, фрагменте сахара или каркасе фосфата, например, для улучшения стабильности молекулы, ее параметров гибридизации и т.д. Полинуклеотид может содержать модифицированный фрагмент основания, который выбран из группы, включая, но не ограничиваясь приведенным, 5-фторурацил, 5-бромурацил, 5-хлорурацил, 5-йодурацил, гипоксантин, ксантин, 4-ацетилцитозин, 5-(карбоксигидроксиметил) урацил, 5-карбоксиметиламинометил-2-тиоуридин, 5-карбоксиметиламинометилурацил, дигидроурацил, бета-D- галактозинквеозин, инозин, N6-изопентиладенин, 1-метилгуанин, 1-метилинозин, 2,2-диметилгуанин, 2-метиладенин, 2-метилгуанин, 3-метилцитозин, 5-метилцитозин, N6-аденин, 7-метилгуанин, 5-метиламинометилурацил, 5-метоксиаминометил-2-тиоурацил, бета-D-маннозилквеозин, 5'-метоксикарбоксиметилурацил, 5-метоксиурацил, 2-метилтио-N6-изопентениладенин, вибутоксозин, псевдоурацил, квеозин, 2-тиоцитозин, 5-метил-2-тиоурацил, 2-тиоурацил, 4-тиоурацил, 5-метилурацил, урацил-5-оксиуксусной кислоты метиловый эфир, урацил-5-оксиуксусную кислоту, 5-метил-2-тиоурацил, 3-(3-амино-3-N-2-карбоксипропил) урацил и 2,6-диаминопурин. Олигонуклеотид может содержать модифицированный сахарный фрагмент (например, 2'-фторрибозу, рибозу, 2'-дезоксирибозу, 2'-O-метилцитидин, арабинозу и гексозу), и/или модифицированный фосфатный фрагмент (например, фосфоротиоаты и 5'-N-фосфорамидитные связи). Нуклеотидная последовательность обычно несет генетическую информацию, включая информацию, используемую клеточным механизмом, чтобы получить белки и ферменты. Эти термины включают двух- или одноцепочечные геномные и кДНК, РНК, любые синтетические и генетически модифицированные полинуклеотиды, и как смысловые, так и антисмысловые полинуклеотиды. Они включают одно- и двухцепочечные молекулы, например, ДНК-ДНК, ДНК-РНК и РНК-РНК гибриды, а также «белки нуклеиновых кислот» (PNA), образованные сопряжением оснований амино кислотного каркаса.
Термин «основание» включает известные пуриновые и пиримидиновые гетероциклические основания, деазапурины и аналоги (включая гетероциклические замещенные аналоги, например, аминоэтиокси феноксазин), производные (например, 1-алкил-, 1-алкенил-, гетероароматические и 1-алкинил производные) и их таутомеры. Примеры пуринов включают аденин, гуанин, инозин, диаминопурин, и ксантин и их аналоги (например, 8-оксо-N6-метиладенин или 7-диазаксантин) и их производные. Пиримидины включают, например, тимин, урацил и цитозин и их аналоги (например, 5-метилцитозин, 5-метилурацил, 5-(1-пропинил)урацил, 5-(1-пропинил)цитозин и 4,4-этаноцитозин). Другие примеры приемлемых оснований включают не пуринильные и не пиримидинильные основания, такие как 2-аминопиридин и триазины.
В предпочтительном осуществлении, нуклеомономеры олигонуклеотида в соответствии с настоящим изобретением являются РНК нуклеотидами. В другом предпочтительном осуществлении, нуклеомономеры олигонуклеотида в соответствии с настоящим изобретением являются модифицированными РНК нуклеотидами. Таким образом, олигонуклеотиды содержат модифицированные РНК нуклеотиды.
Термин «нуклеозид» включает основания, которые ковалентно связаны с сахарным фрагментом, предпочтительно рибозой или дезоксирибозой. Примеры предпочтительных нуклеозидов включают рибонуклеозиды и дезоксирибонуклеозиды. Нуклеозиды также включают основания, связанные с аминокислотами или аналогами аминокислот, которые могут содержать свободные карбоксильные группы, свободные аминогруппы или защитные группы. Приемлемые защитные группы хорошо известны в данной области техники (см. P. G. M. Wuts and T. W. Greene, «Protective Groups in Organic Synthesis», 2nd Ed., Wiley-Interscience, New York, 1999).
Термин «нуклеотид» включает нуклеозиды, которые дополнительно содержат фосфатную группу или фосфатный аналог.
Молекулы нуклеиновой кислоты могут быть связаны с гидрофобным фрагментом для направления и/или доставки молекул в клетку. В определенных осуществлениях, гидрофобный фрагмент связан с молекулой нуклеиновой кислоты через линкер. В определенных осуществлениях, связывание происходит через нековалентные взаимодействия. В других осуществлениях, связывание происходит через ковалентную связь. Любой линкер, известный в данной области техники, может быть использован для связывания нуклеиновой кислоты с гидрофобным фрагментом. Линкеры, известные в данной области техники, описаны в опубликованных международных заявках по процедуре РСТ, WO 92/03464, WO 95/23162, WO 2008/021157, WO 2009/021157, WO 2009/134487, WO 2009/126933, публикации патентной заявки США 2005/0107325, патенте США 5,414,077, патенте США 5,419,966, патенте США 5,512,667, патенте США 5,646,126 и патенте США 5,652,359, которые включены в данную заявку путем ссылки. Линкер может быть также простым, как ковалентная связь, в многоатомных линкерах. Линкер может быть циклическим или ациклическим. Линкер может быть необязательно замещен. В определенных осуществлениях, линкер способен быть отщеплен от нуклеиновой кислоты. В определенных осуществлениях, линкер способен быть гидролизован в физиологических условиях. В определенных осуществлениях, линкер способен быть расщеплен ферментом (например, эстеразой или фосфодиэстеразой). В некоторых осуществлениях, линкер включает спейсерный элемент для разделения нуклеиновой кислоты и гидрофобного фрагмента. Спейсерный элемент может включать от одного до тридцати атомов углерода или гетероатомов. В определенных осуществлениях, линкер и/или спейсерный элемент содержит протонируемые функциональные группы. Такие протонируемые функциональные группы могут способствовать эндосомальному высвобождению молекулы нуклеиновой кислоты. Протонируемые функциональные группы могут также способствовать доставке нуклеиновой кислоты в клетку, например, нейтрализуя общий заряд молекулы. В других осуществлениях, линкер и/или спейсерный элемент является биологически инертным (то есть, не придает биологической активности или функции полученной в результате молекуле нуклеиновой кислоты).
В определенных осуществлениях, молекула нуклеиновой кислоты с линкером и гидрофобным фрагментом имеет формулу, описанную в данной заявке. В определенных осуществлениях, молекула нуклеиновой кислоты имеет формулу:

где
X представляет собой N или CH;
А является связью; замещенным или незамещенным, циклическим или ациклическим, разветвленным или неразветвленным алифатическим фрагментом;
или замещенным или незамещенным, циклическим или ациклическим, разветвленным или неразветвленным гетероалифатическим фрагментом;
R1 представляет собой гидрофобный фрагмент;
R2 представляет собой водород; кислород-защитную группу; циклический или ациклический, замещенный или незамещенный, разветвленный или неразветвленный алифатический фрагмент; циклический или ациклический, замещенный или незамещенный, разветвленный или неразветвленный гетероалифатический фрагмент; замещенный или незамещенный, разветвленный или неразветвленный ацил; замещенный или незамещенный, разветвленный или неразветвленный арил, замещенный или незамещенный, разветвленный или неразветвленный гетероарил, и
R3 представляет собой нуклеиновую кислоту.
В определенных осуществлениях, молекула имеет формулу:
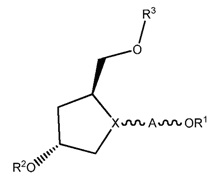
В определенных осуществлениях, молекула имеет формулу:

В определенных осуществлениях, молекула имеет формулу:

В определенных осуществлениях, молекула имеет формулу:

В определенных осуществлениях, X представляет собой N. В определенных осуществлениях, X представляет собой CH.
В определенных осуществлениях, А представляет собой связь. В определенных осуществлениях, A является замещенным или незамещенным, циклическим или ациклическим, разветвленным или неразветвленным алифатическим фрагментом. В определенных осуществлениях, А является ациклическим, замещенным или незамещенным, разветвленным или неразветвленным алифатическим фрагментом. В определенных осуществлениях, A является ациклическим, замещенным, разветвленным или неразветвленным алифатическим фрагментом. В определенных осуществлениях, A является ациклическим, замещенным, неразветвленным алифатическим фрагментом. В определенных осуществлениях, A является ациклическим, замещенным, неразветвленным алкилом. В определенных осуществлениях, A является ациклическим, замещенным, неразветвленным С1-20 алкилом. В определенных осуществлениях, A является ациклическим, замещенным, неразветвленным C1-12 алкилом. В определенных осуществлениях, A является ациклическим, замещенным, неразветвленным C1-10 алкилом. В определенных осуществлениях, A является ациклическим, замещенным, неразветвленным C1-8 алкилом. В определенных осуществлениях, A является ациклическим, замещенным, неразветвленным C1-6 алкилом. В определенных осуществлениях, A является замещенным или незамещенным, циклическим или ациклическим, разветвленным или неразветвленным гетероалифатическим фрагментом. В определенных осуществлениях, A является ациклическим, замещенным или незамещенным, разветвленным или неразветвленным гетероалифатическим фрагментом. В определенных осуществлениях, A является ациклическим, замещенным, разветвленным или неразветвленным гетероалифатическим фрагментом. В определенных осуществлениях, A является ациклическим, замещенным, неразветвленным гетероалифатическим фрагментом.
В определенных осуществлениях, A имеет формулу:
В определенных осуществлениях, A имеет формулу:
 .
.
В определенных осуществлениях, A имеет формулу:
 .
.
В определенных осуществлениях, A имеет формулу:
 .
.
В определенных осуществлениях, A имеет формулу:
 .
.
В определенных осуществлениях, A имеет формулу:
 .
.
В определенных осуществлениях, A имеет формулу:
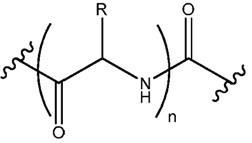
где каждый из R независимо представляет собой боковую цепь природной или неприродной аминокислоты; и
n представляет собой целое число от 1 до 20, включительно. В определенных осуществлениях, A имеет формулу:
 .
.
В определенных осуществлениях, каждый из R независимо представляет собой боковую цепь природной аминокислоты. В определенных осуществлениях, n представляет собой целое число от 1 до 15, включительно. В определенных осуществлениях, n представляет собой целое число от 1 до 10, включительно. В определенных осуществлениях, n представляет собой целое число от 1 до 5, включительно.
В определенных осуществлениях, A имеет формулу:
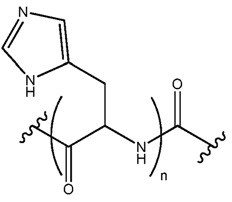
где n представляет собой целое число от 1 до 20, включительно. В определенных осуществлениях, A имеет формулу:
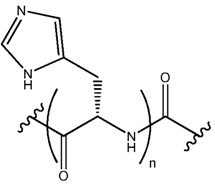 .
.
В определенных осуществлениях, n представляет собой целое число от 1 до 15, включительно. В определенных осуществлениях, n представляет собой целое число от 1 до 10, включительно. В определенных осуществлениях, n представляет собой целое число от 1 до 5, включительно.
В определенных осуществлениях, A имеет формулу:

где n представляет собой целое число от 1 до 20, включительно. В определенных осуществлениях, A имеет формулу:
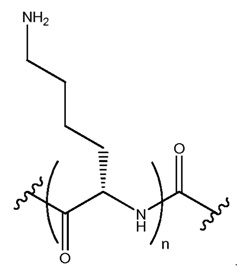
В определенных осуществлениях, n представляет собой целое число от 1 до 15, включительно. В определенных осуществлениях, n представляет собой целое число от 1 до 10, включительно. В определенных осуществлениях, n представляет собой целое число от 1 до 5, включительно.
В определенных осуществлениях, молекула имеет формулу:
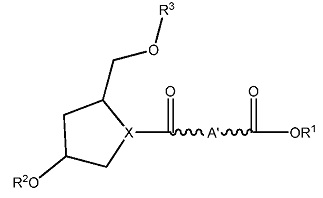
где X, R1, R2 и R3 являются такими, как определено в данной заявке; и
A' является замещенным или незамещенным, циклическим или ациклическим, разветвленным или неразветвленным алифатическим фрагментом; или замещенным или незамещенным, циклическим или ациклическим, разветвленным или неразветвленным гетероалифатическим фрагментом.
В определенных осуществлениях, A' имеет одну из формул:
 .
.
В определенных осуществлениях, A имеет одну из формул:
 .
.
В определенных осуществлениях, A имеет одну из формул:
 .
.
В определенных осуществлениях, A имеет формулу:
 .
.
В определенных осуществлениях, A имеет формулу:
 .
.
В определенных осуществлениях, R1 является стероидом. В определенных осуществлениях, R1 является холестерином. В определенных осуществлениях, R1 является липофильным витамином. В определенных осуществлениях, R1 является витамином A. В определенных осуществлениях, R1 является витамином E.
В определенных осуществлениях, Rl имеет формулу:
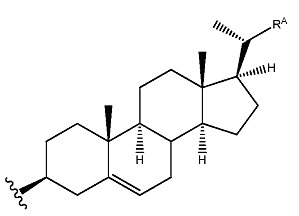 .
.
Где RA является замещенным или незамещенным, циклическим или ациклическим, разветвленным или неразветвленным алифатическим фрагментом; или замещенным или незамещенным, циклическим или ациклическим, разветвленным или неразветвленным гетероалифатическим фрагментом.
В определенных осуществлениях, R1 имеет формулу:

В определенных осуществлениях, R1 имеет формулу:
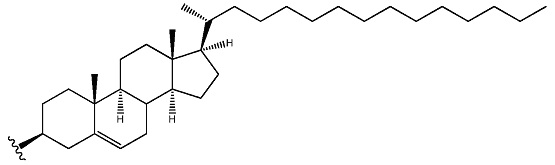
В определенных осуществлениях, R1 имеет формулу:

В определенных осуществлениях, R1 имеет формулу:
 .
.
В определенных осуществлениях, R1 имеет формулу:
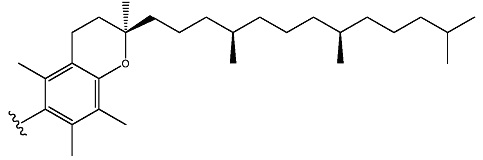 .
.
В определенных осуществлениях, молекула нуклеиновой кислоты имеет формулу:
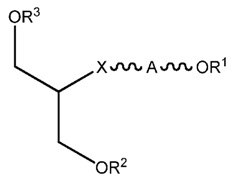
где
X представляет собой N или CH;
A является связью; замещенным или незамещенным, циклическим или ациклическим, разветвленным или неразветвленным алифатическим фрагментом; или замещенным или незамещенным, циклическим или ациклическим, разветвленным или неразветвленным гетероалифатическим фрагментом;
R1 представляет собой гидрофобный фрагмент;
R2 является водородом; кислород-защитной группой; циклическим или ациклическим, замещенным или незамещенным, разветвленным или неразветвленным алифатическим фрагментом; циклическим или ациклическим, замещенным или незамещенным, разветвленным или неразветвленным гетероалифатическим фрагментом; замещенным или незамещенным, разветвленным или неразветвленным ацилом; замещенным или незамещенным, разветвленным или неразветвленным арилом; замещенным или незамещенным, разветвленным или неразветвленным гетероарилом; и
R3 представляет собой нуклеиновую кислоту.
В определенных осуществлениях, молекула нуклеиновой кислоты имеет формулу:
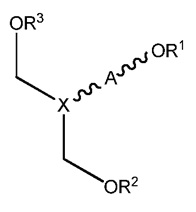
где
X представляет собой N или CH;
A является связью; замещенным или незамещенным, циклическим или ациклическим, разветвленным или неразветвленным алифатическим фрагментом; или замещенным или незамещенным, циклическим или ациклическим, разветвленным или неразветвленным гетероалифатическим фрагментом;
R1 представляет собой гидрофобный фрагмент;
R2 является водородом; кислород-защитной группой; циклическим или ациклическим, замещенным или незамещенным, разветвленным или неразветвленным алифатическим фрагментом; циклическим или ациклическим, замещенным или незамещенным, разветвленным или неразветвленным гетероалифатическим фрагментом; замещенным или незамещенным, разветвленным или неразветвленным ацилом; замещенным или незамещенным, разветвленным или неразветвленным арилом; замещенным или незамещенным, разветвленным или неразветвленным гетероарилом; и
R3 представляет собой нуклеиновую кислоту.
В определенных осуществлениях, молекула нуклеиновой кислоты имеет формулу:
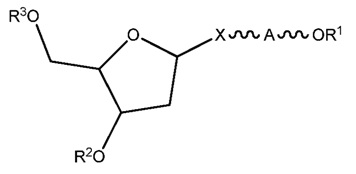
где
X представляет собой N или CH;
A является связью; замещенным или незамещенным, циклическим или ациклическим, разветвленным или неразветвленным алифатическим фрагментом; или замещенным или незамещенным, циклическим или ациклическим, разветвленным или неразветвленным гетероалифатическим фрагментом;
R1 представляет собой гидрофобный фрагмент;
R2 является водородом; кислород-защитной группой; циклическим или ациклическим, замещенным или незамещенным, разветвленным или неразветвленным алифатическим фрагментом; циклическим или ациклическим, замещенным или незамещенным, разветвленным или неразветвленным гетероалифатическим фрагментом; замещенным или незамещенным, разветвленным или неразветвленным ацилом; замещенным или незамещенным, разветвленным или неразветвленным арилом; замещенным или незамещенным, разветвленным или неразветвленным гетероарилом; и
R3 представляет собой нуклеиновую кислоту. В определенных осуществлениях, молекула нуклеиновой кислоты имеет формулу:
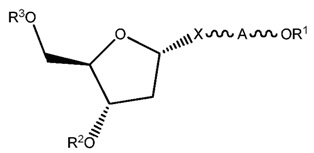 .
.
В определенных осуществлениях, молекула нуклеиновой кислоты имеет формулу:
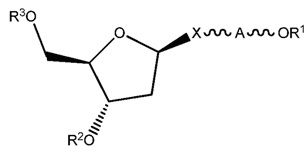
В определенных осуществлениях, молекула нуклеиновой кислоты имеет формулу:
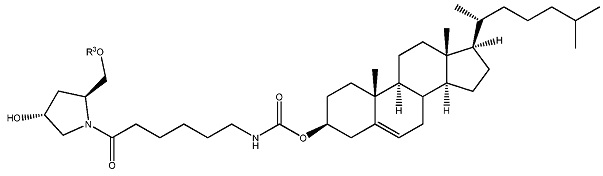
где R3 представляет собой нуклеиновую кислоту.
В определенных осуществлениях, молекула нуклеиновой кислоты имеет формулу:

где R3 представляет собой нуклеиновую кислоту; и
n представляет собой целое число от 1 до 20, включительно.
В определенных осуществлениях, молекула нуклеиновой кислоты имеет формулу:

В определенных осуществлениях, молекула нуклеиновой кислоты имеет формулу:

В определенных осуществлениях, молекула нуклеиновой кислоты имеет формулу:

В определенных осуществлениях, молекула нуклеиновой кислоты имеет формулу:
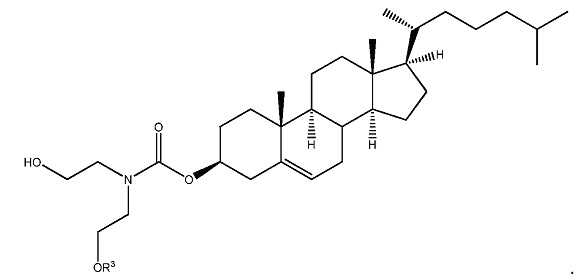
В определенных осуществлениях, молекула нуклеиновой кислоты имеет формулу:

