Область, к которой относится изобретение
Изобретение относится к молекулярной биологии, биотехнологии, генной инженерии и медицине и может быть использовано для коррекции патологических состояний клеток различных органов и тканей, а также собственно органов и тканей человека, связанных с уменьшением уровня экспрессии гена Р4НА2 и/или уменьшением количества белка пролил 4- гидрокислаза альфа 2, а именно, в терапевтических целях.
Предшествующий уровень. Коллаген - фибриллярный белок молекулярной массой около 300 кДа, который является самым распространенным белком в организме человека. Коллаген является основой соединительной ткани, структурной основой кожи, хрящей, синовиальной жидкости суставов, бронхов, легочной ткани, сосудистой стенки, межпозвонковых дисков, стенок кишечника и желудка. В коллагене, который является гликопротеином, одна треть аминокислотных остатков приходится на глицин и еще одна треть на пролин и гидроксипролин, который является стабилизатором и протектором молекулы белка от действия протеаз. Коллаген обеспечивает структурную основу ткани, придает ей необходимые механические свойства.
Коллаген синтезируется в фибробластах в виде высокомолекулярного предшественника - проколлагена. Полипептидные цепи коллагенов поступают в просвет эндоплазматического ретикулума в форме пропептидов с сигнальной последовательностью. Далее происходит гидроксилирование остатков пролина и лизина, что необходимо для корректного фолдинга вновь синтезированных цепочек проколлагена.
Образование гидроксипролила и гидроксилизила катализируют железосодержащие ферменты - пролилгидроксилаза и лизилгидроксилаза, находящиеся в микросомальных фракциях многих тканей. Эти ферменты являются пептидилгидроксилазами, поскольку гидроксилирование происходит только после включения пролина или лизина в полипептидную цепь.
Пролил-4-гидроксилаза - фермент класса оксидоредуктаз, который катализирует гидроксилирование в положении 4 остатков пролина в синтезируемых цепях проколлагена. Фермент требует активности Fe2 +, аскорбата и α-кетоглутарата для активности. Выделенный из всех изученных источников фермент позвоночных обладает структурой тетрамера "альфа2бета2", в результате клонирования и экспрессии альфа-субъединицы фермента С. elegans было обнаружено, что пролил-4-гидроксилаза последней представляет собою димер "альфабета". α-субъединица 2 содержит каталитические и пептид-субстратные связывающие домены. Эта субъединица неактивна в отсутствие β-субъединицы. Считается, что не существует принципиальных различий в тканевом распределении обеих альфа-субъединиц. Фермент является оксигеназой со смешанной функцией и активен при участии молекулярного кислорода.
Экспрессия in vitro активной рекомбинантной пролил-4-гидроксилазы из ее субъединиц была успешно осуществлена путем трансдукции клеток насекомых Spodoptera frugiperda и Trichoplusia ni (Vuori, K., et al. ( 1992) Proc Natl Acad Sci USA 89, 7467-7470) рекомбинантными бакуловирусами или котрансфекцией экспрессирующих векторов в клеточных линиях млекопитающих COS-1 (John, DC и Bulleid, NJ (1996) Biochem J 317 (Pt 3), 659-665) и HEK293 (Wagner, K, et al. ( 2000) Biochem J352 Pt 3, 907-911), в дрожжах Pichia pastoris (Vuorela, A., et al. (1997) Embo J16, 6702-6712) и Saccharomyces cerevisiae (Toman, P.D. и др. ( 2000) J Biol Chem 275, 23303-23309).
Системами экспрессии для получения рекомбинантных коллагенов могут служить также клетки Е. coli, трансгенные животные и растения. Однако почти все вышеназванные системы экспрессии имеют свои недостатки. Например, при получении фермента в клетках кишечной палочки не происходит посттрансляционной модификации белка, а при получении в клетках дрожжей рекомбинантный белок не обладает пролил-4-гидроксилазной активностью. Система экспрессии в клетках млекопитающих обладает недостаточной продукцией. Что касается трансгенных животных (шелковый червь или мыши) или растений (табак), в них продуцируется коллаген, молекулы которого связаны поперечными сшивками, вследствие чего очистка такого продукта сильно затруднена. Поэтому создание эффективной системы экспрессии для получения коллагенов с высоким выходом очень актуально.
Известен ген Р4НА2 пролил 4-гидроксилазы человека, кодирующий каталитическую α (II) субъединицу основного изофермента (https://www.ncbi.nlm.nih.gov/gene/8974). Ген Р4НА2 локализуется в положении 5q31.1, имеет длину 37716 п. о. и состоит из 18 экзонов. Продукт гена катализирует образование 4-гидроксипролина, который необходим для правильного фолдинга синтезированных цепей проколлагена.
Показано, что миопия высокой степени, являясь одной из главных причин развития слепоты, может быть обусловлена мутациями в гене Р4НА2, кодирующем субъединицу альфа-2 пролил 4-гидроксилазы человека (Guo H, Tong P, Liu Y, Xia L, Wang Т, Tian Q, Li Y, Hu Y, Zheng Y, Jin X, Li Y, Xiong W, Tang B, Feng Y, Li J, Pan Q, Hu Z, Xia K. Mutations of Р4НА2 encoding prolyl 4-hydroxylase 2 are associated with nonsyndromic high myopia. Genet Med. 2015 17(4):300-6.) В результате полногеномного скрининга 186 человек, в том числе и с семейной миопатией, было выснено, что мутация c.871G>A приводит к вышеуказанному фенотипу. Был предложен механизм развития патологии, согласно которому происходит деградация матричной РНК, несущей мутацию.
В заявке WO 2014170460 (А2) описан метод получения коллагена из морских губок (Chondrosia reniformis) в дрожжах Pichia pastoris. Были созданы три векторные конструкции:
- с кДНК, кодирующей альфа-субъединицу пролил-4-гидроксилазы (pPinkHC и pPinkLC векторы),
с кДНК, кодирующей бета-субъединицу пролил-4-гидроксилаза (pPIC6B \ PDI-вектор),
- с кДНК, кодирующей нефибриллярный коллагеновый белок С. Reniformis (pPICZB \ ColCH).
В заявке описан способ, который предусматривает совместную трансформацию дрожжевого штамма двумя различными векторами, содержащими: первый - кодирующую последовательность альфа-субъединицы, второй - кодирующую последовательность бета-субъединицы фермента пролил-4-гидроксилазы, с последующей трансформацией с помощью вектора, содержащего одну последовательность, кодирующую коллаген морской губки.
Недостатком данного подхода является то, что для экспрессии генов субъединиц пролил-4-гидроксилазы морской губки выбрана система экспрессии дрожжевых клеток, которая согласно литературным не обладает значимой пролил-4-гидроксилазной активностью. В данной патентной заявке не рассматривается возможность экспрессии в клетках человека в качестве генно-терапевтического средства с учетом индивидуальных характеристик пациента, в связи с которыми может понадобиться группа вариаций для данного средства.
За прототип авторами было принято решение по патенту US 7759090 (В2), где описана экспрессионная система для получения коллагена. Показаны стабильно трансфицированные клеточные линии насекомых, содержащие кДНК, кодирующие α и β-субъединицы пролил-4-гидроксилазы человека в клетках Trichoplusia ni и Drosophila melanogaster S2. Дополнительно охарактеризовано участие пролил-4-гидроксилазы в сборке трех альфа-цепей с образованием миниколлагена XXI тримерного типа, который содержит интактный С-терминальный неколлагеновый домен (NC1) и коллагеновый домен (COL1) в системе Drosophila. Миниколлаген XXI, коэкспрессированный пролил-4-гидроксилазой, содержал достаточное количество гидроксипролина для образования термостойких устойчивых к пепсину тройных спиралей.
Один из вариантов осуществления изобретения описывает клетки насекомого, трансфицированные геном, кодирующим пролил-4-гидроксилазу. Этот ген состоит из α субъединицы и / или β-субъединицы пролил-4-гидроксилазы. Клетки насекомого дополнительно трансфицируются геном, кодирующим коллагеновый (COL1) домен и С1-терминальный домен (NC1) коллагена типа XXI.
Гены кодирующие α субъединицу и β-субъединицу пролил-4-гидроксилазы человека были клонированы совместно в экспрессионном векторе plZ/V5-His под контролем промотора OplE2. В системе Drosophila inducible expression (DES®) (Invitrogen) гены кодирующие α субъединицу и β-субъединицу пролил-4-гидроксилазы были клонированы раздельно в экспрессионном векторе рМТ/5-HisA под контролем промотора металлотионеина, который индуцируется добавлением ионов меди или кадмия.
Показано, что Р4Нα и Р4Нβ субъединицы способны собираться в активный тетрамер α2β2. Полученная пролил-4-гидроксилаза обеспечивает синтез корректно фолдированного коллагена. Сравнение уровней экспрессии Р4Нα и Р4Нβ в лизатах клеток Trichoplusia ni показало, что уровень экспрессии белка Р4Нα примерно в 5 раз меньше, чем Р4Нβ. Для экспрессии Р4Н в индуцибельной Drosophila системе экспрессии клетки Drosophila S2 были совместно трансфицированы рМТ/Р4Нα, рМТ/Р4Нβ и pCoHygro в соотношении 10:10:1 с использованием реагента для трансфекции Effectene (Qiagen). Домены коллагена были введены в клетки путем трансформации вектором pMT/BiP-mC21. Таким образом, рекомбинантный коллаген получен с использованием системы экспрессии трех генов в клетках Drosophila melanogaster.
Недостатком данного подхода является то, что для экспрессии генов субъединиц пролил-4-гидроксилазы человека выбрана система экспрессии клеток насекомых, которая согласно литературным данным имеет низкую активность пролил-4- гидроксилазы. Кроме того, есть данные о недостаточной эффективности и корректности посттрансляционных модификаций в клетках насекомых по сравнению с клетками человека. Также не рассматривается возможность экспрессии в клетках человека в качестве генно-терапевтического средства с учетом индивидуальных характеристик пациента, в связи с которыми может понадобиться группа вариаций для данного средства.
Раскрытие изобретения
Задачей данного изобретения является создание генетической конструкции на основе невирусной векторной плазмиды, включающей кДНК гена Р4НА2, для купирования проявлений состояний человеческого организма, связанных с уменьшением уровня экспрессии гена Р4НА2 и/или уменьшением количества белка пролил 4- гидрокислаза альфа 2, выбранной из группы генетических конструкций с кДНК гена Р4НА2, при использовании которых с учетом индивидуальных особенностей пациента, происходит повышение уровня экспрессии гена Р4НА2 и/или повышение количества белка пролил 4- гидрокислаза альфа 2 в клетках органов и тканей и/или органах и тканях организма.
Указанная задача решается за счет того, что создана генетическая конструкция на основе невирусного ДНК-вектора, включающего кДНК гена Р4НА2, для купирования проявлений состояний человеческого организма, связанных с уменьшением экспрессии гена Р4НА2 и/или уменьшением количества белка пролил 4- гидрокислаза альфа 2, выбранная из группы генетических конструкций, каждая из которых представляет генетическую конструкцию на основе ДНК-вектора, включающего кДНК гена Р4НА2, с кодирующей последовательностью белка пролил 4- гидрокислаза альфа 2, с делениями 5' и 3'-нетранслируемых областей, а именно, полученной на основе участка нативной немодифицированной кДНК гена Р4НА2 SEQ ID No: 1, или модифицированной кДНК гена Р4НА2, при этом в качестве модифицированной кДНК гена Р4НА2 используют SEQ ID No: 2, или SEQ ID No: 3, или SEQ ID No: 4, или SEQ ID No: 5, или SEQ ID No: 6, или SEQ ID No: 7 или сочетание этих генетических конструкций, каждая из которых содержит также регуляторные элементы, обеспечивающие высокий уровень экспрессии гена Р4НА2 в эукариотических клетках, а именно, в клетках органов и тканей человека, и способную увеличить количество белка пролил 4- гидрокислаза альфа 2, в клетках органов и тканей и/или органах и тканях человека, а именно, в гемопоэтических клетках, или мезенхимальных стволовых клетках, или хондробластах, или миоцитах, или миобластах, или фибробластах, или остеобластах, или кератоцитах, или эпителиальных клетках, или клетках роговицы, или эндотелиальных клетках, или эпителиальных клетках, в сочетании с транспортной молекулой или без нее при трансфекции этими генетическими конструкциями клеток органов и тканей человека, и/или в органах и тканях человека, а именно, в соединительно-тканных структурах, или коже, или суставах, или хрящевой ткани, или костной ткани, или мышечной ткани, или сухожилиях, или связках, или кровеносных сосудах, или стенке артерий, или роговице глаза, или склере, или плаценте, или дентине, или строме внутренних органов в сочетании с транспортной молекулой или без нее при введении этих генетических конструкций в органы и ткани человека. При этом каждая генетическая конструкция, выбранная из группы генетических конструкций с модифицированной кДНК гена Р4НА2 содержит последовательность нуклеотидов, включающую в себя белок-кодирующую область кДНК гена Р4НА2, которая несет модификации, не затрагивающие структуру белка пролил 4- гидрокислаза альфа 2, или затрагивающие структуру белка пролил 4- гидрокислаза альфа 2 и влияющие на кодируемую этой последовательностью аминокислотную последовательность, а именно: нуклеотидные замены, приводящие к аминокислотным заменам или обрыву аминокислотной цепи, и/или делеции, или комбинации вышеперечисленных модификаций. Причем в качестве транспортной молекулы используют липосомы, или дендримеры 5-го и выше поколений, или амфифильные блоксополимеры, или реагент на основе полиэтиленимина.
Способ получения генетической конструкции заключается в получении каждой генетической конструкции, выбранной из группы генетических конструкций, при этом получают кДНК гена Р4НА2, затем помещают кДНК в векторную плазмиду, способную обеспечить высокий уровень экспрессии этой кДНК в клетках различных органов и тканей человека, наращивают и выделяют необходимое количество генетической конструкции, затем комбинируют генетическую конструкцию с транспортной молекулой для трансфекции полученной генетической конструкцией клеток органов и тканей и/или введения полученной генетической конструкции в органы и ткани человека, при этом используют кДНК гена Р4НА2 с кодирующей последовательностью белка пролил 4- гидрокислаза альфа 2, с делениями 5' и 3'-нетранслируемых областей, а именно, полученной на основе участка нативной немодифицированной кДНК гена Р4НА2 SEQ ID No: 1, или модифицированной кДНК гена Р4НА2, при этом в качестве модифицированной кДНК гена Р4НА2 используют, или SEQ ID No: 2, или SEQ ID No: 3, или SEQ ID No: 4, или SEQ ID No: 5, или SEQ ID No: 6, или SEQ ID No: 7 или сочетание этих генетических конструкций.
Способ использования генетической конструкции заключается в трансфекции генетической конструкцией или несколькими генетическими конструкциями, выбранными из группы созданных генетических конструкций клеток органов и тканей пациента и/или во введении в органы и ткани пациента аутологичных клеток пациента, трансфицированных генетической конструкцией или несколькими генетическими конструкциями, выбранными из группы созданных генетических конструкций, и/или во введении в органы и ткани пациента генетической конструкции или нескольких генетических конструкций выбранных из группы созданных генетических конструкций, или сочетанием обозначенных способов. При этом выбор генетической конструкции из созданных генетических конструкций производят индивидуально для каждого пациента, после предварительно выполненного определения наиболее эффективной генетической конструкции для данного пациента.
Перечень фигур
На фиг. 1
Представлена нуклеотидная последовательность немодифицированной кДНК гена Р4НА2, последовательность которой гомологична приводимой в базе данных GenBank по/ номером NM_004199 Р4НА2 SEQ ID No: 1.
На фиг. 2
Модифицированная таким образом нуклеотидная последовательность кДНК Р4НА2 SEQ ID No: 2 содержит 2 нуклеотидных замены G→C в позициях 1089, 1119; 1 нуклеотидную замену T→G в позиции 1239, 1 нуклеотидную замену A→G в позиции 1290; не приводящие к изменениям в аминокислотной последовательности белка, кодируемого последовательностью гена Р4НА2 и имеет первичную структуру, приведенную на фиг. 2:
На фиг. 3
Модифицированная таким образом нуклеотидная последовательность кДНК Р4НА2 SEQ ID No: 3 содержит 3 нуклеотидных замены G→C в позициях 1089, 1119, 1347; 4 нуклеотидных замены T→G в позициях 1239, 1449, 1476, 1665, 2 нуклеотидных замены A→G в позициях 1290, 1653; не приводящие к изменениям в аминокислотной последовательности белка, кодируемого последовательностью гена Р4НА2 и имеет первичную структуру, приведенную на фиг. 3:
На фиг. 4
Модифицированная таким образом нуклеотидная последовательность кДНК Р4НА2 SEQ ID No: 4 содержит 4 нуклеотидных замены G→C в позициях 1089, 1119, 1347, 1905; 6 нуклеотидных замен T→G в позициях 1239, 1449, 1476, 1665, 1764, 1914; 6 нуклеотидных замен A→G в позициях 1290, 1653, 1755, 1881, 2058, 2127; не приводящие к изменениям в аминокислотной последовательности белка, кодируемого последовательностью гена Р4НА2 и имеет первичную структуру, приведенную на фиг. 4:
На фиг. 5
Модифицированная таким образом нуклеотидная последовательность кДНК Р4НА2 SEQ ID No: 5 длиной 1602 н.п. содержит делецию в 6 нуклеотидных пар, а также замену 8 аминокислотных остатков, приводящие к изменениям в аминокислотной последовательности белка пролил 4- гидрокислаза альфа 2 и имеет первичную структуру, приведенную на фиг. 5.
На фиг. 6
Модифицированная таким образом нуклеотидная последовательность кДНК Р4НА2 SEQ ID No: 6 длиной 1602 н.п. содержит 3 нуклеотидных замены G→C в позициях 1089, 1119, 1347; 2 нуклеотидных замены T→G в позициях 1239, 1449, 1 нуклеотидную замену A→G в позиции 1290; делецию в 6 нуклеотидных пар, а также замену 8 аминокислотных остатков, приводящие к изменениям в аминокислотной последовательности белка пролил 4- гидрокислаза альфа 2 и имеет первичную структуру, приведенную на фиг. 6:
На фиг. 7
Модифицированная таким образом нуклеотидная последовательность кДНК Р4НА2 SEQ ID No: 7 длиной 1602 н.п. содержит 3 нуклеотидных замены G→C в позициях 1089, 1119, 1347; 5 нуклеотидных замен T→G в позициях 1239, 1449, 1476, 1665, 1764; 5 нуклеотидных замен A→G в позициях 1290, 1653, 1755, 2058, 2127; делецию в 6 нуклеотидных пар, а также замену 8 аминокислотных остатков, приводящие к изменениям в аминокислотной последовательности белка пролил 4- гидрокислаза альфа 2 и имеет первичную структуру, приведенную на фиг. 7:
На фиг. 8
С целью последующего корректного определения генно-терапевтического эффекта после трансфекции фибробластов генетической конструкцией с кДНК гена Р4НА2 - VTvaf17 - P4HA2 SEQ ID No: 1 проводили анализ эндогенной экспрессии гена P4HA2 в культуре первичных фибробластов. На фигуре представлены графики накопления продуктов полимеразной цепной реакции (ПЦР), соответствующих:
1 - кДНК гена P4HA2, фибробласты со сниженной экспрессией гена P4HA2
2 - кДНК гена P4HA2, фибробласты с нормальной экспрессией гена P4HA2
3 - кДНК гена В2М, фибробласты со сниженной экспрессией гена P4HA2
4 - кДНК гена В2М, фибробласты с нормальной экспрессией гена P4HA2
В качестве референтного гена использовали ген В2М (Бета-2-микроглобулин) приведенного в базе данных GenBank под номером NM 004048.2.
На фиг. 9
С целью подтверждения увеличения экспрессии гена P4HA2 в клеточной культуре фибробластов со сниженной экспрессией гена P4HA2 при трансфекции данных клеток генетической конструкцией VTvaf17 - P4HA2 SEQ ID No: 1 с кДНК гена P4HA2 представлены графики накопления ПЦР-продуктов, соответствующих:
1 - кДНК гена Р4НА2 в фибробластах с нормальной экспрессией гена Р4НА2,
2 - кДНК гена Р4НА2 в фибробластах со сниженной экспрессией гена Р4НА2 до трансфекции конструкцией с кДНК гена Р4НА2
3 - кДНК гена Р4НА2 в фибробластах со сниженной экспрессией гена Р4НА2 после трансфекции конструкцией с кДНК гена Р4НА2
4 - кДНК гена Р4НА2 в фибробластах со сниженной экспрессией гена Р4НА2 после трансфекции вектором без кДНК гена Р4НА2
5 - кДНК гена В2М в фибробластах с нормальной экспрессией гена Р4НА2,
6 - кДНК гена В2М в фибробластах со сниженной экспрессией гена Р4НА2 до трансфекции конструкцией с кДНК гена Р4НА2
7 - кДНК гена В2М в фибробластах со сниженной экспрессией гена Р4НА2 после трансфекции конструкцией с кДНК гена Р4НА2
8 - кДНК гена В2М в фибробластах со сниженной экспрессией гена Р4НА2 после трансфекции вектором без кДНК гена Р4НА2
Из графиков следует, что в случае трансфекции вектором без вставки кДНК гена Р4НА2 уровень кДНК гена Р4НА2 в фибробластах не изменился, а в случае трансфекции конструкцией VTvaf17 - Р4НА2 SEQ ID No: 1 с кДНК Р4НА2 - уровень кДНК фибробластов со сниженной экспрессией гена Р4НА2 значительно увеличился.
На фиг. 10
С целью подтверждения увеличения экспрессии гена Р4НА2 в клеточной культуре фибробластов со сниженной экспрессией гена Р4НА2 при трансфекции данных клеток генетической конструкцией VTvaf17- P4HA2 SEQ ID No: 2 с кДНК гена Р4НА2 представлены графики накопления ПЦР-продуктов, соответствующих:
1 - кДНК гена P4HA2 в фибробластах с нормальной экспрессией гена P4HA2,
2 - кДНК гена P4HA2 в фибробластах со сниженной экспрессией гена P4HA2 до трансфекции конструкцией с кДНК гена P4HA2
3 - кДНК гена P4HA2 в фибробластах со сниженной экспрессией гена P4HA2 после трансфекции конструкцией с кДНК гена P4HA2
4 - кДНК гена P4HA2 в фибробластах со сниженной экспрессией гена P4HA2 после трансфекции вектором без кДНК гена P4HA2
5 - кДНК гена В2М в фибробластах с нормальной экспрессией гена P4HA2,
6 - кДНК гена В2М в фибробластах со сниженной экспрессией гена P4HA2 до трансфекции конструкцией с кДНК гена P4HA2
7 - кДНК гена В2М в фибробластах со сниженной экспрессией гена P4HA2 после трансфекции конструкцией с кДНК гена P4HA2
8 - кДНК гена В2М в фибробластах со сниженной экспрессией гена P4HA2 после трансфекции вектором без кДНК гена P4HA2
Из графиков следует, что в случае трансфекции вектором без вставки кДНК гена P4HA2 уровень кДНК гена P4HA2 в фибробластах не изменился, а в случае трансфекции конструкцией VTvaf17- P4HA2 SEQ ID No: 2 с кДНК P4HA2 - уровень кДНК фибробластов со сниженной экспрессией гена P4HA2 значительно увеличился.
На фиг. 11
С целью подтверждения увеличения экспрессии гена P4HA2 в клеточной культуре фибробластов со сниженной экспрессией гена Р4НА2 при трансфекции данных клеток генетической конструкцией VTvaf17- Р4НА2 SEQ ID No: 3 с кДНК гена Р4НА2 представлены графики накопления ПЦР-продуктов, соответствующих:
1 - кДНК гена Р4НА2 в фибробластах с нормальной экспрессией гена Р4НА2,
2 - кДНК гена Р4НА2 в фибробластах со сниженной экспрессией гена Р4НА2 до трансфекции конструкцией с кДНК гена Р4НА2
3 - кДНК гена Р4НА2 в фибробластах со сниженной экспрессией гена Р4НА2 после трансфекции конструкцией с кДНК гена Р4НА2
4 - кДНК гена Р4НА2 в фибробластах со сниженной экспрессией гена Р4НА2 после трансфекции вектором без кДНК гена Р4НА2
5 - кДНК гена В2М в фибробластах с нормальной экспрессией гена Р4НА2,
6 - кДНК гена В2М в фибробластах со сниженной экспрессией гена Р4НА2 до трансфекции конструкцией с кДНК гена Р4НА2
7 - кДНК гена В2М в фибробластах со сниженной экспрессией гена Р4НА2 после трансфекции конструкцией с кДНК гена Р4НА2
8 - кДНК гена В2М в фибробластах со сниженной экспрессией гена Р4НА2 после трансфекции вектором без кДНК гена Р4НА2
Из графиков следует, что в случае трансфекции вектором без вставки кДНК гена Р4НА2 уровень кДНК гена Р4НА2 в фибробластах не изменился, а в случае трансфекции конструкцией VTvaf17- Р4НА2 SEQ ID No: 3 с кДНК Р4НА2 - уровень кДНК фибробластов со сниженной экспрессией гена Р4НА2 значительно увеличился.
На фиг. 12
С целью подтверждения увеличения экспрессии гена Р4НА2 в клеточной культуре фибробластов со сниженной экспрессией гена Р4НА2 при трансфекции данных клеток генетической конструкцией VTvaf17- Р4НА2 SEQ ID No: 4 с кДНК гена Р4НА2 представлены графики накопления ПЦР-продуктов, соответствующих:
1 - кДНК гена Р4НА2 в фибробластах с нормальной экспрессией гена Р4НА2,
2 - кДНК гена Р4НА2 в фибробластах со сниженной экспрессией гена Р4НА2 до трансфекции конструкцией с кДНК гена Р4НА2
3 - кДНК гена Р4НА2 в фибробластах со сниженной экспрессией гена Р4НА2 после трансфекции конструкцией с кДНК гена Р4НА2
4 - кДНК гена Р4НА2 в фибробластах со сниженной экспрессией гена Р4НА2 после трансфекции вектором без кДНК гена Р4НА2
5 - кДНК гена В2М в фибробластах с нормальной экспрессией гена Р4НА2,
6 - кДНК гена В2М в фибробластах со сниженной экспрессией гена Р4НА2 до трансфекции конструкцией с кДНК гена Р4НА2
7 - кДНК гена В2М в фибробластах со сниженной экспрессией гена Р4НА2 после трансфекции конструкцией с кДНК гена Р4НА2
8 - кДНК гена В2М в фибробластах со сниженной экспрессией гена Р4НА2 после трансфекции вектором без кДНК гена Р4НА2
Из графиков следует, что в случае трансфекции вектором без вставки кДНК гена Р4НА2 уровень кДНК гена Р4НА2 в фибробластах не изменился, а в случае трансфекции конструкцией VTvaf17- Р4НА2 SEQ ID No: 4 с кДНК Р4НА2 - уровень кДНК фибробластов со сниженной экспрессией гена Р4НА2 значительно увеличился.
На фиг. 13
С целью подтверждения увеличения экспрессии гена Р4НА2 в клеточной культуре фибробластов со сниженной экспрессией гена Р4НА2 при трансфекции данных клеток генетической конструкцией VTvaf17- Р4НА2 SEQ ID No: 5 с кДНК гена Р4НА2 представлены графики накопления ПЦР-продуктов, соответствующих:
1 - кДНК гена Р4НА2 в фибробластах с нормальной экспрессией гена Р4НА2,
2 - кДНК гена Р4НА2 в фибробластах со сниженной экспрессией гена Р4НА2 до трансфекции конструкцией с кДНК гена Р4НА2
3 - кДНК гена Р4НА2 в фибробластах со сниженной экспрессией гена Р4НА2 после трансфекции конструкцией с кДНК гена Р4НА2
4 - кДНК гена Р4НА2 в фибробластах со сниженной экспрессией гена Р4НА2 после трансфекции вектором без кДНК гена Р4НА2
5 - кДНК гена В2М в фибробластах с нормальной экспрессией гена Р4НА2,
6 - кДНК гена В2М в фибробластах со сниженной экспрессией гена Р4НА2 до трансфекции конструкцией с кДНК гена Р4НА2
7 - кДНК гена В2М в фибробластах со сниженной экспрессией гена Р4НА2 после трансфекции конструкцией с кДНК гена Р4НА2
8 - кДНК гена В2М в фибробластах со сниженной экспрессией гена Р4НА2 после трансфекции вектором без кДНК гена Р4НА2
Из графиков следует, что в случае трансфекции вектором без вставки кДНК гена Р4НА2 уровень кДНК гена Р4НА2 в фибробластах не изменился, а в случае трансфекции конструкцией VTvaf17- Р4НА2 SEQ ID No: 5 с кДНК Р4НА2 - уровень кДНК фибробластов со сниженной экспрессией гена Р4НА2 значительно увеличился.
На фиг. 14
С целью подтверждения увеличения экспрессии гена Р4НА2 в клеточной культуре фибробластов со сниженной экспрессией гена Р4НА2 при трансфекции данных клеток генетической конструкцией VTvaf17- Р4НА2 SEQ ID No: 6 с кДНК гена Р4НА2 представлены графики накопления ПЦР-продуктов, соответствующих:
1 - кДНК гена Р4НА2 в фибробластах с нормальной экспрессией гена Р4НА2,
2 - кДНК гена Р4НА2 в фибробластах со сниженной экспрессией гена Р4НА2 до трансфекции конструкцией с кДНК гена Р4НА2
3 - кДНК гена Р4НА2 в фибробластах со сниженной экспрессией гена Р4НА2 после трансфекции конструкцией с кДНК гена Р4НА2
4 - кДНК гена Р4НА2 в фибробластах со сниженной экспрессией гена Р4НА2 после трансфекции вектором без кДНК гена Р4НА2
5 - кДНК гена В2М в фибробластах с нормальной экспрессией гена Р4НА2,
6 - кДНК гена В2М в фибробластах со сниженной экспрессией гена Р4НА2 до трансфекции конструкцией с кДНК гена Р4НА2
7 - кДНК гена В2М в фибробластах со сниженной экспрессией гена Р4НА2 после трансфекции конструкцией с кДНК гена Р4НА2
8 - кДНК гена В2М в фибробластах со сниженной экспрессией гена Р4НА2 после трансфекции вектором без кДНК гена Р4НА2
Из графиков следует, что в случае трансфекции вектором без вставки кДНК гена Р4НА2 уровень кДНК гена Р4НА2 в фибробластах не изменился, а в случае трансфекции конструкцией VTvaf17- P4HA2 SEQ ID No: 6 с кДНК Р4НА2 - уровень кДНК фибробластов со сниженной экспрессией гена P4HA2 значительно увеличился.
