ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к антителам, которые направлены на EGF рецептор (HER1) для назначения главным образом человеку, и, в частности, для терапевтического использования при опухолях. Антитела представляют собой модифицированные антитела, в которых модификация приводит к уменьшенной склонности антитела вызывать иммунный ответ при введении человеку. Изобретение, в частности, относится к модификации анти-EGFR антитела 425 в его различных формах и их фрагментах, которая приводит к получению вариантов Mab 425, которые являются существенно неиммуногенными или менее иммуногенными, чем любой их немодифицированный эквивалент, при использовании in vivo. Изобретение также относится к пептидному эпитопу Т-клеток, который имеет происхождение от указанного немодифицированного антитела, с помощью которого возможно создать модифицированные варианты Mab 425 с уменьшенной иммуногенностью.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
Рецептор эпидермального фактора роста (EGF рецептор или EGFR), также известный как c-erbB I/Her 1, и продукт neu онкогена (также известный как с-erbB2/Her 2) являются членами суперсемейства EFG рецептора, которое принадлежит большому семейству рецепторов тирозинкиназы. Они взаимодействуют на поверхности клетки с определенными факторами роста или природными лигандами, такими как EGF или альфа TGF, таким образом активируя рецептор тирозинкиназы. Каскад нисходящего потока, сигнализирующих белков активируется, что в общем приводит к изменению экспрессии гена и увеличению скорости роста.
C-erbB2 (Her 2) - трансмембранная тирозинкиназа, имеющая молекулярную массу около 185000, со значительной гомологией с рецептором EGF (Her 1), хотя специфический лиганд к Her 2 пока еще четко не идентифицирован.
EGF рецептор представляет собой трансмембранный гликопротеин, который имеет молекулярную массу 170000 и найден во многих типах клеток эпителия. Он активируется по крайней мере тремя лигандами, EGF, TGF-α (преобразующий фактор роста альфа) и амфирегулином. Как было показано, эпидермальный фактор роста (EGF) так и преобразующий фактор роста альфа (TGF-α), связывается с EGF рецептором и приводит к клеточной пролиферации и росту опухоли. Эти факторы роста не связываются с Her 2 (Ulrich и Schlesinger, 1990, Cell 61, 203). В отличие от нескольких семейств факторов роста, которые вызывают димеризацию рецептора в силу их димерной природы (например, PDGF), мономерные факторы роста, такие как EGF, содержат два связующих места для их рецепторов, и, поэтому, могут кросс-связывать два соседних EGF рецептора (Lemmon и др., 1997, EMBO J. 16, 281). Димеризация рецептора является существенной для стимулирования внутренней каталитической активности и для автофосфорилирования рецепторов фактора роста. Следует отметить, что рецепторы протеин тирозинкиназ (PTKs) способны претерпевать как гомо-, так и гетеродимеризацию.
Клинические исследования показали, что как EGF рецептор так и C-erbB2 с избытком присутствуют в определенных типах опухолей, особенно, груди, яичника, пузыря, ободочной кишки, почки, рака головы и шеи, сквамозного рака легкого (Mendelsohn, 1989, Cancer Cells 7, 359; Mendelsohn, 1990, Cancer Biology 1, 339). Поэтому эти наблюдения стимулировали до клинические исследования, направленные на ингибирование функции EGF рецепторов человека или C-erbB2, как новые терапевтические подходы к лечению рака (см., например, Baselga и др., 1996, J. Clin. Oncol. 14, 737; Fan и Mendelsohn, 1998, Curr. Opin. Oncol. 10, 67). Сообщалось, что, например, антитела анти-EGF рецептора, а так же антитела анти-Her 2 показывают многообещающие результаты в терапии рака человека. Так, гуманизированное моноклональное антитело 4D5 (hMAb 4D5, HERCEPTIN®) - уже является продажным препаратом.
Было показано, что антитела анти-EGF рецептора, блокируя EGF и TGF-a связываясь с рецептором, подавляют пролиферацию опухолевой клетки. Принимая во внимание эти результаты, было произведено некоторое количество мышиных и крысиных моноклональных антител против EGF рецептора и проверена их способность подавлять рост опухолевых клеток in vitro и in vivo (Modjtahedi и Dean, 1994, J. Oncology 4, 277).
Гуманизированное моноклональное антитело 425 (hMAb 425) (US 5558864; ЕР 0531 472) и химерное моноклональное антитело 225 (cMAb 225) (Naramura и др., 1993. Cancer Immunol. Immunother. 37, 343-349.WO 96/40210), направленные на EGF рецептор, показали свою эффективность в клинических экспериментах.
С225 антитело продемонстрировало способность ингибировать EGF-опосредованный рост опухолевой клетки in vitro и подавлять формирование опухоли человека in vivo на бестимусных мышах. Антитело, кроме того, проявляет синергизм, главным образом, с некоторыми химиотерапевтическими средствами, (например, доксорубицин, адриамицин, таксол и цисплатин), чтобы ликвидировать опухоли человека in vivo на ксенотрансплантантных моделях мышей. Ye и др. (1999, Oncogene 18, 731) сообщили, что овариальные раковые клетки яичника человека могут успешно лечиться комбинацией cMAb 225 и hMAb 4D5.
Существует много случаев, при которых эффективность терапевтических белков ограничена нежелательной иммунной реакцией на терапевтический белок. Было показано, что некоторые мышиные моноклональные антитела являются перспективными в качестве терапевтических средств при лечении ряда заболеваний человека, однако в некоторых случаях они являются неудачными в связи с индукцией значительного уровня человеческого антимышиного антитела (НАМА) (SchroffR.W. и др. (1985), Cancer Res. 45: 879-885; Shawler D.L. и др. (1985) J.Immunol. 135: 1530-1535). Для моноклональных антител было предложено много способов уменьшения НАМА ответа (WO 89/09622; ЕР 0239400; ЕР 0438310; WO 91/06667). Эти подходы на основе рекомбинантной ДНК обычно ослабляют мышиную генетическую информацию при конечной конструкции антитела, усиливая тем самым генетическую информацию человека в полученной конструкции. Тем не менее, полученные "гуманизированные" антитела все же вызывают в ряде случаев иммунный ответ у пациентов (Issacs J.D. (1990) Sem.Immunol. 2: 449, 456: Rebello, P.R. и др. (1999) Transplantation 68: 1417-1420).
Антитела не являются единственным классом полипептидных молекул, которые назначаются в качестве терапевтического агента и против которых может развиваться иммунный ответ. Даже белки человеческого происхождения и те, которые имеют существующие у человека аминокислотные последовательности, могут индуцировать иммунный ответ у человека. Показательные примеры включают терапевтическое использование колониестимулирующего фактора гранулоцитов-макрофагов (Wadhwa M. и др. (1999) din. Cancer. Res. 5.: 1353-1361) и интерферон альфа-2 (Russo D. и др. (1996) Bri. J. Haem. 94: 300-305; Stein R. и др. (1998) New Engl. J. Med. 318: 1409-1413) среди прочих.
Принципиальным фактором в индукции иммунного ответа является присутствие в белке пептидов, которые могут стимулировать активность Т-клеток через презентацию молекул класса II МНС, так называемые "эпитопы Т-клеток". Такие потенциальные эпитопы Т-клеток обычно определяют как любую последовательность аминокислотных остатков, которая обладает способностью связываться с молекулой класса II МНС. Такие эпитопы Т-клеток можно измерять для установления МНС-связывания. Конечно, "эпитоп Т-клеток" означает эпитоп, который при связывании с молекулами МНС может узнаваться рецептором Т-клеток (TCR) и который может, по крайней мере, в принципе вызывать активацию этих Т-клеток посредством сцепления TCR для улучшения Т-клеточного ответа. Это, однако, обычно понимается как то, что определенные пептиды, которые являются такими, что связываются с молекулами класса II МНС, могут удерживаться в белковой последовательности, поскольку такие пептиды узнаются как "собственные" для организма, в который вводится конечный белок.
Известно, что определенные пептиды Т-клеточных эпитопов могут высвобождаться во время разрушения пептидов, полипептидов или белков внутри клеток и, следовательно, принимать участие в презентации молекулами основного комплекса гистосовместимости (МНС) для того, чтобы инициировать активацию Т-клеток. Для пептидов, представленных классом II МНС такая активация Т-клеток может потом вызывать, например, ответ антител путем прямой стимуляции В-клеток для получения таких антител.
Молекулы класса II МНС представляют собой группу высокополиморфных белков, которые играют центральную роль в отборе хелперных Т-клеток и активации. Человеческая лейкоцитарная антигенная группа DR (HLA-DR) представляет собой преобладающий изотип этой группы белков и является базисной точкой данного изобретения. Однако, изотипы HLA-DQ и HLA-DP выполняют сходные функции, таким образом, данное изобретение равно применимо к этим двум изотипам. Молекулы класса II МНС DR понимаются как альфа- и бета-цепи, которые встроены своими С-терминальными концами в клеточную мембрану. Каждый гетеродимер обладает лиганд-связывающим доменом, который связывается с пептидами и имеет длину, варьирующую от 9 до 20 аминокислот, несмотря на то, что связывающий желобок может включать максимум 11 аминокислот. Лиганд-связывающий домен включает аминокислоты с 1 по 85 альфа-цепи, и аминокислоты с 1 по 94 бета-цепи. DQ-молекулы, как было показано недавно, имеют гомологичную структуру и DP семейство белков, как ожидается, также является очень подобным. У человека известно около 70 различных аллотипов DR изотипа, для DQ известно 30 различных аллотипов, а для DP - 47 различных аллотипов. Каждая особь несет от двух до четырех аллелей DR, два аллеля DQ и два DP аллеля. Структура ряда DR молекул изучена, и эти структуры указывают на связывающий желобок пептида с открытой концевой структурой с рядом гидрофобных карманов, которые вовлекают гидрофобные остатки (остатки кармана) пептида (Brown и др. Nature (1993) 364: 33, Stern и др. (1994) Nature 368: 215). Полиморфизм, идентифицирующий различные аллотипы класса II молекул вносит свой вклад в широкое разнообразие различных связывающих поверхностей для пептидов в пределах связывающего желобка пептида и на популяционном уровне обеспечивает максимальную гибкость в отношении способности узнавать чужеродные белки и вызывать иммунный ответ к патогенным организмам.
Существует значительный полиморфизм в пределах лиганд-связывающего домена с четкими "семействами" в пределах различных географических популяций и этнических групп. Этот полиморфизм влияет на характеристики связывания пептид-связывающего домена, и таким образом, различные "семейства" DR молекул будут иметь специфичность для пептидов с различными свойствами последовательности, несмотря на то, что они могут в некоторой степени перекрываться. Эта специфичность определяет узнавание Th-клеточных эпитопов (класс II Т-клеточного ответа), которые в конечном счете ответственны за стимулирование выработки антител к Р-клеточным эпитопам, которые присутствуют в том же белке, из которого происходит Th-клеточный эпитоп. Таким образом, иммунный ответ на белок в индивидуальном человеческом организме в значительной степени находится под влиянием узнавания Т-клеточного эпитопа, что является функцией пептид-связывающей специфичности такого индивидуального HLA-DR аллотипа. Таким образом, для того, чтобы идентифицировать Т-клеточные эпитопы в пределах белка или пептида в контексте глобальной популяции, желательно рассматривать связывающие свойства такого широкого набора HLA-DR аллотипов, насколько это возможно, покрывая таким образом настолько высокий процент мировой популяции, насколько это возможно.
Иммунный ответ на терапевтический белок, такой, как белок в соответствии с изобретением, проходит via путь презентации пептида класса II МНС. Тут экзогенные белки вовлекаются в процесс и подвергаются обработке для презентации в ассоциации с молекулами класса II МНС DR, DQ или DP типа. Молекулы класса II МНС экспрессируются профессиональными антиген-презентирующими клетками (АРС), такими, как макрофаги и дендритные клетки, среди прочих. Сцепление пептидного комплекса класса II МНС с родственным Т-клеточным рецептором на поверхности Т-клеток, вместе с перекрестным связыванием определенных других сорецепторов, таких, как CD4 молекула, может индуцировать активированное состояние в Т-клетках. Активация ведет к высвобождению цитокинов, активирующих в дальнейшем другие лимфоциты, такие как В-клетки, для продуцирования антител или активирующих клеток Т-киллеров в качестве полного клеточного иммунного ответа. Способность пептида связывать данные молекулы класса II МНС для презентации на поверхности АРС зависит от числа факторов, в особенности от их первичной последовательности. Это будет влиять как на их склонность к протеолитическому расщеплению, так и на их сродство к связыванию в пределах связывающего желобка молекулы класса II МНС. Комплекс класс II МНС/пептид на поверхности АРС представляет собой центральную часть связывания для частного рецептора Т-клеток (TCR), способного узнавать детерминанты, которые обеспечиваются как открытыми остатками пептида, так и молекулой класса II МНС.