Как используют в данной заявке, термин «связь» включает встречающийся в природе, немодифицированый фосфодиэфирный фрагмент (-O-(PO2-)-O-), который ковалентно связывает прилегающие нуклеомономеры. Как используют в данной заявке, термин «замещающая связь» включает любой аналог или производное природной фосфодиэфирной группы, которое ковалентно связывает прилегающие нуклеомономеры. Замещающие связи включают фосфодиэфирые аналоги, например, фосфоротиоат, фосфородитиоат и P-этиоксифосфодиэфир, P-этоксифосфодиэфир, P-алкилоксифосфотриэфир, метилфосфонат и несодержащие фосфор связи, например, ацетали и амиды. Такие замещающие связи известны из уровня техники (например, Bjergarde et al., 1991. Nucleic Acids Res. 19:5843; Caruthers et al., 1991. Nucleosides Nucleotides. 10:47). В определенных осуществлениях, негидролизуемые связи предпочтительны, например, фосфоротиоатные связи.
В определенных осуществлениях, олигонуклеотиды в соответствии с настоящим изобретением содержат гидрофобно модифицированные нуклеотиды или «гидрофобные модификации». Как используют в данной заявке «гидрофобные модификации» относится к основаниям, которые модифицированы таким образом, что (1) общая гидрофобность основания значительно возрастает, и/или (2) основание все еще способно образовывать близкое к обычному взаимодействие Уотсона-Крика. Некоторые неограничивающие примеры модификаций оснований включают 5-положение уридиновые и цитидиновые модификации, такие как фенил, 4-пиридил, 2-пиридил, индолил и изобутил, фенил (C6H5OH); триптофанил (C8H6N)CH2CH(NH2)CO), изобутил, бутил, аминобензил, фенил и нафтил.
Другой тип конъюгатов, которые могут быть присоединены к концу (3' или 5' концу), области петли, или любой другой части миниРНК, может включать стерин, молекулу типа стерина, пептид, малую молекулу, белок си т.д. В некоторых осуществлениях, миниРНК могут содержать более одного конъюгата (одинаковой или различной химической природы). В некоторых осуществлениях, конъюгатом является холестерин.
Другой способ увеличения специфичности гена-мишени, либо уменьшения эффекта нецелевого сайленсинга, является введение 2'-модификации (например, 2'-O метил модификации) в положении, соответствующему второму 5'-концу нуклеотида ведущей последовательности. Это позволяет позиционирование данной 2'-модификации в Dicer устойчивые шпилечные структуры, таким образом, позволяя разрабатывать лучшие PHKi конструкты с меньшим или отсутствующим нецелевым сайленсингом.
В одном осуществлении, шпилечный полинуклеотид в соответствии с настоящим изобретением может содержать одну часть нуклеиновой кислоты, которая является ДНК и одну часть нуклеиновой кислоты, которая является РНК. Антисмысловые (ведущие) последовательности в соответствии с настоящим изобретением могут быть «химерными олигонуклеотидами», которые содержат РНК-подобные и ДНК-подобные области. Выражение «область, активирующая РНКазу Н» включает область олигонуклеотида, например, химерного олигонуклеотида, которая способна рекрутировать РНКазу Н для того, чтобы расщеплять нить РНК-мишени, с которой связывается олигонуклеотид. Как правило, область, активирующая РНКазу, содержит минимальное ядро (из по меньшей мере приблизительно 3-5, типично приблизительно 3-12, более типично приблизительно 5-12 и более предпочтительно приблизительно 5-10 смежных нуклеомономеров) ДНК или ДНК-подобных нуклеомономеров. (См., например, патент США №5,849,902). Предпочтительно, область, активирующая РНКазу Н, включает приблизительно девять смежных дезоксирибоза-содержащих нуклеомономеров.
Выражение «неактивирующая область» включает область антисмысловой последовательности, например, химерного олигонуклеотида, которая не рекрутирует или не активирует РНКазу Н. Предпочтительно, неактивирующая область не содержит фосфоротиоатную ДНК. Олигонуклеотиды в соответствии с настоящим изобретением содержат по меньшей мере одну неактивирующую область. В одном осуществлении, неактивирующая область может быть стабилизирована против нуклеаз или может обеспечить специфичность мишени будучи комплементарной к мишени и образовывая водородные связи с молекулой-мишенью нуклеиновой кислоты, которая должна быть связана олигонуклеотидом.
В одном осуществлении, по меньшей мере часть смежных полинуклеотидов связана замещающей связью, например, фосфоротиоатной связью.
В определенных осуществлениях, большинство или все нуклеотиды ведущей последовательности (2'-модифицированной или нет) связаны фосфоротиоатными связями. Такие конструкты, как правило, имеют улучшенную фармакокинетику в связи с их высоким сродством к белкам сыворотки. Фосфоротиоатные связи в области неведущей последовательности полинуклеотида, как правило, не препятствуют активности ведущей нити после того, как последняя загружается в RISC.
Антисмысловые (ведущие) последовательности в соответствии с настоящим изобретением могут включать «морфолино олигонуклеотиды». Морфолино олигонуклеотиды являются неионными и функционирующими с помощью РНКаза Н-независимого механизма. Каждое из 4 генетических оснований (аденин, цитозин, гуанин и тимин/урацил) морфолино олигонуклеотидов связано с 6-членным морфолиновым кольцом. Морфолино олигонуклеотиды получены, путем ввода 4 различных типов субъединиц путем, например, неионного фосфородиамидатного взаимодействия субъединиц связи. Морфолино олигонуклеотиды имеют много преимуществ, включая: полное сопротивление нуклеазам (Antisense & Nucl. Acid Drug Dev. 1996. 6: 267); предсказуемость достижения мишени (Biochemica Biophysica Acta. 1999. 1489: 141); надежную активность в клетках (Antisense & Nucl. Acid Drug Dev. 1997. 7: 63); отличную специфичность последовательности (Antisense & Nucl. Acid Drug Dev. 1997. 7: 151); минимальную не антисмысловую активность (Biochemica Biophysica Acta 1999 1489: 141), а также простую осмотическую доставку или доставку через царапины (Antisense & Nucl. Acid Drug Dev. 1997. 7: 291). Морфолино олигонуклеотиды также являются предпочтительными из-за отсутствия токсичности в высоких дозах. Обсуждение получения морфолино олигонуклеотидов можно найти в Antisense & Nucl. Acid Drug Dev. 1997. 7: 187.
Химические модификации, описанные в данной заявке, как полагают, на основе данных, описанных в данной заявке, в целях способствования загрузке одноцепочечного полинуклеотида в RISC. Одноцепочечные полинуклеотиды, как было показано, имеют активность в загрузке в RISC и индукции генного сайленсинга. Однако, уровень активности одноцепочечных полинуклеотидов, кажется, составляет от 2 до 4 порядков ниже по сравнению с дуплексным полинуклеотидом.
Настоящее изобретение относится к описанию моделей химической модификации, которые могут (а) значительно увеличить стабильность одноцепочечного полинуклеотида (б) способствовать эффективной загрузке полинуклеотида в RISC комплекс и (в) улучшить поглощение одноцепочечного нуклеотида клеткой. Фигура 5 обеспечивает некоторые не ограничивающие примеры моделей химической модификации, которые могут быть полезны для достижения эффективности одноцепочечного полинуклеотида внутри клетки. Модели химической модификации могут включать сочетание рибозы, каркаса, гидрофобного нуклеозида и сопряженных типов модификаций. Дополнительно, в некоторых осуществлениях 5' конец одного полинуклеотида может быть химически фосфорилирован.
В еще одном осуществлении настоящее изобретение относится к описанию моделей химической модификации, которые улучшают функциональность RISC ингибирования полинуклеотидов. Одноцепочечные полинуклеотиды, как было показано, ингибируют активность предварительно загруженного RISC комплекса через механизм субстратной конкуренции. Для этих типов молекул, традиционно называемых антагомерами, активность обычно требует высокой концентрации и in vivo доставка не очень эффективна. Настоящее изобретение обеспечивает описание моделей химической модификации, которые могут (а) значительно увеличить стабильность одноцепочечного полинуклеотида (б) способствовать эффективному распознаванию полинуклеотида RISC в качестве субстрата и/или (в) улучшить поглощение одноцепочечного нуклеотида клеткой. Фигура 6 обеспечивает некоторые не ограничивающие примеры моделей химической модификации, которые могут быть полезны для достижения эффективности одноцепочечного полинуклеотида внутри клетки. Модели химической модификация могут включать сочетание рибозы, каркаса, гидрофобного нуклеозида и сопряженных типов модификаций.
Модификации, предусмотренные настоящим изобретением, применимы ко всем полинуклеотидам. Это включает одноцепочечные RISC входящие полинуклеотиды, одноцепочечные RISC ингибирующие полинуклеотиды, обычные дуплексные полинуклеотиды переменной длины (15-40 bp), асимметричные дуплексные полинуклеотиды, и тому подобное. Полинуклеотиды могут быть модифицированы широким разнообразием моделей химической модификации, включая 5' конец, рибозу, каркас и гидрофобные модификации нуклеозидов.
Синтез
Олигонуклеотиды в соответствии с настоящим изобретением могут быть синтезированы любым способом, известным в данной области техники, например, с помощью ферментативного синтеза и/или химического синтеза. Олигонуклеотиды могут быть синтезированы in vitro (например, с помощью ферментативного синтеза и химического синтеза) или in vivo (с использованием технологии рекомбинантной ДНК, хорошо известной в данной области техники).
В предпочтительном осуществлении, химический синтез используются для модифицированных полинуклеотидов. Химический синтез линейных олигонуклеотидов хорошо известен в данной области техники и может быть достигнут путем методов в растворе или твердой фазе. Предпочтительно, синтез осуществляется твердофазными способами. Олигонуклеотиды могут быть получены любым из нескольких различных синтетических процедур, включая фосфорамидитный, фосфиттриэфирный, H-фосфонатный и фосфотриэфирный способы, типично, автоматизированные способы синтеза.
Протоколы олигонуклеотидного синтеза хорошо известны в данной области техники и могут быть найдены, например, в патенте США. № 5,830,653, WO 98/13526; Stec et al. 1984. J. Am. Chem. Soc. 106:6077; Stec et al., 1985. J. Org. Chem. 50:3908; Stec et al., J. Chromatog. 1985. 326:263; LaPlanche et al., 1986. Nucl. Acid. Res. 1986. 14:9081; Fasman G. D., 1989. Practical Handbook of Biochemistry and Molecular Biology. 1989. CRC Press, Boca Raton, Fla.; Lamone. 1993. Biochem. Soc. Trans. 21:1; патенте США № 5,013,830; патенте США № 5,214,135; патенте США № 5,525,719; Kawasaki et al., 1993. J. Med. Chem. 36:831; WO 92/03568; патенте США № 5,276,019; и патенте США № 5,264,423.
Выбранный способ синтеза может зависеть от длины желаемого олигонуклеотида и такой выбор находится в компетенции специалиста в данной области техники. Например, фосфорамидитный и фосфиттриэфирный способ может привести к получению олигонуклеотидов, имеющих 175 или более нуклеотидов, в то время как H-фосфонатный способ хорошо работает для олигонуклеотидов из менее чем 100 нуклеотидов. В случае, если модифицированные основания были включены в олигонуклеотид, и особенно, если модифицированные фосфодиэфирные связи используются, то синтетические процедуры меняются по мере необходимости в соответствии с известными процедурами. В связи с этим, Uhlmann et al. (1990, Chemical Reviews 90:543-584) предоставляют ссылки и описывают процедуры получения олигонуклеотидов с модифицированными основаниями и модифицированными фосфодиэфирными связями. Другие примеры способов получения олигонуклеотидов описаны в Sonveaux. 1994. «Protecting Groups in Oligonucleotide Synthesis»; Agrawal. Methods in Molecular Biology 26:1. Иллюстративные способы синтеза также приведены в «Oligonucleotide Synthesis - A Practical Approach» (Gait, M. J. IRL Press at Oxford University Press. 1984). Дополнительно, линейные олигонуклеотиды определенной последовательности, включая некоторые последовательности с модифицированными нуклеотидами, легко доступны из нескольких коммерческих источников.
Олигонуклеотиды могут быть очищены с помощью электрофореза в полиакриламидном геле, или любого из ряда хроматографических способов, включая гель-хроматографию и жидкостную хроматографию высокого давления. Для подтверждения нуклеотидной последовательности, особенно немодифицированных нуклеотидных последовательностей, олигонуклеотиды могут быть подвергнуты секвенированию ДНК с помощью любого из известных методов, включая Максам и Гилберт секвенирование, Сангер секвенирование, секвенирование капиллярным электрофорезом, процедуру секвенирования блуждающим пятном или с помощью селективного химического распада олигонуклеотидов, связанных с Hybond бумагой. Последовательности коротких олигонуклеотидов также могут быть проанализированы масс-спектроскопией лазерной десорбции или бомбардировкой быстрыми атомами (McNeal, et al., 1982, J. Am. Chem. Soc. 104:976; Viari, et al., 1987, Biomed. Environ. Mass Spectrom. 14:83; Grotjahn et al., 1982, Nuc. Acid Res. 10:4671). Способы секвенирования также доступны для РНК олигонуклеотидов.
Качество синтезированных олигонуклеотиды может быть проверено путем тестирования олигонуклеотида методом капиллярного электрофореза и денатурирации сильным анионом HPLC (SAX-HPLC), используя, например, метод Берго и Эган. 1992. J. Chrom. 599:35.
Другие иллюстративные методы синтеза хорошо известны в данной области техники (Sambrook et al., Molecular Cloning: a Laboratory Manual, Second Edition (1989); DNA Cloning, Volumes I and II (DN Glover Ed. 1985); Oligonucleotide Synthesis (M J Gait Ed, 1984; Nucleic Acid Hybridisation (B D Hames and S J Higgins eds. 1984); Practical Guide to Molecular Cloning (1984); или the series, Methods in Enzymology (Academic Press, Inc.)).
В определенных осуществлениях, рассматриваемые РНКi конструкты или по меньшей мере их части транскрибируются из векторов экспрессии, кодирующих рассматриваемые конструкты. Любые признанные в данной области техники векторы могут быть использованы для этой цели. Транскрибированные РНКi конструкты могут быть выделены и очищены перед осуществлением желаемых модификаций (например, путем замещения немодифицированной смысловой нити модифицированной, и т.д.).
Доставка/Носитель
Поглощение олигонуклеотидов клетками
Олигонуклеотиды и олигонуклеотидные композиции контактируют с (например, их приводят в контакт с, также известными в данной заявке как вводят или доставляют) и поглощаются одной или несколькими клетками или клеточным лизатом. Термин «клетки» включает прокариотические и эукариотические клетки, предпочтительно клетки позвоночных, и, более предпочтительно, клетки млекопитающих. В предпочтительном осуществлении, олигонуклеотидные композиции в соответствии с настоящим изобретением контактируют с клетками человека.
Олигонуклеотидные композиции в соответствии с настоящим изобретением могут контактировать с клетками in vitro, например, в тестовой пробирке или культуральной планшете, (и могут или не могут быть введены субъекту) или in vivo, например, в субъекте, таком как млекопитающее. Олигонуклеотиды поглощаются клетками медленными темпами путем эндоцитоза, но эндоцитарные олигонуклеотиды, как правило, поглощены и не доступны, например, для гибридизации молекулы-мишени нуклеиновой кислоты. В одном осуществлении, поглощение клетками может быть облегчено путем электропорации или осаждения фосфатом кальция. Однако, эти процедуры полезные только для in vitro или ex vivo осуществлений, не удобны, и в некоторых случаях, связаны с клеточной токсичностью.
В другом осуществлении, доставка олигонуклеотидов в клетки может быть повышена с помощью приемлемых признанных в уровне техники методов, включая фосфат кальция, ДМСО, глицерин или декстран, электропорацию, или трансфекцию, например, с использованием катионных, анионных или нейтральных липидных композиций или липосом используя методы, известные в данной области техники (см., например, WO 90/14074, WO 91/16024, WO 91/17424;. Патент США №4,897,355; Bergan et al., 1993. Nucleic Acids Research. 21: 3567). Повышенная доставка олигонуклеотидов также может быть опосредована использованием векторов (см., например, Shi, Y. 2003. Trends Genet 2003 Jan. 19: 9; Reichhart J M et al., Genesis. 2002. 34(1-2): 1604, Yu et al., 2002. Proc. Natl. Acad Sci. USA 99: 6047; Sui et al., 2002. Proc. Natl. Acad Sci. USA 99: 5515), вирусов, полиаминных или поликатионных конъюгатов с использованием соединений, таких как полилизин, протамин, или Ni, N12-бис-(этил) спермин (см., например, Bartzatt, R. et al., 1989. Biotechnol. Appl. Biochem. 11: 133; Wagner E. et al., 1992. Proc. Natl. Acad. Sci. 88: 4255).
В определенных осуществлениях, sd-PHK настоящего изобретения могут быть доставлены с помощью различных частиц, содержащих бета-глюканы, называемые GeRPs (глюкан инкапсулированные РНК загруженные частицы), которые описаны в, и включены в качестве ссылки на, предварительную заявку США 61/310,611, поданную 4 марта 2010 года под названием «Препараты и методы для адресной доставки в фагоцитирующие клетки». Такие частицы также описаны, и включены путем ссылки на публикации США патент США 2005/0281781 А1 и патент США 2010/0040656, а также в РСТ публикации WO 2006/007372 и WO 2007/050643. sd-PHK молекула может быть гидрофобно модифицирована и необязательно может быть связана с липидом и/или амфифильным пептидом. В определенных осуществлениях, бета-глюкановые частицы получены из дрожжей. В определенных осуществлениях, нагрузка захватной молекулы представляет собой полимер, например, с молекулярной массой по меньшей мере приблизительно 1000 Da, 10000 Da, 50000 Da, 100 kDa, 500 kDa, и т.д. Предпочтительные полимеры включают (без ограничения) катионные полимеры, хитозаны или PEI (полиэтиленимин) и т.д.
Глюкановые частицы могут быть получены из нерастворимых компонентов грибковых клеточных стенок, таких, как клеточные стенки дрожжей. В некоторых осуществлениях, дрожжами являются пекарские дрожжи. Полученные из дрожжей молекулы глюкана могут включать одно или несколько из β-(1,3)-глюкана, β-(1,6)-глюкана, маннана и хитина. В некоторых осуществлениях, глюкановая частица содержит полую клеточную стенку дрожжей, где частица сохраняет трехмерную структуру, напоминающую ячейку, в рамках которой она может связываться в комплекс с или инкапсулировать молекулу, такую, как РНК молекула. Некоторые из преимуществ, связанных с использованием дрожжевой частицы клеточной стенки, представляют собой наличие компонентов, их биоразлагаемый характер, и их способность быть направленными на фагоцитные клетки.
В некоторых осуществлениях, глюкановые частицы могут быть получены путем экстракции нерастворимых компонентов из клеточных стенок, например путем экстракции пекарских дрожжей (Fleischmann) с 1М NaOH/рН 4,0 H2O, с последующим промыванием и сушкой. Способы получения дрожжевой частицы клеточной стенки обсуждены, и включены путем ссылки на патенты США 4,810,646, 4,992,540, 5,082,936, 5,028,703, 5,032,401, 5,322,841, 5,401,727, 5,504,079, 5,607,677, 5,968,811, 6,242,594, 6,444,448, 6,476,003, публикации патента США 2003/0216346, 2004/0014715 и 2010/0040656 и опубликованной заявке PCT WO02/12348.
Протоколы получения глюкановых частиц также описаны, и включены путем ссылки на, следующие ссылки: Soto and Ostroff (2008), «Characterization of multilayered nanoparticles encapsulated in yeast cell wall particles for DNA delivery.» Bioconjug Chem 19(4):840-8; Soto and Ostroff (2007), «Oral Macrophage Mediated Gene Delivery System,» Nanotech, Volume 2, Chapter 5 («Drug Delivery»), pages 378-381; and Li et al., (2007), «Yeast glucan particles activate murine resident macrophages to secrete proinflammatory cytokines via MyD88-and Syk kinase-dependent pathways,» Clinical Immunology 124(2):170-181.
Глюкан-содержащие частицы, такие как дрожжевые частицы клеточной стенки, также могут быть получены в коммерческих целях. Некоторые не ограничивающие примеры включают: Nutricell MOS 55 от Biorigin (Sao Paolo, Brazil), SAF-Mannan (SAF Agri, Minneapolis, Minn.), Nutrex (Sensient Technologies, Milwaukee, Wis.), экстрагированные щелочью частицы, такие, как полученные от Nutricepts (Nutricepts Inc., Burnsville, Minn.) и ASA Biotech, экстрагированные кислотой WGP частицы от Biopolymer Engineering, и экстрагированные органическим растворителем частицы, такие как Adjuvax™ от Alpha-beta Technology, Inc. (Worcester, Mass.) и глюкан в виде микрочастиц от Novogen (Stamford, Conn.).
Глюкановые частицы, такие, как дрожжевые частицы клеточных стенок, могут иметь различные уровни чистоты в зависимости от способа получения и/или экстракции. В некоторых случаях, частицы являются экстрагированными щелочью, экстрагированными кислотой или экстрагированными органическими растворителями для удаления внутриклеточных компонентов и/или внешнего слоя маннопротеиновой клеточной стенки. Такие протоколы могут получать частицы, которые имеют глюкановое (мас./мас.) содержание в диапазоне 50% - 90%. В некоторых случаях, частица низкой чистоты, то есть с меньшим содержанием глюкана мас./мас. может быть предпочтительной, в то время как в других осуществлениях, частица высокой чистоты, то есть с большим содержанием глюкана мас./мас. может быть предпочтительной.
Глюкановые частицы, такие как дрожжевые частицы клеточной стенки, могут иметь природное содержание липидов. Например, частицы могут содержать 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 11%, 12%, 13%, 14%, 15%, 16%, 17%, 18%, 19%, 20% или более чем на 20% мас./мас. липидов. В разделе Примеры, эффективность двух партий глюкановых частиц проходит тестирование: YGP SAF и YGP SAF + L (содержащая природные липиды). В некоторых случаях, наличие природных липидов может оказать помощь в комплексообразовании или захвате РНК молекул.
Глюкан-содержащие частицы типично имеют диаметр приблизительно 2-4 микрон, хотя частицы с диаметром менее чем 2 микрон или более чем 4 микрон также совместимы с аспектами настоящего изобретения.
РНК молекула(ы), которые будут доставляться, образуют комплекс или «заперты» внутри оболочки глюкановой частицы. Оболочка или РНК компонент частицы могут быть помечены для визуализации, как описано в, и включены путем ссылки на Soto and Ostroff (2008) Bioconjug Chem 19: 840. Способы загрузки GeRPs обсуждаются ниже.
Такие системы доставки на основе бета-глюкана могут быть сформулированы для пероральной доставки, где перорально доставленные бета-глюканы/sd-PHK конструкты могут быть захвачены макрофагами или другими родственными фагоцитирующими клетками, которые в свою очередь могут освободить sd-PHK конструкты в выбранных in vivo сайтах. Альтернативно или в дополнение sd-PHK может изменять экспрессию определенных генов-мишеней макрофагов.
Оптимальный протокол для поглощения олигонуклеотидов будет зависеть от ряда факторов, наиболее важным является тип клеток, которые используются. Другие факторы, которые играют важную роль в поглощении, включают, но не ограничиваясь приведенным, природу и концентрацию олигонуклеотида, слияние клеток, тип культуры клеток (например, суспендирование или покрытие культуры) и тип сред, в которых клетки выращивают.
Инкапсулирующие агенты
Инкапсулирующие агенты захватывают олигонуклеотиды в пузырьках. В другом осуществлении изобретения олигонуклеотид может быть связан с носителем или основой, например, липосомами или мицеллами, хотя и другие носители могут быть использованы, как будет понятно специалистам в данной области техники. Липосомы являются пузырьками, полученными из липидного двойного слоя, имеющего структуру, похожую на биологические мембраны. Такие носители используются, чтобы облегчить поглощение клетками или направление олигонуклеотида, или улучшения фармакокинетических или токсикологических свойств олигонуклеотидов.
Например, олигонуклеотиды в соответствии с настоящим изобретением можно также вводить инкапсулированными в липосомы, фармацевтические композиции, в которых содержится активный ингредиент, либо диспергированными или они разнообразно присутствуют в корпускулах, состоящих из концентрических водных слоев, имеющих сродство к липидным слоям. Олигонуклеотиды, в зависимости от растворимости, могут быть представлены как в водном слое, так и в липидном слое, или в том, что обычно называют лимосомной суспензией. Гидрофобный слой, как правило, но не исключительно, включает фосфолипиды, такие как лецитин и сфингомиелин, стероиды, такие как холестерин, более или менее ионные поверхностно-активные вещества, такие как диацетилфосфат, стеариламин, или фосфатидная кислота или другие вещества гидрофобной природы. Диаметр липосом обычно находится в диапазоне от приблизительно 15 нм до приблизительно 5 микрон.
Использование липосом в качестве основ доставки лекарственных препаратов имеет ряд преимуществ. Липосомы увеличивают внутриклеточную стабильность, повышают эффективность поглощения и повышают биологическую активность. Липосомы представляют собой полые сферические пузырьки, состоящие из липидов, расположенных таким же образом, как и липиды, образующие клеточные мембраны. Они имеют внутреннее водное пространство для улавливания растворимых в воде соединений и варьируются в размерах от 0,05 до нескольких микрон в диаметре. Несколько исследований показали, что липосомы могут доставлять нуклеиновые кислоты в клетки и, что нуклеиновые кислоты остаются биологически активными. Например, основа доставки липидов первоначально разработана в качестве инструмента исследования, такого, как Lipofectin или LIPOFECTAMINE™ 2000, и может доставить интактные молекулы нуклеиновой кислоты в клетки.
Конкретные преимущества использования липосом включают следующее: они являются не токсичными и биораспадающимися композициями; они проявляют длительный период полураспада, и распознающие молекулы могут быть легко присоединены к их поверхности для направления к тканям. Наконец, экономически эффективное получение лекарственных препаратов на основе липосом, либо в жидкой суспензии или лиофилизированном продукте, продемонстрировало жизнеспособность этой технологии в качестве приемлемой системы доставки лекарственных препаратов.
В некоторых аспектах, композиции, связанные с изобретением могут быть выбраны для класса встречающихся в природе или химически синтезированных или модифицированных насыщенных и ненасыщенных остатков жирных кислот. Жирные кислоты могут существовать в форме триглицеридов, диглицеридов или отдельных жирных кислот. В другом осуществлении, хорошо подтвержденные смеси жирных кислот и/или жировых эмульсий, которые в настоящее время используются в фармакологии для парентерального питания, могут быть использованы.
Композиции на основе липосом широко используются для доставки олигонуклеотидов. Однако, большинство из коммерчески доступных липидных или липосомных композиций содержат по меньшей мере один положительно заряженный липид (катионные липиды). Наличие этого положительно заряженного липида, как полагают, необходимо для получения высокой степени загрузки олигонуклеотидов и для повышения липосомно сливающих свойств. Несколько способов были проведены и опубликованы для определения оптимальной химии положительно заряженных липидов. Однако, коммерчески доступные липосомные композиции, содержащие катионные липиды, характеризуются высоким уровнем токсичности. In vivo ограниченные терапевтические индексы показали, что липосомные композиции, содержащие положительно заряженные липиды, связаны с токсичностью (например, повышение активности печеночных ферментов) в концентрациях, лишь незначительно более высоких, чем концентрации, необходимые для достижения РНК сайленсинга.
Нуклеиновые кислоты, связанные с изобретением, могут быть гидрофобно модифицированы и могут быть охвачены нейтральными нанотранспортерами. Дополнительное описание нейтральных нанотранспортеров включено путем ссылки на заявку РСТ PCT/US 2009/005251, поданную 22 сентября 2009 года под названием «Нейтральные нанотранспортеры». Такие частицы позволяют количественное включение олигонуклеотида в незаряженные смеси липидов. Отсутствие токсичных уровней катионных липидов в таких нейтральных композициях нанотранспортера является важной особенностью.
Как показано в PCT/US 2009/005251, олигонуклеотиды могут быть эффективно включены в смесь липидов, свободную от катионных липидов, и такая композиция может эффективно доставлять терапевтический олигонуклеотид в клетку таким образом, чтобы она являлась функциональной. Например, высокий уровень активности наблюдается, если жирная смесь состоит из фосфатидилхолинового основания жирной кислоты и стерина, такого как холестерин. Например, одна предпочтительная композиция нейтральной жирной смеси состоит из по меньшей мере 20% DOPC или DSPC и по меньшей мере 20% стерина, такого как холестерин. Даже для столь низкого, как 1:5 соотношения липидов в олигонуклеотиде было показано, что его достаточно, чтобы получить полную инкапсуляцию олигонуклеотида в незаряженной композиции.
Композиции нейтральных нанотранспортеров позволяют эффективную загрузку олигонуклеотида в нейтральную жировую композицию. Композиция включает олигонуклеотид, который модифицирован таким образом, что гидрофобность молекулы увеличивается (например, гидрофобная молекула присоединяется (ковалентно или не-ковалентно) к гидрофобной молекуле на олигонуклеотидном конце или не-концевому нуклеотиду, основанию, сахару или каркасу), модифицированный олигонуклеотид смешивают с нейтральной жировой композицией (например, содержащей по меньшей мере 25% холестерина и 25% DOPC или их аналогов). Молекула-носитель, такая, как другой липид, также может быть включена в композицию. Эта композиция, если часть композиции встроена в олигонуклеотид сама по себе, позволяет эффективную инкапсуляцию олигонуклеотида в нейтральных частицах липидов.
В некоторых аспектах, стабильные частицы размером от 50 до 140 нм могут быть сформированы после комплексообразования гидрофобных олигонуклеотидов с предпочтительными композициями. Интересно отметить, что композиция сама по себе типично не образует мелкие частицы, а, скорее, образует агломераты, которые превращаются в стабильные 50-120 нм частицы при добавлении гидрофобно модифицированного олигонуклеотида.
Композиции нейтральных нанотранспортеров в соответствии с настоящим изобретением включают гидрофобно модифицированный полинуклеотид, смесь нейтральных жиров и, необязательно, молекулу-носитель. «Гидрофобно модифицированный полинуклеотид», используемый в данной заявке, представляет собой полинуклеотид в соответствии с настоящим изобретением (например, sd-PHK), который имеет по меньшей мере одну модификацию, которая делает полинуклеотид более гидрофобным, чем полинуклеотид был до модификации. Модификация может быть достигнута путем присоединения (ковалентного или нековалентного) гидрофобной молекулы полинуклеотида. В некоторых случаях гидрофобная молекула является или включает липофильную группу.
Термин «липофильная группа» означает группу, которая имеет более высокое сродство к липидам, чем сродство к воде. Примеры липофильных групп включают, но не ограничиваясь приведенным, холестерин, холестерид или модифицированный холестериновый остаток, адамантин, дигидротестостерон, длинноцепочечный алкил, длинноцепочеченый алкенил, длинноцепочечный алкинил, олеил-литохолил, холенил, олеоил-холенил, пальмитил, гептадецил, миризитил, желчную кислоту, холевую кислоту или таурохолевую кислоту, дезоксихолат, олеил литохолевую кислоту, олеоил холеновую кислоту, гликолипиды, фосфолипиды, сфинголипиды, изопреноиды, такие, как стероиды, витамины, такие как витамин Е, жирные кислоты, либо насыщенные или ненасыщенные, сложные эфиры жирных кислот, такие как триглицериды, пирены, порфирины, тексафирин, адамантан, акридины, биотин, кумарин, флуоресцеин, родамин, Техас-красный, дигоксигенин, диметокситритил, трет-бутилдиметилсилил, трет-бутилдифенилсилил, цианиновые красители (например, Cy3 или Cy5), Hoechst 33258 краситель, псорален или ибупрофен. Холестериновый фрагмент может быть уменьшен (например, как в холестане) или может быть замещен (например, галогеном). Сочетание различных липофильных групп в одной молекуле также возможно.
Гидрофобная молекула может быть присоединена в различных положениях полинуклеотида. Как описано выше, гидрофобная молекула может быть связана с концевым остатком полинуклеотида, таким как 3' 5'-конец полинуклеотида. Альтернативно, это может быть связано с внутренним нуклеотидом или нуклеотидом на ветке полинуклеотида. Гидрофобная молекула может быть присоединена, например, в 2'-положении нуклеотида. Гидрофобная молекула также может быть связана с гетероциклическим основанием, сахаром или каркасом нуклеотида полинуклеотида.
Гидрофобная молекула может быть связана с полинуклеотидом линкерным фрагментом. Необязательно линкерный фрагмент является не-нуклеотидным линкерным фрагментом. Не-нуклеотидные линкеры представляют собой, например, абазиновые остатки (dSpacer), олигоэтиленгликоль, такой как триэтиленгликоль (спейсер 9) или гексаэтиленгликоль (спейсер 18), или алкан-диол, такой как бутандиол. Спейсерные единицы, предпочтительно, связаны фосфодиэфирными или фосфоротиоатными связями. Линкерные единицы могут появиться только один раз в молекуле или могут быть включены несколько раз, например, через фосфодиэфирные, фосфоротиоатные, метилфосфонатные или амидные связи.
Типичные протоколы сопряжения связаны с синтезом полинуклеотидов, несущих аминолинкер в одном или нескольких положениях последовательности, однако, линкер не требуется. Аминогруппа затем реагирует с молекулой, которая является сопряженной с использованием соответствующей связи или активирующих реагентов. Реакция сопряжения может быть выполнена либо с полинуклеотидом, который по-прежнему связан с твердой подложкой или следующим расщеплением полинуклеотида в фазе раствора. Очистка модифицированного полинуклеотида при помощи HPLC типично приводит к получения чистого вещества.
В некоторых осуществлениях гидрофобная молекула является конъюгатом стеринного типа, фитостерин конъюгатом, холестериновым конъюгатом, конъюгатом стеринного типа с измененной длинной боковой цепи, конъюгатом жирных кислот, любым другим гидрофобным конъюгатом и/или гидрофобными модификациями врутреннего нуклеозида, который обеспечивает достаточную гидрофобность для включения в мицеллы.
Для целей в соответствии с настоящим изобретением термин «стерины», относится к стероидным спиртам, которые являются подгруппой стероидов с гидроксильной группой в 3-положении А-кольца. Они являются амфипатическими липидами, которые синтезируются из ацетил-кофермента А через маршрут HMG-CoA-редуктазы. В целом молекула довольно плоская. Гидроксильная группа на А кольце полярная. Остальная часть алифатической цепи неполярная. Обычно стерины, как считается, имеют 8 углеродных цепей в положении 17.
Для целей в соответствии с настоящим изобретением термин «молекулы стеринного типа», относится к стероидным спиртам, которые сходны по структуре со стеринами. Основное отличие состоит в структуре кольца и количестве атомов углерода в положении 21 присоединенной боковой цепи.
Для целей в соответствии с настоящим изобретением термин «фитостерины» (также называемые растительными стеринами) относится к группе стероидных спиртов, фитохимически природно встречающихся в растениях. Существует более чем 200 различных известных фитостеринов.
Для целей в соответствии с настоящим изобретением термин «стеринная боковая цепь» относится к химической композиции боковой цепи, присоединенной в положении 17 молекулы стеринного типа. В стандартном определении стерины ограничены до 4 кольцевой структуры, имеющей 8 углеродных цепей в положении 17. В настоящем изобретении, описаны молекулы стеринного типа с боковой цепью длиннее и короче, чем обычная. Боковая цепь может быть разветвленной или содержать двойные обратные связи.
Таким образом, стерины, полезные в настоящем изобретении, например, включают холестерины, а также уникальные стерины, в которых положение 17 имеет боковую цепь из 2-7 или более чем 9 атомов углерода. В конкретном осуществлении, длина полиуглеродного хвоста варьируется от 5 до 9 атомов углерода. Такие конъюгаты могут иметь значительно лучшую in vivo эффективность, в частности, доставку в печень. Эти типы молекул, как ожидают, будут работать при концентрациях от 5 до 9 раз меньше, чем олигонуклеотиды, сопряженные с обычными холестеринами.
Альтернативно, полинуклеотид может быть связан с белком, пептидом или положительно заряженным химическим веществом, которое функционирует как гидрофобная молекула. Белки могут быть выбраны из группы, состоящей из протамина, dsРНК связывающего домена и аргинин-обогащенных пептидов. Иллюстративные положительно заряженные химические вещества включают спермин, спермидин, кадаверин и путресцин.
В другом осуществлении гидрофобная молекула конъюгатов может продемонстрировать еще более высокую эффективность при сочетании с оптимальными моделями химической модификации полинуклеотида (описанных в данной заявке подробно), содержащими, но не ограничиваясь приведенным, гидрофобные модификации, фосфоротиоатные модификации, и 2' рибо модификации.
В другом осуществлении молекула стеринного типа может быть встречающимся в природе фитостерином. Полиуглеродная цепь может быть длиннее чем 9, и может быть линейной, разветвленной и/или содержать двойные связи. Некоторые фитостерин содержащие полинуклеотидные конъюгаты могут быть значительно более эффективными и активными в доставке полинуклеотидов к различным тканям. Некоторые фитостерины могут продемонстрировать предпочтение тканей и таким образом быть использованы в качестве способа доставки РНКi в конкретные ткани.
Гидрофобно модифицированный полинуклеотид смешивают с нейтральной жирной смесью с образованием мицеллы. Нейтральная смесь жирных кислот представляет собой смесь жиров, которые имеют чистый нейтральный или слегка отрицательный заряд в или вокруг физиологического значения рН, которые могут образовывать мицеллу с гидрофобно модифицированным полинуклеотидом. Для целей в соответствии с настоящим изобретением термин «мицелла» относится к небольшой наночастице, образованной из смеси не заряженных жирных кислот и фосфолипидов. Нейтральная жирная смесь может включать катионные липиды до тех пор, пока они присутствуют в количестве, которое не вызывает токсичности. В предпочтительных осуществлениях нейтральная жирная смесь не содержит катионных липидов. Смесь, свободная от катионных липидов имеет менее чем 1% и предпочтительно 0% от общего липида, который является катионным липидом. Термин «катионный липид» включает липиды и синтетические липиды, имеющие положительный заряд в или вокруг физиологического значения рН. Термин «анионный липид» включает липиды и синтетические липиды, имеющие чистый отрицательный заряд в или вокруг физиологического значения рН.
Нейтральные жиры связываются с олигонуклеотидами в соответствии с настоящим изобретением сильной, но нековалентной связью (например, электростатической, ван дер Ваальса, пи-стэкингом и т.п. взаимодействием).
Нейтральная жировая смесь может включать композиции, выбранные из класса встречающихся в природе или химически синтезированных или модифицированных насыщенных и ненасыщенных остатков жирных кислот. Жирные кислоты могут существовать в форме триглицеридов, диглицеридов или отдельных жирных кислот. В другом осуществлении использование хорошо подтвержденных смесей жирных кислот и/или жировых эмульсий, которые в настоящее время используются в фармакологии для парентерального питания, может быть применено.
Нейтральная жирная смесь предпочтительно представляет собой смесь жирной кислоты на основе холина и стерина. Жирные кислоты на основе холина включают, например, синтетические производные фосфохолина, такие, как DDPC, DLPC, DMPC, DPPC, DSPC, DOPC, POPC и DEPC. DOPC (химический регистрационный номер 4235-95-4) является диолеилфосфатидилхолином (также известным как диелаидоилфосфатидилхолин, диолеоил-PC, диолеоилфосфохолин, диолеоил-Sn-глицеро-3-фосфохолин, диолеилфосфатидилхолин). DSPC (химический регистрационный номер 816-94-4) является дистеароилфосфатидилхолином (также известен как 1,2-дистеароил-Sn-глицеро-3-фосфохолин).
Стерин в нейтральной жирной смеси может быть, например холестерином. Нейтральная жирная смесь может быть составлена полностью из холина на основе жирной кислоты и стерина или она может необязательно включать молекулу-носитель. Например, нейтральная жирная смесь может иметь по меньшей мере 20% или 25% жирной кислоты и 20% или 25% стерина.
Для целей в соответствии с настоящим изобретением термин «жирные кислоты» относится к обычному описанию жирной кислоты. Они могут существовать как отдельные вещества или в виде ди- и триглицеридов. Для целей в соответствии с настоящим изобретением термин «жировые эмульсии» относится к безопасным жировым композициям, которые вводят внутривенно, субъектам, которые не могут получить достаточное количество жира в своем рационе. Это фосфолипиды эмульсии соевого масла (или других природных масел) и яиц. Жировые эмульсии используют для композиций некоторых нерастворимых анестетиков. В этом описании, жировые эмульсии могут быть частью коммерчески доступных препаратов, как Интралипид, Липозин, Нутрилипид, модифицированных коммерческих препаратов, где они обогащены конкретными жирными кислотами или полностью заново сформулированными сочетаниями жирных кислот и фосфолипидов.
В одном осуществлении, клетки для приведения в контакт с олигонуклеотидной композицией в соответствии с настоящим изобретением, контактируют со смесью, содержащей олигонуклеотид и смесью, содержащей липид, например, один из липидов и липидных композиций, описанных выше на срок от приблизительно 12 часов до приблизительно 24 часов. В другом осуществлении, клетки для приведения в контакт с олигонуклеотидной композицией, контактируют со смесью, содержащей олигонуклеотид и смесью, содержащей липид, например, один из липидов и липидных композиций, описанных выше, в течение от приблизительно 1 до приблизительно пяти дней. В одном осуществлении, клетки контактируют со смесью, содержащей липид и олигонуклеотид в течение от приблизительно трех дней до приблизительно 30 дней. В другом осуществлении, смесь, содержащая липид остается в контакте с клетками в течение по меньшей мере от приблизительно пяти до приблизительно 20 дней. В другом осуществлении, смесь, содержащая липид, остается в контакте с клетками в течение по меньшей мере от приблизительно семи до приблизительно 15 дней.
50% - 60% композиции может быть необязательно любым другим липидом или молекулой. Такой липид или молекула называются в данной заявке липидом-носителем или молекулой-носителем. Молекулы-носители включают, но не ограничиваясь приведенным, интралипид, малые молекулы, фузогенные пептиды или липиды или другие малые молекулы, которые могут быть добавлены для изменения клеточного поглощения, эндосомального высвобождения или свойств распределения в тканях. Способность быть молекулами-носителями имеет важное значение для модуляции свойств этих частиц, если такие свойства являются желательными. Например, присутствие некоторых тканеспецифичных метаболитов может радикально изменить профили распределения в тканях. Например, использование интралипидных композиций, обогащенных более короткими или длинными жирными цепями с различной степенью насыщенности влияет на профили распределения в тканях этих типов композиций (и их нагрузок).
Примером липида-носителя, полезного в соответствии с настоящим изобретением, является фузогенный липид. Например, цвиттерионный DOPE липид (химический регистрационный номер 4004-5-1, 1,2-диолеоил-Sn-глицеро-3-фосфоэтаноламин) является предпочтительным липидом-носителем.
Интралипид может иметь следующий состав: 1000 мл содержит: очищенное соевое масло 90 г, очищенные яичные фосфолипиды 12 г, глицерин безводный 22 г, вода для инъекций дост.кол. 1000 мл. рН регулируют гидроксидом натрия до рН приблизительно 8. Содержание энергии/л: 4,6 МДж (190 ккал). Осмотическое давление (прибл.): 300 мОсм/кг воды. В другом осуществлении, жировой эмульсией является липозин, который содержит 5% сафлорового масла, 5% соевого масла, до 1,2% яичных фосфатидов, добавленных в качестве эмульгатора и 2,5% глицерина в воде для инъекций. Она может также содержать гидроксид натрия для регулирования рН. рН 8,0 (6,0 – 9,0). Липозин имеет осмолярность 276 мОсмоль/л (фактическая).
Изменение идентичности, количества и соотношения липидов-носителей влияет на клеточное поглощение и характеристики распределения в тканях этих соединений. Например, длина хвоста липидов и уровень насыщаемости влияет на дифференциальное поглощение в печени, легких, жире и кардиомиоцитах. Добавление специальных гидрофобных молекул, таких как витамины или различные формы стеринов, может способствовать распределению в конкретных тканях, которые участвуют в метаболизме конкретных соединений. Комплексы образуются при различных концентрациях олигонуклеотидов, с более высокими концентрациями, способствующими более эффективному образованию комплекса (Фигуры 21-22).
В другом осуществлении, жирная эмульсия основана на смеси липидов. Такие липиды могут включать природные соединения, химически синтезированные соединения, очищенные жирные кислоты или любые другие липиды. В еще одном осуществлении композиция жирной эмульсии является полностью искусственной. В конкретном осуществлении, жирная эмульсия содержит более чем 70% линолевой кислоты. В еще одном конкретном осуществлении жирная эмульсия содержит по меньшей мере 1% кардиолипина. Линолевая кислота (LA) является ненасыщенной омега-6 жирной кислотой. Она представляет собой бесцветную жидкость, полученную из карбоновой кислоты с 18-углеродной цепью и двумя цис-двойными связями.
В еще одном осуществлении в соответствии с настоящим изобретением, изменение композиции жирной эмульсии используется как способ изменить распределение в тканях гидрофобно модифицированных полинуклеотидов. Эта методика обеспечивает конкретную доставку полинуклеотидов в конкретные ткани (Фигура 12).
В другом осуществлении жирные эмульсии молекулы-носителя содержат более чем 70% линолевой кислоты (C18H32O2) и/или кардиолипина, полезного для конкретной доставки РНКi в сердечную мышцу.
Жирные эмульсии, такие как интралипид, были использованы ранее как композиция доставки для некоторых нерастворимых в воде лекарственных препаратов (таких как пропофол, переформулированный так диприван). Уникальные особенности в соответствии с настоящим изобретением включают (а) концепцию объединения модифицированных полинуклеотидов с гидрофобным соединением(ями), поэтому они могут быть включены в жирные мицеллы и (б) смешивание их с жирными эмульсиями, чтобы обеспечить обратимый носитель. После инъекции в кровоток, мицеллы обычно связываются с белками сыворотки, включая альбумин, HDL, LDL и др. Такое связывание обратимо и в конечном счете жир поглощается клетками. Полинуклеотид, включенный в качестве части мицеллы, затем будет доставлен близко к поверхности клеток. После этого может происходить клеточное поглощение через переменные механизмы, в том числе, но не ограничиваясь приведенным, доставку стеринового типа.
Комплексообразователи
Комплексообразователи связывают олигонуклеотиды в соответствии с настоящим изобретением сильной, но нековалентной связью (например, электростатической, ван дер Ваальса, пи-стэкингом, и др. взаимодействием). В одном осуществлении, олигонуклеотиды в соответствии с настоящим изобретением могут быть связаны с комплексообразователем для повышения клеточного поглощения олигонуклеотидов. Пример комплексообразователя включает катионные липиды. Катионные липиды могут быть использованы для доставки олигонуклеотидов в клетки. Однако, как обсуждено выше, композиции, не содержащие катионных липидов, являются предпочтительными в некоторых осуществлениях.
Термин «катионный липид» включает липиды и синтетические липиды, имеющие как полярные, так и неполярные домены, и которые способны быть положительно заряженными в или вокруг физиологических значений рН и которые связываются с полианионами, такими как нуклеиновые кислоты и облегчают доставку нуклеиновых кислот в клетки. В общем, катионные липиды включают насыщенные и ненасыщенные алкильные и алициклические простые и сложные эфиры, амины, амиды, или их производные. Неразветвленные и разветвленные алкильные и алкенильные группы катионных липидов могут содержать, например, от 1 до приблизительно 25 атомов углерода. Предпочтительные неразветвленные или разветвленные алкильные и алкеновые группы имеют шесть или более атомов углерода. Алициклические группы включают холестерин и другие стероидные группы. Катионные липиды могут быть получены с различными противоионами (анионами), включая, например, Cl-, Br-, I-, F-, ацетат, трифторацетат, сульфат, нитрит и нитрат.
Примеры катионных липидов включают полиэтиленимин, полиамидоамин (РАМАМ) дендримеры, липофектин (комбинацию DOTMA и DOPE), липофектазу, LIPOFECTAMINE™ (например, LIPOFECTAMINE™ 2000), DOPE, цитофектин (Gilead Sciences, Foster City, Calif.) и эуфектины (JBL, San Luis Obispo, Calif.). Иллюстративные катионные липосомы могут быть получены из N-[1-(2,3-деолеолокси)-пропил]-N,N,N-триметиламмоний хлорида (DOTMA), N-[1-(2,3-деолеолокси)-пропил]-N,N,N-триметиламмоний метилсульфата (DOTAP), 3β-[N-(N',N'-диметиламиноэтан)карбамоил]холестерина (DC-Chol), 2,3,-деолеилокси-N-[2(сперминкарбоксмидо)этил]-N,N-диметил-1-пропанаминий трифторацетата (DOSPA), 1,2-димиристилоксипропил-3-диметил-гидроксиэтил аммоний бромида и диметилдиоктадециламмоний бромида (DDAB). Катионный липид N-(1-(2,3-деолеилокси)пропил)-N,N,N-триметиламмоний хлорид (DOTMA), например, как было найдено, в 1000 раз увеличивает антисмысловой эффект фосфоротиоатного олигонуклеотида. (Vlassov et al., 1994, Biochimica et Biophysica Acta 1197:95-108). Олигонуклеотиды также могут быть связаны в комплексы с, например, поли(L-лизином) или авидином и липиды могут или не могут быть включены в эту смесь, например, стерил-поли (L-лизин).
Катионные липиды были использованы в данной области техники для доставки олигонуклеотидов в клетки (см, например, патенты США №№ 5,855,910; 5,851,548; 5,830,430; 5,780,053; 5,767,099; Lewis et al., 1996. Proc. Natl. Acad. Sci. USA 93:3176; Hope et al., 1998. Molecular Membrane Biology 15:1). Другие липидные композиции, которые могут быть использованы для облегчения поглощения рассматриваемых олигонуклеотидов, могут быть использованы в связи с заявленными способами. В дополнение к перечисленным выше, другие липидные композиции также известны в данной области техники и включают, например, описанные в патенте США № 4,235,871; патентах США №№ 4,501,728; 4,837,028; 4,737,323.
В одном осуществлении липидные композиции могут дополнительно содержать агенты, например, вирусные белки для повышения липид-опосредованной трансфекции олигонуклеотидов (Kamata, et al., 1994. Nucl. Acids. Res. 22:536). В другом осуществлении, олигонуклеотиды контактируют с клетками как часть композиции, содержащей олигонуклеотид, пептид и липид как описано, например, в патенте США 5,736,392. Улучшенные липиды были также описаны, которые резистентны к сыворотке (Lewis, et al., 1996. Proc. Natl. Acad. Sci. 93:3176). Катионные липиды и другие комлпексообразователи действуют для увеличения количества олигонуклеотидов, перенесенных в клетки путем эндоцитоза.
В другом осуществлении N-замещенные глициновые олигонуклеотиды (пептоиды) могут быть использованы для оптимизации поглощения олигонуклеотидов. Пептоиды использовали для создания катионных липидообразных соединений для трансфекции (Murphy, et al., 1998. Proc. Natl. Acad. Sci. 95:1517). Пептоиды могут быть синтезированы стандартными способами (например, Zuckermann, R. N., et al. 1992. J. Am. Chem. Soc. 114:10646; Zuckermann, R. N., et al. 1992. Int. J. Peptide Protein Res. 40:497). Комбинации катионных липидов и пептоидов, липтоидов, могут быть также использованы для оптимизации поглощения рассматриваемых олигонуклеотидов (Hunag, et al., 1998. Chemistry and Biology. 5:345). Липтоиды могут быть синтезированы путем разработки пептоидных олигонуклеотидов и связывания аминоконцевого субмономера с липидом через аминогруппу (Hunag, et al., 1998. Chemistry and Biology. 5:345).
Известно из уровня техники, что позитивно заряженные аминокислоты могут быть использованы для создания высокоактивных катионных липидов (Lewis et al. 1996. Proc. Natl. Acad. Sci. USA. 93:3176). В одном осуществлении, композиция для доставки олигонуклеотидов в соответствии с настоящим изобретением содержит ряд аргининовых, лизиновых, гистидиновых или орнитиновых остатков, связанных с липофильным фрагментом (см., например, патент США № 5,777,153).
В другом осуществлении, композиция для доставки олигонуклеотидов в соответствии с настоящим изобретением включает пептид, имеющий приблизительно от одного до приблизительно четырех основных остатков. Эти основные остатки могут быть расположены, например, на амино конце, C-конце или внутренней области пептида. Семейства аминокислотных остатков, имеющих схожие боковые цепи были определены в уровне техники. Эти семейства включают аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), кислыми боковыми цепями (например, аспарагиновой кислотой, глутаминовой кислотой), незаряженными полярными боковыми цепями (например, глицин (можно также считать неполярным), аспарагин, глутамин, серин, треонин, тирозин, цистеин), неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), бета-разветвленными боковыми цепями (например, треонин, валин, изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин). Помимо основных аминокислот, большинство или все прочие остатки пептида могут быть выбраны из неосновных аминокислот, например, аминокислот, кроме лизина, аргинина или гистидина. Предпочтительно используются в основном нейтральные аминокислоты с длинными нейтральными боковыми цепями.
В одном осуществлении, композиция для доставки олигонуклеотидов в соответствии с настоящим изобретением содержит природный или синтетический полипептид, имеющий один или несколько гамма карбоксиглутаминовых кислотных остатков, или γ-Gla остатков. Такие гамма карбоксиглутаминовые кислотные остатки могут позволить полипептиду связываться друг с другом и с мембранными поверхностями. Иными словами, полипептид, имеющий ряд γ-Gla, может быть использован в качестве общего метода доставки, который помогает РНКi конструкту прилипать к любой мембране, с которой он вступает в контакт. Это может по меньшей мере замедлить очистку РНКi конструктов из кровотока и повысить их шансы направления на мишень.
Гамма-карбоксиглутаминовые кислотные остатки могут существовать в природных белках (например, протромбин имеет 10 γ-Gla остатков). Альтернативно, они могут быть введены в очищенные, рекомбинантные или химически синтезированные полипептиды путем карбоксилирования, используя, например, витамин К-зависимую карбоксилазу. Гамма-карбоксиглутаминовые кислотные остатки могут быть последовательными или не последовательными, а общее количество и расположение таких гамма-карбоксиглутаминовых кислотных остатков в полипептиде можно регулировать/тонко настраивать для достижения различных уровней «липкости» полипептида.
В одном осуществлении, клетки для контакта с олигонуклеотидной композицией в соответствии с настоящим изобретением вводят в контакт со смесью, содержащей олигонуклеотид и смесью, содержащей липид, например, одним из липидов или липидных композиций, описанных выше на срок от приблизительно 12 часов до приблизительно 24 часов. В другом осуществлении, клетки для контакта с олигонуклеотидной композицией приводят в контакт со смесью, содержащей олигонуклеотид и смесью, содержащей липид, например, одним из липидов или липидных композиций, описанных выше в течение от приблизительно 1 до приблизительно пяти дней. В одном осуществлении, клетки контактируют со смесью, содержащей липид и олигонуклеотид в течение от приблизительно трех дней до приблизительно 30 дней. В другом осуществлении, смесь, содержащая липид, остается в контакте с клетками в течение по меньшей мере от приблизительно пяти до приблизительно 20 дней. В другом осуществлении, смесь, содержащая липид, остается в контакте с клетками в течение по меньшей мере от приблизительно семи до приблизительно 15 дней.
Например, в одном осуществлении, олигонуклеотидная композиция может контактировать с клетками в присутствии липида, такого как цитофектин CS или GSV (доступный от Glen Research, Sterling, Va), GS3815, GS2888 в течение длительного периода инкубации, описанного в данной заявке.
В одном осуществлении, инкубация клеток со смесью, содержащей липид и олигонуклеотидную композицию, не снижает жизнеспособность клеток. Предпочтительно, после периода трансфекции клетки по существу жизнеспособны. В одном осуществлении, после трансфекции клетки имеют от по меньшей мере приблизительно 70% до по меньшей мере приблизительно 100% жизнеспособности. В другом осуществлении, клетки имеют от по меньшей мере приблизительно 80% до по меньшей мере приблизительно 95% жизнеспособности. В еще одном осуществлении, клетки имеют от по меньшей мере приблизительно 85% до по меньшей мере приблизительно 90% жизнеспособности.
В одном осуществлении, олигонуклеотиды модифицированы путем присоединения пептидной последовательности, которая транспортирует олигонуклеотид в клетку, называемой в данной заявке «транспортирующий пептид». В одном осуществлении, композиция содержит олигонуклеотид, который является комплементарным к молекуле-мишени нуклеиновой кислоты, кодирующей белок, и ковалентно присоединенным транспортирующим пептидом.
Выражение «транспортирующий пептид» включает последовательность аминокислоты, которая облегчает транспортировку олигонуклеотида в клетку. Примерами пептидов, которые облегчают транспортировку фрагментов, с которыми они связаны в клетках, известны в данной области техники и включают, например, HIV TAT транскрипционный фактор, лактоферрин, герпес VP22 белок, и фактор роста фибробластов 2 (Pooga et al. 1998. Nature Biotechnology. 16:857; and Derossi et al. 1998, Trends in Cell Biology. 8:84; Elliott and O'Hare. 1997. Cell 88:223).
Олигонуклеотиды могут быть присоединены к транспортирующему пептиду с использованием известных методик, например, (Prochiantz, A. 1996. Curr. Opin. Neurobiol. 6:629; Derossi et al. 1998. Trends Cell Biol. 8:84; Troy et al 1996. J. Neurosci. 16:253), Vives et al. 1997. J. Biol. Chem. 272:16010). Например, в одном осуществлении, олигонуклеотиды, имеющие активированную тиольную группу, связаны через такую тиольную группу с цистеином, который присутствует в транспортирующем пептиде (например, цистеин, присутствующий в β очереди между второй и третьей спиралью Antennapedia гомеодомен как описано, например, в Derossi et al. 1998. Trends Cell Biol. 8:84; Prochiantz. 1996. Current Opinion in Neurobiol. 6:629; Allinquant et al. 1995. J Cell Biol. 128:919). В другом осуществлении, Boc-Cys-(Npys)OH группа может быть связана с транспортным пептидом, как последняя (N-концевая) аминокислота и олигонуклеотид, имеющий группу SH, может быть связан с пептидом (Troy et al. 1996. J. Neurosci. 16:253).
В одном осуществлении, связывающая группа может быть присоединена к нуклеомономеру и транспортирующий пептид может быть ковалентно связан с линкером. В одном осуществлении, линкер может функционировать как сайт присоединения для транспортирующего пептида и может обеспечить стабильность против нуклеаз. Примеры приемлемых линкеров включают замещенные или незамещенные С1-С20алкильные цепи, С2-С20алкенильные цепи, С2-С20алкинильные цепи, пептиды и гетероатомы (например, S, О, NH и др.). Другие примеры линкеров включают бифункциональные перекрестносшивающие агенты, такие как сульфосукцинимидил-4-(малеимидофенил)-бутират (SMPB) (см., например, Smith et al. Biochem J 1991. 276: 417-2).
В одном осуществлении, олигонуклеотиды в соответствии с настоящим изобретением синтезированы в качестве молекулярных конъюгатов, которые используют рецептор-опосредованные эндоцитозные механизмы для доставки генов в клетки (см., например, Bunnell et al. 1992. Somatic Cell and Molecular Genetics. 18: 559, и ссылки в них).
Направляющие агенты
Доставка олигонуклеотидов также может быть улучшена при помощи направления олигонуклеотидов к клеточному рецептору Направляющие фрагменты могут быть сопряжены с олигонуклеотидами или присоединены к группе-носителю (например, поли(L-лизин) или липосомы), связанной с олигонуклеотидами. Этот способ хорошо подходит для клеток, которые отображают специфический рецептор-опосредованный эндоцитоз.
Например, олигонуклеотидные конъюгаты с 6-фосфоманнозилированными белками интернализованы 20-кратно более эффективно клетками, экспрессирующими манноза-6-фосфат специфические рецепторы, чем свободные олигонуклеотиды. Олигонуклеотиды могут также быть связаны с лигандом для клеточного рецептора с использованием биоразлагаемого линкера. В другом примере, конструкт доставки является маннозилированным стрептавидином, который образует плотный комплекс с биотинилированными олигонуклеотидами. Маннозилированный стрептавидин был обнаружен как увеличивающий в 20 раз интернализацию биотинилированных олигонуклеотидов. (Vlassov et al. 1994. Biochimica et Biophysica Acta 1197: 95-108).
Дополнительно специфические лиганды могут быть сопряжены с полилизиновым компонентом полилизиновых систем доставки. Например, трансферрин-полилизин, аденовирус-полилизин, и гемагглютинин вируса гриппа НА-2 N-концевых фузогенных пептид-полилизин конъюгатов значительно повышают рецептор-опосредованную ДНК доставку в эукариотических клетках. Маннозилированный гликопротеин сопряженный с поли(L-лизин) в альвеолярных макрофагах был использован для повышения клеточного поглощения олигонуклеотидами. Liang et al. 1999. Pharmazie 54: 559-566.
Поскольку злокачественные клетки имеют повышенную потребность в основных питательных веществах, таких как фолиевая кислота и трансферрин, эти питательные вещества могут быть использованы для направления олигонуклеотидов в раковые клетки. Например, когда фолиевая кислота связана с поли(L-лизин), то повышение олигонуклеотидного поглощения наблюдается в промиелоцитарном лейкозе (HL-60) клеток и клеток меланомы человека (М-14). Ginobbi et al. 1997. Anticancer Res. 17: 29. В другом примере, липосомы, покрытые малеилированным бычьим сывороточным альбумином, фолиевой кислотой, или железным протопорфирином IX, показывают повышенное клеточное поглощение олигонуклеотидов в мышиных макрофагах, клетках KB и 2.2.15 человеческих клетках гепатомы. Liang et al. 1999. Pharmazie 54: 559-566.
Липосомы природно накапливаются в печени, селезенке и ретикулоэндотелиальной системе (так называемое, пассивное направление). Путем связи липосом с различными лигандами, такими как антитела, белок А, они могут быть активнее направлены на конкретные клеточные популяции. Например, белок А-несущие липосомы могут быть предварительно обработаны Н-2К специфическими антителами, которые направлены на мышиный основной комплекс гистосовместимости в кодировке Н-2К белка, экспрессированного на L клетках. (Vlassov et al. 1994. Biochimica et Biophysica Acta 1197: 95-108).
Другая in vitro и/или in vivo доставка PHKi реагентов известна из уровня техники и может быть использована для доставки рассматриваемых PHKi конструктов. См, например, патентные заявки США 20080152661, 20080112916, 20080107694, 20080038296, 20070231392, 20060240093, 20060178327, 20060008910, 20050265957, 20050064595, 20050042227, 20050037496, 20050026286, 20040162235, 20040072785, 20040063654, 20030157030, WO 2008/036825, WO 04/065601 и AU 2004206255 B2, только в качестве перечисления нескольких из них (все включены путем ссылки).
Введение
Оптимальный курс введения или доставки олигонуклеотидов может варьироваться в зависимости от желаемого результата и/или данного субъекта, подлежащего лечению. Используемый в данной заявке термин «введение» относится к контактированию клеток с олигонуклеотидами и может быть выполнен in vitro или in vivo. Дозировка олигонуклеотидов может быть регулирована для оптимального снижения экспрессии белка преобразованного из молекулы-мишени нуклеиновой кислоты, например, согласно измерениям считывания стабильности РНК или терапевтического ответа, без излишнего экспериментирования.
Например, экспрессия белка, кодируемого нуклеиновой кислотой-мишенью, может быть измерена, чтобы определить является или нет режим дозирования соответствующим образом отрегулированным. Дополнительно, увеличение или уменьшение в уровнях РНК или белка в клетке или полученной в клетке может быть измерено с помощью любого метода, известного из уровня техники. Путем определением того, была ли транскрипция снижена, эффективность олигонуклеотида в стимулировании расщепления РНК-мишени может быть определена.
Любая из описанных выше олигонуклеотидных композиций может быть использована отдельно или в сочетании с фармацевтически приемлемым носителем. Как используется в данной заявке, «фармацевтически приемлемый носитель» включает соответствующие растворители, дисперсионные среды, покрытия, антибактериальные и противогрибковые агенты, изотонические и задерживающие абсорбцию агенты и тому подобное. Использование таких сред и агентов для фармацевтически активных веществ хорошо известно в данной области техники. За исключением случаев, когда любая традиционная среда или агент несовместимы с активным ингредиентом, они могут быть использованы в терапевтических композициях. Дополнительные активные ингредиенты также могут быть включены в композиции.
Олигонуклеотиды могут быть включены в липосомы, или липосомы, модифицированные полиэтиленгликолем, или в смеси с катионными липидами для парентерального введения. Включение дополнительных веществ в липосомы, например, антитела, реагирующие на мембранные белки, обнаруженные для конкретных клеток-мишеней, может помочь направлению олигонуклеотидов на специфические типы клеток.
Дополнительно, настоящее изобретение предусматривает введение рассматриваемых олигонуклеотидов при помощи осмотического насоса с обеспечением непрерывной инфузии таких олигонуклеотидов, например, как описано в Rataiczak et al. (1992 Proc. Natl. Acad. Sci. USA 89:11823-11827). Такие осмотические насосы являются коммерчески доступными, например, от Alzet Inc (Palo Alto, Calif.). В некоторых случаях, местное введение и парентеральное введение в носитель катионного липида являются предпочтительными.
Что касается применений in vivo, композиции в соответствии с настоящим изобретением можно вводить пациенту в различных формах, адаптированных к выбранному маршруту введения, например, парентерально, перорально или внутрибрюшинно. Парентеральное введение, которое в некоторых случаях предпочтительнее, включает введение следующими маршрутами: внутривенное, внутримышечное, интранодальное, внутриартериальное, подкожное, внутриглазное; внутрисуставное; трансэпителиальное, включая трансдермальное; легочное путем ингаляции; офтальмологическое; сублингвальное и буккальное; местное, включая офтальмологическое; кожное; глазное; ректальное и ингаляции через нос с помощью инсуффляции. Как показано в разделе Примеры, sd-PHK молекулы могут быть эффективно введены различными методами, включая внутривенную инъекцию, подкожное введение и инсуффляцию.
sd-PHK молекулы, когда желательно, чтобы доставить их системно, могут быть получены для парентерального введения путем инъекции, например, путем болюсной инъекции или непрерывной инфузии. Композиции для инъекции могут быть представлены в единичной дозированной форме, например, в ампулах или в многодозовых контейнерах, с добавлением консерванта. Композиции могут принимать такие формы, как суспензии, растворы или эмульсии в масляных или водных основах, и могут содержать вспомогательные вещества, такие как суспендирующие, стабилизирующие и/или диспергирующие агенты.
Фармацевтические препараты для парентерального введения включают водные растворы активных соединений в водорастворимой или вододиспергируемой форме. Дополнительно, можно вводить суспензии активных соединений в виде соответствующих масляных суспензий для инъекций. Приемлемые липофильные растворители или основы включают жирные масла, например, кунжутное масло, или синтетические эфиры жирных кислот, например, этилолеат или триглицериды. Водные суспензии для инъекций могут содержать вещества, которые повышают вязкость суспензии и включают, например, натрий карбоксиметилцеллюлозу, сорбит или декстран, необязательно, суспензия может также содержать стабилизаторы. Олигонуклеотиды в соответствии с настоящим изобретением могут быть сформулированы в жидких растворах, предпочтительно в физиологически совместимых буферах, таких как раствор Хэнка или раствор Рингера. Дополнительно, олигонуклеотиды могут быть сформулированы в твердом виде и их растворяют или суспендируют непосредственно перед использованием. Лиофилизированные формы также включены в настоящее изобретение.
Фармацевтические препараты для местного введения включают трансдермальные пластыри, мази, лосьоны, кремы, гели, капли, спреи, свечи, жидкости и порошки. Дополнительно, традиционные фармацевтические носители, водные, порошковые или масляные основы, или загустители могут быть использованы в фармацевтических препаратах для местного введения.
Фармацевтические препараты для перорального введения включают порошки или гранулы, суспензии или растворы в воде или неводной среде, капсулы, саше или таблетки. Дополнительно, загустители, ароматизаторы, разбавители, эмульгаторы, диспергирующие вспомогательные средства, или связующие вещества могут быть использованы в фармацевтических препаратах для перорального введения.
Для трансмукозального или трансдермального введения, проникающие агенты для соответствующего барьера для проникновения через него, используют в композиции. Такие проникающие агенты известны в данной области техники и включают, например, для трансмукозального введения соли желчных кислот и производные фузидиевой кислоты, и моющих веществ. Трансмукозальное введение может быть через назальные спреи или с помощью свечей. Для перорального введения олигонуклеотиды сформулированы в обычные пероральные формы введения, такие как капсулы, таблетки и тоники. Для местного введения олигонуклеотиды в соответствии с настоящим изобретением сформулированы в мази, бальзамы, гели или кремы, как известно в данной области техники.
Для введения путем ингаляции, например, путем инсуффляции, sd-PHK молекулы для использования в соответствии с настоящим изобретением могут быть традиционно доставлены в форме презентации аэрозоля из упаковок под давлением или распылителя с использованием приемлемого пропеллента, например, дихлордифторметана, трихлорфторметана, дихлортетрафторэтана, двуокиси углерода или другого приемлемого газа. В случае аэрозоля под давлением единица дозы может быть определена с помощью клапана для доставки отмеренного количества. Капсулы и картриджи, например, желатин для использования в ингаляторе или инсуффляторе, могут быть сформулированы как содержащие порошковую смесь соединения и приемлемую порошковую основу, такую как лактоза или крахмал.
Дополнительно в данной заявке предусмотренна легочная доставка sd-PHK молекул. sd-PHK молекула доставляется в легкие млекопитающего на вдохе и проходит через эпителиальный слой легких в кровоток. Другие доклады про ингаляционные молекулы включают Adjei et al., 1990, Pharmaceutical Research, 7: 565-569; Adjei et al., 1990, International Journal of Pharmaceutics, 63: 135-144 (лейпролид ацетат); Braquet et al., 1989, Journal of Cardiovascular Pharmacology, 13 (suppl. 5): 143-146 (эндотелии-1); Hubbard et al., 1989, Annals of Internal Medicine, Vol. III, pp. 206-212 (a1 - антитрипсин); Smith et al., 1989, J. Clin. Invest. 84:1145-1146 (a-1-протеиназа); Oswein et al., 1990, «Aerosolization of Proteins», Proceedings of Symposium on Respiratory Drag Delivery II, Keystone, Colorado, March, (рекомбинантный человеческий гормон роста); Debs et al., 1988, J. Immunol. 140:3482-3488 (интерферон-g и фактор некроза опухолей альфа) и Platz et al., патент США № 5,284,656 (гранулоцитраный колоинестимулирующий фактор). Способ и композиция для легочной доставки лекарственных препаратов для системного эффекта описана в, и включена путем ссылки на патент США № 5,451,569, выданный 19 сентября 1995 г. Wong et al.
Предполагаемым для использования в практике настоящего изобретения является широкий диапазон механических устройств, предназначенных для легочной доставки терапевтических продуктов, включая, но не ограничиваясь приведенным, небулайзеры, ингаляторы отмеренных доз, и порошковые ингаляторы, все из которых известны специалистам в данной области техники.
Некоторые конкретные примеры коммерчески доступных устройств, приемлемых для практики в соответствии с настоящим изобретением, представляют собой Ultravent небулайзер, производства Mallinckrodt, Inc., St. Louis, Missouri; Acorn II небулайзер, произведенный Marquest Medical Products, Englewood, Colorado; Ventolin ингалятор отмеренных доз, произведенный Glaxo Inc., Research Triangle Park, North Carolina; и Spinhaler порошковый ингалятор, произведенный Fisons Corp., Bedford, Massachusetts.
Все такие устройства требуют использования композиций, приемлемых для распределения олигонуклеотида (или производного). Как правило, каждая композиция является специфической для типа устройства, которое используют, и может включать использование соответствующих пропеллентных материалов, в дополнение к обычным разбавителям, вспомогательным веществам и/или носителям, полезным в терапии. Дополнительно, использование липосом, микрокапсул или микросфер, комплексов включения или других типов носителей, предполагается. Химически модифицированный олигонуклеотид также может быть получен в различных композициях в зависимости от типа химической модификации или типа устройства, которое используют.
Композиции, приемлемые для использования с небулайзером, либо струей или ультразвуком, будут типично содержать олигонуклеотид (или производное), растворенный в воде в концентрации от приблизительно 0,1 до 25 мг биологически активного олигонуклеотида на мл раствора. Композиция может также включать буфер и простой сахар (например, для олигонуклеотидной стабилизации и регуляции осмотического давления). Композиция небулайзера может также содержать поверхностно-активное вещество, чтобы уменьшить или предотвратить поверхносно индуцированную агрегацию олигонуклеотида, вызванную распылением раствора при формировании аэрозоля.
Композиции для использования в устройстве ингалятора отмеренных доз будут обычно содержать тонко измельченный порошок, такой, как композиция сухого порошка, содержащая sd-PHK молекулу, суспендированную в пропелленте с помощью поверхностно-активного вещества. Пропеллентом может быть любой обычный материал, применяемый для этой цели, например, хлорфторуглерод, гидрохлорфторуглерод, гидрофторуглерод, или углеводороды, включая трихлорфторметан, дихлордифторметан, дихлортетрафторэтанол и 1,1,1,2-тетрафторэтан, или их комбинации. Приемлемые поверхностно-активные вещества включают сорбитантриолеат и соевый лецитин. Олеиновая кислота может также быть полезной в качестве поверхностно-активного вещества.
Композиции для распределения из устройства порошкового ингалятора будут содержать тонко измельченный сухой порошок, содержащий олигонуклеотид (или производное), а также могут содержать наполнитель, такой как лактоза, сорбит, сахароза или маннит в количествах, которые облегчают распределение порошка из устройства, например, от 50 до 90% по массе композиции. sd-PHK молекула может быть получена в виде частиц со средним размером частиц менее чем 10 мм (или микрон), наиболее предпочтительно от 0,5 до 5 мм, для наиболее эффективной доставки в отдаленные части легких.
Интраназальная доставка фармацевтической композиции в соответствии с настоящим изобретением также рассматривается. Интраназальная доставка позволяет проходить фармацевтической композиции в соответствии с настоящим изобретением в кровоток непосредственно после введения терапевтических продуктов в нос, без необходимости осаждения продукта в легких. Композиции для интраназальной доставки включают препараты с декстраном или циклодекстраном.
Для интраназального введения, полезное устройство представляет собой небольшую твердую бутылку, к которой присоединен опрыскиватель отмеренных доз. В одном осуществлении, отмеренная доза доставляется путем вытяжения раствора фармацевтической композиции в соответствии с настоящим изобретением в камеру определенного объема, где камера имеет отверстие размерами, соответствующими аэрозолю и аэрозольной композиции, путем образования спрея, когда жидкость в камере сжимается. Камера сжимается для введения фармацевтической композиции в соответствии с настоящим изобретением. В конкретном осуществлении, камера представляет собой поршневое устройство. Такие устройства коммерчески доступны.
Альтернативно, используют пластиковые бутылки сжатия с отверстием, размером соответствующим аэрозолю или аэрозольному препарату, путем образования спрея при сжатии. Отверстие обычно находится в верхней части бутылки, а сверху, как правило, сужено к концу, для частичного соответствия носовым ходам для эффективного введения аэрозольной композиции. Предпочтительно, интраназальный ингалятор обеспечит дозированное количество аэрозольной композиции для введения отмеренной дозы лекарственного препарата.
Основы для доставки лекарственных препаратов могут быть выбраны, например, для in vitro, для системного, или для местного введения. Эти основы могут быть разработаны, чтобы служить в качестве резервуара медленного высвобождения или чтобы доставлять их содержимое прямо в клетки-мишени. Преимуществом использования некоторых основ для прямой доставки лекарственных препаратов является то, что много молекул доставляется за один прием. Такие основы, как было показано, увеличивают полураспад в крови лекарственных препаратов, которые могли бы быстро выводиться из кровотока. Некоторые примеры таких специализированных основ для доставки лекарственных препаратов, которые попадают в эту категорию, включают липосомы, гидрогели, циклодекстрины, биоразлагаемые нанокапсулы, и биоадгезивные микросферы.
Описанные олигонуклеотиды могут быть введены системно субъекту. Системная абсорбция относится к поступлению лекарственных препаратов в кровоток с последующим распределением по всему организму. Маршруты введения, которые приводят к системной абсорбции, включают: внутривенные, подкожные, внутрибрюшинные и интраназальные. Каждый из этих маршрутов введения доставляет олигонуклеотид в доступные больные клетки. После подкожного введения терапевтический агент стекает в местные лимфатические узлы и проходит через лимфатическую сеть в кровоток. Скорость поступления в кровоток, как было показано, является функцией молекулярной массы или размера. Использование липосомы или другого носителя лекарственного препарата локализует олигонуклеотид в лимфатических узлах. Олигонуклеотид может быть модифицирован, чтобы диффундировать в клетку, или липосомы могут непосредственно участвовать в доставке немодифицированного или модифицированного олигонуклеотида в клетку.
Выбранный способ доставки приведет к поступлению в клетки. Предпочтительные способы доставки включают липосомы (10-400 нм), гидрогели, полимеры контролируемого высвобождения, и другие фармацевтически применимые основы, а также микроинъекции или электропорации (для ex vivo лечения).
Фармацевтические препараты в соответствии с настоящим изобретением могут быть получены и сформулированы в виде эмульсий. Эмульсии, как правило, представляют собой гетерогенные системы из одной жидкости, диспергированной в другой жидкости в виде капель, как правило, превышающих 0,1 мкм в диаметре. Эмульсии в соответствии с настоящим изобретением могут содержать эксципиенты, такие как эмульгаторы, стабилизаторы, красители, жиры, масла, воски, жирные кислоты, жирные спирты, эфиры жирных кислот, увлажнители, гидрофильные коллоиды, консерванты и антиоксиданты, также могут присутствовать в эмульсии, если необходимо. Эти эксципиенты могут быть представлены в виде раствора в водной фазе, масляной фазе, или сами по себе в качестве отдельной фазы.
Примеры встречающихся в природе эмульгаторов, которые могут быть использованы в композициях эмульсий в соответствии с настоящим изобретением, включают ланолин, пчелиный воск, фосфатиды, лецитин и гуммиарабик. Мелко измельченные твердые вещества были также использованы как хорошие эмульгаторы, особенно в сочетании с поверхностно-активными веществами и вязкими препаратами. Примеры мелко измельченных твердых частиц, которые могут быть использованы в качестве эмульгаторов, включают полярные неорганические твердые вещества, такие, как гидроксиды тяжелых металлов, неразбухающие глины, такие как бентонит, аттапульгит, гекторит, каолин, мортмориллонит, коллоидный силикат алюминия и коллоидный силикат магния алюминия, пигменты и неполярные твердые вещества, такие как уголь или глицерин тристеарат.
Примеры консервантов, которые могут быть включены в композиции эмульсий, включают метилпарабен, пропилпарабен, соли четвертичного аммония, хлорид бензалкония, сложные эфиры п-гидроксибензойной кислоты и борную кислоту. Примеры антиоксидантов, которые могут быть включены в композиции эмульсий, включают свободные радикальные поглотители, такие как токоферолы, алкил галлаты, бутилированный гидроксианизол, бутилированный гидрокситолуол, или восстановители, такие как аскорбиновая кислота и метабисульфит натрия, и антиоксидантные синергисты, такие как лимонная кислота, винная кислота и лецитин.
В одном осуществлении, композиции олигонуклеотидов сформулированы как микроэмульсии. Микроэмульсия представляет собой систему воды, масла и амфифила, которая является одним оптически изотропным и термодинамически стабильным жидким раствором. Типично микроэмульсии получают путем изначального диспергирования масла в водном растворе поверхностно-активного вещества, а затем добавляют достаточное количество 4-го компонента, как правило, спирта промежуточной длины цепи, чтобы сформировать прозрачную систему.
Поверхностно-активные вещества, которые могут быть использованы при получении микроэмульсий включают, но не ограничиваясь приведенным, ионные поверхностно-активные вещества, неионные поверхностно-активные вещества, Brij 96, полиоксиэтилен олеиловые эфиры, полиглицериновые эфиры жирных кислот, тетраглицерин монолаурат (ML310), тетраглицерин моноолеат (MO310), гексаглицерин моноолеат (PO310), гексаглицерин пентаолеат (PO500), декаглицерин монокапрат (MCA750), декаглицерин моноолеат (MO750), декаглицерин секвиолеат (S0750), декаглицерин декаолеат (DA0750), отдельно или в комбинации с ко-поверхностно-активными веществами. Ко-поверхностно-активное вещество, как правило, является коротокцепочечным спиртом, таким, как этанол, 1-пропанол и 1-бутанол, служит для увеличения межфазной текучести, проникая в пленку поверхностно-активного вещества и, следовательно, создавая неупорядоченную пленку, из-за полостей пор, образованных среди молекул поверхностно-активного вещества.
Микроэмульсии могут, однако, быть получены без использования ко-поверхностно-активных веществ и не содержащие спирт самоэмульгирующиеся микроэмульсионные системы известны в данной области техники. Водной фазой типично, могут быть, но не ограничиваются приведенным, вода, водный раствор лекарственного препарата, глицерин, ПЭГ300, ПЭГ400, полиглицерины, пропилен гликоли и производные этиленгликоля. Масляная фаза может включать, но не ограничиваясь приведенным, вещества, такие, как Captex 300, Captex 355, Capmul MCM, сложные эфиры жирных кислот, моно-, ди- и триглицериды со средней цепью (С8-C12), полиоксиэтилированные глицериновые сложные эфиры жирных кислот, жирные спирты, полигликолизированные глицериды, насыщенные полигликолизированные С8-С10 глицериды, растительные масла и силиконовое масло.
Микроэмульсии представляют особый интерес с точки зрения солюбилизации лекарственного препарата и усиленной абсорбции лекарственных препаратов. Микроэмульсии на основе липидов (как масло/вода, так и вода/масло) были предложены для повышения пероральной биодоступности лекарственных препаратов.
Микроэмульсии предлагают улучшенную солюбилизацию лекарственных препаратов, защиту лекарственного препарата от ферментативного гидролиза, возможное повышение абсорбции лекарственного препарата в связи с изменениями, индуцированными поверхностно-активным веществом, текучести и проницаемости мембран, простоту получения, удобство перорального введения более твердых форм дозирования, улучшение клинической эффективности, снижение токсичности (Constantinides et al., Pharmaceutical Research, 1994, 11:1385; Ho et al., J. Pharm. Sci., 1996, 85:138-143). Микроэмульсии были также эффективны при трансдермальной доставке активных компонентов в косметической и фармацевтической промышленности. Ожидается, что композиции микроэмульсий и композиции в соответствии с настоящим изобретением будут способствовать повышению системной абсорбции олигонуклеотидов из желудочно-кишечного тракта, а также улучшать местное клеточное поглощение олигонуклеотидов в пределах желудочно-кишечного тракта, влагалища, ротовой полости и других областях введения.
В одном осуществлении в соответствии с настоящим изобретением используются различные усилители проникновения для влияния на эффективную доставку нуклеиновых кислот, в частности олигонуклеотидов, в кожу животных. Даже не липофильные лекарственные препараты могут пересекать клеточные мембраны, если мембрану для пересечения обрабатывают усилителем проникновения. В дополнение к увеличению диффузии не липофильных лекарственных препаратов через клеточные мембраны, усилители проникновения также действуют для повышения проницаемости липофильных лекарственных препаратов.
Пять категорий усилителей проникновения, которые могут быть использованы в настоящем изобретении, включают: поверхностно-активные вещества, жирные кислоты, соли желчных кислот, хелаторы, и не хелатирующие не-поверхностно-активные вещества. Другие агенты, которые могут быть использованы для повышения проникновения вводимых олигонуклеотидов, включают: гликоли, такие как этиленгликоль и пропиленгликоль, пирролы, такие как 2-15 пиррол, азоны, и терпены, такие как лимонен, и ментон.
Олигонуклеотиды, особенно в липидных композициях, также можно вводить путем покрытия медицинского устройства, например, катетера, такого, как катетер ангиопластического баллона, композицией катионных липидов. Покрытие может быть достигнуто, например, путем погружения медицинского устройства в композицию липида или смесь композиции липида и приемдемого растворителя, например, буфера на водной основе, водного растворителя, этанола, метиленхлорида, хлороформа и тому подобное. Количество композиции природно прилипает к поверхности устройства, которое затем вводят пациенту, по мере необходимости. Альтернативно, лиофилизированная смесь липидной композиции может специфично связываться с поверхностью устройства. Такие связывающие методы описаны, например, в K. Ishihara et al., Journal of Biomedical Materials Research, Vol. 27, pp. 1309-1314 (1993), описания которых включены в данную заявку путем ссылки в полном объеме.
Полезная доза для введения и конкретный маршрут введения будут варьироваться в зависимости от таких факторов, как тип клеток, или для использования in vivo, возраст, вес и особенности животного и области, которые нужно лечить, конкретного олигонуклеотида и способа доставки, который используется, предусмотренного терапевтического или диагностического использования, и формы композиции, например, суспензия, эмульсия, мицелла или липосомы, как будет очевидно специалистам в данной области техники. Как правило, доза вводится на более низких уровнях и увеличивается до достижения желаемого эффекта. Если липиды используются для доставки олигонуклеотидов, то количество липидного соединения, которое вводят, может меняться и, как правило, зависит от количества олигонуклеотидного агента, который вводят. Например, массовое соотношение липидного соединения и олигонуклеотидного агента предпочтительно составляет от приблизительно 1:1 до приблизительно 15:1, массовое соотношение от приблизительно 5:1 до приблизительно 10:1 является более предпочтительным. Как правило, количество катионного липидного соединения, которое вводят, будет варьироваться от приблизительно 0,1 миллиграмма (мг) до приблизительно 1 грамма (г). В качестве общего руководства, типично, от приблизительно 0,1 мг до приблизительно 10 мг конкретного олигонуклеотидного агента, и от приблизительно 1 мг до приблизительно 100 мг липидных композиций, каждый на килограмм массы тела пациента, вводят, хотя более высокие и низкие количества могут быть использованы.
Агенты в соответствии с настоящим изобретением вводят субъектам или контактируют с клетками в биологически совместимой форме, приемлемой для фармацевтического введения. Под «биологически совместимой формой, приемлемой для введения» подразумевают, что олигонуклеотид вводят в форме, в которой любые токсические эффекты перевешивают терапевтический эффект олигонуклеотида. В одном осуществлении, олигонуклеотиды могут вводиться субъектам. Примеры субъектов включают млекопитающих, например, человека и других приматов; коров, свиней, лошадей и сельскохозяйственных животных; собак, кошек и других домашних животных; мышей, крыс и трансгенных не-человеческих животных.
Введение активного количества олигонуклеотида в соответствии с настоящим изобретением определяется как эффективное количество, в дозах и в течение периодов времени, необходимых для достижения желаемого результата. Например, активное количество олигонуклеотида может варьироваться в зависимости от таких факторов, как тип клеток, олигонуклеотид, который используется, и для in vivo, использует болезненное состояние, возраст, пол и массу человека, и способность олигонуклеотида вызвать желаемый ответ в человеке. Создание терапевтических уровней олигонуклеотидов в клетке зависит от скорости поглощения и оттока или распада. Уменьшение степени распада продлевает внутриклеточное полувыведение олигонуклеотида. Таким образом, химически модифицированные олигонуклеотиды, например, с изменением фосфатного каркаса, могут потребовать различные дозировки.
Точная доза олигонуклеотида и количество введенных доз будет зависеть от данных, полученных экспериментально и в клинических испытаниях. Несколько факторов, таких как желаемый эффект, средство доставки, симптом болезни, и маршрут введения, будут влиять на дозу. Дозы могут быть легко определены специалистом в данной области техники и сформулированы в рассматриваемых фармацевтических композициях. Предпочтительно, продолжительность лечения будет распространяться по меньшей мере на ход симптомов заболевания.
Режимы дозировок могут быть скорректированы для обеспечения оптимального терапевтического эффекта. Например, олигонуклеотид может быть повторно введен, например, несколько доз можно вводить ежедневно или доза может быть пропорционально уменьшена, как указано в конкретной терапевтической ситуации. Специалист в данной области техники легко сможет определить соответствующие дозы и схемы введения рассматриваемых олигонуклеотидов, будут ли олигонуклеотиды введены в клетки или субъектам.
Как описано в разделе Примеры, внутривенное введение sd-PHKs может быть оптимизировано путем тестирования режимов дозирования. В некоторых случаях, внутривенное введение достигается путем инфузии, например, с помощью инфузионного насоса для вливания молекулы в кровоток субъекта. Инфузия может быть непрерывной или прерывистой. В некоторых случаях предпочтительно, если режим дозирования включает повторные введения краткосрочной непрерывной инфузии. Например, непрерывная инфузия может длиться в течение приблизительно 5 мин, 10 мин, 20 мин, 30 мин, 40 мин, 50 мин, 1,0 часа, 1,1 часа, 1,2 часа, 1,3 часа, 1,4 часа, 1,5 часа, 1,6 часа, 1,7 часа, 1,8 часа, 1,9 часа, 2,0 часов, 2,1 часа, 2,2 часа, 2,3 часа, 2,4 часа, 2,5 часа, 2,6 часа, 2,7 часа, 2,8 часа, 2,9 часа, 3,0 часов, 3,1 часа, 3,2 часа, 3,3 часа, 3,4 часов, 3,5 часа, 3,6 часа, 3,7 часа, 3,8 часа, 3,9 часа, 4,0 часов, 4,1 часа, 4,2 часа, 4,3 часа, 4,4 часа, 4,5 часа, 4,6 часа, 4,7 часа, 4,8 часа, 4,9 часа, 5,0 часов, 5,1 часа, 5,2 часа, 5,3 часа, 5,4 часа, 5,5 часа, 5,6 часа, 5,7 часа, 5,8 часа, 5,9 часа, 6,0 часов или более чем 6,0 часов, включая любые промежуточные значения.
Инфузия может быть повторяющейся. В некоторых случаях ее вводят ежедневно, два раза в неделю, еженедельно, раз в две недели, раз в три недели, ежемесячно, раз в два месяца, раз в три месяца, раз в четыре месяца, раз в пять месяцев, раз в шесть месяцев или реже чем раз в шесть месяцев. В некоторых случаях, ее вводят несколько раз в день, неделю, месяц и/или год. Например, она может быть введена приблизительно раз в час, 2 часа, 3 часа, 4 часа, 5 часов, 6 часов, 7 часов, 8 часов, 9 часов 10 часа, 12 часов или более чем двенадцать часов. Она может быть введена 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более чем 10 раз в день.
Аспекты настоящего изобретения относятся к введению sd-PHK молекул субъекту. В некоторых случаях субъектом является пациент и введение sd-PHK молекулы включает введение sd-PHK молекулы через непрерывную инфузию в кабинете врача. Не желая быть связанными теорией, непрерывная инфузия может насытить нормальный механизм клиренса и поддерживать относительно высокие уровни соединения в крови, чтобы обеспечить распределение в тканях. sd-PHK хорошо подходят для такого подхода в связи с их низкими уровнями токсичности.
Фигура 1 показывает клиренс крови и поглощение в печени sd-PHK после sd-PHK внутривенного введения при 50 мг/кг и 10 мг/кг. Фигуры 2 и 3 демонстрируют режимы дозирования. Фигуры 4 и 5 показывают распределение sd-PHK в тканях, включая печень, селезенку, сердце и легкие после внутривенного введения. Фигура 6 демонстрирует повышение sd-PHK экспрессии в печени после введения нескольких доз в течение 24 часов.
Фигура 7 показывает, что при эквивалентных дозах 80 мг/кг sd-PHK вводили внутривенным и подкожным способами, в некоторых случаях, подкожное введение привело к более высокой эффективности sd-PHK экспрессии в печени, чем внутривенное введение. Фигура 8 подытоживает эффект маршрута введения на кинетику клиренса и уровни крови. Фигуры 9 и 10 показывают, что через 24 часа после дозирования, большее количество sd-PHK было обнаружено в печени после подкожного введения, чем после внутривенного введения. Эффективная доставка sd-PHK в кожу через подкожное введение была также продемонстрирована (Фигура 11).
В некоторых случаях, эффективное количество sd-PHK, которое доставляется путем внутривенного или подкожного введения, составляет по меньшей мере приблизительно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 100 или более чем 100 мг/кг, включая любые промежуточные значения.
Подкожное введение также может быть повторяющимся. В некоторых случаях его вводят ежедневно, два раза в неделю, еженедельно, раз в две недели, раз в три недели, ежемесячно, раз в два месяца, раз в три месяца, раз в четыре месяца, раз в пять месяцев, раз в шесть месяцев или реже чем раз в шесть месяцев. В некоторых случаях, его вводят несколько раз в день, неделю, месяц и/или год. Например, оно может быть введено приблизительно раз в час, 2 часа, 3 часа, 4 часа, 5 часов, 6 часов, 7 часов, 8 часов, 9 часов, 10 часов, 12 часов или более чем двенадцать часов. Оно может быть введено 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более чем 10 раз в день.
В некоторых случаях, sd-PHK вводят путем инсуффляции. Фигуры 13-14 показывают, что sd-PHK эффективно доставлена в легкие мыши при 1-44 мг/кг и легкие крысы при 1-10 мг/кг. В некоторых случаях, эффективное количество sd-PHK, которая доставляется путем инсуффляции, составляет по меньшей мере приблизительно 0,1, 0,2, 0,3, 0,4, 0,5, 0,6, 0,7, 0,8, 0,9, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 100 или более чем 100 мг/кг, включая любые промежуточные значения.
Введение путем инсуффляции также может быть повторяющимся. В некоторых случаях его вводят ежедневно, два раза в неделю, еженедельно, раз в две недели, раз в три недели, ежемесячно, раз в два месяца, раз в три месяца, раз в четыре месяца, раз в пять месяцев, раз в шесть месяцев или реже чем раз в шесть месяцев. В некоторых случаях, его вводят несколько раз в день, неделю, месяц и/или год. Например, оно может быть введено приблизительно раз в час, 2 часа, 3 часа, 4 часа, 5 часов, 6 часов, 7 часов, 8 часов, 9 часов, 10 часов, 12 часов или более чем двенадцать часов. Оно может быть введено 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более чем 10 раз в день.
sd-PHK молекулы, которые вводят способами, описанными в данной заявке, включая внутривенное, подкожное введение и инсуффляцию, могут быть направлены в различные отдаленные ткани организма, включая печень, сердце, легкие, почки, селезенку и кожу.
Физические методы введения нуклеиновых кислот включают инъекцию раствора, содержащего нуклеиновую кислоту, бомбардировку частицами, покрытыми нуклеиновой кислотой, просачивание клеток или организма в растворе нуклеиновой кислоты, или электропорацию клеточных мембран в присутствии нуклеиновой кислоты. Вирусный конструкт, упакованный в вирусную частицу, выполняет эффективное введение конструкта экспрессии в клетку и транскрипцию нуклеиновой кислоты, кодируемой конструктом экспрессии. Другие способы, известные в данной области техники, для введения нуклеиновых кислот в клетки могут быть использованы, например, липид-опосредованный транспорт носителей, химико-опосредованный транспорт, такой, как фосфат кальция, и тому подобное. Таким образом, нуклеиновая кислота может быть введена вместе с компонентами, которые выполняют одну или несколько из следующих функций: повышают поглощение нуклеиновой кислоты в клетке, ингибируют гибридизацию одной нити, стабилизируют одну нить, или иным образом увеличивают ингибирование гена-мишени.
Нуклеиновая кислота может быть введена непосредственно в клетку (например, внутриклеточно), или внеклеточно введена в полость, интерстициальное пространство, в кровоток организма, введена перорально или путем вдыхания, или может быть введена путем погружения клетки или организма в раствор, содержащий нуклеиновую кислоту. Сосудистая или внесосудистая циркуляция, кровь или лимфатическая система, и спинномозговая жидкость, являются сайтами, куда нуклеиновая кислота может быть введена.
Клетка с геном-мишенью может быть получена из или содержаться в любом организме. Организм может быть растительным, животным, простейшим, бактерией, вирусом или грибком. Растение может быть однодольным, двудольным или голосеменным, животное может быть позвоночным или беспозвоночным. Предпочтительные микробы, представляют собой те, которые используются в сельском хозяйстве или промышленности, и те, которые являются патогенными для растений или животных.
Альтернативно, векторы, например, трансгены, кодирующие siРНК в соответствии с настоящим изобретением, могут быть разработаны в клетке-хозяине или трансгенных животных с использованием признанных в данной области методов.
Другое применение нуклеиновых кислот в соответствии с настоящим изобретением (или векторов или трансгенов кодирующих их) представляет собой функциональный анализ, который будет проводиться в эукариотических клетках, или эукариотических нечеловеческих организмах, предпочтительно клетках или организмах млекопитающих и наиболее предпочтительно клетках человека, например, клеточных линиях, таких как HeLa или 293 или грызунах, например, крыс и мышей. При введении приемлемой нуклеиновой кислоты в соответствии с настоящим изобретением, которая достаточно комплементарна целевой мРНК последовательности для прямой мишень-специфической РНК интерференции, конкретные нокаут или нокдаун фенотипы могут быть получены в клетке-мишени, например, в клетке культуры или в организме-мишени.
Таким образом, дополнительным объектом настоящего изобретения является эукариотическая клетка или эукариотический не-человеческий организм, проявляющий ген-мишень специфичный нокаут или нокдаун фенотип, содержащий полностью или, по меньшей мере частично, недостаточную экспрессию по меньшей мере одного эндогенного гена-мишени, в котором указанную клетку или организм трансфицируют по меньшей мере одним вектором, содержащим ДНК, кодирующий РНКi агент, способный ингибировать экспрессию гена-мишени. Следует отметить, что настоящее изобретение позволяет мишень-специфичный нокаут или нокдаун нескольких различных эндогенных генов в силу специфики РНКi агента.
Ген-специфичные нокаут или нокдаун фенотипы клеток или не-человеческих организмов, в частности клеткок человека или не-человеческого млекопитающего, могут быть использованы в аналитических процедурах, например, в функциональном и/или фенотипическом анализе сложных физиологических процессов, таком как анализ профилей экспрессии генов и/или протеомов. Предпочтительно анализ проводят методами высокой производительности с использованием чипов на основе олигонуклеотидов.
Анализы олигонуклеотидной стабильности
В некоторых осуществлениях, олигонуклеотиды в соответствии с настоящим изобретением стабилизованы, например, по существу устойчивы к эндонуклеазному и экзонуклеазному распаду. Олигонуклеотид определяется как, по существу, устойчивый к нуклеазам, когда по меньшей мере приблизительно 3-кратно более устойчив к воздействию эндогенных клеточных нуклеаз, и высоко устойчив к нуклеазам, когда по меньшей мере приблизительно 6-кратно более устойчив, чем соответствующий олигонуклеотид. Это может быть продемонстрировано, показав, что олигонуклеотиды в соответствии с настоящим изобретением, по существу, устойчивы к нуклеазам с использованием методов, которые известны в данной области техники.
Один из способов, в которых существенная стабильность может быть продемонстрирована, состоит в демонстрации того, что олигонуклеотиды в соответствии с настоящим изобретением функционируют при доставке в клетку, например, что они снижают транскрипцию или трансляцию молекул-мишеней нуклеиновой кислоты, например, путем измерения уровней белка или путем измерения расщепления мРНК. Анализы, которые измеряют стабильность РНК мишени, могут быть выполнены приблизительно через 24 часа после трансфекции (например, используя методы норзерн-блоттинг, анализы защиты от растепления РНКазой, или QC-ПЦР анализы, как известно в данной области техники). Альтернативно, уровни белка-мишени могут быть измерены. Предпочтительно, в дополнение к тестированию РНК или рассматриваемого белка, уровни РНК или белка контрольного, нецелевого гена будут измерены (например, актин, или, предпочтительно, контроль со сходством последовательности мишени), как контроль специфичности. Измерения РНК или белка можно выполнять с помощью любого метода, признанного в данной области техники. Предпочтительно, измерения будут выполнены, начиная приблизительно через 16-24 часа после трансфекции. (М.Y. Chiang, et al. 1991. J Biol Chem. 266: 18162-71; T. Fisher, et al. 1993. Nucleic Acids Research. 21 3857).
Способность олигонуклеотидной композиции в соответствии с настоящим изобретением ингибировать синтез белка может быть измерена с помощью методов, известных в данной области техники, например, путем определения ингибирования транскрипции генов или синтеза белка. Например, может быть выполнено нуклеаза S1 картирование. В другом примере анализ норзерн-блоттинг может быть использован для определения наличия РНК, кодирующей конкретный белок. Например, общая РНК может быть получена для прокладки хлорида цезия (см., например, Ausebel et al, 1987. Current Protocols in Molecular Biology (Greene & Wiley, New York)). Норзерн-блоттинг может быть выполнен с использованием РНК и исследован (см., например, Id.). В другом примере, уровень конкретной мРНК, произведенной белком-мишенью, может быть измерен, например, с помощью ПЦР. В еще одном примере, вестерн-блоттинг может быть использован для измерения количества присутствующего белка-мишени. В еще одном осуществлении, фенотип зависит от количества белка, которое может быть обнаружено. Методы выполнения вестерн-блоттинга хорошо известны в данной области техники, см., например, Chen et al. J. Biol. Chem. 271: 28259.
В другом примере, последовательность промотора гена-мишени может быть связана с геном-репортером и можно контролировать транскрипцию гена-репортера (например, как описано более подробно ниже). Альтернативно, олигонуклеотидные композиции, которые не нацелены на промотор, могут быть идентифицированы путем слияния часть молекулы-мишени нуклеиновой кислоты с геном-репортером, так что ген-репортер транскрибируется. Путем контроля изменения экспрессии гена-репортера в присутствии олигонуклеотидной композиции, можно определить эффективность олигонуклеотидной композиции при ингибировании экспрессии гена-репортера. Например, в одном осуществлении, эффективная олигонуклеотидная композиция приведет к снижению экспрессии гена-репортера.
«Ген-репортер» является нуклеиновой кислотой, которая экспрессирует детектируемый генный продукт, который может быть РНК или белком. Детекция мРНК экспрессии может быть осуществлена при помощи норзерн-блоттинга и детекция белка может быть достигнута путем окрашивания антител, специфичных к белку. Предпочтительные гены-репортеры производят легко детектируемый продукт. Ген-репортер может быть функционально связан с регуляторной последовательностью ДНК, где детекция продукта гена-репортера представляет собой меру транскрипционной активности регуляторной последовательности. В предпочтительных осуществлениях генный продукт гена-репортера детектируется внутренней активностью, связанной с этим продуктом. Например, ген-репортер может кодировать генный продукт, который, через ферментативную активность, приводит к детектируемому сигналу на основе цвета, флуоресценции или люминесценции. Примеры генов-репортеров включают, но не ограничиваясь приведенным, кодирующие хлорамфеникол ацетил-трансферазу (CAT), люциферазу, бета-галактозидазу и щелочную фосфатазу.
Специалист в данной области техники легко признает многочисленные гены-репортеры, которые приемлемы для использования в настоящем изобретении. Они включают, но не ограничиваясь приведенным, хлорамфениколацетилтрансферазу (CAT), люциферазу, гормон роста человека (hGH), и бета-галактозидазу. Примеры таких генов-репортеров могут быть найдены в F. A. Ausubel et al., Eds., Current Protocols in Molecular Biology, John Wiley & Sons, New York, (1989). Любой ген, который кодирует детектируемый продукт, например, любой продукт, имеющий детектируемую ферментативную активность или в отношении которого специфические антитела могут быть подняты, может быть использован в качестве гена-репортера в данных способах.
Одна система гена-репортера представляет собой систему репортеров люцифераз светляков. (Gould, S. J., and Subramani, S. 1988. Anal, Biochem., 7:404-408, включены в данную заявку путем ссылки). Люциферазный анализ быстрый и чувствительный. В этом анализе, лизат тестовой клетки получен и скомбинирован с АТФ и субстратным люциферином. Кодированный люциферазный фермент катализирует быстрое, АТФ зависимое окисление субстрата для создания светоизлучающего продукта. Обще количество света измеряется и пропорционально количеству люциферазы, присутствует в более широком диапазоне концентраций ферментов.
CAT является другой часто используемой системой гена-репортера, основным преимуществом этой системы является то, что она было широко подтверждена и получила широкое признание в качестве меры активности промотора. (Gorman С M., Moffat, L. F., and Howard, B. H. 1982. Mol. Cell. Biol., 2:1044-1051). В этой системе, тестовые клетки трансфицировали векторами CAT экспресии и инкубировали с кандидатным веществом в течение 2-3 дней после начальной трансфекции. После этого, клеточные экстракты получали. Экстракты инкубировали с ацетил-CoA и радиоактивным хлорамфениколом. После инкубации, ацетилированный хлорамфеникол отделяли от неацетилированной формы с помощью тонкослойной хроматографии. В этом анализе, степень ацетилирования отражает активность гена CAT с конкретным промотором.
Другая приемлемая система гена-репортера основана на иммунологическом обнаружении hGH. Эта система также быстрая и простая в использовании. (Selden, R., Burke-Howie, K. Rowe, M. E., Goodman, H. M., and Moore, D. D. (1986), Mol. Cell, Biol., 6:3173-3179, включена в данную заявку путем ссылки). hGH система выгодна тем, что экспрессированный hGH полипептид анализировали в средах, а не в клеточном экстракте. Таким образом, эта система не требует разрушения тестовых клеток. Следует иметь в виду, что принцип этой системы гена-репортера не ограничивается hGH, а адаптирован для использования с любым полипептидом, для которого антитело приемлемой специфичности доступно или может быть получено.
В одном осуществлении, нуклеазная стабильность двухцепочечного олигонуклеотида в соответствии с настоящим изобретением измеряется и ее сравнивают с контролем, например, РНКi молекулой, которая типично используется в данной области техники (например, дуплексный олигонуклеотид менее чем 25 нуклеотидов в длину и содержащий 2 нуклеотидных выступающих основания) или немодифицированный дуплекс РНК с тупыми концами.
Реакция расщепления РНК-мишени достигается с помощью siРНКs в соответствии с настоящим изобретением и является очень специфичной последовательностью. Идентичность последовательностей может определяться сравнением последовательностей и алгоритмы согласования известны в данной области техники. Для определения процента идентичности двух последовательностей нуклеиновых кислот (или двух аминокислотных последовательностей), последовательности выравниваются для целей оптимального сравнения (например, гэпы могут быть введены в первую последовательность или вторую последовательность для оптимального выравнивания). Предпочтительным, не ограничивающим примером алгоритма локального выравнивания, который используется для сравнения последовательностей, является алгоритм Karlin and Altschul (1990) Proc. Natl. Acad. Sci. USA 87:2264-68, модифицированный как в Karlin and Altschul (1993) Proc. Natl. Acad. Sci. USA 90:5873-77. Такой алгоритм включен в программы BLAST (версия 2.0) из Altschul, et al. (1990) J. Mol. Biol. 215:403-10. Дополнительно, многочисленные коммерческие структуры, такие, как Dharmacon, и Invitrogen обеспечивают доступ к алгоритмам на их сайте. Whitehead Institute также предлагает бесплатную программу выбора siРНК. Более 90% идентичность последовательности, например, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или даже 100% идентичность последовательности, между siРНК и частью гена-мишени является предпочтительной. Альтернативно, siРНК может быть определена как функциональная нуклеотидная последовательность (или олигонуклеотидная последовательность), которая способна к гибридизации с частью транскрипта гена-мишени. Примеры жестких условий гибридизации полинуклеотида представлены в Sambrook, J., E. F. Fritsch, and T. Maniatis, 1989, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., chapters 9 and 11, and Current Protocols in Molecular Biology, 1995, F. M. Ausubel et al., eds., John Wiley & Sons, Inc., sections 2.10 and 6.3-6.4, включены в данную заявку путем ссылки.
Терапевтическое использование
Путем ингибирования экспрессии гена, олигонуклеотидные композиции в соответствии с настоящим изобретением могут быть использованы для лечения любых заболеваний, связанных с экспрессией белка. Примеры заболеваний, которые можно лечить с помощью олигонуклеотидных композиций, только для иллюстрации, включают: рак, ретинопатии, аутоиммунные заболевания, воспалительные заболевания (например, ICAM-1 связанные расстройства, псориаз, язвенный колит, болезнь Крона), вирусные заболевания (например, ВИЧ, гепатит С), miРНК расстройства и сердечно-сосудистые заболевания.
Как уже обсуждалсь выше, sd-rxРНК молекулы, введеные способами, описанными в данной заявке, включая внутривенное, подкожное и инсуффляцию, могут быть направлены на различные отдаленные ткани организма, включая печень, сердце, легкие, почки, селезенку, кожу и жир.
В некоторых случаях, sd-rxРНК направляется на печень и используется для улучшения по меньшей мере одного из симптомов состояния или расстройства, связанных с печенью. Некоторые не ограничивающие примеры состояний или расстройств, связанных с печенью, включают: цирроз печени, болезнь печени, вызванную употреблением алкоголя, наркотиков, гепатотоксических лекарств, радиацией, физическими травмами, воздействим гепатотоксических веществ, травматические повреждения печени, инфекции (включая вирусную, бактериальную, микоплазменную, грибковую, протозойную, паразитарные инфекции или гельминтоз), гипертрофию печени (гепатомегалию), жировую дистрофию печени, безалкогольный стеатогепатит, первичный желчный цирроз печени, билиарную атрезию, гемохроматоз, альфа-1-антитрипсиновый дефицит, болезнь накопления тип-1 гликогена, порфирию, тирозинемию, болезнь Вильсона-Коновалова, аутоиммунный гепатит, неонатальный гепатит, синдром Рейе, саркоидоз, кистозную болезнь печени (включая общего желчного протока кист, синдром Кароли, врожденный фиброз печени, и поликистоз печени), воспалительные заболевания печени (например, первичный склерозирующий холангит), муковисцидоз, туберкулез, болезнь Байлера, болезнь Ниман-Пика, гепатит (например, хронический гепатит, острый гепатит, волчаночный гепатит, аутоиммунный гепатит или вирусный гепатит), гепатит, вызванный вирусом, выбранным из группы, состоящей гепатита А, гепатита В, гепатита С, гепатита D, гепатита Е, гепатита А-Е, цитомегаловируса, вируса Эпштейна-Барр, и их комбинации, гепатита, вызванного бактериальной инфекцией, например, вызванного бактерией, выбранной из группы состоящей из лептоспир, риккетсий, и стрептококковых видов, и их комбинаций, микоплазменной инфекции, протистовой инфекции, или гельминтной инфекции. Не ограничивающие примеры состояний или расстройств, влияющих на печень, включены путем ссылки на патентные публикации США US2002/0142986 и US2007/0128294.
sd-rxРНК, которая направлена на печень, может, в некоторых случаях, направлять печень-специфичный ген или ген, который экспрессируется на более высоких уровнях в печени, чем в других тканях. В некоторых случаях, sd-rxРНК могут направлять гены, участвующие в развитии печени. Не ограничивающие примеры генов, участвующих в развитии печени, включены путем ссылки на патент США 5,955,594, и включают, например, эльф 1-3, liyor-1 (145), pk, белок 106 и praja -1. Специалисту в данной области техники должно быть понятно, что общедоступные базы данных могут быть использованы для идентификации генов, которые имеют печень-специфичную экспрессию или повышенную экспрессию в печени по сравнению с другими тканями. Не ограничивающие примеры печень-специфичных генов включены путем ссылки на патентную публикацию США 2004/0229233.
В некоторых случаях, sd-rxРНК направлена на сердце и используется для улучшения по меньшей мере одного из симптомов состояния или расстройства, связанного с сердцем. Некоторые не ограничивающие примеры состояний или расстройств, связанных с сердцем, включают: заболевания, причиненные сердечной недостаточностью, ишемическую болезнь сердца, кардиомиопатию, гипертоническую болезнь сердца, пороки сердца, врожденные пороки сердца, миокардит, аритмию и заболевания, связанные с сердечной гипертрофией и/или тахикардией, ишемическую болезнь сердца (например, инфаркт миокарда), кардиомиопатию (например, застойную кардиомиопатию, гипертрофическую кардиомиопатию, рестриктивную кардиомиопатию), гипертоническую болезнь сердца, заболевания клапанов (например, атрезию аортального клапана, митрального клапана т.п., или стеноз таковых), врожденные пороки сердца, миокардит и т.п., аритмию, включая брадиаритмию и тахиаритмию, а также примеры тахиаритмии включают суправентрикулярную аритмию (например, синусовую тахикардию, суправентрикулярную экстрасистолу, фибрилляцию предсердий, трепетание предсердий, суправентрикулярную тахикардию), желудочковую аритмию (например, желудочковую экстрасистолию, желудочковую тахикардию, фибрилляцию желудочков, трепетание-мерцание), WPW синдром, синдром удлиненного QT, синдром Бругада, аритмия-вызывающую дисплазию правого желудочка (ARVD, ARVC). Не ограничивающие примеры состояний или расстройств, влияющих на сердце, включены путем ссылки на патентную публикацию США 2007/0161584.
sd-PHK, которая направлена на сердце, может, в некоторых случаях направлять сердечно-специфичный ген или ген, который экспрессируется при более высоких уровнях в сердце, чем в других тканях. Не ограничивающие примеры сердечно-специфичных генов приведены путем ссылки на патентную публикацию США 2004/0229233, и включают, например, миозин легких цепей 2а, тропонин I, кардиотоник, предшественник натрийуретического пептида В, тропонин Т2, кардиотоник, и нервные молекулы клеточной адгезии 1. Как оценит специалист в данной области техники, общедоступные базы данных могут быть использованы для идентификации генов, которые имеют сердечно-специфичную экспрессию или повышенную экспрессию в сердце по отношению к другим тканям.
В некоторых случаях, sd-PHK направлена на легкие и используется для улучшения по меньшей мере одного из симптомов состояния или расстройства, связанных с легкими. Не ограничивающие примеры состояний или расстройств, связанных с легкими, включенные путем ссылки на патентную публикацию США 20090325958, включают: бронхиальную астму, хроническую обструктивную болезнь легких; заболевания дыхательных путей, вызванные респираторно-синцитиальным вирусом; легочную артериальную гипертензию, острый респираторный дистресс-синдром и вентилятор-индуцированное повреждение легких; муковисцидоз; бронхоэктаз; дефицит альфа-1-антитрипсина; ринит; риносинусит; первичную цилиарную дискинезию; пневмонию; бронхит, вызванный агентами, кроме респираторно-синцитиального вируса; интерстициальные заболевания легких, включая лимфангилеомиоматоз; идиопатический легочный фиброз; облитерирующий бронхиолит или облитерирующий бронхиолит, организующий пневмонии, вызванные трансплантациями легких или HSCT; неспецифическую интерстициальную пневмонию; криптогенную организацию пневмонии; острую интерстициальную пневмонию; дыхательный бронхиолит, связанный с интерстициальным заболеванием легких, или легочный саркоидоз.
sd-PHK, которая направлена на легкие, может, в некоторых случаях, направлять легкие-специфичный ген или ген, который экспрессируется при более высоких уровнях в легких, а не в других тканях. Не ограничивающие примеры легкие-специфичных генов включены путем ссылки на патентную публикацию США 2004/0229233. Как оценит специалист в данной области техники, общедоступные базы данных могут быть использованы для идентификации генов, которые имеют сердце-специфичную экспрессию или повышенную экспрессию в сердце по отношению к другим тканям.
В некоторых случаях, sd-PHK направлена на селезенку и используется для улучшения по меньшей мере одного из симптомов состояния или расстройства, связанных с селезенкой. Не ограничивающие примеры состояний или расстройств, связанных с селезенкой, включены путем ссылки на патентную публикацию США 2006/0134109 и включают: аномальные иммунобластные пролиферации неизвестного происхождения, острые инфекции, острую паразитемию, идиопатическую миелоидную метаплазию, амилоидоз, ангиоиммунобластную лимфаденопатию, антитело-покрытые клетки, асплению, аутоиммунные заболевания, аутоиммунную гемолитическую анемию, В-клеточный хронический лимфолейкоз и полиморфцитратный лейкоз, бабезиоз, карциному костного мозга, бруцеллез, карциному, цероидный гистиоцитоз, хронический алкоголизм, хроническое гранулематозное заболевание, хроническую гемолитическую анемию, хронические гемолитические расстройства, хронические воспалительные расстройства, иммунологические расстройства, хронические инфекции, хронический лимфолейкоз, хронический миелолейкоз, хроническую паразитемию, хроническую уремию, цирроз печени, болезнь холодного агглютинина, застойную спленомегалию, криоглобулинемию, диссеминированный туберкулез, диспротенемии, эндокринные расстройства, эритробластический лейкоз, эритропоэз, эссаенциальную тромбоцитемию, экстрамедуллярный гематопоэтоз, синдром Фелти, фиброконгестивную спленомегалию, грибковые инфекции, заболевание гамма тяжелой цепи, болезнь Гоше, отторжение трансплантанта, гранулематозную инфильтрацию, волосатый клеточный лейкоз, гамартомы, болезнь Ханд-Шуллер-Кристиана, гемангиомы, гемангиосаркомы, гематологические нарушения, гемоглобинопатии, гемолитическую анемию, наследственный овалоцитоз, наследственный сфероцитоз, гистиоцитарный медуллярный ретикулез, гистиоцитоз X, болезнь Ходжкина, реакции гиперчувствительности, гиперспленизм, гипоспленизм, идиопатическую тромбоцитопеническую пурпуру, IgA дефицит, иммунные гранулемы, иммунную тромбоцитопению, иммунную тромбоцитопеническую пурпуру, иммунодефицитные заболевания, инфекции, связанные с гемофагоцитарным синдромом, инфекционные гранулемы, инфекционный мононуклеоз, инфекционный эндокардит, инфильтративные спленомегалии, воспалительные псевдоопухли, лейшманиоз, болезнь Летереа-Сиве, лейкемию, липогрануломы, лимфоцитарную лейкемию, лимфомы, синдромы мальабсорбции, малярию, злокачественные лимфомы, мегакариобластный лейкоз, метастатические опухоли, моноцитарные лейкозы, мукополисахаридозы, многоцентровое заболевание Кастельмана, множественную миелому, миелоцитарные лейкозы, миелофиброз, миелопролиферативные синдромы, новообразования, болезнь Ниман-Пика, не-Ходжкинковскую лимфому, паразитарные заболевания, заражения эритроцитов, пелиоз, стойкую красную полицитемию, конгестию воротной вены, стеноз воротной вены, тромбоз воротной вены, гипертензию воротной вены, ревматоидный артрит, правостороннюю сердечную недостаточность, саркоидоз, саркому, вторичный амилоидоз, вторичную миелоидную метаплазию, сывороточную болезнь, серповидно-клеточную анемию, кисты селезенки, миокард селезенки, гипертонию селезеночной вены, стеноз селезеночной вены, тромбоз селезеночной вены, спленомегалию, заболевания накопления, системную красную волчанку, системный васкулит, Т-клеточный хронический лимфолейкоз, таласемию, тромбоцитопеническую пурпуру, тиреотоксикоз, захват незрелых гематологических клеток, туберкулез, опухолеподобные заболевания, брюшной тиф, сосудистые опухоли, васкулит и вирусные инфекции.
sd-PHK, которая направлена на селезенку, может, в некоторых случаях, направлять селезенка-специфичный ген или ген, который экспрессируется при более высоких уровнях в селезенке, чем в других тканях. Не ограничивающие примеры селезенка-специфичных генов включены путем ссылки на патентную публикацию США 2004/0229233. Специалист в данной области техники оценит, что общедоступные базы данных могут быть использованы для идентификации генов, которые имеют селезенка-специфичную экспрессию или повышенную экспрессию в селезенке по сравнению с другими тканями.
В некоторых осуществлениях, sd-PHK направлена на жир.
В некоторых осуществлениях, sd-PHK направлена против МАР4К4, PCSK9, АроВ, PPIB, KSP или VEGF.
МАР4К4 является серин/треонин протеинкиназой млекопитающих, которая относится к группе протеинкиназ, связанных с Saccharomyces cerevisiae Sterile 20 (STE20). МАР4К4 (также известна как NIK для Nck взаимодействующей киназы) была впервые выявлена в мышином скрининге белков, которые взаимодействуют с SH3 доменом Nck (Su et al. (1997). Co времени своего открытия, МАР4К4 была и продолжает быть связана с широким спектром физиологических функций.
МАР4К4, как предполагают, связана с протеинтирозинкиназными сигналами для активации JNK и может играть определенную роль в регуляции цитоскелета (Xue et al., Development (2000), 128, 1559-1572). МАР4К4, как было установлено, имеет важное значение для развития в клетках млекопитающих. Nik-/- мышиные эмбрионы, как было показано, умирают после гаструляции. Модельные эксперименты на мышах показали, что NIK играет важную и особую роль в регулировании миграции клеток, которые возникают из области первичной полоски позади узла (Xue et al., Development (2000), 128, 1559-1572). Эти эксперименты также привели к предположению, что МАР4К4 может регулировать мезодермальную миграцию, что способствует удлинению оси тела. Xue et al. дополнительно предположил, что NCK/MAP4K4 комплекс может быть необходим для сегментации пресомитной мезодермы в сомиты.
Совсем недавно, МАР4К4, также было показано, участвует в метаболических расстройствах. Например, сайленсинг МАР4К4 привел к увеличению экспрессии ядерных рецепторов гормона, PPARγ, которые регулируют экспрессию генов, ответственных за дифференциацию адипоцитов (Tang et al., Proc. Natl. Acad, Sci. (2006), 103, 2087-2092). Такие гены включают, например, инсулин-чувствительные стимулирующие транспортеры глюкозы изоформы 4 (GLUT4), которые выступают посредником инсулин-зависимого транспорта глюкозы в обе мышцы и жировую ткань. Действительно, ряд исследований показал, что МАР4К4-зависимый сигнальный путь эффективно ингибирует PPARγ чувствительную генную экспрессию, адипогенез, и инсулин-стимулированный транспорт глюкозы. Дальнейшее обсуждение PHKi, направленных на МАР4К4 описано, и включено путем ссылки на предварительную заявку США серийный №61/199,661, под названием «Ингибирование МАР4К4 посредством РНКi», поданной 19 ноября 2008 года, и заявку РСТ PCT/US2009/006211, поданной 19 ноября 2009 года под названием «Ингибирование MAP4K4 посредством РНКi».
Пропротеин конвертаза субтилизин кексин 9 (PCSK9) является членом семейства субтилизинсеринпротеаз. Другие восемь субтилизин протеаз млекопитающих, PCSK1-PCSK8 (также имеющие название PC1/3, PC2, фурин, PC4, PC5/6, PACE4, PC7 и S1P/SKI-1) являются слитыми пропротеинами, обрабатывающими широкое разнообразие белков в секреторном маршруте и играют роль в различных биологических процессах (Bergeron, F. (2000) J, Mol. Endocrinol 24, 1-22, Gensberg, K., (1998) Semin. Cell Dev. Biol. 9, 11-17, Seidah, N. G. (1999) Brain Res. 848, 45-62, Taylor, N. A., (2003) FASEB J. 17, 1215-1227, и Zhou, A., (1999) J. Biol. Chem. 274, 20745-20748). PCSK9, как было предложено, играет роль в холестериновом метаболизме. PCSK9 мРНК экспрессия уменьшается питанием диетического холестерина у мышей (Maxwell, K, N., (2003) J. Lipid Res. 44, 2109-2119), увеличивается статинами HepG2 клеток (Dubuc, G., (2004) Arterioscler. Thromb. Vase. Biol. 24, 1454-1459), и увеличивается стерин регуляторный элемент, связывающий белки (SREBF) трансгенных мышей (Horton, J, D., (2003) Proc. Natl. Acad. Sci. USA 100, 12027-12032), аналогично холестерин биосинтетическим ферментам и рецептору липопротеина низкой плотности (LDLR). Дополнительно, PCSK9 бессмысленные мутации, как было найдено, связаны с формой аутосомальной доминатной гиперхолистеринемии (Hchola3) (Abifadel, M, et at (2003) Nat. Genet, 34, 154-156, Timms, К. М., (2004) HuM. Genet 114, 349-353, Leren, T. P. (2004) Clin. Genet. 65, 419-422), PCSK9 может также играть роль в определении LDL холестериновых уровней в общей популяции, поскольку однонуклеотидные полиморфизмы (SNPs) были связаны с уровнями холестерина в японской популяции (Shioji, K., (2004) J. HuM. Genet. 49, 109-114).
Аутосомальные доминантные гиперхолестеринемии (ADHs) являются моногенными заболеваниями, в которых пациенты проявляют повышенные уровни общего и LDL холестерина, ксантомы сухожилий и ранний атеросклероз (Rader, D. J., (2003) J. Clin. Invest. Ill, 1795-1803). Патогенез ADHs и рецессивная форма, аутосомальная рецессивная гиперхолестеринемия (ARH) (Cohen, J. C, (2003) Curr. Opin. Lipidol. 14, 121-127), вызвана дефектами поглощения LDL печенью, ARH может быть вызвана LDLR мутациями, предотвращающими поглощение LDL или мутациями в белке на LDL, аполипопротеине B, который связывает LDLR. ARH вызвана мутациями в ARM белке, необходимыми для эндоцитоза LDLR-LDL комплекса посредством его взаимодействия с клатрином. Поэтому, если PCSK9 мутации являются каузативными в семействах Hchola3, то возможно, что PCSK9 играет роль в рецептор-опосредованном
поглощении LDL.
Исследования избыточной экспрессии подчеркивают роль PCSK9 в контроле LDLR уровней, из-за LDL поглощения печенью (Maxwell К.N. (2004) Proc. Natl. Acad, Sci. USA 101, 7100-7105, Benjannet, S., et al. (2004) J. Biol. Chem. 279, 48865-18875, Park, S.W., (2004) J. Biol. Chem. 279, 50630-50638). Аденовирус-опосредованная избыточная экспрессия мышиного или человеческого PCSK9 в течение 3 или 4 дней у мышей привела к повышенным уровням общего и LDL холестерина; данный эффект не наблюдали у LDLR нокаутированных животных (Maxwell К.N. (2004) Proc. Natl. Acad. Sci. USA 101, 7100-7105, Benjannet, S., et al. (2904) J. Biol. Chem. 279, 48865-48875, Park, S.W., (2004) J. Biol. Chem. 279, 50630-50638). Дополнительно, PCSK9 избыточная экспрессия приводит к сильному уменьшению гепатического LDLR белка, не влияя на IDLE мРНК уровни, уровни SREBP белка, или SREBP белок ядерное и цитоплазмовое соотношение. Эти результаты указывают, что PCSK9, прямо или опосредованно, уменьшает уровни LDLR белка по пост-транскрипицонному механизму.
Потеря функциональных мутаций в PCSK9 была разработана в мышиных моделях (Rashid et al., (2005) PNAS, 102, 5374-5379, и идентифицирована у человеческих особей Cohen et al., (2005), Nature Genetics., 37. 161-165. В обоих случаях потеря PCSK9 функции приводит к понижению общего и LDLc холетерина. В ретроспективном итоговом исследовании в течении более 15 лет, потеря одной копии PCSK9 была показана как сдвигающая LDLc ниже, что приводит к повышенной защите риск-полезный эффект от развития сердечнососудистого заболевания сердца (Cohen et al. 2006 N. Engl. J. Med., 354, 1264-1272.). Четко свидетельство данных указывает на то, что понижение PCSK9 уровней понижает LDLc.
Несмотря на значительный прогресс в области PHKi и успехи в лечении патологического процесса, который может быть опосредован нижней регуляцией PCSK9 генной экспресии, остается необходимость в агентах, которые могут ингибировать PCSK9 генную экспрессию и лечить заболевания, связанные с PCSK9 генной экспрессией.
Дополнительное обсуждение PHKi, направленной на PCSK9, описано в, и включено путем ссылки на предварительную заявку США серийный номер 61/204,348, под названием «ИНГИБИРОВАНИЕ PCSK9 ПОСРЕДСТВОМ PHKI», поданной 5 января, 2009 г., и заявку РСТ PCT/US 2010/000019, поданной 5 января, 2010, под названием «Ингибирование PCSK9 посредством PHKi».
АроВ (аполипопротеин В) является аполипопротеином, задействованным в транспорте холестерина. Человеческий ApoB представлен номером GenBank NM_000384.2. АроВ был связан с коронарным заболеванием сердца, атеросклерозом, метаболическими расстройствами и гиперхолистеринемией. PPIB (пептидил-пропил цис-транс изомераза В) связан с секреторной системой и был связан с раком и иммуносупрессией. PPIB представлен номером GenBank NM_000942. KSP (кинезин веретенообразный белок, также имеющий название кинезин член семейства 11 (KIF11), EG5, HKSP, KNSL1 и TRIP5), связан с раком. Использование KSP в качестве мишени представляет антимитотический подход к лечению рака. VEGF (сосудистый эндотелиальный фактор роста) относится к трамбоцитарному семейству факторов роста. Он связан с ангиогенезом и васулогенезом.
В некоторых случаях, sd-PHK направлена на новообразование или опухолевые ткани и используется для улучшения по меньшей мере одного из симптомов состояния или расстройства, связанного с неоплазией. Неоплазия относится к аномальной пролиферации клеток, что часто приводит к ненормальной массе ткани (например, новообразованию). Новообразование может быть доброкачественным, предзлокачественным (например, карцинома in situ), или злокачественным (раковым). Доброкачественные новообразования включают фибромиомы матки и меланоцитарные родимые пятна (например, невоидные опухоли), которые не переходят в рак. Потенциально злокачественные или предраковые, новообразования включают карциному in situ, которая является ранней формой карциномы, которая не вторгается в окружающие ткани, а размножается в их обычной среде. Злокачественные новообразования обычно называют раком, и они вторгаются и разрушают окружающие ткани, могут образовывать метастазы, и в конечном итоге это может быть фатальным для хозяина.
В некоторых случаях, sd-PHK направлена на новообразование или опухолевые клетки эпителиального происхождения. Эпителиальные клетки находятся в одном или нескольких слоях, которые покрывают всю поверхность тела и выстилают большинство полых структур организма, за исключением кровеносных сосудов, лимфатических сосудов и внутренней части сердца, которые выстланы эндотелием, и груди и брюшной полости, которые выстланы мезотелием.
Эпителиальные новообразования включают, но не ограничиваясь приведенным, доброкачественные и предзлокачественные эпителиальные опухоли, такие, как фиброаденома молочной железы и аденома толстой кишки и злокачественные эпителиальные опухоли. Злокачественные эпителиальные опухоли включают первичные опухоли, также известные как карцинома, и вторичные опухоли, также известные как метастазы эпителиального происхождения. Карциномы включают, но не ограничиваясь приведенным, железистую карциному, аденокарциному, альвеолярную аденокарциному (также называются аденокистозным раком, аденомиоэпителиомой, решетчатой карциномой и цилиндроаденомой), карциному аденоматоз, аденокарциному, карциному коры надпочечников, альвеолярную карциному клеток (также называются бронхиолярной карциномой, альвеолярными опухолевыми клетками и легочным аденоматозом), базально-клеточную карциному, базально-клеточную карциному (также называемую базалома, или базилома, и базально-клеточная эпителиома), базалоидную карциному, базасквамозную карциному, рак молочной железы, бронхоальвеолярную карциному, бронхиолярную карциному, бронхогенную карциному, медуллярную карциному, холангиоцеллюлярную карциному (также называется холангиома и холангиокарцинома), хорионическую карциному, коллоидную карциному, карциному черного угря, карциному тела, решетчатый рак, распространенную карциному, карциному кожи, цилиндрическую карциному, карциному цилиндрических клеток, карциному протоков, раковую опухоль, эмбриональную карциному, энцефалоидную карциному, эпибульбарную карциному, плоскоклеточный рак, эпителиальную карциному аденоидов, карциному изъязвлений, фиброзную карциному, желеобразную карциному, коллоидный рак, гигантоклеточную карциному, гигантоклеточный рак, железистую карциному, рак гранулезных клеток, базально-клеточную эпителиому, гематоидную карциному, гепатоцеллюлярную карциному (также называется гепатома, злокачественная гепатома и гепатокарцинома), карциному Гюртле, гиалиновую карциному, гипернефроидную карциному, детскую эмбриональную карциному, карциному in situ, интраэпидермальную карциному, интраэпителиальную карциному, карциному Кромпехера, карциному клеток Кульчицкого, линзовидную карциному, чечевицеобразную карциному, липоматозную карциному, лимфоэпителиальную карциному, карциному mastitoides, медуллярную карциному, мозговидный рак, карциному melanodes, меланотическую карциному, коллоидную карциному, карциному muciparum, слизеобразующую карциному, слизеэпидермоидную карциному, карциному mucosum, слизистую карциному, карциному myxomatodes, карциному носоглотки, карциному nigram, овсяноклеточную карциному, оссифицирующую карциному, остеоидную карциному, карциному яичников, папиллярную карциному, перипортальную карциному, преинвазивную карциному, карциному простаты, карциному почечных клеток почек (также называется аденокарциномой почек и гипернефороидной карциномой), карциному резервных клеток, саркомоподобный рак, карциному scheinderian, скиррозную карциному, карциному scroti, карциному кольцевидного трофозоита, недифференцированный рак, мелкоклеточный рак, карциному solanoid, карциному сфероидальных клеток, карциному веретенообразных клеток, губчатую карциному, сквамозную карциному, плоскоклеточный рак, гемонемную карциному, остеогенную кациному с сосудистым компонентом, гемангиому, переходно-клеточный рак, карциному tuberosum, туберозный рак, бородавчатый рак, папилломную карциному.
В других случаях, sd-PHK направлена на новобразование или опухолевые клетки мезенхимального происхождения, например, опухолевые клетки, образующие саркому. Саркомы являются редкими мезенхимальными новообразованиями, которые возникают в костях и мягких тканях. Различные типы сарком, признаны и включают липосаркомы (включая миксоидные липосаркомы и плеоморфические липосаркомы), лейомиосаркомы, рабдомиосаркомы, злокачественные периферические опухоли оболочки нерва (также называются злокачественная шваннома, нейрофибросаркома, или нейрогенная саркома), опухоли Юинга (включая саркому кости Юинга, внемышечную [не кости] саркому Юинга и примитивную нейроэктодермальную опухоль [PNET]), синовиальную саркому, ангиосаркому, гемангиосаркомы, лимфангиосаркомы, саркому Капоши, гемангиоэндотелиому, фибросаркому, десмоидную опухоль (также называемые агрессивный фиброматоз), дерматофибросаркому protuberans (DFSP), злокачественную фиброзную гистиоцитому (MFH), гемангиоперицитому, злокачественную мезенхимому, альвеолярную саркому мягких частей, эпителиоидную саркому, саркому паренхиматозных клеток, десмопластическую мелкоклеточную саркому, желудочно-кишечную стромальную опухоль (GIST) (также известную как стромальная саркома GI), остеосаркому (также известную как остеогенная саркома) мышечную и внемышечную и хондросаркому.
В других случаях, sd-PHK направлена на новообразования или опухолевые клетки меланоцитарного происхождения. Меланомы представляют собой опухоли, возникающие из меланоцитарной системы кожи и других органов. Примеры включают пигментную злокачественную меланому, поверхностную распространенную меланому, узловую меланому, и Acral lentiginous меланому.
В других случаях, sd-PHK направлена на злокачественные новообразования или опухолевые клетки, включая, но не ограничиваясь приведенным, те, которые находят в раке желчного тракта, раке эндометрия, раке пищевода, раке желудка, интраэпителиальных новообразованиях, включая болезнь Боуэна и болезнь Педжета, раке печени, раке ротовой полости, включая плоскоклеточный рак, саркоме, включая фибросаркому и остеосаркому, раке кожи, включая меланому, саркоме Капоши, раке яичка, включая зародышевые опухоли (семиномы, не семиномы (тератомы, хориокарциномы)), стромальных опухолях, опухолях половых клеток, раке щитовидной железы, включая аденокарциномы щитовидной железы и рак мозга, и раке почек, включая аденокарциному и опухоль Вильмса.
В других случаях, sd-PHK направлена на новообразования или опухолевые клетки, происходящие из костной, мышечной или соединительной ткани. Опухолевые клетки могут быть найдены в первичных опухолях (например, саркоме) костной и соединительной ткани.
В некоторых случаях, sd-PHK доставляется непосредственно в новообразование, например, путем инъекции с помощью иглы и шприца. Инъекция в новообразование позволяет большим количествам sd-PHK быть доставленными непосредственно в клетки-мишени при сведении к минимуму доставки в системные сайты. При прямой инъекции в новообразование, эффективное количество для способствования РНК интерференции при помощи sd-PHK распределяется по всему по меньшей мере значительному объему новообразования. В некоторых случаях, доставка sd-PHK требует однократной инъекции в новообразование. В других случаях, доставка sd-PHK требует множество инъекций в отдельных областях новообразования так, что вся масса новообразования наделена эффективным количеством для способствования РНК интерференции при помощи sd-PHK. См. патенты США №№5,162,115 и 5,051,257, и Livraghi et al, Tumori 72 (1986), pp. 81-87, каждый из которых включен в данную заявку путем ссылки.
Общая доза, концентрация, объем доставленной sd-PHK, и скорость доставки могут быть оптимизированы для данного типа, размера и архитектуры новообразования. Зону РНК интерференции можно контролировать с помощью оптимизации этих параметров. Объем и концентрация sd-PHK, доставленной в новообразование, должны быть достаточными для способствования РНК интерференции по всей опухоли. В зависимости от количества инъекций, и их размещения по отношению к архитектуре новообразования, может быть полезно вводить объемы всей sd-PHK меньше, чем объем новообразования, больше, чем объем новообразования, или приблизительно равные объему новообразования.
В некоторых случаях, sd-PHK доставляется непосредственно в новообразование с использованием имплантируемого устройства.
В некоторых случаях sd-PHK инъекция в новообразование может сопровождаться ультразвуковым контролем.
В других случаях, sd-PHK вводят системно, например, внутривенно, внутриартериально, внутримышечно или подкожно.
sd-PHK, которая направлена на новообразование, в некоторых случаях направляет пролиферативный ген или ген, который экспрессируется на более высоких уровнях в тканях новообразования, чем в других тканях. «Пролиферативной ген», как называют в данной заявке, может быть любым геном, который способствует, прямо или косвенно, увеличению темпов роста или репликации клеток, приводя к образованию новообразования или опухолевых клеток. Увеличение темпов роста или репликации в результате экспрессии/функции пролиферативного гена соответствует темпам роста или репликации неопухолевых тканей аналогичного происхождения (например, новообразования кожи по сравнению с неопухолевой кожей). Некоторые не ограничивающие примеры пролиферативных генов или генов, которые экспрессируются на более высоких уровнях в опухолевой ткани, чем в других тканях, включают VEGF/VEGFR, HER2, PDGF/PDGFR, HDAC, MET, c-kit, CDK, FLT-1, IGF/IGFR, FGF/FGFR, Ras/Raf, Abl, Bcl-2, Src, mTOR, PKC, МАРК, BIRC5, FAS, HIF1A, CDH16, MYC, HRAS и CTNNB1.
Фактор роста эндотелия сосудов (VEGF) является членом PDGF/VEGF семейства фактора роста и кодирует белок, который часто встречается в виде дисульфидно связанного гомодимера. Этот белок является гликозилированным митогеном, который специфично действует на эндотелиальные клетки и имеет различные эффекты, включая опосредование повышенной проницаемости сосудов, вызывая ангиогенез, васкулогенез и рост эндотелиальных клеток, способствуя миграции клеток и ингибируя апоптоз. Повышенные уровни этого белка связаны с синдромом POEMS, также известным как Кроу-Фукас синдром. Мутации в этом гене были связаны с пролиферативной и непролиферативной диабетической ретинопатией. Альтернативно сплайсированные транскриптные варианты, кодирующие либо свободно выделяемые, или клеточно-ассоциированные изоформы, были охарактеризованы и могут быть направлены с sd-PHKs в соответствии с настоящим изобретением. Существует также доказательство использования не-AUG (CUG) сайтов инициации трансляции сверху, и в каркасе первого AUG, которые приводят к дополнительным изоформам. Иллюстративный пример варианта транскрипта человеческого VEGFA представляет собой Genbank номер доступа NM_001025366.2. Его соответствующий белок представляет собой Genbank номер доступа NP_001020537.2.
Тромбоцитарный фактор роста (PDGFA/PDGFB) является членом семейства тромбоцитарного фактора роста. Четыре члена этого семейства являются митогенными факторами для клеток мезенхимального происхождения и характеризуются фрагментом из восьми цистеинов. Генный продукт PDGF может существовать либо как гомодимер или как гетеродимер с тромбоцитарным фактором роста бета-полипептида, где димеры связаны дисульфидными связями. Исследования с использованием нокаутированных мышей показали клеточные дефекты в олигодендроцитах, альвеолярных клетках гладких мышц и клетках Лейдига в яичках; нокаутированные мыши умирают либо в виде эмбрионов, либо вскоре после рождения. Два сплайсированных варианта были определены для PDGF, и могут быть направлены sd-PHK в соответствии с настоящим изобретением. Репрезентативные примеры человеческих PDGF транскриптов представляют собой GenBank номера доступа NM_002607.5 и NM_011057.3. Их соответствующие белки представляют собой Genbank номера доступа NP_002598.4 и NP_03187.2, соответственно. PDGF связывается со своим рецептором, PDGFR. Репрезентативный пример человеческого PDGFR транскрипта представляет собой Genbank номер доступа NM_006206.4, и соответствующий белок NP_006197.1.
Человеческий эпидермальный фактор роста 2 (HER2, также известный как HER-2, NEU, NGL, TKR1, CD340, MLN19 и ERBB2) кодирует член семейства рецепторов эпидермального фактора роста (EGF) рецепторов тирозинкиназы. Этот белок не имеет лиганд-связывающего собственного домена и, следовательно, не может связывать факторы роста. Однако, он сильно связывает другие лиганд-связанные EGF рецепторы членов семейства с образованием гетеродимера, стабилизируя связывание лигандов и повышая киназа-опосредованную активацию нижних сигнальных путей, таких, как те, которые вовлекают митоген-активированную протеинкиназу и фосфатидилинозитол-3-киназу. Аллельные изменения в положениях аминокислот 654 и 655 изоформы (положения 624 и 625 изоформы b) были сообщены, с самым распространенным аллелем Ile654/Ile655. Амплификация и/или избыточная экспрессия данного гена сообщены для многочисленных видов рака, включая рак молочной железы и опухоли яичников. Альтернативный сплайсинг приводит в результате к нескольким дополнительным вариантам транскриптов, некоторые кодируют различные изоформы. Каждый вариант транскрипта может быть мишенью sd-PHK в соответствии с настоящим изобретением. Репрезентативный пример варианта транскрипта HER2 представляет собой GenBank номер доступа NM_004448.2. Его соответствующий белок представляет собой Genbank номер доступа NP_004439.2.
Гистон деацителаза 1 (HDAC1), относится к семейству гистон деацетилаза/acuc/альфа и является компонентом комплекса гистон деацителаз. Она взаимодействует с ретинобластомным опухоль-супрессорным белком и этот комплекс является ключевым элементом в контроле клеточной пролиферации и дифференциирования. Вместе с метастазы-ассоциированным белком-2, он деацетилирует р53 и модулирует его влияние на рост клеток и апоптоз. В некоторых случаях, sd-PHKs могут направлять HDAC1, ретинобластомный опухоль-супрессорный белок, и/или метастазы-ассоциированный белок-2. В других случаях, sd-PHK может направлять р53. Репрезентативный пример человеческого HDAC1 транскрипта представляет собой Genbank номер доступа NM_004964.2, и соответствующий белок представляет собой Genbank номер доступа NP_004955.2.
Met прото-онкоген (МЕТ), является рецептором фактора роста гепатоцитов и кодирует тирозин-киназную активность. Основной одинарный белок предшественник цепи посттрансляционно расщепляется для получения альфа- и бета-субъединиц, которые связаны дисульфидными связями для формирования зрелого рецептора. Различные мутации в гене МЕТ связаны с папиллярной карциномой почки. Два варианта транскрипта, кодирующие разные изоформы, были найдены для этого гена, каждый из которых может быть направлен sd-PHK. Репрезентативный пример человеческого МЕТ транскрипта представляет собой Genbank номер доступа NM_000245.2, и соответствующий белок представляет собой Genbank номер доступа NP_000236.2.
V-комплект Харди-Цукерман 4 вирусного онкогена кошачьей саркомы (KIT, также известный как РВТ, SCFR, C-Kit или CD117), кодирует человеческий гомолог с-комплекта прото-онкогена. С-комплект был впервые идентифицирован как клеточный гомолог V-комплекта вирусного онкогена кошачьей саркомы. Этот белок является типом 3 трансмембранного рецептора для MGF (фактор роста тучных клеток, также известный как фактор стволовых клеток). Мутации в этом гене связаны с желудочно-кишечными стромальными опухолями, заболеваниями тучных клеток, острой миелоидной лейкемией и альбинизмом. Несколько вариантов транскриптов, кодирующих разные изоформы, были найдены для этого гена, каждый из которых может быть направлен sd-PHKs. Репрезентативный пример человеческого транскрипта KIT представляет собой Genbank номер доступа NM_000222.2, и соответствующий белок представляет собой NP_000213.1.
Циклин-зависимые киназы (CDKs) играют важную роль в клеточном цикле контроля эукариотических клеток, фосфорилируются, и таким образом активируют CDK-активирующую киназу (САК). САК является мультисубъединичным белком, который включает CDK7 (MIM 601955), циклин Н (CCNH; MIM 601953) и МАТ1. МАТ1 (для упорядочивания Trois-1) участвует в сборке САК комплекса. Репрезентативный пример человеческого транскрипта CDK представляет собой Genbank номер доступа NM_001177963.1, и соответствующий белок представляет собой NP_001171434.1.
Fms-связанная тирозинкиназа 1 (FLT-1, также известная как FLT, VEGFR1, FLT1) кодирует член семейства рецепторов фактора сосудистого эндотелиального роста (VEGFR). Члены семейства VEGFR представляют собой рецепторы тирозинкиназы (RTK), которые содержат внеклеточную лиганд-связывающую область из семи иммуноглобулинов (Ig)-подобных доменов, трансмембранный сегмент, и тирозинкиназный домен (ТК) в цитоплазматическом домене. Этот белок связывается с VEGFR-A, VEGFR-B и плацентарным фактором роста и играет важную роль в ангиогенезе и васкулогенезе. Экспрессия этого рецептора находится в клетках эндотелиальных сосудов, плацентарных трофобластных клетках и моноцитах периферической крови. Несколько вариантов транскриптов, кодирующих разные изоформы, были найдены для этого гена. Изоформы включают полноразмерную изоформу трансмембранного рецептора и укороченные, растворимые изоформы. Растворимые изоформы связаны с началом пре-эклампсии. Каждый вариант транскрипта FLT-1 может быть мишенью sd-PHK. Репрезентативный пример человеческого FLT-1 транскрипта представляет собой Genbank номер доступа NM_001159920.1, и соответствующий белок представляет собой NP_00115392.1.
Инсулиноподобные факторы роста (IGFs) аналогичны инсулину по функциям и структуре и являются членами семейства белков, участвующих в опосредовании роста и развития. IGFI белок, например, обрабатывается из предшественника, связанного со специфическим рецептором, и выделяется. Дефекты в этом гене являются причиной недостаточности инсулин-подобного фактора роста I. Несколько вариантов транскриптов, кодирующих разные изоформы, были найдены для этих генов, каждый из которых может быть мишенью sd-PHK. Иллюстративный пример человеческго транскрипта IGF представляет собой Genbank номер доступа NM_000618.3, и соответствующий белок представляет собой NP_000609.1.
Члены семейства фактора роста фибробластов (FGF) обладают широким спектром митогенной активности и активности выживаемости клеток и участвуют в различных биологических процессах, включая эмбриональное развитие, рост клеток, морфогенез, восстановление тканей, рост опухолей и инвазию. FGF1, например, функционирует в качестве модификатора миграции эндотелиальных клеток и пролиферации, а также фактора кровеносных сосудов. Он действует как митоген для различных мезодерм- и нейроэктодерм-производных клеток, полученных in vitro, поэтому как считают, они участвуют в органогенезе. Альтернативно варианты сплайсинга транскриптов, кодирующие различимые изоформы нескольких FGFs, были сообщены, каждый из которых может быть мишенью sd-PHK. Репрезентативный пример человеческого FGF1 транскрипта представляет собой Genbank номер доступа NM_000800.3, и соответствующий белок представляет собой NP_000791.1.
Члены семейства фактора роста рецептора фибробластов (FGFR), имеющие высоко консервативные аминокислотные последовательности между членами и по всей эволюции, отличаются друг от друга афинностями лиганда и распределением в тканях. Полноразмерный репрезентативный белок состоит из внеклеточной области, состоящей из трех иммуноглобулин-подобных доменов, одного гидрофобного мембранно-распространенного сегмента и цитоплазматического домена тирозинкиназы. Внеклеточная часть белка взаимодействует с факторами роста фибробластов, приводя в движение каскад сигналов ниже, в конечном счете влияя на митогенез и дифференциацию. FGFR1, например, связывает как кислотные, так и основные факторы роста фибробластов и участвует в индукции конечностей. Мутации в этом гене были связаны с синдромом Пфайффера, синдромом Джексона-Вейсса, синдромом Антли-Бикслера, остеоглофоновой дисплазией, и аутосомно-доминантным синдромом Кальмана 2. Хромосомные аберрации с участием FGFR1 связаны с миелопролиферативным расстройством стволовых клеток и синдромом лейкемии лимфомы стволовых клеток. Альтернативно сплайсированные варианты, которые кодируют разные изоформы белка членов семейства FGFR1 были описаны, каждый из которых может быть мишенью sd-PHK. Репрезентативный пример человеческого FGFR1 представляет собой Genbank номер доступа NM_001174063.1, и соответствующий белок представляет собой NP_001167534.1.
Ras подсемейство (аббревиатура саркомы крыс) является подсемейством белка малых GTPaз, которые участвуют в клеточной передаче сигнала, а также используются для обозначения генов подсемейства генов, кодирующих эти белки. Активация Ras сигнализации вызывает клеточный рост, дифференцирование и выживание. Ras является прототипическим членов суперсемейства Ras белков, которые все связаны в структуру и регулируют разнообразное поведение клетки. Поскольку Ras передает сигналы из вне клетки к ядру, мутации в генах Ras могут постоянно активировать его и привести к нарушению передачи внутри клетки, даже в отсутствие внеклеточных сигналов. Поскольку эти сигналы приводят в результате к росту и делению клеток, нарушение регуляции Ras сигнализации в конечном итоге может привести к онкогенезу и раку. Активирующие мутации в Ras находятся в 20-25% всех опухолей человека и до 90% в конкретных типах опухолей.
KRAS, онкоген Кирстен Ras гомолога млекопитающих семейства генов Ras, кодирует белок, который является членом небольшого суперсемейства GTPaз. Одна аминокислотная замена несет ответственность за активирующую мутацию. Трансформирующий белок, который получают в результате, участвует в различных злокачественных новообразованиях, включая аденокарциному легких, муцинозную аденому, карциному эпителия протоков поджелудочной железы и колоректальную карциному. Альтенативный сплайсинг приводит к вариантам, кодирующим две изоформы, которые отличаются в С-концевой области. Каждый вариант гена KRAS может быть мишенью sd-PHK. Репрезентативный пример человеческого транскрипта KRAS представляет собой Genbank номер доступа NM_004985.3, и соответствующий белок представляет собой NP_04976.2.
HRAS, v-HA-ras вирусный гомолог онкогена саркомы крыс Харви млекопитающих семейства генов ras, кодирует белок, который подвергается непрерывному циклу де- и повторного пальмитоилирования, который регулирует его быстрый обмен между плазматической мембраной и аппаратом Гольджи. Мутации в этом гене вызывают синдром Костелло, заболевание, характеризующеемя увеличением роста на пренатальной стадии, дефицитом роста на послеродовой стадии, предрасположенностью к образованию опухолей, умственной отсталостью, нарушениями кожи и опорно-двигательного аппарата, характерной внешностью и сердечно-сосудистыми нарушениями. Дефекты этого гена вовлечены в различные виды рака, включая рак мочевого пузыря, фолликулярный рак щитовидной железы и плоскоклеточную карциному ротовой полости. Несколько вариантов транскриптов, которые кодируют разные изоформы, были определены для этого гена. Каждый вариант транскрипта может быть мишенью sd-PHK. Репрезентативный пример человеческого транскрипта HRAS представляет собой Genbank номер доступа NM_001130442.1, и соответствующий белок представляет собой NP_001123914.1.
RAF прото-онкогенная серин/треонин-протеинкиназа также известна как прото-онкоген c-RAF или просто c-Raf и является ферментом, который у человека кодируется геном RAF1. c-Raf белок функционирует в MAPK/ERK пути передачи сигнала как часть каскада протеинкиназ. c-Raf является членом семейства Raf серин/треонин-специфичных протеинкиназ, и МАР-киназы киназы киназы (МАР3К), которая функционирует ниже подсемейства Ras мембраны связанной GTPaз, с которыми она связана непосредственно. После активации Raf-1 может фосфорилировать для активации двойной специфичности протеинкиназ МЕК1 и МЕК2, которая, в свою очередь, фосфорилирует, чтобы активировать серин/треонин-специфичные протеинкиназы ERK1 и ERK2. Активированные ERKs являются плейотропными эффекторами физиологии клетки и играют важную роль в регуляции экспрессии генов, участвующих в цикле деления клеток, апоптозе, дифференциировании клеток и миграции клеток. Любая одна или несколько из c-Raf (RAF1), МЕК1, МЕК2, ERK1 и ERK2 могут быть мишенями sd-PHK. Репрезентативный пример человеческого RAF1 транскрипта представляет собой NM_002880.3, и соответствующий белок представляет собой NP_00287.1.
Митоген-активированная протеинкиназа 1 (МАРК1) (также известная как ERK, р38, р40, р41, ERK2, ERT1, МАРК2, PRKM1, PRKM2, Р42МАРК, или p41mapk) кодирует член семейства MAP киназ. MAP киназы, также известные как внеклеточные сигнал-регулируемые киназы (ERK), выступают в качестве точки интеграции для нескольких биохимических сигналов, а также участвуют в различных клеточных процессах, таких как пролиферация, дифференциация, регуляция транскрипции и развитие. Активация данной киназы требует ее фосфорилирования верхними киназами. После активации эта киназа транслоцируется к ядру стимулированных клеток, где она фосфорилирует ядерные мишени. Два альтернативно сплайсированных варианта транскриптов, кодирующих тот же белок, но различающихся по UTR, были зарегистрированы для этого гена. Каждый вариант транскрипта МАРК1 может быть мишенью sd-PHK. Репрезентативный пример человеческого МАРК1 транскрипта представляет собой NM_002745.4, и его соответствующий белок представляет собой NP_002736.3.
C-abl онкоген 1, не-рецепторная тирозинкиназа (ABL1), кодирует цитоплазматические и ядерные протеинтирозинкиназы, которые были вовлечены в процесы дифференциации клеток, деления клеток, клеточной адгезии и стрессовой реакции. Активность с-Abl белка негативно регулируется с помощью его SH3 домена и делеция SH3 домена направляет ABL1 в онкоген. t(9;22) транслокация приводит к голова к хвосту слиянию BCR (MIM:151410) и ABL1 гены присутствуют во многих случаях хронической миелогенной лейкемии. ДНК-связывающая активность повсеместно экспрессированной ABL1 тирозинкиназы регулируется CDC2-опосредованным фосфорилированием, предлагая функции клеточного цикла для ABL1. Ген ABL1 экспрессируется либо в виде 6- или 7- kb мРНК транскрипта, с альтернативно сплайсированным первым экзоном, сплайсированным с общими экзонами 2-11. Каждый вариант транскрипта ABL1 может быть мишенью sd-PHK. Репрезентативный пример человеческого транскрипта ABL1 представляет собой Genbank номер доступа NM_005057.4, и его соответствующий белок представляет собой NP_005148.2.
В-клеточная CLL/лимфома 2 (Bcl-2) кодирует неотъемлемый внешний белок митохондриальной мембраны, который блокирует апоптоз некоторых клеток, таких как лимфоциты. Конститутивная экспрессия BCL2, например, в случае транслокации BCL2 в Ig локус тяжелой цепи, считается причиной фолликулярной лимфомы. Два варианта транскрипта, производимые поочередным сплайсингом, различаются их С-терминальными концами, каждый из которых может быть мишенью sd-PHK. Репрезентативный пример человекого Bcl-2 транскрипта представляет собой NM_000633.2, и его соответствующий белок представляет собой NP_00624.2.
V-src онкогенный гомолог вирусной саркомы (SRC) очень похож на v-src ген вируса саркомы Рауса. Этот прото-онкоген может играть определенную роль в регуляции эмбрионального развития и роста клеток. Белок, кодируемый этим геном, является тирозин-протеинкиназой, чья активность может быть ингибирована путем фосфорилирования при помощи c-SRC-киназы. Мутации в этом гене могут быть вовлечены в злокачественную прогрессию рака толстой кишки. Два варианта транскриптов кодирующих тот же белок были найдены для этого гена, каждый из которых может быть мишенью sd-PHK. Репрезентативный пример человеческого транскрипта SRC представляет собой NM_005417.3, и его соответствующий белок представляет собой NP_005408.1.
Механистическая мишень рапамицина (серин/треонин киназа) (MTOR) кодирует белок, принадлежащий к семейству фосфатидилинозитол киназа-связанных киназ. Эти киназы опосредуют клеточные реакции на стрессы, такие как повреждение ДНК и лишение питания. Этот белок выступает в качестве мишени для остановки клеточного цикла и иммуносупрессивных эффектов FKBP12-рапамицин комплекса. Репрезентативный пример человеческого транскрипта MTOR представляет собой NM_004958.3, и соответствующий белок представляет собой NP_004949.1.
Протеинкиназа С (РКС) кодирует семейство ферментов, которые участвуют в контроле функции других белков через фосфорилирование гидроксильных групп сериновых и треониновых аминокислотных остатков на этих белках. РКС ферменты, в свою очередь активируются сигналами, такими как увеличение концентрации диацилглицерина или Са2+. Поэтому РКС ферменты играют важную роль в нескольких каскадах передачи сигнала. РКС семейство состоит из приблизительно десяти изоферментов. Они делятся на три подсемейства, на основе требований к своему второму мессенджеру: традиционные (или классические), новые и нетипичные. Традиционные (с)РКС содержат изоформы, α, βI, βII и γ. Они требуют Са2+, диацилглицерин (DAG) и фосфолипид, такой как фосфатидилсерин, для активации. Новые (n)РКС включают δ, ε, η, и θ изоформы, и требуют DAG, но не требуют Са2+ для активации. Таким образом, традиционные и новые РКС активируется через те же пути передачи сигнала, что и фосфолипаза С. С другой стороны, атипичная (а)РКС (включая протеинкиназу Мζ и υ/λ изоформы) не требуют ни Са2+, ни диацилглицерина для активации. Термин «протеинкиназа С» относится ко всему семейству изоформ. Любой один или несколько традиционных, новых и атипичных генов РКС может быть мишенью sd-PHK. Репрезентативный пример человеческого РКС транскрипта представляет собой NM_005400.2, и его соответствующий белок представляет собой NP_005391.1.
Бакуловирусый повтор IAP содержит 5 (BIRC5) (также известен как API4 или EPR-1) и является членом семейства генов ингибиторов апоптоза (IAP), которые кодируют негативные регуляторные белки, которые препятствуют апоптозу клеток. Члены семейства IAP обычно содержат несколько доменов бакуловирусных IAP повторов (BIR), но этот ген кодирует белки только с одним доменом BIR. Кодируемые белки также не содержат С-конец RING домен отпечатков пальцев. Экспрессия генов является высокой в период внутриутробного развития и в большинстве опухолей и еще низкой во взрослых тканях. Антисмысловые транскрипты участвуют в регуляции экспрессии этого гена. По меньшей мере четыре варианта транскриптов, кодирующих различные изоформы, были найдены для этих генов, каждый из которых может быть мишенью sd-PHK. Репрезентативный пример человеческого BIRC5 транскрипта представляет собой NM_001012270.1, и его соответствующий белок представляет собой NP_001012270.1.
Fas (надсемейство TNF рецепторов, член 6) (FAS, также известный как АРТ1, CD95, FAS1, АРО-1, FASTM, ALPS1A или TNFRSF6) кодирует член TNF-рецепторного надсемейства. Этот рецептор содержит домен смерти. Было показано, что он играет центральную роль в физиологической регуляции запрограммированной клеточной смерти, и был вовлечен в патогенез различных злокачественных опухолей и заболеваний иммунной системы. Взаимодействие этого рецептора с лигандом позволяет формировать смерть-вызывающие сигнализации комплекса, который включает Fas-связанный белок домена смерти (FADD), каспазу 8 и каспазу 10. Аутопротеолитическая обработка каспаз в комплексе запускает вниз каскад каспаз, и приводит к апоптозу. Этот рецептор был также показан для активации NF-kappaB, MAPK3/ERK1 и MAPK8/JNK, и участвует в трансдукции пролиферирующих сигналов в нормальных фибробластах диплоидных и Т-клеток. Несколько альтернативно сплайсированных вариантов транскриптов были описаны, некоторые из которых являются кандидатами для несмыслово-опосредованного распада мРНК (NMD). Изоформы, которые не содержат трансмембранный домен, могут негативно регулировать апоптоз при посредничестве полноразмерной изоформы. Каждый вариант транскрипта может быть мишенью sd-PHK. В некоторых случаях, sd-PHK мишенью является FADD, каспаза 8, и/или каспаза 10. В других случаях, sd-PHK мишенью является NF-kappaB, MAPK3/ERK1 и/или MAPK8/JNK. Репрезентативный пример человеческого BIRC5 транскрипта представляет собой NM_001012270.1, и его соответствующий белок представляет собой NP_001012270.1.
Гипоксия индуцибельный фактор 1, альфа-субъединица (HIF1A), является фактором транскрипции, найденным в клетках млекопитающих, которые культивировали при пониженном давлении кислорода, который играет важную роль в клеточных и системных гомеостатических реакциях на гипоксию. HIF1 представляет собой гетеродимер, который состоит из альфа-субъединицы и бета-субъединицы. Бета-субъединица была определена как арилуглеводородный рецептор ядерного транслокатора (ARNT). Этот ген кодирует альфа-субъединицу HIF-1. Избыточная экспрессия природного антисмыслового транскрипта (aHIF) этого гена была показана как связанная с непапиллярной почечной карциномой. Два альтернативных транскрипта, кодирующие разные изоформы, выявлено не было. Каждый вариант транскрипта и/или природного антисмыслового транскрипта может быть мишенью sd-PHK. Репрезентативный пример человеческого транскрипта HIF1A представляет собой NM_001530.3, и его соответствующий белок представляет собой NP_001521.1.
Кадгерин 16, KSP-кадгерин (CDH16) является членом надсемейства кадгеринов, генов, кодирующих кальций-зависимые, ассоциированные с мембранами гликопротеины. Картированный с ранее выявленным кластером кадгериновых генов в хромосоме 16q22.1, ген локализуется с членами надсемейства CDH1, CDH3, CDH5, CDH8 и CDH11. Белок состоит из внеклеточного домена, содержащего 6 кадгериновых доменов, трансмембранной области и усеченного цитоплазматического домена, но не содержит пропоследовательность и трипептид адгезия HAV распознающую последовательность, которая типична для большинства классических кадгеринов. Экспрессия происходит исключительно в почках, где белок функционирует как основной посредник распознавания гомотипичных клеток, играет роль в морфогенетическом направлении развития тканей. Альтернативно сплайсированные транскриптные варианты, кодирующие различные изоформы были определены, каждый из которых может быть мишенью sd-PHK. Репрезентативный пример человеческого CDH16 транскрипта представляет собой NM_004062.3, и его соответствующий белок представляет собой NP_004053.1.
Катенин (кадгерин-связанный белок), бета 1 (CTNNB1) кодирует белок, который является частью комплекса белков, которые составляют соединения адгеренов (AJs). AJs необходимы для создания и поддержания эпителиальных клеточных слоев, регулируя клеточный рост и адгезию между клетками. Кодируемый белок также фиксирует актиновый цитоскелет и может быть ответственным за передачу сигнала контактного ингибирования, что заставляет клетки прекращать деление после завершения эпителиальной выстилки. Этот белок связывается с продуктом гена АРС, который мутирует в аденоматозный полипоз толстой кишки. Мутации в этом гене являются причиной колоректального рака (КРР), пиломатриксомы (PTR), медуллобластомы (MDB), и рака яичников. Три варианта транскриптов, кодирующих тот же белок, были найдены для этого гена, каждый из которых может быть мишенью sd-PHK. Репрезентативный пример человеческого транскрипта CTNNB1 представляет собой NM_001098209.1, и его соответствующий белок представляет собой NP_001091679.1.
V-myc миелоцитоматоз вирусного гомолога онкогена (МУС) кодирует многофункциональный ядерный фосфопротеин, который играет важную роль в прогрессии клеточного цикла, апоптозе и клеточной трансформации. Он функционирует как транскрипционный фактор, который регулирует транскрипцию специфических генов-мишеней. Мутации, избыточная экспрессия, перестановка и перемещение этого гена были связаны с различными гематопоэтическими опухолями, лейкемией и лимфомами, включая лимфому Беркитта. Существует доказательство того, что альтернативные инициации трансляции сверху, в рамке, не-AUG (CUG) и ниже AUG стартового сайта приводят в результате к получению двух изоформ с различными N-концами. Синтез не-AUG инициированного белка подавляется при лимфоме Беркитта, предлагая свое значение в нормальной функции этого гена. Каждый вариант транскрипта, включая мутантные варианты, может быть мишенью sd-PHK. Репрезенативный пример человеческого МУС транскрипта представляет собой NM_002467.4, и его соответствующий белок представляет собой NP_002458.2.
Дальнейшие аспекты настоящего изобретения относятся к доставке sd-PHK в нервную систему. Например, sd-PHK может быть доставлена в мозг или спинной мозг. Любой соответствующий механизм доставки для доставки sd-PHK в головной или спинной мозг может быть применен. В некоторых осуществлениях, доставка в головной или спинной мозг происходит путем инфузии, интратекальной доставки, паренхиматозной доставки, внутривенной доставки или прямой инъекции в головной или спинной мозг. В некоторых осуществлениях, sd-PHK доставляется в конкретную область мозга. sd-PHK может быть модифицирована или сформулирована, чтобы надлежащим образом проходить через гематоэнцефалический барьер. В других осуществлениях sd-PHK вводится таким образом, что не нужно пересекать гематоэнцефалический барьер.
В некоторых осуществлениях, sd-PHK доставляется с помощью насоса или катетерной системы в головной или спинной мозг. Примеры такой доставки включены путем ссылки на патент США №6,093,180 (Elsberry). Методы инфузии лекарственных препаратов в мозг также включены путем ссылки на патент США №5,814014 (Elsberry et al.).
Аспекты настоящего изобретения относятся к использованию sd-PHK в лечении расстройств, влияющих на нервную систему. В некоторых осуществлениях, sd-PHK используют для лечения нейродегенеративных расстройств. Как используется в данной заявке, термин «нейродегенеративное расстройство» относится к расстройствам, заболеваниям или состояниям, вызванным ухудшением клеточных и тканевых компонентов нервной системы. Некоторые не ограничивающие примеры нейродегенеративных расстройств включают инсульт, болезнь Альцгеймера, болезнь Паркинсона, болезнь Хантингтона, перивентрикулярную лейкомаляцию (PVL), боковой амиотрофический склероз (ALS, «болезнь Лу Герига»), ALS-Паркинсон-деменция комплекс Гуама, атаксию Фридриха, болезнь Вильсона-Коновалова, рассеянный склероз, церебральный паралич, прогрессирующий супрануклеарный паралич (Стил-Ричардсона синдром), бульбарный и псевдобульбарный паралич, диабетическую ретинопатию, мульти-инфарктную деменцию, дегенерацию желтого пятна, болезнь Пика, болезнь диффузных телец Леви, прионные заболевания, такие как болезнь Крейтцфельда-Якоба, болезнь Герштманна-Штраусслера-Шейнкера, Куру и фатальную семейную бессонницу, первичный боковой склероз, дегенеративные атаксии, болезнь Мачадо-Джозефа/спиноцеребеллярную атаксию типа 3 и оливомостомозжечковые вырождения, спинную и спинобульбарную мышечную атрофию (болезнь Кеннеди), семейную спастическую параплегию, болезнь Вольфарта-Кугельберга-Веландера, болезнь Тея-Сах, мультисистемную дегенерацию (синдром Шу-Дагера), болезнь Жиля де ля Туретта, семейную дизавтоновмию (синдром Рилея-Дея), болезнь Кугельберг-Веландера, подострый склерозирующий панэнцефалит, болезнь Верднига-Гоффмана, синуклеопатии (включая множественную системную атрофию), болезнь Сандхоффа, корковую базальную дегенерацию, спастический парапарез, первичную прогрессивную афазию, прогрессирующую мультифокальную лейкоэнцефалопатию, стриатонигральную дегенерацию, семейные спастические заболевания, хронические эпилептические состояния, связанные с нейродегенерацией, болезнь Бинсвангера и деменции (включая все основные этиологии деменции).
В некоторых осуществлениях, расстройство является болезнью Паркинсона, болезнью Хантингтона или ALS. В определенных осуществлениях, расстройство представляет собой ALS и sd-PHK направлена на SOD1, супероксиддисмутазу.
Нейродегенеративные расстройства также могут быть результатом повреждения головного мозга или травмы, включая то, что вызывает инсульт, повреждения головы или спинного мозга, или острого ишемического повреждения. Ишемические повреждения относятся к состояниям, которые возникают, когда мозг получает недостаточное кровообращение. В некоторых осуществлениях, повреждение головного мозга или нервной системы может возникнуть в результате травмы, или может быть результатом инфекции, радиации, химических или токсичных веществ. Травма в мозгу и нервной системе, которая может быть диффузной или локализованной, включает внутричерепное или интравертебральное поражение или кровоизлияние, церебральную ишемию или инфаркт, включая эмболическую окклюзию и тромботическую окклюзию, перинатальное гипоксически-ишемическое повреждение, травму шейного отдела позвоночника, потрясенный детский синдром, реперфузию при острой ишемии, или остановку сердца.
В одном осуществлении, in vitro лечение клеток олигонуклеотидами может быть использовано для ex vivo терапии клеток, удаленных из субъекта (например, для лечения лейкемии или вирусной инфекции) или для лечения клеток, которые не происходят из субъекта, но должны быть введены субъекту (например, для устранения трансплантации экспрессии антигенов на клетках, которые должны быть пересажены субъекту). Дополнительно, in vitro лечение клеток может быть использовано при немедицинских параметрах, например, для оценки функции генов, изучения регуляции генов и синтеза белков или для оценки улучшения, внесенного в олигонуклеотиды, предназначенные для модуляции экспрессии генов и синтеза белка. In vivo лечение клеток может быть полезно в определенных клинических условиях, где желательно подавлять экспрессию белка. Существует множество медецинских состояний, для которых антисмысловая терапия, как сообщается, является приемлемой (см., например, патент США номер 5,830,653), а также респираторно-синцитиальная вирусная инфекция (WO 95/22,553), вирус гриппа (WO 94/23,028) и злокачественные новообразования (WO 94/08,003). Другие примеры клинических применений антисмысловых последовательностей рассматриваются, например, в Glaser. 1996. Genetic Engineering News 16:1. Илюстративные мишени для расщепления олигонуклеотидов, включают, например, протеинкиназу Ca, ICAM-1, C-Raf киназу p53, с-MYB, и BCR/ABL слияния генов, найденных при хронической миелоидной лейкемии.
Рассматриваемые нуклеиновые кислоты могут быть использованы в РНКi терапии в любом животном, имеющем РНКi маршрут, например, человеке, нечеловеческом примате, нечеловеческом млекопитающем, нечеловеческих позвоночных, грызунах (мышах, крысах, хомяках, кроликах и т.д.), домашнем скоте, домашних животных (котах, собаках, и т.д.), Xenopus, рыбах, насекомых (Drosophila, и т.д.), и червях (C. elegans), и т.д.
Настоящее изобретение относится к способам ингибирования или предотвращения у субъекта, заболевания или состояния, связанного с аномальной или нежелательной экспрессией гена-мишени или активности, путем введения субъекту нуклеиновой кислоты в соответствии с настоящим изобретением. При необходимости, субъектов сначала лечат сенсибилизирующим агентом таким образом, чтобы более чутко реагировать на последующее лечение РНКi. Субъекты, имеющие риск заболевания, которое причинило или содействовало аберрантной или нежелательной экспрессии гена-мишени или активности, могут быть определены, например, любым или комбинацией диагностических и прогностических анализов, известных в данной области техники. Введение профилактического агента может произойти до проявления симптомов, характерных для аберрации гена-мишени, таким образом, чтобы заболевание или расстройство предотвратить или, альтернативно, задержать их развитие. В зависимости от типа аберрации гена-мишени, например, ген-мишень, агонист гена-мишени или агент антагонист гена-мишени может быть использован для лечения данного субъекта.
В другом аспекте настоящее изобретение относится к способам модуляции экспрессии гена-мишени, экспрессии или активности белка для терапевтических целей. Соответственно, в иллюстративом осуществлении способы настоящего изобретения включают контактирование клетки, способной экспрессировать ген-мишень, с нуклеиновой кислотой в соответствии с настоящим изобретением, которая является специфичной для гена-мишени или белка (например, является специфичной для мРНК, кодируемой указанным геном или указанной аминокислотной последовательностью указанного белка) так, что модулируется экспрессия или один или более видов активности белка-мишени. Эти способы могут быть выполнены in vitro (например, путем культивирования клетки с агентом), in vivo (например, путем введения вещества субъекту), или ex vivo. Субъекты могут быть сначала пролечены сенсибилизирующим агентом, таким образом, чтобы более чутко реагировать на последующее лечение РНКi, если это необходимо. Таким образом, настоящее изобретение обеспечивает способы лечения пациента, страдающего заболеванием или растройством, характеризующимся аберрантной или нежелательной экспрессией или активностью гена-мишени полипептида или молекулы нуклеиновой кислоты. Ингибирование активности гена-мишени желательно в ситуациях, в которых ген-мишень является аномально нерегулируемым и/или в которых снижение активности гена-мишени может иметь положительный эффект.
Таким образом, терапевтические агенты в соответствии с настоящим изобретением можно вводить субъектам для лечения (профилактического или терапевтического) расстройств, связанных с аномальной или нежелательной активностью гена-мишени. В связи с таким лечением, фармакогеномика (например, изучение взаимосвязи между генотипом индивида и ответом индивида на чужеродные соединения или лекарственные препараты) может быть рассмотрена. Различия в метаболизме терапевтических средств могут привести к серьезной токсичности или терапевтической неудаче, изменяя соотношение между дозой и концентрацией в крови фармакологически активного лекарственного препарата, Таким образом, врач или клиницист может рассмотреть вопрос о применении знаний, полученных в соответствующих исследованиях фармакогеномики, при определении того, вводить ли терапевтический агент, а также определении дозировки и/или терапевтической схемы лечения терапевтическим агентом. Фармакогеномика имеет дело с клинически значимыми наследственными изменениями в ответ на лекарственные препараты в связи с измененным расположением лекарственных препаратов и аномальной активностью у пострадавших лиц.
Для целей настоящего изобретения, диапазон может быть выражен в данной заявке как от «приблизительно» одного конкретного значения, и/или до «приблизительно» второго конкретного значения. Если такой диапазон выражается, другое осуществление включает от одного конкретного значения и/или до другого конкретного значения. Аналогичным образом, когда значения выражаются в приближении, при использовании предшествующих «приблизительно», будет подразумеваться, что конкретное значение образует другое осуществление. Дополнитлеьно будет понятно, что концевые точки каждого из диапазонов являются значимыми как по отношению к другой конечной точке, так и независимо к другой конечной точке.
Дополнительно, для целей настоящего изобретения единственное число относится к единственному или множественному числу; например, «белок» или «молекула нуклеиновой кислоты» относится к одному или более из этих соединений или по меньшей мере одному соединению. Таким образом, «один или более» и «по меньшей мере один» могут быть использованы в данной заявке взаимозаменяемо. Следует также отметить, что термины «содержащий», «включающий» и «имеющий» могут быть использованы как взаимозаменяемые. Дополнительно, соединение «выбранное из группы, состоящей из», относится к одному или более соединениям в списке, который следует, включая смеси (например, комбинации) двух или более соединений. В соответствии с настоящим изобретением, выделенный, или биологически чистый белок или нуклеиновая кислота представляет собой соединение, которое было удалено из своей природной среды. Таким образом, «выделенный» и «биологически чистый» не обязательно отражает степень, в которой соединение было очищено. Выделенное соединение в соответствии с настоящим изобретением может быть получено из природного источника, может быть получено с использованием методов молекулярной биологии или может быть получено путем химического синтеза.
Настоящее изобретение далее иллюстрируется следующими примерами, которые никоим образом не должны рассматриваться как дополнительное ограничение. Все содержания всех ссылок (включая ссылки на литературу, выданные патенты, опубликованные патентные заявки, и совместно рассматриваемые заявки на патенты), приведенные в данной заявке, специально включены путем ссылки.
ПРИМЕРЫ
Пример 1: Системная доставка sd-PHK посредством внутривенного введения
sd-PHK молекулы были эффективно системно доставлены в отдаленные ткани-мишени, такие как печень и сердце, с использованием внутривенного введения. Системная доставка привела к эффективному поглощению соединения и сайленсинге в тканях-мишенях. Как показано на ФИГ. 1, клиренс крови sd-PHK молекулы зависит от концентрации молекул. Периоды полураспада sd-PHK молекул, доставленных при 10 мг/кг или 50 мг/кг показаны на ФИГ. 1. sd-PHK была обнаружена в печени через 6 часов, но существенный сигнал не был обнаружен через 24 часа.
Ежедневные режимы дозирования были протестированы для внутривенной доставки sd-PHK, мишенью которой являлась МАР4К4 и сайленсинг МАРК4К в печени был определен (ФИГ. 2). Самцы мышей BALB/c6J, 6-8 недельного возраста, были использованы, по 5 мышей в каждой тестовой группы. 50 мг/кг sd-PHK вводили через хвостовую вену путем инъекции ежедневно в течение 3 дней, а затем пробы были собраны на 4 день.
sd-PHK молекулы с поколением I и II химии были проверены, включая активные молекулы и несоответствие или нецелевой контроль. Экспрессия МАР4К4 была определена QPCR и нормирована на экспрессию гена домашнего хозяйства. Экспрессия также была нормирована по отношению к контрольной PBS-группе, получавшей лечение.
Одной из целей данной работы было определение того, усиливают ли повторяющиеся дозировки в течение 24-часового периода поглощение в печени. Примеры повторяющихся режимов дозирования проверяли на sd-PHK доставке и они приведены на ФИГ. 3. sd-PHK (мишенью которой является МАР4К4) вводили 1-4 раза в течение 24-часового периода. sd-PHK молекулы вводили путем внутривенной инъекции в хвост при 50 мг/кг на инъекцию. Пробы были проанализированы в нескольких моментах времени, включая 6 часов после последней дозы и 30 часов после первой дозы. Конфокальная микроскопия и анализ гибридизации были выполнены. Концентрацию в крови определяли путем гибридизационного анализа. Пробы анализировали в нескольких моментах времени, включая 24 часа после последней дозы.
ФИГ. 4 демонстрирует поглощение в печени после 1-4 доз sd-PHK. Мышам вводили дозы sd-PHK 1, 2, 3 или 4 раза по 50 мг/кг в течение 24 часов. Изображения были получены с использованием 20-кратного объектива. Улучшенное поглощение в печени была отмечено с несколькими дозами в течение 24 часов. Выраженной токсичности не было отмечено. Важно отметить, что эффективная доставка была также отмечена в селезенке и сердце после нескольких доз sd-PHK в течение 24-часового периода (ФИГ. 5). Повторяющееся дозирование привело к увеличению поглощения в печени sd-PHK без накопления в крови (ФИГ. 6).
Фигура 37 также демонстрирует эффективную системную sd-PHK доставку в печень. BALB/c мыши-самцы (n=4/группа) получили 100 мг/кг инъекций в хвостовую вену sd-PHK один раз в день в течение 3 дней. На 4-й день, 3 мм биопсии было собрано для получения РНК. Генная экспрессия была проанализирована QPCR, по сравнению с PBS, нормированной на ген домашнего хозяйства. Статистически значимый сайленсинг генов-мишеней наблюдается в печени, согласно измерениям qPCR через 24 часа после последней дозы. Эти данные демонстрируют потенциал sd-PHKs для использования в системных клинических показаниях. Предварительная доза и кровотечения из конечностей были также собраны для ALT/AST уровней. Важно отметить, что системное введение sd-PHKs не приводит к повышенной функции ферментов печени, например, ALT и AST.
Пример 2: подкожное введение sd-PHK
Эффективная доставка sd-PHK молекул была также достигнута путем подкожной доставки. В условиях, описанных в данной заявке, равные дозы введенных подкожно sd-PHK молекул привели к более высокому sd-PHK присутствию в тканях-мишенях, чем достигается путем внутривенного введения. ФИГ. 7 показывает сравнение подкожного и внутривенного введения для эффективности в печени. Для внутривенного и подкожного введения, вводили один болюс 80 мг/кг sd-PHK. Уровень в крови определяли с помощью гибридизационного анализа и флуоресцентная визуализация была проведена с помощью конфокальной микроскопии.
Маршрут введения, как было установлено, имел вляние на кинетику клиренса, уровни в крови и поглощение в тканях (ФИГ. 8). Клиренс соединения (обозначенной розовым мочи) был обнаружен в течение 30 минут после внутривенного введения и в течение 6-8 часов после подкожного введения. Концентрация в крови увеличивается в течение по меньшей мере 4 часов после подкожной инъекции. Подкожное введение привело к увеличению sd-PHK уровней в печени в течение 24 часов, по сравнению с внутривенным введением (ФИГ. 9-10).
sd-PHK также эффективно была доставлена в кожу с помощью подкожного введения. Одна подкожная инъекция привела к sd-PHK доставке в месте инъекции. Наличие sd-PHK экспрессии было обнаружено через 24 часа после инъекции (ФИГ. 11). Подкожная площадь кожи в месте инъекции сохранила целостность в течение по меньшей мере 24 часов после инъекции и sd-PHK была обнаружена во всех клетках (ФИГ. 12).
Пример 3: дыхательные применения sd-PHK
sd-PHK молекулы, как было обнаружено, эффективно доставляются в различные типы клеток в легких путем инсуффляции. ФИГ. 13-14 раскрывает эффективную доставку sd-PHK для мышиных легких при 1-44 мг/кг и легких крыс при 1-10 мг/кг. sd-PHK, предпочтительно, поглощается альвеолярными макрофагами. В тканях мыши, флуоресцентный сигнал был обнаружен в основном в макрофагах при до 20 мг/кг, а в тканях и макрофагах при 44 мг/кг. ФИГ. 15 раскрывает эффективную доставку sd-PHK соединений в альвеолярные макрофаги. Идентификация типа клеток была основана на визуализации. Некоторая доставка в другие типы клеток наблюдались, но в проанализированных условиях, не наблюдали видимой доставки в мерцательный эпителий.
Визуализация sd-PHK-DY547, доставленной инсуффляцией в выделенные макрофаги легких, показана на ФИГ. 16. BALB/C легкие мышей-самцов были измельчены в фарш и инкубированы в коллагеназе в течение 30 минут. Клетки пропускали через 70 мкм фильтр, промывали в PBS и высевали в 10 см тарелки в DMEM. Через 3 часа, клетки промывали для удаления тромбоцитов. Животных наблюдали после дозировок и они не проявляли вялость или обезвоживание (ФИГ. 17). Легкие были нормальными и здоровыми.
Способы
Процедура трахеальной инсуффляции
Животных анестезировали при помощи аэрозоля изофлурана (5% при индукции с 2 л/м потока О2) через носовой конус. Животные лежали, грудной клеткой вверх. Грудная клетка была поднята приблизительно под углом 45° путем размещения рулона марли под шеей. Источник света, оснащенный гибким волоконно-оптическим рычагом, был отрегулирован, чтобы обеспечить оптимальную освещенность трахеи. Небольшая стерильная ватная палочка была использована, чтобы открыть нижнюю челюсть животного и затупленные щипцы были использованы, чтобы помочь вытеснить язык для максимального орофарингеального воздействия. После получения четкого вида трахеи, MicroSprayer (Penn Century MSA-250) наконечник был вставлен эндотрахеально и максимальный объем 50 мл (мышам) или 250 мл (крысам) из ТА (тестовое изделие) было доставлен с помощью распылителя вниз от трахеи в легочное пространство. После того, как дозирование было достигнуто, наконечник был немедленно отведен и животное было выведено из наркоза. Стетоскоп был использован, чтобы подтвердить наличие жидкости в легких, и животное было перенесено в чистую клетку, помещенную на грелку для наблюдения и восстановления. Период наблюдения после дозировок стоставлял не менее 1 часа, после введения дозы,
Самцам крыс SD вводили путем легочной инсуффляции дозу 250 мкл DY547 меченой sd-PHK (олигонуклеотид номер 13766.3) при 0,1-10 mpk. Образцы были собраны для визуализации через 24 или 48 часов. При 5 и 7,7 дозах mpk, значительное sd-PHK поглощение в легких наблюдается, включая предпочтительную доставку соединения в альвеолярные макрофаги. При некоторых условиях испытаний, уровни дозы 5-7,5 mpk были найдены предпочтительными.
Пример 4: Идентификация sd-PHK последовательностей, мишенями которых являются hApoB, PCSK9 и PPIB
Оптимальные последовательности в АроВ, PCSK9 и PPIB для sd-PHK развития были определены с использованием алгоритма выбора последовательности. Алгоритм выбора последовательностей основан на следующих критериях: GC содержание более 32%, но менее 47%, гомология к конкретным животным моделям (например, мышей или крыс), избегание 5 и более U/U участков и/или 2 или более G/C участков, не попадание в цель со счетом меньше чем 500, и избегание последовательностей, содержащихся в 5' UTR.
Последовательности были разработаны изначально как 25 нуклеотидные дуплексы с тупыми концами с О-метил модификацией. Эти последовательности были показаны в различных клеточных линиях для выявления тех, которые были наиболее эффективными в снижении экспрессии генов. Несколько концентраций РНК молекул, таких как 0,025, 0,1 и 0,25 нм, были протестированы, и приемлемые концентрации были определены. Полный скрининг был затем проведен при желаемой концентрации. Кривые доза-ответ были созданы с целью определения наиболее эффективных последовательностей. Попадания рассматривали как гиперфункциональные, если они имели ЕС50 значения менее чем 100 пМ при липидной трансфекции. Оптимальные последовательности затем были разработаны в sd-PHK молекулах на основе параметров, описанных в данной заявке. sd-PHKs были использованы для вторичного скрининга.
Таблица 1 отображает последовательности, определенные в АроВ с помощью алгоритма, как описано в данной заявке выше. Представлены данные процента экспрессии для каждой последовательности, указывающие на эффективность при достижении генного сайленсинга, при использовании в концентрации 0,1 нМ. В Таблице 2 представлены АроВ sd-PHK последовательности. В Таблице 3 представлены PCSK9 sd-PHK последовательности и в Таблице 4 представлены PPIB sd-PHK последовательности.
Пример 5: Химия линкера
Фигура 36 показывает, что изменение химии линкера не влияет на активность сайленсинга sd-PHKs in vitro. Две различные химии линкера были оценены, гидроксипролиновый линкер и рибо линкер, на множестве sd-PHKs (мишенью которых является Map4k4 или PPIB) в анализах пассивного поглощения, чтобы определить линкеры, которые способствуют самостоятельной доставке. HeLa клетки трансфицировали в отсутствие основы доставки (пассивная трансфекция) с sd-PHKs при 1 мкМ, 0,1 мкМ или 0,01 мкМ в течение 48 часов. Использовали оба результата линкеров при эффективной доставке sd-PHK.
Рибо линкер, который использовали в Примере 5, имел следующую структуру:

