ЗАЯВЛЕНИЕ О ГОСУДАРСТВЕННЫХ ИНТЕРЕСАХ
Данное изобретение было сделано при государственной поддержке грантом СА136551, присужденным Национальным институтом здравоохранения. Правительство имеет определенные права на изобретение.
ЗАЯВЛЕНИЕ ОТНОСИТЕЛЬНО ПЕРЕЧНЯ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Перечень последовательностей, связанных с этой заявкой, предоставляется в текстовом формате вместо бумажной копии и тем самым включается посредством ссылки в описание. Имя текстового файла, содержащего список последовательностей, - 360056_445WO_SEQUENCE_LISTING.txt. Размер текстового файла составляет 14,4 КБ, он был создан 16 февраля 2018 года и отправляется в электронном виде через EFS-Web.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Иммунная система человека обычно защищает организм от проникновения посторонних веществ и патогенов. В-лимфоциты, также называемые В-клетками и компонентом иммунной системы, продуцируют антитела, которые связываются с чужеродным веществом или патогеном и в некоторых случаях обеспечивают его уничтожение. Однако, в некоторых случаях иммунная система может выходить из-под контроля и приводить к заболеваниям, которые включают неконтролируемую пролиферацию В-клеток, таким как рак, аутоиммунное заболевание и воспалительное заболевание.
Зрелые В-клетки и их дифференцированное потомство могут быть идентифицированы молекулами на их клеточной поверхности, такими как B-клеточный антиген созревания (ВСМА, также известный как член 17 надсемейство рецепторов факторов некроза опухоли (TNFRSF17), TNFRSF13A и CD269), который экспрессируется на плазматические клетки и некоторые зрелые В-клетки. Было показано, что ВСМА специфично связывается с фактором активации В-клеток B-клеток (BAFF, также известным как TNFSF13B, TALL-1 и CD257) и лигандом, индуцирующим пролиферацию (APRIL, также известным как TNFSF13, TALL-2 и CD256), который может привести к активации NF-κВ. ВСМА-таргетированная терапия, включая адаптивный перенос модифицированных ВСМА-специфичных химерных антигенных рецепторов (CAR) Т-клеток, специфичных антител к оголенному ВСМА или введение ВСМА-специфичных антител, конъюгированных с терапевтическим фрагментом (конъюгат антитело-лекарственное средство, ADC, такой как радиоактивная метка) может использоваться для лечения некоторых типов B-клеточных злокачественных новообразований, таких как множественная миелома, но их эффективность может быть ограничена количеством молекул ВСМА, экспрессируемых на поверхности опухолевых клеток, и/или наличием растворимого ВСМА в кровотоке. Низкая поверхностная экспрессия ВСМА на раковых клетках или растворимые ВСМА могут ограничивать и предотвращать эффективность терапевтических агентов из-за неадекватного связывания с ВСМА, присутствующим на поверхности опухолевых клеток. Было показано, что низкие уровни других молекул-мишеней в опухолевых клетках (например, CD19, CD20), на которые таргетированы антитела, конъюгаты лекарственного средства или Т-клетки химерного антигенного рецептора ограничивают эффективность этих методов лечения и позволяют опухолевым клеткам, которые экспрессируют низкие уровни молекулы-мишени, чтобы избежать ликвидации. В случае ВСМА короткая внеклеточная часть молекулы отщепляется от поверхности клетки и выделяется под действием гамма-секретазы (γ-секретазы), клеточного фермента, локализованного в мембране, участвующего в расщеплении белка. Это расщепление снижает плотность ВСМА на клетках, таких как клетки опухоли миеломы, которые экспрессируют молекулу, и приводит к повышенным уровням растворимого ВСМА (sBCMA) в сыворотке пациентов с определенными аутоиммунными заболеваниями (например, системной красной волчанкой) и раком (например, множественная миелома).
В настоящее время в области иммунотерапии сохраняется потребность в альтернативных или улучшенных композициях и способах для более эффективного лечения аутоиммунного заболевания и рака.
КРАТКОЕ СОДЕРЖАНИЕ
В определенных аспектах настоящее изобретение относится к лечению рака с помощью ВСМА-специфичного CAR Т-клетки и ингибитора γ-секретазы.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
Фиг. 1А-1Р показывают конструкцию и функциональное тестирование типичных молекул химерного антигенного рецептора (CAR) данного изобретения.
(A) Иллюстрации типичных CAR, имеющих внеклеточный компонент, состоящий из ВСМА-специфичного scFv, полученного из антитела A7D12.2 («А7») или антитела C11D5.3 («С11»), и спейсерной области (состоящей из шарнирной области IgG4), гидрофобная часть (состоящая из трансмембранного домена CD28) и внутриклеточный компонент, состоящий из эффекторного домена CD3ζ и ко стимулирующего домена 4-1ВВ. ScFv были сконструированы с C-терминальным концом VH-области, связанной (линкер вариабельной области «G4S» (SEQ ID NO: 30)) с N-терминальным концом VL-области (ориентация «HL») или с C-терминальным концом области VL, связанной с N-терминальным концом области VH (ориентация «LH»).
(B) Данные проточной цитометрии (окрашивание карбоксифлуоресцеин сукцинимидиловым эфиром (CFSE)), показывающие пролиферацию Т-клеток человека, экспрессирующих CAR, показанные на Фиг. 1А, и контрольные Т-клетки, не содержащие CAR, в ответ на ВСМА-экспрессирующие опухолевые клетки.
(C) Продукция цитокинов (IFN-γ) трансдуцированными CAR Т-клетками, показанными на Фиг. 1В, при культивировании in vitro с указанными линиями опухолевых клеток ВСМА- (K562) или ВСМА+ (K562 ВСМА, U266, MM1.S).
(D) Специфичный лизис указанных клеток-мишеней Т-клетками ВСМА CAR.
(E) Иллюстрация двух примерных CAR данного тзобетения, где спейсерная область (Spacer+) может включать в себя кассету с метками, модуль линкера, шарнир, спейсерную аминокислоту и любую их комбинацию. Если CAR содержит одну или несколько меток в области спейсера, он будет упоминаться как T-ChARM (меченая молекула химерного антигенного рецептора), как описано в данном документе. Верхний CAR содержит внеклеточный компонент (состоящий из специфичного для ВСМА scFv и спейсерной области, которая необязательно содержит другие элементы, такие как метка или линкер), гидрофобную часть (состоящую из трансмембранного домена CD28) и внутриклеточный компонент (состоящий из эффекторного домена CD3ζ и костимулирующего домена 4-1ВВ). Нижний CAR содержит внеклеточный компонент (состоящий из специфичного для ВСМА scFv и спейсерной области, которая необязательно содержит другие элементы, такие как метка или линкер), гидрофобную часть (состоящую из трансмембранного домена CD28) и внутриклеточный компонент (состоящий из эффекторного домена CD3ζ и костимулирующий домена CD28). Обе ВСМА-специфичные конструкции CAR/T-ChARM содержат генный маркер для трансдукции, включающий усеченный EGFR человека (EGFRt), который отделен от ВСМА-специфичных конструкций CAR/T-ChARM саморасщепляющимся пептидом вируса Thoseaasigna 2А (Т2А) последовательность. Также можно использовать другие известные саморасщепляющиеся пептиды, такие как свиной тешовирус-1 2А (Р2А), вирус ринита А лошади (ERAV) 2А (Е2А) и вирус ящура (FMDV) 2А (F2A).
(F) Типичные конструкции CAR/T-ChARM, имеющие разделительные области разной длины, sh представляет собой 12 аминокислотный короткий спейсер; 2ST представляет собой 48 аминокислотный спейсер с двумя кассетами Strep-меток; 3ST представляет собой 66 аминокислотный спейсер с тремя кассетами Strep-меток; 2ST Int представляет собой 157 аминокислотный спейсер промежуточной длины с двумя кассетами Strep-меток; и Lo, который представляет собой 228 аминокислотный спейсер длинный спейсер. Короткий и длинный спейсер может необязательно содержать кассету с метками, такую как Strep-метка.
(G) Дополнительные иллюстрации примерных конструкций CAR/T-ChARM с С11 или А7 HL scFv и имеющих разные спейсерные области, необязательно, включая метки STII.
(H) Способность человеческих Т-клеток, модифицированных ВСМА-специфичными CAR, распознавать ВСМА и пролиферировать, измеряется путем мечения Т-клеток карбоксифтороцеином (CFSE), культивирования CAR-T-клеток или контроля нетрансдуцированных меченных CFSE Т-клеток с трансдуцированными клетками K562 с полинуклеотидом, кодирующим полноразмерный ВСМА (K562/ВСМА), и с использованием проточной цитометрии для измерения разведения CFSE с каждым клеточным делением. Меченные CFSE Т-клетки, содержащие разные ВСМА-специфичные CAR с разной длиной спейсера, но не контролирующие нетрансдуцированные Т-клетки (UT), разведенные CFSE после совместного культивирования с клетками K562/ВСМА. Клетки CAR-T, содержащие 2 ST или более длинные спейсеры, пролиферировали лучше, чем клетки CAR-T, экспрессирующие короткий спейсер.
(I,J) Высвобождение цитокинов (IFN-γ, I; IL-2, J) анти-ВСМА CAR Т-клетками с длиной спейсера в ответ на ВСМА-экспрессирующие клетки миеломы U266 и 8266.
(K) Экспрессия EGFRt и STII на клеточной поверхности на CD8+ Т-клетках, трансдуцированных анти-ВСМА С11 T-ChARM или ВСМА-2 CAR конструкциями после выделения и размножения.
(L, М) Высвобождение цитокинов (IFN-γ, L; IL-2, М) CD4 Т-клетками человека (верхние панели) и CD8 (нижние панели), сконструированными для экспрессии T-ChARM С11 по настоящему изобретению, которые включают ко стимулирующий домен 41ВВ или костимулирующий домен CD28, или с помощью ранее описанного анти-ВСМА CAR («ВСМА-2»; см. Carpenter и др. Clin. Cancer Res. 19: 2048, 2013).
(N) Пролиферация человеческих Т-клеток, сконструированных для экспрессии описанных T-CHARM С11 или ВСМА-2 CAR, при совместном культивировании с указанными клетками, экспрессирующими ВСМА.
(О) Лизис ВСМА-негативных клеток K562 CD8 Т-клетками, сконструированными для экспрессии T-ChARM С11 по настоящему изобретению (кружок отображает костимулирующий домен 41ВВ, квадрат отображает костимулирующий домен CD28) или ВСМА-2 CAR (треугольник) в указанном Е: Т соотношении.
(Р) Лизис клеток K562, трансдуцированных для экспрессии ВСМА, с помощью сконструированных CD8 Т-клеток, указанных на Фиг. 1O, при различных соотношениях Е:Т (ось X).
Фиг. 2A-2Q показывают продуцирование ВСМА и PD-L1 клетками множественной миеломы (ММ) в культуре и влияние растворимого ВСМА, поверхностно-связанного ВСМА и поверхностно-связанного PD-L1 на способность анти-ВСМА ChARM-T Т-клеток распознавать опухолевые клетки и продуцировать IFN-γ.
(A) Клетки миеломы U266 промывали и высевали в культуральную среду на 1, 3, 5 и 24 часа. Супернатант среды собирали и анализировали на растворимую ВСМА с помощью ИФА. Данные показывают зависящее от времени увеличение уровней растворимого ВСМА (sBCMA) в супернатанте.
(B) Гистограммы, показывающие экспрессию ВСМА эталонными клетками ММ (RPMI 8226) или примерными первичными клетками ММ пациента, имеющими высокую (Pt. 1), промежуточную (Pt. 2) или низкую/отрицательную экспрессию (Pt. 3), измеренную с помощью ИФА (черный отображает антитело против ВСМА; серая линия отображает контроль изотипа).
(C) Диаграмма, показывающая процент пациентов с миеломой (n равно 19), имеющих высокую, промежуточную или низкую/отрицательную экспрессию ВСМА в клетках ММ.
(D) Продукция IFN-γ анти-ВСМА С11 T-ChARM+ CD8+ Т-клетками в ответ на стимуляцию пациентами ММ клетками с высокой (слева, n равно 5) или низкой (справа, n равно 4) экспрессией ВСМА (24 часа в соотношении 2:1 Е: Т (ИФА)). Значение было проверено с использованием непарного двустороннего Т-критерия, и столбцы представляют среднюю продукцию IFN-γ + SEM.
(E) Гистограммы, показывающие экспрессию PD-L1 клетками миеломы RPMI 8226 (крайний левый график) и первичными клетками ММ у 3 пациентов, имеющих высокую, низкую/отрицательную или промежуточную экспрессию PD-L1 по данным ИФА (диагональная штриховка отображает окрашивание с антителом против PD-L1; пустая гистограмма отображает контрольный изотип).
(F) Диаграмма, показывающая процент пациентов (n равно 19), имеющих высокую, промежуточную или низкую/отрицательную экспрессию PD-L1 в клетках ММ.
(G) Продукция IFN-γ анти-ВСМА C11 T-ChARM+ CD8+ клетками данного описания в ответ на клетки ММ пациента с высокой (слева, n равно 4) или низкой (справа, n равно 5) экспрессией PD-L1 (24 часа при соотношении 2:1 Е: Т (ИФА)). Значение было проверено с использованием непарного двустороннего Т-критерия, и столбцы представляют среднюю продукцию IFN-γ + SEM.
(H) Продукция IFN-γ ВСМА-специфичными T-ChARM Т-клетками в присутствии экзогенного растворимого ВСМА. sBCMA добавляли к ко-культурам Т-клеток, экспрессирующих ВСМА-специфичные клетки T-ChARM и K562, трансдуцированные полинуклеотидом, кодирующим полноразмерную ВСМА (K562/ВСМА+). Существует дозозависимое ингибирование эффекторной функции ВСМА-специфичных T-ChARM Т-клеток, измеренное по высвобождению IFN-γ в супернатант среды.
(I) Данные другого эксперимента по титрованию дозы, показывающие влияние sBCMA на продукцию IFN-γ ВСМА-специфичных T-ChARM Т-клетками, распознающими клетки ВСМА+ K562, которые включали добавление sBCMA в культуру в другой, более высокой концентрации (5000 нг/мл).
(J) Выделение ВСМА указанными клетками ММ, культивируемыми in vitro.
(K) sBCMA, измеренная в сыворотках костного мозга (ВМ) от пациентов с более низким (менее 2% CD138+ клетками) или более высоким (более 2% CD138+ клетками) заболеванием.
(L) sBCMA-связывание с C113ST-ChARM Т-клетками по настоящему изобретению. Т-клетки инкубировали с указанными уровнями рекомбинантного sBCMA (правая часть диаграммы) и затем окрашивали слитый BCMA-Fc, который конъюгировали с АРС.
(М) Данные проточной цитометрии, показывающие окрашивание поверхности (АРС-конъюгированное BCMA-Fc и анти-EGFRt антитело) и экспрессию CD4 клеток C113ST T-ChARM и FMC63 CAR-T.
(N) Данные проточной цитометрии, показывающие продукцию IFN-γ (ось Y) и экспрессию CD4 (ось X) Т-клетками, экспрессирующими или C113ST-ChARM («C113ST») по настоящему изобретению, или контрольный анти-CD19 CAR («FMC632») в совместной культуре с клетками-экспрессирующими мишенями K562 и введенным слитым белком BCMA-Fc (левые панели: 0 нг/мл BCMA-Fc; правые панели: 1000 нг/мл BCMA-Fc.
(О) Титрование дозы, показывающее высвобождение IFN-γ Т-клетками CAR в ответ на клеточные линии-мишени, как указано в присутствии или отсутствии экзогенного рекомбинантного ВСМА (слитый BCMA-Fc). FMC63 (анти-CD 19) в сравнении с K562 CD19+ (контроль; треугольник, обращенный вниз); C113ST T-ChARM Т клетки против RPMI 8226 (обращенный вверх треугольник), U266 (квадрат) и K562 ВСМА+ клетки (круг). Данные представляют 2 независимых эксперимента. Столбцы представляют среднее значение +SEM. Значение Р составляет 0,05 или менее, как определено односторонним ANOVA с пост-тестом. MFI означает среднюю интенсивность флуоресценции.
(Р) Продукция IFN-γ (нормализованная средняя интенсивность флуоресценции, MFI) Т-клетками, показанными на фигуре 2O, в совместной культуре с указанными антиген-экспрессирующими клетками и в присутствии или отсутствии BCMA-Fc (ось х).
(Q) Цитолитическая активность CD8+ C113ST T-ChARM Т-клеток против клеток-мишеней ВС56+ K562 и FMC63-CAR-T-клеток против клеток-мишеней CD56+ K562 при различных концентрациях рекомбинантного ВСМА, проанализированных с помощью 4-часового CRA при соотношении Е:Т 10:1. Данные представляют 2 независимых эксперимента. Столбцы представляют среднее значение +SEM. Значение Р составляет 0,05 или менее, как определено одним способом ANOVA с пост-тестом. MFI означает среднюю интенсивность флуоресценции.
Фиг. 3A-3R показывают влияние ингибитора γ-секретазы (GSI) RO4929097 на уровни ВСМА на клеточной поверхности и других молекул клеточной поверхности на клеточных линиях миеломы или клетках первичной миеломы.
(А) ВСМА клеточной поверхности измеряли на четырех клеточных линиях миеломы (8226, U266B1, MM1.R, Н929) методом проточной цитометрии с моноклональным анти-ВСМА антителом до и через 5 часов после (см. диаграммы) инкубации клеточной линии миеломы с GSI RO4929097 в диапазоне концентраций от 0 мкМ (контроль ДМСО) до 1,0 мкМ.
(B) поверхностная экспрессия ВСМА клетками MM.1R, культивируемыми с указанными концентрациями RO4929097; окрашивание антителом против ВСМА (черные линии) по сравнению с контрольным изотипом (серая линия).
(C) кратное изменение поверхностной экспрессии ВСМА клеточными линиями ММ при культивировании с указанными концентрациями RO4929097; кратное изменение указано относительно необработанных клеток ММ той же линии.
(D) Кинетика кратного изменения поверхностной экспрессии ВСМА с течением времени указанными клетками ММ в культуре с 1 мкМ RO4929097.
(E) Клетки миеломы U266 инкубировали в течение 1, 3, 5 и 24 часов в присутствии различных концентраций GSI RO429097 (0,01 мкМ, 0,1 мкМ и 1,0 мкМ) и оценивали экспрессию поверхностного ВСМА методом проточной цитометрии. Экспрессия ВСМА возрастала дозозависимым образом в присутствии GSI с увеличением пика, наблюдаемым после 5 часов воздействия.
(F) Изменение кратности поверхностной экспрессии ВСМА на клеточных линиях (MM1R отображается треугольником; U266 отображается квадратом; 8226 отображается кругом), культивируемых в 1 мкМ GSI с течением времени. GSI повторно вводили как половину смены среды каждые 2 дня. Изменение сгиба в ВСМА определяется как Обработанный (MFIBCMA-MFIiso)/Контроль (MFIBCMA-MFIiso). Данные представляют по меньшей мере 2 независимых эксперимента.
(G, Н) Концентрация супернатанта культуры sBCMA в клетках линии клеток ММ, культивированных в присутствии указанных концентраций RO4929097.
(I) Клетки миеломы U266 промывали и высевали в культуральные среды в присутствии различных концентраций GSI RO429097 (0,01 мкМ, 0,1 мкМ и 1,0 мкМ). Супернатант среды собирали через 1, 3, 5 и 24 часа и анализировали на растворимую ВСМА (sBCMA) с помощью ИФА. Данные показывают, что количество ВСМА, высвобождаемого из опухолевых клеток в супернатант с течением времени, уменьшалось, когда GSI присутствовал в концентрации, по меньшей мере, приблизительно 0,01 мкМ.
(J, K) Изменение кратности экспрессии ВСМА (J) и концентрации супернатанта sBCMA (K) в различные моменты времени после удаления 1 мкМ GSI из культур линии миеломных клеток (GSI +/-) по сравнению с культурами с постоянным присутствием GSI (GSI +/+).
(L) Жизнеспособность указанных ММ-экспрессирующих клеток, измеренная путем окрашивания йодидом пропидия клеточных линий, культивируемых в 1 мкМ GSI.
(М) Поверхностная экспрессия ВСМА первичными клетками ММ пациента, культивируемыми с указанными концентрациями R04929097. Окрашивание было таким, как описано в отношении Фиг. 3В.
(N) Изменение кратности ВСМА на клетках первичной миеломы (n равно 7), культивируемых с различными количествами GSI в течение 4 часов. Первичные и клеточные линии культивировали при 0,5×106 клеток/мл. Изменение кратности в ВСМА определяется как Обработанный (MFIBCMA-MFIiso)/Контроль (MFIBCMA-MFIiso). Данные представляют 3 независимых эксперимента с Т-клетками, полученными от разных доноров.
(О, Р) Совместное культивирование клеток первичной миеломы с различными концентрациями GSI в течение 4 часов не влияет на уровни некоторых других молекул клеточной поверхности в опухолевых клетках, включая CS1, CD86, PD-L1, CD80 и CD38.
(Q) Окрашивание различных поверхностных маркеров на клетках MM1R в присутствии (черный) или отсутствии (серый) 1 мкМ GSI в культуральной среде. Окрашивание изотипа показано в виде открытого участка.
(R) CD 138+ клетки первичной миеломы обогащали из образцов костного мозга пациента, инкубировали в течение 3 часов в присутствии различных концентраций GSI RO429097 (от 0,01 мкМ до 10 мкМ) и оценивали на предмет экспрессии ВСМА на поверхности методом проточной цитометрии. Средняя интенсивность флуоресценции ВСМА (MFI) на опухолевых клетках представлена в виде увеличения кратности по сравнению с опухолевой клеткой, инкубированной без RO429097. Наблюдается установленное дозозависимое повышение экспрессии ВСМА.
Фиг. 4А-4С показывают, что высвобождение цитокинов после распознавания клеток первичной миеломы ВСМА-специфичными CAR-T клетками увеличивается, когда клетки миеломы предварительно обрабатывают GSI.
(A) Продукция IL-2 клетками ВСМА CAR-T (ВСМА-специфичные T-ChARM С11 3ST-CD28 и ВСМА-специфичные T-ChARM С11 3ST-41BB) или контрольные CD19sh CAR (короткий спейсер) Т-клетки, совместно культивированные с первичными клетками опухоли миеломы человека в течение 24 часов отдельно или с различными концентрациями GSI RO429097 (от 0,003 мкМ до 3,0 мкМ).
(B) Продукция IFN-γ ВМСА T-ChARM Т-клетками, совместно культивированными с клетками миеломы в различных концентрациях RO429097.
(C) Пролиферация меченных CFSE ВСМА-специфичных T-ChARM Т-клеток увеличивалась дозозависимым образом после совместного культивирования в течение 3 дней с первичными опухолевыми клетками миеломы человека в среде отдельно или в среде, содержащей GSI RO492097, в указанных концентрациях.
