Настоящая заявка подана 20 августа 2013 г. в виде международной патентной заявки согласно РСТ и заявляет приоритет по заявке на патент США с серийным номером 61/691117, поданной 20 августа 2012 г., раскрытие которой включено в данный документ с помощью ссылки во всей ее полноте.
Область изобретения
Настоящее изобретение относится к области биомедицины и, конкретно, к способам, пригодным для терапии рака. В частности, варианты осуществления настоящего изобретения, относятся к способам и композициям для проведения клеточной иммунотерапии, включающей Т-клетки, модифицированные рецепторами, нацеливающимися на опухоль.
Положение касательно исследования, финансируемого из государственного бюджета
Настоящее изобретение было создано при государственной поддержке по грантам СА 136551 и СА 114536, присужденным Национальными институтами здравоохранения. Правительство имеет определенные права на настоящее изобретение.
Предпосылки изобретения
Адоптивный перенос Т-лимфоцитов человека, сконструированных с помощью переноса генов для экспрессии химерных антигенных рецепторов (химерных рецепторов), специфичных к поверхностным молекулам, экспрессируемым на опухолевых клетках, имеет потенциал в эффективном лечении запущенных злокачественных опухолей. Химерные рецепторы представляют собой синтетические рецепторы, которые включают внеклеточный лиганд-связывающий домен, чаще всего одноцепочечный вариабельный фрагмент моноклонального антитела (scFv), соединенный с внутриклеточными сигнальными компонентами, чаще всего CD3ζ в отдельности или в комбинации с одним или несколькими костимулирующими доменами. Большая часть исследования по разработке химерного рецептора была сосредоточена на определении scFv и других лиганд-связывающих элементов, которые нацеливаются на опухолевые клетки, не вызывая серьезную токсичность в отношении важнейших нормальных тканей, и на определении оптимального состава внутриклеточных сигнальных модулей для активации эффекторных функций Т-клеток. Однако, неясно, будут ли разновидности конструкции химерного рецептора, которые опосредуют более эффективное функционирование in vitro, воспроизводимо преобразовываться в улучшенную терапевтическую активность in vivo при применении в клинической практике Т-клеток, модифицированных химерным рецептором.
Имеется необходимость в нахождении способов определения элементов конструкции химических рецепторов, которые важны для терапевтической активности и клеточных популяций, с целью генетической модификации и адоптивного переноса, которые обеспечивают повышенную выживаемость и эффективность in vivo.
Краткое описание изобретения
В одном аспекте настоящее раскрытие относится к способам и композициям для обеспечения и/или усиления иммунных ответов, опосредованных клеточной иммунотерапией, как, например, посредством адоптивного переноса опухолеспецифических генетически модифицированных субпопуляций Т-клеток CD8+ или CD4+ отдельно или в комбинации. Настоящее раскрытие предусматривает нуклеиновые кислоты, кодирующие химерный рецептор, а также векторы и клетки-хозяева, содержащие такие нуклеиновые кислоты. Последовательность нуклеиновых кислот, которая кодирует химерный рецептор, связывает вместе ряд модульных компонентов, которые можно вырезать и заменить другими компонентами для создания индивидуального химерного рецептора для эффективной активации Т-клеток и распознавания специфичной молекулы-мишени или эпитопа на молекуле-мишени.
В вариантах осуществления нуклеиновая кислота, кодирующая химерный рецептор, содержит полинуклеотид, кодирующий лиганд-связывающий домен, где лигандом является молекула, экспрессируемая на опухолевых или инфицированных клетках, полинуклеотид, кодирующий полипептидный спейсер, при этом полипептидный спейсер содержит приблизительно 200 аминокислот или менее, полинуклеотид, кодирующий трансмембранный домен, и полинуклеотид, кодирующий внутриклеточные сигнальные домены. В вариантах осуществления полипептидный спейсер включает модифицированный шарнирный участок IgG4, содержащий аминокислотную последовательность Х1РРХ2Р, которая может быть связана с другими аминокислотными последовательностями, включающими, без ограничения, последовательности СН2 и СН3 или только СН3 Fc Ig. Было обнаружено, что длина спейсерного участка, который предположительно не имеет способности к передаче сигнала, влияет на in vivo эффективность Т-клеток, модифицированных для экспрессии химерного рецептора, и она должна быть индивидуальной для отдельных молекул-мишеней для обеспечения оптимального распознавания опухолевой клетки или клетки-мишени.
В другом аспекте настоящего раскрытия предлагается выделенная нуклеиновая кислота, кодирующая химерный рецептор, содержащая: полинуклеотид, кодирующий лиганд-связывающий домен, где лигандом является опухолеспецифический антиген, вирусный антиген или любая другая молекула, экспрессируемая в популяции клеток-мишеней, пригодная для опосредования распознавания и удаления лимфоцитом; полинуклеотид, кодирующий полипептидный спейсер, где полипептидный спейсер имеет индивидуальную длину, которая специфична для каждого лиганда-мишени, где спейсер обеспечивает повышенные пролиферацию Т-клеток и/или выработку цитокинов по сравнению с эталонным химерным рецептором; полипептид, кодирующий трансмембранный домен; и полипептид, кодирующий один или несколько внутриклеточных сигнальных доменов. В вариантах осуществления используется длинный спейсер, если эпитоп на лиганде-мишени находится в мембранно-проксимальном положении, и используется короткий спейсер, если эпитоп на лиганде-мишени находится в мембранно-дистальном положении. Настоящее раскрытие включает векторы экспрессии и клетки-хозяева, содержащие выделенный химерный рецептор, описанный в данном документе.
В другом аспекте настоящего раскрытия предлагается полипептидный химерный рецептор, содержащий лиганд-связывающий домен, где лигандом является опухолеспецифический антиген, вирусный антиген или любая другая молекула, которая экспрессируется в популяции клеток-мишеней и на которую можно нацеливаться для опосредования распознавания и удаления лимфоцитами; полипептидный спейсер, где полипептидный спейсер содержит приблизительно 20-229 аминокислот; трансмембранный домен и один или несколько внутриклеточных сигнальных доменов. В вариантах осуществления полипептидный спейсер содержит модифицированный шарнирный участок IgG, содержащий аминокислотную последовательность Х1РPХ2Р.
В другом аспекте настоящее раскрытие предлагает композиции для обеспечения и/или усиления иммунных ответов, опосредованных клеточной иммунотерапией, как, например, посредством адоптивного переноса опухолеспецифических генетически модифицированных Т-клеток CD4+ с субпопуляционной специфичностью, где Т-клетки CD4+ обеспечивают и/или усиливают способность Т-клеток CD8+ к поддержанию противоопухолевой реактивности и увеличению и/или максимизации пролиферации опухолеспецифических клеток. В вариантах осуществления клетки CD4+ генетически модифицированы для экспрессии нуклеиновой кислоты, кодирующей химерный рецептор, и/или полипептидного химерного рецептора, описанных в данном документе.
В другом аспекте настоящее раскрытие предлагает композиции для обеспечения и/или усиления иммунных ответов, опосредованных клеточной иммунотерапией, как, например, посредством адоптивного переноса опухолеспецифических генетически модифицированных Т-клеток CD8+ с субпопуляционной специфичностью. В вариантах осуществления клетки CD8+ экспрессируют нуклеиновую кислоту, кодирующую химерный рецептор, и/или полипептидный химерный рецептор, описанные в данном документе.
В другом варианте осуществления настоящее изобретение предлагает композицию для адоптивной клеточной иммунотерапии, содержащую клеточный препарат генетически модифицированных цитотоксических Т-лимфоцитов CD8+ для обеспечения и/или усиления иммунных ответов, где клеточный препарат цитотоксических Т-лимфоцитов содержит Т-клетки CD8+, которые экспрессируют химерный рецептор, содержащий лиганд-связывающий домен для лиганда, ассоциированного с заболеванием или нарушением, индивидуальный спейсерный участок, трансмембранный домен и внутриклеточный сигнальный домен Т-клеточного или другого рецептора, такой как костимулирующий домен, и/или клеточный препарат генетически модифицированных Т-лимфоцитов-хелперов, где клеточный препарат Т-лимфоцитов-хелперов имеет клетки CD4+, которые экспрессируют химерный рецептор, содержащий вариабельный домен антитела, специфичный к лиганду, ассоциированному с заболеванием или нарушением, индивидуальный спейсерный участок, трансмембранный домен и один или несколько внутриклеточных сигнальных доменов.
В одном варианте осуществления настоящее изобретение предлагает способ проведения клеточной иммунотерапии у субъекта, имеющего заболевание или нарушение, путем введения субъекту клеточного препарата генетически модифицированных цитотоксических Т-лимфоцитов, который обеспечивает клеточный иммунный ответ, где клеточный препарат цитотоксических Т-лимфоцитов содержит Т-клетки CD8+, которые имеют химерный рецептор, содержащий полинуклеотид, кодирующий лиганд-связывающий домен, где лигандом является опухолеспецифический антиген, вирусный антиген или любая другая молекула, экспрессируемая в популяции клеток-мишеней, пригодная для опосредования распознавания и удаления лимфоцитом; полинуклеотид, кодирующий полипептидный спейсер, где полипептидный спейсер имеет индивидуальную длину, которая специфична для каждого лиганда-мишени, где спейсер обеспечивает повышенные пролиферацию Т-клеток и/или выработку цитокинов по сравнению с эталонным химерным рецептором; полинуклеотид, кодирующий трансмембранный домен; и полинуклеотид, кодирующий один или несколько внутриклеточных сигнальных доменов. В варианте осуществления лиганд-связывающим доменом является внеклеточный вариабельный домен антитела, специфичный к лиганду, ассоциированному с заболеванием или нарушением. Вариант осуществления включает клеточный препарат генетически модифицированных Т-лимфоцитов-хелперов, где клеточный препарат Т-лимфоцитов-хелперов содержит Т-клетки CD4+, которые имеют химерный рецептор, содержащий полинуклеотид, кодирующий лиганд-связывающий домен, где лигандом является опухолеспецифический антиген, вирусный антиген или любая другая молекула, экспрессируемая в популяции клеток-мишеней, пригодная для опосредования распознавания и удаления лимфоцитом; полинуклеотид, кодирующий полипептидный спейсер, где полипептидный спейсер имеет индивидуальную длину, которая специфична для каждого лиганда-мишени, где спейсер обеспечивает повышенные пролиферацию Т-клеток и/или выработку цитокинов по сравнению с эталонным химерным рецептором; полинуклеотид, кодирующий трансмембранный домен; и полинуклеотид, кодирующий один или несколько внутриклеточных сигнальных доменов. В вариантах осуществления популяции генетически модифицированных клеток CD8+ и генетически модифицированных клеток CD4+ вводят совместно. В вариантах осуществления Т-клетки являются аутологическими или аллогенными Т-клетками.
Возможны различные модификации вышеупомянутого способа. Например, химерный рецептор, который экспрессируется Т-клеткой CD4+ и Т-клеткой CD8+, может быть одним и тем же или разным.
В другом аспекте настоящее изобретение предлагает способ производства композиции для адоптивной иммунотерапии посредством получения модифицированного химерным рецептором клеточного препарата опухолеспецифических цитотоксических Т-лимфоцитов CD8+, который вызывает клеточный иммунный ответ и экспрессирует антиген-реактивный химерный рецептор, где модифицированный клеточный препарат цитотоксических Т-лимфоцитов содержит Т-клетки CD8+, которые имеют химерный рецептор, содержащий лиганд-связывающий домен, где лигандом является опухолеспецифический антиген, вирусный антиген или любая другая молекула, экспрессируемая в популяции клеток-мишеней, пригодная для опосредования распознавания и удаления лимфоцитом; полипептидный спейсер, где полипептидный спейсер имеет индивидуальную длину, которая специфична для каждого лиганда-мишени, где спейсер обеспечивает повышенные пролиферацию Т-клеток и/или выработку цитокинов по сравнению с эталонным химерным рецептором; трансмембранный домен и один или несколько внутриклеточных сигнальных доменов; и/или получения модифицированной наивной клетки или клетки памяти Т-хелпера CD4+, где модифицированный клеточный препарат Т-лимфоцитов-хелперов содержит клетки CD4+, которые имеют химерный рецептор, содержащий лиганд-связывающий домен, где лигандом является опухолеспецифический антиген, вирусный антиген или любая другая молекула, экспрессируемая в популяции клеток-мишеней, пригодная для опосредования распознавания и удаления лимфоцитом; полипептидный спейсер, где полипептидный спейсер имеет индивидуальную длину, которая специфична для каждого лиганда-мишени, где спейсер обеспечивает повышенную пролиферацию Т-клеток или выработку цитокинов по сравнению с эталонным химерным рецептором; трансмембранный домен и один или несколько внутриклеточных сигнальных доменов.
Эти и другие варианты осуществления настоящего изобретения описаны также в сопутствующих описании, фигурах и формуле изобретения.
Краткое описание графических материалов
Фигура 1. Библиотека спейсерных последовательностей. Была создана плазмидная библиотека, которая содержит подвергнутые оптимизации кодонов последовательности ДНК, кодирующие внеклеточные компоненты, включая шарнирный участок IgG4 в отдельности, шарнирный участок IgG4, связанный с доменами СН2 и СH3, или шарнирный участок IgG4, связанный с доменом СН3. Любая последовательность scFv (VH и VL) может быть клонирована в направлении к 5' от последовательностей, кодирующих данную библиотеку вариабельных спейсерных доменов. Спейсерные домены, в свою очередь, связаны с трансмембранными и внутриклеточными сигнальными доменами CD28 и с CD3-зета. Последовательность Т2А в векторе отделяет химерный рецептор от селективного маркера, кодирующего усеченный рецептор эпидермального фактора роста человека (tEGFR).
Фигура 2. In vitro цитотоксичность, выработка цитокинов и пролиферация Т-клеток, модифицированных для экспрессии химерных рецепторов ROR1 из 2А2 с модифицированной длиной спейсера. (А) Фенотип очищенных линий клеток CD8+, полученных из ТСМ, модифицированных каждым из химерных рецепторов ROR1 из 2А2 с длинным, средним или коротким спейсерным доменом. Окрашивание антителом к F(ab), которое связывается с эпитопом в scFv 2А2, показывает поверхностную экспрессию химерных рецепторов ROR1 с полноразмерным или усеченным спейсером. (В) Цитолитическая активность Т-клеток, экспрессирующих различные химерные рецепторы ROR1 из 2А2 с длинным ( ), средним (
), средним ( ) и коротким спейсером (
) и коротким спейсером (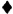 ), или с контрольным лентивирусным вектором, кодирующим tEGFR, относительно ROR1+ (х) и контрольных клеток-мишеней. В столбчатой диаграмме обобщены данные по цитотоксичности из 3 независимых экспериментов (Е : Т =30:1), нормализованные по цитолитической активности под влиянием химерного рецептора ROR1 из 2А2 'длинный' =1, и анализируемые с помощью t-критерия Стьюдента. (С) Разведение красителя CFSE применяли для измерения пролиферации Т-клеток с химерным рецептором ROR1 из 2А2 и контрольных Т-клеток с tEGFR спустя 72 часа после стимуляции клетками Raji/ROR1 (левая панель) и первичными клетками CLL (правая панель) без добавления экзогенных цитокинов. Для анализа объединяли данные по трем лункам и анализировали пролиферацию живых Т-клеток (PI-) CD8+. Числа над каждой гистограммой указывают число клеточных делений, которым подверглась пролиферирующая субпопуляция, и возле каждого графика представлена доля Т-клеток в каждом окне, которые подверглись ≥4/3/2/1 клеточным делениям. (D) Мультиплексный анализ цитокинов надосадочных жидкостей, полученных спустя 24 часа из трех кокультур 5×104 Т-клеток, экспрессирующих различные химерные рецепторы ROR1 из 2А2, с клетками Raji/ROR1 и первичными клетками CLL. Данные мультиплексного анализа цитокинов из трех независимых экспериментов нормализовали (высвобождение цитокинов под влиянием химерного рецептора ROR1 'длинный' = 1) и анализировали с помощью t-критерия Стьюдента (столбчатая диаграмма справа).
), или с контрольным лентивирусным вектором, кодирующим tEGFR, относительно ROR1+ (х) и контрольных клеток-мишеней. В столбчатой диаграмме обобщены данные по цитотоксичности из 3 независимых экспериментов (Е : Т =30:1), нормализованные по цитолитической активности под влиянием химерного рецептора ROR1 из 2А2 'длинный' =1, и анализируемые с помощью t-критерия Стьюдента. (С) Разведение красителя CFSE применяли для измерения пролиферации Т-клеток с химерным рецептором ROR1 из 2А2 и контрольных Т-клеток с tEGFR спустя 72 часа после стимуляции клетками Raji/ROR1 (левая панель) и первичными клетками CLL (правая панель) без добавления экзогенных цитокинов. Для анализа объединяли данные по трем лункам и анализировали пролиферацию живых Т-клеток (PI-) CD8+. Числа над каждой гистограммой указывают число клеточных делений, которым подверглась пролиферирующая субпопуляция, и возле каждого графика представлена доля Т-клеток в каждом окне, которые подверглись ≥4/3/2/1 клеточным делениям. (D) Мультиплексный анализ цитокинов надосадочных жидкостей, полученных спустя 24 часа из трех кокультур 5×104 Т-клеток, экспрессирующих различные химерные рецепторы ROR1 из 2А2, с клетками Raji/ROR1 и первичными клетками CLL. Данные мультиплексного анализа цитокинов из трех независимых экспериментов нормализовали (высвобождение цитокинов под влиянием химерного рецептора ROR1 'длинный' = 1) и анализировали с помощью t-критерия Стьюдента (столбчатая диаграмма справа).
Фигура 3. Химерному рецептору R11 требуется длинный спейсер для распознавания опухолевых клеток ROR1+. Последовательности, кодирующие scFv из моноклонального антитела R11, которое специфично к эпитопу в мембранно-проксимальном крингл-домене орфанного тирозинкиназного рецептора ROR1, клонировали выше последовательностей шарнирного участка IgG4 в отдельности (короткий спейсер), шарнирного участка IgG4/CH3 (средний спейсер) и шарнирного участка IgG4/CH2/CH3 в библиотеке химерных рецепторов, содержащих костимулирующие домены 4-1ВВ, и получали в виде лентивирусных векторов. А). Т-клетки CD8+ человека трансдуцировали, и эффективность трансдукции с каждым из химерных рецепторов с коротким, средним и длинным спейсерами определяли с помощью окрашивания на маркер tEGFR. В). Трансдуцированные Т-клетки, экспрессирующие рецепторы с коротким (вверху), средним (посредине) и длинным (внизу) спейсерами, оценивали в отношении лизиса лейкозных клеток K562 самих по себе или трансфицированных для экспрессии ROR1. Только Т-клетки, экспрессирующие химерный рецептор с длинным спейсером, эффективно уничтожали клетки K562 ROR1+. С). Трансдуцированные Т-клетки, экспрессирующие рецепторы с коротким (вверху), средним (посредине) и длинным (внизу) спейсерами, метили CFSE, стимулировали клетками K562, экспрессирующими ROR1 или CD19 (контроль), и анализировали в отношении пролиферации клеток в течение 72 часов. Происходила специфическая пролиферация Т-клеток, экспрессирующих химерный рецептор с длинным спейсером, в ответ на воздействие клеток K562 ROR1+. D). Трансдуцированные Т-клетки, экспрессирующие рецепторы с коротким (вверху), средним (посредине) и длинным (внизу) спейсерами, стимулировали клетками лимфомы Raji и клетками K562, которые экспрессировали ROR1 или CD19 (контроль), и анализировали в отношении секреции интерферона гамма в надосадочной жидкости в течение 24 часов. Т-клетки, экспрессирующие химерный рецептор с длинным спейсером, пролиферировали и вырабатывали наиболее высокие уровни интерферона гамма в ответ на воздействие ROR1-положительных клеток-мишеней.
Фигура 4. Конструкция химерных рецепторов ROR1 с модифицированной длиной спейсера, полученных из scFv 2А2 и R12 с различной аффинностью.
(А) Конструкция трансгенных вставок в лентивирусный вектор, кодирующих панель химерных рецепторов ROR1, содержащих scFv 2А2, полученный из Fc IgG4 спейсер 'шарнирный участок-СН2-СН3' (длинный спейсер, 229 аминокислот), 'шарнирный участок-СТ13' (средний, 119 аминокислот) или 'шарнирный участок' в отдельности (короткий, 12 аминокислот), а также сигнальный модуль с CD3ζ и CD28. Кассета каждого химерного рецептора содержит усеченный маркер EGFR, расположенный ниже элемента Т2А. (В) Трансгенные вставки в лентивирусный вектор, кодирующие ROR1-специфические химерные рецепторы, полученные из scFv R12 и 2А2, с коротким 'шарнирным участком' Fc IgG4 (12 аминокислот) и сигнальным модулем, содержащим CD28 или 4-1ВВ и CD3ζ, соответственно (всего 4 конструкта).
Фигура 5. Противоопухолевая реактивность Т-клеток, модифицированных химерными рецепторами ROR1, полученными из mAb R12 с более высокой аффинностью, чем 2А2. (А) Экспрессия tEGFR в очищенных поликлональных линиях Т-клеток CD8+, полученных из ТСМ, модифицированных каждым из химерных рецепторов ROR1 из R12 и 2А2 с коротким спейсером 'шарнирный участок' Fc IgG4 и костимулирующим доменом CD28 или 4-1ВВ. (В) Цитотоксичность в отношении ROR1+ и контрольных клеток-мишеней со стороны Т-клеток, экспрессирующих химерные рецепторы RO1 R12 (28-; 4-1ВВ-Δ) и 2А2 (28-; 4-1ВВ-°) или контрольного вектора, кодирующего tEGFR (х). (С) Мультиплексный анализ цитокинов надосадочных жидкостей, полученных спустя 24 часа из кокультур 5×104 Т-клеток, экспрессирующих различные химерные рецепторы ROR1, с опухолевыми клетками Raji/ROR1. Средняя/правая столбчатые диаграммы показывают нормализованные данные мультиплексного анализа из трех независимых экспериментов (высвобождение цитокинов под влиянием химерного рецептора ROR1 из 2А2=1), анализируемые с помощью t-критерия Стьюдента. (D) Пролиферацию Т-клеток с химерным рецептором ROR1 и контрольных Т-клеток с tEGFR спустя 72 часа после стимуляции клетками Raji/ROR1 и без добавления экзогенных цитокинов оценивали с помощью разведения красителя CFSE. Числа над каждой гистограммой указывают число клеточных делений, которым подверглась пролиферирующая субпопуляция, и над каждым графиком представлена доля Т-клеток в каждом окне, которые подверглись ≥4/3/2/1 клеточным делениям.
Фигура 6. Анализ выработки цитокинов и пролиферации линий Т-клеток CD4+, модифицированных химерным рецептором ROR1, полученным из mAb R12 с более высокой аффинностью, чем 2А2. (А-В) Химерные рецепторы ROR1 из 2А2 и R12 имели короткий спейсер и костимулирующий домен CD28. (А) Мультиплексный анализ цитокинов из надосадочных жидкостей, полученных спустя 24 часа после стимуляции 5×104 Т-клеток CD4+, экспрессирующих химерный рецептор ROR1 из 2А2 и R12, опухолевыми клетками Raji/ROR1. (В) Пролиферацию Т-клеток с химерными рецепторами ROR1 из R12 и 2А2 и контрольных Т-клеток с tEGFR спустя 72 часа после стимуляции клетками Raji/ROR1 и без добавления экзогенных цитокинов оценивали с помощью разведения красителя CFSE. Числа над каждой гистограммой указывают число клеточных делений, которым подверглась пролиферирующая субпопуляция, и над гистограммами представлена доля Т-клеток в каждом окне, которые подверглись ≥5/4/3/2/1 клеточным делениям.
Фигура 7. Распознавание первичных клеток CLL Т-клетками, модифицированными химерными рецепторами ROR1 из 2А2 и R12 с оптимальным коротким спейсером и костимулирующим доменом 4-1ВВ или СD19-специфическим химерным рецептором. (А) Экспрессия ROR1/CD19 на первичных клетках CLL и CD80/86 на первичных клетках CLL и опухолевых клетках Raji/ROR1 (точечные графики черного цвета), которые могут взаимодействовать с CD28 на Т-клетках с химерными рецепторами (гистограммы белого цвета). Окрашивание изотипически сходными контрольными mAb показано в виде точечных графиков/гистограмм серого цвета. (В) Цитолитическая активность Т-клеток, экспрессирующих химерный рецептор ROR1 2А2 () и R12 ( ), СD19-специфический химерный рецептор (А), и Т-клеток, модифицированных контрольным вектором, коддирующим tEGFR (х), в отношении первичных клеток CLL (левая диаграмма) и нормальных В-клеток (правая диаграмма), анализируемая с помощью теста с радиоактивным хромом. Данные о цитотоксичности в отношении первичных клеток CLL из четырех независимых экспериментов (Е : Т = 30:1) нормализовали (цитолитическая активность под влиянием химерного рецептора ROR1 из 2А2=1) и анализировали с помощью t-критерия Стьюдента (столбчатая диаграмма). (С) Мультиплексный анализ цитокинов после 24-часовой стимуляции 5×104 Т-клеток с химерным рецептором первичными клетками CLL. Высвобождение цитокинов нестимулированными Т-клетками с химерными рецепторами составляло менее 3,6 пг/мл (предел обнаружения) (левая столбчатая диаграмма). ELISA для оценки выработки TFN-γ 5×104 Т-клетками с химерными рецепторами ROR1 из 2А2 и R12 после 24-часового совместного культивирования с первичными клетками CLL. O.D., составляющая 1, соответствует примерно 250 пг/мл (правая столбчатая диаграмма). (D) Пролиферация Т-клеток CD8+, модифицированных химерными рецепторами ROR1 из 2А2, ROR1 из R12 и CD19, спустя 72 часа после стимуляции первичными клетками CLL. Числа над каждой гистограммой указывают число клеточных делений, и возле каждого графика представлена доля Т-клеток в каждом окне, которые подверглись ≥3/2/1 клеточным делениям.
), СD19-специфический химерный рецептор (А), и Т-клеток, модифицированных контрольным вектором, коддирующим tEGFR (х), в отношении первичных клеток CLL (левая диаграмма) и нормальных В-клеток (правая диаграмма), анализируемая с помощью теста с радиоактивным хромом. Данные о цитотоксичности в отношении первичных клеток CLL из четырех независимых экспериментов (Е : Т = 30:1) нормализовали (цитолитическая активность под влиянием химерного рецептора ROR1 из 2А2=1) и анализировали с помощью t-критерия Стьюдента (столбчатая диаграмма). (С) Мультиплексный анализ цитокинов после 24-часовой стимуляции 5×104 Т-клеток с химерным рецептором первичными клетками CLL. Высвобождение цитокинов нестимулированными Т-клетками с химерными рецепторами составляло менее 3,6 пг/мл (предел обнаружения) (левая столбчатая диаграмма). ELISA для оценки выработки TFN-γ 5×104 Т-клетками с химерными рецепторами ROR1 из 2А2 и R12 после 24-часового совместного культивирования с первичными клетками CLL. O.D., составляющая 1, соответствует примерно 250 пг/мл (правая столбчатая диаграмма). (D) Пролиферация Т-клеток CD8+, модифицированных химерными рецепторами ROR1 из 2А2, ROR1 из R12 и CD19, спустя 72 часа после стимуляции первичными клетками CLL. Числа над каждой гистограммой указывают число клеточных делений, и возле каждого графика представлена доля Т-клеток в каждом окне, которые подверглись ≥3/2/1 клеточным делениям.
Фигура 8. Действие Т-клеток CD8+, модифицированных химерным рецептором ROR1 и химерным рецептором CD19, в отношении первичных клеток CLL усиливается Т-клетками-хелперами CD4+, модифицированными химерными рецепторами. (A) ELISA для оценки выработки IL-2 в трех кокультурах 5×104 Т-клеток CD8+ и CD4+, экспрессирующих химерный рецептор ROR1 из R12 и CD19, соответственно, инкубированных с первичными клетками CLL в течение 24 часов. O.D., составляющая 1, соответствует примерно 800 пг/мл. (В) Пролиферация Т-клеток CD8+, модифицированных химерными рецепторами, в ответ на воздействие первичных клеток CLL усиливается добавлением Т-клеток CD4+, модифицированных химерными рецепторами. Меченные CFSE Т-клетки CD8+, экспрессирующие химерные рецепторы ROR1 из 2А2, ROR1 из R12 и CD19, соответственно, подвергали совместному культивированию с опухолевыми клетками и с трансдуцированными химерными рецепторами ROR1 из 2А2, ROR1 из R12 и CD19 или контрольными нетрансдуцированными Т-клетками CD4+ (CD8+ : CD4+=1:1). Пролиферацию субпопуляции CD8+ анализировали спустя 72 часа после стимуляции. Числа над каждой гистограммой указывают число клеточных делений, и над каждым графиком представлена доля Т-клеток в каждом окне, которые подверглись ≥3/2/1 клеточным делениям.
Фигура 9. In vivo противоопухолевая эффективность Т-клеток с химерными рецепторами ROR1 из 2А2, ROR1 из R12 и CD19. Группам мышей инокулировали 0,5×106 MCL JeKo-1/ffluc посредством инъекции в хвостовую вену, и через 7 дней после инокуляции опухоли вводили 5×106 Т-клеток с химерными рецепторами ROR1 из 2А2, ROR1 из R12 или CD19 или Т-клетки, экспрессирующие контрольный вектор tEGFR. Все конструкты химерных рецепторов имели короткий спейсер 'шарнирный участок в отдельности' IgG4 и костимулирующий домен 4-1ВВ. (А, В) Серийная биолюминесцентная визуализация опухоли в группах мышей, которых обрабатывали Т-клетками, экспрессирующими химерный рецептор ROR1 из 2А2 (▼), химерный рецептор ROR1 из R12 с высокой аффинностью (■), СD19-специфический химерный рецептор (▲), мышей, которых обрабатывали Т-клетками, трансдуцированными только tEGFR (•), и необработанных мышей. Биолюминесцентная визуализация показала манифестацию опухоли в костном мозге и грудной клетке, и поэтому интенсивность сигнала измеряли в исследуемых областях, которые охватывали все тело и грудную клетку каждой отдельно взятой мыши. (С) Анализ выживаемости по Каплану-Мейеру в отдельных группах обработки и контрольных группах. Статистические анализы выполняли с помощью логарифмического рангового критерия. Данные, показанные в А-С, представляют результаты, полученные в двух независимых экспериментах. (D) In vivo пролиферация Т-клеток с химерными рецепторами ROR1 из 2А2, ROR1 из R12 и CD19. Мыши-опухоленосители NSG/JeKo-1 получали однократную дозу 5×106 меченных CFSE Т-клеток с химерными рецепторами ROR1 из 2А2, ROR1 из R12 и CD19 на 7-й день после инокуляции опухоли, а спустя 72 часа у каждой отдельно взятой мыши забирали образцы периферической крови, костного мозга и селезенки. Анализировали частоту и пролиферацию живых (PI-) Т-клеток CD45+ CD8+ tEGFR+. Частота Т- клеток с химерными рецепторами ROR1 из 2А2, ROR1 из R12 и CD19 представлена слева на каждой гистограмме в виде процента живых клеток, и над каждым графиком представлена доля Т-клеток, которая подверглась ≥4/3/2/1 клеточным делениям.
Фигура 10. Экспрессия лигандов ROR1 и NKG2D в линиях клеток эпителиального рака. (А) Экспрессия ROR1 в линиях клеток трижды негативного рака молочной железы MDA-MB-231 и 468 и линиях клеток почечно-клеточного рака FARP, TREP и RWL (гистограммы черного цвета). Окрашивание изотипически сходным контрольным антителом показано в виде гистограмм серого цвета. (В) Экспрессия лигандов MICA/B для CD80/86 и NKG2D на опухолевых клетках MDA-MB-231 и Raji/ROR1, a также NKG2D (CD314) на Т-клетках с химерными рецепторами ROR1 из 2А2 и R12. Окрашивание изотипически сходными контрольными mAb показано в виде точечных графиков/гистограмм серого цвета.
Фигура 11. Модифицированные химерным рецептором ROR1 Т-клетки распознают клетки эпителиальных опухолей ROR1+ in vitro. (А) Тест с радиоактивным хромом для оценки цитолитической активности модифицированных химерным рецептором ROR1 из R12 Т-клеток (короткий спейсер/костимулирующий домен 4-1ВВ, заштрихованные обозначения) и контрольных клеток с tEGFR (незаштрихованные обозначения) по отношению к линиям клеток рака молочной железы и почечно-клеточного рака ROR1+. (A-D) Химерные рецепторы ROR1 из 2А2 и R12 имели оптимальный короткий спейсер и костимулирующий домен 4-1ВВ. (В) Мультиплексный анализ цитокинов после стимуляции Т-клеток, экспрессирующих химерные рецепторы ROR1 из 2А2 и R12, опухолевыми клетками MDA-MB-231 и Raji/ROR1. (С) Пролиферация Т-клеток CD8+, модифицированных химерными рецепторами ROR1 из 2А2 и R12, спустя 72 часа после стимуляции опухолевыми клетками MDA-MB-231. Для анализа объединяли данные по трем лункам и анализировали пролиферацию живых (PI-) Т-клеток CD8+. Числа над каждой гистограммой указывают число клеточных делений, которым подверглась пролиферирующая субпопуляция, и возле каждой гистограммы представлена доля Т-клеток в каждом окне, которые подверглись ≥4/3/2/1 клеточным делениям. (D) ELISA для оценки выработки IL-2 Т-клетками с химерным рецептором ROR1 из R12 после 24-часового совместного культивирования с MDA-MB-231 в среде без испытуемых компонентов и после добавления коктейля из антител, блокирующего путь NKG2D [антитела к NKG2D (клон 1D11), антитела к MICA/B (клон 6D4) и антитела к ULBP], или изотипически сходных контрольных mAb. O.D., составляющая 0,6, соответствует примерно 1900 пг/мл.
Фигура 12. Влияние длины внеклеточного спейсера на распознавание и запуск лизиса опухолевых клеток Т-клетками CD8+ человека, которые экспрессируют Неr2-специфический химерный рецептор. А.) Обозначение расположения эпитопа к Fab герцептина как мембранно-проксимального эпитопа опухолевых клеток в Неr2 человека. В.) Структурные форматы вариантов длины спейсера CAR scFv герцептина в виде полипептидов, связанных через -Т2А- с карбокси-концом маркерного трансмембранного белка EGFRt. С.) Выявление с помощью вестерн-блоттинга экспрессии варианта CAR герцептина с коротким, средним и длинным спейсером в CTL CD8+ человека. D.) Выявление с помощью проточной цитометрии EGFRt с помощью трансдуцированных CTL CD8+, трансдуцированных вариантами. CAR герцептина, а затем подвергнутых иммуномагнитной очистке герцептином, конъюгированным с биотином, и микрогранулами с антителами к биотину. Е.) Различное цитолитическое действие Т-клеток, трансдуцированных для экспрессии вариантов CAR герцептина (короткий - S; средний - М и длинный - L), в отношении линий клеток медуллобластомы человека Med411 FH и D283 Her2+ (D341 является контрольной линией клеток медуллобластомы Неr2-, потоковые графики на вставках представляют опухолевые линии-мишени, окрашенные mAb, специфичным к Неr2). Зеленый цвет = полноразмерный IgG4 (длинный спейсер;  ), синий цвет = шарнирный участок IgG4:CH3 (средний спейсер;
), синий цвет = шарнирный участок IgG4:CH3 (средний спейсер;  ), красный цвет = шарнирный участок IgG4 в отдельности (короткий спейсер;
), красный цвет = шарнирный участок IgG4 в отдельности (короткий спейсер;  ).
