Перекрестная ссылка на родственную заявку
[0001] По настоящей заявке испрашивается приоритет предварительной заявки США № 61/550170, поданной 21 октября 2011 г., которая включена в настоящее описание посредством ссылки во всей полноте.
Уровень техники раскрытия
[0002] Род Brassica включает канолу, одну из наиболее важных мировых масличный культур и важную масличную культуру, выращиваемую в умеренных широтах. Канолу традиционно характеризуют как Brassica napus L. (вид, выведенный в результате межвидового скрещивания Brassica rapa и Brassica oleracea), в который эруковая кислота и глюкозинолаты были устранены или значительно уменьшены посредством обычной селекции. Большая часть масла канолы имеет форму растительных масел, производимых для потребления человеком. Существует также растущий рынок использования масла канолы в промышленных применениях.
[0003] Род Brassica состоит из трех диплоидных видов, каждый из которых обладает уникальным геномом, которые обозначают как геном A, геном B или геном C. Растения Brassica rapa обладают диплоидным геномом A. Растения Brassica nigra обладают диплоидным геномом B. Растения Brassica oleracea обладают диплоидным геномом C. Гибриды данных видов могут быть получены посредством скрещивания между двумя диплоидными видами. Канола является амфидиплоидным видом, который, как полагают, произошел от гибридизации Brassica oleracea, имеющей диплоидный геном C, и Brassica rapa, имеющей диплоидный геном A. Цитогенетические исследования показали, что геномы AA и CC демонстрируют такую степень родства, что являются частично гомологичными друг другу, и полагают, что они произошли от генома общего предка (Prakash and Hinata, 1980). Хотя технически их классифицируют как диплоиды, геномы обоих видов-предшественников содержат большое количество участков, дублирующих друг друга (Song et al., 1991). Генетический анализ показал, что геном AA Brassica rapa внес десять хромосом в Brassica napus, тогда как Brassica oleracea внесла девять хромосом из своего генома CC в качестве материнского донора (Song et al., 1992).
[0004] Качество пищевого и технического масла, получаемого из конкретного сорта семян канолы, определяют по составляющим его жирным кислотам, поскольку тип и уровень ненасыщенности жирных кислот имеет последствия как для пищевых, так и для технических применений. Обычное масло канолы содержит приблизительно 60% олеиновой кислоты (C18:1), 20% линолевой кислоты (C18:2) и 10% линоленовой кислоты (18:3). Количество полиненасыщенной линоленовой кислоты, типичное для обычной канолы, нежелательно, поскольку масло легко подвергается окислению, причем на скорость окисления воздействуют несколько факторов, включая присутствие кислорода, воздействие света и тепла, и присутствие собственных или добавленных антиоксидантов и прооксидантов в масле. Окисление приводит к неприятным привкусам и прогорклому вкусу в результате повторной жарки (индуцированное окисление) или хранения в течение продолжительного периода (автоокисление). Окисление также может изменять смазочные и вязкие свойства масла канолы.
[0005] Профили масла канолы, которые демонстрируют пониженный уровень полиненасыщенных жирных кислот и повышенный уровень мононенасыщенной олеиновой кислоты по сравнению с обычным маслом канолы, связаны с более высокой устойчивостью к окислению. Подверженность отдельных жирных кислот окислению зависит от степени их ненасыщенности. Таким образом, скорость окисления линоленовой кислоты, которая имеет три двойные связи углерод-углерод, в 25 раз выше, чем олеиновой кислоты, которая имеет только одну двойную связь углерод-углерод, и в 2 раза выше, чем линолевой кислоты, которая имеет две двойные связи углерод-углерод. Линолевая и линоленовая кислоты также обладают наибольшим воздействием на запах и аромат, поскольку они легко образуют гидропероксиды. Масло с высоким содержанием олеиновой кислоты (содержание олеиновой кислоты >=70%) менее подвержено окислению во время хранения, жарки и рафинирования, и может нагреваться до более высокой температуры без дымления, что делает его более подходящим в качестве масла для приготовления пищи.
[0006] Качество масла канолы определяют по составляющим его жирным кислотам, таким как олеиновая кислота (C18:1), линолевая кислота (C18:2) и линоленовая кислота (C18:3). Большинство культиваров канолы обычно дают масло с приблизительно 55-65% олеиновой кислоты и 8-12% линоленовой кислоты. Высокие концентрации линоленовой кислоты приводят к нестабильности масла и нетипичному запаху, тогда высокий уровень олеиновой кислоты повышает устойчивость к окислению и пищевую ценность масла. Следовательно, разработка культиваров канолы с повышенным содержанием олеиновой кислоты и пониженным содержанием линоленовой кислоты очень желательна для качества масла канолы.
[0007] Было идентифицировано два локуса, и определена их геномная локализация, из культивара канолы, который обладает повышенным содержанием олеиновой кислоты и пониженным содержанием линоленовой кислоты. Один локус обладает значительным воздействием, и второй локус обладает незначительным воздействием на производство повышенного содержания олеиновой кислоты и пониженного содержания линолевой кислоты. Определено, что основным локусом для высокого содержания олеиновой кислоты (C18:1) является ген десатуразы-2 жирных кислот (fad-2), и он расположен на группе сцепления N5. Второй локус расположен на группе сцепления N1. Одним из основных локусов количественных признаков (QTL) для линоленовой кислоты (C18:3) является ген десатуразы-3 жирных кислот генома C (fad-3c), и он расположен на группе сцепления N14. Второй основной QTL располагается на группе сцепления N4 и представляет собой ген десатуразы-3 жирных кислот генома A (fad-3a). Геномные последовательности генов fad-2 и fad-3c амплифицировали и секвенировали как из индуцированного этилметансульфонатом (ЭМС) мутанта, так и из культивара канолы дикого типа. Сравнение последовательностей аллелей мутанта и дикого типа генов fad-2 и fad-3c показало наличие однонуклеотидных полиморфизмов (SNP) в генах растений, мутировавших под действием ЭМС. На основании различий последовательностей между аллелями мутанта и дикого типа было разработано два SNP-маркера, соответствующих мутациям генов fad-2 и fad-3c (Hu et al., 2006).
[0008] Современные способы производства семян гибрида F1 Brassica имеют ограничения в отношении стоимости и чистоты семян. Обычно для данных способов требуются стабильные, сибс-несовместимые и самонесовместимые, близкие к гомозиготным родительские линии разведения, каковые родительские линии разведения доступны только после повторного самоопыления для создания инбредных линий. Кроме того, инбридинг для развития и сохранения родительских линий осуществляется посредством трудоемких методов, таких как опыление бутонов, поскольку системы производства гибридных семян Brassica, основанные на самонесовместимых признаках, должны использовать строго самонесовместимые растения. Условия окружающей среды во время процесса разведения, такие как температура и влажность, обычно влияют на липидный метаболизм растений, таким образом также влияя на содержание жирных кислот (Harwood, 1999). Изменчивость окружающей среды, следовательно, делает фенотипическую селекцию растений менее надежной. Deng и Scarth (1998) обнаружили, что повышение температуры после цветения значительно снижает уровень C18:3 и повышает C18:1. Аналогичные результаты приводились в других исследованиях (Yermanos and Goodin, 1965; Canvin, 1965).
[0009] Селекция сортов с низким содержанием линоленовой кислоты является особенно сложной, поскольку содержание C18:3 является мультигенным признаком и наследуется рецессивным образом с относительно низкой наследуемостью. Генетический анализ популяции, получаемой от скрещивания между "Stellar" (имеющим низкое содержание C18:3 (3%)) и "Drakkar" (имеющим "обычный" уровень C18:3 (9-10%)), показало, что признак низкого содержания C18:3 управляется двумя основными локусами с аддитивным действием, обозначаемыми L1 и L2 (Jourdren et al., 1996b). Было обнаружено, что эти два основных локуса, управляющих содержанием C18:3, соответствуют двум генам fad-3 (десатуразы-3 жирных кислот); один расположен в геноме A (происходящем от Brassica rapa), а другой в геноме C (происходящем от Brassica olecera) (Jourdren et al., 1996; Barret et al., 1999).
[0010] Признаки, которые непрерывно изменяются из-за генетических (аддитивных, доминантных и эпистатических) и относящихся к окружающей среде факторов, обычно называют "количественными признаками". Количественные признаки можно отличать от "качественных" или "дискретных" признаков на основании двух факторов: воздействия окружающей среды на экспрессию генов, которое производит непрерывное распределение фенотипов; и сложной схемы расщепления, производимой мультигенным наследованием. Идентификация одного или нескольких участков генома, связанных с экспрессией количественного признака, привело к открытию локусов количественных признаков ("QTL"). Thormann et al., (1996) локализовали два QTL, которые объясняли 60% изменчивости содержания линоленовой кислоты, тогда как Somers et al., (1998) идентифицировали три QTL, которые вместе объясняли 51% фенотипической изменчивости содержания C18:3. Chen и Beversdorf (1990) также описали аддитивную модель трех локусов. Rucker и Robbelen (1996) указали, что в стадии десатурации с наибольшей вероятностью принимают участие несколько минорных генов.
[0011] Наследуемость содержания C18:3 была определена равной 26-59% (Kondra and Thomas, 1975) (где изменчивость наследуемости является функцией генетики, а не факторов окружающей среды). Сложность наследования линоленовой кислоты может объясняться тем фактом, что линоленовая кислота может синтезироваться как десатурацией C18:2, так и удлинением C16:3 (Thompson, 1983).
