Связанная информация
По настоящей заявке испрашивается приоритет US 61/075697, поданной 25 июня 2008 года; дополнительно испрашивается приоритет US 61/155041, поданной 24 февраля 2009 года и US61/155105 от 24 февраля 2009 года.
Предпосылки изобретения
Моноклональные антитела, их конъюгаты и производные являются очень важными с коммерческой точки зрения в качестве терапевтических и диагностических средств. Не являющиеся человеческими антитела вызывают сильный иммунный ответ у пациентов, как правило, после однократной инъекции в низкой дозе (Schroff, 1985 Cancer Res. 45:879-85, Shawler. J. Immunol. 1985 135:1530-5; Dillman, Cancer Biother. 1994 9:17-28). Поэтому было разработано несколько способов снижения иммуногенности антител мышей и других грызунов, а также технологии для получения полностью человеческих антител с использованием, например, трансгенных мышей или фагового дисплея. Были сконструированы химерные антитела, в которых скомбинированы вариабельные области грызунов и константные области человека (например, Boulianne Nature 1984 312:643-6), значительно уменьшая проблему иммуногенности (например, LoBuglio, Proc. Natl. Acad. Sci. 1989 86:4220-4; Clark, Immunol. Today 2000 21:397-402). Также сконструированы гуманизированные антитела, в которых собственно последовательность вариабельной области грызуна сконструирована таким образом, чтобы максимально соответствовать последовательности человека, сохраняя по меньшей мере исходные CDR, или где CDR антитела грызуна пересажены в каркас антитела человека (например, Riechmann, Nature 1988 332:323-7; US5693761). Поликлональные антитела кролика широко используют для биологических анализов, таких как ELISA или вестерн-блоттинг. Поликлональные антитела кролика как правило являются предпочтительными среди поликлональных антител грызунов, как правило, из-за их намного более высокой аффинности. Кроме того, у кролика часто можно получить хороший ответ антитела на антигены, которые вызывают слабый иммунный ответ у мышей и/или не являются источником удовлетворительных связывающих молекул при использовании в фаговом дисплее. Вследствие этих хорошо известных преимуществ антитела кролика могут быть идеальными для использования при исследовании и разработки терапевтических антител. Причина, по которой они широко не используются, в основном связана с техническими сложностями при получении моноклональных антител кролика. Так как у кроликов не известны опухоли, подобные миеломам, то для получения моноклональных антител кролика не может использоваться традиционная гибридомная технология. Впервые исследование способа получения клеточной линии-партнера для слияния с экспрессирующими антитела кролика клетки было проведено Knight et al. (Spieker-Polet et al., PNAS 1995, 92:9348-52), а улучшенная клеточная линия-партнер для слияния была описана Pytela et al. in 2005 (см. например патент США № 7429487). Однако эта технология не имеет широкого применения, так как ноу-хау в основном контролируется одной исследовательской группой. Для получения моноклональных антител в литературе описаны альтернативные способы, включающие клонирование антител из выбранных клеток, экспрессирующих антитела, посредством ОТ-ПЦР, но никогда еще не было публикаций об их успешном применении для антител кролика.
Полагают, что антитела кролика, подобно антителам мыши вызывают сильный иммунный ответ при использовании для лечения человека, таким образом, антитела кролика, перед тем, как их можно будет использовать клинически, необходимо гуманизировать. Однако способы, которые используют для получения гуманизированных антител грызунов, нельзя просто экстраполировать для антител кролика вследствие структурных различий между антителами кролика и мыши и, соответственно, между антителами кролика и человека. Например, CDR3 легкой цепи (CDR L3) часто является намного более длинной, чем ранее известные CDR L3 из антител человека или мыши.
На современном этапе развития в данной области было описано несколько подходов для гуманизации антител кролика, которые, однако, не являются классическим подходом для пересадки, в котором CDR не являющийся человеческим донорного антитела пересаживают на акцепторное антитело человека. В WO 04/016740 описан так называемый подход "изменения поверхности". Целью этого подхода "изменения поверхности" является модификация доступных для растворителя остатков не являющегося человеческим каркаса так, чтобы они становились более похожими на остатки человека. В данной области известны способы гуманизации антител кролика, подобные способам, описанным в WO 04/016740. В WO08/144757 и WO05/016950 описаны способы гуманизации моноклонального антитела кролика, включающие сравнение аминокислотных последовательностей исходного антитела кролика с аминокислотными последовательностями подобного антитела человека. Затем аминокислотную последовательность исходного антитела кролика изменяют так, чтобы его каркасные области были более похожи по последовательности с эквивалентными каркасными областями подобного антитела человека. Для получения хороших связывающих способностей для каждой иммуносвязывающей молекулы отдельно необходимо произвести трудоемкие опытно-конструкторские работы.
Возможной проблемой указанных выше подходов является то, что в них используют не каркас человека, а конструируют каркас кролика так, чтобы он выглядел более похожим на каркас человека. У такого подхода существует риск того, что участки аминокислот, находящиеся в сердцевине белка, все еще могут содержать иммуногенные T-клеточные эпитопы.
К настоящему времени заявители не нашли антител кролика, которые были гуманизированы с применением способов пересадки, существующих на современном уровне техники. Это можно объяснить тем, что CDR кролика может полностью отличаться от CDR человека или грызуна. Как известно в данной области, большинство цепей VH кролика относительно вариантов мыши и человека содержат дополнительные спаренные цистеины. В дополнение к консервативному дисульфидному мостику, формирующемуся между cys22 и cys92, также существует мостик cys21-cys79, а также S-S-мостик между CDR, формирующийся в некоторых цепях кролика между последним остатком CDR H1 и первым остатком CDR H2. Кроме того, пары остатков цистеина часто находят в CDR L3. Кроме того, многие CDR антител кролика не подходят ни одной из ранее известных канонических структур. В частности, CDR L3 часто является намного длиннее, чем CDR L3 вариантов человека или мыши.
Таким образом, пересадка CDR из не являющихся человеческими антител в каркас человека является важной задачей белковой инженерии. Перенос антигенсвязывающих петель из эволюционировавшего в природе каркаса в другой искусственно выбранный каркас человека необходимо проводить так, чтобы природные конформации петель сохранялись для связывания антигена. Часто после пересадки петель аффинность связывания антигена значительно снижается или исчезает. Использование тщательно подобранных каркасов человека при пересадке антигенсвязывающих петель максимально повышает вероятность сохранения аффинности связывания у гуманизированной молекулы (Roguzka et al 1996). Хотя для множества экспериментов по пересадке, доступных в литературе, приведено приблизительное руководство для пересадки CDR, обобщить этот принцип невозможно. Характерные проблемы после пересадки петель CDR связаны с потерей специфичности, стабильности или возможности получения.
Таким образом, существует актуальная необходимость в улучшенных способах надежной и быстрой гуманизации антител кролика для применения в качестве терапевтических и диагностических средств. Кроме того, существует необходимость в акцепторных каркасах человека для надежной гуманизации антител кролика, обеспечивающих функциональные антитела и/или фрагменты антител с биофизическими свойствами, подобными лекарственным средствам.
Сущность изобретения
Неожиданно было обнаружено, что каркас высокорастворимого и стабильного антитела человека, идентифицированного анализом "контроль качества" (QC) (как описано в WO 0148017 и в Auf der Maur et al., (2001), FEBS Lett. 508, p. 407-412) особенно удобен для встраивания CDR из других, не являющихся человеком видов животных, например, CDR кролика. Таким образом, в первом аспекте изобретение относится к вариабельным областям легкой и тяжелой цепей конкретного антитела человека (так называемого, антитела "FW1.4"), которые особенно удобны в качестве универсального акцептора для CDR из ряда антител, в частности, из антител кролика, с различной специфичностью связывания, вне зависимости от присутствия или отсутствия в CDR дисульфидного мостика. Кроме того, настоящее изобретение относится к двум мутантным последовательностям каркаса указанного конкретного антитела человека, а именно rFW1.4 и rFW1.4(V2), которые оба являются каркасами, особенно подходящими в качестве универсальных акцепторных каркасов для пересадки CDR кролика. В другом аспекте изобретение относится к повтору каркасных остатков, который делает каркас человека подходящим для встраивания CDR из других, не являющихся человеком видов животных, в частности, CDR кролика.
Гуманизированные иммуносвязывающие молекулы, полученные пересадкой CDR кролика в эти высокосовместимые каркасы вариабельных областей тяжелой и легкой цепей, постоянно и надежно сохраняют пространственную ориентацию антител кролика, из которых получены донорные CDR. Таким образом, нет никакой необходимости вводить в акцепторный каркас какие-либо структурно значимые позиции донорной иммуносвязывающей молекулы. Вследствие этих преимуществ можно достичь высокопроизводительной гуманизации антител кролика без оптимизации или с незначительной оптимизацией связывающей способности.
Таким образом, в другом аспекте изобретение относится к способам пересадки CDR кролика и других не являющихся человеческими CDR, в каркасные последовательности легкой цепи и/или тяжелой цепи растворимого и стабильного антитела человека, описываемые в настоящем документе, таким образом, получая гуманизированные антитела с превосходными биофизическими свойствами. В частности, иммуносвязывающие молекулы, получаемые способами по изобретению, демонстрируют превосходные функциональные свойства, такие как растворимость и стабильность.
Краткое описание рисунков
На фигуре 1 представлено определение CDR H1, используемой в настоящем документе для пересадки антигенсвязывающих участков из моноклональных антител кролика в каркасы высокорастворимого и стабильного антитела человека.
На фигуре 2 схематически представлена B-клетка 1, меченная флуоресцентным антителом 2, взаимодействующая с экспрессирующей мишень клеткой 3, окрашенной внутриклеточным красителем 4. Выбранная мишень: 5; BCR: 6.
Фигура 3: способ отбора FACS B-клеток кролика, связывающихся с растворимой мишенью ESBA903. Фиг. 3A: лимфоциты отбирают по прямому и боковому рассеянию. Фиг. 3B: Среди них выбирают клетки IgG+ IgM- (вероятно B-клетки памяти) (красное окно). Фиг. 3C: Полагают, что клетки с двойным окрашиванием ESBA903-PE и ESBA903-PerCP (зеленое окно) кодируют высокоаффинное IgG против ESBA903. Клетки, демонстрирующие наибольшую яркость флуоресценции (розовое окно), сортировали в 96-луночные планшеты.
Фигура 4. Гранулы, покрытые антителами против TNF-альфа (мечеными PE), связываются с клетками CHO, трансфицированными TNF-альфа (верхняя панель). Контрольные гранулы, покрытые антителами против CD19 (мечеными APC) не связываются с клетками CHO, трансфицированными TNF-альфа (средняя панель). Гранул, покрытые антителами против TNF-альфа (мечеными PE), не связываются с клетками CHO дикого типа (wt) (нижняя панель). Точечные диаграммы слева демонстрируют прямое и боковое рассеяние, которое указывает, соответственно, на размер и степень детализации событий. Группу единичных гранул (≈3 мкм) отбирают в P2. Клетки CHO окончательно связанные с гранулами (≈30 мкм) отбирают в P1. Точечные диаграммы в середине демонстрируют события P1 (клетки CHO) относительно их окрашивания PE или APC. Таким образом, если клетки взаимодействуют с гранулами с антителами против TNF-альфа, они должны появится в окне P3, а если они взаимодействуют с гранулами с антителами против CD19, они должны появится в окне P4. Справа детализирована статистика для каждого из образцов.
Фигура 5. Гранулы, покрытые антителами против TNF-альфа-PE, и гранулы, покрытые антителами против CD19-APC смешивают вместе с клетками CHO, трансфицированными TNF-альфа. Клетки CHO отбирают (P1) и из них клетки, связывающиеся с гранулами, покрытыми антителами против TNF-альфа-PE, или гранулами, покрытыми антителами против CD19-APC показаны в окнах P3 и P4, соответственно. Несвязанные гранулы видны в окне P2.
Фигура 6. Анализ последовательностей антител кролика, извлеченных из базы данных Kabat, подтверждает, что CDR3 вариабельной области тяжелой цепи, как правило, на три аминокислоты длиннее, чем ее вариант мыши.
Подробное описание изобретения
Определения
Для простоты понимания настоящего изобретения ниже определены некоторые термины. На всем протяжении подробного описания указаны дополнительные определения.
Термин "антитело" относится к полноразмерным антителам и к любому антигенсвязывающему фрагменту. Термин "антигенсвязывающий полипептид" и "иммуносвязывающая молекула" используют в настоящем документе взаимозаменяемо. "Антитело" относится к белку, необязательно гликозилированному, содержащему по меньшей мере две тяжелых (H) цепи и две легких (L) цепи, соединенных между собой дисульфидными связями, или к его антигенсвязывающей части. Каждая тяжелая цепь состоит из вариабельной области тяжелой цепи (сокращаемой в настоящем документе как VH) и константной области тяжелой цепи. Константная область тяжелой цепи состоит из трех доменов, CH1, CH2 и CH3. Каждая легкая цепь состоит из вариабельной области легкой цепи (сокращаемой в настоящем документе как VL) и константной области легкой цепи. Константная область легкой цепи состоит из одного домена, CL. Области VH и VL можно дополнительно подразделить на области гипервариабельности, называемые определяющими комплементарность областями (CDR), расположенные между более консервативными областями, называемыми каркасными областями (FR). Каждая VH и VL состоит из трех CDR и четырех FR, расположенными от N-конца до C-конца в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Вариабельные области тяжелых и легких цепей содержат связывающий домен, который взаимодействует с антигеном. Константные области антител могут опосредовать связывание иммуноглобулина с тканями или факторами хозяина, включая различные клетки иммунной системы (например, эффекторные клетки) и первый компонент (C1q) классической системы комплемента.
Термин "антигенсвязывающая часть" антитела (или просто "часть антитела") относится к одному или нескольким фрагментам антитела, которые сохраняют способность специфически связываться с антигеном (например, TNF). Показано, что антигенсвязывающую функцию антитела могут осуществлять фрагменты полноразмерного антитела. Примеры связывающих фрагментов, охватываемых термином "антигенсвязывающая часть" антитела, включают (i) Fab-фрагмент, моновалентный фрагмент, состоящий из доменов VL, VH, CL и CH1; (ii) F(ab')2-фрагмент, бивалентный фрагмент, содержащий два Fab-фрагмента, связанные дисульфидным мостиком в шарнирной области; (iii) Fd-фрагмент, состоящий из доменов VH и CH1; (iv) Fv-фрагмент, состоящий из доменов VL и VH одного плеча антитела; (v) один домен или фрагмент dAb (Ward et al., (1989) Nature 341:544-546), состоящий из домена VH; и (vi) выделенная определяющая комплементарность область (CDR) или (vii) сочетание двух или более выделенных CDR, которые необязательно может соединять синтетический линкер. Кроме того, хотя два домена Fv-фрагмента, VL и VH, кодируются отдельными генами, их, используя рекомбинантные способы, можно соединять синтетическим линкером, который позволяет получать их в виде одной белковой цепи, в которой области VL и VH спарены с формированием моновалентных молекул (известных как одноцепочечные Fv (scFv); см. например, Bird et al., (1988) Science 242:423-426; и Huston et al., (1988) Proc. Natl. Acad. Sci. USA 85:5879-5883). Такие одноцепочечные антитела также следует включать в термин "антигенсвязывающая часть" антитела. Эти фрагменты антител получают общепринятыми способами, известными специалистам в данной области, и фрагменты подвергают скринингу на полезность тем же способом, как и интактные антитела. Антигенсвязывающие части можно получать способами рекомбинантной ДНК, или ферментативным или химическим расщеплением интактных иммуноглобулинов. Антитела могут быть различного изотипа, например, антителом IgG (например, подтипов IgG1, IgG2, IgG3 или IgG4), IgA1, IgA2, IgD, IgE или IgM.
Термин "иммуносвязывающая молекула" относится к молекуле, содержащий весь антигенсвязывающий участок антитела или его часть, например, весь вариабельный домен тяжелой и/или легкой цепи или его часть так, что иммуносвязывающая молекула специфически распознает антиген-мишень. Неограничивающие примеры иммуносвязывающих молекул включают полноразмерные молекулы иммуноглобулинов и scFv, а также фрагменты антител, включая в качестве неограничивающих примеров (i) Fab-фрагмент, моновалентный фрагмент, состоящий из доменов VL, VH, CL и CH1; (ii) F(ab')2-фрагмент, бивалентный фрагмент, содержащий два Fab-фрагмента, связанных дисульфидным мостиком в шарнирной области; (iii) фрагмент Fab', который по существу представляет собой Fab с частью шарнирной области (см., Fundamental Immunology (Paul ed., 3.sup.rd ed. 1993); (iv) Fd-фрагмент, состоящий из доменов VH и CH1; (v) Fv-фрагмент, состоящий из доменов VL и VH одного плеча антитела, (vi) однодоменное антитело, такое как фрагмент dAb (Ward et al., (1989) Nature 341:544-546), состоящий из домена VH или VL, антител верблюдовых (см. Hamers-Casterman, et al., Nature 363:446-448 (1993), и Dumoulin, et al., Protein Science 11:500-515 (2002)) или акул (например, нанотела® Ig-NAR акул; и (vii) нанотело, вариабельная область тяжелой цепи, содержащая один вариабельный домен и два константных домена.
Термин "одноцепочечное антитело", "одноцепочечный Fv" или "scFv" относится к молекуле, содержащий вариабельный домен тяжелой цепи антитела (или область; VH) и вариабельный домен легкой цепи антитела (или область; VL), соединенные линкером. Такие молекулы scFv могут иметь общую структуру: NH2-VL-линкер-VH-COOH или NH2-VH-линкер-VL-COOH. Подходящие линкеры на существующем уровне техники состоят из повторяющихся аминокислотных последовательностей GGGGS или их вариантов. В предпочтительном варианте осуществления настоящего изобретения используют линкер (GGGGS)4 с аминокислотной последовательностью SEQ ID NO:8, но также возможны варианты из 1-3 повторов (Holliger et al., (1993), Proc. Natl. Acad. Sci. USA 90:6444-6448). Другие линкеры, которые можно использовать по настоящему, изобретению описаны в Alfthan et al., (1995), Protein Eng. 8:725-731, Choi et al., (2001), Eur. J. Immunol. 31:94-106, Hu et al., (1996), Cancer Res. 56:3055-3061, Kipriyanov et al., (1999), J. Mol. Biol. 293:41-56 и Roovers et al., (2001), Cancer Immunol.
Как используется в настоящем документе, термин "функциональное свойство" представляет собой свойство полипептида (например, иммуносвязывающей молекулы), улучшение которого (например, относительно обычного полипептида) является желательным и/или предпочтительным для специалиста в данной области, например, для улучшения технологических свойств или терапевтической эффективности полипептида. В одном из вариантов осуществления функциональное свойство представляет собой стабильность (например, термостабильность). В другом варианте осуществления функциональное свойство представляет собой растворимость (например, в условиях клетки). В еще одном варианте осуществления функциональное свойство представляет собой характер агрегации. В другом варианте осуществления функциональное свойство представляет собой экспрессию белка (например, в прокариотической клетке). В еще одном варианте осуществления функциональное свойство представляет собой характер рефолдинга после растворения телец включения в производственном процессе. В определенных вариантах осуществления функциональное свойство не представляет собой улучшения аффинности связывания антигена. В другом предпочтительном варианте осуществления улучшение одного или нескольких функциональных свойств не оказывает значительного эффекта на аффинность связывания иммуносвязывающей молекулы.
Термин "CDR" относится к одной из шести гипервариабельных областей в вариабельных доменах антитела, которые в основном вносят вклад в связывание антигена. Одно из наиболее общеупотребительных определений для шести CDR предоставлено в Kabat E.A. et al., (1991) Sequences of proteins of immunological interest. NIH Publication 91-3242). Как используется в настоящем документе, определение CDR по Kabat используют только для CDR1, CDR2 и CDR3 вариабельного домена легкой цепи (CDR L1, CDR L2, CDR L3 или LI, L2, L3), а также для CDR2 и CDR3 вариабельного домена тяжелой цепи (CDR H2, CDR H3 или H2, H3). Однако, как используют в настоящем документе, CDR1 вариабельного домена тяжелой цепи (CDR H1 или H1) определяют по положениям остатков (нумерация по Kabat), начиная с положения 26 и заканчивая до положения 36. Это определение по существу представляет собой объединение CDR H1, различно определенного по Kabat и Chotia (также см. фигуру 1 для иллюстрации).
Как используется в настоящем документе термин "каркас антитела" относится к части вариабельного домена, VL или VH, которая служит в качестве поддержки для антигенсвязывающих петель (CDR) этого вариабельного домена. По существу он представляет собой вариабельный домен без CDR.
Термин "эпитоп" или "антигенная детерминанта" относится к участку на антигене, с которым специфически связывается иммуноглобулин или антитело (например, специфический участок на молекуле TNF). Как правило, эпитоп включает по меньшей мере 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15 последовательных или не последовательных аминокислот в уникальной пространственной конформации. См., например, Epitope Mapping Protocols in Methods in Molecular Biology, Vol. 66, G. E. Morris, Ed. (1996).
Термины "специфическое связывание", "селективное связывание", "селективно связывается" и "специфически связывается" относятся к связыванию антитела с эпитопом на предопределенном антигене. Как правило, антитело связывается с аффинностью (KD) приблизительно менее 10-7 M, такой как приблизительно менее 10-8 M, 10-9 M или 10-10 M или даже менее.
Термин "KD" или "Kd" относится к равновесной константе диссоциации конкретного взаимодействия антитело-антиген. Как правило, антитела по изобретению связываются с TNF с равновесной константой диссоциации (KD) менее чем приблизительно 10-7 M, такой как менее чем приблизительно 10-8 M, 10-9 M или 10-10 M или даже менее, например, как определяют с применением технологии поверхностного плазмонного резонанса (SPR) на устройстве BIACORE.
Как используется в настоящем документе термин "молекула нуклеиновой кислоты", относится к молекулам ДНК и к молекулам РНК. Молекула нуклеиновой кислоты может быть одноцепочечной или двухцепочечной, но, предпочтительно, представляет собой двухцепочечную ДНК. Нуклеиновая кислота является "функционально связанной", если она находится в функциональной связи с другой последовательностью нуклеиновой кислоты. Например, промотор или энхансер функционально связан с кодирующей последовательностью, если он влияет на транскрипцию последовательности.