В области техники существуют способы для идентификации синтетических пептидов, способных к связыванию молекул класса II МНС (например, WO 98/52976 и WO 00/34317). Такие пептиды могут не функционировать как эпитопы Т-клеток во всех ситуациях, в частности, in vivo, благодаря путям процессирования и другим явлениям. Идентификация эпитопа Т-клеток представляет собой первый этап удаления эпитопа. Идентификация и удаление потенциальных эпитопов Т-клеток из белков была раскрыта ранее. В области техники существуют способы для получения возможности определения эпитопов Т-клеток обычно с помощью вычислительных средств исследования для узнавания участка последовательности в экспериментально определенных эпитопах Т-клеток или, альтернативно, при использовании вычислительных способов для прогнозирования пептидов, связывающих класс II МНС и, в частности, DR-связывающих пептидов. WO 98/52976 и WO 00/34317 обеспечивают вычислительные подходы к идентификации полипептидных последовательностей с потенциалом для связывания подмножества человеческих DR аллотипов класса II МНС. В этих способах, спрогнозированные эпитопы Т-клеток вычленяются при использовании целесообразных аминокислотных замен в первичной последовательности терапевтического антитела или белка, который не представляет собой антитела, имеющих происхождение как от человека, так и нечеловеческого происхождения.
Другие способы, использующие растворимые комплексы рекомбинантных молекул МНС в комбинации с синтетическими пептидами, которые способны к связыванию с клонами Т-клеток образцов периферической крови человека или экспериментальных животных, применяются в области техники (Kern F. и др. (1998) Nature Medicine 4: 975-978; Kwok, W.W. и др. (2001) TRENDS in Immunology 22: 583-588) и могут использоваться в стратегии идентификации эпитопа.
Как показано выше, и как следствие этого, желательно идентифицировать и удалить или, по крайней мере, уменьшить количество Т-клеточных эпитопов из данных, в принципе, терапевтически ценных, но изначально иммуногенных иммуноглобулинов, полипептидов или белков.
Модифицированный Mab 425 был получен ранее (US 5558864; ЕР 0531472) точно также как и химерная и гуманизированная версии с225 (WO 96/40210), но эти подходы были направлены на уменьшение имуногенности путем получения химерных и гуманизированных версий указанных антител из мышиной формы стандартными методами. Однако ни один из этих методов не признает важности эпитопов Т-клеток для иммуногенных свойств белка, не рассматривает прямого влияния указанных свойств на специфический и контролированный путь в соответствии со схемой данного изобретения.
Тем не менее, существует потребность в анти-EGFR антителах с улучшенными свойствами. Желательные усовершенствования включают альтернативные схемы и модальности для экспрессии и очистки указанного терапевтического средства, а также в особенности, улучшения биологических свойств иммуноглобулина. Существует особая потребность в улучшении in vivo характеристик при назначении человеку. В этой связи особенно желательно обеспечить анти-EGFR антитела, особенно MAb 425, с уменьшенным или отсутствующим потенциалом индукции иммунного ответа в человеческом организме.
КРАТКОЕ ИЗЛОЖЕНИЕ И ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Данное изобретение обеспечивает модифицированные формы анти-EGFR антитела, особенно MAb 425, в которых иммунные характеристики модифицированы путем уменьшения или удаления числа потенциальных эпитопов Т-клеток.
Изобретение раскрывает последовательности, идентифицированные в пределах первичной последовательности MAb 425 и являющиеся потенциальными эпитопами Т-клеток на основании потенциала связывания класса II МНС.
Изобретение раскрывает также специфические положения в пределах первичной последовательности молекулы в соответствии с изобретением, которые должны быть изменены посредством замен аминокислот, добавлений или делеций, без затрагивания биологической активности в принципе. В случаях, когда потеря иммуногенности может быть достигнута только путем одновременной потери биологической активности или авидности/специфичности антитела, возможно восстановить указанные параметры путем дальнейших изменений в пределах аминокислотной последовательности варианта антитела.
Изобретение раскрывает также способы получения таких модифицированных антител, главным образом, идентификации эпитопов Т-клеток, которые должны быть изменены для того, чтобы уменьшить или устранить иммуногенетические сайты.
Модифицированное анти-EGFR антитело в соответствии с изобретением, как ожидается, будет демонстрировать увеличенное время циркуляции в человеке и будет особенно полезно при хронических или рецидивирующих заболеваниях, как, например, в случае показаний. Данное изобретение обеспечивает модифицированные формы белков указанного антитела, которые, как ожидается, будут демонстрировать улучшенные свойства in vivo. Эти модифицированные молекулы анти-EGFR антитела могут использоваться в фармацевтических композициях.
Вкратце изобретение относится к следующим результатам:
- модифицированному антителу или его фрагменту, направленному на EGF рецептор (Нег1), которое является существенно неиммуногенным или менее иммуногенным, чем любое исходно иммуногенно немодифицированное антитело, направленное на тот же рецептор, когда подвергнуто иммунной системе определенного вида и по сравнению с немодифицированным антителом, где модифицированное антитело - по сравнению с исходно немодифицированным антителом - не содержит или содержит меньшее количество последовательностей эпитопов Т-клеток и / или МНС аллотипов, обладающих способностью связывать пептиды, производные из указанного немодифицированного антитела;
- соответственно указанному модифицированному антителу, в котором указанные исходно присутствующие эпитопы Т-клеток являются лигандами класса II МНС или пептидными последовательностями, которые демонстрируют способность стимулировать или связывать Т-клетки посредством презентации на классе II;
- соответственно указанному модифицированному антителу, в котором 1-9 аминокислотных остатков, предпочтительно один аминокислотный остаток в любом из исходно присутствующих эпитопов Т-клеток, изменен(ы);
- соответственно указанному модифицированному антителу, в котором изменение аминокислотных остатков представляет собой замену, добавление или делецию исходно присутствующего(их) аминокислотного(ых) остатка(ов) другим(ми) аминокислотным(ми) остатком(ами) в специфическом(их) положении(ях);
- соответственно указанному модифицированному антителу, в котором далее дополнительно производится замена, добавление или делеция специфической(их) аминокислоты(от) для восстановления биологической активности указанной молекулы;
- соответственно указанному модифицированному антителу, в котором изменение аминокислот выполняют в отношении гомологичной последовательности белка и/или в соответствии со способами моделирования in sili со,
- соответственно указанному модифицированному антителу, в котором указанное исходно иммуногенно немодифицированное антитело имеет последовательности, полностью или частично нечеловеческого происхождения;
- соответственно указанному модифицированному антителу, в котором указанное исходно иммуногенно немодифицированное антитело является химерным антителом или нечеловеческим антителом, включающим поверхностные остатки кроме тех, которые не являются близкими к CDR областям, которые происходят от соответствующих структур последовательностей человека (венированное антитело);
- соответственно указанному модифицированному антителу, в котором указанное исходно иммуногенно немодифицированное антитело является мышиным MAb 425, предпочтительно содержащим любую из последовательностей как показано в Таблице 5;
- последовательности ДНК, которая кодирует тяжелую и /или легкую цепь модифицированного антитела, как указано выше;
- фармацевтической композиции, включающей модифицированное анти-EGFR антитело, как указано выше, необязательно вместе с фармацевтически приемлемым носителем, разбавителем или эксципиентом;
- соответствующей фармацевтической композиции или набору, включающему дополнительно фармакологически эффективное лекарственное средство, предпочтительно цитотоксическое средство, более предпочтительно химиотерапевтическое средство;
- способу производства модифицированного антитела или его фрагмента, направленного на EGF рецептор (Heri), которое является существенно неиммуногенным или менее иммуногенным, чем любое исходно иммуногенно немодифицированное антитело, направленное на тот же рецептор, когда подвергнуто иммунной системе определенного вида и по сравнению с немодифицированным антителом, и который включает следующие стадии: (i) определение аминокислотной последовательности тяжелой и /или легкой цепи исходно иммуногенно немодифицированного антитела или его части; (ii) идентификацию одного или более потенциальных эпитопов Т-клеток в пределах аминокислотной последовательности антитела любым способом, включающим определение связывания пептидов с молекулами МНС при использовании способов in vitro или in silico или биологических анализов; (iii) разработка новых вариантов последовательности с одной или более аминокислотами в пределах идентифицированных потенциальных эпитопов Т-клеток, модифицированных таким образом для существенного уменьшения или устранения активности эпитопов Т-клеток, как определено путем связывания пептидов с молекулами МНС при использовании способов in vitro или in silico или биологическими анализами, или путем связывания комплекса пептид-МНС с Т-клетками; (iv) конструирование таких вариантов последовательности с помощью техники рекомбинантных ДНК и тестирование указанных вариантов для того, чтобы идентифицировать один или более вариантов с желаемыми свойствами; и (v) необязательное повторение этапов (ii)-(iv);
- соответственно указанному способу, в котором стадию (iii) выполняют путем замены, добавления или делеции 1-9 аминокислотных остатков в любых исходно присутствующих эпитопах Т-клеток, необязательно в отношении гомологичной последовательности белка и/или в соответствии со способами моделирования in silico,
- соответственно указанному способу, в котором стадию (ii) выполняют с помощью следующих стадий: (а) выбора участка пептида, который имеет известную последовательность аминокислотных остатков; (b) последовательного отбора аминокислотных сегментов перекрывающихся участков аминокислотных остатков с предварительно определенным одинаковым размером и состоящих, по крайней мере, из трех аминокислотных остатков выбранного участка; (с) подсчета значения связывания молекулы класса II МНС для каждого выборочного сегмента путем суммирования определенных значений для каждой гидрофобной аминокислотной боковой цепи, которая присутствует в каждом выборочном сегменте аминокислотного остатка; и (d) идентификации, по крайней мере, одного из указанных сегментов, подходящих для модификации, основываясь на подсчитанной величине связывания молекулы класса II МНС этого сегмента для изменения общего значения связывания класса II МНС для пептида без существенного уменьшения терапевтической полезности пептида;
- соответственно указанному способу, в котором стадию (с) преимущественно выполняют при использовании функции подсчета Bohm, модифицированной для включения 12-6 составляющей отталкивания энергии комплекса лиганд-белок Ван дер Ваальса и составляющей конформационной энергии лиганда путем (1) обеспечения первой базы данных модели молекулы класса II МНС; (2) обеспечения второй базы данных возможного пептидного скелета для моделей молекул указанного класса II МНС; (3) отбора модели из указанной первой базы данных; (4) отбора возможного пептидного скелета из второй базы данных; (5) идентификации боковых цепей аминокислотных остатков, которые присутствуют в каждом выборочном сегменте; (6) определения значения связывающего сродства ко всем боковым цепям, которые присутствуют в каждом выборочном сегменте; и повторения стадий от (1) до (5) для каждой указанной модели и каждого указанного скелета;
- соответственно указанному способу, в котором используют исходно иммуногенно немодифицированное антитело, которое имеет последовательности, полностью или частично нечеловеческого происхождения;
- соответственно указанному способу, в котором указанное исходно иммуногенно немодифицированное антитело является химерным антителом или нечеловеческим антителом, включающим поверхностные остатки кроме тех, которые не являются близкими к CDR областям, которые происходят от соответствующих базовых структур последовательностей человека (венированное антитело);
- соответственно указанному способу, в котором указанное исходно иммуногенно немодифицированное антитело является мышиным антителом, предпочтительно мышиное MAb 425;
- использованию тринадцатимерного пептида эпитопа Т-клеток, предпочтительно по крайней мере 9 последовательных аминокислотных остатков указанного тринадцатимерного пептида эпитопа Т-клеток, имеющего потенциальную связывающую активность класса II МНС и созданного из иммуногенно немодифицированного антитела, для производства иммуногенно модифицированного антитела, которое существенно не имеет иммуногенности или имеет меньшую иммуногенность, при использовании in vivo, по сравнению с указанным немодифицированным антителом, каждое антитело определено выше и в формуле изобретения.