Фиг. 5A-5R показывают влияние различных концентраций GSI на жизнеспособность, рост и функциональную активность клеток CAR-T.
(A) CD 19 окрашивание K562 CD19 + и клеток Raji, которые культивировали с GSI или без GSI в течение ± 12-16 часов. Контрольный изотип показан серой линией.
(B) Первичные человеческие Т-клетки культивировали в GSI RO4929097 в концентрациях от 0,01 до 100 мкМ, и жизнеспособность измеряли путем исключения красителя трипанового синего через 24 часа. При любой концентрации GSI не оказывал влияния на жизнеспособность Т-клеток.
(C) CD19 CAR-T клетки совместно культивировали с клетками-мишенями K562/CD19 в среде, содержащей различные концентрации RO4929097. RO4929097 ингибирует эффекторную функцию CD19 CAR-T-клеток при концентрациях более 3 мкМ при совместном культивировании, что определяется путем измерения продукции IL-2 (верхний график) и IFNγ (нижний график). Во вставке показано соответствующее терапевтическое окно лекарственного средства, которое не ингибирует эффекторную функцию клеток CAR-T.
(D) Продукция IL-2 клетками CD19 CAR-T, культивируемыми с клетками-мишенями в присутствии возрастающих концентраций GSI RO4929097.
(E) Продукция IL-2 CAR-T клетками CD19, культивируемыми с клетками-мишенями («K562 CD19») или контрольными клетками («K562 ВСМА») в присутствии возрастающих концентраций GSI RO4929097. Клеткам вводили указанные количества GSI и затем промывали (пустые столбики) или нет (заполненные столбики).
(F) Данные другого эксперимента, показывающие продукцию IL-2 CAR-T клетками CD19 после совместного культивирования в течение ночи с клетками K562 ВСМА+ или K562 CD19+ после предварительной инкубации с различными концентрациями GSI. После промывания GSI либо добавляли обратно (+/+), либо исключали из совместного культивирования (+/-) для оценки обратимости продукции цитокинов.
(G) Специфичный лизис CAR-T клетками CD19 указанных клеток-мишеней или контрольных клеток в присутствии GSI RO4929097.
(H) Пролиферация клеток CD19 CAR-T, культивируемых с CD19-экспрессирующими клетками-мишенями в присутствии или отсутствии GSI или с контрольными клетками в отсутствие GSI. Клетки окрашивали CFSE и пролиферацию измеряли проточной цитометрией.
(I) Графическое представление количества клеточных делений CAR-T клеток CD19 в присутствии указанных концентраций GSI RO4929097. Ширина горизонтальных полос представляет собой долю CAR-T клеток в культуре, которые разделили указанное число поколений (то есть 5, 4, 3, 2, 1 или 0 поколений) в течение эксперимента.
(J) Подсчет клеток (окрашивание CD8) во время размножения СБ19-специфичных CAR-T клеток с LCL CD19+ ТМ клетками и экзогенным IL-2 в отсутствие GSI (кружок) или в присутствии GSI при 0,5 мкМ (квадратный) или 5 мкМ (треугольник).
(K) Количество клеток CD19-специфичных CAR-T клеток (CD4:CD8 (1:1)), размноженных с клетками CD19+ ТМ. LCL, в отсутствие или в присутствии GSI, как указано, но без добавления экзогенного IL-2.
(L) Концентрации IFN-γ в супернатантах GSI-размноженных анти-CD19 CAR CD8+ Т-клеток после повторной стимуляции K562 клетками (без антигена), K562 CD19+ клетками или клетками Raji.
(М, N) Продукция IFN-γ (М) и IL-2 (N) смесью CD4:CD8 анти-CD19 CAR-T клеток после повторной стимуляции указанными клеточными линиями в отсутствие GSI или в присутствии либо 0,5 мкМ, либо 5 мкМ GSI.
(О) Внутриклеточное окрашивание, показывающее продукцию IFN-γ (ось Y) и экспрессию CD8 (ось X) T-ChARM Т-клетками настоящего изобретения, культивированными с первичными клетками ММ от 2 пациентов в отсутствие (0 мкМ; левые панели) или наличие (1 мкМ; правые рисунки) GSI RO4929097.
(Р) Продукция IFN-γ (средняя геометрическая интенсивность флуоресценции (gMFI)) T-ChARM Т-клетками согласно настоящему изобретению, культивируемыми с первичными клетками ММ, в присутствии указанной концентрации RO4929097 (ось х).
(Q) Продукция IFN-γ (нормализованная MFI; ось у) T-ChARM Т-клетками согласно настоящему изобретению, культивируемыми с первичными клетками ММ, в присутствии указанной концентрации RO4929097 (ось х).
(R) Пролиферация T-ChARM Т-клеток согласно настоящему изобретению в присутствии первичных клеток ММ, которые не были обработаны (серая штриховка) или обработаны 1,0 мкМ GSI RO4929097.
Фиг. 6А-6С показывают влияние GSI RO4929097 на экспрессию ВСМА в доклинической модели множественной миеломы in vivo.
(А) Программа эксперимента для модели мышиной миеломы с распространенным ксенотрансплантатом. Мышей NSG облучали (275 рад) для облегчения приживления опухоли и получали опухолевые клетки ММ человека (5×106 MM.1R) с последующей обработкой GSI (30 мг/кг). Впоследствии мышей подвергали эвтаназии и отбирали образцы крови и ВМ, чтобы определить, повышала ли GSI экспрессию ВСМА на клетках миеломы in vivo.
(B) Изменение кратности поверхностной экспрессии ВСМА на клетках миеломы у мышей, умерщвленных в указанные моменты времени после второго введения GSI.
(C) Уровни sBCMA в сыворотке от мыши, умерщвляемой в указанные моменты времени после введения RO4929097.
Фиг. 7А-7Е показывают влияние GSI RO4929097 на анти-ВСМА CAR Т-клеточную терапию в доклинической мышиной модели ММ.
(A) Программа эксперимента, в которой мыши получали радиацию, за которой следовали опухолевые клетки ММ человека (5×106 MM.1R, экспрессирующих люциферазу светлячка). Двадцать дней спустя мышам вводили GSI (30 мг/кг) в указанные моменты времени и одну субоптимальную дозу T-ChARM Т-клеток против ВСМА (0,33×106 клеток, CD4: CD8 1: 1) в день 0. Визуализация биолюминесценции (BLI) и выживание контролировались на всем протяжении.
(B) Изображения BLI мышей, полученные на 2, 17 и 16 дни после обработки Т-ChARM Т-клетками C113ST (0 мг/кг 0,33×10 6 клеток, CD4:CD8 1:1, левые рисунки; 30 мг/кг, рисунки посередине) или контролируют FM63 анти-CD19 CAR Т-клетки (0,33×106 клеток, 1:1 CD4: CD8, 30 мг/кг, правые рисунки).
(C) Количественные данные люминесценции от BLI, показанные на Фиг. 8В.
(D) Процент выживаемости мышей, показанных на Фиг. 8В, после введения Т-ChARM Т-клеток.
(E) (слева) Количественные данные люминесценции от BLI, показанные на Фиг. 8В; (справа) процент выживания мышей после введения T-ChARM Т-клеток.
Фиг. 8 показывает проточный цитометрический анализ связывания биспецифичной слитой молекулы со специфичностью к ВСМА клеткам Н929 ММ в присутствии или отсутствии GSI. Контрольная биспецифичная слитая молекула, не таргетированная на ВСМА, также была протестирована.
ПОДРОБНОЕ ОПИСАНИЕ
Настоящее изобретение предоставляет композиции и способы лечения аутоиммунного заболевания и рака посредством комбинированного использования В-клеточного антигена созревания (ВСМА)-специфичного связывающего белка, в растворимой форме или экспрессируемого на цитотоксической или другой клетке, и ингибитора γ-секретазы (GSI). Полинуклеотиды, кодирующие такие ВСМА-специфичные связывающие белки, можно использовать для создания модифицированных иммунных клеток-хозяев (например, Т-клеток) для применения, например, в адаптивной иммунотерапии. В определенных аспектах настоящее описание относится к применению такой терапии у субъекта, нуждающегося в лечении, в сочетании с GSI, причем последнее лечение может быть назначено до, одновременно или после адаптивной иммунотерапии (то есть иммунотерапии с модифицированной иммунной клеткой, экспрессирующей ВСМА-специфичный связывающий белок на поверхности клетки). В настоящем документе также предлагаются способы иммунотерапии, включающие введение GSI в комбинации с ВСМА-специфичным связывающим белком, который содержит антитело или его антигенсвязывающую часть, которые могут в некоторых вариантах осуществления быть конъюгированы или иным образом связаны с цитотоксическим лекарственным средством, например, с образованием конъюгата антитело-лекарственное средство (ADC). Полезные терапевтические применения включают лечение пролиферативного заболевания или расстройства (такого как рак), аутоиммунного заболевания или связанного со старением заболевания или расстройства у субъекта, у которого ВСМА-позитивные клетки являются патогенными.
Настоящее изобретение также относится к набору для лечения гематологического заболевания или расстройства, связанного с экспрессией ВСМА, включающему: а) единицу дозирования Т-клетки, экспрессирующей ВСМА-специфичный химерный антигенный рецептор (CAR), и b) единицу дозирования ингибитора γ-секретазы. В определенных аспектах набор дополнительно содержит CD20-специфичный связывающий белок, такой как ритуксимаб, офатумумаб, окрелизумаб; CD19-специфичный связывающий белок; CD45-специфичный связывающий белок; CD38-специфичный связывающий белок; цитокин; хемокин; фактор роста; химиотерапевтический агент или радиотерапевтический агент.
Перед изложением настоящего изобретения более подробно, может быть полезно понять его, чтобы дать определения определенных терминов, которые будут использоваться в данном документе. Дополнительные определения изложены в данном описании.
В настоящем описании любой диапазон концентрации, процентный диапазон, диапазон отношения или целочисленный диапазон следует понимать как включающий значение любого целого числа в указанном диапазоне и, при необходимости, его доли (например, одной десятой и одной сотой целого числа), если не указано иное. Кроме того, любой диапазон чисел, приведенный здесь в отношении любого физического признака, такого как полимерные субъединицы, размер или толщина, следует понимать как включающий любое целое число в указанном диапазоне, если не указано иное. Используемый здесь термин «около» означает ±20% указанного диапазона, значения или структуры, если не указано иное. Следует понимать, что термины, используемые в единственном числе в данном документе, относятся к «одному или нескольким» из перечисленных компонентов. Использование альтернативы (например, «или») следует понимать как означающее либо одну, обе, либо любую их комбинацию альтернатив. Используемые здесь термины «включать», «иметь» и «содержать» используются как синонимы, и их термины и варианты предназначены для использования в качестве неограничивающих.
Кроме того, следует понимать, что отдельные соединения или группы соединений, полученные из различных комбинаций структур и заместителей, описанных в данном документе, описаны в настоящей заявке в той же степени, как если бы каждое соединение или группа соединений были выдвинуты индивидуально. Таким образом, выбор конкретных структур или конкретных заместителей входит в объем настоящего описания.
Термин «состоящий по существу из» ограничивает объем формулы изобретения указанными материалами или стадиями или такими, которые не оказывают существенного влияния на основные характеристики заявленного изобретения. Например, белковый домен, область, модуль или кассета (например, связывающий домен, шарнирная область, линкерный модуль, метка-кассета) или белок (который может иметь один или несколько доменов, областей, модулей или кассет) «по существу состоит» из конкретной аминокислотной последовательности, когда аминокислотная последовательность домена, области, модуля, кассеты или белка включает удлинения, делеции, мутации или их комбинацию (например, аминокислоты на амино- или карбокси-конце или между доменами) которые в комбинации составляют не более 20% (например, не более 15%, 10%, 8%, 6%, 5%, 4%, 3%, 2% или 1%) длины домена, области, модули, кассеты или белка и не оказывают существенного влияния (т.е. не снижают активность более чем на 50%, например, не более чем на 40%, 30%, 25%, 20%, 15%, 10%, 5% или 1%) активность домена (ов), области (ей), модуля (ей), кассеты (кассет) или белка (например, аффинность связывания с мишенью связывающего белка или метки-кассеты).
Используемый здесь термин «пролиферативное расстройство» относится к чрезмерному или иным образом аномальному росту или пролиферации по сравнению с нормальной или небольной клеткой. Чрезмерный или аномальный рост включает, например, нерегулируемый рост или пролиферацию, которые могут происходить быстро (например, гиперпролиферация) или могут происходить медленнее или прогрессивнее с течением времени (например, множественная миелома) в нативной ткани (например, рост плазмоцитомы в костном мозге), а также распространяются в/растут в пределах дистальной ткани или участка тела, который не является нативным для больной клетки. Типичные пролиферативные расстройства включают опухоли, рак, опухолевую ткань, карциному, саркому, злокачественные клетки, предзлокачественные клетки, а также неопухолевые или незлокачественные пролиферативные расстройства (например, аденома, фиброма, липома, лейомиома, гемангиома, фиброз, рестеноз) или аутоиммунные заболевания (такие как ревматоидный артрит, остеоартрит, псориаз, воспалительное заболевание кишечника или тому подобное).
Термин «связывающий белок» (также называемый «связывающим доменом», «связывающей областью» или «связывающим фрагментом») в контексте настоящего описания относится к молекуле, такой как пептид, олигопептид, полипептид или белок, который обладает способность специфично и нековалентно связываться, объединяться или комбинироваться с молекулой-мишенью (например, ВСМА). Связывающий белок включает любого встречающегося в природе, синтетического, полусинтетического или рекомбинантно полученного партнера по связыванию для биологической молекулы, соединения или другой интересующей мишени. В некоторых вариантах осуществления связывающий белок происходит из иммуноглобулина или иммуноглобулиноподобной молекулы, такой как антитело или рецептор Т-клеток (TCR), которая включает функциональный связывающий домен или его антигенсвязывающий фрагмент. Типичные связывающие белки включают вариабельные области одноцепочечных антител (например, доменные антитела, sFv, scFv, Fab), лиганды ВСМА (например, BAFF, APRIL и их связывающие фрагменты), антигенсвязывающие области рецепторов Т-клеток (TCR), такие как одноцепочечные TCR (scTCR) или синтетические полипептиды, выбранные для специфичной способности связываться с биологической молекулой.
Используемый в данном документе термин «специфично связывает» относится к ассоциации или объединению связывающего домена или его слитого белка с молекулой-мишенью с аффиностью или Ka (то есть константой равновесной ассоциации конкретного взаимодействия связывания с единицами 1/М) составляет 105 М-1 или более, при незначительном связывании или объединении с любыми другими молекулами или компонентами в образце. Связывающие домены (или их слитые белки) могут быть классифицированы как связывающие домены с высокой аффинностью (или их слитые белки) или связывающие домены с низкой аффинностью (или их слитые белки). «Связывающие домены с высокой аффинностью» относятся к тем связывающим доменам, у которых Ka составляет не менее 107 М-1, не менее 108 М-1, не менее 109 М-1, не менее 1010 М-1, не менее 1011 М-1, по меньшей мере 1012 М-1 или по меньшей мере 1013 М-1. «Связывающие домены с низкой аффинностью» относятся к таким связывающим доменам с Ka до 107 М-1, до 106 М-1, до 105 М-1. Альтернативно, аффинность может быть определена как константа равновесной диссоциации (Kd) конкретного связывающего взаимодействия с единицами М (например, от 10-5 М до 10-13 М). В некоторых вариантах осуществления домен связывания может иметь «повышенную аффинность», которая относится к выбранному или сконструированному домену связывания с более сильным связыванием с антигеном-мишенью, чем домен связывания дикого типа (или родительский). Например, повышенная аффинность может быть обусловлена тем, что Ka (константа равновесной ассоциации) для целевого антигена выше, чем для связывающего домена дикого типа, или тем, что Kd (константа диссоциации) для целевого антигена меньше, чем для связывающего домена дикого типа, или тем, что скорость диссоциации (Koff) для целевого антигена меньше, чем для связывающего домена дикого типа. Известны множество анализов для идентификации связывающих доменов согласно настоящему изобретению, которые специфично связывают конкретную мишень, а также для определения сродства связывающего домена или слитого белка, таких как вестерн-блот, ИФА и анализ Biacore® (см. также, например, Scatchard и др., Ann. NY Acad. Sci. 51: 660, 1949; и патенты США №5283173, 5468614 или эквивалентные).
Используемый в данном документе термин «гетерологичный», «неэндогенный» или «экзогенный» относится к любому гену, белку, соединению, молекуле или активности, которая не является нативной для клетки-хозяина или субъекта, или представляет собой любой ген, белок, соединение, молекула или активность являются нативными для клетки-хозяина, но были изменены или видоизменены так, что структура, активность или и то, и другое отличается от нативной и измененной молекул. В некоторых вариантах осуществления гетерологичные, неэндогенные или экзогенные молекулы (например, рецепторы, лиганды) могут не быть эндогенными для клетки-хозяина или субъекта, но вместо этого нуклеиновые кислоты, кодирующие такие молекулы, могут быть добавлены в клетку-хозяина путем конъюгации, трансформации, трансфекции, электропорации или тому подобное, где добавленная молекула нуклеиновой кислоты может интегрироваться в геном клетки-хозяина или может существовать в виде внехромосомного генетического материала (например, в виде плазмиды или другого самореплицирующегося вектора). Термин «гомологичный» или «гомолог» относится к молекуле или активности, обнаруженной или полученной из клетки-хозяина, вида или штамма. Например, гетерологичная или экзогенная молекула или ген, кодирующий молекулу, могут быть гомологичными нативной молекуле субъекта или клетки-хозяина или гену, который кодирует молекулу, соответственно, но могут иметь измененную структуру, последовательность, уровень экспрессии или их комбинации. Неэндогенная молекула может быть одного и того же вида, разных видов или их комбинации.
Используемый в данном документе термин «эндогенный» или «нативный» относится к гену, белку, соединению, молекуле или активности, которые обычно присутствуют в клетке-хозяине или хозяине.
Используемый в данном документе термин «кассета-метка» относится к уникальной пептидной последовательности, присоединенной, слитой или являющейся частью представляющего интерес белка, с которой гетерологичная или неэндогенная когнатно-связывающая молекула (например, рецептор, лиганд, антитело, или другого партнера по связыванию) способен специфично связываться, где свойство связывания можно использовать для обнаружения, идентификации, выделения или очистки, отслеживания, обогащения или таргетирования на меченый белок или клетки, экспрессирующие меченый белок, особенно когда меченый белок является частью гетерогенной популяции белков или другого материала, или когда клетки, экспрессирующие меченый белок, являются частью гетерогенной популяции клеток (например, биологического образца, такого как периферическая кровь). В некоторых вариантах осуществления клетка, экспрессирующая меченый белок, может приводиться в контакт с гетерологичной или неэндогенной когнат-связывающей молекулой и индуцировать биологический ответ, такой как стимуляция активации клеток, пролиферация клеток или гибель клеток. В предоставленных слитых белках способность метки-кассеты (кассет) специфично связываться с когнат-связывающей молекулой(ами) отличается от или в дополнение к способности связывающего домена(ов) специфично связываться с молекулой-мишенью(ями). Метка-кассета обычно не является антигенсвязывающей молекулой, например, не является антителом или TCR или его антигенсвязывающей частью.
Используемый в данном документе термин «шарнирная область» или «шарнир» относится к: а) шарнирной последовательности иммуноглобулина (состоящей, например, из верхней и внутренней областей) или ее функциональному фрагменту или варианту, b) типу II междоменной (стеблевой) области C-лектина или ее функциональному фрагменту или варианту, или с) области стебля молекулы кластера дифференцировки (CD) или его функциональному варианту. Используемый в данном документе термин «шарнирная область иммуноглобулина дикого типа» относится к встречающимся в природе аминокислотным последовательностям верхнего и среднего шарниров, вставленным между и соединяющими домены СН1 и СН2 (для IgG, IgA и IgD) или расположенным между и соединяющими домены СН1 и СН3 (для IgE и IgM) обнаружены в тяжелой цепи антитела. В определенных вариантах осуществления шарнирная область является человеческой, и в конкретных вариантах осуществления содержит шарнирную область IgG человека.
Используемый в данном документе термин «спейсерная область» относится к одному или нескольким белкам, полипептидам, олигопептидам, пептидам, доменам, областям, модулям, кассетам, мотивам или любой их комбинации, которые объединяют два или более белков, полипептидов, олигопептидов, пептидов, доменов, областей, модулей, кассет, мотивов или любые их комбинации в слитом белке. Например, спейсерная область может обеспечивать функцию разделения или разнесения для облегчения взаимодействия двух одноцепочечных слитых белков или позиционирования одного или нескольких связывающих доменов, так что получающаяся в результате полипептидная структура сохраняет специфичную аффинность связывания с молекулой-мишенью или поддерживает сигнальную активность (например, активность эффекторного домена) или то и другое. В некоторых вариантах осуществления спейсерная область может содержать «линкерный модуль», который представляет собой аминокислотную последовательность, имеющую от приблизительно двух до приблизительно 500 аминокислот, которая может обеспечить гибкость и пространство для конформационного перемещения между двумя областями, доменами, мотивами, кассетами или модулями, связанными с помощью линкера. Типичные линкерные модули включают в себя модули, имеющие от одного до десяти повторов GlyxSery (SEQ ID NO: 31), где х и у независимо представляют собой целые числа от 0 до 10, при условии, что х и у не равны 0 (например, (Gly4Ser)2 (SEQ ID NO: 32), (Gly3Ser)2 (SEQ ID NO: 33), Gly2Ser или их комбинация, такая как (Gly3Ser)2Gly2Ser)) (SEQ ID NO: 34). В некоторых других вариантах осуществления спейсерная область может иметь линкерный модуль, который содержит одну или несколько константных областей тяжелой цепи иммуноглобулина, таких как только СН3 или СН2СН3. В дополнительных вариантах осуществления спейсерная область может содержать шарнирную область или метку-кассету. Каждый такой компонент соединителя не является взаимоисключающим. Например, спейсерная область может содержать шарнир и один или несколько линкерных модулей, или спейсерная область может содержать шарнир, один или несколько линкерных модулей и одну или более меток-кассет. Типичные спейсерные области могут варьироваться по длине, например, от приблизительно 5 до приблизительно 500 аминокислот, или от приблизительно 10 до приблизительно 350 аминокислот, или от приблизительно 15 до приблизительно 100 аминокислот, или от приблизительно 20 до приблизительно 75 аминокислот, или от приблизительно 25 до приблизительно 35 аминокислот. Типичные короткие спейсеры варьируются от приблизительно 5 до приблизительно 100 аминокислот (например, 12 аминокислот, 15 аминокислот, 48 аминокислот, 50 аминокислот, 66 аминокислот, 70 аминокислот), промежуточные спейсеры варьируются от приблизительно 100 до приблизительно 200 аминокислот кислоты (например, НО аминокислот, 120 аминокислот, 130 аминокислот, 140 аминокислот, 150 аминокислот, 157 аминокислот, 175 аминокислот) и длинные спейсеры варьируются от приблизительно 200 до приблизительно 500 аминокислот (например, 200 аминокислот кислоты, 210 аминокислот, 220 аминокислот, 228 аминокислот, 230 аминокислот, 250 аминокислот, 300 аминокислот, 350 аминокислот, 400 аминокислот, 450 аминокислот).