Термин "вектор" относится к молекуле нуклеиновой кислоты, способной к транспортировке другой нуклеиновой кислоты, с которой он соединен. В одном из вариантов осуществления вектор представляет собой "плазмиду", которая обозначает замкнутую кольцевую двухцепочечную ДНК в которую можно лигировать дополнительные участки ДНК. В другом варианте осуществления вектор представляет собой вирусный вектор, где дополнительные участки ДНК можно лигировать в вирусный геном. Векторы, описываемые в настоящем документе, могут быть способными к автономной репликации в клетке-хозяине, в которую их вводят (например, бактериальные векторы с бактериальным участком начала репликации и эписомные векторы млекопитающих), или могут интегрироваться в геном клетки-хозяина после введения в клетку-хозяина и, таким образом, реплицируются вместе с геномом хозяина (например, неэписомные векторы млекопитающих).
Термин "клетка-хозяин" относится к клетке, в которую вводят вектор экспрессии. Клетки-хозяева включают бактериальные, микробные, растительные или животные клетки, предпочтительно, клетки Escherichia coli, Bacillus subtilis; Saccharomyces cerevisiae, Pichia pastoris, CHO (линии китайского хомяка) или NSO.
Термин "зайцеобразные" относится к представителям таксономического порядка Lagomorpha, включающего семейства Leporidae (например, зайцы и кролики), и Ochotonidae (пищухи). В наиболее предпочтительном варианте осуществления, зайцеобразные представляет собой кролика. Как используется в настоящем документе термин "кролик" относится к животному, принадлежащему к семейству Leporidae.
Как используется в настоящем документе, "идентичность" относится к совпадению последовательностей двух полипептидов, молекул или двух нуклеиновых кислот. Когда положение в двух сравниваемых последовательностях занято одним и тем же основанием или одной и той же аминокислотной мономерной субъединицей (например, если положение в обоих из двух полипептидов занято лизином), тогда соответствующие молекулы являются идентичными по этому положению. "Процент идентичности" двух последовательностей представляет собой функцию количества идентичных положений, выявляемых в последовательностях с учетом количества пропусков и длины каждого пропуска, которые необходимо внести для оптимального выравнивания двух последовательностей. Как правило, сравнение проводят, когда две последовательности выровнены с получением максимума идентичности. Такое выравнивание можно проводить, например, с помощью алгоритма Needleman and Wunsch (J. Mol. Biol. (48):444-453 (1970)), на котором основана программа GAP в программный пакет GCG, с использованием матрицы Blossum 62 или матрицы PAM250, и весом пропуска 16, 14, 12, 10, 8, 6 или 4 и длиной пропуска 1, 2, 3, 4, 5 или 6.
Если не указано иного, все технические и научные термины, используемые в настоящем документе, имеют то же значение, как обычно понимает специалист в области, к которой принадлежит настоящее изобретение. Хотя в практическом осуществлении или тестировании настоящего изобретения можно использовать способы и материалы, сходные или эквивалентные способам и материалам, описываемым в настоящем документе, ниже описаны подходящие способы и материалы. В случае сомнений следует руководствоваться настоящим описанием. Кроме того, материалы, способы и примеры являются только иллюстративными и не предназначены для ограничения.
В приведенных ниже разделах подробнее описаны различные аспекты изобретения. Следует понимать, что различные варианты осуществления, наиболее предпочтительные варианты осуществления и диапазоны можно произвольно комбинировать. Кроме того, в зависимости от конкретного варианта осуществления выбранные определения, варианты осуществления или диапазоны могут не использоваться.
Если не указано иного, положения аминокислот указаны в соответствии со схемой нумерации AHo. Система нумерации AHo дополнительно описана в Honegger, A. and Pluckthun, A. (2001) J. Mol. Biol. 309:657-670). Альтернативно можно использовать нумерацию по системе Kabat, как дополнительно описано в Kabat et al., (Kabat, E. A., et al., (1991) Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No. 91-3242). Таблицы преобразования для двух различных систем нумерации, используемых для идентификации положений аминокислотных остатков в вариабельных областях тяжелых и легких цепей антител, предоставлены в A. Honegger, J. Mol. Biol. 309 (2001) 657-670.
В первом аспекте настоящее изобретение относится к универсальному акцепторному каркасу для пересадки CDR других видов животных, например, от кролика. Ранее описано, что антитела или производные антител, содержащие каркасы человека, идентифицированные в так называемом скрининге "контроль качества" (WO0148017), как правило, характеризуются высокой стабильностью и/или растворимостью. Хотя одноцепочечный каркас человека FW1.4 (комбинация SEQ ID NO:1 (называемого в WO03/097697 a43) и SEQ ID NO:2 (называемого в WO03/097697 KI27)) по характерным параметрам однозначно уступает в анализе "контроль качества", неожиданно обнаружено, что он обладает высокой собственной термодинамической стабильностью и хорошо продуцируется, притом в сочетании с рядом различных CDR. Стабильность этой молекулы в основном можно считать свойством ее каркасных областей. Кроме того, показано, что FW1.4 по существу высоко совместим с антигенсвязывающими участками антител кролика. Таким образом, FW1.4, представляет собой подходящую поддерживающую молекулу для конструирования стабильных гуманизированных фрагментов антител scFv, получаемых при пересадке петель кролика. Таким образом, в одном из аспектов изобретение относится к акцепторному каркасу иммуносвязывающей молекулы, содержащему последовательность VH по меньшей мере на 90% идентичную SEQ ID NO:1, и/или последовательность VL по меньшей мере на 85% идентичную SEQ ID NO:2, более предпочтительно, содержащему последовательность FW1.4 (SEQ ID NO:3), для пересадки CDR кролика, или последовательность по меньшей мере на 60%, более предпочтительно, по меньшей мере на 65%, 70%, 75%, 80%, 85%, 90%, 95% идентичную SEQ ID NO:3.
Кроме того, выявлено, что FW1.4 можно оптимизировать, замещая несколько остатков в положениях в тяжелой цепи FW1.4 и/или замещая остаток в 1 положении в легкой цепи FW1.4. Таким образом, неожиданно обнаружено, что конформация петель большого множества CDR кролика в VH может полностью сохраняться, как правило, вне зависимости от последовательности донорного каркаса. Указанные остатки в тяжелой цепи, а также в 1 положении в легкой цепи FW1.4 являются консервативными в антителах кролика. Консенсусный остаток для положений в тяжелой цепи, а также одного положения в легкой цепи устанавливают на основании репертуара кролика и вводят в последовательность акцепторного каркаса человека.
Как результат, модифицированный каркас 1.4 (далее в настоящем документе, обозначаемый как rFW1.4) совместим практически с любой CDR кролика. Кроме того, rFW1.4, содержащая различные CDR кролика, хорошо экспрессируется и хорошо продуцируется в отличие от одиночных цепей кролика дикого типа и при этом почти полностью сохраняет аффинность исходного донорного антитела кролика.
Таким образом, настоящее изобретение относится к каркасу вариабельной области тяжелой цепи SEQ ID NO:1, дополнительно содержащему один или несколько аминокислотных остатков, которые, как правило, поддерживают конформацию CDR, получаемых из иммуносвязывающей молекулы кролика. В частности, указанные остатки находятся в одном или нескольких положениях аминокислот, выбранных из группы, состоящей из 24H, 25H, 56H, 82H, 84H, 89H и 108H (нумерация AHo). Доказано, что эти положения влияют на конформацию CDR и, таким образом, предусмотрены для мутации для встраивания донорных CDR. Предпочтительно, указанные один или несколько остатков выбраны из группы, состоящей из: треонина (T) в положении 24, валина (V) в положении 25, глицина или аланина (G или A) в положении 56, лизина (K) в положении 82, треонина (T) в положении 84, валина (V) в положении 89 и аргинина (R) в положении 108 (нумерация AHo). Предпочтительно, присутствуют по меньшей мере три, более предпочтительно, четыре, пять, шесть, а наиболее предпочтительно, все семь остатков. Неожиданно выявлено, что присутствие указанных остатков повышает стабильность иммуносвязывающей молекулы.
В предпочтительном варианте осуществления изобретение относится к акцепторному каркасу иммуносвязывающей молекулы, содержащему VH по меньшей мере на 50%, более предпочтительно, по меньшей мере на 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95% и даже, более предпочтительно, на 100% идентичную SEQ ID NO:4, при условии, что присутствуют по меньшей мере один, более предпочтительно, по меньшей мере три, более предпочтительно, четыре, пять, шесть, а наиболее предпочтительно, семь остатков из группы, содержащей треонин (T) в положении 24, валин (V) в положении 25, аланин (A) или глицин (G) в положении 56, треонин (T) в положении 84, лизин (K) в положении 82, валин (V) в положении 89 и аргинин (R) в положении 108 (нумерация AHo). В предпочтительном варианте осуществления акцепторный каркас иммуносвязывающей молекулы представляет собой акцепторный каркас иммуносвязывающей молекулы для CDR кролика.
В предпочтительном варианте осуществления указанный каркас вариабельной области тяжелой цепи представляет собой или содержит SEQ ID NO:4 или SEQ ID NO:6. Оба из указанных каркасов вариабельных областей тяжелых цепей можно, например, комбинировать с любым подходящим каркасом легкой цепи.
Таким образом, настоящее изобретение относится к акцепторному каркасу иммуносвязывающей молекулы, содержащему
(i) каркас вариабельной области тяжелой цепи по меньшей мере на 70% идентичный, предпочтительно, по меньшей мере на 75%, 80%, 85%, 90%, более предпочтительно, по меньшей мере на 95% идентичный SEQ ID NO:4; и/или
(ii) каркас вариабельной области легкой цепи по меньшей мере на 70% идентичный, предпочтительно, по меньшей мере на 75%, 80%, 85%, 90%, более предпочтительно, по меньшей мере на 95% идентичный SEQ ID NO:2.
В более предпочтительном варианте осуществления, каркас вариабельной области тяжелой цепи содержит треонин (T) в положении 24, глицин (G) в положении 56, треонин (T) в положении 84, валин (V) в положении 89 и аргинин (R) в положении 108 (нумерация AHo).
В предпочтительном варианте осуществления вариабельная область легкой цепи содержит треонин (T) в положении 87 (нумерация AHo).
В предпочтительном варианте осуществления указанный акцепторный каркас иммуносвязывающей молекулы содержит
(i) каркас вариабельной области тяжелой цепи, выбранный из группы, состоящей из SEQ ID NO:1, SEQ ID NO:4 и SEQ ID NO:6; и/или
(ii) каркас вариабельной области легкой цепи SEQ ID NO:2 или SEQ ID NO:9.
В предпочтительном варианте осуществления каркас вариабельной области тяжелой цепи соединен с каркасом вариабельной области легкой цепи посредством линкера. Линкер может представлять собой любой подходящий линкер, например линкер, содержащий от 1 до 4 повторов последовательности GGGGS, предпочтительно, пептид (GGGGS)4 (SEQ ID NO:8), или линкер, как описано в Alfthan et al. (1995) Protein Eng. 8:725-731.
В другом предпочтительном варианте осуществления акцепторный каркас иммуносвязывающей молекулы представляет собой последовательность по меньшей мере на 70%, 75%, 80%, 85%, 90%, более предпочтительно, по меньшей мере на 95% идентичную SEQ ID NO:5, с тем условием, что последовательность, предпочтительно, не представляет собой SEQ ID NO:3. Более предпочтительно, акцепторный каркас иммуносвязывающей молекулы содержит или представляет собой SEQ ID NO:5.
В другом предпочтительном варианте осуществления акцепторный каркас иммуносвязывающей молекулы представляет собой последовательность по меньшей мере на 70%, 75%, 80%, 85%, 90%, более предпочтительно, по меньшей мере на 95% идентичную SEQ ID NO:7, с тем условием, что последовательность, предпочтительно, не представляет собой SEQ ID NO:3. Более предпочтительно, акцепторный каркас иммуносвязывающей молекулы содержит или представляет собой SEQ ID NO:7.
Кроме того, неожиданно было обнаружено, что присутствие описанного выше аминокислотного повтора делает каркас, предпочтительно, каркас человека, особенно подходящим для введения CDR других не являющихся человеком видов животных, в частности, CDR кролика. Указанный повтор не оказывает отрицательного влияния на стабильность иммуносвязывающей молекулы. CDR представлены в конформации, сходной с их природной пространственной ориентацией в иммуносвязывающей молекуле кролика; таким образом, необходимости пересадки в акцепторный каркас структурно значимый положений нет. Таким образом, акцепторный каркас иммуносвязывающей молекулы человека или гуманизированный акцепторный каркас иммуносвязывающей молекулы содержит по меньшей мере три аминокислоты, предпочтительно, четыре, пять, шесть, а более предпочтительно, семь аминокислот из группы, состоящей из треонина (T) в положении 24, валина (V) в положении 25, аланина (A) или глицина (G) в положении 56, лизина (K) в положении 82, треонина (T) в положении 84, валина (V) в положении 89 и аргинина (R) в положении 108 (нумерация AHo).
Как описано в настоящем документе, акцепторные каркасы иммуносвязывающих молекул могут содержать повышающую растворимость замену в каркасе тяжелой цепи, предпочтительно, в положениях 12, 103 и 144 (нумерация AHo). Предпочтительно, гидрофобную аминокислоту заменяют более гидрофильной аминокислотой. Гидрофильные аминокислоты представляют собой например аргинин (R), аспарагин (N), аспарагиновую кислоту (D), глутамин (Q), глицин (G), гистидин (H), лизин (K), серин (S) и треонин (T). Более предпочтительно, каркас тяжелой цепи содержит (a) серин (S) в положении 12; (b) серин (S) или треонин (T) в положении 103 и/или (c) серин (S) или треонин (T) в положении 144.
Кроме того, в одном или нескольких положениях из 1, 3, 4, 10, 47, 57, 91 и 103 каркаса вариабельной области легкой цепи (нумерация AHo) могут присутствовать повышающие стабильность аминокислоты. Более предпочтительно, каркас вариабельной области легкой цепи содержит глутаминовую кислоту (E) в положении 1, валин (V) в положении 3, лейцин (L) в положении 4, серин (S) в положении 10; аргинин (R) в положении 47, серин (S) в положении 57, фенилаланин (F) в положении 91 и/или валин (V) в положении 103.
Так как глутамин (Q) подвержен дезаминированию, в другом предпочтительном варианте осуществления VH содержит в положении 141 глицин (G). Эта замена может увеличивать длительность хранения белка.
Например, акцепторные каркасы, описываемые в настоящем документе, можно использовать для получения антитела человека или гуманизированного антитела, сохраняющего свойства связывания не являющегося человеческим антитела, из которого получают не являющиеся человеческими CDR. Таким образом, в предпочтительном варианте осуществления настоящее изобретение относится к акцепторному каркасу иммуносвязывающей молекулы, как описано в настоящем документе, дополнительно содержащему CDR1, CDR2 и CDR3 тяжелой цепи и/или CDR1, CDR2 и CDR3 легкой цепи из донорной иммуносвязывающей молекулы, предпочтительно, из иммуносвязывающей молекулы млекопитающего, более предпочтительно, из иммуносвязывающей молекулы зайцеобразного, и, наиболее предпочтительно, кролика. Таким образом, в одном из вариантов осуществления изобретение относится к иммуносвязывающей молекуле, специфичной к желаемому антигену, содержащей
(i) CDR вариабельной области легкой цепи зайцеобразного; и
(ii) каркас вариабельной области тяжелой цепи человека по меньшей мере на 50%, более предпочтительно, по меньшей мере на 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 95%, а наиболее предпочтительно, на 100% идентичный SEQ ID NO:4.
В одном из предпочтительных вариантов осуществления, существует условие, что в указанной последовательности каркаса вариабельной области тяжелой цепи человека присутствует по меньшей мере одна аминокислота из группы, состоящей из треонина (T) в положении 24, валина (V) в положении 25, аланина (A) или глицина (G) в положении 56, треонина (T) в положении 84, лизина (K) в положении 82, валина (V) в положении 89 и аргинина (R) в положении 108 (нумерация AHo).
Предпочтительно, зайцеобразное представляет собой кролика. Более предпочтительно, иммуносвязывающая молекула содержит CDR1, CDR2 и CDR3 тяжелой цепи и CDR1, CDR2 и CDR3 легкой цепи из донорной иммуносвязывающей молекулы.
Как известно в данной области, многие цепи VH кролика по сравнению с вариантами мыши и человека содержат дополнительные спаренные цистеины. Кроме консервативного дисульфидного мостика, образующемуся между cys22 и cys92, также существует мостик cys21-cys79, а также S-S-мостик между CDR, образующийся в некоторых цепях кролика между последним остатком CDR H1 и первым остатком CDR H2. Кроме того, пары остатков цистеина часто находят в CDR L3. Кроме того, многие CDR антител кролика не подходят ни к одной из ранее известных канонических структур. В частности, CDR L3 часто является намного длиннее, чем CDR L3 вариантов человека или мыши.
Как указано ранее, пересадка не являющихся человеческими CDR в каркасы, описываемые в настоящем документе, дает молекулу, где CDR представлены в правильной конформации. Если необходимо, аффинность иммуносвязывающей молекулы можно улучшать посредством пересадки взаимодействующих с антигеном каркасных остатков не являющейся человеческой донорной иммуносвязывающей молекулы. Эти положения можно идентифицировать, например, посредством
(i) идентификации соответствующей последовательности предшественника зародышевой линии или, альтернативно, в случае высокогомологичных каркасных последовательностей, используя консенсусные последовательности;
(ii) проведения выравнивания последовательности донорного вариабельного домена с последовательностью предшественника зародышевой линии или консенсусной последовательностью из этапа (i); и
(iii) идентификации различающихся остатков.
Различающиеся остатки на поверхности молекулы во многих случаях мутируют в процессе формирования аффинности in vivo, предположительно для формирования аффинности к антигену.
В другом аспекте настоящее изобретение относится к иммуносвязывающей молекуле, которая содержит акцепторный каркас иммуносвязывающей молекулы, описываемый в настоящем документе. Например, указанная иммуносвязывающая молекула может представлять собой антитело scFv, полноразмерный иммуноглобулин, Fab-фрагмент, Dab или нанотело.
В предпочтительном варианте осуществления иммуносвязывающая молекула присоединена к одной или нескольким молекулам, например, терапевтического средства, таким как цитотоксическое средство, цитокин, хемокин, фактор роста или другая сигнальная молекула, визуализирующее средство или второй белок, такой как активатор транскрипции или ДНК-связывающий домен.
Как описано в настоящем документе, иммуносвязывающую молекулу можно использовать, например, в диагностических целях, в терапии, для подтверждения мишени или генотерапии.
Изобретение дополнительно относится к выделенной нуклеиновой кислоте, кодирующей акцепторный каркас иммуносвязывающей молекулы, описываемый в настоящем документе, или иммуносвязывающую молекулу(ы), как описано в настоящем документе.
В другом варианте осуществления изобретение относится к вектору, содержащему нуклеиновую кислоту, описываемую в настоящем документе.
Нуклеиновую кислоту или вектор, как описано в настоящем документе, можно использовать, например, в генотерапии.
Изобретение дополнительно относится к клетке-хозяину, содержащей вектор и/или нуклеиновую кислоту, описываемые в настоящем документе.
Кроме того, предоставлена композиция, содержащая акцепторный каркас иммуносвязывающей молекулы, как описано в настоящем документе, иммуносвязывающую молекулу, как описано в настоящем документе, выделенную нуклеиновую кислоту, как описано в настоящем документе, или вектор, как описано в настоящем документе.
Последовательности, описываемые в настоящем документе, представляют собой следующее (остатки X представляют собой участки вставок CDR):
SEQ ID NO:1: каркас вариабельной области тяжелой цепи FW1.4 (a43)
EVQLVESGGGLVQPGGSLRLSCAAS(X)n=1-50WVRQAPGKGLEWVS(X)n=1-50RFTISRDNSKNTLYLQMNSLRAEDTAVYYCAK(X)n=1-50WGQGTLVTVSS
SEQ ID NO:2: каркас вариабельной области легкой цепи FW1.4 (KI27)
EIVMTQSPSTLSASVGDRVIITC(X)n=1-50WYQQKPGKAPKLLIY(X)n=1-50GVPSRFSGSGSGAEFTLTISSLQPDDFATYYC(X)n=1-50FGQGTKLTVLG
SEQ ID NO:3: каркас FW1.4
EIVMTQSPSTLSASVGDRVIITC(X)n=1-50WYQQKPGKAPKLLIY(X)n=1-50GVPSRFSGSGSGAEFTLTISSLQPDDFATYYC(X)n=1-50FGQGTKLTVLGGGGGSGGGGSGGGGSGGGGSEVQLVESGGGLVQPGGSLRLSCAAS(X)n=1-50WVRQAPGKGLEWVS(X)n=1-50RFTISRDNSKNTLYLQMNSLRAEDTAVYYCAK(X)n=1-50WGQGTLVTVSS
SEQ ID NO:4: каркас вариабельной области тяжелой цепи rFW1.4
EVQLVESGGGLVQPGGSLRLSCTAS(X)n=1-50WVRQAPGKGLEWVG(X)n=1-50RFTISRDTSKNTVYLQMNSLRAEDTAVYYCAR(X)n=1-50WGQGTLVTVSS
SEQ ID NO:5: каркас rFW1.4
EIVMTQSPSTLSASVGDRVIITC(X)n=1-50WYQQKPGKAPKLLIY(X)n=1-50GVPSRFSGSGSGTEFTLTISSLQPDDFATYYC(X)n=1-50FGQGTKLTVLGGGGGSGGGGSGGGGSGGGGSEVQLVESGGGLVQPGGSLRLSCTAS(X)n=1-50WVRQAPGKGLEWVG(X)n=1-50RFTISRDTSKNTVYLQMNSLRAEDTAVYYCAR(X)n=1-50WGQGTLVTVSS
SEQ ID NO:6: каркас вариабельной области тяжелой цепи rFW1.4(V2)
EVQLVESGGGLVQPGGSLRLSCTVS(X)n=1-50WVRQAPGKGLEWVG(X)n=1-50RFTISKDTSKNTVYLQMNSLRAEDTAVYYCAR(X)n=1-50WGQGTLVTVSS
SEQ ID NO:7: каркас rFW1.4(V2)
EIVMTQSPSTLSASVGDRVIITC(X)n=1-50WYQQKPGKAPKLLIY(X)n=1-50GVPSRFSGSGSGTEFTLTISSLQPDDFATYYC(X)n=1-50FGQGTKLTVLGGGGGSGGGGSGGGGSGGGGSEVQLVESGGGLVQPGGSLRLSCTVS(X)n=1-50WVRQAPGKGLEWVG(X)n=1-50RFTISKDTSKNTVYLQMNSLRAEDTAVYYCAR(X)n=1-50WGQGTLVTVSS
SEQ ID NO:8: линкер
GGGGSGGGGSGGGGSGGGGS
SEQ ID NO:9: замещенный каркас вариабельной области легкой цепи FW1.4
EIVMTQSPSTLSASVGDRVIITC(X)n=1-50WYQQKPGKAPKLLIY(X)n=1-50GVPSRFSGSGSGTEFTLTISSLQPDDFATYYC(X)n=1-50FGQGTKLTVLG
В другом аспекте изобретение относится к способам гуманизации не являющихся человеческими антител посредством пересадки CDR не являющихся человеческими донорных антител в каркасы стабильных и растворимых антител. В особенно предпочтительном варианте осуществления CDR, происходящие из антител кролика, и каркасы являются CDR и каркасами, описанными выше.