Пример 6: Химическая оптимизация sd-PHKs для оптимальной доставки
Влияние химической модификации на стабильность и системную доставку sd-PHK было оценено. Фигура 38 представляет не ограничивающие примеры модифицированных последовательностей для АроВ и PPIB sd-PHKs. Примечательно, что путем обширных исследований моделей химической модификации, было установлено, что повышение стабильности ведущей нити путем химических модификаций приводит к увеличению системной эффективности. В примерах, приведенных на Фигуре 38, АроВ sd-PHK была сильнее модифицирована по сравнению с последовательностью PPIB (12 2'F по сравнению с 8, соответственно). Дополнительно, последовательность PPIB содержала участок > 3 2'ОН в ряд, потенциальный сайт для эндонуклеаз. Увеличенная схема модификации АроВ sd-PHK придает стабильность соединению и обеспечивает максимальное количество соединения, которое поступает в рассматриваемую мишень, которая в данном примере представляла собой печень. Увеличение стабильности олигонуклеотида, с помощью химической модификации 2'ОН (2'F или 2'ОМе), привело к увеличению сайленсинга после системного введения в печень.
В целях улучшения биораспределения sd-PHKs после системного введения, была исследована панель химических и структурных модификаций. Химию, известную, как придающую стабильность, использовали для всех sd-PHK вариантов химических структур; 2'ОМе, 2'дезокси и фосфоротиоатов. Как показано на Фигуре 39, повышение стабильности sd-PHK приводит к более чем 140-кратному улучшению sd-PHK распределения в печени по сравнению с обычной sd-PHK. (В некоторых случаях, химические варианты не являются активными соединениями в связи со схемами модификации, которые используют, чтобы значительно повысить стабильность) Было также отмечено, что изменение фосфоротиоатного содержания существенно влияет на распределение в печени.
Стабилизация sd-PHK вариантов также привела к распределению соединения в других тканях, такие как сердце и жир, как показано на Фигуре 40. В целом, повышение уровня стабилизирующих модификаций, как было найдено, приводит к: i) увеличенному сайленсингу (mApoB sd сайленсинг в печени) по сравнению с менее стабильными sd-PHK (PPIB); ii) увеличенному биораспределению в печени, а также распределению в других тканях, таких как сердце и жир.
Соединения III поколения
Целью создания нового поколения sd-PHK было модулирование PK/PD поведения sd-PHKs путем включения гидрофобных фрагментов в положении 5 уридинов. Увеличение гидрофобности, как известно, повышает клеточное поглощение (например, добавление холестерина в siPHK соединения). Таким образом, было высказано предположение, что включение гидрофобных фрагментов в sd-PHK может придать дополнительную стабильность при сохранении эффективной sd-PHK активности.
Фигура 41 показывает, что добавление октильных модификаций в ведущую нить sd-PHK, мишенью которой является Map4k4 привело в результате к увеличению времени удержания на 2 минуты или больше (19403-19405) по сравнению с немодифицированной ведущей нитью (12984). Этот сдвиг означает, что эти соединения являются более гидрофобными, чем немодифицированное соединение. В связи с увеличением гидрофобного характера пассажирской нити, включение фрагментов GIII в эту нить не приводит к изменениям времени удержания.
Фигура 42 показывает, что для PCSK9, октильные модификации не были хорошо переносимы, если они включены как в пассажирскую, так и ведущую нити одновременно. Фигура 43 показывает, что для PCSK9, октильные модификации переносятся в пассажирской нити (19719 и 20333) при образовании дуплекса с не-октил модифицированной ведущей нитью. Октильные модификации в ведущей нити не были хорошо переносимы, две октильные модифицированные ведущие нити продемонстрировали значительное снижение активности (19722 и 19723).
Фигура 44 показывает, что тиофеновые модификации были хорошо переносимы в пассажирской нити (20329 и 20333) при образовании дуплекса с не-тиофен модифицированной ведущей нитью. Тиофеновые модификации в ведущей нити не были хорошо переносимы. Только одна тиофеновая модифицированная ведущая нить продемонстрировала значительное снижение активности (20339).
Фигура 45 показывает, что октин-1-ильные модификации были хорошо переносимы в пассажирской нити (20344 и 20348) при образовании дуплекса с не-октин-1-ил модифицированной ведущей нитью. Октин-1-ильные модификации ведущей нити не были хорошо переносимы. Только одна октин-1-ильная модифицированная ведущая нить продемонстрировала значительное снижение активности (20354). Фигура 46 также показывает, что октин-1-ильные модификации ведущей нити не были хорошо переносимы для PCSK9 (20295).
Фигура 47 показывает, что этинильные модификации были хорошо переносимы в пассажирской нити (20344 и 20348) для МАР4К4 при образовании дуплекса с не-этинильно модифицированной ведущей нитью. Включение 2 этинильных модификаций в ведущую нить были переносимы и приводили лишь к незначительному снижению активности (20369 и 20370). Включение 3-этинильных модификаций в ведущую нить не было хорошо переносимо и приводило к значительному снижению активности (20368).
Фигура 48 показывает, что этинильные модификации не были переносимы в пассажирской нити (с 20298 по 20302) для PCSK9 при образовании дуплекса с модифицированной или не этинильно модифицированной ведущей нитью. Включение 2 этинильных модификаций в ведущую нить было переносимо и продемонстрировало лишь незначительное снижение активности (20309 и 20310).
Фигура 49 показывает, что для Map4k4, включение пиридил амидных модификаций не было переносимо в любой пассажирской или ведущей нити. Фигура 50 показывает, что сокращение количества пиридильных включений в смысловую нить приводило в результате к активному соединению (21428). Включение (2) пиридил амидов на 3' конце смысловой нити не снижает активности, однако, включение (2) пиридил амидов на 5' конце смысловой нити, в некоторых случаях, привело к полной потере активности (21427).
Фигура 51 показывает, что для PCSK9, включение пиридил амидных модификаций не было хорошо переносимо в любой пассажирской или ведущей нити. Уменьшение количества пиридильных включений в смысловой нити привело в результате к получению активных соединений (21424-21426).
Фигура 53 показывает, что для Map4k4, фенильные модификации были переносимы в пассажирской нити (21412) при образовании дуплекса с не-фенильно модифицированной ведущей нитью. Фенильные модификации в ведущей нити не были переносимы. Фигура 54 показывает, что фенильные модификации были переносимы в пассажирской нити (21399) при образовании дуплекса с не-фенильно модифицированной ведущей нитью. Фенильные модификации в ведущей нити не были хорошо переносимы. Только одна фенильная модификация в ведущей нити продемонстрировала активность (21405).
Фигура 55 предусматривает краткое описание данных 5-уридильной модификации. Было установлено, что переносимость 5-уридильных модификаций была последовательность-специфичной. Например, Map4k4 были более склонны к 5-уридильным модификациям, чем PCSK9. Для Map4k4, 5-уридильные модификации были лучше переносимы в смысловой нити, чем в антисмысловой нити. В общем, включение 5-уридильных модификаций в ведущей нити приводило к получению соединений с повышенной гидрофобностью, как видно по изменению времен удержания согласно HPLC.
Способы GIII скрининга
Через 48 часов после добавления sd-PHKs, HeLa клетки лизировали и мРНК урони количественно определяли при помощи анализа разветвленной ДНК (р-ДНК) (Affymetrix). Для определения относительной гидрофобности соединений GIII, использовали HPLC с обращенной фазой. Колонка: Hamilton PRP-1, 150×4,6 мм, 10 мкм), подвижная фаза А - 5 мМ фосфата калия, 2% ацетонитрила (рН 7,4). Подвижная фаза В - 85% ацетонитрила в H2O. В следующей таблице приведены репрезентативные результаты такого анализа:

Фигура 56 показывает, что модуляция химии в положении 5 уридина приводит к измененному профилю распределения в ткани печени. BALB/c мыши-самцы (n=2/группа) получали одну 50 мг/кг инъекцию в хвостовую вену sd-PHK. Поглощение было проанализировано гибридизационным анализом. Выводы из этого анализа были такими, что включение уридильных модификаций увеличивало уровни соединения в печени через 24 часа после введения дозы, включение тиофеновых модификаций увеличивало sd-PHK уровни в печени более чем в 10 раз, и октильные модификации как в пассажирской нити, так и в ведущей нити (без холестерина) приводили в результате к распределению sd-PHK в печени.
Аналогично, Фигура 57 демонстрирует, что модуляция химии в положении 5 уридина приводит в результате к измененному профилю распределение в тканях сердца и жире. BALB/c мыши-самцы (n=2/группа) получали одну 50 мг/кг инъекцию в хвостовую вену sd-PHK. Поглощение было проанализировано гибридизационным анализом. Выводы из этого анализа были такими, что включение октил уридильных модификаций увеличивало уровни sd-PHK в сердце и жире через 24 часа после введения дозы, и октильные модификации как в пассажирской нити, так и в ведущей нити (без холестерина) приводили в результате к распределению sd-PHK как в сердце, так и в жире через 24 часа после введения дозы.
Синтоны были разработаны на основе О-метил и 2'F. GIII химии, которые показали обещающие результаты, были выбраны, и 2'-дезокси химии была заменена на 2'-O метил и 2'-фтор. Несколько неограничивающих примеров синтонов приведены на Фигурах 58-60.
Эндосомальное высвобождение
Модификации в sd-PHK были также разработаны для содействия эндосомальному высвобождению. Например, как показано на Фигуре 61, sd-PHK может быть присоединена к линкеру, такому, как протонируемый линкер. В одном примере, гистидин может быть сплайсирован на существующий TEG линкер. В некоторых случаях, между 2-5 протонируемыми амин-содержащими химиями, такими как гистидиновые, может быть эффективным для способствованию эндосомальному высвобождению. Фигуры 62 и 63 представляют несколько не ограничивающих примеров улучшения siPHK эндосомального высвобождения путем включения протонируемых функций. Фигура 64 обеспечивает пример GIV, «клик» химию для быстрого доступа к разнообразию. При таком подходе, один олигонуклеотид может быть быстро диверсифицирован, вместо того, чтобы производить различные олигонуклеотиды с исходного положения.
Фосфоротиоатные модификации
Исследовали влияние размещения фосфоротиоатного хвоста на sd-PHK на активность сайленсинга (Фигуры 65-71). На Фигуре 65, серии sd-PHKs с дифференциально размещеными фосфоротиоатными хвостами были исследованы. Размещение фосфоротиоатного хвоста на смысловой нити sd-PHK, как было установлено, оказывает минимальное влияние на активность. sd-PHK соединения, содержащие 6 нуклеотидов с не спаренными основаниями, на любом из 5', 3' или 5' и 3' конце смысловой нити, сохраняли высокую активность (20109, 20111 и 20113, соответственно) (Фигура 66). Ведущие нити содержат (6) фосфоротиоаты на 3' конце. Добавление двух дезокси нуклеотидов, содержащих фосфоротиоаты на 3' конце ведущей нити в дополнение к фосфоротиоатному хвосту в смысловых нитях, привело к снижению активности (20115, 20117 и 20119, соответственно) (Фигура 67) по сравнению с sd-PHK содержащими 8 фосфоротиоатов на ведущих нитях отдельно (20133). В некоторых случаях, добавление двух дезокси нуклеотидов, содержащих фосфоротиоаты на 5' конце ведущей нити привело к полной потере активности (с 20121 по 20132). Не желая быть связанными теорией, это может происходить потому, что участок затравки (seed region) sd-PHK сдвигается на 2 основания.
Фигура 69 показывает, что для PPIB также, размещение фосфоротиоатного хвоста на смысловой нити sd-PHK оказывает минимальное влияние на активность. sd-PHK соединения, содержащие 6 нуклеотидов с не спаренными основаниями, либо на 5', 3' или на 5' и 3' конце смысловой нити демонстрируют снижение активности с увеличением содержания PS (20136, 20138 и 20140, соответственно). Ведущие нити содержат (6) фосфоротиоатов на 3' конце. Добавление двух дезокси нуклеотидов, содержащих фосфоротиоаты на 3' конце ведущей нити в дополнение к фосфоротиоатному хвосту в смысловых нитях привело к снижению активности (20142, 20144 и 20146, соответственно) по сравнению с sd-PHK, содержащей 8 фосфоротиоатов в ведущей нити отдельно (20160). В некоторых случаях, добавление двух дезокси нуклеотидов, содержащих фосфоротиоаты на 5' конце ведущей нити, привело к полной потере активности (с 20148 по 20159).
Влияние изменения содержания фосфоротиоатов как на ведущую, так и на пассажирскую нити PHKi соединений, было затем исследовано. Полностью фосфоротиоатные соединения (21552), которые являются весьма активными, были определены. Интересно, что соединения, содержащиеся на 21 Mer ведущей нити, которая была полностью фосфоротиоатно модифицирована, были активными, однако, уменьшение длины ведущей нити на два нуклеотида, например, 19 Mer ведущей нити (21550), привело к снижению активности. Не желая быть связанными любой теорией, увеличение ведущей нити от 19 до 21 нуклеотидов (полностью фосфоротиоатной ведущей нити) может увеличить температуру плавления между ведущей нитью и мРНК, в результате повышенной активности сайленсинга. Изменение содержания фосфоротиоатов на 13 Мер пассажирской нити такое, что она либо полностью фосфоротиоатно модифицирована, либо содержала 6 фосфоротиоатных модификаций, не приводило к изменению активности (21551 по сравнению с 21556).
Несколько групп ранее пытались разработать полностью фосфоротиоатные PHKi соединения. Полностью фосфоротиоатные дуплексы, на содержащие одноцепочечных областей, были разработаны и протестированы ранее, однако эти соединения не демонстрируют правильный фармакокинетический профиль. Полностью фосфоротиоатные одноцепочечные PHKi соединения также были разработаны ранее, однако, эти соединения не эффективно вводили RISC. Примечательно, что в данной заявке были разработаны гибридные PHKi соединения, которые эффективно вводят RISC и содержат полностью фосфоротиоатные каркасы.
Фигура 71 показывает, что увеличение содержания фосфоротиоатов на пассажирской нити за счет включения нуклеотидов с неспренными основаниями на 5' и 3' конце нити привело к несколько сниженной активности. AML12 клеточные линии трансфицировали с различными концентрациями PHKs (с разным фосфоротиоатным содержанием) с использованием липофектамина PHKiMAX (Invitrogen). Через 48 часов после трансфекции клетки лизировали и уровни гена-мишени были количественно определены, используя анализ разветвленной ДНК (Affymetrix) в соответствии с рекомендациями производителя.
Как показано на Фигуре 72, в in vitro изучения повышения дозы проводились для определения уровней sd-PHK цитотоксичности. Включение стабилизирующих модификаций (т.е. полного 2'Ome на ведущей нити, дуплекс 20407) приводит к снижению цитотоксичности по сравнению с 13966 (2'F С и U на ведущей нити). HeLa клетки обрабатывали различными концентрациями sd-PHK в течение 48 часов. Жизнеспособность клеток оценивали с помощью анализа Alamar Blue. Фигура 73 показывает, что активность sd-PHKs сохраняется при разнообразной химии линкеров.
Фигуры 74-78 представляют не ограничивающие примеры химической оптимизации sd-PHK, мишенью которой является PPIB. Фигуры 79 и 80 показывают, что оптимизированные sd-PHKs, мишенью которых является АроВ, являются активной. Фигура 81 показывает, что химически оптимизированная sd-PHK против Apob была более эффективной в индукции генного сайленсинга в печени, чем большинство традиционных соединений. При очень стабилизированных соединениях АроВ, даже 50 мг/кг инъекции привели к более чем 70% сайленсингу в печени. Эти данные подтверждают, что стабилизация является критически важной для in vivo функциональности.
Фигура 82 показывает стабильность sd-PHK. В присутствии 80% сыворотки, РНК была все еще детектируемой через 48 часов.
Фигура 83 показывает доставку sd-PHK в спинной мозг после инъекции в центральную нервную систему. Фигура 84 показывает проникновение в спинной мозг после доставки sd-PHK в центральную нервную систему. Поглощение sd-PHK происходит, по-видимому, как в тканях, так и в клетках. Фигура 85 показывает результаты экспериментального исследования, указывающие на sd-PHK доставку в ствол головного мозга. В то время как была отмечена вариабельность в зависимости от животного, эти данные свидетельствуют о том, что заболевания ЦНС ствола мозга (такие, как болезнь Хантингтона) могут происходить при прямой доставке sd-PHK.
Пример 7: Новый фосфорамидитный синтез
Введение новых химий в олигонуклеотиды требует синтеза предшественников - фосфорамидитов. Синтез каждого нового модифицированного фосфорамидита может быть сложным. Чтобы упростить процесс, начальный химический скрининг был ограничен контекстом 5-dU.
Это оптимальная отправная точка, поскольку: (1) dU предоставляет возможность для самых простых химических схем синтеза и не требует дополнительной защиты групп; (2) положение 5 уридина находится не на поверхности Уотсона и Крика и модификация этого положения, как было показано, легко переносится большинством олигонуклеотид-взаимодействующих белков, и (3) мы определили, что дезокси U хорошо переносится в нескольких положениях sd-PHK молекулы, что позволяет включение во множестве положений как ведущей, так и пассажирской нити sd-PHK направляющих. Синтезировано 10 химий, которые показаны ниже. Отборы проводили на основе: (1) простоты синтеза и коммерческой доступности предшественников и (2) разнообразия масштабов и структуры гидрофобных модификаций.
Структуры липофильной 2'-дезокси уридиновой модификации:

Главным образом, реакции сочетания Соногаширы использовали для сборки основного химического фрагмента, хотя некоторые мишени будут использовать перекрестное сочетание Сузуки или палладий. Другие химические реакции, которые использовали, хорошо разработаны в литературе и являются надежными и приводят к высоким выходам.
Дизайн и синтез соединений
Как уже говорилось выше, каркасные последовательности, мишенью которых являются МАР4К4 и PCSK9, показали эффективность двух различных sd-PHK соединений с и без дезокси модификации U в нескольких местах.
Варианты пассажирской и ведущей нити синтезируются с и без гидрофобной модификации концов. Это позволяет оценку воздействия гидрофобной модификации на свойства поглощения сами по себе и в сочетании с гидрофобной модификацией концов, которая используется в настоящее время.
Синтезирован массив из 108 олиго вариантов, мишенью которых является МАР4К4 и PCSK9, с различными степенями липофильности, что приводит к более чем 288 различным соединениям (12 олигонуклеотидов на химию). Панель включает от 3 до 7 различных липофильных мономеров, в 12 различных моделей модификаций. Использование различных конфигураций позволит «титрование» количества липофильности и оценку влияния позиционирования в PHKi. Соединения, для которых установлена активность в клеточной культуре, получены с флуоресцентной меткой для оценки клеточного поглощения.
Одноцепочечные олигорибонуклеотиды синтезировали с использованием MerMade 12 олигонуклеотидного синтезатора и снимали защиту с использованием стандартных методик. Потенциально, поскольку задействованные химии не являются стандартными в связи с методологией, некоторые изменения могут быть сделаны. Олигорибонуклеотиды очищали с использованием стандартной С18 методологии (>70% чистоты) и анализировали с использованием аналитической обращеннофазной ионно-спаривающей хроматографии и подмножества MALDI-TOF. Одноцепочечные пассажирскую и ведущую нити затем дуплексировали. Влияние дополнительных химий на олигонуклеотидную дуплексную гидрофобность оценивали хроматографически с использованием методологии Dagle et al. после установления корреляции сравнительно с ко-коэффициентом разделения октанол-вода. Целью является повышение logP дуплексов на порядок величины по сравнению с текущей версией sd-PHK химии. Наилучшие соединения были радиоактивно мечены и ко-коэффициенты разделения оценивали обычным протоколом разделения октанол-вода.
Эффективность синтезированных соединений затем оценивали в анализах культуры тканей для клеточного поглощения, токсичности и эффективности. Следующие стандартные анализы использовали для оценки соединения:
Оценка поглощения
Флуоресцентно меченые sd-PHK (DY547) варианты оценивали по поглощению. Поглощение исходных sd-PHK соединений происходило в течение нескольких минут воздействия, так что оценивали как эффективность, так и кинетику. Клетки инкубировали в культуральной среде в присутствии различных концентраций sd-PHK вариантов и фиксировали в соответствующие моменты времени. Использовали Leica SP5 конфокальный микроскоп, оснащенный 405 синим диодным лазером, 458, 476 и 514 аргоновыми лазерами, 561 желтым диодным лазером и 633 HeNe лазером. 405 лазер использовали для сбора DAPI или Hoechst изображения. Конфокальный микроскоп также имеет возможность захвата множества изображений из сканирования множества лазеров, и в то же время позволяет генерацию объединенных изображений. Оптимальные соединения дополнительно оценивали для визуализации кинетики и механизма поглощения с использованием TIRF микроскопии.
Оценка токсичности in vitro
Анализ Alamar Blue использовали в соответствии с протоколами производителей для оценки общей токсичности соединения. Некоторые из химий, как ожидается, были токсичными и будут по-прежнему включены в оценку эффективности тканевой культуры, но исключены из любого in vivo анализа. Для того, чтобы определить потенциал соединений для IFN индукции, соединения добавляют в HelaS3 и первичные макрофаги и уровень экспрессии мРНК для IFIT1, IL-6, CD45 и других маркеров воспаления оценивали с помощью Q-PCR и В-ДНК. Эти два типа клеток ранее были чувствительны к токсичности соединений. Poly IC, 45 mer дуплекс и другие положительные контроли использовали. Соединения, которые не показывали клеточной токсичности и IFN индукции в анализах тканевых культур, дополнительно оценивали на предмет активации воспалительного ответа с использованием анализа высвобождения цитокинов цельной крови.
Анализы генного сайленсинга
Hela клетки и/или первичные гепатоциты (и другие клеточные линии, если это необходимо) использовали для оценки эффективности соединения. Для оценки совместимости с PHKi клеточным механизмом (RISC загрузкой и расщеплением) sd-PHK конструкты трансфицировали в клетки с помощью липофектамин PHKiMAX (Invitrogen, Carlsbad, СА) в соответствии с инструкциями производителя. Дополнительно, эффективность при пассивном поглощении позволяет оценить как поглощение, так и RISC поступление. Проведение обоих анализов позволяет дифференцирование между влиянием химии на RISC загрузку и клеточным поглощением. Через 24-48 часов инкубации клетки лизировали и активность генного сайленсинга измеряли при помощи qPCR или анализов разветвленной ДНК (Affymetrix). Уровни экспрессии генов-мишеней нормировали на экспрессию генов домашнего хозяйства. Для анализов самодоставки (без трансфекции реагента) sd-PHK просто добавляли к культуральным средам и инкубировали в течение 24-48 часов до лизиса клеток для получения РНК. Данный анализ позволяет идентифицировать соединения, которые не токсичны и имеют по меньшей мере в 3-10 раз более высокие ЕС50 значения по сравнению с оригинальной sd-PHK при пассивном поглощении (ЕС50 <10 нМ in vitro).
Анализ in vivo
Не токсичные и эффективные соединения дополнительно оценивали in vivo в BALB/c мышах-самцах. Короче говоря, животным IV делали инъекции при уровнях целевых доз и пробы крови отбирали в соответствующих точках для оценки удержания крови и скорости клиренса. Типичные конечные точки включали количественное определение соединений в крови и ткани, химию крови, ферменты печени, гистологическое исследование всех основных органов и DY547, помеченный как sd-PHK поглощение (конфокальная микроскопия). Ранее подтвержденную гибридизацию на основе анализа использовали для количественного анализа крови и тканей. Для наиболее перспективных соединений, исследования оценивали уровень нокаутированной мРНК мишени по сравнению с контролем для несоответствия контроля и PBS обработанных групп. N-5 с 3 биопсиями в ткани собирали и хранили с использованием PHKlaler® (Qiagen). Все анализы хорошо известны. Помимо системной оценки, панель лучших нетоксичных соединений испытывали при различных концентрациях локально (внутри-кожные инъекции) и по сравнению с оригинальной sd-PHK.
Эндосомальное высвобождение
В то время как текущая версия sd-PHK химии дает несколько порядков улучшения клеточного поглощения по сравнению с обычными siPHKs, в некоторых случаях, относительно высокие концентрации соединений необходимы для достижения эффективности (5-50 нм ЕС50 значения). Если те же самые sd-PHK вводят путем липидной опосредованной доставки, ЕС50 значение снижается в по меньшей мере 10-50 раз, что свидетельствует о количестве пассивно интернализованных олигонуклеотидов, которые не достигают RISC комплекса. Визуализация и совместное окрашивание показывают, что поглощение происходит через механизм эндоцитоза. Большинство sd-PHK проходят через мульти-везикулярное тело (MVB) и становятся пойманными в лизосомах через 24 часа, в то время как лишь небольшая фракция высвобождает эндосомы и лизосомы и доступна для загрузки RISC.
Не желая быть связанными теорией, эндосомальный захват может быть эффективность-ограничивающим фактором. Дополнительный протонируемый амин, содержащий модификации, может улучшать sd-PHK эндосомальное и лизосомальное высвобождение. Инкапсуляция олигонуклеотидов в протонируемых аминах, содержащих липиды, является основным маршрутом олигонуклеотидов, которым в настоящее время они доставляются в клетки (большинство коммерчески доступных реагентов трансфекции используют данный подход). Включение протонируемых аминов в контексте динамических поликонъюгатов было использовано ранее (Rozema, Lewis et al. 2007) и хорошая эффективность была продемонстрирована в печени. К сожалению, накопление неразлагаемого полимера, используемого в качестве каркаса и другие свойства в результате ограничивают профиль токсичности. При изменении рН при созревании эндосом, амин протонирован, что приводит к разрыву эндосом и олиго-высвобождению.
В данной заявке, различные протонируемые амины включены в контекст sd-PHK непосредственно. Путем добавления разного количества и типа аминовых химий в разных местах sd-PHK, существенное увеличивается олиго эндосомальное высвобождение и, таким образом, эффективность может быть достигнута. Это позволит обеспечить более эффективную самостоятельную доставку РНК конструктов с улучшенной фармакологией, которая может быть использована для разработки кандидатных PHKi лекарственных препаратов в нескольких терапевтических областях, как для местного, так и системного применений. Эти новые конструкции также обеспечивают самостоятельную доставку РНК в in vivo функциональных геномных исследованиях. Такой подход является весьма новым. Стартовой точкой являются соединения, которые продемонстрировали беспрецедентные уровни клеточного поглощения, где эффективность ограничена внутриклеточной компартментализацией (эндосомальным захватом). Включение протонируемых аминов в смысловую нить (часть молекулы, которая удаляется после RISC загрузки) используется для значительного увеличения эффективности. Дополнительно, эти исследования предоставляют данные о структуре/функции протонируемых аминов, содержащих дуплексные PHKs, которые помогут в проектировании PHKi соединений и симулировании микроРНК.
Синтез протонируемых аминов, содержащих предшественники, для способствования эндосомального высвобождения
В настоящее время, в некоторых осуществлениях, sd-PHK смысловая нить использует TEG линкер для прикрепления гидрофобных фрагментов. Это трифункционально, что позволяет присоединение к твердой подложке, на которой осуществляется синтез олигонуклеотидов, к химически модифицированной РНК (которая образует одну половину siPHK дуплекса) и к одной из гидрофобных функциональных групп (генерически холестерин). Включение нескольких протонируемых аминов, содержащих химии, было сначала исследовано в контексте линкера. Хотя присутствие положительно заряженного линкера может вернуть в исходное положение, наличие гидрофобного ядра может предотвратить это.
Карбонильно-азотная связь карбаматной функциональной группы может быть разработана, чтобы включить аминокислоты, такие как гистидин. Гистидин имеет рКВН+ имидазольной боковой цепи ~6, что означает, что он является преимущественно нейтральным при физиологических значениях рН, но преимущественно протонирован в эндосомальном пространстве. Тозилатная защитная группа предложена для имидазольной функциональной группы, чтобы освободить гистидин или метильную группу для использования. Дополнительно, имидазольный цикл может быть замещен электроноакцепторной группой для оптимизации рКВН+ в случае необходимости.
Имидазолильная функциональная группа в своей природной или синтетически модифицированной форме также может быть включена в уридиновое нуклеооснование с использованием опубликованных методов (Vaught, Bock et al.; Holmes SC 2005).
Другие химические функциональные группы могут быть синтезированы в последующих попытках для протонирования в диапазоне рН 5-7, включая электронно-обогащенные пиридины. Пиридильные варианты с различной степенью метильного замещения могут быть синтезированы из коммерчески доступных хлорметильных предшестенников.
Панель соединений, которые образуются, будет включать 2 типа протонируемых аминов с различными номерами в контексте гидрофобного линкера и внутренне в соединении. Этот материал позволяет синтез панели sd-PHK вариантов с разными уровнями включения протонируемых аминов.
Создана панель олигонуклеотидов, содержащих 1-5 протонируемых аминов в контексте гидрофобного линкера, прикрепленного к 3' концу смысловой нити, а также распределенных по всей смысловой нити. Чтобы свести к минимуму воздействие смещения последовательности, используют две каркасные последовательности (мишенью одной является МАР4К4 и мишенью другой является PCSK9). Обе последовательности позволяют включение 3-4 протонируемых аминов в контексте смысловой нити. Первоначально внутреннее включение ограничивается смысловой нитью, поскольку наличие этих типов химии в ведущей нити может в некоторых случаях влиять на RISC функцию.
Синтезирован массив из 50-70 вариантов олигонуклеотидов, мишенью которых являются МАР4К4 и PCSK9, с различными степенями, типами и положениями химии протонируемых аминов. Панель включает от 1 до 5 различных мономеров, с несколькими моделями модификации. Использование этих различных конфигураций позволит «титрование» количества и положения (конечное или внутреннее) протонируемых аминов и оценку влияния на RISC сборку и клеточное поглощение. Соединения, для которых установлена активность в клеточной культуре, получены с флуоресцентной меткой для оценки клеточного поглощения.
Одноцепочечные олигорибонуклеотиды синтезировали с использованием MerMade 12 олигонуклеотидного синтезатора (BioAutomation, Piano, ТХ) и снимали защиту с использованием стандартных методик. Поскольку задействованные химии не являются стандартными, некоторые изменения в сочетании и условиях снятия защиты, возможно, необходимо будет выполнить. Неочищенные олигорибонуклеотиды затем очищали, используя стандартную методологию С18 (>70% чистоты) и анализировали с использованием аналитической обращеннофазной хроматографии ионного спаривания и подмножества по MALDI-TOF. Одноцепочечные пассажирскую и ведущую нити затем дуплексировали. Влияние дополнительных химий на связывание в олигонуклеотидный дуплекс оценивали при помощи гель-электрофореза. В результате, включено максимальное количество протонируемых аминов, не препятствуя образованию дуплекса и целостности соединения. Следующие стандартные анализы будут использованы для оценки соединения.
Оценка поглощения
Флуоресцентно меченые sd-PHK (DY547) варианты оценивали по поглощению. Исходные sd-PHK соединения поглощались клетками в течение нескольких минут воздействия, так что оценивали как эффективность, так и кинетику. Клетки инкубировали в культуральной среде в присутствии различных концентраций sd-PHK вариантов и фиксировали в соответствующих моментах времени. Использовали Leica SP5 конфокальный микроскоп, оснащенный 405 синим диодным лазером, 458, 476 и 514 аргоновыми лазерами, 561 желтым диодным лазером и 633 HeNe лазером. 405 лазер используется для сбора DAPI или Hoechst изображения. Конфокальный микроскоп также имеет возможность захвата множества изображений со сканирования множества лазеров и в то же время позволяет генерацию объединенных изображений. Наиболее активные соединения дополнительно оценивали для визуализации кинетики и механизма поглощения с использованием TIRF микроскопии.
Оценка токсичности in vitro
Alamar Blue анализы использовали в соответствии с протоколом производителя для оценки общей токсичности соединений. В случае, если некоторые соединения являются токсичными, они включены в оценку эффективности тканевой культуры, но исключены из любого in vivo анализа. Соединения добавлены в HelaS3 и первичные макрофаги и уровень мРНК экспрессии для IFN, IFIT1, IL-6, CD45 и других маркеров воспаления оценивали с помощью Q-PCR и В-ДНК. Эти два типа клеток ранее были чувствительны к провоспалительным эффектам siPHK. Poly IC, 45 mer дуплекс и другие позитивные контроли использовали. Соединения, которые не показывали клеточной токсичности и IFN индукции в анализах тканевой культуры, дополнительно оценивали на предмет активации воспалительного ответа с использованием анализа цитокинов цельной крови (Lankveld, Van et al. 2010). Токсичность оценивали, поскольку новые химии могут привести к увеличению токсичности.
Анализы генного сайленсинга
Hela клетки и/или первичные гепатоциты (и другие клеточные линии, если это необходимо) использовали для оценки эффективности соединения. Для оценки совместимости с PHKi клеточным механизмом (RISC загрузка и расщепление), sd-PHK конструкты трансфицировали в клетки с помощью липофектамин PHKiMAX (Invitrogen, Carlsbad, СА) в соответствии с инструкциями производителя. Дополнительно, эффективность при пассивном поглощении позволяет оценить как поглощение, так и RISC поступление. Проведение обоих анализов позволяет различать влияние химии на RISC загрузку и клеточное поглощение. Через 24-48 часов инкубации клетки лизировали и активность генного сайленсинга измеряли при помощи qPCR или анализов разветвленной ДНК (Affymetrix). Уровни экспрессии генов-мишеней нормированы на экспрессию генов домашнего хозяйства. Для анализов самодоставки (без трансфекции реагента) sd-PHK просто добавляли к культуральным средам и инкубировали в течение 24-48 часов до лизиса клеток для получения мРНК. Этот анализ приводит к идентификации соединений, которые не являются токсичными и имеют по меньшей мере в 3-10 раз более высокие ЕС50 значения по сравнению с оригинальной sd-PHK при пассивном поглощении (ЕС50 <2 нм in vitro).
Тестирование in vivo
Соединения, показанные как нетоксичные и эффективные в культуре клеток дополнительно оценивали in vivo у BALB/м мышей-самцов. Короче говоря, животным IV вводили путем инъекции при уровнях целевых доз и пробы крови отбирали в соответствующих точках для оценки удержания крови и скорости клиренса. Типичные конечные точки включали количественное определение соединения в крови и ткани, химии крови, ферменты печени, гистологическое исследование всех основных органов и поглощение DY547 меченой sd-PHK (конфокальная микроскопия). Дополнительно, оценивали внутрикожную и внутриглазную эффективность и токсичность. Ранее подтвержденный гибридизационный анализ использовали для количественного анализа крови и тканей. Исследования оценивали уровень целевой нокаутированной мРНК по сравнению с контрольным несоответствием и PBS обработанными группами. N-5 с 3 биопсиями в ткани собирали и хранили PHKlater® (Qiagen). В дополнение к системной оценке, панель наиболее активных соединений анализировали в различных концентрациях локально (внутри-кожные инъекцие) по сравнению с оригинальной sd-PHK.