).
Фигура 13. Векторы экспрессии химерного рецептора CD19 и образование Т-клеток с химерным рецептором CD19.
(А) Конструкция трансгенных вставок в лентивирусный вектор, кодирующих панель СD19-специфических химерных рецепторов, которые отличаются длиной внеклеточного спейсера и внутриклеточным костимулирующим доменом. Каждый химерный рецептор содержит CD19-специфический одноцепочечный вариабельный фрагмент, полученный из mAb FMC63 в ориентации VL-VH, полученный из IgG4 спейсерный домен «шарнирный участок-СН2-СН3» (длинный спейсер, 229 аминокислот) или «шарнирный участок в отдельности» (короткий спейсер, 12 аминокислот) и сигнальный модуль, содержащий CD3ζ с CD28 или 4-1ВВ в отдельности или в тандеме. Кассета каждого химерного рецептора содержит усеченный маркер EGFR, расположенный ниже расщепляемого элемента 2А. (В) Поликлональные линии Т-клеток, модифицированные каждым из конструктов химерного рецептора CD19, были получены из очищенных центральных Т-клеток памяти CD8+ CD45RO+ CD62L+ (ТCM) нормальных доноров. После транедукции лентивирусом трансген-положительные Т-клетки в каждой линии клеток очищали с помощью маркера tEGFR и размножали для экспериментов in vitro и in vivo. (С) MFI после окрашивания на маркер tEGFR показал одинаковую экспрессию трансгена в Т-клетках, модифицированных каждым из химерных рецепторов CD19.
Фигура 14. In vitro цитотоксичность, выработка цитокинов и пролиферация Т-клеток, модифицированных различными химерными рецепторами CD19.
(А) Цитолитическая активность Т-клеток, экспрессирующих различные химерные рецепторы CD19, в отношении CD19+ и контрольных клеток-мишеней. (В) Мультиплексный анализ цитокинов надосадочных жидкостей, полученных спустя 24 часа из трех кокультур Т-клеток, экспрессирующих различные химерные рецепторы CD19, и клеток K562, трансфицированных с помощью CD19, а также клеток Raji CD19+. (С) Сравнение выработки цитокинов Т-клетками, экспрессирующими различные химерные рецепторы CD19. Данные мультиплексного анализа цитокинов из шести независимых экспериментов нормализовали (высвобождение цитокинов CTL с химерным рецептором CD19 'короткий/СD28'=1) и анализировали с помощью t-критерия Стьюдента. (D) Разведение красителя CFSE применяли для измерения пролиферации Т-клеток с химерным рецептором CD19 спустя 72 часа после стимуляции клетками K562/CD19 (верхняя панель) и опухолевыми клетками Raji CD19+ (нижняя панель) без добавления экзогенных цитокинов. Для анализа объединяли данные по трем лункам и анализировали пролиферацию живых (PI-) Т-клеток CD8+. Числа над каждой гистограммой указывают число клеточных делений, которым подверглась пролиферирующая субпопуляция, и в левом верхнем углу каждого графика представлена доля Т-клеток в каждом окне, которые подверглись ≥4/3/2/1 клеточным делениям. (Е) Окрашивание PI выполняли в конце 72-часового совместного культивирования Т-клеток, экспрессирующих различные химерные рецепторы CD19, с опухолевыми клетками Raji. На каждой гистограмме представлен процент клеток PI+ в линии Т-клеток (CD3+) с химерными рецепторами.
Фигура 15. Т-клетки с химерным рецептором CD19 с коротким внеклеточным спейсерным доменом уничтожают опухоли из Raji у мышей NOD/SCID. (А) Группам мышей инокулировали Raji-ffluc посредством инъекции в хвостовую вену, и Т-клетки, трансдуцированные химерными рецепторами CD19, содержащими длинные и короткие спейсерные домены, или только tEGFR, вводили через 2 дня и 9 дней после инокуляции опухоли путем инъекции в хвостовую вену. Прогрессирование и распространение опухоли оценивали с помощью серийной биолюминесцентной визуализации после инъекции субстрата люциферина. (В) Серийная биолюминесцентная визуализация опухоли в группах мышей, обработанных Т-клетками, экспрессирующими химерные рецепторы CD19 с коротким спейсерным ('короткий/СD28' и 'короткий/4-1ВВ') и длинным спейсерным ('длинный/СD28' и 'длинный/4-1ВВ') доменами, мышей, обработанных Т-клетками, трансдуцированными контрольным вектором, кодирующим tEGFR, или необработанных мышей. На каждой диаграмме, представляющей группы мышей, обработанных Т-клетками с химерным рецептором CD19 или Т-клетками, трансдуцированными tEGFR, также показано для сравнения среднее значение прогрессирования опухоли у необработанных мышей (треугольники красного цвета). (С) Анализ выживаемости по Каплану-Мейеру необработанных мышей и мышей, обработанных Т-клетками, экспрессирующими химерные рецепторы CD19 с коротким спейсерным ('короткий/СD28' и 'короткий/4-1ВВ') и длинным спейсерным ('длинный/СD28' и 'длинный/4-1ВВ') доменами, а также контрольным tEGFR. Статистические анализы выполняли с помощью логарифмического рангового критерия. Данные, показанные в В и С, представляют результаты, полученные в трех независимых экспериментах.
Фигура 16. Т-клетки с химерным рецептором CD19 с коротким спейсером (короткий/4-1ВВ) дозозависимым образом уничтожают развившиеся опухоли из Raji у мышей NSG. (А) Мышам инокулировали Raji-ffluc посредством инъекции в хвостовую вену и на 6-й день приживление опухоли подтверждали с помощью биолюминесцентной визуализации. На 7-й день мыши получили однократную внутривенную инъекцию различных доз Т-клеток, трансдуцированных химерным рецептором CD19 'короткий/4-1ВВ' или контрольным лентивирусом, кодирующим tEGFR. (В, С) Дозозависимая противоопухолевая эффективность Т-клеток, экспрессирующих химерный рецептор CD19 'короткий/4-1ВВ'. Контрольная группа мышей получала однократную высокую дозу Т-клеток, модифицированных только tEGFR. (D) Персистирование Т-клеток с химерным рецептором CD19 после адоптивного переноса мышам NSG/Raji. Анализ периферической крови (кровянистые выделения из глаз) по методу проточной цитометрии в группе мышей, обработанных 2,5×106 Т-клетками с химерным рецептором 'короткий/4-1ВВ'. Частота Т-клеток CD8+ tEGFR+ показана в виде процента живых клеток периферической крови.
Фигура 17. Т-клетки, экспрессирующие химерные рецепторы CD19 с коротким спейсером и CD28 или 4-1ВВ, более эффективны в отношении развившейся лимфомы, чем Т-клетки, экспрессирующие химерные рецепторы CD19 с длинным спейсером. (А) Мышам NSG на 0-й день инокулировали Raji-ffluc, а на 7-й день их обрабатывали одной дозой 2,5×106 Т-клеток, экспрессирующих химерный рецептор CD19 с коротким или длинным спейсером и костимулирующим доменом CD28 или 4-1ВВ. (В) Анализ выживаемости мышей по Каплану-Мейеру в каждой из групп обработки. Статистические анализы выполняли с помощью логарифмического рангового критерия. (С) Биолюминесцентная визуализация групп мышей, обработанных Т-клетками, экспрессирующими химерные рецепторы CD19 с короткими спейсерами ('короткий/СD28' и 'короткий/4-1ВВ') и длинными спейсерами ('длинный/СD28 и 'длинный/4-1ВВ'). Для сравнения на каждой диаграмме показана средняя опухолевая нагрузка у необработанных мышей в каждый момент времени (треугольники). (D) In vivo сохранение Т-клеток, экспрессирующих рецептор CD19 с коротким спейсерным доменом, улучшено по сравнению с Т-клетками, экспрессирующими химерные рецепторы CD19 с длинным спейсерным доменом. Частоту Т-клеток CD8+ tEGFR+ в периферической крови, полученной на 3-й и 10-й день после переноса, определяли посредством проточной цитометрии, и она показана в виде процента живых (PI-) клеток периферической крови. Статистические анализы выполняли с помощью t-критерия Стьюдента. Данные, показанные в B-D, представляют результаты, полученные в трех независимых экспериментах.
Фигура 18. Повышение дозы Т-клеток с химерным рецептором или усиление костимулирующих сигналов не повышает противоопухолевую эффективность химерных рецепторов CD19 с длинным спейсерным доменом по отношению к развившейся лимфоме. (А) Цитолитическая активность Т-клеток, экспрессирующих химерные рецепторы CD19 'длинный/СD28', 'длинный/4-1ВВ' и 'длинный/СD28 4-1ВВ', в отношении CD19+и контрольных клеток-мишеней. (В) Мультиплексный анализ цитокинов надосадочной жидкости, полученной спустя 24 часа из трех кокультур K562/CD19 и опухолевых клеток Raji с Т-клетками, экспрессирующими различные рецепторы CD19. (С) Оценка пролиферации Т-клеток с химерным рецепторов CD19 спустя 72 часа после стимуляции опухолевыми клетками CD19+ (K562/CD19 - левая панель; Raji - правая панель) с помощью разведения красителя CFSE. Для анализа объединяли данные по трем лункам и анализировали пролиферацию живых (PI-) Т-клеток CD8+. Числа над каждой гистограммой указывают число клеточных делений, которым подверглась пролиферирующая субпопуляция, и в левом верхнем углу каждого графика представлена доля Т-клеток в каждом окне, которые подверглись ≥4/3/2/1 клеточным делениям. (D) Анализ выживаемости по Каплану-Мейеру мышей, обработанных Т-клетками, экспрессирующими химерные рецепторы CD19 с коротким ('короткий/СD28' и 'короткий/4-1ВВ') и длинным ('длинный/СD28' и 'длинный/4-1ВВ') спейсерными доменами, или Т-клетками, модифицированными контрольным лентивирусным вектором, кодирующим tEGFR. Статистические анализы выполняли с помощью логарифмического рангового критерия. (Е) Биолюминесцентная визуализация групп мышей, обработанных Т-клетками, экспрессирующими химерные рецепторы CD19 с коротким спейсером ('короткий/СD28') и длинными спейсерами ('длинный/СD28' и 'длинный/4-1ВВ'). Для сравнения на диаграммах показано среднее прогрессирование опухоли у необработанных мышей (треугольники красного цвета). (F) In vivo персистирование Т-клеток, экспрессирующих различные рецепторы CD19. Частоту Т-клеток CD8+ tEGFR+ в периферической крови, полученной на 3-й и 10-й день после переноса, определяли посредством проточной цитометрии, и она показана в виде процента живых (PI-) клеток периферической крови. Статистические анализы выполняли с помощью t-критерия Стьюдента.
Фигура 19. Т-клетки с химерным рецептором CD19 с длинным спейсерным доменом активируются опухолью in vivo, но не способны увеличиваться в численности. (А) Экспрессия CD69 и CD25 на Т-клетках, модифицированных каждым химерным рецептором CD19, до переноса мышам NSG/Raji. (В) Группам мышей инокулировали опухолевые клетки Raji-ffluc, и через 7 дней получали меченные CFSE трансдуцированные химерным рецептором CD19 или контрольные Т-клетки. Образцы костного мозга и селезенки забирали из подгрупп мышей через 24 и 72 часа после введения Т-клеток. (С, D) Мультипараметрический анализ по методу проточной цитометрии мононуклеарных клеток костного мозга, полученных через 24 часа (С) и 72 часа (D) после переноса Т-клеток. Точечные графики показывают окрашивание антителами к CD3 и антителами к CD45 после прохождения клеток PI- через окно для обнаружения жизнеспособных Т-клеток человека. Окно для CD3- CD45+ содержит опухолевые клетки Raji. На гистограммах показана экспрессия CD25 и CD69 на живых (PI-) Т-клетках CD3+ CD45+. (Е) Частота Т-клеток CD3+ CD45+ в селезенках, полученных спустя 24 и 72 часа после переноса Т-клеток. На точечных графиках показано окно для живых спленоцитов PI-, и на каждом графике показан процент Т-клеток CD3+ CD45+. (F) Окрашивание PI костного мозга и спленоцитов спустя несколько часов после переноса Т-клеток мышам NSG/Raji. Числа на гистограммах показывают процент клеток PI+ в популяции CD3+. (G) Биолюминесцентная визуализация групп мышей, обработанных Т-клетками, экспрессирующими химерные рецепторы CD19 с коротким спейсером ('короткий/СD28' и 'короткий/4-1ВВ') и длинными спейсерами ('длинный/СD28 и 'длинный/4-1ВВ'), а также контрольными Т-клетками.
Фигура 20. Т-клетки, экспрессирующие химерные рецепторы с 4-1ВВ и CD3-зета, а также модифицированным шарнирным участком Fc IgG4, проявляют более эффективное функционирование in vitro и in vivo по сравнению с Т-клетками, экспрессирующими химерные рецепторы CD19 с 4-1ВВ и СD3-зета, а также шарнирным участком СD8-альфа. А. Цитолитическая активность модифицированных химерным рецептором CD19 Т-клеток с шарнирным участком Fc IgG4 и шарнирным участком СD8-альфа, а также контрольных Т-клеток в отношении меченых Сr51 клеток K562, трансфицированных с помощью CD19, клеток лимфомы Raji, которые экспрессируют CD19, и контрольных клеток K562. Лизис показан при различных соотношениях Е/Т в 4-часовом тесте с радиоактивным Сr51. В. Выработка интерферона гамма 5×104 Т-клетками, экспрессирующими химерный рецептор CD19 с шарнирным участком Fc IgG4 или шарнирным участком СD8-альфа через 24 часа совместного культивирования с опухолевыми клетками Raji. O.D., составляющая 1, соответствует ~500 пг/мл интерферона гамма. С. Тест с разведением красителя CFSE для измерения пролиферации Т-клеток, экспрессирующих химерный рецептор CD19 с шарнирным участком Fc IgG4 или шарнирным участком СD8-альфа, и Т-клеток, экспрессирующих только tEGFR (контроль), через 72 часа совместного культивирования с СD19-положительными клетками лимфомы Raji. Числа над каждой гистограммой указывают число клеточных делений, которым подверглась пролиферирующая субпопуляция. Возле каждого графика представлена доля Т-клеток в каждом окне, которые подверглись ≥3/2/1 клеточным делениям. D. In vivo противоопухолевая активность Т-клеток, экспрессирующих химерный рецептор CD19 с шарнирным участком Fc IgG4 (группа 1) или шарнирным участком CD8-альфа (группа 2), и Т-клеток, экспрессирующих только tEGFR (группа 3), у мышей NSG, которым инокулировали опухолевые клетки Raji, экспрессирующие люциферазу светлячка (ffluc). Визуализацию у мышей проводили спустя 17 дней после инокуляции опухоли и спустя 10 дней после инокуляции Т-клеток. Данные показывают более высокую опухолевую нагрузку у мышей, обработанных контрольными Т-клетками с tEGFR (группа 3) или Т-клетками с химерным рецептором CD19 с шарнирным участком СD8-альфа (группа 2), по сравнению с мышами, обработанными Т-клетками с химерным рецептором CD19 с шарнирным участком Fc IgG4 (группа 1).
Подробное описание
Если не определено иное, все технические и научные термины, применяемые в данном документе, имеют то же значение, какое обычно понимается специалистом в данной области, к которой относится настоящее изобретение.
Подразумевается, что термин "приблизительно", применяемый в данном документе при обозначении измеряемой величины, охватывает изменения, составляющие ±20% или ±10%, более предпочтительно ±5%, еще более предпочтительно ±1% и наиболее предпочтительно ±0,1% от установленной величины.
Термин "активация", применяемый в данном документе, относится к состоянию Т-клетки, которая была в достаточной степени стимулирована для индукции выявляемой клеточной пролиферации, выработки цитокинов или экспрессии маркеров клеточной поверхности, таких как CD69 и CD25, или выявляемых эффекторных функций.
Термин "индуцируемая активацией клеточная гибель", применяемый в данном документе, относится к состоянию Т-клетки, которая является активированной, но не может пролиферировать на протяжении более 2 поколении и характеризуется маркерами апоптоза.
Термин "антиген" или "Ag", применяемый в данном документе, относится к молекуле, которая вызывает иммунный ответ. Этот иммунный ответ может включать выработку антител, или активацию специфических иммунологически компетентных клеток, или и то и другое. Очевидно, что антиген может быть создан синтетически, получен рекомбинантным путем или может происходить из биологического образца. Такой биологический образец может включать, без ограничения, образец ткани, образец опухоли, клетку или биологическую жидкость.
Термин "противоопухолевый эффект", применяемый в данном документе, относится к биологическому эффекту, который может проявляться в уменьшении объема опухоли, снижении числа опухолевых клеток, снижении числа метастазов, увеличении ожидаемой продолжительности жизни или ослаблении различных физиологических симптомов, ассоциированных с раковым состоянием. "Противоопухолевый эффект" также может проявляться в снижении частоты рецидивов или увеличении времени до рецидива.
Термин "химерный рецептор", применяемый в данном документе, относится к рецептору, сконструированному синтетическим путем, содержащему лиганд-связывающий домен антитела или другую белковую последовательность, которая связывается с молекулой, ассоциированной с заболеванием или нарушением, и соединена посредством спейсерного домена с одним или несколькими внутриклеточными сигнальными доменами Т-клеточных или других рецепторов, такими как костимулирующий домен.
Термин "костимулирующий домен", применяемый в данном документе, относится к сигнальному фрагменту, который передает Т-клеткам сигнал, который, в дополнение к первичному сигналу, привносимому, например, зета-цепью CD3 комплекса TCR/CD3, опосредует Т-клеточный ответ, в том числе, без ограничения, активацию, пролиферацию, дифференцировку, секрецию цитокинов и т.п. Костимулирующий домен может включать всю молекулу или часть молекулы, без ограничения, CD27, CD28, 4-1ВВ, ОХ40, CD30, CD40, ICOS, функционально-связанного антигена лимфоцитов-1 (LFA-1), CD2, CD7, LIGHT, NKG2C, В7-Н3 и лиганда, который специфически связывается с CD83. В вариантах осуществления костимулирующим доменом является внутриклеточный сигнальный домен, который взаимодействует с другими внутриклеточными медиаторами для опосредования клеточного ответа, в том числе активации, пролиферации, дифференцировки и секреции цитокинов и т.п.
Термин "кодирующий" применяется в данном документе для обозначения свойства конкретных последовательностей нуклеотидов в полинуклеотиде, таком как ген, кДНК или мРНК, выступать в качестве матриц для синтеза других макромолекул, таких как определенная последовательность аминокислот. Таким образом, ген кодирует белок, если в результате транскрипции и трансляции мРНК, соответствующей этому гену, получают белок в клетке или другой биологической системе. Термин "последовательность нуклеиновой кислоты, кодирующая полипептид" включает все нуклеотидные последовательности, которые являются вырожденными вариантами друг друга и которые кодируют одну и ту же аминокислотную последовательность.
Термин "цитотоксический Т-лимфоцит" (CTL), применяемый в данном документе, относится к Т-лимфоциту, который экспрессирует CD8 на своей поверхности (например, Т-клетка CD8+). В некоторых вариантах осуществления такими клетками являются предпочтительно Т-клетки "памяти" (клетки ТМ), которые "обучены" антигеном.
Термин "центральная Т-клетка памяти" (или "ТСМ"), применяемый в данном документе, относится к "обученному" антигеном CTL, который экспрессирует CD62L или CCR-7 и CD45RO на своей поверхности и не экспрессирует или характеризуется сниженной экспрессией CD45RA по сравнению с наивными клетками. В вариантах осуществления центральные клетки памяти являются положительными в отношении экспрессии CD62L, CCR7, CD28, CD 127, CD45RO и CD95 и характеризуются сниженной экспрессией CD54RA по сравнению с наивными клетками.
Термин "эффекторная Т-клетка памяти" (или "ТЕМ"), применяемый в данном документе, относится к "обученному" антигеном CTL, который не экспрессирует или характеризуется сниженной экспрессией CD62L на своей поверхности по сравнению с центральными клетками памяти и не экспрессирует или характеризуется сниженной экспрессией CD45RA по сравнению с наивными клетками. В вариантах осуществления эффекторные клетки памяти являются отрицательными в отношении экспрессии CD62L и CCR7 по сравнению с наивными клетками или центральными клетками памяти и характеризуются изменчивой экспрессией CD28 и CD45RA.
Термин "наивные Т-клетки", применяемый в данном документе, относится к "необученному" антигеном Т-лимфоциту, который экспрессирует CD62L и CD45RA и не экспрессирует CD45RO по сравнению с центральными или эффекторными клетками памяти. В некоторых вариантах осуществления наивные Т-лимфоциты CD8+ характеризуются экспрессией фенотипических маркеров наивных Т-клеток, в том числе CD62L, CCR7, CD28, CD127 и CD45RA.
Термин "эффекторные ("ТЕ") Т-клетки", применяемый в данном документе, относится к клеткам, являющимся "обученными" антигеном цитотоксическими Т-лимфоцитами, которые не экспрессируют или характеризуются сниженной экспрессией CD62L, CCR7, CD28 и являются положительными по гранзиму В и перфорину по сравнению с центральными Т-клетками памяти или наивными Т-клетками.
Термины "обогащенный" и "истощенный", применяемые в данном документе для описания количеств типов клеток в смеси, относятся к воздействию на смесь клеток способа или этапа, которое приводит к увеличению числа клеток "обогащенного" типа и снижению числа клеток "истощенного" типа. Таким образом, в зависимости от источника исходной популяции клеток, подвергаемых способу обогащения, смесь или композиция может содержать приблизительно 60, 70, 80, 90, 95 или 99 процентов или более (по числу или содержанию) клеток "обогащенного" типа и около 40, 30, 20, 10, 5 или 1 процента или менее (по числу или содержанию) клеток "истощенного" типа.
Термин "эпитоп", применяемый 6 данном документе, относится к части антигена или молекулы, которая распознается иммунной системой, в том числе антителами, Т-клетками и/или В-клетками. Эпитопы обычно имеют по меньшей мере 7 аминокислот и могут быть линейными или конформационными.
Термин "выделенный", применяемый для описания различных полипептидов, раскрытых в данном документе, означает полипептид или нуклеиновую кислоту, которые были идентифицированы и отделены от и/или извлечены из компонента своего естественного окружения. Предпочтительно, выделенный полипептид или нуклеиновая кислота не связаны ни с какими из компонентов, с которыми они связаны в естественных условиях. Загрязняющими компонентами их естественного окружения являются соединения, которые обычно препятствуют диагностическим или терапевтическим путям применения полипептида или нуклеиновой кислоты и могут включать ферменты, гормоны и другие белковые или небелковые растворенные вещества.
Термин "внутриклеточный сигнальный домен", применяемый в данном документе, относится ко всему одному или всем нескольким доменам молекулы или части одного или нескольких доменов молекулы (в данном случае молекулы химерного рецептора), обеспечивающим активацию лимфоцита. Внутриклеточные домены таких молекул опосредуют передачу сигнала путем взаимодействия с клеточными медиаторами, в результате чего осуществляются пролиферация, дифференцировка, активация и другие эффекторные функции. В вариантах осуществления такие молекулы включают все CD28, CD3, 4-1ВВ или их части и их комбинации.
Термин "лиганд", применяемый в данном документе, относится к веществу, которое специфически связывается с другим веществом с образованием комплекса. Примеры лигандов включают эпитопы на антигенах, молекулы, которые связываются с рецепторами, субстраты, ингибиторы, гормоны и активаторы.
Термин "лиганд-связывающий домен", применяемый в данном документе, относится к веществу или часхи вещества, которое связывается с лигандом. Примеры лиганд-связывающих доменов включают антиген-связывающие части антител, внеклеточные домены рецепторов и активные центры ферментов.
Термин "функционально связанный", применяемый в данном документе, относится к функциональной связи между регуляторной последовательностью и последовательностью гетерологичной нуклеиновой кислоты, обуславливающей экспрессию последней. Например, первая последовательность нуклеиновой кислоты функционально связана со второй последовательностью нуклеиновой кислоты, если первая последовательность нуклеиновой кислоты находится в функциональной связи со второй последовательностью нуклеиновой кислоты. Например, промотор функционально связан с кодирующей последовательностью, если промотор влияет на транскрипцию или экспрессию кодирующей последовательности. В целом, функционально связанные последовательности ДНК являются смежными и при необходимости соединяют два участка, кодирующих белок, в одной рамке считывания.
Термин "процент (%) идентичности аминокислотных последовательностей" в отношении полипептидных последовательностей химерного рецептора, идентифицируемых в данном документе, определяется как процент аминокислотных остатков в последовательности-кандидате, которые идентичны аминокислотным остаткам в эталонной последовательности для каждого из лиганд-связывающего домена, спейсера, трансмембранного домена и/или домена, активирующего лимфоциты, после выравнивания последовательностей и, при необходимости, введения гэпов для достижения максимального процента идентичности последовательности и без учета любых консервативных замен как составляющих идентичности последовательностей. Выравнивание с целью определения процента идентичности аминокислотной последовательности может быть достигнуто различными способами, которые находятся в пределах компетенции специалиста в данной области, например, с помощью общедоступного компьютерного программного обеспечения, такого как программное обеспечение BLAST, BLAST-2, ALIGN, ALIGN-2 или Megalign (DNASTAR). Специалисты в данной области могут определять соответствующие параметры для измерения выравнивания, включая любые алгоритмы, необходимые для достижения максимального выравнивания по всей длине последовательностей, которые сравниваются. Например, значения % идентичности аминокислотных последовательностей получали с помощью компьютерной программы WU-BLAST-2 [Altschul et al., Methods in Enzymology, 266:460-480 (1996)], в которой используются несколько параметров поиска, большинству из которых присваиваются значения по умолчанию. Параметры, которым не присваиваются значения по умолчанию (например, регулируемые параметры), определяют следующими значениями: длина перекрывания = 1, доля перекрывания = 0,125, пороговый вес слова (Т) = 11 и матрица замен = BLOSUM62. Значение % идентичности аминокислотных последовательностей определяется путем деления (а) числа совпадающих идентичных аминокислотных остатков между каждой в отдельности или всеми в совокупности аминокислотными последовательностями полипептида из эталонных последовательностей химерного рецептора, представленных в таблице 2, и сравниваемой аминокислотной последовательностью, представляющей интерес, определяемого с помощью WU-BLAST-2, на (b) общее число аминокислотных остатков полипептида, представляющего интерес.
Термин "вариант полинуклеотида, кодирующего химерный рецептор" или "вариант последовательности нуклеиновой кислоты, кодирующей химерный рецептор", применяемый в данном документе, относится к молекуле кодирующей полипептид нуклеиновой кислоты, определенной ниже, характеризующейся по меньшей мере приблизительно 80% идентичностью последовательности нуклеиновой кислоты с последовательностью полинуклеотида, показанной в таблице 1, или определенным образом полученным из нее фрагментом, такой как полинуклеотид, кодирующий антиген-связывающий домен, полинуклеотид, кодирующий спейсерный домен, полинуклеотид, кодирующий трансмембранный домен, и/или полинуклеотид, кодирующий лимфоцит-стимулирующий домен. Как правило, вариант полинуклеотида, кодирующего химерный рецептор, или его фрагмент будут характеризоваться по меньшей мере приблизительно 80% идентичностью последовательности нуклеиновой кислоты, более предпочтительно по меньшей мере приблизительно 81% идентичностью последовательности нуклеиновой кислоты, более предпочтительно по меньшей мере приблизительно 82% идентичностью последовательности нуклеиновой кислоты, более предпочтительно по меньшей мере приблизительно 83% идентичностью последовательности нуклеиновой кислоты, более предпочтительно по меньшей мере приблизительно 84% идентичностью последовательности нуклеиновой кислоты, более предпочтительно по меньшей мере приблизительно 85% идентичностью последовательности нуклеиновой кислоты, более предпочтительно по меньшей мере приблизительно 86% идентичностью последовательности нуклеиновой кислоты, более предпочтительно по меньшей мере приблизительно 87% идентичностью последовательности нуклеиновой кислоты, более предпочтительно по меньшей мере приблизительно 88% идентичностью последовательности нуклеиновой кислоты, более предпочтительно по меньшей мере приблизительно 89% идентичностью последовательности нуклеиновой кислоты, более предпочтительно по меньшей мере приблизительно 90% идентичностью последовательности нуклеиновой кислоты, более предпочтительно по меньшей мере приблизительно 91% идентичностью последовательности нуклеиновой кислоты, более предпочтительно по меньшей мере приблизительно 92% идентичностью последовательности нуклеиновой кислоты, более предпочтительно по меньшей мере приблизительно 93% идентичностью последовательности нуклеиновой кислоты, более предпочтительно по меньшей мере приблизительно 94% идентичностью последовательности нуклеиновой кислоты, более предпочтительно по меньшей мере приблизительно 95% идентичностью последовательности нуклеиновой кислоты, более предпочтительно по меньшей мере приблизительно 96% идентичностью последовательности нуклеиновой кислоты, более предпочтительно по меньшей мере приблизительно 97% идентичностью последовательности нуклеиновой кислоты, более предпочтительно по меньшей мере приблизительно 98% идентичностью последовательности нуклеиновой кислоты и еще более предпочтительно по меньшей мере приблизительно 99% идентичностью последовательности нуклеиновой кислоты с последовательностью нуклеиновой кислоты, показанной в таблице 1, или полученным из нее фрагментом. Варианты не охватывают нативную нуклеотидную последовательность. В связи с этим из-за вырожденности генетического кода специалист в данной области незамедлительно обнаружит, что большое количество вариантов полинуклеотидов, кодирующих химерный рецептор, характеризующихся по меньшей мере приблизительно 80% идентичностью последовательности нуклеиновой кислоты с нуклеотидной последовательностью из таблицы 1, будут кодировать полипептид, имеющий аминокислотную последовательность, которая идентична аминокислотной последовательности из таблицы 2.
Термин "в значительной степени очищенный" относится к молекуле, которая практически свободна от других типов молекул, или клетке, которая практически свободна от других типов клеток. В значительной степени очищенная клетка также означает клетку, которая была отделена от других типов клеток, с которыми она обычно связана в своем встречающемся в природе состоянии. В некоторых примерах популяция в значительной степени очищенных клеток относится к гомогенной популяции клеток.
Термин "обнаруженный в незначительном количестве" при использовании в отношении наличия опухолевого антигена или других молекул на нормальных клетках относится к процентной доле нормальных клеток определенного типа, имеющих антиген или молекулу, и/или плотности антигена на клетках. В вариантах осуществления обнаруженный в незначительном количестве означает, что антиген или молекула обнаружены на менее 50% нормальных клеток определенного типа и/или при плотности менее 50% по сравнению с количеством клеток или антигена, обнаруживаемым в случае опухолевых клеток или других больных клеток.
"Т-клетки" или "Т-лимфоциты", применяемые в данном документе, могут происходить из любого вида млекопитающих, предпочтительно приматов, в том числе от обезьян, собак и людей. В некоторых вариантах осуществления Т-клетки являются аллогенными (от одного и того же вида, но другого донора) по отношению к субъекту-реципиенту; в некоторых вариантах осуществления Т-клетки являются аутологическими (донор и реципиент являются одним и тем же индивидуумом); в некоторых вариантах осуществления Т-клетки являются сингенными (донор и реципиент являются разными индивидуумами, но являются идентичными близнецами).
Принципы раскрытия
Настоящее раскрытие предусматривает нуклеиновые кислоты, кодирующие химерный рецептор, а также векторы и клетки-хозяева, содержащие такие нуклеиновые кислоты. Нуклеиновая кислота, кодирующая химерный рецептор, содержит ряд модульных компонентов, которые можно вырезать и заменить другими компонентами для создания индивидуального химерного рецептора для конкретной молекулы-мишени. Настоящее раскрытие предусматривает, что один из модульных компонентов является спейсерным компонентом. Было обнаружено, что длина спейсерного участка, который предположительно не имеет способности к передаче сигналов, влияет на in vivo эффективность Т-клеток, модифицированных для экспрессии химерного рецептора, и она должна быть индивидуальной для отдельных молекул-мишеней для обеспечения повышенной терапевтической активности.
В одном аспекте предлагаются способы и конструкты нуклеиновой кислоты для разработки химерного рецептора, который обеспечивает улучшенное распознавание опухолей, повышенные пролиферацию Т-клеток и/или выработку цитокинов в ответ на воздействие лиганда по сравнению с эталонным химерным рецептором. В вариантах осуществления предлагается библиотека нуклеиновых кислот, в которой каждая нуклеиновая кислота кодирует спейсерный участок, который отличается от других по последовательности и длине. Каждую из нуклеиновых кислот затем можно применять для образования конструкта нуклеиновой кислоты, кодирующей химерный рецептор, который можно испытывать in vivo (в животной модели) и/или in vitro с тем, чтобы можно было выбрать спейсер, который обеспечивает улучшенное распознавание опухолей, повышенные пролиферацию Т-клеток и/или выработку цитокинов в ответ на воздействие лиганда.
В вариантах осуществления нуклеиновая кислота, кодирующая химерный рецептор, содержит полинуклеотид, кодирующий лиганд-связывающий домен, где лигандом является опухолеспецифический или вирусоспецифический антиген или молекула, полинуклеотид, кодирующий индивидуальный полипептидный спейсер, при этом спейсер обеспечивает повышенную пролиферацию Т-клеток; полинуклеотид, кодирующий трансмембранный домен, и полинуклеотид, кодирующий один или несколько внутриклеточных сигнальных доменов. В вариантах осуществления используется длинный спейсер, если эпитоп молекулы-мишени является мембранно-проксимальным на клетке-мишени, и используется короткий спейсер, если эпитоп молекулы-мишени является мембранно-дистальным на клетке-мишени.
Конструкцию химерного рецептора можно разработать индивидуально в зависимости от типа опухоли или вируса, антигена-мишени или молекулы-мишени, присутствующих на опухоли, аффинности антитела к молекуле-мишени, гибкости, необходимой для антиген-связывающего домена, и/или внутриклеточного сигнального домена. В вариантах осуществления ряд конструктов, кодирующих химерные рецепторы, испытывают в моделях in vitro и in vivo для определения способности Т-клеток, модифицированных рецептором, к уничтожению опухолевых клеток у мышей с иммунодефицитом и пролиферации и персистированию после адоптивного переноса. В вариантах осуществления выбирают химерный рецептор, который обеспечивает способность по меньшей мере 30% клеток к пролиферации на протяжении по меньшей мере двух поколений in vitro и/или в течение 72 часов после введения in vivo. В вариантах осуществления не выбирают химерный рецептор, который приводит к тому, что более 50% клеток подвергаются индуцируемой активацией клеточной гибели (AICD) в течение 72 часов in vivo у мышей с иммунодефицитом, и не способен обеспечивать уничтожение опухолевых клеток.В зависимости от того, присутствует ли молекула-мишень на опухолевых клетках субъекта, химерный рецептор содержит лиганд-связывающий домен, который специфически связывается с молекулой-мишенью. В вариантах осуществления опухолевые клетки субъекта характеризуются опухолевыми молекулами на клеточной поверхности. Молекулу-мишень можно выбрать на основании определения ее присутствия на опухолевых клетках определенного субъекта. В вариантах осуществления молекулу-мишень выбирают так, что она является молекулой клеточной поверхности, встречающейся преимущественно на опухолевых клетках и не встречающейся в нормальных тканях в какой-либо значительной степени. В вариантах осуществления выбирают антитело для связывания с эпитопом на клеточной поверхности молекулы-мишени. В некоторых случаях эпитоп характеризуется своей близостью к клеточной мембране. Эпитоп характеризуется как проксимальный по отношению к мембране, когда, как предполагается или известно на основании структурного анализа, он располагается ближе к мембране клетки-мишени, чем другие эпитопы, которые, как предполагается или известно на основании структурного анализа, располагаются на большем расстоянии от мембраны клетки-мишени. В вариантах осуществления аффинность антитела, из которого конструируют scFv, сравнивают посредством анализов связывания, и антитела с различными аффинностями исследуют в форматах химерных рецепторов, экспрессируемых в Т-клетках, для определения аффинности, которая обеспечивает оптимальное распознавание опухоли, на основании более высоких цитотоксичности в отношении клеток-мишеней и/или выработки цитокинов Т-клетками и пролиферации последних.