На фиг. 15
С целью подтверждения увеличения экспрессии гена P4HA2 в клеточной культуре фибробластов со сниженной экспрессией гена P4HA2 при трансфекции данных клеток генетической конструкцией VTvaf17- P4HA2 SEQ ID No: 7 с кДНК гена P4HA2 представлены графики накопления ПЦР-продуктов, соответствующих:
1 - кДНК гена P4HA2 в фибробластах с нормальной экспрессией гена P4HA2,
2 - кДНК гена P4HA2 в фибробластах со сниженной экспрессией гена P4HA2 до трансфекции конструкцией с кДНК гена P4HA2
3 - кДНК гена P4HA2 в фибробластах со сниженной экспрессией гена P4HA2 после трансфекции конструкцией с кДНК гена P4HA2
4 - кДНК гена P4HA2 в фибробластах со сниженной экспрессией гена P4HA2 после трансфекции вектором без кДНК гена P4HA2
5 - кДНК гена В2М в фибробластах с нормальной экспрессией гена P4HA2,
6 - кДНК гена В2М в фибробластах со сниженной экспрессией гена P4HA2 до трансфекции конструкцией с кДНК гена P4HA2
7 - кДНК гена В2М в фибробластах со сниженной экспрессией гена P4HA2 после трансфекции конструкцией с кДНК гена P4HA2
8 - кДНК гена В2М в фибробластах со сниженной экспрессией гена P4HA2 после трансфекции вектором без кДНК гена P4HA2
Из графиков следует, что в случае трансфекции вектором без вставки кДНК гена P4HA2 уровень кДНК гена P4HA2 в фибробластах не изменился, а в случае трансфекции конструкцией VTvaf17- P4HA2 SEQ ID No: 7 с кДНК Р4НА2 - уровень кДНК фибробластов со сниженной экспрессией гена P4HA2 значительно увеличился.
На фиг. 16
С целью подтверждения увеличения количества белка пролил 4- гидрокислаза альфа 2 в клеточной культуре фибробластов с нормальной экспрессией гена P4HA2 при трансфекции данных клеток генетической конструкцией содержащей кДНК гена P4HA2, представлен график изменения количества белка пролил 4- гидрокислаза альфа 2 нетрансфицированных фибробластов (культура А), трансфицированных вектором pCDNA 3.1 (+) не содержащим кДНК P4HA2 (культура В) и трансфицированных генетической конструкцией pCDNA 3.1-P4HA2 SEQ ID No: 2 (культура С). Из графика следует, что при трансфекции фибробластов генетической конструкцией с кДНК гена P4HA2 происходит увеличение количества белка пролил 4- гидрокислаза альфа 2 в клеточном лизате.
На фиг. 17
С целью подтверждения увеличения количества белка пролил 4- гидрокислаза альфа 2 в коже человека при введении в кожу клеточной культуры фибробластов, трансфицированной генетической конструкцией содержащей кДНК гена P4HA2, представлена диаграмма изменения количества белка пролил 4- гидрокислаза альфа 2 в коже пациентов. При этом пациентам вводили три варианта культуры аутологичных фибробластов -нетрансфицированные (зона А), трансфицированные вектором VTvaf17 (зона В) и трансфицированные генетической конструкцией VTvaf17- P4HA2 SEQ ID No: 7 (зона С) - в кожу предплечья. Также анализировали количественный уровень белка пролил 4- гидрокислаза альфа 2 в интактной коже. Показано повышение количества белка пролил 4- гидрокислаза альфа 2 в коже пациента в области введения фибробластов, трансфицированных генетической конструкцией с кДНК гена P4HA2 (С).
На фиг. 18
С целью подтверждения увеличения количества белка пролил 4- гидрокислаза альфа 2 до различного индивидуального уровня в клеточных культурах фибробластов пациентов при трансфекции данных клеток генетическими конструкциями с модифицированными и нативной кДНК гена P4HA2 в зависимости от наличия и типа в них той или иной модификации кДНК гена P4HA2, представлен анализ изменения количественного уровня белка пролил 4- гидрокислаза альфа 2 в культурах фибробластов кожи человека в зависимости от наличия и типа модификаций в кДНК гена P4HA2, используемой для трансфекции фибробластов.
Культуры фибробластов 50 пациентов делили на 8 частей каждую с (А) по (Z); первые части (А) клеточных культур пациентов трансфицировали генетической конструкцией pCMV6- XL5 P4HA2 SEQ ID No: 1, части (В) трансфицировали генетической конструкцией pCMV6- XL5 P4HA2 SEQ ID No: 2, части (С) трансфицировали генетической конструкцией pCMV6- XL5 P4HA2 SEQ ID No: 3, части (D) трансфицировали генетической конструкцией pCMV6- XL5 P4HA2 SEQ ID No: 4, части (Е) трансфицировали генетической конструкцией pCMV6- XL5 P4HA2 SEQ ID No: 5, части (F) трансфицировали генетической конструкцией pCMV6- XL5 P4HA2 SEQ ID No: 6, части (G) трансфицировали генетической конструкцией pCMV6- XL5 Р4НА2 SEQ ID No: 7, части (Н) трансфицировали векторной плазмидой pCMV6- XL5, не содержащей кДНК гена Р4НА2.
По итогам анализа количественного уровня белка пролил 4- гидрокислаза альфа 2 выбрали показатели, касательно каждой части клеточной культуры от каждого пациента, продемонстрировавшие максимальное количество белка пролил 4- гидрокислаза альфа 2 и объединили их в группы, исходя из следующего критерия:
В группе 1 максимальное количество белка пролил 4- гидрокислаза альфа 2 наблюдалась при трансфекции pCMV6- XL5 Р4НА2 SEQ ID No: 1,
в группе 2 максимальное количество белка пролил 4- гидрокислаза альфа 2 наблюдалась при трансфекции pCMV6- XL5 Р4НА2 SEQ ID No: 2,
в группе 3 максимальное количество белка пролил 4- гидрокислаза альфа 2 наблюдалась при трансфекции pCMV6- XL5 Р4НА2 SEQ ID No: 3,
в группе 4 максимальное количество белка пролил 4- гидрокислаза альфа 2 наблюдалась при трансфекции pCMV6- XL5 Р4НА2 SEQ ID No: 4,
в группе 5 максимальное количество белка пролил 4- гидрокислаза альфа 2 наблюдалась при трансфекции pCMV6- XL5 Р4НА2 SEQ ID No: 5,
в группе 6 максимальное количество белка пролил 4- гидрокислаза альфа 2 наблюдалась при трансфекции pCMV6- XL5 Р4НА2 SEQ ID No: 6,
в группе 7 максимальное количество белка пролил 4- гидрокислаза альфа 2 наблюдалась при трансфекции pCMV6- XL5 Р4НА2 SEQ ID No: 7.
Ни в одной из клеточных культур не наблюдалось того, что максимальное количество белка пролил 4- гидрокислаза альфа 2 присутствует при трансфекции вектором pCMV6- XL5 без вставки кДНК гена Р4НА2.
На фигуре 18 для каждой группы клеточных культур приведена таблица показателей концентрации белка пролил 4- гидрокислаза альфа 2 (усредненных в рамках группы, в случае, если в группу входит более одной клеточной культуры) применительно ко всем, участвующим в эксперименте генетическим конструкциям после трансфекции этих клеточных культур генетическими конструкциями, содержащими модифицированные и нативную кДНК гена Р4НА2.
Из фигуры следует, что достижение максимального количества белка пролил 4- гидрокислаза альфа 2 в культурах фибробластов кожи различных пациентов при их трансфекции генетическими конструкциями связано с индивидуальными особенностями пациентов и зависит от наличия и типа модификаций в кДНК гена Р4НА2, встроенных в генетические конструкции.
Каждая генетическая конструкция из группы генетических конструкций является эффективной в некоторой значительной группе пациентов. Следовательно, для выбора наиболее эффективной генетической конструкции из группы генетических конструкций для терапевтических целей необходимо предварительное персонализированное исследование пациента.
На фиг. 19
С целью подтверждения увеличения экспрессии гена Р4НА2 в клеточной культуре кератоцитов и эпителиальных клеток роговицы глаза при трансфекции данных клеток генетической конструкцией pCMV6-Kan/Neo P4HA2 SEQ ID No: 2 с кДНК гена Р4НА2 приведены графики накопления ПЦР-продуктов, соответствующих:
1 - кДНК гена P4HA2, кератоциты до трансфекции конструкцией с кДНК гена P4HA2
2 - кДНК гена P4HA2, эпителий роговицы до трансфекции конструкцией с кДНК гена P4HA2
3 - кДНК гена P4HA2, кератоциты после трансфекции конструкцией с кДНК гена P4HA2
4 - кДНК гена P4HA2, эпителий роговицы после трансфекции конструкцией с кДНК гена P4HA2
5 - кДНК гена В2М, кератоциты до трансфекции конструкцией с кДНК гена P4HA2
6 - кДНК гена В2М, эпителий роговицы до трансфекции конструкцией с кДНК гена P4HA2
7 - кДНК гена В2М, кератоциты после трансфекции конструкцией с кДНК гена P4HA2
8 - кДНК гена В2М, эпителий роговицы после трансфекции конструкцией с кДНК гена P4HA2
Ген В2М использовали в качестве референтного.
Из фигуры следует, что в результате трансфекции генетической конструкцией pCMV6-Kan/Neo P4HA2 SEQ ID No: 2 уровень специфической кДНК гена P4HA2 в культуре кератоцитов и в культуре эпителия роговицы многократно вырос.
На фиг. 20
С целью подтверждения увеличения экспрессии гена P4HA2 в первичной культуре гладкомышечных клеток мочевого пузыря человека HBdSMC (ATCC PCS-420-012) при трансфекции данных клеток генетической конструкцией GDTT1.8NAS1-P4HA2 SEQ ID No: 3 с кДНК гена Р4НА2 приведены графики накопления ПЦР-продуктов, соответствующих:
1 - кДНК гена Р4НА2 до трансфекции генетической конструкцией GDTT1.8NAS1-Р4НА2 SEQ ID No: 3, несущей участок гена Р4НА2;;
2 - кДНК гена Р4НА2 после трансфекции генетической конструкцией GDTT1.8NAS1-Р4НА2 SEQ ID No: 3, несущей участок гена Р4НА2;
3 - кДНК гена В2М до трансфекции генетической конструкцией GDTT1.8NAS1-Р4НА2 SEQ ID No: 3, несущей участок гена Р4НА2;
4 - кДНК гена В2М после трансфекции генетической конструкцией GDTT1.8NAS1-Р4НА2 SEQ ID No: 3, несущей участок гена Р4НА2;
Ген В2М использовали в качестве референтного.
Из фигуры следует, что в результате трансфекции генетической конструкцией GDTT1.8NAS1-Р4НА2 SEQ ID No: 3 уровень специфической кДНК гена Р4НА2 в культуре гладкомышечных клеток мочевого пузыря человека HBdSMC многократно вырос.
На фиг. 21
С целью подтверждения увеличения экспрессии гена Р4НА2 в клеточной культуре хондробластов при трансфекции данных клеток генетической конструкцией pCMV6-Kan/Neo Р4НА2 SEQ ID No: 4 с кДНК гена Р4НА2 приведены графики накопления ПЦР-продуктов, соответствующих:
1 - кДНК гена Р4НА2, до трансфекции pCMV6-Kan/Neo Р4НА2 SEQ ID No: 4
2 - кДНК гена Р4НА2, после трансфекции pCMV6-Kan/Neo Р4НА2 SEQ ID No: 4
3 - кДНК гена В2М, до трансфекции pCMV6-Kan/Neo Р4НА2 SEQ ID No: 4
4 - кДНК гена В2М, после трансфекции pCMV6-Kan/Neo Р4НА2 SEQ ID No: 4
Ген В2М использовали в качестве референтного.
Из фигуры следует, что в результате трансфекции хондробластов генетической конструкцией pCMV6-Kan/Neo Р4НА2 SEQ ID No: 3 уровень специфической кДНК гена Р4НА2 вырос многократно.
На фиг. 22
С целью подтверждения увеличения экспрессии гена Р4НА2 в первичной культуре эндотелиальных клеток из пупочной вены человека HUVEC (ATCC® CRL-1730™) при трансфекции данных клеток генетической конструкцией GDTT1.8NAS3-P4HA2 SEQ ID No: 4 с кДНК гена Р4НА2 приведены графики накопления ПЦР-продуктов, соответствующих:
1 - кДНК гена Р4НА2 до трансфекции генетической конструкцией GDTT1.8NAS3-P4HA2 SEQ ID No: 4, несущей участок гена Р4НА2;;
2 - кДНК гена Р4НА2 после трансфекции генетической конструкцией GDTT1.8NAS3-P4HA2 SEQ ID No: 4, несущей участок гена Р4НА2;
3 - кДНК гена В2М до трансфекции трансфекции генетической конструкцией GDTT1.8NAS3-P4HA2 SEQ ID No: 4, несущей участок гена Р4НА2;;
4 - кДНК гена В2М после трансфекции генетической конструкцией GDTT1.8NAS3-P4HA2 SEQ ID No: 4, несущей участок гена Р4НА2;
Ген В2М использовали в качестве референтного.
Из фигуры следует, что в результате трансфекции генетической конструкцией GDTT1.8NAS3-P4HA2 SEQ ID No: 4 уровень специфической кДНК гена Р4НА2 в первичной культуре эндотелиальных клеток из пупочной вены человека HUVEC (АТСС® CRL-1730™) многократно вырос.
На фиг. 23
С целью подтверждения увеличения количества белка пролил-4- гидрокислаза альфа 2 в коже человека при введении в кожу генетической конструкции представлен анализ изменения количественного уровня белка пролил 4- гидрокислаза альфа 2 в коже. При этом пациенту вводили генетическую конструкцию PCMV6-XL5 Р4НА2 SEQ ID No: 5 с кДНК гена Р4НА2 (зона В) и параллельно вводили плацебо, представляющее собой комбинацию векторной плазмиды pCMV6-XL5 не содержащей кДНК гена Р4НА2 с транспортной молекулой (зона А) - в кожу предплечья. Показано увеличение количества белка в биоптате кожи пациента в зоне В, в которую вводилась генетическая конструкция pCMV6- XL5 Р4НА2 SEQ ID No: 5, что говорит об эффективности данной генетической конструкции.
На фиг. 24
С целью подтверждения увеличения количества белка пролил-4- гидрокислаза альфа 2 в хрящевой ткани человека при введении в эту ткань генетической конструкции VTvaf17-P4HA2 SEQ ID No: 6 с кДНК гена Р4НА2, представлен анализ изменения количественного уровня белка пролил 4- гидрокислаза альфа 2 в хрящевой ткани. При этом пациенту вводили генетическую конструкцию VTvaf17-P4HA2 SEQ ID No: 6 с кДНК гена Р4НА2 (зона В) и параллельно вводили плацебо, представляющее собой комбинацию векторной плазмиды VTvaf17 не содержащей кДНК гена Р4НА2 (зона А) - в хрящевую ткань.
Показано увеличение количественного уровня белка пролил 4- гидрокислаза альфа 2 в лизате биоптата хрящевой ткани пациента в зоне В, в которую вводились генетическая конструкция VTvaf17-P4HA2 SEQ ID No: 6, с кДНК гена Р4НА2, что говорит об эффективности данной генетической конструкции.
На фиг. 25
С целью подтверждения увеличения количества белка пролил-4- гидрокислаза альфа 2 в мышечной ткани человека при введении в эту ткань генетической конструкции GDTT1.8NAS1-Р4НА2 SEQ ID No: 7 с кДНК гена Р4НА2, представлен анализ изменения количества белка пролил 4- гидрокислаза альфа 2 в мышечной ткани.
При этом пациенту вводили генетическую конструкцию с кДНК гена Р4НА2 - GDTT1.8NAS1-P4HA2 SEQ ID No: 7 (зона В) и параллельно вводили плацебо, представляющее собой комбинацию векторной плазмиды GDTT1.8NAS1 не содержащей кДНК гена Р4НА2 с транспортной молекулой (зона А) - в мышечную ткань предплечья. Показано увеличение количества белка пролил-4- гидрокислаза альфа 2 в биоптате мышечной ткани пациента в зоне В, в которую вводилась генетическая конструкция GDTT1.8NAS1-P4HA2 SEQ ID No: 7 с кДНК гена Р4НА2, что говорит об эффективности данной генетической конструкции.
На фиг. 26
С целью подтверждения увеличения количества белка пролил-4-гидрокислазы альфа 2 до различного индивидуального уровня при введении в кожу пациентов генетической конструкции с модифицированными и нативной кДНК гена Р4НА2 анализировали количественный уровень белка пролил 4-гидрокислазы альфа 2 в коже человека в зависимости от наличия и типа модификаций в кДНК гена Р4НА2.
Каждому из 32-х пациентов, отобранных в случайном порядке, вводили в кожу предплечья 7 генетических конструкций pCMV6- XL5 Р4НА2 SEQ ID No: 1, pCMV6- XL5 Р4НА2 SEQ ID No: 2, pCMV6- XL5 Р4НА2 SEQ ID No: 3, pCMV6- XL5 Р4НА2 SEQ ID No: 4, pCMV6- XL5 Р4НА2 SEQ ID No: 5, pCMV6- XL5 Р4НА2 SEQ ID No: 6, pCMV6- XL5 Р4НА2 SEQ ID No: 7, и плацебо pCMV6- XL5.
По итогам анализа количества белка пролил 4-гидрокислазы альфа 2 в биоптатах выбрали показатели, касательно каждого биоптата от каждого пациента, продемонстрировавшие максимальные количественные уровни белка пролил 4-гидрокислазы альфа 2 и объединили их в группы, исходя из следующего критерия:
В группе 1 максимальное количество белка пролил 4-гидрокислазы альфа 2 наблюдалась при введении pCMV6- XL5 Р4НА2 SEQ ID No: 1,
в группе 2 максимальное количество белка пролил 4-гидрокислазы альфа 2 наблюдалась при введении pCMV6- XL5 Р4НА2 SEQ ID No: 2,
в группе 3 максимальное количество белка пролил 4-гидрокислазы альфа 2 наблюдалась при введении pCMV6- XL5 Р4НА2 SEQ ID No: 3,
в группе 4 максимальное количество белка пролил 4-гидрокислазы альфа 2 наблюдалась при введении pCMV6- XL5 Р4НА2 SEQ ID No: 4,
в группе 5 максимальное количество белка пролил 4-гидрокислазы альфа 2 наблюдалась при введении pCMV6- XL5 Р4НА2 SEQ ID No: 5,
в группе 6 максимальное количество белка пролил 4-гидрокислазы альфа 2 наблюдалась при введении pCMV6- XL5 Р4НА2 SEQ ID No: 6,
в группе 7 максимальное количество белка пролил 4-гидрокислазы альфа 2 наблюдалась при введении pCMV6- XL5 Р4НА2 SEQ ID No: 7.
Ни в одном из биоптатов не наблюдалось того, что максимальное количество белка пролил 4-гидрокислазы альфа 2 присутствует в случае введения плацебо.
На фигуре 26 для каждой группы биоптатов приведены диаграммы показателей концентрации белка пролил 4-гидрокислазы альфа 2 (усредненных в рамках группы, в случае, если в группу входит более одного биоптата) применительно ко всем, участвующим в эксперименте генетическим конструкциям, после введения пациентам этих генетическмх конструкций, содержащих модифицированные и нативную кДНК гена Р4НА2.
Из данного примера следует, что достижение максимального количества белка пролил 4-гидрокислазы альфа 2 в биоптатах кожи различных пациентов при введении им в кожу генетических конструкций, связано с индивидуальными особенностями пациентов и зависит от наличия и типа модификаций в кДНК гена Р4НА2, входящих в генетическую конструкцию.
Каждая генетическая конструкция из группы генетических конструкций является эффективной в некоторой значительной группе пациентов. Следовательно, для выбора наиболее эффективной генетической конструкции из группы генетических конструкций для терапевтических целей, необходимо предварительное персонализированное исследование пациента.
Обозначения:
 биоптаты пациентов после введения ГК Р4НА2 SEQ ID No: 1 (A)
биоптаты пациентов после введения ГК Р4НА2 SEQ ID No: 1 (A)
 биоптаты пациентов после введения ГК Р4НА2 SEQ ID No: 2 (B)
биоптаты пациентов после введения ГК Р4НА2 SEQ ID No: 2 (B)
 биоптаты пациентов после введения ГК Р4НА2 SEQ ID No: 3 (C)
биоптаты пациентов после введения ГК Р4НА2 SEQ ID No: 3 (C)
 биоптаты пациентов после введения ГК Р4НА2 SEQ ID No: 4 (D)
биоптаты пациентов после введения ГК Р4НА2 SEQ ID No: 4 (D)
 биоптаты пациентов после введения ГК Р4НА2 SEQ ID No: 5 (E)
биоптаты пациентов после введения ГК Р4НА2 SEQ ID No: 5 (E)
 биоптаты пациентов после введения ГК Р4НА2 SEQ ID No: 6 (F)
биоптаты пациентов после введения ГК Р4НА2 SEQ ID No: 6 (F)
 биоптаты пациентов после введения ГК Р4НА2 SEQ ID No: 7 (G)
биоптаты пациентов после введения ГК Р4НА2 SEQ ID No: 7 (G)
 биоптаты пациентов после введения плацебо (Н)
биоптаты пациентов после введения плацебо (Н)
На фиг. 27
С целью определения наиболее эффективной применительно к конкретному пациенту генетической конструкции с кДНК гена Р4НА2 анализировали количественный уровень белка пролил 4- гидрокислаза альфа 2 в клеточных лизатах фибробластов этого пациента, трансфицированных разными генетическими конструкциями, содержащими нативную и модифицированные кДНК гена Р4НА2.
По итогам анализа количества белка пролил 4- гидрокислаза альфа 2 в культуре фибробластов пациента выделили вариант генетической конструкции, при трансфекции которой выявляется максимальная концентрация белка пролил 4- гидрокислаза альфа 2 в лизате клеточной культуры. В данном эксперименте максимальная концентрация белка пролил 4- гидрокислаза альфа 2 в лизате наблюдается при трансфекции генетической конструкцией pCMV6- XL5 Р4НА2 SEQ ID No: 4, содержащей модифицированную кДНК Р4НА2, что показано на фигуре.
Таким образом, выбрана наиболее эффективная применительно к данному пациенту генетическая конструкция для последующей трансфекции клеток пациента в рамках терапевтической процедуры.
Обозначения:
1 - клеточный лизат после трансфекции pCMV6- XL5 Р4НА2 SEQ ID No: 1 (A)
2 - клеточный лизат после трансфекции pCMV6- XL5 Р4НА2 SEQ ID No: 2 (В)
3 - клеточный лизат после трансфекции pCMV6- XL5 Р4НА2 SEQ ID No: 3 (С)
4 - клеточный лизат после трансфекции pCMV6- XL5 Р4НА2 SEQ ID No: 4 (D)
5 - клеточный лизат после трансфекции PCMV6- XL5 Р4НА2 SEQ ID No: 5 (E)
6 - клеточный лизат после трансфекции PCMV6- XL5 Р4НА2 SEQ ID No: 6 (F)
7 - клеточный лизат после трансфекции PCMV6- XL5 Р4НА2 SEQ ID No: 7 (G)
8 - клеточный лизат после трансфекции плацебо (Н)
На фиг. 28
С целью определения наиболее эффективной применительно к конкретному пациенту генетической конструкции с кДНК гена Р4НА2 анализировали количество белка пролил-4- гидрокислаза альфа 2 в супернатанте биоптатов кожи этого пациента, после введения ему генетических конструкций, содержащих нативную и модифицированные кДНК гена Р4НА2.
По итогам анализа количества белка пролил 4- гидрокислаза альфа 2 в супернатанте биоптатов кожи пациента выделили вариант генетической конструкции, при введении которой выявляется максимальная концентрация белка пролил-4- гидрокислаза альфа 2 в супернатанте биоптата. В данном эксперименте максимальная концентрация белка пролил-4- гидрокислаза альфа 2 отмечена при введении генетической конструкции на базе pCMV6-XL5 Р4НА2 SEQ ID No: 1, содержащей нативную кДНК Р4НА2, что показано на фигуре 64.
Таким образом, выбрана наиболее эффективная применительно к данному пациенту генетическая конструкция для ее последующего введения пациенту в рамках терапевтической процедуры.
Обозначения:
1 - супернатант биоптата после введения PCMV6- XL5 Р4НА2 SEQ ID No: 1 (А)
2 - супернатант биоптата после введения PCMV6- XL5 Р4НА2 SEQ ID No: 2 (В)
3 - супернатант биоптата после введения PCMV6- XL5 Р4НА2 SEQ ID No: 3 (С)
4 - супернатант биоптата после введения PCMV6-XL5 Р4НА2 SEQ ID No: 4 (D)
5 - супернатант биоптата после введения PCMV6- XL5 Р4НА2 SEQ ID No: 5 (E)
6 - супернатант биоптата после введения PCMV6-XL5 Р4НА2 SEQ ID No: 6 (F)
7 - супернатант биоптата после введения PCMV6- XL5 Р4НА2 SEQ ID No: 7 (G)
8 - супернатант биоптата после введения плацебо (Н)
Реализация изобретения.
При снижении экспрессии генов, кодирующих белки синтеза пролил-4- гидрокислаза альфа-1 и альфа- 2, например, гена Р4НА2, наблюдается изменение структуры органов и тканей организма. Несмотря на то, что мыши, с нокаутированными обоими аллелями гена Р4НА2 (P4HA2-/-) не демонстрировали аномалий в развитии, у мышей с двойным нокаутом по альфа-1 (P4HA1-/+) и альфа-2 (P4HA2-/-) субъединицам наблюдалось задержка в росте, умеренная хондродисплазия, кифоз. На клеточном уровне было замечено радикальное изменение структуры внеклеточного матрикса (Aro E, Salo AM, Khatri R,  M, Miinalainen I, Sormunen R, Pakkanen O, Holster T, Soininen R, Prein C, Clausen-Schaumann Н,
M, Miinalainen I, Sormunen R, Pakkanen O, Holster T, Soininen R, Prein C, Clausen-Schaumann Н,  A, Tuukkanen J, Kivirikko Kl, Schipani E, Myllyharju J. Severe Extracellular Matrix Abnormalities and Chondrodysplasia in Mice Lacking Collagen Prolyl 4-Hydroxylase Isoenzyme II in Combination with a Reduced Amount of Isoenzyme I. J Biol Chem. 2015.290(27):16964-78.)
A, Tuukkanen J, Kivirikko Kl, Schipani E, Myllyharju J. Severe Extracellular Matrix Abnormalities and Chondrodysplasia in Mice Lacking Collagen Prolyl 4-Hydroxylase Isoenzyme II in Combination with a Reduced Amount of Isoenzyme I. J Biol Chem. 2015.290(27):16964-78.)
Преимущества использования генетической конструкции с геном Р4НА2 для коррекции экспрессии гена Р4НА2 и количества белка пролил-4-гидрокислаза альфа 2 в клетках органов и тканей человека:
1) легче обеспечить более высокий и стабильный уровень целевого белка в клетках,
2) обеспечивается транспортировка генетической конструкции в больший спектр клеток органов и тканей человека, а также более эффективная внутриклеточная транспортировка генетической конструкции.
3) учитываются индивидуальные характеристики пациента. Таким образом, для купирования проявлений состояний человеческого организма, связанных с уменьшением экспрессии гена Р4НА2 и/или уменьшением количества белка пролил 4-гидрокислаза альфа 2, вместо применения препаратов, содержащих белок пролил 4-гидрокислаза альфа 2, по данному изобретению создана генетическая конструкция, выбранная из группы генетических конструкций с одной из модифицированных кДНК гена Р4НА2 или с участком нативной немодифицированной кДНК гена Р4НА2, кодирующих белок пролил 4-гидрокислаза альфа 2.
Для получения генетической конструкции на основе невирусной векторной плазмиды, включающей кДНК гена Р4НА2, для купирования проявлений состояний человеческого организма, связанных с уменьшением экспрессии гена Р4НА2 и/или уменьшением количества белка пролил 4-гидрокислаза альфа 2, выбранной из группы генетических конструкций с кДНК гена Р4НА2, осуществляют следующие действия:
1. Получение участка немодифицированной кДНК гена Р4НА2, содержащей белок-кодирующую область гена Р4НА2 и делеции 5ʹ нетранслируемых областей и делеции 3ʹ-нетранслируемых областей, клонирование его в промежуточнуюплазмиду, переклонирование его в экспрессионные векторные плазмиды под контроль эукариотических регуляторных элементов для экспрессии целевого гена таким образом, что полученные генетические конструкции содержат нуклеотидную последовательность, кодирующую аминокислотную последовательность белка пролил 4- гидрокислаза альфа 2, которая является участком немодифицированной кДНК гена Р4НА2, и которая используется для дальнейших модификаций.
2. Внесение в последовательность нуклеотидов кДНК гена Р4НА2 модификаций с целью создания ряда кДНК гена Р4НА2, обеспечивающих достаточный уровень трансляции белка пролил 4- гидрокислаза альфа 2 для коррекции патологических состояний.
3. Клонирование участка немодифицированной кДНК гена Р4НА2 и полученных на его основе модифицированных кДНК гена Р4НА2 в векторные плазмиды, способные обеспечить эффективную экспрессию этой кДНК в клетках органов и тканей человека.
4. Трансформация каждой из полученных генетических конструкций бактериальных клеток E.coli, анализ трансформированных клонов на предмет наличия, ориентации и копийности вставки кДНК и наращивание отобранных клонов для получения необходимого для дальнейшей работы количества вариантов плазмидной ДНК.
5. Выделение ряда генетических конструкций, содержащих модифицированные кДНК гена Р4НА2 и участок немодифицированной кДНК гена Р4НА2 для создания на их базе группы высокоэффективных генетических конструкций.
6. Создание высокоэффективных генетических конструкций, каждая из которых включает генетическую конструкцию с модифицированными кДНК гена Р4НА2 и участок немодифицированной кДНК гена Р4НА2, или комбинацию такой конструкции с транспортной молекулой для эффективной трансфекции различных типов клеток органов и тканей и/или введения в органы и ткани человека.
Для доказательства эффективности созданных генетических конструкций, приводящих к увеличению экспрессии гена Р4НА2, проводят следующие исследования:
A) Выращивание культур различных типов клеток из биоптатов различных органов и тканей человека или с использованием коммерческих клеточных линий, например, фибробластов кожи, или хондробластов, или кератоциов или эпителиальных клеток роговицы или эндотелиальных клеток из пупочной вены человека.
B) Выделение РНК из культуры клеток, например, фибробластов кожи, или хондробластов, или кератоциов или эпителиальных клеток роговицы или эндотелиальных клеток из пупочной вены человека, и анализ экспрессии гена Р4НА2 из генома этих клеток.
C) Трансфекция культуры клеток, например, фибробластов кожи, или хондробластов, или кератоциов или эпителиальных клеток роговицы или кератоциты и эпителиальные клетки роговицы или эндотелиальных клеток из пупочной вены человека, генетическими конструкциями содержащими немодифицированную и/или модифицированные кДНК гена Р4НА2 и параллельная трансфекция культуры этих же клеток векторной плазмидой, не содержащей кДНК гена Р4НА2 в различных комбинациях.