[0012] В отличие от линоленовой кислоты наследование олеиновой кислоты является менее сложным, и наследуемость олеиновой кислоты относительно высока. Сообщалось, что высокое содержание олеиновой кислоты контролируется основным локусом, называемым ген fad-2 (десатуразы жирных кислот 2), который кодирует фермент, отвечающий за десатурацию олеиновой кислоты до линолевой кислоты (C18:2) (Tanhuanpaa et al., 1998; Schierholt et al., 2001). Все копии функционального гена fad-2, о которых было сообщено, и которые были локализованы к настоящему времени, расположены на происходящей из генома A группы сцепления N5 (Scheffler et al., 1997; Schierholt et al., 2000). Chen и Beversdorf (1990) сообщали, что накопление олеиновой кислоты контролируется двумя генетическими системами расщепления, причем одна действует на удлинение цепи, а другая использует десатурацию. Наследуемость содержания C18:1 определяли равной от 53% до 78% (Kondra and Thomas 1975) и 94% (Schierholt and Becker, 1999) соответственно. Благодаря более высокой наследуемости экспрессия содержания C18:1 менее подвержена воздействию окружающей среды и относительно стабильна (Schierholt and Becker, 1999).
[0013] В зародышевой плазме канолы Nexera™ от 1 до 2 генов, как обнаружено, контролируют содержание C18:1, и по меньшей мере 3 гена участвуют в экспрессии C18:3 (Nexera™ является товарным знаком Dow AgroSciences, LLC). В потомстве после расщепления распределение содержания C18:3 в семенах является непрерывным, что делает сложной идентификацию генотипических классов с желаемым уровнем C18:3. Кроме того, низка корреляция содержания жирных кислот между растениями, выращиваемыми в оранжерее (GH) и в поле, что делает еще более сложным надежную селекцию растений с желаемым уровнем C18:3 в GH.
[0014] Для детектирования наличия определенного гена в образце ткани растения можно использовать различные способы. Одним примером является метод пиросеквенирования, описанный Winge (Innov. Pharma. Tech. 00:18-24, 2000). В данном способе конструируют олигонуклеотид, который перекрывает вставленную последовательность ДНК и геномную ДНК, примыкающую к ней. Олигонуклеотид подвергают гибридизации с одноцепочечным продуктом ПЦР ("ампликоном") от представляющего интерес участка (т.е. один праймер во вставленной последовательности и один во фланкирующей геномной последовательности) и инкубируют в присутствии ДНК-полимеразы, АТФ-сульфурилазы, люциферазы, апиразы, аденозин-5'-фосфосульфата и люцеферина. dNTP добавляют по отдельности, и их включение приводит к световому сигналу, который измеряют. Световой сигнал указывает на наличие трансгенной вставки/фланкирующей последовательности за счет успешной амплификации, гибридизации и удлинения на одно или несколько оснований. (Данный метод обычно применяют для начального секвенирования, не для детектирования конкретного гена, когда он известен).
[0015] Флуоресцентная поляризация является другим способом, который можно использовать для детектирования ампликона. При следовании данному способу, олигонуклеотид конструируют так, чтобы перекрывать соединение геномной фланкирующей и вставленной ДНК. Олигонуклеотид подвергают гибридизации с одноцепочечным продуктом ПЦР от представляющего интерес участка (один праймер во вставленной ДНК и один во фланкирующей геномной последовательности ДНК) и инкубируют в присутствии ДНК-полимеразы и флуоресцентно-меченного ddNTP. Удлинение на одно основание приводит к включению ddNTP. Включение можно измерять как изменение поляризации с применением флуориметра. Изменение поляризации указывает на наличие трансгенной вставки/фланкирующей последовательности за счет успешной амплификации, гибридизации и удлинения на одно основание.
[0016] Описано применение молекулярных маяков при определении последовательности. Кратко говоря, молекулярные маяки содержат олигонуклеотидный зонд FRET (резонансный перенос энергии флуоресценции), который можно конструировать так, чтобы FRET-зонд перекрывал соединение фланкирующей геномной ДНК и ДНК вставки. Уникальная структура FRET-зонда приводит к тому, что он имеет вторичную структуру, которая удерживает флуоресцентный и гасящий фрагменты в непосредственной близости. FRET-зонд и праймеры ПЦР (один праймер в последовательности ДНК вставки и один во фланкирующей геномной последовательности) проходят цикл в присутствии термостабильной полимеразы и dNTP. После успешной ПЦР-амплификации гибридизация FRET-зонда с мишеневой последовательностью приводит к исчезновению вторичной структуры зонда и пространственного разделения флуоресцентного и гасящего фрагментов. Флуоресцентный сигнал указывает на наличие фланкирующей геномной последовательности/последовательности трансгенной вставки за счет успешной амплификации и гибридизации.
[0017] Анализ гидролиза зондов, также известный как TaqMan® ПЦР (TaqMan® является зарегистрированным торговым знаком Roche Molecular Systems, Inc.), предлагает способ детектирования и количественного определения наличия последовательности ДНК. Кратко говоря, TaqMan® ПЦР использует олигонуклеотидный FRET-зонд, который сконструирован так, чтобы часть олигомера была расположена в трансгене, а другая часть олигомера во фланкирующей геномной последовательности для детектирования по событию. FRET-зонд и праймеры ПЦР (один праймер в последовательности ДНК вставки и один во фланкирующей геномной последовательности) проходят цикл в присутствии термостабильной полимеразы и dNTP. Гибридизация FRET-зонда и последующее расщепление во время стадии ПЦР-амплификации вследствие 5'-экзонуклеазной активности Taq-полимеразы приводит к отщеплению и освобождению флуоресцентного фрагмента от гасящего фрагмента на FRET-зонде. Флуоресцентный сигнал указывает на наличие фланкирующей последовательности/последовательности трансгенной вставки за счет успешной гибридизации и амплификации.
[0018] Для специфической к последовательности идентификации ДНК также используют молекулярные маркеры. Селекция по молекулярным маркерам основана на генотипах и, следовательно, не зависит от воздействия окружающей среды. Молекулярные маркеры помогают уменьшить проблему ненадежности селекции растения в оранжерее, возникающую из-за низкой корреляции содержания жирных кислот между выращиваемыми в оранжерее растениями и выращиваемыми в поле растениями. Важно отметить, что молекулярные маркеры, тесно связанные с генами, контролирующими содержание C18:1 и C18:3, могут облегчать раннюю селекцию растений, несущих гены для высокого содержания C18:1 и низкого содержания C18:3. Маркерзависимая селекция на ранней стадии может способствовать сохранению пространства в оранжерее, повышает эффективность использования оранжереи и уменьшает трудоемкость разведения в поле.
[0019] В более общем смысле, преимущества молекулярных маркеров над морфологическими маркерами заключаются в том, что: молекулярные маркеры могут быть высоко полиморфными, тогда как морфологические маркеры строго зависят от фенотипа; морфологические маркеры могут вмешиваться в подсчет некоторых количественных фенотипов, тогда как молекулярные маркеры демонстрируют взаимосвязь между генотипом и фенотипом 1:1 (тем самым делая возможным однозначный подсчет всех возможных генотипов для заданного локуса); и эпистатические взаимодействия способствуют ограничению количества морфологических маркеров, подходящих для популяции, тогда как молекулярные маркеры не взаимодействуют эпистатически.
[0020] Было определено, что различные типы молекулярных маркеров, такие как маркеры RAPD (случайно амплифицируемая полиморфная ДНК) (Tanhuanpaa et al., 1995; Hu et al., 1995; Rajcan et al., 1999; Jourdren et al., 1996), маркеры RFLP (полиморфизм длины рестрикционых фрагментов) (Thormann et al., 1996) и маркеры SCAR (амплифицированный участок с известной последовательностью) (Hu et al., 1999), связаны с низким уровнем C18:3 в Brassica napus. Также были определены молекулярные маркеры для высокого содержания C18:1. Было определено, что маркер RAPD связан с QTL, воздействующим на концентрацию олеиновой кислоты в весенней репе масличной (B. rapa ssp. Oleifera), и позже он был превращен в маркер SCAR (Tanhuanpaa et al., 1996). Schierholt et al., (2000) идентифицировали три маркера AFLP (полиморфизм длины амплифицированных фрагментов), связанных с мутацией высокого содержания олеиновой кислоты в озимом масличном рапсе (B. napus L). Tanhuanpaa et al., (1998) разработали аллель-специфический маркер ПЦР для олеиновой кислоты посредством сравнения дикого типа и аллеля высокого содержания олеиновой кислоты локуса гена fad-2 в весенней репе масличной (B. rapa ssp. oleifera). Однако, большинство этих маркеров являются низкопроизводительными маркерами, такими как RAPD, AFLP и RFLP, и не подходят для масштабного скрининга с помощью автоматики.
Краткая сущность изобретения
[0021] Рассматриваемое раскрытие частично относится к TaqMan® ПЦР-анализам по конечной точке для детектирования гена fad-3c в каноле и высокопроизводительного анализа его зиготности. Рассматриваемое раскрытие, кроме того, относится, частично, к применению гена fad-3c дикого типа в каноле в качестве эталона для применения в определении зиготности. Можно использовать те или иные соответствующие процедуры, для того чтобы однозначно устанавливать зиготность и сорт линий канолы, содержащих рассматриваемый ген.
[0022] Рассматриваемое раскрытие также предлагает соответствующие наборы для определения зиготности и сорта по образцу (канолы, например).