«Гидрофобная часть», как используется в данном документе, означает любую аминокислотную последовательность, имеющую трехмерную структуру, которая является термодинамически стабильной в клеточной мембране и обычно имеет длину от около 15 аминокислот до около 30 аминокислот. Структура гидрофобного домена может содержать альфа-спираль, бета-бочку, бета-лист, бета-спираль или любую их комбинацию.
Используемый в данном документе «эффекторный домен» представляет собой внутриклеточную часть слитого белка или рецептора, которая может прямо или косвенно стимулировать биологический или физиологический ответ в клетке при получении соответствующего сигнала. В некоторых вариантах осуществления эффекторный домен является частью белка или белкового комплекса, который получает сигнал при связывании, или он связывается непосредственно с молекулой-мишенью, которая запускает сигнал от эффекторного домена. Эффекторный домен может непосредственно стимулировать клеточный ответ, когда он содержит один или несколько сигнальных доменов или мотивов, таких как иммунорецепторный мотив активации на основе тирозина (ITAM), и такому клеточному ответу может быть помогать или улучшать с помощью костимулирующего домена или его функциональной части. Типичным белком, имеющим эффекторный домен, является CD3ζ. В других вариантах осуществления эффекторный домен будет косвенно стимулировать клеточный ответ путем ассоциации с одним или несколькими другими белками, которые непосредственно стимулируют клеточный ответ.
Термин «костимулирующий домен», используемый в данном документе, относится к сигнальному фрагменту, который предоставляет Т-клеткам сигнал, который, в дополнение к первичному (эффекторному) сигналу, предоставляемому, например, цепью CD3ζ комплекса TCR/CD3, опосредует ответ Т-клеток, включая активацию, пролиферацию, дифференцировку, секрецию цитокинов или тому подобное. В некоторых вариантах осуществления внутриклеточный компонент содержит эффекторный домен или его функциональную часть, костимулирующий домен или его функциональную часть или любую их комбинацию.
«Линкер вариабельной области», в частности, относится к аминокислотной последовательности от 5 до 35, которая связывает вариабельную область тяжелой цепи иммуноглобулина с вариабельной областью легкой цепи иммуноглобулина или соединяет цепи Vα/β и Сαβ рецептора Т-клеток (например, Vα-Cα, Vβ-Cβ, Vα-Vβ) или соединяет каждую пару Vα-Cα, Vβ-Cβ, Vα-Vβ с шарнирным или гидрофобным доменом, который обеспечивает спейсерную функцию и гибкость, достаточные для взаимодействия двух субсвязывающих доменов, так что полученный одноцепочечный полипептид сохраняет специфичную аффинность связывания с той же молекулой-мишенью, что и антитело или рецептор Т-клеток. В некоторых вариантах осуществления линкер вариабельной области содержит от приблизительно 10 до приблизительно 30 аминокислот или от приблизительно 15 до приблизительно 25 аминокислот. В конкретных вариантах осуществления линкерный пептид вариабельной области включает от одного до десяти повторов GlyxSery, где х и у независимо представляют собой целое число от 1 до 5 (например, Gly4Ser (SEQ ID NO: 1), Gly3Ser (SEQ ID NO: 2), Gly2Ser, или (Gly3Ser)n(Gly4Ser), (SEQ ID NO: 3), (Gly3Ser)n(Gly4Ser)n (SEQ ID NO:4), или (Gly4Ser)n (SEQ ID NO: 5), где n представляет собой целое число 1, 2, 3, 4 или 5), и где связанные вариабельные области образуют домен функционального связывания (например, scFv, scTCR).
«Соединительные аминокислоты» или «соединительные аминокислотные остатки» относятся к одному или нескольким (например, около 2-20) аминокислотным остаткам между двумя соседними мотивами, областями или доменами полипептида, такими как между связывающим доменом и соседней линкерной областью или между гидрофобным доменом и смежным эффекторным доменом или на одном или обоих концах линкерной области, которая связывает два мотива, области или домена (например, между линкером и смежным доменом связывания и/или между линкером и смежным шарниром). Соединительные аминокислоты могут быть результатом разработки конструкции слитого белка (например, аминокислотные остатки, полученные в результате использования сайта рестрикционного фермента во время конструирования молекулы нуклеиновой кислоты, кодирующей слитый белок).
Термины, понятные специалистам в области техники антител, имеют значение, приобретенное в данной области техники, если в настоящем документе не указано иное. Термин «антитело» относится к интактному антителу, содержащему, по меньшей мере, две тяжелые (Н) цепи и две легкие (L) цепи, соединенные дисульфидными связями, а также антигенсвязывающую часть интактного антитела, которое имеет или сохраняет способность связывать целевую молекулу. Моноклональное антитело или его антигенсвязывающая часть могут быть нечеловеческим, химерным, гуманизированным или человеческим, предпочтительно гуманизированным или человеческим. Структура и функция иммуноглобулина рассмотрены, например, в Harlow и др., Eds., Antibodies: А Laboratory Manual, глава 14 (Cold Spring Harbor Laboratory, Cold Spring Harbor, 1988).
Например, термины «VL» и «VH» относятся к вариабельной области связывания легкой и тяжелой цепи антитела, соответственно. Области вариабельного связывания состоят из дискретных, четко определенных подобластей, известных как «определяющие комплементарность области» (CDR) и «каркасные области» (FR). Термин «CL» относится к «константной области легкой цепи иммуноглобулина» или «константной области легкой цепи», то есть константной области легкой цепи антитела. Термин «СН» относится к «константной области тяжелой цепи иммуноглобулина» или «константной области тяжелой цепи», которая дополнительно делится в зависимости от изотипа антитела на CH1, СН2 и СН3 (IgA, IgD, IgG) или домены CH1, СН2, СН3 и СН4 (IgE, IgM). «Fab» (фрагмент, связывающий антиген) представляет собой часть антитела, которая связывается с антигенами и включает вариабельную область и СН1 тяжелой цепи, связанные с легкой цепью через межцепочечную дисульфидную связь.
Используемый в данном документе термин «часть области Fc» относится к сегменту константной области тяжелой цепи фрагмента Fc («кристаллизуемой фрагментом» области или области Fc) антитела, который может включать один или несколько константных доменов, таких как СН2, СН3, СН4 или любая их комбинация. В некоторых вариантах осуществления часть области Fc включает домены СН2 и СН3 антитела IgG, IgA или IgD или любую их комбинацию или домены СН3 и СН4 антитела IgM или IgE и любую их комбинацию. В других вариантах осуществления структура СН2СН3 или СН3СН4 имеет субобластные домены из одного и того же изотипа антитела и являются человеческими, такими как человеческие IgG1, IgG2, IgG3, IgG4, IgAl, IgA2, IgD, IgE или IgM (например, CH2CH3 из человеческого IgG1). В качестве фона, область Fc отвечает за эффекторные функции иммуноглобулина, такие как ADCC (антителозависимая клеточная цитотоксичность), CDC (комплементзависимая цитотоксичность) и фиксация комплемента, связывание с рецепторами Fc (например, CD16)., CD32, FcRn), большее время полужизни in vivo по сравнению с полипептидом, лишенным области Fc, связывания белка А и, возможно, даже переноса плаценты (см. Capon и др., Nature 337: 525, 1989). В некоторых вариантах осуществления часть области Fc, обнаруженный в слитых белках согласно настоящему изобретению, будет способен опосредовать одну или несколько из этих эффекторных функций или будет не обладать одной или несколькими или всеми этими активностями, например, одной или несколькими мутациями, известными в области технике.
Кроме того, антитела имеют шарнирную последовательность, которая обычно расположена между областью Fab и Fc (но нижняя часть шарнира может включать аминоконцевую часть области Fc). В качестве фона, шарнир иммуноглобулина действует как гибкая прокладка, позволяющая Fab-части свободно перемещаться в пространстве. В отличие от константных областей, шарниры структурно разнообразны, варьируются как по последовательности, так и по длине между классами иммуноглобулинов и даже среди подклассов. Например, шарнирная область человеческого IgG1 является свободно гибкой, что позволяет фрагментам Fab вращаться вокруг своих осей симметрии и перемещаться в сфере, центрированной в первом из двух дисульфидных мостиков между тяжелыми цепями. Для сравнения, шарнир человеческого IgG2 является относительно коротким и содержит жесткую двойную спираль из полипролина, стабилизированную четырьмя дисульфидными мостиками между тяжелыми цепями, что ограничивает гибкость. Шарнир человеческого IgG3 отличается от других подклассов своей уникальной расширенной шарнирной областью (приблизительно в четыре раза длиннее шарнира IgGl), содержащей 62 аминокислоты (включая 21 пролин и 11 цистеинов), образуя негибкую двойную спираль полипролина и обеспечивая большую гибкость, поскольку фрагменты Fab находятся относительно далеко от фрагмента Fc. Шарнир человеческого IgG4 короче, чем IgG1, но имеет ту же длину, что и IgG2, и его гибкость является промежуточной между гибкостью IgG1 и IgG2.
«Т-клеточный рецептор» (TCR) относится к молекуле, обнаруженной на поверхности Т-клеток (или Т-лимфоцитов), которая, в связи с CD3, обычно ответственна за распознавание антигенов, связанных с молекулами главного комплекса гистосовместимости (МНС). TCR имеет дисульфид-связанный гетеродимер сильно изменчивых α и β цепей (также известных как TCRα и TCRβ соответственно) в большинстве Т-клеток. В небольшой подпопуляции Т-клеток TCR состоит из гетеродимера с переменными γ и δ цепями (также известных как TCRγ и TCRδ соответственно). Каждая цепь TCR является членом суперсемейства иммуноглобулинов и обладает одним N-терминальным вариабельным доменом иммуноглобулина, одним константным доменом иммуноглобулина, трансмембранной областью и коротким цитоплазматическим хвостом на С-терминальном конце (см. Janeway и др., Immunobiology: The immune system and disease, 3-е изд., Current Biology Publications, стр. 4:33, 1997). TCR, как используется в настоящем документе, может быть от различных видов животных, включая человека, мышь, крысу, кошку, собаку, козу, лошадь или других млекопитающих. TCR могут быть связаны с клетками (то есть иметь трансмембранную область или домен) или в растворимой форме.
«Молекулы главного комплекса гистосовместимости» (молекулы МНС) относятся к гликопротеинам, которые доставляют пептидные антигены на клеточную поверхность. Молекулы МНС класса I представляют собой гетеродимеры, состоящие из α-цепи, охватывающей мембрану (с тремя α-доменами), и нековалентно связанного β2-микроглобулина. Молекулы МНС класса II состоят из двух трансмембранных гликопротеинов, α и β, которые охватывают мембрану. Каждая цепочка имеет два домена. Молекулы МНС класса I доставляют пептиды, происходящие из цитозоля, на клеточную поверхность, где комплекс пептид: МНС распознается CD8+ Т-клетками. Молекулы МНС класса II доставляют пептиды, происходящие из везикулярной системы, на клеточную поверхность, где они распознаются CD4+ Т-клетками. Молекула МНС может быть из различных видов животных, включая человека, мышь, крысу или других млекопитающих.
«Вектор» представляет собой молекулу нуклеиновой кислоты, которая способна транспортировать другую нуклеиновую кислоту. Векторы могут быть, например, плазмидами, космидами, вирусами или фагом. «Вектор экспрессии» представляет собой вектор, который способен направлять экспрессию белка, кодируемого одним или несколькими генами, переносимыми вектором, когда он присутствует в соответствующей среде.
«Ретровирусы» представляют собой вирусы, имеющие РНК геном. «Гаммаретровирус» относится к роду семейства Retro viridae. Типичные гаммаретровирусы включают вирус стволовых клеток мыши, вирус лейкоза мыши, вирус лейкоза кошки, вирус саркомы кошки и вирусы птичьего ретикулоэндотелиоза.
«Лентивирус» относится к роду ретровирусов, которые способны инфицировать делящиеся и неделящиеся клетки. Несколько примеров лентивирусов включают ВИЧ (вирус иммунодефицита человека: включая ВИЧ типа 1 и ВИЧ типа 2); вирус инфекционной анемии лошадей; вирус иммунодефицита кошек (FIV); вирус иммунодефицита крупного рогатого скота (BIV); и вирус иммунодефицита обезьян (SIV).
«Т-клетки» или «клетки линии Т-клеток» относятся к клеткам, которые проявляют по меньшей мере одну фенотипическую характеристику Т-клетки или ее предшественника, который отличает клетки от других лимфоидных клеток, и клетки эритроидных или миелоидных линий. Такие фенотипические характеристики могут включать экспрессию одного или нескольких белков, специфичных для Т-клеток (например, CD3+, CD4+, CD8+), или физиологических, морфологических, функциональных или иммунологических признаков, специфичных для Т-клеток. Например, клетки линии Т-клеток могут быть клетками-предшественниками или клетками-предшественниками, связанными с линией Т-клеток; CD25+ незрелые и инактивированные Т-клетки; клетки, которые подверглись связыванию CD4 или CD8; клетки-предшественники тимоцитов, которые представляют собой дважды положительные CD4+ CD8+; одноположительные CD4+ или CD8+; TCRαβ или TCR γδ; или зрелые и функциональные или активированные Т-клетки.
«Молекула нуклеиновой кислоты» или «полинуклеотид» может быть в форме РНК или ДНК, которая включает кДНК, геномную ДНК и синтетическую ДНК. Молекула нуклеиновой кислоты может быть двухцепочечной или одноцепочечной, и, если одноцепочечная, может быть кодирующей цепью или некодирующей (антисмысловой цепью). Кодирующая молекула может иметь кодирующую последовательность, идентичную кодирующей последовательности, известной в данной области техники, или может иметь другую кодирующую последовательность, которая в результате избыточности или вырожденности генетического кода или путем сплайсинга может кодировать один и тот же полипептид.
«Лечить» или «лечение» или «улучшать состояние» относится к медицинским мерам борьбы с заболеванием, расстройством или состоянием субъекта (например, человека или млекопитающего, не являющегося человеком, такого как примат, лошадь, собака, мышь, крыса). Например, подходящую дозу или схему лечения, включающую специфичный для ВСМА-специфичный связывающий белок, или клетку-хозяина, экспрессирующую ВСМА-специфичный связывающий белок, используемый в сочетании с ингибитором у-секретазы (GSI) согласно настоящему изобретению, и, необязательно, адъювантом или режимом предварительного кондиционирования, вводят для выявления терапевтического или профилактического эффекта. Терапевтическая или профилактическая/профилактическая польза включает улучшение клинического исхода; уменьшение или облегчение симптомов, связанных с заболеванием; уменьшение появления симптомов; улучшение качества жизни; более длительный статус без болезней; уменьшение степени заболеваемости, стабилизацию болезненного состояния; замедления прогрессирования заболевания; ремиссию; выживание; длительное выживание; или любую их комбинацию.
«Терапевтически эффективное количество» или «эффективное количество» ВСМА-специфичного связывающего белка (также называемого ВСМА-специфичной или ВСМА-таргетированной иммунотерапией), ингибитора γ-секретазы, клеток-хозяев, экспрессирующая ВСМА-специфичный связывающий белок или клетка-хозяин, экспрессирующих ингибитор γ-секретазы согласно настоящему изобретению (например, ВСМА-специфичный CAR, анти-γ-секретазное антитело), относится к такому количеству соединения или клеток, которое достаточно для ослабления одного или нескольких симптомов заболевания, которое лечат достоверным образом. Когда речь идет об отдельном активном ингредиенте или клетке, экспрессирующей один активный ингредиент в отдельности, вводимой отдельно, терапевтически эффективная доза относится к действию этого ингредиента или клетки, экспрессирующей этот ингредиент в отдельности. Когда речь идет о комбинации, терапевтически эффективная доза относится к объединенным количествам активных ингредиентов или объединенного вспомогательного активного ингредиента с клеткой, экспрессирующей активный ингредиент, что приводит к терапевтическому эффекту при введении последовательно, параллельно или одновременно. Другая комбинация может представлять собой клетку, экспрессирующую более одного активного ингредиента, таких как два или более различных ВСМА-специфичных связывающих белка или тому подобное.
Дополнительные определения предоставлены по всему настоящему описанию.
Белок или молекулы, связывающие антиген созревания В-клеток (ВСМА)
В определенных аспектах настоящее описание предоставляет способы лечения пролиферативного или аутоиммунного заболевания или расстройства у субъекта, имеющего или подозреваемого на наличие заболевания или расстройства, включающие введение субъекту терапевтически эффективного количества ВСМА-специфичного связывающего белка (или ВСМА-таргетированная иммунотерапия) и терапевтически эффективного количества ингибитора γ-секретазы. Типичным ВСМА-специфичным связывающим белком является химерный антигенный рецептор, содержащий внеклеточный компонент и внутриклеточный компонент, связанный гидрофобной частью, где внеклеточный компонент содержит ВСМА-специфичный связывающий домен (например, ВСМА-специфичный scFv, лиганд ВСМА или его связывающую часть, например, BAFF или APRIL), и необязательно содержит спейсерную область или шарнир, и где внутриклеточный компонент содержит эффекторный домен и, необязательно, костимулирующий домен.
В некоторых вариантах осуществления настоящее описание предоставляет ВСМА-таргетированную иммунотерапию, для применения с ингибитором γ-секретазы для лечения пролиферативного или аутоиммунного заболевания или расстройства, содержащую ВСМА-специфичное антитело или его антигенсвязывающую часть, химерный антигенный рецептор (CAR) или меченую молекулу химерного антигенного рецептора (T-ChARM). В некоторых вариантах осуществления ВСМА-специфичное антитело или его антигенсвязывающая часть являются человеческими или гуманизированными.
Типичные ВСМА-специфические антитела включают антитела J22.0-xi, J22.9-xi, J6M0, J6M1, J6M2, J9M0, J9M1, J9M2, 11D5-3, СА8, A7D12.2, СП D5.3, С12А3.2, C13F12.1, 13С2, 17А5, 83А10, 13А4, 13D2, 14В11, 14Е1, 29В11, 29F3, 13А7, СА7, SGI, S307118G03, S332121F02, S332126E04, S322110D07, S336105A07, S335115G01, S335122F05, ЕТ140-3, ЕТ140-24, ЕТ140-37, ЕТ140-40, ЕТ140-54, TBL-CLN1, С4.Е2.1, Vicky-1, pSCHLI333, pSCHLI372 и pSCHLI373 и их антигенсвязывающие части. Различные варианты ВСМА-специфичных антител и их антигенсвязывающих частей, включая гуманизированные варианты, описаны, например, в публикациях РСТ №WO 2002/066516, WO 2007/062090, WO 2010/104949, WO 2011/108008, WO 2012/163805, WO 2014/068079, WO 2015/166073, WO 2014/122143, WO 2014/089335, WO 2016/090327 и WO 2016/079177; Ryan и др., Mol. Рак. Ther. 6 (11): 3009, 2007; и Abbas и др., Blood 128: 1688, 2016, в которых ВСМА-специфичные антитела, их антигенсвязывающие части и гуманизированные варианты включены в настоящее описание в качестве ссылки во всей их полноте. Вариабельные домены и молекулы scFv из этих ВСМА-специфичных антител можно использовать в качестве связывающего домена в любом из упомянутых здесь белков T-ChARM и CAR.
Антигенсвязывающие части или домены, полученные из ВСМА-специфичных антител согласно настоящему изобретению и применимые в способах, описанных в настоящем документе, включают, например, доменные антитела, sFv, одноцепочечные фрагменты Fv (scFv), Fabs, F (ab ')2, нанотела, тандемные scFvs, scFv-Fcs, димеры scFv, "застежки-молнии" scFv, диатела, минитела, триатела, тетратела, Fabs, F (ab) '2s, scFabs, миниантитела, нанотела, нанотела-HSA, биспецифичные Т-клетки Engagers (BiTEs), DAR, sc-диатела, sc-диатела-СН3 или scFv-СН3 сборки выступы-во-впадины (KIH).
В некоторых вариантах осуществления ВСМА-специфичный связывающий белок содержит биспецифичное или мультиспецифичное антитело (или его антигенсвязывающую часть), содержащее первую область связывания (например, вариабельную область тяжелой цепи, вариабельную область легкой цепи или обе), которая является специфичной для ВСМА и, по меньшей мере, одной другой области связывания, которая специфична для второй мишени (например, эпитопа ВСМА, который отличается от эпитопа первой области связывания, или эпитопа мишени не-ВСМА, такой как, например, ассоциированный с опухолью антиген, который не является ВСМА (например, CD19 (например, блинатумомаб, MOR-208, SGN-19A, SAR3419, колтуксимабравтанзин, денитузумабмафодотин, таплитумомабпаптокс, XmAb 5574, MDI-551, патент Мерк анти-CD19, известный также как В4, известный также как DI-B4, XmAb 5871, MDX-1342, AFM11), CD20 (например, ритуксимаб, офатумумаб, окрелизумаб), CD38 (например, даратумумаб или изатуксимаб (SAR650984)), CD45 или белок клеточной поверхности, экспрессируемый на иммунноффекторной клетке, такой как Т-клетка (например, CD3), NK-клетка (естественная клетка киллер) (например, CD56) или NK Т-клетка (например, NK1.1) или другой, не связанный с опухолью антиген или мишень.
В конкретных вариантах осуществления настоящее описание предоставляет ВСМА-специфичный связывающий белок, один или экспрессируемый в виде T-ChARM в клетке, для использования с ингибитором у-секретазы (GSI). Пример T-ChARM содержит внеклеточный компонент и внутриклеточный компонент, соединенные гидрофобной частью, где внеклеточный компонент содержит связывающий домен, который специфично связывает ВСМА, необязательную спейсерную область, метку-кассету и шарнирную область, и где внутриклеточный компонент содержит эффекторный домен и, необязательно, ко стимулирующий домен (например, функциональный домен или часть из 4-1 ВВ, функциональный домен или часть из CD28 или оба). В некоторых вариантах осуществления связывающий домен T-ChARM содержит ВСМА-специфичный scFv, ВСМА-специфичный scTCR или лиганд ВСМА или его связывающую часть (например, BAFF, APRIL), необязательно, где ВСМА-специфичный scFv является человеческим или гуманизированным. Различные варианты осуществления T-ChARM описаны в публикации РСТ №WO 2015/095895, в которой скаффолды T-ChARM полностью включены в настоящее описание посредством ссылки.