D) Сравнительный анализ изменения уровня кДНК и/или изменения количества белка пролил 4- гидрокислаза альфа 2 после трансфекции в различных комбинациях клеток, например, фибробластов кожи, или хондробластов, или кератоциов или эпителиальных клеток роговицы или эндотелиальных клеток из пупочной вены человека, генетическими конструкциями, содержащими немодифицированную и/или модифицированные кДНК гена Р4НА2 и параллельной трансфекции культуры этих же клеток векторной плазмидой, не содержащей кДНК гена Р4НА2. Данный анализ проводят, а именно, перед предполагаемой терапией с целью определения наиболее эффективной для пациента генетической конструкции.
Е) Введение пациентам в органы и ткани культуры клеток с кДНК гена Р4НА2, например, введение, в кожу человека, аутологичных фибробластов, трансфицированных генетической конструкцией с кДНК гена Р4НА2 и параллельное введение пациентам в органы и ткани, например, в кожу, культур этих же клеток нетрансфицированных и трансфицированных вектором без вставки кДНК гена Р4НА2 и/или введение пациентам в органы и ткани генетической конструкции, например, введение в кожу человека генетических конструкций, содержащих немодифицированную и/или модифицированные кДНК гена Р4НА2, а также плацебо в различных комбинациях.
F) Сравнительный анализ количества белка пролил-4- гидрокислаза альфа 2 в органах и тканях человека, а именно в коже человека, после введения клеток нетрансфицированных и трансфицированных генетическими конструкциями, содержащими кДНК гена Р4НА2 и векторными плазмидами, не содержащими кДНК гена Р4НА2 и/или после введения генетических конструкций, содержащих немодифицированную или модифицированные кДНК гена Р4НА2, а также плацебо.
G) Введение пациентам в органы и ткани, например, в кожу, в хрящевую ткань, или в мышечную ткань генетических конструкций, содержащих немодифицированную и/или модифицированные кДНК гена Р4НА2, а также плацебо.
Н) Сравнительный анализ количества белка пролил-4- гидрокислаза альфа 2 в органах и тканях человека, а именно в коже, в хрящевой ткани, или в мышечной ткани человека, после введения генетических конструкций, содержащих немодифицированную и/или модифицированные кДНК Р4НА2 также плацебо.
I) Трансфекция культуры клеток, например, фибробластов кожи пациентов генетическими конструкциями, содержащими немодифицированную и модифицированные кДНК гена Р4НА2; введение пациентам в органы и ткани, например, введение в кожу пациента генетических конструкций, содержащих немодифицированную и модифицированные кДНК гена Р4НА2.
J) Сравнительный анализ количества белка пролил-4- гидрокислаза альфа 2 в клеточных лизатах, например лизатах фибробластов кожи после трансфекции этих клеток генетическими конструкциями, содержащих немодифицированную и модифицированные кДНК гена Р4НА2; или в супернатантах биоптатов органов и тканей пациента, например, в супернатантах биоптатов кожи, после введения в кожу генетических конструкций, содержащих немодифицированную и модифицированные кДНК гена Р4НА2. Данный анализ проводят, а именно перед предполагаемой терапевтической процедурой с целью выбора наиболее эффективной для пациента генетической конструкции, выбранной из группы.
Пример 1.
Получение участка нативной (немодифицированной) кДНК гена Р4НА2.
Немодифицированная кДНК гена Р4НА2 представляет собой последовательность нуклеотидов, гомологичную белок кодирующей последовательности, приводимой в базе даных GenBank под номером NM_004199; нуклеотиды с 565 по 2172 транслируются в аминокислотную последовательность, соответствующую приведенной в GenBank под номером NP_004190.1.
Суммарную РНК получают из клеток человеческой крови с помощью набора PAXGeneBlood RNA Kit (Qiagen, Germany) ииспользуют для получения суммарной кДНК путем обратной транскрипции с помощью обратной транскриптазы RevertAid (ThermoScientific, USA) и случайных 9-нуклеотидных праймеров по методике производителя обратной транскриптазы. Затем, используя суммарную кДНК в качестве матрицы и специфичные, предварительно кинированные праймеры, комплементарные нуклеотидам
160-179 5' GAGCGCTGTGCTGGAAGGGA 3' (P4HA2-F1)
и
2371-2352 5' GGACAGCAGAGCCAGCACTTGA 3' (P4HA2-R1)
из последовательности мРНК Р4НА2 (GenBank NM_004199), получают участок нативной (немодифицированной) кДНК Р4НА2, содержащий белок-кодирующую область гена Р4НА2 и частичные делеции 5' и 3'-нетранслируемых областей. Полимеразную цепную реакцию (ПЦР) проводят с помощью ДНК-полимеразы Phusion (ThermoScientific, USA), дающей продукты с тупыми концами. ПЦР проводят с помощью амплификатора Master CyclerGradient (Eppendorf, USA) в 50 мкл. реакционной смеси, содержащей 0,5 мкл суммарной кДНК первой цепи, по 0,1 мкМ каждого праймера Р4НА2 F1 и Р4НА2 R1, 250 мкМ каждого дезоксинуклеотидтрифосфата (дАТФ, дЦТФ, дТТФ и дГТФ), 100 мМтрис-HCl (рН 8,85 при 20°С), 50 мМ сульфата аммония, 250 мМ хлористого калия, 0,01% Твин-20 и 5 ед. Pfu ДНК-полимеразы (PhusionThermoScientific.USA) при следующих условиях: первоначальная денатурация при +98°С в течение 30 сек., 35 циклов, включающих денатурацию при +98°С в течение 10 сек., отжиг праймеров при +64°С в течение 30 сек. и элонгацию при +68°С течение 4 минут.
Продукт амплификации выделяют из агарозного геля с помощью набора QIAquickGelExtractionKit (Qiagen, Germany)
Амплифицированный фрагмент кДНК длиной 1608 н.п, несущий участок нативной (немодифицированной) кДНК Р4НА2, клонируют в плазмидном векторе pUC19 (NEB, USA, кат номер N3041S). ДНК векторной плазмиды pUC19 (NEB, USA, кат. номер N3041S) гидролизуют рестриктазой рестриктазой, дающей тупые концы, например, Smal (NEB, USA), и обрабатывают щелочной фосфатазой. Амплифицированный фрагмент кДНК и линеаризованную плазмиду pUC19 лигируют с помощью ДНК-лигазы фага Т4 400 000 ед./ мл. (NEB, USA, кат номер M0202S) из расчета 1 мкл фермента на 1 мкг ДНК. Лигирование проводят в объеме 20 мкл в присутствии 2 мМ АТФ, 50 мМ трис-HCl, рН 7,6, 10 мМ MgCl2, 10 мМ DTT в течение 10 ч при +160С.
Полученной смесью трансформируют компетентные клетки Е.coliTop10 (http://molbiol.ru/protocol/03_04.html), которые высевают на чашки Петри с L-агаром, содержащих 100 мкг/мл ампициллина и по 40 мкл на чашку растворов 100 мМ ИПТГ и 2% X-gal. Отдельные колонии анализируют на наличие вставки с помощью ПЦР с праймерами P4HA2-F1 и P4HA2-R1 и подтверждают секвенированием по методу Сэнгера.
Пример 2.
Получение генетических конструкций с участком немодифицированной кДНК гена Р4НА2
Для экспрессии участка немодифицированной кДНК гена Р4НА2 в клетках органов и тканей человека вариант кДНК гена Р4НА2 по Примеру 1 помещают в векторную плазмиду, при выборе которой используют следующие критерии выбора:
1) Плазмида должна обязательно реплицироваться в E.coli, желательно с высокой копийностью;
2) В плазмиде должен быть бактериальный фактор селекции, либо другой фактор селекции не содержащий генов резистентности к антибиотикам;
3) Плазмида должна содержать эукариотические регуляторные элементы - промотор и терминатор, сигнала полиаденилирования, энхансер и интронный(е) элемент(ы);
4) В плазмиде должно быть наличие удобного полилинкера для клонирования.
5) Плазмида может содержать нуклеотидные последовательности с регуляторными элементами для репликации в клетках млекопитающих, например, такие как SV40 ori из вируса обезьян SV40 или ori P/EBNA-1 из вируса Эпштейн-Барра человека.
6) Плазмида также может содержать дополнительные регуляторные элементы, усиливающие трансляцию белка, например, участки связывания с транскрипционным фактором NFkB, обеспечивающим активный транспорт плазмидной ДНК в клеточное ядро для более эффективной транскрипции целевого гена.
Примерами плазмид, отвечающим таким критериям могут быть pCMV6-XL5, pCMV6-Kan/Neo (OriGene, USA), VTvaf17 (Cell and Gene Therapy Ltd, UK), GDTT1.8NAS1, GDTT1.8NAS3, GDTT1.8NAS7 (Genetic Diagnostics and Therapy 21 LTD, UK) или pCDNA 3.1 (+) (ThermoFisherScientific, USA), либо плазмидные векторы вирусного происхождения - например, аденовирус человека 5-го серотипа.
При клонировании участка немодифицированой кДНК гена Р4НА2 в pCDNA 3.1 (+) кодирующая нуклеотидная последовательность помещается под контроль промотора цитомегаловируса человека CMV, эффективного в эукариотических клетках, и сигнала полиаденилирования гена бычьего гормона роста. Также данная плазмида обладает бактериальным фактором селекции - геном устойчивости к ампициллину, и эукариотическим фактором селекции - геном устойчивости к неомицину.
При клонировании участка немодифицированой кДНК гена Р4НА2 в pCMV6-XL5, pCMV6-Kan/Neo кодирующая нуклеотидная последовательность помещается под контроль промотора цитомегаловируса человека CMV, эффективного в эукариотических клетках, и сигнала полиаденилирования гена гормона роста человека. Также данная плазмида обладает бактериальным фактором селекции - геном устойчивости к ампициллину.
При клонировании участка немодифицированной кДНК гена Р4НА2 в VTvaf17 кодирующая нуклеотидная последовательность помещается под контроль промотора гена человеческого фактора элонгации EF1A с собственным энхансером, эффективного в эукариотических клетках и сигнала полиаденилирования гена фактора роста человека. Также данная плазмида обладает фактором селекции, обеспечивающим возможность положительной селекции без использования антибиотиков.
При клонировании участка немодифицированной кДНК гене Р4НА2 в GDTT1.8NAS1, GDTT1.8NAS3, GDTT1.8NAS7 кодирующая нуклеотидная последовательность помещается по/ контроль промоторов генов десмина, эндоглина и нефринс человека соответственно, имеющих собственный энхансер и сигнал полиаденилирования гена фактора роста человека. Данньк плазмиды также обладает фактором селекции, обеспечивающим возможность положительной селекции без использована антибиотиков.
Для получения генетических конструкций с участком немодифицированной кДНК гена Р4НА2 векторную плазмиду pUC19-P4HA2 по Примеру 1 с участком немодифицированной кДНК Р4НА2, содержащим белок-кодирующую область гена Р4НА2 и частичные делеции 5' и 3'-нетранслируемых областей, используют в качестве матрицы для амплификации со следующими парами праймеров:
Р4НА2 F2 5' GGAGATCTGCCACCATGAAACTCTGGGTG 3'
Р4НА2 R2 5' CCTCTAGATCAGTCAACTTCTGTTGATCC 3'
Амплификацию проводят при условиях, приведенных в Примере 1. Далее продукты амплификации переосаждают этиловым спиртом, осадки промывают 70% этанолом и растворяют в 1× ТЕ буфере, после чего подвергают рестрикции с эндонуклеазами EcoRI и XbaI.
Продукты рестрикции разделяют с помощью электрофореза в геле 1% агарозы, окрашивают раствором бромистого этидия и фрагменты ДНК, соответствующие гену-вставке EcoRI -Р4НА2- XbaI, и линеаризованной плазмиде PCMV6-XL5 и pCMV6-Kan/Neo и pCDNA 3.1(+), вырезают, выделяют из геля с помощью набора для выделения из геля QIAquickGelExtractionKit (Qiagen, Germany) и смешивают. Полученную смесь рестрикционных фрагментов лигируют с помощью ДНК-лигазы фага Т4. Лигирование проводят в течение 10-15 мин при комнатной температуре, а затем - при 12°С в течение 10 ч. Лигированную ДНК используют для трансформации клеток Е. coli по стандартной методике с использованием хлористого кальция (Маниатис Т., и др. Молекулярное клонирование. М., Мир, 1984). Трансформированные клетки отбирают на агаризованной среде LB с ампициллином (100 мкг/мл). Плазмидную ДНК из ампициллин-устойчивых колоний выделяют с помощью набора для выделения плазмид QiagenSpinMiniprepKit (Qiagen, Germany), анализируют с помощью гидролиза рестриктазами EcoRI и XbaI и отбирают генетическую (ие) конструкцию (ии) с помощью секвенирования ДНК генетических конструкций по методу Сэнгера.
Для клонирования гена-вставки EcoRI -Р4НА2- XbaI в вектора VTvaf17 и GDTT1.8NAS1, GDTT1.8NAS3, GDTT1.8NAS7 после выделения ее из геля дополнительно обрабатывают 2 ед.акт. Т4 ДНК полимеразой E.coli и 2 ед. акт. фрагментом Кленова ДНК полимеразы I E.coli с добавлением 10 мМ dNTP, a затем лигируют в предварительно лианеризованный эндонуклеазой рестрикции EcoRM вектора VTvaf17 и GDTT1.8NAS1, GDTT1.8NAS3, GDTT1.8NAS7. Лигированную ДНК используют для трансформации клеток штамма Е, coli SCS110-AF и Е. coli JM-110-NAS с помощью электропорации. Для приготовления электрокомпетентных клеток штамма Escherichia coli SCS 110-AF и Escherichia coli JM-110-NAS, одиночной колонией заражают 10 мл среды LB и культивируют ночь в орбитальном шейкере при 150 об/мин и 37°С. На следующий день пересеивают 1/20 в 100 мл среды LB и культивируют в орбитальном шейкере при 150 об/мин и 37°С до достижения OD600=0,5. По достижении необходимой оптической плотности, клетки охлаждают до 0°С и центрифугируют 10 мин при 4000 g. Далее, удаляют среду, клетки отмывают от остатков среды охлажденной во льду 100 мл бидистилированной водой дважды, затем промывают 20 мл 10% глицерина. После этого, клетки ресуспендируют в 1 мл 10% глицерина и используют для трансформации методом электропорации. Электропорацию проводят в кюветах 1 мм при 2 кВ, 200 Ом, 25 мкФ на приборе Gene Pulser Xcell (Bio-Rad, США). Время импульса составляет от 4,9 до 5,1 мсек, с использованием 1-10 нг вектора. После чего клетки культивируют в течение 2,5 часов в среде SOC в шейкере-инкубаторе при 30°С. Затем, клетки высеивают на чашки Петри с агаризованной селективной средой, содержащей дрожжевой экстракт, пептон, 6% сахарозы, а также 10 мкг/мл хлорамфеникола.
Плазмидную ДНК из устойчивых колоний выделяют с помощью набора для выделения плазмид QiagenSpinMiniprepKit (Qiagen, Germany), анализируют с помощью гидролиза рестриктазами BamHI и SaIl и отбирают генетическую (ие) конструкцию (ии) с помощью секвенирования ДНК генетических конструкций по методу Сэнгера.
Бактерии, содержащие полученные генетические конструкции на базе векторов pCMV6-XL5, pCMV6-Kan/Neo и pCDNA 3.1(+), выращивают на жидкой среде LB с ампициллином. Бактерии штамма Escherichia соli SCS110-AF/VTvaf17, Escherichia coli JM-110-NAS/GDTT1.8NAS1, Escherichia coli JM-110-NAS/ GDTT1.8NAS3, Escherichia coli JM-110-NAS/GDTT1.8NAS7, содержащие полученные генетические конструкции на базе соответственно векторов VTvaf17, GDTT1.8NAS1, GDTT1.8NAS3, GDTT1.8NAS7, выращивают на селективной среде, содержащей дрожжевой экстракт, пептон, 6% сахарозы, а также 10 мкг/мл хлорамфеникола, и культивируют ночь в орбитальном шейкере при 150 об/мин и 37°С, лизируют и выделяют плазмидную ДНК для дальнейшей трансфекции специальным набором для выделения плазмид QiagenMaxiKit.
Таким образом, получают экспрессионые генетические конструкции на базе векторов pCMV6-XL5, pCMV6-Kan/Neo и pCDNA 3.1(+), VTvaf17, GDTT1.8NAS1, GDTT1.8NAS3, GDTT1.8NAS7 которые содержат нуклеотидную последовательность длиной 1608 н.п., которая включает белок-кодирующую область гена Р4НА2 и не содержит нетранслируемые 5' и 3' области гена с первичной структурой, приведенной на фиг. 1 Seq ID No 1.
Пример 3.
Проведение модификации кДНК гена Р4НА2 в генетических конструкциях.
Для получения модифицированных кДНК гена Р4НА2 с целью повышения эффективности транскрипции и трансляции белка пролил 4- гидрокислаза альфа 2 и/или персонализированного подбора для конкретного пациента наиболее эффективного для него генетической конструкции, в клетках тканей и органов человека в участок немодифицированной последовательности кДНК гена Р4НА2, используя принцип оптимизации кодонов, вносят модификации путем замены минорных кодонов на синонимичные мажорные, или осуществляют аминокислотные замены, или осуществляют делеции таким образом, чтобы модификации не ухудшили процессы транскрипции и трансляции.
Все модификации осуществляют в несколько этапов методом ПЦР мутагенеза набором QuikChP4HA2e II XL kit или QuikChP4HA2e Multi Site-Directed Mutagenesis Kit (AgilentTechnologies, USA,) согласно рекомендациям производителя. http://www.agilent.com/cs/library/usermanuals/Public/200513.
К плазмидной ДНК, полученной по Примеру 2, в количестве 10-50 нг добавляют 50-100 нг праймера, или двухцепочечный фрагмент ДНК, содержащего необходимую замену, инсерцию или делецию, и проводят ПЦР в буфере QuikChP4HA2e Multi reaction buffer (AgilentTechnologies, USA) со смесью ферментов QuikChP4HA2e Multi mix (AgilentTechnologies, USA) при следующих условиях: 95°С, 1 мин; далее от 30 до 35 циклов: 95°С 1 минута, 55°С 1 минута, 65°С 2 минуты/1000 н.п. После проведения ПЦР к амплификационной смеси добавляют 1-2 мкл рестриктазы Dpn I и инкубируют 1-2 часа при 37°С. Полученной смесью для pCMV6-XL5, pCMV6-Kan/Neo и pCDNA 3.1 (+) трансформируют компетентные клетки E.coli Тор10 (http://molbiol.ru/protocol/03_04.html), которые высевают на чашки с L-агаром, содержащих 100 мкг/мл ампициллина. Трансформированные колонии анализируют на содержание мутаций секвенированием плазмидной ДНК по методу Сэнгера. Для VTvaf17 и GDTT1.8NAS1, GDTT1.8NAS3, GDTT1.8NAS7 полученной смесью трансформируют компетентные клетки соответственно Escherichia coli SCS110-AF (Cell and gene therapy Ltd.) и Escherichia coli JM-110-NAS/GDTT1.8NAS1, Escherichia coli JM-110-NAS/GDTT1.8NAS3, Escherichia coli JM-110-NAS/GDTT1.8NAS7 (Genetic Diagnostics and Therapy 21 LTD, UK), которые высевают на чашки Петри с агаризованной селективной средой, содержащей дрожжевой экстракт, пептон, 6% сахарозы, а также 10 мкг/мл хлорамфеникола. Трансформированные колонии анализируют на содержание мутаций секвенированием плазмидной ДНК по методу Сэнгера.
Для получения модифицированной кДНК гена Р4НА2 SEQ ID No 2 проводят последовательный ПЦР-мутагенез на плазмидах VTvaf17-P4HA2 Seq ID No 1, GDTT1.8NAS1-Р4НА2 Seq ID No 1, GDTT1.8NAS3-P4HA2 Seq ID No 1, GDTT1.8NAS7 Seq ID No 1, pCMV6-XL5-P4HA2 Seq ID No 1, pCMV6-Kan/Neo-P4HA2 Seq ID No 1 и pCDNA 3.1(+)- Р4НА2 Seq ID No1 с праймерами, комплементарными участкам нативной кДНК Р4НА2, приведенной в GenBank под номером NM_004199:
1. Р4НА2 1075-1105 F:
5' GGGATGGGCCGCTCCGCCTACAATGAAGGGG 3',
Комплементарный нуклеотидам 1075-1105, с заменой G→C в позиции 1089;
2. Р4НА2 1108-1133 F:
5' TATTATCATACCGTGTTGTGGATGGA 3',
Комплементарный нуклеотидам 1108-1133, с заменой G→C в позиции 1119;
3. Р4НА2 1228-1256 F:
5' GATCTGCACCGGGCCCTGGAGCTCACCC 3',
Комплементарный нуклеотидам 1228-1256, с заменой T→G в позиции 1239;
4. Р4НА2 1279-1308 F:
5' AGCCACGAACGGGCTGGAGGGAATCTGCGG 3',
Комплементарный нуклеотидам 1279-1308, с заменой A→G в позиции 1290;
Модифицированная таким образом нуклеотидная последовательность кДНК Р4НА2 SEQ ID No: 2 содержит 2 нуклеотидных замены G→C в позициях 1089, 1119; 1 нуклеотидную замену T→G в позиции 1239, 1 нуклеотидную замену A→G в позиции 1290; не приводящие к изменениям в аминокислотной последовательности белка, кодируемого последовательностью гена Р4НА2 и имеет первичную структуру, приведенную на фиг. 2:
Для получения модифицированной кДНК гена Р4НА2 SEQ ID No 3 проводят последовательный ПЦР-мутагенез на плазмидах VTvaf17 Seq ID No 1, GDTT1.8NAS1-Р4НА2 Seq ID No 1, GDTT1.8NAS3-P4HA2 Seq ID No 1, GDTT1.8NAS7-P4HA2 Seq ID No 1, pCMV6-XL5-P4HA2 Seq ID No 1, pCMV6-Kan/Neo-P4HA2 Seq ID No1 и pCDNA 3.1(+)- Р4НА2 Seq ID No 1 с праймерами, комплементарными участкам нативной кДНК Р4НА2, приведенной в GenBank под номером NM_004199:
1. Р4НА2 1075-1105 F:
5' GGGATGGGCCGCTCCGCCTACAATGAAGGGG 3',
Комплементарный нуклеотидам 1075-1105, с заменой G→C в позиции 1089;
2. Р4НА2 1108-1133 F:
5' TATTATCATACCGTGTTGTGGATGGA 3',
Комплементарный нуклеотидам 1108-1133, с заменой G→C в позиции 1119;
3. Р4НА2 1228-1256 F:
5' GATCTGCACCGGGCCCTGGAGCTCACCC 3',
Комплементарный нуклеотидам 1228-1256, с заменой T→G в позиции 1239;
4. Р4НА2 1279-1308 F:
5' AGCCACGAACGGGCTGGAGGGAATCTGCGG 3',
Комплементарный нуклеотидам 1279-1308, с заменой A→G в позиции 1290;
5. Р4НА2 1336-1365 F:
5' AGAGAAAAAACCTTAACAAATCAGACAGAA 3',
Комплементарный нуклеотидам 1336-1365, с заменой G→C в позиции 1347;
6. Р4НА2 1433-1464 F:
5' ACGAGAGCCTCTGTCGGGGGGAGGGTGTCAAA 3',
Комплементарный нуклеотидам 1433-1464, с заменой T→G в позиции 1449;
7. Р4НА2 1463-1489 F:
5' AACTGACACCCCGGAGACAGAAGAGGC 3',
Комплементарный нуклеотидам 1463-1489, с заменой T→G в позиции 1476;
8. Р4НА2 1648-1681 F:
5' GCACGGGCCACCGTTCGGGATCCCAAGACAGGAG 3',
Комплементарный нуклеотидам 1648-1681, с заменами A→G в позиции 1653 и T→G в позиции 1665;
Модифицированная таким образом нуклеотидная последовательность кДНК Р4НА2 SEQ ID No: 3 содержит 3 нуклеотидных замены G→C в позициях 1089, 1119, 1347; 4 нуклеотидных замены T→G в позициях 1239, 1449, 1476, 1665, 2 нуклеотидных замены A→G в позициях 1290, 1653; не приводящие к изменениям в аминокислотной последовательности белка, кодируемого последовательностью гена Р4НА2 и имеет первичную структуру, приведенную на фиг. 3:
Для получения модифицированной кДНК гена Р4НА2 SEQ ID No 4 проводят последовательный ПЦР-мутагенез на плазмидах VTvaf17-Р4НА2 Seq ID No 1, GDTT1.8NAS1-Р4НА2 Seq ID No 1, GDTT1.8NAS3-P4HA2 Seq ID No 1, GDTT1.8NAS7 Seq ID No 1, pCMV6-XL5-P4HA2 Seq ID No 1, pCMV6-Kan/Neo-P4HA2 Seq ID No1 и pCDNA 3.1(+)- Р4НА2 Seq ID No1 с праймерами, комплементарными участкам нативной кДНК Р4НА2 приведенной в GenBank под номером NM_004199:
1. Р4НА2 1075-1105 F:
5' GGGATGGGCCGCTCCGCCTACAATGAAGGGG 3',
Комплементарный нуклеотидам 1075-1105, с заменой G→C в позиции 1089;
2. Р4НА2 1108-1133 F:
5' TATTATCATACCGTGTTGTGGATGGA 3',
Комплементарный нуклеотидам 1108-1133, с заменой G→C в позиции 1119;
3. Р4НА2 1228-1256 F:
5' GATCTGCACCGGGCCCTGGAGCTCACCC 3',
Комплементарный нуклеотидам 1228-1256, с заменой T→G в позиции 1239;
4. Р4НА2 1279-1308 F:
5' AGCCACGAACGGGCTGGAGGGAATCTGCGG 3',
Комплементарный нуклеотидам 1279-1308, с заменой A→G в позиции 1290;
5. Р4НА2 1336-1365 F:
5' AGAGAAAAAACCTTAACAAATCAGACAGAA 3',
Комплементарный нуклеотидам 1336-1365, с заменой G→C в позиции 1347;
6. Р4НА2 1433-1464 F:
5' ACGAGAGCCTCTGTCGGGGGGAGGGTGTCAAA 3',
Комплементарный нуклеотидам 1433-1464, с заменой T→G в позиции 1449;
7. Р4НА2 1463-1489 F:
5' AACTGACACCCCGGAGACAGAAGAGGC 3',
Комплементарный нуклеотидам 1463-1489, с заменой T→G в позиции 1476;
8. Р4НА2 1648-1681 F:
5' GCACGGGCCACCGTTCGGGATCCCAAGACAGGAG 3',
Комплементарный нуклеотидам 1648-1681, с заменами A→G в позиции 1653 и T→G в позиции 1665;
9. Р4НА2 1750-1777 F:
5' GCCCGGGTAAATCGGCGGATGCAGCATA 3',
Комплементарный нуклеотидам 1750-1777, с заменами A→G в позиции 1755 и T→G в позиции 1764;
10. Р4НА2 1873-1898 F:
5' GATGAGCGGGATACTTTCAAGCATTT 3',
Комплементарный нуклеотидам 1873-1898, с заменой A→G в позиции 1881;
11. Р4НА2 1899-1928 F:
5' AGGGACCGGGAATCGGGTGGCTACTTTCTT 3',
Комплементарный нуклеотидам 1899-1928, с заменами G→C в позиции 1905 и T→G в позиции 1914
12. Р4НА2 2049-2078 F:
5' TGACTACCGGACAAGACATGCTGCCTGCCC 3',
Комплементарный нуклеотидам 2049-2078, с заменой A→G в позиции 2058;
13. Р4НА2 2119-2145 F:
5' CATGAACGGGGACAGGAGTTCTTGAGA 3',
Комплементарный нуклеотидам 2119-2145, с заменой A→G в позиции 2127;
Модифицированная таким образом нуклеотидная последовательность кДНК Р4НА2 SEQ ID No: 4 содержит 4 нуклеотидных замены G→C в позициях 1089, 1119, 1347, 1905; 6 нуклеотидных замен T→G в позициях 1239, 1449, 1476, 1665, 1764, 1914; 6 нуклеотидных замен A→G в позициях 1290, 1653, 1755, 1881, 2058, 2127; не приводящие к изменениям в аминокислотной последовательности белка, кодируемого последовательностью гена Р4НА2 и имеет первичную структуру, приведенную на фиг. 4:
Для получения модифицированной кДНК гена Р4НА2 SEQ ID No 5 проводят ПЦР-мутагенез на плазмидах VTvaf17-P4HA2 Seq ID No 1, GDTT1.8NAS1-P4HA2 Seq ID No 1, GDTT1.8NAS3-P4HA2 Seq ID No 1, GDTT1.8NAS7-P4HA2 Seq ID No 1, pCMV6-XL5-P4HA2 Seq ID No 1, pCMV6-Kan/Neo-P4HA2 Seq ID No 1 и pCDNA 3.1(+)- Р4НА2 Seq ID No 1 с синтезированным двухцепочечным фрагментом ДНК длиной 245 н.п.

Модифицированная таким образом нуклеотидная последовательность кДНК Р4НА2 SEQ ID No: 5 длиной 1602 н.п. имеет первичную структуру, приведенную на фиг. 5.
Для получения модифицированной кДНК гена Р4НА2 SEQ ID No 6 проводят последовательный ПЦР-мутагенез на плазмидах VTvaf17-P4HA2 Seq ID No 1, GDTT1.8NAS1-Р4НА2 Seq ID No 1, GDTT1.8NAS3-P4HA2 Seq ID No 1, GDTT1.8NAS7-P4HA2 Seq ID No 1, pCMV6-XL5-P4HA2 Seq ID No 1, pCMV6-Kan/Neo-P4HA2 Seq ID No 1 и pCDNA 3.1(+)- Р4НА2 Seq ID No 1 сначала с двухцепочечным фрагментом ДНК длиной 245 н.п.