Общий способ в соответствии с данным изобретением, ведущий к модифицированным анти-EGFR антителам, включает следующие стадии:
(a) определение аминокислотной последовательности антитела или его части;
(b) идентификацию одного или более потенциальных эпитопов Т-клеток в пределах аминокислотной последовательности иммуноглобулина с помощью любого метода, включающего определение связывания пептидов с молекулами МНС при использовании способов in vitro или in silico или биологическими анализами;
(c) конструирование вариантов с новыми последовательностями с одной или более аминокислотами в пределах идентифицированных потенциальных эпитопов Т-клеток, модифицированных таким образом для существенного уменьшения или устранения активности Т-клеточного эпитопа, как определено связыванием пептидов с молекулами МНС при использовании способов in vitro или in silico или биологическими анализами. Такие варианты последовательностей создаются таким путем для избежания создания новых потенциальных эпитопов Т-клеток путем вариации последовательности, до тех пор, пока не будет существовать новых потенциальных эпитопов Т-клеток, в свою очередь, модифицированных таким путем для существенного уменьшения или устранения активности эпитопов Т-клеток;и
(d) конструирование таких вариантов последовательности с помощью способов рекомбинантной ДНК и тестирование таких вариантов для того, чтобы идентифицировать один или более вариантов с желаемыми свойствами в соответствии с хорошо известными способами рекомбинантных ДНК;
Идентификация потенциальных эпитопов Т-клеток в соответствии со стадией (b) может быть выполнена в соответствии со способами, описанными ранее в уровне техники. Приемлемые способы раскрыты в WO 98/59244; WO 98/52976; WO 00/34317; и могут быть предпочтительно использованы для идентификации предрасположенности пептидов, производных от ТРО, связываться с молекулой класса II МНС.
Другой очень эффективный способ для идентификации эпитопов Т-клеток путем вычисления описан в примере, который является предпочтительным воплощением в соответствии с изобретением.
На практике ряд вариантов иммуноглобулинов анти-EGFR, предпочтительно MAb 425, будет продуцироваться и тестироваться на желаемые иммунные и функциональные характеристики. Вариантные антитела будут наиболее предпочтительно продуцироваться с помощью техники рекомбинантных ДНК, несмотря на то, что другие процедуры, включающие химический синтез фрагментов антител, могут также рассматриваться.
Изобретение относится к аналогам антитела, в которых замены, по крайней мере, одного аминокислотного остатка сделаны в положениях, приводящих к существенному уменьшению активности или устранению одного или более потенциальных эпитопов Т-клеток из белка. Одна или более аминокислотных замен в определенных точках в пределах любых потенциальных лигандов класса II МНС, идентифицированные в Таблице 1, могут приводить к образованию MAb 425 с уменьшенным иммуногенным потенциалом при введении в качестве терапевтического средства для человека. Предпочтительно, когда аминокислотные замены делаются в приемлемых точках в пределах пептидной последовательности, предсказанной для достижения существенного уменьшения или устранения активности эпитопа Т-клеток. На практике приемлемая точка будет предпочтительно совпадать с аминокислотным остатком связывания в пределах одного из карманов, которые обеспечиваются в пределах желобка связывания класса II МНС.
Наиболее предпочтительно изменять связывание в пределах первого кармана углубления, в так называемых Р1 или Р1 анкерных положениях пептида. Качество связывающего взаимодействия между Р1 анкерным остатком пептида и желобком связывания первого кармана класса II МНС признается основной детерминантой общей связывающей аффинности для целого пептида. Приемлемая замена в этом положении пептида будет осуществляться для остатка, который является менее легко приспосабливаемым в пределах кармана, например, замена на более гидрофильный остаток. Аминокислотные остатки в пептиде в положениях, приравниваемых к связыванию внутри других карманных участков в пределах углубления связывания МНС, также учитываются и подпадают под объем настоящего изобретения.
Понятно, что единственные аминокислотные замены в пределах данного потенциального эпитопа Т-клеток являются более предпочтительным путем, посредством которого эпитоп может устраняться. Комбинации замен в пределах одного эпитопа могут предполагаться и, например, могут быть особенно приемлемыми в том случае, если индивидуально определенные эпитопы перекрываются друг с другом. Кроме того, аминокислотные замены или по отдельности в пределах данного эпитопа, или в комбинации в пределах единственного эпитопа могут быть сделаны в положениях, не приравниваемых к "остаткам кармана" в отношении связывающего желобка класса П МНС, но в любой точке в пределах пептидной последовательности. Замены могут быть сделаны в отношении гомологичных структур или структурным способом, который проводится при использовании способов in silico, известных в данной области техники, эти замены могут основываться на известных структурных признаках молекулы в соответствии с данным изобретением. Все такие замены подпадают под объем данного изобретения.
Могут предполагаться аминокислотные замены иные, чем в пределах пептидов, идентифицированных выше, в особенности тогда, когда они сделаны в комбинации с заменой(ами), осуществленными в пределах приведенного пептида. Например, может предполагаться изменение для восстановления структуры или биологической активности вариантной молекулы. Такие компенсаторные изменения и изменения для включения делеции или добавления особых аминокислотных остатков из анти-EGFR антитела, приводящие к образованию вариантов с желаемой активностью, и в комбинации с изменениями в любом из раскрытых пептидов подпадают под объем данного изобретения.
Поскольку изобретение относится к модифицированным анти-EGFR антителам, предпочтительно к MAb 425, композиции, содержащие такие модифицированные иммуноглобулины или их фрагменты их белков слияния, и родственные им композиции считаются такими, которые также подпадают под объем данного изобретения. В другом аспекте данное изобретение относится к нуклеиновым кислотам, кодирующим модифицированные анти-EGFR антитела, предпочтительно MAb 425. В еще одном аспекте данное изобретение относится к способам терапевтического лечения человека при использовании модифицированных иммуноглобулинов.
ДЕТАЛЬНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ:
Следует указать, что термин "уменьшенная или сниженная иммуногенность", используемый ранее и здесь далее является относительным термином и касается иммуногенности соответствующего первоначального исходного антитела, когда оно подвергается воздействию in vivo тем же видам по сравнению с антителом, модифицированным согласно изобретению. Таким образом, исходное антитело может быть полностью нечеловеческим антителом, таким как антитело мыши или крысы. Однако изобретение также касается антител, которые уже содержат последовательности человека, таких как химерные или гуманизированные антитела. В этих искусственно созданных антителах, полученных классическими стандартными методами, есть последовательности в пределах первичной структуры, которые являются все еще иммуногенными. Также возможно, что при создании таких химерных или гуманизированных версий известными методами получают новые эпитопы Т-клеток. Даже полностью человеческие антитела или их фрагменты могут быть иммуногенными для некоторых человеческих индивидуумов или групп индивидуумов, имеющих генетически иммунологически различную структуру. Термин "химерное антитело" означает антитела, у которых часть тяжелой и/или легкой цепи является идентичной с, или гомологична по отношению к соответствующим последовательностям в антителах, полученных от определенных видов или принадлежащих конкретному классу или подклассу антител, тогда как остаточный член цепочки(ек) идентичен с или гомологичен соответствующим последовательностям в антителах, полученных от других видов или принадлежащих другому классу или подклассу антител, также как и фрагменты этих антител, поскольку они проявляют ожидаемую биологическую активность (например, US 4816567; Morrison и др., Proc. Nat. Acad. Sci. USA, 81:6851-6855 (1984)). Методы для создания химерных и гуманизированных антител также известны. Например, методы для создания химерных антител включают описание в патентах Boss (Celltech) и Cabilly (Genentech) (US 4816397; US 4816567). "Гуманизированные антитела" являются формами химерных антител, полученных не от человека (например, грызун), которые содержат минимальную последовательность, полученную из не человеческого иммуноглобулина. В большей части гуманизированные антитела это иммуноглобулины человека (антитело реципиента), в котором остатки от гиперпеременной области (CDRs) реципиента заменены остатками из гиперпеременной области нечеловеческого вида (антитело донора), например мыши, крысы, кролика или примата, обладающего желательной специфичностью, сродством и способностью. В некоторых случаях рабочая область (FR) остатков иммуноглобулина человека заменена соответствующими нечеловеческими остатками. Вообще, гуманизированное антитело включает в себя по крайней мере один, а обычно два изменяемых домена, в которых все или почти все гиперпеременные петли соответствуют таковым в не человеческом иммуноглобулине и все или почти все FRs которые обладают последовательностями иммуноглобулина человека. Гуманизированное антитело необязательно также будет включать в себя, по крайней мере часть иммуноглобулина с постоянной областью (Fc), обычно из иммуноглобулина человека. Методы для создания гуманизированных антител описаны, например, Winter (US 5225539) и Boss (Celltech, US 4816397). "Фрагменты антитела" включают часть интактного антитела, предпочтительно, включая его антиген-связанную или переменную область. Примерами фрагментов антитела являются Fab, Fab', F(ab')2, Fv и Fc фрагменты, диатела, линейные антитела, молекулы одноцепочного антитела; и мультиспецифические антитела, образованные из фрагмента(ов) антитела. "Интактное" антитело - то, которое содержит антигенсвязывающую переменную область, а так же неизменный домен легкой цепи (CL) и неизменный домен тяжелой цепи, СН1, СН2 и СН3. Предпочтительно, интактное антитело имеет одну или более эффекторных функций. Усвоение папаина антител производит два идентичных антигенсвязывающих фрагмента, называемые "Fab" фрагменты, каждый включает единичный антигенсвязывающий участок и CL и СН1 область, и остаточный "Fc" фрагмент, название которого отражает его способность быстро кристаллизоваться. "Fc" область антитела включает, как правило, СН2, СН3 и область петли IgGI или IgG2 антитела основного класса. Обработка пепсином дает "F(ab')2" фрагмент, который имеет два антигенсвязывающих участка и все еще способен к антиген крое-сшиванию. "Fv" является минимальным фрагментом антитела, который содержит полный антиген-узнавающий и антигенсвязывающий участок. Эта область состоит из димера одной тяжелой цепи и одной легкой цепи переменного домена с плотной, нековалентной связью. "Одиночная цепь Fv" или "scFv" фрагментов антитела включает VH, и VL, домены антитела, где эти домены присутствуют в Единичной полипептидной цепи. Предпочтительно, Fv полипептид дальше включает полипептидный линкер между VH и VL доменами, которые позволяют scFv образовывать желаемую структуру для связывания антигена. Одноцепочные FV антитела известны, например, из Pluckthun (Pharmacology of Monoclonal Antibodies, Vol. 113, Rosenburg and Moore eds.. Springer-Verlag, New York, pp. 269-315 (1994)), WO 93/16185; US 5571894; US 5587458; Huston и др. (1988, Proc.Natl. Acad. Sci. 85, 5879) или Skerra и Plueckthun (1988, Science 240, 1038).
Термин "эпитоп Т-клеток" означает в соответствии с пониманием данного изобретения аминокислотную последовательность, которая способна связывать МСН II, способна стимулировать Т-клетки и/или также связывать (без неизбежной до определенной степени активации) с Т-клетками в комплексе с МСН II. Термин "пептид", как используется в контексте данной заявки и приложенных пунктах формулы, представляет собой соединение, которое содержит две или более аминокислот. Аминокислоты связаны вместе при помощи пептидной связи (определено ниже). Существует 20 различных природных аминокислот, вовлеченных в биологическую продукцию пептидов, и любое их количество может связываться в любом порядке для образования пептидной цепи или кольца. Существующие в природе аминокислоты, используемые в биологическом производстве пептидов, все имеют L-конфигурацию. Синтетические пептиды могут быть получены с применением приемлемых способов синтеза при использовании L-аминокислот, D-аминокислот, различных комбинаций аминокислот двух различных конфигураций. Некоторые пептиды содержат только несколько аминокислотных единиц. Короткие пептиды, например, имеющие менее чем десять аминокислотных единиц, иногда называют "олигопептидами". Другие пептиды содержат большое число аминокислотных остатков, например до 100 или более, и называются "полипептидами". По договоренности "полипептидом" можно считать любую пептидную цепь, которая содержит три или более аминокислот, в то время, как "олигопептид" обычно считается частным типом "короткого" полипептида. Таким образом, как используется в контексте данной заявки, понятно, что любое упоминание "полипептида" также включает и олигопептид. Кроме того, любая ссылка на "пептид" включает полипептиды, олигопептиды и белки. Каждое различное распределение аминокислот образует различные полипептиды или белки. Число полипептидов, и отсюда, число различных белков, которые могут быть образованы, является практически неограниченным. Термин "модифицированный белок/антитело", как используется в контексте данного изобретения, описывает белок/антитело, которое имеет уменьшенное количество эпитопов Т-клеток и поэтому выявляет сниженную иммуногенность, по сравнению с родительским белком, когда подвергается иммунной системе определенных видов. Термин "немодифицированный белок", как используется в данном изобретении, описывает "родительский" белок, который в сравнении с "модифицированным белком" имеет большее количество эпитопов Т-клеток и, поэтому увеличенную иммуногенность относительно модифицированного белка, когда подвергается иммунной системе определенных видов. "Альфа-углерод (Сα)" представляет собой углеродный атом углеводородного (СН) компонента, который находится в пептидной цепи. "Боковая цепь" представляет собой подвешенную группу к Сα, которая может включать простую или сложную группу или остаток, и которая имеет физические размеры, которые могут значительно отличаться по сравнению с размерами пептида.