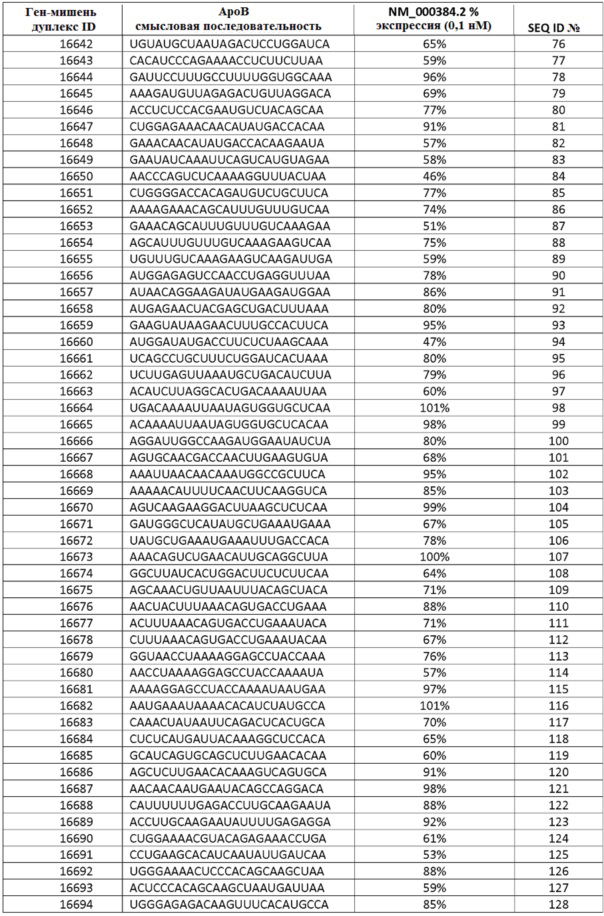

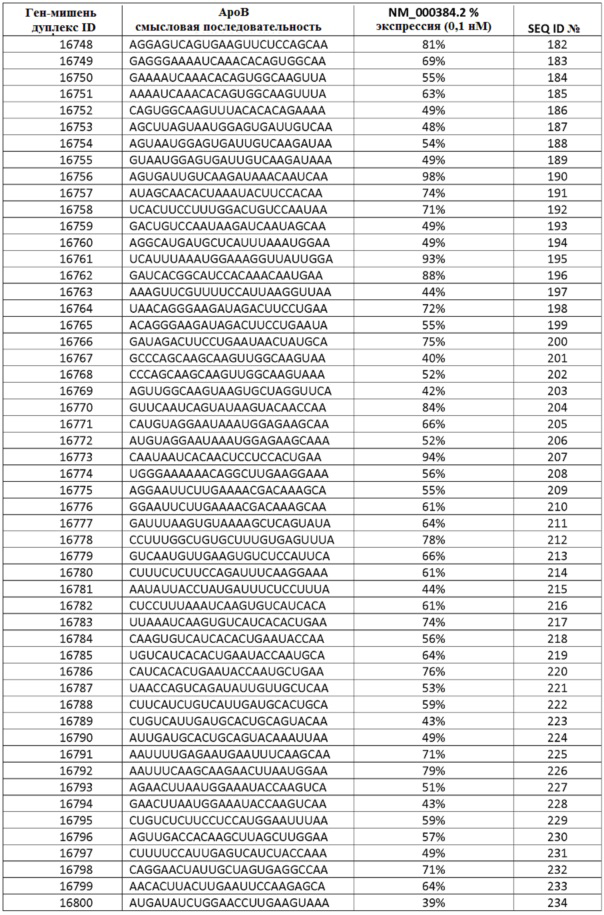

Таблица 2: hApoB sd-rxРНК последовательности
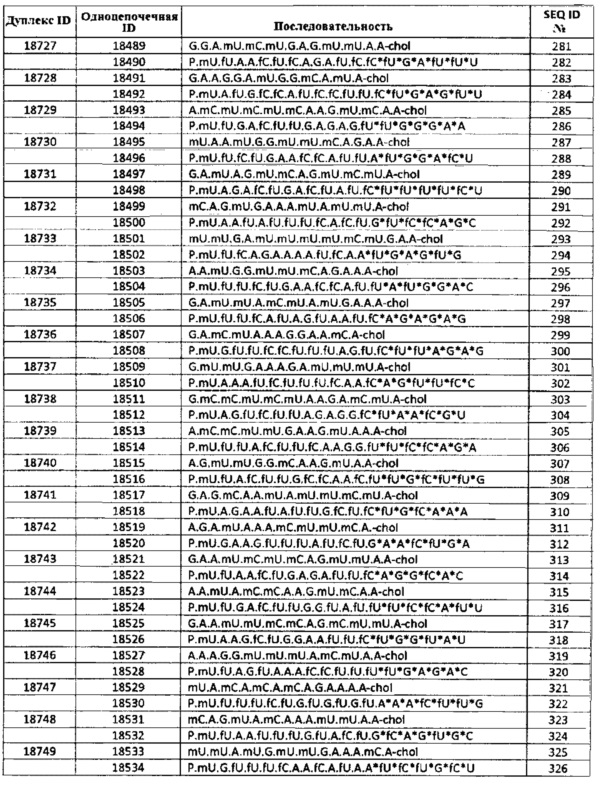
Таблица 3: PCSK9 sd-rxРНК последовательности