а затем с праймерами, комплементарными участкам нативной кДНК Р4НА2, приведенной в GenBank под номером NM_004199:
1. Р4НА2 1075-1105 F:
5' GGGATGGGCCGCTCCGCCTACAATGAAGGGG 3',
Комплементарный нуклеотидам 1075-1105, с заменой G→C в позиции 1089;
2. Р4НА2 1108-1133 F:
5' TATTATCATACCGTGTTGTGGATGGA 3',
Комплементарный нуклеотидам 1108-1133, с заменой G→C в позиции 1119;
3. Р4НА2 1228-1256 F:
5' GATCTGCACCGGGCCCTGGAGCTCACCC 3',
Комплементарный нуклеотидам 1228-1256, с заменой T→G в позиции 1239;
4. Р4НА2 1279-1308 F:
5' AGCCACGAACGGGCTGGAGGGAATCTGCGG 3',
Комплементарный нуклеотидам 1279-1308, с заменой A→G в позиции 1290;
5. Р4НА2 1336-1365 F:
5' AGAGAAAAAACCTTAACAAATCAGACAGAA 3',
Комплементарный нуклеотидам 1336-1365, с заменой G→C в позиции 1347;
6. Р4НА2 1433-1464 F:
5' ACGAGAGCCTCTGTCGGGGGGAGGGTGTCAAA 3',
Комплементарный нуклеотидам 1433-1464, с заменой T→G в позиции 1449;
Модифицированная таким образом нуклеотидная последовательность кДНК Р4НА2 SEQ ID No: 6 длиной 1602 н.п. содержит 3 нуклеотидных замены G→C в позициях 1089, 1119, 1347; 2 нуклеотидных замены T→G в позициях 1239, 1449, 1 нуклеотидную замену A→G в позиции 1290; делецию в 6 нуклеотидных пар, а также замену 8 аминокислотных остатков и имеет первичную структуру, приведенную на фиг. 6:
Для получения модифицированной кДНК гена Р4НА2 SEQ ID No 7 проводят последовательный ПЦР-мутагенез на плазмидах VTvaf17-P4HA2 Seq ID No 1, GDTT1.8NAS1-Р4НА2 Seq ID No 1, GDTT1.8NAS3-P4HA2 Seq ID No 1, GDTT1.8NAS7-P4HA2 Seq ID No 1, pCMV6-XL5-P4HA2 Seq ID No 1, pCMV6-Kan/Neo-P4HA2 Seq ID No 1 и pCDNA 3.1(+)- P4HA2 Seq ID No 1 сначала с двухцепочечным фрагментом ДНК длиной 245 н.п.

а затем с праймерами, комплементарными участкам нативной кДНК P4HA2, приведенной в GenBank под номером NM_004199:
1. P4HA2 1075-1105 F:
5' GGGATGGGCCGCTCCGCCTACAATGAAGGGG 3',
Комплементарный нуклеотидам 1075-1105, с заменой G→C в позиции 1089;
2. P4HA2 1108-1133 F:
5' TATTATCATACCGTGTTGTGGATGGA 3',
Комплементарный нуклеотидам 1108-1133, с заменой G→C в позиции 1119;
3. P4HA2 1228-1256 F:
5' GATCTGCACCGGGCCCTGGAGCTCACCC 3',
Комплементарный нуклеотидам 1228-1256, с заменой T→G в позиции 1239;
4. P4HA2 1279-1308 F:
5' AGCCACGAACGGGCTGGAGGGAATCTGCGG 3',
Комплементарный нуклеотидам 1279-1308, с заменой A→G в позиции 1290;
5. Р4НА2 1336-1365 F:
5' AGAGAAAAAACCTTAACAAATCAGACAGAA 3',
Комплементарный нуклеотидам 1336-1365, с заменой G→C в позиции 1347;
6. Р4НА2 1433-1464 F:
5' ACGAGAGCCTCTGTCGGGGGGAGGGTGTCAAA 3',
Комплементарный нуклеотидам 1433-1464, с заменой T→G в позиции 1449;
7. Р4НА2 1463-1489 F:
5' AACTGACACCCCGGAGACAGAAGAGGC 3',
Комплементарный нуклеотидам 1463-1489, с заменой T→G в позиции 1476;
8. Р4НА2 1648-1681 F:
5' GCACGGGCCACCGTTCGGGATCCCAAGACAGGAG 3',
Комплементарный нуклеотидам 1648-1681, с заменами A→G в позиции 1653 и T→G в позиции 1665;
9. Р4НА2 1750-1777 F:
5' GCCCGGGTAAATCGGCGGATGCAGCATA 3',
Комплементарный нуклеотидам 1750-1777, с заменами A→G в позиции 1755 и T→G в позиции 1764;
10. Р4НА2 2049-2078 F:
5' TGACTACCGGACAAGACATGCTGCCTGCCC 3',
Комплементарный нуклеотидам 2049-2078, с заменой A→G в позиции 2058;
11. Р4НА2 2119-2145 F:
5' CATGAACGGGGACAGGAGTTCTTGAGA 3',
Комплементарный нуклеотидам 2119-2145, с заменой A→G в позиции 2127;
Модифицированная таким образом нуклеотидная последовательность кДНК Р4НА2 SEQ ID No: 7 длиной 1602 н.п. содержит 3 нуклеотидных замены G→C в позициях 1089, 1119, 1347; 5 нуклеотидных замен T→G в позициях 1239, 1449, 1476, 1665, 1764; 5 нуклеотидных замен A→G в позициях 1290, 1653, 1755, 2058, 2127; делецию в 6 нуклеотидных пар, а также замену 8 аминокислотных остатков и имеет первичную структуру, приведенную на фиг. 7:
Таким образом, получают генетические конструкции (экспрессионные векторы) на базе плазмид VTvaf17, GDTT1.8NAS1, GDTT1.8NAS3, GDTT1.8NAS7, pCMV6-XL5, pCMV6-Kan/Neo и pCDNA 3.1(+), в которых клонированы участок немодифицированной кДНК гена Р4НА2 (SEQ ID No: 1) и модифицированные нуклеотидные последовательности кДНК гена Р4НА2 (SEQ ID No: 2-7), кодирующие белок пролил 4- гидрокислаза альфа 2.
Бактерии Escherichia coli, содержащие полученные генетические конструкции на базе векторов pCMV6-XL5, pCMV6-Kan/Neo и pCDNA 3.1(+), выращивают на жидкой среде LB с ампициллином, лизируют и выделяют плазмидную ДНК для трансфекции специальным набором для выделения плазмид QiagenMaxiKit (Qiagen, Germany) для получения соответствующих генетических конструкций.
Бактерии штамма Escherichia coli SCS110-AF/VTvaf17, Escherichia coli JM-110-NAS/GDTT1.8NAS1, Escherichia coli JM-110-NAS/GDTT1.8NAS3, Escherichia coli JM-110-NAS/GDTT1.8NAS7, содержащие полученные генетические конструкции на базе векторов VTvaf17, GDTT1.8NAS1, GDTT1.8NAS3, GDTT1.8NAS7 выращивают на селективной среде, содержащей дрожжевой экстракт, пептон, 6% сахарозы, а также 10 мкг/мл хлорамфеникола, затем лизируют и выделяют плазмидную ДНК для трансфекции специальным набором для выделения плазмид Endo Free Plasmid Maxi Kit (Qiagen, Germany).
Полученные генетические конструкции применяют для трансфекции эукариотических клеток органов и тканей и/или введения в органы и ткани человека. В качестве контрольных плазмид используют исходный вектор VTvaf17, GDTT1.8NAS1, GDTT1.8NAS3, GDTT1.8NAS7, pCMV6 - XL5, pCMV6-Kan/Neo, или pCDNA 3.1 (+) без вставки кДНК гена Р4НА2.
Пример 4.
Наращивание генетической конструкции в бактериальной культуре.
Для получения препаративных количеств векторной плазмиды, содержащей один из вариантов кДНК гена Р4НА2, полученными конструкциями по примеру 2 и 3 трансформируют бактериальные клетки E.coli - для конструкций на базе pCMV6 -XL5, pCMV6-Kan/Neo или pCDNA 3.1 (+) (Маниатис Т., и др. Молекулярное клонирование. М., Мир, 1984). Для конструкции на базе VTvaf17 трансформируют клетки линии Escherichia coli SCS110-AF. Для конструкций на базе GDTT1.8NAS1, GDTT1.8NAS3, GDTT1.8NAS7 трансформируют клетки линии Escherichia coli JM-110-NAS. Полученные клоны бактерий, содержащие конструкцию, выращивают в присутствии необходимого для каждого вида конструкций фактора селекции -для конструкций на базе pCMV6 - XL5, pCMV6-Kan/Neo или pCDNA 3.1 (+) - ампициллина, для конструкций на базе VTvaf17 и GDTT1.8NAS1, GDTT1.8NAS3, GDTT1.8NAS7 - сахарозы.
Лигазную смесь по примеру 2 и 3 используют для трансформации компетентных клеток E.coli штамма XLblue, SCS110-AF или JM-110-NAS. Предварительный отбор клонов, содержащих вставку кДНК гена Р4НА2 в правильной ориентации и единичной копии, осуществляют путем гидролиза плазмидной ДНК рестриктазами EcoRI и XbaI или BamHI и SalI и анализа продуктов рестрикции путем электрофореза в 1.2% агарозном геле. Отобранные клоны используют для препаративного наращивания плазмидной ДНК с целью дальнейшей трансфекции культуры фибробластов (кератоцитов, эпителиальных клеток роговицы, эндотелиальных клеток из пупочной вены, хондроцитов человека и др.).
Для получения препаративных количеств генетических экспрессионных конструкций по Примеру 2 и 3 в векторах pCMV6 -XL5, pCMV6-Kan/Neo или pCDNA 3.1 (+) выращивали 20 мл ночной культуры E.coli в LB-среде с 150 мкг/мл ампициллина. Этой культурой инокулировали 500 мл LB-среды с 100 мкг/мл ампициллина, рост осуществляли в шейкере-инкубаторе в течение 16 часов при 37°С и 200 об/мин.
Для получения препаративных количеств генетических экспрессионных конструкций по Примеру 2 и 3 векторы VTvaf17 и GDTT1.8NAS1, GDTT1.8NAS3, GDTT1.8NAS7 выращивали на селективной среде, содержащей дрожжевой экстракт, пептон, 6% сахарозы, а также 10 мкг/мл хлорамфеникола Выделение плазмидной ДНК проводили с помощью набора EndoFreePlasmidMaxiKit (Qiagen, Germany). Выход составил 5 мг ДНК.
Пример 5.
Культивирование эукариотических клеток, например, фибробластов для последующих трансфекции генетической конструкцией и анализа экспрессии гена Р4НА2.
Для последующей трансфекции генетической конструкцией, содержащей один из вариантов кДНК гена Р4НА2 по примеру 3, выращивают первичные культуры фибробластов человека из биоптатов кожи пациентов.
Используя устройство для взятия биопсии кожи Epitheasy 3.5 (MedaxSRL) берут образец биоптата кожи из зоны, защищенной от действия ультрафиолета, например, за ушной раковиной или с боковой внутренней поверхности в зоне локтевого сустава, объемом не менее 10 куб. мм, массой - не менее 11 мг. Кожу пациента предварительно промывают стерильным физиологическим раствором и анестезируют раствором лидокаина. Выращивание первичной культуры клеток осуществляют на чашках Петри при 37°С в атмосфере, содержащей 5% CO2 в среде DMEM с 10% фетальной телячьей сывороткой и ампициллином 100 Ед/мл. Трипсинизацию и пересев производят каждые 5 дней, для трипсинизации клетки промывают раствором PBS и инкубируют 30 минут при 37°С в растворе, содержащем 0.05% трипсина и 1 мМ ЭДТА. Для нейтрализации трипсина к клеткам добавляют 5 мл культуральной среды и центрифугируют суспензию при 600 об/мин 5 минут. Рост культуры фибробластов после 4-5 пассажей осуществляют в культуральных флаконах емкостью 75 мл в среде, содержащей 10 г/л DMEM, 3.7 г/л Na2CO3, 2.4 г/л HEPES, 10% фетальной сыворотки и 100 Ед/мл ампициллина. Смену культуральной среды осуществляют каждые 2 дня. Общая продолжительность роста культуры не превышала 25-30 дней.
Из культуры клеток отбирают аликвоту, содержащую 106 клеток.
Часть культуры фибробластов, предназначенную для выделения РНК с последующим анализом транскрипции гена Р4НА2, центрифугируют при 1000 об/мин 20 минут и дважды отмывают в буферном растворе PBS центрифугированием при 600 об/мин в течение 5 минут. Затем клетки ресуспендируют в 1.5 мл стабилизирующего реагента RNAIater и хранят при 4°С в течение 10 дней для последующего выделения РНК.
Выделение РНК проводят с помощью набора RNeasyMiniKit. (Qiagen, Germany). Выделенную РНК анализируют спектрофотометрически, измеряя соотношение оптической плотности при 260 и 280 нм, а также с помощью капиллярного электрофореза на приборе QIAxcel (Qiagen, Germany), используя картридж RNA Qiality Control. Для дальнейшей работы используют только те образцы, для которых общее количество выделенной РНК было не менее 50 мкг РНК, соотношение D260: D280 - не менее 1,8, а соотношение полос 28S: 18S на капиллярном электрофорезе - не ниже 1:1.
Синтез суммарной кДНК первой цепи проводят, используя обратную транскриптазу RevertAid (Thermo Scientific, USA), согласно рекомендациям изготовителя. 1-2 мкг суммарной РНК используют в качестве матрицы для синтеза первой цепи кДНК. В реакционную смесь для проведения обратной транскрипции вносят 100-200 ед. обратной транскриптазы и 10 пМ случайного 9-нуклеотидного праймера.
Пример 6.
Анализ эндогенной экспрессии гена Р4НА2 в культуре первичных фибробластов
Анализ экспрессии гена Р4НА2 из генома фибробластов проводят с целью последующего корректного определения генно-терапевтического эффекта после трансфекции этих клеток генетическими конструкциями с кДНК гена Р4НА2 VTvaf17- Р4НА2 SEQ ID No: 1.
Для анализа используют клеточные линии фибробластов кожи, фибробласты с низкой экспрессией гена Р4НА2 и фибробласты с нормальной экспрессией гена Р4НА2. Культивирование фибробластов проводят по примеру 5. Для трансфекции клеток используют раствор дендримера SuperFect Transfection Reagent 6-го поколения (Qiagen, Германия).
Приготовление комплексов ДНК-дендример проводят по методике производителя (QIAGEN, SuperFect Transfection Reagent Handbook, 2002) с некоторыми изменениями: к 15 мкл раствора дендримера (с концентрацией 3 мкг/мкл) добавляют 5 мкл (с концентрацией 0,5 мкг/мкл) раствора каждой плазмидной ДНК и тщательно перемешивают. Затем инкубируют каждую смесь ДНК с дендримером в течение 5-10 минут при температуре 15-25°С.
Далее из лунок планшетов с клеточный культурами осторожно отбирают среду (не нарушая клеточного слоя) и промывали клетки 3 мл буфера PBS. К растворам, каждый из которых содержал комплексы ДНК-дендример, добавляют 600 мкл среды DMEM с фетальной сывороткой и переносят эти смеси в лунки планшетов с клетками. Клетки инкубируют с комплексами 2-3 часа при 37°С и в атмосфере, содержащей 5% CO2. Затем осторожно удаляют среду и промывают клеточный слой 3 мл буфера PBS. Затем добавляют культуральную среду и инкубируют 24-48 часов при 37°С и в атмосфере, содержащей 5% CO2, после чего оценивают уровень кДНК гена Р4НА2 и контрольной кДНК гена В2М с помощью амплификации в режиме реального времени (SYBR GreenRealTimePCR).
Выделение РНК из фибробластов и синтез первой цепи кДНК проводят по примеру 5.
Выделенную РНК для каждой клеточной линии фибробластов, трансфицированных генетической конструкцией VTvaf17- Р4НА2 SEQ ID No: 1, анализируют методом ПЦР в режиме реального времени (SYBR GreenRealTimePCR).
Для амплификации кДНК (1608 н.п. ), специфичной для конструкции VTvaf17- Р4НА2 SEQ ID No: 1, используют
прямой праймер Р4НА2 -
FA1: TGTGGCCCGAGTAAATCGTC
и
обратный праймер
Р4НА2 -RA1: AAGTAGCCACACGATTCCCC.
Длина продукта амплификации - 179 п.н. В качестве референтного гена используют ген бета-2-микроглобулина В2М, уровень экспрессии которого в фибробластах сопоставим с таковым в норме для гена Р4НА2.
ПЦР-амплификацию проводят с помощью набора реагентов SYBR GreenQuantitect RT-PCR Kit (Qiagen, USA) или другого набора для ПЦР в режиме реального времени в 20 мкл амплификационной смеси, содержащей: 10 мкл QuantiTect SYBR Green RT-PCR MasterMix, 2,5 mM хлорида магния, по 0,5 мкМ каждого праймера, 5 мкл суммарной РНК. Реакцию осуществляют на амплификаторе CFX96 (Bio-Rad, USA) при следующих условиях: 1 цикл обратной транскрипции при 50°С - 30 минут, денатурация 98°С - 15 мин., затем 40 циклов, включающих денатурацию 94°С - 15 сек., отжиг праймеров 59°С - 30 сек. и элонгацию 72°С - 30 сек.
В качестве положительного контроля используют концентрацию ампликонов, получаемых при ПЦР на матрицах, представляющих собой плазмиды в известных концентрациях, содержащие последовательности кДНК генов Р4НА2 и В2М. В качестве отрицательного контроля используют деионизированную воду (кривые накопления ПЦР продуктов отрицательного и положительных контролей для упрощения визуализации на фигуре 8 не показаны). Количество ПЦР продуктов - кДНК генов Р4НА2 и В2М, полученных в результате амплификации, оценивают в режиме реального времени с помощью программного обеспечения амплификатора Bio-RadCFXManager 2.1.
По итогам анализа экспрессии гена Р4НА2 в разных культурах фибробластов (с низкой экспрессией гена Р4НА2 и с нормальной экспрессией гена Р4НА2) были отобраны клоны от пациентов одного возраста и одного типа старения, в которых количество ПЦР-продукта, соответствующего кДНК гена Р4НА2, была в 10-20 раз ниже таковой кДНК гена В2М. Кривые накопления специфических ПЦР-продуктов приведены на фигуре 8.
Пример 7.
Трансфекция клеточной культуры фибробластов со сниженной экспрессией гена Р4НА2 генетическими конструкциями, выбранными из группы, с целью подтверждения увеличения экспрессии гена Р4НА2 в культуре данных клеток.
Для оценки эффективности генетических конструкций, содержащих немодифицированную и модифицированные кДНК гена Р4НА2 по примеру 2 и 3, каждой из них трансфицируют первичную культуру фибробластов человека со сниженной экспрессией гена Р4НА2, полученную по примеру 5.
Данную культуру фибробластов выращивают в культуральных флаконах (25 см2) до момента, когда клетки покрывают 50-70% поверхности флакона. Общее количество клеток не менее 1×107. Выращенные клетки делят на 8 частей. Каждую часть полученных клеток трансфицируют одной генетической конструкцией, выбранной из группы: VTvaf17- Р4НА2 SEQ ID No: 1 или VTvaf17- Р4НА2 SEQ ID No: 2 или VTvaf17- Р4НА2 SEQ ID No: 3 или VTvaf17- Р4НА2 SEQ ID No: 4 или VTvaf17- Р4НА2 SEQ ID No: 5 или VTvaf17- Р4НА2 SEQ ID No: 6 или VTvaf17- Р4НА2 SEQ ID No: 7 в присутствии транспортной молекулы (например, SuperFect Transfection Reagent 6-го поколения (Qiagen, Германия) и вектором VTvaf17 в качестве контроля.
6-луночный планшет с DMEM-средой с 10% фетальной телячьей сывороткой (1.6 мл в лунке) засевают фибробластами из расчета 1×106 клеток на лунку. Инкубируют клетки в течение 18 часов при 37°С и в атмосфере, содержащей 5% CO2.
Приготовление комплексов ДНК-дендример и трансфекцию клеточной культуры проводят по по примеру 6.
Выделение РНК и синтез первой цепи кДНК проводят по примеру 5. Оценивают уровень кДНК гена Р4НА2 и контрольной кДНК гена В2М с помощью амплификации в режиме реального времени (SYBR GreenRealTimePCR).
Для амплификации кДНК (1608 н.п.), специфичной для конструкции VTvaf17- Р4НА2 SEQ ID No: 2, используют
прямой праймер
Р4НА2 FA1: CTAAACTTGCACGAGCCACC
и обратный праймер
Р4НА2 -RA1: AGAAGTCGAAGTGCGGTTCAT
Длина продукта амплификации - 225 п.н., условия амплификации, как для VTvaf17- Р4НА2 SEQ ID No: 1.
Для амплификации кДНК (1608 н.п. ), специфичной для конструкции VTvaf17- Р4НА2 SEQ ID No: 3, используют
прямой праймер
Р4НА2 FA1: TGTGGCCCGAGTAAATCGTC
и обратный праймер
Р4НА2 -RA1: AGAAGTCGAAGTGCGGTTCA
Длина продукта амплификации - 119 п.н., условия амплификации, как для VTvaf17- Р4НА2 SEQ ID No: 1.
Для амплификации кДНК (1608 н.п.), специфичной для конструкции VTvaf17- Р4НА2 SEQ ID No: 4, используют
прямой праймер
Р4НА2 FA1: CGGCGGATGCAGCATATCAC
и обратный праймер
Р4НА2 -RA1: GTCGAAGTGCGGTTCATACTGT
Длина продукта амплификации - 99 п.н., условия амплификации, как для VTvaf17- Р4НА2 SEQ ID No: 1.
Для амплификации кДНК (1602 н.п.), специфичной для конструкции VTvaf17- Р4НА2 SEQ ID No: 5, используют
прямой праймер
Р4НА2 FA1: CCTGTTGTGGCCCGAGTAAAT
и обратный праймер
Р4НА2 -RA1: GCTGTCAAAAGGTCGCCTAGA
Длина продукта амплификации - 144 нуклеотид, условия амплификации, как для VTvaf17- Р4НА2 SEQ ID No: 1.
Для амплификации кДНК (1602 н.п.), специфичной для конструкции VTvaf17- Р4НА2 SEQ ID No: 6, используют
прямой праймер
Р4НА2 FA1 GCCCGAGTAAATCGTCGGA
и обратный праймер
Р4НА2 -RA1 GCTGTCAAAAGGTCGCCTAGA
Длина продукта амплификации - 135 п.н., условия амплификации, как для VTvaf17- Р4НА2 SEQ ID No: 1.
Для амплификации кДНК (1602 н.п.), специфичной для конструкции VTvaf17- Р4НА2 SEQ ID No: 7, используют
прямой праймер
Р4НА2 FA1 ACTTCGACTTCTCTAGGCGAC
и обратный праймер
Р4НА2 -RA1 TTCTTAGGCCAAATTGCAGCC
Длина продукта амплификации - 145 нуклеотид, условия амплификации, как для VTvaf17- Р4НА2 SEQ ID No: 1.
В качестве референтного гена используют ген бета-2-микроглобулина В2М, уровень экспрессии которого в фибробластах сопоставим с таковым в норме для гена Р4НА2. В качестве положительного контроля используют концентрацию ампликонов, получаемых при ПЦР на матрицах, представляющих собой плазмиды в известных концентрациях, содержащие последовательности кДНК генов Р4НА2 и В2М. В качестве отрицательного контроля используют деионизированную воду (кривые накопления ПЦР продуктов отрицательного и положительных контролей для упрощения визуализации на фигуре 8 не показаны). Количество ПЦР продуктов - кДНК генов Р4НА2 и В2М, полученных в результате амплификации, оценивают в режиме реального времени с помощью программного обеспечения амплификатора Bio-RadCFXManager 2.1. Результаты анализа приведены на фигурах 9-15.
Из графиков следует, что в случае трансфекции вектором без вставки кДНК гена Р4НА2 уровень кДНК гена Р4НА2 в фибробластах не изменился, а в случае трансфекции генетическими конструкциями, выбранными из группы - уровень кДНК фибробластов со сниженной экспрессией гена Р4НА2 значительно увеличился.
Показано, что в каждом случае в клетках трансфицированных генетическими конструкциями VTvaf17- Р4НА2 SEQ ID No: 1 или VTvaf17- P4HA2 SEQ ID No: 2 или VTvaf17- P4HA2 SEQ ID No: 3 или VTvaf17- P4HA2 SEQ ID No: 4 или VTvaf17- P4HA2 SEQ ID No: 5 или VTvaf17- P4HA2 SEQ ID No: 6 или VTvaf17- P4HA2 SEQ ID No: 7 наблюдается усиление экспрессии целевого гена Р4НА2.
Пример 8.
Трансфекция клеточной культуры фибробластов с нормальной экспрессией гена Р4НА2 генетической конструкцией с кДНК гена Р4НА2 с целью подтверждения увеличения количества белка пролил 4- гидрокислаза альфа 2 в культуре данных клеток.
Для оценки изменения количества пролил 4- гидрокислаза альфа 2, использовали культуру фибробластов кожи человека по Примеру 5, в качестве транспортной молекулы - дендримеры SuperFect Transfection Reagent 6-го поколения (Qiagen, Германия), в качестве контроля - водный раствор дендримеров без плазмидной ДНК (А), векторную плазмиду pCDNA 3.1(+), не содержащую кДНК гена Р4НА2 (В), в качестве трансфицирующих агентов - генетическую конструкцию на основе векторной плазмиды pCDNA 3.1(+), содержащую кДНК гена Р4НА2 - pCDNA 3.1 Р4НА2 SEQ ID No: 2 (С), Приготовление комплекса ДНК-дендример и трансфекцию фибробластов проводили согласно Примеру 7.
После трансфекции к 0,5 мл культуральной жидкости приливали 0,1 мл 1N HCl, тщательно перемешивали и инкубировали 10 минут при комнатной температуре. Затем нейтрализовывали смесь, добавляя 0,1 мл 1.2N NaOH/0.5M HEPES (рН 7-7,6) и тщательно перемешивали.
Отбирали супернатант и использовали его для количественного определения целевого белка. Количественное определение продукта кДНК гена Р4НА2 проводили методом твердофазного иммуноферментного анализа (ELISA), используя набор Human Р4НА2 ELISA Kit Prolyl 4-hydroxylase subunit alpha-2, ELISA Kit (#MBS2885550) (MyBioSource, США), согласно методике производителя https://www.mybiosource.com/prods/ELISA-Kit/Human/Prolyl-4-hydroxylase-subunit-alpha-2/P4HA2/datasheet.php?products id=2885550. Оптическая плотность измерялась при длине волны 450 нм при помощи автоматического биохимического и иммуноферментного анализатора ChemWell (Awareness TechNology Inc., США) и пропорциональна концентрации белка в пробе. Численное значение концентрации определяется с помощью калибровочной кривой, построенной по калибраторам с известной концентрацией белка гена Р4НА2, входящим в состав набора. Разница между значениями оптической плотности (ОП) калибраторов поставленных в трех повторностях не превышала 10%.
Статистическую обработку полученных результатов, осуществляли с помощью программного обеспечения для статистической обработки и визуализации данных R, версия 3.0.2 (https://www.r-project.org/). Таким образом, показано, что транфекция фибробластов полученной генетической конструкцией pCDNA 3.1 Р4НА2 SEQ ID No: 2, несущей кДНК гена Р4НА2, приводит к увеличению количества белка пролил 4- гидрокислаза альфа 2 в клетках фибробластов. Результаты показаны на фигуре 16.
Пример 9.
Введение в кожу человека клеточной культуры фибробластов, трансфицированной генетической конструкцией с кДНК гена Р4НА2 с целью подтверждения увеличения количества белка пролил 4- гидрокислаза альфа 2 в биоптате кожи человека.
С целью анализа изменения количества белка пролил 4- гидрокислаза альфа 2 в тканях человека, пациентам в кожу предплечья вводили три варианта культуры аутологичных фибробластов - нетрансфицированных (зона А), трансфицированных вектором без вставки, например, VTvaf17 (зона В) и трансфицированных генетической конструкцией VTvaf17-Р4НА2 SEQ ID No: 7 (зона С) в комбинации с транспортными молекулами - липосомами TRANSFAST ТМ Transfection Reagent (PROMEGA, USA).
Приготовление суспензии липосом проводили по методике производителя (Promega, TransFast Transfection Reagent, Instruction for Use of Product, E2431) с некоторыми изменениями: к 0,4 мг порошка липосом добавляли 0,4 мл стерильной воды степени очистки Nuclease-Free. Суспензию хорошо перемешивали взбалтыванием. Полученный полупродукт суспензии липосом выдерживали в течение 18 часов при температуре - 20°С, затем оттаивали при комнатной температуре, ресуспендировали встряхиванием или на настольном миксере типа «ВОРТЕКС».
Для приготовления комплекса ДНК-липосомы к 400 мкл раствора липосом (с концентрацией 1 мг/мл) добавляли 400 мкл раствора плазмидной ДНК (с концентрацией 150 мкг/мл) и тщательно перемешивали. Затем инкубировали смесь ДНК с липосомами в течение 5-10 минут при температуре 15-25°С и использовали далее в качестве генетической конструкции.
Для последующей трансфекции клеток генетической конструкцией выращивали первичные культуры фибробластов человека из биоптатов кожи пациентов по примеру 5.
Для трансфекции из лунок планшета с клеточной культурой осторожно отбирали среду (не нарушая клеточного слоя) и промывали клетки 3 мл буфера PBS. К раствору, содержащему комплексы ДНК-липосомы, добавляли 600 мкл среды DMEM с фетальной сывороткой и переносили эту смесь в лунки планшета с клетками. Клетки инкубировали с комплексами 2-3 часа при 37°С и в атмосфере, содержащей 5% CO2. Затем осторожно удаляли среду и промывали клеточный слой 3 мл буфера PBS. Затем добавляли культуральную среду и инкубировали 24-48 часов при 37°С в атмосфере, содержащей 5% CO2.
Часть клеточной культуры центрифугировали при 700 об/мин, дважды отмывали клетки физиологическим раствором, ресуспендировали в физиологическом растворе из расчета 1 млн клеток в 200 мкл и использовали для введения пациенту. Введение осуществляли тоннельным методом иглой 30G на глубину 3 мм. Очаги введения культур фибробластов располагались на расстоянии 3-5 см друг от друга.
Определение количества белка пролил 4- гидрокислаза альфа 2 проводили в лизатах биоптатов кожи пациента по методу, описанному в Примере 8.
Биопсийные образцы брали на 3 сутки после введения суспензии фибробластов. Взятие биопсии осуществляли из участков введения суспензии фибробластов, а также из интактной кожи, используя устройство для взятия биопсии кожи Epitheasy 3.5 (MedaxSRL). Кожу пациента предварительно промывали стерильным физиологическим раствором и анестезировали раствором лидокаина. Объем биопсийного образца - не менее 10 куб. мм, массой - не менее 11 мг. Образец помещали в буферный раствор, содержащий 50 мМ Трис-HCl рН 7.6, 100 мМ NaCl, 1 мМ ЭДТА и 1 мМ фенилметилсульфонилфторид, и гомогенизировали до получения однородной суспензии. Полученную суспензию центрифугировали в течение 10 минут при 14000 об/мин. Отбирали супернатант и использовали его для количественного определения целевого белка.
Как видно из фигуры 17, в коже пациента в области введения фибробластов, трансфицированных генетической конструкцией VTvaf17- Р4НА2 SEQ ID No: 7, произошло увеличение количества белка пролил 4- гидрокислаза альфа 2 что может свидетельствовать об усилении экспрессии гена Р4НА2. Тогда как при введении контрольных культур фибробластов (нетрансфицированных и трансфицированных вектором без вставки кДНК гена Р4НА2 VTvaf17) количество пролил 4- гидрокислаза альфа 2, в коже не изменялась.