В последующих разделах, изобретение описано более подробно для моноклонального анти-EGFR антитела 425, которое, как было показано, имеет большую терапевтическую ценность. Однако изобретение не ограничено этим антителом и его несколькими существующими формами и может быть расширено на другие анти-EGFR антитела, прежде всего химерное антитело 225, которое имеет очень подобные свойства.
Если не указано иначе, все аминокислоты в переменной тяжелой и легкой цепях нумеруются как в Kabat и др., 1991 (Sequences of Proteins of Immunological Interest, US Department of Health and Human Services). Потенциальные эпитопы Т-клеток нумеруются с линейным номером первой аминокислоты эпитопа, считая от первой аминокислоты тяжелой и легкой цепей.
1. Сравнение со Структурами Подгруппы Мыши
Аминокислотные последовательности мышиных 425 VH (тяжелая цепь) и VK (легкая цепь) были сравнены с последовательностями общих типичных элементов структуры для Kabat мышиной тяжелой и легкой подгрупп цепи. 425 VH может быть отнесен к подгруппе ИВ тяжелых цепей мыши. Сравнение с последовательностью общих типичных элементов структуры для этой подгруппы показывает, что серин для положения 94 в 425 VH является несвойственным. Самый распространенный остаток для этого положения - аргинин. 425 VK может быть отнесен к подгруппе VI каппа цепей мыши. Сравнение с последовательностью общих типичных элементов структуры для этой подгруппы показывает, что остатки в положениях 45-47, 60 и 100 в 425 VK являются несвойственными для этой подгруппы. Аминокислотные остатки пронумерованы как в Kabat.
2. Сравнение со Структурами Человека
Аминокислотные последовательности мышиных 425 VH (переменная тяжелая цепь) и VK (переменная каппа легкая цепь) были сравнены с последовательностями справочника зародышевых линий человека (germline) VH (Tomlinson, I.M и др., (1992) J. Mol. Biol. 227: 776-798) и VK (Cox, J.P.L. и др., (1994) Eur. J. Immunol. 24:827-836) последовательностями, а также с областью последовательностей зародышевых линий J человека (Routledge, E.G. и др., в Protein Engineering of Antibody Molecules/or Prophylactic and Therapeutic Applications in Man, dark, M. ed. Academic Titles, Nottingham, UK, pp. 13-44, 1991). Мышиные 425 последовательности тяжелых и легких цепей могут быть взяты, например, из ЕР 0531472.
Структурой человека, выбранной для 425 VH, на которую ссылались, была VH1GRR с JH6 человека. Последовательность VH1GRR в справочнике заканчивается на остатке 88. Поэтому нет никакого соответствующего остатка для несвойственного сирина в положении 94 из мышиной последовательности. Эта последовательность зародышевой линии была найдена в перестроенном зрелом гене антитела с изменениями 4 аминокислот. Структурой человека, выбранной для 425 VK, на которую ссылались, была L6/vg с JK2 человека. Эта последовательность зародышевой линии была найдена в перестроенной зрелой тяжелой цепи антитела без изменений аминокислот.
3. Разработка "венированных" ("veneered") последовательностей
После идентификации базовых последовательностей структуры человека некоторые неидентичные аминокислотные остатки в пределах 425 VH и VK структур были изменены на соответствующую аминокислоту в базовых последовательностях структуры человека. Остатки, которые, как полагают, являются критическими для структуры антитела и связывания, были исключены из этого процесса и не изменялись. Мышиные остатки, которые были сохранены на этой стадии, являлись в значительной степени неповерхностными, внутренними остатками, кроме остатков на N-концах, например, которые являются близкими к CDR-ам в целевом антителе (1-8, предпочтительно 1-5 аминокислотных остатков). Этот процесс приводит к последовательности, которая является определенно подобной "венированному" антителу, поскольку поверхностные остатки являются главным образом человеческими, а внутренние остатки, такими как в первоначальной мышиной последовательности.
4. Пептид пронизывающий анализ
Мышиные и венированные 425 VH и VK последовательности были проанализированы с использованием способа согласно изобретению. Последовательности аминокислот были разделены на все возможные тринадцатимеры. Тринадцатимерные белки последовательно представлены моделям связывающей бороздки аллотипов HLA-DR и значения связывания соотносились каждому пептиду для каждой аллели. Было рассчитано конформационное значение для каждой карманой-связи боковой цепи пептида. Это значение базируется на стерическом наложении, потенциальных водородных связях между белком и остатками в связывающей бороздке, электростатических взаимодействиях и благоприятных контактах между белком и остатками карманна. Конформацию каждой боковой цепи изменили, значение повторно рассчитали. Определив самое высокое конформационное значение, пересчитывали значение связывания основываясь на бороздковой-связи гидрофобных остатков, небороздковых гидрофильных остатках и количестве остатков, которые умещаются в бороздку связывания. Пептиды, которые являются известными связывающими класса II МНС человека, достигают высокого значения связывания почти без неверных результатов. Таким образом, пептиды, которые достигают значительного значения связывания в текущем анализе, как полагают, являются потенциальными эпитопами Т-клеток. Результаты пептид пронизывающего анализа показаны в Таблице 1 для 425 VH и 425 VK. Потенциальные эпитопы Т-клеток указаны линейным номером первого остатка тринадцатимера.
Таблица 1: Потенциальные эпитопы Т-клеток в мышиной и венированной 425 последовательностях
75(10), 92(10), 93(17)
5. Удаление потенциальных эпитопов Т-клеток
Аминокислотные остатки для замены пронумерованы как в Kabat.
Потенциальные эпитопы Т-клеток указаны линейным номером первого остатка тринадцатимера.
Необходимые замены аминокислот для удаления потенциальных эпитопов Т-клеток из венированной 425 областей переменной тяжелой цепи были следующие:
- Замена пролина на аланин в остатке 41 (Kabat номер 41) удаляет потенциальный эпитоп в остатке номер 31.
- Замена пролина на лейцин в остатке 45 (Kabat номер 45) удаляет потенциальный эпитоп в остатке номер 43. Пролин в положении 45 найден в человеческой последовательности VH зародышевой линии, DP52.
- Замена аланина на изолейцин в остатке 48 (Kabat номер 48) удаляет потенциальный эпитоп в остатке номер 46.
- Замена валина на аланин в остатке 68 (Kabat номер 67) удаляет потенциальный эпитоп в остатке номер 58.
- Замена изолейцина на лейцин в остатке 70 (Kabat номер 69) удаляет потенциальный эпитоп в остатке номер 62.
- Замена треонина на серин в остатке 91 (Kabat номер 87) удаляет потенциальные эпитопы в остатках под номерами 81 и 84.
Необходимые замены аминокислот для удаления потенциальных эпитопов Т-клеток из венированной 425 областей переменной легкой цепи были следующие:
- Замена гистидина на тирозин в остатке 35 (Kabat номер 36) удаляет потенциальный эпитоп в остатке номер 27.
- Замена аланина на треонин в остатке 50 (Kabat номер 51) удаляет потенциальный эпитоп в остатке номер 43. Этот остаток является частью CDR2. Аланин обычно находится в этом положении и в человеческих и в мышиных антителах. Альтернативной заменой для удаления этого эпитопа является замена аланина на лейцин в положении 45 (Kabat номер 46). Нет никакой предохраняющей замены, которая устранит потенциальный эпитоп. Аланин находится в этом положении в некоторых антителах.
- Замена пролина на изолейцин в остатке 94 (Kabat номер 95) удаляет потенциальный эпитоп в остатке номер 92. Kabat остаток 95 является частью CDRL3. Пролин является общим в этом положении в последовательностях антитела мыши и нет никакого изменения с CDR, которое устраняет потенциальный эпитоп.
- Замена валина на лейцин в остатке 103 (Kabat номер 104) удаляет потенциальный эпитоп в остатке номер 93.
6. Разработка деиммунизированных последовательностей
Деиммунизированные последовательности области переменной тяжелой и легкой цепи были разработаны с учетом замен, требуемых для удаления потенциальных эпитопов Т-клеток и анализа структуры остатков, которые могли бы быть критичными для структуры антитела и связывания. В дополнение к деиммунизированным последовательностям, которые основаны на венированной последовательности, дополнительная последовательность была разработана для каждого VH и VK, основанного на мышиной последовательности, названные Белок Мыши Прошитая (Мо РТ) версия. Для этой версии замены были сделаны непосредственно в мышиной последовательности, чтобы устранить эпитопы Т-клеток, но изменения сделаны только с CDR-ами, которые не рассматривают как нежелательные для закрепления. Никакой попытки удалять поверхностные (В-клетки) эпитопы не было сделано для этой версии деиммунизированной последовательности.
Первичная деиммунизированная VH включает замены 1-6, описанные в разделе 5 выше, и не включает никаких потенциальных эпитопов Т-клеток. Дополнительные 4 деиммунизированных последовательности VH были разработаны, чтобы проверить эффект необходимых различных замен на связывание антитела. Совокупные изменения сделаны для первой деиммунизированной последовательности (425 VHlGRR-VH-vl) и остающиеся потенциальные эпитопы Т-клеток отображены в Таблице 2. Мышиная прошитая версия включена для сравнения.
Таблица 2: Аминокислотные замены и потенциальные эпитопы в деиммунизированном 425 VH
Первичная деиммунизированная VK включает замены 1-4, описанные в разделе 5 выше, и не включает никаких потенциальных эпитопов Т-клеток. Дополнительные 4 деиммунизированных последовательности VK были разработаны, чтобы проверить эффект необходимых различных замен на связывание антитела. Версия 2 является альтернативой Версии 1, в которой использовалась альтернативная замена, чтобы удалить тот же самый потенциальный эпитоп Т-клетки. Совокупные изменения сделаны для первой деиммунизированной последовательности (425 L6-vg-VK-vl) и остающиеся потенциальные эпитопы Т-клеток отображены в Таблице 3. Мышиная прошитая версия включена для сравнения.