Кроме того, спейсерный участок химерного рецептора можно изменять с целью оптимизации распознавания Т-клетками лиганда на клетке-мишени. В вариантах осуществления, когда антитело связывается с эпитопом на клетке-мишени, который находится в наиболее проксимальном положении по отношению к мембране, выбирают спейсер, длина которого превышает приблизительно 15 аминокислот. Например, в вариантах осуществления, когда эпитоп или его часть в антигене-мишени находятся среди первых 100 аминокислот линейной последовательности внеклеточного домена, прилежащих к трансмембранному домену, можно выбрать длинный спейсерный участок. В вариантах осуществления, когда антитело связывается с эпитопом на клетке-мишени, который находится в дистальном положении по отношению к мембране, выбирают спейсер, длина которого составляет приблизительно 119 или 15 аминокислот или менее. Например, в вариантах осуществления, когда эпитоп или его часть находятся среди 150 аминокислот от конца линейной последовательности внеклеточного домена, можно использовать короткий или средний спейсер. В вариантах осуществления спейсер содержит аминокислотную последовательность X1PPX2P.
Для повышения in vivo эффективности химерного рецептора можно использовать ряд комбинаций первичного и костимулирующего внутриклеточного сигнального домена. В вариантах осуществления различные конструкты химерного рецептора можно испытывать в животной модели in vivo для определения эффективности уничтожения опухоли. В вариантах осуществления костимулирующий внутриклеточный сигнальный домен выбирают из группы, включающей CD28 и его модифицированные варианты, 4-1ВВ и его модифицированные варианты и их комбинации. Можно включать другие костимулирующие домены, такие как ОХ40.
Центральные Т-клетки памяти CD8+ имеют внутреннее программирование, которое позволяет им персистировать в течение продолжительных периодов после введения, что делает их предпочтительной субпопуляцией Т-клеток CD8+ для иммунотерапии. В вариантах осуществления модифицированные специфическим химерным рецептором CD19 цитотоксические Т-клетки, полученные из центральных Т-клеток памяти CD8+, очищенных путем сортировки, вводят в присутствии или в отсутствие Т-клеток CD4+, модифицированных специфическим химерным рецептором CD19. В вариантах осуществления опухолеспецифические Т-клетки CD4+ проявляют противоопухолевую реактивность и содействуют опухолеспецифическим Т-клеткам CD8+ in vitro и in vivo. В конкретном варианте осуществления опухолеспецифические Т-клетки CD4+ или Т-клетки CD4+, выбранные из субпопуляций наивных клеток или центральных клеток памяти, используют отдельно или в комбинации с ТСМ CD8+.
Нуклеиновые кислоты, векторы и полипептиды
Настоящее раскрытие предлагает нуклеиновую кислоту, кодирующую химерный рецептор, пригодную для трансформации или трансдукции лимфоцитов для применения в адоптивной иммунотерапии. В вариантах осуществления нуклеиновая кислота содержит ряд модульных компонентов, которые предусматривают легкую замену элементов нуклеиновой кислоты. Не предполагая ограничение области применения настоящего раскрытия, считают, что химерный рецептор для каждого опухолевого антигена предпочтительно разработан индивидуально в отношении компонентов с целью обеспечения эффективности и эффективной экспрессии in vivo в клетках млекопитающих. Например, в конкретном варианте осуществления для обеспечения эффективности химерного рецептора, содержащего scFv, который связывается с эпитопом ROR1, расположенным в мембранно-дистальном домене Ig/Frizzled, используется спейсер, длина которого составляет приблизительно 15 аминокислот или менее. В другом конкретном варианте осуществления для обеспечения эффективности химерного рецептора, содержащего scFv, который связывается с эпитопом ROR1, расположенным в мембранно-проксимальном крингл-домене, используется спейсер, длина которого превышает 15 аминокислот. В другом варианте осуществления для обеспечения эффективности химерного рецептора, содержащего scFv, который связывается с CD19, используется спейсер, длина которого составляет 15 аминокислот или менее.
В вариантах осуществления выделенная нуклеиновая кислота, кодирующая химерный рецептор, содержит полинуклеотид, кодирующий лиганд-связывающий домен, где молекулой-мишенью является опухолеспецифический антиген, полинуклеотид, кодирующий полипептидный спейсер, при этом полипептидный спейсер содержит приблизительно 229 аминокислот или менее, полинуклеотид, кодирующий трансмембранный домен, и полинуклеотид, кодирующий внутриклеточный сигнальный домен. В вариантах осуществления вектор экспрессии содержит химерную нуклеиновую кислоту, описанную в данном документе. В данный документ также включены полипептиды, кодируемые всей нуклеиновой кислотой, кодирующей химерный рецептор, или ее частью.
Лиганд-связывающий домен
В вариантах осуществления нуклеиновая кислота, кодирующая химерный рецептор, содержит полинуклеотид, кодирующий лиганд-связывающий домен. В вариантах осуществления лиганд-связывающий домен специфически связывается с опухолеспецифическим или вирусоспецифическим антигеном. В вариантах осуществления лиганд-связывающим доменом является антитело или его фрагмент. Последовательность нуклеиновой кислоты, кодирующая антитело или фрагмент антитела, может быть без труда определена. В конкретном варианте осуществления полинуклеотид кодирует одноцепочечный Fv, который специфически связывается с CD19. В других конкретных вариантах осуществления полинуклеотид кодирует одноцепочечный Fv, который специфически связывается с ROR1. Последовательности этих антител известны и могут быть без труда определены специалистами в данной области.
Опухолевые антигены являются белками, которые вырабатываются опухолевыми клетками, вызывающими иммунный ответ. Выбираемый лиганд-связывающий домен по настоящему изобретению будет зависеть от типа рака, подлежащего лечению, и он может нацеливаться на опухолевые антигены или другие поверхностные молекулы опухолевых клеток. Образец опухоли от субъекта может быть охарактеризован с точки зрения присутствия некоторых биомаркеров или маркеров клеточной поверхности. Например, раковые клетки молочной железы от субъекта могут быть положительными или отрицательными по каждому из Her2Neu, рецептора эстрогенов и/или рецептора прогестерона. Выбирают опухолевый антиген или молекулу клеточной поверхности, которые встречаются на отдельных опухолевых клетках субъекта. Опухолевые антигены и молекулы клеточной поверхности хорошо известны в данной области и включают, например, раковый эмбриональный антиген (СEА), простатический специфический антиген, PSMA, Her2/neu, рецептор эстрогенов, рецептор прогестерона, эфрин В2, CD19, CD20, CD22, CD23, CD123, CS-1, ROR1, мезотелин, c-Met, GD-2 и TCR MAGE A3. В вариантах осуществления молекулой-мишенью является молекула клеточной поверхности, которая обнаруживается на опухолевых клетках и обнаруживается в незначительной степени в нормальных тканях, или ограничена в своей экспрессии нормальными тканями, не являющимися жизненно важными.
Другие молекулы-мишени включают, без ограничения, антигены, полученные из возбудителей инфекций, таких как HIV (вирус иммунодефицита человека), HBV (вирус гепатита В), HPV (вирус папилломы человека) и вирус гепатита С.
В одном варианте осуществления молекула-мишень в опухоли содержит один или несколько эпитопов, ассоциированных со злокачественной опухолью. Злокачественные опухоли экспрессируют ряд белков, которые могут выступать в качестве антигенов-мишеней для Т-клеточного рецептора или химерного рецептора, опосредующих распознавание. Другие молекулы-мишени принадлежат к группе молекул, связанных с трансформацией клеток, таких как онкоген HER-2/Neu/ErbB2. В вариантах осуществления опухолевый антиген избирательно экспрессируется или сверхэкспрессируется на опухолевых клетках по сравнению с контрольными клетками того же типа ткани. В других вариантах осуществления опухолевым антигеном является полипептид клеточной поверхности.
Непосредственно после идентификации поверхностной молекулы опухолевой клетки, на которую может нацеливаться химерный рецептор, выбирают и характеризуют эпитоп молекулы-мишени. В вариантах осуществления выбирают эпитоп, который является проксимальным по отношению к мембране опухолевой клетки. В других вариантах осуществления выбирают эпитоп, который является дистальным по отношению к мембране опухолевой клетки. Эпитоп характеризуется как проксимальный по отношению к мембране, когда, как предполагается или известно на основании структурного анализа, он располагается ближе к мембране клетки-мишени, чем другие эпитопы, которые, как предполагается или известно на основании структурного анализа, располагаются на большем расстоянии от мембраны клетки-мишени.
Антитела, которые специфически связываются с поверхностной молекулой опухолевой клетки, могут быть получены с помощью способов получения моноклональных антител, способов фагового дисплея, способов создания антител человека или гуманизированных антител или способов с применением трансгенного животного или растения, сконструированных для выработки антител человека. Фаг-дисплейные библиотеки частично или полностью синтетических антител доступны и могут быть подвергнуты скринингу в отношении антитела или его фрагмента, которые могут связываться с молекулой-мишенью. Также доступны фаг-дисплейные библиотеки антител человека. В вариантах осуществления антитела специфически связываются с поверхностной молекулой опухолевой клетки и не реагируют перекрестно с неспецифическими компонентами, такими как бычий сывороточный альбумин и другие неродственные антигены. Непосредственно после идентификации аминокислотная последовательность или последовательность полинуклеотида, кодирующая антитело, могут быть выделены и/или определены.
Антитела или антиген-связывающие фрагменты включают поликлональные антитела, моноклональное антитело, антитело человека, гуманизированное антитело, синтетическое антитело, химерное антитело, биспецифическое антитело, миниантитело и линейное антитело - всю молекулу или ее часть. Фрагменты антитела включают в себя часть интактного антитела, предпочтительно антиген-связывающий или вариабельный участок интактного антитела, и могут быть без труда получены. Примеры фрагментов антител включают фрагменты Fab, Fab', F(ab')2 и Fv; диатела; линейные антитела; одноцепочечных молекулы антител и полиспецифические антитела, образованные из фрагментов антител.
В вариантах осуществления может быть выделен и охарактеризован ряд различных антител, которые связываются с определенными поверхностными молекулами опухолевой клетки. В вариантах осуществления антитела характеризуются на основании специфичности к эпитопу молекулы-мишени. Кроме того, в некоторых случаях антитела, которые связываются с одним и тем же эпитопом, могут быть выбраны на основании аффинности антитела к этому эпитопу. В вариантах осуществления антитело имеет аффинность по меньшей мере 1 мМ и предпочтительно <50 нМ. В вариантах осуществления выбирают антитело, которое имеет более высокую аффинность к эпитопу по сравнению с другими антителами.
Например, выбирают антитело, которое имеет по меньшей мере в 2 раза, по меньшей мере в 5 раз, по меньшей мере в 10 раз, по меньшей мере в 20 раз, по меньшей мере в 30 раз, по меньшей мере в 40 раз или по меньшей мере в 50 раз большую аффинность, чем эталонное антитело, которое связывается с тем же эпитопом.
В вариантах осуществления молекулы-мишени выбирают из группы, включающей CD19, CD20, CD22, CD23, CD123, CS-1, ROR1, мезотелин, Her2, c-Met, PSMA, GD-2, TCR MAGE A3 и их комбинации.
В конкретных вариантах осуществления антигеном-мишенью является CD19. Ряд антител, специфичных к CD19, известен специалистам в данной области и может быть без труда охарактеризован в отношении последовательности, связывания с эпитопом и аффинности. В конкретном варианте осуществления конструкт химерного рецептора содержит последовательность scFv из антитела FMC63. В других вариантах осуществления scFv представляет собой scFv человека или гуманизированный scFv, содержащий вариабельный участок легкой цепи, который содержит последовательность CDRL1 RASQDISKYLN, последовательность CDRL2 SRLHSGV и последовательность CDRL3 GNTLPYTFG. В других вариантах осуществления scFv представляет собой scFv человека или гуманизированный scFv, содержащий вариабельный участок тяжелой цепи, который содержит последовательность CDRH1 DYGVS, последовательность CDRH2 VIWGSETTYYNSALKS и последовательность CDRH3 YAMDYWG. Настоящее раскрытие также охватывает вариабельные участки, которые характеризуются по меньшей мере 90% идентичностью аминокислотной последовательности по отношению к аминокислотной последовательности scFv FMC63 и которые имеют по меньшей мере такую же аффинность к CD19. В вариантах осуществления химерный рецептор имеет короткий или средний спейсер из 119 аминокислот или менее или 12 аминокислот или менее. В конкретном варианте осуществления спейсер имеет длину 12 аминокислот или менее и имеет последовательность SEQ ID NO: 4.
В вариантах осуществления CDR-участки находятся в пределах участков антитела, пронумерованных согласно Кабат следующим образом: для легкой цепи - аминокислоты CDRL1 24-34; аминокислоты CDRL2 50-56; аминокислоты CDRL3 89-97; для тяжелой цепи - аминокислоты CDRH1 31-35; аминокислоты CDRH2 50-65 и аминокислоты CDRH3 95-102. CDR-участки в антителах можно без труда определить.
В конкретных вариантах осуществления антигеном-мишенью является ROR1. Ряд антител, специфичных к ROR1, известен специалистам в данной области и может быть без труда охарактеризован в отношении последовательности, связывания с эпитопом и аффинности. В конкретном варианте осуществления конструкт химерного рецептора содержит последовательность scFv из антитела R12. В других вариантах осуществления scFv представляет собой scFv человека или гуманизированный scFv, содержащий вариабельный участок легкой цепи, который содержит последовательность CDRL1 ASGFDFSAYYM, последовательность CDRL2 TIYPSSG и последовательность CDRL3 ADRATYFCA. В других вариантах осуществления scFv представляет собой scFv человека или гуманизированный scFv, содержащий вариабельный участок тяжелой цепи, который содержит последовательность CDRH1 DTDDWY, последовательность CDRH2 VQSDGSYTKRPGVPDR и последовательность CDRH3 YIGGYVFG. Настоящее раскрытие также охватывает вариабельные участки, которые характеризуются по меньшей мере 90% идентичностью аминокислотной последовательности по отношению к аминокислотной последовательности scFv R12 и которые имеют по меньшей мере такую же аффинность к ROR1. В вариантах осуществления химерный рецептор имеет короткий или средний спейсер из 119 аминокислот или менее или 12 аминокислот или менее. В конкретном варианте осуществления спейсер имеет длину 12 аминокислот или менее и имеет последовательность SEQ ID NO: 4.
В конкретных вариантах осуществления антигеном-мишенью является ROR1. Ряд антител, специфичных к ROR1, известен специалистам в данной области и может быть без труда охарактеризован в отношении последовательности, связывания с эпитопом и аффинности. В конкретном варианте осуществления конструкт химерного рецептора содержит последовательность scFv из антитела R11. В других вариантах осуществления scFv представляет собой scFv человека или гуманизированный scFv, содержащий вариабельный участок легкой цепи, который содержит последовательность CDRL1 SGSDINDYPIS, последовательность CDRL2 INSGGST и последовательность CDRL3 YFCARGYS. В других вариантах осуществления scFv представляет собой scFv человека или гуманизированный ScFv, содержащий вариабельный участок тяжелой цепи, который содержит последовательность CDRH1 SNLAW, последовательность CDRH2 RASNLASGVPSRFSGS и последовательность CDRH3 NVSYRTSF. Настоящее раскрытие также охватывает вариабельные участки, которые характеризуются по меньшей мере 90% идентичностью аминокислотной последовательности по отношению к аминокислотной последовательности scFv R11 и которые имеют по меньшей мере такую же аффинность к ROR1. В вариантах осуществления химерный рецептор имеет длинный спейсер из 229 аминокислот или менее. В конкретном варианте осуществления спейсер имеет длину 229 аминокислот и имеет последовательность SEQ ID NO: 50.
В конкретных вариантах осуществления антигеном-мишенью является Неr2. Ряд антител, специфичных к Неr2, известен специалистам в данной области и может быть без труда охарактеризован в отношении последовательности, связывания с эпитопом и аффинности. В конкретном варианте осуществления конструкт химерного рецептора содержит последовательность scFv из антитела герцептина. В других вариантах осуществления scFv представляет собой scFv человека или гуманизированный scFv, содержащий вариабельный участок легкой цепи, который содержит последовательность CDRL1, последовательность CDRL2 и последовательность CDRL3 антитела герцептина. В других вариантах осуществления scFv представляет собой scFv человека или гуманизированный scFv, содержащий вариабельный участок тяжелой цепи, который содержит последовательность CDRH1, последовательность CDRH2 и последовательность CDRH3 герцептина. Последовательности CDR могут быть без труда определены на основании аминокислотной последовательности герцептина. Настоящее раскрытие также охватывает вариабельные участки, которые характеризуются по меньшей мере 90% идентичностью аминокислотной последовательности по отношению к аминокислотной последовательности scFv герцептина и которые имеют по меньшей мере такую же аффинность к Неr2. В вариантах осуществления химерный рецептор имеет длинный спейсер из 229 аминокислот или менее. В конкретном варианте осуществления спейсер имеет длину 229 аминокислот и имеет последовательность SEQ ID NO: 50.
В вариантах осуществления полинуклеотид, кодирующий лиганд-связывающий домен, функционально связан с полинуклеотидом, кодирующим спейсерный участок. В вариантах осуществления полинуклеотид, кодирующий лиганд-связывающий домен, может также иметь один или несколько сайтов для фермента рестрикции на 5'- и/или 3'-конце кодирующей последовательности для обеспечения легкого вырезания и замены полинуклеотида другим полинуклеотидом, кодирующим лиганд-связывающий домен, связывающийся с другим антигеном или имеющим другие характеристики связывания. Например, сайт рестрикции NheI расположен выше лидерной последовательности; а 3' RsrII, расположенный в пределах шарнирного участка, обеспечивает субклонирование любого предпочтительного scFv в вектор химерного рецептора. В вариантах осуществления полинуклеотид подвергнут оптимизации кодонов для экспрессии в клетках млекопитающих.
В вариантах осуществления полинуклеотид, кодирующий лиганд-связывающий домен, функционально связан с полинуклеотидом, кодирующим сигнальный пептид. В вариантах осуществления сигнальным пептидом является сигнальный пептид гранулоцитарного колониестимулирующего фактора. Могут использоваться полинуклеотиды, кодирующие другие сигнальные пептиды, такие как СD8-альфа.
В вариантах осуществления полинуклеотид, кодирующий лиганд-связывающий домен, функционально связан с промотором. Выбирают промотор, который обеспечивает экспрессию рецептора химерного антигена в клетке млекопитающего. В конкретном варианте осуществления промотором является промотор гена фактора элонгации (EF-1). Другим примером подходящего промотора является последовательность немедленно-раннего промотора цитомегаловируса (CMV). Однако, можно использовать другие последовательности конститутивных промоторов, включая, без ограничения, ранний промотор вируса обезьян 40 (SV40), промотор длинного концевого повтора (LTR) вируса рака молочных желез мышей (MMTV) и вируса иммунодефицита человека (HIV), промотор MuMoLV, промотор вируса лейкоза птиц, немедленно-ранний промотор вируса Эпштейна-Барр, промотор вируса саркомы Рауса, а также промоторы генов человека, такие как, без ограничения, промотор гена актина, промотор гена миозина, промотор гена гемоглобина и промотор гена креатинкиназы. Также рассматриваются индуцируемые промоторы. Примеры индуцируемых промоторов включают, без ограничения, промотор гена металлотионеина, промотор гена глюкокортикоидного рецептора, промотор гена рецептора прогестерона и промотор гена устойчивости к тетрациклину.
Конкретный вариант осуществления полинуклеотида, кодирующего лиганд-связывающий домен, показан в таблице 1 в виде scFv из антитела, которое специфически связывается с CD19, такого как FMC63. Полинуклеотид, кодирующий гибкий линкер, содержащий аминокислоты GSTSGSGKPGSGEGSTKG (SEQ ID NO: 36), разделяет цепи VH и VL в scFv. Аминокислотная последовательность scFv, содержащая линкер, показана в таблице 2 (SEQ ID NO: 11). Известны другие антитела, нацеливающиеся на CD19, такие как SJ25C1 и HD37 (SJ25C1: Bejcek et al. Cancer Res 2005, PMID 7538901; HD37: Pezutto et al. JI 1987, PMID 2437199).
Спейсер
В вариантах осуществления нуклеиновая кислота, кодирующая химерный рецептор, содержит полинуклеотид, кодирующий спейсерный участок. Было обнаружено, что длина спейсерного участка, который предположительно не имеет способности к передаче сигналов, влияет на in vivo эффективность Т-клеток, модифицированных для экспрессии химерного рецептора, и она должна быть индивидуальной для отдельных молекул-мишеней для обеспечения оптимального распознавания опухолевой клетки или клетки-мишени. В вариантах осуществления нуклеиновая кислота, кодирующая химерный рецептор, содержит полинуклеотид, кодирующий индивидуальный спейсерный участок, выбранный из библиотеки полинуклеотидов, кодирующих спейсерные участки. В вариантах осуществления длину спейсера выбирают на основании расположения эпитопа, аффинности антитела к эпитопу и/или способности Т-клеток, экспрессирующих химерный рецептор, пролиферировать in vitro и/или in vivo в ответ на распознавание антигена.
Как правило, спейсерный участок находится между лиганд-связывающим доменом и трансмембранным доменом химерного рецептора. В вариантах осуществления спейсерный участок обеспечивает гибкость лиганд-связывающего домена, способствует высоким уровням экспрессии в лимфоцитах. СD19-специфический химерный рецептор, имеющий спейсерный домен из приблизительно 229 аминокислот, обладал меньшей противоопухолевой активностью, чем CD19-специфический химерный рецептор с коротким спейсерным участком, содержащим только модифицированный шарнирный участок IgG4. Другим химерным рецепторам, таким как химерные рецепторы, сконструированные из scFv R12 или 2А2, также требуется короткий спейсер для оптимального запуска эффекторных функций Т-клеток, в то время как химерному рецептору, сконструированному из scFv R11, специфичного к ROR1, требуется длинный спейсерный домен из приблизительно 229 аминокислот для распознавания опухоли.
В вариантах осуществления спейсерный участок имеет по меньшей мере приблизительно от 10 до 229 аминокислот, приблизительно от 10 до 200 аминокислот, приблизительно от 10 до 175 аминокислот, приблизительно от 10 до 150 аминокислот, приблизительно от 10 до 125 аминокислот, приблизительно от 10 до 100 аминокислот, приблизительно от 10 до 75 аминокислот, приблизительно от 10 до 50 аминокислот, приблизительно от 10 до 40 аминокислот, приблизительно от 10 до 30 аминокислот, приблизительно от 10 до 20 аминокислот или приблизительно от 10 до 15 аминокислот, и включает любое целое число между конечными значениями любого из приведенных диапазонов. В вариантах осуществления спейсерный участок имеет приблизительно 12 аминокислот или менее, приблизительно 119 аминокислот или менее или приблизительно 229 аминокислот или менее.
В некоторых вариантах осуществления спейсерный участок получен из шарнирного участка иммуноглобулин-подобной молекулы. В вариантах осуществления спейсерный участок содержит весь шарнирный участок из IgG1 человека, IgG2 человека, IgG3 человека или IgG4 человека или его часть и может содержать одну или несколько аминокислотных замен. Иллюстративные последовательности шарнирных участков приведены в таблице 8. В вариантах осуществления часть шарнирного участка включает аминокислоты верхней области шарнирного участка, находящиеся между вариабельным участком тяжелой цепи и сердцевиной, и аминокислоты сердцевины шарнирного участка, включающие в себя полипролиновый участок. Как правило, верхняя область шарнирного участка имеет приблизительно от 3 до 10 аминокислот. В некоторых случаях спейсерный участок содержит аминокислотную последовательность X1PPX2P (SEQ ID NO: 1). В вариантах осуществления X1 является цистеином, глицином или аргинином, а Х2 является цистеином или треонином.
В вариантах осуществления последовательности шарнирного участка могут быть модифицированы по одной или нескольким аминокислотам во избежание нежелательных структурных взаимодействий, таких как димеризация. В конкретном варианте осуществления спейсерный участок содержит часть модифицированного шарнирного участка IgG4 человека, например, как показано в таблице 2 или таблице 8 (SEQ ID NO: 21). Типичный полинуклеотид, кодирующий часть модифицированного шарнирного участка IgG4, представлен в таблице 1 (SEQ ID NO: 4). В вариантах осуществления шарнирный участок может характеризоваться по меньшей мере приблизительно 90%, 92%, 95% или 100% идентичностью последовательности с аминокислотной последовательностью шарнирного участка, идентифицированной в таблице 2 или таблице 8. В конкретном варианте часть шарнирного участка IgG4 человека имеет аминокислотную замену в аминокислотах сердцевины от CPSP до СРРС.
В некоторых вариантах осуществления весь шарнирный участок или его часть объединяют с одним или несколькими доменами константного участка иммуноглобулина. Например, часть шарнирного участка можно объединять со всем доменом СН2 или СН3 или его вариантом или его частью. В вариантах осуществления спейсерный участок не содержит последовательность шарнирного участка длиной 47-48 аминокислот из СD8-альфа или же спейсерный участок содержит внеклеточную часть молекулы CD28.
В вариантах осуществления короткий спейсерный участок имеет приблизительно 12 аминокислот или менее и содержит всю последовательность шарнирного участка IgG4 или его варианта или часть таковой, средний спейсерный участок имеет приблизительно 119 аминокислот или менее и содержит всю последовательность шарнирного участка IgG4 и весь участок СН3 или его вариант или части таковых, и длинный спейсер имеет приблизительно 229 аминокислот или менее и содержит всю последовательность шарнирного участка IgG4, весь участок СН2 и весь участок СН3 или их варианты или части таковых.
Полинуклеотид, кодирующий спейсерный участок, может быть без труда получен с помощью синтетических или рекомбинантных способов из аминокислотной последовательности. В вариантах осуществления полинуклеотид, кодирующий спейсерный участок, функционально связан с полинуклеотидом, кодирующим трансмембранный участок. В вариантах осуществления полинуклеотид, кодирующий спейсерный участок, может также иметь один или несколько сайтов для фермента рестрикции на 5'- и/или 3'-конце кодирующей последовательности для обеспечения легкого вырезания и замены полинуклеотида другим полинуклеотидом, кодирующим другой спейсерный участок. В вариантах осуществления полинуклеотид, кодирующий спейсерный участок, подвергнут оптимизации кодонов для экспрессии в клетках млекопитающих.
В вариантах осуществления предлагается библиотека полинуклеотидов, каждый из которых кодирует различный спейсерный участок. В варианте осуществления спейсерный участок выбирают из группы, включающей последовательность шарнирного участка из IgG1, IgG2, IgG3 или IgG4 или часть таковой, последовательность шарнирного участка из IgG1, IgG2, IgG3 или IgG4 в комбинации со всем участком СН2 или его вариантом или частью таковых, последовательность шарнирного участка из IgG1, IgG2, IgG3 или IgG4 в комбинации со всем участком СН3 или его вариантом или частью таковых и последовательность шарнирного участка из IgG1, IgG2, IgG3 или IgG4 в комбинации со всем участком СН2 или его вариантом или частью таковых и со всем участком СН3 или его вариантом или частью таковых. В вариантах осуществления короткий спейсерный участок представляет собой модифицированную последовательность шарнирного участка IgG4 (SEQ ID NO: 4), имеющую 12 аминокислот или менее, средняя последовательность представляет собой последовательность шарнирного участка IgG4 вместе с последовательностью СН3, имеющую 119 аминокислот или менее (SEQ ID NO: 49); или же последовательность шарнирного участка IgG4 вместе с участками СН2 и СН3 имеет 229 аминокислот или менее (SEQ ID NO: 50).
В вариантах осуществления в данном документе предлагается способ выбора спейсерного участка для химерного рецептора. Некоторые конструкты химерных рецепторов, несмотря на эффективность в отношении активации Т-клеток и управления уничтожением ими опухолевых клеток in vitro, не были эффективными in vivo. Кроме того, профиль побочных эффектов модификации Т-клеток химерным рецептором может быть таким, что будет приводить к тому, что больше клеток будет подвергаться индуцируемой активацией клеточной гибели, или вызывать повышение цитокинеза in vivo. В вариантах осуществления способ включает обеспечение множества нуклеиновых кислот, кодирующих химерные рецепторы, где химерные рецепторы, кодируемые нуклеиновыми кислотами, отличаются только спейсерным участком; введение каждой из нуклеиновых кислот, кодирующих химерные рецепторы, в отдельную популяцию Т-лимфоцитов; размножение каждой отдельной популяции лимфоцитов in vitro, и введение каждой популяции лимфоцитов животному, несущему опухоль, для определения противоопухолевой эффективности каждого из химерных рецепторов при экспрессии в Т-клетках, и выбор химерного рецептора, который обеспечивает противоопухолевую эффективность по сравнению с каждой из других отдельных популяций лимфоцитов, модифицированных каждым из других химерных рецепторов.
Известны животные модели различных опухолей. Противоопухолевую эффективность можно измерить путем определения снижения объема опухоли, путем определения смертности животных, персистирования генетически модифицированных Т-клеток in vivo, активации генетически модифицированных Т-клеток (например, путем выявления повышения экспрессии CD25 и/или CD69) и/или пролиферации генетически модифицированных Т-клеток in vivo. В варианте осуществления выбирают химерный рецептор, который обеспечивает наилучшую противоопухолевую эффективность in vivo, определенную с помощью одного или нескольких из этих параметров. Отсутствие противоопухолевой эффективности можно определить по отсутствию персистирования генетически модифицированных лимфоцитов in vivo, гибели животных, увеличению интенсивности апоптоза, измеренному по повышению индукции каспазы-3, и/или снижению пролиферации генетически модифицированных лимфоцитов.
В других вариантах осуществления способ выбора спейсера включает выбор эпитопа молекулы-мишени и характеристику положения эпитопа по отношению к клеточной мембране, выбор спейсерного участка, который является длинным или коротким в зависимости от положения эпитопа по отношению к клеточной мембране, выбор антитела или его фрагмента, которые имеют большую или меньшую аффинность к эпитопу по сравнению с эталонным антителом, и определение того, обеспечивает ли конструкт химерного рецептора повышенную пролиферацию Т-клеток или выработку цитокинов in vitro и/или in vivo. В некоторых вариантах осуществления, если эпитоп-мишень или его часть расположены проксимально по отношению к мембране, они находятся среди первых 100 аминокислот линейной последовательности внеклеточного домена, прилежащих к трансмембранному домену. Если эпитоп расположен проксимально по отношению к мембране, выбирают длинный спейсер (например, 229 аминокислот или менее и более 119 аминокислот). В некоторых вариантах осуществления, если эпитоп-мишень расположен дистально по отношению к мембране, он находится среди первых 150 аминокислот от конца линейной последовательности внеклеточного домена. Если эпитоп расположен дистально по отношению к мембране, выбирают средний или короткий спейсер (например, 119 аминокислот или менее или 12-15 аминокислот или менее). Альтернативно, определить, является ли эпитоп проксимальным или дистальным по отношению к мембране, можно с помощью моделирования трехмерной структуры или на основании анализа кристаллической структуры.
В некоторых вариантах осуществления выбирают химерный рецептор, который обеспечивает пролиферацию по меньшей мере 30% клеток на протяжении двух поколений in vitro и/или in vivo. В некоторых вариантах осуществления не выбирают химерный рецептор, если он приводит к тому, что по меньшей мере 50% клеток подвергаются индуцируемой активации клеточной гибели в течение 72 часов. В вариантах осуществления выбирают короткий спейсер (например, 15 аминокислот или менее), если эпитоп является дистальным по отношению к мембране. В вариантах осуществления выбирают длинный спейсер (например, 229 аминокислот или менее и более 119 аминокислот), если эпитоп является проксимальным по отношению к мембране.
В вариантах осуществления обеспечение множества нуклеиновых кислот, кодирующих химерные рецепторы, где химерные рецепторы, кодируемые нуклеиновыми кислотами, отличаются только спейсерным участком, включает обеспечение конструкта химерного рецептора, содержащего полинуклеотид, кодирующий лиганд-связывающий домен, где лигандом является опухолеспецифический антиген, вирусный антиген или любая другая молекула, экспрессируемая в популяции клеток-мишеней, пригодная для опосредования распознавания и удаления лимфоцитом; полинуклеотид, кодирующий первый полипептидный спейсер, имеющий определенный сайт рестрикции на 5'- и 3'- концах кодирующей последовательности для первого полипептидного спейсера; полинуклеотид, кодирующий трансмембранный домен; и полинуклеотид, кодирующий один или несколько внутриклеточных сигнальных доменов.
В вариантах осуществления способ дополнительно включает обеспечение одного или нескольких полинуклеотидов, каждый из которых кодирует различный спейсерный участок. В варианте осуществления различные спейсерные участки выбирают из группы, включающей последовательность шарнирного участка из IgG1, IgG2, IgG3 или IgG4 или его варианта или часть таковой, последовательность шарнирного участка из IgG1, IgG2, IgG3 или IgG4 в комбинации со всем участком СН2 или его вариантом или частью таковых, последовательность шарнирного участка из IgG1, IgG2, IgG3 или IgG4 в комбинации со всем участком СН3 или его вариантом или частью таковых и последовательность шарнирного участка из IgG1, IgG2, IgG3 или IgG4 в комбинации со всем участком СН2 или его вариантом или частью таковых и со всем участком СН3 или его вариантом или частью таковых. В вариантах осуществления участки СН2 или СН3 можно модифицировать с помощью одной или нескольких делеций или аминокислотных замен с целью обеспечения экспрессии в лимфоцитах и/или с целью минимизации взаимодействий с другими молекулами. В вариантах осуществления часть шарнирного участка содержит по меньшей мере аминокислоты верхней области и последовательность сердцевины. В вариантах осуществления шарнирный участок содержит последовательность X1PPX2P.
В вариантах осуществления способ дополнительно включает замену полинуклеотида, кодирующего спейсерный участок, полинуклеотидом, кодирующим другой спейсерный участок, с образованием нуклеиновой кислоты, кодирующей химерный рецептор с другим спейсерным участком. Способ можно повторять для образования любого количества нуклеиновых кислот, кодирующих химерные рецепторы, каждый из которых отличается спейсерным участком. В вариантах осуществления химерные рецепторы, кодируемые нуклеиновыми кислотами, отличаются друг от друга только спейсерным участком.