Таким образом, показана эффективность генетической конструкции при введении в кожу человека клеточной культуры фибробластов, трансфицированных генетической конструкцией, несущей модифицированную кДНК гена Р4НА2, что приводит к повышению уровня экспрессии гена Р4НА2, а именно - к увеличению количества белка пролил 4- гидрокислаза альфа 2 в коже в зоне введения.
Пример 10.
Трансфекция культур фибробластов из биоптатов группы различных пациентов генетическими конструкциями, содержащими модифицированные кДНК гена Р4НА2 и участок нативной немодифицированной кДНК гена Р4НА2.
С целью подтверждения индивидуального характера увеличения экспрессии гена Р4НА2 до различного уровня при трансфекции клеточных культур фибробластов пациентов генетическими конструкциями с модифицированными кДНК гена Р4НА2 и участком нативной кДНК гена Р4НА2 анализировали количественный уровень белка в клеточных лизатах фибробластов полученных из биоптатов группы пациентов, трансфицированных разными генетическими конструкциями по примеру 2 и 3, содержащими модифицированные кДНК гена Р4НА2 и участок нативной немодифицированной кДНК гена Р4НА2 в зависимости от наличия и типа модификаций в кДНК гена Р4НА2.
Последовательности участка нативной немодифицированной кДНК гена Р4НА2 приведена на фигуре 1, SEQ ID No: 1., последовательности модифицированных кДНК гена Р4НА2 приведены на фигурах 2-7 (SEQ ID No: 2, SEQ ID No: 3, SEQ ID No: 4, SEQ ID No: 5, SEQ ID No: 6, SEQ ID No: 7).
Вырастили культуры фибробластов из однотипных биоптатов кожи, отобранных из зоны внутренней боковой поверхности в зоне локтевого сустава или из биоптата полученного при косметических операциях 50 пациентов, выбранных случайным образом по примеру 5, отобрали аликвоты и оценили уровень белка пролил 4- гидрокислаза альфа 2, в клеточных лизатах. Клеточный осадок, соответствующий 1×107 клеток, промывали фосфатным буфером и ресуспендировали во льду в лизирующем буфере, содержащем 25 мМ ХЕПЕС рН 7.9, 100 MMNaCl, 1мМ ЭДТА, 1% Тритон Х-100, 10% глицерин и 1 мМ фенилметилсульфонилфторид. Лизат центрифугировали 5 минут при 14000 об/мин, концентрацию белка в супернатанте определяли методом Брэдфорда ([Bradford М.М. (1976) Anal. Biochem. 72: 248-254.]).
Затем каждую из 50-ти культур фибробластов разделили на 27 частей.
Одну часть, обозначенную (А), трансфицировали по примеру 7 генетической конструкцией pCMV6- XL5 SEQ ID No: 1, содержащей немодифицированную кДНК гена Р4НА2 (SEQ ID No: 1.) по примеру 2 в сочетании с транспортной молекулой -дендримером по примеру 7.
Вторую часть, обозначенную (В), трансфицировали по примеру 7 генетической конструкцией pCMV6- XL5 SEQ ID No: 2, содержащей вариант модифицированной кДНК гена Р4НА2 (SEQ ID No: 2.) по примеру 3 в сочетании с транспортной молекулой - дендримером по примеру 7.
3-ю часть, обозначенную (С), трансфицировали по примеру 7 генетической конструкцией pCMV6- XL5 SEQ ID No: 3, содержащей вариант модифицированной кДНК гена Р4НА2 (SEQ ID No: 3.) по примеру 3 в сочетании с транспортной молекулой - дендримером по примеру 7.
4-ю часть, обозначенную (D), трансфицировали по примеру 7 генетической конструкцией вектора pCMV6- XL5 SEQ ID No: 4, содержащей вариант модифицированной кДНК гена Р4НА2 (SEQ ID No: 4.) по примеру 3 в сочетании с транспортной молекулой - дендримером по примеру 7.
5-ю часть, обозначенную (Е), трансфицировали по примеру 7 генетической конструкцией pCMV6- XL5 SEQ ID No: 5, содержащей вариант модифицированной кДНК гена Р4НА2 (SEQ ID No: 5.) по примеру 3 в сочетании с транспортной молекулой - дендримером по примеру 7.
6-ю часть, обозначенную (F), трансфицировали по примеру 7 генетической конструкцией pCMV6- XL5 SEQ ID No: 6, содержащей вариант модифицированной кДНК гена Р4НА2 (SEQ ID No: 6) по примеру 3 в сочетании с транспортной молекулой - дендримером по примеру 7.
7-ю часть, обозначенную (G), трансфицировали по примеру 7 генетической конструкцией pCMV6- XL5 SEQ ID No: 7, содержащей вариант модифицированной кДНК гена Р4НА2 (SEQ ID No: 7) по примеру 3 в сочетании с транспортной молекулой - дендримером по примеру 7.
8-ю часть, обозначенную (Н), трансфицировали по примеру 7 векторной плазмидой pCMV6-XL5, не содержащей кДНК гена Р4НА2 в сочетании с транспортной молекулой - дендримером.
Количественное определение продукта кДНК гена Р4НА2 проводили по методу, описанному в Примере 8.
По итогам определения количественного уровня белка пролил 4- гидрокислаза альфа 2 выбрали показатели, касательно каждой части клеточной культуры от каждого пациента, продемонстрировавшие максимальные количественные уровни белка и объединили их в группы, исходя из следующего критерия:
В группе 1 максимальное количество белка пролил 4- гидрокислаза альфа 2, наблюдалось при трансфекции немодифицированной кДНК гена Р4НА2 SEQ ID No: 1. В эту группу вошло 11 культур из 50.
В группе 2 максимальное количество белка пролил 4- гидрокислаза альфа 2, наблюдалось при трансфекции модифицированной кДНК гена Р4НА2 SEQ ID No: 2. В эту группу вошло 5 культуры из 50.
В группе 3 максимальное количество белка пролил 4- гидрокислаза альфа 2, наблюдалось при трансфекции модифицированной кДНК гена Р4НА2 SEQ ID No: 3. В эту группу вошло 8 культуры из 50.
В группе 4 максимальное количество белка пролил 4- гидрокислаза альфа 2, наблюдалось при трансфекции модифицированной кДНК гена Р4НА2 SEQ ID No: 4. В эту группу вошли 5 культуры из 50.
В группе 5 максимальное количество белка пролил 4- гидрокислаза альфа 2, наблюдалось при трансфекции модифицированной кДНК гена Р4НА2 SEQ ID No: 5. В эту группу вошло 6 культуры из 50.
В группе 6 максимальное количество белка пролил 4- гидрокислаза альфа 2, наблюдалось при трансфекции модифицированной кДНК гена Р4НА2 SEQ ID No: 6. В эту группу вошли 8 культуры из 50.
В группе 7 максимальное количество белка пролил 4- гидрокислаза альфа 2, наблюдалось при трансфекции модифицированной кДНК гена Р4НА2 SEQ ID No: 7. В эту группу вошло 7 культуры из 50.
Ни в одной из клеточных культур, трансфицированных генетической конструкцией с векторной плазмидой, не содержащей кДНК гена Р4НА2 не был обнаружен максимальный количественный уровень белка пролил 4- гидрокислаза альфа 2, в отличие от клеточных культур, трансфицированных генетическими конструкциями с кДНК гена Р4НА2.
На фигуре 18 приведена таблица показателей количества белка пролил 4- гидрокислаза альфа 2, (усредненных в рамках группы, в случае, если в группу входит более одной клеточной культуры), применительно ко всем, участвующим в эксперименте генетическим конструкциям и для каждой группы клеточных культур после трансфекции этих клеточных культур генетическими конструкциями, содержащими модифицированные кДНК гена Р4НА2 и участок нативной кДНК гена Р4НА2.
Из данного примера следует, что достижение максимального количества белка пролил 4- гидрокислаза альфа 2 в культурах фибробластов кожи различных пациентов при их трансфекции генетическими конструкциями, связано с индивидуальными особенностями пациентов и зависит от наличия и типа модификаций в кДНК гена Р4НА2.
Каждая генетическая конструкция из группы является эффективной в некоторой значительной группе пациентов. Следовательно, для выбора наиболее эффективной генетической конструкции из группы для терапевтических целей необходимо предварительное персонализированное исследование биоптатов пациента, либо клеток, выращенных из этих биоптатов на предмет максимальной эффективности терапевтического воздействия, созданных генетических конструкций в рамках группы.
Пример 11.
Трансфекции клеточной культуры кератоцитов и эпителиальных клеток глаза генетической конструкцией без транспортной молекулы с целью подтверждения при этом увеличения экспрессии гена Р4НА2 в данных культурах клеток.
С целью выяснения эффективности генетической конструкции, содержащей кДНК Р4НА2, в различных типах клеток, трансфицировали кератоциты и эпителиальные клетки роговицы человека, в комбинации с транспортными молекулами -липосомами TRANSFAST ТМ Transfection Reagent (PROMEGA, USA) по Примеру 8.
Биопсийный материал из области лимба в виде круга диаметром 1-1.5 мм помещали в чашку Петри в сбалансированный солевой раствор Хэнкса, добавляли диспазу (20 мкл 25 ед/мл) и гентамицин и инкубировали 24 часа при +4. Затем отделяли эпителиальный слой от стромы и нарезали обе ткани кусочками 1-2 мм3.
Суспензию эпителиальных клеток получали путем инкубирования кусочков тканей в растворе Трипсин-ЭДТА в течение 15 минут при +37°С. Трипсинизацию останавливали добавлением соевого ингибитора трипсина в PBS. Клеточные суспензии отмывали центрифугированием при 700 об/мин и ресуспендировали в бессывороточной среде DMEM с гентамицином. Клетки растили в 25 см2 флаконах с 5 мл среды при 37°С и в атмосфере, содержащей 5% CO2. Через 24 часа среду с не прикрепившимися клетками удаляли и добавляли 5 мл свежей среды. Пересев производили каждые 7 дней в соотношении 1:2-1:3.
Суспензию кератоцитов получали путем инкубирования в среде RPMI-1640 с коллагеназой 300 ед/мл в течение 2 часов. Затем отмывали клетки и ресуспендировали в среде DMEM с 20% телячьей сывороткой и антибиотиком-антимикотиком. Клетки растили в 25 см2 флаконах с 1 мл среды при +37°С и в атмосфере, содержащей 5% CO2. Пересев производили каждые 7 дней в соотношении 1:4.
Поскольку эффективность трансфекции кератоцитов низкая, использовали генетическую конструкцию на основе плазмиды pCMV6-Kan/Neo Р4НА2 SEQ ID No: 2, содержащую дополнительный фактор селекции - ген neor, придающий устойчивость к антибиотику генетицину.
Генетическую конструкцию добавляли к клеткам и инкубировали клетки 1 час при 37°С. После инкубации добавляли бессывороточную среду DMEM к эпителиальным клеткам и среду DMEM, содержащую 10% фетальной телячьей сыворотки к кератоцитам и продолжали инкубировать 24 часа. После 24-часового инкубирования клетки высевали в новую среду DMEM, содержащую 400 μg/mL фактора селекции - антибиотика G418 (Geneticin). Выжившие клетки культивировали в течение 2 недель в 25 см3 флаконах с 5 мл среды с G418. Всего выращивали 24 образца культур эпителиальных клеток и 24 образца культур кератоцитов.
Каждые 24 часа из каждой культуры отбирали аликвоту по 500 мкл суспензии с целью выделения РНК и анализа уровня специфической кДНК. Выделение РНК проводили с помощью набора RNeasyMiniKit (Qiagen, Germany). Образцы РНК были сгруппированы в 3 группы по 8 образцов с близкой концентрацией и объединены. Анализ уровня специфической кДНК гена Р4НА1 проводили с помощью амплификации в режиме реального времени по методике и с праймерами по примеру 6, используя набор реагентов iTaqUniversalSYBRGreenSupermix (Bio-Rad, USA), амплификатор CFX96 (Bio-RadUSA) и программное обеспечение Bio-RadCFXManager 2.1. Максимальное увеличение экспрессии (транскрипции) гена Р4НА2 наблюдали на 3 сутки для эпителиальных клеток и на 5 сутки - для кератоцитов.
На фигуре 19 приведены кривые накопления специфического продукта амплификации, соответствующего кДНК гена Р4НА2, в кератоцитах и эпителиальных клетках роговицы.
Показано, что в кератоцитах и эпителиальных клетках роговицы человека, трансфицированных генетической конструкцией на основе pCMV6- Kan/Neo Р4НА2 SEQ ID No: 2, в комбинации с транспортными молекулами - липосомами TRANSFAST ТМ Transfection Reagent, наблюдается усиление экспрессии целевого гена Р4НА2.
Пример 12.
Трансфекция первичной культуры гладкомышечных клеток мочевого пузыря человека генетической конструкцией с транспортной системой в виде липосом Lipofectamine 3000 (ThermoFisher Scientific, США) с целью подтверждения при этом увеличения экспрессии гена Р4НА2 в данных культурах клеток.
С целью выяснения эффективности генетической конструкции, содержащей кДНК Р4НА2, в различных типах клеток, трансфицировали генетической конструкцией клетки первичной культуры гладкомышечных клеток мочевого пузыря человека HBdSMC (АТСС PCS-420-012) человека, с использованием липосомальных транспортных молекул Lipofectamine 3000 (ThermoFisher Scientific, США). В эксперименте использовали генетическую конструкцию на основе плазмиды GDTT1.8NAS1 Р4НА2 SEQ ID No: 3. Клетки первичной культуры гладкомышечных клеток мочевого пузыря человека HBdSMC (АТСС PCS-420-012) выращивают в среде, входящей в состав Vascular Smooth Muscle Cell Growth Kit (ATCC® PCS-100-042™) в атмосфере 5% CO2 при 37°C. Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевают в 24-луночный планшет из расчета 5*104 клеток/лунку.
Трансфекцию генетической конструкцией GDTT1.8NAS1-Р4НА2 SEQ ID No: 3 проводят следующим образом. В пробирке №1 к 25 мкл среды Opti-MEM (Gibco) добавляют 1 мкл раствора генетической конструкции (концентрация 500 нг/мкл) и 1 мкл реагента Р3000. Аккуратно перемешивают легким встряхиванием. В пробирке №2 к 25 мкл среды Opti-MEM (Gibco) добавляют 1 мкл раствора Lipofectamine 3000. Аккуратно перемешивают легким встряхиванием. Добавляют содержимое пробирки №1 к содержимому пробирки №2, инкубируют 5 мин при комнатной температуре. Полученный раствор по каплям добавляют к клеткам в объеме 40 мкл.
В качестве контроля используют клетки мочевого пузыря человека HBdSMC, трансфицированные ДНК-вектором GDTT1.8NAS1, не содержащим вставку целевого гена Р4НА2. Подготовку контрольного вектора GDTT1.8NAS1 для трансфекции проводят как описано выше.
Выделение суммарной РНК из трансфицированных клеток и ПЦР-амплификацию проводят по примеру 6. В качестве референтного гена используют ген бета-2-микроглобулина В2М. В качестве отрицательного контроля используют деионизированную воду. Количество ПЦР продуктов - кДНК генов Р4НА2 и В2М, полученных в результате амплификации, оценивают в режиме реального времени с помощью программного обеспечения амплификатора Bio-RadCFXManager 2.1.
С целью подтверждения увеличения экспрессии гена Р4НА2 в клетках мочевого пузыря человека HBdSMC человека после трансфекции данных клеток генетической конструкцией GDTT1.8NAS1-P4HA2 Р4НА2 SEQ ID No: 3 на фиг. 20 приведены графики накопления ПЦР-продуктов, соответствующих:
1 - кДНК гена Р4НА2 после трансфекции вектором GDTT1.8NAS1;
2 - кДНК гена Р4НА2 после трансфекции генетической конструкцией GDTT1.8NAS1 Р4НА2 SEQ ID No: 3, несущей участок гена Р4НА2;
3 - кДНК гена В2М после трансфекции вектором GDTT1.8NAS1
4 - кДНК гена В2М после трансфекции генетической конструкцией GDTT1.8NAS1 Р4НА2 SEQ ID No: 3, несущей участок гена Р4НА2;
Из фигуры следует, что в результате трансфекции клеток мочевого пузыря человека HBdSMC генетической конструкцией GDTT1.8NAS1 Р4НА2 SEQ ID No: 3, уровень специфической мРНК гена Р4НА2 человека вырос многократно.
Пример 13.
Трансфекции клеточной культуры хондробластов генетической конструкцией с кДНК гена Р4НА2 с целью подтверждения при этом увеличения экспрессии гена Р4НА2 в данных культуре клеток.
С целью выяснения эффективности генетической конструкции, содержащей кДНК гена Р4НА2, в различных типах клеток, а также с целью оценки влияния транспортной молекулы на успешность трансфекции этой генетической конструкцией -трансфицировали хондробласты человека, используя в качестве транспортной молекулы амфифильные блок-сополимеры.
Биоптаты из позвоночника эмбрионов человека 12-недельного возраста массой 50-150 мг измельчали на фрагменты до 10 мм3 и инкубировали в буферном растворе с коллагеназой, гиалуронидазой и трипсином в течение 48 часов при +37°С. Клетки отмывали бессывороточной средой DMEM, ресуспендировали в той же среде с гентамицином и выращивали при +37°С и в атмосфере, содержащей 5% CO2 во флаконах 75 см2 с 15 мл среды. Рост осуществлялся в бессыворотчной среде, так как в монослойной культуре хондроциты меняют форму и биохимические свойства. Пересев производили каждые 4 дня в соотношении 1:3, клетки снимали с поверхности флакона раствором трипсин-ЭДТА с 0.02% раствором Версена.
Для трансфекции клеток генетической конструкцией содержащей кДНК гена Р4НА2 использовали амфифильные блок-сополимеры. Применяли методику по (IntJPharm, 2012, 427, 80-87) или в (Macromol. Biosci.2011, 11, 652-661), с некоторыми изменениями. Блок-сополимер синтезировали из смеси линейного полиэтиленимина (ПЭИ) (Polyscience Inc., США) и бифункционального полиэтиленгликоля (ПЭГ) N-гидроксисукцинимидил-75-N-(3-малеимидопропионил)-амидо-4, 7, 10, 13, 16, 19, 22, 25, 28, 31, 34, 37, 40, 43, 46, 49, 52, 55, 58, 61, 64, 67, 70, 73 - тетракосаоксапента-гептаконтаноата (MAL-dPEG™-NHS ester, Quanta BioDesign, Ltd., США) в боратном буфере. Полиплекс готовили за 1 час до введения в клетки, смешивая раствор блок-сополимера с ДНК генетической конструкции pCMV6- Kan/Neo Р4НА2 SEQ ID No: 3 с модифицированной кДНК Р4НА2 SEQ ID No: 3. Смесь для трансфекции добавляли к суспензии клеток в 1 мл культуральной среды DMEM с 10% фетальной сывороткой и ампициллином.
Так как для трансфекции использовали конструкцию, содержащую фактор селекции (устойчивость к генетицину), то клетки после трансфекции выращивали в 25 см3 флаконах с 5 мл среды с генетицином в течение 12 дней, меняя среду каждые 4 дня. Каждые 24 часа из каждой культуры отбирали аликвоту по 500 мкл суспензии с целью выделения РНК и анализа уровня специфической кДНК. Выделение РНК производили с помощью набора RNeasyMiniKit (Qiagen, Germany). Образцы РНК были сгруппированы в 3 группы по 8 образцов с близкой концентрацией и объединены. Анализ уровня специфической кДНК Р4НА2 проводили с помощью амплификации в режиме реального времени по методике и с праймерами по примеру 6, используя набор реагентов iTaqUniversalSYBRGreenSupermix (Bio-Rad, USA), амплификатор CFX96 (Bio-Rad, USA) и программное обеспечение Bio-RadCFXManager 2.1. Максимальное увеличение транскрипции Р4НА2 наблюдали на 5 сутки. На фигуре 21 приведены кривые накопления специфического продукта амплификации, соответствующего кДНК гена Р4НА2.
Показано, что в хондробластах трансфицированных генетической конструкцией pCMV6- Kan/Neo Р4НА2 SEQ ID No: 3, используя в качестве транспортной молекулы амфифильные блок-сополимеры, наблюдается усиление экспрессии целевого гена Р4НА2.
Пример 14.
Трансфекция культуры эндотелиальных клеток из пупочной вены человека HUVEC (АТСС® CRL-1730™) генетической конструкцией с транспортной системой Lipofectamine 3000 (ThermoFisher Scientific, США) с целью подтверждения при этом увеличения экспрессии гена Р4НА2 в данных культурах клеток.
С целью выяснения эффективности генетической конструкции, содержащей кДНК Р4НА2, в различных типах клеток, трансфицировали генетической конструкцией первичную культуру эндотелиальных клеток из пупочной вены человека HUVEC (АТСС® CRL-1730™) человека, с использованием транспортных молекул Lipofectamine 3000 (ThermoFisher Scientific, США). В эксперименте использовали генетическую конструкцию на основе плазмиды GDTT1.8NAS3-P4HA2 Р4НА2 SEQ ID No: 4.
Линию первичной культуры эндотелиальных клеток из пупочной вены человека HUVEC (АТСС® CRL-1730™) выращивают в среде F-12K Medium (АТСС) с добавлением 0.05 мг/мл аскорбиновой кислоты, 0.01 мг/мл инсулина, 0.01 мг/мл трансферрина, 10 нг/мл селенита натрия, 0.03 мг/мл Endothelial Cell Growth Supplement (ECGS), 10% сыворотки эмбрионов коров в атмосфере 5% CO2 при 37°С. Для получения 90% конфлюэнтности, за 24 часа до постановки трансфекции клетки высевают в 24-луночный планшет из расчета 5*104 клеток/лунку.
Трансфекцию генетической конструкцией GDTT1.8NAS3-Р4НА2 Р4НА2 SEQ ID No: 4, экспрессирующей ген Р4НА2 человека проводят следующим образом. В пробирке №1 к 25 мкл среды Opti-MEM (Gibco) добавляют 1 мкл раствора генетической конструкции (концентрация 500 нг/мкл) и 1 мкл реагента Р3000. Аккуратно перемешивают легким встряхиванием. В пробирке №2 к 25 мкл среды Opti-MEM (Gibco) добавляют 1 мкл раствора Lipofectamine 3000. Аккуратно перемешивают легким встряхиванием. Добавляют содержимое пробирки №1 к содержимому пробирки №2, инкубируют 5 мин при комнатной температуре. Полученный раствор по каплям добавляют к клеткам в объеме 40 мкл.
В качестве контроля используют культуру эндотелиальных клеток из пупочной вены человека HUVEC, трансфицированную ДНК-вектором GDTT1.8NAS3, не содержащим вставку целевого гена Р4НА2. Подготовку контрольного вектора GDTT1.8NAS3 для трансфекции проводят как описано выше.
Выделение суммарной РНК из трансфицированных клеток и ПЦР-амплификацию проводят по примеру 6. В качестве референтного гена используют ген бета-2-микроглобулина В2М. В качестве отрицательного контроля используют деионизированную воду. Количество ПЦР продуктов - кДНК генов Р4НА2 и В2М, полученных в результате амплификации, оценивают в режиме реального времени с помощью программного обеспечения амплификатора Bio-RadCFXManager 2.1.
С целью подтверждения увеличения экспрессии гена Р4НА2 в культуре эндотелиальных клеток из пупочной вены человека HUVEC (АТСС® CRL-1730™) после трансфекции данных клеток генетической конструкцией GDTT1.8NAS3-P4HA2 Р4НА2 SEQ ID No: 4 на фиг. 22 приведены графики накопления ПЦР-продуктов.
Из фигуры следует, что в результате трансфекции культуры эндотелиальных клеток из пупочной вены человека HUVEC генетической конструкцией GDTT1.8NAS3-P4HA2 Р4НА2 SEQ ID No: 4, уровень специфической мРНК гена Р4НА2 человека вырос многократно.
Пример 15.
Введение в кожу человека генетической конструкции с кДНК гена Р4НА2 с целью подтверждения при этом увеличения количества белка пролил 4- гидрокислаза альфа 2 в коже человека.
С целью анализа увеличения количественного уровня белка пациенту вводили генетическую конструкцию pCMV6-XL5 Р4НА2 SEQ ID No: 5 с кДНК гена P4HA2 (зона В) и параллельно вводили плацебо, представляющее собой комбинацию векторной плазмиды не содержащей кДНК гена Р4НА2 с транспортной молекулой (зона А) - в кожу предплечья.
В качестве транспортной системы использовали полиэтиленимин Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, France - USA). Генетическую конструкцию pCMV6-XL5 P4HA2 SEQ ID No: 5, которая содержит модифицированную кДНК гена Р4НА2 (SEQ ID No: 5) и плазмиду pCMV6-XL5, используемую в качестве плацебо - которая не содержит кДНК гена Р4НА2, каждую из которых растворяли в стерильной воде степени очистки Nuclease-Free. Для получения генетической конструкции готовили комплексы ДНК- cGMP grade in-vivo-jetPEI в соответствие с рекомендациями производителя.
Расчет количества реагента осуществляли по формуле, предложенной производителем:
V(мкл)=(pDNA (мкг)×3×N/Р)/150
где N/P - соотношение между количеством азотистых остатков в jetPEI и количеством фосфатных групп нуклеиновых кислот. В данном эксперименте было использовано соотношение N/P=6, то есть на 20 мкг pDNA - 2,4 мкл jetPEI.
Полученные генетическую конструкцию и плацебо использовали для введения пациенту. Введение осуществляли тоннельным методом иглой 30G на глубину 3 мм. Объем вводимого раствора генетической конструкции и плацебо - около 0,3 мл для каждого. Очаги введения генетической конструкции и плацебо располагались на расстоянии 3-5 см друг от друга.
Количество белка пролил 4- гидрокислаза альфа 2 оценивали в лизатах биоптатов кожи пациента по методу, описанному в Примере 8.
Биопсийные образцы брали на 4 сутки после введения генетической конструкции. Взятие биопсии осуществляли из участков кожи в зоне введения генетической конструкции и плацебо, а также из интактной кожи, используя устройство для взятия биопсии Epitheasy 3.5 (Medax SRL) далее, как описано в примере 9.
Показано, что в коже пациента в области введения генетической конструкции pCMV6-XL5 Р4НА2 SEQ ID No: 5 с кДНК гена Р4НА2, произошло увеличение количества белка пролил 4- гидрокислаза альфа 2, тогда как при введении плацебо, количество белка пролил 4- гидрокислаза альфа 2 в коже не изменялась. Результат свидетельствует об усилении экспрессии гена Р4НА2 при использовании генетической конструкции pCMV6-XL5 Р4НА2 SEQ ID No: 5 с кДНК гена Р4НА2 и соответственно - об эффективности данной генетической конструкции. Результаты отражены на фигуре 23.
Пример 16.
Введение в хрящевую ткань человека генетической конструкции с кДНК гена Р4НА2 с целью подтверждения при этом увеличения количества белка пролил 4- гидрокислаза альфа 2 в хрящевой ткани человека.
С целью анализа изменения количества белка пролил 4- гидрокислаза альфа 2 пациентам вводили генетическую конструкцию VTvaf17 Р4НА2 SEQ ID No: 6 с кДНК гена Р4НА2 без транспортной системы (зона В) и параллельно вводили плацебо, представляющее собой комбинацию векторной плазмиды VTvaf17 не содержащей кДНК гена Р4НА2 без транспортной системы (зона А) - в хрящевую ткань ушных раковин с тыльной стороны.
Генетическую конструкцию VTvaf17 Р4НА2 SEQ ID No: 6, с кДНК гена Р4НА2 (SEQ ID No: 6), и плазмиду VTvaf17, используемую в качестве плацебо, которая не содержит кДНК гена Р4НА2, растворяли в стерильной воде степени очистки Nuclease-Free.
Полученные генетическую конструкцию и плацебо использовали для введения пациенту. Введение осуществляли тоннельным методом иглой 30G на глубину 1,5-2 мм в хрящевую ткань ушных раковин с тыльной стороны. Объем вводимого раствора генетической конструкции и плацебо - около 0,2 мл для каждого, концентрация генетической конструкции 0.5 мг/мл. Очаги введения генетической конструкции и плацебо располагались на расстоянии 2-3 см друг от друга.
Количество белка пролил 4- гидрокислаза альфа 2 оценивали в лизатах биоптатов хрящевой ткани пациента по методу, описанному в Примере 8.
Биопсийные образцы брали на 3 сутки после введения генетической конструкции. Взятие биопсии осуществляли из участков хрящевой ткани в зоне введения генетической конструкции, а также из интактных участков и в зоне введения плацебо, методом чрескожной пункционной биопсии при помощи одноразовой ручной биопсийной иглы, далее по примеру 9.
Показано, что в хрящевой ткани пациента в области введения генетической конструкции VTvaf17 Р4НА2 SEQ ID No: 6 с кДНК гена Р4НА2, произошло увеличение количества белка пролил 4- гидрокислаза альфа 2, тогда как при введении плацебо количество белка пролил 4- гидрокислаза альфа 2 в хрящевой ткани не изменялось. Результат свидетельствует об усилении экспрессии гена Р4НА2 при использовании генетической конструкции VTvaf17 Р4НА2 SEQ ID No: 6 и соответственно - об эффективности данной конструкции. Зезультаты отражены на фигуре 24
Пример 17.
Введение в мышечную ткань человека генетической конструкции с кДНК гена Р4НА2 с целью подтверждения при этом увеличения количества белка пролил 4- гидрокислаза альфа 2 в мышечной ткани человека.
С целью анализа изменения количества белка пациенту вводили генетическую конструкцию GDTT1.8NAS1-P4HA2 Р4НА2 SEQ ID No: 7 с кДНК гена Р4НА2 (зона В) и параллельно вводили плацебо, представляющее собой комбинацию векторной плазмиды GDTT1.8NAS1 не содержащей кДНК гена Р4НА2 с транспортной молекулой (зона А) - в мышечную ткань в зоне верхней трети предплечья.
В качестве транспортной системы использовали полиэтиленимин transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, France - USA).
Генетическую конструкцию GDTT1.8NAS1-P4HA2 P4HA2 SEQ ID No: 7, с кДНК гена P4HA2 (SEQ ID No: 7), и плазмиду GDTT1.8NAS1, используемую в качестве плацебо, которая не содержит кДНК гена Р4НА2, растворяли в стерильной воде степени очистки Nuclease-Free. Для получения генетической конструкции готовили комплексы ДНК- cGMP grade in-vivo-jetPEI в соответствие с рекомендациями производителя.
Расчет количества реагента осуществляли по формуле, предложенной производителем:
V(мкл)=(pDNA (мкг)×3×N/Р)/150
где N/P - соотношение между количеством азотистых остатков в jetPEI и количеством фосфатных групп нуклеиновых кислот.
Полученные генетическую конструкцию и плацебо использовали для введения пациенту. Введение осуществляли тоннельным методом иглой 30G на глубину 15-20 мм. Объем вводимого раствора генетической конструкции и плацебо - около 0,5 мл для каждого. Очаги введения генетической конструкции и плацебо располагались на расстоянии 5-7 см друг от друга
Количество белка пролил 4- гидрокислаза альфа 2 измеряли в лизатах биоптатов мышечной ткани пациента по методу, описанному в Примере 8.