Таблица 3: Аминокислотные замены и потенциальные эпитопы в деиммунизированном 425 VK
Таблица 4: первоначальные и "венированные" последовательности VH и VK мышиной MAb 425
425 VH мышиная
QVQLQQPGAELVKPGASVKLSCKASGYTFTSHWMHWVKQRAGQGLEWIGEFNPS NGRTNYNEKFK SKATLTVDKSSSTAYMQLSSLTSEDSAVYYCASRDYDYDGRYFDYWGQGTTLTV
ss
425 VK мышиная
QIVLTQSPAIMSASPGEKVTMTCSASSSVTYMYWYQQKPGSSPRLLIYDTSNLASG
VPVRFSGSG
SGTSYSLTISRMEAEDAATYYCQQWSSHIFTFGSGTKLEIK
425 VH венированная:
QVQLVQSGAELVKPGASVKLSCKASGYTFTSHWMHWVKQAAGQGLEWIGEFNPS
NGRTNYNEKFK
SRATLTVDKSTSTAYMQLSSLTSEDSAVYYCASRDYDYDGRYFDYWGQGTTLTV
ss
425 VK венированная:
QIVLTQSPATLSASPGERATMSCSASSSVTYMYWYQQKPGQSPRLLIYDTSNLASG VPARFSGSG SGTSYTLTISSLEAEDAATYYCQQWSSHIFTFGQGTKLEIK
Таблица 5: деиммунизированные последовательности переменных тяжелой и легкой цепи MAb 425
425 деиммунизированная VH1
QVQLVQSGAELVKPGASVKLSCKASGYTFTSHWMHWVKQAPGQGPEWAGEFNP SNGRTNYNEKFK
SRVTITVDKSTSTAYMQLSSLTSEDTAVYYCASRDYDYDGRYFDYWGQGTTLTVS S
425 деиммунизированная VK1
QIVLTQSPATLSASPGERATMSCSASSSVTYMYWHQQKPGQSPRLLIYDASNLASG VPARFSGSG SGTSYTLTISSLEAEDAATYYCQQWSSHPFTFGQGTKVEIK 425 деиммунизированная VH2
QVQLVQSGAELVKPGASVKLSCKASGYTFTSHWMHWVKQAPGQGPEWIGEFNPS NGRTNYNEKFK
SRVTITVDKSTSTAYMQLSSLTSEDTAVYYCASRDYDYDGRYFDYWGQGTTLTVS S
425 деиммунизированная VK2
QIVLTQSPATLSASPGERATMSCSASSSVTYMYWHQQKPGQSPRALIYDTSNLASG VPARFSGSG
SGTSYTLTISSLEAEDAATYYCQQWSSHPFTFGQGTKVEIK 425 деиммунизированная VH3
QVQLVQSGAELVKPGASVKLSCKASGYTFTSHWMHWVKQAPGQGLEWIGEFNPS NGRTNYNEKFK
SRVTITVDKSTSTAYMQLSSLTSEDTAVYYCASRDYDYDGRYFDYWGQGTTLTVS
S
425 деиммунизированная VK3
QIVLTQSPATLSASPGERATMSCSASSSVTYMYWHQQKPGQSPRLLIYDTSNLASG
VPARFSGSG
SGTSYTLTISSLEAEDAATYYCQQWSSHPFTFGQGTKVEIK
425 деиммунизированная VH4
QVQLVQSGAELVKPGASVKLSCKASGYTFTSHWMHWVKQAPGQGLEWIGEFNPS
NGRTNYNEKFK
SRATLTVDKSTSTAYMQLSSLTSEDTAVYYCASRDYDYDGRYFDYWGQGTTLTV
ss
425 деиммунизированная VK4
QIVLTQSPATLSASPGERATMSCSASSSVTYMYWHQQKPGQSPRLLIYDTSNLASG
VPARFSGSG
SGTSYTLTISSLEAEDAATYYCQQWSSHIFTFGQGTKVEIK
425 деиммунизированная VH5
QVQLVQSGAELVKPGASVKLSCKASGYTFTSHWMHWVKQAAGQGLEWIGEFNPS
NGRTNYNEKFK
SRATLTVDKSTSTAYMQLSSLTSEDTAVYYCASRDYDYDGRYFDYWGQGTTLTV
SS
425 деиммунизированная VK5
QIVLTQSPATLSASPGERATMSCSASSSVTYMYWYQQKPGQSPRLLIYDTSNLASG
VPARFSGSG
SGTSYTLTISSLEAEDAATYYCQQWSSHIFTFGQGTKVEIK
425 VH мышиная, белок прошитая (Мо РТ)
QVQLQQPGAELVKPGASVKLSCKASGYTFTSHWMHWVKQAPGQGLEWIGEFNPS
NGRTNYNEKFK
SRVTITVDKSSSTAYMQLSSLTSEDTAVYYCASRDYDYDGRYFDYWGQGTTLTVS
S
425 VK мышиная, белок прошитая (Мо РТ)
QIVLTQSPATLSASPGEKATMTCSASSSVTYMYWYQQKPGSSPRLLIYDTSNLASG
VPVRFSGSG
SGTSYSLTISRLEAEDAATYYCQQWSSHIFTFGQGTKVEIK
Как уже было указано, модифицированные анти-EGFR антитела, согласно изобретению, предпочтительно MAb 425, могут быть предпочтительно использованы в фармацевтических композициях и фармацевтических наборах для лечения рака. Термин "рак" и "опухоль" относится к или описывает физиологическое состояние млекопитающих, которое обычно характеризуется нерегулируемым ростом клетки. Посредством фармацевтических композиций, в соответствии с настоящим изобретением, могут лечиться опухоли такого типа:
опухоли груди, сердца, легкого, тонкой кишки, ободочной кишки, селезенки, почки, пузыря, головы и шеи, яичника, простаты, мозга, поджелудочной железы, кожи, кости, костного мозга, крови, тимуса, матки, яичников, гортани и печени.
Фармацевтические композиции изобретения могут включать средства, которые уменьшают или исключают побочные факторы, связанные с комбинированной терапией данного изобретения ("добавочная терапия"), включая, но не ограничиваясь, теми средствами, например, которые уменьшают токсическое действие антираковых препаратов, например, ингибиторы поглощения кости, кардиопротекторные средства. Упомянутые добавочные средства предотвращают или уменьшают побочные эффекты, тошноту и рвоту, связанную с химиотерапией, лучевой терапией или операцией, или уменьшают действие инфекции, связанной с назначением миелосупрессорных антираковых препаратов. Добавочные средства известны из уровня техники. Модифицированные антитела средства, согласно изобретению, могут дополнительно назначаться со стимуляторами, такими как BCG и другими иммунностимуляторами. Кроме того, композиции могут включать химиотерапевтические средства, описанные выше, которые содержат цитотоксически эффективные радиомеченные изотопы или другие цитотоксические средства, такие как цитотоксические пептиды (например, цитокинины) или цитотоксические препараты и т.п. Фармацевтический набор для лечения опухолей или метастаз опухоли относится к упаковке и, как правило, инструкции использования реагентов в методах лечения опухолей и метастаз опухоли. Реактив в наборе этого изобретения обычно сформулирован как терапевтическая композиция, как описано здесь, и поэтому может быть в любых различных формах, подходящих для размещения в наборе. Такие формы могут включать жидкость, порошок, таблетку, суспензию и подобный состав для обеспечения антагониста и/или белок слияния данного изобретения. Реактивы могут находиться в отдельных контейнерах, подходящих для назначения отдельно, согласно данным методам, или альтернативно могут быть объединены в композиции в отдельном контейнере в упаковке. Упаковка может содержать количество, достаточное для одной или более дозировок реагента, согласно методам лечения, описанным здесь. Набор этого изобретения также содержит "инструкцию для использования" материалов, содержавшихся в упаковке. Фармацевтически приемлемые носители, разбавители и эксципиенты, а также их грамматические вариации, являются материалами допустимыми к применению млекопитающим без проявления нежелательных физиологических последствий, таких как тошнота, головокружение, желудочное расстройство и т.п.
Приготовление фармакологической композиции, которая содержит активные ингредиенты, в растворенном или диспергированном виде понятны в технологии и не должны ограничиваться на основании состава. Как правило, такие композиции приготовлены как инъекционные или как жидкие растворы или суспензии, однако, твердые формы, подходящие для раствора, или суспензии могут также быть приготовлены в виде раствора до использования. Препарат может также быть в виде эмульсии. Активный ингредиент может быть смешан с эксципиентами, которые являются фармацевтически приемлемыми и совместимыми с активным ингредиентом и в количествах, подходящих для использования в терапевтических методах, описанных здесь. Подходящими эксципиентами являются, например, вода, солевой раствор, декстроза, глицерин, этанол или подобные и их комбинации. Кроме того, если необходимо, композиция может содержать незначительные количества вспомогательных веществ, таких как смачивающие или эмульгирующие агенты, буфферные средства и т.п., которые усиливают эффективность активного ингредиента. Терапевтическая композиция данного изобретения может включать фармацевтически приемлемые соли компонентов.
Как правило, терапевтически эффективное количество антитела анти-ErbB - это такое количество, применяемое в физиологически приемлемой композиции, которое является достаточным, чтобы достигнуть концентрации в плазме приблизительно от 0,01 микрограмма (мкг) в миллилитре (мл) до приблизительно 100 мкг/мл, предпочтительно, от приблизительно 1 мкг/мл приблизительно до 5 мкг/мл и обычно около 5 мкг/мл. Дозировка может быть назначена по-другому и изменяться приблизительно от 0,1 мг/кг до приблизительно 300 мг/кг, предпочтительно, приблизительно от 0,2 мг/кг до приблизительно 200 мг/кг, наиболее предпочтительно приблизительно от 0,5 мг/кг до приблизительно 20 мг/кг, в одной или большем количестве доз введения ежедневно в течение одного или нескольких дней. Как правило, меньшая дозировка, как указано выше, может быть применена с достижением той же эффективности, если иммуногенно модифицированные антитела согласно изобретению применяют вместо соответствующих немодифицированных версий.
В случаях комбинированной терапии, например с химиотерапевтическим средством, необходимая или рекомендуемая обычная доза такого активного ингредиента составляет от 10 мг до 1000 мг, предпочтительно, приблизительно 20-200 мг, и более предпочтительно, 50-100 мг на килограмм веса тела в сутки.
Следующий пример в общей форме описывает способ идентификации согласно изобретению эпитопов Т-клеток, присутствующих на исходном антителе с немодифицированным иммуногенным потенциалом. Тем не менее, идентификация указанных последовательностей эпитопов может быть выполнена известными методами, как описано выше.
ПРИМЕР
Существует целый ряд факторов, которые играют важную роль в определении общей структуры белка, полипептида или иммуноглобулина. Во-первых, пептидная связь, т.е. та связь, которая связывает вместе аминокислоты в цепи, является ковалентной связью. Эта связь является плоской по структуре, по сути замещенным амидом. "Амид" представляет собой любую группу органических соединений, содержащих группировку -CONH-.
Плоская пептидная связь, соединяющая Сα соседних аминокислот, может быть представлена так, как показано ниже:
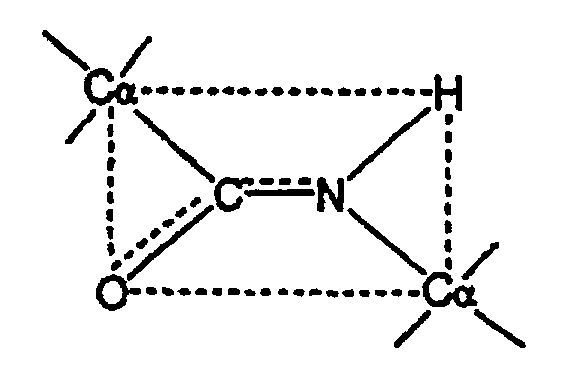
Поскольку O=С и C-N атомы лежат в относительно неподвижной плоскости, свободное вращение не происходит вокруг этих осей. Следовательно, плоскость, схематически показанная пунктирной линией, иногда называется как "амидная" или "пептидная плоскость", в которой лежат атомы кислорода (О), углерода (С), азота (N) и водорода (Н) пептидного скелета. В противоположных углах этой 30 амидной плоскости располагаются Сα атомы. Поскольку не происходит существенного вращения вокруг O=С и C-N атомов в пептидной или амидной плоскости, полипептидная цепь, таким образом, включает серию плоскостных пептидных связей, соединяющих Сα атомы. Вторым фактором, который играет важную роль в определении общей структуры или конформации полипептида или белка, является угол вращения каждой амидной плоскости вокруг общей Со, связи. Термин "угол вращения" и "угол закручивания" употребляются в дальнейшем как эквивалентные термины. Предполагая, что атомы О, С, N и Н остаются в амидной плоскости (что обычно является реальным предположением, несмотря на то, что могут существовать некоторые слабые отклонения от плоскости этих атомов для некоторых конформации), эти углы вращения определяют N и R конформацию полипептидного скелета, то есть структуру в том виде, в котором она существует между смежными остатками. Эти два угла известны, как φ и ψ. Набор углов φ1 и V1, где индекс 1 представляет собой отдельный остаток полипептидной цепи, таким образом, эффективно определяет вторичную структуру полипептида. Договоренности, используемые в определении φ и ψ углов, то есть базисные точки, в которых амидная плоскость образует угол, равный нулю градусов, и определение того, что представляет собой угол φ, и что представляет собой угол ψ, для данного полипептида, приведено в литературе. Смотри, например, Ramachandran и др. Adv.Prot.Chem. 23: 283-437 (1968), а именно, на страницах 285-94, которые введены в данную заявку как ссылки. Данный способ может быть применен к любому белку и основывается частично на открытии того факта, что у человека анкерное положение первичного кармана 1 связывающих желобков молекулы класса II МНС имеет четко соответствующую специфичность для определенных аминокислотных боковых цепей. Специфичность этого кармана определяется идентичностью аминокислоты в положении 86 бета-цепи молекулы класса II МНС. Этот сайт располагается на дне кармана 1 и определяет размер боковой цепи, которая может подвергаться аккомодации этим карманом (Marshall, K.W., J. Immunol., 152: 4946-4956 (1994)). Если этот остаток представляет собой глицин, то все гидрофобные алифатические и ароматические аминокислоты (гидрофобными алифатическими аминокислотами являются: валин, лейцин, изолейцин, метионин, а ароматическими являются: фенилаланин, тирозин и триптофан) могут подвергаться аккомодации в кармане, предпочтение отдается ароматическим боковым цепям. Если этот остаток кармана представляет собой валин, то боковая цепь этой аминокислоты выдается в карман и ограничивает размер боковых цепей пептида, которые могут подвергаться аккомодации таким образом, что только гидрофобные алифатические боковые цепи могут подвергаться аккомодации. Таким образом, в последовательности аминокислотных остатков, где найдена аминокислота с гидрофобной алифатической или ароматической боковой цепью, существует потенциал для присутствующего ограниченного эпитопа Т-клеток класса II МНС. Даже если боковая цепь является гидрофобной алифатической, тем не менее она приблизительно в два раза более ассоциирована с эпитопом Т-клеток, чем ароматическая боковая цепь (даже предполагая приблизительное распределение типов кармана 1 в общей популяции).