Общий способ пересадки CDR в акцепторные каркасы человека описан Winter в патенте США № 5225539 и Queen et al. в WO9007861 A1, которые, таким образом, в полном объеме включены в качестве ссылки. Общая стратегия пересадки CDR из моноклональных антител кролика в выбранные каркасы сходна со стратегией Winter et al. и Queen et al., но расходится по некоторым ключевым аспектам. В частности, способы по изобретению отличаются от обычной методологии Winter и Queen, известной в данной области тем, что каркасы антител человека, как описано в настоящем документе, являются особенно подходящими в качестве универсальных акцепторов для донорные антител человека или не являющихся человеческими донорных антител. Таким образом, в отличие от общего способа Winter и Queen, каркасная последовательность, используемая для способов гуманизации по изобретению не обязательно представляет собой каркасную последовательность, демонстрирующую наибольшее сходство последовательности с последовательностью не являющегося человеческим антитела (например, кролика) из которого получают донорные CDR. Кроме того, для поддержки конформации CDR не требуется пересадки остатков каркаса из донорной последовательности. По большей мере вводить можно расположенные в каркасе антигенсвязывающие аминокислоты или другие мутации, которые происходят при соматическом гипермутировании.
Ниже описаны конкретные варианты способов пересадки для получения гуманизированных антител с высокой растворимостью и стабильностью, производных из антител кролика.
Таким образом, изобретение относится к способу гуманизации иммуносвязывающей молекулы-донора CDR кролика, которая содержит последовательности CDR1, CDR2 и CDR3 тяжелой цепи и/или последовательности CDR1, CDR2 и CDR3 легкой цепи. Способ включает этапы:
(i) пересадки в тяжелую цепь по меньшей мере одной, предпочтительно, двух, более предпочтительно, трех CDR из группы, состоящей из последовательностей CDR1, CDR2 и CDR3 в акцепторный каркас тяжелой цепи человека по меньшей мере на 50%, предпочтительно, по меньшей мере на 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, более предпочтительно, по меньшей мере на 95% идентичный SEQ ID NO:1; и/или
(ii) пересадка в легкую цепь по меньшей мере одной, предпочтительно, двух, более предпочтительно, трех CDR из группы, состоящей из последовательности CDR1, CDR2 и CDR3 в акцепторный каркас легкой цепи человека, где каркас легкой цепи человека по меньшей мере на 50%, предпочтительно, по меньшей мере на 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, более предпочтительно, по меньшей мере на 95% идентичен SEQ ID NO:2.
В предпочтительном варианте осуществления акцепторный каркас вариабельной цепи содержит (i) каркас тяжелой цепи человека, содержащий каркасную аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:1, SEQ ID NO:4 и SEQ ID NO:6 и (ii) каркас легкой цепи человека, содержащий каркасную аминокислотную последовательность SEQ ID NO:2 или SEQ ID NO:9.
В более предпочтительном варианте осуществления способ включает этап (i) пересадки последовательностей CDR1, CDR2 и CDR3 тяжелой цепи в тяжелую цепь и (ii) пересадки последовательностей CDR1, CDR2 и CDR3 легкой цепи в легкую цепь иммуносвязывающей молекулы по меньшей мере на 75%, 80%, 85%, 90%, более предпочтительно, по меньшей мере на 95% идентичной SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:7. Более предпочтительно, иммуносвязывающая молекула представляет собой или содержит SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:7.
В другом варианте осуществления для улучшения связывания антигена способ может дополнительно включать этап замещения каркасных остатков акцептора остатками донора, которые вовлечены в связывание антигена.
В иллюстративных вариантах осуществления способов по изобретению аминокислотную последовательность антитела-донора CDR идентифицируют впервые, а последовательности выравнивают традиционными способами выравнивания последовательностей (например, алгоритма Нидлмана-Вунша и матриц Blossum). Введение пропусков и обозначение положений остатков можно проводить с использованием традиционной системы нумерации в антителах. Например, можно использовать систему нумерации AHo для вариабельных доменов иммуноглобулинов. Также можно использовать схему нумерации по Kabat, так как она является наиболее широко принятым стандартом нумерации остатков в антителе. Например, нумерацию по Kabat можно определять с применением программы SUBIM. Эта программа анализирует вариабельные области последовательности антитела и нумерует последовательность в соответствии с системой, разработанной Kabat и коллегами (Deret et al 1995). Определение областей каркаса и CDR, как правило, проводят в соответствии с определением Kabat, которое основано на вариабельности последовательности и является чаще всего используемым. Однако для CDR H1 обозначение предпочтительно, представляет собой комбинацию определений Kabat, средних данных о контактах, получаемых посредством анализа контактов антитела и антигена подмножества трехмерных сложных структур (MacCallum et al., 1996), и Chotia, которое основано на определении положения областей структурных петель (также см. фиг. 1). Таблицы преобразования для различных систем нумерации, используемых для идентификации положений аминокислотных остатков в вариабельных областях тяжелых и легких цепей антител предоставлены в A. Honegger, J. Mol. Biol. 309 (2001) 657-670. Нумерация по системе Kabat дополнительно описана в Kabat et al. (Kabat, E. A., et al., (1991) Sequences of Proteins of Immunological Interest, Fifth Edition, U.S. Department of Health and Human Services, NIH Publication No. 91-3242). Система нумерации AHo дополнительно описана в Honegger, A. and Pluckthun, A. (2001) J. Mol. Biol. 309:657-670.
Например, вариабельные домены моноклональных антител кролика можно классифицировать в соответствующие подгруппы человека с применением, например, реализации алгоритмов анализа последовательностей EXCEL и способов классификации на основе анализа репертуара антител человека (Knappik et al., 2000, J. Mol. Biol. Feb 11;296(1):57-86).
В донорных антигенсвязывающих областях можно определять конформацию CDR, затем также можно определять положения остатков, необходимых для поддержания различных канонических структур. Канонические структуры CDR для пяти из шести гипервариабельных областей антител в антителах кролика (L1, L2, L3, H1 и H2) определяют с применением определения Chothia (1989).
В предпочтительном варианте осуществления CDR получают, идентифицируют и выделяют следующим способом: B-клетки, предпочтительно, B-клетки кролика, инкубируют с (i) антигенами-мишенями (предпочтительно, очищенными) или (ii) с клетками, экспрессирующими на своей поверхности антиген-мишень.
В случае (ii) указанные клетки, экспрессирующие антиген-мишень, могут представлять собой, например, клетки млекопитающих, предпочтительно, клетки CHO или HEK293, дрожжевые клетки, предпочтительно, сферобласты дрожжей, или бактериальные клетки, которые в природе экспрессируют выбранную мишень или трансформированы так, чтобы экспрессировать на своей поверхности белок-мишень. При экспрессии антиген-мишень может экспрессироваться на клеточной поверхности или интегрированным в клеточную мембрану или прикрепленным к ней. Клетки можно культивировать, например, в виде изолированных линий в клеточной культуре или выделять из их природной среды, например, из ткани, органа или организма.
Предоставление антигена-мишени, экспрессированным на поверхности клеток, т.е. случай (ii), особенно предпочтительно, для трансмембранных белов, даже более предпочтительно, для трансмембранных белков с несколькими трансмембранными доменами, таких как GPCR (сопряженные с G-белками рецепторы), или ионных каналов или любого другого белка, у которого после рекомбинантной экспрессии и очистки трудно поддерживать природную конформацию. Традиционная иммунизация рекомбинантным белком в этих случаях нецелесообразна или невозможна вследствие потери интегральными мембранными белками/комплексами в течение способа очистки природной конформации или вследствие недостаточных количеств чистого белка. В предпочтительном варианте осуществления изобретения, млекопитающее, более предпочтительно, кролика, вместо рекомбинантного белок иммунизируют ДНК, например, по протоколу ДНК-вакцинации, как описано в WO/2004/087216. ДНК-вакцинация индуцирует быстрый иммунный ответ на природный антиген. Так как нет необходимости в рекомбинантном белке, эта технология с одной стороны является очень экономически эффективной, с другой стороны, и что более важно, этот способ обеспечивает природную экспрессию интегральных мембранных комплексов и/или трансмембранных белков с несколькими трансмембранными доменами. B-клетки могут быть выделенными у указанного иммунизированного млекопитающего, предпочтительно, указанного кролика, или, альтернативно, являться наивными B-клетками.
На следующем этапе указанного способа из лимфатических органов (таких как селезенка или лимфоузлы) иммунизированных животных, предпочтительно, иммунизированных кроликов, выделяют B-клетки, предпочтительно, B-клетки памяти. B-клетки инкубируют в смеси с клетками, экспрессирующими на своей поверхности антиген, или с флуоресцентно меченным растворимым антигеном. Выделяют B-клетки, которые экспрессируют на своей поверхности специфичные к мишени антитела и, таким образом, связываются с антигеном-мишенью или с антигеном-мишенью, экспрессированным на клеточной поверхности. В более предпочтительном варианте осуществления, B-клетки и/или клетки-мишени окрашивают для обеспечения выделения посредством проточной цитометрии на основе сортировки комплексов B-клетка/клетка-мишень или B-клетка/антиген. Как правило, при проточной цитометрии измеряют флуоресценцию, испускаемую отдельными клетками при пересечении лазерного луча. Однако некоторые исследователи уже используют цитометры для исследования взаимодействий клетка-клетка, например адгезии, опосредованной кадгеринами (Panorchan et al., 2006, J. Cell Science, 119, 66-74; Leong et Hibma, 2005, J. Immunol. Methods, 302, 116-124) или интегринами (Gawaz et al, 1991, J. Clin. Invest, 88, 1128-1134). Таким образом, в случае (ii), клетки, экспрессирующие выбранную мишень окрашивают внутриклеточным флуоресцентным красителем (например, кальцеином). B-клетки окрашивают флуоресцентными антителами, связывающимися с маркерами, специфичными для клеточной поверхности. Таким образом, можно отбирать двухцветные "события", состоящие из двух клеток, соединяющихся друг с другом посредством взаимодействий B-клеточный рецептор-мишень (см. фиг. 2).
Так как IgG, как правило, обладают большей аффинностью, чем IgM, предпочтительно, отбирают положительные B-клетки, экспрессирующие на своей поверхности IgG, а не IgM (что характерно для B-клеток памяти). Для указанной цели предпочтительно, используют многоцветное окрашивание, где антитела, специфичные для IgG и IgM метят различным образом, например, APC и FITC, соответственно.
В одном конкретном варианте осуществления данные для сортировки B-клеток также можно получать по способности этого взаимодействия функционально блокировать/активировать передачу сигнала рецептором. Например, B-клетки можно инкубировать с клетками, которые функционально экспрессируют GPCR (сопряженный с G-белком рецептор). К смеси для индукции опосредованного GPCR выхода Ca2+ из эндоплазматического ретикулума можно добавлять агонист, передающий сигнал через GPCR. В том случае, если антитело, презентированное не B-клетке, функционально блокирует передачу сигнала агонистом, таким образом, этим взаимодействием клетка-клетка также будет блокирован выход Ca2+. Выход Ca2+ можно количественно измерять посредством проточной цитометрии. Таким образом, можно отсортировывать только конгломераты B-клетка/клетка-мишень, которые демонстрируют или увеличение, или снижение выхода Ca2+.
За этапом отбора следует культивирование B-клеток в подходящих условиях так, чтобы в среду для культивирования секретировались антитела. Продуцируемые антитела представляют собой моноклональные антитела. Культивирование может включать использование вспомогательной клеточной линии, такой как вспомогательной клеточной линии тимомы (например, EL4-B5, см. Zubler et al, 1985, J. Immunol., 134(6): 3662-3668). Предпочтительно, проводят этап подтверждения, тестируя полученные антитела на специфическое связывание с мишенью, например, на исключение антител, которые направлены к экспрессируемому на клеточной поверхности белку, отличному от белка-мишени. Например, подходящим для указанной цели является CELISA, т.е. модифицированный твердофазный иммуноферментный анализ (ELISA), где стадию покрытия проводят с применением целых клеток. Указанный способ позволяет оценивать селективность и способность антител конкурировать с лигандом.
Затем полученные на указанном выше этапе антитела анализируют для идентификации CDR указанных антител. Это может включать такие этапы, как очистка антител, определение их аминокислотной последовательности и/или последовательности нуклеиновой кислоты.
В результате CDR можно затем пересаживать в акцепторные каркасы, например, посредством генетического синтеза, способом элонгации олигонуклеотидов, предпочтительно, в акцепторные каркасы, описанные выше. В одном из вариантов осуществления для перенесения донорных CDR в акцепторные каркасы можно использовать общепризнанный в области способ пересадки CDR. В большинстве случаев все три CDR из тяжелой цепи пересаживают из донорного антитела в один акцепторный каркас, а все три CDR из легкой цепи пересаживают в другой акцепторный каркас. Полагают, что пересаживание всех трех CDR необходимо не всегда, так как некоторые CDR могут не участвовать в связывании антигена, а CDR с другими последовательностями (и такой же длиной) могут иметь такую же укладку (и, таким образом, несмотря на различные последовательности, можно сохранять контакты антигена с контактами основной цепи). Фактически антигенсвязывающей активностью могут обладать отдельные домены (Ward et al, 1989, Nature 341, pp.544-546) или даже отдельные CDR (R. Taub et al, 1989, J. Biol Chem 264, pp.259-265). Однако, вне зависимости от того пересаживают ли все или только некоторые из CDR, целью пересадки CDR является пересадка в антитела человека того же или почти того же антигенсвязывающего участка животного (см., например, патент США 5225539 (Winter)).
В другом варианте осуществления CDR донорного антитела до или после их введения в акцепторный каркас можно изменять.
Альтернативно, характеристику антител можно проводить только в их конечном формате иммуносвязывающей молекулы. При этом подходе последовательности CDR антител, экспрессируемых на отсортированных B-клетках, восстанавливают посредством ОТ-ПЦР или из культивируемых отсортированных B-клеток, или непосредственно из единичных отсортированных B-клеток. Для указанной цели размножение B-клеток и/или этап подтверждения, описанные выше, и/или этап анализа, как описано выше, могут быть необязательными. Комбинация двух групп частично перекрывающихся олигонуклеотидов, в которых одна группа олигонуклеотидов кодирует CDR, а вторая группа кодирует каркасные области подходящей иммуносвязывающей поддерживающей молекулы, позволяет получать гуманизированные иммуносвязывающие молекулы способом одноэтапной ПЦР. Высокопроизводительное секвенирование, клонирование и получение позволяют проводить отбор клонов на основе характеристик очищенных гуманизированных иммуносвязывающих молекул, а не характеристики секретируемых в супернатант клеточной культуры IgG. В ее предпочтительном варианте осуществления иммуносвязывающая молекула представляет собой scFv.
Однако пересадка CDR может приводить к ухудшению аффинности получаемой иммуносвязывающей молекулы к антигену вследствие остатков каркаса, которые контактируют с антигеном. Такие взаимодействия могут быть результатом соматического гипермутирования. Таким образом, все еще может требоваться пересадка таких аминокислот из каркаса донора в каркас гуманизированного антитела. Аминокислотные остатки из не являющейся человеческой иммуносвязывающей молекулы, участвующие в связывании антигена, можно идентифицировать, исследуя последовательности и структуры вариабельных областей моноклональных антител кролика. Каждый остаток в каркасе донора CDR, отличающийся от зародышевой линии можно рассматривать как значимый. Если ближайшую зародышевую линию установить невозможно, последовательность можно сравнивать с консенсусом подгруппы или консенсусом последовательностей кролика с высоким процентом сходства. Редкие каркасные остатки рассматривают как возможный результат соматического гипермутирования и, таким образом, как играющие роль в связывании.
Антитела по изобретению можно дополнительно оптимизировать так, чтобы они проявляли улучшенные функциональные свойства, например, увеличенную растворимость и/или стабильность. В определенных вариантах осуществления антитела по изобретению оптимизируют в соответствии с методологией "функционального консенсуса", описанной в заявке PCT с серийным № PCT/EP2008/001958, озаглавленной "Sequence Based Engineering and Optimization of Single Chain Antibodies", поданной 12 марта 2008 года, включенной в настоящий документ в качестве ссылки.
Например, иммуносвязывающие молекулы по изобретению можно сравнивать с базой данный функционально отобранных scFv, используемой для идентификации положений аминокислотных остатков, которые или более, или менее устойчивы к изменению, чем соответствующие положения в иммуносвязывающей молекуле, что, таким образом, означает, что такие идентифицированные положения остатков могут быть подходящими для инженерии с улучшением функциональности, такой как стабильность и/или растворимость.
Характерные положения остатков каркаса для замены и характерные замены в каркасе описаны в заявке PCT № PCT/CH2008/000285, озаглавленной "Methods of Modifying Antibodies, and Modified Antibodies with Improved Functional Properties", поданной 25 июня 2008 года, и заявке PCT № PCT/CH2008/000284, озаглавленной "Sequence Based Engineering and Optimization of Single Chain Antibodies", поданной 25 июня 2008 года. Например, в положениях аминокислот (для каждого из положений аминокислот, перечисленных ниже, указана нумерация AHo) в вариабельной области тяжелой цепи иммуносвязывающей молекулы по изобретению можно проводить одну или несколько из указанных ниже замен:
(a) Q или E в положении аминокислоты 1;
(b) Q или E в положении аминокислоты 6;
(c) T, S или A в положении аминокислоты 7, более предпочтительно, T или A, даже более предпочтительно, T;
(d) A, T, P, V или D, более предпочтительно, T, P, V или D, в положении аминокислоты 10,
(e) L или V, более предпочтительно, L, в положении аминокислоты 12,
(f) V, R, Q, M или K, более предпочтительно, V, R, Q или M в положении аминокислоты 13;
(g) R, M, E, Q или K, более предпочтительно, R, M, E или Q, даже более предпочтительно, R или E, в положении аминокислоты 14;
(h) L или V, более предпочтительно, L, в положении аминокислоты 19;
(i) R, T, K или N, более предпочтительно, R, T или N, даже более предпочтительно, N, в положении аминокислоты 20;
(j) I, F, L или V, более предпочтительно, I, F или L, даже более предпочтительно, I или L, в положении аминокислоты 21;
(k) R или K, более предпочтительно, K, в положении аминокислоты 45;
(l) T, P, V, A или R, более предпочтительно, T, P, V или R, даже более предпочтительно, R, в положении аминокислоты 47;
(m) K, Q, H или E, более предпочтительно, K, H или E, даже более предпочтительно, K, в положении аминокислоты 50;
(n) M или I, более предпочтительно, I, в положении аминокислоты 55;
(o) K или R, более предпочтительно, K, в положении аминокислоты 77;
(p) A, V, L или I, более предпочтительно, A, L или I, даже более предпочтительно, A, в положении аминокислоты 78;
(q) E, R, T или A, более предпочтительно, E, T или A, даже более предпочтительно, E, в положении аминокислоты 82;
(r) T, S, I или L, более предпочтительно, T, S или L, даже более предпочтительно, T, в положении аминокислоты 86;
(s) D, S, N или G, более предпочтительно, D, N или G, даже более предпочтительно, N, в положении аминокислоты 87;
(t) A, V, L или F, более предпочтительно, A, V или F, даже более предпочтительно, V, в положении аминокислоты 89;
(u) F, S, H, D или Y, более предпочтительно, F, S, H или D, в положении аминокислоты 90;
(v) D, Q или E, более предпочтительно, D или Q, даже более предпочтительно, D, в положении аминокислоты 92;
(w) G, N, T или S, более предпочтительно, G, N или T, даже более предпочтительно, G, в положении аминокислоты 95;
(x) T, A, P, F или S, более предпочтительно, T, A, P или F, даже более предпочтительно, F, в положении аминокислоты 98;
(y) R, Q, V, I, M, F, или L, более предпочтительно, R, Q, I, M, F или L, даже более предпочтительно, Y, даже более предпочтительно, L, в положении аминокислоты 103; и
(z) N, S или A, более предпочтительно, N или S, даже более предпочтительно, N, в положении аминокислоты 107.