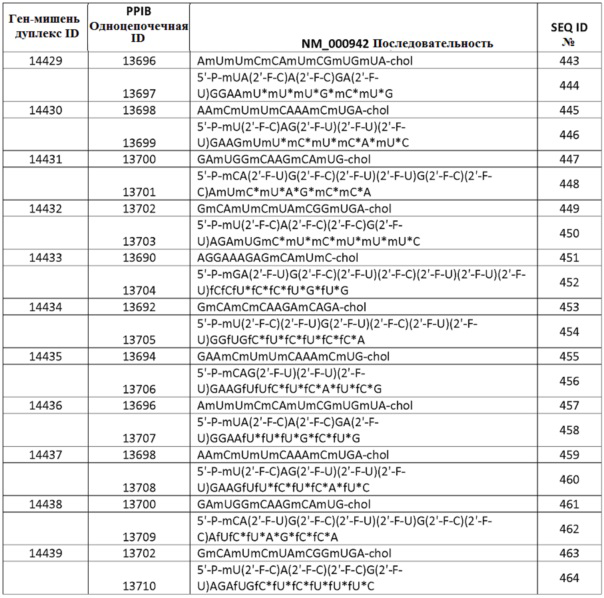

Имея, таким образом, описанные несколько аспектов по меньшей мере одного осуществления настоящего изобретения, специалистам в данной области техники должно быть понятно, что будут легко возникать различные изменения, модификации и усовершенствования. Такие изменения, модификации и усовершенствования призваны быть частью данной заявки, и предназначены, чтобы не выходить за суть и объем настоящего изобретения. Таким образом, приведенное выше описание и чертежи приведены только для примера.
ЭКВИВАЛЕНТЫ
Специалистам в данной области техники будет понятно, или они будут иметь возможность убедиться, используя не более чем рутинные эксперименты, что возможно многие эквиваленты конкретных осуществлений настоящего изобретения описаны в данной заявке. Такие эквиваленты предназначены для того, чтобы быть охваченными в приведенной ниже формуле изобретения.
Все ссылки, включая патентные документы, описанные в данной заявке, включены в качестве ссылки в полном объеме. Данная заявка включает ссылки на все содержимое, включая все чертежи и все части заявки (включая перечень последовательностей или последовательности аминокислот/полинуклеотидов) РСТ публикации № WO 2010/033247 (заявка № PCT/US 2009/005247), поданной 22 сентября 2009 года под названием «САМОДОСТАВЛЯЮЩИЕСЯ PHKI СОЕДИНЕНИЯ УМЕНЬШЕННОГО РАЗМЕРА» и РСТ публикации № WO 2009/102427 (заявка № PCT/US 2009/000852), поданной 11 февраля 2009 года, под названием «МОДИФИЦИРОВАННЫЕ PHKI ПОЛИНУКЛЕОТИДЫ И ИХ ИСПОЛЬЗОВАНИЕ».























































































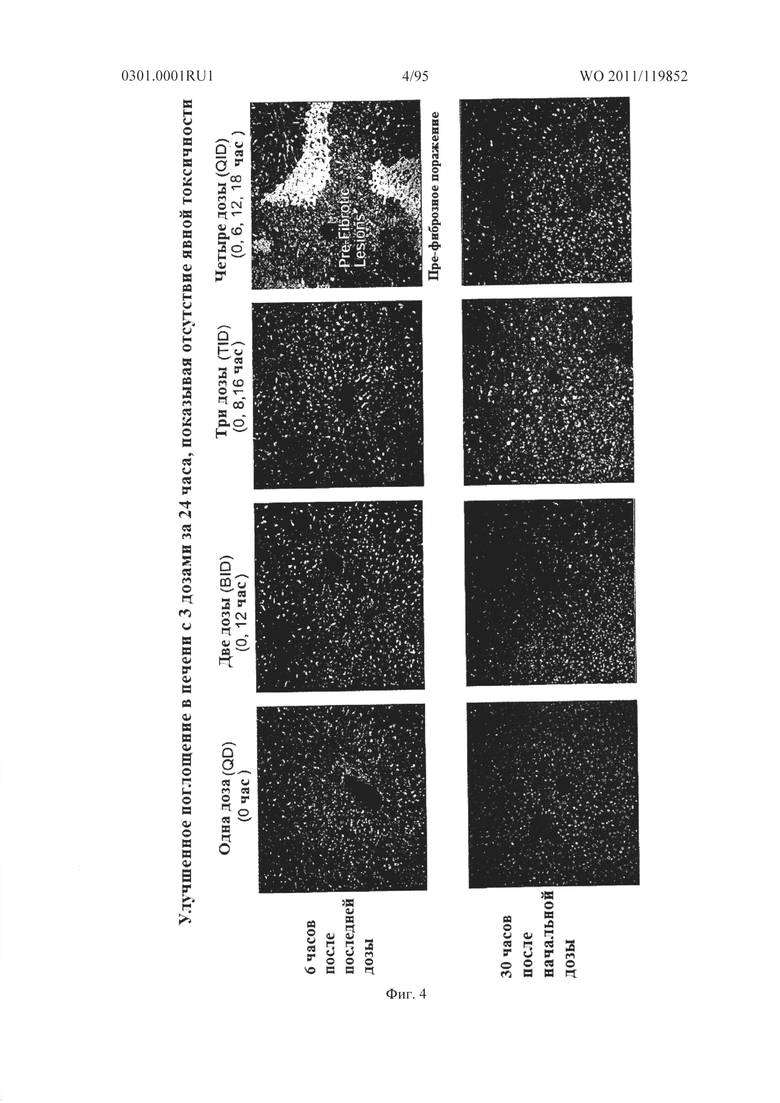


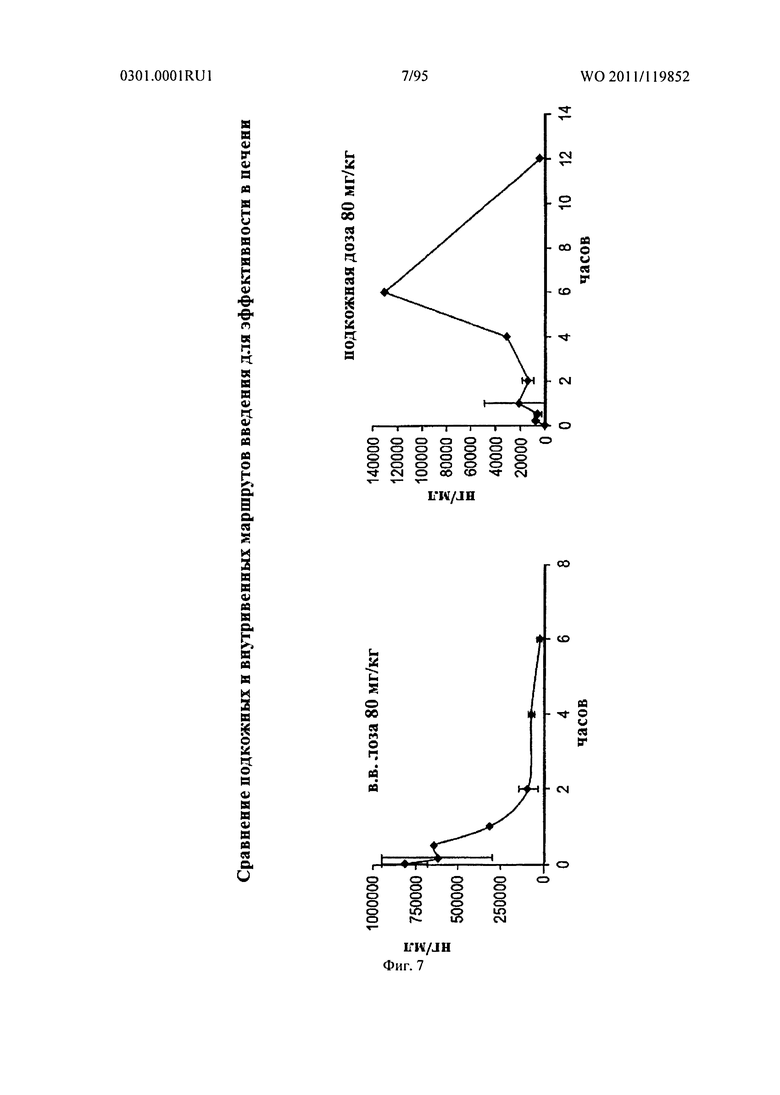














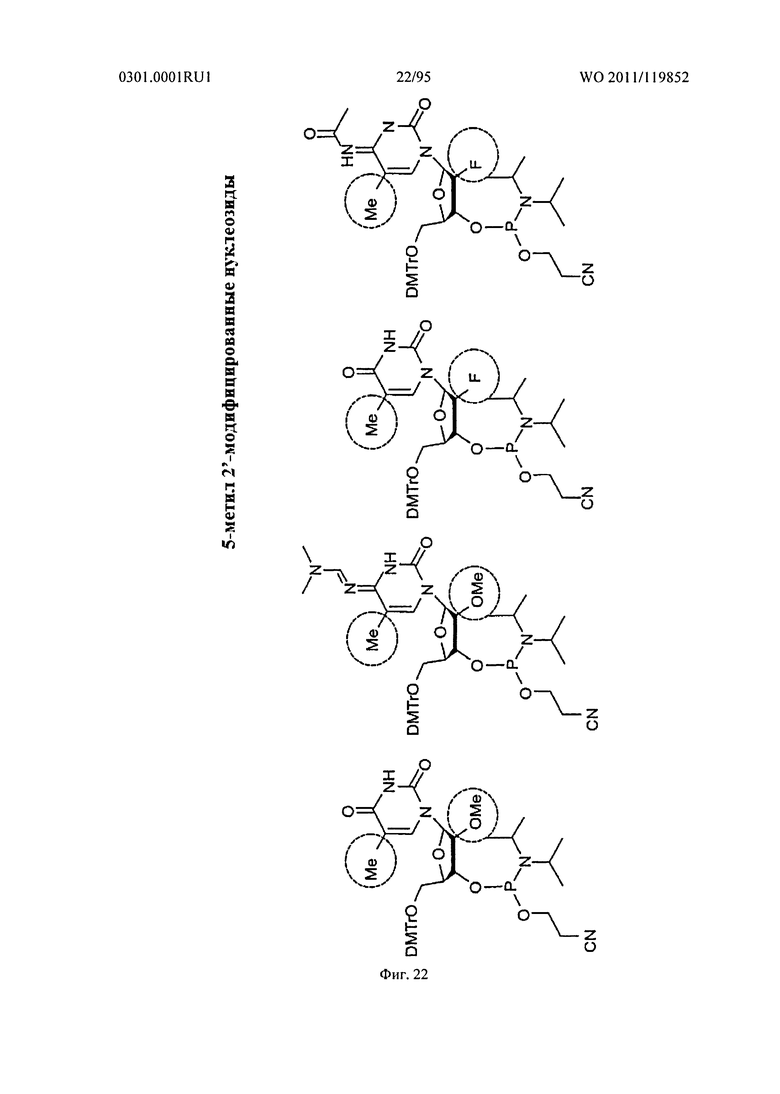
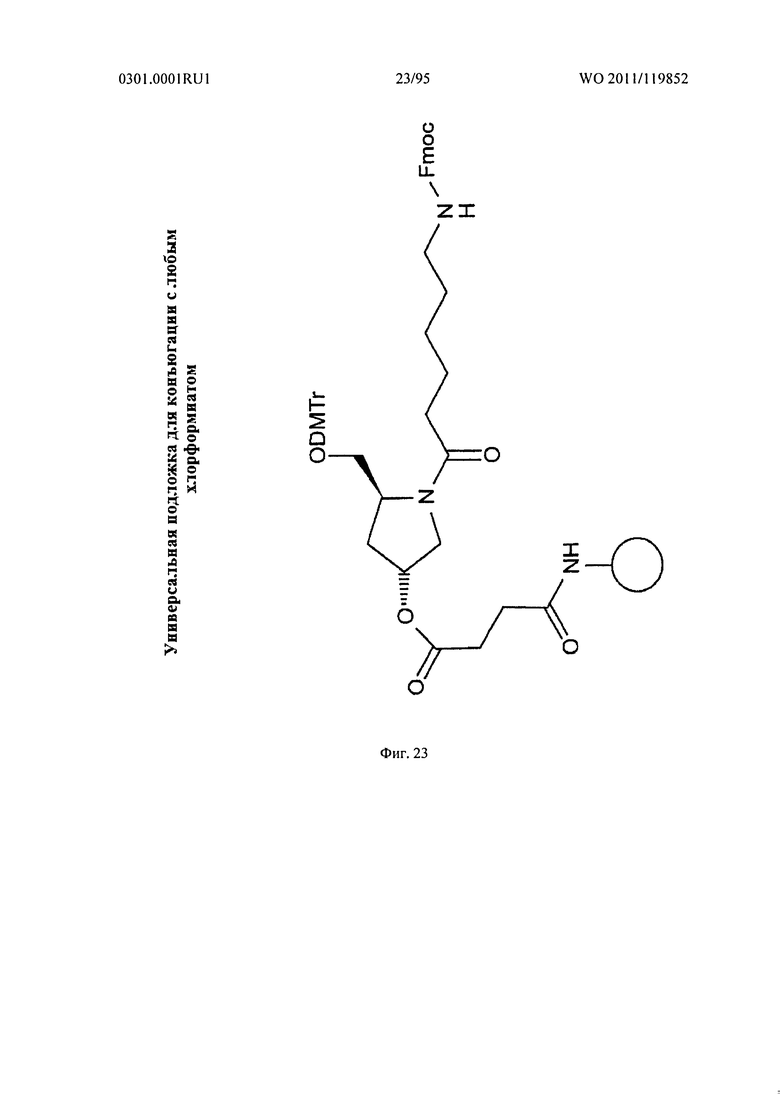


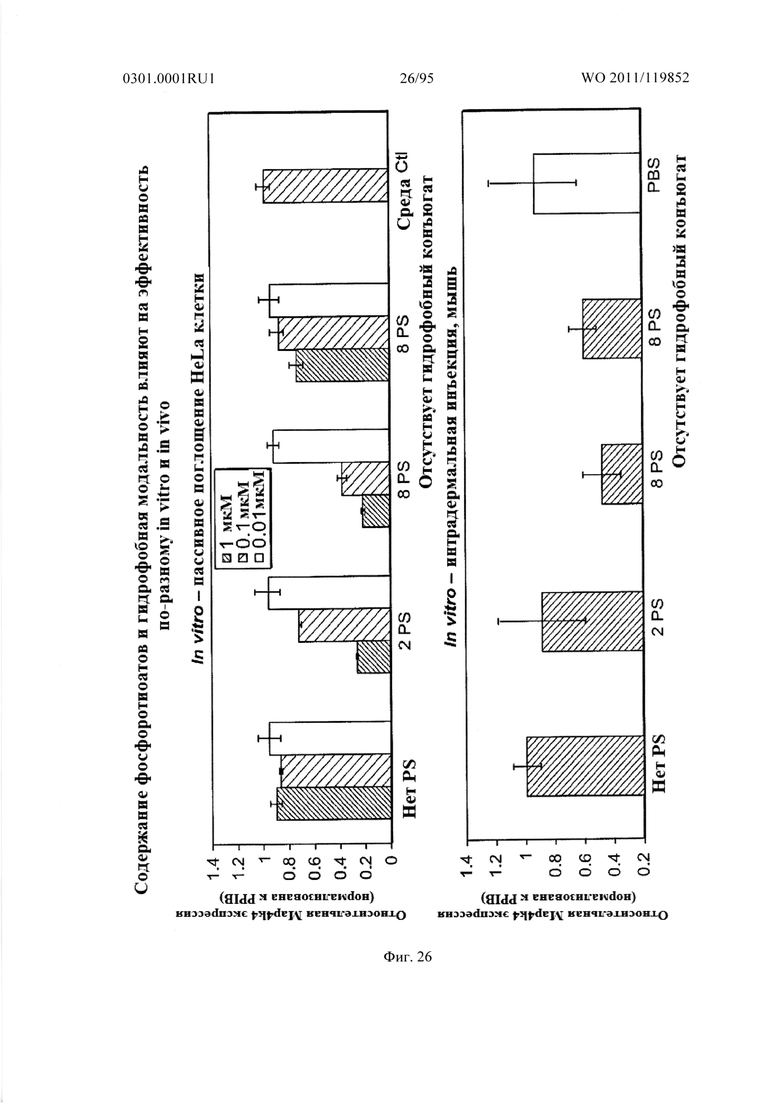

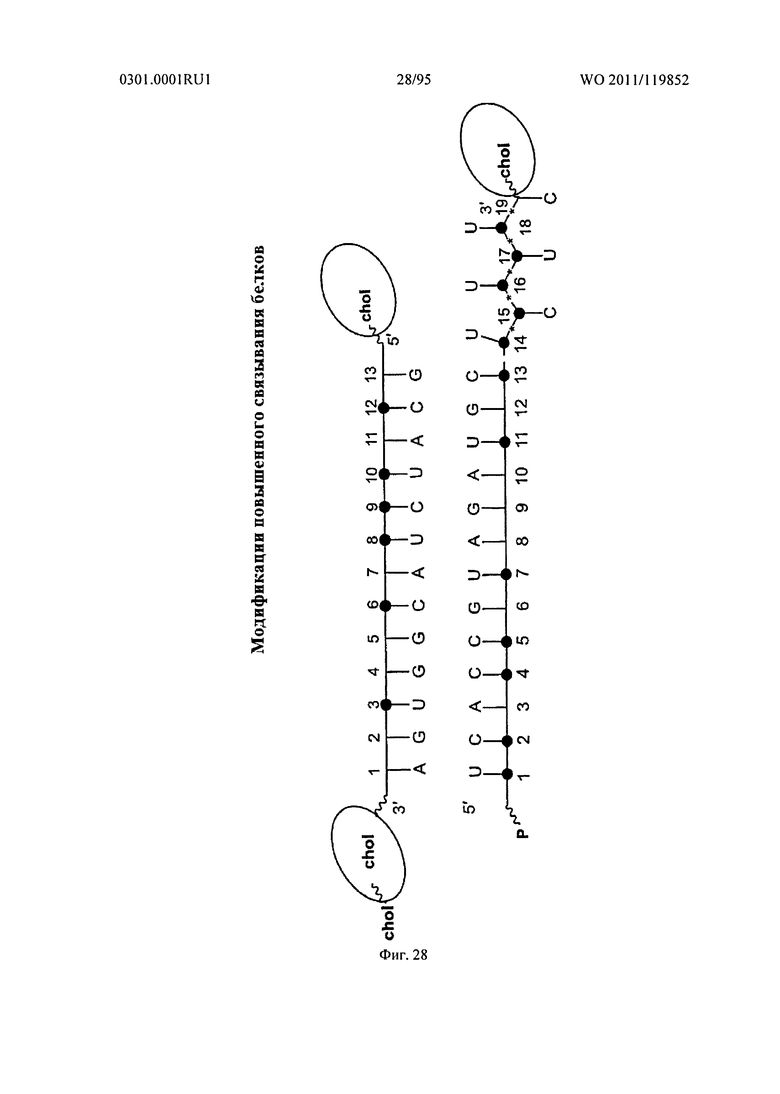


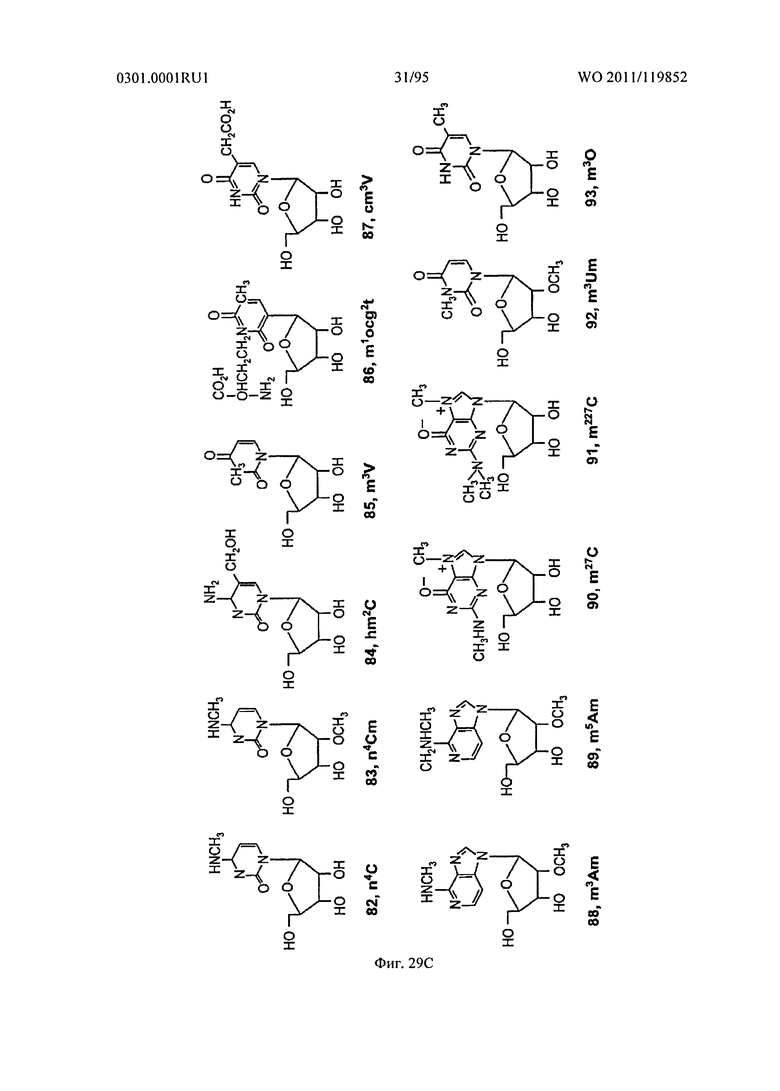

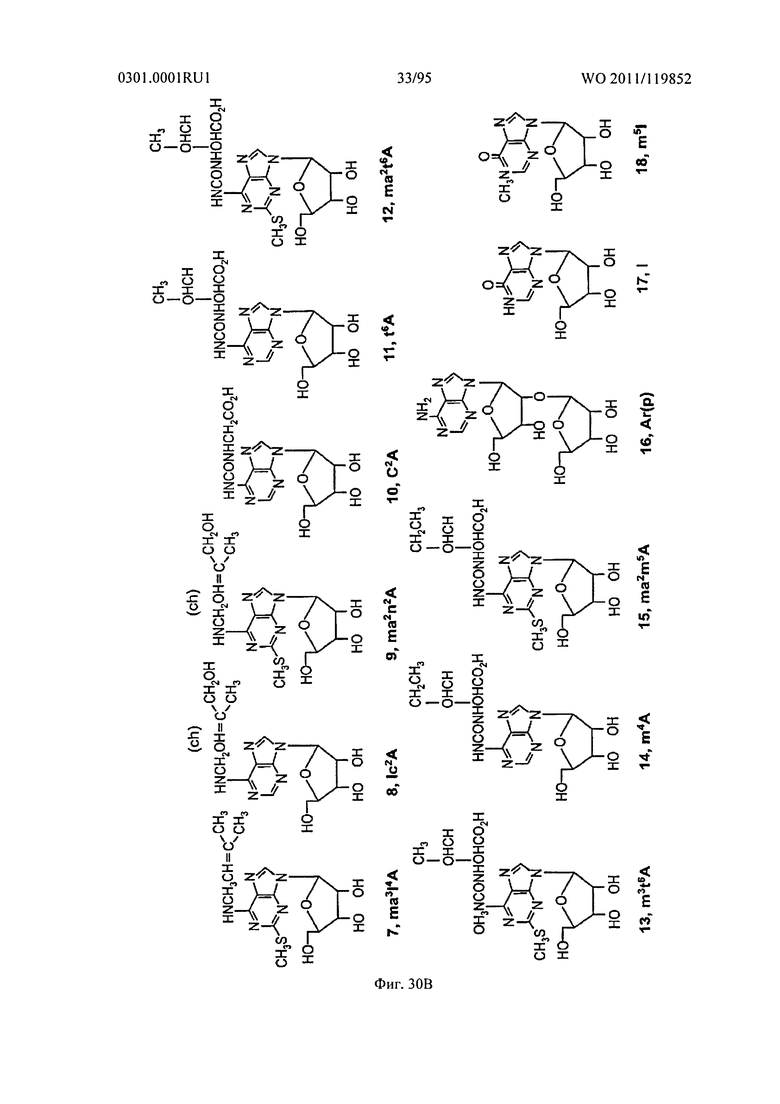

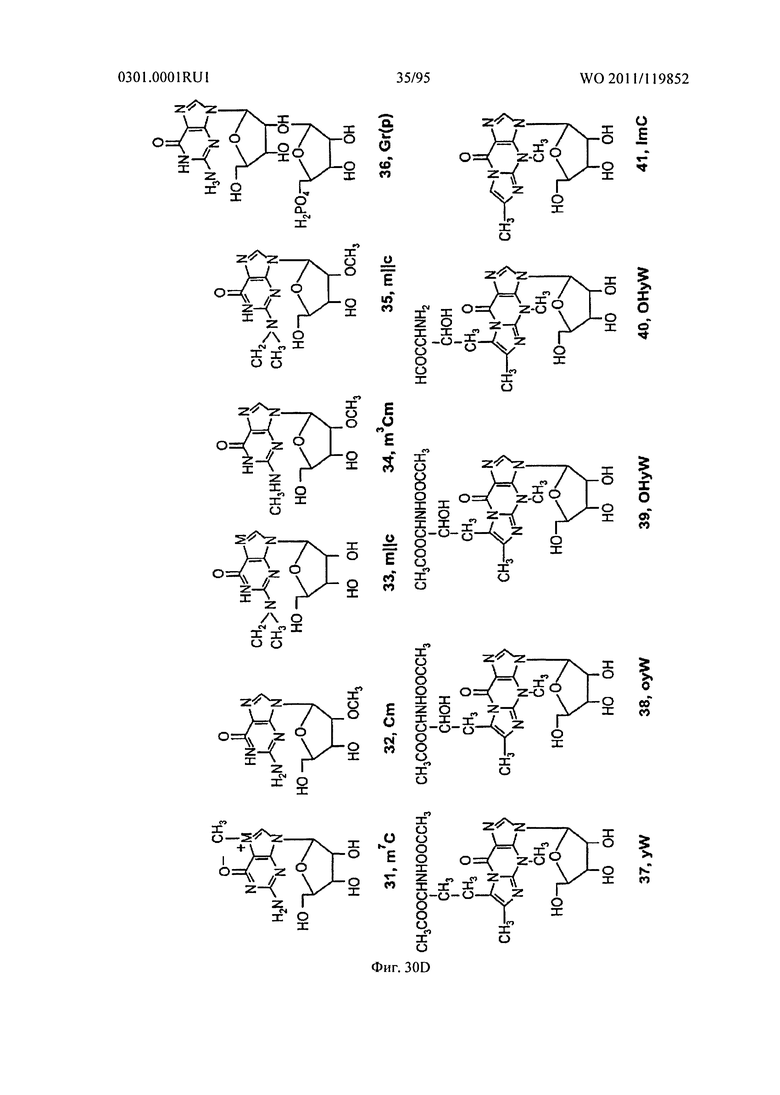




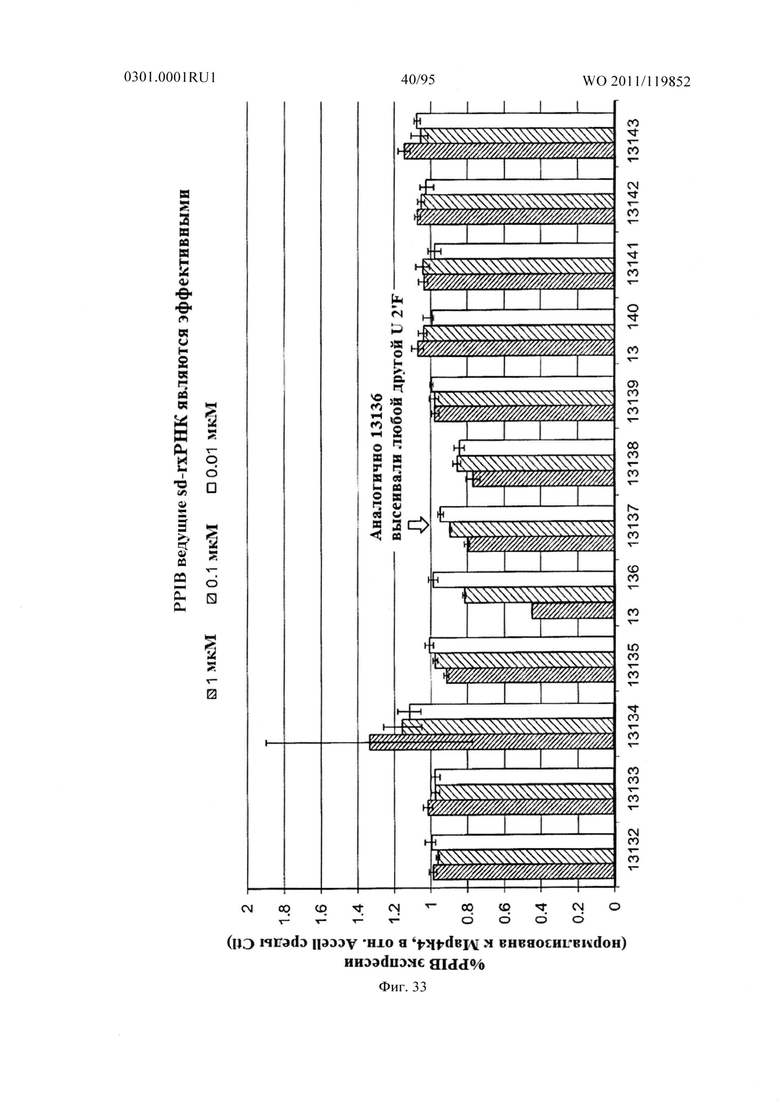




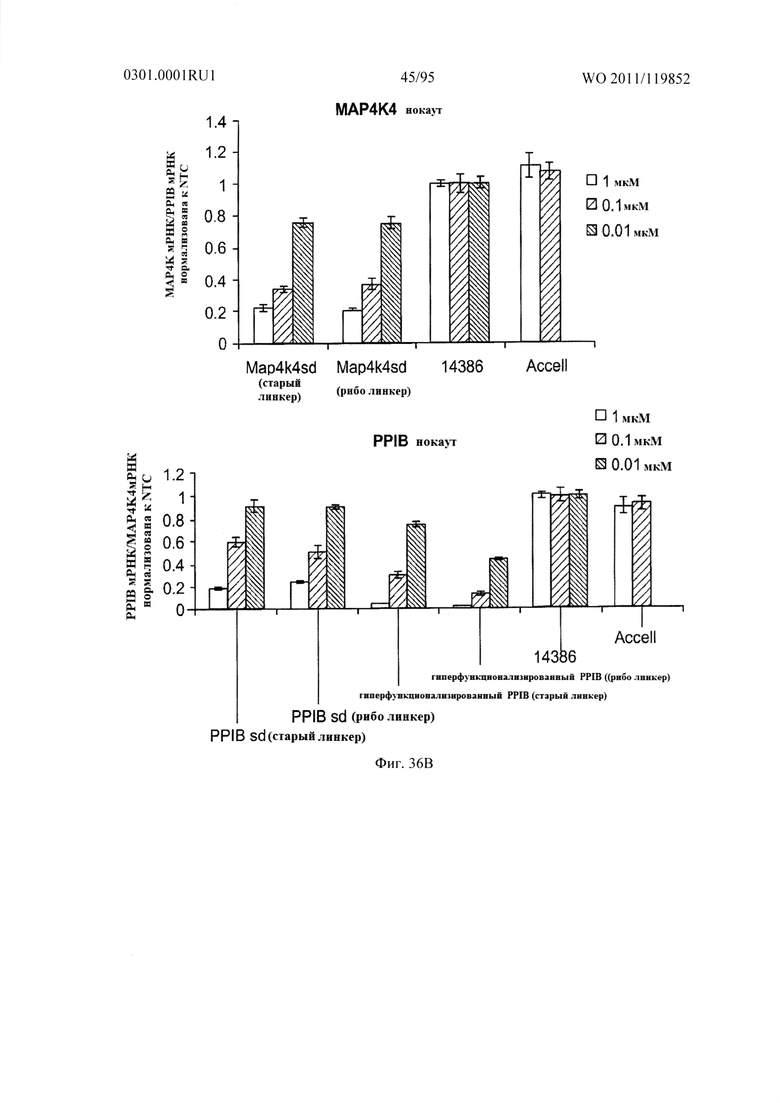
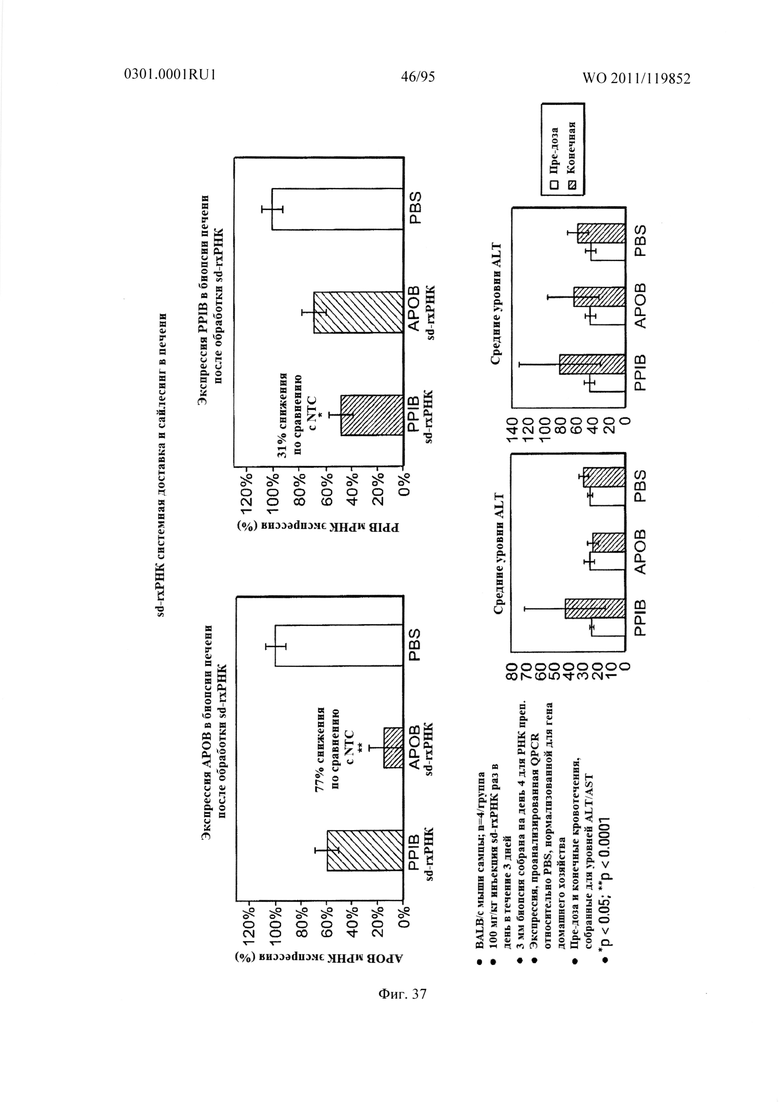
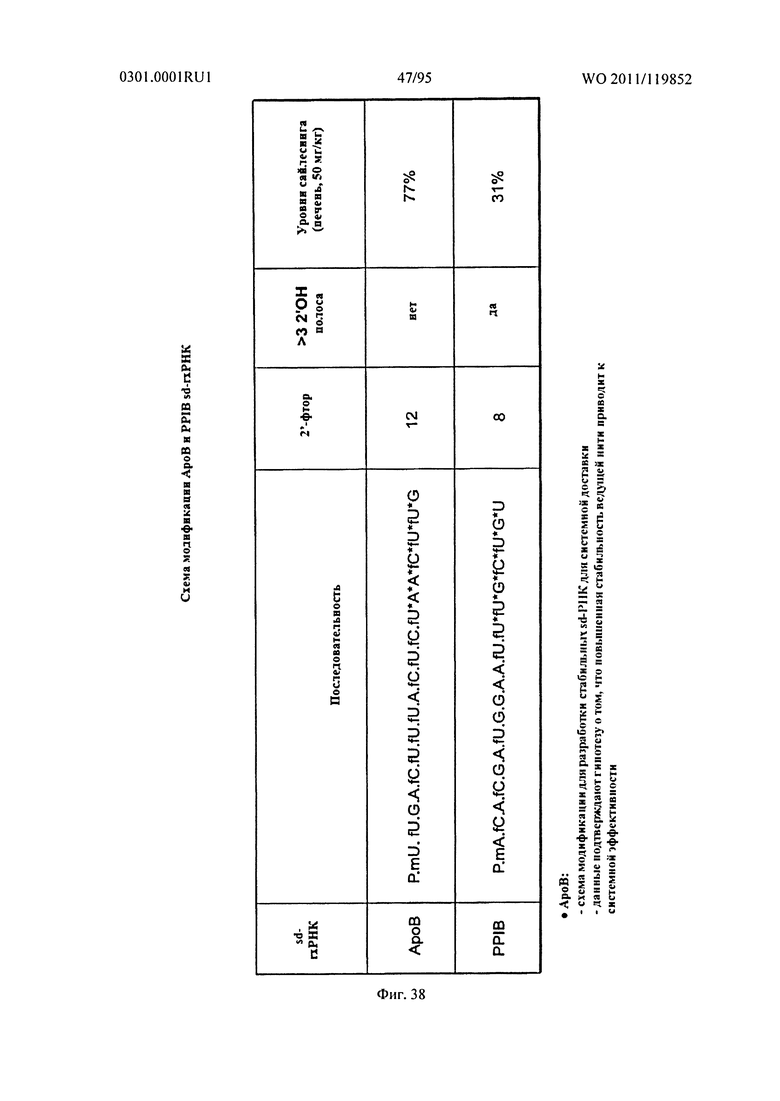