Трансмембранный домен
В вариантах осуществления нуклеиновая кислота, кодирующая химерный рецептор, содержит полинуклеотид, кодирующий трансмембранный домен. Трансмембранный домен обеспечивает заякоривание химерного рецептора в мембране.
В варианте осуществления применяется трансмембранный домен, который в естественных условиях связан с одним из доменов в химерном рецепторе. В некоторых случаях трансмембранный домен может быть выбран или модифицирован с помощью аминокислотной замены во избежание связывания таких доменов с трансмембранными доменами одинаковых или разных поверхностных мембранных белков с целью минимизации взаимодействий с другими членами рецепторного комплекса.
Трансмембранный домен может быть получен из природного или синтетического источника. Если источник является природным, домен может быть получен из любого мембраносвязанного или трансмембранного белка. Трансмембранные участки включают в себя по меньшей мере трансмембранный(трансмембранные) участок(участки) альфа-, бета- или зета-цепи Т-клеточного рецептора, CD28, CD3, CD45, CD4, CD8, CD9, CD16, CD22; CD33, CD37, CD64, CD80, CD86, CD134, CD137 и CD154. В конкретном варианте осуществления трансмембранный домен содержит аминокислотную последовательность трансмембранного домена CD28, как показано в таблице 2. Типичная полинуклеотидная последовательность, кодирующая трансмембранный домен CD28, показана в таблице 1(SEQ ID NO: 5).
Трансмембранный домен может быть синтетическим или вариантом трансмембранного домена, встречающегося в природе. В вариантах осуществления синтетические трансмембранные домены или варианты трансмембранных доменов содержат преимущественно гидрофобные остатки, такие как лейцин и валин. В вариантах осуществления трансмембранный домен может характеризоваться по меньшей мере приблизительно 80%, 85%, 90%, 95% или 100% идентичностью аминокислотной последовательности с трансмембранным доменом, показанным в таблице 2 или таблице 6. Варианты трансмембранных доменов предпочтительно имеют рассчитанный по шкале Кайта-Дулиттла индекс гидрофобности, составляющий по меньшей мере 50.
Полинуклеотид, кодирующий трансмембранный домен, может быть без труда получен с помощью синтетических или рекомбинантных способов. В вариантах осуществления полинуклеотид, кодирующий трансмембранный домен, функционально связан с полинуклеотидом, кодирующим внутриклеточный сигнальный участок. В вариантах осуществления полинуклеотид, кодирующий трансмембранный домен, может также иметь один или несколько сайтов для фермента рестрикции на 5'- и/или 3'-конце кодирующей последовательности для обеспечения легкого вырезания и замены полинуклеотида, кодирующего трансмембранный домен, другим полинуклеотидом, кодирующим другой трансмембранный домен. В вариантах осуществления полинуклеотид, кодирующий трансмембранный домен, подвергнут оптимизации кодонов для экспрессии в клетках млекопитающих.
Внутриклеточный сигнальный домен
В вариантах осуществления нуклеиновая кислота, кодирующая химерный рецептор, содержит полинуклеотид, кодирующий внутриклеточный сигнальный домен. Внутриклеточный сигнальный домен обеспечивает активацию одной функции трансдуцированной клетки, экспрессирующей химерный рецептор, при связывании с лигандом, экспрессируемым на опухолевых клетках. В вариантах осуществления внутриклеточный сигнальный домен содержит один или несколько внутриклеточных сигнальных доменов. В вариантах осуществления внутриклеточный сигнальный домен представляет собой часть и/или вариант внутриклеточного сигнального домена, обеспечивающие активацию по меньшей мере одной функции трансдуцированной клетки.
Примеры внутриклеточных сигнальных доменов для применения в химерном рецепторе по настоящему раскрытию включают цитоплазматические последовательности зета-цепи CD3 и/или корецепторы, которые действуют совместно для инициирования передачи сигнала после привлечения химерных рецепторов, а также любое производное или вариант этих последовательностей и любую синтетическую последовательность, которая имеет такие же функциональные возможности. Можно сказать, что активация Т-клеток опосредована двумя различными классами цитоплазматических сигнальных последовательностей: цитоплазматическими сигнальными последовательностями, которые инициируют антиген-зависимую первичную активацию и обеспечивают передачу сигнала, индуцируемого молекулами, подобными Т-клеточным рецепторам (первичные цитоплазматические сигнальные последовательности), и цитоплазматическими сигнальными последовательностями, которые действуют антиген-независимым образом для обеспечения передачи вторичного или костимулирующего сигнала (вторичные цитоплазматические сигнальные последовательности). Первичные цитоплазматические сигнальные последовательности, которые действуют стимулирующим образом, могут содержать сигнальные мотивы, которые известны как рецепторные тирозиновые активирующие мотивы или ITAM. Примеры ITAM, содержащих первичные цитоплазматические сигнальные последовательности, включают ITAM, полученные из CD3-3eTa, FcR-гамма, CD3-гамма, CD3-дельта, CD3-эпсилон, CD5, CD22, CD79a, CD79b и CD66d. В вариантах осуществления первичный сигнальный внутриклеточный домен может характеризоваться по меньшей мере приблизительно 80%, 85%, 90% или 95% идентичностью последовательности с CD3-зета, имеющей последовательность, представленную в таблице 2. В вариантах осуществления СD3-зета сохраняет по меньшей мере один, два, три или все ITAM-участки, показанные в таблице 7.
В предпочтительном варианте осуществления можно разработать внутриклеточный сигнальный домен химерного рецептора, содержащий сигнальный домен СD3-зета сам по себе или в комбинации с любым(любыми) другим(другими) предпочтительным(предпочтительными) цитоплазматическим(цитоплазматическими) доменом(доменами). Например, внутриклеточный сигнальный домен химерного рецептора может содержать зета-цепь CD3 и костимулирующий сигнальный домен. Костимулирующий сигнальный участок относится к части химерного рецептора, содержащей внутриклеточный домен костимулирующей молекулы. Костимулирующая молекула является молекулой клеточной поверхности, отличной от антигенного рецептора или его лигандов, которая требуется для ответа лимфоцитов на антиген. Примеры таких молекул включают CD27, CD28, 4-1ВВ (CD137), ОХ40, CD30, CD40, функционально-связанный антиген лимфоцитов-1 (LFA-1), CD2, CD7, LIGHT, NKG2C, В7-Н3 и лиганд, который специфически связывается с CD83. В вариантах осуществления костимулирующий сигнальный домен может характеризоваться по меньшей мере 80%, 85%, 90% или 95% идентичностью аминокислотной последовательности с внутриклеточным доменом CD28, показанным в таблице 5, или с 4-1ВВ, имеющим последовательность, представленную в таблице 2. В варианте осуществления вариант внутриклеточного домена CD28 содержит аминокислотную замену в положениях 186-187, в которых LL заменены на GG.
Внутриклеточные сигнальные последовательности химерного рецептора могут быть соединены друг с другом случайным или определенным образом. Связь может необязательно быть образована коротким олиго- или полипептидным линкером, предпочтительно длиной от 2 до 10 аминокислот. В одном варианте осуществления внутриклеточный сигнальный домен содержит весь сигнальный домен СD3-зета или его вариант или часть таковых и весь сигнальный домен CD28 или его вариант или часть таковых. В другом варианте осуществления внутриклеточный сигнальный домен содержит весь сигнальный домен СD3-зета или его вариант или часть таковых и весь сигнальный домен 4-1ВВ или его вариант или часть таковых. В еще одном варианте осуществления внутриклеточный сигнальный домен содержит весь сигнальный домен СD3-зета или его вариант или часть таковых, весь сигнальный домен CD28 или его вариант или часть таковых и весь сигнальный домен 4-1ВВ или его вариант или часть таковых. В конкретном варианте осуществления аминокислотная последовательность внутриклеточного сигнального домена, содержащего вариант СD3-зета и часть внутриклеточного сигнального домена 4-1ВВ, представлена в таблице 2. Типичная последовательность нуклеиновой кислоты представлена в таблице 1 (SEQ ID NO: 6; SEQ ID NO: 7).
В варианте осуществления внутриклеточный сигнальный домен, кодируемый полинуклеотидом, содержит внутриклеточный домен 4-1ВВ, соединенный с частью домена СD3-зета. В других вариантах осуществления внутриклеточный домен 4-1ВВ и внутриклеточный домен CD28 соединены с частью домена СD3-зета.
Полинуклеотид, кодирующий внутриклеточный сигнальный домен, может быть без труда получен с помощью синтетических или рекомбинантных способов из аминокислотной последовательности. В вариантах осуществления полинуклеотид, кодирующий внутриклеточный сигнальный домен, может также иметь один или несколько сайтов для фермента рестрикции на 5'- и/или 3'-конце кодирующей последовательности для обеспечения легкого вырезания и замены полинуклеотида, кодирующего внутриклеточный сигнальный домен, другим полинуклеотидом, кодирующим другой внутриклеточный сигнальный домен. В вариантах осуществления полинуклеотид, кодирующий внутриклеточный сигнальный домен, подвергнут оптимизации кодонов для экспрессии в клетках млекопитающих.
Маркерные последовательности
В вариантах осуществления нуклеиновая кислота, кодирующая химерный рецептор, необязательно дополнительно содержит полинуклеотидную последовательность, кодирующую маркерную последовательность. Маркерная последовательность может обеспечивать выбор трансдуцированных клеток и идентификацию трансдуцированных клеток. В вариантах осуществления полинуклеотидная последовательность, кодирующая маркерную последовательность, функционально связана с полинуклеотидной последовательностью, кодирующей линкерную последовательность. В вариантах осуществления линкерной последовательностью является расщепляемая линкерная последовательность.
Можно использовать ряд различных маркерных последовательностей. Как правило, маркерная последовательность имеет функциональную характеристику, которая позволяет производить выбор трансдуцированных клеток и/или выявление трансдуцированных клеток. В вариантах осуществления маркерная последовательность совместима с трансдукцией лимфоцитов человека.
Положительным селективным маркером может быть ген, который при введении в клетку-хозяина экспрессирует доминантный фенотип, обеспечивающий положительную селекцию клеток, несущих этот ген. Гены этого типа известны в данной области техники и включают, среди прочих, ген гигромицин-В-фосфотрансферазы (hph), который придает устойчивость к гигромицину В, ген аминогликозидфосфотрансферазы (nео или aph) из Тn5, который отвечает за устойчивость к антибиотику G418, ген дигидрофолатредуктазы (DHFR), ген аденозиндезаминазы (ADA) и ген множественной лекарственной устойчивости (MDR).
В варианте осуществления нуклеиновая кислота, кодирующая химерный рецептор, дополнительно содержит полинуклеотид, кодирующий маркерную последовательность. В варианте осуществления маркерной последовательностью является усеченный рецептор эпидермального фактора роста, показанный в таблице 2. Иллюстративный полинуклеотид для усеченного рецептора эпидермального фактора роста показан в таблице 1 (SEQ ID NO: 9). В вариантах осуществления полинуклеотид, кодирующий маркерную последовательность, функционально связан с полинуклеотидом, кодирующим линкерную последовательность. В конкретном варианте осуществления линкерной последовательностью является расщепляемая линкерная последовательность Т2А, показанная в таблице 2. Иллюстративная полинуклеотидная последовательность, кодирующая линкер Т2А, представлена в таблице 1 (SEQ ID NO: 8).
Полинуклеотид, кодирующий маркерную последовательность, может быть без труда получен с помощью синтетических или рекомбинантных способов из аминокислотной последовательности. В вариантах осуществления полинуклеотид, кодирующий маркерную последовательность, функционально связан с полинуклеотидом, кодирующим внутриклеточный сигнальный домен. В вариантах осуществления полинуклеотид, кодирующий маркерную последовательность, может также иметь один или несколько сайтов для фермента рестрикции на 5'-и/или 3'-конце кодирующей последовательности для обеспечения легкого вырезания и замены полинуклеотида, кодирующего маркерную последовательность, другим полинуклеотидом, кодирующим другую маркерную последовательность. В вариантах осуществления полинуклеотид, кодирующий маркерную последовательность, подвергнут оптимизации кодонов для экспрессии в клетках млекопитающих.
Векторы, клетки и способы трансдукции клеток
Селекция и сортировка популяций Т-лимфоцитов
Композиции, описанные в данном документе, представляют Т-лимфоциты CD4+ и/или CD8+. Т-лимфоциты могут быть собраны в соответствии с известными методиками и подвергнуты обогащению или истощению с помощью известных методик, таких как аффинное связывание с антителами, таких как проточная цитометрия и/или иммуномагнитная селекция. После этапов обогащения и/или истощения может быть проведено in vitro размножение предпочтительных Т-лимфоцитов в соответствии с известными методиками (включая, без ограничения, методики, описанные в патенте США №6040177 Riddell et al.) или их вариантами, которые будут очевидны специалистам в данной области. В вариантах осуществления Т-клетки являются аутологическими Т-клетками, полученными от пациента.
Например, предпочтительную популяцию или субпопуляцию Т-клеток можно размножать путем добавления исходной популяции Т-лимфоцитов в среду для культивирования in vitro, и затем добавления в среду для культивирования питающих клеток, таких как неделящиеся мононуклеарные клетки периферической крови (РВМС) (например, так, что образующаяся популяция клеток содержит по меньшей мере 5, 10, 20 или 40 или более питающих РВМС-клеток для каждого Т-лимфоцита в исходной популяции, которую необходимо размножать); и инкубирования культуры (например, в течение времени, достаточного для размножения большого количества Т-клеток). Неделящиеся питающие клетки могут содержать облученные гамма-лучами питающие РВМС-клетки. В некоторых вариантах осуществления РВМС облучают гамма-лучами в диапазоне от приблизительно 3000 до 3600 рад для предотвращения деления клеток. При необходимости порядок добавления Т-клеток и питающих клеток в среду для культивирования может быть обратным. Культуру, как правило, можно инкубировать в таких условиях температуры и т.п., которые являются подходящими для роста Т-лимфоцитов. Для роста Т-лимфоцитов человека, например, температура обычно будет составлять по меньшей мере приблизительно 25 градусов Цельсия, предпочтительно приблизительно 30 градусов, более предпочтительно приблизительно 37 градусов.
Размножаемые Т-лимфоциты включают цитотоксические Т-лимфоциты CD8+ (CTL) и Т-лимфоциты-хелперы CD4+, которые могут быть специфичными к антигену, присутствующему в опухоли или патогене человека.
Необязательно, способ размножения может также включать этап добавления неделящихся EBV-трансформиррванных лимфобластоидных клеток (LCL) в качестве питающих клеток. LCL можно облучать гамма-лучами в диапазоне от приблизительно 6000 до 10000 рад. Питающие LCL-клетки могут быть представлены в любом подходящем количестве, как, например, в соотношении питающих LCL-клеток и исходных Т-лимфоцитов, составляющем по меньшей мере приблизительно 10:1.
Способ размножения необязательно может также включать этап добавления антитела к CD3 и/или антитела к CD28 в среду для культивирования (например, при концентрации по меньшей мере приблизительно 0,5 нг/мл). Способ размножения необязательно может также включать этап добавления IL-2 и/или IL-5 в среду для культивирования (например, где концентрация IL-2 составляет по меньшей мере приблизительно 10 единиц/мл).
После выделения Т-лимфоцитов как цитотоксические Т-лимфоциты, так и Т-лимфоциты-хелперы можно сортировать по субпопуляциям наивных Т-клеток, Т-клеток памяти и эффекторных Т-клеток до или после размножения.
Клетки CD8+ можно получить с помощью стандартных способов. В некоторых вариантах осуществления клетки CD8+ дополнительно сортируют по группам наивных клеток, центральных клеток памяти и эффекторных клеток памяти путем идентификации антигенов клеточной поверхности, которые ассоциированы с каждым из этих типов клеток CD8+. В вариантах осуществления Т-клетки памяти присутствуют как в субпопуляции CD62L+ , так и в субпопуляции CD62L-лимфоцитов периферической крови CD8+. После окрашивания антителами к CD8 и антителами к CD62L РВМС сортируют по фракциям CD62L- CD8+ и CD62L+ CD8+. В некоторых вариантах осуществления центральные клетки памяти ТСМ экспрессируют фенотипические маркеры, включающие GD45RO, CD62L, CCR7, CD28, CD3 и CD127, и являются отрицательными в отношении экспрессии гранзима В или характеризуются низким ее уровнем. В некоторых вариантах осуществления центральные Т-клетки памяти представляют собой Т-клетки CD8+ CD45RO+, CD62L+ . В некоторых вариантах осуществления эффекторные ТЕ являются отрицательными по CD62L, CCR7, CD28 и CD127 и положительными по гранзиму В и перфорину. В некоторых вариантах осуществления наивные Т-лимфоциты CD8+ характеризуются экспрессией фенотипических маркеров наивных Т-клеток, в том числе CD62L, CCR7, CD28, CD3, CD127 и CD45RA.
Определить, является ли клетка или популяция клеток положительной по определенному маркеру клеточной поверхности, можно посредством проточной цитометрии с помощью окрашивания антителом, специфичным к поверхностному маркеру, и изотипически сходным контрольным антителом. Популяцию клеток, отрицательную по маркеру, обозначает отсутствие значительного окрашивания популяции клеток специфичным антителом на уровне, превышающем таковой для изотопического контроля, положительную обозначает равномерное окрашивание популяции клеток на уровне, превышающем таковой для изотопического контроля. В некоторых вариантах осуществления снижение экспрессии одного из маркеров относится к потере 1 log 10 в средней интенсивности флуоресценции и/или снижению процентной доли клеток, характеризующихся экспрессией маркера, на по меньшей мере приблизительно 20% клеток, 25% клеток, 30% клеток, 35% клеток, 40% клеток, 45% клеток, 50% клеток, 55% клеток, 60% клеток, 65% клеток, 70% клеток, 75% клеток, 80% клеток, 85% клеток, 90% клеток, 95% клеток, и 100% клеток, и любой % от 20 до 100% по сравнению с эталонной популяцией клеток. В некоторых вариантах осуществления популяция клеток, положительная по одному из маркеров, относится к процентной доле клеток, характеризующихся экспрессией маркера, которая составляет по меньшей мере приблизительно 50% клеток, 55% клеток, 60% клеток, 65% клеток, 70% клеток, 75% клеток, 80% клеток, 85% клеток, 90% клеток, 95% клеток, и 100% клеток, и любой % от 50 до 100% по сравнению с эталонной популяцией клеток.
Т-клетки-хелперы CD4+ сортируют по группам наивных клеток, центральных клеток памяти и эффекторных клеток путем идентификации клеточных популяций, которые имеют антигены клеточной поверхности. Лимфоциты CD4+ можно получить с помощью стандартных способов. В некоторых вариантах осуществления наивными Т-лимфоцитами CD4+ являются Т-клетки CD4+ CD45RO-, CD45RA+, CD62L+ . В некоторых вариантах осуществления центральными клетками памяти CD4+ являются CD62L+ и CD45RO+. В некоторых вариантах осуществления эффекторными клетками CD4+ являются CD62L- и CD45RO-.
В вариантах осуществления популяции CD4+ и CD8+, которые являются антиген-специфическими, могут быть получены с помощью стимуляции наивных или антиген-специфических Т-лимфоцитов антигеном. Например, можно получить линии или клоны Т-клеток, являющихся антиген-специфическими по отношению к антигенам цитомегаловируса, путем выделения Т-клеток от инфицированных субъектов и стимуляции клеток in vitro этим же антигеном. Можно также применять наивные Т-клетки. В качестве мишеней для вызова Т-клеточных ответов можно использовать любое число антигенов из опухолевых клеток. В некоторых вариантах осуществления для лечения заболевания или нарушения, включающего солидную опухоль, гематологическую злокачественную опухоль, рак молочной железы или меланому, пригодны композиции для адоптивной клеточной иммунотерапии.
Модификация популяций Т-лимфоцитов
В некоторых вариантах осуществления может быть предпочтительным введение функциональных генов в Т-клетки для применения в иммунотерапии в соответствии с настоящим раскрытием. Например, введенный ген или гены могут повышать эффективность терапии путем повышения жизнеспособности и/или содействия функционированию перенесенных Т-клеток; или они могут представлять собой генетический маркер для обеспечения селекции и/или оценки in vivo выживаемости или миграции; или они могут иметь функции, которые улучшают безопасность иммунотерапии, например, делая клетку восприимчивой к отрицательной селекции in vivo, как описано в Lupton S.D. et al., Mol. and Cell Biol., 11:6 (1991); и Riddell et al., Human Gene Therapy 3:319-338 (1992); см. также публикации PCT/US 91/08442 и PCT/US 94/05601 Lupton et al., в которых описано применение бифункциональных селективных гибридных генов, полученных в результате слияния доминантного положительного селективного маркера с отрицательным селективным маркером. Это можно выполнить в соответствии с известными методиками (см., например, патент США №6040177 Riddell et al. в столбцах 14-17) или их вариантами, которые будут очевидными для специалистов в данной области на основании настоящего раскрытия.
В вариантах осуществления Т-клетки модифицированы химерными рецепторами, описанными в данном документе. В некоторых вариантах осуществления Т-клетки получают от субъекта, подлежащего лечению, в других вариантах осуществления лимфоциты получают от аллогенных доноров-людей, предпочтительно, от здоровых доноров-людей.
В некоторых вариантах осуществления химерные рецепторы содержат лиганд-связывающий домен, который специфически связывается с поверхностной молекулой опухолевой клетки, полипептидный спейсерный участок, трансмембранный домен и внутриклеточный сигнальный домен, описанные в данном документе. В вариантах осуществления лиганд-связывающим доменом является одноцепочечный фрагмент антитела (scFv), полученный из вариабельного участка тяжелой (VH) и вариабельного участка легкой (VL) цепей моноклонального антитела (mAb). Костимулирующие сигналы также могут передаваться химерным рецептором благодаря слиянию костимулирующего домена CD28 и/или 4-1ВВ с ζ-цепью CD3. Химерные рецепторы являются специфичными к молекулам клеточной поверхности независимо от HLA, что, таким образом, преодолевает ограничения распознавания для TCR, включающие рестрикцию по HLA и низкие уровни экспрессии HLA на опухолевых клетках.
Химерные рецепторы можно конструировать со специфичностью к любому маркеру клеточной поверхности с использованием антиген-связывающих фрагментов или вариабельных доменов антител, например, молекул антител. Антиген-связывающие молекулы могут быть соединены с одним или несколькими сигнальными модулями. В вариантах осуществления сигнальные модули клеток включают трансмембранный домен CD3, внутриклеточные сигнальные домены CD3 и трансмембранные домены CD28. В вариантах осуществления внутриклеточный сигнальный домен содержит трансмембранный и сигнальный домены CD28, соединенные с внутриклеточным доменом СD3-зета. В некоторых вариантах осуществления химерный рецептор может также содержать маркер трансдукции, такой как tEGFR.
В вариантах осуществления в каждую из популяций Т-лимфоцитов CD4+ и CD8+ можно вводить один и тот же или разные химерные рецепторы. В вариантах осуществления химерный рецептор в каждой из этих популяций имеет лиганд-связывающий домен, который специфически связывается с одним и тем же лигандом на опухолевой или инфицированной клетке. Сигнальные модули клетки могут различаться. В вариантах осуществления внутриклеточный сигнальный домен цитотоксических Т-клеток CD8+ и внутриклеточный сигнальный домен Т-клеток-хелперов CD4+ являются одинаковыми. В других вариантах осуществления внутриклеточный сигнальный домен цитотоксических Т-клеток CD8+ отличается от внутриклеточного сигнального домена Т-клеток-хелперов CD4+.
В вариантах осуществления каждый из Т-лимфоцитов CD4+ или CD8+ можно сортировать по группам наивных клеток, центральных клеток памяти, эффекторных клеток памяти или эффекторных клеток перед трансдукцией, как описано в данном документе. В альтернативных вариантах осуществления каждый из Т-лимфоцитов CD4+ или CD8+ можно сортировать по группам наивных клеток, центральных клеток памяти, эффекторных клеток памяти или эффекторных клеток после трансдукции.
Были разработаны различные методики трансдукции, в которых используют рекомбинантные инфекционные вирусные частицы для доставки генов. Они представляют современный предпочтительный подход к трансдукции Т-лимфоцитов по настоящему изобретению. Вирусные векторы, которые применяли в этом направлении, включают вирусные векторы, полученные из вируса обезьян 40, аденовирусов, аденоассоциирдванного вируса (AAV), лентивирусных векторов и ретровирусов. Таким образом, способы переноса и экспрессии генов многочисленны, но главным образом используются для введения генетического материала в клетки млекопитающих и экспрессии его там. Некоторые из упомянутых выше методик применяли для трансдукции гемопоэтических или лимфоидных клеток, включая кальций-фосфатную трансфекцию, слияние протопластов, электропорацию и инфицирование рекомбинантным аденовирусом, аденоассоциированным вирусом и ретровирусными векторами. Первичные Т-лимфоциты успешно трансдуцировали с помощью электропорации и инфицирования ретровирусом или лентивирусом.
Ретровирусные и лентивирусные векторы обеспечивают высокоэффективный способ переноса генов в эукариотические клетки. Кроме того, интеграция ретровируса или лентивируса происходит контролируемо и приводит к стабильной интеграции одной или нескольких копий новой генетической информации в каждую клетку.
Предполагается, что сверхэкспрессия стимулирующего фактора (например, лимфокина или цитокина) может быть токсической для обрабатываемого индивида. Таким образом, включение генных сегментов, которые вызывают восприимчивость Т-клеток по настоящему изобретению к отрицательной селекции in vivo, находится в пределах объема настоящего изобретения. Под "отрицательной селекцией" подразумевается то, что инфузированная клетка может быть удалена в результате изменения состояния индивида in vivo. Отрицательный селективный фенотип может образовываться в результате вставки гена, который придает чувствительность к вводимому средству, например, соединению. Отрицательные селективные гены известны в данной области техники и включают, среди прочего, следующие: ген тимидинкиназы вируса герпеса типа I (ТK HSV-1), который придает чувствительность к ганцикловиру; ген клеточной гипоксантинфосфорибозилтрансферазы (HPRT), ген клеточной аденинфосфорибозилтрансферазы (APRT) и ген цитозиндезаминазы бактерий.
В некоторых вариантах осуществления может быть пригодным включение в Т-клетки положительного маркера, который обеспечивает селекцию клеток с отрицательным селективным фенотипом in vitro. Положительным селективным маркером может быть ген, который при введении в клетку-хозяина экспрессирует доминантный фенотип, обеспечивающий положительную селекцию клеток, несущих этот ген. Гены этого типа известны в данной области техники и включают, среди прочих, ген гигромицин-В-фосфотрансферазы (hph), который придает устойчивость к гигромицину В, ген аминогликозидфосфотрансферазы (neо или aph) из Тn5, который отвечает за устойчивость к антибиотику G418, ген дигидрофолатредуктазы (DHFR), ген аденозиндезаминазы (ADA) и ген множественной лекарственной устойчивости (MDR).
Для трансдукции Т-лимфоцитов можно использовать ряд способов, хорошо известных в данной области техники. В вариантах осуществления трансдукцию проводят с помощью лентивирусных векторов.
В вариантах осуществления для образования определенных популяций каждая из групп клеток CD4+ и CD8+ может быть модифицирована по отдельности с помощью вектора экспрессии, кодирующего химерный рецептор. В вариантах осуществления эти клетки затем дополнительно сортируют по субпопуляциям наивных клеток, центральных клеток памяти и эффекторных клеток, как описано выше, путем сортировки относительно антигенов клеточной поверхности, уникальных для каждой из этих клеточных популяций. Кроме того, среди популяций клеток CD4+ или CD8+ можно производить отбор по их цитокиновому профилю или пролиферативной активности. Например, можно отбирать Т-лимфоциты CD4+, которые характеризуются повышенной выработкой цитокинов, таких как IL-2, IL-4, IL-10, TNFα и IFNγ, по сравнению с клетками после ложной трансдукции или трансдуцированными клетками CD8+ при стимуляции антигеном. В других вариантах осуществления отбирают наивные Т-клетки или центральные Т-клетки памяти CD4+, которые характеризуются повышенной выработкой IL-2 и/или TNFα. Подобным образом отбирают клетки CD8+, которые характеризуются повышенной выработкой IFNγ по сравнению с клетками CD8+ после ложной трансдукции.
В вариантах осуществления отбирают клетки CD4+ и CD8+, которые пролиферируют в ответ на воздействие антигенов-мишеней или опухолей-мишеней. Например, отбирают клетки CD4+, которые интенсивно пролиферируют при стимуляции антигенами-мишенями или опухолями-мишенями по сравнению с клетками после ложной трансдукции или трансдуцированными клетками CD8+. В некоторых вариантах осуществления отбирают клетки CD4+ и CD8+, которые являются цитотоксическими для клеток, несущих антиген. В вариантах осуществления предполагается, что клетки CD4+ характеризуются слабой цитотоксичностью по сравнению с клетками CD8+.
В предпочтительном варианте осуществления отбирают трансдуцированные лимфоциты, такие как центральные клетки памяти CD8+, которые обеспечивают уничтожение опухолевых клеток in vivo при использовании животной модели, созданной для определенного типа рака. Такие животные модели хорошо известны специалистам в данной области, и они не включают людей. Как описано в данном документе, не все конструкты химерных рецепторов, трансдуцированные в лимфоциты, придают способность к уничтожению опухолевых клеток in vivo, несмотря на способность к активации и уничтожению опухолевых клеток in vitro. В частности, для некоторых молекул-мишеней Т-клетки, имеющие конструкты химерных рецепторов с длинным спейсерным участком, были менее эффективными в уничтожении опухолевых клеток in vivo по сравнению с Т-клетками, имеющими химерный рецептор с коротким спейсерным участком. Для других молекул-мишеней Т-клетки, имеющие конструкты химерных рецепторов с коротким спейсерным участком, были менее эффективными в уничтожении опухолевых клеток in vivo по сравнению с Т-клетками, имеющими химерные рецепторы с длинным спейсерным участком.
В еще нескольких вариантах осуществления отбирают трансдуцированные Т-клетки, экспрессирующие химерные рецепторы, которые могут персистировать in vivo при использовании животной модели, созданной для определенного типа рака. В вариантах осуществления было показано, что центральные клетки памяти CD8+, трансдуцированные химерными рецепторами с коротким спейсерным участком, персистируют in vivo после введения животному в течение приблизительно 3 дней или более, 10 дней или более, 20 дней или более, 30 дней или более, 40 дней или более или 50 дней или более.
В настоящем раскрытии предполагается, что в композициях будут использоваться комбинации Т-клеток CD4+ и CD8+. В одном варианте осуществления комбинации клеток CD4+, трансдуцированных химерными рецепторами, можно комбинировать с клетками CD8+, трансдуцированными химерными рецепторами, с такой же специфичностью к лигандам или комбинировать с Т-клетками CD8+, характеризующимися специфичностью к другому опухолевому лиганду. В других вариантах осуществления клетки CD8+, трансдуцированные химерными рецепторами, комбинируют с клетками CD4+, трансдуцированными химерными рецепторами, специфичными к другому лиганду, экспрессируемому в опухоли. В еще одном варианте осуществления комбинируют клетки CD4+ и CD8+, модифицированные химерным рецептором. В вариантах осуществления клетки CD8+ и CD4+ можно комбинировать в различных соотношениях, например, в соотношении CD8+ и CD4+ 1:1, в соотношении CD8+ и CD4+ 10:1 или в соотношении CD8+ и CD4+ 100:1. В вариантах осуществления комбинированную популяцию исследуют в отношении пролиферации клеток in vitro и/или in vivo, и выбирают соотношение клеток, которое обеспечивает пролиферацию клеток.
Как описано в данном документе, в настоящем раскрытии предполагается, что клетки CD4+ и CD8+ можно дополнительно разделить на субпопуляции, такие как популяции наивных клеток, центральных клеток памяти и эффекторных клеток памяти. Как описано в данном документе, в некоторых вариантах осуществления наивные клетки CD4+ являются Т-клетками CD4+ CD45RO-, CD45RA+, CD62L+ . В некоторых вариантах осуществления центральные клетки памяти CD4+ являются СD62b-положительными и СБ45RО-положительными. В некоторых вариантах осуществления эффекторные клетки CD4+ являются СD62L-отрицательными и СD45RО-положительными. Каждая из этих популяций может быть независимо модифицирована химерным рецептором.
Как описано в данном документе, в вариантах осуществления Т-клетки памяти присутствуют как в субпопуляции CD62L+ , так и в субпопуляции CD62L-лимфоцитов периферической крови CD8+. После окрашивания антителами к CD8 и антителами к CD62L РВМС сортируют по фракциям CD62L-CD8+ и CD62L+ CD8+. В некоторых вариантах осуществления центральные Т-клетки памяти (ТСM) экспрессируют фенотипические маркеры, включающие CD62L, CCR7, CD28, CD3 и CD127, и являются отрицательными в отношении экспрессии гранзима В или характеризуются низким ее уровнем. В некоторых вариантах осуществления центральные Т-клетки памяти представляют собой Т-клетки CD8+ CD45RO+, CD62L+ . В некоторых вариантах осуществления эффекторные Т-клетки (ТЕ) являются отрицательными по CD62L, CCR7, CD28 и CD127 и положительными по гранзиму В и перфорину. В некоторых вариантах осуществления наивные Т-лимфоциты CD8+ характеризуются CD8+, CD62L+ , CD45RO+, CCR7+, CD28+, CD17+ и CD45RO+. Каждая из этих популяций может быть независимо модифицирована химерным рецептором.
После трансдукции и/или отбора клеток, несущих химерные рецепторы, популяции клеток предпочтительно размножают in vitro до получения достаточного числа клеток для обеспечения по меньшей мере одной инфузии субъекту-человеку, как правило, приблизительно от 104 клеток/кг до 109 клеток/кг. В вариантах осуществления трансдуцированные клетки культивируют в присутствии клеток, несущих антиген, антител к CD3, антител к CD28 и IL-2, IL-7, IL-15, IL-21 и их комбинаций.
Каждую из субпопуляций клеток CD4+ и CD8+ можно комбинировать друг с другом. В конкретном варианте осуществления модифицированные наивные клетки или центральные клетки памяти CD4+ комбинируют с модифицированными центральными клетками памяти CD8+ для обеспечения синергического цитотоксического эффекта в отношении клеток, несущих антиген, таких как опухолевые клетки.
Композиции
Настоящее раскрытие предлагает композицию для адоптивной клеточной иммунотерапии, содержащую клеточный препарат генетически модифицированных Т-лимфоцитов, описанных в данном документе.
В вариантах осуществления клеточный препарат Т-лимфоцитов содержит Т-клетки CD4+, которые имеют химерный рецептор, содержащий внеклеточный вариабельный домен антитела, специфичный к лиганду, ассоциированному с заболеванием или нарушением, индивидуализируемый спейсерный участок, трансмембранный домен и внутриклеточный сигнальный домен Т-клеточного рецептора или других рецепторов, описанных в данном документе. В других вариантах осуществления композиция для адоптивной клеточной иммунотерапии дополнительно содержит клеточный препарат опухолеспецифических цитотоксических Т-лимфоцитов CD8+, модифицированных химерным рецептором, который обеспечивает клеточный иммунный ответ, где клеточный препарат цитотоксических Т-лимфоцитов содержит Т-клетки CD8+, которые имеют химерный рецептор, содержащий внеклеточное одноцепочечное антитело, специфичное к лиганду, ассоциированному с заболеванием или нарушением, индивидуализируемый спейсерный участок, трансмембранный домен и внутриклеточный сигнальный домен Т-клеточного рецептора, описанные в данном документе. В вариантах осуществления популяция Т-клеток, модифицированных химерными рецепторами, по настоящему раскрытию может персистировать in vivo в течение по меньшей мере приблизительно 3 дней или дольше.