[0023] Таким образом, вариант осуществления рассматриваемого раскрытия относится к TaqMan® ПЦР, гибкой платформе для высокопроизводительного анализа зиготности и скрещивания. Использование способа применения TaqMan® ПЦР по конечной точке, представляемого данным раскрытием, предлагает надежный, точный и высокопроизводительный способ применения для анализа зиготности fad-3c и скрещивания у канолы.
Краткое описание фигур
[0024] Фигура 1. представляет собой участок последовательности гена fad-3c (SEQ ID NO:1), иллюстрирующий положение мутации fad-3c, идентифицированной Hu et al., (2006) (стрелка). Интрон 6 напечатан более светлым текстом, а второй полиморфизм отмечен с помощью звездочки.
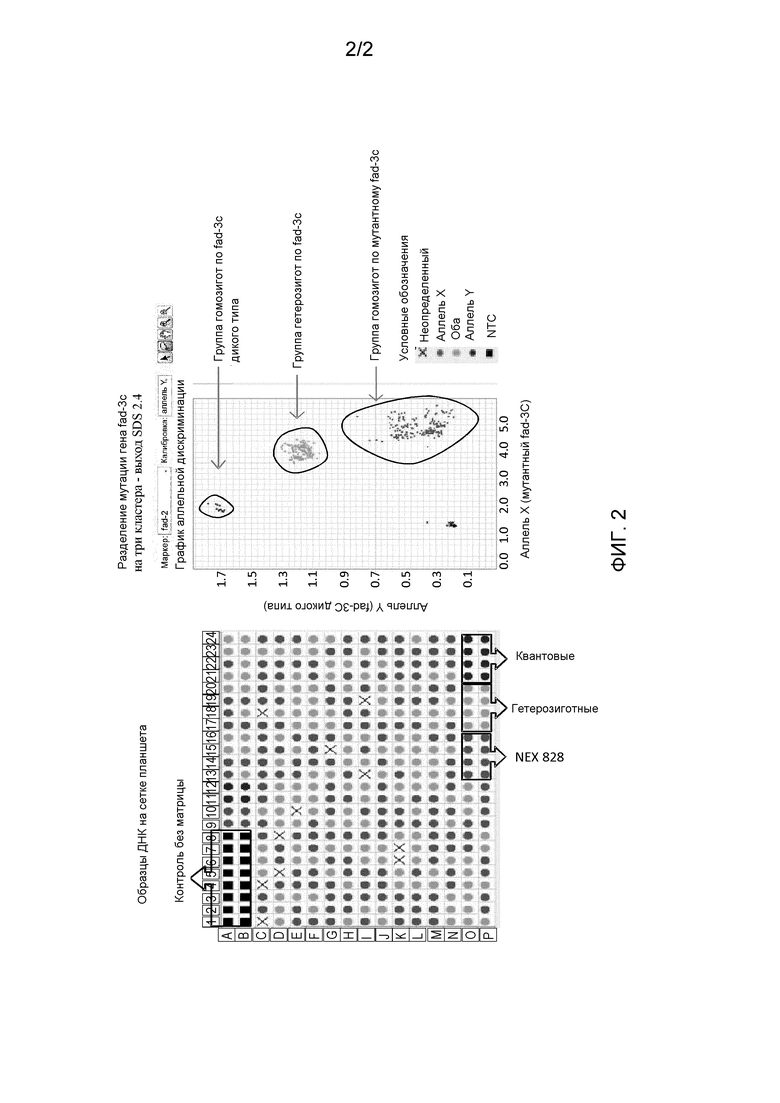
[0025] Фигура 2. представляет собой пример результатов анализа зиготности (канолы), демонстрирующий три генотипа fad-3c после TaqMan®-анализа по конечной точке (результаты получали с применением программного обеспечения SDS 2.4, доступного от Applied Biosystems, Foster City, CA, США).
Краткое описание последовательностей
[0026] SEQ ID NO:1 предлагает участок последовательности гена fad-3c, иллюстрирующий положение мутации fad-3c.
[0027] SEQ ID NO:2 предлагает прямой праймер D-CL-FAD3C-F (который связывается с фланкирующей геномной последовательностью).
[0028] SEQ ID NO:3 предлагает обратный праймер D-CL-FAD3C-R2 (который связывается с последовательностью вставки).
[0029] SEQ ID NO:4 предлагает зонд D-CL-FAD3C-FAM для преимущественного связывания мутировавшего гена fad-3c с однонуклеотидным полиморфизмом G->A.
[0030] SEQ ID NO:5 предлагает зонд D-CL-FAD3C-VIC для детектирования гена fad-3c дикого типа.
Подробное описание раскрытия
[0031] Рассматриваемое раскрытие частично относится к TaqMan® ПЦР-анализам по конечной точке для детектирования гена fad-3c в каноле и высокопроизводительного анализа его зиготности. Рассматриваемое раскрытие, кроме того, относится, частично, к применению гена fad-3c дикого типа в каноле в качестве эталона для применения в определении зиготности. Можно использовать те или иные соответствующие процедуры, для того чтобы однозначно устанавливать зиготность и сорт линий канолы, содержащих рассматриваемый ген. Рассматриваемое раскрытие также предлагает соответствующие наборы для определения зиготности и сорта по образцу (канолы, например). Таким образом, вариант осуществления рассматриваемого раскрытия относится к TaqMan® ПЦР, гибкой платформе для высокопроизводительного анализа зиготности и скрещивания. Использование способа применения TaqMan® ПЦР по конечной точке, представляемого данным раскрытием, предлагает надежный, точный и высокопроизводительный способ применения для анализа зиготности fad-3c и скрещивания у канолы.
[0032] Были разработаны новые анализы рассматриваемого изобретения, частично основанные на мутации однонуклеотидного полиморфизма (SNP) аллеля fad-3c в соответствии с данными Hu et al., (2006). Анализ использует два участка праймеров и два MGB-зонда для детектирования мутантных и дикого типа аллелей fad-3c (см. таблица 1). Праймеры и зонды TaqMan® для детектирования данной SNP-мутации конструировали частично посредством программного обеспечения Primer express (Applied Biosystems, Austin, Tx) с применением последовательностей гена fad-3c. Данный новый fad-3c TaqMan®-анализ проверяли с применением ДНК, экстрагированной из гомозиготных, гемизиготных и дикого типа (без мутации) по гену fad-3c растений канолы. Также частично оптимизировали fad-3c TaqMan®-анализ по эффективности с помощью системы ПЦР в реальном времени Applied Biosystems 7900HT как в 96, так и в 384-луночном формате с применением условий быстрого теплового цикла ПЦР.
Последовательности праймеров и зондов, применяемых в fad-3c TaqMan®-анализе
[0034] В анализе использовали образцы листьев NEX828 и Quantum. Для проверки данного анализа использовали ДНК из разводимых популяций канолы.
[0035] Аспекты рассматриваемого раскрытия включают способы конструирования и/или производства диагностических молекул нуклеиновой кислоты, которые приведены в качестве примера и/или предложены в настоящем описании. Конкретные TaqMan® праймеры и зонд были сконструированы, как подробно описано в настоящем описании, частично в соответствии с последовательностями ДНК, расположенными у конкретных SNP в гене fad-3c, указанных в настоящем описании, или поблизости от них против хода транскрипции или по ходу транскрипции.
[0036] Таким образом, в некоторых вариантах осуществления данное раскрытие относится к определению зиготности растений канолы, производящих масло. Рассматриваемое раскрытие частично относится к детектированию наличия SNP, указанных в настоящем описании, для того чтобы определять, содержит ли потомство от полового скрещивания представляющие интерес SNP, и зиготность потомства. Кроме того, включены способы детектирования зиготности, и они полезны, например, для соблюдения правил, требующих предпродажного разрешения и обозначения продуктов питания, получаемых из рекомбинантных сельскохозяйственных культур.
[0037] Рассматриваемое раскрытие частично относится к основанному на флуоресценции TaqMan® ПЦР-анализу по конечной точке, использующему эндогенный немутантный ген fad-3c в качестве контроля для высокопроизводительного анализа зиготности растений канолы.
[0038] Рассматриваемое раскрытие также частично относится к разработке биплексной TaqMan® ПЦР по конечной точке для анализа зиготности канолы. Кроме того, рассматриваемое раскрытие частично относится к разработке наборов для анализа селекции по гену канолы fad-3c.
[0039] В целом TaqMan®-анализы по конечной точке основаны на стратегии плюс/минус, в которой "плюс" обозначает, что образец положителен по анализируемому гену, а "минус" обозначает, что образец отрицателен по анализируемому гену. Данные анализы, как правило, используют один набор олигонуклеотидных праймеров и два олигонуклеотидных зонда, причем один зонд преимущественно гибридизуется с SNP мутировавшего fad-3c, а другой зонд преимущественно гибридизуется с последовательностью fad-3c дикого типа соответственно.
[0040] Преимущества, связанные с рассматриваемым раскрытием, включают уменьшение зависимости от качества и количества ДНК. Кроме того, рассматриваемое раскрытие не требует продолжительной начальной стадии денатурации, которая, если проведена неправильно, часто может приводить к неудаче других способов детектирования SNP. Дополнительно рассматриваемое раскрытие предлагает способ эффективного анализа большого количества образцов канолы высокопроизводительным образом в коммерческих условиях. Другим преимуществом рассматриваемого раскрытия является экономия времени. Рассматриваемый TaqMan®-анализ по конечной точке для анализа зиготности и скрещивания канолы предоставляет преимущества по сравнению с другими применяемыми форматами, особенно когда анализу подвергается большое количества образцов.