Дополнительно или альтернативно в вариабельной области легкой цепи иммуносвязывающей молекулы по изобретению можно проводить одну или несколько из перечисленных ниже замен:
(aa) Q, D, L, E, S, или I, более предпочтительно, L, E, S или I, даже более предпочтительно, L или E, в положении аминокислоты 1;
(bb) S, A, Y, I, P или T, более предпочтительно, A, Y, I, P или T, даже более предпочтительно, P или T в положении аминокислоты 2;
(cc) Q, V, T или I, более предпочтительно, V, T или I, даже более предпочтительно, V или T, в положении аминокислоты 3;
(dd) V, L, I или M, более предпочтительно, V или L, в положении аминокислоты 4;
(ee) S, E или P, более предпочтительно, S или E, даже более предпочтительно, S, в положении аминокислоты 7;
(ff) T или I, более предпочтительно, I, в положении аминокислоты 10;
(gg) A или V, более предпочтительно, A, в положении аминокислоты 11;
(hh) S или Y, более предпочтительно, Y, в положении аминокислоты 12;
(ii) T, S или A, более предпочтительно, T или S, даже более предпочтительно, T, в положении аминокислоты 14;
(jj) S или R, более предпочтительно, S, в положении аминокислоты 18;
(kk) T или R, более предпочтительно, R, в положении аминокислоты 20;
(ll) R или Q, более предпочтительно, Q, в положении аминокислоты 24;
(mm) H или Q, более предпочтительно, H, в положении аминокислоты 46;
(nn) K, R или I, более предпочтительно, R или I, даже более предпочтительно, R, в положении аминокислоты 47;
(oo) R, Q, K, E, T, или M, более предпочтительно, Q, K, E, T или M, в положении аминокислоты 50;
(pp) K, T, S, N, Q или P, более предпочтительно, T, S, N, Q или P, в положении аминокислоты 53;
(qq) I или M, более предпочтительно, M, в положении аминокислоты 56;
(rr) H, S, F или Y, более предпочтительно, H, S или F, в положении аминокислоты 57;
(ss) I, V или T, более предпочтительно, V или T, R, даже более предпочтительно, T, в положении аминокислоты 74;
(tt) R, Q или K, более предпочтительно, R или Q, даже более предпочтительно, R, в положении аминокислоты 82;
(uu) L или F, более предпочтительно, F, в положении аминокислоты 91;
(vv) G, D, T или A, более предпочтительно, G, D или T, даже более предпочтительно, T, в положении аминокислоты 92;
(xx) S или N, более предпочтительно, N, в положении аминокислоты 94;
(yy) F, Y или S, более предпочтительно, Y или S, даже более предпочтительно, S, в положении аминокислоты 101; и
(zz) D, F, H, E, L, A, T, V, S, G или I, более предпочтительно, H, E, L, A, T, V, S, G или I, даже более предпочтительно, A или V, в положении аминокислоты 103.
В других вариантах осуществления иммуносвязывающие молекулы по изобретению содержат одну или несколько повышающих стабильность мутаций, описанных в предварительной заявке США с серийным № 61/075692, озаглавленной "Solubility Optimization of Immunobinders", поданной 25 июня 2008 года. В определенных предпочтительных вариантах осуществления иммуносвязывающая молекула содержит повышающую растворимость мутацию в положении аминокислоты, выбранном из группы положений аминокислот тяжелой цепи, состоящей из 12, 103 и 144 (соглашение по нумерации AHo). В одном из предпочтительных вариантов осуществления иммуносвязывающая молекула содержит одну или несколько замен, выбранных из группы, состоящей из: (a) серина (S) в положении аминокислоты тяжелой цепи 12; (b) серина (S) или треонина (T) в положении аминокислоты тяжелой цепи 103 и (c) серина (S) или треонина (T) в положении аминокислоты тяжелой цепи 144. В другом варианте осуществления иммуносвязывающая молекула содержит следующие замены: (a) серин (S) в положении аминокислоты тяжелой цепи 12; (b) серин (S) или треонин (T) в положении аминокислоты тяжелой цепи 103 и (c) серин (S) или треонин (T) в положении аминокислоты тяжелой цепи 144.
В определенных предпочтительных вариантах осуществления иммуносвязывающая молекула содержит повышающие стабильность мутации в остатках каркаса акцепторного каркаса легкой цепи по меньшей мере в одном из положений 1, 3, 4, 10, 47, 57, 91 и 103 вариабельной области легкой цепи в соответствии с системой нумерации AHo. В предпочтительном варианте осуществления акцепторный каркас легкой цепи содержит одну или несколько замен, выбранных из группы, состоящей из (a) глутаминовой кислоты (E) в положении 1, (b) валина (V) в положении 3, (c) лейцина (L) в положении 4; (d) серина (S) в положении 10; (e) аргинина (R) в положении 47; (e) серина (S) в положении 57; (f) фенилаланина (F) в положении 91 и (g) валина (V) в положении 103.
Для получения гуманизированного антитела, содержащего мутацию, как описано выше, можно использовать любой из множества доступных способов.
Таким образом, настоящее изобретение относится к иммуносвязывающей молекуле, гуманизированной способом, описываемым в настоящем документе.
В определенных предпочтительных вариантах осуществления антиген-мишень указанной иммуносвязывающей молекулы представляет собой VEGF или TNFα.
Полипептиды, описанные в настоящем изобретении или полученные способом по настоящему изобретению, можно синтезировать, например, известными в данной области способами. Альтернативно, можно синтезировать молекулы нуклеиновой кислоты, кодирующие желаемые вариабельные области, и полипептиды получать рекомбинантными способами.
Например, после того, как последовательность гуманизированной вариабельной области определена, вариабельную область или полипептид, содержащий ее, можно получать известными в данной области способами молекулярной биологии. Более конкретно, можно использовать способы рекомбинантной ДНК для получения широкого диапазона полипептидов посредством трансформации клетки-хозяина последовательностью нуклеиновой кислоты (например, последовательностью ДНК, кодирующей желаемую вариабельную область (например, модифицированные тяжелую или легкую цепи; их вариабельные домены, или другие их антигенсвязывающие фрагменты)).
В одном из вариантов осуществления можно получить вектор экспрессии, содержащий промотор, функционально связанный с последовательностью ДНК, кодирующей по меньшей мере VH или VL. Если необходимо или желательно, можно получить второй вектор экспрессии, содержащий промотор, который функционально связан с последовательностью ДНК, кодирующей комплементарный вариабельный домен (т.е., когда исходный вектор экспрессии кодирует VH, второй вектор экспрессии кодирует VL и наоборот). Затем одним или обоими векторами экспрессии можно трансформировать клеточную линию (например, иммортализованную клеточную линию млекопитающего) и культивировать в условиях, обеспечивающих экспрессию химерного вариабельного домена или химерного антитела (см., например, международную патентную заявку № PCT/GB85/00392 Neuberger et. al.
В одном из вариантов осуществления можно получить вариабельные области, содержащие аминокислотные последовательности CDR донора и FR акцептора, а затем для осуществления замен аминокислот CDR вносить изменения в молекулы нуклеиновой кислоты.
Характерные общепризнанные в данной области способы получения молекулы нуклеиновой кислоты, кодирующей вариант аминокислотной последовательности полипептида, включают в качестве неограничивающих примеров, получение посредством сайт-специфического (или опосредованного олигонуклеотидами) мутагенеза, ПЦР-мутагенеза и кассетного мутагенеза ранее полученной ДНК, кодирующей полипептид.
Сайт-специфический мутагенез представляет собой предпочтительный способ получения вариантов с заменами. Этот способ хорошо известен в данной области (см., например, Carter et al., Nucleic Acids Res. 13:4431-4443 (1985) и Kunkel et al., Proc. Natl. Acad. Sci. USA 82:488 (1987)). В кратком изложении, при проведении сайт-специфического мутагенеза ДНК исходную ДНК изменяют посредством первой гибридизации олигонуклеотида, кодирующего желаемую мутацию, с одиночной цепью такой исходной ДНК. После гибридизации используют ДНК-полимеразу для синтеза целой второй цепи с использованием гибридизующегося олигонуклеотида в качестве праймера и с использованием одиночной цепи исходной ДНК в качестве матрицы. Таким образом, в получаемую двухцепочечную ДНК вводят олигонуклеотид, кодирующий желаемую мутацию.
Также для получения вариантов аминокислотной последовательности полипептидов подходит ПЦР-мутагенез. См. Higuchi, в PCR Protocols, pp. 177-183 (Academic Press, 1990); и Vallette et al., Nuc. Acids Res. 17:723-733 (1989). В кратком изложении, когда в качестве исходного вещества для ПЦР используют небольшие количества матричной ДНК, можно использовать праймеры, которые немного отличаются по последовательности от соответствующей области в матричной ДНК с получением относительно больших количеств конкретного фрагмента ДНК, который отличается от матричной последовательности только по положениям, где праймеры отличаются от матрицы.
Другой способ получения вариантов, кассетный мутагенез, основан на способе, описанном Wells et al., Gene 34:315-323 (1985). Исходным материалом является плазмида (или другой вектор), содержащая ДНК для мутирования. Кодон(ы) в исходной ДНК для мутирования являются идентифицированными. С каждой стороны от идентифицированного участка(ов) мутирования должен находиться уникальный участок узнавания рестрикционной эндонуклеазы. Если таких участков рестрикции не существует, их можно получать с использованием описанного выше мутагенеза, опосредованного олигонуклеотидами, с введением их в подходящие положения кодирующей полипептид ДНК. Плазмидную ДНК разрезают по этим участкам для ее линеаризации. Стандартными способами синтезируют двухцепочечный олигонуклеотид, кодирующий последовательность ДНК между участками рестрикции, но содержащий желаемую мутацию(и), где две цепи олигонуклеотида синтезируют раздельно, а затем стандартными способами гибридизуют вместе. Этот двухцепочечный олигонуклеотид называют кассетой. Эту кассету конструируют так, чтобы она имела 5'- и 3'-концы, которые совместимы с концами линеаризованной плазмиды так, что его можно было непосредственно лигировать в плазмиду. Теперь эта плазмида содержит мутантную последовательность ДНК.
Вариабельную область, полученную способами по изобретению можно перестраивать и дополнительно изменять с дальнейшим усилением связывания антигена. Таким образом, перед описанными выше этапами или после них можно проводить этапы, включающие, например созревание аффинности. Кроме того, для дальнейшей оптимизации можно использовать эмпирические данные о связывании.
Специалисту в данной области должно быть понятно, что полипептиды по изобретению можно дополнительно модифицировать так, чтобы они отличались по аминокислотной последовательности, но при этом сохраняли желаемую активность. Например, для белка можно проводить дополнительные замены нуклеотидов, приводящие к заменам аминокислот в "несущественных" аминокислотных остатках. Например, несущественный аминокислотный остаток в полипептиде иммуноглобулина можно заменять на другой аминокислотный остаток из того же семейства боковых цепей. В другом варианте осуществления можно заменять участок аминокислот структурно сходным участком, который отличается порядком и/или составом представителей семейства боковых цепей, т.е., можно проводить консервативную замену, при которой аминокислотный остаток заменяют аминокислотным остатком со сходной боковой цепью.
В данной области определены семейства аминокислотных остатков со сходными боковыми цепями, включая основные боковые цепи (например, лизин, аргинин, гистидин), кислые боковые цепи (например, аспарагиновая кислота, глутаминовая кислота), незаряженные полярные боковые цепи (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин), неполярные боковые цепи (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), разветвленные по бета-положению боковые цепи (например, треонин, валин, изолейцин) и ароматические боковые цепи (например, тирозин, фенилаланин, триптофан, гистидин).
Кроме замен аминокислот, по настоящему изобретению предусмотрены другие модификации, например, в аминокислотных последовательностях Fc-области для получения варианта Fc-области с измененной эффекторной функцией. Например, можно удалять один или несколько аминокислотных остатков Fc-области для ослабления или усиления связывания с FcR. В одном из вариантов осуществления для получения такого варианта Fc-области можно модифицировать один или несколько остатков Fc-области. Как правило, в соответствии с этим вариантом осуществления изобретения удаляют не более чем от одного до приблизительно десяти остатков Fc-области. Fc-область по настоящему документу, содержащая одну или несколько делеций аминокислот предпочтительно, сохраняет по меньшей мере приблизительно 80%, а предпочтительно, по меньшей мере приблизительно 90%, а наиболее предпочтительно, по меньшей мере приблизительно 95% исходной Fc-область или природной последовательности Fc-области человека.
Также можно получать варианты Fc-областей с инсерциями аминокислот, где у этих вариантов изменена эффекторная функция. Например, рядом с одним или несколькими положениями в Fc-области, идентифицированными в настоящем документе, как влияющие на связывание FcR, можно вводить по меньшей мере один аминокислотный остаток (например, от одного до двух аминокислотных остатков и, как правило, не более десяти остатков). Под "рядом" подразумевают в пределах от одного до двух аминокислотных остатков от остатка Fc-области, идентифицированного в настоящем документе. Такие варианты Fc-областей могут демонстрировать увеличенное или уменьшенное связывание FcRn.
Такие варианты Fc-области, как правило, содержать по меньшей мере одну модифицированную аминокислоту в Fc-области. В одном из вариантов осуществления модификации аминокислот можно комбинировать. Например, вариант Fc-области может содержать два, три, четыре, пять и т.д. замен, например, в конкретных положениях в Fc-области, идентифицированных в настоящем документе. В другом варианте осуществления полипептид может обладать измененным связыванием с FcRn и с другим рецептором Fc.
В одном из вариантов осуществления полипептиды, описанные в настоящем изобретении или полученные способом по настоящему изобретению, например, гуманизированные вариабельные области Ig и/или полипептиды, содержащие гуманизированные вариабельные области Ig, можно получать рекомбинантными способами. Например, полинуклеотидную последовательность, кодирующую полипептид, можно вставлять в подходящий вектор экспрессии для рекомбинантной экспрессии. Когда полипептид представляет собой антитело, полинуклеотиды, кодирующие дополнительные вариабельные области легких и тяжелых цепей, необязательно соединенные с константными областями, можно вставлять в тот же или другой вектор экспрессии. Для облегчения последующей очистки к полипептидной последовательности необязательно можно присоединять или в полипептидную последовательность необязательно можно включать последовательность аффинной метки (например, метки His(6)). Участки ДНК, кодирующие цепи иммуноглобулинов функционально связаны с контролирующими последовательностями в векторе(ах) экспрессии, которые обеспечивают экспрессию иммуноглобулиновых полипептидов. Контролирующие экспрессию последовательности включают в качестве неограничивающих примеров промоторы (например, ассоциированные в природе или гетерологичные промоторы), сигнальные последовательности, энхансерные элементы и последовательности терминации транскрипции. Предпочтительно, контролирующие экспрессию последовательности представляют собой эукариотические промоторные системы в векторах, способных к трансформации или трансфекции эукариотических клеток-хозяина. После введения вектора в подходящего хозяина, хозяина содержат в условиях, подходящих для высокого уровня экспрессии нуклеотидных последовательностей и сборки и очистки полипептида.
Эти векторы экспрессии, как правило, реплицируются в организмах-хозяевах в виде эписом или в виде интегральной части хромосомной ДНК хозяина. Как правило, векторы экспрессии содержат селективные маркеры (например, устойчивости к ампициллину, устойчивости к гигромицину, устойчивости к тетрациклину или устойчивости к неомицину) для обеспечения детекции клеток, трансформированных желаемыми последовательностями ДНК (см., например, патент США № 4704362).
Одним из особенно подходящих прокариотических хозяев для клонирования полинуклеотидов (например, последовательностей ДНК) по настоящему изобретению является E. coli. Другие микробные хозяева, подходящие для использования, включают бациллы, такие как Bacillus subtilus, и другие Enterobacteriaceae, такие как Salmonella, Serratia и различные виды Pseudomonas.
Также для экспрессии подходят другие микробы, такие как дрожжи. Примерами дрожжевых хозяев являются Saccharomyces и Pichia, с соответствующими векторами с контролирующими экспрессию последовательностями (например, промоторами), участками начала репликации, терминации последовательности и т.п., как желательно. Обычные промоторы включают промоторы 3-фосфоглицераткиназы и других ферментов гликолиза. Индуцибельные дрожжевые промоторы включают среди прочих промоторы алкогольдегидрогеназы, изоцитохрома C и ферментов, ответственных за утилизацию метанола, мальтозы и галактозы.
В объеме настоящего изобретения предпочтительными клетками-хозяевами являются E. coli и S. cerevisiae.
В дополнение к микроорганизмам для экспрессии и получения полипептидов по настоящему изобретению, (например, полинуклеотидов, кодирующих иммуноглобулины или их фрагменты) можно использовать тканевую культуру млекопитающих. См. Winnacker, From Genes to Clones, VCH Publishers, N.Y., N.Y. (1987). Фактически, эукариотические клетки являются предпочтительными, поскольку в данной области разработан ряд подходящих линий клеток-хозяев, способных к секреции гетерологичных белков (например, интактных иммуноглобулинов), и он включает клеточные линии CHO, различные клеточные линии Cos, клетки HeLa, клетки 293, миеломные клеточные линии, трансформированные B-клетки и гибридомы. Векторы экспрессии для этих клеток могут включать контролирующие экспрессию последовательности, такие как участок начала репликации, промотор и энхансер (Queen et al., Immunol. Rev. 89:49 (1986)), и необходимые для процессинга информационные участки, такие как участки связывания рибосом, участки сплайсинга РНК, участки полиаденилирования и последовательности терминации транскрипции. Предпочтительными контролирующими экспрессию последовательностями являются промоторы, получаемые из генов иммуноглобулинов, SV40, аденовирусов, вируса папилломы крупного рогатого скота, цитомегаловируса и т.п. См. Co et al., J. Immunol 148:1149 (1992).
Векторы, содержащие представляющие интерес полинуклеотидные последовательности (например, последовательности, кодирующие тяжелые и легкие цепи, и контролирующие экспрессию последовательности) можно вносить в клетку-хозяина хорошо известными способами, которые варьируют в зависимости от типа клетки-хозяина. Например, для прокариотических клеток, как правило, используют трансфекцию с хлоридом кальция, тогда как для других клеток-хозяев можно использовать обработку фосфатом кальция, электропорацию, липофекцию, биолистическую или основанную на вирусах трансфекцию. (в основном, см. Sambrook et al., Molecular Cloning: A Laboratory Manual (Cold Spring Harbor Press, 2nd ed., 1989). Другие способы, используемые для трансформации клеток млекопитающих, включают использование полибрена, слияния протопластов, липосом, электропорации и микроинъекции (в основном, см. Sambrook et al., выше). Для получения трансгенных животных трансгены посредством микроинъекции можно вводить в оплодотворенные яйцеклетки или можно вводить в геном эмбриональных стволовых клеток, и ядра таких клеток пересаживать в лишенные ядер яйцеклетки.
Рассматриваемый полипептид также можно встраивать в трансгены для введения в геном трансгенного животного и последующей экспрессии, например, в молоке трансгенного животного (см., например, Deboer et al. 5741957; Rosen 5304489 и Meade 5849992. Подходящие трансгены включают кодирующие последовательности легких и/или тяжелых цепей в функциональной связи с промотором и энхансером специфичного для молочной железы гена, такого как казеин или бета-лактоглобулин.
Полипептиды можно экспрессировать с использованием одного вектора или двух векторов. Например, тяжелые и легкие цепи антитела можно клонировать в раздельных векторах экспрессии и совместно трансфицировать в клетки.
В одном из вариантов осуществления для облегчения экспрессии полипептидов по изобретению можно использовать сигнальные последовательности.
После экспрессии полипептиды можно очищать стандартными для данной области способами, включающими осаждение сульфатом аммония, аффинные колонки (например, с белком A или белком G), колоночную хроматографию, очистку ВЭЖХ, электрофорез в геле и т.п. (в основном, см. Scopes, Protein Purification (Springer-Verlag, N.Y., (1982)).
Гуманизированные вариабельные области Ig или содержащие их полипептиды можно экспрессировать в клетках-хозяевах или в клеточных линиях в культуре. Их также можно экспрессировать в клетках in vivo. Клеточная линия, которая трансформирована (например, трансфицирована) для продукции измененного антитела, может представлять собой иммортализованную клеточную линию млекопитающего, такую как клеточные линии лимфоидного происхождения (например, миеломные, гибридомные, триомные или квадромные клеточные линии). Клеточная линия также может включать нормальные лимфоидные клетки, такие как B-клетки, иммортализованные посредством трансформации вирусом (например, вирусом Эпштейна-Барр).
Хотя, как правило, клеточная линия, используемая для продукции полипептида, представляет собой клеточную линию млекопитающего, также можно использовать клеточные линии из других источников (таких как бактерии и дрожжи). В частности, можно использовать бактериальные штаммы, полученные из E. coli, в особенности, например, фаговый дисплей.
Некоторые иммортализованные лимфоидные клеточные линии, такие как миеломные клеточные линии, в их нормальном состоянии секретируют отдельные легкие или тяжелые цепи Ig. Если такая клеточная линия трансформирована вектором, экспрессирующим измененное антитело, полученное способом по изобретению, оставшиеся этапы способа проводить не обязательно, при условии, что секретируемая в норме цепь комплементарна вариабельному домену цепи Ig, кодируемой полученным ранее вектором.
Если иммортализованная клеточная линия не секретирует или не секретирует комплементарную цепь, необходимо ввести в клетки вектор, кодирующий соответствующую комплементарную цепь или ее фрагмент.
В том случае, когда иммортализованная клеточная линия секретирует комплементарную легкую или тяжелую цепь, линию трансформированных клеток можно получать, например, трансформируя подходящую бактериальную клетку вектором, а затем сливая бактериальную клетку с иммортализованной клеточной линией (например, посредством слияния сферопластов). Альтернативно, ДНК можно вводить в иммортализованную клеточную линию напрямую посредством электропорации.
В одном из вариантов осуществления гуманизированная вариабельная область Ig, как описано в настоящем изобретении или полученная способом по настоящему изобретению, может присутствовать в антигенсвязывающем фрагменте любого антитела. Фрагменты можно получать рекомбинантно и конструировать, синтезировать или получать посредством расщепления антитела протеолитическим ферментом. Например, фрагмент может представлять собой Fab-фрагмент; расщепление папаином разделяет антитело в области перед межцепочечной (т.е., VH-VH) дисульфидной связью, которая соединяет две тяжелые цепи. Это приводит к формированию двух идентичных фрагментов, содержащих легкую цепь и домены VH и CH1 тяжелой цепи. Альтернативно, фрагмент может представлять собой фрагмент F(ab')2. Эти фрагменты можно получать, расщепляя антитело пепсином, который расщепляет тяжелую цепь после межцепочечной дисульфидной связи и приводит к образованию фрагмента, содержащего оба антигенсвязывающих участка. Еще одной альтернативой является использование "одноцепочечного" антитела. Одноцепочечные фрагменты Fv (scFv) можно получать множеством способов. Например, C-конец VH можно присоединять к N-кону VL. Как правило, между VH и VL помещают линкер (например, (GGGGS)4). Однако порядок, в котором можно соединять цепи можно изменять на обратный, и можно включать метки, которые облегчают детекцию или очистку (например, метки Myc, His или FLAG-) (метки, такие как эти, можно присоединять к любому антителу или фрагменту антитела по изобретению; их использование не ограничено scFv). Таким образом, и как отмечено ниже, меченые антитела входят в объем настоящего изобретения. В альтернативных вариантах осуществления антитела, описываемые в настоящем документе, или полученные способами, описываемыми в настоящем документе, могут представлять собой димеры тяжелых цепей или димеры легких цепей. Кроме того, можно использовать легкую или тяжелую цепь антитела, или их части, например, однодоменные антитела (DAb).