В некоторых вариантах осуществления композиция для адоптивной клеточной иммунотерапии содержит клеточный препарат опухолеспецифических цитотоксических Т-лимфоцитов CD8+, модифицированных химерным рецептором, который обеспечивает клеточный иммунный ответ, где клеточный препарат цитотоксических Т-лимфоцитов содержит Т-клетки CD8+, которые имеют химерный рецептор, содержащий внеклеточное одноцепочечное антитело, специфичное к лиганду, ассоциированному с заболеванием или нарушением, индивидуализируемый спейсерный участок, трансмембранный домен и внутриклеточный сигнальный домен Т-клеточного рецептора, в комбинации с модифицированными химерным рецептором антиген-реактивными наивными Т-клетками CD4+, полученными из Т-клеток CD4+ CD45RO- CD62L+ , и фармацевтически приемлемым носителем.
В других вариантах осуществления композиция для адоптивной клеточной иммунотерапии содержит клеточный препарат антиген-специфических цитотоксических Т-лимфоцитов CD8+, полученных от пациента, который обеспечивает клеточный иммунный ответ, в комбинации с антиген-реактивными наивными Т-клетками-хелперами CD4+, модифицированными химерным рецептором, которые усиливают иммунный ответ, опосредованный CD8+, при этом клеточный препарат Т-лимфоцитов-хелперов содержит Т-клетки CD4+, которые имеют химерный рецептор, содержащий внеклеточный вариабельный домен антитела, специфичный к лиганду, ассоциированному с заболеванием или нарушением, индивидуализируемый спейсерный участок, трансмембранный домен и внутриклеточный сигнальный домен Т-клеточного рецептора. В дополнительном варианте осуществления композиция для адоптивной клеточной иммунотерапии содержит антиген-реактивные наивные Т-клетки-хелперы CD4+, модифицированные химерным рецептором, которые усиливают иммунный ответ, опосредованный CD8+, при этом клеточный препарат Т-лимфоцитов-хелперов содержит Т-клетки CD4+, которые имеют химерный рецептор, содержащий внеклеточный вариабельный домен антитела, специфичный к лиганду, ассоциированному с заболеванием или нарушением, индивидуализируемый спейсерный участок, трансмембранный домен и внутриклеточный сигнальный домен Т-клеточного рецептора.
В вариантах осуществления Т-лимфоцит-хелпер CD4+ выбирают из группы, включающей наивные Т-клетки CD4+, центральные Т-клетки памяти CD4+, эффекторные Т-клетки памяти CD4+ или общую популяцию Т-клеток CD4+. В некоторых вариантах осуществления лимфоцит-хелпер CD4+ представляет собой наивную Т-клетку CD4+, при этом наивная Т-клетка CD4+ включает в себя Т-клетку CD4+ CD45RO-, CD45RA+, CD62L+ . В вариантах осуществления цитотоксический Т-лимфоцит CD8+ выбирают из группы, включающей наивные Т-клетки CD8+, центральные Т-клетки памяти CD8+, эффекторные Т-клетки памяти CD8+ или общую популяцию Т-клеток CD8+. В некоторых вариантах осуществления цитотоксический Т-лимфоцит CD8+ представляет собой центральную Т-клетку памяти, при этом центральная Т-клетка памяти включает в себя Т-клетку CD8+ CD45RO+, CD62L+ . В еще нескольких вариантах осуществления цитотоксический Т-лимфоцит CD8+ представляет собой центральную Т-клетку памяти, а Т-лимфоцит-хелпер CD4+ представляет собой наивную Т-клетку или центральную Т-клетку памяти CD4+.
Способы
Настоящее раскрытие предлагает способы получения композиций для адоптивной иммунотерапии и пути применения или способы применения этих композиций для проведения клеточной иммунотерапии у субъекта, имеющего заболевание или нарушение. В вариантах осуществления Т-клетки, модифицированные химерными рецепторами, описанные в данном документе, способны персистировать in vivo в течение по меньшей мере 3 дней или по меньшей мере 10 дней. В вариантах осуществления Т-клетки, модифицированные химерными рецепторами, описанные в данном документе, могут пролиферировать in vivo на протяжении по меньшей мере 2 или по меньшей мере 3 поколений, как определяется с помощью разведения красителя CFSE. Пролиферацию и персистирование Т-клеток, модифицированных химерными рецепторами, можно определить с помощью животной модели заболевания или нарушения, а также путем введения клеток и определения персистирования и/или пролиферативной способности перенесенных клеток. В других вариантах осуществления пролиферацию и активацию можно исследовать in vitro путем выполнения нескольких циклов активации с помощью клеток, несущих антиген.
В вариантах осуществления способ производства композиций включает получение модифицированной наивной Т-клетки-хелпера CD4+, при этом клеточный препарат модифицированных Т-лимфоцитов-хелперов содержит Т-клетки CD4+, которые имеют химерный рецептор, содержащий лиганд-связывающий домен, специфичный к поверхностной молекуле опухолевой клетки, индивидуальный спейсерный домен, трансмембранный домен и внутриклеточный сигнальный домен, описанные в данном документе.
В другом варианте осуществления способ дополнительно включает получение модифицированных цитотоксических Т-клеток CD8+, при этом клеточный препарат модифицированных цитотоксических Т-лимфоцитов содержит клетки CD8+, которые имеют химерный рецептор, содержащий лиганд-связывающий домен, специфичный к поверхностной молекуле опухолевой клетки, индивидуальный спейсерный домен, трансмембранный домен и внутриклеточный сигнальный домен, описанные в данном документе.
В другом варианте осуществления способ включает получение модифицированной цитотоксической Т-клетки CD8+, при этом клеточный препарат модифицированных цитотоксических Т-лимфоцитов содержит Т-клетки CD8+, которые имеют химерный рецептор, содержащий лиганд-связывающий домен, специфичный к поверхностной молекуле опухолевой клетки, индивидуальный спейсерный домен, трансмембранный домен и внутриклеточный сигнальный домен, описанные в данном документе, и дополнительно включает комбинирование модифицированных цитотоксических Т-клеток CD8+ с клеточным препаратом лимфоцитов-хелперов CD4+.
Препарат клеток CD4+ и CD8+, которые модифицированы химерным рецептором, описан выше, а также в примерах. Антиген-специфические Т-лимфоциты могут быть получены от пациента, имеющего заболевание или нарушение, или могут быть получены с помощью in vitro стимуляции Т-лимфоцитов в присутствии антигена. Субпопуляции Т-лимфоцитов CD4+ и CD8+, среди которых не был произведен отбор по специфичности к антигену, могут также быть выделены, как описано в данном документе, и комбинированы в способах производства. В вариантах осуществления комбинацию клеточных популяций можно оценивать в отношении однородности маркеров клеточной поверхности, способности к пролиферации на протяжении по меньшей мере двух поколений, наличия однородного дифференцировочного статуса клеток. Контроль качества можно выполнять путем совместного культивирования линии клеток, экспрессирующей лиганд-мишень, с Т-клетками, модифицированными химерными рецепторами, для определения того, распознают ли Т-клетки, модифицированные химерными рецепторами, линию клеток, с помощью анализов цитотоксичности, пролиферации или выработки цитокинов, которые известны в данной области. Дифференцировочный статус клеток и маркеры клеточной поверхности в Т-клетках, модифицированных клеточными рецепторами, могут быть определены с помощью проточной цитометрии. В вариантах осуществления маркеры и дифференцировочный статус клеток в клетках CD8+ включают CD3, CD8, CD62L, CD28, CD27, CD69, CD25, PD-1, CTLA-4, CD45RO и CD45RA. В вариантах осуществления маркеры дифференцировочный статус клеток в клетках CD4+ включают CD3, CD4, CD62L, CD28, CD27, CD69, CD25, PD-1, CTLA-4, CD45RO и CD45RA.
В вариантах осуществления в данном документе предлагается способ выбора спейсерного участка для химерного рецептора. Некоторые конструкты химерных рецепторов, несмотря на эффективность в отношении активации Т-клеток in vitro, не были эффективными in vivo. В вариантах осуществления способ включает обеспечения множества нуклеиновых кислот, кодирующих химерные рецепторы, где химерные рецепторы, кодируемые нуклеиновыми кислотами, отличаются только спейсерным участком; введение каждой из нуклеиновых кислот, кодирующих химерные рецепторы, в отдельную популяцию Т-лимфоцитов; размножение каждой отдельной популяции лимфоцитов in vitro, и введение каждой популяции лимфоцитов животному-опухоленосителю, для определения противоопухолевой эффективности каждой из Т-клеток, модифицированных химерными рецепторами, и выбор химерного рецептора, который обеспечивает противоопухолевую эффективность по сравнению с каждой из других отдельных популяций лимфоцитов, модифицированных Т-клетками, модифицированными каждым из других химерных рецепторов.
Известны животные модели различных опухолей. Противоопухолевую эффективность можно измерить путем определения снижения объема опухоли, путем определения смертности животных, персистирования генетически модифицированных Т-клеток in vivo, активации генетически модифицированных Т-клеток (например, путем выявления повышения экспрессии CD25 и/или CD69) и/или пролиферации генетически модифицированных Т-клеток in vivo. В варианте осуществления выбирают химерный рецептор, который обеспечивает наилучшую противоопухолевую эффективность in vivo, определенную с помощью одного или нескольких из этих параметров. Отсутствие противоопухолевой эффективности можно определить по отсутствию персистирования генетически модифицированных лимфоцитов in vivo, гибели животных, увеличению интенсивности апоптоза, измеренному по повышению индукции каспазы-3, и/или снижению пролиферации генетически модифицированных лимфоцитов.
В вариантах осуществления обеспечение множества нуклеиновых кислот, кодирующих химерные рецепторы, где химерные рецепторы, кодируемые нуклеиновыми кислотами, отличаются только спейсерным участком, включает обеспечение конструкта химерного рецептора, содержащего полинуклеотид, кодирующий лиганд-связывающий домен, где лигандом является опухолеспецифический антиген, вирусный антиген или любая другая молекула, экспрессируемая в популяции клеток-мишеней, пригодная для опосредования распознавания и удаления лимфоцитом; полинуклеотид, кодирующий первый полипептидный спейсер, имеющий определенный сайт рестрикции на 5'- и 3'-концах кодирующей последовательности для первого полипептидного спейсера; полинуклеотид, кодирующий трансмембранный домен; и полинуклеотид, кодирующий внутриклеточный сигнальный домен.
Настоящее раскрытие также предлагает способы проведения клеточной иммунотерапии у субъекта, имеющего заболевание или нарушение, включающие введение композиции лимфоцитов, экспрессирующих химерный рецептор, описанный в данном документе. В других вариантах осуществления способ включает введение субъекту клеточного препарата генетически модифицированных цитотоксических Т-лимфоцитов, который обеспечивает клеточный иммунный ответ, при этом клеточный препарат цитотоксических Т-лимфоцитов содержит Т-клетки CD8+, которые имеют химерный рецептор, содержащий лиганд-связывающий домен, специфичный к поверхностной молекуле опухолевой клетки, индивидуальный спейсерный домен, трансмембранный домен и внутриклеточный сигнальный домен, описанные в данном документе, и клеточного препарата генетически модифицированных Т-лимфоцитов-хелперов, который вызывает прямое распознавание опухоли и усиливает способность клеточных препаратов генетически модифицированных цитотоксических Т-лимфоцитов опосредовать клеточный иммунный ответ, при этом клеточный препарат Т-лимфоцитов-хелперов содержит Т-клетки CD4+, которые имеют химерный рецептор, содержащий лиганд-связывающий домен, специфичный к поверхностной молекуле опухолевой клетки, индивидуальный спейсерный домен, трансмембранный домен и внутриклеточный домен, описанные в данном документе.
Не ограничивая объем настоящего раскрытия, считают, что выбор популяции Т-клеток, модифицированных химерными рецепторами, которые могут персистировать и пролиферировать in vivo до введения, может приводить к возможности применения более низкой дозы Т-клеток и обеспечения более однородной терапевтической активности. В вариантах осуществления дозы Т-клеток могут быть снижены по меньшей мере на 10%, 20% или 30% или больше. Снижение дозы Т-клеток может быть благоприятным для снижения риска развития синдрома лизиса опухоли и цитокинового шторма.
В другом варианте осуществления способ проведения клеточной иммунотерапии у субъекта, имеющего заболевание или нарушение, включает введение субъекту клеточного препарата генетически модифицированных Т-лимфоцитов-хелперов, при этом клеточный препарат модифицированных Т-лимфоцитов-хелперов содержит Т-клетки CD4+, которые имеют химерный рецептор, содержащий лиганд-связывающий домен, специфичный к поверхностной молекуле опухолевой клетки, индивидуальный спейсерный домен, трансмембранный домен и внутриклеточный сигнальный домен, описанные в данном документе. В вариантах осуществления способ дополнительно включает введение субъекту клеточного препарата генетически модифицированных цитотоксических Т-лимфоцитов, при этом клеточный препарат модифицированных цитотоксических Т-лимфоцитов содержит клетки CD8+, которые имеют химерный рецептор, содержащий лиганд-связывающий домен, специфичный к поверхностной молекуле опухолевой клетки, индивидуальный спейсерный домен, трансмембранный домен и внутриклеточный сигнальный домен, описанные в данном документе.
В другом варианте осуществления описан способ проведения клеточной иммунотерапии у субъекта, имеющего заболевание или нарушение, включающий анализ биологического образца субъекта на наличие молекулы-мишени, ассоциированной с заболеванием или нарушением, и введение композиций для адоптивной иммунотерапии, описанных в данном документе, в котором химерный рецептор специфически связывается с молекулой-мишенью.
В некоторых вариантах осуществления Т-лимфоцит-хелпер CD4+ выбирают до введения химерного рецептора из группы, включающей наивные Т-клетки CD4+, центральные Т-клетки памяти CD4+, эффекторные Т-клетки памяти CD4+ или общую популяцию Т-клеток CD4+. В конкретном варианте осуществления лимфоцит-хелпер CD4+ представляет собой наивную Т-клетку CD4+, при этом наивная Т-клетка CD4+ включает в себя Т-клетку CD4+ CD45RO-, CD45RA+, CD62L+ . В еще нескольких вариантах осуществления цитотоксический Т-лимфоцит CD8+ выбирают до введения химерного рецептора из группы, включающей наивные Т-клетки CD8+, центральные Т-клетки памяти CD8+, эффекторные Т-клетки памяти CD8+ или общую популяцию Т-клеток CD8+. В конкретном варианте осуществления цитотоксический Т-лимфоцит CD8+ представляет собой центральную Т-клетку памяти, при этом центральная Т-клетка памяти включает в себя Т-клетку CD8+ CD45RO+, CD62L+ . В конкретном варианте осуществления цитотоксический Т-лимфоцит CD8+ представляет собой центральную Т-клетку памяти, а Т-лимфоцит-хелпер CD4+ представляет собой наивную Т-клетку.
В вариантах осуществления как Т-клетка CD8+, так и Т-клетка CD4+ являются генетически модифицированными химерным рецептором, содержащим домен тяжелой цепи антитела, который специфически связывается с опухолеспецифической молекулой клеточной поверхности. В других вариантах осуществления внутриклеточный сигнальный домен цитотоксических Т-клеток CD8+ и внутриклеточный сигнальный домен Т-клеток-хелперов CD4+ являются одинаковыми. В еще нескольких вариантах осуществления внутриклеточный сигнальный домен цитотоксических Т-клеток CD8+ отличается от внутриклеточного сигнального домена Т-клеток-хелперов CD4+.
Субъекты, которых можно лечить при помощи настоящего изобретения, являются, как правило, людьми и другими субъектами-приматами, такими как обезьяны и высшие обезьяны в контексте ветеринарной медицины. Субъекты могут быть самцами или самками и могут быть любого подходящего возраста, в том числе детенышами, подростками, молодыми особями, взрослыми и пожилыми субъектами.
Способы являются пригодными в лечении, например, гематологической злокачественной опухоли, меланомы, рака молочной железы и других эпителиальных злокачественных образований или солидных опухолей. В некоторых вариантах осуществления молекулу, ассоциированную с заболеванием или нарушением, выбирают из группы, включающей орфанный тирозинкиназный рецептор ROR1, Her2, CD19, CD20, CD22, мезотелин, СЕА и поверхностный антиген вируса гепатита В.
Субъекты, которых можно лечить, включают субъектов, пораженных раком, в том числе, без ограничения, раком кишечника, легких, печени, молочной железы, почек, предстательной железы, яичников, кожи (включая меланому), костей и мозга и т.п. В некоторых вариантах осуществления известны антигены или молекулы, ассоциированные с опухолью, такой как меланома, рак молочной железы, плоскоклеточная карцинома, рак кишечника, лейкоз, миелома и рак предстательной железы. В других вариантах осуществления на молекулы, ассоциированные с опухолью, могут нацеливаться генетически модифицированные Т-клетки, экспрессирующие сконструированный химерный рецептор. Примеры включают, без ограничения, В-клеточную лимфому, рак молочной железы, рак предстательной железы и лейкоз.
Клетки, полученные, как описано выше, можно использовать в способах и композициях для адоптивной иммунотерапии в соответствии с известными методиками или их вариантами, которые будет очевидны специалистам в данной области на основании настоящего раскрытия.
В некоторых вариантах осуществления получают состав на основе клеток, вначале собирая их из среды для культивирования, а затем отмывая и концентрируя клетки в среде и системе содержания, подходящей для введения ("фармацевтически приемлемом" носителе), в эффективном для лечения количестве. Подходящей средой для инфузии может быть любой состав в виде изотонической среды, как правило, физиологический раствор, Normosol R (Abbott) или Plasma-Lyte А (Baxter), но также можно использовать и 5% водный раствор декстрозы или лактат Рингера. Среда для инфузии может быть дополнена сывороточным альбумином человека, фетальной бычьей сывороткой или другими компонентами сыворотки человека.
Эффективное для лечения количество клеток в композиции составляет по меньшей мере 2 субпопуляции клеток (например, 1 субпопуляция центральных Т-клеток памяти CD8+ и 1 субпопуляция Т-клеток-хелперов CD4+) или, более типично, более 102 клеток, и до 106, до 108 или 109 клеток включительно, и может составлять более 1010 клеток. Число клеток будет зависеть от конечного применения, для которого предполагается композиция, а также от типа клеток, включенных в нее. Например, если предпочтительными являются клетки, специфичные к определенному антигену, то популяция будет содержать более 70%, как правило, более 80%, 85% и 90-95% таких клеток. Для путей применения, представленных в данном документе, клетки, как правило, содержатся в объеме, который составляет один литр или менее, может составлять 500 мл или менее и даже 250 мл или 100 мл или менее. Поэтому плотность предпочтительных клеток составляет, как правило, более 104 клеток/мл и, как правило, более 107 клеток/мл, как правило, 108 клеток/мл или более. Клинически значимое количество иммунных клеток может быть распределено на несколько инфузий и в сумме составляет или превышает 106, 107, 108, 109, 1010 или 1011 клеток.
В некоторых вариантах осуществления лимфоциты по настоящему изобретению можно применять для обеспечения иммунитета индивидам. Под "иммунитетом" подразумевается уменьшение проявлений одного или нескольких физических симптомов, ассоциированных с отсветом на инфекцию, вызываемую патогеном, или на опухоль, на которые направлен ответ лимфоцитов. Количество вводимых клеток обычно находится в диапазоне, присутствующем у нормальных индивидов с иммунитетом к данному патогену. Таким образом, клетки обычно вводят посредством инфузии, при этом количество клеток, вводимых с каждой инфузией, находится в диапазоне от 2 клеток до по меньшей мере от 106 до 3×1010 клеток, предпочтительно в диапазоне от по меньшей мере 107 до 109 клеток. Т-клетки могут быть введены посредством однократной инфузии или посредством многократных инфузий в течение определенного промежутка времени. Однако, поскольку предполагается, что различные индивиды различаются по отвечаемости, тип и количество инфузированных клеток, а также число инфузий и промежуток времени, в течение которого осуществляют многократные инфузии, устанавливаются лечащим врачом и могут быть определены путем стандартного обследования. Получение достаточных уровней Т-лимфоцитов (в том числе цитотоксических Т-лимфоцитов и/или Т-лимфоцитов-хелперов) легко достигается с помощью способа быстрого размножения по настоящему изобретению, как показано на примерах в данном документе. См., например, патент США №6040177 Riddell et al. в столбце 17.
В вариантах осуществления композицию, описанную в данном документе, вводят внутривенно, внутрибрюшинно, внутрь опухоли, в костный мозг, в лимфатический узел и/или в спинномозговую жидкость. В вариантах осуществления композиции сконструированных химерных рецепторов доставляют в место локализации опухоли. Альтернативно, композиции, описанные в данном документе, можно комбинировать с соединением, которое направляет клетки к опухоли или компартментам иммунной системы и избегает таких мест, как легкие.
В вариантах осуществления композиции, описанные в данном документе, вводят с химиотерапевтическими средствами и/или иммунодепрессантами. В варианте осуществления пациента обрабатывают сначала химиотерапевтическим средством, которое подавляет функции других иммунных клеток или разрушает их, а затем композициями, описанными в данном документе. В некоторых случаях химиотерапии можно избежать полностью.
Настоящее изобретение дополнительно проиллюстрировано в примерах, изложенных ниже.
ЭКСПЕРИМЕНТАЛЬНАЯ ЧАСТЬ
Пример 1. Индивидуализация длины спейсерного домена и аффинности scFv для оптимального распознавания ROR1 Т-клетками, модифицированными химерными рецепторами
Были сконструированы химерные рецепторы, специфичные к молекуле ROR1, которая экспрессируется в большом числе злокачественных опухолей человека, включающих хронический лимфоцитарный лейкоз, мантийноклеточную лимфому, острый лимфобластный лейкоз, а также рак молочной железы, легких, предстательной железы, поджелудочной железы и яичников. Химерные рецепторы ROR1 были разработаны из ROR1-специфических scFv с различными аффинностями и содержали внеклеточные спейсерные домены Fc IgG4 разной длины. Анализировали способность Т-клеток, экспрессирующих каждый ROR1-специфический рецептор, к распознаванию гемопоэтических и эпителиальных опухолей RO1+ in vitro и устранению мантийноклеточной лимфомы, трансплантированной мышам с иммунодефицитом.
Материалы и способы
Субъекты-люди
Мононуклеарные клетки периферической крови (РВМС) получали от здоровых доноров и пациентов после письменного информированного согласия на участие в протоколах исследования, утвержденных ученым советом Онкологического исследовательского центра Фреда Хатчинсона (FHCRC).
Линии клеток
Линии клеток K562, Raji, JeKo-1, MDA-MB-231, MDA-MB-468 и 293T получали из Американской коллекции типовых культур. Д-ром Эдусом X. Уорреном (FHCRC) были любезно предоставлены линии клеток почечно-клеточного рака FARP, TREP и RWL. K562/ROR1 и Raji/ROR1 получали с помощью трансдукции лентивирусом с геном ROR1 полной длины. Для получения JeKo-1/ffluc нативные клетки JeKo-1 трансдуцировали лентивирусным вектором, содержащим ген люциферазы светлячка (ffluc), расположенный выше последовательности Т2А и eGFP. Трансдуцированные клетки JeKo-1 сортировали относительно экспрессии eGFP и размножали для экспериментов in vivo.
Иммунофенотипирование
РВМС и линии клеток окрашивали следующими конъюгированными mAb: CD3, CD4, CD5, CD8, CD19, CD28, CD45RO, CD62L, CD314 (NKG2D), MICA/B и изотипически сходными контролями (BD Biosciences). Окрашивание йодидом пропидия (PI) выполняли для установления различия между живыми и мертвыми клетками. Экспрессию ROR1 на клеточной поверхности анализировали с помощью поликлонального антитела козы к ROR1 человека (R&D Systems).
Поверхностную экспрессию химерного рецептора ROR1 из 2А2 анализировали с помощью поликлонального антитела козы к IgG мыши (Fab-специфического) (Jackson ImmunoResearch). Проточные анализы выполняли на FACSCanto®, очистку путем сортировки на FACSAriaII® (Becton Dickinson), и данные анализировали с помощью программного обеспечения FlowJo® (Treestar).
Конструирование вектора и получение лентивируса, кодирующего химерный рецептор
ROR1-специфические и СD19-специфические химерные рецепторы конструировали с применением сегментов цепей VL и VH mAb 2А2, R12 и R11 (ROR1) и mAb FMC63 (CD19) (последовательности вариабельных участков для R11 и R12 представлены в Yang et al, Plos One 6(6):e21018, June 15, 2011). Каждый scFv был соединен с помощью пептида (G4S)3 (SEQ ID NO: 12) со спейсерным доменом, полученным из Fc IgG4 (база данных Uniprot: Р01861, SEQ ID NO: 13), содержащим последовательности 'шарнирный участок-СН2-СН3' (229 аминокислот, SEQ ID NO:), 'шарнирный участок-СН3' (119 аминокислот, SEQ ID NO:) или 'шарнирный участок' в отдельности (12 аминокислот, SEQ ID NO: 4) (фигура 1). Все спейсеры содержали замену S→Р в домене 'шарнирный участок', находящуюся в положении 108 нативного белка Fc IgG4, и были соединены с трансмембранным доменом CD28 человека длиной 27 аминокислот (Uniprot: Р10747, SEQ ID NO: 14) и с сигнальным модулем, содержащим (i) цитоплазматический домен CD28 человека длиной 41 аминокислота с заменой LL→GG, находящейся в положениях 186-187 нативного белка CD28 (SEQ ID NO: 14), или (ii) цитоплазматический домен 4-1ВВ человека длиной 42 аминокислоты (Uniprot: Q07011, SEQ ID NO: 15), каждый из которых был соединен с цитоплазматическим доменом изоформы 3 CD3ζ человека длиной 112 аминокислот (Uniprot: Р20963, SEQ ГО NO: 16). Конструкт кодировал элемент рибосомного перепрыгивания Т2А (SEQ ID NO: 8) и последовательность tEGFR (SEQ ID NO: 9) ниже химерного рецептора. Подвергнутые оптимизации кодонов нуклеотидные последовательности, содержащие каждый трансген, были синтезированы (Life Technologies) и клонированы в лентивирусный вектор epHIVT.
Лентивирусы, кодирующие химерный рецептор ROR1, химерный рецептор CD19 или tEGFR, получали в клетках 293Т с помощью упаковочных векторов pCHGP-2, pCMV-Rev2 и pCMV-G, а также реагента для трансфекции Calphos® (Clontech).
Создание линий Т-клеток, экспрессирующих химерные рецепторы ROR1 и CD19
Центральные Т-клетки памяти (ТСM) CD8+ CD45RO+ CD62L+ или общую популяцию Т-клеток CD4+ отсортировывали из РВМС нормальных доноров, активировали гранулами с антителами к CD3/CD28 (Life Technologies) и трансдуцировали на 3-й день после активации путем центрифугирования при 800 g в течение 45 минут при 32°С надосадочной жидкостью, содержащей лентивирус (MOI=3), дополненной 1 мкг/мл полибрена (Millipore). Т-клетки размножали в RPMI с 10% сывороткой человека, 2 мМ L-глутамином и 1% пенициллином/стрептомицином (среде для CTL), дополненной рекомбинантным IL-2 человека до конечной концентрации 50 Ед/мл. Субпопуляцию tEGFR+ каждой линии Т-клеток обогащали посредством иммуномагнитной селекции с mAb к EGFR, конъюгированным с биотином (ImClone Systems), и гранулами стрептавидина (Miltenyi). Т-клетки с химерным рецептором ROR1 и контрольные Т-клетки с tEGFR размножали согласно протоколу быстрого размножения (Riddell SR, Greenberg PD, The use of anti-CD3 and anti-CD28 monoclonal antibodies to clone and expand human antigen-specific T cells J Immunol Methods. 1990; 128(2):189-201. Epub 1990/04/17.), и Т-клетки, модифицированные химерным рецептором CD19, размножали путем стимуляции облученными (8000 рад) B-LCL в соотношении Т-клетки: LCL 1:7. Т-клетки культивировали в среде для CTL с 50 Ед/мл IL-2.
Анализы цитотоксичности, секреции цитокинов и пролиферации
Клетки-мишени метили 51Сr (PerkinElmer), отмывали и инкубировали в трех повторностях при концентрации 1-2×103 клеток/лунка с эффекторными Т-клетками, модифицированными химерными рецепторами, при различных соотношениях эффектора и мишени (Е : Т). Надосадочные жидкости собирали для у-радиометрии после 4-часового инкубирования, и специфический лизис рассчитывали с помощью стандартной формулы. Для анализа секреции цитокинов 5×104 Т-клеток высевали на чашки в трех повторностях с клетками-мишенями в соотношении Е : Т 1:1 (первичные СLL), 2:1 (Raji ROR1; JeKo-1), 4:1 (K562/ROR1, K562/CD19 и K562) или 10:1 (MDA-MB-231), и измеряли уровень IFN-γ, TF-α и IL-2 с помощью ELISA или мультиплексного иммунологического анализа цитокинов (Luminex) в надосадочной жидкости, удаленной после 24-часового инкубирования. В экспериментах с блокированием передачи сигналов с помощью NKG2D, применяли антитело к NKG2D (клон 1D11), антитело к MICA/B (клон 6D4, все от BD) и антитело к ULBP (любезно предоставленное д-ром Вероникой Грох, FHCRC) в насыщающих концентрациях. Для анализа пролиферации Т-клетки метили 0,2 мкМ сложным сукцинимидиловым эфиром карбоксифлуоресцеина (CFSE, Invitrogen), отмывали и высевали на чашки в трех повторностях с клетками-стимуляторами в среде без экзогенных цитокинов. После 72-часового инкубирования клетки метили с помощью mAb к CD8 и PI и анализировали с помощью проточной цитометрии для оценки клеточного деления живых Т-клеток CD8+.
Эксперименты на мышах NOD/SCID/γc-/-(NSG)
Институциональный комитет по содержанию и использованию животных одобрил все эксперименты с мышами. Самок мышей NOD.Cg-Prkdcscid II2rgtmlWjl/SzJ (NSG) в возрасте 6-8 месяцев приобретали в Jackson Laboratory или разводили в условиях собственного производства. Мышам инъецировали 0,5×106 опухолевых клеток JeKo-1 /ffluc в хвостовую вену, и после этого они получали инъекцию модифицированных химерными рецепторами или контрольных Т-клеток в хвостовую вену.
Для биолюминесцентной визуализации роста опухолей мыши получали внутрибрюшинные инъекции субстрата люциферина (Caliper Life Sciences), ресуспендированного в PBS (15 мкг/г массы тела). Мышей подвергали анестезии изофлураном, и у них проводили визуализацию с помощью системы визуализации Xenogen IVIS (Caliper) через 10, 12 и 14 минут после инъекции люциферина в режиме малого коэффициента биннинга при времени экспозиции от 1 секунды до 1 минуты для получения ненасыщенных изображений. Люциферазную активность анализировали с помощью программного обеспечения Living Image (Caliper), и в областях, представляющих интерес, которые охватывали все тело или грудную клетку каждой отдельной мыши, анализировали поток фотонов.
Статистические анализы
Статистические анализы выполняли с помощью программного обеспечения Prism (GraphPad®). t-критерий Стьюдента применяли в виде двустороннего парного критерия с 95% доверительным интервалом, а результаты с р-значением р<0,05 считали значимыми. Статистический анализ выживаемости выполняли с помощью логарифмического рангового критерия, а результаты с р-значением р<0,05 считали значимыми.
Результаты
Усечение спейсерного домена химерного рецептора ROR1 2А2 обеспечивает лучшее распознавание опухолей ROR1+. Ранее была описано конструирование ROR1-специфического химерного рецептора с помощью scFv 2А2, который связывается с эпитопом в №12-концевой, мембранно-дистальной Ig-подобной/Frizzled-части ROR1-1. Исходный химерный рецептор ROR1 из 2А2 имел длинный спейсер длиной 229 аминокислот, который содержал участок 'шарнирный участок-СН2-СН3' Fc IgG4, а также включал в себя костимулирующий домен CD28 и сигнальный домен CD3ζ (Hudecek М et al. Blood, 2010). Этот химерный рецептор обеспечивал специфическое распознавание опухолей ROR1+, однако, было выдвинуто предположение, что в связи с мембранно-дистальным положением эпитопа ROR1 усечение спейсерного домена могло усиливать распознавание опухоли и передачу сигналов Т-клеткам. В связи с этим были сконструированы два дополнительных химерных рецептора, в которых спейсерный домен Fc IgG4 подвергали последовательному удалению с получением вариантов 'шарнирный участок-СН3' (119 аминокислот, средний) и 'шарнирный участок в отдельности' (12 аминокислот, короткий). Каждый из новых рецепторов содержал идентичные сигнальные модули scFv 2А2, а также CD28 и CD3ζ. Кассета трансгена включала в себя усеченный EGFR (tEGFR), служащий в качестве маркера для трансдукции, селекции и in vivo отслеживания Т-клеток, модифицированных химерными рецепторами.
Очищенные ТСМ CD8+ трансдуцировали химерными рецепторами ROR1 из 2А2, содержащими полноразмерные или усеченные спейсеры Fc IgG4, и контрольным вектором, кодирующим tEGFR. Средняя эффективность трансдукции составляла 15% (диапазон 9-22%), и трансген-положительные Т-клетки обогащали до однородной чистоты (>90%) на 10-й день путем селекции в отношении экспрессии tEGFR и размножали (фигура 2А). Поверхностную экспрессию каждого из химерных рецепторов подтверждали окрашиванием F(аb)-специфическими антителами (фигура 2А).
Анализ in vitro функционирования Т-клеток CD8+, модифицированных для экспрессии каждого из химерных рецепторов ROR1 из 2А2, показал, что каждый рецептор обеспечивал специфический лизис клеток JeKo-1 MCL и первичных клеток CLL, которые в естественных условиях экспрессируют ROR1, и клеток K562, которые были трансдуцированы с помощью ROR1, но не обеспечивал распознавание контрольных мишеней ROR1- (фигура 2В). Т-клетки, экспрессирующие химерный рецептор ROR1 из 2А2 с коротким спейсером 'шарнирный участок в отдельности', обладали максимальной цитолитической активностью, и четко прослеживалась иерархия лизиса опухолей (короткий > средний > длинный) в отношении всех опухолей-мишеней ROR1+ (фигура 2В), что показывает важность длины спейсерного домена в распознавании опухолевых клеток ROR1+.