[0041] Данное раскрытие частично относится к анализу скрещивания растений. Данное раскрытие включает новые способы детектирования SNP в растениях канолы, которые влияют на уровни олеиновой и линоленовой кислот в рассматриваемых растениях.
[0042] Кроме того, может оказаться возможным детектировать наличие рассматриваемых SNP посредством других известных способов детектирования нуклеиновой кислоты, таких как ПЦР или гибридизация ДНК с применением зондов нуклеиновой кислоты, описанных в настоящем описании. В настоящем описании рассмотрены специфические к событию ПЦР-анализы (См. также Windels et al., (Med. Fac. Landbouww, Univ. Gent 64/5b:459462, 1999).
[0043] Как применяется в настоящем описании, термин "потомство" обозначает потомка любого поколения родительского растения.
[0044] Методы детектирования рассматриваемого раскрытия в сочетании с селекцией растений особенно полезны, например, для определения зиготности растений-потомков, после того как родительское растение, содержащее представляющий интерес SNP, скрещивают с другим растением. Рассматриваемые применение и способы полезны для программ селекции канолы, а также процессов контроля качества. Теперь можно изготавливать и применять наборы для детектирования с помощью ПЦР для линий канолы, использующие способы и анализы, раскрытые в настоящем описании. Кроме того, рассматриваемое раскрытие может быть полезно для регистрации продуктов и учета продуктов.
[0045] Растение канолы, содержащее желаемую генетическую композицию fad-3c, можно вывести посредством сначала полового скрещивания первого родительского растения канолы, являющегося растением канолы, выращенным из семени любой из линий, упомянутых в настоящем описании, и второго родительского растения канолы, посредством чего создается множество растений первого потомства; и затем отбора растений первого потомства, несущих желаемые гены fad-3c, как раскрыто в рассматриваемом раскрытии; и самоопыления растения первого потомства, посредством чего создается множество растений второго потомства; и затем отбора из растений второго потомства растения, которое несет желаемые гены fad-3c в соответствии с рассматриваемым раскрытием. Данные стадии могут дополнительно включать возвратное скрещивание растения первого потомства или растения второго потомства со вторым родительским растением канолы или третьим родительским растением канолы. Затем можно сеять урожай канолы, содержащий семена канолы рассматриваемого раскрытия или их потомство.
[0046] Данное раскрытие дополнительно включает способы осуществления скрещиваний с использованием растения канолы, содержащего желаемую генетическую композицию fad-3c в качестве по меньшей мере одного родителя. Например, рассматриваемое раскрытие включает F1-гибридное растение, имеющее в качестве одного или обоих родителей любое растение канолы, содержащее желаемую генетическую композицию fad-3c. Также в пределах рассматриваемого раскрытия лежат семена, получаемые посредством таких гибридов F1. Данное раскрытие включает способ идентификации семян гибрида F1 посредством скрещивания образца растения с другим (например инбредным родительским) растением и сбора и анализа получаемых семян гибрида с применением способа рассматриваемого раскрытия. Растения канолы, которые применяют для получения гибрида F1, могут представлять собой или материнскую форму, или отцовскую форму.
[0047] Также следует понимать, что можно получать трансгенные растения, содержащие гены fad-3c, раскрытые в настоящем описании. Дополнительно, трансгенные растения, имеющие характеристики гена fad-3c, раскрытые в настоящем описании, можно скрещивать с растением, содержащим другую генетическую композицию, тем самым создавая потомство, содержащее независимо сегрегирующиеся экзогенные гены. Самоопыление соответствующего потомства может производить растения, которые являются гомозиготными по добавленным экзогенным генам. Также предусмотрены возвратное скрещивание с родительским растением и ауткроссинг с нетрансгенным растением, как и вегетативное размножение. В данной области техники известны другие способы разведения, обычно используемые для различных признаков и культур. Возвратное скрещивание применяют для переноса генов легко интрогрессируемых высоко наследуемых признаков в желаемый гомозиготный культивар или инбредную линию, представляющие собой рекуррентного родителя. Источник признака, который должен быть перенесен, называют родителем-донором. Ожидается, что получаемое растение имеет атрибуты рекуррентного родителя (например, культивар) и желаемый признак, перенесенный от родителя-донора. После первоначального скрещивания выбирают особи, обладающие фенотипом родителя-донора, и повторно скрещивают их (подвергают возвратному скрещиванию) с рекуррентным родителем. Ожидается, что получаемый родитель имеет атрибуты рекуррентного родителя (например, культивар) и желаемый признак, перенесенный от родителя-донора. Способ рассматриваемого раскрытия предлагает высокопроизводительный основанный на флуоресценции TaqMan® ПЦР-анализ по конечной точке для детектирования трансгена fad-3c в растениях-потомках и для определения уровня зиготности растений-потомков.
[0048] Способы настоящего раскрытия, например олигонуклеотидные праймеры и зонды, можно использовать для способов маркерзависимой селекции (MAB). Способы настоящего раскрытия, например олигонуклеотидные праймеры и зонды, можно использовать с родственными способами анализа (анализами полиморфизма длины амплифицированных фрагментов (AFLP), анализами полиморфизма длины рестрикционых фрагментов (RFLP), анализами случайно амплифицируемой полиморфной ДНК (RAPD)), которые идентифицируют генетически связанные агрономически полезные признаки посредством детектирования SNP или простых повторяющихся последовательностей (SSR) с применением публично доступных протоколов, которые известны в данной области техники. SNP, раскрытые в настоящем описании, можно отслеживать в потомстве от скрещивания с растением канолы рассматриваемого раскрытия (или потомства от него и любого другого культивара или сорта канолы) с применением MAB-способов. Молекулы ДНК можно использовать в качестве маркеров для данного признака, и MAB-способы, которые хорошо известны в данной области техники, можно использовать для отслеживания SNP в растениях канолы, где по меньшей мере одно растение канолы рассматриваемого раскрытия или его потомство представляет собой родителя или предка. Способы настоящего раскрытия можно использовать для идентификации любого сорта канолы, имеющего рассматриваемые SNP, раскрытые в настоящем описании.
[0049] Способы рассматриваемого раскрытия включают способ создания растения канолы, содержащего комбинацию SNP, указанных в настоящем описании, причем упомянутый способ содержит скрещивание с растением рассматриваемого раскрытия. Более конкретно, упомянутые способы могут содержать скрещивание двух растений рассматриваемого раскрытия или одного растения рассматриваемого раскрытия и любого другого растения. Иллюстративные способы могут дополнительно содержать отбор потомства от упомянутого скрещивания посредством анализа упомянутого потомства на SNP рассматриваемого раскрытия, детектируемого в соответствии с рассматриваемым раскрытием. Например, рассматриваемое раскрытие можно использовать для отслеживания зиготности растений канолы на протяжении циклов скрещивания с растениями, имеющими другие желаемые признаки, такие как агрономические признаки, такие как исследованные в настоящем описании в различных примерах. Растения, содержащие рассматриваемые SNP и желаемые признаки, можно также, например, детектировать, идентифицировать, отбирать и быстро использовать в последующих циклах скрещивания. Рассматриваемые SNP/признаки можно также комбинировать во время скрещивания и отслеживать в соответствии с рассматриваемым раскрытием с другими признаками, например с возможным признаком(ами) устойчивости к насекомым-вредителям и/или с признаками устойчивости к гербицидам. Одним вариантом осуществления последнего является растение, содержащее один или несколько рассматриваемых SNP в сочетании с геном, кодирующим устойчивость к гербициду, такому как глифосат.
[0050] В некоторых вариантах осуществления настоящее раскрытие включает последовательности ДНК, которые содержат смежный фрагмент, применимые в качестве последовательностей праймера для создания ампликонового продукта, диагностического для одного или нескольких растений канолы fad-3c.
[0051] Родственные варианты осуществления имеют отношение к последовательностям ДНК, которые содержат по меньшей мере 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25 или более смежных нуклеотидов части последовательностей ДНК, указанных в настоящем описании, или их комплементов. Такие последовательности можно использовать в качестве праймеров ДНК в способах амплификации ДНК. Ампликоны, получаемые с применением данных праймеров, могут быть диагностическими для любой комбинации и зиготности сортов канолы fad-3c, упоминаемых в настоящем описании. Следовательно, данное раскрытие также включает ампликоны, производимые такими праймерами ДНК и гомологичными праймерами.
[0052] В следующих вариантах осуществления рассматриваемое раскрытие включает способы создания SNP fad-3c рассматриваемого раскрытия, причем упомянутый способ содержит стадии: (a) полового скрещивания первой родительской линии канолы, содержащей один из SNP, раскрытых в настоящем описании и несущих один из признаков олеиновой и/или линоленовой кислоты, раскрытых в настоящем описании, и второй родительской линии канолы (в которой отсутствуют данные SNP), посредством чего создают множество растений-потомков; и (b) отбор растения-потомка с применением молекулярных маркеров. Такие способы могут необязательно содержать дополнительную стадию возвратного скрещивания растения-потомка со второй родительской линией канолы для получения растения канолы чистого разведения или гомозиготного, которое содержит упомянутые признаки fad-3c.