Биопсийные образцы брали на 3 сутки после введения генетической конструкции. Взятие биопсии осуществляли из участков мышечной ткани в зоне введения генетической конструкции, а также из интактных участков мышцы и в зоне введения плацебо, используя автоматическое устройство для взятия биопсии MAGNUM (компания BARD, USA) в объеме не менее 25 куб.мм, далее по примеру 9.
Показано, что, в мышечной ткани пациента в области введения генетической конструкции GDTT1.8NAS1-P4HA2 Р4НА2 SEQ ID No: 7 произошло увеличение количества белка пролил 4- гидрокислаза альфа 2, тогда как при введении плацебо количество белка пролил 4- гидрокислаза альфа 2 в мышечной ткани не изменялось. Результат свидетельствует об усилении экспрессии гена Р4НА2 при использовании генетической конструкции GDTT1.8NAS1-P4HA2 Р4НА2 SEQ ID No: 7 и соответственно - об эффективности данной генетической конструкции. Результаты отражены на фигуре 25.
Пример 18.
Введение группе различных пациентов генетических конструкций, содержащих модифицированные кДНК гена Р4НА2 и участок нативной немодифицированной кДНК гена Р4НА2.
С целью подтверждения индивидуального характера увеличения уровня экспрессии целевого гена Р4НА2 до различного уровня при введении в кожу пациентов генетических конструкций с модифицированными и немодифицированной кДНК гена Р4НА2 анализировали уровень количества белка пролил 4-гидрокислазы альфа 2, в лизате биоптатов кожи группы пациентов в зависимости от наличия и типа модификаций в кДНК гена Р4НА2.
Последовательность участка нативной немодифицированной кДНК гена Р4НА2 приведена на фигуре 1, SEQ Р4НА2 ID No: 1, последовательности модифицированных кДНК гена Р4НА2 приведены на фигурах 2-7 (SEQ Р4НА2 ID No: 2, SEQ Р4НА2 ID No: 3, SEQ P4HA2 ID No: 4, SEQ P4HA2 ID No: 5, SEQ P4HA2 ID No: 6, SEQ P4HA2 ID No: 7).
При этом генетические конструкции на базе VTvaf17, одна из которых содержит участок нативной немодифицированной кДНК гена Р4НА2 (SEQ ID No: 1), вторая генетическая конструкция содержит участок модифицированной кДНК гена Р4НА2 (SEQ ID No: 2), третья генетическая конструкция содержит модифицированную кДНК гена Р4НА2 (SEQ ID No: 3), четвертая генетическая конструкция содержит модифицированную кДНК гена Р4НА2 (SEQ ID No: 4), пятая генетическая конструкция содержит модифицированную кДНК гена Р4НА2 (SEQ ID No: 5), шестая генетическая конструкция содержит модифицированную кДНК гена Р4НА2 (SEQ ID No: 6), седьмая генетическая конструкция содержит модифицированную кДНК гена Р4НА2 (SEQ ID No: 7), восьмая генетическая конструкция (плацебо), представляет собой вектор VTvaf17, растворяли в стерильной воде степени очистки Nuclease-Free.Для получения генетических конструкций готовили комплексы ДНК в сочетании с транспортной молекулой - Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, France - USA) no примеру 15.
Полученные семь вариантов генетических конструкций и плацебо использовали для введения в кожу пациентам. Введение осуществляли тоннельным методом иглой 30G на глубину 3 мм. Объем вводимого раствора каждой генетической конструкции составлял около 0,3 мл. Очаги введения генетических конструкций и плацебо располагались на расстоянии 3-5 см друг от друга.
Каждому из 32-х пациентов, которые были отобраны в случайном порядке, вводили 7 генетических конструкций и плацебо в кожу предплечья.
Биопсийные образцы брали через 72 часа после введения генетических конструкций. Взятие биопсии осуществляли из участков введения генетических конструкций, плацебо, а также из интактной кожи, используя устройство для взятия биопсии кожи Epitheasy 3.5 (Medax SRL). Кожу пациента предварительно промывали стерильным физиологическим раствором и анестезировали раствором лидокаина. Объем каждого биопсийного образца - не менее 10 куб. мм, масса - не менее 11 мг. Образец помещали в буферный раствор, содержащий 50 мМ Трис-HCI рН 7.6, 100 мМ NaCl, 1 мМ ЭДТА и 1 мМ фенилметилсульфонилфторид и гомогенизировали до получения однородной суспензии. Полученную суспензию центрифугировали в течение 10 минут при 14000 об/мин. Отбирали супернатант и использовали его для количественного определения целевого белка.
А - биоптат, полученный после введения генетической конструкции на базе VTvaf17 Р4НА2 SEQ ID No: 1, содержащий немодифицированную кДНК гена Р4НА2 (SEQ ID No: 1.) по примеру 2 в сочетании с транспортной молекулой - Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, France - USA) no примеру 15.
В - биоптат, полученный после введения генетической конструкции на базе VTvaf17 Р4НА2 SEQ ID No: 2, содержащий модифицированную кДНК гена Р4НА2 (SEQ ID No: 2.) по примеру 3 в сочетании с транспортной молекулой - Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, France - USA) по примеру 15.
С - биоптат, полученный после введения генетической конструкции на базе VTvaf17 Р4НА2 SEQ ID No: 3, содержащий модифицированную кДНК гена Р4НА2 (SEQ ID No: 3.) по примеру 3 в сочетании с транспортной молекулой - Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, France- USA) по примеру 15.
D - биоптат, полученный после введения генетической конструкции на базе VTvaf17 Р4НА2 SEQ ID No: 4, содержащий модифицированную кДНК гена Р4НА2 (SEQ ID No: 4.) по примеру 3 в сочетании с транспортной молекулой - Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, France - USA) по примеру 15.
E - биоптат, полученный после введения генетической конструкции на базе VTvaf17 Р4НА2 SEQ ID No: 5, содержащий модифицированную кДНК гена Р4НА2 (SEQ ID No: 5) по примеру 3 в сочетании с транспортной молекулой - Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, France - USA) по примеру 15.
F - биоптат, полученный после введения генетической конструкции на базе VTvaf17 Р4НА2 SEQ ID No: 6, содержащий модифицированную кДНК гена Р4НА2 (SEQ ID No: 6) по примеру 3 в сочетании с транспортной молекулой - Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, France - USA) по примеру 15.
G - биоптат, полученный после введения генетической конструкции на базе VTvaf17 Р4НА2 SEQ ID No: 7, содержащий модифицированную кДНК гена Р4НА2 (SEQ ID No: 7.) по примеру 3 в сочетании с транспортной молекулой - Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, France - USA) по примеру 15.
H - биоптат, полученный после введения векторной плазмиды VTvaf17, не содержащей кДНК гена Р4НА2 по примеру 3 в сочетании с транспортной молекулой - Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, France- USA) по примеру 15.
Количество белка пролил 4-гидрокислазы альфа 2 определяли в девяти биоптатах кожи от каждого пациента (от А до Н и в биоптате интактной кожи) через 72 часа после введения генетических конструкций методом твердофазного иммуноферментного анализа (ИФА ELISA), используя набор Human Р4НА2 ELISA Kit (MyBioSource, США), как описано в Примере 8.
По итогам количественного анализа белка пролил 4-гидрокислазы альфа 2, выбрали показатели, касательно каждого биоптата от каждого пациента, продемонстрировавшие максимальные уровни количества белка пролил 4-гидрокислазы альфа 2 и объединили их в группы, исходя из следующего критерия:
В группе 1 максимальное количество белка пролил 4-гидрокислазы альфа 2, наблюдалось при введении немодифицированной кДНК гена Р4НА2 SEQ ID No: 1. В эту группу вошел 7 биоптат из 32.
В группе 2 максимальное количество белка пролил 4-гидрокислазы альфа 2, наблюдалось при введении модифицированной кДНК гена Р4НА2 SEQ ID No: 2. В эту группу вошли 6 биоптата из 32.
В группе 3 максимальное количество белка пролил 4-гидрокислазы альфа 2, наблюдалось при введении модифицированной кДНК гена Р4НА2 SEQ ID No: 3. В эту группу вошли 4 биоптата из 32.
В группе 4 максимальное количество белка пролил 4-гидрокислазы альфа 2, наблюдалось при введении модифицированной кДНК гена Р4НА2 SEQ ID No: 4. В эту группу вошли 4 биоптата из 32.
В группе 5 максимальное количество белка пролил 4-гидрокислазы альфа 2, наблюдалось при введении модифицированной кДНК гена Р4НА2 SEQ ID No: 5. В эту группу вошли 3 биоптата из 32
В группе 6 максимальное количество белка пролил 4-гидрокислазы альфа 2, наблюдалось при введении модифицированной кДНК гена Р4НА2 SEQ ID No: 6. В эту группу вошли 4 биоптата из 32.
В группе 7 максимальное количество белка пролил 4-гидрокислазы альфа 2, наблюдалось при введении модифицированной кДНК гена Р4НА2 SEQ ID No: 7. В эту группу вошли 4 биоптата из 32.
Ни в одном из биоптатов, в которые были введены плацебо в виде векторной плазмидой, не содержащей кДНК гена Р4НА2, не было обнаружено максимальное количество белка пролил 4-гидрокислазы альфа 2, в отличие от биоптатов, в которые были введены генетические конструкции, содержаще кДНК гена Р4НА2.
На фигуре 26 для каждой группы биоптатов приведены диаграммы показателей концентраций белка (усредненных в рамках группы, в случае, если в группу входит более одного биоптата) применительно ко всем, участвующим в эксперименте генетическим конструкциям, после введения пациентам этих генетических конструкций, содержащих модифицированные и участок нативной кДНК гена Р4НА2.
Из данного примера следует, что достижение максимального уровня количества белка пролил 4- гидрокислазы альфа 2 в биоптатах кожи различных пациентов при введении им в кожу генетических конструкций, связано с индивидуальными особенностями пациентов и зависит от наличия и типа модификаций в кДНК гена Р4НА2, входящих в генетические конструкции.
Каждая генетическая конструкция из группы генетических конструкций является эффективной в некоторой значительной группе пациентов. Следовательно, для выбора наиболее эффективной генетической конструкции из группы генетических конструкций для терапевтических целей необходимо предварительное персонализированное исследование биоптатов пациента, либо клеток, выращенных из этих биоптатов на предмет максимальной эффективности терапевтического воздействия, созданных генетические конструкции в рамках группы генетических конструкций.
Пример 19.
Трансфекция клеточной линии фибробластов пациента разными генетическими конструкциями, содержащими модифицированные кДНК гена Р4НА2 и участок нативной немодифицированной кДНК гена Р4НА2 с целью персонализированного выбора из группы конструкций наиболее эффективной генетической конструкции применительно к данному пациенту для последующей трансфекции этой генетической конструкцией клеток пациента, либо введению этой генетической конструкции пациенту в рамках терапевтической процедуры,.
С целью определения наиболее эффективной применительно к конкретному пациенту генетической конструкции анализировали количество белка пролил 4- гидрокислаза альфа 2, в клеточных лизатах фибробластов этого пациента, трансфицированных разными генетическими конструкциями, содержащими участок нативной немодифицированной кДНК гена Р4НА2 и модифицированные кДНК гена Р4НА2.
Последовательность участка нативной немодифицированной кДНК гена Р4НА2 приведена на фигуре 1, SEQ Р4НА2 ID No: 1, последовательности модифицированных кДНК гена Р4НА2 приведены на фигурах 2-7 (SEQ Р4НА2 ID No: 2, SEQ Р4НА2 ID No: 3, SEQ P4HA2 ID No: 4, SEQ P4HA2 ID No: 5, SEQ P4HA2 ID No: 6, SEQ P4HA2 ID No: 7).
Культуры фибробластов из биоптата пациента вырастили по примеру 5, отобрали аликвоты и провели клеточный лизис: клеточный осадок, соответствующий 1×106 клеток, промывали фосфатным буфером и ресуспендировали во льду в лизирующем буфере, содержащем 25 мМ ХЕПЕС рН 7.9, 100 мМ NaCl, 1мМ ЭДТА, 1% Тритон Х-100, 10% глицерин и 1 мМ фенилметилсульфонилфторид. Лизат центрифугировали 5 минут при 14000 об/мин.
Затем культуру фибробластов пациента разделили на 27 частей. Одну часть, обозначенную (А), трансфицировали по примеру 7 генетической конструкцией pCMV6-XL5 SEQ ID No: 1, содержащей немодифицированную кДНК гена Р4НА2 (SEQ ID No: 1.) по примеру 2 в сочетании с транспортными молекулами -липосомами по примеру 9.
Вторую часть, обозначенную (В), трансфицировали по примеру 7 генетической конструкцией pCMV6- XL5 SEQ ID No: 2, содержащей вариант модифицированной кДНК гена Р4НА2 (SEQ ID No: 2.) по примеру 3 в сочетании с транспортными молекулами -липосомами по примеру 9.
3-ю часть, обозначенную (С), трансфицировали по примеру 7 генетической конструкцией pCMV6- XL5 SEQ ID No: 3, содержащей вариант модифицированной кДНК гена Р4НА2 (SEQ ID No: 3.) по примеру 3 в сочетании с транспортными молекулами - липосомами по примеру 9.
4-ю часть, обозначенную (D), трансфицировали по примеру 7 генетической конструкцией pCMV6- XL5 SEQ ID No: 4, содержащей вариант модифицированной кДНК гена Р4НА2 (SEQ ID No: 4.) по примеру 3 в сочетании с транспортными молекулами - липосомами по примеру 9.
5-ю часть, обозначенную (Е), трансфицировали по примеру 7 генетической конструкцией pCMV6- XL5 SEQ ID No: 5, содержащей вариант модифицированной кДНК гена Р4НА2 (SEQ ID No: 5.) по примеру 3 в сочетании с транспортными молекулами - липосомами по примеру 9.
6-ю часть, обозначенную (F), трансфицировали по примеру 7 генетической конструкцией pCMV6- XL5 SEQ ID No: 6, содержащей вариант модифицированной кДНК гена Р4НА2 (SEQ ID No: 6) по примеру 3 в сочетании с транспортными молекулами - липосомами по примеру 9.
7-ю часть, обозначенную (G), трансфицировали по примеру 7 генетической конструкцией pCMV6- XL5 SEQ ID No: 7, содержащей вариант модифицированной кДНК гена Р4НА2 (SEQ ID No: 7) по примеру в сочетании с транспортными молекулами - липосомами по примеру 9.
8-ю часть, обозначенную (Н), трансфицировали по примеру 7 векторной плазмидой pCMV6-XL5, не содержащей кДНК гена Р4НА2 в сочетании с транспортными молекулами - липосомами по примеру 9.
Определение количества белка пролил 4- гидрокислаза альфа 2, в клеточных лизатах фибробластов пациента проводили через 72 часа после трансфекции по методу, описанному в Примере 8.
По итогам определения количества белка пролил 4- гидрокислаза альфа 2, в культуре фибробластов пациента выделили вариант генетической конструкции, при трансфекции которой наблюдается максимальное количество белка пролил 4- гидрокислаза альфа 2 в лизате клеточной культуры, что в итоге свидетельствует о максимальной экспрессии гена Р4НА2. В данном эксперименте максимальное количество белка в лизате отмечена при трансфекции генетической конструкцией pCMV6-XL5 Р4НА2 SEQ ID No: 4, содержащей модифицированную кДНК гена Р4НА2, что показано на фигуре 27.
Таким образом, выбрана наиболее эффективная применительно к данному пациенту генетическая конструкция для последующей трансфекции клеток пациента в рамках терапевтической процедуры.
Пример 20.
Введение в кожу пациента различных генетических конструкций, содержащих модифицированные кДНК гена Р4НА2 или участок нативной немодифицированной кДНК гена Р4НА2 с целью выбора из группы наиболее эффективной генетической конструкции применительно к данному пациенту для последующего введения этой конструкции пациенту в рамках терапевтической процедуры.
С целью определения наиболее эффективной применительно к конкретному пациенту генетической конструкции анализировали количество белка в супернатанте биоптатов кожи этого пациента, после введения ему в кожу генетических конструкций, содержащих модифицированные кДНК гена Р4НА2 или участок нативной кДНК гена Р4НА2.
Последовательность участка нативной немодифицированной кДНК гена Р4НА2 приведена на фигуре 1, SEQ Р4НА2 ID No: 1, последовательности модифицированных кДНК гена Р4НА2 приведены на фигурах 2-7 (SEQ Р4НА2 ID No: 2, SEQ Р4НА2 ID No: 3, SEQ P4HA2 ID No: 4, SEQ P4HA2 ID No: 5, SEQ P4HA2 ID No: 6, SEQ P4HA2 ID No: 7).
Пациенту вводили 7 генетических конструкций и плацебо в кожу предплечья.
1-я генетическая конструкция (A) pCMV6-XL5 SEQ ID No: 1, содержащая участок нативной немодифицированной кДНК гена Р4НА2 (SEQ ID No: 1.) по примеру 2 в сочетании с транспортной молекулой - Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, France - USA) по примеру 15.
2-я генетическая конструкция (В) pCMV6- XL5 SEQ ID No: 2, содержащая участок модифицированной кДНК гена Р4НА2 (SEQ ID No: 2.) по примеру 3 в сочетании с транспортной молекулой - Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, France- USA) по примеру 15.
3-я генетическая конструкция (С) pCMV6- XL5 SEQ ID No: 3, содержащая участок модифицированной кДНК гена Р4НА2 (SEQ ID No: 3.) по примеру 3 в сочетании с транспортной молекулой - Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, France - USA) по примеру 15.
4-я генетическая конструкция (D) pCMV6- XL5 SEQ ID No: 4, содержащая участок модифицированной кДНК гена Р4НА2 (SEQ ID No: 4.) по примеру 3 в сочетании с транспортной молекулой - Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, France- USA) по примеру 15.
5-я генетическая конструкция (Е) pCMV6- XL5 SEQ ID No: 5, содержащая участок модифицированной кДНК гена Р4НА2 (SEQ ID No: 5.) по примеру 3 в сочетании с транспортной молекулой - Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, France- USA) по примеру 15.
6-я генетическая конструкция (F) pCMV6- XL5 SEQ ID No: 6, содержащая участок модифицированной кДНК гена Р4НА2 (SEQ ID No: 6.) по примеру 3 в сочетании с транспортной молекулой - Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, France- USA) по примеру 15.
7-я генетическая конструкция (G) pCMV6- XL5 SEQ ID No: 7, содержащая участок модифицированной кДНК гена Р4НА2 (SEQ ID No: 7.) по примеру 3 в сочетании с транспортной молекулой -Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, France- USA) по примеру 15.
8-я векторная плазмида pCMV6-XL5 (Н), не содержащая кДНК гена Р4НА2 по примеру 3 в сочетании с транспортной молекулой - Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, France- USA) по примеру 15.
При этом генетические конструкции, одна из которых содержит участок нативной немодифицированной кДНК гена Р4НА2 (SEQ ID No: 1), вторая генетическая конструкция содержит модифицированную кДНК гена Р4НА2 (SEQ ID No: 2), третья генетическая конструкция содержит модифицированную кДНК гена Р4НА2 (SEQ ID No: 3), четвертая генетическая конструкция содержит модифицированную кДНК гена Р4НА2 (SEQ ID No: 4), пятая генетическая конструкция содержит модифицированную кДНК гена Р4НА2 (SEQ ID No: 5), шестая генетическая конструкция содержит модифицированную кДНК гена Р4НА2 (SEQ ID No: 6), седьмая генетическая конструкция содержит модифицированную кДНК гена Р4НА2 (SEQ ID No: 7), восьмая плазмида (плацебо), представляет собой вектор pCMV6 XL5, растворяли в стерильной воде степени очистки Nuclease-Free. Для получения генетической конструкции готовили комплексы ДНК-полиэтиленимин Transfection reagent cGMP grade in-vivo-jetPEI (Polyplus Transfection, France- USA) по примеру 15.
Полученные семь вариантов генетических конструкций и плацебо использовали для введения пациенту. Введение осуществляли тоннельным методом иглой 30G на глубину 3 мм. Объем вводимого раствора каждой генетической конструкции составлял около 0,2 мл. Кожу в месте введения маркировали, очаги введения генетических конструкций и плацебо располагались на расстоянии 3-4 см друг от друга.
Биопсийные образцы брали через 72 часа после введения генетических конструкций. Взятие биопсии осуществляли из участков введения генетических конструкций, плацебо, а также из интактной кожи используя устройство для взятия биопсии кожи Epitheasy 3.5 (Medax SRL). Кожу пациента предварительно промывали стерильным физиологическим раствором и анестезировали раствором лидокаина. Размер каждого биопсийного образца был не менее 10 куб. мм, масса - не менее 11 мг.
Каждый образец помещали в буферный раствор, содержащий 50 мМ Трис-HCl рН 7.6, 100 мМ NaCl, 1 мМ ЭДТА и 1 мМ фенилметилсульфонилфторид и гомогенизировали до получения однородной суспензии. Полученную суспензию центрифугировали в течение 10 минут при 14000 об/мин. Отбирали супернатант и использовали его для количественного определения целевого белка.
Определение количества белка пролил 4- гидрокислаза альфа 2 проводили в девяти биоптатах кожи пациента через 72 часа после введения генетических конструкций по методу, описанному в Примере 8.
По итогам определения количества белка пролил 4- гидрокислаза альфа 2, в супернатанте биоптатов кожи пациента выделили вариант генетической конструкции, при введении которой в кожу наблюдается максимальное количество белка в супернатанте биоптата, что в итоге свидетельствует о максимальной экспрессии гена Р4НА2. В данном эксперименте максимальное количество целевого белка пролил 4- гидрокислаза альфа 2 в супернатанте биоптата кожи отмечено при введении генетической конструкции pCMV6-XL5 Р4НА2 SEQ ID No: 1, содержащей нативную кДНК гена Р4НА2, что показано на фигуре 28.
Таким образом, выбрана наиболее эффективная применительно к данному пациенту генетическая конструкция для Создана генетическая конструкция на основе невирусной векторной плазмиды, включающей кДНК гена Р4НА2, для купирования проявлений состояний человеческого организма, связанных с уменьшением экспрессии гена Р4НА2 и/или уменьшением количества белка пролил 4- гидрокислаза альфа 2, способ ее получения и использования.
Созданная генетическая конструкция для купирования проявлений состояний человеческого организма, связанных с уменьшением экспрессии гена Р4НА2 и/или уменьшением количества белка пролил 4- гидрокислаза альфа 2, выбрана из группы генетических конструкций, каждая из которых создана на основе векторной невирусной плазмиды, включающей нативную или модифицированную кДНК гена Р4НА2, с кодирующей последовательностью белка пролил 4- гидрокислаза альфа 2.
Созданная генетическая конструкция на основе невирусного ДНК-вектора, включающего кДНК гена Р4НА2 для купирования проявлений состояний человеческого организма, связанных с уменьшением экспрессии гена Р4НА2 и/или уменьшением количества белка пролил 4- гидрокислаза альфа 2, выбранная из группы генетических конструкций с одной из модифицированных кДНК гена Р4НА2 или с участком нативной немодифицированной кДНК гена Р4НА2, позволяет на практике повышать до необходимого уровня экспрессию белка пролил 4- гидрокислаза альфа 2, в различных клетках органов и тканей и/или органах и тканях человека путем увеличения экспрессии гена Р4НА2.
В результате проведения предварительных исследований биоптата пациента, либо клеток, выращенных из этих биоптатов, определяют, какой вариант генетической конструкции из созданной группы генетических конструкций необходимо выбрать для данного пациента для того, чтобы повысить количество белка пролил 4- гидрокислаза альфа 2, в клетках этих органов и тканей и/или органах и тканях до необходимого уровня применительно к конкретному пациенту. В тех случаях, когда использование генетической конструкции с участком нативной немодифицированной кДНК гена Р4НА2 не приводит к желаемому изменению количественного уровня белка пролил 4- гидрокислаза альфа 2, в клетках органов и тканей и/или органах и тканях, необходимо применять генетическую конструкцию с модифицированной кДНК гена Р4НА2, приводящей к более эффективной экспрессии гена Р4НА2.
Генетическая конструкция, выбранная из группы генетических конструкций с одной из модифицированных кДНК гена Р4НА2 или с участком нативной немодифицированной кДНК гена Р4НА2, обеспечивает высокий уровень экспрессии гена Р4НА2, повышая количество белка пролил 4- гидрокислаза альфа 2, в клетках органов и тканей и/или органах и тканях человека, а именно, в гемопоэтических клетках, или мезенхимальных стволовых клетках, или хондробластах, или миоцитах, или миобластах, или фибробластах, или остеобластах, или кератоцитах, или эпителиальных клетках, или клетках роговицы, или эндотелиальных клетках, или эпителиальных клетках, в сочетании с транспортной молекулой или без нее при трансфекции этими генетическими конструкциями клеток органов и тканей человека и/или в органах и тканях человека, а именно, в соединительнотканных структурах, или коже, или суставах, или хрящевой ткани, или костной ткани, или мышечной ткани, или сухожилиях, или связках, или кровеносных сосудах, или стенке артерий, или роговице глаза, или склере, или плаценте, или дентине, или строме внутренних органов в сочетании с транспортной молекулой или без нее при введении этих генетических конструкций в органы и ткани человека.
Таким образом, приведенные примеры подтверждают выполнение поставленной задачи, а именно, создание генетической конструкции на основе невирусной векторной плазмиды, включающей кДНК гена Р4НА2, для купирования проявлений состояний человеческого организма, связанных с уменьшением экспрессии гена Р4НА2 и/или уменьшением количества белка пролил 4- гидрокислаза альфа 2, выбранной из группы генетических конструкций, каждая из которых создана на основе векторной невирусной плазмиды, включающей нативную или модифицированную кДНК гена Р4НА2, при использовании которой с учетом индивидуальных особенностей пациента, происходит повышение уровня экспрессии гена Р4НА2 и/или повышение количества белка пролил 4- гидрокислаза альфа 2, в клетках органов и тканей и/или органах и тканях организма.
Кроме этого при введении генетических конструкций в отличие от введения непосредственно белка, снижается требуемая частота их введения в связи с пролонгированным действием, а также облегчается внутриклеточная доставка.
При использовании заявленной выбранной из группы генетической конструкции не происходит встраивания экзогенного генетического материала в геном клетки. В случае использования генетической конструкции на базе векторов VTvaf17 и GDTT1.8NAS1, GDTT1.8NAS3, GDTT1.8NAS7 абсолютно исключается вероятность встраивания элементов вирусных геномов в геном пациента и возникновения у пациента вследствие выполненной терапевтической процедуры антибиотикорезистентности.
Повышение эффективности коррекции количественного уровня белка пролил 4- гидрокислаза альфа 2 в клетках органов и тканей человека достигают за счет того, что при недостаточном количестве этого белка вследствие дефекта или недостаточности экспрессии гена Р4НА2 используют не белок, а генетическую конструкцию с кДНК гена Р4НА2, кодирующую этот белок.
Промышленная применимость.
Все приведенные примеры по созданию и использованию созданной генетической конструкции, выбираемой из группы генетических конструкций, подтверждают ее промышленную применимость.