Противоопухолевая эффективность адоптивной Т-клеточной терапии коррелирует с пролиферацией и выживаемостью перенесенных Т-клеток, которые можно изменять с помощью передачи сигнала посредством химерного рецептора. Для анализа пролиферации Т-клеток, модифицированных каждым из химерных рецепторов ROR1 2А2, применяли анализы разведения CFSE после привлечения Raji/ROR1 или CLL и обнаружили, что конструкт с коротким спейсером способствовал наибольшей пролиферации Т-клеток после стимуляции (фигура 2С). Для подтверждения того, что повышенная пролиферация не была связана с большей индуцируемой активацией клеточной гибелью (AICD), также анализировали долю Т-клеток, модифицированных химерными рецепторами ROR1 из 2А2, которые окрашивались йодидом пропидия (PI) после стимуляции опухолевыми клетками Raji/ROR1 и JeKo-1. В линии Т-клеток, модифицированных короткими спейсерами (Raji/ROR1: 17,2%/JeKo-1: 20,2%), выявили намного более низкую частоту Т-клеток CD8+ PI+ по сравнению с Т-клетками, модифицированными средними (41,6%/42,4%) и длинными (44,5%/48,5%) спейсерами.
Количественный анализ выработки цитокинов в ответ на стимуляцию клетками Raji/ROR1 и первичными клетками CLL показал выработку IFN-γ, TNF-α и IL-2 Т-клетками, экспрессирующими каждый из химерных рецепторов ROR1 из 2А2. По наблюдениям в анализах цитотоксичности конструкт с коротким спейсером более эффективно опосредовал секрецию цитокинов после распознавания опухоли (фигура 2D). Таким образом, данный анализ показывает, что усечение внеклеточного спейсерного домена Fc IgG4 химерного рецептора ROR1 из 2А2 приводит к значительному усилению цитотоксичности, пролиферации и эффекторных функций in vitro после распознавания опухоли.
scFv R11, специфичному к мембранно-проксимальному эпитопу в крингл-домене ROR1, требуется длинный внеклеточный спейсерный домен.
Очищенные Т-клетки CD8+ трансдуцировали химерными рецепторами ROR1, содержащими scFv R11, специфичный к крингл-домену ROR1, и содержащими полноразмерные или усеченные спейсеры Fc IgG4 (СН3 и шарнирный участок в отдельности). Измеренная по экспрессии EGFR эффективность трансдукции каждым из векторов с коротким (шарнирный участок IgG4 в отдельности), средним (шарнирный участок IgG4/CH3) и длинным (шарнирный участок IgG4/CH2/CH3) спейсерами была сопоставимой (45-51%) (фигура 3А). Т-клетки, трансдуцированные каждым из векторов, анализировали в отношении цитолиза (фигура 3В), пролиферации (фигура 3С) и выработки цитокинов (фигура 3D) в ответ на воздействие лейкозных клеток или клеток лимфомы, которые экспрессировали или не экспрессировали ROR1. Как показано, только Т-клетки, трансдуцированные химерным рецептором R11, содержащим длинную спейсерную последовательность, были способны эффективно распознавать опухоли ROR1+ и опосредовать эффекторные функции.
Химерные рецепторы ROR1, полученные из mAb R12 с более высокой аффинностью, чем 2А2, опосредуют более высокую противоопухолевую реактивность
Далее было изучено, может ли повышение аффинности scFv, применяемого для конструирования химерного рецептора ROR1, влиять на распознавание опухоли и функционирование Т-клеток. Были получены ROR1-специфические химерные рецепторы из mAb R12, которое, подобно 2А2, связывается с эпитопом в NH2-концевом домене Ig/Frizzled ROR1, но с > в 50 раз более высокой аффинностью моновалентного связывания.
Химерные рецепторы ROR1 из R12 конструировали как с длинными, так и с короткими спейсерами Fc IgG4 для определения того, отличается ли оптимальная конструкция спейсера для данного scFv с более высокой аффинностью от таковой для scFv с более низкой аффинностью. Было обнаружено, что химерный рецептор ROR1 из R12 с коротким спейсером, подобно таковому из 2А2, обеспечивал улучшенные цитолитическую активность, секрецию цитокинов и пролиферацию (данные не показаны), что позволяет предположить, что более короткая длина спейсера обеспечивает более эффективное пространственное привлечение Т-клетки и клетки-мишени ROR1+ для активации Т-клеток.
Затем были разработаны химерные рецепторы ROR1 из R12 и 2А2, которые содержали оптимальный (короткий) внеклеточный спейсер, а также, для сравнения, костимулирующий домен CD28 или 4-1ВВ в тандеме с СD3ζ (4 конструкта) (фигура 4А, В). Эти конструкты химерных рецепторов ROR1 экспрессировали в очищенных ТCМ CD8+ здоровых доноров, и с помощью окрашивания tEGFR была подтверждена одинаковая экспрессия трансгена (фигура 5А). Т-клетки, модифицированные каждым из химерных рецепторов ROR1 из 2А2 и R12, специфично лизировали опухолевые клетки K562/ROR1 и Raji/ROR1 с приблизительно одинаковой эффективностью (фигура 5В). Однако, анализ выработки цитокинов показал, чтр химерные рецепторы ROR1 из R12 с высокой аффинностью, которые содержали CD28 или 4-1ВВ, обеспечивали значительно более высокую выработку IFN-γ, TNF-α и IL-2 по сравнению с соответствующими конструктами из 2А2 (фигура 5С). Было обнаружено, что Т-клетки, экспрессирующие химерные рецепторы с костимулирующим доменом CD28, вырабатывали больше IFN-γ, TNF-α и IL-2 по сравнению с Т-клетками, экспрессирующими химерные рецепторы с костимулирующим доменом 4-1ВВ.
Эксперименты по анализу пролиферации Т-клеток с химерными рецепторами ROR1 показал более высокую процентную долю пролиферирующих Т-клеток и большее число клеточных делений у Т-клеток, экспрессирующих химерные рецепторы ROR1 из R12 с высокой аффинностью с доменами CD28 и 4-1ВВ по сравнению с Т-клетками, экспрессирующими соответствующие аналоги из 2А2 (фигура 4D). В Т-клетках, экспрессирующих химерные рецепторы с доменом CD28, наблюдалась более интенсивная пролиферация, соответствующая более высокой выработке IL-2, индуцированной этими рецепторами. В линиях Т-клеток, модифицированных химерными рецепторами ROR1 из R12, наблюдалась более низкая частота AICD, измеренная по окрашиванию PI, по сравнению с линиями Т-клеток, модифицированными химерными рецепторами ROR1 из 2А2, после стимуляции опухолевыми клетками Raji/ROR1 и JeKo-1, соответственно (R12: 5,6%/6,9% по сравнению с 2А2: 10%/9,65%). Линии Т-клеток, которые экспрессировали химерные рецепторы с доменом CD28, также характеризовались более низкой AICD по сравнению с линиями Т-клеток, которые экспрессировали химерные рецепторы с доменом 4-1ВВ, в ответ на воздействие опухолевых клеток Raji/ROR1 и JeKo-1, соответственно (R12: 16,4%/18,4% по сравнению с 2А2: 38,1%/39,6%).
Для определения того, распространялось ли усиление функционирования, наблюдаемое в Т-клетках CD8+ с химерными рецепторами ROR1 из R12, на Т-клетки CD4+, общую популяцию Т-клеток CD4+ трансдуцировали химерными рецепторами ROR1 из R12 и 2А2, содержащими короткий спейсер и костимулирующий домен CD28. В ответ на воздействие опухолевых клеток Raji/ROR1+ Т-клетки CD4+, которые экспрессировали scFv R12 с высокой аффинностью, вырабатывали IFN-γ, TNF-α, IL-2, IL-4 и IL-10 на более высоких уровнях и подвергались большей пролиферации, чем Т-клетки CD4+, которые экспрессировали 2А2 (фигура 5А, В). Как выработка цитокинов, так и пролиферация были выше для Т-клеток CD4+ по сравнению с Т-клетками CD8+, модифицированными теми же химерными рецепторами ROR1. В заключение, данные показывают, что как длина несигнального внеклеточного спейсерного домена химерных рецепторов, так и аффинность scFv, подбираемые индивидуально, являются независимыми параметрами, которые влияют на функцию Т-клеток с химерными рецепторами ROR1.
Т-клетки CD8+, модифицированные химерным рецептором ROR1 с высокой аффинностью, и химерный рецептор CD19 имеют сопоставимую активность в отношении первичных CLL in vitro
На всех первичных CLL наблюдается однородная экспрессия ROR1 и CD19 (фигура 6А), однако, абсолютное число молекул ROR1 в каждой опухолевой клетке по оценкам составляет в 10 раз меньше, чем абсолютное число молекул CD19, на которые в клинических испытаниях успешно нацеливались Т-клетками с химерным рецептором CD19. Было проведено сравнение распознавания первичных CLL Т-клетками CD8+, экспрессирующими оптимизированные химерные рецепторы ROR1 из R12 и 2А2, и химерным рецептором CD19, полученным из scFv FMC63.
Для модификации химерного рецептора применяли очищенные ТСМ CD8+ с целью обеспечения однородного клеточного продукта, и каждый химерный рецептор содержал короткий спейсер 'шарнирный участок в отдельности' Fc IgG4 и костимулирующий домен 4-1ВВ. Было подтверждено, что химерный рецептор CD19 (шарнирный участок IgG4) был по меньшей мере таким же и более эффективным в распознавании опухолей CD19+ по сравнению с химерным рецептором CD19 со спейсером, представляющим собой шарнирный участок CD8a, и костимулирующим доменом 4-1ВВ, который применяется в текущих клинических испытаниях (фигура 20). Т-клетки, экспрессирующие химерные рецепторы с 4-1ВВ и СD3-зета, а также модифицированным шарнирным участком Fc IgG4, проявляют более эффективное in vitro и in vivo функционирование по сравнению с Т-клетками, экспрессирующими химерные рецепторы CD19 с 4-1ВВ и СD3-зета, а также шарнирным участком СD8-альфа. На фигуре 20D сравнивали in vivo противоопухолевую активность Т-клеток, экспрессирующих химерный рецептор CD19 с шарнирным участком Fc IgG4 (группа 1) или шарнирным участком СD8-альфа (группа 2), и Т-клеток, экспрессирующих только tEGFR (группа 3), у мышей NSG, которым инокулировали опухолевые клетки Raji, экспрессирующие люциферазу светлячка (ffluc). Визуализацию у мышей проводили спустя 17 дней после инокуляции опухоли и 10 дней после инокуляции Т-клеток. Данные показывают более высокую опухолевую нагрузку у мышей, обработанных контрольными Т-клетками с tEGFR (группа 3) или Т-клетками с химерным рецептором CD19 с шарнирным участком СD8-альфа (группа 2), по сравнению с мышами, обработанными Т-клетками с химерным рецептором CD19 с шарнирным участком IgG4 Fc (группа 1).
Цитолитическая активность Т-клеток с химерным рецептором ROR1 из R12 в отношении первичных опухолевых клеток от нескольких пациентов с CLL (n=4) была выше по сравнению с Т-клетками, модифицированными химерным рецептором ROR1 из 2А2 с более низкой аффинностью, и эквивалентной лизису, наблюдаемому в присутствии Т-клеток с химерным рецептором CD19 (фигура 6В). Мультиплексный анализ цитокинов показал практически аналогичную выработку IFN-γ и TNF-α, но меньшую выработку IL-2 Т-клетками CD8+, экспрессирующими ROR1 из R12, по сравнению с Т-клетками CD8+, экспрессирующими химерный рецептор CD19, после совместного культивирования с первичными CLL (фигура 6С). Т-клетки с химерным рецептором ROR1 из 2А2 вырабатывали все цитокины в меньших количествах, чем Т-клетки с химерным рецептором ROR1 из R12, как отмечено ранее. Выработка цитокинов всеми Т-клетками, трансдуцированными химерными рецепторами, после стимуляции с помощью CLL была значительно меньшей, чем в присутствии Raji/ROR1, которые, в отличие CLL, экспрессируют как CD80, так и CD86, которые могут привлекать CD28, экспрессируемый на Т-клетках с химерными рецепторами (фигура 6А, С).
Наблюдалась меньшая пролиферация Т-клеток, экспрессирующих химерный рецептор ROR1 из R12 и 2А2, по сравнению с Т-клетками, экспрессирующими химерный рецептор CD19, после стимуляции с помощью CLL (CD19>R12>2A2) (фигура 6D). Было выдвинуто предположение, что пролиферация Т-клеток CD8+ с химерным рецептором ROR1 в ответ на воздействие CLL может усиливаться в присутствии Т-клеток CD4+, модифицированных химерными рецепторами, в связи с характерной для них более высокой секрецией IL-2 по сравнению с ТСМ CD8+ (фигура 4А; фигура 8А). Для проверки этой возможности выполняли эксперименты по совместному культивированию in vitro, в которых ТСМ CD4+ и CD8+ по отдельности модифицировали химерными рецепторами ROR1 из R12, ROR1 из 2А2 и CD19, соответственно, обогащали по экспрессии химерных рецепторов и комбинировали в соотношении 1:1 для обеспечения равной пропорции Т-клеток CD8+ и CD4+, модифицированных каждым из этих векторов. Эти клетки метили CFSE и стимулировали первичными CLL. Наблюдалось значительное повышение пролиферации Т-клеток CD8+ с химерным рецептором ROR1 из R12 после добавления Т-клеток CD4+, трансдуцированных химерным рецептором, но не являющихся нетрансдуцированными (фигура 8В). Примечательно, что при содействии CD4+ наблюдалась равная пролиферация Т-клеток CD8+ с химерными рецепторами ROR1 из R12 и CD19 в ответ на воздействие CLL, в то время как пролиферация Т-клеток CD8+, экспрессирующих химерный рецептор ROR1 из 2А2 с более низкой аффинностью, оставалась более низкой. В совокупности данные показывают, что химерный рецептор ROR1 из R12 с высокой аффинностью обеспечивает более высокую реактивность по сравнению с таковым из 2А2 в отношении первичных клеток CLL in vitro.
Т-клетки с химерным рецептором ROR1 опосредуют in vivo противоопухолевую активность в модели системной мантийноклеточной лимфомы на мышах
Оставалось неясным, могла ли более высокая in vitro активность Т-клеток, модифицированных химерным рецептором из R12 с более высокой аффинностью, обуславливать улучшенную противоопухолевую активность in vivo, и как нацеливание на ROR1 можно сравнивать с нацеливанием на CD19. Для рассмотрения этих вопросов группам мышей NSG с иммунодефицитом инокулировали линию клеток JeKo-1/ffluc MCL человека путем инъекции в хвостовую вену, и через семь дней, когда произошла диссеминация опухоли, мышей обрабатывали однократной внутривенной дозой Т-клеток CD8+ с химерными рецепторами ROR1 из R12, ROR1 из 2А2 или CD19. Контрольных мышей обрабатывали Т-клетками с tEGFR или не обрабатывали. Все химерные рецепторы имели оптимальный короткий спейсер и костимулирующий домен 4-1ВВ.
У необработанных мышей NSG/JeKo-1 развивалась быстро прогрессирующая системная лимфома, приводящая к необходимости эвтаназии приблизительно через 4 недели после инокуляции опухоли (фигура 9А-С).
Наблюдали регрессию опухоли и улучшение выживаемости у всех мышей, обработанных Т-клетками с химерными рецепторами ROR1 из R12, ROR1 из 2А2 и CD19. Мыши, обработанные Т-клетками с химерным рецептором ROR1 из R12, характеризовались лучшими противоопухолевым ответом и выживаемостью по сравнению с мышами, обработанными Т-клетками с химерным рецептором ROR1 из 2А2 (р<0,01), и сопоставимой противоопухолевой активностью по сравнению с мышами, обработанными Т-клетками с химерным рецептором CD19 (фигура 9А-С).
Анализировали частоту Т-клеток с химерными рецепторами в периферической крови после адоптивного переноса и выявляли более высокие количества Т-клеток с tEGFR+у мышей, обработанных химерным рецептором ROR1 из R12, по сравнению с химерным рецептором ROR1 из 2А2, что позволяет предположить, что более интенсивная пролиферация in vivo улучшала подавление опухоли. Для подтверждения этого вводили меченные CFSE Т-клетки с химерным рецептором CD19, химерными рецепторами ROR1 из R12 и 2А2 группам мышей NSG, несущих JeKo-1/ffluc, и анализировали пролиферацию Т-клеток в периферической крови, костном мозге и селезенке спустя 72 часа после переноса. Т-клетки с химерными рецепторами R12 и CD19 характеризовались более высокой процентной долей пролиферирующих клеток и подвергались большему числу клеточных делений по сравнению с Т-клетками с химерным рецептором ROR1 из 2А2 (фигура 9D). В конечном итоге наблюдался рецидив опухоли JeKo-1 у всех мышей, обработанных Т-клетками с химерными рецепторами ROR1 или CD19 (фигура 9А-С). Рецидив опухоли не являлся результатом отбора вариантов с утратой ROR1 или CD19, поскольку рецидивирующие опухоли были положительными по обеим молекулам.
Для сравнения проанализировали противоопухолевую эффективность Т-клеток с химерным рецептором CD19 у мышей NSG, которым трансплантировали опухоли Raji, и наблюдали полное уничтожение опухоли, указывающее на то, что рецидив JeKo-1 отражает сложность уничтожения этой опухоли (данные не показаны). Таким образом, эти данные впервые показывают, что Т-клетки с химерными рецепторами ROR1 обладают противоопухолевой эффективностью in vivo, и позволяют предположить, что для В-клеточных злокачественных опухолей оптимизированный химерный рецептор ROR1, такой как рецептор из R12, может быть эффективным и не затрагивать нормальные В-клетки CD19+, которые не характеризуются экспрессией ROR1.
Т-клетки, экспрессирующие химерный рецептор ROR1 из R12, характеризуются более высокой реактивностью по сравнению с 2А2 в отношении клеток эпителиальных опухолей ROR1+
ROR1 был обнаружен на многих клетках эпителиальных опухолей, хотя неизвестно, является ли экспрессия ROR1 достаточной для распознавания Т-клетками с химерными рецепторами ROR1. С помощью проточной цитометрии была подтверждена экспрессия ROR1 в линиях клеток рака молочной железы MDA-MB-231 и 468 и линиях клеток почечно-клеточного рака FARP, TREP и RWL (фигура 10А). Затем анализировали распознавание опухолей Т-клетками CD8+, трансдуцированными химерными рецепторами ROR1 из R12 с оптимальным коротким спейсером и доменом 4-1ВВ, и наблюдали эффективное распознавание MDA-MB-231, MDA-MB-468, FARP, TREP и RWL (фигура 11А). Анализировали секрецию цитокинов и пролиферацию Т-клеток, модифицированных химерными рецепторами ROR1 из R12 и 2А2, после совместного культивирования с MDA-MB-231, и наблюдали более высокую выработку цитокинов и пролиферацию в присутствии химерного рецептора ROR1 из R12 (фигура 11В, С). Подобно тому, что наблюдали в отношении В-клеточных злокачественных опухолей ROR1+, лучшая активация Т-клеток с химерными рецептором ROR1 из R12 после стимуляции MDA-MB-231 не была связана с повышенной AICD (R12: 9,8% по сравнению с 2А2: 10,9%).
Обсуждение
ROR1 привлекал интерес в качестве потенциальной мишени для иммунотерапии рака в связи с его экспрессией на поверхности клеток многих видов В-клеточных лимфоидных и эпителиальных опухолей, в том числе подвидов рака легких, кишечника и почечно-клеточного рака. Ранее было показано, что CLL и MCL специфически распознавались Т-клетками, модифицированными для экспрессии ROR1 - специфического химерного рецептора (Hudecek М, et al. Blood. 2010; 1 16(22):4532-41. Epub 2010/08/13). Конструкция и функционирование химерных рецепторов ROR1 были улучшены путем модификации внеклеточного спейсерного домена и получения химерного рецептора из scFv с более высокой аффинностью, и было показано, что Т-клетки, модифицированные разработанными химерными рецепторами ROR1, характеризуются in vivo активностью в отношении В-клеточной лимфомы ROR1+ и in vitro активностью в отношении широкого спектра эпителиальных опухолей.
Сравнивали функционирование Т-клеток, модифицированных химерными рецепторами ROR1, полученными из mAb 2А2, которые содержали исходный длинный спейсер 'шарнирный участок-СН2-СН3' Fc IgG4, который, как было показано, способствует высокому уровню экспрессии на клеточной поверхности, или усеченные средний 'шарнирный участок-СН3' и короткий 'шарнирный участок в отдельности' варианты спейсера. Домен, представляющий собой шарнирный участок длиной 12 аминокислот, был сохранен в конструкте с коротким спейсером на основании предварительных данных о том, что для отделения scFv от мембраны Т-клетки и обеспечения распознавания антигена на опухолевых клетках требовался гибкий спейсер (Fitzer-Attas CJ, et al., Harnessing Syk family tyrosine kinases as signaling domains for chimeric single chain of the variable domain receptors: optimal design for T cell activation. J Immunol. 1998; 160(1): 145-54. Epub 1998/04/29.)
Исследования с химерным рецептором ROR1 из 2А2 показывают, что секреция цитокинов Т-клетками и пролиферация последних после распознавания опухолевых клеток более эффективны с конструктами со средним и коротким спейсером по сравнению с конструктом с длинным спейсером. Окрашивание Аb к F(ab) показало одинаковую экспрессию химерных рецепторов для всех трех химерных рецепторов, что демонстрирует, что улучшение функционирования Т-клеток с химерным рецептором с коротким спейсером не было обусловлено различиями в плотности химерных рецепторов. Эти данные подтверждают принцип, заключающийся в том, что конструкцию внеклеточных спейсеров следует подбирать индивидуально для каждой молекулы-мишени и эпитопа.
Аффинность scFv, выбранного для разработки химерного рецептора, является дополнительным параметром, который может влиять на распознавание Т-клетками. Была получена и охарактеризована панель ROR1-специфических mAb с различными аффинностями и выбрано mAb R12, которое, как и 2А2, распознает эпитоп в Ig-подобном/Frizzled-участке. R12 имело более высокую аффинность к белку ROR1 в связи с намного более медленной диссоциацией. Химерный рецептор R12, разрабатываемый с коротким внеклеточным доменом, подобно химерному рецептору 2А2, обеспечивал оптимальное распознавание Т-клетками и их функционирование. Непосредственное сравнение пролиферации и выработки цитокинов после привлечения к опухоли Т-клеток, модифицированных химерными рецепторами 2А2 и R12, показало, что химерный рецептор R12, полученный из mAb с более высокой аффинностью, был более эффективным. Беспокойство вызывало то, что более медленная диссоциация R12 от ROR1 могла продлевать активацию Т-клеток и обеспечивать повышенную восприимчивость к AICD. Однако, был обнаружен более низкий уровень AICD для Т-клеток, модифицированных химерным рецептором ROR1 из R12, по сравнению с Т-клетками, модифицированными химерным рецептором ROR1 из 2А2, что указывало на то, что повышенная аффинность R12 не оказывала вредного эффекта на выживаемость Т-клеток в моделях для доклинических исследований.
ROR1 имеет потенциальное преимущество над CD19 в качестве мишени для CLL и MCL, поскольку он не экспрессируется на нормальных зрелых наивных В-клетках и В-клетках памяти. Однако, на клетках В-клеточных опухолей имеется меньшее число молекул ROR1 по сравнению с CD19, и неясно, будет ли оптимизированный химерный рецептор ROR1 таким же эффективным, так и химерный рецептор CD19, подобный по конструкции химерным рецепторам, применяемым в клинической практике. К сожалению, ксенотрансплантатные модели В-клеточных опухолей, применяемые ранее у мышей NSG для оценки функции Т-клеток с химерным рецептором CD19, включающие Raji, Daudi и Nalm-6, не были получены из CLL или MCL и не характеризуются конститутивной экспрессией ROR1. Поэтому для сравнения нацеливания на CD19 и ROR1 in vivo применяли клеточную линию MCL JeKo-1, которая в естественных условиях экспрессирует как CD19, так и ROR1, и трансплантировали ее мышам NSG. Для обеспечения клинической значимости модели клетки лимфомы JeKo-1 инокулировали внутривенно для получения системных опухолей, и мышей обрабатывали продуктами Т-клеток однородной консистенции непосредственно после формирования развившихся опухолей. Было обнаружено, что Т-клетки, экспрессирующие химерный рецептор из R12 с высокой аффинностью, обеспечивали противоопухолевую активность in vivo, аналогичную таковой Т-клеток с химерным рецептором CD19. В соответствии с анализом in vitro, химерный рецептор ROR1 из R12 также опосредовал более высокую активность in vivo по сравнению с оптимизированным химерным рецептором ROR1 из 2А2. Эти результаты следует интерпретировать с осторожностью, поскольку модели опухолей на мышах могут и не прогнозировать эффективность адоптивной терапии в клинических условиях. Однако, результаты позволяют предположить, что требуется рассмотрение ROR1 в качестве альтернативы CD19 или обеспечение дополнительной мишени для минимизации возможности появления вариантов с утратой CD19.
По-видимому, ROR1 играет решающую роль в выживании некоторых эпителиальных опухолей. Таким образом, преимуществом нацеливания на ROR1 является то, что один химерный рецептор может быть пригодным для лечения пациентов с большим числом гемопоэтических и негемопоэтических опухолей.
Данные впервые показывают, что Т-клетки, которые экспрессируют разработанный химерный рецептор ROR1, эффективно распознают виды эпителиальных опухолей in vitro. Секреция цитокинов и пролиферация Т-клеток, индуцируемые раковыми клетками молочной железы ROR1+, были более высокими, чем таковые, индуцируемыми лейкозными клетками, несмотря на отсутствие костимулирующего лиганда CD80/86. Исследования, описанные в данном документе, показывают, что конструкция внеклеточного спейсерного домена и аффинность химерных рецепторов являются параметрами, которые можно модулировать для усиления распознавания гематологических и эпителиальных опухолей ROR1+in vitro и in vivo Т-клетками, модифицированными химерным рецептором ROR1. Разработка химерных рецепторов ROR1 с повышенной противоопухолевой реактивностью предоставляет возможность для клинических путей применения при ряде форм рака у человека.
Пример 2. Влияние длины внеклеточного спейсерного домена на запуск лизиса опухолевых клеток с Неr2-специфическим химерным рецептором, который распознает эпитоп, расположенный проксимально по отношению к мембране опухолевой клетки
Влияние длины спейсера CAR на распознавание и запуск распознавания опухолевых клеток Т-лимфоцитами человека CD8+, которые экспрессировали Неr2-специфический химерный рецептор, исследовали с помощью способов, аналогичных описанным выше для ROR1. Неr2-специфические химерные рецепторы конструировали с применением сегментов цепей VL и VH Неr2-специфического mAb, которое распознавало мембранно-проксимальный эпитоп Неr2 (фигура 12А), и scFv были соединены с внеклеточными спейсерными доменами шарнирный участок IgG4/CH2/CH3, шарнирный участок IgG4/CH3 и шарнирный участок IgG4 в отдельности и трансмембранным доменом CD28, сигнальными доменами 4-1ВВ и СD3-зета (фигура 12В). Первичные Т-клетки CD8+ трансдуцировали каждым из химерных рецепторов Неr2, и среди них производили отбор по экспрессии,маркера трансдукции EGFR (фигура 12D).
Экспрессию химерных рецепторов Неr2 и размер каждого рецептора подтверждали с помощью вестерн-блоттинга (фигура 12С). Затем Т-клетки размножали с mAb к CD3 и питающими клетками и исследовали в отношении их способности к распознаванию опухолевых клеток Неr2+. Как наблюдалось в отношении ROR1-специфического химерного рецептора из R11, химерный рецептор Неr2, который содержал длинный внеклеточный спейсерный домен, обеспечивал более эффективное распознавание Т-клетками опухолевых клеток Неr2+ (фигура 12Е).
Обсуждение
В этом примере о влиянии длины внеклеточного спейсера на распознавание опухолевых клеток Т-клетками, модифицированными химерными рецепторами, применяли химерный рецептор, содержащий scFv, созданный на основе последовательностей VH+L химерного mAb герцептина. В исследованиях Cho et al (Nature 421:756, 2003) определили положение эпитопа герцептина как мембранно-проксимальное во внеклеточном домене Her2 (ERRB2) (фигура 12А). На основании понимания структуры вариантов шарнирного участка Fc IgG4 человека (фигура 12В) было выдвинуто предположение о том, что эпитоп-мишень в мембранно-проксимальном положении во внеклеточном антигене опухолевой клетки наилучшим образом распознавался бы эффекторными Т-клетками, которые экспрессируют химерный рецептор, содержащий длинный спейсер. Данные показывают градиент цитолитической активности от приблизительно исходной активности у Т-клеток, экспрессирующих химерный рецептор из герцептина с коротким спейсером, до средней активности у Т-клеток, экспрессирующих химерный рецептор со средней длиной спейсера, и максимального лизиса у Т-клеток, которые экспрессировали химерный рецептор с длинным спейсером. Таким образом, внеклеточный спейсер оказывает определяющее влияние на распознавание опухолей Т-клетками, и эти данные дополнительно подтверждают необходимость в индивидуальном подборе конструкции химерных рецепторов на основании положения эпитопа в молекулах-мишенях, экспрессируемых в опухолях.
Пример 3. Индивидуализация длины и последовательности спейсера для оптимального распознавания и in vivo эффективности Т-клеток, модифицированных химерным рецептором CD19
Материалы и способы
Субъекты-люди
Образцы крови получали от здоровых доноров, которые предоставляли письменное информированное согласие на участие в протоколах исследования, утвержденных ученым советом Онкологического исследовательского центра Фреда Хатчинсона (FHCRC). Мононуклеарные клетки периферической крови (РВМС) выделяли путем центрифугирования на Ficoll-Hypaque (Sigma, Сент-Луис, Миссури) и подвергали криоконсервации в RPMI, 20% сыворотке человека и 10% диметилсульфоксиде.
Линии клеток
Линии клеток K562, Raji, JeKo-1 и 293Т получали из Американской коллекции типовых культур (Манассас, Виргиния) и культивировали согласно указаниям. Лентивирус, содержащий ген ffluc выше последовательности Т2А и eGFP, получали в клетках 293Т и применяли для трансдукции опухолевых клеток Raji и JeKo-1. Клетки Raji и JeKo-1 размножали после трансдукции лентивирусом, и eGFP-положительную субпопуляцию очищали путем сортировки.
Иммунофенотипирование
РВМС и линии Т-клеток окрашивали одним или несколькими из следующих конъюгированных моноклональных антител: CD3, CD4, CD8, CD25, CD45RA, CD45RO, CD62L, CD69 и изотипически сходный контроль (BD Biosciences). Окрашивание йодидом пропидия (PI, BD Biosciences) выполняли для установления различия между живыми и мертвыми клетками согласно указаниям производителя. Проточные анализы выполняли на FACSCanto®, очистку путем сортировки на FACSAriaII® (Becton Dickinson), и данные анализировали с помощью программного обеспечения FlowJo® (Treestar).
Конструирование вектора и получение лентивируса, кодирующего химерный рецептор CD19
СD19-специфические химерные рецепторы конструировали с применением: (1) сегментов цепей VL и VH СD19-специфического mAb FMC63 (SEQ ID NO: 3), соединенных линкерным пептидом (G4S)3 (SEQ ID NO: 12) (VL-линкер-VH); (2) спейсерного домена, полученного из Fc IgG4 (база данных Uniprot: Р01861, (SEQ ID NO: 13)), содержащего часть 'шарнирный участок'-СН2-СН3' (229 аминокислот, (SEQ ID NO:)) или 'шарнирный участок в отдельности' (12 аминокислот; (SEQ ID NO: 4)), при этом оба спейсера содержали замену S→Р в домене, представляющем собой шарнирный участок, находящуюся в положении 108 нативного белка Fc IgG4; (3) трансмембранного домена CD28 человека длиной 27 аминокислот (база данных Uniprot: P10747, (SEQ ID NO: 14); (4) сигнального модуля, содержащего (i) цитоплазматический домен CD28 человека длиной 41 аминокислота с заменой LL→GG, находящейся в положениях 186-187 нативного белка CD28 (SEQ ID NO: 14); или (ii) цитоплазматический домен 4-1ВВ человека длиной 42 аминокислоты (база данных Uniprot: Q07011, (SEQ ID NO: 15)); соединенный с (iii) цитоплазматическим доменом изоформы 3 CD3ζ человека длиной 112 аминокислот (база данных Uniprot: Р20963, (SEQ ID NO: 16)); (5) последовательности саморасщепляющегося пептида Т2А (SEQ ID NO: 8) и (6) усеченной последовательности рецептора эпидермального фактора роста (EGFR) (SEQ ID NO: 9).
Подвергнутые оптимизации кодонов нуклеотидные последовательности, кодирующие каждый трансген, синтезировали (LifeTechnologies, Карлсбад, Калифорния) и клонировали в лентивирусный вектор epHIV7 с применением сайтов рестрикции Nhe1 и Notl. Лентивирусный вектор epHIV7 получали из вектора pHIV7 путем замены промотора цитомегаловируса pHIV7 промотором EF-1.
Лентивирус, кодирующий химерный рецептор CD19 или tEGFR, получали в клетках 293Т, котрансфицировацных лентивирусным вектором и упаковочными векторами pCHGP-2, pCMV-Rev2 и pCMV-G с помощью реагента для трансфекции Calphos (Clontech). Среду меняли через 16 часов после трансфекции, и лентивирус собирали спустя 24, 48 и 72 часа.
Создание линий Т-клеток, экспрессирующих химерные рецепторы CD19
Очищенные путем сортировки центральные Т-клетки памяти (ТСM) CD8+ CD45RA-CD45RO+CD62L+ здоровых доноров активировали гранулами с антителами к CD3/CD28 (Life Technologies) согласно указаниям производителя и трансдуцировали надосадочной жидкостью, содержащей лентивирус (MOI=3), дополненной 1 пг/мл полибрена (Millipore), на 3-й день после активации путем центрифугирования при 2100 об./мин. в течение 45 минут при 32°С. Т-клетки размножали в RPMI, 10% сыворотке человека, 2 мМ L-глутамине и 1% пенициллине/стрептомицине (среде для CTL), дополняемых рекомбинантным IL-2 человека (rh) до конечной концентрации 50 Ед/мл каждые 48 часов. После размножения аликвоту каждой трансдуцированной линии Т-клеток окрашивали конъюгированным с биотином антителом к EGFR (рецептору эпителиального фактора роста) и гранулами стрептавидина (Miltenyi), и Т-клетки tEGFR+выделяли посредством иммуномагнитной селекции.
Затем субпопуляцию Т-клеток tEGFR+стимулировали облученными (8000 рад) ТМ ЕВV-LCL в соотношении T-клетки:LCL 1:7 и размножали в течение 8 дней в среде для CTL с добавлением 50 Ед/мл rh IL-2 каждые 48 часов.
Тесты с радиоактивным хромом, анализы секреции цитокинов и пролиферации с применением CFSE
Клетки-мишени метили 51Cr (PerkinElmer) в течение ночи, отмывали и инкубировали в трех повторностях при концентрации 1-2×103 клеток/лунка с эффекторными Т-клетками при различных соотношениях эффектора и мишени (Е : Т). Надосадочные жидкости собирали для γ-радиометрии после 4-часового инкубирования, и специфический лизис рассчитывали с помощью стандартной формулы. Для анализа секреции цитокинов клетки-мишени и эффекторные клетки высевали в лунки чашек в трех повторностях в соотношении Е : Т 2:1 (Raji) или 4:1 (K562/CD19 и K562), и измеряли IFN-γ, TF-α, IL-2, IL-4, IL-6 и IL-10 с помощью мультиплексного иммунологического анализа цитокинов (Luminex) в надосадочной жидкости, удаленной после 24-часового инкубирования.