[0053] В соответствии с другим аспектом данного раскрытия предлагаются способы определения зиготности потомства от скрещивания с упомянутыми растениями канолы fad-3c. Упомянутые способы могут включать в себя приведение образца, содержащего ДНК канолы, в контакт с набором праймеров рассматриваемого раскрытия. Упомянутые праймеры при использовании в реакции амплификации нуклеиновой кислоты с геномной ДНК от по меньшей мере одного из упомянутых растений канолы fad-3c производят первый ампликон, который является диагностическим для по меньшей мере одного из упомянутых SNP канолы или генов дикого типа. Такие способы дополнительно включают в себя осуществление реакции амплификации нуклеиновой кислоты, посредством чего производится первый ампликон, и детектирование первого ампликона с помощью зондов, специфичных к SNP fad-3c, раскрытым в настоящем описании, и генам дикого типа. Данные способы дополнительно включают в себя осуществление применений аллельного дискриминантного плавления ампликонов, к которым отожжены раскрытые зонды, и сравнение относительной флуоресценции зондов, использованных в применении аллельного дискриминантного плавления. Относительная флуоресценция зондов указывает, содержит ли образец представляющий интерес SNP, и если так, является ли образец гетерозиготным или гомозиготным по данному SNP.
[0054] Могут быть разработаны наборы для детектирования ДНК с применением композиций, раскрытых в настоящем описании, в соединении со способами, хорошо известными в области техники детектирования ДНК. Данные наборы полезны для идентификации рассматриваемых SNP канолы в образце и могут применяться в способах селекции растений канолы, содержащих данную ДНК. Данные наборы содержат последовательности ДНК, гомологичные или комплементарные ампликонам, например раскрытым в настоящем описании. Данные последовательности ДНК можно использовать в реакциях амплификации ДНК или в качестве зондов в способах гибридизации ДНК. Данные наборы могут также содержать реагенты и материалы, необходимые для осуществления способа детектирования.
[0055] "Зонд" представляет собой выделенную молекулу нуклеиновой кислоты, к которой прикреплена обычная детектируемая метка или репортерная молекула (такая как радиоактивный изотоп, лиганд, хемилюминесцентное средство или фермент). Такой зонд комплементарен цепочке мишеневой нуклеиновой кислоты, в случае настоящего раскрытия цепочке геномной ДНК от одного из упомянутых растений канолы, содержащих представляющие интерес гены fad-3c, или от растения канолы, или от образца, который содержит ДНК от трансформанта. Зонды в соответствии с настоящим раскрытием включают не только дезоксирибонуклеиновы или рибонуклеиновые кислоты, но также полиамиды и другие материалы для зондов, которые специфически связываются с мишеневой последовательностью ДНК и могут использоваться для детектирования наличия мишеневой последовательности ДНК.
[0056] Были сконструированы специфичные зонды, содержащие флуоресцентный репортер (флуорофор) и гаситель, который гибридизуется с мишеневой ДНК между праймерами ПЦР. Молекулу флуорофора присоединяют к олигонуклеотидному зонду во время синтеза олигонуклеотидного зонда, посредством чего помечают олигонуклеотидный зонд. К олигонуклеотидному зонду можно присоединять другие молекулы, такие как молекула гасителя. Присоединение данных молекул к олигонуклеотидному зонду не нарушает функцию олигонуклеотидного зонда при гибридизации с одноцепочечной ДНК и получении новой цепочки ДНК в процессе амплификации.
[0057] Были разработаны многочисленные флуорофоры, которые возбуждаются при определенных длинах волн и известны в данной области техники. Возбуждение флуорофора приводит к испусканию флуорофором флуоресцентного сигнала, который может быть погашен гасителем, расположенным в непосредственной близости от флуорофора. Когда гаситель отсоединяется от флуорофора, флуоресцентный сигнал больше не гасится, и сумму флуоресцентного сигнала, которая прямо коррелирует с количеством мишеневой ДНК, можно детектировать в реальном времени с помощью автоматического флуориметра. Флуорофоры можно использовать в комбинации, причем спектры возбуждения и испускания значительно отличаются, делая возможным множественное детектирование двух или нескольких флуорофоров. Некоторые предпочтительные варианты осуществления флуорофоров включают: флуоресцентный краситель HEX, флуоресцентный краситель TET, флуоресцентный краситель Cy3, флуоресцентный краситель Cy3.5, флуоресцентный краситель Cy5, флуоресцентный краситель Cy5.5, флуоресцентный краситель Cy7, или флуоресцентный краситель ROX. Один предпочтительный вариант осуществления флуорофора для использования со способом, включающим систему детектирования для гомогенного анализа для процесса ПЦР с применением FRET рассматриваемого изобретения, включает флуоресцентный краситель FAM флуоресцентного красителя JOE.
[0058] Были разработаны гасители для гашения флуорофоров на определенной длине волны, и они известны в данной области техники. Когда гаситель расположен в непосредственной близости к флуорофору, флуорофор передает энергию гасителю. Гаситель передает данную энергию и возвращается в исходное основное состояние посредством спада излучения или безызлучательно. При безызлучательном или темновом спаде энергия, передаваемая от флуорофора, выделяется в виде молекулярных колебаний. При выборе гасителя принимают во внимание такие качества, как низкая фоновая флуоресценция, высокая чувствительность и максимальное спектральное перекрытие, для получения гасителя, который может сделать возможным широкое использование флуорофоров. Некоторые предпочтительные варианты осуществления гасителей включают: гасители Dabcyl, гасители Tamra, гаситель Qxl, гаситель Iowa black FQ, гаситель Iowa black RQ или гаситель IR Dye QC-1. Особенно предпочтительный вариант осуществления гасителя включает гаситель Blackhole, используемый в качестве метки на олигонуклеотидном праймере, который сконструирован как антисмысловой FAM-меченному олигонуклеотиду.
[0059] "Праймеры" представляют собой выделенные/синтезированные нуклеиновые кислоты, которые подвергают отжигу на комплементарной мишеневой цепочке ДНК посредством гибридизации нуклеиновой кислоты, посредством чего образуется гибрид между праймером и мишеневой цепочкой ДНК, и затем удлиняют вдоль мишеневой цепочки ДНК посредством полимеразы, например ДНК-полимеразы. Пары праймеров настоящего раскрытия относятся к их применению для амплификации мишеневой последовательности нуклеиновой кислоты, например посредством полимеразной цепной реакции (ПЦР) или других обычных способов амплификации нуклеиновых кислот.
[0060] Зонды и праймеры обычно имеют длину 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 100, 101, 102, 103, 104, 105, 106, 107, 108, 109, 110, 111, 112, 113, 114, 115, 116, 117, 118, 119, 120, 121, 122, 123, 124, 125, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 150, 151, 152, 153, 154, 155, 156, 157, 158, 159, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182, 183, 184, 185, 186, 187, 188, 189, 190, 191, 192, 193, 194, 195, 196, 197, 198, 199, 200, 201, 202, 203, 204, 205, 206, 207, 208, 209, 210, 211, 212, 213, 214, 215, 216, 217, 218, 219, 220, 221, 222, 223, 224, 225, 226, 227, 228, 229, 230, 231, 232, 233, 234, 235, 236, 237, 238, 239, 240, 241, 242, 243, 244, 245, 246, 247, 248, 249, 250, 251, 252, 253, 254, 255, 256, 257, 258, 259, 260, 261, 262, 263, 264, 265, 266, 267, 268, 269, 270, 271, 272, 273, 274, 275, 276, 277, 278, 279, 280, 281, 282, 283, 284, 285, 286, 287, 288, 289, 290, 291, 292, 293, 294, 295, 296, 297, 298, 299, 300, 301, 302, 303, 304, 305, 306, 307, 308, 309, 310, 311, 312, 313, 314, 315, 316, 317, 318, 319, 320, 321, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 336, 337, 338, 339, 340, 341, 342, 343, 344, 345, 346, 347, 348, 349, 350, 351, 352, 353, 354, 355, 356, 357, 358, 359, 360, 361, 362, 363, 364, 365, 366, 367, 368, 369, 370, 371, 372, 373, 374, 375, 376, 377, 378, 379, 380, 381, 382, 383, 384, 385, 386, 387, 388, 389, 390, 391, 392, 393, 394, 395, 396, 397, 398, 399, 400, 401, 402, 403, 404, 405, 406, 407, 408, 409, 410, 411, 412, 413, 414, 415, 416, 417, 418, 419, 420, 421, 422, 423, 424, 425, 426, 427, 428, 429, 430, 431, 432, 433, 434, 435, 436, 437, 438, 439, 440, 441, 442, 443, 444, 445, 446, 447, 448, 449, 450, 451, 452, 453, 454, 455, 456, 457, 458, 459, 460, 461, 462, 463, 464, 465, 466, 467, 468, 469, 470, 471, 472, 473, 474, 475, 476, 477, 478, 479, 480, 481, 482, 483, 484, 485, 486, 487, 488, 489, 490, 491, 492, 493, 494, 495, 496, 497, 498, 499 или 500 полинуклеотидов или более. Такие зонды и праймеры специфически гибридизуются с мишеневой последовательностью в условиях гибридизации высокой строгости. Предпочтительно, зонды и праймеры в соответствии с настоящим раскрытием имеют полное сходство последовательности с мишеневой последовательностью, хотя можно с помощью обычных способов конструировать зонды, отличающиеся от мишеневой последовательности, которые сохраняют способность гибридизоваться с мишеневой последовательностью.
[0061] Способы получения и применения зондов и праймеров описаны, например, в Molecular Cloning: A Laboratory Manual, 2nd ed., vol. 1-3, ed. Sambrook et al., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., 1989. Пары ПЦР-праймеров можно получать из известных последовательностей, например, с применением компьютерных программ, предназначенных для этой цели.
[0062] Праймеры и зонды, основанные на последовательностях ДНК, расположенных против хода транскрипции и по ходу транскрипции от SNP, раскрытых в настоящем описании, можно использовать для подтверждения (и, если необходимо, исправления) раскрытых последовательностей посредством обычных способов, например посредством повторного клонирования и секвенирования таких последовательностей.