Метки-кассеты включают в себя Strep-метку (которая относится к оригинальной Strep®-метке, Strep®-метке II или любому ее варианту; см., например, патент США №7981322, Strep-метки которого включены в настоящее описание посредством ссылки), His метка, Flag метка, метка Xpress, метка Avi, метка кальмодулина, метка полиглутамата, метка НА, метка Мус, метка Nus, метка S, метка SBP, метка Softag 1, метка Softag 3, метка V5, CREB-связывающий белок (СВР), глутатион-S-трансфераза (GST), мальтозосвязывающий белок (МВР), зеленый флуоресцентный белок (GFP), тиоредоксиновый тег или любая их комбинация. В некоторых вариантах осуществления метка-кассета может представлять собой генетически сконструированный аффинный сайт, такой как сайт минимального хелатирования (например, HGGHHG, SEQ ID NO: 6). В некоторых вариантах осуществления метка-кассета представляет собой Strep-метку, имеющую аминокислотную последовательность Trp-Ser-His-Pro-Gln-Phe-Glu-Lys (SEQ ID NO: 7) или Trp-Arg-His-Pro-Gln-Phe-Gly-Gly (SEQ ID NO: 8).
Метки-кассеты могут присутствовать в одной или нескольких копиях во слитых белках данного описания. Например, ВСМА-специфичный связывающий белок согласно настоящему изобретению может иметь одну, две, три, четыре или пять меток-кассет (например, Strep-метка). В некоторых вариантах осуществления внеклеточный компонент ВСМА-специфичного T-ChARM включает одну метку-кассету с метками, две метки-кассеты, три метки-кассеты, четыре метки-кассеты или пять меток-кассет. Каждая из множества меток-кассет может быть одинаковой или разной.
В некоторых вариантах осуществления кассета-метка содержит от около 5 до около 500 аминокислот, или от около 6 до около 100 аминокислот, или от около 7 до около 50 аминокислот, или от около 8 до около 20 аминокислот. В некоторых вариантах осуществления метка-кассета содержит от семи до десяти аминокислот. Предпочтительно, метка-кассета является неиммуногенной или минимально иммуногенной. По существу, метка-кассета может функционировать в качестве указателя или маяка для идентификации, обогащения, выделения, стимулирования пролиферации, активации, отслеживания или удаления клеток, экспрессирующих ВСМА-специфичный T-ChARM.
В дополнителных вариантах осуществления настоящее описание предоставляет ВСМА-специфичный связывающий белок для использования с ингибитором γ-секретазы, который представляет собой химерный антигенный рецептор (CAR), который включает внеклеточный компонент и внутриклеточный компонент, соединенные гидрофобной частью, где внеклеточный компонент содержит связывающий домен, который специфично связывает ВСМА, и шарнирную область, и где внутриклеточный компонент содержит эффекторный домен и, необязательно, костимулирующий домен. В некоторых вариантах осуществления домен связывания CAR содержит ВСМА-специфичный scFv, ВСМА-специфичный scTCR, ВСМА-специфичный домен связывания TCR (см., например, Walseng и др., Scientific Reports 7: 10713 (2017), CAR TCR конструкции, включенные в настоящее описание в качестве ссылки во всей их полноте), или лиганд ВСМА или его связывающая часть (например, BAFF, APRIL), необязательно, где ВСМА-специфичный scFv является человеческим или гуманизированным. В любом из этих вариантов осуществления ВСМА-специфичный связывающий белок в форме CAR может экспрессироваться на поверхности клетки, такой как клетка иммунной системы (например, Т-клетка).
ВСМА-специфичный T-ChARM или CAR может быть связан с клеткой (например, экспрессирован на поверхности клетки) или находиться в растворимой форме. В некоторых вариантах осуществления полинуклеотиды, кодирующие ВСМА-специфичные белки T-ChARM или CAR, могут быть оптимизированы по кодонам для усиления или максимизации экспрессии в клетке-хозяине, такой как Т-клетка (Scholten и др., Clin. Immunol. 119: 135,2006),
В некоторых вариантах осуществления шарнир, присутствующий в ВСМА-специфичном T-ChARM или CAR согласно настоящему изобретению, может представлять собой шарнирную область иммуноглобулина, такую как шарнирная область иммуноглобулина дикого типа или измененная шарнирная область иммуноглобулина. В некоторых вариантах осуществления шарнир представляет собой шарнирную область иммуноглобулина человека дикого типа. В некоторых других вариантах осуществления один или несколько аминокислотных остатков могут быть добавлены на амино- или карбоксиконце шарнирной области иммуноглобулина дикого типа как часть конструкции слитого белка. Например, один, два или три дополнительных аминокислотных остатка могут присутствовать на амино-конце или карбокси-конце шарнира, или шарнир может содержать терминальную или внутреннюю делецию и добавлять обратно один, два или три дополнительные аминокислотных остатка.
В некоторых вариантах осуществления шарниром является измененный шарнир иммуноглобулина, в котором один или несколько остатков цистеина в шарнирной области иммуноглобулина дикого типа замещены одним или несколькими другими аминокислотными остатками. Типичные измененные шарниры иммуноглобулина включают в себя шарнирную область иммуноглобулина человека IgG1, IgG2 или IgG4, имеющую один, два или три остатка цистеина, обнаруженные в шарнире человеческого дикого типа IgG1, IgG2 или IgG4, замещенной одним, двумя или тремя различными аминокислотными остатками (например, серином или аланин). В некоторых вариантах осуществления шарнирный полипептид содержит или представляет собой последовательность, которая составляет, по меньшей мере, 80%, по меньшей мере, 81%, по меньшей мере, 82%, по меньшей мере, 83%, по меньшей мере, 84%, по меньшей мере, 85%, по меньшей мере, 86%, по меньшей мере, 87%, по меньшей мере, 88%, по меньшей мере, 89%, по меньшей мере, 90%, по меньшей мере, 91%, по меньшей мере, 92%, по меньшей мере, 93%, по меньшей мере, 94%, по меньшей мере, 95%, по меньшей мере, 96%, по меньшей мере, 97%, по меньшей мере, 98%, по меньшей мере, на 99% идентичны шарнирной области иммуноглобулина дикого типа, такой как шарнир человеческого IgG1 дикого типа, шарнир человеческого IgG2 дикого типа или шарнир человеческого IgG4 дикого типа.
В дополнительных вариантах осуществления шарнир, присутствующий в ВСМА-специфичном T-ChARM или CAR согласно настоящему изобретению, может представлять собой шарнир, который не основан на шарнире иммуноглобулина или не получен из него (т.е. не является шарниром иммуноглобулина дикого типа или измененным шарниром иммуноглобулина), примеры таких шарниров включают в себя пептиды, содержащие от приблизительно пяти до приблизительно 150 аминокислот области стебля С-лектинов типа II или молекул CD, включая пептиды, содержащие от приблизительно 8 до приблизительно 25 аминокислот, или пептиды, содержащие от приблизительно 7 до приблизительно 18 аминокислот, или их варианты.
Термин «область стебля» молекулы С-лектина типа II или молекулы CD относится к части внеклеточного домена молекулы С-лектина типа II или молекулы CD, которая расположена между лектиноподобным доменом С-типа (CTLD; например, похожим на CTLD рецепторов естественных клеток-киллеров) и гидрофобной частью (трансмембранный домен). Например, внеклеточный домен человеческого CD94 (номер GenBank ААС50291.1) соответствует аминокислотным остаткам 34-179, но CTLD соответствует аминокислотным остаткам 61-176, поэтому область стебля молекулы CD94 человека содержит аминокислотные остатки 34-60, которые расположены между гидрофобной частью (трансмембранный домен) и CTLD (см. Boyington и др., Immunity 10:75, 1999; для описания других областей стебля, см. также Beavil и др., Proc. Nat. '1. Acad. Sci. USA 89: 753, 1992; и Figdor и др., Nat. Rev. Immunol. 2:77, 2002). Эти молекулы С-лектина типа II или CD молекулы могут также иметь соединительные аминокислоты между областью стебля и трансмембранной областью или CTLD. В другом примере 233-аминокислотный белок NKG2A человека (номер в GenBank Р26715.1) имеет гидрофобную часть (трансмембранный домен) в пределах аминокислот 71-93 и внеклеточный домен в пределах аминокислот 94-233. CTLD содержит аминокислоты 119-231, а область стебля содержит аминокислоты 99-116, которые могут быть фланкированы дополнительными соединительными аминокислотами. В данной области известны молекулы С-лектин типа II или другие CD молекулы, а также их внеклеточные лиганд-связывающие домены, области стебля и CTLD (см., например, номера в GenBank NP_001993.2; ААН07037.1; NP_001773.1; AAL65234.1; САА04925.1; для последовательностей человеческих CD23, CD69, CD72, NKG2A и NKG2D и их описания соответственно).
«Производное» шарнира области стебля или его фрагмент, молекулы С-лектина типа II или молекулы CD включает от приблизительно 8 до приблизительно 150 аминокислотных последовательностей, в которых одна, две или три аминокислоты области стебля являются аминокислотами дикого типа молекулы С-лектина типа II или молекулы CD имеют делецию, вставку, замену или любую их комбинацию. Например, производное может содержать одну или несколько аминокислотных замен и/или аминокислотную делецию. В некоторых вариантах осуществления производное области стебля является более устойчивым к протеолитическому расщеплению по сравнению с последовательностью области стебля дикого типа, такой как последовательности, происходящие от приблизительно восьми до приблизительно 20 аминокислот NKG2A, NKG2D, CD23, CD64, CD72, или CD94.
В некоторых вариантах осуществления шарниры области стебля могут содержать от приблизительно 7 до приблизительно 18 аминокислот и могут образовывать α-спиральную суперспиральную структуру. В некоторых вариантах осуществления шарниры области стебля содержат 0, 1, 2, 3 или 4 цистеина. Типичные шарниры области стебля включают фрагменты областей стебля, такие как те части, которые содержат от приблизительно 10 до приблизительно 150 аминокислот из областей стебля CD69, CD72, CD94,NKG2AhNKG2D.
Альтернативные шарниры, которые можно использовать в ВСМА-специфичных Т-CliARM и CAR по данному изобретению, относятся к частям рецепторов клеточной поверхности (междоменные области), которые соединяют V-подобные иммуноглобулиновые или С-подобные иммуноглобулиновые домены. Области между Ig V-подобными доменами, где рецептор клеточной поверхности содержит несколько Ig V-подобных доменов в тандеме, и между Ig С-подобными доменами, где рецептор клеточной поверхности содержит множественные тандемные Ig С-подобные области, также рассматриваются как шарниры, используемые в ВСМА-специфичных T-ChARM и CAR по данному изобретению. В некоторых вариантах осуществления шарнирные последовательности, состоящие из междоменных областей рецептора клеточной поверхности, могут дополнительно содержать встречающийся в природе или добавленный мотив, такой как стержневая последовательность ядра IgG, для обеспечения одной или нескольких дисульфидных связей для стабилизации ВСМА-специфичного T-ChARM или образования димера CAR. Примеры шарниров включают междоменные области между Ig V-подобными и Ig С-подобными областями CD2, CD4, CD22, CD33, CD48, CD58, CD66, CD80, CD86, CD150, CD166 или CD244.
В некоторых вариантах осуществления шарнирные последовательности содержат от приблизительно 5 до приблизительно 150 аминокислот, от приблизительно 5 до приблизительно 10 аминокислот, от приблизительно 10 до приблизительно 20 аминокислот, от приблизительно 20 до приблизительно 30 аминокислот, от приблизительно 30 до приблизительно 40 аминокислот, приблизительно 40 до приблизительно 50 аминокислот, от приблизительно 50 до приблизительно 60 аминокислот, от приблизительно 5 до приблизительно 60 аминокислот, от приблизительно 5 до приблизительно 40 аминокислот, например, от приблизительно 8 до приблизительно 20 аминокислот или от приблизительно 10 до приблизительно 15 аминокислот. Шарниры могут быть в основном гибкими, но могут также обеспечивать более жесткие характеристики или могут содержать в основном а-спиральную структуры с минимальным количеством структур β-листа.
В некоторых вариантах осуществления шарнирная последовательность является стабильной в плазме и сыворотке и устойчива к протеолитическому расщеплению. Например, первый лизин в верхней шарнирной области IgG1 может быть мутирован или удален, чтобы минимизировать протеолитическое расщепление, и шарниры могут включать соединительные аминокислоты. В некоторых вариантах осуществления шарнирная последовательность может содержать встречающийся в природе или добавленный мотив, такой как структура ядра шарнира иммуноглобулина СРРСР (SEQ ID NO: 9), которая придает способность образовывать дисульфидную связь или множественные дисульфидные связи для стабилизации образования димера.
Гидрофобная часть, содержащаяся в ВСМА-специфичном связывающем белке согласно настоящему изобретению (например, ВСМА-специфичный T-ChARM или CAR), позволит ВСМА-специфичному связывающему белку согласно настоящему изобретению связываться с клеточной мембраной, так что часть связывающего белка будет локализована внеклеточно (например, метка-кассета, связывающий домен), а часть будет локализована внутриклеточно (например, эффекторный домен, костимулирующий домен). Гидрофобная часть обычно располагается внутри фосфолипидного бислоя клеточной мембраны. В некоторых вариантах осуществления одна или несколько соединительных аминокислот могут быть расположены между и соединять гидрофобную часть с эффекторным доменом, или могут располагаться между и соединять гидрофобную часть со спейсерной областью, или располагаться между и соединять гидрофобную часть с меткой-кассетой.
В некоторых вариантах осуществления гидрофобный домен представляет собой трансмембранный домен, такой как домен, полученный из интегрального мембранного белка (например, рецептор, молекула кластера дифференцировки (CD), фермент, транспортер, молекула клеточной адгезии или тому подобное). В конкретных вариантах осуществления гидрофобная часть представляет собой трансмембранный домен из CD4, CD8, CD27 или CD28.
Внутриклеточный компонент, содержащийся в ВСМА-специфичном связывающем белке согласно настоящему изобретению (например, ВСМА-специфичном T-ChARM или CAR), будет способен передавать функциональные сигналы в клетку. В некоторых вариантах осуществления ВСМА-специфичные T-ChARM или CAR будут димеризоваться со вторым одноцепочечным T-ChARM или CAR, соответственно, где димеризация позволяет внутриклеточному компоненту, содержащему эффекторный домен и, необязательно, костимулирующий домен, находиться в непосредственной близости и способствовать передаче сигнала при воздействии правильного сигнала. В дополнение к образованию таких димерных белковых комплексов эффекторные домены и необязательные костимулирующие домены могут дополнительно ассоциироваться с другими сигнальными факторами, такими как костимулирующие факторы, с образованием мультипротеиновых комплексов, которые продуцируют внутриклеточный сигнал. В некоторых вариантах осуществления эффекторный домен будет косвенно стимулировать клеточный ответ путем ассоциации с одним или несколькими другими белками, которые непосредственно стимулируют клеточный ответ. Внутриклеточный компонент может включать один, два, три или более рецепторных сигнальных доменов (например, эффекторных доменов), костимулирующие домены или их комбинации. Любой внутриклеточный компонент, содержащий эффекторный домен или его функциональную часть, костимулирующий домен или его функциональную часть или любую их комбинацию из любой из множества сигнальных молекул (например, рецепторов сигнальной трансдукции), можно использовать в ВСМА-специфичных связывающих белках по настоящему изобретению.
Внутриклеточный компонент может иметь эффекторный или костимулирующий домен, полезный в ВСМА-специфичных связывающих белках данного описания, которые могут основываться на белке сигнального пути Wnt (например, LRP, Ryk, ROR2) или сигнальном пути NOTCH (например, NOTCH 1, NOTCH2, NOTCH3, NOTCH4), сигнальном пути Hedgehog (например, PTCH, SMO), рецепторных тирозинкиназах (RTK) (например, семейство рецепторов эпидермального фактора роста (EGF), семейство рецепторов фактора роста фибробластов (FGF), гепатоциты семейство рецепторов фактора роста (HGF), семейство рецепторов инсулина (IR), семейство рецепторов тромбоцитарного фактора роста (PDGF), семейство рецепторов сосудистого эндотелиального фактора роста (VEGF), семейство рецепторов тропинациновой киназы (Trk), рецептор эфрина (Eph) семейство, семейство рецепторов AXL, семейство рецепторов лейкоцитарной тирозинкиназы (LTK), тирозинкиназа с семейством рецепторов иммуноглобулиноподобных и EGF-подобных доменов 1 (TIE), семейство тирозинкиназоподобных орфан-рецепторов (ROR), семейство дискоидиновых доменов (DDR), семейство рецепторов Ly, перестроенные во время семейства рецепторов трансфекции (RET), семейство рецепторов тирозин-протеинкиназы (РТК7), относящихся к семейству рецепторов рецепторной тирозинкиназы (RYK), семейству рецепторов мышечно-специфичной киназы (MuSK); рецепторы, связанные с G-белком, GPCR (Frizzled, Smoothened); рецепторы серин/треонинкиназы (BMPR, TGFR); или цитокиновые рецепторы (IL-1R, IL-2R, IL-7R, IL-15R).
В некоторых вариантах осуществления эффекторный домен содержит сигнальный домен рецептора лимфоцитов или содержит аминокислотные последовательности, имеющие один или несколько иммунорецепторных мотивов активации на основе тирозина (ITAM). В дополнительных вариантах осуществления эффекторный домен содержит цитоплазматическую часть, которая ассоциируется с цитоплазматическим сигнальным белком, где цитоплазматический сигнальный белок представляет собой рецептор лимфоцита или его сигнальный домен, белок, содержащий множество ITAM, костимулирующий фактор или любую их комбинацию.
Типичные эффекторные и костимулирующие домены включают домены, основанные или полученные из 4-1ВВ, CD3e, CD38, CD3ξ, CD27, CD28, CD79A, CD79B, CARD11, DAP10, FcRα, FcRβ, FcRγ, Fyn, HVEM, ICOS, Lck, LAG3, LAT, LRP, NOTCH1, Wnt, NKG2D, OX40, ROR2, Ryk, SLAMF1, Slp76, pTα, TCRα, TCRβ, TRIM, Zap70, PTCH2 или любой их комбинации.
В конкретных вариантах осуществления ВСМА-специфичные связывающие белки согласно настоящему изобретению содержат (а) эффекторный домен из CD3ξ или его функциональной части и костимулирующий домен из CD28 или его функциональной части, (b) эффекторный домен из CD3ξ или его функциональной части и костимулирующий домен из 4-1ВВ или его функциональной части, или (с) (а) эффекторный домен из CD3ξ или его функциональной части и костимулирующий домен из CD28 и 4-1ВВ или их функциональных частей.
Ингибиторы γ-секретазы (GSI)
В качестве основы, γ-секретаза представляет собой мультисубъединичный интегральный мембранный протеазный комплекс, включающий пресенилин (PS), никастрин (NCT), белок дефекта передней глотки 1 (anterior pharynx-defective 1)(АРН-1) и энхансер презенилина 2 (PEN-2). PS представляет собой каталитическую субъединицу, которая представляет собой аспартатпротеазу, способную образовывать гидрофильную каталитическую пору, утопленную в мембране (Takasugi и др., Nature 422: 438-41, 2003), которая расщепляет однопроходные трансмембранные белки внутри трансмембранного домена. NCT представляет собой мембранный гликопротеин типа I с большим внеклеточным доменом (ECD), который фиксирует аминоконец субстрата в качестве первичного субстратного рецептора для γ-секретазы (Shah и др., Cell 122: 435-47, 2005)., Комплекс у-секретазы играет роль в процессинге различных субстратов, включая Notch, CD44, кадгерины и эфрин В2, а также расщеплении белка-предшественника амилоида до амилоидного бета-пептида, который участвует в болезни Альцгеймера. Известно также, что комплекс γ-секретазы расщепляет антиген созревания В-клеток (ВСМА) (Laurent и др., Nature Communications 6, 2015), который является терапевтической мишенью при различных формах рака, включая множественную миелому.
Типичные ингибиторы γ-секретазы (GSI) включают малые молекулы, пептидомиметические соединения или специфичные для γ-секретазы связывающие белки. GSI может нацеливаться на любой один или несколько белков комплекса γ-секретазы, включая пресенилин 1 (PS1), пресенилин 2 (PS2), никастрин (NCT), белок дефекта передней глотки 1 (anterior pharynx-defective 1) (АРН-1) и энхансер презенилина 2 (PEN-2) при условии, что активность расщепления γ-секретазой снижена по сравнению с неингибированной γ-секретазой. В некоторых вариантах осуществления активность γ-секретазы снижается, по меньшей мере, приблизительно на 50%, по меньшей мере, приблизительно на 60%, по меньшей мере, приблизительно 70%, по меньшей мере, приблизительно 75%, по меньшей мере, приблизительно 80%, по меньшей мере, приблизительно 85%, по меньшей мере, приблизительно 90%, по крайней мере, около 95% или 100%. Анализы для измерения активности γ-секретазы известны в данной области (см., например, Laurent и др., 2015). Например, уровень растворимого ВСМА может быть суррогатной мерой активности γ-секретазы. Репрезентативные низкомолекулярные GSI для использования с ВСМА-таргетированной иммуннотерапией для лечения пролиферативного или аутоиммунного заболевания или расстройства включают авагацестат, DAPT, BMS-906024, BMS-986115, LY411575, МК-0752, PF-03084014, RO4929097, семагацестат YO-01027 и любую их комбинацию.
Другими GSI являются γ-секретаза-специфичные связывающие белки, такие как антитела или их антигенсвязывающие части, такие как комплекс γ-секретазы или белок-γ-секретаза, такие как пресенилин 1 (PS1), пресенилин 2 (PS2), никастрин (NCT), дефект передней глотки 1 (anterior pharynx-defective 1) (АРН-1) и энхансер пресенилина 2 (PEN-2). Типичным γ-секретаза-специфичным связывающим белком является никастрин-специфичный связывающий белок, такой как антитела scFvG9, А5226А, 2Н6, 10С11 и их антигенсвязывающие фрагменты.