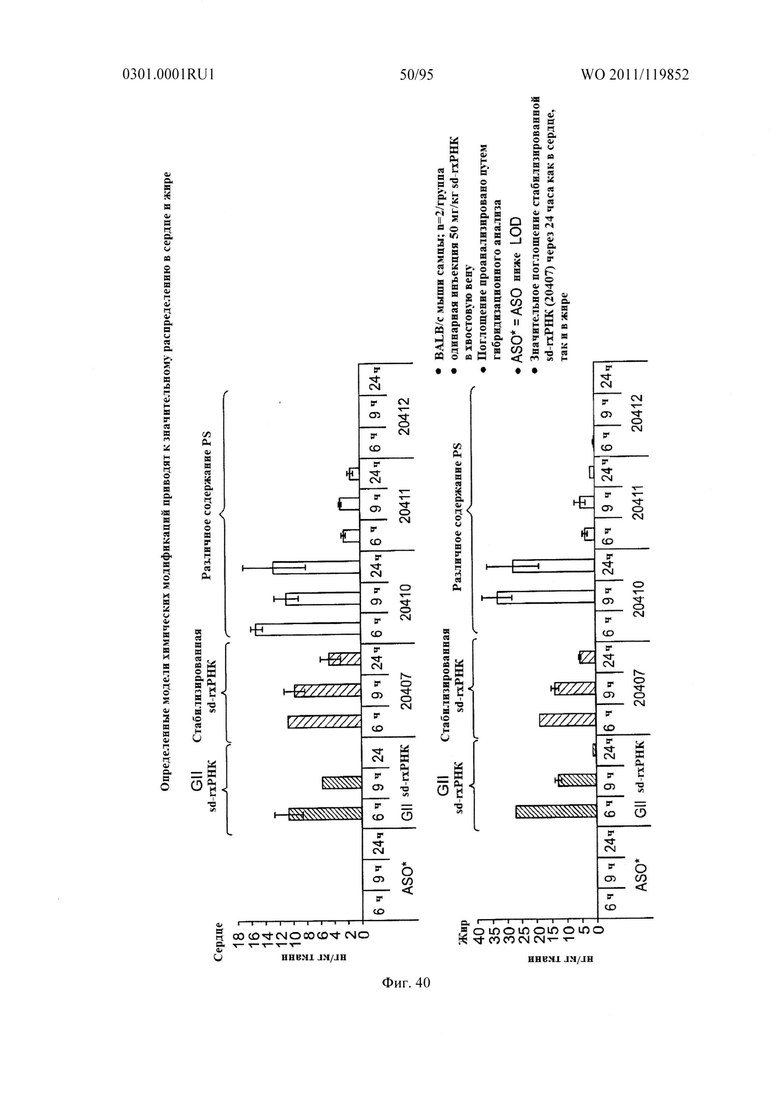




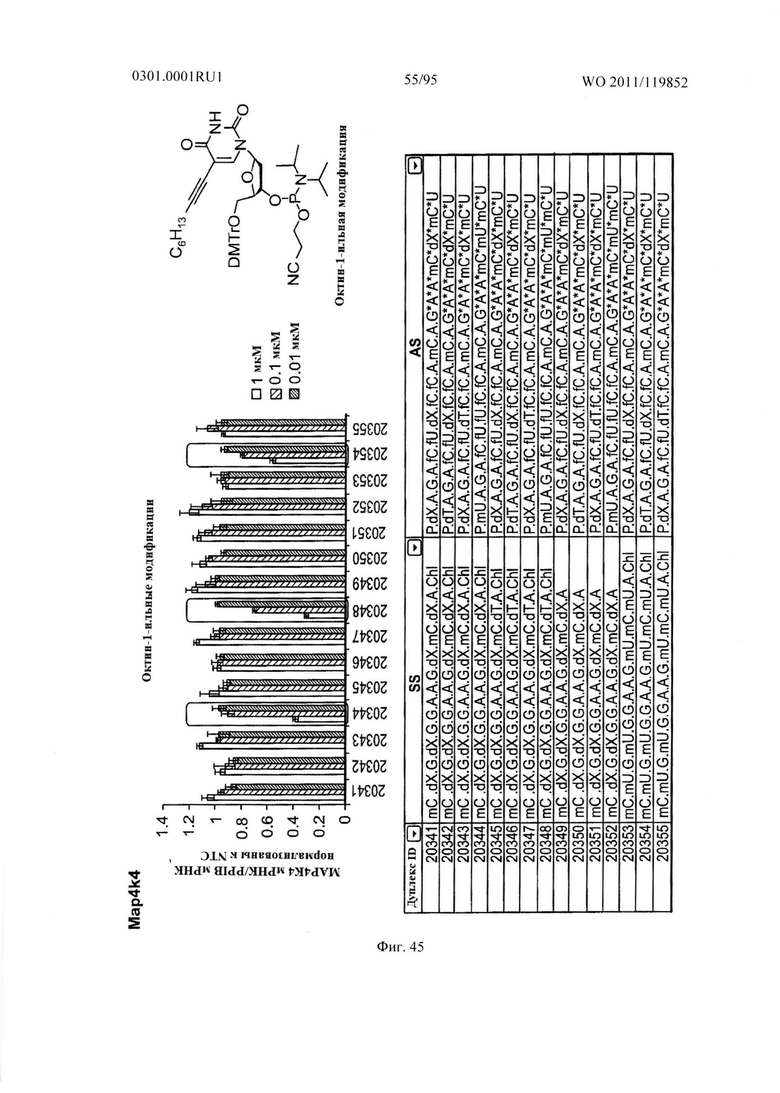
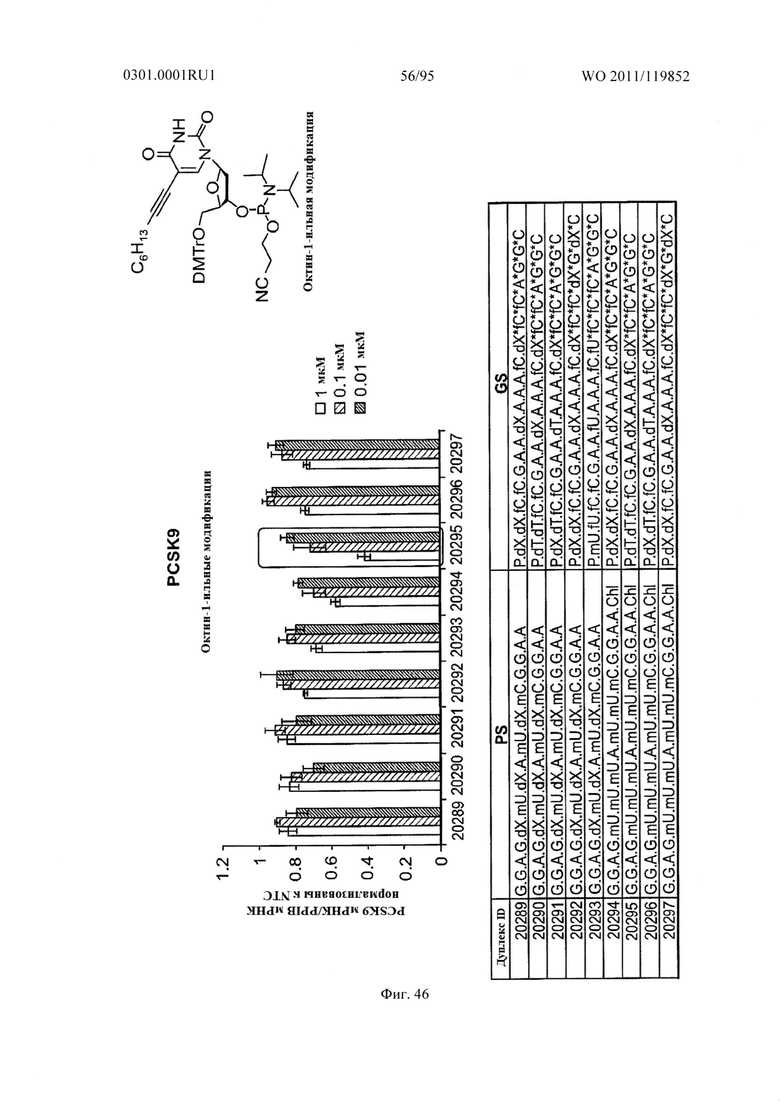
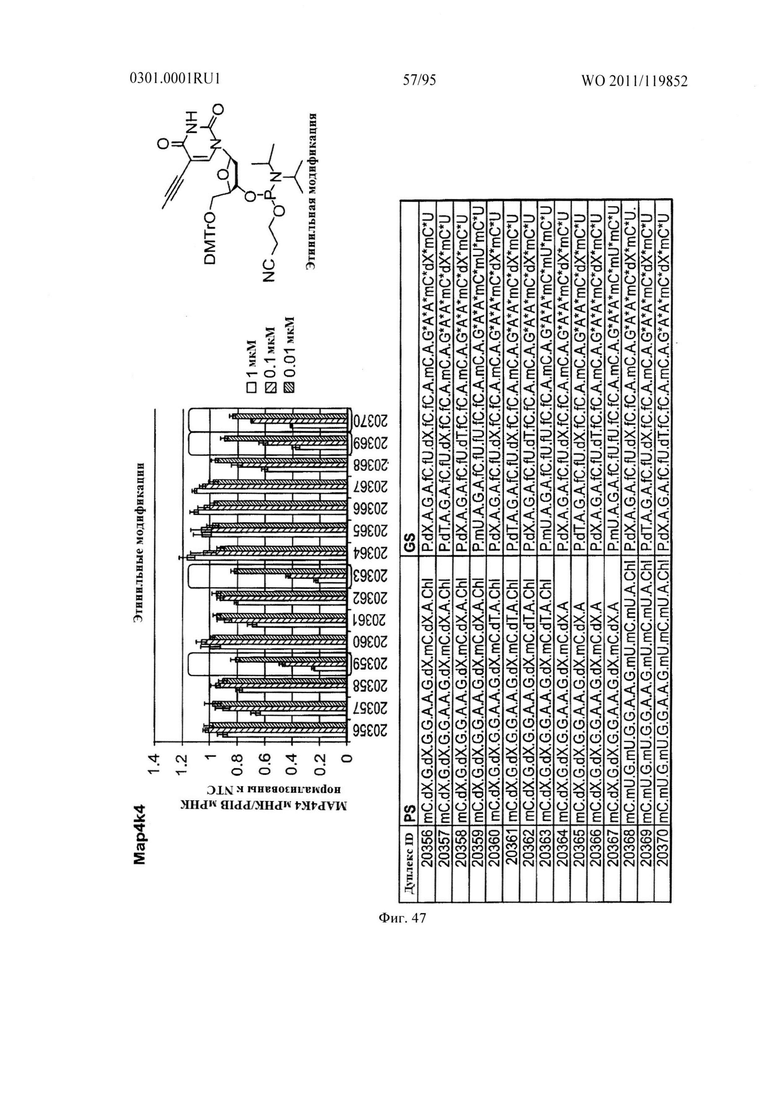

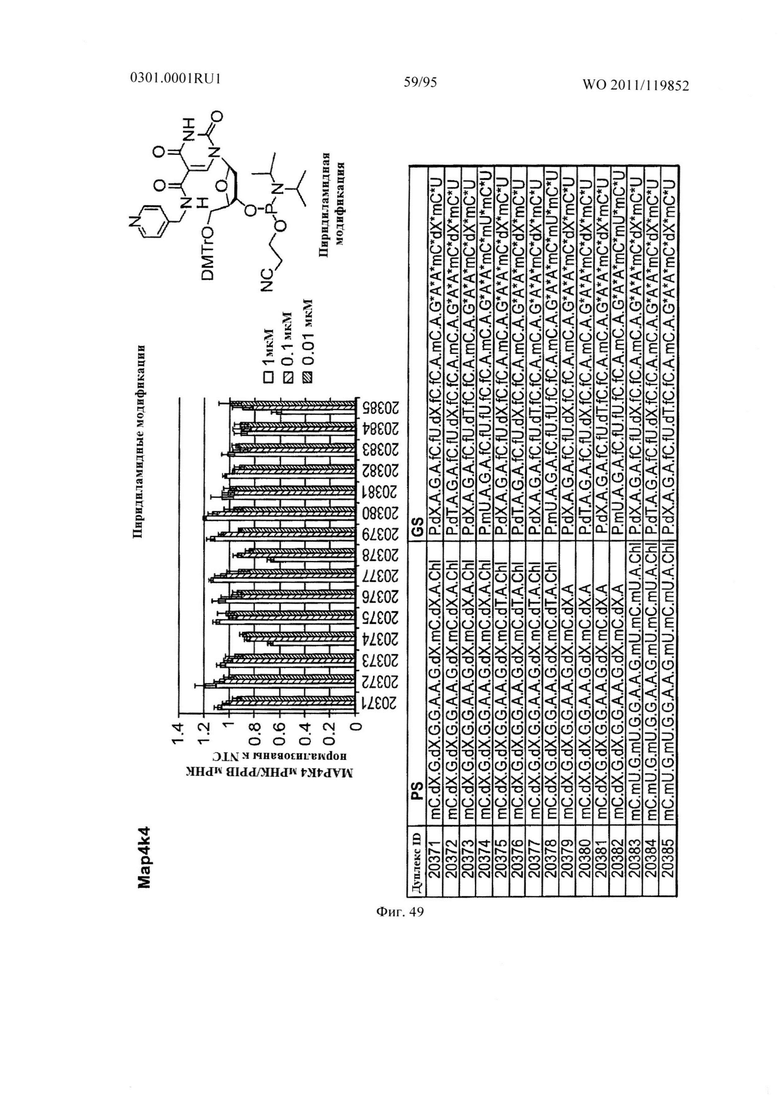


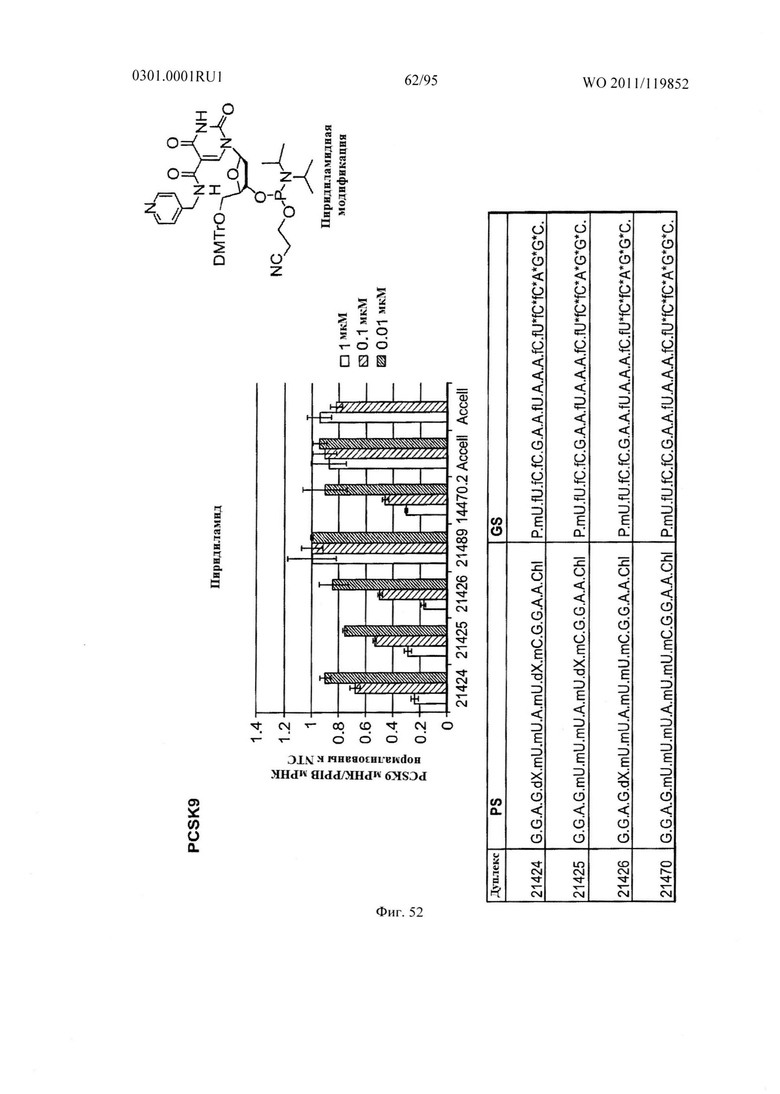
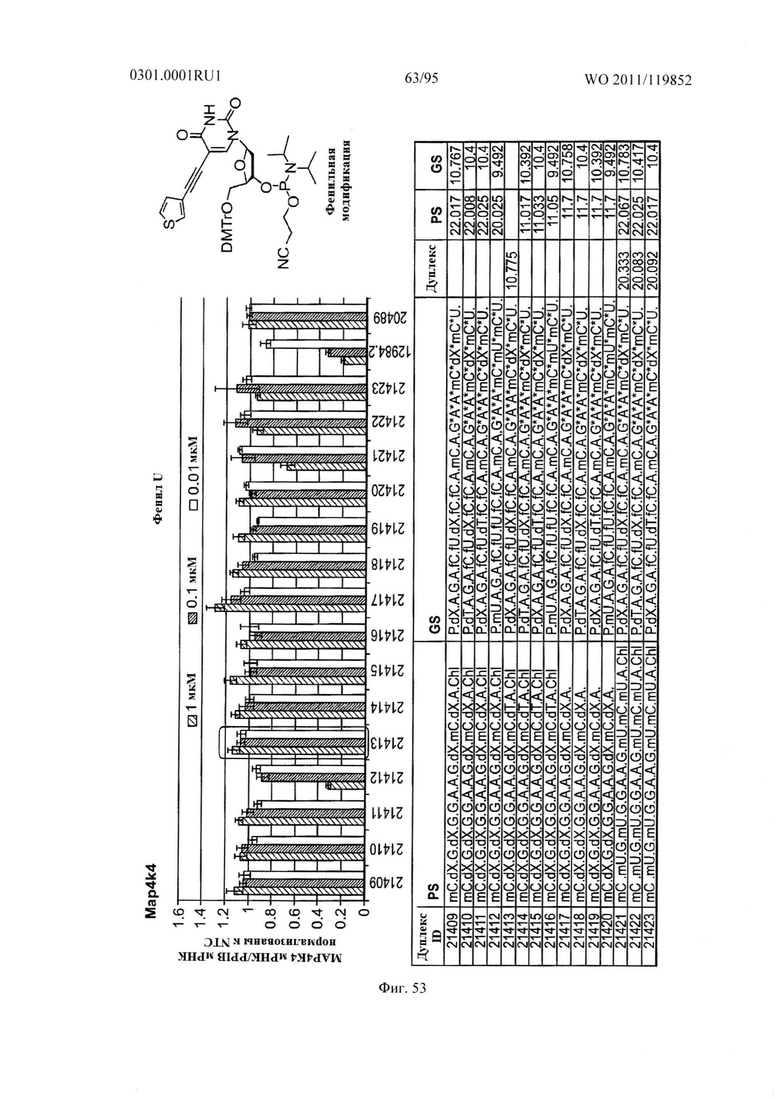
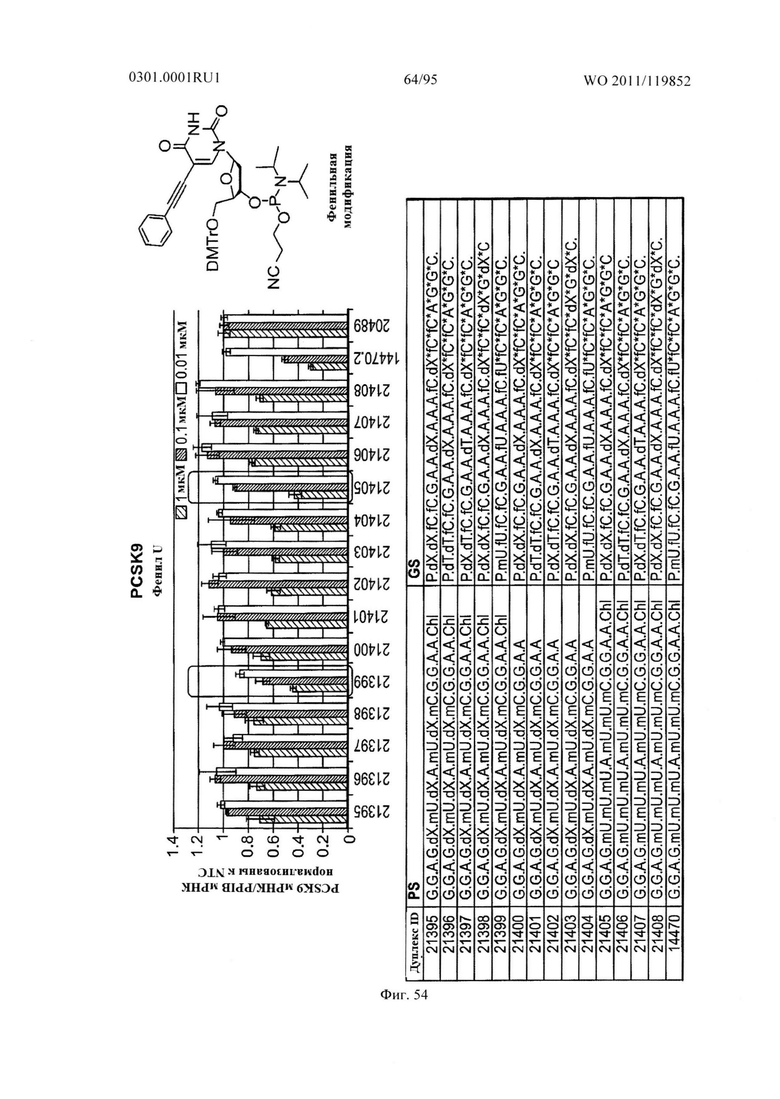

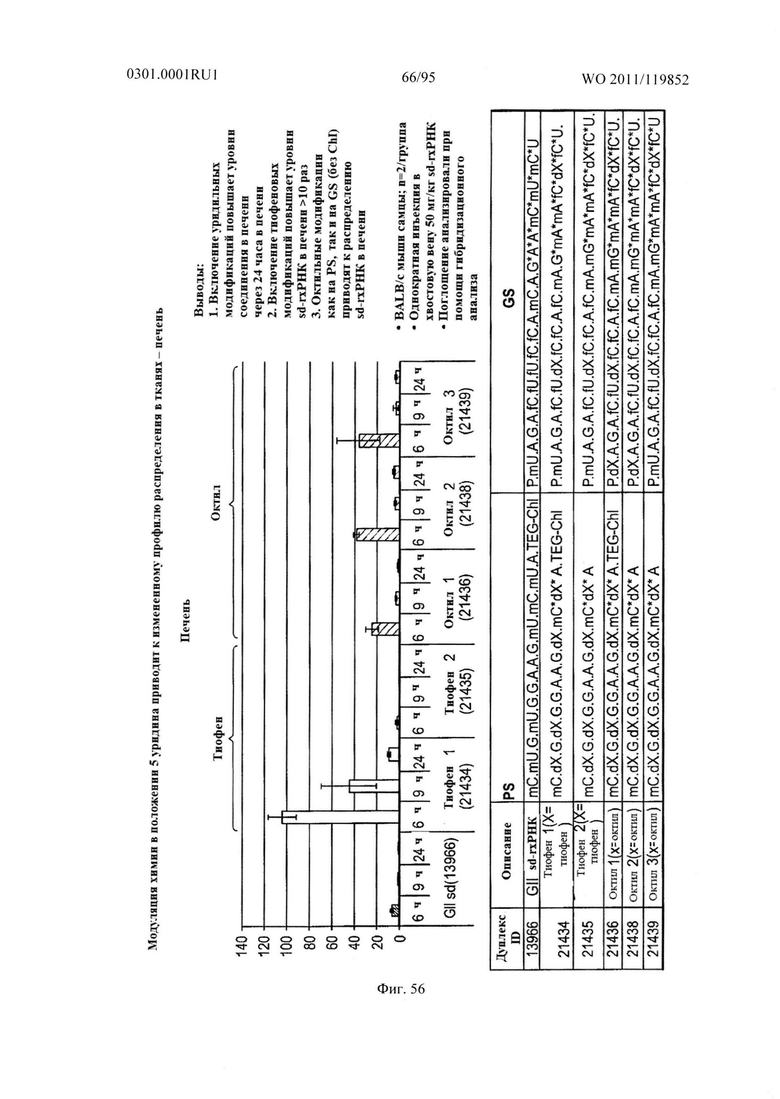



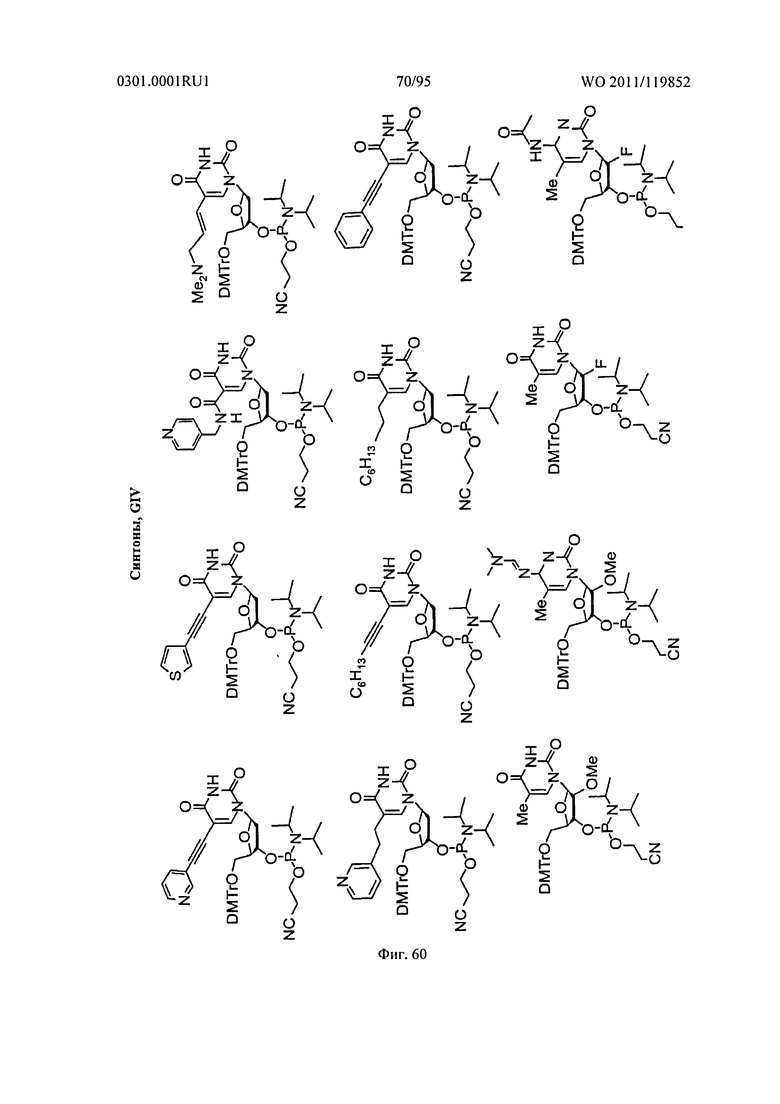

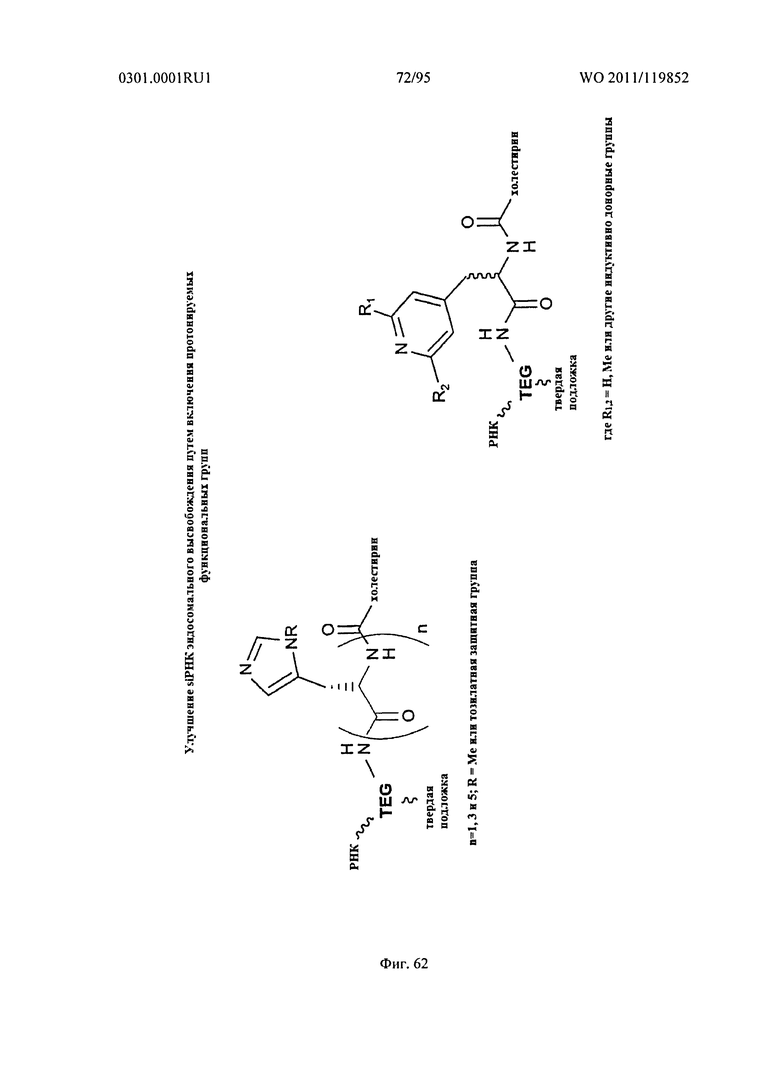
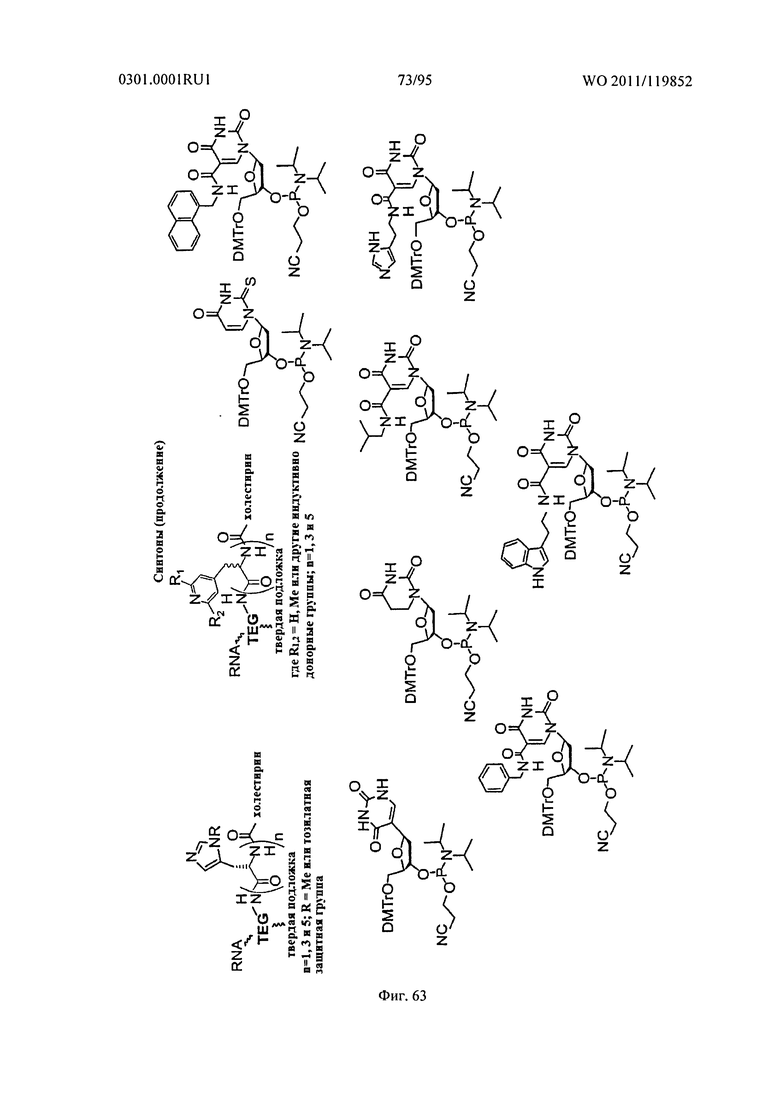





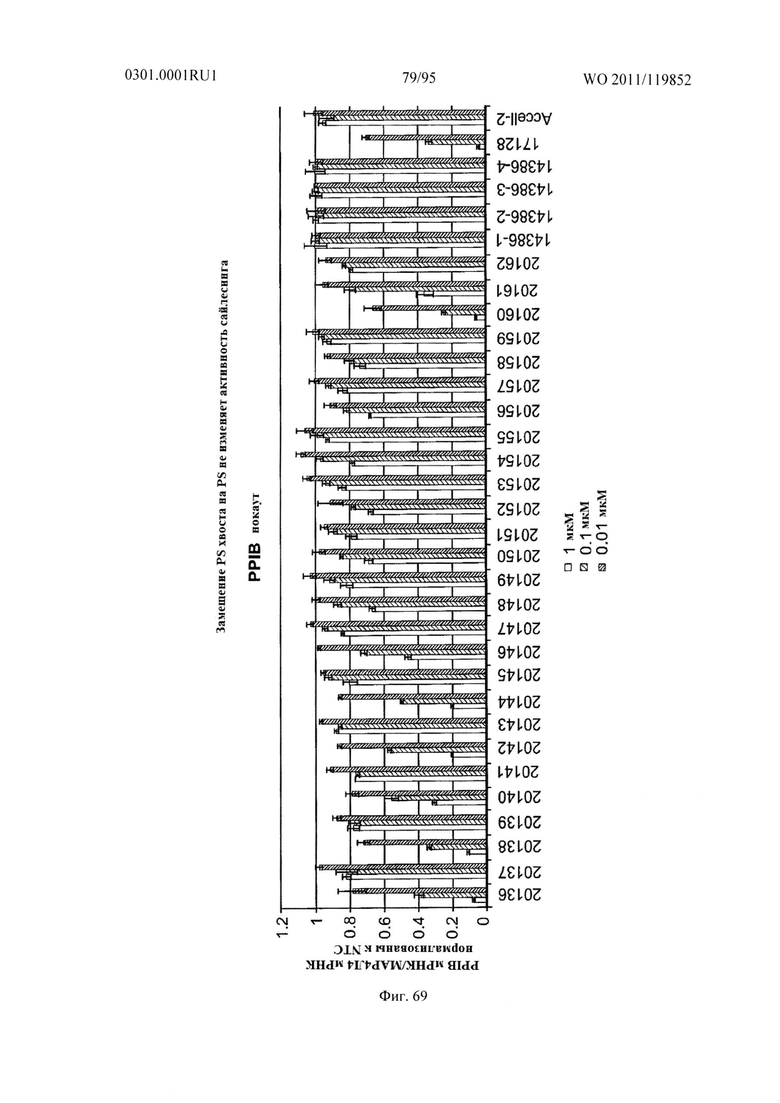
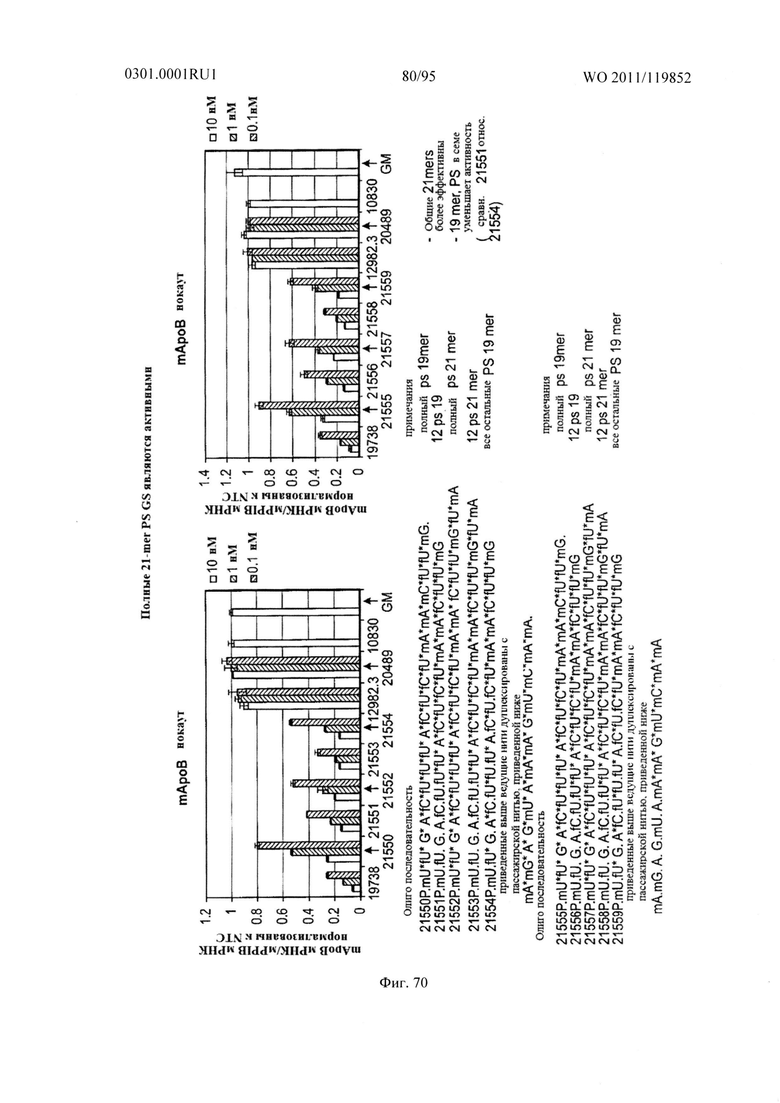














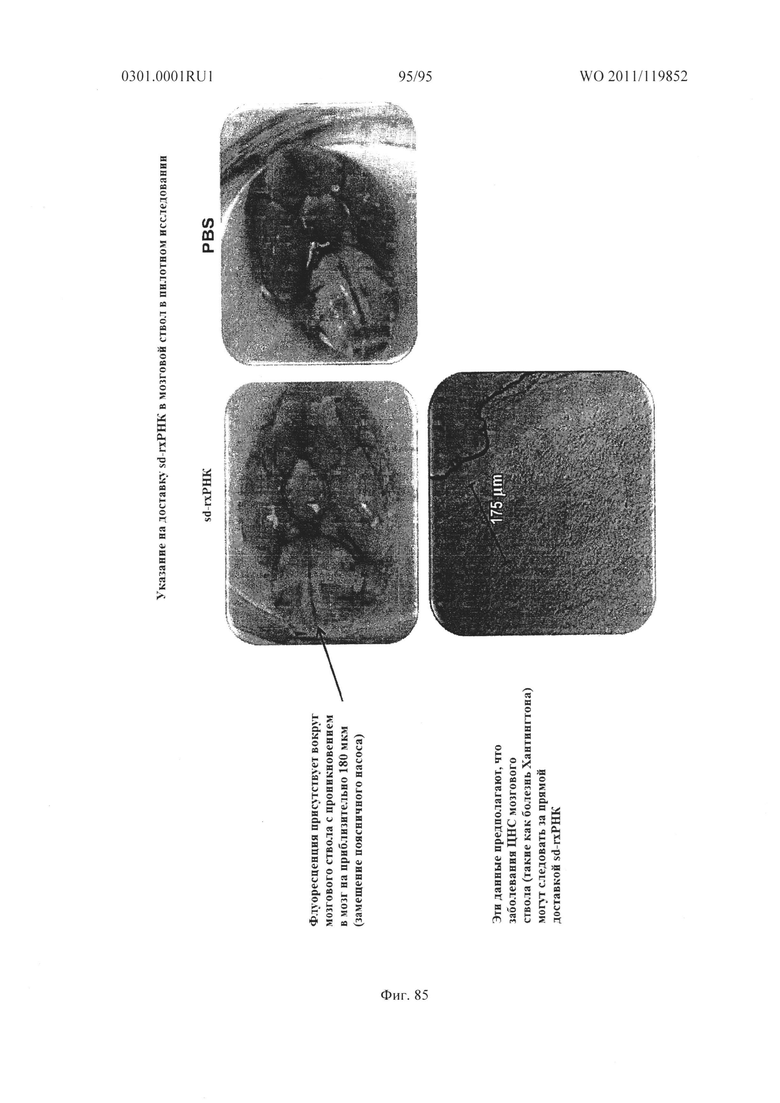
Изобретение относится к самодоставляющейся интерферирующей РНК (sd-РНК), которая может использоваться в медицине. Самодоставляющаяся интерферирующая sd-РНК содержит ведущую нить и пассажирскую нить, причем sd-РНК содержит двухцепочечную область и одноцепочечную область, где одноцепочечная область расположена на 3'-конце ведущей нити, двухцепочечная область имеет длину 8-15 нуклеотидов, а одноцепочечная область имеет длину 4-12 нуклеотидов, по меньшей мере 40% нуклеотидов sd-РНК модифицированы, причем модификации выбраны из группы, включающей 5'-фосфорилирование, фосфоротиоатную модификацию каркаса, 2'-дезокси-, 2'F- и 2'O-метильную модификацию рибозы, гидрофобную модификацию U и/или С, причем гидрофобная модификация находится в положении 4 или 5 U и/или С, по меньшей мере два U и/или С включают гидрофобную модификацию, выбранную из группы, состоящей из метильной, октильной, тиофеновой, октин-1-ильной, этинильной, пиридиламидной, изобутильной, фенильной, 4-пиридильной, 2-пиридильной, индолильной, бутильной, нафтильной и имидазольной модификации; одноцепочечная область содержит 3, 4, 5, 6, 7, 8, 9, 10, 11 или 12 фосфоротиоатных модификаций; и модификации включают по меньшей мере одну 2'F- или 2'O-метильную модификацию; sd-РНК необязательно содержит по меньшей мере одну дополнительную одноцепочечную область и sd-РНК необязательно содержит холестерин или длинноцепочечный алкилхолестериновый аналог, присоединенный к 3'-концу непосредственно или посредством линкерного фрагмента. Предложены новые самодоставляющиеся интерферирующие РНК и композиции на их основе, эффективные для сайленсинга генов. 2 н. и 8 з.п. ф-лы, 85 ил., 1 табл., 7 пр., список последовательностей.
1. Самодоставляющаяся интерферирующая РНК (sd-РНК), содержащая:
ведущую нить и пассажирскую нить, причем sd-РНК содержит двухцепочечную область и одноцепочечную область, причем одноцепочечная область расположена на 3'-конце ведущей нити, причем двухцепочечная область имеет длину 8-15 нуклеотидов, а одноцепочечная область имеет длину 4-12 нуклеотидов,
причем по меньшей мере 40% нуклеотидов sd-РНК модифицированы, причем модификации выбраны из группы, включающей 5'-фосфорилирование, фосфоротиоатную модификацию каркаса, 2'-дезокси-, 2'F- и 2'O-метильную модификацию рибозы, гидрофобную модификацию U и/или С, причем гидрофобная модификация находится в положении 4 или 5 U и/или С, причем:
по меньшей мере два U и/или С включают гидрофобную модификацию, выбранную из группы, состоящей из метильной, октильной, тиофеновой, октин-1-ильной, этинильной, пиридиламидной, изобутильной, фенильной, 4-пиридильной, 2-пиридильной, индолильной, бутильной, нафтильной и имидазольной модификации;
одноцепочечная область содержит 3, 4, 5, 6, 7, 8, 9, 10, 11 или 12 фосфоротиоатных модификаций; и
модификации включают по меньшей мере одну 2'F- или 2'O-метильную модификацию;
причем sd-РНК необязательно содержит по меньшей мере одну дополнительную одноцепочечную область;
и причем sd-РНК необязательно содержит холестерин или длинноцепочечный алкилхолестериновый аналог, присоединенный к 3'-концу непосредственно или посредством линкерного фрагмента.
2. sd-РНК по п. 1, отличающаяся тем, что по меньшей мере 60% нуклеотидов модифицированы.
3. sd-РНК по любому из пп. 1, 2, отличающаяся тем, что ведущая нить содержит одноцепочечную область, содержащую по меньшей мере 5 фосфоротиоатных модификаций.
4. sd-РНК по п. 1, отличающаяся тем, что ведущая нить имеет длину 19, 20, 21, 22, или 23 нуклеотида и пассажирская нить имеет длину 13-15 нуклеотидов и по меньшей мере 50% нуклеотидов в молекуле sd-РНК является фосфоротиоатно модифицированными.
5. sd-РНК по п. 1, отличающаяся тем, что по меньшей мере 50% нуклеотидов в молекуле sd-РНК является фосфоротиоатно модифицированными.
6. Фармацевтическая композиция, полезная для сайленсинга генов, содержащая эффективное количество sd-РНК по любому из пп. 1-5 и фармацевтически приемлемый носитель или разбавитель.
7. Композиция по п. 6, отличающаяся тем, что композиция содержит две или более разные sd-РНК.
8. Композиция по п. 6, отличающаяся тем, что композиция составлена для внутривенного введения.
9. Композиция по п. 6, отличающаяся тем, что композиция составлена для подкожного введения.
10. Композиция по п. 6, отличающаяся тем, что композиция составлена для интратекального введения.
| WO 2010011346 A1, 28.01.2010 | |||
| RHED CHIP COMPANIES, REDCHIP BLOGGER ON LINE, 15.05.14 | |||
| RHED CHIP SMALL-CAP INVESTOR`S CONFERENCE, 16.06.2009, p | |||
| Топка с несколькими решетками для твердого топлива | 1918 |
|
SU8A1 |
| US 20080311040 A1, 18.12.2008 | |||
| RU 2007110823 A, 10.10.2008 | |||
| WO 2009045457 A2, 09.04.2009 | |||
| WO 2010008582 A2, 21.01.2010. | |||