[0063] Нуклеиново-кислотные зонды и праймеры настоящего раскрытия гибридизуются в строгих условиях с мишеневой последовательностью ДНК. В целом, для идентификации наличия ДНК из образца fad-3c можно использовать любой обычный способ гибридизации или амплификации нуклеиновой кислоты. Молекулы нуклеиновой кислоты или ее фрагменты способны специфически гибридизоваться с другими молекулами нуклеиновой кислоты в определенных условиях. Как применяется в настоящем описании, говорят, что две молекулы нуклеиновой кислоты способны специфически гибридизоваться друг с другом, если две молекулы способны к образованию антипараллельной двухцепочечной нуклеиново-кислотной структуры. Говорят, что молекула нуклеиновой кислоты является "комплементом" другой молекулы нуклеиновой кислоты, если они демонстрируют полную комплементарность. Как применяется в настоящем описании, говорят, что молекулы демонстрируют "полную комплементарность", когда каждый нуклеотид одной из молекул комплементарен нуклеотиду из другой. Говорят, что две молекулы являются "минимально комплементарными", если они могут гибридизоваться друг с другом со стабильностью, достаточной для того, чтобы позволять им оставаться отожженными друг на друге по меньшей мере в обычных условиях "низкой строгости". Аналогично, говорят, что молекулы являются "комплементарными", если они могут гибридизоваться друг с другом со стабильностью, достаточной для того, чтобы позволять им оставаться отожженными друг на друге в обычных условиях "высокой строгости". Обычные условия строгости описаны в Sambrook et al., 1989. Следовательно, отклонения от полной комплементарности допустимы до тех пор, пока такие отклонения не устраняют полностью способность молекул образовывать двухцепочечную структуру. Для того чтобы молекула нуклеиновой кислоты служила в качестве праймера или зонда, она должна только иметь достаточно комплементарную последовательность для того, чтобы быть способной образовывать стабильную двухцепочечную структуру при конкретных используемых концентрациях растворителя и соли.
[0064] Как применяется в настоящем описании, по существу гомологичная последовательность представляет собой последовательность нуклеиновой кислоты, которая будет специфически гибридизоваться с комплементом последовательности нуклеиновой кислоты, с которой ее сравнивают, в условиях высокой строгости. Термин "строгие условия" функционально определяют в отношении гибридизации нуклеиново-кислотного зонда с мишеневой нуклеиновой кислотой (т.е. с конкретной представляющей интерес последовательностью нуклеиновой кислоты) посредством процедуры специфической гибридизации, рассмотренной в Sambrook et al., 1989, на 9.52-9.55. См. также Sambrook et al., 1989 на 9.47-9.52 и 9.56-9.58. Соответственно, нуклеотидные последовательности данного раскрытия можно применять ввиду их способности селективно образовывать двойные молекулы с комплементарными отрезками фрагментов ДНК.
[0065] В зависимости от предполагаемого применения можно использовать различные условия гибридизации для достижения различной степени селективности зонда в отношении мишеневой последовательности. Для применений, требующих высокой селективности, для образования гибридов будут, как правило, использоваться относительно строгие условия, например, будут выбраны условия относительно низкой концентрации соли и/или высокой температуры, такие как получаемые при от приблизительно 0,50 мМ до приблизительно 02,00 мМ MgCl2 при температурах, составляющих от приблизительно 50°C до приблизительно 75°C. Можно изменять как температуру, так и соль, или либо температуру, либо концентрацию соли можно поддерживать постоянными, в то же время изменяя другую переменную. Такие условия селективности допускают небольшое число, если вообще допускают, несовпадений между зондом и матричной или мишеневой цепочкой. Детектирование последовательностей ДНК посредством гибридизации хорошо известно специалистам в данной области техники, и идеи патентов США №№ 4965188 и 5176995 являются примерами способов гибридизационных анализов.
[0066] В одном иллюстративном варианте осуществления нуклеиновая кислота настоящего раскрытия специфически гибридизуется с одним или несколькими из праймеров (или ампликонов или других последовательностей), приведенных в качестве примера или предлагаемых в настоящем описании, включая комплементы и их фрагменты, в условиях высокой строгости. В одном аспекте настоящего раскрытия маркерная молекула нуклеиновой кислоты настоящего раскрытия имеет последовательность нуклеиновой кислоты, приведенную в настоящем описании в одном из примеров последовательностей, или их комплементов и/или фрагментов.
[0067] В другом аспекте настоящего раскрытия маркерная молекула нуклеиновой кислоты настоящего раскрытия имеет от 80% до 100% или от 90% до 100% сходства последовательности с такими последовательностями нуклеиновой кислоты. В следующем аспекте настоящего раскрытия маркерная молекула нуклеиновой кислоты настоящего раскрытия имеет от 95%, 96%, 97%, 98% и/или 99% до 100% сходства последовательности с такой последовательностью. Такие последовательности можно применять в качестве маркеров в способах селекции растений для идентификации потомства генетических скрещиваний. Гибридизацию зонда с мишеневой молекулой ДНК можно детектировать посредством любого числа способов, известных специалистам в данной области техники, они могут включать, но без ограничения, флуоресцентные метки, радиоактивные метки, метки, основанные на антителах, и хемилюминесцентные метки.
[0068] В отношении амплификации мишеневой последовательности нуклеиновой кислоты (например, посредством ПЦР) с применением конкретной пары праймеров для амплификации, "строгие условия" представляют собой условия, которые позволяют паре праймеров гибридизоваться в первую очередь и с высоким предпочтением с их мишеневыми последовательностями нуклеиновой кислоты, что позволяет паре праймеров связываться и, предпочтительно, производить уникальный ампликон.
[0069] Термин "специфичный к (мишеневой последовательности)" указывает на то, что зонд или праймер гибридизуется в строгих условиях гибридизации в первую очередь и с высоким предпочтением с последовательностью нуклеиновой кислоты в образце, содержащем мишеневую последовательность.
[0070] Как применяется в настоящем описании, "амплифицированная ДНК" или "ампликон" относится к продукту нуклеиново-кислотной амплификации мишеневой последовательности нуклеиновой кислоты, которая является частью нуклеиново-кислотной матрицы. Например, для того чтобы определить, содержит ли растение канолы, получаемое половым скрещиванием, представляющий интерес SNP, как раскрыто в настоящем описании. ДНК, экстрагированную из образца ткани растения канолы, можно подвергать способу амплификации нуклеиновой кислоты с применением пары праймеров, которая включает праймер, происходящий от последовательности, расположенной в геноме растения канолы против хода транскрипции или по ходу транскрипции от участка SNP, и второй праймер, происходящий от другого конца последовательности, расположенной в геноме растения канолы против хода транскрипции или по ходу транскрипции от участка SNP, благодаря чему создается ампликон, который является диагностическим для наличия SNP. Ампликон имеет длину и последовательность, которые также являются диагностическими для гена дикого типа или мутировавшего. Длина ампликона может лежать в диапазоне от общей длины пар праймеров плюс одна пара нуклеотидных оснований и/или общей длины пар праймеров плюс приблизительно 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 100, 101, 102, 103, 104, 105, 106, 107, 108, 109, 110, 111, 112, 113, 114, 115, 116, 117, 118, 119, 120, 121, 122, 123, 124, 125, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139, 140, 141, 142, 143, 144, 145, 146, 147, 148, 149, 150, 151, 152, 153, 154, 155, 156, 157, 158, 159, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179, 180, 181, 182, 183, 184, 185, 186, 187, 188, 189, 190, 191, 192, 193, 194, 195, 196, 197, 198, 199, 200, 201, 202, 203, 204, 205, 206, 207, 208, 209, 210, 211, 212, 213, 214, 215, 216, 217, 218, 219, 220, 221, 222, 223, 224, 225, 226, 227, 228, 229, 230, 231, 232, 233, 234, 235, 236, 237, 238, 239, 240, 241, 242, 243, 244, 245, 246, 247, 248, 249, 250, 251, 252, 253, 254, 255, 256, 257, 258, 259, 260, 261, 262, 263, 264, 265, 266, 267, 268, 269, 270, 271, 272, 273, 274, 275, 276, 277, 278, 279, 280, 281, 282, 283, 284, 285, 286, 287, 288, 289, 290, 291, 292, 293, 294, 295, 296, 297, 298, 299, 300, 301, 302, 303, 304, 305, 306, 307, 308, 309, 310, 311, 312, 313, 314, 315, 316, 317, 318, 319, 320, 321, 322, 323, 324, 325, 326, 327, 328, 329, 330, 331, 332, 333, 334, 335, 336, 337, 338, 339, 340, 341, 342, 343, 344, 345, 346, 347, 348, 349, 350, 351, 352, 353, 354, 355, 356, 357, 358, 359, 360, 361, 362, 363, 364, 365, 366, 367, 368, 369, 370, 371, 372, 373, 374, 375, 376, 377, 378, 379, 380, 381, 382, 383, 384, 385, 386, 387, 388, 389, 390, 391, 392, 393, 394, 395, 396, 397, 398, 399, 400, 401, 402, 403, 404, 405, 406, 407, 408, 409, 410, 411, 412, 413, 414, 415, 416, 417, 416, 419, 420, 421, 422, 423, 424, 425, 423, 427, 428, 429, 430, 431, 432, 433, 434, 435, 433, 437, 438, 439, 440, 441, 442, 443, 444, 445, 443, 447, 448, 449, 450, 451, 452, 453, 454, 455, 456, 457, 458, 459, 460, 461, 462, 463, 434, 435, 433, 437, 438, 439, 470, 471, 472, 473, 474, 475, 473, 477, 478, 479, 480, 481, 482, 483, 484, 485, 486, 487, 488, 489, 490, 491, 492, 493, 494, 495, 496, 497, 498, 499, или 500, 750, 1000, 1250, 1500, 1750, 2000 или более пар нуклеотидных оснований (плюс или минус любое из приращений, перечисленных выше). Член пары праймеров, происходящих от геномной последовательности растения, может быть расположен на расстоянии от последовательности SNP. Данное расстояние может лежать в диапазоне от одной пары нуклеотидных оснований вплоть до приблизительно двадцати тысяч пар нуклеотидных оснований. Применение термина "ампликон" прямо исключает димеры праймеров, которые могут образовываться в реакции тепловой амплификации ДНК.