Связывающие Домены
Связывающий домен может представлять собой любой пептид, который специфично связывает ВСМА или специфично ингибирует активность у-секретазы, как описано в настоящем документе. Источники связывающих доменов включают вариабельные области антител различных видов (которые могут быть в форме антител, sFv, scFv, Fabs, grababody на основе scFv или растворимых доменов VH или доменов антител), включая антитела человека, грызунов, птиц или овец. Дополнительные источники связывающих доменов включают вариабельные области антител других видов, таких как верблюд (от верблюдов, дромадеров или лам; Ghahroudi и др., FEBS Lett. 414: 521, 1997; Vincke и др., J. Biol. Chem. 284: 3273, 2009; Hamers-Casterman и др., Nature 363: 446, 1993 и Nguyen и др., J. Mol. Biol. 275: 413, 1998), акулы-медсестры (Roux и др., Ргос.Nat. 'L. Acad. Sci. (USA) 95: 11804, 1998), американский гидролаг (Nguyen и др., Immunogen. 54:39, 2002) или минога (Herrin и др., Ргос.Natl. Acad. Sci. (USA) 105: 2040, 2008 и Alder и др. Nat. Immunol. 9: 319, 2008). Эти антитела могут образовывать антигенсвязывающие области, используя только вариабельную область тяжелой цепи, т.е. эти функциональные антитела являются гомодимерами только тяжелых цепей (называемых «антителами тяжелой цепи») (Jespers и др., Nat. Biotechnol. 22: 1161, 2004; Cortez-Retamozo и др., Cancer Res. 64: 2853, 2004; Baral и др., Nature Med. 12: 580, 2006; и Barthelemy и др., J. Biol. Chem. 283: 3639, 2008).
Альтернативный источник связывающих доменов этого описания включает последовательности, которые кодируют случайные пептидные библиотеки или последовательности, которые кодируют сконструированное разнообразие аминокислот в шарнирных областях альтернативных не-антител каркасов, таких как scTCR (см., например, Lake и др., Int. Immunol.11: 745, 1999; Maynard и др., J. Immunol. Methods 306: 51, 2005; патент США №8,361,794), домены фибриногена (см., например, Weisel и др., Science 230: 1388, 1985), домены Kunitz (см., например, патент США №6433298), сконструированные повторные анкириновые белки (DARP) (Binz и др., J. Mol. Biol. 332: 489, 2003 и Binz и др., Nat. Biotechnol. 22: 575, 2004), фибронектин-связывающие домены (аднектины или монотела) (Richards и др., J. Mol. Biol. 326: 1475, 2003; Parker и др., Protein Eng. Des. Selec. 18: 435, 2005 и Hackel и др. (2008) J. Mol. Biol. 381: 1238-1252), цистеин-узелковые минипротеины (Vita и др. (1995), Proc. Natl. Acad. Sci. (USA) 92: 6404- 6408; Martin и др. (2002) Nat. Biotechnol. 21:71, 2002 и Huang и другие. (2005) Structure 13: 755, 2005), тетратрикопептидные повторные домены (Main и др., Structure 11: 497, 2003 и Cortajarena и др., ACS Chem. Biol. 3: 161, 2008), богатые лейцином повторные домены (Stumpp и др., J. Mol. Biol. 332: 471, 2003), липокалиновые домены (см., например, WO 2006/095164, Beste и др., Proc. Natl. Acad. Sci. (USA) 96: 1898, 1999 и Schonfeld и др., Proc. Natl. Acad. Sci. (USA) 106: 8198, 2009), V-подобные домены (см., например, публикацию заявки на патент США №2007/0065431), лектиновые домены типа С (Zelensky and Gready, FEBS J. 272: 6179, 2005; Beavil и др., Proc. Natl. Acad. Sci. (США) 89: 753, 1992 и Sato и др., Proc. Nat. 'L. Acad. Sci. (USA) 100: 7779, 2003), mAb2 или Fcab™ (см., например, публикацию патентной заявки РСТ №WO 2007/098934; WO 2006/072620), повторяющиеся белки броненосца (см., например, Madhurantakam и др., Protein Sci. 21: 1015, 2012; публикация патентной заявки РСТ №WO 2009/040338), аффилин (Ebersbach и др., J. Mol. Biol. 372: 172, 2007), аффитело, авимеры, ноттинс, финомеры, атримеры, цитотоксический белок-4, ассоциированный с Т-лимфоцитами (Weidle и др., Cancer Gen. Proteo. 10: 155, 2013) или тому подобное (Nord и др., Protein Eng. 8: 601, 1995; Nord и др., Nat. Biotechnol. 15: 772, 1997; Nord и др., Euro. J. Biochem. 268: 4269, 2001; Binz и др., Nat. Biotechnol. 23: 1257, 2005; Boersma and Pluckthun, Curr. Opin. Biotechnol. 22: 849, 2011).
Связывающие домены по данному изобретению могут быть получены, как описано здесь, или различными способами, известными в данной области (см., например, патенты США №№6291161 и 6291158). Например, связывающие домены по данному изобретению могут быть идентифицированы путем скрининга библиотеки Fab-фагов на наличие фрагментов Fab, которые специфично связываются с интересующей мишенью (например, ВСМА, компонентом комплекса γ-секретазы, таким как пресенилин или никастрин) (см. Hoet и др., Nat. Biotechnol. 23: 344, 2005). Кроме того, традиционные стратегии развития гибридомы с использованием представляющей интерес мишени (например, ВСМА, компонента комплекса γ-секретазы, такого как презенилин или никастрин) в качестве иммуногена в удобных системах (например, мыши, (HuMAb-мышь®, ТС-мышь™, KM-мышь®), ламы, курица, крысы, хомяки, кролики и т.д.) могут быть использованы для разработки связывающих доменов по данному изобретению.
В некоторых вариантах осуществления связывающий домен представляет собой одноцепочечный фрагмент Fv (scFv), который содержит области VH и VL, специфичные для интересующей мишени (например, ВСМА, компонента комплекса γ-секретазы, такой как пресенилин или никастрин). В определенных вариантах осуществления области VH и VL являются человеческими. Типичные области VH и VL включают сегменты анти-ВСМА-специфичных антител J22.0-xi, J22.9-xi, J6M0, J6M1, J6M2, J9M0, J9M1, J9M2, 11D5-3, СА8, A7D12.2, С11 D5.3, С12А3.2, C13F12.1, 13С2, 17А5, 83А10, 13А4, 13D2, 14 В11, 14Е1, 29В11, 29F3, 13А7, СА7, SGI, S307118G03, S332121F02, S332126E04, S322110D07, S336105A07, S335115G01, S335122F05, ЕТ140-3, ЕТ140-24, ЕТ140-37, ЕТ140-40, ЕТ140-54, TBL-CLN1, С4.Е2.1, Vicky-1, pSCHLI333, pSCHLI372 или pSCHLI373. Другие типичные области VH и VL включают сегменты антител против никастрина или их антигенсвязывающие фрагменты из scFvG9, А5226А, 2Н6 или 10С11.
В некоторых вариантах осуществления связывающий домен содержит или представляет собой последовательность, которая составляет по меньшей мере на 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, при по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99%, по меньшей мере 99,5% или 100% идентична аминокислотной последовательности вариабельной области легкой цепи (VL) (например, из анти-ВСМА J22.0-xi, J22.9-xi, J6M0, J6M1, J6M2, J9M0, J9M1, J9M2, 11D5-3, СА8, A7D12.2, C11D5.3, С12А3.2 или C13F12.1, 13С2, 17А5, 83А10, 13А4, 13D2, 14 В11, 14Е1, 29В11, 29F3, 13А7, СА7, S307118G03, S332121F02, S332126E04, S322110D07, S336105A07, S335115G01, S335122F05, ЕТ140-3, ЕТ140-24, ЕТ40-37, ЕТ140-40, ЕТ140-54, TBL-CLN1, С4.Е2.1, Vicky-1, pSCHLI333, pSCHLI372 или pSCHLI373 или из анти-никастрина scFvG9, А5226А, 2Н6 или 10С11) или в вариабельной области тяжелой цепи (VH) (например, из анти-ВСМА J22.0-xi, J22.9-xi, J6M0, J6M1, J6M2, J9M0, J9M1, J9M2, 11D5-3, СА8, A7D12.2, C11D5.3, С12А3.2, C13F12.1, 13С2, 17А5, 83А10, 13А4, 13D2, 14 В11, 14Е1, 29В11, 29F3, 13А7, СА7, SGI, S307118G03, S332121F02, S332126E04, S322110D07, S336105A07, S335115G01, S335122F05, ЕТ140-3, ЕТ140-24, ЕТ140-37, ЕТ140-40, ЕТ140-54, TBL-CLN1, C4.E2.1, Vicky-1, pSCHLI333, pSCHLI372 или pSCHLI373; или анти-никастрин scFvG9, A5226A, 2H6 или 10С11), или обеих, где каждый CDR не содержит изменений или самое большее одно, два или три изменения от моноклонального антитела или его фрагмента или производного, которое специфично связывается с представляющей интерес мишенью (например, ВСМА, компонент комплекса γ-секретазы, такой как пресенилин или никастрин).
В некоторых вариантах осуществления область VH связывающего домена согласно настоящему изобретению может быть получена из или основана на VH известного моноклонального антитела и содержит одну или несколько (например, 2, 3, 4, 5, 6, 7, 8, 9, 10) вставки, одну или несколько (например, 2, 3, 4, 5, 6, 7, 8, 9, 10) делеций, одну или несколько (например, 2, 3, 4, 5, 6, 7, 8, 9, 10) аминокислотных замен (например, консервативные аминокислотные замены или неконсервативные аминокислотные замены) или комбинацию вышеуказанных изменений по сравнению с VH известного моноклонального антитела. Вставка, делеция или замена могут быть где угодно в области VH, в том числе на амино- или карбокси-конце или на обоих концах этой области, при условии, что каждый CDR не содержит изменений или самое большее одно, два или три изменения, и при условии, что связывающий домен, содержащий модифицированную область VH, все еще может специфично связывать свою мишень со сродством, сходным с связывающим доменом дикого типа.
В дополнительных вариантах осуществления область VL в связывающем домене согласно настоящему изобретению получена из или основана на VL известного моноклонального антитела и содержит одну или несколько (например, 2, 3, 4, 5, 6, 7, 8, 9, 10) вставки, одну или несколько (например, 2, 3, 4, 5, 6, 7, 8, 9, 10) делеций, одну или несколько (например, 2, 3, 4, 5, 6, 7, 8, 9, 10) аминокислотных замен (например, консервативные аминокислотные замены) или комбинацию вышеуказанных изменений по сравнению с VL известного моноклонального антитела. Вставка, делеция или замена могут быть где угодно в области VL, в том числе на амино- или карбокси-конце или на обоих концах этой области, при условии, что каждый CDR не содержит изменений или самое большее одно, два или три изменения, и при условии, что связывающий домен, содержащий модифицированную область VL, все еще может специфично связывать свою мишень со сродством, сходным с связывающим доменом дикого типа.
Домены VH и VL могут быть расположены в любой ориентации (то есть от амино-конца до карбокси-конца, VH - VL или VL - VH) и могут быть соединены аминокислотной последовательностью (например, имеющей длину от около пяти до около 35 аминокислот), способной обеспечивать функцию спейсера, так что два субсвязывающих домена могут взаимодействовать с образованием функционального связывающего домена. В некоторых вариантах осуществления линкер вариабельной области, который присоединяется к доменам VH и VL, включает в себя домены, принадлежащие к семейству (GlynSer), такие как (Gly3Ser)n(Gly4Ser)1 (SEQ ID NO:3), (Gly3Ser)1(Gly4Ser)n (SEQ ID NO:10), (Gly3Ser)n(Gly4Ser)n (SEQ ID NO:4), или (Gly4Ser)n (SEQ ID NO: 5), где n является целым числом от 1 до 5. В некоторых вариантах осуществления линкер представляет собой (Gly-Gly-Gly-Gly-Ser)3 (SEQ ID NO.: 12) или (Gly-Gly-Gly-Ser)4 (SEQ ID NO.: 13). В некоторых вариантах осуществления эти линкеры на основе (GlynSer) используются для связывания доменов VH и VL в связывающем домене, и эти линкеры также могут использоваться для связывания связывающего домена связывания с меткой-кассетой или для связывания метки-кассеты с гидрофобной частью или внутриклеточным компонентом.
В некоторых вариантах осуществления связывающий домен представляет собой одноцепочечный Т-клеточный рецептор (scTCR), содержащий цепи Vα/β и Сα/β (например, Vα-Cα, Vβ-Cβ, Vα-Vβ) или содержащий Vα-Cα, Vβ-Cβ, Vα-Vβ пару, специфичные для интересующей мишени (например, комплекс пептид-МНС).
В некоторых вариантах осуществления связывающий домен содержит или представляет собой последовательность, которая составляет, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, при по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99%, по меньшей мере на 99,5% или на 100% идентична аминокислотной последовательности TCR Vα, Vβ, Cα, или Сβ, где каждый CDR не содержит изменений или не более одного, двух или трех изменений относительно TCR или его фрагмента или производного, которое специфично связывается с интересующей мишенью (например, ВСМА, компонент комплекса γ-секретазы, такой как пресенилин или никастрин).
В некоторых вариантах осуществления связывающий домен Vα, Vβ, Cα или Сβ согласно настоящему изобретению может быть получен из или основа на Vα, Vβ, Cα или Сβ известного TCR (например, TCR с высоким сродством) и содержит одну или несколько (например, 2, 3, 4, 5, 6, 7, 8, 9, 10) вставок, одну или несколько (например, 2, 3, 4, 5, 6, 7, 8, 9, 10) делеций, одну или несколько (например, 2, 3, 4, 5, 6, 7, 8, 9, 10) аминокислотных замен (например, консервативных аминокислотных замен или неконсервативных аминокислотных замен) или комбинации отмеченных выше изменений по сравнению с Vα, Vβ, Cα или Сβ известного TCR. Вставка, делеция или замена могут присутствовать где угодно в области Vα, Vβ, Cα или Сβ, в том числе на амино- или карбокси-конце или на обоих концах этих областей, при условии, что каждый CDR содержит ноль изменений или самое большее одно, два или три изменения и при условии, что связывающий домен, содержащий модифицированную область Vα, Vβ, Cα или Сβ, все еще может специфично связывать свою мишень со сродством, сходным с доменом дикого типа.
ВСМА, усекретаза или оба могут быть обнаружены в интересующей клетке («клетке-мишени») или связаны с ней. Типичные клетки-мишени включают раковую клетку, клетку, связанную с аутоиммунным заболеванием или расстройством или с воспалительным заболеванием или расстройством, и инфекционный организм или клетку (например, бактерии, вирус, инфицированные вирусом клетки).
Цитотоксические конъюгаты
Конъюгаты антитело-лекарственное средство (ADC) используют для селективной доставки цитотоксического элемента к клетке-мишени, например, к опухолевой или раковой клетке. Конъюгаты антитело-лекарственное средство и связанные с ними методики и химические составы описаны, например, в Nasiri и др., J. Cell. Physiol. (2018), Hedrich и др., Clin. Pharmacokinet. (2017), Drake and Rabuka, BioDrugs 31 (6): 521 (2017), Meyer и др., Bioconj. Химреагент 27 (12): 2791 (2016), Moek и др., J. Nucl. Med. 58: 83S (2017), Nareshkumar и др., Pharm. Местожительство 32: 3526 (2015), Parslow и др., Biomedicines 4:14 (2016) и Green и др., Blood 131:611 (2018), из которых форматы антител, цитотоксические полезные нагрузки, химические соединения линкера и конъюгации, схемы дозирования, способы лечения, фармакокинетика и принципы построения АЦП включены в настоящее описание в качестве ссылки во всей их полноте.
В некоторых вариантах осуществления способов, представленных в настоящем документе, ВСМА-специфичное антитело или его антигенсвязывающая часть, химерный антигенный рецептор (CAR) или меченая молекула химерного антигенного рецептора (Т-ChARM) конъюгированы или иным образом связаны с цитотоксическим агентом, таким как в качестве химиотерапевтического агента. Химиотерапевтический агент включает, но не ограничивается ими, ингибитор функции хроматина, ингибитор топоизомеразы, лекарственное средство, ингибирующее микротрубочки, агент, повреждающий ДНК, антиметаболит (такой как антагонисты фолата, аналоги пиримидина, аналоги пурина и аналоги, модифицированные сахаром), ингибитор синтеза ДНК, интерактивный агент ДНК (такой как интеркалирующий агент) и ингибитор репарации ДНК. Иллюстративные химиотерапевтические агенты включают, без ограничения, следующие группы: антиметаболитные/противораковые агенты, такие как аналоги пиримидина (5-фторурацил, флоксуридин, капецитабин, гемцитабин и цитарабин) и аналоги пурина, антагонисты фолата и родственные ингибиторы (меркаптопурин, тиогуанин, пентостатин и 2-хлордезоксиаденозин (кладрибин)), антипролиферативные/антимитотические агенты, включающие природные продукты, такие как алкалоиды барвинка (винбластины, винкристины, и винорелбина), разрушители микротрубочек, такие как таксан (паклитаксла, доцетаксла), винкристины, винбластины, нокодазол, эпотилоны и навелбин, эпидиподофиллотоксины (этопозид, тенипозид), ДНК-повреждающие агенты (актиномицин, амсакрин, антрациклины, блеомицин, бусульфан, камптотецин, карбоплатин, хлорамбуцил, цисплатин, циклофосфамид, цитоксан, дактиномицин, даунорубицин, доксорубицин, эпирубицин, гексаметилмеламинеоксалиплатин, ифосфамид, мелфалан, мерхлорехтамин, митомицин, митоксантрон, нитрозомочевина, пликамицин, прокарбазин, таксол, таксотер, темозоламид, тенипозид, триэтилентиофосфорамид и этопозид (VP 16)); антибиотики, такие как дактиномицин (актиномицин D), даунорубицин, доксорубицин (адриамицин), идарубицин, антрациклины, митоксантрон, блеомицины, пликамицин (митрамицин) и митомицин; ферменты (L-аспарагиназа, которая системно метаболизирует L-аспарагин и элиминирует клетки, которые не способны синтезировать свой собственный аспарагин); антиагреганты; антипролиферативные/антимитотические алкилирующие агенты, такие как азотистые иприты (мехлорэтамин, циклофосфамид и аналоги, мелфалан, хлорамбуцил), этиленимины и метилмеламины (гексаметилмеламин и тиотепа), алкилсульфонаты -бусульфан, нитрозомочевина (аналог кармустозина, уксусная кислота) (аналоги кармустозина) (ВС) и др. (DTIC); антипролиферативные/антимитотические антиметаболиты, такие как аналоги фолиевой кислоты (метотрексат); координационные комплексы платины (цисплатин, карбоплатин), прокарбазин, гидроксимочевина, митотан, аминоглутетимид; гормоны, аналоги гормонов (эстроген, тамоксифен, госерелин, бикалутамид, нилутамид) и ингибиторы ароматазы (летрозол, анастрозол); антикоагулянты (гепарин, синтетические соли гепарина и другие ингибиторы тромбина); фибринолитические агенты (такие как тканевый активатор плазминогена, стрептокиназа и урокиназа), аспирин, дипиридамол, тиклопидин, клопидогрел, абциксимаб; антимиграционные агенты; антисекреторные средства (бревелдин); иммунодепрессанты (циклоспорин, такролимус (FK-506), сиролимус (рапамицин), азатиоприн, микофенолат мофетил); антиангиогенные соединения (TNP470, генистеин) и ингибиторы фактора роста (ингибиторы фактора роста эндотелия сосудов (VEGF), ингибиторы фактора роста фибробластов (FGF)); блокатор ангиотензиновых рецепторов; доноры оксида азота; антисмысловые олигонуклеотиды; антитела (трастузумаб, ритуксимаб); химерных антигенных рецепторов; ингибиторы клеточного цикла и индукторы дифференцировки (третиноин); ингибиторы mTOR (мишень рапамицина в клетках), ингибиторы топоизомеразы (доксорубицин (адриамицин), амсакрин, камптотецин, даунорубицин, дактиномицин, eniposide, эпирубицин, этопозид, идарубицин, иринотеканно (СРТ-11) и митоксантрон, топотеканны, иринотеканен), кортикостероиды (кортизон, дексаметазон, гидрокортизон, метилпреднизолон, преднизон и преднизолон); ингибиторы киназы сигнальной трансдукции фактора роста; индукторы митохондриальной дисфункции, токсины, такие как токсин холеры, рицин, экзотоксин Pseudomonas, токсин аденилатциклазы Bordetella pertussis или токсин дифтерии и активаторы каспазы; и хроматиновые разрушители.
Типичными ADC, который специфично связывается с ВСМА, является J6M0-mcMMAF (GSK2857916), описанный в Tai и др., Blood 123 (20): 3128-3138 (2014), этот ADC и способы его использования включены в настоящее описание посредством ссылки во всей их полноте. Другой ВСМА-специфичный ADC (SG1-MMAF) был описан Ryan и др. (Mol. Cancer Ther., 6 (11): 3009-3018, 2007), он включен в настоящий документ в качестве ссылки.
Клетки-хозяева и нуклеиновые кислоты
В определенных аспектах настоящее изобретение относится к молекулам нуклеиновой кислоты, которые кодируют любой один или более ВСМА-специфичных белков (включая мультиспецифичные и биспецифичные связывающие белки, содержащие, по меньшей мере, один домен связывания ВСМА), или ингибиторы γ-секретазы, описанные в данном документе. Такие молекулы нуклеиновой кислоты могут быть вставлены в соответствующий вектор (например, вирусный вектор или невирусный плазмидный вектор) для введения в интересующую клетку-хозяина (например, Т-клетку).
Используемый в данном документе термин «рекомбинантный» или «неприродный» относится к организму, микроорганизму, клетке, молекуле нуклеиновой кислоты или вектору, который включает в себя по меньшей мере одно генетическое изменение или был модифицирован путем введения экзогенной молекулы нуклеиновой кислоты, где такие изменения или модификации вносятся генной инженерией. Генетические изменения включают, например, модификации, вводящие экспрессируемые молекулы нуклеиновых кислот, кодирующие белки, слитые белки или ферменты, или другие добавления, делеции, замены молекул нуклеиновых кислот или другие функциональные нарушения генетического материала клетки. Дополнительные модификации включают, например, некодирующие регуляторные области, в которых модификации изменяют экспрессию гена или оперона. В некоторых вариантах осуществления клетка, такая как Т-клетка, полученная от субъекта, может быть превращена в неприродную или рекомбинантную клетку (например, неприродную или рекомбинантную Т-клетку) путем введения нуклеиновой кислоты, которая кодирует ВСМА- специфичный связывающий белок или ингибитор γ-секретазы согласно настоящему изобретению (например, ВСМА-специфичный T-ChARM или CAR; или анти-γ-секретаза), как описано в настоящем документе, и посредством этого клетка экспрессирует локализованный на клеточной поверхности ВСМА-специфичный связывающий белок.