Для анализа пролиферации Т-клетки метили 0,2 мкМ сложным сукцинимидиловым эфиром карбоксифлуоресцеина (CFSE, Invitrogen), отмывали и высевали в лунки чашек в трех повторностях с клетками-стимуляторами в соотношении 2:1 (Raji) или 4:1 (K562/CD19 и K562) в среде для CTL без экзогенных цитокинов. После 72-часового инкубирования клетки метили mAb к CD3 и йодидом пропидия (PI) для исключения мертвых клеток из анализа. Образцы анализировали с помощью проточной цитометрии, и клеточное деление живых Т-клеток CD3+оценивали путем разведения CFSE.
Эксперименты на мышах NOD/SCID и NOD/SCID/γc-/- (NSG)
Все эксперименты на мышах были одобрены Институциональным комитетом по содержанию и использованию животных FHCRC. Самок мышей NOD.CBI7-Prkdcscid/J (NOD/SCID) и NOD.Cg-PrkdcscidII2rgtmlwjl/SzJ (NSG) приобретали в Jackson Laboratory или разводили в условиях собственного производства (FHCRC). Мышам инъецировали внутривенно (i.v.) 0,5×106 опухолевых клеток Raji-ffluc посредством инъекции в хвостовую вену, и они получали инъекции Т-клеток, модифицированных химерными рецепторами, контрольных Т-клеток или PBS посредством инъекции в хвостовую вену согласно указаниям.
Для биолюминесцентной визуализации мыши получали внутрибрюшинные (i.p.) инъекции свежеприготовленного субстрата люциферина (Caliper Life Sciences, Массачусетс), ресуспендированного в PBS (15 мкг/г массы тела), и затем их подвергали анестезии изофлураном в индукционной камере. После индукции глубокой анестезии у мышей проводили визуализацию с помощью системы in vivo визуализации Xenogen IVIS (Caliper Life Sciences, Массачусетс) через 10, 12 и 14 минут после i.p. нъекции люциферина при времени экспозиции от 1 секунды до 1 минуты в режиме малого коэффициента биннинга для получения ненасыщенных изображений. Люциферазную активность анализировали с помощью программного обеспечения Living Image (Caliper Life Sciences, Массачусетс), и в областях, представляющих интерес, которые охватывали все тело каждой отдельной мыши, анализировали поток фотонов.
Статистические анализы
Статистические анализы выполняли с помощью программного обеспечения Prism (GraphPad, Калифорния), t-критерий Стьюдента применяли в виде двустороннего парного критерия с 95% доверительным интервалом, а результаты с р-значением р<0,05 считали значимыми. Статистический анализ выживаемости выполняли с помощью логарифмического рангового критерия, а результаты с р-значением р<0,05 считали значимыми.
Результаты
Получение политональных линий клеток CD8+, полученных из ТСM, которые экспрессируют химерные рецепторы CD19 с длинными и короткими внеклеточными спейсерами
Были сконструированы отдельные лентивирусные векторы, содержащие панель подвергнутых оптимизации кодонов генов химерного рецептора CD19, для исследования влияния длины внеклеточного спейсера на in vitro функционирование и in vivo противоопухолевую активность Т-клеток, модифицированных химерными рецепторами CD19. Каждый химерный рецептор содержал одноцепочечный вариабельный фрагмент, соответствующий последовательности СD19-специфического mAb FMC63 (scFv: VL-VH), спейсер, полученный из Fc IgG4, содержащий домен 'шарнирный участок-СН2-СН3' (229 аминокислот, длинный спейсер) или только домен 'шарнирный участок' (12 аминокислот, короткий спейсер), и сигнальный модуль CD3ζ с мембранно-проксимальными костимулирующими доменами CD28 или 4-1ВВ, в отдельности или в тандеме (фигура 13А). Кассета трансгена включала в себя усеченный EGFR (tEGFR) ниже гена химерного рецептора, отделенный расщепляемым элементом Т2А, служащий в качестве маркера трансдукции, селекции и in vivo отслеживания для Т-клеток, модифицированных.химерными рецепторами.
Выделяли популяцию центральных Т-клеток памяти (ТСM) CD8+ CD45RO+CD62L+ путем сортировки клеток из крови здоровых доноров для трансдукции и размножения в связи с лучшей способностью ТСМ к персистированию in vivo после адоптивного переноса. Т-клетки CD8+ стимулировали гранулами с антителами к CD3/28, трансдуцировали каждым из лентивирусных векторов и размножали в культуре в течение 18 дней до применения в экспериментах in vitro и in vivo (фигура 13 В). Сходные значения эффективности трансдукции получали с каждым из лентивирусных векторов (среднее значение 25%), и трансген-положительные Т-клетки обогащали до однородной чистоты путем иммуномагнитной селекции с помощью биотинилированных mAb к EGFR и гранул стрептавидина. После обогащения tEGFR каждую линию Т-клеток с химерными рецепторами CD19 размножали путем однократной стимуляции с помощью B-LCL CD19+ без видимых различий в кинетике роста in vitro между линиями Т-клеток, экспрессирующих различные конструкты химерных рецепторов CD19. После размножения маркер tEGFR экспрессировался на одинаковых уровнях на >90% Т-клеток, трансдуцированных каждым из векторов (фигура 13С).
Химерные рецепторы CD19 с длинным и коротким внеклеточным спейсерным доменом обеспечивают специфическую противоопухолевую реактивность in vitro
Проводили сравнение эффекторной функции полученных из ТСМ линий Т-клеток, модифицированных для экспрессии химерных рецепторов CD19 с костимулирующими сигнальными фрагментами CD28 и 4-1ВВ и коротким ('короткий/СD28; 'короткий/4-1ВВ') или длинным ('длинный/СD28'; 'длинный/4-1ВВ') внеклеточным спейсерным доменом, соответственно. Т-клеткам, экспрессирующим каждый из 4 конструктов химерных рецепторов CD19, придавали специфическую цитолитическую активность по отношению к клеткам лимфомы Raji и JeKo-1 CD19+ и к клеткам K562, которые были подвергнуты стабильной трансфекции с помощью CD19, но не к нативным клеткам K562 CD19-(фигура 14А). Количественные анализы выработки цитокинов в ответ на стимуляцию опухолевыми клетками K562/CD19 или Raji с помощью мультиплексного анализа цитокинов (Luminex) показали выработку IFN-γ, TNF-α, IL-2, IL-4, IL-6 и IL-10 Т-клетками, экспрессирующими каждый из химерных рецепторов CD19 (фигура 14 В). Т-клетки, экспрессирующие химерные рецепторы CD19 с костимулирующим доменом CD28, вырабатывали IFN-γ, TNF-α, IL-2 и IL-10 на значительно более высоких уровнях по сравнению с Т-клетками, экспрессирующими соответствующие конструкты с костимулирующим доменом 4-1ВВ (фигура 14В, С). Наблюдалась значительно более высокая выработка IFN-γ и значительно более низкая выработка IL-4 Т-клетками, экспрессирующими химерный рецептор CD19 'длинный/СD28', по сравнению с Т-клетками, экспрессирующими химерный рецептор CD19 'короткий/СD28'. Среди химерных рецепторов CD19 с костимулирующим сигнальным доменом 4-1ВВ были обнаружены значительно более высокие уровни секреции IFN-γ, TNF-α, IL-2, IL-4 и IL-10 в Т-клетках, экспрессирующих конструкт с коротким спейсерным доменом (фигура 14В, С).
Для анализа пролиферации Т-клеток, модифицированных каждым из химерных рецепторов CD19, после привлечения опухолевых клеток CD19+ применяли разведение красителя CFSE. Специфическую и интенсивную пролиферацию каждой из линий Т-клеток с химерным рецептором CD19 наблюдали спустя 72 часа после стимуляции с помощью K562/CD19 или Raji. Среднее число клеточных делений было выше для Т-клеток с химерным рецептором CD19 с костимулирующим доменом CD28 по сравнению с Т-клетками с химерным рецептором CD19 с костимулирующим доменом 4-1ВВ, что соответствовало более высокой выработке IL-2 Т-клетками, экспрессирующими химерный рецептор, содержащий CD28 (фигура 14B-D). Путем совместного окрашивания культуры с помощью CD3+ и PI также анализировали долю Т-клеток с химерными рецепторами, которые подвергались индуцируемой активацией клеточной гибели после стимуляции опухолевыми клетками K562/CD19 и Raji в конце 72-часового периода. Была обнаружена более высокая частота Т-клеток CD3+ CD8+ PI+ в линии клеток с химерным рецептором CD19 'длинный/4-1 ВВ', однако, наблюдалось немного клеток PI+с другими химерными рецепторами CD19 (фигура 14Е).
Этот анализ эффекторных функций in vitro находился в соответствии с предыдущими исследованиями, в которых сравнивали костимулирующие домены CD28 и 4-1ВВ, и не выявил различий в функционировании Т-клеток, что позволяет предположить, что в случае применения определенного конструкта химерного рецептора CD19 из этой панели будет отсутствовать противоопухолевая эффективность in vivo.
Т-клетки, экспрессирующие химерные рецепторы CD19 с короткими внеклеточными спейсерными доменами, но не с длинными внеклеточными спейсерными доменами, уничтожают опухоли Raji в моделях на мышах с иммунодефицитом Далее оценивали in vivo противоопухолевую эффективность Т-клеток, модифицированных каждым из химерных рецепторов CD19, у мышей с иммунодефицитом (NOD/SCID), которым трансплантировали клетки Raji, трансфицированные люциферазой светлячка (Raji-ffluc), что обеспечивает последовательные количественные анализы опухолевой нагрузки и распространения опухоли с помощью биолюминесцентной визуализации. У мышей NOD/SCID, которым инокулировали 0,5×106 клеток Raji-ffluc посредством инъекции в хвостовую вену, развивалась диссеминированная лимфома, которая без лечения приводила к параличу задних конечностей через приблизительно 3,5 недели, что влекло за собой необходимость эвтаназии. Мышей-опухоленосителей обрабатывали 2 дозами полученных из ТCM Т-клеток CD8+, модифицированных каждым из химерных рецепторов CD19 или контрольным вектором, кодирующим tEGFR, вводимыми в день 2 и день 9 после инокуляции опухоли (фигура 15А).
В этой модели только Т-клетки, модифицированные для экспрессии химерных рецепторов CD19 с коротким внеклеточным спейсерным доменом ('короткий/СD28' и 'короткий/4-1ВВ'), уничтожали опухоли Raji, в то время как у мышей, обработанных Т-клетками, экспрессирующими химерные рецепторы CD19 с длинным спейсером ('длинный/СD28' и 'длинный/4-1ВВ'), развивались системная лимфома и паралич задних конечностей с примерно идентичной кинетикой по сравнению с необработанными мышами или мышами, обработанными контрольными Т-клетками tEGFR+ (фигура 15В, С). Четко выраженное различие в противоопухолевой активности между химерными рецепторами CD19 с короткими и длинными спейсерными доменами было высокозначимым и воспроизводимым в нескольких экспериментах с линиями Т-клеток с химерными рецепторами, полученными от 3 разных здоровых доноров.
Модель лимфомы на NOD/SCID может быть субоптимальной для прогноза противоопухолевой активности в клинических условиях ввиду короткого интервала между инокуляцией опухоли и введением Т-клеток и большей устойчивости к трансплантации клеток человека по сравнению с линиями мышей с более тяжелым иммунодефицитом, такими как NOD/SCID/γc-/- (NSG). Поэтому оценку противоопухолевой активности при адоптивной терапии проводили в более клинически значимой модели, в которой мыши NSG имели развившуюся лимфому Raji-ffluc, а Т-клетки с химерным рецептором CD19 вводили спустя 7 дней, когда опухоль можно было легко выявить в костном мозге с помощью биолюминесцентной визуализации (фигура 16А). Для определения минимальной дозы Т-клеток, трансдуцированных химерным рецептором CD19 'короткий/4-IBB', которая требовалась для уничтожения развившихся опухолей Raji, проводили эксперименты по начальному титрованию дозы. Однократная доза 2,5×106 Т-клеток, экспрессирующих химерный рецептор CD19 'короткий/4-1ВВ', способствовала полной регрессии развившихся опухолей Raji и приводила к длительной безопухолевой выживаемости у 100% мышей (фигура 16В, С). При уровне дозы 2,5×106 Т-клетки легко выявляли в периферической крови мышей NSG в течение по меньшей мере трех недель после адоптивного переноса и уничтожения опухоли. Поэтому эта модель делала возможными сравнительные исследования как противоопухолевой активности, так и персистирования Т-клеток, модифицированных каждым из химерных рецепторов CD19 в панели (фигура 16D).
Затем группы мышей NSG, которым трансплантировали лимфому Raji, обрабатывали PBS в отдельности, однократной дозой 2,5×106 Т-клеток, экспрессирующих каждый из химерных рецепторов CD19, или Т-клетками, модифицированными контрольным вектором, кодирующим tEGFR (фигура 17А). В этой модели развившейся лимфомы Т-клетки, экспрессирующие химерные рецепторы CD19 с коротким внеклеточным спейсерным доменом и костимулирующими доменами 4-1ВВ или CD28 ('короткий/СD28' и 'короткий/4-1ВВ'), опосредовали полную регрессию опухоли в течение 7-10 дней, и все мыши проявляли безопухолевую выживаемость в течение >56 дней. В отличие от этого, мыши, обработанные Т-клетками, модифицированными для экспрессии химерных рецепторов CD19 с длинным спейсерным доменом ('длинный/СD28' и 'длинный/4-1ВВ'), проявляли прогрессирование опухоли, и их, а также мышей, обработанных контрольными Т-клетками с tEGFR, приходилось умерщвлять в сходное время (фигура 17В, С). Отсутствие in vivo противоопухолевой активности конструктов химерных рецепторов с длинными спейсерами было неожиданным, если учитывать способность Т-клеток, экспрессирующих эти конструкты, к лизису опухолевых клеток in vitro, и повышенную выработку IL-2 и пролиферацию после привлечения Т-клеток, экспрессирующих химерный рецептор CD19 'длинный/СD28', по сравнению с конструктами с 4-1ВВ.
Для получения представления о том, на чем основано отсутствие эффективности, выполняли последовательную проточную цитометрию на образцах периферической крови мышей с определенными интервалами после инфузии Т-клеток. Все мыши, обработанные Т-клетками, экспрессирующими химерные рецепторы CD19 'короткий/СD28’ и 'короткий/4-1ВВ', имели значительно более высокие уровни перенесенных Т-клеток CD19 в крови во все моменты времени после адоптивного переноса по сравнению с мышами, обработанными Т-клетками, экспрессирующими соответствующие химерные рецепторы CD19 с длинным внеклеточным спейсером (р<0,01) (фигура 17D). Значимых различий в персистировании Т-клеток в периферической крови мышей, которые получали Т-клетки, экспрессирующие химерные рецепторы CD19 с костимулирующими доменами CD28 или 4-1ВВ и короткими спейсерными доменами, не наблюдалось (фигура 17D).
In vivo противоопухолевая эффективность химерных рецепторов CD19 с длинными спейсерами не улучшалась путем повышения дозы Т-клеток или обеспечения дополнительного костимулирующего домена
Отсутствие in vivo противоопухолевой эффективности и более низкий уровень персистирующих Т-клеток с химерными рецепторами у мышей, обработанных Т-клетками, модифицированными химерными рецепторами CD19 с длинными спейсерными доменами, позволяло предположить, что эффективность можно улучшать путем повышения дозы Т-клеток с химерным рецептором или включения как домена CD28, так и домена 4-1BB в химерный рецептор для усиления передачи костимулирующего сигнала. Для оценки этой возможности ТСМ CD8+ модифицировали векторами химерных рецепторов CD19 'длинный/СD28', 'короткий/СD28' и 'длинный/СD28_4-1ВВ', и подтвердили, что химерный рецептор CD19 'длинный/СD28_4-1ВВ' обеспечивал специфический лизис и выработку цитокинов in vitro после распознавания клеток-мишеней CD19+ (фигура 18А-С). В соответствии с предыдущими исследованиями химерных рецепторов CD19, уровень выработки цитокинов Т-клетками, экспрессирующих химерный рецептор CD19 'CD28_4-1ВВ', и их пролиферации in vitro был ниже по сравнению с Т-клетками с идентичным конструктом с CD28 в отдельности и выше по сравнению с Т-клетками, экспрессирующими химерный рецептор CD19 'длинный 4-1ВВ' (фигура 18В, С).
Группы мышей NSG с развившимися опухолями Raji затем обрабатывали высокой дозой Т-клеток (10×106), экспрессирующих химерный рецептор CD19 'длинный/СD28', химерный рецептор CD19 'длинный/СD28_4-1ВВ', химерный рецептор CD19 'короткий/СD28' и только tEGFR. Опухолевую нагрузку измеряли с помощью биолюминесцентной визуализации и серийных анализов по методу проточной цитометрии образцов периферической крови, выполненных для определения частоты перенесенных Т-клеток. В соответствии с результатами предыдущих исследований с применением гораздо меньших доз Т-клеток, опухоли Raji были полностью уничтожены у мышей, обработанных Т-клетками, экспрессирующими химерный рецептор CD19 'короткий/СD28'. Однако, даже при в 4 раза более высокой дозе Т-клеток обработка Т-клетками, экспрессирующими химерный рецептор CD19 'длинный CD28' или химерный рецептор CD19 'длинный CD28 4-1ВВ', не обеспечивала ощутимого противоопухолевого эффекта (фигура 18D, E).
Поэтому повышение дозы Т-клеток с химерными рецепторами и добавление костимулирующего домена 4-1ВВ в химерные рецепторы CD19 оказались не в состоянии преодолеть отрицательное влияние более длинного спейсерного домена на противоопухолевую активность in vivo. Поэтому в данной модели противоопухолевая реактивность химерных рецепторов CD19 в значительной степени обусловлена длиной внеклеточного спейсерного домена, а не внутриклеточными костимулирующими сигнальными модулями.
Т-клетки, модифицированные химерными рецепторами CD19, которые имеют длинные внеклеточные спейсеры, подвергаются индуцируемой активацией клеточной гибели in vivo
Стремились определить потенциальные механизмы, лежащие в основе пониженной in vivo противоопухолевой активности Т-клеток, которые экспрессируют химерные рецепторы CD19 с длинными спейсерными доменами. Поскольку в крови присутствовали более низкие количества перенесенных Т-клеток, модифицированных для экспрессии химерных рецепторов CD19 с длинными спейсерными доменами, рассматривали возможность того, что Т-клетки не подвергались эффективной активации опухолевыми клетками in vivo или, наоборот, что они подвергались индуцируемой активацией Т-клеточной гибели in vivo. Поэтому Т-клетки, модифицированные химерными рецепторами CD19, и соответствующие контрольные Т-клетки метили с помощью CFSE, и эти Т-клетки вводили мышам-опухоленосителям NSG/Raji для исследования активации, пролиферации и выживаемости Т-клеток, модифицированных каждым из конструктов химерного рецептора CD19, в места локализации опухолей in vivo (фигура 19А). В конце их размножения in vitro и непосредственно перед мечением CFSE и инфузией мышам NSG, несущим развившиеся опухоли Raji, Т-клетки, трансдуцированные каждым из химерных рецепторов CD19, экспрессировали маркеры активации CD69 и CD25 на низких уровнях (фигура 19В).
Костный мозг получали из подгрупп мышей спустя 24 и 72 часа после инфузии Т-клеток для исследования частоты, активации и пролиферации перенесенных Т-клеток. Через 24 часа опухолевые клетки (CD45+CD3-) присутствовали в костном мозге во всех группах обработки, и большая часть Т-клеток с химерными рецепторами, но не контрольных Т-клеток, имела повышенную экспрессию CD69 и CD25. Измеримого разведения CFSE в перенесенных Т-клетках с химерными рецепторами не наблюдалось (фигура 19С). Как CD69, так и CD25 экспрессировались в более высокой доле Т-клеток, модифицированных химерными рецепторами CD19 с 'длинным спейсером', что позволяло предположить, что эти клетки могли получать более сильный стимул по сравнению с Т-клетками с химерными рецепторами CD19 с 'коротким спейсером' (фигура 19С). Несмотря на доказательство активации Т-клеток, через 24 часа в костном мозге мышей, обработанных Т-клетками, модифицированными конструктами 'длинный спейсер' с CD28 и 4-1ВВ, наблюдалось значительно более низкое количество Т-клеток с химерными рецепторами по сравнению с костным мозгом мышей, обработанных Т-клетками, модифицированными конструктами 'короткий спейсер' с CD28 и 4-1ВВ или контрольным вектором, кодирующим tEGFR (фигура 19С, Е).
Через 72 часа после переноса Т-клеток частота Т-клеток, экспрессирующих химерные рецепторы CD19 'короткий CD28' и 'короткий/4-1ВВ', возрастала в от 3 до >10 раз в костном мозге и селезенке, и эти клетки подвергались нескольким клеточным делениям (фигура 19D, Е). Контрольные Т-клетки tEGFR+ сохранялись в костном мозге и селезенке через 72 часа на уровне, сходном с уровнем, наблюдаемым через 24 часа, и не делились, что определяли по разведению CFSE. В отличие от этого, количество Т-клеток, экспрессирующих химерные рецепторы CD19 'длинный/СD28' и 'длинный/4-1ВВ', не повышалось в костном мозге и селезенке (фигура 19D, Е). В соответствии с более низким количеством клеток, анализ окрашивания с помощью CFSE в живых Т-клетках PI- с химерными рецепторами CD19 'длинный/СD28' и 'длинный/4-1ВВ' показал, что эти клетки подвергались гораздо меньшему числу клеточных делений по сравнению с Т-клетками с химерными рецепторами CD19 'короткий/СD28' и 'короткий/4-1ВВ' (фигура 19D). При анализе данных проточной цитометрии для включения Т-клеток PI+ была выявлена гораздо более высокая частота Т-клеток PI+ CD3+ в костном мозге и селезенке мышей, которые получали Т-клетки с химерным рецептором CD19 с 'длинными спейсерными' доменами, что демонстрирует, что значительная доля Т-клеток, несмотря на активацию опухолью in vivo, подверглась клеточной гибели (фигура 19F). В соответствии с биолюминесцентной визуализацией, опухолевые клетки Raji CD45+CD3- присутствовали в больших количествах в костном мозге мышей, обработанных Т-клетками, экспрессирующими химерные рецепторы CD19 с длинными спейсерными доменами или экспрессирующими только tEGFR, по сравнению с мышами, обработанными химерными рецепторами CD19 с короткими спейсерными доменами (фигура 19D, Е, G).
В целом, эти данные представляют доказательство того, что химерные рецепторы CD19 с длинным внеклеточным спейсерным доменом, несмотря на опосредование аналогичных или лучших эффекторной функции и распознавания опухолей in vivo, вызывают индуцируемую активацией клеточную гибель in vivo на высоком уровне и не в состоянии уничтожать развившуюся лимфому.
Обсуждение
Химерные рецепторы представляют собой искусственные рецепторы, которые включают в себя внеклеточный антиген-связывающий scFv, спейсерный домен, который обеспечивает отделение scFv от клеточной мембраны, и внутриклеточный сигнальный модуль, который опосредует активацию Т-клеток. Химерные рецепторы, которые содержат scFv, полученный из СD19-специфического mAb FMC63, изучаемого в данном документе, перешли на этап исследования в клинических испытаниях у пациентов с В-клеточными злокачественными опухолями. В различных испытаниях противоопухолевая активность и персистирование Т-клеток значительно варьировали. Каждое из этих клинических испытаний отличалось по потенциально определяющим переменным, включая различные векторы для переноса генов, методики культивирования клеток и режимы кондиционирования перед переносом Т-клеток с химерным рецептором CD19.
Была исследована возможность того, что внеклеточный спейсерный домен химерных рецепторов CD19 может быть важным определяющим фактором противоопухолевой активности in vivo, независимо от передачи костимулирующего сигнала, обеспечиваемой химерным рецептором. Были получены спейсерные домены из Fc IgG4, обеспечивающие высокие уровни экспрессии химерных рецепторов на клеточной поверхности и с меньшей вероятностью вызывающие распознавание клетками врожденной иммунной системы по сравнению с другими изотопами IgG. В разработке конструктов с длинным (229 аминокислот) спейсером применяли 'шарнирный участок-СН2-СН3' IgG4, а в химерных рецепторах с коротким (12 аминокислот) спейсером применяли домен IgG4 'шарнирный участок'. Для сравнения отдельных конструктов химерных рецепторов применяли очищенные (>90%) положительные по химерному рецептору Т-клетки CD8+, полученные из ТСМ, для устранения различий в клеточном составе и частоте трансдукции как потенциального источника погрешности при анализе in vitro и in vivo функционирования. Было показано, что ТСМ CD8+ имеют лучшие характеристики для адоптивной иммунотерапии по сравнению с другими более распространенными субпопуляциями Т-клеток в крови, которые плохо персистируют и неэффективны в терапии опухолей. Т-клетки с химерным рецептором CD19 получали с помощью стандартизированного протокола культивирования, который подобен протоколу культивирования, применяемому для получения' Т-клеток с химерными рецепторами для клинических испытаний. Данные показывают, что химерные рецепторы CD19 с коротким спейсером 'шарнирный участок' IgG4 обеспечивали сильную противоопухолевую реактивность in vitro и in vivo, в то время как соответствующие химерные рецепторы CD19 с длинным спейсером 'шарнирный участок-СН2-СН3' IgG4, несмотря на аналогичную или более высокую реактивность in vitro, не были в состоянии обеспечивать значительные противоопухолевые эффекты в моделях лимфомы на мышах. Длина спейсерного домена оказалась решающим элементом для in vivo противоопухолевой активности, и отсутствие эффективности химерного рецептора с 'длинным спейсером' нельзя было преодолеть путем повышения дозы Т-клеток.
Также были обнаружены основные различия в секреции цитокинов и пролиферации in vitro между Т-клетками, экспрессирующими химерные рецепторы CD19, содержащие костимулирующие домены CD28 и 4-1ВВ, при этом CD28 усиливал секрецию IFN-γ, IL-2 и TNF-α по сравнению с 4-1ВВ. Химерные рецепторы CD19, которые имели тандем CD28_4-1BB, также обеспечивали выработку этих цитокинов на более высоких уровнях по сравнению с химерными рецепторами, содержащими только 4-1ВВ. Однако, данные показывают, что эти различия в in vitro функционировании не были прогностическими для in vivo противоопухолевой эффективности, поскольку химерные рецепторы CD19 с костимулирующим доменом CD28 или 4-1ВВ и коротким спейсером были аналогичным образом эффективными в уничтожении запущенных развившихся опухолей Raji у мышей NSG. В отличие от этого, химерные рецепторы CD19 с субоптимальной длиной спейсера и CD28, 4-1ВВ или обоими костимулирующими доменами, несмотря на обеспечение функционирования in vitro, подобного таковому в случае применения идентичного конструкта химерного рецептора с коротким спейсерным доменом, не имели значительной противоопухолевой активности in vivo, что демонстрирует роль длины спейсера в in vivo функционировании Т-клеток с химерными рецепторами.
Исследования дают представление о механизме, ответственном за отсутствие in vivo эффективности химерных рецепторов CD19 с длинными спейсерными доменами. Т-клетки, экспрессирующие химерные рецепторы CD19 как с коротким, так и с длинным спейсерными доменами, можно было выявить в костном мозге и селезенке после адоптивного переноса мышам NSG, несущим развившуюся лимфому Raji, и большинство из них активировалось, как было показано по повышению экспрессии CD25 и CD69. Однако, для Т-клеток, модифицированных для экспрессии химерного рецептора CD19 с длинным спейсерным доменом, проявлялось резкое падение количества клеток в отличие от выраженного in vivo размножения Т-клеток, экспрессирующих химерные рецепторы CD19 с коротким спейсерным доменом. Падение количества Т-клеток являлось следствием гораздо более высоких уровней клеточной гибели в первые 72 часа после адоптивного переноса по сравнению с Т-клетками с короткими спейсерными доменами и контрольными Т-клетками, которые не экспрессировали химерный рецептор CD19. В целом, эти данные указывают, что распознавание опухолевых клеток in vivo приводило к гибели Т-клеток, экспрессирующих химерные рецепторы CD19 с длинными спейсерными доменами. Подобным механизмом можно объяснить короткую продолжительность и низкие уровни персистирования Т-клеток в клинических испытаниях, в которых использовали химерные рецепторы CD19 с длинным спейсером (14).
Исследования, описанные в данном документе, впервые показывают, что спейсерные домены химерных рецепторов CD19, которые не имели присущих свойств передачи сигналов, оказывали существенные эффекты на in vivo противоопухолевую активность независимо от передачи костимулирующего сигнала, и определяют важность анализа оптимального состава этого участка при разработке химерных рецепторов для клинических путей применения.
Вышеизложенное является иллюстрацией настоящего изобретения и не подразумевает его ограничение. Настоящее изобретение определено следующей формулой изобретения, при этом эквиваленты пунктов формулы изобретения подлежат включению в нее. Все литературные источники и документы, упомянутые в данном документе, включены в данный документ с помощью ссылки.