Вычислительный способ воплощения параметров данного изобретения касательно вероятности пептидных участков, которые содержат эпитопы Т-клеток, заключается в следующем:
(1) Исследуется первичная последовательность пептидного сегмента предварительно определенной длины и идентифицируются все присутствующие гидрофобные алифатические и ароматические боковые цепи. (2) Гидрофобные алифатические боковые цепи оцениваются значением большим, чем для ароматических боковых цепей; приблизительно в два раза большим значением, чем для ароматический боковых цепей, например, значение 2 для гидрофобной алифатической боковой цепи и значение 1 для ароматической боковой цепи. (3) Определенные значения суммируются для каждого перекрывающегося сегмента аминокислотных остатков (окна) предварительно определенной постоянный длины в пределах пептида, и общее значение для частного сегмента (окна) определяется для одного аминокислотного остатка в промежуточном положении сегмента (окна), предпочтительно для остатка, который находится в центре выборочного сегмента (окна). Эта процедура повторяется для каждого выборочного перекрывающегося сегмента аминокислотных остатков (окна). Таким образом, каждый аминокислотный остаток пептида определяется значением, которое имеет отношение к вероятности эпитопа Т-клеток, который присутствует в этом частном сегменте (окне). (4) Подсчитанные и определенные как описано в Этапе 3 значения могут быть нанесены против координат аминокислот цельной последовательности аминокислот, которая подвергается анализу. (5) Все части последовательности, которые имеют счет предварительно определенного значения, например, значение 1, считаются вероятно такими, которые содержат эпитоп Т-клеток и могут быть при желании модифицированы. Этот особый аспект данного изобретения обеспечивает общий способ, с помощью которого могут быть описаны участки пептида, вероятностно содержащие эпитопы Т-клеток. Модификации пептида в этих участках имеют потенциал для модификации связывающих характеристик класса II МНС.
В соответствии с другим аспектом данного изобретения эпитопы Т-клеток могут быть предсказаны с большей точностью при использовании более сложного вычислительного способа, который учитывает взаимодействия пептидов с моделями аллелей класса II МНС. Вычислительное прогнозирование эпитопов Т-клеток, присутствующих в пределах пептида в соответствии с частным аспектом изобретения, рассматривает конструкцию моделей, по крайней мере, 42 аллелей класса II МНС, основываясь на структурах хорошо известных молекул класса II МНС и способе для использования этих моделей в вычислительной идентификации эпитопов Т-клеток, конструкции библиотек пептидных скелетов для каждой модели для того, чтобы получить возможность для известной вариабельности в родственных положениях альфа-углерода (Сα) пептидного скелета, конструкции библиотек структур аминокислотных боковых цепей для каждого скелета с каждой моделью для каждой из 20 аминокислотных альтернатив в критических положениях для каждого взаимодействия между пептидом и молекулой класса II МНС, использовании этих библиотек скелетов и структур боковых цепей в сочетании со счетной функцией для выбора оптимального скелета и структуры боковой цепи для частного пептида, состыкованного с частной молекулой класса II МНС, и получения значения связывания из результатов этого взаимодействия.
Модели молекул класса II МНС могут быть получены посредством гомологичного моделирования из числа подобных структур, находящихся в банке данных белков Брукхавена ("PDB"). Это может быть сделано с использованием полуавтоматического программного обеспечения гомологичного моделирования (Modeller, Sali A. & Blundell TL., 1993. J.Mol.Biol. 234: 779-815), которое вводит воспроизводящую отжиговую функцию в сопряжение с CHARMm силовым полем для минимизации энергии (доступно из Molecular Simulations Inc., San Diego, Cα). Альтернативные способы моделирования также могут использоваться.
Настоящий способ значительно отличается от других вычислительных способов, которые используют библиотеки экспериментально полученных данных относительно связывания каждой аминокислотной альтернативы в каждом положении связывающего желобка для небольшого набора молекул класса II МНС (Marshall, K.W и др., Biomed. Pept. Proteins Nucleic Acids, i (3): 157-162) (1995)) или даже других вычислительных способов, которые используют простые экспериментальные данные по связыванию для того, чтобы определить характеристики связывания частных типов карманов связывания в пределах бороздки, снова же используя относительно небольшие подмножества молекул класса II МНС, и потом "перемешивая и подбирая" типы карманов из этой библиотеки карманов для искусственного создания дальнейших "виртуальных" молекул класса II МНС (Sturniolo Т. и др., Nat. Biotech. 17(6): 555-561 (1999)). Оба предыдущих способа страдают от основных недостатков, которые, благодаря сложности анализов и необходимости синтезировать большое количество вариантов пептидов, дают возможность экспериментально исследовать только небольшое количество молекул класса II МНС. Таким образом, первый способ уровня техники может только дать возможность предсказать свойства только для небольшого числа молекул класса II МНС. Второй способ уровня техники также дает возможность сделать предположение, что карман, покрытый подобными аминокислотами в одной молекуле, будет иметь такие же связывающие характеристики в контексте различных аллелей класса II и страдать также теми недостатками, что и те молекулы класса II МНС, которые могут быть виртуально созданы и которые содержат карманы, содержащиеся в пределах библиотеки карманов. Используя моделирующий подход, описанный в данной заявке, структура любого числа и типа исследованных молекул класса II МНС могут быть дедуцированы, и, таким образом, аллели могут быть специфично выбраны как представители общей популяции. В дополнение к этому, число исследованных молекул класса II МНС может быть увеличено путем создания дополнительных моделей для получения дополнительных данных посредством комплексных экспериментов.
Использование библиотеки скелетов предусматривает вариации положений Сα атомов различных исследованных пептидов при стыковке с частными молекулами класса II МНС. Это снова-таки отличается от альтернативных вычислительных способов уровня техники, описанных выше, которые полагаются на использование упрощенных пептидых скелетов для исследования аминокислотного связывания в частных карманах. Эти упрощенные скелеты, вероятно, не являются представительными структурами скелета, которые находятся в "реальных" пептидах, что ведет к погрешностям в прогнозировании пептидного связывания. Данная библиотека скелетов создается путем накладывания скелетов всех пептидов, связанных с молекулами класса II МНС, найденными в пределах белкового банка данных и с помощью фиксации исходного значения площади отклонения (RMC) между Сα атомами каждой из одиннадцати аминокислот, размещенных в пределах связывающего желобка. Поскольку эта библиотека может быть получена из небольшого количества приемлемых мышиных или человеческих структур (в настоящее время 13), то для того, чтобы предусмотреть возможность даже большей вариабельности, количественные данные RMS для каждого Сα положения увеличивается на 50%. Потом определяют среднюю величину положения Сα каждой аминокислоты и очерчивают сферу вокруг этой точки, радиус которой равен отклонению RMS в этом положении плюс 50%. Эта сфера представляет все возможные Сα положения. Используя Сα с, по крайней мере, отклонением RMS (те аминокислоты в кармане 1, как упомянуто выше, которые эквивалентны положению 2 одиннадцати остатков в связывающем желобке), сфера имеет пространственную привязку, и каждая вершина в пределах решетки потом используется как возможное расположение для Сα аминокислоты. Полученная амидная плоскость, соответствующая пептиду, связанному с последующей аминокислотой, перемещается на каждый из этих Сα, и углы φ и ψ поворачиваются пошагово на ряд интервалов для того, чтобы переместить последующие Сα. Если последовательные Сα попадают в пределы "сферы разрешенных положений" для этих Сα, то ориентация дипептида принимается, в то время, если она попадает за пределы сферы, то дипептид отвергается. Этот процесс потом повторяют для каждого последующего Сα положения так, что дипептид растет от исходного Сα кармана 1, пока все 9 последующих Сα не будут позиционированы от всех возможных перестановок предыдущих Сα. Процесс потом повторяют еще раз для одного Сα предыдущего кармана 1 для создания библиотеки положений Сα скелета, размещенных в пределах желобка связывания. Количество полученных скелетов зависит от нескольких факторов: размера "сферы возможных положений"; качества привязки "первичной сферы" в положении кармана 1; качества пошагового вращения углов φ и ψ, используемых для позиционирования последующих Сα. При использовании этого процесса может быть создана большая библиотека скелетов. Для большей библиотеки скелетов более вероятно, что она будет такой, что оптимальная подгонка будет найдена для частного пептида в пределах желобка связывания молекулы класса II МНС. Поскольку все скелеты не будут приемлемыми для стыковки со всеми моделями молекул класса II МНС по причине дисгармонии с аминокислотами доменов связывания, то для каждого аллеля создается подмножество библиотеки, включая скелеты, которые могут быть подвергаться аккомодации этими аллелями. Использование библиотеки скелетов в сочетании с моделями молекул класса II МНС создает исчерпывающую базу данных, состоящую из возможных структур боковой цепи для каждой аминокислоты в каждом положении связывающего желобка для каждой молекулы класса II МНС, которая стыкуется с каждым возможным скелетом. Этот набор данных получают при использовании простой функции стерического перекрывания, где молекула класса II МНС стыкуется со скелетом, и аминокислотная боковая цепь перемещается в скелет в желаемом положении. Каждая из способных к вращению связей боковой цепи вращается пошагово на набор интервалов, и полученные положения атомов зависят от записанной связи. Взаимодействие атома с атомами боковых цепей связывающего желобка записывается и положения принимаются либо отвергается в зависимости от следующих критериев: общая сумма перекрывания всех атомов, позиционированных к настоящему моменту времени, не должна превышать предварительно определенного значения. Таким образом, жесткость конформационного поиска является функцией интервала, используемого при пошаговом вращении связи и предварительно определенной границы для общего перекрывания. Таким образом, последнее значение может быть малым, если известно, что определенный карман является жестким, однако жесткость может быть ослаблена, если известно, что положения боковых цепей кармана являются относительно гибкими. Таким образом, может быть сделано допущение для имитации вариаций гибкости в пределах карманов связывающего желобка. Этот конформационный поиск потом повторяют для каждой аминокислоты в каждом положении каждого скелета при стыковке с каждой из молекул класса II МНС для создания исчерпывающей базы данных конформаций боковых цепей.
Приемлемое математическое выражение используется для оценки энергии связывания между моделями молекул класса II МНС в сочетании с конформациями петидных лигандов, которые эмпирически получены путем исследования большой базы данных конформаций скелета/боковых цепей, описанных выше. Таким образом, белок исследуется на потенциальные эпитопы Т-клеток путем исследования каждой возможной длины пептида, которая варьирует между 9 и 20 аминокислотами (несмотря на то, что длина сохраняется постоянной для каждого исследования) для следующих вычислений: молекулу класса II МНС выбирают вместе с пептидным скелетом, возможным для этой молекулы и боковых цепей, соответствующих желаемой пептидной последовательности, на которую их переносят. Тождественность атомов и данные относительно межатомного расстояния, относящиеся к определенной боковой цепи в определенном положении в скелете, собирают для каждой возможной конформации этой аминокислоты (полученой из базы данных, описанной выше). Это повторяют для каждой боковой цепи вдоль скелета и получают пептидные значения, используя функцию подсчета. Лучшее значение для этого скелета запоминают и процесс повторяют для каждого возможного скелета для выбранной модели. Значения всех возможных скелетов сравнивают и самое высокое значение считают как пептидное значение для желаемого пептида в этой модели класса II МНС. Этот процесс потом повторяют для каждой модели с каждым возможным пептидом, который имеет происхождение от исследуемого белка, и показывают значения для пептидов против моделей.