[0071] Амплификацию нуклеиновой кислоты можно осуществлять посредством любого из различных способов амплификации нуклеиновой кислоты, известных в данной области техники, включая ПЦР. Множество способов амплификации известно в данной области техники и описано, в том числе, в патенте США № 4683195 и патенте США № 4683202. Были разработаны способы амплификации с помощью ПЦР для амплификации вплоть до 22 т.п.о. геномной ДНК. Данные способы, а также другие известные в данной области техники способы амплификации ДНК, можно применять при осуществлении на практике настоящего раскрытия. Последовательность SNP fad-3c можно верифицировать посредством амплификации таких последовательностей с применением праймеров, происходящих от последовательностей, предлагаемых в настоящем описании, с последующим стандартным секвенированием ДНК ПЦР-ампликона или клонированной ДНК.
[0072] Ампликон, получаемый в данных способах, можно детектировать посредством множества методов. Электрофорез на агарозном геле и окрашивание с помощью бромида этидия является обычным хорошо известным способом детектирования ампликонов ДНК. Другим таким способом является генетический бит-анализ, в котором конструируют олигонуклеотид ДНК, который перекрывает как прилегающую фланкирующую геномную последовательность ДНК, так и вставленную последовательность ДНК. Олигонуклеотид иммобилизуют в лунках микролуночного планшета. После ПЦР представляющего интерес участка (с применением одного праймера во вставленной последовательности и одного в прилегающей фланкирующей геномной последовательности) одноцепочечный продукт ПЦР можно гибризовать с иммобилизованным олигонуклеотидом, и он может служить в качестве матрицы для реакции удлинения на одно основание с применением ДНК-полимеразы и меченых ddNTP, специфичных к следующему ожидаемому основанию. Считывание результата может быть основано на флуоресценции или ELISA. Сигнал указывает на наличие вставленной/фланкирующей последовательности за счет успешной амплификации, гибридизации и удлинения на одно основание.
[0073] TaqMan® ПЦР представляет собой способ детектирования и количественного определения наличия последовательности ДНК. Кратко говоря, конструируют олигонуклеотидный FRET-зонд, который перекрывает представляющий интерес SNP. FRET-зонд и праймеры ПЦР (по меньшей мере один, расположенный против хода транскрипции, и по меньшей мере один, расположенный по ходу транскрипции от представляющего интерес SNP) проходят цикл в присутствии термостабильной полимеразы и dNTP.
[0074] После амплификации может быть осуществлен аллельный дискриминантный анализ (с применением гидролиза зонда TaqMan®, описанного выше) для определения наличия представляющего интерес SNP и зиготности образца. Во время аллельного дискриминантного анализа два различных гибридизационных зонда (один зонд включает нуклеотид, комплементарный последовательности SNP, а другой зонд несет нуклеотид, комплементарный последовательности дикого типа) гибридизуются с ампликоном и расщепляются, посредством чего из зонда высвобождаются гасящие фрагменты благодаря 5'-экзонуклеазной активности taq-полимеразы, и это приводит к флуоресценции. Сравнение относительной флуоресценции зонда, специфичного к гену дикого типа, с зондом, специфичному к SNP, обеспечивает индикацию наличия и зиготности представляющего интерес SNP.
[0075] Все патенты, патентные заявки, предварительные заявки и публикации, на которые ссылается настоящее описание, или которые перечислены в настоящем описании, включены посредством ссылки во всей полноте до той степени, в которой они не являются несовместимыми с явно выраженными идеями настоящего описания изобретения.
[0076] Следующие примеры включены для иллюстрации способов осуществления данного раскрытия на практике и для демонстрации некоторых предпочтительных вариантов осуществления данного раскрытия. Данные примеры не следует рассматривать как ограничивающие. Специалистам в данной области техники должно быть понятно, что методы, раскрытые в следующих примерах, представляют конкретные подходы, применяемые для иллюстрации предпочтительных вариантов их практического осуществления. Однако, специалистам в данной области техники в свете настоящего раскрытия должно быть понятно, что можно осуществлять множество изменений в данных конкретных вариантах осуществления, получая при этом такие же или схожие результаты, без отклонения от сущности и объема данного раскрытия. Если не указано иное, все процентные доли приведены по весу, и все соотношения смесей растворителей приведены по объему, если не отмечено иное.
[0077] Если не указано иное, используются следующие сокращения.
п. о. - пара оснований
°C - градусы Цельсия
ДНК - дезоксирибонуклеиновая кислота
FRET - резонансный перенос энергии флуоресценции
DIG - дигоксигенин
ЭДТА - этилендиаминтетрауксусная кислота
т.п.о. - тысяча пар нуклеотидов
мкг - микрограмм
мкл - микролитр
мл - миллилитр
M - молярная масса
OLP - перекрывающий зонд
ПЦР - полимеразная цепная реакция
PTU - транскрипционная единица растения
ДСН - додецилсульфат натрия
SNP - однонуклеотидный полиморфизм
SOP - стандартный порядок действий
SSC - буферный раствор, содержащий смесь хлорида натрия и цитрата натрия, pH 7,0
TBE - буферный раствор, содержащий смесь основания Tris, борной кислоты и ЭДТА, pH 8,3
V - вольты
Примеры
Пример 1: FAD-3c TaqMan®-анализ по конечной точке
[0078] Был разработан TaqMan-анализ по конечной точке для детектирования мутации однонуклеотидного полиморфизма fad-3c и для определения состояния зиготности растений канолы, содержащих мутацию гена fad-3c в разводимых популяциях. Сконструировали два праймера для связывания с высоко консервативными последовательностями ДНК, расположенными на экзоне 6 и 7 гена fad-3c. Данные праймеры амплифицировали фрагмент ДНК размером 154 п. о., который перекрывал однонуклеотидный полиморфизм fad-s3c в мутировавших и не мутировавших растениях канолы. Мутация fad-3c в каноле описана Hu et al., (2006) и расположена у соединения экзона 6 и интрона 6 в сайте сплайсинга данного гена (фиг. 1). Сконструировали два зонда TaqMan®, связывающихся с малой бороздкой и несущих нефлуоресцентный гаситель (MGBNFQ), с FAM и VIC в качестве репортерных красителей для детектирования наличия гена fad-3c дикого типа и мутировавшего гена fad-3c (который содержит однонуклеотидный полиморфизм, SNP) соответственно. Данные два зонда были сконструированы с особым вниманием к тому, чтобы избежать близлежащего полиморфизма, расположенного в интроне 6 и в непосредственной близости от однонуклеотидного полиморфизма fad-3c (фиг. 1). Избегание второго полиморфизма приводит к возрастанию специфичности зондов в детектировании fad-3c-мутантных растений. Способ TaqMan®-детектирования растений канолы, содержащих SNP fad-3c, тестировали на сорте канолы "NEX 828" (содержащем SNP fad-3c), контрольном сорте канолы "Quantum" (не содержит SNP fad-3c) и образце ДНК, выделенном из растений, о которых известно, что они являются гетерозиготными по SNP fad-3c. TaqMan®-анализ по конечной точке использовали для определения наличия SNP fad-3c, а также для определения зиготности тестируемых растений при высокопроизводительном способе применения, например в формате 96 и 384-луночного планшета.
Пример 1.1: Выделение гДНК
[0079] В данном исследовании тестировали образцы геномной ДНК (гДНК) 156 различных растений канолы, содержащих мутацию fad-3c, и контрольных растений канолы. Геномную ДНК экстрагировали с применением модифицированного набора Qiagen MagAttract plant DNA (Qiagen, Valencia, CA). Для экстракции гДНК использовали свежие листовые диски канолы по 4 на образец. гДНК количественно характеризовали с помощью метода Pico Green в соответствии с инструкциями производителя (Molecular Probes, Eugene, OR). Образцы разбавляли свободной от ДНКазы водой, получая концентрацию 5 нг/мкл для данного исследования.
Пример 1.2: TaqMan®-анализ и результаты
[0080] Были разработаны специфичные праймеры и зонды TaqMan® для использования в TaqMan®-анализе по конечной точке. Данные праймеры и зонды конструировали так, чтобы амплифицировать и детектировать участок гена fad-3c, содержащий представляющий интерес SNP. Данные реагенты можно использовать в условиях, перечисленных ниже, для детектирования мутировавшего гена fad-3c в растениях канолы. Таблица 2 перечисляет последовательности праймеров и зондов, которые были разработаны специально для детектирования SNP fad-3c в растениях канолы.