Вектор, который кодирует ядро вируса, упоминается здесь как «вирусный вектор». Существует большое количество доступных вирусных векторов, подходящих для использования с композициями согласно настоящему изобретению, включая идентифицированные для применения в генной терапии человека (см. Pfeifer and Verma, Ann. Rev. Genomics Hum. Genet. 2: 177, 2001). Подходящие вирусные векторы включают векторы на основе РНК-вирусов, такие как векторы, полученные из ретровируса, например векторы, полученные из вируса мышиного лейкоза Молони (MLV), и включают более сложные векторы, полученные из ретровируса, например векторы, полученные из лентивируса. Векторы, полученные из ВИЧ-1, относятся к этой категории. Другие примеры включают лентивирусные векторы, полученные из ВИЧ-2, FIV, вирус инфекционной анемии лошадей, SIV и вирус Maedi-Visna (лентивирус овец). Способы использования ретровирусных и лентивирусных вирусных векторов и упаковочных клеток для трансформации клеток-хозяев млекопитающих вирусными частицами, содержащими трансгены химерного антигенного рецептора, известны в данной области техники и были ранее описаны, например, в патенте США 8,119,772; Walchli и др., PLoS One 6:327930, 2011; Zhao и др., J. Immunol. 174:4415, 2005; Engels и др., Hum. Gene Ther. 14:1155, 2003; Frecha и др., Mol. Ther. 18:1748, 2010; Verhoeyen и др., Methods Mol. Biol. 506:97, 2009. Ретровирусные и лентивирусные векторные конструкции и системы экспрессии также коммерчески доступны.
В некоторых вариантах осуществления вирусный вектор используется для введения неэндогенного полинуклеотида, кодирующего ВСМА-специфичный связывающий белок или ингибитор γ-секретазы. Вирусный вектор может представлять собой ретровирусный вектор или лентивирусный вектор. Вирусный вектор также может включать полинуклеотиды, кодирующие маркер для трансдукции. Маркеры трансдукции для вирусных векторов известны в данной области и включают маркеры селекции, которые могут придавать лекарственную устойчивость, или обнаруживаемые маркеры, такие как флуоресцентные маркеры или белки клеточной поверхности, которые можно обнаружить такими методами, как проточная цитометрия. В конкретных вариантах осуществления вирусный вектор дополнительно содержит маркер гена для трансдукции, содержащий зеленый флуоресцентный белок, внеклеточный домен человеческого CD2 или усеченный EGFR человека (huEGFRt; см. Wang и др., Blood 118: 1255, 2011). Когда геном вирусного вектора содержит множество последовательностей нуклеиновых кислот, которые должны экспрессироваться в клетке-хозяине в виде отдельных транскриптов, вирусный вектор может также содержать дополнительные последовательности между двумя (или более) транскриптами, обеспечивающими бицистронную или мультицистронную экспрессию. Примеры таких последовательностей, используемых в вирусных векторах, включают сайты проникновения внутренней рибосомы (IRES), сайты расщепления фурином, пептид 2А вируса (например, Р2А, Т2А, Е2А, F2A) или любую их комбинацию.
Другие векторы также могут быть использованы для доставки полинуклеотидов, включая вирусные векторы ДНК, включая, например, векторы на основе аденовируса и векторы на основе аденоассоциированного вируса (AAV); векторы, полученные из вирусов простого герпеса (HSV), включая векторы ампликонов, дефектные по репликации HSV и ослабленные HSV (Krisky и др., Gene Ther. 5: 1517, 1998).
Другие векторы, недавно разработанные для применения в генной терапии, также могут быть использованы с композициями и способами согласно настоящему изобретению. Такие векторы включают векторы, полученные из бакуловирусов и α-вирусов. (Jolly, D.J. 1999. Emerging Viral Vectors. P. 209-40, Friedmann T. ed. Development of Human Gene Therapy. New York: Cold Spring Harbor Lab) или плазмидные векторы (такие как sleeping beauty или другие транспозоновые векторы). В некоторых вариантах осуществления вирусный или плазмидный вектор дополнительно содержит генный маркер для трансдукции (например, зеленый флуоресцентный белок, huEGFRt).
В некоторых вариантах осуществления гематопоэтические клетки-предшественники или эмбриональные стволовые клетки модифицированы для включения неэндогенного полинуклеотида, который кодирует ВСМА-специфичный белок (например, ВСМА-специфичный T-ChARM или CAR) согласно настоящему изобретению. Гематопоэтические клетки-предшественники могут включать клетки-предшественники тимоцитов или индуцированные плюрипотентные стволовые клетки, которые могут происходить или получаться из ткани печени плода, костного мозга, пуповинной крови или периферической крови. Кроветворные клетки-предшественники могут быть от человека, мыши, крысы или других млекопитающих. В конкретных вариантах осуществления используют клетки-предшественники тимоцитов CD24lo Lin- CD117+.
В определенных вариантах осуществления условия культивирования влекут за собой культивирование гемопоэтических клеток-предшественников, экспрессирующих слитые белки этого раскрытия, в течение времени, достаточного для индукции пролиферации или дифференцировки. Клетки поддерживают в культуре, как правило, в течение от приблизительно 3 дней до приблизительно 5 дней, или от приблизительно 4 до приблизительно 10 дней, или от приблизительно 5 до приблизительно 20 дней. Понятно, что клетки могут поддерживаться в течение соответствующего периода времени, необходимого для достижения желаемого результата, то есть желаемой клеточной композиции или уровня пролиферации. Например, для создания клеточной композиции, содержащей в основном незрелые и инактивированные Т-клетки, клетки могут поддерживаться в культуре в течение от приблизительно 5 до приблизительно 20 дней. Клетки могут поддерживаться в культуре в течение от приблизительно 20 до приблизительно 30 дней для генерирования клеточной композиции, содержащей в основном зрелые Т-клетки. Неприлипающие клетки также могут быть собраны из культуры в различные моменты времени, например, от приблизительно нескольких дней до приблизительно 25 дней. В некоторых вариантах осуществления гемопоэтические стволовые клетки совместно культивируют на линиях стромальных клеток (патент США №7575925; Schmitt и др., Nat. Immunol. 5: 410, 2004; Schmitt и др., Immunity 17: 749, 2002).
Один или несколько цитокинов, которые способствуют фиксации или дифференцировке гемопоэтических клеток-предшественников, могут быть добавлены в культуру. Цитокины могут быть человеческими или нечеловеческими. Репрезентативные примеры цитокинов, которые можно использовать, включают всех членов семейства FGF, включая FGF-4 и FGF-2; Flt-3-лиганд, фактор стволовых клеток (SCF), тромбопоэтин (ТРО) и IL-7. Цитокины могут быть использованы в комбинации с гликозаминогликаном, таким как сульфат гепарина.
В некоторых вариантах осуществления клетки-хозяева, способные экспрессировать ВСМА-специфичный белок, описанный в данном описании, на поверхности клетки, представляют собой Т-клетки, включая первичные клетки или клеточные линии, полученные от человека, мыши, крысы или других млекопитающих. При получении от млекопитающего Т-клетка может быть получена из многочисленных источников, включая кровь, костный мозг, лимфатический узел, тимус или другие ткани или жидкости. Т-клетка или ее субпопуляции (например, наивная, центральная память, эффекторная память) могут быть обогащены или очищены. Т-клеточные линии хорошо известны в данной области, некоторые из которых описаны в Sandberg и др., Leukemia 21: 230, 2000. В некоторых вариантах осуществления используются Т-клетки, в которых отсутствует эндогенная экспрессия TCRα и β цепей. Такие Т-клетки могут, естественно, не иметь эндогенной экспрессии TCRα и β-цепей или могут быть модифицированы для блокирования экспрессии (например, Т-клетки от трансгенной мыши, которая не экспрессирует TCR α и β-цепи, или клетки, которыми манипулировали для ингибирования экспрессии TCR α и β цепей) или для нокаута цепи TCRα, цепи TCRP или обоих генов. В некоторых вариантах осуществления клетки, способные экспрессировать ВСМА-специфичный белок, описанный в данном описании, на клеточной поверхности, не являются Т-клетками или клетками линии Т-клеток, но клетками, которые являются клетками-предшественниками, стволовыми клетками или клетками, которые были модифицированы для экспрессии анти-CD3 клеточной поверхности.
В любом из представленных здесь вариантов осуществления клетка-хозяин может представлять собой «универсальную донорскую» клетку, которая модифицирована для уменьшения или устранения экспрессии одного или нескольких эндогенных генов, участвующих в иммунном ответе. Например, Т-клетка может быть модифицирована для уменьшения или устранения экспрессии одного или нескольких полипептидов, выбранных из PD-1, LAG-3, CTLA4, TIGIT, TIM3, комплексного компонента HLA или комплексного компонента TCR или TCR. Не желая быть связанными теорией, некоторые эндогенно экспрессируемые белки иммунных клеток могут быть распознаны как чужеродные аллогенным хозяином, который получает модифицированные иммунные клетки, что может привести к элиминации модифицированных иммунных клеток (например, аллеля HLA), или может подавлять иммунную активность модифицированных иммунных клеток (например, PD-1, LAG-3, CTLA4, TIGIT) или могут влиять на активность связывания гетерологически экспрессируемого связывающего белка согласно настоящему изобретению (например, эндогенного TCR, который связывает антиген, не ассоциированный с опухолью, и взаимодействует с антигенспецифичным рецептором модифицированной иммунной клетки, специфично связывающейся с антигеном, ассоциированным с опухолью). Соответственно, уменьшение или устранение экспрессии или активности таких эндогенных генов или белков может улучшить активность, толерантность и персистенцию модифицированных иммунных клеток в условиях аллогенного хозяина и может позволить универсальное введение клеток (например, любому реципиенту независимо от типа HLA).
В определенных вариантах осуществления клетка-хозяин (например, модифицированная иммунная клетка) согласно настоящему изобретению содержит хромосомный ген-нокаут одного или нескольких генов, кодирующих PD-1, LAG-3, CTLA4, TIM3, TIGIT, компонент комплекса HLA (например, ген, который кодирует α1 макроглобулин, α2 макроглобулин, α3 макроглобулин, β1 микроглобулин или β2 микроглобулин), компонент TCR (например, ген, который кодирует вариабельную область TCR или константную область TCR) (см., например, Torikai и др., Nature Sci. Rep.6: 21757 (2016); Torikai и др., Blood 119 (24): 5697 (2012) и Torikai и др., Blood 122 (8): 1341 (2013), методы редактирования генов, композиции и адаптивные клеточные терапии, которых полностью включены в настоящее описание посредством ссылки). Например, в некоторых вариантах осуществления нокаут хромосомного гена получают с использованием системы CRISPR/Cas9 и могут включать трансфекцию модифицированной иммунной клетки лентивирусом (например, pLentiCRISPRv2; Torikai и др., Blood (2016)), экспрессирующим систему CRISPR/Cas9, таргетированную на PD-1, LAG-3, CTLA4, компонент HLA или компонент TCR, или любую их комбинацию. Праймеры, полезные для конструирования лентивируса, который экспрессирует систему CRISPR/Cas9 для ингибирования эндогенно экспрессируемого белка иммунных клеток, включают, например, пары праймеров, содержащие прямой и обратный праймеры, имеющие нуклеотидные последовательности, представленные в SEQ ID NO: 22 и 23, 24 и 25. 26 и 27 и 28 и 29.
В некоторых вариантах осуществления Т-клетка-хозяин трансфицирована для экспрессии ВСМА-специфичного связывающего белка (например, T-ChARM, CAR, мультиспецифичных связывающих белков, содержащих по меньшей мере один ВСМА-связывающий домен, биспецифичных связывающих белков, содержащих по меньшей мере один домен ВСМА-связывающий) этого описания представляет собой функциональную Т-клетку, такую как вирус-специфичная Т-клетка, опухоле-антиген-специфичная цитотоксическая Т-клетка, наивная Т-клетка, Т-клетка памяти, центральная Т-клетка памяти или эффекторная Т-клетка памяти или CD4+ CD25+ регуляторная Т-клетка.
В культуру может быть добавлен один или несколько цитокинов фактора роста, которые способствуют пролиферации Т-клеток, экспрессирующих ВСМА-специфичный белок, описанный в данном описании. Цитокины могут быть человеческими или нечеловеческими. Типичные цитокины фактора роста, которые могут использоваться, способствуют пролиферации Т-клеток, включают IL-2, IL-15, IL-21 или тому подобное.
Применение
Заболевания, которые можно лечить с помощью ВСМА-таргетированной иммунотерапии или клеток, экспрессирующих ВСМА-специфичный связывающий белок, в сочетании с ингибитором γ-секретазы (GSI), как описано в настоящем описании, включают рак (например, рак, который экспрессирует ВСМА), иммунные заболевания (например, аутоиммунные) или связанные со старением заболевания (например, старение). Адаптивная иммунная и генная терапия являются перспективными методами лечения различных видов рака (Morgan и др., Science 314: 126, 2006; Schmitt и др., Hum. Gene Ther. 20: 1240, 2009; июнь, J. Clin. Invest. 117: 1466, 2007) и инфекционных заболеваний (Kitchen и др., PLoS One 4: 38208, 2009; Rossi и др., Nat. Biotechnol. 25: 1444, 2007; Zhang и др., PLoS Pathog. 6: el001018, 2010; Luo и др., J. Mol. Med. 89: 903, 2011).
Виды рака, которые поддаются воздействию композиций и способов, описанных в данном документе, представляют собой те, которые экспрессируют или способны экспрессировать ВСМА на своей клеточной поверхности. Типичные виды рака, которые можно лечить, включают миеломы (такие как множественная миелома), лейкозы (такие как лейкоз плазматических клеток), лимфомы (такие как лимфоплазмоцитарная лимфома), плазмоцитомы, макроглобулинемию Вальденстрема. Другие виды рака, которые могут экспрессировать ВСМА, включают аденокарциному молочной железы и бронхогенную карциному легкого. В некоторых вариантах осуществления пролиферативные расстройства, поддающиеся комбинированной терапии ВСМА-специфичного связывающего белка и GSI, представляют собой определенные типы В-клеточного рака, включая болезни плазматических клеток (такие как, например, множественная миелома). Воспалительные и аутоиммунные заболевания включают артрит, ревматоидный артрит, ювенильный ревматоидный артрит, остеоартрит, полихондрит, псориатический артрит, псориаз, дерматит, полимиозит/дерматомиозит, миозит с включениями телец, воспалительных миозит, токсический эпидермальный некролиз, системная склеродермия и склероз, синдром CREST, воспалительное заболевание кишечника, болезнь Крона, язвенный колит, респираторный дистресс-синдром, респираторный дистресс-синдром взрослых (ARDS), менингит, энцефалит, увеит, колит, гломерулонефрит, аллергические состояния, экзему, астму, состояния, включающие инфильтрацию Т-клеток и хронические воспалительные реакции, атеросклероз, аутоиммунный миокардит, дефицит адгезии лейкоцитов, системную красную волчанку (СКВ), подострую кожную красную волчанку, дискоидную волчанку, миелит при СКВ, энцефалит при СКВ, ювенильный диабет, рассеянный склероз, аллергический энцефаломиелит, оптикомиелит, острую ревматическую лихорадку, хорею Сиденгама, реакции, связанные с острой и отсроченной гиперчувствительностью, опосредованной цитокинами и Т-лимфоцитами, туберкулез, саркоидоз, гранулематоз, включая гранулематоз Вегенера и синдром Чарга-Стросса, агранулоцитоз, васкулит (включая аллергический васкулит/ангиит, ANCA-ассоциированные иревматоидные васкулиты), апластичную анемию, анемию Даймонда-Блекфена, анемию, иммунную гемолитическую анемию, включая аутоиммунную гемолитическую анемию (AIHA), пернициозную анемию, парциальную красноклеточную аплазию костного мозга (PRCA), дефицит фактора VIII, гемофилию А, аутоиммунную нейтропению, панцитопению, лейкопению, заболевания, включающие диапедезо лейкоцитов, воспалительные расстройства центральной нервной системы (ЦНС), полиорганную недостаточность, миастению грависа, заболевания, связанные с комплексом антиген-антитело, анти-ОВМ-болезнь, синдром антифосфолипидных антител, аллергический неврит, болезнь Бехчета, синдром Каслмана, синдром Гудпасчера, миастенический синдром Ламберта-Итона, синдром Рейно, синдром Рейно, синдром Шегрена, синдром Стивенса-Джонсона, отторжение трансплантата твердого органа, реакцию трансплантат против хозяина (GVHD), буллезный пемфигоид, пузырчатку, аутоиммунные полиэндокринопатии, серонегативную спондилоартропатию, болезнь Рейтера, синдром мышечной скованности, гигантонлеточный артериит, нефрит иммунного комплекса, IgA-нефропатию, IgM-полиневропатию или IgM-опосредованную невропатию, идиопатическую тромбоцитопеническую пурпуру (ИТП), тромботическую тромбоцитопеническую пурпуру (ТТП), болезнь Шенлейн-Геноха, аутоиммунную тромбоцитопению, аутоиммунное заболевание яичка и яичника, включая аутоиммунный орхит и оофорит, первичный гипотиреоз; аутоиммунные эндокринные заболевания, включая аутоиммунный тиреоидит, хронический тиреоидит (тиреоидит Хашимото), подострый тиреоидит, идиопатический гипотиреоз, болезнь Аддисона, болезнь Грейвса, аутоиммунные полигландулярные синдромы (или синдромы полигландулярной эндокринопатии), диабет типа I, который также упоминается как инсулин-зависимый сахарный диабет (ИЗСД), и синдром Шихана; аутоиммунный гепатит, лимфоидный интерстициальный пневмонит (HIV), облитерирующий бронхиолит (без трансплантации), неспецифическую интерстициальная пневмония (NSIP), синдром Гийена-Барре, васкулиты крупных сосудов (включая ревматическую полимиалгию и гигантоклеточный артериит (синдром Такаясу)), васкулиты сосудов среднего размера (включая болезнь Кавасаки и узелковый полиартериит), узелковый полиартериит (ПАН), анкилозирующий спондиалоартрит, болезнь Бергера (IgA-нефропатия), быстро прогрессирующий гломерулонефрит, первичный билиарный цирроз печени, целиалкию спру (глютеновая энтеропатия), криоглобулинемию, криоглобулинемии, связанные с гепатитом, боковой амиотрофический склероз (БАС), ишемическую болезнь сердца, семейную средиземноморскую лихорадку, микроскопический полиангиит, синдром Когана, синдром Вискотта-Олдрича и облитерирующий тромбоангиит.
В конкретных вариантах осуществления способ лечения субъекта ВСМА-связывающим белком в комбинации с ингибитором γ-секретазы, как описано в настоящем документе, включает лечение множественной миеломы, плазмоцитомы, лейкемии плазматических клеток, макроглобулинемии Вальденстрема, В-клеточной лимфомы и лимфоплазмлактической лимфомы.
ВСМА-специфичный связывающий белок, такой как CAR или T-ChARM, согласно настоящему изобретению может вводиться субъекту в клеточной форме (например, генная терапия популяцией клеток-мишеней (зрелые Т-клетки (например, CD8+ или CD4+ клетки) или другие клетки линии Т-клеток)). В конкретном варианте осуществления клетки Т-клеточной линии, экспрессирующие ВСМА-специфичный связывающий белок (например, ВСМА-специфичный CAR или T-ChARM), вводимые субъекту, представляют собой сингенные, аллогенные или аутологичные клетки.
Способ согласно настоящему изобретению включает стадии введения ВСМА-специфичной молекулы, связывающейся с поверхностью иммунных клеток (например, Т-клеток), и введения ингибитора γ-секретазы (GSI). В определенных вариантах осуществления комбинацию можно вводить одновременно, вместе в одном и том же фармацевтически приемлемом носителе или в отдельных составах (но одновременно). Параллельное введение означает, что каждый компонент вводится одновременно или в пределах 8-12 часов друг относительно друга. Введение второго компонента более чем через 12 часов после первого компонента будет считаться последовательным введением. В других вариантах осуществления ВСМА-специфичная иммунотерапия и GSI могут вводиться последовательно (например, через один, два, три, четыре, пять, шесть, семь, восемь или девять дней друг от друга; одну, две, три или четыре недели друг от друга; одну, две, три, четыре, пять, шесть, семь, восемь или девять недель друг от друга, или один, два, три, четыре, пять, шесть или более лет друг от друга, или тому подобное), в любом порядке и в любом сочетании. В конкретных вариантах осуществления при последовательном введении GSI вводят первым, а ВСМА-таргетированную иммунотерапию (растворимую или клеточную форму) вводят второй. В других вариантах осуществления ВСМА-таргетированная иммунотерапия (растворимая или клеточная форма) вводится первой, a GSI второй. В конкретных вариантах осуществления ВСМА-таргетированная иммунотерапия, включающая модифицированную иммунную клетку, которая специфично связывается с ВСМА (например, ВСМА-специфичная Т-клетка, такая как CAR-T, T-ChARM, биспецифичная, мультиспецифичная Т-клетка), вводится первой, и GSI вводят второй (например, в течение часов, дней, недель, месяцев или лет после иммунотерапии ВСМА). В дополнительных вариантах осуществления ВСМА-таргетированная иммунотерапия включает модифицированную иммунную клетку, содержащую связывающий белок, который специфично связывается с ВСМА (например, ВСМА-специфичная Т-клетка, такая как CAR-T, T-ChARM, биспецифичная, мультиспецифичная Т-клетка) (а) вводится субъекту; (b) после определенного периода времени (например, от около 5 дней до около одной недели, от около одной недели до около двух недель, от около двух недель до около трех недель, от одной недели до около одного месяца, от одного месяца до около шести месяцев от приблизительно трех месяцев до примерно одного года или дольше по мере необходимости) субъект или ткань субъекта исследуют или исследуют на предмет наличия или устойчивости ранее введенной модифицированной иммунной клетки, содержащей связывающий белок, который специфично связывается с ВСМА, и (с) введение GSI через секунду после обнаружения наличия или устойчивости иммунотерапии ВСМА.
В некоторых вариантах осуществления GSI вводят субъекту по меньшей мере один раз до, одновременно и по меньшей мере один раз (например, по меньшей мере два, три или четыре раза) после ВСМА-таргетированной иммунотерапии. В одном варианте осуществления, GSI вводят по меньшей мере 2, по меньшей мере 3, по меньшей мере 4, по меньшей мере 5, по меньшей мере 6, по меньшей мере 7, по меньшей мере 8, по меньшей мере 9, по меньшей мере 10, по меньшей мере 15, по меньшей мере 20, по меньшей мере 25, по меньшей мере 30, по меньшей мере 35, по меньшей мере 40, по меньшей мере 45 или по меньшей мере 50 раз после первого введения Т-клетки, которая экспрессирует ВСМА-специфичный химерный антигенный рецептор (CAR). В конкретных вариантах осуществления комбинированная терапия согласно настоящему изобретению включает ВСМА-таргетированную иммунотерапию и введение от около 0,01 мкМ GSI до около 5 мкМ GSI, от около 0,03 мкМ GSI до около 1,5 мкМ GSI, от около 0,05 мкМ GSI до около 2,5 мкМ GSI или от приблизительно 0,1 мкМ GSI до приблизительно 1,0 мкМ GSI. В некоторых вариантах осуществления комбинированная терапия, включающая модифицированную иммунную клетку, которая специфично связывает ВСМА, и GSI содержит меньшее количество иммунной клетки, которая специфично связывает ВСМА, GSI или обе, по сравнению с введением этих методов лечения индивидуально.