Перечень сокращений
ДНК - дезоксирибонуклеиновая кислота
кДНК - комплементарная дезоксирибонуклеиновая кислота
РНК - рибонуклеиновая кислота
мРНК - матричная рибонуклеиновая кислота
ПЦР - полимеразная цепная реакция
мл - миллилитр,
мкл - микролитр
л - литр
куб. мм - кубический миллиметр
мкг - микрограмм
мг - миллиграмм
г - грамм
мкМ - микромоль
мМ - миллимоль
об/мин - обороты в минуту
нм - нанометр
см - сантиметр
мВт - милливатт
о.е. ф-относительная единица флуоресценции
ГК - генетическая конструкция












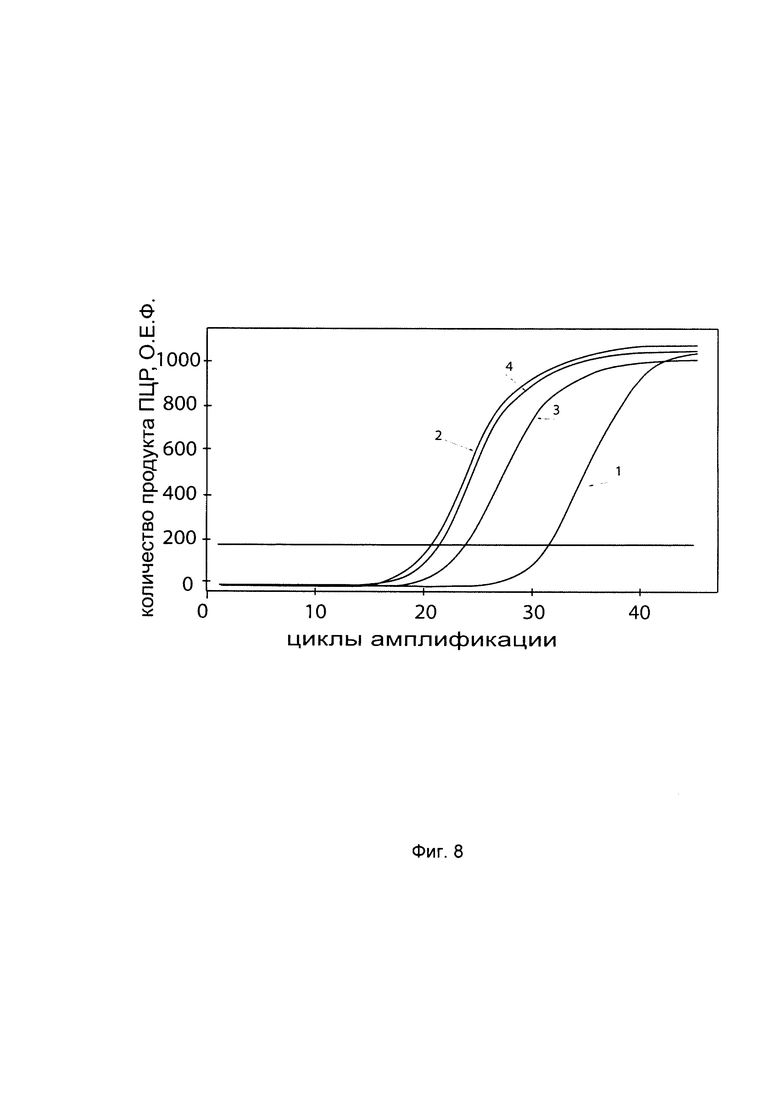
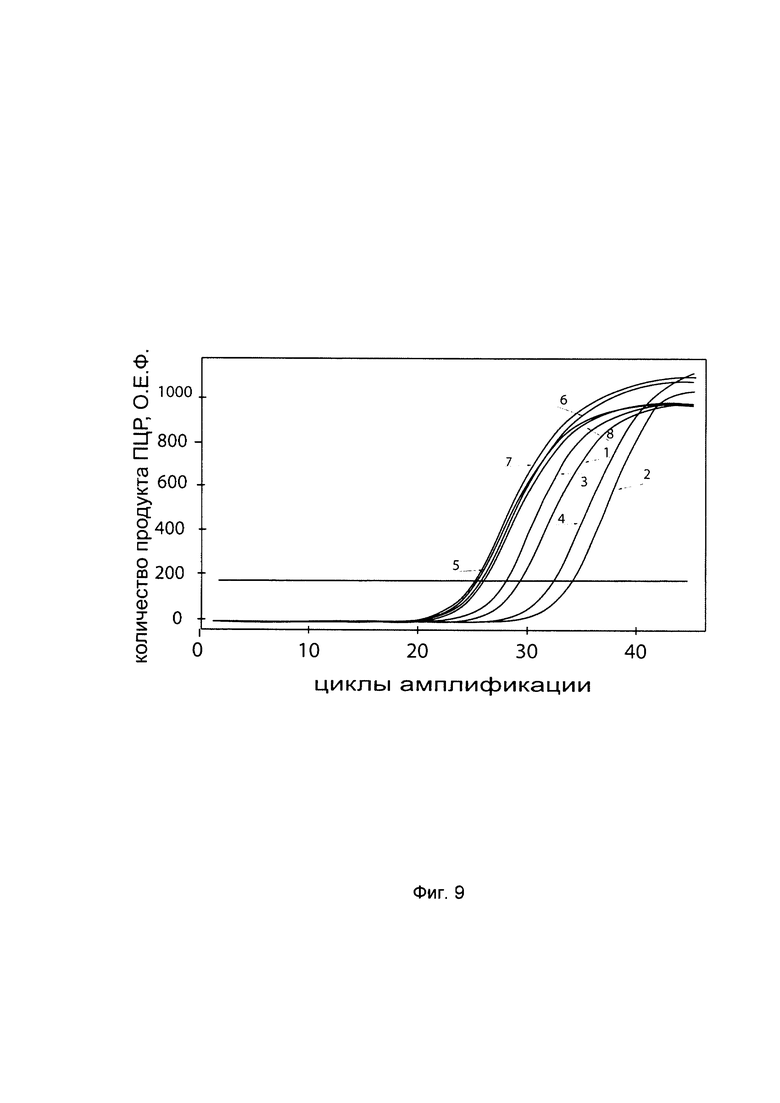
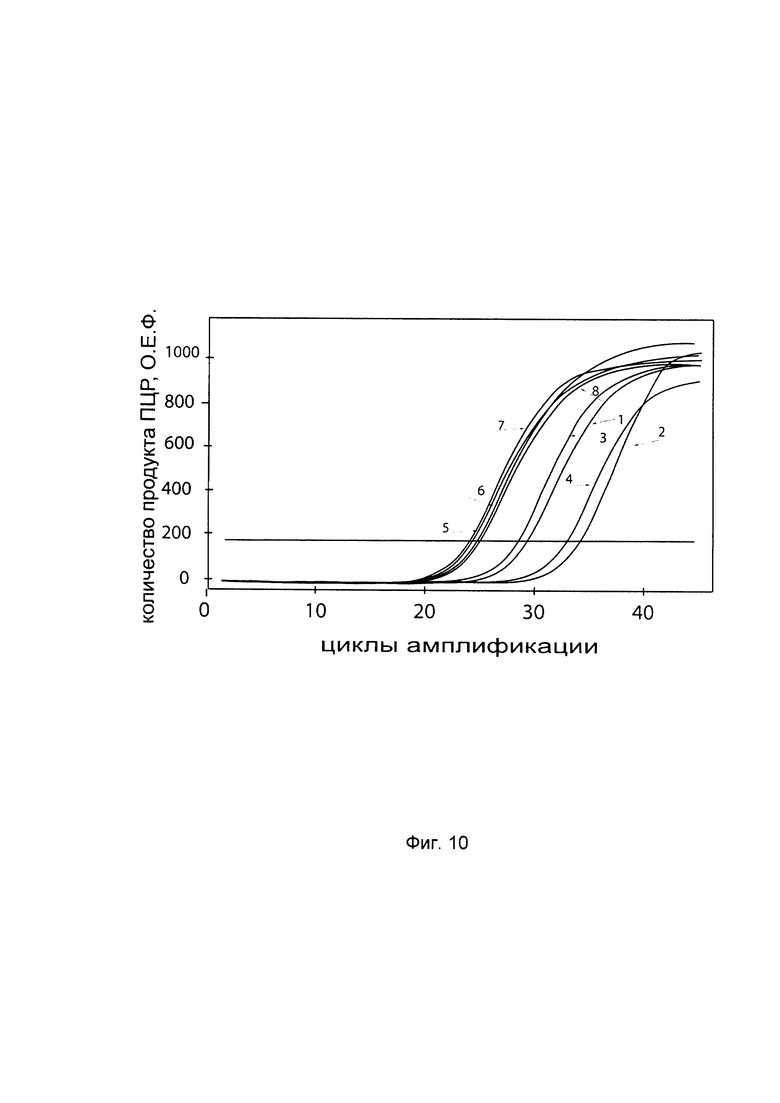
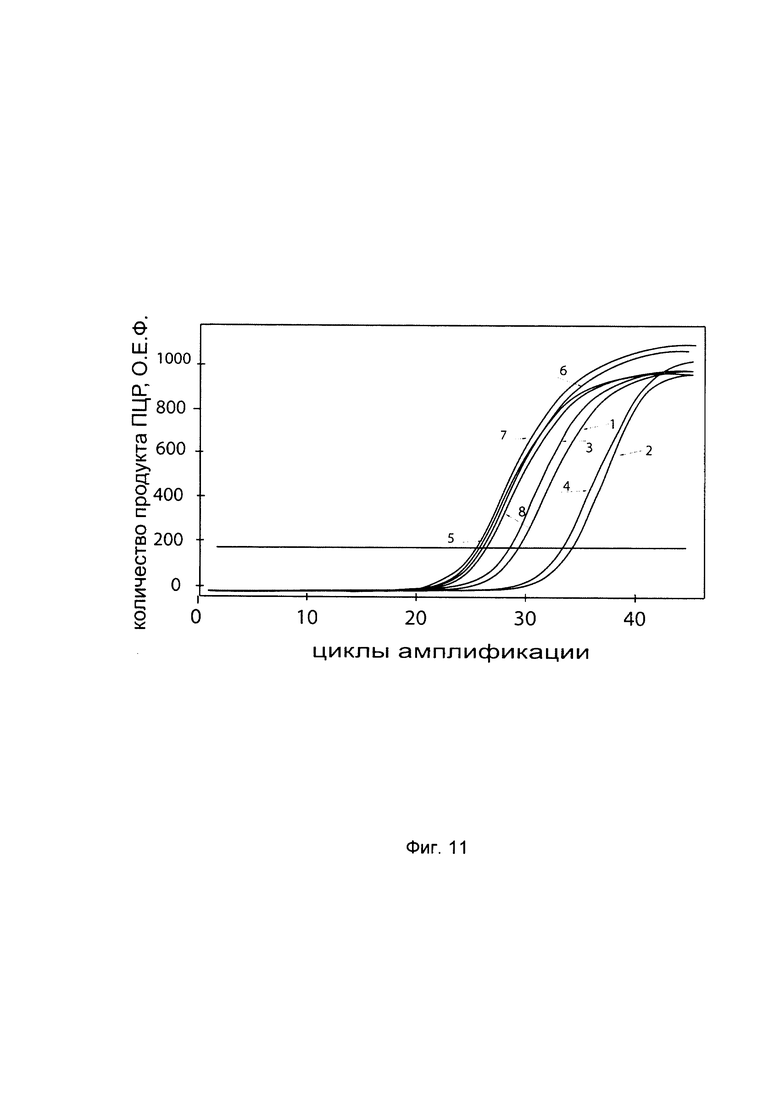
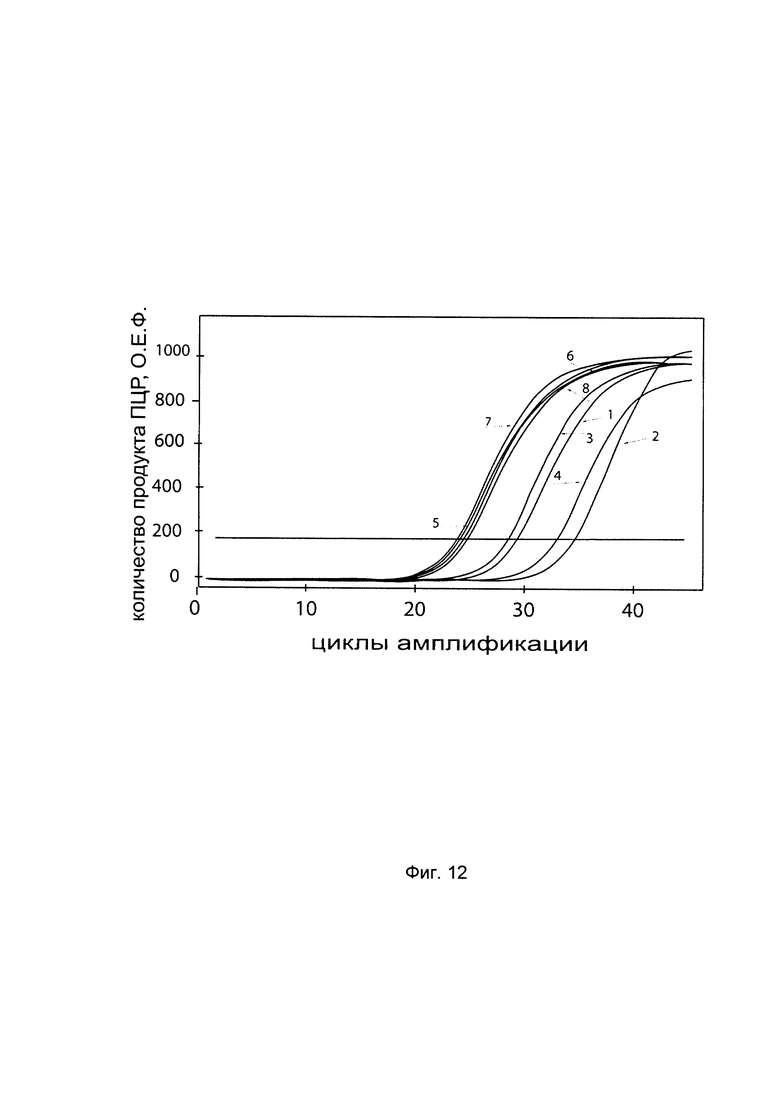
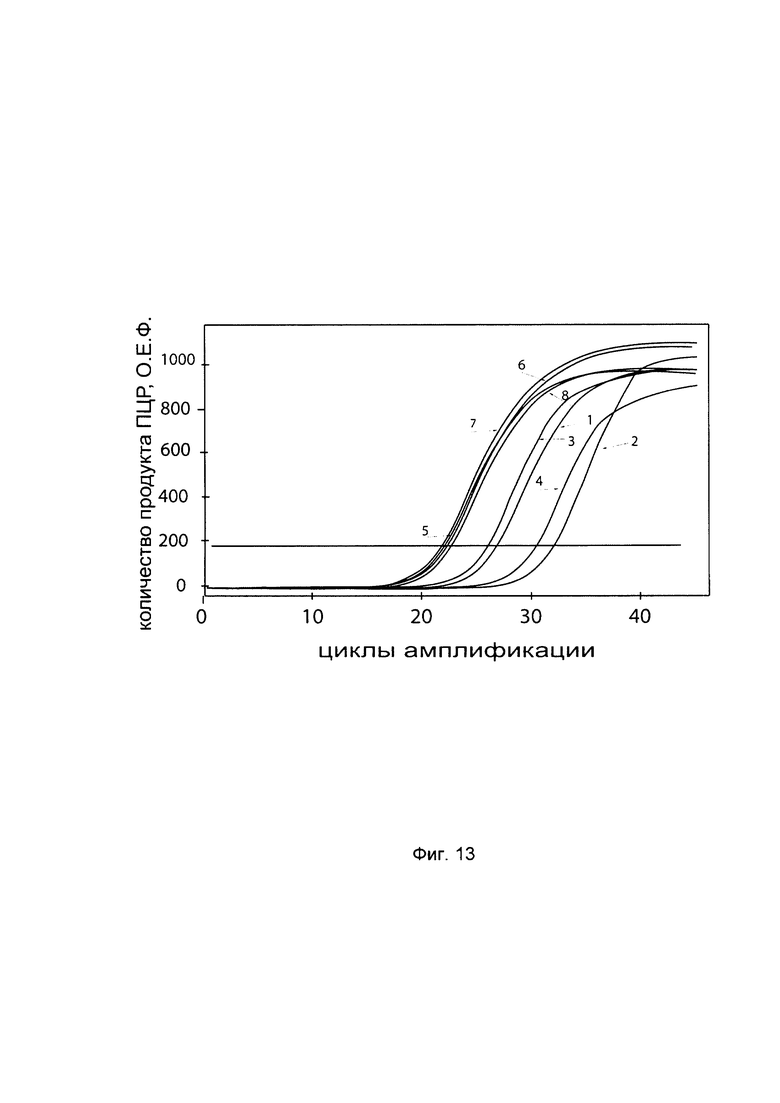
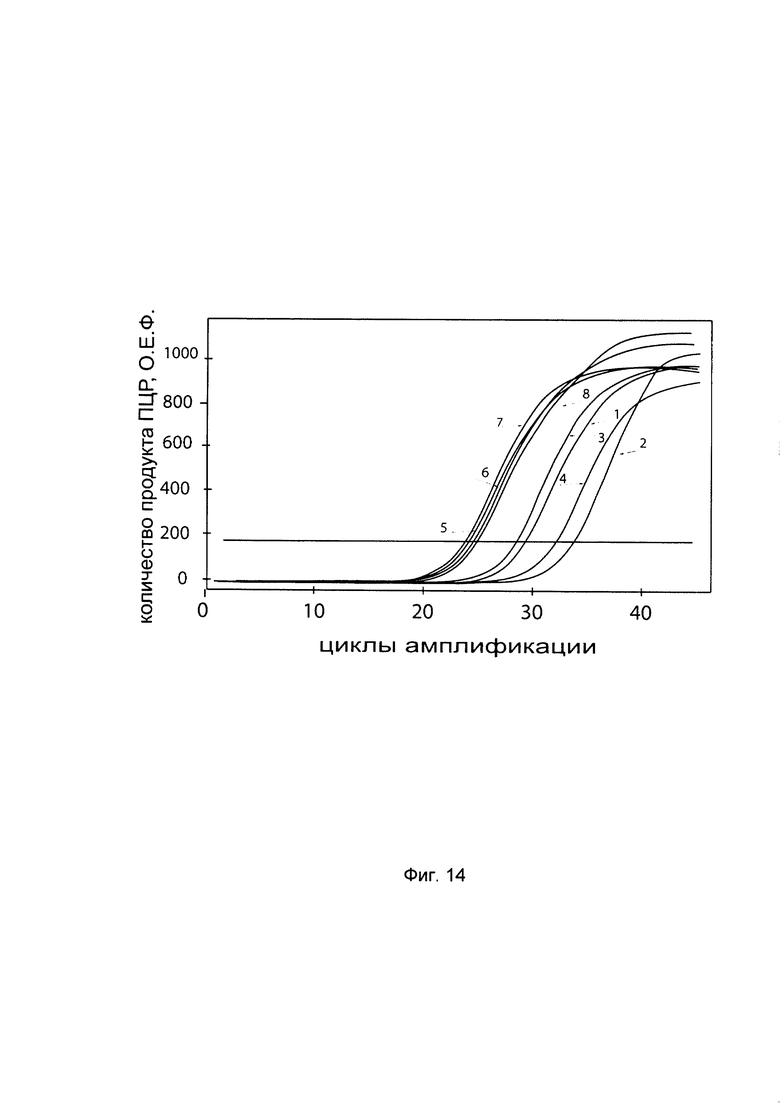
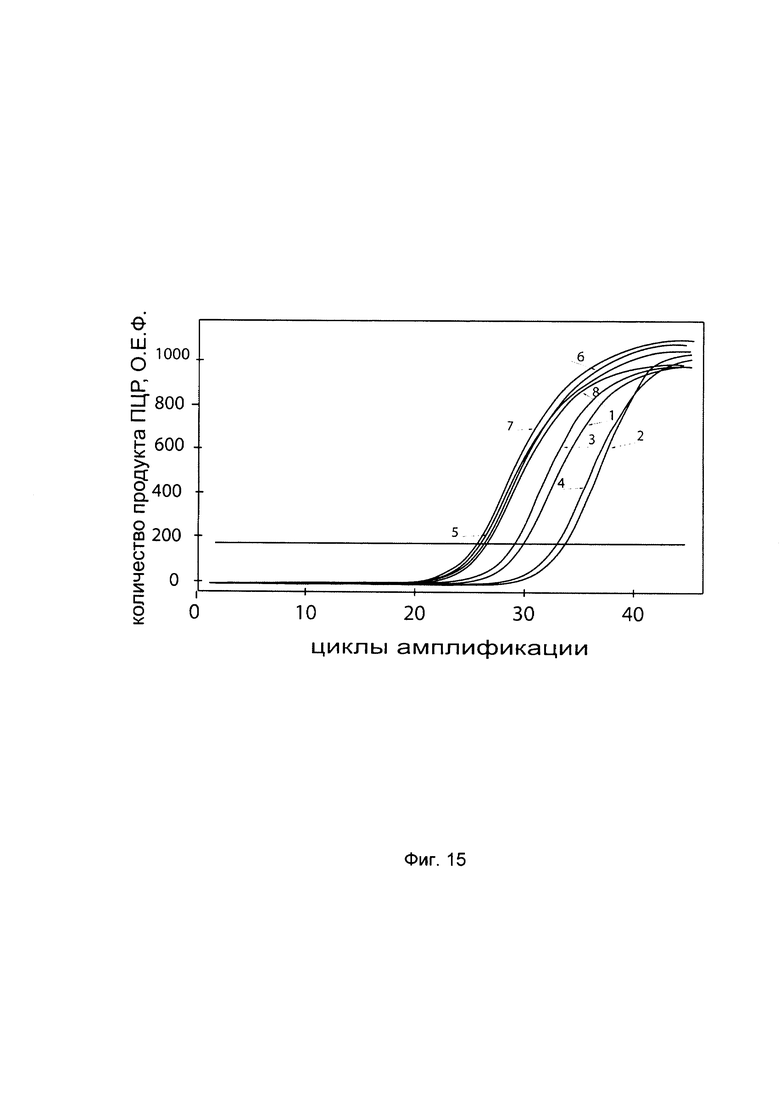
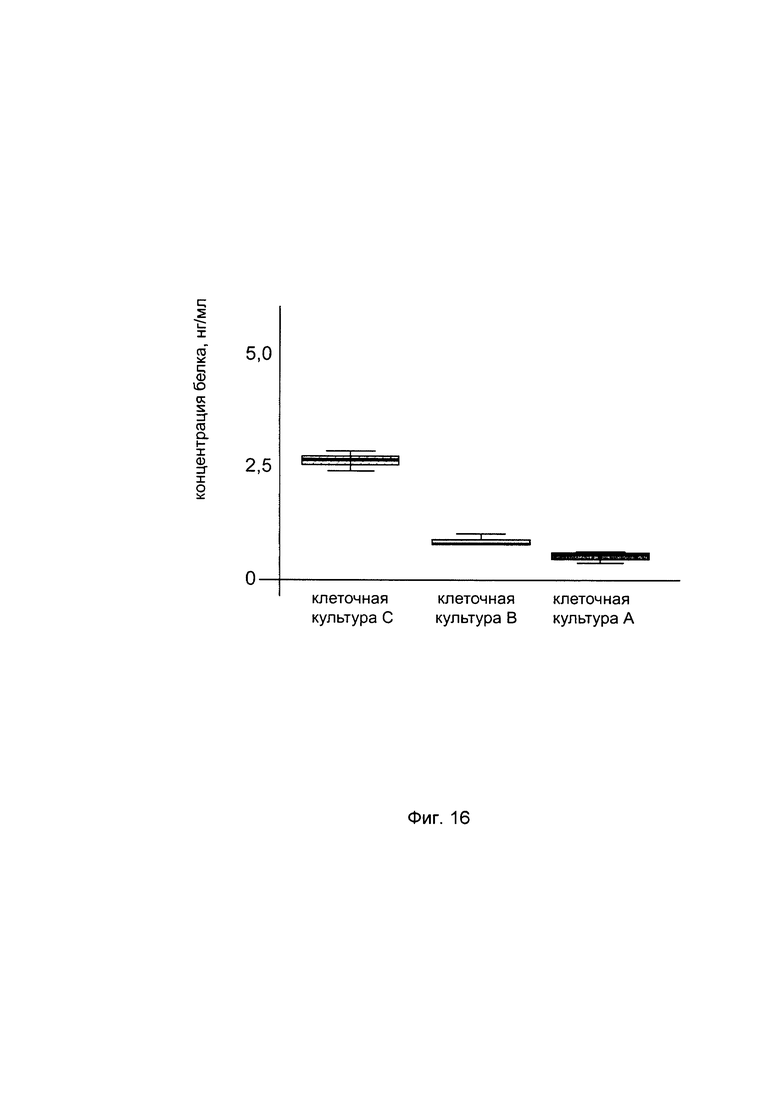
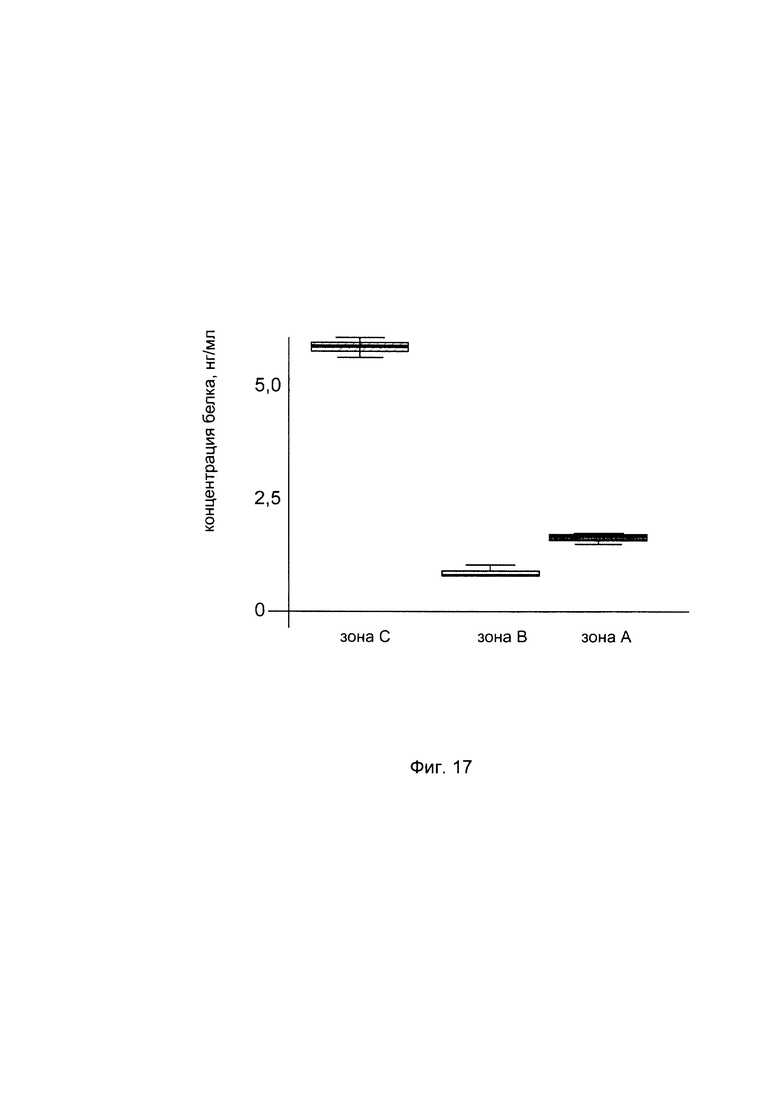
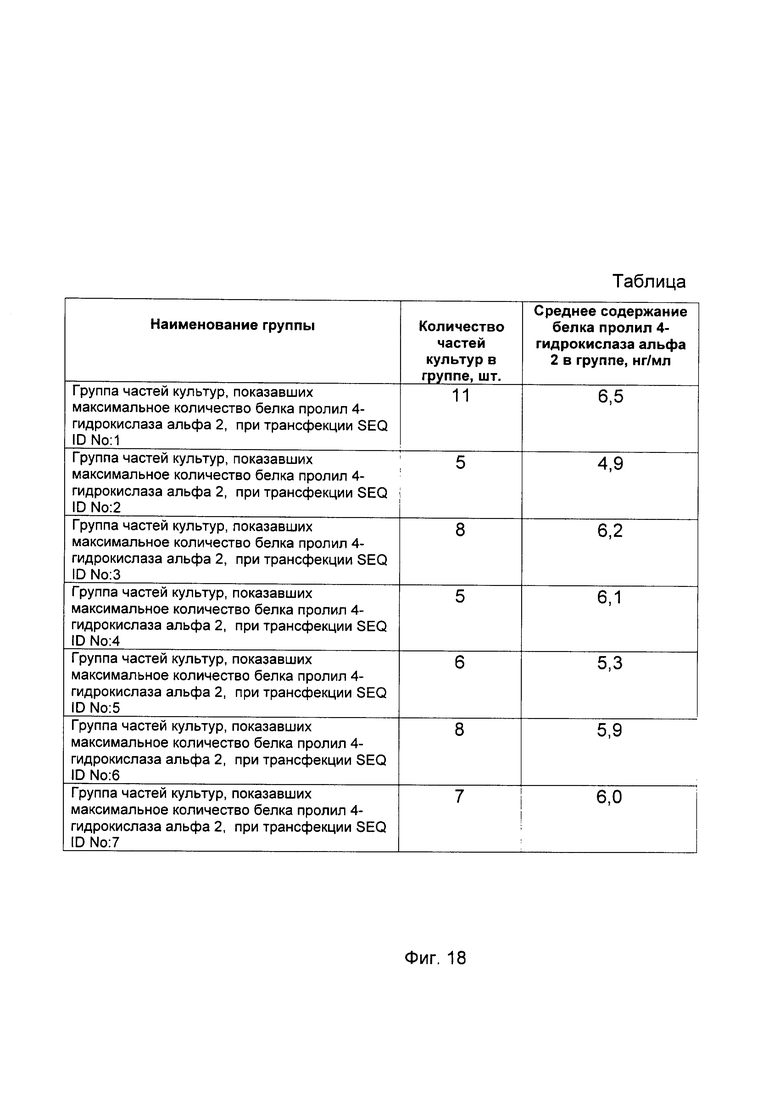
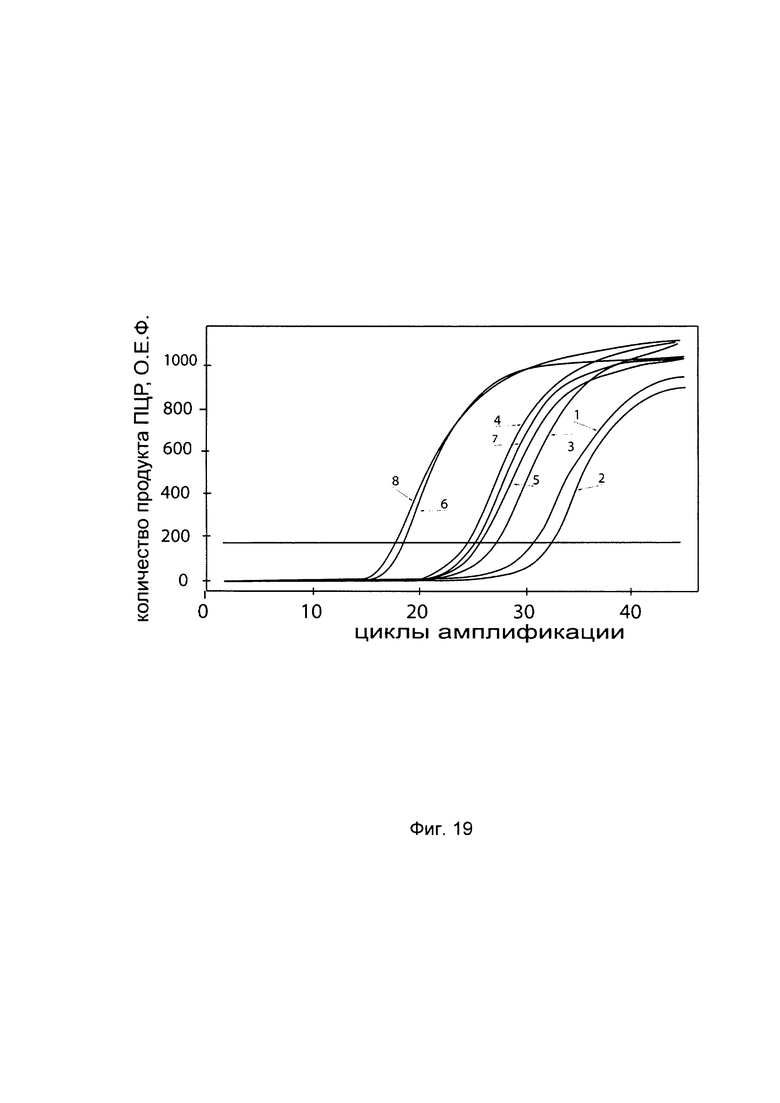
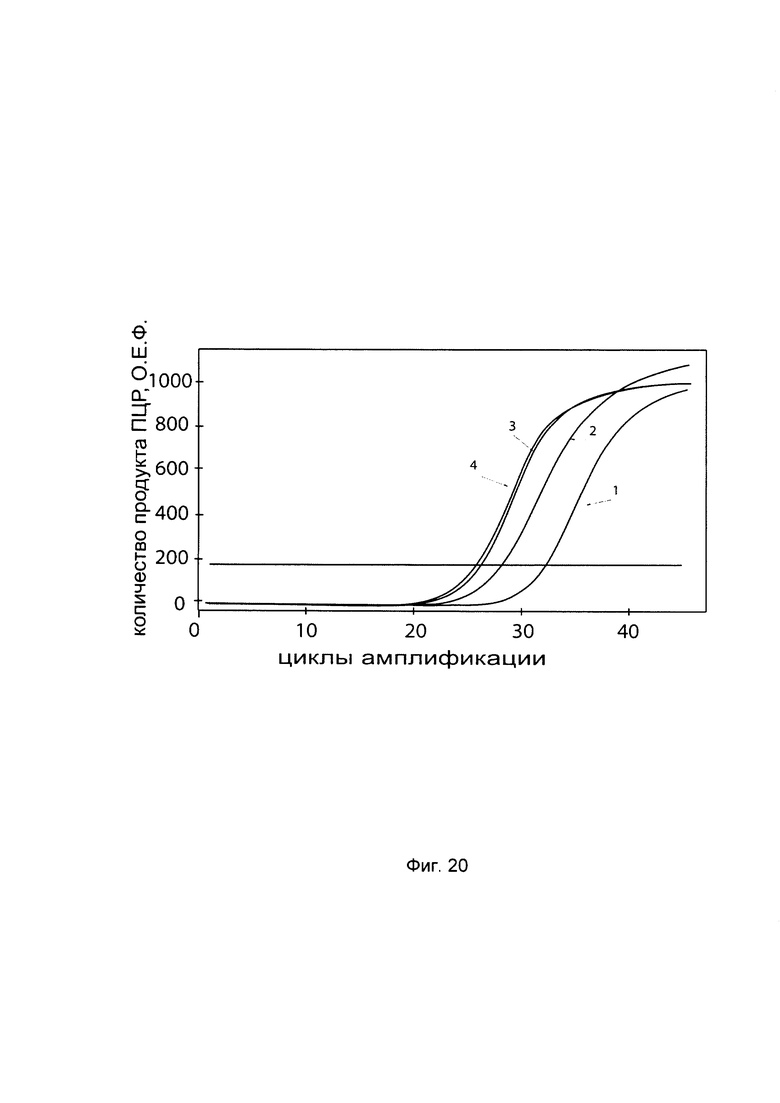
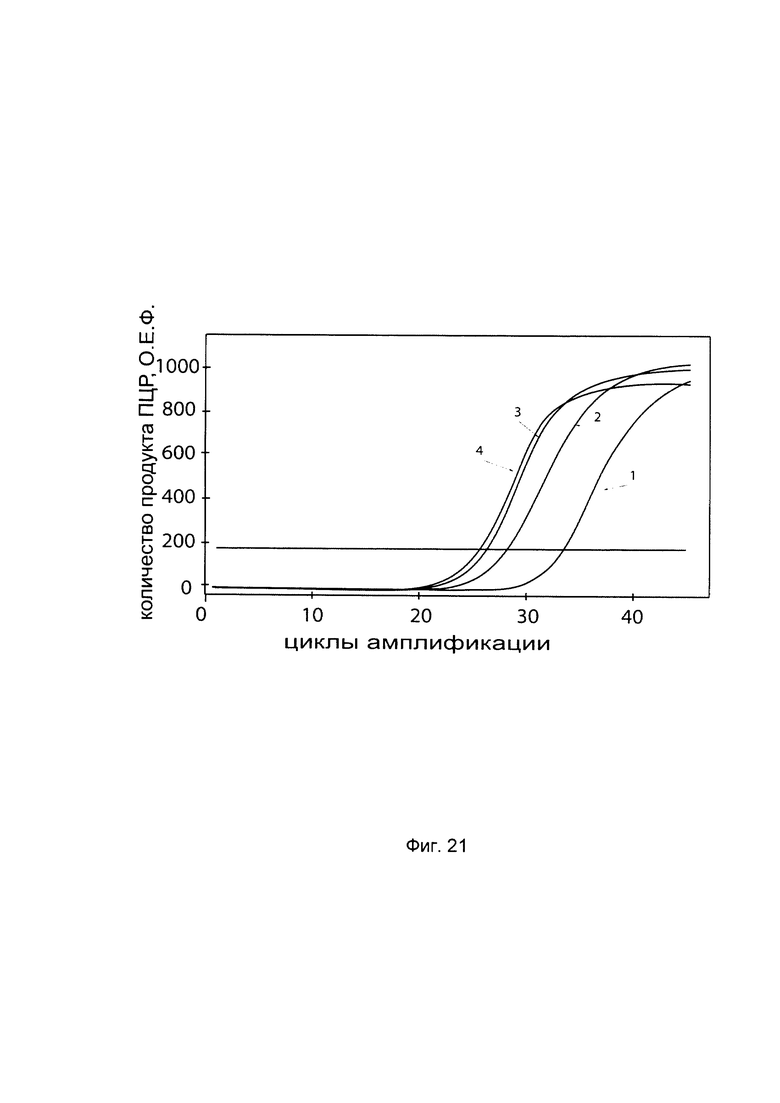
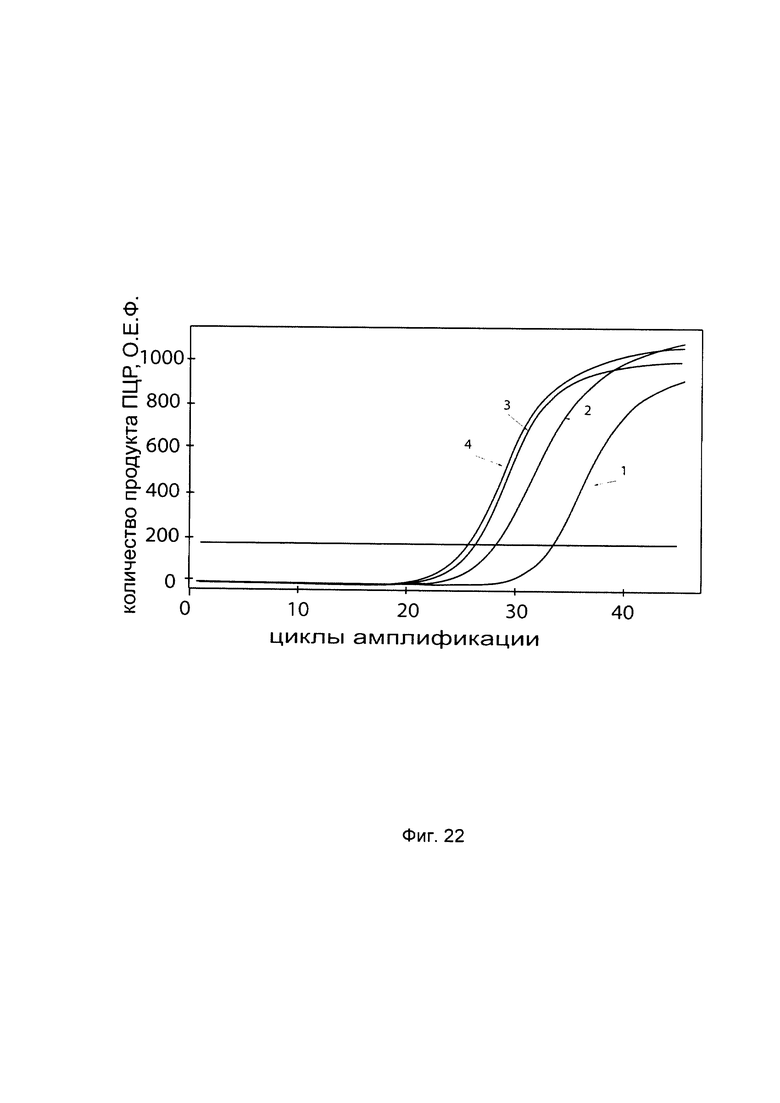
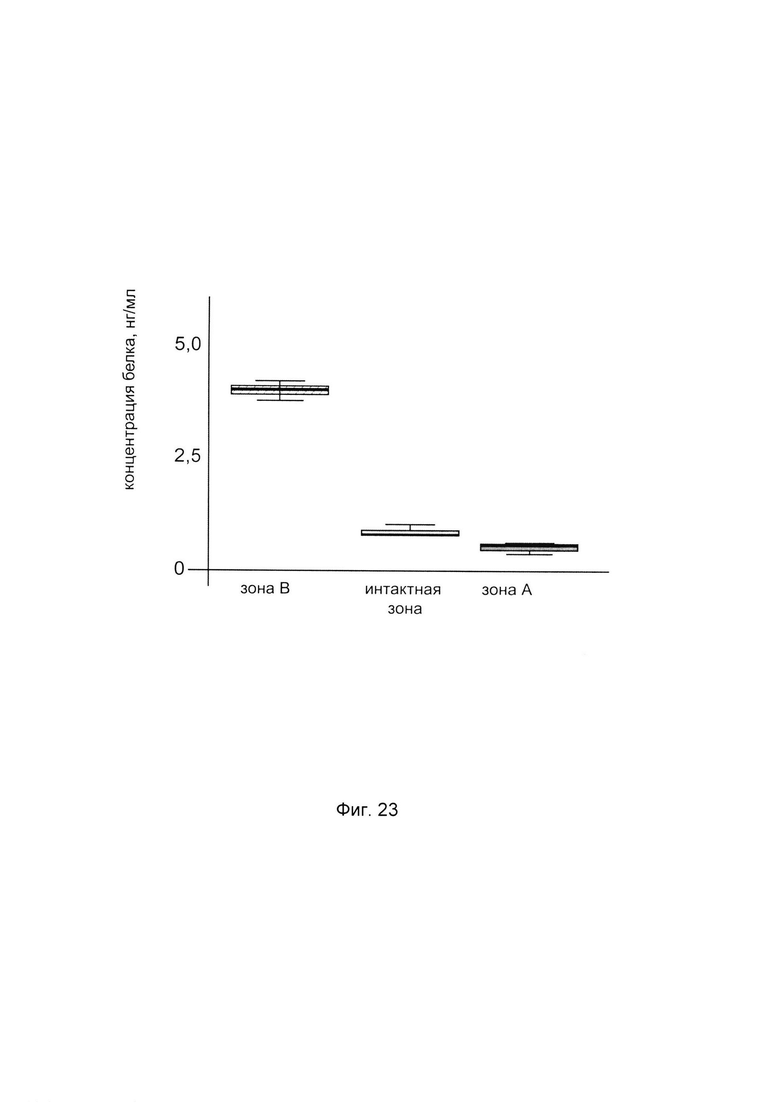
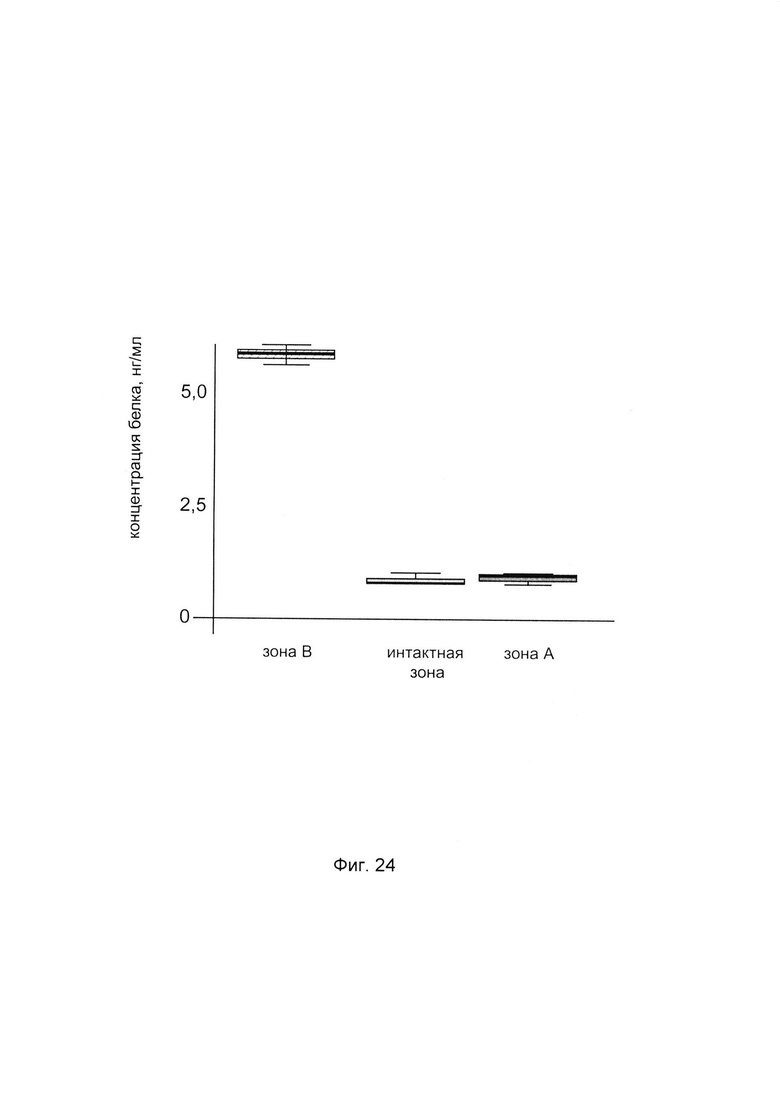
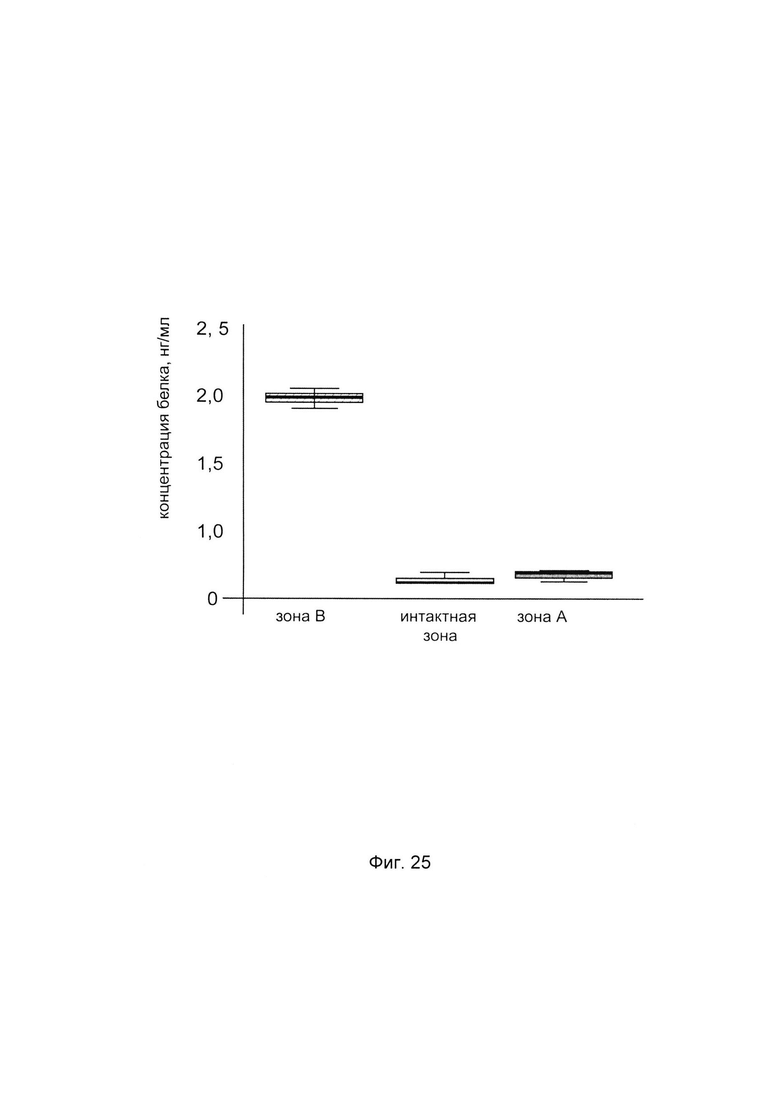
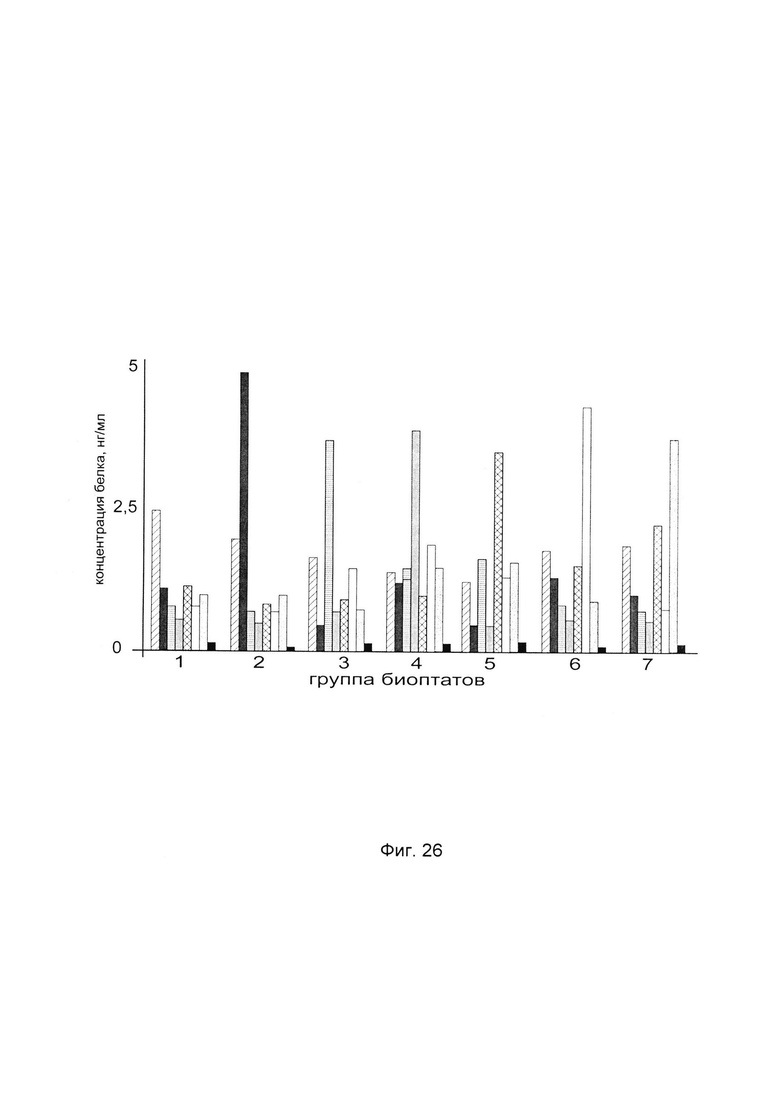
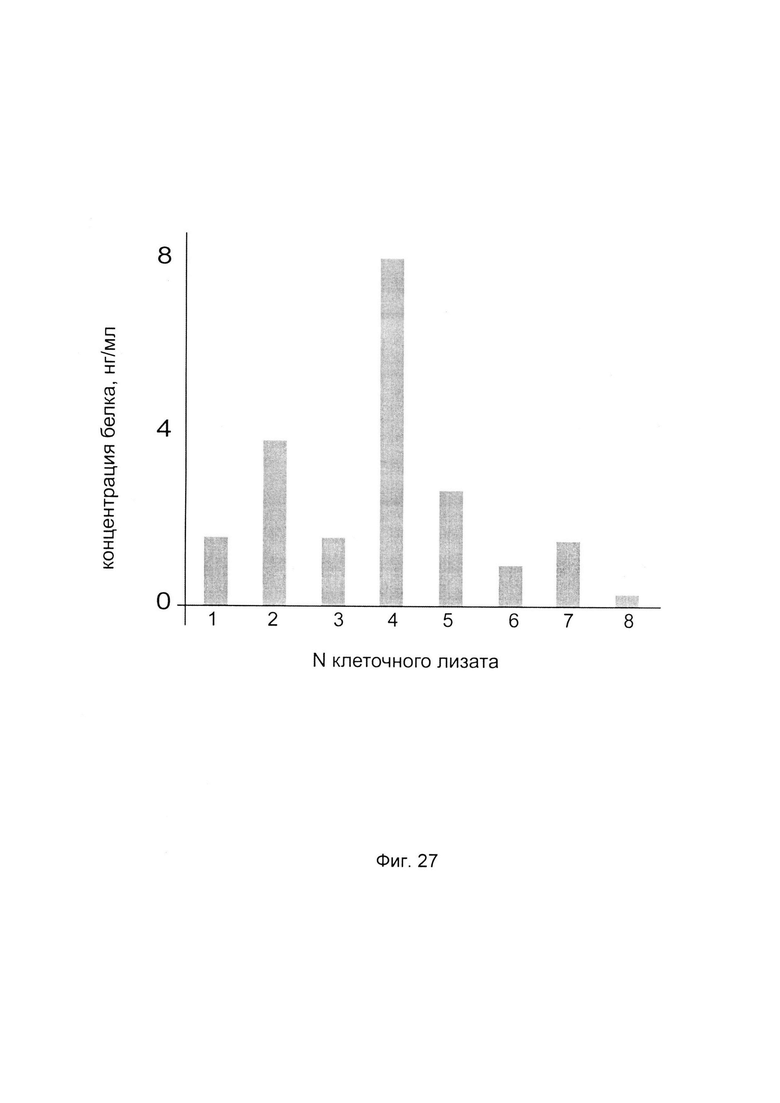
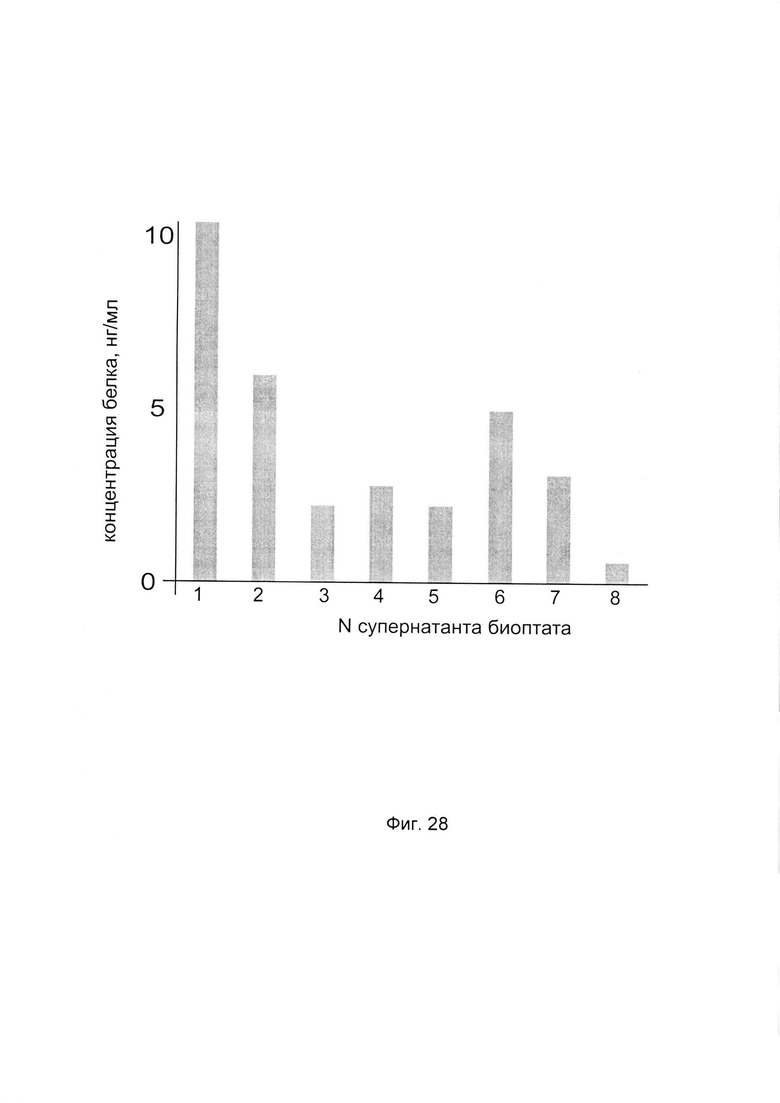
Изобретение относится к молекулярной биологии, биотехнологии, генной инженерии и медицине и может быть использовано для купирования проявлений состояний человеческого организма, связанных с уменьшением экспрессии гена Р4НА2 и/или уменьшением количества белка пролил 4-гидрокислаза альфа 2. Предложена генетическая конструкция на основе невирусного ДНК-вектора, включающего кДНК гена Р4НА2, выбранная из генетических конструкций, каждая из которых представляет генетическую конструкцию на основе ДНК-вектора, включающего кДНК гена Р4НА2, с кодирующей последовательностью белка пролил 4-гидрокислаза альфа 2, с делециями 5' и 3'-нетранслируемых областей, а именно полученной на основе участка нативной немодифицированной кДНК гена Р4НА2 SEQ ID No: 1 или модифицированной кДНК гена Р4НА2, при этом в качестве модифицированной кДНК гена Р4НА2 используют SEQ ID No: 2, или SEQ ID No: 3, или SEQ ID No: 4, или SEQ ID No: 5, или SEQ ID No: 6, или SEQ ID No: 7, или сочетание этих генетических конструкций, каждая из которых содержит также регуляторные элементы, обеспечивающие высокий уровень экспрессии гена Р4НА2 в эукариотических клетках. Предложены также способ получения генетической конструкции и ее применение. Изобретение позволяет увеличить количество белка пролил 4-гидроксилазы альфа 2 в эукариотических клетках. 3 н. и 3 з.п. ф-лы, 28 ил., 20 пр.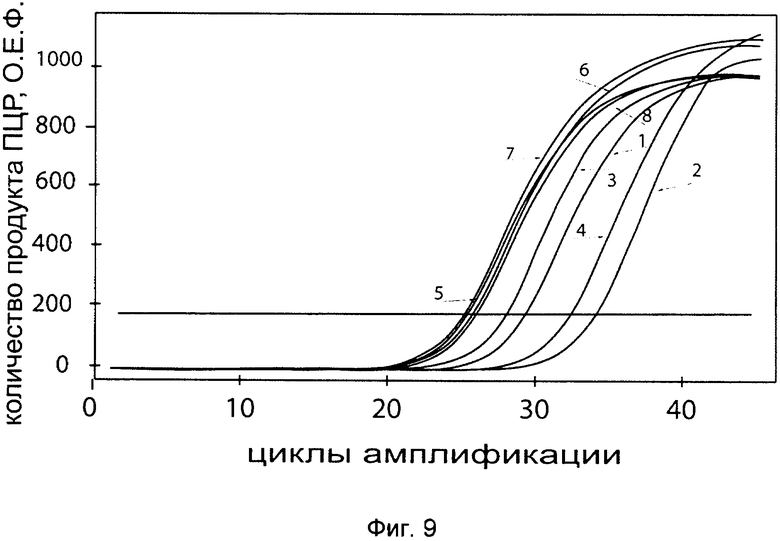
1. Генетическая конструкция на основе невирусного ДНК-вектора, включающего кДНК гена Р4НА2, для купирования проявлений состояний человеческого организма, связанных с уменьшением экспрессии гена Р4НА2 и/или уменьшением количества белка пролил 4- гидрокислаза альфа 2, выбранная из генетических конструкций, каждая из которых представляет генетическую конструкцию на основе ДНК-вектора, включающего кДНК гена Р4НА2, с кодирующей последовательностью белка пролил 4-гидрокислаза альфа 2, с делециями 5'- и 3'-нетранслируемых областей, а именно полученной на основе участка нативной немодифицированной кДНК гена Р4НА2 SEQ ID No:1 или модифицированной кДНК гена Р4НА2, при этом в качестве модифицированной кДНК гена Р4НА2 используют SEQ ID No:2, или SEQ ID No:3, или SEQ ID No:4, или SEQ ID No:5, или SEQ ID No:6, или SEQ ID No:7, или сочетание этих генетических конструкций, каждая из которых содержит также регуляторные элементы, обеспечивающие высокий уровень экспрессии гена Р4НА2 в эукариотических клетках, а именно в клетках органов и тканей человека, и способную увеличить количество белка пролил 4-гидрокислаза альфа 2 в клетках органов и тканей, а именно в гемопоэтических клетках, или мезенхимальных стволовых