В другом варианте осуществления гуманизированная вариабельная область Ig, как описано в настоящем изобретении или полученная способом по настоящему изобретению присутствует в одноцепочечном антителе (scFv) или минителе (см. например, патент США № 5837821 или WO 94/09817A1). Минитела представляют собой димерные молекулы, полученные из двух полипептидных цепей, каждая из которых содержит молекулу scFv (отдельный полипептид, содержащий один или несколько антигенсвязывающих участков, например, домен VL, соединенный подвижным линкером с доменом VH, слитым с доменом CH3 посредством соединительного пептида). Молекулы scFv можно конструировать в ориентации VH-линкер-VL или в ориентации VL-линкер-VH. Подвижный шарнир, который соединяет домены VL и VH, образующие антигенсвязывающий участок, предпочтительно, содержит приблизительно от 10 до приблизительно 50 аминокислотных остатков. Иллюстративный соединительный пептид для этой цели представляет собой (Gly4Ser)3 (Huston et al., (1988). PNAS, 85:5879). В данной области известны другие соединительные пептиды.
Способы получения одноцепочечных антител хорошо известны в данной области, например, Ho et al., (1989), Gene, 77:51; Bird et al., (1988), Science 242:423; Pantoliano et al., (1991), Biochemistry 30:10117; Milenic et al., (1991), Cancer Research, 51:6363; Takkinen et al., (1991), Protein Engineering 4:837. Минитела можно получать, конструируя компонент scFv и присоединяя компонент пептид-CH3 способами, описанными в данной области (см., например, патент США 5837821 или WO 94/09817A1). Эти компоненты можно выделять из отдельных плазмид в качестве рестрикционных фрагментов, а затем лигировать и повторно клонировать в подходящий вектор. Соответствующую конструкцию можно проверять посредством рестрикционного расщепления и анализа последовательности ДНК. В одном из вариантов осуществления минитело по изобретению содержит соединительный пептид. В одном из вариантов осуществления соединительный пептид содержит линкер Gly/Ser, например, GGGSSGGGSGG.
В другом варианте осуществления можно конструировать четырехвалентное минитело. Четырехвалентные минитела можно конструировать таким же способом, как и минитела, за исключением того, что с применением подвижного линкера, например, с аминокислотной последовательностью (G4S)4G3AS, связывают две молекулы scFv.
В другом варианте осуществления гуманизированная вариабельная область, как описано в настоящем изобретении или полученная способом по настоящему изобретению, может присутствовать в диателе. Диатела подобны молекулам scFv, но, как правило, содержат короткий (менее 10, а предпочтительно, 1-5) линкер из аминокислотных остатков, соединяющий оба вариабельных домена так, что домены VL и VH на одной полипептидной цепи не могут взаимодействовать. Вместо этого, домены VL и VH одной полипептидной цепи взаимодействуют с доменами VH и VL (соответственно) на второй полипептидной цепи (WO 02/02781).
В другом варианте осуществления гуманизированная вариабельная область по изобретению может находиться в иммунореактивных фрагменте или части антитела (например, молекулы scFv, минитела, четырехвалентного минитела или диатела), функционально связанных с частью, связывающей FcR. В иллюстративном варианте осуществления, связывающая FcR часть представляет собой целую Fc-область.
Предпочтительно, способы гуманизации, описываемые в настоящем документе приводят к получению вариабельных областей Ig, у которых аффинность к антигену по сравнению с донорным антителом по существу не изменена.
В одном из вариантов осуществления полипептиды, содержащие вариабельные домены по настоящему изобретению связываются с антигенами с аффинностью связывания большей (или равной) аффинности с константой ассоциации Ka равной приблизительно 105 М-1, 106 М-1, 107 М-1, 108 М-1, 109 М-1, 1010 М-1, 1011 М-1 или 1012 М-1, (включая аффинности между этими значениями).
Аффинность, авидность и/или специфичность можно измерять множеством способов. Как правило, и независимо от определенного способа, которым определяют или измеряют аффинность, способы по изобретению, когда ими получают антитело, которое в любом аспекте его клинического применения превосходит антитело (или антитела) из которого его получали (например, способы по изобретению рассматривают как эффективные или успешные, когда модифицированное антитело можно вводить с меньшей дозой или менее часто или более удобным способом введения, чем антитело (или антитела) из которого его получали) улучшают аффинность антитела.
Более конкретно, аффинность антитела к антигену, с которой оно связывается, можно измерять различными анализами, включая, например, анализ ELISA, анализ BiaCore или анализ KinExA™ 3000 (доступный в Sapidyne Instruments (Boise, ID)). В кратком изложении, гранулы сефарозы покрывают антигеном (антиген, используемый в способах по изобретению, может представлять собой любой представляющий интерес антиген (например, антиген злокачественной опухоли; белок клеточной поверхности или секретируемый белок; антиген патогенного организма (например, бактериальный или вирусный антиген (например, антиген ВИЧ, антиген гриппа или антиген гепатита)) или аллерген) посредством ковалентного связывания. Получают разведения антител для тестирования и каждое разведение добавляют в определенные лунки на планшете. Затем в каждую лунку добавляют детектирующее антитело (например, конъюгат антител козы к IgG человека с HRP) с последующим добавлением хромогенного субстрата (например, HRP). Затем планшет считывают в спектрофотометре ELISA для чтения планшетов при 450 нМ, и рассчитывают значения EC50. (однако следует понимать, что описанные в настоящем документе способы применимы вообще; они не ограничены получением антител, которые связывают любой конкретный антиген или класс антигенов.)
Специалистам в данной области понятно, что определение аффинности не всегда является настолько простым, как выглядит в простейшем случае. Так как антитела имеют два плеча, их видимая аффинность, как правило, намного выше, чем собственная аффинность вариабельной области к антигену (полагают, что это происходит вследствие авидности). Собственную аффинность можно измерять с использованием фрагментов scFv или Fab.
В другом аспекте настоящее изобретение относится к биспецифическим молекулам, содержащим гуманизированное антитело кролика или его фрагмент по изобретению. С антителом по изобретению или с его антигенсвязывающими частями можно образовывать производные или их можно соединять с другой функциональной молекулой, например, другим пептидом или белком (например, другое антитело или лиганд рецептора) с получением биспецифической молекулы, которая связывает по меньшей мере два различных участка связывания или две молекулы-мишени. С антителом по изобретению можно образовывать производные или его можно соединять более чем с одной другой функциональной молекулой с получением полиспецифических молекул, которые связывают более двух различных участков связывания и/или молекул-мишеней; такие полиспецифичные молекулы также предназначены для включения в термин "биспецифическая молекула" как используют в настоящем документе. Для получения биспецифической молекулы по изобретению антитело по изобретению можно функционально связывать (например, посредством химического связывания, генетического слияния, нековалентного связывания или иным образом) с одной или несколькими другими связывающими молекулами, такими как другое антитело, фрагмент антитела, опухолеспецифичные или патогенспецифичные антигены, пептидомиметик или имитатор связывания, так, что образуется биспецифическая молекула. Таким образом, настоящее изобретение относится к биспецифическим молекулам, содержащим по меньшей мере одну первую связывающую молекулу со специфичностью к первой мишени и вторую связывающую молекулу со специфичностью к одной или несколькими дополнительным эпитопам-мишеням.
В одном из вариантов осуществления биспецифические молекулы по изобретению содержат в качестве специфически связывающейся молекулы по меньшей мере одно антитело, или его фрагмент, включая, например, Fab, Fab', F(ab')2, Fv или одноцепочечный Fv. Антитело также может представлять собой димер легких цепей или тяжелых цепей, или его минимальный фрагмент, такой как Fv или одноцепочечная конструкция, как описано в Ladner et al., патенте США № 4946778, содержимое которых явно включено в качестве ссылки.
Хотя предпочтительными являются моноклональные антитела человека, другие антитела, которые можно использовать в биспецифических молекулах по изобретению представляют собой моноклональные антитела мыши, химерные и гуманизированные моноклональные антитела.
Биспецифические молекулы по настоящему изобретению можно получать, конъюгируя составляющие специфически связывающиеся молекулы известными в данной области способы. Например, каждую специфически связывающуюся молекулу биспецифической молекулы можно получать раздельно, а затем конъюгировать друг с другом. Когда специфически связывающиеся молекулы представляют собой белки или пептиды, для ковалентной конъюгации можно использовать ряд связывающих или сшивающих средств. Примеры сшивающих средств включают белок A, карбодиимид, N-сукцинимидил-S-ацетилтиоацетат (SATA), 5,5'-дитиобис(2-нитробензойную кислоту) (DTNB), о-фенилендималеимид (oPDM), N-сукцинимидил-3-(2-пиридилдитио)пропионат (SPDP) и сульфосукцинимидил-4-(N-малеимидометил)циклогексан-1-карбоксилат (сульфо-SMCC) (см. например, Karpovsky et al., (1984) J. Exp. Med. 160:1686; Liu, MA et al., (1985) Proc. Natl. Acad. Sci. USA 82:8648). Другие способы включают способы, описанные в Paulus (1985) Behring Ins. Mitt. No. 78, 118-132; Brennan et al., (1985) Science 229:81-83) и Glennie et al., (1987) J. Immunol. 139: 2367-2375). Предпочтительные конъюгирующие средства представляют собой SATA и сульфо-SMCC, оба доступные в Pierce Chemical Co. (Rockford, IL).
Когда специфически связывающиеся молекулы представляют собой антитела, их можно конъюгировать посредством образования сульфгидрильных связей между C-концевыми шарнирными областями двух тяжелых цепей. В особенно предпочтительном варианте осуществления шарнирную область модифицируют так, чтобы она до конъюгации содержала нечетное количество сульфгидрильных остатков, предпочтительно, один.
Альтернативно, обе специфически связывающиеся молекулы можно кодировать на одном векторе и экспрессировать и собирать в одной клетке-хозяине. Этот способ особенно пригоден, когда биспецифическая молекула представляет собой слитый белок mAb×mAb, mAb×Fab, Fab×F(ab')2 или лиганд×Fab. Биспецифическая молекула по изобретению может представлять собой одноцепочечную молекулу, содержащую одно одноцепочечное антитело и одну связывающую детерминанту, или одноцепочечную биспецифическую молекулу, содержащую две связывающих детерминанты. Биспецифические молекулы могут содержать по меньшей мере две одноцепочечных молекулы. Способы получения биспецифических молекул описаны, например, в патенте США № 5260203; патенте США № 5455030; патенте США № 4881175; патенте США № 5132405; патенте США № 5091513; патенте США № 5476786; патенте США № 5013653; патенте США № 5258498 и в патенте США № 5482858.
Связывание биспецифических молекул с их специфическими мишенями можно подтверждать, например, твердофазным иммуноферментным анализом (ELISA), радиоиммунологическим анализом (RIA), проточной цитометрией, на основе сортировки единичных клеток (например, анализ FACS), биологическим анализом (например, ингибирование роста), или анализом вестерн-блоттинг. В каждом из этих анализов, как правило, определяют присутствие комплексов белок-антитело, представляющих конкретный интерес, используя метящий реагент (например, антитело), специфичный к представляющему интерес комплексу. Например, комплексы антител можно детектировать с применением, например, связанного с ферментом антитела или фрагмента антитела, которые распознают и специфически связываются с комплексами антитело-VEGF. Альтернативно, комплексы можно детектировать с применением любого из множества других иммунологических анализов. Например, антитело можно радиоактивно метить и использовать в радиоиммунологическом анализе (RIA) (см., например, Weintraub, B., Principles of Radioimmunoassays, Seventh Training Course on Radioligand Assay Techniques, The Endocrine Society, March, 1986, включенную в настоящий документ в качестве ссылки). Радиоактивный изотоп можно детектировать такими способами, как применение γ-счетчика или сцинтилляционного счетчика или посредством авторадиографии.
В другом аспекте настоящее изобретение относится к гуманизированному антителу кролика или его фрагменту, конъюгированным с терапевтической молекулой, такой как цитотоксин, лекарственное средство (например, иммуносупрессор) или радиотоксин. Такие конъюгаты обозначают в настоящем документе как "иммуноконъюгаты". Иммуноконъюгаты, которые включают один или несколько цитотоксинов обозначают как "иммунотоксины". Цитотоксин или цитотоксическое средство включает любое средство, которое причиняет вред (например, убивает) клеткам. Примеры включают таксол, цитохалазин B, грамицидин D, бромистый этидий, эметин, митомицин, этопозид, тенопозид, винкристин, винбластин, колхицин, доксорубицин, даунорубицин, дигидрокси антрациндион, митоксантрон, митрамицин, актиномицин D, 1-дегидротестостерон, глюкокортикоиды, прокаин, тетракаин, лидокаин, пропранолол и пуромицин и их аналоги или гомологи. Терапевтические средства также могут включать, например, антиметаболиты (например, метотрексат, 6-меркаптопурин, 6-тиогуанин, цитарабин, 5-фторурацил декарбазин), алкилирующие средства (например, мехлоретамин, тиотепа, хлорамбуцил, мелфалан, кармустин (BSNU) и ломустин (CCNU), циклофосфамид, бусульфан, дибромманнит, стрептозотоцин, митомицин C, и цис-дихлордиамин платины (II) (DDP) цисплатин), антрациклины (например, даунорубицин (ранее дауномицин) и доксорубицин), антибиотики (например, дактиномицин (ранее актиномицин), блеомицин, митрамицин и антрамицин (AMC)) и антимитотические средства (например, винкристин и винбластин).
Другие предпочтительные примеры терапевтических цитотоксинов, которые можно конъюгировать с антителом по изобретению, включают дуокармицины, калихеамицины, майтанзины и ауристатины и их производные. Пример конъюгата калихеамицина с антителом является коммерчески доступным (милотарг™; Wyeth-Ayerst).
Цитотоксины можно конъюгировать с антителами по изобретению с применением технологии линкеров, доступной в данной области. Примеры типов линкеров, которые использовали для конъюгации цитотоксина с антителом, включают в качестве неограничивающих примеров гидразоны, простые тиоэфиры, сложные эфиры, дисульфиды и содержащие пептиды линкеры. Можно выбирать линкер, который, например, чувствителен к расщеплению при низком pH в условиях лизосомального компартмента или чувствителен к расщеплению протеазами, такими как протеазы, преимущественно экспрессируемые в опухолевой ткани, такие как катепсины (например, катепсины B, C, D).
Для дополнительных сведений о типах цитотоксинов, линкерах и способах конъюгирования терапевтических средств с антителами, также см. Saito, G. et al., (2003) Adv. Drug Deliv. Rev. 55:199-215; Trail, P.A. et al., (2003) Cancer Immunol. Immunother. 52:328-337; Payne, G. (2003) Cancer Cell 3:207-212; Allen, T.M. (2002) Nat. Rev. Cancer 2:750-763; Pastan, I. and Kreitman, R. J. (2002) Curr. Opin. Investig. Drugs 3:1089-1091; Senter, P.D. and Springer, C.J. (2001) Adv. Drug Deliv. Rev. 53:247-264.
Антитела по настоящему изобретению также можно конъюгировать с радиоактивным изотопом с получением цитотоксических радиофармацевтических средств, также обозначаемых как радиоиммуноконъюгаты. Примеры радиоактивных изотопов, которые можно конъюгировать с антителами для диагностического или терапевтического применения включают в качестве неограничивающих примеров йод131, индий111, иттрий90 и лютеций177. Способ получения радиоиммуноконъюгатов широко известен в данной области. Примеры радиоиммуноконъюгатов коммерчески доступны, включая зевалин™ (IDEC Pharmaceuticals) и бексар™ (Corixa Pharmaceuticals), и для получения радиоиммуноконъюгатов с применением антител по изобретению можно использовать сходные способы.
Конъюгаты антител по изобретению можно использовать для модификации имеющегося биологического ответа, и молекулу лекарственного средства не следует рассматривать как ограниченную классическими химическими терапевтическими средствами. Например, молекула лекарственного средства может представлять собой белок или полипептид, обладающие желаемой биологической активностью. Такие белки могут включать, например, ферментативно активный токсин или его активный фрагмент, такой как абрин, рицин A, экзотоксин Pseudomonas или дифтерийный токсин; белок, такой как фактор некроза опухоли или интерферон-γ; или модификаторы биологического ответа, такие как, например, лимфокины, интерлейкин-1 ("IL-1), интерлейкин-2 ("IL-2"), интерлейкин-6 ("IL-6"), гранулоцитарно-макрофагальный колониестимулирующий фактор ("GM-CSF"), гранулоцитарный колониестимулирующий фактор ("G-CSF") или другие факторы роста.
Способы конъюгации таких терапевтических молекул с антителами хорошо известны, см., например, Arnon et al., "Monoclonal Antibodies For Immunotargeting Of Drugs In Cancer Therapy", in Monoclonal Antibodies And Cancer Therapy, Reisfeld et al., (eds.), pp. 243-56 (Alan R. Liss, Inc. 1985); Hellstrom et al., "Antibodies For Drug Delivery", in Controlled Drug Delivery (2nd Ed.), Robinson et al., (eds.), pp. 623-53 (Marcel Dekker, Inc. 1987); Thorpe, "Antibody Carriers Of Cytotoxic Agents In Cancer Therapy: A Review", в Monoclonal Antibodies '84: Biological And Clinical Applications, Pinchera et al., (eds.), pp. 475-506 (1985); "Analysis, Results, And Future Prospective Of The Therapeutic Use Of Radiolabeled Antibody In Cancer Therapy", в Monoclonal Antibodies For Cancer Detection And Therapy, Baldwin et al., (eds.), pp. 303-16 (Academic Press 1985), и Thorpe et al., "The Preparation And Cytotoxic Properties Of Antibody-Toxin Conjugates", Immunol. Rev., 62:119-58 (1982).
В одном из аспектов изобретение относится к фармацевтическим составам, содержащим гуманизированные антитела кролика для лечения заболевания. Термин "фармацевтический состав" относится к препаратам, которые находятся в такой форме, которая допускает определенную эффективную биологическую активность антитела или производного антитела, и которые не содержат дополнительных компонентов, которые токсичны для индивидуумов, которым вводят этот состав. "Фармацевтически приемлемые" эксципиенты (носители, добавки) представляют собой эксципиенты, которые можно в разумных пределах вводить рассматриваемому млекопитающему для обеспечения эффективной дозы используемого активного ингредиента.
"Стабильный" состав представляет собой состав, в котором антитело или производное антитела по существу сохраняет свою физическую стабильность, и/или химическую стабильность, и/или биологическую активность при хранении. В данной области доступны и рассмотрены, например, в Peptide and Protein Drug Delivery, 247-301, Vincent Lee Ed., Marcel Dekker, Inc., New York, N.Y., Pubs. (1991) и Jones, A. Adv. Drug Delivery Rev. 10: 29-90 (1993) различные аналитические способы измерения стабильности белка. Стабильность можно измерять при выбранной температуре в течение выбранного периода времени. Предпочтительно, состав стабилен при комнатной температуре (приблизительно 30°C) или при 40°C в течение по меньшей мере 1 месяца и/или стабилен приблизительно при 2-8°C в течение по меньшей мере 1 года, в течение по меньшей мере 2 лет. Кроме того, состав предпочтительно, стабилен после замораживания (например, до -70°C) и оттаивания состава.
Антитело или производное антитела "сохраняет свою физическую стабильность" в фармацевтическом составе, если он не демонстрирует признаков агрегации, осаждения и/или денатурации при визуальном исследовании цвета и/или прозрачности, или как измеряют посредством рассеяния УФ-света или посредством гель-хроматографии.
Антитело или производное антитела "сохраняет свою химическую стабильность" в фармацевтическом составе, если химическая стабильность в данный момент времени такова, что белок рассматривают как все еще сохраняющий биологическую активность, как определено ниже. Химическую стабильность можно оценивать посредством детекции и количественного определения химически измененных форм белка. Химическое изменение может включать изменение размера (например, сжимание), которое можно оценивать, например, с применением гель-хроматографии, SDS-PAGE и/или времяпролетной масс-спектрометрии с лазерной десорбцией/ионизацией с помощью матрицы (MALDI/TOF MS). Другие типы химического изменения включают изменение заряда (например, происходящее в результате дезамидирования), которое можно оценивать, например, посредством ионообменной хроматографии.
Антитело или производное антитела "сохраняет свою биологическую активность" в фармацевтическом составе, если биологическая активность антитела в данный момент времени находится в пределах приблизительно 10% (в пределах ошибки анализа) от биологической активности, демонстрируемой в момент времени, когда был получен фармацевтический состав, как определяют, например, в анализе связывания антигена. Другие анализы "биологической активности" антител указаны ниже в настоящем документе.
Под "изотоническим" подразумевают, что представляющий интерес состав обладает по существу тем же осмотическим давлением, как и кровь человека. Изотонические составы, как правило, обладают осмотическим давлением приблизительно от 250 до 350 мОсм. Изотоничность можно измерять с использованием, например, осмометра давления пара или криоскопического типа.