В контексте данного изобретения каждый лиганд, представленный для подсчета связывающей аффинности, представляет собой аминокислотный сегмент, выбранный из группы пептидов или белков, которые обсуждались выше. Таким образом, лиганд представляет собой выбранную цепь аминокислот, которая содержит от приблизительно 9 до 20 аминокислот в длину и которая имеет происхождение от пептида, полипептида или белка известной последовательности. Термины "аминокислоты" и "остатки" в данной заявке употребляются как эквивалентные термины. Лиганд в форме последовательных аминокислот пептида, который исследуется, перемещают в скелет из библиотеки скелетов, позиционируют в связывающем желобке молекулы класса II МНС из библиотеки моделей молекул класса II МНС через координаты Сα атомов пептидного скелета и возможные структуры для каждой боковой цепи выбирают из базы данных возможных конформации. Тождественности релевантных атомов и межатомные расстояния также извлекаются из этой базы данных и используются для подсчета пептидного показателя связывания. Лиганды с высокой связывающей аффинностью для связывающего кармана класса П МНС отмечаются как кандидаты для сайт-направленного мутагенеза. Аминокислотные замены делают в отмеченных лигандах (и, таким образом, в белке, который представляет интерес) и который потом повторно тестируют, используя счетную функцию для того, чтобы определить изменения, которые уменьшают связывающую аффинность ниже предварительно определенного значения. Эти изменения могут быть потом введены в белок, который представляет интерес, для удаления эпитопов Т-клеток. Связывание между пептидным лигандом и желобком связывания молекул класса II МНС вовлекает нековалентные взаимодействия, включая, но не ограничиваясь, следующие: водородные связи, электростатические взаимодействия, гидрофобные (липофильные) взаимодействия и Ван-дер-ваальсовые взаимодействия. Это включают в пептидную счетную функцию так, как описано более детально ниже. Понятно, что водородная связь представляет собой нековалентную связь, которая может образовываться между полярными или заряженными группами и состоит из водородного атома, поделенного между двумя другими атомами. Водород донора водорода имеет позитивный заряд, в то время как акцептор водорода имеет частичный негативный заряд. Для целей пептидных/белковых взаимодействий доноры водородной связи могут быть или азотами с присоединенным водородом, или водородами, присоединенными к кислороду или азоту. Акцептор водородной связи может быть кислородами, не присоединенными к водороду, азотами без присоединенных водородов и одной или двумя связями, или серой только с одной связью. Определенные атомы, такие как кислород, присоединенный к водороду или азоту имина (например, C=NH), могут быть как акцепторами, так и донорами водорода. Энергия водородной связи колеблется от 3 до 7 ккал/мол и является более сильной, чем Ван-дер-ваальсовые связи, но слабее, чем ковалентные связи. Водородные связи являются также высоко направленными и являются наиболее сильными, когда донорный атом, атом водорода и акцепторный атом колинеарны. Электростатические связи образуются между противоположно заряженными парами ионов, и сила взаимодействия является обратно пропорциональной квадрату расстояния между атомами в соответствии с законом Кулона. Оптимальное расстояние между парами ионов составляет приблизительно 2,8 Å. В белок/пептидных взаимодействиях электростатические связи могут образовываться между аргинином, гистидином или лизином и аспартатом или глутаматом. Сила связи будет зависеть от рКа ионизирующих групп и диэлектрической постоянной среды, несмотря на то, что они являются приблизительно подобными по силе гидрофобным связям. Липофильные взаимодействия являются благоприятными гидрофобно-гидрофобными контактами, которые возникают между белком и пептидным лигандом. Обычно, они возникают между гидрофобными боковыми аминокислотными цепями пептида, спрятанными внутри карманов желобка связывания, так, что они не подвергаются действию растворителя. Подверженность гидрофобных остатков действию растворителя является крайне неблагоприятной, поскольку окружающие молекулы растворителя оказывают влияние на водородную связь с другими клеткоподобными клатрат-формирующими структурами. Полученное в результате уменьшение энтропии является крайне неблагоприятным. Липофильные атомы могут быть серой, которая не является ни полярным, ни водородным акцептором, и атомами углерода, которые не являются полярными. Ван-дер-ваальсовые связи являются неспецифическими силами, существующими между атомами, которые находятся на расстоянии 3-4 Å. Они слабее и менее специфичны, чем водородные и электростатические связи. Распределение электронного заряда вокруг атома меняется со временем, и в любом случае распределение заряда не есть симметричным. Эта кратковременная асимметрия в электронном заряде индуцирует подобную асимметрию в соседних атомах. Проистекающие отсюда силы притяжения между атомами достигают максимума при контактном расстоянии Ван-дер-Ваальса, но очень быстро убывают при расстоянии от приблизительно 1 Å до приблизительно 2 Å. Наоборот, как только атомы становятся разделенными менее чем контактным расстоянием, в значительной степени сильные силы отталкивания становятся доминантными тогда, как внешние электронные облака атомов перекрываются. Несмотря на то, что силы притяжения являются относительно более слабыми по сравнению с электростатическими и водородными связями (приблизительно 0,6 ккал/мол), силы отталкивания, в частности, могут быть более важными в определении будет ли пептидный лиганд успешно связываться с белком.
В одном воплощении функция показателя Bohm (SCORE1 подход) используется для оценки связывающего контакта (Bohm H.J., J. Comput Aided Mol. Des 8(3): 243-256 (1994), ссылка введена в данную заявку в своей целостности). В другом воплощении функция показателя (SCORE2 подход) используется для оценки связывающей аффинности как индикатор лиганд-содержащего эпитопа Т-клеток (Bohm H.J., J. Comput Aided Mol. Des. 12(4): 309-323 (1998), ссылка введена в данную заявку в своей целостности). Однако функция оценки Bohm, как описано в указанных выше ссылках, используется для оценки связывающей аффинности лиганда с белком, где заранее известно, что лиганд успешно связывается с белком и комплекс белок/лиганд имеет определенную структуру, определенная структура представлена в банке данных белков ("PDB"). Таким образом, функция подсчета была развита далее с преимуществом известных позитивных данных связывания. Для того чтобы иметь возможность различения между позитивными и негативными связующими, составляющая отталкивания должна прибавляться к равенству. В дополнение, более удовлетворительная оценка связывающей энергии достигается путем вычисления липофильных взаимодействий попарным образом, более точно, чем при использовании площади, основанной на составляющей энергии приведенных функций Bohm. Таким образом, в предпочтительном воплощении энергия связывания оценивается при использовании модифицированной счетной функции Bohm. В модифицированной счетной функции Bohm энергия связывания между белком и лигандом (AGbind) оценивается при учете следующих параметров: уменьшение энергии связывания благодаря абсолютной потере трансляционной и вращательной энтропии лиганда (AGo); вклады идеальных водородных связей (AGhb), где, по крайней мере, один партнер является нейтральным; вклады невозмущенных ионных взаимодействий (AGionic); липофильные взаимодействия между липофильными атомами лиганда и липофильными атомами акцептора (AGupo); потеря связывающей энергии благодаря застыванию внутренних степеней свободы в лиганде, то есть свобода вращения вокруг каждой связи С-С уменьшена (AGrot); энергия взаимодействия между белком и лигандом (Eydw). Анализ этих составляющих дает равенство 1:
(ΔGbind)=(ΔG0)+(ΔGhb×Nhb)+(ΔGionic×Nionic)+(ΔGiipo×Nupo)+(ΔGrot+Nrot)+(Evdw).
где N представляет собой количество установленных взаимодействий для специфической составляющей и в одном воплощении, ΔG0, ΔGhb, ΔGionic, ΔGupo и ΔGrot являются константами, которые имеют значения: 5,4, -4,7, -4,7, -0,17 и 1,4, соответственно.
Составляющую Nhb подсчитывают в соответствии с равенством 2:
Nhb=Σh-bondf(ΔR, Δα)×f(Nneighb)×fpcs
f(ΔR, Δα) представляет собой штрафную функцию, которая подсчитывается для больших отклонений водородных связей от идеальных и рассчитывается в соответствии с равенством 3:
f(ΔR, Δα)=f1(ΔК)×f2(Δ(x), где f1(ΔR)=1, если ΔR<=TOL
или =1-(ΔR-TOL)/0,4, если ΔR <=0,4 +TOL
или =0, если ΔR> 0,4 + TOL и: f2(Δα)=1, если Δα<30°
или =1-(Δα-30)/50, если Δα<=80° или=0, если Δα>80°
TOL представляет собой допустимое отклонение в длине водородной связи =0,25 Å.
ΔR представляет собой отклонение длины ∠N/O-H...O/N водородной связи от идеального значения =1,9 Å.
Δα представляет собой отклонение угла водородной связи идеального значения 180°.
f(Nneighb) проводит отличие между вогнутой и выгнутой частями поверхности белка и, таким образом, устанавливает больший вес для полярных взаимодействий, найденных в карманах, больше, чем таковые для тех, которые найдены на поверхности белка. Эта функция рассчитывается в соответствии с равенством 4, приведенным ниже:
f(Nneighb)=(Nneighb/Nneighb.o)α, где α=0,5;
Nneighb представляет собой число негидрофобных атомов белка, которые находятся ближе, чем 5 Å к любому атому белка;
Nneighb,o представляет собой константу, равную 25.
fpcs является функцией, которая допустима для площади полярной поверхности контакта на одну водородную связь и, таким образом, проводит различия между сильными и слабыми водородными связями, ее значение определяется в соответствии со следующими критериями:
fpcs=β, когда Apoiar/NHB<10 Å2
или fpcs=1, когда Apoiar/NHB>10 Å2
Apoiar представляет собой размер площади полярного контакта белок-лиганд NHB - число водородных связей;
Р является константой, значение которой =1,2
Для осуществления модифицированной функции подсчета Bohm вклады ионных взаимодействий ΔGionic подсчитываются тем же способом, что и для водородных связей, описанных выше, поскольку предполагается одинаковая геометрическая зависимость.
Составляющую Nnpo подсчитывают в соответствии с равенством 5, приведенным ниже:
Nlipo=ΣILf(rIL)
f(rIL) подсчитывается для всех липофильных лигандных атомов, 1, и всех липофильных белковых атомов, L, в соответствии со следующими критериями:
f(rIL)=1, когда rIL<=R1f(rIL)=(rIL-R1)/(R2-R1), когда R2<rIL>R1 f(rIL)=0, когда rIL>=R2,
где: R1=r1 vdw+rL vdw+0,5
и R2=R1+3,0
и: r1 vdw представляет собой радиус Ван-дер-Ваальса атома 1;
и rL vdw представляет собой радиус Ван-дер-Ваальса атома L.
Составляющая Nrot представляет собой число способных к вращению связей аминокислотной боковой цепи и берется для того, чтобы представлять число ациклических sp3-sp и sp3-sp2 связей. Вращения терминального конца -СН3 или -МН3 не берутся в расчет.
Конечная составляющая EVDW подсчитывается в соответствии с равенством 6 ниже:
EVDW=ε1ε2((r1 vdw+r2 vdw)12/r12-(r1 vdw+r2 vdw)6/r6,
где ε1 и ε2 являются константами, зависимыми от идентичности атомов r1 vdw+r2 vdw, являются атомными радиусами Ван-дер-Ваальса, r представляет собой расстояние между парой атомов.
Что касается равенства 6, то в одном воплощении константы ε1 и ε2 имеют следующие значения для атомов: С:0,245, N:0,283, О:0,316, S:0,316, соответственно (то есть атомы углерода, азота, кислорода и серы, соответственно). Что касается равенств 5 и 6, то радиусы Ван-дер-Ваальса имеют следующие значения для атомов: С:1,85, N:1,75, О:1,60, S:2,00 Å.
Понятно, что все предварительно определенные значения и константы, приведенные в равенствах выше, определены в пределах ограниченных условий понимания белок-пептидных взаимодействий в особой связи с типом вычисления, которое проводится в данной заявке. Таким образом возможно, что эта счетная функция будет усовершенствована в дальнейшем, и эти значения и константы могут изменяться, поэтому любое приемлемое числовое значение, которое дает желаемые результаты в составляющих оценки связывающей энергии белка с лигандом, может использоваться и, следовательно, подпадает под объем данного изобретения.
Как описано выше, счетная функция применяется к данным, извлеченным из базы данных конформаций боковых цепей, идентичных атомов и межатомных расстояний. Для целей данного описания число молекул класса II МНС, включенных в эту базу, составляет 42 модели плюс 4 установленные структуры. Из приведенного выше описания понятно, что модульная природа построения вычислительного способа в соответствии с данным изобретением означает, что новые модели могут просто прибавляться и исследоваться совместно с библиотекой пептидных скелетов и функцией поиска конформаций боковых цепей для создания дополнительных наборов данных, которые могут быть обработаны с помощью пептидной счетной функции, как описано выше. Это дает возможность легко увеличить набор исследованных молекул класса II МНС или структур и ассоциированных данных, которые замещаются, если данные являются доступными для создания более точных моделей существующих аллелей.