Например, в качестве фона и не желая быть связанными какой-либо теорией, во время адаптивной иммунотерапии приживление введенных клеток, экспрессирующих ВСМА-специфичный белок может быть облегчено предварительным иммуносупрессивным кондиционированием одним или несколькими химиотерапевтическими агентами или методами лечения. В любом из вариантов осуществления, описанных в настоящем документе, способ дополнительно включает предварительную подготовку субъекта одновременно или до введения ВСМА-таргетированной иммунотерапии, или иммунной клетки, экспрессирующей ВСМА-специфичный белок и GSI. В конкретных вариантах осуществления режим предварительного кондиционирования представляет собой иммуносупрессивное кондиционирование, включающее истощающие эндогенные лимфоциты (также называемые лимфодеплецией, которые могут быть не миелоаблативными или миелоаблативными). Примерная лимфодеструкция может быть достигнута с помощью одного циклофосфамида, циклофосфамида в комбинации с флударабином, путем использования других агентов или методов лечения, которые цитотоксичны для лимфоцитов (например, общего облучения организма) или любой их комбинации.
Связывающая молекула, ингибитор или комбинированные композиции могут вводиться перорально, местно, трансдермально, парентерально, посредством ингаляционного спрея, вагинально, ректально или путем внутричерепной инъекции или любой их комбинации. При отдельном назначении ВСМА-таргетированная иммунотерапия и GSI могут проводиться одним и тем же путем или разными путями. Например, в некоторых вариантах осуществления ВСМА-таргетированную иммунотерапию вводят парентерально, a GSI вводят перорально, что может быть одновременно или последовательно. Используемый здесь термин «парентеральный» включает подкожные инъекции, внутривенные, внутримышечные, внутрицистернальные инъекции или инфузии. Предполагается также введение внутривенной, внутрикожной, внутримышечной, внутримаммарной, внутрибрюшинной, интратекальной, ретробульбарной, внутрилегочной инъекции и/или хирургической имплантации в конкретном месте. Как правило, композиции по существу не содержат пирогенов, а также других примесей, которые могут быть вредными для реципиента. Инъекция или инфузия, особенно внутривенная, являются предпочтительными для введения ВСМА-таргетированной иммунотерапии.
В других вариантах осуществления ВСМА-специфичный связывающий белок, GSI или оба могут быть введены субъекту в растворимой форме (например, антитело). Растворимые TCR также известны в данной области (см., например, Molloy и др., Curr. Opin. Pharmacol. 5: 438, 2005; патент США №6759243).
Фармацевтические композиции, включающие комбинированную терапию ВСМА-таргетированной иммунотерапии и ингибитора γ-секретазы согласно настоящему изобретению, могут вводиться способом, соответствующим заболеванию или состоянию, которое нужно лечить (или предотвращать), как определено специалистами в данной области техники. Подходящая доза, подходящая продолжительность и частота введения композиций будут определяться такими факторами, как состояние пациента, размер, тип и тяжесть заболевания, конкретная форма активного ингредиента и способ введения. Настоящее изобретение относится к фармацевтическим композициям, содержащим клетки, экспрессирующие ВСМА-специфичный связывающий белок, такой как CAR или T-ChARM, и фармацевтически приемлемый носитель, разбавители или наполнитель. Подходящие наполнители включают воду, физиологический раствор, декстрозу, глицерин или тому подобное и их комбинации.
Преимущество настоящего изобретения заключается в том, что клетки, экспрессирующие ВСМА-специфичные связывающие белки, такие как CAR или Т-ChARM, вводимые пациенту, могут быть истощены с использованием партнера по родственному связыванию с меткой-кассетой. В некоторых вариантах осуществления настоящее описание обеспечивает способ истощения Т-клетки, экспрессирующей ВСМА-специфичный T-ChARM, с использованием антитела, специфичного для метки-кассеты, с использованием партнера по родственному связыванию, специфичного для метки-кассеты, или с использованием второй Т клетка, экспрессирующей CAR и обладающей специфичностью для метки-кассеты. В некоторых вариантах осуществления метка-кассета допускает иммунодефицит Т-клетки, экспрессирующей ВСМА-специфичный T-ChARM, согласно настоящему описанию. Элиминация сконструированных Т-клеток может быть выполнена с использованием агентов истощения, специфичных для метки-кассеты. Например, если используется Strep-метка, то можно использовать анти-Strep антитело, scFv анти-Strep или Streptactin, каждое из которых слито или конъюгировано с клеточно-токсическим реагентом (таким как токсин, радиометалл) или можно использовать анти-Strep-метку/анти-CD3-биспецифичный scFv или Т-клетку с анти-Strep-меткой CAR.
В другом аспекте настоящее описание относится к способу селективного стимулирования пролиферации рекомбинантной Т-клетки, экспрессирующей ВСМА-специфичный T-ChARM согласно данному описанию. В некоторых вариантах осуществления способ включает селективную пролиферацию ex vivo Т-клеток, экспрессирующих ВСМА-специфичный T-ChARM, с использованием партнера по связыванию метки, такой как антитело. В других вариантах осуществления способ включает размножение функциональных Т-клеток (например, вирус-специфичный, ТАА (опухолевого антигена) -специфических CTL или специфических подмножеств Т-клеток, таких как наивные Т-клетки, Т-клетки памяти, центральные Т-клетки памяти, или эффекторную Т-клетку памяти, CD4+ CD25+ регуляторные Т-клетки) с партнером по связыванию метки, таких как антитело, что необязательно можно сделать в присутствии партнера по связыванию костимулирующей молекулы (такой как антитело против CD27 или против CD28).
В еще других вариантах осуществления ВСМА-специфичный T-ChARM позволяет селективно стимулировать пролиферацию Т-клеток in vivo при экспрессии ВСМА-специфичного T-ChARM данного описания. В некоторых вариантах осуществления Т-клетка, экспрессирующая CAR, содержащая метку-кассету, позволяет экспансию Т-клеток CAR in vivo при контакте с клетками, экспрессирующими лиганд (например, включая лиганды клеток-супрессоров Т-клеток PD-L1, PD-L2). Такие размноженные Т-клетки полезны в описанных здесь способах лечения заболеваний. В некоторых вариантах осуществления пролиферация или размножение клеток, экспрессирующих ВСМА-специфичный T-ChARM, как описано в настоящем документе, индуцируется in vivo, что может быть индуцировано партнером по связыванию с меткой-кассетой (такой как антитело-антитело) и необязательно партнером по связыванию с костимулирующей молекулой (например, антитело против CD27 или против CD28).
В некоторых дополнительных вариантах осуществления клетки, экспрессирующие ВСМА- специфичный T-ChARM, как описано в настоящем документе, активируются in vivo, например, в месте опухоли. Например, композиция (например, альгинат, матрица базальной мембраны (Matrigel®), биополимер или другая матрица) или носитель (например, микрошарик, наночастица или другая твердая поверхность), содержащая партнера по связыванию метки-зкассеты (такого как анти-кассетное антитело) и партнер, связывающий костимулирующую молекулу (такой как анти-CD27 или анти-CD28-антитело), могут быть использованы для локальной активации в месте опухоли (например, солидной опухоли) Т-клетки, экспрессирующей ВСМА-специфичный T-ChARM, как описано здесь.
В некоторых вариантах осуществления рекомбинантные клетки, экспрессирующие ВСМА-специфичный T-ChARM, могут быть обнаружены или отслежены in vivo с использованием антител, которые специфично связываются с меткой-кассетой (например, антител против метки), или с помощью других родственных связывающих белков, которые специфично связывают последовательность метки-кассеты (например, стрептактин-связывающая Strep-метка), партнеры по связыванию которой с меткой-кассетой конъюгированы с флуоресцентным красителем, радиоактивным изотопом, наночастицами оксида железа или другим визуализирующим агентом, известным в данной области техники, для обнаружения с помощью рентгеновского излучения, КТ-сканирования, МРТ-сканирования, ПЭТ-сканирования, ультразвука, проточной цитометрии, систем визуализации в ближней инфракрасной области или других методов визуализации (см., например, Yu и др., Theranostics 2: 3, 2012).
В других вариантах осуществления клетки, экспрессирующие ВСМА-специфичный T-ChARM настоящего изобретения, могут использоваться в диагностических способах или методах визуализации, включая способы, применяемые в отношении показаний или состояний, определенных в настоящем документе.
В некоторых вариантах осуществления способ включает введение GSI в комбинации (например, одновременно, отдельно или последовательно) с ВСМА-специфично связывающим белком, который может включать, например, ВСМА-специфичное антитело или его антигенсвязывающую часть, такую как конъюгат антитело-лекарственное средство или биспецифичныйе или мультиспецифичный связывающий белок, такой как те, которые полезны для предварительно таргетированной лучевой иммунотерапии (см., например, Green и др., Blood 131: 611 (2018)).
ПРИМЕРЫ
Пример 1
Разработка и тестирование ВСМА-специфичных химерных антигенных рецепторов
Анти-ВСМА CAR были подготовлены для изучения их полезности для иммунотерапии множественной миеломы и других заболеваний. Анти-ВСМА CAR были сконструированы с scFv, состоящими из областей VH и VL из антитела С115 D5.3 («С11») и антитела A7D12.2 («А7»), которые включали шарнирную область IgG4 (спейсер), трансмембранный домен CD28, костимулирующий домен 4-1ВВ и эффекторный домен CD3ζ. ScFv были получены как в «HL», так и в «LH» ориентации (см. Фиг. 1А). Т-клетки человека трансдуцировали экспрессирующими конструкциями, кодирующими Анти-ВСМА CAR, и исследовали на функциональные характеристики. Как показано на Фиг. 1B, Т-клетки, экспрессирующие CAR С11 и А7 CAR, пролиферировали при совместном культивировании с клетками-мишенями, экспрессирующими ВСМА, хотя CAR С11, по-видимому, пролиферируют несколько более устойчиво, чем Т-клетки, экспрессирующие CAR А7. Клетки CAR-T С11 также продуцировали больше цитокинов в ответ на клеточные линии-мишени (либо трансфицированные антигеном клетки К562, либо клеточные линии ММ, экспрессирующие ВСМА; Фиг. 1С) и обладали большей специфичной активностью уничтожения клеток-мишеней по сравнению с CAR АН LH -Т клетки (Фиг. 1D).
Были получены дополнительные CAR А7 и С11, имеющие различные внутриклеточные компоненты, включая CAR, содержащие костимулирующий домен 4-1 ВВ (Фиг. 1Е, верхняя иллюстрация) и костимулирующий домен CD28 (Фиг. 1Е, нижняя иллюстрация). Длина внеклеточного спейсерного домена также варьировалась для улучшения взаимодействия между CAR-экспрессирующей Т-клеткой и клеткой-мишенью ВСМА+, включая короткий (например, 12 аминокислот, 48 аминокислот и 66 аминокислот в длину), промежуточный (например, 157 аминокислот в длину) и длинный (например, 228 аминокислот в длину) спейсеры. Каждый из 48 аминокислотных спейсеров и 157 аминокислотных спейсеров CAR включал две (2) кассеты Strep-метки, в то время как спейсер 66 аминокислот включал три (3) кассеты Strep-метки (см. Фиг. 1F). Меченые химерные антигенные рецепторы, такие как CAR, содержащие кассету Strep-метки, показанные на Фиг. 1F и 1G, обозначаются здесь как T-ChARM.
Т-клетки человека трансдуцировали для экспрессии T-ChARM (см. Пример 2) и анализировали на функциональность. Длина спейсера оказала влияние на конструкции Т-ChARM, содержащие scFv, полученный из анти-ВСМА антител С11 D5.3, где промежуточное соединение (около 65 аминокислот; см. С11 3ST, С11 2STint и C11Lo на Фиг. 1Н) является длинным (около 200 аминокислот) спейсеры работали лучше всего для этого связывающего домена. C113ST_4-1BB и C113ST_CD28 были выбраны в качестве наиболее эффективных конструкций (Фиг. 1H-1J) и сравнивались с описанными анти-ВСМА CAR («ВСМА-2»; Carpenter и др. Clin. Cancer Res. 19: 2048, 2013). Экспрессия EGFRt в первичных Т-клетках была сходной для каждой конструкции T-ChARM / CAR, и поверхностная экспрессия T-ChARM была подтверждена для тех, которые содержали последовательности STII, как показано окрашиванием моноклональным антителом против STII (Фиг. 1К). Т-клетки, экспрессирующие каждый C113ST T-ChARM, продуцировали больше цитокинов (Фиг. 1L, 1М) и пролиферировали более устойчиво (Фиг. 1N), чем Т-клетки, экспрессирующие ВСМА-2, при совместном культивировании с клетками-мишенями. Также было определено, что Т-клетки, экспрессирующие каждый C113ST Т-ChARM, могут распознавать и лизировать CD 138 + клетки ММ пациента (данные не показаны). Т-клетки, экспрессирующие либо C113ST T-ChARM, либо CAR ВСМА-2, не обладали цитолитической активностью в отношении неэкспрессирующих клеток К562 (Фиг. 10), но эффективно лизировали клетки К562, трансдуцированные для экспрессии антигена ВСМА (Фиг. 1Р), демонстрируя что сконструированные Т-клетки специфично распознают антиген.
Пример 2
Производство рекомбинантных Т-клеток и
экспрессия ВСМА-специфичных T-ChARM
CD8+ и CD4+ Т-клетки выделяли из РВМС нормальных человеческих доноров с использованием набора для выделения CD8+/CD4+ Т-клеток (Miltenyi Biotec), активировали анти-CD3/CD28 шариками (Life Technologies) в соответствии с инструкциями производителя и трансдуцировали с помощью CAR-кодирующего супернатанта лентивируса (epHIB7), полученного путем временной трансфекции клеток HEK293T с использованием упаковочных плазмид PsPAX2 и pMD2G (MOI=3) с добавлением 0,8 мкг/мл полибрена (Millipore, Bedford, MA) на 3-й день после активации центрифугированием при 2100 об/мин в течение 45 минут мин при 32°С. Т-клетки размножали в RPMI, 10% человеческой сыворотке, 2 мМ L-глутамина и 1% пенициллин-стрептомицине (среда CTL), с добавлением рекомбинантного человеческого (rh) IL-2 до конечной концентрации 50 ед/мл каждые 48 часов. После размножения аликвоту каждой трансдуцированной линии Т-клеток окрашивали конъюгированным с биотином анти-EGFR антителом и стрептавидином- РЕ (Miltenyi, Auburn, СА). TEGFR+ Т-клетки выделяли путем сортировки на FACS-клеточном сортировщике Aria (Becton Dickinson). Подмножество tEGFR+ Т-клеток затем стимулировали облученным (8000 рад) CD19+ В-LCL при соотношении Т-клетка : LCL 1:7 и размножали в течение 8 дней в среде CTL с добавлением 50 ед/мл rh IL-2 каждый 48 часов или с использованием протокола быстрой экспансии для клеток, экспрессирующих T-ChARM (Riddell and Greenberg, J. Immunol. Methods 128: 189, 1990), где T-ChARM содержит scFv, полученный из анти-ВСМА антитела A7 или С11 D5. 3.
Следующие конъюгированные антитела были использованы для фенотипирования и анализа проточной цитометрии: CD4, CD8, CD25, CD137, CD45, аннексии V, CD62L, CD27, CD28 (BD Biosciences), анти-Streptag II-антитело (Genscript), EGFR-антитело (ImClone Systems Incorporated, Branchburg, NJ); стрептавидин-РЕ (BD Biosciences, Сан-Хосе, Калифорния). Окрашивание йодидом пропидия (PI, BD Biosciences) проводили для распознавания живых/мертвых клеток в соответствии с указаниями производителя. Анализ потока проводился на FACS Canto II, сортировка с помощью FACS AriaII (Becton Dickinson, Franklin Lakes, NJ) и анализ данных с использованием программного обеспечения FlowJo (Treestar, Ashland, OR).
Чтобы исследовать экспрессию на клеточной поверхности ВСМА-специфичного Т-ChARM, трансдуцированные Т-клетки сортировали по экспрессии EGFRt и оценивали путем окрашивания меченным флуорохромом анти-Streptag mAb. Средняя интенсивность флуоресценции (MFI) окрашивания EGFR была сходной на Т-клетках, трансдуцированных каждым из ВСМА-специфичных T-ChARM и CD19-Short CAR. Введение метки в CAR для получения T-ChARM не влияло на экспрессию трансгена (данные не показаны). Специфично окрашенные анти-Strep-метка mAb Т-клетки, трансдуцированные различными ВСМА-специфичесными T-ChARM, независимо от положения или количества последовательностей меток в каждом T-ChARM.
Пример 3
Растворимый ВСМА (sBCMA) ингибирует активность ВСМА-специфичных CAR Т-клеток
Была исследована потенциальная эффективность ВСМА-таргетированной Т-клеточной терапии против большинства миелом. Поскольку выделение поверхностного ВСМА клетками миеломы может препятствовать распознаванию Т-клеток, уровни растворимого ВСМА измерялись в супернатантах во время культивирования линии клеток миеломы. Клетки миеломы U266 промывали и высевали в культуральную среду на 1, 3, 5 и 24 часа. Супернатант среды собирали и анализировали на sBCMA с помощью ИФА. Данные показывают зависящее от времени увеличение уровней sBCMA в супернатанте (Фиг. 2А). Затем была исследована степень экспрессии ВСМА в клетках первичной пациентской множественной миеломы (ММ) и продукции цитокинов ВСМА-специфичными T-ChARM Т-клетками, связанными с клетками ММ пациента. На Фиг. 2В показана поверхностная экспрессия ВСМА контрольной клеточной линией (RPMI, левая панель) и образцами из первичных клеток ММ пациента с различными уровнями экспрессии ВСМА. Круговая диаграмма на Фиг. 2С показывает, что экспрессия ВСМА на поверхности клеток ММ у девятнадцати (19) разных пациентов была положительной для большинства пациентов, хотя были некоторые образцы пациентов, которые показали промежуточную или низкую/нулевую поверхностную экспрессию ВСМА (~25%). Кроме того, на Фиг. 2D показано, что T-ChARM Т-клетки продуцировали больше IFN-γ при культивировании с клетками ММ пациента, экспрессирующими высокие уровни поверхностного ВСМА, по сравнению с культурой с клетками ММ пациента с низким уровнем ВСМА.
Клетки миеломы часто экспрессируют PD-L1, который, как полагают, ингибирует функцию Т-клеток, связываясь с PD-1 на Т-клетках (Freeman и др. J. Exp. Med. 192 (7): 1027 (2000)). Была также исследована возможность того, что PD-L1 повлиял на ВСМА-специфичные T-ChARM Т-клетки. Проточная цитометрия образцов пациентов показала, что PD-L1 также был экспрессирован на 79% образцов, с 47% PD-L1hi, 32% PD-L1int и 21% PD-L1low/neg (Фиг. 2E и 2F). Корреляции между экспрессией ВСМА и PD-L1 не наблюдалось. Фиг. 2G показывает, что T-ChARM Т-клетки продуцировали больше IFN-γ при культивировании с клетками ММ пациента, экспрессирующими низкие уровни PD-L1 по сравнению с высокими уровнями PD-L1, хотя эта тенденция не была статистически значимой.
Эффект sBCMA также исследовали путем добавления sBCMA к ко-кулыурам Т-клеток, экспрессирующих ВСМА-специфичные T-ChARM, и K562 клетки, трансдуцированные полинуклеотидом, кодирующим полноразмерный ВСМА (K562/ВСМА+). Дозозависимое ингибирование ВСМА-специфичной эффекторной функции Т-ChARM Т-клеток было обнаружено как мера высвобождения IFN-γ в супернатант среды после введения экзогенного sBCMA (Фиг. 2Н, 2I). Кроме того, клеточные линии ММ показали значительное увеличение уровня sBCMA в супернатанте культуры в течение 24 часов (Фиг. 2J).
Чтобы исследовать, могут ли высокие уровни sBCMA ингибировать активность ВСМА-специфиченых T-ChARM Т-клеток, сыворотки костного мозга (ВМ) были исследованы на уровни sBCMA. Было установлено, что уровень sBCMA в ВМ у пациентов высокий и приблизительно коррелирует с бременем заболевания, определяемым процентом присутствующих клеток CD138+ (Фиг. 2K). Чтобы проверить, будет ли sBCMA связываться с T-ChARM Т-клетками, Т-клетки инкубировали с повышающимися уровнями рекомбинантного ВСМА и затем окрашивали с использованием BCMA-Fc, конъюгированного с АРС. Как показано на Фиг. 2L, окрашивание уменьшилось с более высокими уровнями рекомбинантного ВСМА, что указывает на то, что sBCMA потенциально может оказывать вредное влияние на ВСМА-специфичную функцию Т-ChARM Т-клеток.
Для подтверждения экспрессии T-ChARM Т-клетки C113ST T-ChARM и контрольные анти-CD19 CAR Т-клетки ("FMC63") инкубировали с Fc-BCMA и окрашивали на EGFRt (маркер трансдукции CAR/T-ChARM) и CD4, показывая, что Т-ChARMs были экспрессированы клетками (Фиг. 2М). Окрашивание EGFRt и других молекул поверхности Т-клеток не было затронуто (данные не показаны).
Чтобы определить, ингибирует ли выделение и связывание sBCMA с T-ChARM функцию Т-клеток, BCMA-Fc добавляли к совместным культурам Т-клеток/линий клеток-мишеней, что приводило к значительно уменьшенному распознаванию (продуцированию IFN-γ и экспрессии CD4) путем T-ChARM Т-клетки, таргетированными на ВСМА, но не контрольные анти-С019 CAR Т-клетки (культивируемые с клетками K562, экспрессирующими антиген CD 19 (Фиг. 2N). Воздействие на ВСМА T-ChARM-клеток былр дозозависимым (Фиг. 2O, 2Р). Добавление рекомбинантного ВСМА, не слитого с Fc, также ингибировало продукцию IFN-γ в зависимости от дозы, но не ингибировало лизис клеток K562 ВСМА+ T-ChARM Т-клетками (Фиг. 2Q), которые могут иметь место из-за высокой плотности антигена на поверхности клеток-мишеней.
Пример 4
Влияние ингибитора γ-секретазы на уровни ММ ВСМА
Для изучения влияния ингибирования γ-секретазы на уровни ВСМА в клетках миеломы был использован ингибитор γ-секретазы (GSI) RO4929097. ВСМА быстро активируется на различных клеточных линиях миеломы при инкубации с ингибитором γ-секретазы (GSI) RO4929097 (используемая концентрация варьировалась от 0,001 мкМ до 1,0 мкМ) (Фиг. 3A-3D).