"Полиол" представляет собой вещество с несколькими гидроксильными группами и включает сахара (восстанавливающие и невосстанавливающие сахара), сахарные спирты и сахарные кислоты. Предпочтительные полиолы по настоящему документу имеют молекулярную массу, составляющую менее чем приблизительно 600 кДа (например, в диапазоне приблизительно от 120 до приблизительно 400 кДа). "Восстанавливающий сахар" представляет собой сахар, содержащий полуацетальную группу, которая может восстанавливать ионы металлов или ковалентно реагировать с лизином и другими аминогруппами в белках, а "невосстанавливающий сахар" представляет собой сахар у которого нет этих свойств восстанавливающего сахара. Примеры восстанавливающих сахаров представляют собой фруктозу, маннозу, мальтозу, лактозу, арабинозу, ксилозу, рибозу, рамнозу, галактозу и глюкозу. Невосстанавливающие сахара включают сахарозу, трегалозу, сорбозу, мелицитозу и рафинозу. Примерами сахарных спиртов являются маннит, ксилит, эритритол, треитол, сорбит и глицерин. В отношении сахарных кислот, они включают L-глюконат и его соли металлов. Когда желательно, чтобы состав был стабилен к замораживанию-оттаиванию, полиол предпочтительно, представляет собой полиол, который не кристаллизуется при температурах замораживания (например, -20°C) так как, это дестабилизирует антитело в составе. Предпочтительными полиолами по настоящему документу являются невосстанавливающие сахара, такие как сахароза и трегалоза, где трегалоза является более предпочтительной, чем сахароза, по причине лучшей стабильности трегалозы в растворах.
Как используют в настоящем документе, "буфер" относится к буферному раствору, препятствующему изменениям pH действием его компонентов конъюгированных кислот-оснований. pH буфера по настоящему изобретению находится в диапазоне приблизительно от 4,5 до приблизительно 6,0; предпочтительно, приблизительно от 4,8 до приблизительно 5,5; а наиболее предпочтительно, pH составляет приблизительно 5,0. Примеры буферов, которые поддерживают pH в этом диапазоне, включают ацетат (например, ацетат натрия), сукцинат (такой как сукцинат натрия), глюконат, гистидин, цитрат и другие буферы органических кислот. Когда желателен состав, стабильный при замораживании-оттаивании, буфер предпочтительно, не является фосфатным.
В фармакологическом смысле, в контексте настоящего изобретения "терапевтически эффективное количество" антитела или производного антитела относится к количеству, эффективному для предотвращения или лечения нарушения, при лечении которого эффективно антитело или производное антитела. "Заболевание/нарушение" представляет собой любое состояние, на которое можно оказывать благоприятное воздействие при лечении антителом или производным антитела. Оно включает хронические и острые нарушения или заболевания, включая патологические состояния, которые предрасполагают млекопитающего к соответствующему нарушению.
"Консервант" представляет собой соединение, которое можно включать в состав для существенного снижения действия в нем бактерий, таким образом, например, облегчая получение состава для многократного использования. Примеры возможных консервантов включают хлорид октадецилдиметилбензиламмония, хлорид гексаметония, хлорид бензалкония (смесь хлоридов алкилбензилдиметиламмония в которых алкильные группы представляют собой длинноцепочечные соединения) и хлорид бензетония. Другие типы консервантов включают ароматические спирты, такие как фенол, бутиловый и бензиловый спирт, алкилпарабены, такие как метил- или пропилпарабен, катехол, резорцин, циклогексанол, 3-пентанол и м-крезол. Наиболее предпочтительным консервантом по настоящему документу является бензиловый спирт.
Настоящее изобретение также относится к фармацевтическим композициям, содержащим соединения одного или нескольких антител или производных антител по меньшей мере вместе с одним физиологически приемлемым носителем или эксципиентом. Фармацевтические композиции могут содержать, например, одно или несколько из воды, буферов (например, нейтральный буферный физиологический раствор или фосфатно-солевой буфер), этанола, минерального масла, растительного масла, диметилсульфоксида, углеводов (например, глюкозы, маннозы, сахарозы или декстранов), маннита, белков, адъювантов, полипептидов или аминокислот, таких как глицин, антиоксидантов, хелатирующих средств, таких как ЭДТА или глутатион, и/или консервантов. Как указано выше, в фармацевтические композиции, предоставляемые по настоящему документу можно включать другие активные ингредиенты may (но не обязательно).
Носитель представляет собой вещество, которое может быть ассоциированным с антителом или производным антитела до введения пациенту, часто с целью регуляции стабильности или биодоступности соединения. Носители для применения в таких составах, как правило, являются биологически совместимыми, а также могут являться биологически разрушаемыми. Носители включают, например, моновалентные или поливалентные молекулы, такие как сывороточный альбумин (например, человека или быка), яичный альбумин, пептиды, полилизин и полисахариды, такие как аминодекстран и полиамидоамины. Носители также включают вещества твердых носителей, таких как гранулы и микрочастицы, содержащие, например, полилактат-полигликолат, сополимер лактида с гликолидом, полиакрилат, латекс, крахмал, целлюлозу или декстран. Носитель может нести соединения множеством способов, включая ковалентное связывание (напрямую или посредством линкерной группы), нековалентное взаимодействие или смешивание.
Фармацевтические композиции можно формулировать для любого подходящего способа введения, включая, например, топическое пероральное, назальное, ректальное или парентеральное введение. В определенных вариантах осуществления предпочтительными являются композиции в форме, подходящей для перорального применения. Такие формы включают, например, пилюли, таблетки, пастилки, таблетки-леденцы, водные или масляные суспензии, диспергируемые порошки или гранулы, эмульсию, твердые или мягкие капсулы или сиропы или эликсиры. В других вариантах осуществления композиции, предоставляемые по настоящему документу, можно формулировать в виде лиофилизата. Как используется в настоящем документе термин парентеральный включает подкожную, интрадермальную, внутрисосудистую (например, внутривенную), внутримышечную, спинальную, внутричерепную, интратекальную и интраперитонеальную инъекцию, а также любой подобный способ инъекции или инфузии.
Композиции, предназначенные для перорального применения можно получать любым способом, известным в области получения фармацевтических композиций, и они могут содержать одно или несколько средств, таких как подсластители, ароматизаторы, красители и консерванты для придания привлекательности и вкусовых качеств препаратам. Таблетки содержат активный ингредиент в смеси с физиологически приемлемыми эксципиентами, подходящими для получения таблеток. Такие эксципиенты включают, например, инертные разбавители (например, карбонат кальция, карбонат натрия, лактозу, фосфат кальция или фосфат натрия), гранулирующие и дезинтегрирующие средства (например, кукурузный крахмал или альгиновую кислоту), связывающие средства (например, крахмал, желатин или гуммиарабик) и смазки (например, стеарат магния, стеариновую кислоту или тальк). Таблетки могут быть непокрытыми, или они могут быть покрытыми известными способами для задержки дезинтеграции и всасывания в желудочно-кишечном тракте и, таким образом, обеспечивать пролонгированное действие в течение более длительного периода. Например, можно использовать такое вещество для задержки времени, как глицерилмоностеарат или глицерилдистеарат.
Составы для перорального применения также можно предоставлять в виде твердых желатиновых капсул, где активный ингредиент смешан с твердым инертным разбавителем (например, карбонатом кальция, фосфатом кальция или каолином), или в виде мягких желатиновых капсул, где активный ингредиент смешан с водной или масляной средой (например, арахисовое масло, парафиновое масло или оливковое масло). Водные суспензии содержат антитело или производное антитела в смеси с эксципиентами, подходящими для получения водных суспензий. Такие эксципиенты включают суспендирующие средства (например, карбоксиметилцеллюлозу натрия, метилцеллюлозу, гидроксипропилметилцеллюлозу, альгинат натрия, поливинилпирролидон, трагакантовую камедь и гуммиарабик) и диспергирующие средства или увлажнители (например, природные фосфолипиды, такие как лецитин, продукты конденсации алкиленоксидов с жирными кислотами, такие как полиоксиэтиленстеарат, продукты конденсации этиленоксида с длинноцепочечными алифатическими спиртами, такие как гептадекаэтиленоксикетанол, продукты конденсации этиленоксида с частичными сложными эфирами, получаемыми из жирных кислот и гексита, такие как полиоксиэтиленсорбитмоноолеат, или продукты конденсации этиленоксида с частичными сложными эфирами, получаемыми из жирных кислот и ангибдридов гексита, такие как полиэтиленсорбитанмоноолеат). Водные суспензии также могут содержать один или несколько консервантов, например этил- или н-пропил-п-гидроксибензоат, один или несколько красителей, один или несколько ароматизаторов и один или несколько подсластителей, таких как сахароза или сахарин. Сиропы и эликсиры можно формулировать с подсластителями, такими как глицерин, пропиленгликоль, сорбит или сахароза. Такие составы также могут содержать один или несколько из уменьшающих раздражение средств, консервантов, ароматизаторов и/или красителей.
Масляные суспензии можно формулировать, суспендируя активные ингредиенты в растительном масле (например, арахисовом масле, оливковом масле, сезамовом масле или кокосовом масле) или в минеральном масле, таком как парафиновое масло. Масляные суспензии могут содержать загуститель, такой как пчелиный воск, твердый парафин или цетиловый спирт. Для предания пероральным препаратам приятного вкуса можно добавлять подсластители, такие как подсластители, указанные выше, и/или ароматизаторы. Такие суспензии можно предохранять добавлением антиоксиданта, такого как аскорбиновая кислота.
Диспергируемые порошки и гранулы, подходящие для получения водной суспензии посредством добавления воды, обеспечивают доставку активного ингредиента в смеси с диспергирующим средством или увлажнителем, суспендирующим средством и одним или несколькими консервантами. Подходящие диспергирующие средства или увлажнители и суспендирующие средства проиллюстрированы средствами, уже указанными выше. Также могут присутствовать дополнительные эксципиенты, например подсластители, ароматизаторы и красители.
Фармацевтические композиции также могут находиться в форме эмульсий "масло-в-воде". Масляная фаза может представлять собой растительное масло (например, оливковое масло или арахисовое масло), минеральное масло (например, парафиновое масло) или их смесь. Подходящие эмульгаторы включают природные камеди (например, гуммиарабик или трагакантовую камедь), природные фосфолипиды (например, соевые, лецитин и сложные эфиры или частичные сложные эфиры, получаемые из жирных кислот и гексита), ангидриды (например, сорбитанмоноолеат) и продукты конденсации частичных сложных эфиров, получаемые из жирных кислот и гексита с этиленоксидом (например, полиоксиэтиленсорбитанмоноолеат). Эмульсия также может содержать один или несколько подсластителей и/или ароматизаторов.
Фармацевтическую композицию можно получать в виде стерильной инъецируемой водной или масляной суспензии, в которой в зависимости от используемых носителя и концентрации модулятор или суспендирован, или растворен в носителе. Такую композицию можно формулировать в соответствии с известным уровнем техники с применением подходящих диспергирующих средств, увлажнителей и/или суспендирующих средств, таких как указанные выше средства. В числе приемлемых носителей и растворителей, которые можно использовать, находятся вода, 1,3-бутандиол, раствор Рингера и изотонический раствор хлорида натрия. Кроме того, в качестве растворителя или среды для суспендирования можно использовать стерильные жирные масла. Для этой цели можно использовать любое мягкое жирное масло, включая синтетические моно- или диглицериды. Кроме того, для получения инъецируемых композиций можно использовать жирные кислоты, так как олеиновая кислота, а вспомогательные вещества, такие как местные анестетики, консерванты и/или буферные средства можно растворять в носителе.
Фармацевтические композиции можно формулировать в виде составов с пролонгированным высвобождением (т.е., состав, такой как капсула, осуществляющий замедленное высвобождение модулятора после введения). Как правило, такие составы можно получать с применением хорошо известной технологии и вводить, например, посредством перорального, ректального или подкожного введения или посредством введения в желаемый участок-мишень. Носители для применения в таких составах являются биосовместимыми, а также могут быть биологически разрушаемыми; предпочтительно, состав обеспечивает относительно постоянный уровень высвобождения модулятора. Количество антитела или производного антитела, содержащееся в составе с пролонгированным высвобождением, зависит, например, от участка введения, скорости и ожидаемой длительности высвобождения и характера заболевания/нарушения, подлежащего лечению или предотвращению.
Антитела или производные антител, предоставляемые по настоящему документу, как правило, вводят в таком количестве, чтобы достичь такой концентрации в биологической жидкости (например, крови, плазме, сыворотке, CSF, синовиальной жидкости, лимфе, межклеточной жидкости, слезах или моче), которой достаточно для детектируемого связывания с мишенью, например, такой как VEGF, и для предотвращения или подавления опосредуемых такой мишенью заболеваний/нарушений, например опосредованных VEGF заболеваний/нарушений. Полагают, что доза является эффективной, если она приводит к видимым улучшениям у пациента, как описано в настоящем документе. Предпочтительный диапазон системных доз составляет приблизительно от 0,1 мг до приблизительно 140 мг на килограмм массы тела в сутки (приблизительно от 0,5 мг до приблизительно 7 г одному пациенту в сутки), с пероральными дозами, как правило, составляющими приблизительно в 5-20 раз выше, чем внутривенные дозы. Количество антитела или производного антитела, которое можно комбинировать с веществами носителей для получения одной лекарственной формы варьирует в зависимости от подлежащего лечению организма и конкретного способа введения. Стандартные лекарственные формы, как правило, содержат приблизительно от 1 мг до приблизительно 500 мг активного ингредиента.
Фармацевтические композиции можно упаковывать для лечения состояний, реагирующих на антитело или производное антитела, направленное, например, к VEGF. Упакованные фармацевтические композиции могут включать контейнер, содержащий эффективное количество по меньшей мере одного антитела или производного антитела, как описано в настоящем документе, и инструкции (например, этикетку), указывающие, что содержащуюся композицию следует использовать для лечения заболевания/нарушения, реагирующего на это одно антитело или производное антитела после введения пациенту.
Антитела или производные антител по настоящему изобретению также можно модифицировать химически. Предпочтительные модифицирующие группы представляют собой полимеры, например, необязательно замещенные полиалкеновые, полиалкениленовые или полиоксиалкиленовые полимеры с неразветвленной или разветвленной цепью или неразветвленные или разветвленные полисахариды. Такая эффекторная группа может увеличивать период полувыведения антитела in vivo. Конкретные примеры синтетических полимеров включают необязательно замещенные поли(этиленгликоль) (PEG), поли(пропиленгликоль), поли(виниловый спирт) с неразветвленной или разветвленной цепью или их производные. Конкретные природные полимеры включают лактозу, амилозу, декстран, гликоген или их производные. Размер полимера можно варьировать по желанию, но, как правило, его средняя молекулярная масса находится в диапазоне от 500 Да до 50000 Да. Для местного применения, когда антитело конструируют так, чтобы проникать в ткань, предпочтительная молекулярная масса полимера составляет приблизительно 5000 Да. Полимерную молекулу можно присоединять к антителу, в частности, к C-концу тяжелой цепи Fab-фрагмента посредством ковалентно связанного шарнирного пептида, как описано в WO0194585. Относительно присоединения молекул PEG, можно привести ссылку на "Poly(ethyleneglycol) Chemistry, Biotechnological and Biomedical Applications", 1992, J. Milton Harris (ed), Plenum Press, New York и "Bioconjugation Protein Coupling Techniques for the Biomedical Sciences", 1998, M. Aslam and A. Dent, Grove Publishers, New York.
После получения представляющего интерес антитела или производного антитела, как описано выше, получают содержащий его фармацевтический состав. Антитело для формулирования не подвергают предшествующей лиофилизации и представляющий интерес состав по настоящему документу представляет собой водный состав. Предпочтительно, антитело или производное антитела в составе представляет собой фрагмент антитела, такой как scFv. Терапевтически эффективное количество антитела, находящееся в составе, определяют, например, принимая в расчет желаемые объемы доз и способ(ы) введения. Иллюстративной концентрацией антитела в составе является приблизительно от 0,1 мг/мл до приблизительно 50 мг/мл, предпочтительно, приблизительно от 0,5 мг/мл до приблизительно 25 мг/мл, а наиболее предпочтительно, приблизительно от 2 мг/мл до приблизительно 10 мг/мл.
Получают водный состав, содержащий антитело или производное антитела в раствор с забуференным pH. pH буфера по настоящему изобретению находится в диапазоне приблизительно от 4,5 до приблизительно 6,0, предпочтительно, приблизительно от 4,8 до приблизительно 5,5, а наиболее предпочтительно, pH составляет приблизительно 5,0. Примеры буферов, которые поддерживают pH в этом диапазоне включают ацетат (например, ацетат натрия), сукцинат (такой как сукцинат натрия), глюконат, гистидин, цитрат и другие буферы органических кислот. Концентрация буфера может составлять приблизительно от 1 мМ до приблизительно 50 мМ, предпочтительно, приблизительно от 5 мМ до приблизительно 30 мМ, в зависимости, например, от буфера и желаемой изотоничности состава. Предпочтительным буфером является ацетат натрия (приблизительно 10 мМ), pH 5,0.
В состав включают полиол, который действует в качестве придающего тоничность средства и может стабилизировать антитело. В предпочтительных вариантах осуществления состав не содержит придающего тоничность количества соли, такой как хлорид натрия, так как это может вызывать осаждение антитела или производного антитела и/или может приводить к окислению при низком pH. В предпочтительных вариантах осуществления полиол представляет собой невосстанавливающий сахар, такой как сахароза или трегалоза. Полиол добавляют к составу в количестве, которое можно варьировать в соответствии с желаемой изотоничностью состава. Предпочтительно, водный состав является изотоническим, в случае чего подходящие концентрации полиола в составе находятся, например, в диапазоне приблизительно от 1% до приблизительно 15% масс./об., предпочтительно, в диапазоне приблизительно от 2% до приблизительно 10% масс./об. Однако подходящими также могут являться гипертонические или гипотонические составы. Количество добавляемого полиола также можно изменять в соответствии с молекулярной массой полиола. Например, можно добавлять меньшее количество моносахарида (например, маннита) по сравнению с дисахаридом (таким как трегалоза).
В состав антитела или производного антитела также добавляют поверхностно-активное вещество. Примеры поверхностно-активных веществ включают неионные поверхностно-активные вещества, такие как полисорбаты (например, полисорбаты 20, 80 и т.д.) или полоксамеры (например, полоксамер 188). Количество добавляемого поверхностно-активного вещества является таким, чтобы оно снижало агрегацию формулируемого антитела/производного антитела, и/или минимизировало образование частиц в составе, и/или снижало адсорбцию. Например, поверхностно-активное вещество может находиться в составе в количестве приблизительно от 0,001% до приблизительно 0,5%, предпочтительно, приблизительно от 0,005% до приблизительно 0,2%, а наиболее предпочтительно, приблизительно от 0,01% до приблизительно 0,1%.
В одном из вариантов осуществления состав содержит указанные выше средства (т.е. антитело или производное антитела, буфер, полиол и поверхностно-активное вещество) и по существу не содержит один или несколько консервантов, таких как бензиловый спирт, фенол, м-крезол, хлорбутанол и хлорид бензетония. В другом варианте осуществления в состав можно включать консервант, особенно когда состав представляет собой состав для многократного дозирования. Концентрация консерванта может находиться в диапазоне приблизительно от 0,1% до приблизительно 2%, наиболее предпочтительно, приблизительно от 0,5% до приблизительно 1%. В состав можно включать один или несколько других фармацевтически приемлемых носителей, эксципиентов или стабилизаторов, таких как описаны в Remington's Pharmaceutical Sciences 21st edition, Osol, A. Ed. (2006), при условии, что они не оказывают негативного воздействия на желаемые характеристики состава. Приемлемые носители, эксципиенты или стабилизаторы являются нетоксическими для реципиента в применяемых дозах и концентрациях и включают дополнительные буферные средства; вспомогательные растворители; антиоксиданты, включая аскорбиновую кислоту и метионин; хелатирующие средства, такие как ЭДТА; комплексные соединения с металлами (например, комплексы Zn-белок); биологически разрушаемые полимеры, такие как сложные полиэфиры; и/или образующие соли противоионы, такие как натрий.
Составы для использования для введения in vivo должны быть стерильными. Это легко осуществлять посредством фильтрации через стерильные фильтровальные мембраны до или после получения состава.
Состав вводят нуждающемуся в лечении антителом млекопитающему, предпочтительно, человеку, известными способами, такими как внутривенное введение в виде болюсной или непрерывной инфузии в течение периода времени, внутримышечным, интраперитонеальным, интрацереброспинальным, подкожным, внутрисуставным, интрасиновиальным, интратекальным, пероральным, топическим или ингалационным способами. В предпочтительных вариантах осуществления состав вводят млекопитающему посредством внутривенного введения. С этой целью состав можно инъецировать с применением, например, шприца или капельницы.
Подходящая доза ("терапевтически эффективное количество") антитела зависит, например, от подлежащего лечению состояния, тяжести и течения состояния, вводят ли антитело с профилактической или терапевтической целью, предшествующего лечения, истории болезни пациента и его реакции на антитело, типа используемого антитела, решения лечащего врача. Антитело или производное антитела целесообразно вводить пациенту один раз или посредством нескольких введений и в дальнейшем можно в любое время вводить пациенту для диагностики. Антитело или производное антитела можно вводить в качестве единственного способа лечения или в сочетании с другими лекарственными средствами или способами лечения, пригодными для лечения соответствующего заболевания.
В качестве общего предложения терапевтически эффективное количество вводимого антитела или производного антитела находится в диапазоне приблизительно от 0,1 до приблизительно 50 мг/кг массы тела пациента при одном или нескольких введениях, например, с обычным диапазоном используемого антитела, вводимого ежедневно, составляющим приблизительно от 0,3 до приблизительно 20 мг/кг, более предпочтительно, приблизительно от 0,3 до приблизительно 15 мг/кг. Однако пригодными могут являться и другие режимы дозирования. Ход этого лечения легко контролировать общепринятыми способами.
В другом варианте осуществления изобретение относится к промышленному изделию, содержащему контейнер, содержащий фармацевтический состав по настоящему изобретению, предпочтительно, водный состав, и необязательно содержащее инструкции для его использования. Подходящие контейнеры включают, например, бутылки, ампулы и шприцы. Контейнер можно получать из ряда материалов, таких как стекло или пластик. Пример контейнера представляет собой стеклянную ампулу для однократного применения объемом 3-20 см3. Альтернативно, для составов с несколькими дозами, контейнер может представлять собой стеклянную ампулу объемом 3-100 см3. Контейнер содержит состав, и на ярлыке на контейнере, или приложенном к нему, могут находиться указания для использования. Промышленное изделие может дополнительно включать материалы, желательные с коммерческой и пользовательской точки зрения, включая другие буферы, разбавители, фильтры, иглы, шприцы и вкладыши в упаковку с инструкциями по применению.