Праймеры для секвенирования


NVFSCSVMHE ALHNHYTQKS LSLSLGK
1-98 CH1
99-110 Шарнирный участок
111-220 CH2
221-327 СН3
Положение 108 S→P

SKRSRLLHSD YMNMTPRRPG PTRKHYQPYA PPRDFAAYRS
1-18 сигнальный пептид
19-152 внеклеточный домен
153-179 трансмембранный домен
180-220 внутриклеточный домен
Положения 186-187 LL→GG

CSCRFPEEEE GGCEL
1-23 сигнальный пептид
24-186 внеклеточный домен
187-213 трансмембранный домен
214-255 внутриклеточный домен

EAYSEIGMKG ERRRGKGHDG LYQGLSTATK DTYDALHMQA LPPR
1-21 сигнальный пептид
22-30 внеклеточный домен
31-51 трансмембранный домен
52-164 внутриклеточный домен
61-89 ITAM1
100-128 ITAM2
131-159 ITAM3






































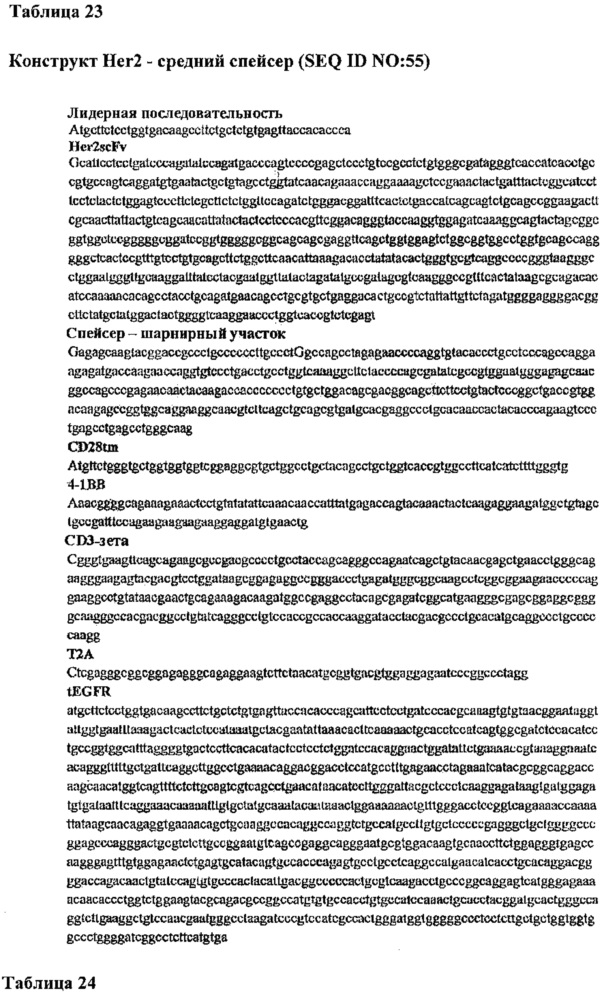





























































































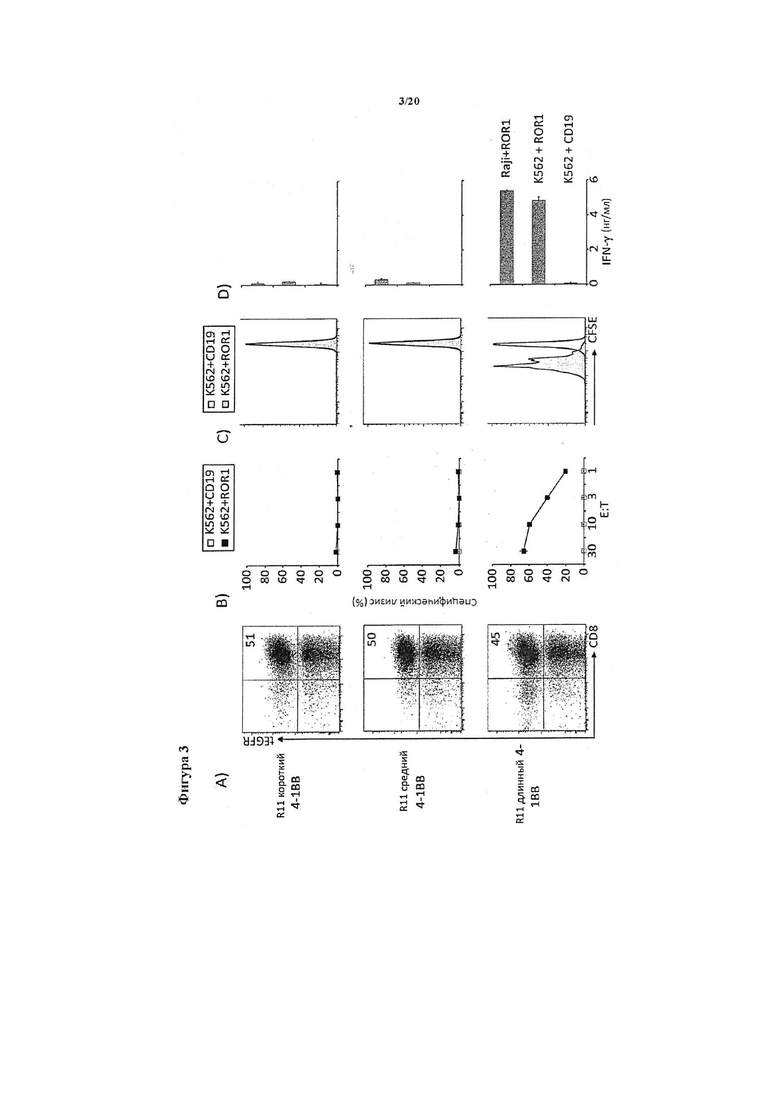

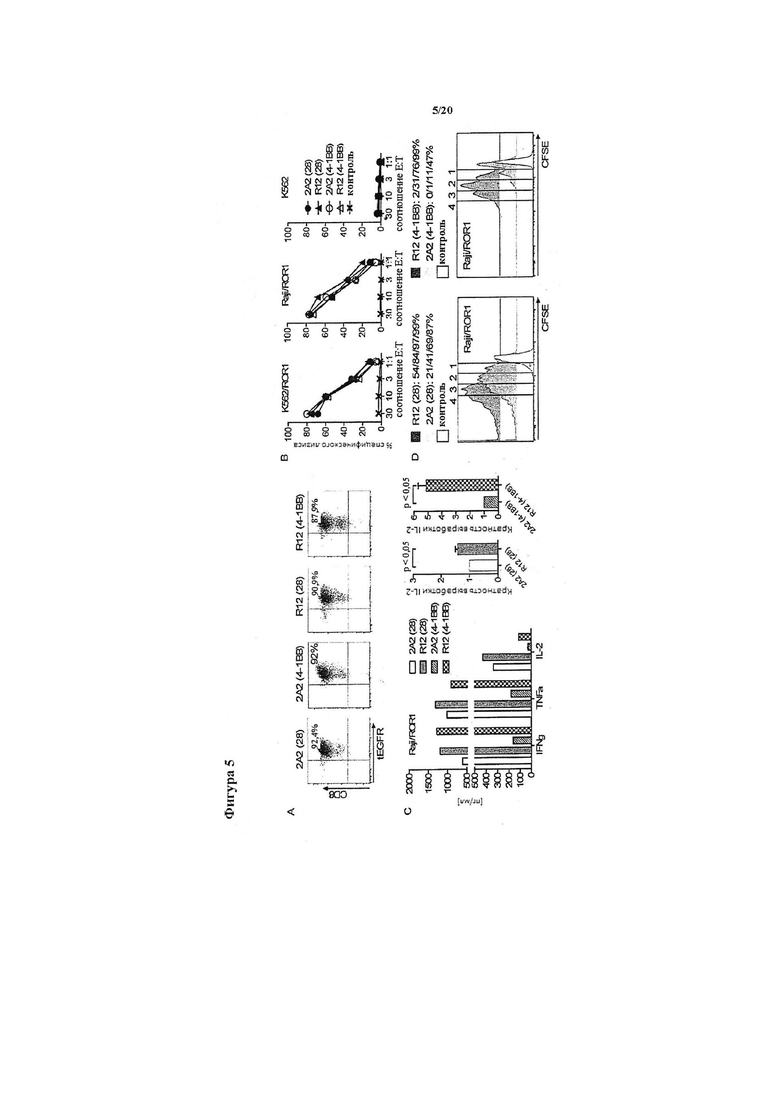



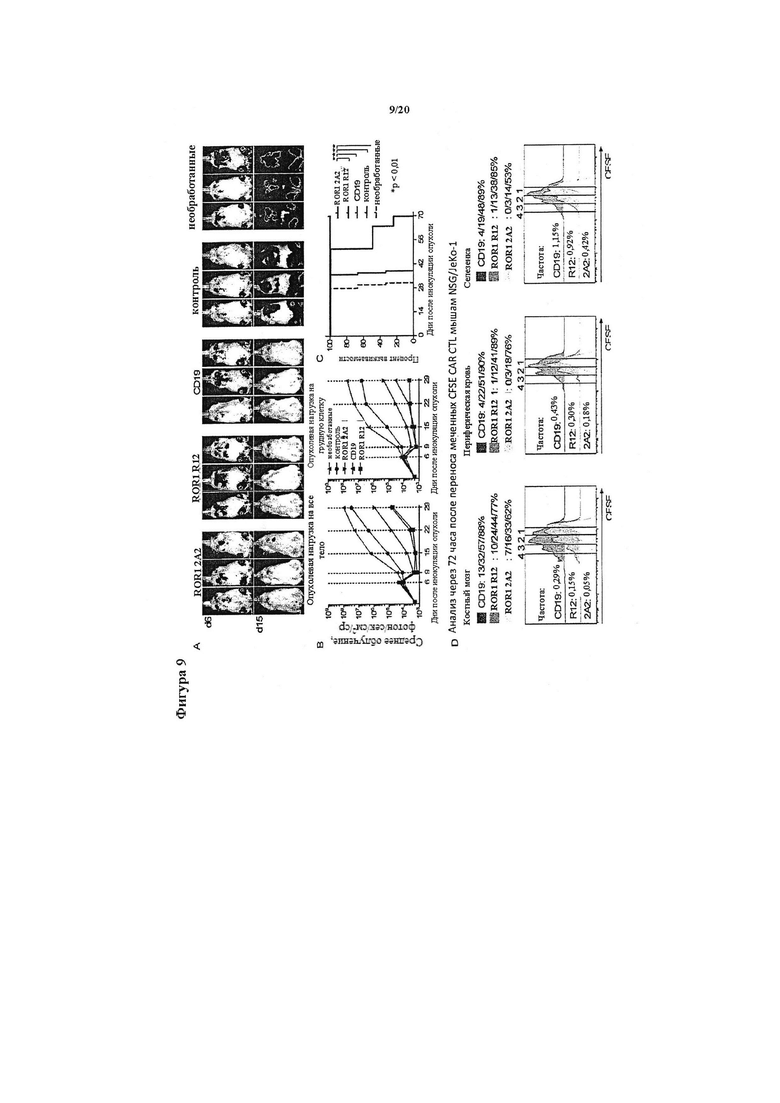



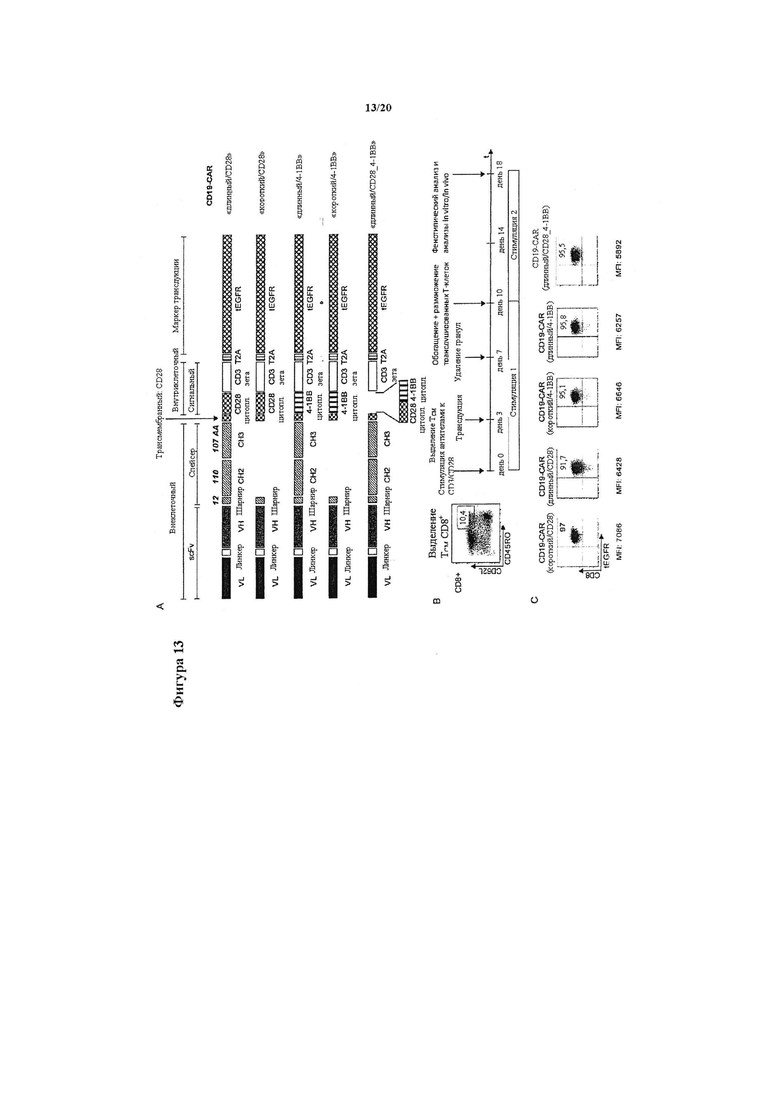





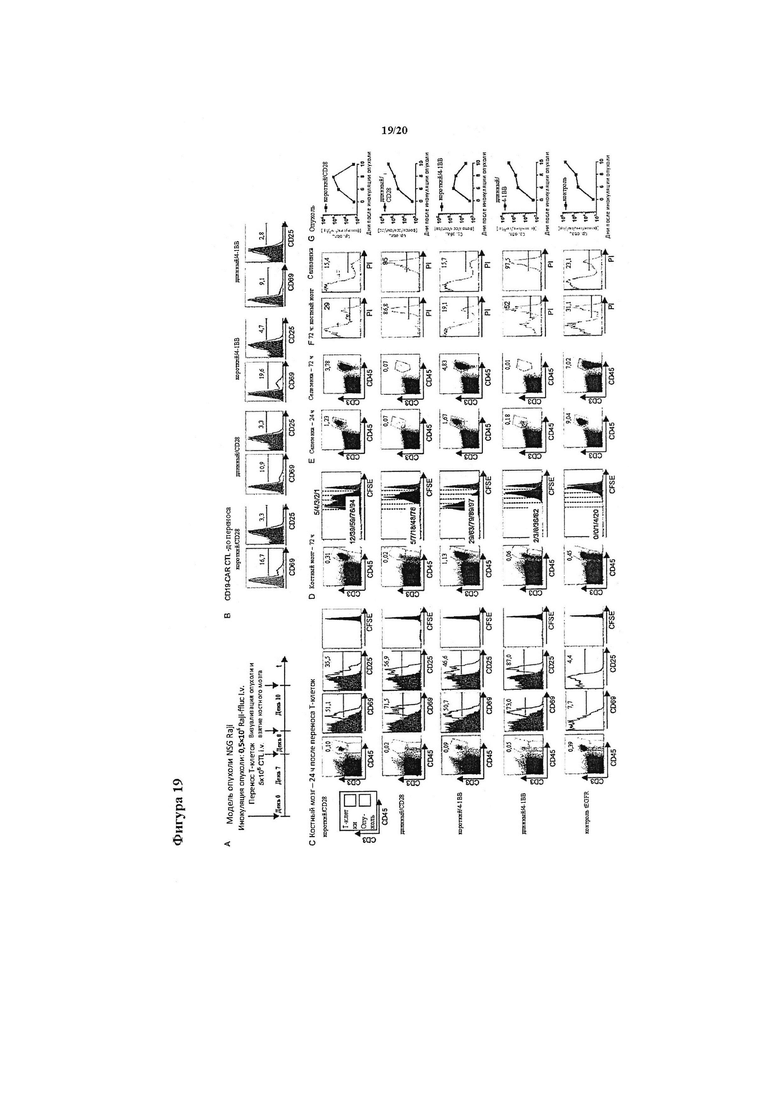
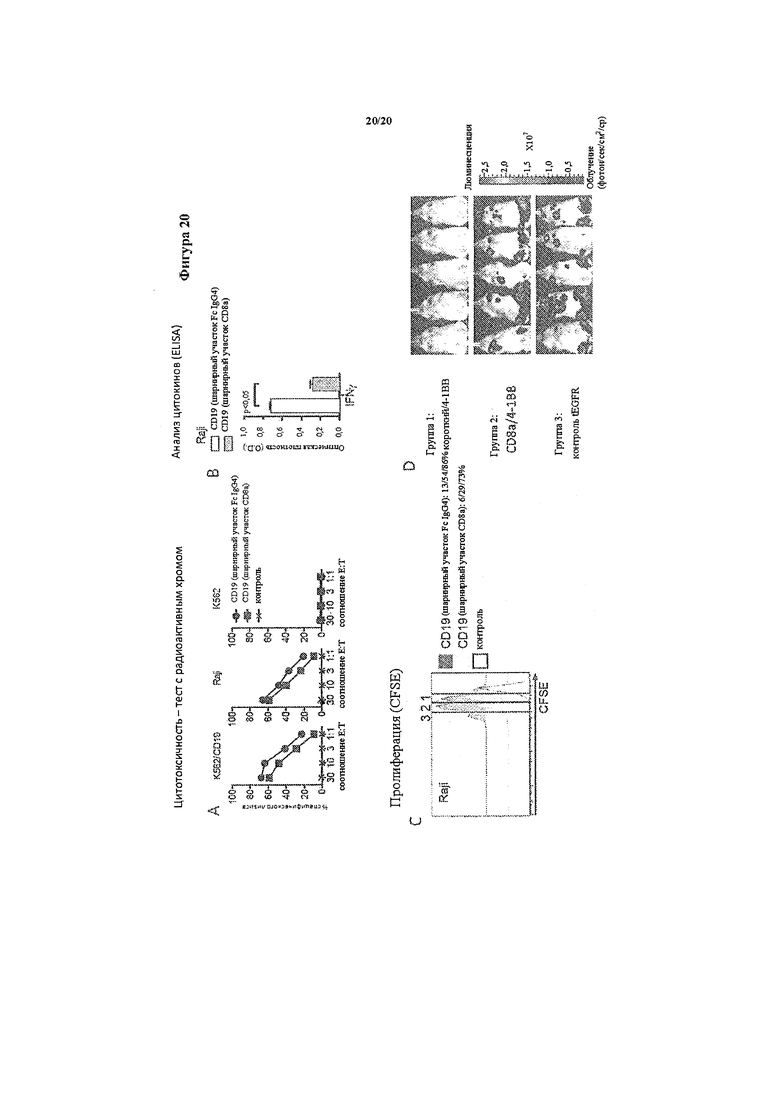
Изобретение относится к области биотехнологии. Представлена нуклеиновая кислота, кодирующая химерный рецептор, специфично связывающийся с CD19. Нуклеиновая кислота содержит полинуклеотид, кодирующий лиганд-связывающий домен, полинуклеотид, кодирующий трансмембранный домен, полинуклеотид, кодирующий полипептидный спейсер между лиганд-связывающим и трансмембранным доменами, и полинуклеотид, кодирующий внутриклеточный сигнальный домен. Полипептидный спейсер имеет длину 15 аминокислот или менее и содержит аминокислотную последовательность X1PPX2P, где X1 представляет собой цистеин, глицин или аргинин, а X2 – цистеин или треонин. Также предложены полипептидные химерные рецепторы, экспрессионные векторы, клетки-хозяева, способы и композиции для обеспечения и/или усиления иммунных ответов, опосредованных клеточной иммунотерапией, например, посредством адоптивного переноса центральных T-клеток памяти CD8+ или комбинаций центральных T-клеток памяти и Т-клеток CD4+, генетически модифицированных для экспрессии химерного рецептора. Изобретение обеспечивает повышенную выживаемость и in vivo эффективность Т-клеток, модифицированных химерным рецептором, и может быть использовано для терапии рака. 9 н. и 46 з.п. ф-лы, 20 ил., 24 табл., 3 пр.
1. Нуклеиновая кислота, кодирующая химерный рецептор, который специфично связывается с CD19, содержащая:
(a) полинуклеотид, кодирующий лиганд-связывающий домен, который специфично связывается с CD19;
(b) полинуклеотид, кодирующий трансмембранный домен;
(c) полинуклеотид, кодирующий полипептидный спейсер, расположенный между лиганд-связывающим доменом и трансмембранным доменом, где полипептидный спейсер имеет длину 15 аминокислот или менее и содержит аминокислотную последовательность X1PPX2P, где X1 представляет собой цистеин, глицин или аргинин, а X2 представляет собой цистеин или треонин (SEQ ID NO:1); и
(d) полинуклеотид, кодирующий внутриклеточный сигнальный домен, содержащий сигнальный домен CD3ζ и костимулирующий домен.
2. Нуклеиновая кислота по п. 1, где лиганд-связывающий домен представляет собой или содержит одноцепочечный вариабельный фрагмент (scFv).
3. Нуклеиновая кислота по п. 2, где scFv содержит вариабельный домен тяжелой цепи (VH) и/или вариабельный домен легкой цепи (VL), разделенные пептидным линкером.
4. Нуклеиновая кислота по любому из пп. 1-3, где полипептидный спейсерный участок содержит весь шарнирный участок из IgG1, IgG2 или IgG4 человека или его часть, или его модифицированную версию, при этом его модифицированная версия имеет длину 15 аминокислот или менее и содержит аминокислотную последовательность X1PPX2P, где X1 представляет собой цистеин, глицин или аргинин, а X2 представляет собой цистеин или треонин (SEQ ID NO:1).
5. Нуклеиновая кислота по любому из пп. 1-4, где полипептидный спейсер содержит модифицированный шарнирный участок из IgG4 человека, при этом модифицированный шарнирный участок из IgG4 человека имеет длину 15 аминокислот или менее и содержит аминокислотную последовательность X1PPX2P, где X1 представляет собой цистеин, глицин или аргинин, а X2 представляет собой цистеин или треонин (SEQ ID NO:1).
6. Нуклеиновая кислота по любому из пп. 1-4, где полипептидный спейсер содержит аминокислотную последовательность SEQ ID NO:17, SEQ ID NO:18, SEQ ID NO:21, SEQ ID NO:51, SEQ ID NO:52 или SEQ ID NO:53.
7. Нуклеиновая кислота по любому из пп. 1-6, где полипептидный спейсер состоит из аминокислотной последовательности SEQ ID NO:51.
8. Нуклеиновая кислота по любому из пп. 1-6, где полипептидный спейсер состоит из аминокислотной последовательности SEQ ID NO:21.
9. Нуклеиновая кислота по любому из пп. 1-8, где полипептидный спейсер не содержит внеклеточную часть CD8 и не содержит внеклеточную часть CD28.
10. Нуклеиновая кислота по любому из пп. 1-9, где полипептидный спейсер имеет длину 12 аминокислот или менее.
11. Нуклеиновая кислота по любому из пп. 1-10, где полипептидный спейсер имеет длину 12 аминокислот.
12. Нуклеиновая кислота по любому из пп. 1-11, где лиганд-связывающий домен содержит вариабельный домен тяжелой цепи (VH), который характеризуется по меньшей мере 90% идентичностью по отношению к VH участку антитела FMC63 или последовательности домена цепи VH, присутствующей в аминокислотной последовательности, кодируемой SEQ ID NO:3, и/или вариабельный домен легкой цепи (VL), который характеризуется по меньшей мере 90% идентичностью по отношению к VL участку антитела FMC63 или последовательности домена цепи VL, присутствующей в аминокислотной последовательности, кодируемой SEQ ID NO:3.
13. Нуклеиновая кислота по любому из пп. 1-11, где лиганд-связывающий домен содержит вариабельный домен легкой цепи (VL), содержащий CDRL1, CDRL2 и CDRL3 антитела FMC63 или последовательность домена цепи VL, присутствующую в аминокислотной последовательности, кодируемой SEQ ID NO:3, и вариабельный домен тяжелой цепи (VH), содержащий CDRH1, CDRH2 и CDRH3 антитела FMC63 или последовательность домена цепи VH, присутствующую в аминокислотной последовательности, кодируемой SEQ ID NO:3.
14. Нуклеиновая кислота по любому из пп. 1-13, где лиганд-связывающий домен содержит вариабельный домен легкой цепи (VL), содержащий последовательности RASQDISKYLN, SRLHSGV и GNTLPYTFG, вариабельный домен тяжелой цепи (VH), содержащий последовательности DYGVS, VIWGSETTYYNSALKS и YAMDYWG.
15. Нуклеиновая кислота по любому из пп. 1-14, где:
лиганд-связывающий домен связывается с эпитопом CD19, расположенным на таком же расстоянии от мембраны клетки или ближе к ней, по сравнению с эпитопом, связанным FMC63; и/или
лиганд-связывающий домен проявляет по меньшей мере такую же аффинность к CD19 по сравнению с FMC63.
16. Нуклеиновая кислота по любому из пп. 3-15, где пептидный линкер содержит аминокислотную последовательность SEQ ID NO:36.
17. Нуклеиновая кислота по любому из пп. 3-16, где scFv имеет ориентацию VL-линкер-VH.
18. Нуклеиновая кислота по любому из пп. 1-17, где трансмембранный домен содержит трансмембранный домен CD8 или CD28, а костимулирующий домен содержит сигнальный домен 4-1BB.
19. Нуклеиновая кислота по любому из пп. 1-18, где трансмембранный домен содержит трансмембранный домен CD8 или CD28, или полинуклеотид, кодирующий трансмембранный домен, содержит или состоит из полинуклеотида, представленного в SEQ ID NO:5.
20. Нуклеиновая кислота по любому из пп. 1-17 и 19, где:
костимулирующий домен содержит весь сигнальный домен 4-1BB или его часть, необязательно аминокислоты 214-255 SEQ ID NO:15; или
костимулирующий домен содержит весь сигнальный домен CD28 или его часть, необязательно аминокислоты 180-220 SEQ ID NO:14.
21. Нуклеиновая кислота по любому из пп. 1-5, 9-17, 19 и 20, где:
полипептидный спейсер содержит модифицированный шарнирный участок из IgG4 человека, при этом модифицированный шарнирный участок из IgG4 человека имеет длину 15 аминокислот или менее и содержит аминокислотную последовательность X1PPX2P, где X1 представляет собой цистеин, глицин или аргинин, а X2 представляет собой цистеин или треонин (SEQ ID NO:1);
трансмембранный домен содержит трансмембранный домен CD28; и
костимулирующий домен содержит сигнальный домен 4-1BB или сигнальный домен CD28.
22. Нуклеиновая кислота по любому из пп. 1-5, 9-17 и 19-21, где:
полипептидный спейсер содержит аминокислотную последовательность SEQ ID NO:21;
трансмембранный домен содержит трансмембранный домен CD28; и
костимулирующий домен содержит сигнальный домен 4-1BB или сигнальный домен CD28.
23. Нуклеиновая кислота по любому из пп. 1-5 и 9-21, где:
лиганд-связывающий домен содержит scFv, имеющий ориентацию VL-линкер-VH;
полипептидный спейсер содержит модифицированный шарнирный участок из IgG4 человека, при этом модифицированный шарнирный участок из IgG4 человека имеет длину 15 аминокислот или менее и содержит аминокислотную последовательность X1PPX2P, где X1 представляет собой цистеин, глицин или аргинин, а X2 представляет собой цистеин или треонин (SEQ ID NO:1);
трансмембранный домен содержит трансмембранный домен CD28; и
костимулирующий домен содержит сигнальный домен 4-1BB.
24. Нуклеиновая кислота по любому из пп. 1-23, где внутриклеточный сигнальный домен состоит из костимулирующего домена, который представляет собой сигнальный домен 4-1BB, и сигнального домена CD3ζ.
25. Нуклеиновая кислота по любому из пп. 18 и 20-24, где сигнальный домен 4-1BB содержит аминокислотную последовательность, кодируемую SEQ ID NO:6.
26. Нуклеиновая кислота по любому из пп. 1-25, где сигнальный домен CD3ζ содержит аминокислотную последовательность, кодируемую SEQ ID NO:7.
27. Нуклеиновая кислота по любому из пп. 1-5 и 9-26, где:
лиганд-связывающий домен содержит scFv, имеющий ориентацию VL-линкер-VH;
полипептидный спейсер содержит модифицированный шарнирный участок из IgG4 человека, где модифицированный шарнирный участок имеет длину 15 аминокислот или менее и содержит аминокислотную последовательность X1PPX2P, где X1 представляет собой цистеин, глицин или аргинин, а X2 представляет собой цистеин или треонин (SEQ ID NO:1);
трансмембранный домен содержит аминокислотную последовательность, кодируемую полинуклеотидной последовательностью SEQ ID NO:5; и
внутриклеточный сигнальный домен содержит аминокислотную последовательность, кодируемую полинуклеотидной последовательностью SEQ ID NO:6, и аминокислотную последовательность, кодируемую SEQ ID NO:7.
28. Нуклеиновая кислота по п. 27, где:
модифицированный шарнирный участок из IgG4 человека содержит аминокислотную последовательность SEQ ID NO:51 или аминокислотную последовательность SEQ ID NO:21.
29. Нуклеиновая кислота по любому из пп. 1-11 и 15-28, где:
лиганд-связывающий домен содержит scFv, содержащий домен цепи VH, который характеризуется по меньшей мере 90% идентичностью последовательности по отношению к последовательности домена цепи VH, присутствующей в аминокислотной последовательности, кодируемой SEQ ID NO:3, или домен цепи VH антитела FMC63, и домен цепи VL, который характеризуется по меньшей мере 90% идентичностью последовательности по отношению к последовательности домена цепи VL, присутствующей в аминокислотной последовательности, кодируемой SEQ ID NO:3, или домен цепи VL антитела FMC63;
полипептидный спейсер содержит аминокислотную последовательность SEQ ID NO:51;
трансмембранный домен содержит аминокислотную последовательность, кодируемую SEQ ID NO:5; и
внутриклеточный сигнальный домен содержит аминокислотную последовательность, кодируемую SEQ ID NO:7, и содержит аминокислотную последовательность, кодируемую SEQ ID NO:6.
30. Нуклеиновая кислота по любому из пп. 2-11 и 15-29, где:
scFv содержит домен цепи VH, который характеризуется по меньшей мере 90% идентичностью последовательности по отношению к последовательности домена цепи VH, присутствующей в аминокислотной последовательности, кодируемой полинуклеотидной последовательностью SEQ ID NO:3, и домен цепи VL, который характеризуется по меньшей мере 90% идентичностью последовательности по отношению к последовательности домена цепи VL, присутствующей в аминокислотной последовательности, кодируемой SEQ ID NO:3;
полипептидный спейсер состоит из аминокислотной последовательности, кодируемой SEQ ID NO:4, и/или состоит из аминокислотной последовательности SEQ ID NO:21;
трансмембранный домен содержит аминокислотную последовательность, кодируемую SEQ ID NO:5; и
внутриклеточный сигнальный домен содержит аминокислотную последовательность, кодируемую SEQ ID NO:6, и аминокислотную последовательность, кодируемую SEQ ID NO:7.
31. Нуклеиновая кислота по любому из пп. 1-30, где нуклеиновая кислота кодирует химерный рецептор, кодируемый конструкцией нуклеиновой кислоты, содержащей последовательность SEQ ID NO:10 или полинуклеотидную последовательность, которая характеризуется по меньшей мере 90% идентичностью по отношению к ней.
32. Нуклеиновая кислота по любому из пп. 1-31, где нуклеиновая кислота содержит:
(a) последовательность SEQ ID NO:3, кодирующую анти-CD19 scFv;
(b) последовательность SEQ ID NO:4, кодирующую шарнирный спейсер IgG4;
(c) последовательность SEQ ID NO:5, кодирующую трансмембранный домен CD28;
(d) последовательность SEQ ID NO:6, кодирующую костимулирующий домен 4-1BB;
(e) последовательность SEQ ID NO:7, кодирующую сигнальный домен CD3ζ.
33. Полипептидный химерный рецептор, который специфически связывается с CD19, кодируемый нуклеиновой кислотой по любому из пп. 1-32.
34. Полипептидный химерный рецептор, который специфически связывается с CD19, кодируемый конструкцией нуклеиновой кислоты, содержащей последовательность SEQ ID NO:10.
35. Вектор экспрессии, содержащий нуклеиновую кислоту по любому из пп. 1-32 или нуклеиновую кислоту, кодирующую полипептидный химерный рецептор по п. 33 или 34, необязательно содержащий полинуклеотид, кодирующий маркерную последовательность.
36. Вектор экспрессии по п. 35, дополнительно содержащий полинуклеотид, кодирующий маркерную последовательность, где полинуклеотид, кодирующий маркерную последовательность, функционально связан в рамке с полинуклеотидами, кодирующими химерный рецептор.
37. Вектор экспрессии по п. 36, где полинуклеотиды, кодирующие химерный рецептор, и полинуклеотид, кодирующий маркерную последовательность, разделены линкерной последовательностью.
38. Вектор экспрессии по п. 36 или 37, где маркерная последовательность представляет собой усеченную последовательность EGFR.
39. Вектор экспрессии по п. 38, где усеченная последовательность EGFR кодируется полинуклеотидом SEQ ID NO:9.
40. Вектор экспрессии по любому из пп. 37-39, где линкерная последовательность содержит пептид Т2А.
41. Вектор экспрессии по п. 40, где пептид Т2А кодируется полинуклеотидом SEQ ID NO:8.
42. Вектор экспрессии, кодирующий аминокислотную последовательность
MLLLVTSLLLCELPHPAFLLIPDIQMTQTTSSLSASLGDRVTISCRASQDISKYLNWYQQKPDGTVKLLIYHTSRLHSGVPSRFSGSGSGTDYSLTISNLEQEDIATYFCQQGNTLPYTFGGGTKLEITGSTSGSGKPGSGEGSTKGEVKLQESGPGLVAPSQSLSVTCTVSGVSLPDYGVSWIRQPPRKGLEWLGVIWGSETTYYNSALKSRLTIIKDNSKSQVFLKMNSLQTDDTAIYYCAKHYYYGGSYAMDYWGQGTSVTVSSESKYGPPCPPCPMFWVLVVVGGVLACYSLLVTVAFIIFWVKRGRKKLLYIFKQPFMRPVQTTQEEDGCSCRFPEEEEGGCELRVKFSRSADAPAYQQGQNQLYNELNLGRREEYDVLDKRRGRDPEMGGKPRRKNPQEGLYNELQKDKMAEAYSEIGMKGERRRGKGHDGLYQGLSTATKDTYDALHMQALPPRLEGGGEGRGSLLTCGDVEENPGPRMLLLVTSLLLCELPHPAFLLIPRKVCNGIGIGEFKDSLSINATNIKHFKNCTSISGDLHILPVAFRGDSFTHTPPLDPQELDILKTVKEITGFLLIQAWPENRTDLHAFENLEIIRGRTKQHGQFSLAVVSLNITSLGLRSLKEISDGDVIISGNKNLCYANTINWKKLFGTSGQKTKIISNRGENSCKATGQVCHALCSPEGCWGPEPRDCVSCRNVSRGRECVDKCNLLEGEPREFVENSECIQCHPECLPQAMNITCTGRGPDNCIQCAHYIDGPHCVKTCPAGVMGENNTLVWKYADAGHVCHLCHPNCTYGCTGPGLEGCPTNGPKIPSIATGMVGALLLLLVVALGIGLFM.
43. Клетка-хозяин для экспрессии химерного рецептора, содержащая нуклеиновую кислоту по любому из пп. 1-32, полипептидный химерный рецептор по п. 33 или 34 или вектор экспрессии по любому из пп. 35-42.
44. Клетка-хозяин по п. 43, где клетка-хозяин представляет собой цитотоксический Т-лимфоцит CD8+ или лимфоцит CD4+.
45. Фармацевтическая композиция для лечения, профилактики или облегчения рака у субъекта, содержащая эффективное количество клетки-хозяина по п. 43 или 44 и фармацевтически приемлемый наполнитель.
46. Фармацевтическая композиция по п. 45, где фармацевтическая композиция содержит множество клеток-хозяев, которые представляют собой Т-лимфоциты CD8+, необязательно выбранные из группы, состоящей из наивных Т-клеток CD8+, центральных Т-клеток памяти CD8+, эффекторных Т-клеток памяти CD8+ и общей популяции T-клеток CD8+; и/или фармацевтическая композиция содержит множество клеток-хозяев, которые представляют собой T-лимфоцит-хелперы CD4+, необязательно выбранные из группы, состоящей из наивных Т-клеток CD4+, центральных Т-клеток памяти CD4+, эффекторных Т-клеток памяти CD4+ и общей популяции T-клеток CD4+.
47. Фармацевтическая композиция по п. 46, где множество Т-лимфоцитов CD8+ и множество Т-лимфоцитов CD4+ присутствуют в композиции в соотношении 1:1.
48. Фармацевтическая композиция по любому из пп. 45-47, где через 72 ч после введения композиции в животной модели опухоли, экспрессирующей CD19, (a) Т-лимфоциты композиции, экспрессирующей химерный рецептор, проявляют пониженную степень индуцируемой активацией клеточной гибели (AICD), повышенное персистирование, и/или пониженный апоптоз в животной модели, и/или (b) наблюдается более значительное уменьшение объема опухоли, более значительное уменьшение количества опухолевых клеток и/или повышенная выживаемость животных по сравнению с последствиями введения композиции, содержащей Т-лимфоциты, экспрессирующие эталонный химерный рецептор, который отличается от кодированного химерного рецептора только полипептидным спейсером, где эталонный химерный рецептор содержит полипептидный спейсер длиной более 119 аминокислот.
49. Фармацевтическая композиция по любому из пп. 45-48, где по меньшей мере 30% Т-лимфоцитов в композиции, экспрессирующей химерный рецептор, могут пролиферировать на протяжении по меньшей мере двух поколений in vitro.
50. Фармацевтическая композиция по любому из пп. 45-49, где после введения композиции животным из животной модели, установленной для опухоли, экспрессирующей CD19,
у животных из животной модели наблюдается повышенное уничтожение клеток опухоли, повышенная регрессия опухоли, повышенная выживаемость животных и/или повышенная безопухолевая выживаемость животных по сравнению с контрольными животными модели, которым не вводили композицию; и/или
Т-лимфоциты композиции, экспрессирующей химерный рецептор, проявляют персистирование in vivo в крови животного из животной модели в течение 3, 10, 20, 30, 40 или 50 дней или более или пролиферацию в течение по меньшей мере 2 или по меньшей мере 3 циклов деления клеток.
51. Фармацевтическая композиция по любому из пп. 45-50, где рак представляет собой лимфому.
52. Фармацевтическая композиция по любому из пп. 45-51, где рак представляет собой лейкоз.
53. Фармацевтическая композиция по п. 52, где лейкоз представляет собой хронический лимфоцитарный лейкоз (CLL).
54. Способ проведения клеточной иммунотерапии у субъекта, имеющего рак, включающий введение субъекту эффективного количества клетки-хозяина по п. 43 или 44, или эффективного количества фармацевтической композиции по любому из пп. 45-53.
55. Способ получения клетки-хозяина для экспрессии химерного рецептора in vitro, включающий:
введение нуклеиновой кислоты по любому из пп. 1-32 или вектора экспрессии по любому из пп. 35-42 в клетки популяции лимфоцитов и культивирование клеток в присутствии анти-CD3 и/или анти-CD28, и по меньшей мере одного гомеостатического цитокина.
| KOCHENDERFER J.N | |||
| et al., Construction and Pre-clinical Evaluation of an Anti-CD19 Chimeric Antigen Receptor, J Immunother., 2009, vol.32, no.7, pp.689-702; | |||
| TILL G.B | |||
| et al., Adoptive immunotherapy for indolent non-Hodgkin lymphoma and mantle cell lymphoma using genetically modified autologous CD20-specific T cells, Blood, 2008, v.112, n.6, pp.2261-2271 | |||
| KOEHLER P | |||
| et al., Engineered T Cells for the Adoptive Therapy of B-Cell Chronic Lymphocytic Leukaemia, 2011, Advances in Hematology, v.2012, pp.1-13 | |||
| CHEADLE E.J | |||
| et al., Natural Expression of the CD19 Antigen Impacts the Long-Term Engraftment but Not Antitumor Activity of CD19-Specific Engineered T Cells, The Journal of Immunology, 2010, v.184, pp.1885-1896 | |||
| Топчак-трактор для канатной вспашки | 1923 |
|
SU2002A1 |
| Топливный насос для двигателей внутреннего горения | 1919 |
|
SU42760A1 |
| СПОСОБ НАПРАВЛЕНИЯ (ИНДУКЦИИ) ИММУННОГО ОТВЕТА, ДНК, ВЕКТОР, СПОСОБ ЛЕЧЕНИЯ | 1992 |
|
RU2161044C2 |