Праймеры ПЦР и зонды Taqman
GTTGTTAATCCTCCACGT
[0081] Реакционные смеси для ПЦР для амплификации имеют следующий вид: 1X TaqMan® GTExpress Master Mix, 0,9 мкМ прямого праймера (SEQ ID NO:2), 0,9 мкМ обратного праймера (SEQ ID NO:3), 0,2 мкМ зонда для мутантного FAD-3C (SEQ ID NO:4), 0,2 мкМ зонда для дикого типа (SEQ ID NO:5), 15 нг гДНК при общем объеме реакции, равном 6 мкл. Реакционную смесь подвергали амплификации, используя следующие условия теплового цикла: в начале две стадии при 50°C в течение 2 минут и 95°C в течение 30 секунд; затем 40 циклов по 3 секунды при 95°C и 30 секунд при 62°C. Затем реакцию держали при 10°C до тех пор, пока не вынимали из термоциклера. Тепловые циклы ПЦР можно осуществлять с применением или системы для ПЦР в реальном времени ABI-Applied Biosystems 7900 HT, или термоциклера Applied Biosystems Verity (Life Technologies, Carlsbad, CA). Планшеты для образцов содержали контрольную ДНК из растений канолы, которые являются гомозиготными по мутантному fad-3c ("NEX 828"), гетерозиготными по мутантному fad-3c или гомозиготными по fad-3c дикого типа ("Quantum"). Дополнительно включали контроль без матрицы, который не содержал ДНК. После амплификации считывали флуоресцентные сигналы в конечной точке (VIC и FAM) с применением системы для ПЦР в реальном времени Applied Biosystems 7900 HT в соответствии с аллельным дискриминантным способом считывания планшетов, как описано производителем. Затем данные анализировали с применением программного обеспечения SDS 2.4 (Life Technologies, Carlsbad, CA) для определения относительной флуоресценции каждого образца (фиг. 2).
[0082] Был протестирован способ детектирования мутации fac-3c в каноле с помощью TaqMan® по образцам, о которых известно, что они являются гомозиготными, гемизиготными и дикого типа. Анализ флуоресценции, создаваемой каждым зондом (в реакции с образцом), с флуоресценцией, создаваемой зондами контрольных образцов, способствовал определению зиготности каждого образца. Данный анализ продемонстрировал высокую специфичность детектирования однонуклеотидного полиморфизма fad-3c с мутацией и дикого типа в каноле и не создавал или не увеличивал никаких детектируемых ложно-положительных результатов от контрольных образцов. Специфические к событию праймеры и зонды можно использовать для детектирования мутантного гена fad-3c и гена fad-3c дикого типа в каноле, и данные условия и реагенты применимы для анализов зиготности.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ОПРЕДЕЛЕНИЯ ЗИГОТНОСТИ ГЕНА FAD-2 КАНОЛЫ С ИСПОЛЬЗОВАНИЕМ ПЦР С ДЕТЕКЦИЕЙ ПО КОНЕЧНОЙ ТОЧКЕ | 2012 |
|
RU2639508C2 |
| ДЕТЕКТИРОВАНИЕ AAD-12-СОБЫТИЯ 416 У СОИ | 2010 |
|
RU2573898C2 |
| ДЕТЕКЦИЯ AAD-1 ОБЪЕКТА DAS-40278-9 | 2010 |
|
RU2577143C2 |
| МАТЕРИАЛЫ И МЕТОДЫ ДЛЯ ОБНАРУЖЕНИЯ ГЕНА АРИЛОКСИАЛКАНОАТДИОКСИГЕНАЗЫ (AAD-12) В РАСТЕНИЯХ | 2012 |
|
RU2644249C2 |
| ПАКЕТИРОВАННОЕ СОБЫТИЕ 8264.44.06.1 С УСТОЙЧИВОСТЬЮ К ГЕРБИЦИДАМ, СВЯЗАННЫЕ ТРАНСГЕННЫЕ ЛИНИИ СОИ И ИХ ДЕТЕКТИРОВАНИЕ | 2011 |
|
RU2608650C2 |
| ПРИМЕНЕНИЕ СПЕЦИФИЧЕСКИХ МАРКЕРОВ ГЕНА КОРИЧНЕВОЙ СРЕДНЕЙ ЖИЛКИ 3 (BROWN MIDRIB-3) У КУКУРУЗЫ ДЛЯ ИНТРОГРЕССИИ ПРИЗНАКОВ | 2011 |
|
RU2593958C2 |
| ТРАНСГЕННЫЙ ОБЪЕКТ СОИ MON 87708 И СПОСОБЫ ЕГО ПРИМЕНЕНИЯ | 2010 |
|
RU2624025C2 |
| ЛИНИИ ТРАНСГЕННОЙ СОИ, ГЕНЕТИЧЕСКОЕ СОБЫТИЕ 8264.42.32.1, УСТОЙЧИВОЕ К ГЕРБИЦИДАМ С ПАКЕТИРОВАННЫМИ ГЕНАМИ НА ЕГО ОСНОВЕ, И ИХ ДЕТЕКТИРОВАНИЕ | 2012 |
|
RU2636021C2 |
| МОЛЕКУЛЯРНЫЕ МАРКЕРЫ ГЕНА RLM2 РЕЗИСТЕНТНОСТИ К ЧЕРНОЙ НОЖКЕ BRASSICA NAPUS И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2014 |
|
RU2717017C2 |
| ГЕН-СПЕЦИФИЧЕСКИЙ АНАЛИЗ НА FLUORY2 В МАИСЕ ДЛЯ ИНТРОГРЕССИИ МУЧНИСТОГО ПРИЗНАКА (FL2) | 2013 |
|
RU2661110C2 |



Изобретение относится к области биохимии, в частности к способу определения зиготности гена fad-3c в растении канолы. Также раскрыт набор для осуществления указанного способа. Изобретение позволяет эффективно определять зиготность гена fad-3c в растении канолы. 2 н. и 12 з.п. ф-лы, 2 ил., 2 табл., 1 пр.
1. Способ определения зиготности гена fad-3с в растении канолы, причем упомянутый способ включает в себя:
получение образца геномной ДНК от растения канолы;
гибридизацию образца геномной ДНК с первым праймером и вторым праймером, где упомянутый первый праймер и упомянутый второй праймер содержат SEQ ID NO: 2 и SEQ ID NO: 3;
подвергание упомянутого образца условиям полимеразной цепной реакции (ПЦР), причем производится ампликон;
предоставление возможности каждому из первого зонда и второго зонда гибридизоваться с упомянутым ампликоном в течение периода времени и при температуре 50-70 градусов Цельсия, где упомянутый первый зонд и упомянутый второй зонд содержат SEQ ID NO: 5 и SEQ ID NO: 4, где каждый из упомянутого первого зонда и упомянутого второго зонда мечен флуоресцентным красителем и гасителем;
повышение упомянутой температуры после периода времени;
измерение флуоресценции упомянутого первого зонда, упомянутого второго зонда или их комбинации; и
определение зиготности гена fad-3с в упомянутом растении канолы.
2. Способ по п. 1, в котором упомянутый ампликон состоит из 91-154 пар оснований.
3. Способ по п. 1, в котором образец геномной ДНК содержит мутированную fad-3с последовательность, имеющую однонуклеотидный полиморфизм, где упомянутый однонуклеотидный полиморфизм состоит из полиморфизма G->A.
4. Способ по п. 3, в котором образец геномной ДНК дополнительно содержит fad-3с последовательность дикого типа.
5. Способ по п. 1, в котором упомянутый способ применяют для верификации интрогрессии при селекции у кроссбредных растений канолы.
6. Способ по п. 1, в котором упомянутый первый зонд гибридизуется с областью мутированной fad-3с последовательности, имеющей однонуклеотидный полиморфизм (SNP), и упомянутый второй зонд гибридизуется с областью fad-3с последовательности дикого типа.
7. Способ по п. 6, в котором упомянутый первый зонд содержит FAM в качестве упомянутого флуоресцентного красителя на 5'-конце упомянутого первого зонда и MGB-гаситель на 3'-конце упомянутого первого зонда.
8. Способ по п. 6, в котором упомянутый второй зонд помечен VIC на 5'-конце упомянутого второго зонда и MGB-гасителем на 3'-конце упомянутого второго зонда.
9. Способ по п. 1, в котором упомянутый второй зонд содержит SEQ ID NO: 4.
10. Способ по п. 1, в котором результаты флуоресценции упомянутого способа анализируют непосредственно в планшет-ридере.
11. Способ по п. 1, в котором упомянутый образец ДНК получают от растения канолы в поле.
12. Способ по п. 1, в котором стадия повышения включает в себя повышение упомянутой температуры с по существу однородными приращениями температуры за период времени.
13. Способ по п. 1, в котором упомянутую флуоресценцию, создаваемую каждым из упомянутого первого зонда и упомянутого второго зонда во время стадии повышения, измеряют во время каждого приращения на стадии повышения упомянутой температуры.
14. Набор для осуществления способа по п. 1, причем упомянутый набор содержит первый праймер, состоящий из SEQ ID NO: 2, второй праймер, состоящий из SEQ ID NO: 3, первый зонд, состоящий из SEQ ID NO: 5, и второй зонд, состоящий из SEQ ID NO: 4.
| WO 20040072259 A2 от 26.08.2004 | |||
| WO 2001025453 A2 от 12.04.2001 | |||
| US 20100005548 A1 от 07.01.2010 | |||
| US 20110191904 A1 от 04.08.2011 | |||
| СОЕВОЕ МАСЛО С ВЫСОКОЙ ОКИСЛИТЕЛЬНОЙ СТАБИЛЬНОСТЬЮ | 1997 |
|
RU2162642C2 |