В определенных предпочтительных вариантах осуществления промышленное изделие содержит лиофилизированную иммуносвязывающую молекулу, как описано в настоящем документе, или полученную способами, описываемыми в настоящем документе.
Примеры
Настоящее описание дополнительно проиллюстрировано приведенными ниже примерами, которые не следует рассматривать как дополнительное ограничение. Содержание всех фигур и всех ссылок, патентов и опубликованных патентных заявок, цитируемых на всем протяжении настоящей заявки в явном виде полностью включено в настоящий документ в качестве ссылки.
Материалы и методы
В основном, в практическом осуществлении настоящего изобретения, если не указано иначе, используют общепринятые способы химии, молекулярной биологии, технологии рекомбинантных ДНК, иммунологии (в частности, например, технологии антител) и стандартные способы получения полипептидов. См., например, Sambrook, Fritsch and Maniatis, Molecular Cloning: Cold Spring Harbor Laboratory Press (1989); Antibody Engineering Protocols (Methods in Molecular Biology), 510, Paul, S., Humana Pr (1996); Antibody Engineering: A Practical Approach (Practical Approach Series, 169), McCafferty, Ed., Irl Pr (1996); Antibodies: A Laboratory Manual, Harlow et al., C.S.H.L. Press, Pub. (1999); и Current Protocols in Molecular Biology, eds. Ausubel et al., John Wiley & Sons (1992). Способы пересадки CDR из моноклональных антител кролика и других не являющихся человеческими моноклональных антител в выбранные каркасы антител человека подробно описан выше. Примеры экспериментов по такой пересадке приведены ниже.
С целью лучшего понимания, пересадки, обозначенные как "мин" представляют собой пересадки, где CDR пересажены на каркас 1.4 или его вариабельный домен, тогда как пересадки, обозначенные как "макс" представляют собой пересадки, где в каркас 1.4 или его вариабельный домен пересажены CDR, и где каркас дополнительно содержит донорные каркасные остатки, которые взаимодействуют с антигеном.
Пример 1: Конструкция rFW1.4
1.1. Анализ первичной последовательности и поиск в базах данных
1.1.1. Сбор последовательностей иммуноглобулинов кролика
Последовательности вариабельных доменов зрелых антител и зародышевой линии кролика собирали из различных общедоступных баз данных (например, база данных Kabat и IMGT) и вводили в собственную базу данных в виде однобуквенного кода аминокислотных последовательностей. Для полного анализа авторы использовали только часть аминокислот, соответствующую V (вариабельной) области. Последовательности в базе данных KDB, содержащие менее 70% полной последовательности или содержащие множество неопределенных остатков в каркасных областях, удаляли. Последовательности, более чем на 95% идентичные другим последовательностям в базе данных, также исключали во избежание случайных помех при анализе.
1.1.2. Выравнивание и нумерация последовательностей кролика
Последовательности антител кролика выравнивали с использованием традиционного средства для выравнивания последовательностей на основе алгоритма Нидлмана-Вунша и матриц Blossum. Введение пропусков и обозначение положений остатков проводили в соответствии с системой нумерации AHo для вариабельных доменов иммуноглобулинов (Honegger and Pluckthun, 2001). Также параллельно применяли схему нумерации по Kabat, так как она представляет собой наиболее широко принятый стандарт нумерации остатков в антителе. Нумерацию по Kabat присваивали с применением программы SUBIM. Эта программа анализирует вариабельные области последовательности антитела и нумерует последовательность в соответствии с системой, установленной Kabat и с коллегами (Deret et al 1995).
Определение областей каркаса и CDR проводили по определению Kabat, которое основано на вариабельности последовательностей и является наиболее часто применяемым. Однако обозначение CDR H1 представляло собой компромисс между различными определениями, включающими AbM, Kabat, средние данные о контактах, получаемые посредством анализа контактов антитела и антигена подмножества трехмерных сложных структур (MacCallum et al., 1996) и Chotia, которое основано на определении положения областей структурных петель (описано выше и представлено на фиг. 1).
1.1.3. Частота и консервативность положений остатков
Вариабельность аминокислотной последовательности анализировали с использованием набора из 423 последовательностей кролика из базы данных Kabat. Частоту остатков, f(r), для каждого положения, i, в зрелых последовательностях кролика рассчитывали по количеству раз, когда конкретный остаток наблюдали в наборе данных, деленому на общее количество последовательностей. Степень консервативности для каждого положения, i, рассчитывали с использованием индекса Симпсона, который учитывает количество различных встречающихся аминокислот, а также относительное содержание каждого остатка.
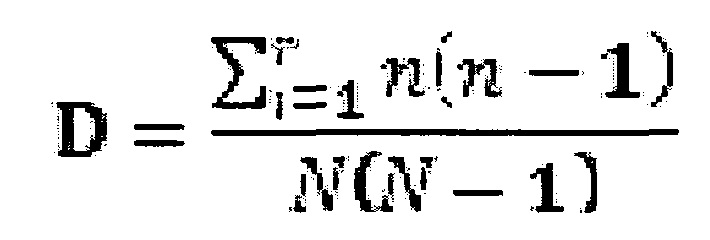
где: N представляет собой общее количество аминокислот, r представляет собой количество различных встречающихся аминокислот в каждом положении, и n представляет собой количество остатков конкретного типа аминокислоты.
1.1.4. Генеалогический анализ V-области кролика
Для исследования репертуара кролика использовали средства филогенетического анализа. Последовательности аминокислот V-области кластеризовали с применением алгоритма кластеризации и топологического алгоритма. Для всего массива рассчитывали матрицу расстояний и использовали как показатель использования зародышевой линии. Рассчитывали консенсусную последовательность всего кластера и идентифицировали ближайший вариант последовательности зародышевой линии кролика. Также из полного набора последовательностей получали общую консенсусную последовательность.
1.1.5. Распределение по подгруппам человека.
Для каждой характерной последовательности кролика из различных кластеров идентифицировали наиболее гомологичную подгруппу человека с применением реализации алгоритмов анализа последовательностей EXCEL и способов классификации на основе анализа репертуара антител человека (Knappik et al., 2000).
1.2. Проектирование акцепторного каркаса человека
С копией репертуара кролика из анализа последовательностей описанного выше, определяли остатки в каркасе, как правило, участвующие в расположении CDR кролика. Среди каркасов с высокой гомологией с репертуаром кролика и с соответствующими кластерами из совокупности полностью человеческих последовательностей выбирали одну с хорошими биофизическими свойствами. Каркас, выбранный для того, чтобы служить акцепторным каркасом, принадлежит вариабельной области легкой цепи подгруппы каппа 1 и вариабельной области тяжелой цепи подгруппы III с ID ESBATech KI27, a43 соответственно. Этот стабильный и растворимый каркас антитела идентифицировали в скрининге библиотеки scFv селезенки человека с использованием основанного на применении дрожжей способа скрининга, названного система "контроль качества" (Auf der Maur et al., 2004) и обозначили как "FW1.4". Хотя стабильная и растворимая каркасная последовательность FW1.4 демонстрирует высокую гомологии, она не является наиболее гомологичной доступной последовательностью. Идентифицированные остатке ввели в указанный акцепторный каркас с получением rFW1.4.
С информацией о вариабельности аминокислотной последовательности, использованием зародышевой линией и структурных характеристик антител кролика авторы анализировали FW1.4 на совместимость положений остатков, необходимую для сохранения конформации CDR в новом каркасе человека. Авторы исследовали вариабельные области FW1.4 на совместимость по следующим характеристикам:
i. Остатки, которые являются частью канонических последовательностей для петлеобразных структур.
ii. Каркасные остатки, расположенные на поверхности контакта VL/VH.
iii. Основание из остатков, расположенных непосредственно под CDR
iv. Остатки верхней и нижней сердцевины
v. Каркасные остатки, определяющие подтип
1.3. Пересадка CDR кролика
Пересадки получали посредством простого комбинирования последовательностей CDR (по указанному выше определению) из одного антитела с каркасной последовательностью FW1.4 или rFW1.4. Идентифицировали остатки, потенциально участвующие в связывании. Для каждой последовательности вариабельного домена кролика, определяли ближайший вариант зародышевой линии. Если ближайшую зародышевую линию не устанавливали, последовательность сравнивали с консенсусом подгруппы или консенсусом последовательностей кролика с высоким процентом сходства. Редкие каркасные остатки рассматривали как возможный результат соматического гипермутирования и, таким образом, как играющие роль в связывании. Таким образом, такие остатки пересаживали в акцепторный каркас.
1.4 Результаты
Посредством анализа репертуара антител кролика, исходя из структуры, вариабельности аминокислотных последовательностей и с использованием зародышевой линии, найдено 5 положений остатков в легкой цепи FW1.4, которые модифицировали для поддержания конформации петель CDR кролика. Эти положения в антителах кролика являются высококонсервативными. Для этих 5 положений на основе репертуара кролика получали консенсусные остатки и вводили в акцепторный каркас человека 1.4. При модификации этих консервативных положений указанный каркас фактически становился совместимым со всеми шестью определяющими комплементарность областями (CDR) любых CDR кролика. Базовая rFW1.4, содержащая различные CDR кролика хорошо экспрессируется и хорошо продуцируется в отличие от отдельных цепей дикого типа. Получили 16 представителей, полученных на основе комбинации этого каркаса и CDR кролика, подробная характеристика продемонстрировала функциональность.
Пример 2: система скрининга B-клеток
Для отбора B-клеток, связывающих представляющую интерес мишень посредством своих B-клеточных рецепторов (BCR) адаптировали основанную на FACS (проточная цитометрия на основе сортировки единичных клеток) систему скрининга на ESBATech. Например, одной из мишеней являлся, растворимый белок, а именно, одноцепочечное антитело (ESBA903), меченое флуоресцентным красителем (PE и PerCP). Из селезенки кроликов, иммунизированных рекомбинантной мишенью, получали суспензию лимфоцитов. Затем клетки инкубировали с меченым PE и PerCP ESBA903, а также с антителами специфичными к IgG (мечеными APC) или IgM (мечеными FITC). ESBA903-позитивные B-клетки, которые экспрессируют на своей поверхности IgG, но не IgM отсортировывают в 96-луночные планшеты (фигура 3; таблица 2). Посредством вспомогательной клеточной линии тимомы (EL4-B5: см. Zubler et al, 1985, J. Immunol, 134(6): 3662-3668), отобранные B-клетки пролиферировали, дифференцировались в плазматические клетки и секретировали антитела. Аффинность этих IgG к мишени подтверждали посредством измерений ELISA и Biacore. Кинетические параметры для семи отобранных клонов приведены в таблице 1. Эти клоны из совокупности из ≈200 отсортированных клеток демонстрируют аффинности связывания в диапазоне низких концентраций от наномолярных до пикомолярных. В заключение из 6 представляющих интерес клонов выделяли мРНК и CDR пересаживали в одноцепочечный каркас FW1.4.
Кинетические значения для 7 супернатантов культур B-клеток
Статистические параметры сортировки
Пример 3: Детекция взаимодействия между гранулами, покрытыми антителами против TNF-альфа и клетками CHO, экспрессирующими мембраносвязанный TNF-альфа.
Для оценки того, разрушает ли поток при высоком давлении при проточной цитометрии или нет нековалентное связывание двух клеток, проводили следующий эксперимент. Клетки CHO стабильно трансфицированные мембраносвязанным TNF-альфа (клетки B-220) инкубировали с гранулами, покрытыми мечеными PE антителами против TNF. В этих условиях гранулы имитировали B-клетки памяти (они имеют более или менее такой же размер). В качестве отрицательного контроля использовали нетрансфицированные клетки CHO, а гранулы, покрытые несвязывающимся антителом (против CD19), меченым APC. Через 2 часа инкубации при 4°C с перемешиванием суспензию клеток-гранул анализировали посредством FACS (с использованием 130 мкм форсунок). На фигуре 4 показано, что специфическое связывание между гранулами с антителами против TNF-альфа и клетками CHO, трансфицированными TNF-альфа четко детектируемо FACS. Действительно, в этом образце (верхняя панель) приблизительно два трети гранул связаны с клетками (585 связанных относительно 267 несвязанных). В отличие от этого в контрольных образцах (средняя и нижняя панели), гранулы почти не связываются с клетками CHO. Кроме того, обе совокупности гранул (с антителами против TNF-альфа-PE и с антителами против CD19-APC) смешивали вместе с клетками CHO, трансфицированными TNF-альфа. На фигуре 5 и в таблице 4 показано, что приблизительно половина из гранул с антителами против TNF-альфа связывается с клетками CHO, тогда как подавляющее большинство гранул с антителами против CD19 остаются несвязанными. Процент связывания гранул с клетками в каждом образце подробно представлен в таблице 5. Таким образом, проведена демонстрация того, что специфический отбор одиночных B-клеток, которые связываются с интегральным мембранным белком-мишенью посредством своего B-клеточного рецептора с применением проточной цитометрии возможен.
Статистические параметры сортировки (также см. фиг. 4a)
Статистические параметры сортировки (также см. фиг. 4b)
Статистические параметры сортировки (также см. фиг. 4c)
Таблица 4
Статистические параметры сортировки (также см. фиг. 5)
Доля гранул, связанных с клетками CHO в каждом образце
Пример 4: Пересадка CDR и функциональная гуманизация донорных антител кролика против TNFα
Для пересадки CDR выбирали четыре антитела кролика против TNFα "Rabmab" (EPI-1, EPI-15, EP-34, EP-35 и EP-42). Общая схема эксперимента по пересадке CDR, гуманизации и предварительной характеристики гуманизированных донорных антител кролика проводили, как изложено в описании. В отличие от традиционных способов гуманизации, в которых используют акцепторный каркас антитела человека, обладающий наибольшей гомологией последовательности с не являющимся человеческим донорным антителом, CDR кролика пересаживали в каркас человека (FW1.4), который предварительно выбрали на основании желаемых функциональных свойств (растворимости и стабильности) с использованием анализа "контроль качества" (WO0148017). Эта стабильная и растворимая каркасная последовательность демонстрировала высокую гомологию с RabMab.
Для каждого из RabMab получали пересадки CDR с использованием описываемого в настоящем документе принципа. В пересадках "мин" пересаживали только CDR кролика из доменов VL и VH донорного антитела кролика в акцепторный каркас человека FW1.4. Пересадки "макс" относятся к пересадке CDR кролика в rFW1.4.
ScFv, описываемые и характеризуемые в настоящем документе, получали указанным далее образом. Последовательности гуманизированных VL соединяли с последовательностями гуманизированных VH последовательности посредством линкера SEQ ID NO:8 с получением scFv в следующей ориентации: NH2-VL-линкер-VH-COOH. Во многих случаях последовательности ДНК, кодирующую различные scFv, синтезировали de novo у поставщика услуг Entelechon GmbH (www.entelechon.com). Полученные вставки ДНК клонировали в бактериальный вектор экспрессии pGMP002 посредством участков рестрикции NcoI и HindIII, вводимых на 5'- и 3'-концах последовательности ДНК scFv, соответственно. Между последовательностями ДНК домена VL и домена VH, расположен участок BamHI. В некоторых случаях ДНК, кодирующую scFv, не синтезировали de novo, а конструкции, экспрессирующие scFv, клонировали посредством перестановки доменов. Таким образом, домены VL вырезали и вставляли в новые конструкции посредством участков рестрикции NcoI и BamHI, а домены VH посредством участков рестрикции BamHI и HindIII. В других случаях в домены VH и/или VL вносили точечные мутации с использованием способов сборки ПЦР на существующем уровне техники. Клонирование GMP002 описано в примере 1 WO2008006235. Получение scFv проводили аналогично ESBA105, как описано в примере 1 WO2008006235.
В таблице 3 приведены обобщенные данные подробной характеристики четырех моноклональных антител кролика (EP6, EP19, EP34, EP35 и EP43) и их вариантов с пересаженными CDR. Хотя пересадки CDR в анализах связывания BIACore и в анализах опосредованной TNFα цитотоксичности L929, демонстрировали широкий диапазон активности, 3 из 4 максимальных ("макс") пересадок демонстрировали терапевтически значимую активность. EP43макс демонстрировал наиболее подходящую аффинность связывания (Kd 0,25 нМ) и превосходную EC50 в анализе цитотоксичности. Эти данные демонстрируют, что FW1.4 (SEQ ID NO:1 и 2) представляет собой иллюстративную растворимую и стабильную область акцепторного каркаса человека для пересадки CDR кролика.
Анализ эффективности
Нейтрализующую активность связывающих антител против TNFα оценивали в анализе опосредованной TNFα цитотоксичности L929. Токсичность клеток фибробластов L929 мыши, обработанных актиномицином индуцировали рекомбинантным TNF человека (hTNF). Определяли, что 90% максимальной индуцированной hTNF цитотоксичности достигается при концентрации TNF 1000 пг/мл. Все клетки L929 культивировали в RPMI 1640 с феноловым красным, со средой с L-глутамином, дополненной эмбриональной телячьей сывороткой (10% об./об.). Нейтрализующую активность связывающих антител против TNFα оценивали в RPMI 1640 без фенолового красного и 5% эмбриональной телячьей сыворотки. К клеткам L929 в присутствие 1000 пг/мл hTNF для определения концентрации, при которой антагонистический эффект достигал половины максимального ингибирования (% EC50), добавляли различные концентрации (0-374 нг/мл) связывающих антител против TNF. Кривую доза-ответ аппроксимировали нелинейной сигмовидной регрессией переменным угловым коэффициентом и рассчитывали EC50.
Анализ связывания scFv против TNF Biacore
Для измерения аффинности связывания проводили измерения поверхностного плазмонного резонанса на BIAcore™-T100 с использованием чипа сенсора NTA и меченого His TNF (производимого ESBATech). Поверхность чипа сенсора NTA состоит из карбоксиметилированного декстранового матрикса предварительно иммобилизованного нитрилтриуксусной кислотой (NTA) для захвата меченых гистидином молекулы посредством хелатирования Ni2+NTA. Тримеры TNFα человека N-his (5 нМ) захватываются никелем посредством их N-концевых меток his и в различных концентрациях в диапазоне от 30 нМ до 0,014 нМ с шагом 3-кратного серийного разведения инъецируют ESBA105 (анализируемое вещество). На этапе регенерации комплекс, образуемый никелем, лигандом и анализируемым веществом смывают. Это позволяет использовать одинаковые условия регенерации для различных образцов. Ответный сигнал получают посредством технологии поверхностного плазмонного резонанса (SPR) и измеряют в резонансных единицах (RU). Все измерения проводят при 25°C. Сенсограммы получают для каждого из образцов scFv против TNF после встроенной коррекции на клетки эталона с последующим вычитанием образца буфера. Кажущуюся константу скорости диссоциации (kd), кажущуюся константу скорости ассоциации (ka) и кажущуюся равновесную константу диссоциации (KD) рассчитывали с применением модели связывания Лэнгмюра один к одному с использованием программного обеспечения для анализа BIAcore T100 версии 1.1.
Второе поколение связывающих TNF-альфа молекул
**(мг/л раствора для рефолдинга); *** Не определяли
Пример 5: Пересадка CDR и функциональная гуманизация донорных антител кролика против VEGF
Для пересадки CDR отбирали восемь Rabmab против VEGF (375, 435, 509, 511, 534, 567, 578 и 610). В отличие от традиционных способов гуманизации, в которых используют акцепторный каркас антитела человека, обладающий наибольшей гомологией последовательности с не являющимся человеческим донорным антителом, CDR кролика пересаживали в акцепторный каркас человека FW1.4 (SEQ ID NO:1 и 2), который предварительно выбрали на основании желаемых функциональных свойств (растворимости и стабильности) с использованием анализа "контроль качества" (WO0148017).
Для каждого из RabMab (антител кролика) получали несколько пересадок CDR с использованием описываемого в настоящем документе принципа (см. пример 4). Пересадки "мин" включали минимальную пересадку, где из доменов VL и VH донорного антитела кролика в акцепторный каркас человека FW1.4 (SEQ ID NO:1) пересаживали только CDR кролика. Пересадки "макс" содержали не только CDR кролика из VL и VH, но также некоторые дополнительные каркасные остатки из донорного антитела кролика, для которых был получен прогноз, что они являются важными для связывания антигена. В случае 578макс, каркасная область FW1.4 вариабельного домена тяжелой цепи содержала дополнительные изменения аминокислот в положениях по Kabat 23H, 49H, 73H, 78H и 94H.
В таблице 4 приведены обобщенные данные подробной характеристики вариантов "мин" и "макс" с пересаженными CDR. Описана их эффективность в качестве ингибиторов VEGF, которую измеряют с использованием конкурентного ELISA VEGFR и/или анализа HUVEC. Эти данные демонстрируют, что FW1.4 (SEQ ID NO:1 и 2) представляет собой иллюстративную растворимую и стабильную область акцепторного каркаса человека для пересадки CDR кролика.
Анализ связывания scFv против VEGF Biacore
Тестировали способность к связыванию scFv в Biacore и аффинность связывания измеряли с использованием иллюстративного способа поверхностного плазмонного резонанса на BIAcore™-T100. Белки VEGF, которые тестировали на связывание этими scFv-кандидатами, в этом примере и дальнейших примерах включают очищенные экспрессируемые Escherichia coli рекомбинантный VEGF165 человека (PeproTech EC Ltd.), рекомбинантный VEGF121 человека (PeproTech EC Ltd.), рекомбинантный VEGF110 человека (ESBATech AG), рекомбинантный VEGF164 мыши (PeproTech EC Ltd.), рекомбинантный VEGF164 крысы (Biovision), рекомбинантный VEGF110 кролика (ESBATech AG) и рекомбинантный PLGF человека (PeproTech EC Ltd.). Для эксперимента с поверхностным плазмонным резонансом карбоксиметилированные декстрановые чипы биосенсора (CM4, GE Healthcare) активировали гидрохлоридом N-этил-N'-(3-диметиламинопропил)карбодиимидом и N-гидроксисукцинимидом по инструкциям поставщика. Каждую из 6 различных форм VEGF, как проиллюстрировано выше, связывали с различными проточными ячейками на чипе сенсора CM4 в количестве от 1 до 4 стандартным способом связывания аминов. Диапазон ответов, полученных с этими иммобилизованными молекулами VEGF после связывания и блокирования составлял ≈250-500 единиц ответа (RU) для hVEGF165, ≈200 RU для hVEGF110, hVEGF121, VEGF164 мыши, VEGF164 крысы и VEGF110 кролика, и ≈400 RU для PLGF. 4-ую проточную ячейку каждого чипа обрабатывали сходным образом, за исключением того, что до блокирования не добавляли белков и проточную ячейку использовали в качестве встроенного эталона. В проточные ячейки инъецировали различные концентрации scFv против VEGF (например, 90 нМ, 30 нМ, 10 нМ, 3,33 нМ, 1,11 нМ, 0,37 нМ, 0,12 нМ и 0,04 нМ) в буфере HBS-EP (0,01 М HEPES, pH 7,4, 0,15 М NaCl, 3 мМ ЭДТА, 0,005% поверхностно-активное вещество P20) при скорости потока 30 мкл/мин в течение 5 мин. Диссоциации scFv к VEGF и VEGF на чипе CM4 позволяли проходить в течение 10 мин при 25°C. Сенсограммы получают для каждого из образцов scFv к VEGF после встроенной коррекции на клетки эталона с последующим вычитанием образца буфера. Кажущуюся константу скорости диссоциации (kd), кажущуюся константу скорости ассоциации (ka) и кажущуюся равновесную константу диссоциации (KD) рассчитывали с применением модели связывания Лэнгмюра один к одному с использованием программного обеспечения для анализа BIAcore T100 версии 1.1.