клетках, или хондробластах, или миоцитах, или миобластах, или фибробластах, или остеобластах, или кератоцитах, или эпителиальных клетках, или клетках роговицы, или эндотелиальных клетках, или эпителиальных клетках, в сочетании с транспортной молекулой или без нее при трансфекции этими генетическими конструкциями клеток органов и тканей человека, и/или в органах и тканях человека, а именно в соединительно-тканных структурах, или коже, или суставах, или хрящевой ткани, или костной ткани, или мышечной ткани, или сухожилиях, или связках, или кровеносных сосудах, или стенке артерий, или роговице глаза, или склере, или плаценте, или дентине, или строме внутренних органов, в сочетании с транспортной молекулой или без нее при введении этих генетических конструкций в органы и ткани человека.
2. Генетическая конструкция по п.1, отличающаяся тем, что каждая генетическая конструкция, выбранная из одной или нескольких генетических конструкций с модифицированной кДНК гена Р4НА2, содержит последовательность нуклеотидов, включающую в себя белок-кодирующую область кДНК гена Р4НА2, которая несет модификации, не затрагивающие структуру белка пролил 4-гидрокислаза альфа 2 или затрагивающие структуру белка пролил 4-гидрокислаза альфа 2 и влияющие на кодируемую этой последовательностью аминокислотную последовательность, а именно нуклеотидные замены, приводящие к аминокислотным заменам или обрыву аминокислотной цепи, и/или делеции, или комбинации вышеперечисленных модификаций.
3. Генетическая конструкция по п.1, отличающаяся тем, что в качестве транспортной молекулы используют липосомы, или дендримеры 5-го и выше поколений, или амфифильные блоксополимеры, или реагент на основе полиэтиленимина.
4. Способ получения генетической конструкции по п.1, заключающийся в получении каждой генетической конструкции, выбранной из одной или нескольких генетических конструкций по п.1, при этом получают кДНК гена Р4НА2, затем помещают кДНК в векторную плазмиду, способную обеспечить высокий уровень экспрессии этой кДНК в клетках различных органов и тканей человека, наращивают и выделяют необходимое количество генетической конструкции, затем комбинируют генетическую конструкцию с транспортной молекулой для трансфекции полученной генетической конструкцией клеток органов и тканей и/или введения полученной генетической конструкции в органы и ткани человека, при этом используют кДНК гена Р4НА2 с кодирующей последовательностью белка пролил 4-гидрокислаза альфа 2, с делециями 5'- и 3'-нетранслируемых областей, а именно полученной на основе участка нативной немодифицированной кДНК гена Р4НА2 SEQ ID No:1 или модифицированной кДНК гена Р4НА2, при этом в качестве модифицированной кДНК гена Р4НА2 используют или SEQ ID No:2, или SEQ ID No:3, или SEQ ID No:4, или SEQ ID No:5, или SEQ ID No:6, или SEQ ID No:7, или сочетание этих генетических конструкций.
5. Способ применения генетической конструкции по п.1 для купирования проявлений состояний человеческого организма, связанных с уменьшением экспрессии гена Р4НА2 и/или уменьшением количества белка пролил 4-гидрокислаза альфа 2, заключающийся в трансфекции одной или несколькими генетическими конструкциями по п.1, путем введения в органы и ткани пациента аутологичных клеток пациента, трансфицированных одной или несколькими генетическими конструкциями по п.1, и/или во введении в органы и ткани пациента одной или нескольких генетических конструкций по п.1, или в сочетании обозначенных способов.
6. Способ применения генетической конструкции по п.5, заключающийся в том, что выбор генетической конструкции из созданных по п.1 генетических конструкций производят индивидуально для каждого пациента после предварительно выполненного определения наиболее эффективной генетической конструкции для данного пациента.
| US 7759090 B2, 20.07.2010 | |||
| US 20160075747 A1, 17.03.2016 | |||
| WO 2003046164 A1, 05.06.2003 | |||
| WAGNER K | |||
| ET AL | |||
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |
| RU 2008123706 A, 27.12.2009. | |||