Настоящий способ прогнозирования может быть откалиброван в соответствии с набором данных, включающим большое количество пептидов, аффинность которых для различных молекул класса II МНС была ранее экспериментально определена. Путем сравнения подсчитанных данных и экспериментальных данных как определено выше можно сократить число значений, если известно, что все экспериментально определенные эпитопы Т-клеток были правильно предсказаны.
Понятно, что, несмотря на то, что описанная выше счетная функция является относительно простой по сравнению с некоторыми сложными методологиями, которые доступны, подсчеты проводятся чрезвычайно быстро. Понятно, что цель не заключается в том, чтобы подсчитать реальную энергию связывания саму по себе для каждого пептида, который стыкуется со связывающим желобком выбранного белка класса II МНС. Основная цель заключается в том, чтобы получить данные относительно энергии связывания для претендентов как цели для прогнозирования размещения эпитопов Т-клеток, основанных на первичной структуре (то есть, аминокислотной последовательности) выбранного белка. Относительно высокая энергия связывания или связывающая энергия выбранного значения предполагает присутствие эпитопов Т-клеток в лиганде. Лиганд можно потом подвергать, по крайней мере, одному кругу аминокислотных замен и повторно подсчитывать энергию связывания. Благодаря быстроте подсчета эта обработка пептидной последовательности может быть проведена при использовании компьютерной технологии в пределах интерфейса пользователя программы на эффективной доступной компьютерной технике. Основное вложение в компьютерную технику, таким образом, не требуется.
Любому специалисту в данной области понятно, что другие доступные программные обеспечения могут использоваться для тех же целей. В частности, более сложное программное обеспечение, которое способно к состыковке лигандов в белок-связывающих сайтах, может использоваться в соединение с энергетической минимизацией. Примерами стыковочных программных продуктов являются следующие: DOCK (Kuntz и др., J. Mol. Biol., 161: 269-288 (1982)), LUDI (Bohm H.J., J. Comput Aided Mol. Des., 8: 623-632 (1994) и FLEXX (Rarey M. И др. ISMB, 3: 300-308 (1995)). Примеры программного обеспечения молекулярного моделирования и манипулирования включают: AMBER (Tripos) и CHARMm (Molecular Simulations Inc.). Использование этих вычислительных методов будет строго ограничивать пропускную мощность способа в соответствии с данным изобретением из-за продолжительности времени обработки, которое необходимо для проведения необходимых вычислений. Однако является вероятным, что такие способы могут использоваться как "вторичный скрининг" для получения более точных вычислений энергии связывания для пептидов, которые определены как "позитивные связующие" при использовании способа в соответствии с данным изобретением. Ограничение времени обработки для сложных молекулярно-механических или молекулярно-динамических вычислений является одной из причин, которая определяется программным обеспечением и возможностями современной технологии компьютерного оборудования. Можно предвидеть, что в будущем с написанием более рационального кода и продолжающегося увеличения скорости процессоров компьютера станет возможным проводить такие вычисления в пределах, более поддающихся управлению временных рамок. Дальнейшая информация относительно энергетических функций применимо к макромолекулам и рассмотрение различных взаимодействий, которые имеют место внутри собранной белковой структуры, может быть найдена в: Brooks B.R. и др. J. Comput Chem., 4: 187-217 (1983) и дополнительная информация, касающаяся общих взаимодействий белок-лиганд, может быть найдена в: Dauber-Osguthrpe и др., Proteins 4 (I): 31-47 (1988), эти ссылки введены в данную заявку в своей целостности. Полезная информация также может быть найдена, например, в Fasman G.D. ed.. Prediction of Protein Structure of Protein Conformation, Plenum Press, New York, ISBN: 0-306, 4313-9.
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБ ИДЕНТИФИКАЦИИ ЭПИТОПОВ Т-КЛЕТОК И ЕГО ПРИМЕНЕНИЕ ДЛЯ ПОЛУЧЕНИЯ МОЛЕКУЛ СО СНИЖЕННОЙ ИММУНОГЕННОСТЬЮ | 2002 |
|
RU2303264C2 |
| МОДИФИЦИРОВАННЫЕ ЭПИТОПЫ ДЛЯ УСИЛЕНИЯ ОТВЕТОВ CD4+ Т-КЛЕТОК | 2013 |
|
RU2724994C2 |
| ИСКУССТВЕННЫЕ БЕЛКИ С ПОНИЖЕННОЙ ИММУНОГЕННОСТЬЮ | 2002 |
|
RU2363707C2 |
| ФАКТОР КОАГУЛЯЦИИ VIII С УМЕНЬШЕННОЙ ИММУНОГЕННОСТЬЮ | 2013 |
|
RU2631801C2 |
| НОВЫЕ ИММУНОГЕННЫЕ ПЕПТИДЫ | 2015 |
|
RU2709711C2 |
| ПРЕДСКАЗАНИЕ ИММУНОГЕННОСТИ Т-КЛЕТОЧНЫХ ЭПИТОПОВ | 2014 |
|
RU2724370C2 |
| СПОСОБ КАРТИРОВАНИЯ И УСТРАНЕНИЯ ЭПИТОПОВ Т-КЛЕТОК | 2003 |
|
RU2334235C2 |
| СПОСОБЫ ПРОГНОЗИРОВАНИЯ ПРИМЕНИМОСТИ СПЕЦИФИЧНЫХ ДЛЯ ЗАБОЛЕВАНИЯ АМИНОКИСЛОТНЫХ МОДИФИКАЦИЙ ДЛЯ ИММУНОТЕРАПИИ | 2018 |
|
RU2799341C2 |
| RA АНТИГЕННЫЕ ПЕПТИДЫ | 2004 |
|
RU2359974C2 |
| ИММУНОКОНЪЮГАТЫ ДЛЯ СПЕЦИФИЧЕСКОЙ ИНДУКЦИИ ЦИТОТОКСИЧНОСТИ Т-КЛЕТОК ПРОТИВ КЛЕТОК-МИШЕНЕЙ | 2016 |
|
RU2746021C2 |
Настоящее изобретение относится к антителам, которые направлены на EGF рецептор (HER1) для назначения главным образом человеку, и, в частности, для терапевтического использования при опухолях. Антитела представляют собой модифицированные антитела, в которых модификация приводит к уменьшенной склонности антитела вызывать иммунный ответ при введении человеку. Изобретение, в частности, относится к модификации анти-EGFR антитела 425 в его различных формах и их фрагментах, которая приводит к получению вариантов Mab 425, которые являются существенно неиммуногенными или менее иммуногенными, чем любой их немодифицированный эквивалент, при использовании in vivo. Преимущество изобретения заключается в снижении иммуногенности антитела, 5 н. и 5 з.п. ф-лы, 5 табл.
VH1:
QVQLVQSGAELVKPGASVKLSCKASGYTFTSHWMHWVKQAPGQGPEWAGEFNPSNGRTNYNEKFKSRVTITVDKSTSTAYMQLSSLTSEDTAVYYCASRDYDYDGRYFDYWGQGTTLTVSS
VK1:
QIVLTQSPATLSASPGERATMSCSASSSVTYMYWHQQKPGQSPRLLIYDASNLASGVPARFSGSGSGTSYTLTISSLEAEDAATYYCQQWSSHPFTFGQGTKVEIK
VH2:
QVQLVQSGAELVKPGASVKLSCKASGYTFTSHWMHWVKQAPGQGPEWIGEFNPSNGRTNYNEKFKSRVTITVDKSTSTAYMQLSSLTSEDTAVYYCASRDYDYDGRYFDYWGQGTTLTVSS
VK2:
QIVLTQSPATLSASPGERATMSCSASSSVTYMYWHQQKPGQSPRALIYDTSNLASGVPARFSGSGSGTSYTLTISSLEAEDAATYYCQQWSSHPFTFGQGTKVEIK
VH3:
QVQLVQSGAELVKPGASVKLSCKASGYTFTSHWMHWVKQAPGQGLEWIGEFNPSNGRTNYNEKFKSRVTITVDKSTSTAYMQLSSLTSEDTAVYYCASRDYDYDGRFDYWGQGTTLTVSS
VK3:
QIVLTQSPATLSASPGERATMSCSASSSVTYMYWHQQKPGQSPRLLIYDTSNLASGVPARFSGSGSGTSYTLTISSLEAEDAATYYCQQWSSHPFTFGQGTKVEIK
VH4:
QVQLVQSGAELVKPGASVKLSCKASGYTFTSHWMHWVKQAPGQGLEWIGEFNPSNGRTNYNEKFKSRATLTVDKSTSTAYMQLSSLTSEDTAVYYCASRDYDYDGRYFDYWGQGTTLTVSS
VK4:
QLVLTQSPATLSASPGERATMSCSASSSVTYMYWHQQKPGQSPRLLIYDTSNLASGVPARFSGSGSGTSYTLTISSLEAEDAATYYCQQWSSHIFTFGQGTKVEIK
VH5:
QVQLVQSGAELVKPGASVKLSCKASGYTFTSHWMHWVKQAAGQGLEWIGEFNPSNGRTNYNEKFKSRATLTVDKSTSTAYMQLSSLTSEDTAVYYCASRDYDYDGRYFDYWGQGTTLTVSS
VK5:
QIVLTQSPATLSASPGERATMSCSASSSVTYMYWYQQKPGQSPRLLIYDTSNLASGVPARFSGSGSGTSYTLTISSLEAEDAATYYCQQWSSHIFTFGQGTKVEIK.
(i) определение аминокислотной последовательности тяжелой и/или легкой цепи исходно иммуногенно немодифицированного антитела или его части;
(ii) идентификацию одного или более потенциальных эпитопов Т-клеток в пределах аминокислотной последовательности антитела, выбранной из Таблицы 1 посредством (а) выбора участка пептида, который имеет известную последовательность аминокислотных остатков; (b) последовательного отбора аминокислотных сегментов перекрывающихся участков аминокислотных остатков с предварительно определенным одинаковым размером и состоящих, по крайней мере, из трех аминокислотных остатков выбранного участка; (с) подсчета значения связывания молекулы класса II МНС для каждого выбранного сегмента путем суммирования определенных значений для каждой гидрофобной аминокислотной боковой цепи, которая присутствует в каждом выбранном сегменте аминокислотного остатка; и (d) идентификации по крайней мере одного из указанных сегментов, подходящих для модификации, основываясь на подсчитанной величине связывания молекулы класса II МНС этого сегмента для изменения общего значения связывания класса II МНС для пептида без существенного уменьшения терапевтической полезности пептида;
(iii) разработка новых вариантов последовательности с одной или более аминокислотами в пределах идентифицированных потенциальных эпитопов Т-клеток, модифицированных посредством замены 1-9 аминокислотных остатков в любом исходном эпитопе Т-клеток;
(iv) конструирование таких вариантов последовательности с помощью техники рекомбинантных ДНК и тестирование указанных вариантов для того, чтобы идентифицировать один или более вариантов с желаемыми свойствами;
и (v) необязательное повторение этапов (ii)-(iv).
(i) определение аминокислотной последовательности тяжелой и/или легкой цепи исходно иммуногенно немодифицированного антитела или его части;
(ii) идентификацию одного или более потенциальных эпитопов Т-клеток в пределах аминокислотной последовательности антитела, выбранной из Таблицы 1, любым способом, включающим определение связывания пептидов с молекулами МНС при использовании способов in vitro или биологических анализов;
(iii) разработка новых вариантов последовательности с одной или более аминокислотами в пределах идентифицированных потенциальных эпитопов Т-клеток, модифицированных посредством замены 1-9 аминокислотных остатков для существенного уменьшения или устранения активности эпитопов Т-клеток, как определено путем связывания пептидов с молекулами МНС при использовании способов in vitro или биологическими анализами, или путем связывания комплекса пептид-МНС с Т-клетками;
(iv) конструирование таких вариантов последовательности с помощью техники рекомбинантных ДНК и тестирование указанных вариантов для того, чтобы идентифицировать один или более вариантов с желаемыми свойствами;
и (v) необязательное повторение этапов (ii)-(iv).
| Автоматический огнетушитель | 0 |
|
SU92A1 |
| RODECKU et al., Tumo grouth modulation by monoclonal antibogy to the epidermal grow factor receptot immunologically mediated and effector cellindependent effects., Cancer Reserch, 1987, v.47, №14, pp.3692-3696 | |||
| Приспособление в пере для письма с целью увеличения на нем запаса чернил и уменьшения скорости их высыхания | 1917 |
|
SU96A1 |