Анализ HUVEC ингибирования VEGF
Анализ HUVEC представляет собой способ измерения эффективности описанных scFv к VEGF в качестве кандидатов в ингибиторы VEGF.
Использовали эндотелиальные клетки пупочной вены человека (HUVEC) (Promocell), объединенные от нескольких доноров, с пассажа 2 до пассажа 14. Клетки высевали при 1000 клеток/лунку в 50 мкл полной среды для выращивания эндотелиальных клеток (ECGM) (Promocell), содержащей 0,4% ECGS/H, 2% эмбриональную телячью сыворотку, 0,1 нг/мл эпидермального фактора роста, 1 мкг/мл гидрокортизона, 1 нг/мл основного фактора фибробластов и 1% пенициллин/стрептомицин (Gibco). Через период от 7 до 8 часов в клетки добавляли 50 мкл обедненной среды (ECGM без добавок, содержащей 0,5% инактивированной нагреванием FCS и 1% пенициллин/стрептомицин) и клетки поддерживали на обедненной среде в течение периода от 15 до 16 часов. В обедненной среде получали 3-кратные серийные разведения к scFv VEGF (0,023-150 нМ) и одного из следующего: рекомбинантного VEGF165 человека (0,08 нМ), рекомбинантного VEGF164 мыши (0,08 нМ) или рекомбинантного VEGF164 крысы (0,3 нМ), и предварительно инкубировали в течение 30-60 мин при комнатной температуре. Использовали различные концентрации VEGF для компенсации их различной относительной биологической активности. Использовали концентрации, которые стимулируют субмаксимальную индуцированную VEGF пролиферацию (EC90). 100 мкл смесей добавляли в 96-луночные планшеты для культивирования тканей, содержащие суспензию HUVEC, и инкубировали в течение 4 суток во влажном инкубаторе с 37°C/5% CO2. Пролиферацию HUVEC оценивали посредством измерения оптической плотности при 450 нм (в качестве эталонной длины волны использовали 620 нм) после добавления 20 мкл/лунку реагента WST-1 для клеточной пролиферации (Roche) с использованием микроспектрофотометра для чтения планшетов Sunrise (Tecan). Данные анализировали с использованием аппроксимации 4-параметрической логистической кривой, и по кривым ингибирования устанавливали концентрации scFv к VEGF, необходимые для ингибирования пролиферации HUVEC на 50% (EC50).
ЭКВИВАЛЕНТЫ
Специалистам в данной области, принимая во внимание приведенное выше описание, очевидны многочисленные модификации и альтернативные варианты осуществления настоящего изобретения. Таким образом, это описание следует рассматривать только как иллюстративное и предназначенное для указания специалистам в данной области наилучшего способа осуществления настоящего изобретения. Подробности структуры по существу могут варьировать без отклонения от сущности изобретения, и резервируется исключительное использование всех модификаций, входящих в объем приложенной формулы изобретения. Следует понимать, что настоящее изобретение ограничено только до той степени, которая требуется приложенной формулой изобретения и применимыми правовыми нормами.
Все литературные и сходные документы, цитируемые в этой заявке, включая патенты, патентные заявки, статьи, книги, монографии, диссертации, веб-страницы, фигуры и/или приложения, вне зависимости от формата таких литературных и сходных документов, явным образом включены в полном объеме в качестве ссылки. В случае если один или несколько из включенных литературных и сходных документов отличается от данного описания или противоречит ему, включая определенные термины, использование терминов, описанные способы или т.п., следует руководствоваться настоящей заявкой.


























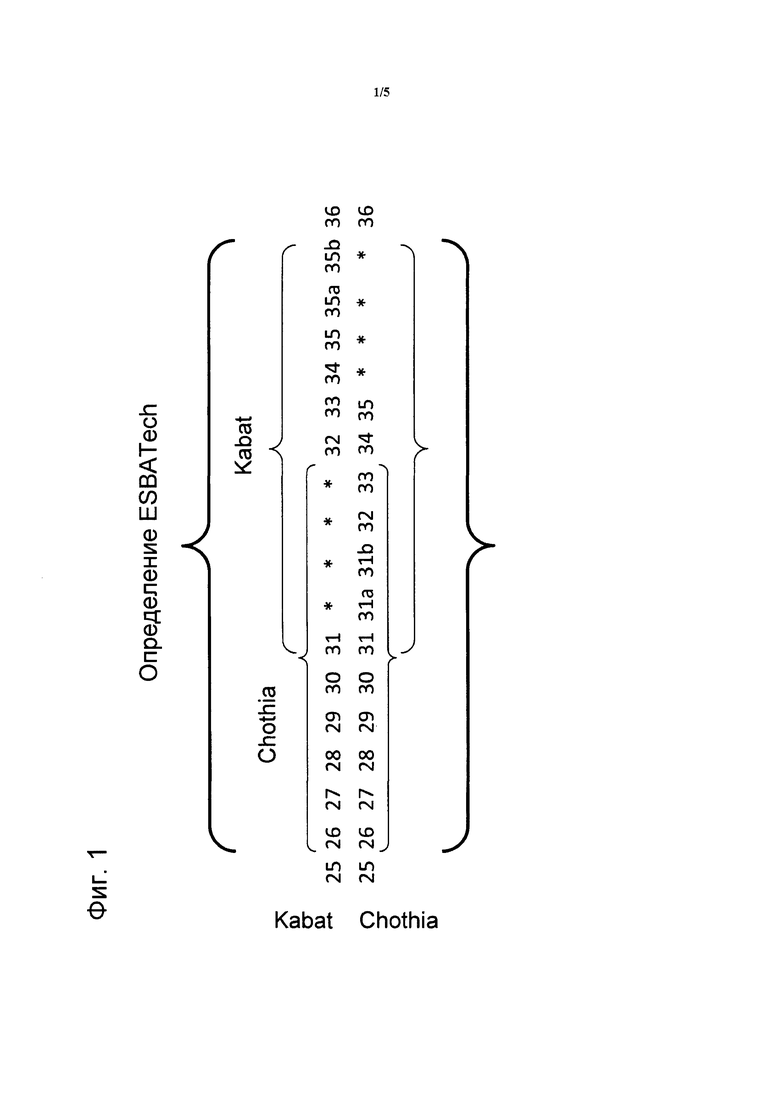
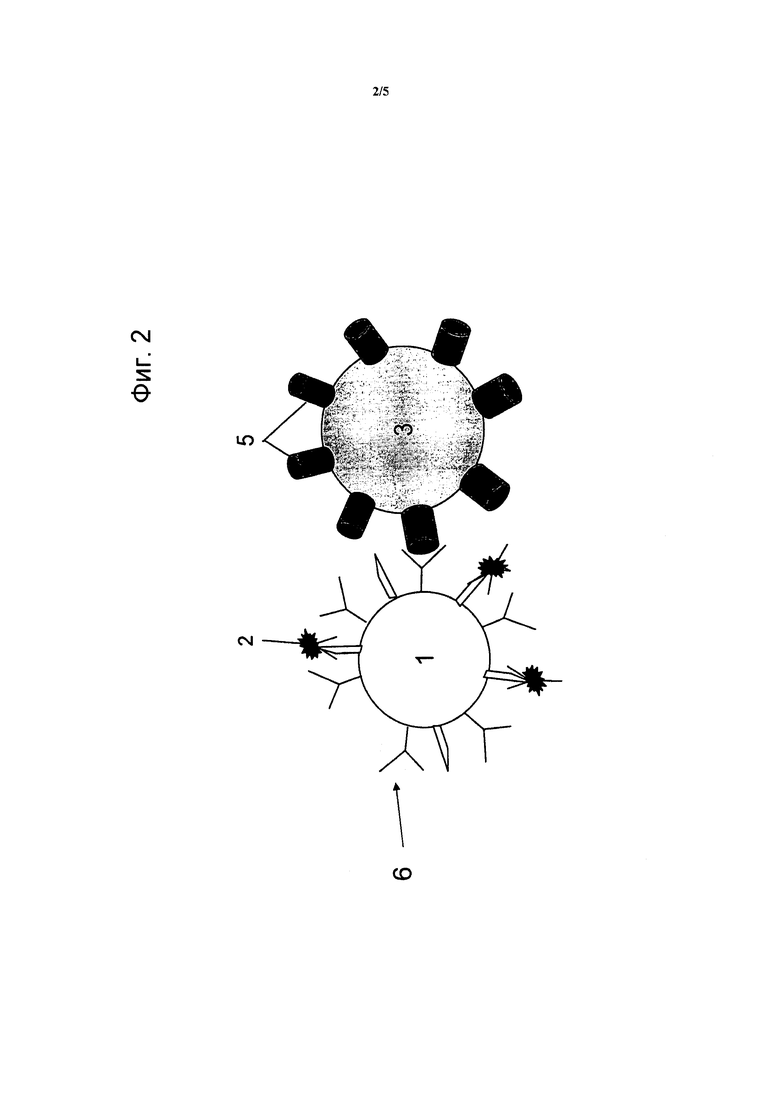
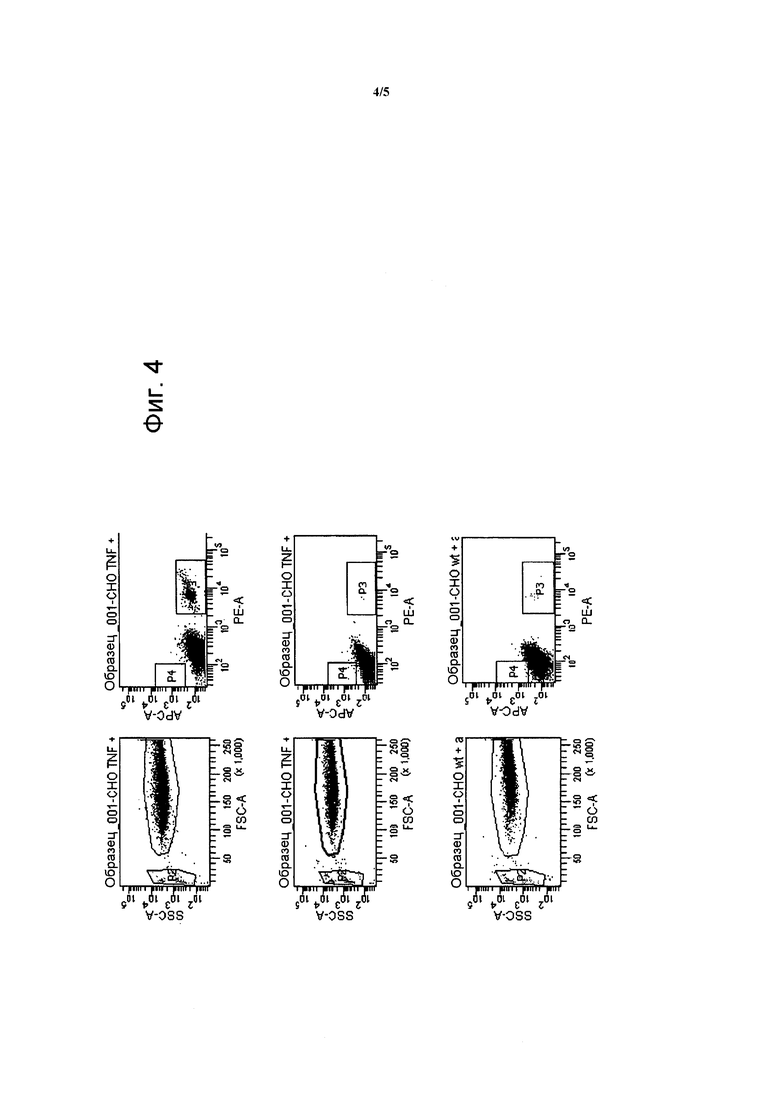
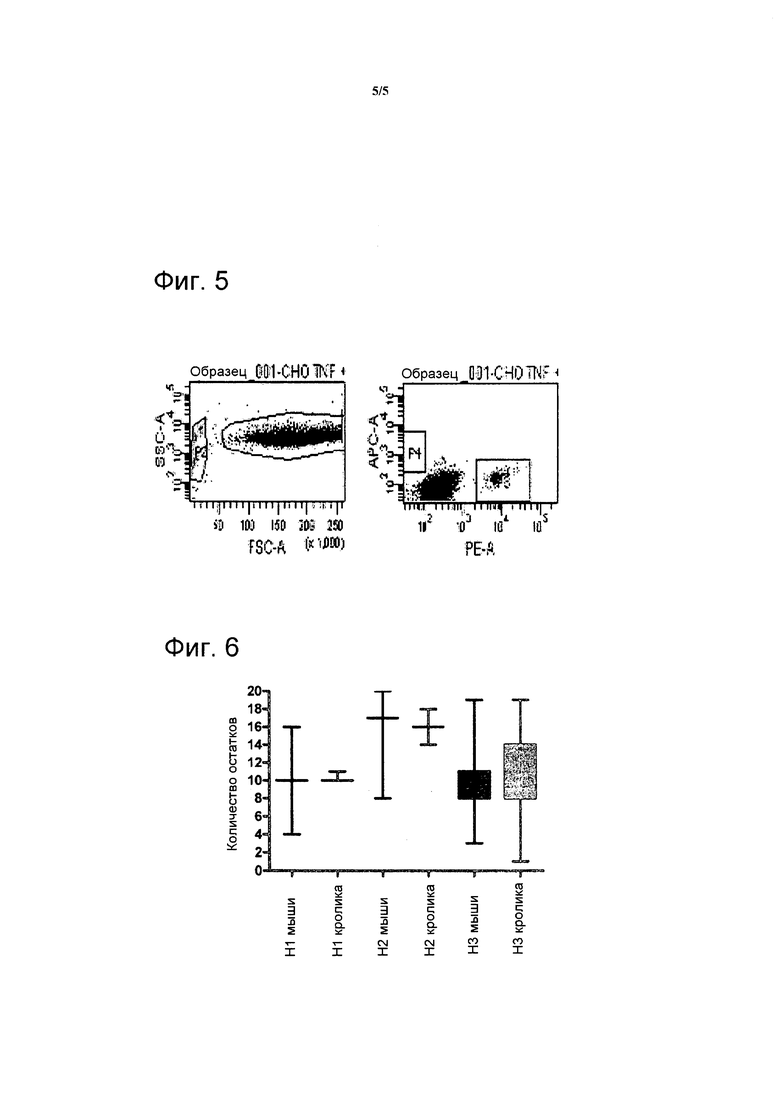
Изобретение относится к биотехнологии. Описана иммуносвязывающая молекула, содержащая: CDR1, CDR2 и CDR3 легкой цепи из донорной иммуносвязывающей молекулы зайцеобразного и CDR1, CDR2 и CDR3 тяжелой цепи из донорной иммуносвязывающей молекулы зайцеобразного, каркас вариабельной области легкой цепи человека и каркас вариабельной области тяжелой цепи человека, содержащий по меньшей мере четыре следующих аминокислоты: треонин (T) в положении 24, аланин (A) или глицин (G) в положении 56, треонин (T) в положении 84, валин (V) в положении 89 и аргинин (R) в положении 108 (нумерация AHo). Также представлена фармацевтическая композиция и биспецифическая иммуносвязывающая молекула, содержащие указанную молекулу. Антитела, получаемые способами по изобретению, пригодны для ряда диагностических и терапевтических применений. 3 н. и 19 з.п. ф-лы, 6 ил., 4 табл., 5 пр.
1. Иммуносвязывающая молекула, содержащая:
(i) CDR1, CDR2 и CDR3 легкой цепи из донорной иммуносвязывающей молекулы зайцеобразного и CDR1, CDR2 и CDR3 тяжелой цепи из донорной иммуносвязывающей молекулы зайцеобразного;
(ii) каркас вариабельной области легкой цепи человека; и
(iii) каркас вариабельной области тяжелой цепи человека, содержащий по меньшей мере четыре следующих аминокислоты: треонин (T) в положении 24, аланин (A) или глицин (G) в положении 56, треонин (T) в положении 84, валин (V) в положении 89 и аргинин (R) в положении 108 (нумерация AHo).
2. Иммуносвязывающая молекула по п.1, где аминокислотная последовательность каркаса вариабельной области тяжелой цепи по меньшей мере на 85% идентична последовательности SEQ ID NO:4 и содержит по меньшей мере четыре следующих аминокислоты: треонин (T) в положении 24, аланин (A) или глицин (G) в положении 56, треонин (T) в положении 84, валин (V) в положении 89 и аргинин (R) в положении 108 (нумерация AHo).
3. Иммуносвязывающая молекула по п.1, где указанный каркас вариабельной области тяжелой цепи содержит треонин (T) в положении 24 (нумерация AHo).
4. Иммуносвязывающая молекула по п.1, где указанный каркас вариабельной области тяжелой цепи содержит треонин (T) в положении 84 (нумерация AHo).
5. Иммуносвязывающая молекула по п.1, где указанный каркас вариабельной области тяжелой цепи содержит валин (V) в положении 89 (нумерация AHo).
6. Иммуносвязывающая молекула по п.1, где указанный каркас вариабельной области тяжелой цепи дополнительно содержит по меньшей мере одну из следующих аминокислот: серин (S) в положении 12; серин (S) или треонин (T) в положении 103; и серин (S) или треонин (T) в положении 144 (нумерация AHo).
7. Иммуносвязывающая молекула по п.1, где указанный каркас вариабельной области тяжелой цепи дополнительно содержит глицин (G) в положении 141 (нумерация AHo).
8. Иммуносвязывающая молекула по п.1, где указанный каркас вариабельной области тяжелой цепи содержит аминокислотную последовательность, которая выбрана из группы, состоящей из SEQ ID NO:4 и SEQ ID NO:6.
9. Иммуносвязывающая молекула по п.1, где указанный каркас вариабельной области легкой цепи содержит аминокислотную последовательность, которая по меньшей мере на 85% идентична последовательности SEQ ID NO:2.
10. Иммуносвязывающая молекула по п.9, где указанный каркас вариабельной области легкой цепи содержит SEQ ID NO:2.
11. Иммуносвязывающая молекула по п.9, где указанный каркас вариабельной области легкой цепи содержит треонин (T) в положении 87 (нумерация AHo).
12. Иммуносвязывающая молекула по п.11, где указанный каркас вариабельной области легкой цепи содержит SEQ ID NO:9.
13. Иммуносвязывающая молекула по п.9, где указанный каркас вариабельной области тяжелой цепи и указанный каркас вариабельной области легкой цепи связаны через линкерную последовательность, которая содержит SEQ ID NO:8.
14. Иммуносвязывающая молекула по п.9, содержащая аминокислотную последовательность, которая по меньшей мере на 85% идентична последовательности SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:7.
15. Иммуносвязывающая молекула по п.14, содержащая аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:3, SEQ ID NO:5 или SEQ ID NO:7.
16. Иммуносвязывающая молекула по п.9, где указанный каркас вариабельной области легкой цепи дополнительно содержит по меньшей мере одну из следующих аминокислот: глутаминовую кислоту (E) в положении 1, валин (V) в положении 3, лейцин (L) в положении 4, серин (S) в положении 10, аргинин (R) в положении 47, серин (S) в положении 57, фенилаланин (F) в положении 91 и валин (V) в положении 103 (нумерация AHo).
17. Иммуносвязывающая молекула по п.9, где указанная иммуносвязывающая молекула представляет собой scFv.
18. Фармацевтическая композиция для лечения состояний, реагирующих на иммуносвязывающую молекулу, содержащая иммуносвязывающую молекулу по п.1 и фармацевтически приемлемый эксципиент.
19. Иммуносвязывающая молекула по п.1, где каркас вариабельной области тяжелой цепи дополнительно содержит валин (V) в положении 25 и/или лизин (K) в положении 82 (нумерация AHo).
20. Биспецифическая иммуносвязывающая молекула, содержащая иммуносвязывающую молекулу по любому из пп.1-17 или 19.
21. Биспецифическая молекула по п.20, которая связывается по меньшей мере с двумя различными сайтами связывания или мишеневыми молекулами.
22. Биспецифическая молекула по п.20 или 21, где иммуносвязывающая молекула связана с антителом, фрагментом антитела (включая Fab, Fab’, F(ab’)2, Fv или одноцепочечный Fv), опухолеспецифичными или патогенспецифичными антигенами, пептидом или связывающим миметиком.
| ПРИБОР ДЛЯ ЗАПИСЫВАНИЯ ИЗМЕНЕНИЯ НАТЯЖЕНИЙ ГИБКИХ ТЯГ ВО ВРЕМЯ ПЕРЕВОДА СТРЕЛОК, СЕМАФОРОВ И Т. П. | 1925 |
|
SU7005A1 |
| МОДИФИЦИРОВАННЫЕ АНТИ-EGFR АНТИТЕЛА С УМЕНЬШЕННОЙ ИММУНОГЕННОСТЬЮ | 2002 |
|
RU2297245C2 |