Для изучения влияния GSI на кинетику экспрессии ВСМА на поверхности клетки миеломы U266 инкубировали в течение 1, 3, 5 и 24 часов в присутствии различных концентраций GSI RO429097 (0,01 мкМ, 0,1 мкМ и 1,0 мкМ) и оценивали экспрессию ВСМА на клеточной поверхности методом проточной цитометрии. Экспрессия ВСМА возрастала в зависимости от дозы в присутствии GSI (Фиг. 3Е). Повышенная регуляция сохранялась в течение 7 дней культивирования в 1,0 мкМ GSI (Фиг. 3F).
Чтобы проверить влияние GSI на выделение ВСМА в ММ, три разные клеточные линии миеломы (MM1.R, U266 и 8226) промывали и высевали в культуральную среду на 24 часа в присутствии различных концентраций GSI RO429097 (0,01 мкМ, 0,1). мкМ и 1,0 мкМ). Супернатант среды собирали и анализировали на sBCMA с помощью ИФА. Уровни sBCMA снижались в супернатанте в присутствии GSI в зависимости от дозы (Фиг. 3G и 3Н).
Чтобы исследовать влияние на уровни растворимого ВСМА (sBCMA) с течением времени, клетки U266 промывали и высевали в культуральную среду на 1, 3, 5 и 24 часа в присутствии различных концентраций GSI RO429097 (0,01 мкМ, 0,1 мкМ и 1,0 мкМ). Супернатант среды собирали и анализировали на растворимый ВСМА (sBCMA) с помощью ИФА. Данные показывают дозозависимое снижение уровней растворимого ВСМА в супернатанте, когда присутствует GSI (Фиг. 3I).
Затем был изучен эффект удаления GSI из культуры клеток ММ. Как показано на Фиг. 3J и ЗК, уровни ВСМА на поверхности снижались после удаления 1,0 мкМ RO429097, в то время как содержание sBCMA в супернатанте увеличивалось при удалении GSI, но не увеличивалось при сохранении GSI. Эти данные указывают на обратимое ингибирующее влияние GSI на выделение поверхностного ВСМА. Наконец, на жизнеспособность протестированных клеточных линий ММ не влияло добавление 1,0 мкМ RO429097 в культуру (Фиг. 3L).
Влияние GSI на экспрессию ВСМА было затем проверено на образцах первичной миеломы от пациентов. Клетки миеломы CD138+ обогащали из образцов костного мозга пациента, инкубировали в течение 3 часов в присутствии различных концентраций GSI RO429097 (от 0,01 мкМ до 10 мкМ) и оценивали на предмет экспрессии ВСМА на поверхности методом проточной цитометрии (Фиг. 3М и 3R). Средняя интенсивность флуоресценции ВСМА (MFI) на опухолевых клетках представлена как кратное увеличение по сравнению с опухолевыми клетками, инкубированными без RO4929097. Наблюдается дозозависимая активация ВСМА на опухолевых клетках (Фиг. 3N), в то время как не было влияния на уровни некоторых других молекул клеточной поверхности, включая CS1, CD86, PD-L1, CD80 и CD38 (Фиг. 3O-3Q).
Пример 5
Ингибитор γ-секретазы улучшает распознавание Bcma+ миеломных клеток Т-клетками, экспрессирующими ВСМА-специфичный T-ChAR
Для того, чтобы изучить влияние GSI на активность CAR-T-клеток против ВСМА+ множественной миеломы, продукцию IL-2 ВСМА CAR-T клетками (ВСМА-специфичные T-ChARM С11 3ST-CD28 и ВСМА-специфичные T-ChARM С11 3ST-41BB) или контрольными CD19sh CAR Т-клетками (короткий спейсер) измеряли после совместного культивирования с первичными опухолевыми клетками миеломы человека в течение 24 часов в среде, содержащей в различных концентрациях GSI RO429097 (от 0,003 мкМ до 3,0 мкМ) (Фиг. 4А). Лечение GSI приводило к дозозависимому увеличению продукции IL-2 Т-клетками ВСМА T-ChARM. Кроме того, продукция IFNγ Т-клетками ВМСА Т-ChARM, совместно культивированными с клетками миеломы при различных концентрациях RO429097, была измерена и также увеличилось в присутствии GSI (Фиг. 4В). Эти данные показывают, что клетки множественной миеломы лучше стимулируют ВСМА-специфичные T-ChARM Т-клетки при предварительной обработке ингибитором γ-секретазы, который усиливает экспрессию ВСМА в опухолевой клетке. Наконец, пролиферация меченных CFSE ВСМА-специфичных T-ChARM Т-клеток увеличивалась дозозависимым образом после совместного культивирования в течение 3 дней с первичными опухолевыми клетками миеломы человека в присутствии GSI RO492097 (Фиг. 4С).
Не желая быть связанными какой-либо теорией, отметим, что потенциальное предостережение от использования GSI для увеличения ВСМА в клетках миеломы для улучшения распознавания клеток CAR-T заключается в том, что высокие концентрации GSI могут ингибировать передачу сигналов Т-клетками и эффекторные функции. См. Eagar и др., Immunity 20 (4): 407, 2004. Чтобы оценить потенциальные эффекты GSI на CAR Т-клетки в условиях, когда экспрессия лиганда-мишени остается стабильной, CD 19-CAR-T-клетки совместно культивировали с Raji или K562/CD19+ клетки-мишени в присутствии GSI (от 0,01 мкМ до 100 мкМ) и измеряли жизнеспособность и эффекторную функцию. В этом диапазоне концентраций не было влияния на экспрессию антигена CD 19 (Фиг. 5А) или на жизнеспособность клеток CAR-T через 24 часа (Фиг. 5В). Было обнаружено, что GSI RO4929097 ингибирует эффекторную функцию CD 19 CAR-T-клеток при концентрациях более 1 мкМ при совместном культивировании с клетками K562 CD 19+, как было определено путем измерения продукцию IL-2 (Фиг. 5С, верхняя панель) и IFNγ (Фиг. 5С, нижняя панель) (см. также Фиг. 5D-5F). Однако на способность клеток CD 19 CAR-T специфично убивать клетки-мишени не влияла концентрация GSI (Фиг. 5G). Клетки CD 19 CAR-T также пролиферировали в ответ на стимуляцию клеток-мишеней в присутствии 10 мкМ GSI RO4929097, хотя и менее эффективно, чем в отсутствие GSI RO4929097 (Фиг. 5Н). Для дальнейшего исследования дозированного влияния GSI на деление клеток CAR-T, дозированное титрование GSI RO4929097 было добавлено к совместным культурам CD 19 CAR-T и клеток-мишеней. При добавлении терапевтических доз GSI (0,01 мкМ - 1 мкМ) деление клеток CAR-T практически не затрагивалось или изменялось лишь незначительно, тогда как GSI в более высокой нетерапевтической дозе (10 мкМ) действительно заметно ингибировал пролиферацию (Фиг. 5I). Клетки CD 19 CAR-T, культивируемые в присутствии GSI, не демонстрировали снижения экспансии в течение 8 дней в культуре с (Фиг. 5J) или без экзогенного IL-2 (Фиг. 5K). Кроме того, Т-клетки, которые были размножены в присутствии 5 мкМ, 0,5 мкМ или в отсутствие GSI, не демонстрировали существенной разницы в продукции IFN-γ при последующей повторной стимуляции клетками-мишенями после размножения в присутствии (Фиг. 5L) или в отсутствие (Фиг. 5М) экзогенного IL-2. Продукция IL-2 между группами клеток CAR-T также не заметно различалась (Фиг. 5N). В целом, GSI в терапевтических уровнях не влияет на функцию Т-клеток, включая продукцию цитокинов, активность лизиса клеток и пролиферацию.
Затем было исследовано влияние GSI на Т-клеточную функциональность Т-клеток, содержащих ВСМА-специфичный T-ChARM. Обработка GSI первичных образцов опухоли миеломы значительно улучшила продукцию IFN-γ (Фиг. 5O, 5Р и 5Q) и экспрессию CD8 (Фиг. 5O) совместно культивированными T-ChARM Т-клетками, с эффектами, наблюдаемыми после обработки 0,1 мкМ GSI или выше, а также увеличивало пролиферации Т-клеток (Фиг. 5R).
Эти данные указывают на клиническую полезность сочетания ВСМА-специфичных CAR Т-клеток и GSI для повышения уровня ВСМА в клетках множественной миеломы. Это может обеспечить преимущество или даже оказать синергетический эффект при лечении множественной миеломы путем облегчения элиминации опухолевых клеток, которые экспрессируют низкие уровни ВСМА. Данные также показывают, что диапазон концентраций ингибитора γ-секретазы способствует экспрессии ВСМА опухолевыми клетками, не влияя на активность Т-клеток.
Пример 6
Исследование активности GSI in vivo на модели ксенотрансплантата ММ
Чтобы исследовать, могут ли sBCMA-ингибирующие эффекты GSI RO4929097, описанные в Примере 4, реплицироваться in vivo, тест на мышиной модели ксенотрансплантата ММ человека проводили, как показано на Фиг. 6А. Вкратце, иммунодефицитные мыши NOD/SCID (NSG) (The Jackson Laboratory) получали сублетальное облучение (275 рад) в день -1, за которым следовало 5×106 клеток MM. 1R на следующий день (день 0). GSI RO4929097 (30 мг/кг) вводили через желудочный зонд на 19-й и 20-й день. Мышей умерщвляли в разное время после повторного введения GSI через желудочный зонд и отбирали образцы крови и костного мозга для анализа. Дозирование GSI сопровождалось быстрым увеличением (более 3-кратного на 4 часа после второго зондирования) экспрессии поверхностного ВСМА в опухолевых клетках (Фиг. 6В) и одновременным снижением sBCMA (Фиг. 6С). Наблюдаемые эффекты, как правило, были снижены в течение 48 часов после второй обработки через желудочный зонд.
Пример 7
Исследование in vivo CAR Т-клеток+ GSI комбинированной терапии с таргетингом на ММ
Была исследована способность GSI-терапии улучшать анти-ВСМА CAR Т-клеточную терапию in vivo. В эксперименте, аналогичном описанному в примере 6, мышей NSG облучали сублетально в день -21, после чего вводили клетки ММ человека (5×106 MM.lRffluc) в день -20 (см. Фиг. 7А). Первую дозу GSI (30 мг/кг RO4929097) вводили через желудочный зонд в день -1. В день 0 мышам вводили субоптимальную дозу С11 3ST T-ChARM Т-клеток (0.33×106 клеток; CD4 : CD8 1:1) и они получали вторую дозу GSI. Дальнейшие дозы GSI вводили в день +1, день +8 и день +9. Биолюминесцентная визуализация (BLI) выполнялась на протяжении всего эксперимента, и мышей контролировали на выживаемость. Как показано на Фиг. 7В, 7С и 7Е (левый график), у группы, получавшей GSI + Т-клетки, было пониженное свечение и больше мышей без обнаруживаемых опухолей по сравнению с группой, получавшей только Т-клетки (данные BLI из Т-клеток только группа не показана). Количественные данные люминесценции (Фиг. 7С и 7Е (левый график)) показали, что первоначальный эффект T-ChARM Т-клеток начал проявляться приблизительно на 9-й день, когда опухоли начали расти («Mock + ВСМА Т-клетки»). Противоопухолевый эффект был продлен путем комбинации с GSI RO4929097 («RO49 + ВСМА Т-клетки»), но опухоли в этой группе лечения также в конечном итоге выросли. Данные визуализации соответствовали выживаемости мышей (Фиг. 7D и 7Е (правый график)), при этом мыши, получавшие комбинированную терапию, выживали дольше, чем группа, получающая Т-клетки только до смерти. В некоторых вариантах осуществления лечение GSI повторяют (по меньшей мере, от 2 до, по меньшей мере, приблизительно 5, по меньшей мере, приблизительно 25 раз) при использовании в комбинации с анти-ВСМА CAR Т-клетками, причем лечение GSI можно вводить одновременно, до или после введения анти-ВСМА CAR Т-клеток.
Пример 8
GSI улучшает связывание с клетками ВСМА + ММ
Биспецифичный анти-ВСМА слитый белок
Был сконструирован биспецифичный гибридный белок, обладающий специфичностью к ВСМА и другому антигену и имеющий метку YFP (желтый флуоресцентный белок). Биспецифичный гибридный белок добавляли к клеткам, экспрессирующим ВСМА Н929 ММ, в культуре (0.5×106 клеток) либо с GSI, либо без него. Связывание оценивали с помощью проточной цитометрии. Как показано на Фиг. 8, добавление GSI улучшало связывание ВСМА-связывающим биспецифичным слитым белком, но не оказывало влияния на связывание контрольным биспецифичным слитым белком, который не таргетирован на ВСМА. Эти данные показывают, что иммунотерапия с участием биспецифичных молекул, таргетированных на ВСМА, может быть усилена или улучшена с помощью GSI.
Различные варианты осуществления, описанные выше, могут быть объединены для обеспечения дополнительных вариантов осуществления. Все патенты США, публикации патентных заявок США, патентные заявки США, иностранные патенты, зарубежные патентные заявки и непатентные публикации, упомянутые в этом описании и/или перечисленные в листе данных заявки, включены в настоящий документ в качестве ссылки во всей их полноте. Аспекты вариантов осуществления могут быть модифицированы, если необходимо, чтобы использовать концепции различных патентов, заявок и публикаций для обеспечения еще дополнительных вариантов осуществления.
Эти и другие изменения могут быть сделаны в вариантах осуществления в свете вышеприведенного подробного описания. В общем, в следующей формуле изобретения используемые термины не должны толковаться как ограничивающие формулу изобретения конкретными вариантами осуществления, описанными в описании и формуле изобретения, но должны толковаться как включающие в себя все возможные варианты осуществления вместе с полным объемом эквивалентов, которым такая формула изобретения имеет отношения. Соответственно, формула изобретения не ограничивается описанием.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Фред Хатчинсон Кэнсер Рисерч Сентер
РИДДЕЛЛ Стенли Р.
ГРИН Демиан
ХИЛЛ Тайлер
<120> КОМБИНИРОВАННАЯ ТЕРАПИЯ ДЛЯ ЛЕЧЕНИЯ
СВЯЗАННЫХ С BCMA РАКОВЫХ ЗАБОЛЕВАНИЙ И АУТОИММУННЫХ РАССТРОЙСТВ
<130> 360056.445WO
<140> PCT
<141> 2018-02-16
<150> US 62/582,270
<151> 2017-11-06
<150> US 62/406,612
<151> 2017-02-17
<160> 34
<170> FastSEQ для Windows Версии 4.0
<210> 1
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 1
Gly Gly Gly Gly Ser
1 5
<210> 2
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 2
Gly Gly Gly Ser
1
<210> 3
<211> 25
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<220>
<221> ВАРИАНТ
<222> (5)...(20)
<223> любая одна или все аминокислоты c 5 по 20 могут
присутствовать или отсутствовать.
<400> 3
Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Ser
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser
20 25
<210> 4
<211> 45
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<220>
<221> ВАРИАНТ
<222> (5)...(0)
<223> любая одна или все аминокислоты c 5 по 20 могут
присутствовать или отсутствовать
<220>
<221> ВАРИАНТ
<222> (26)...(45)
<223> любая одна или все аминокислоты c 5 по 20 могут
присутствовать или отсутствовать.
<400> 4
Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Ser
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
20 25 30
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
35 40 45
<210> 5
<211> 25
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<220>
<221> ВАРИАНТ
<222> (6)...(20)
<223> любая одна или все аминокислоты c 5 по 20 могут
присутствовать или отсутствовать.
<400> 5
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser
20 25
<210> 6
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 6
His Gly Gly His His Gly
1 5
<210> 7
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 7
Trp Ser His Pro Gln Phe Glu Lys
1 5
<210> 8
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 8
Trp Arg His Pro Gln Phe Gly Gly
1 5
<210> 9
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 9
Cys Pro Pro Cys Pro
1 5
<210> 10
<211> 29
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<220>
<221> ВАРИАНТ
<222> (10)...(29)
<223> любая одна или все аминокислоты c 5 по 20 могут
присутствовать или отсутствовать.
<400> 10
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
1 5 10 15
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
20 25
<210> 11
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 11
Gly Gly Gly
1
<210> 12
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 12
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 13
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 13
Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Ser
1 5 10 15
<210> 14
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 14
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Arg Ala Ser Glu Ser Val Ser Val Ile
20 25 30
Gly Ala His Leu Ile His Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Leu Ala Ser Asn Leu Glu Thr Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Ala Glu Asp Ala Ala Ile Tyr Tyr Cys Leu Gln Ser Arg
85 90 95
Ile Phe Pro Arg Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 15
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 15
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Asn Cys Arg Ala Ser Glu Ser Val Ser Val Ile
20 25 30
Gly Ala His Leu Ile His Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Leu Ala Ser Asn Leu Glu Thr Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Ala Glu Asp Ala Ala Ile Tyr Ser Cys Leu Gln Ser Arg
85 90 95
Ile Phe Pro Arg Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 16
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 16
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Met Ser Leu Gly
1 5 10 15
Glu Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Ser Val Ile
20 25 30
Gly Ala His Leu Ile His Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Leu Ala Ser Asn Leu Glu Thr Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Arg Val Gln Ala Glu Asp Ala Ala Ile Tyr Ser Cys Leu Gln Ser Arg
85 90 95
Ile Phe Pro Arg Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 17
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 17
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Ser Ile Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Thr Glu Thr Arg Glu Pro Ala Tyr Ala Tyr Asp Phe
50 55 60
Arg Gly Arg Phe Val Phe Ser Leu Asp Thr Ser Val Ser Thr Ala Tyr
65 70 75 80
Leu Gln Ile Ser Ser Leu Lys Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Tyr Ser Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 18
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 18
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Glu
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Ser Ile Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Thr Glu Thr Arg Glu Pro Ala Tyr Ala Tyr Asp Phe
50 55 60
Arg Gly Arg Phe Val Phe Ser Leu Asp Thr Ser Val Ser Thr Ala Tyr
65 70 75 80
Leu Gln Ile Ser Ser Leu Lys Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Leu Asp Tyr Ser Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 19
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 19
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Glu
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Ser Ile Asn Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Trp Ile Asn Thr Glu Thr Arg Glu Pro Ala Tyr Ala Tyr Asp Phe
50 55 60
Arg Gly Arg Phe Val Phe Ser Leu Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Leu Gln Ile Ser Ser Leu Lys Ala Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Leu Asp Tyr Ser Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 20
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 20
Gln Val Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Glu
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Ser Ile Asn Trp Val Lys Gln Ala Pro Gly Gln Gly Leu Lys Trp Met
35 40 45
Gly Trp Ile Asn Thr Glu Thr Arg Glu Pro Ala Tyr Ala Tyr Asp Phe
50 55 60
Arg Gly Arg Phe Val Phe Ser Leu Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Leu Gln Ile Ser Ser Leu Lys Ala Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Leu Asp Tyr Ser Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 21
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 21
Gln Ile Gln Leu Val Gln Ser Gly Ser Glu Leu Lys Lys Pro Gly Glu
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Ser Ile Asn Trp Val Lys Gln Ala Pro Gly Gln Gly Leu Lys Trp Met
35 40 45
Gly Trp Ile Asn Thr Glu Thr Arg Glu Pro Ala Tyr Ala Tyr Asp Phe
50 55 60
Arg Gly Arg Phe Val Phe Ser Leu Asp Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Leu Gln Ile Ser Ser Leu Lys Ala Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Leu Asp Tyr Ser Tyr Ala Met Asp Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser
115
<210> 22
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 22
caccggagaa tcaaaatcgg tgaat
25
<210> 23
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 23
aaacattcac cgattttgat tctcc
25
<210> 24
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 24
caccgcagtt gtgtgacacg gaag
24
<210> 25
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 25
aaaccttccg tgtcacacaa ctgc
24
<210> 26
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 26
caccggcaaa ggtgagtgag acttt
25
<210> 27
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 27
aaacaaagtc tcactcacct ttgcc
25
<210> 28
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 28
caccggtttc tgcagccgct ttggg
25
<210> 29
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 29
aaaccccaaa gcggctgcag aaacc
25
<210> 30
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность G4S линкера
<400> 30
Gly Gly Gly Gly Ser
1 5
<210> 31
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<220>
<221> ВАРИАНТ
<222> (1)...(20)
<223> любая из аминокислот с 1 по 2 может отсутствовать или может
присутствовать до 10 раз, при условии, что аминокислоты 1
и 2 не отсутствуют одновременно
<400> 31
Gly Gly Gly Gly Gly Gly Gly Gly Gly Gly Ser Ser Ser Ser Ser Ser
1 5 10 15
Ser Ser Ser Ser
20
<210> 32
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 32
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10
<210> 33
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 33
Gly Gly Gly Ser Gly Gly Gly Ser
1 5
<210> 34
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетическая последовательность
<400> 34
Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Ser
1 5 10
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ, ТЕРАПИЯ И ПРИМЕНЕНИЕ ДЛЯ ЛЕЧЕНИЯ ЗЛОКАЧЕСТВЕННОГО НОВООБРАЗОВАНИЯ | 2021 |
|
RU2831425C1 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ, НАЦЕЛЕННЫЕ НА АНТИГЕН СОЗРЕВАНИЯ B-КЛЕТОК | 2013 |
|
RU2766608C2 |
| АНТИТЕЛО, НАЦЕЛЕННОЕ НА ВСМА, И ЕГО ПРИМЕНЕНИЕ | 2018 |
|
RU2799655C2 |
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО К BCMA И BCMA-CAR-T-КЛЕТКИ | 2020 |
|
RU2762942C1 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ ДЛЯ BCMA И ПУТИ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2785658C2 |
| АНТИТЕЛА К BCMA, СОДЕРЖАЩИЕ ТОЛЬКО ТЯЖЕЛУЮ ЦЕПЬ | 2018 |
|
RU2816994C2 |
| МОЛЕКУЛЫ, СВЯЗЫВАЮЩИЕ BCMA, И ПУТИ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2812322C2 |
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО К BCMA И BCMA-CAR-NK-КЛЕТКИ | 2020 |
|
RU2795443C2 |
| ГЕНЕТИЧЕСКИ МОДИФИЦИРОВАННЫЙ ОНКОЛИТИЧЕСКИЙ ВИРУС ПРОСТОГО ГЕРПЕСА, ДОСТАВЛЯЮЩИЙ ХЕМОКИН И ОПУХОЛЬ-АССОЦИИРОВАННЫЙ/СПЕЦИФИЧЕСКИЙ АНТИГЕН | 2022 |
|
RU2827338C1 |
| АНТИ-BCMA АНТИТЕЛА, СОДЕРЖАЩИЕ ТОЛЬКО ТЯЖЁЛУЮ ЦЕПЬ | 2017 |
|
RU2781301C2 |
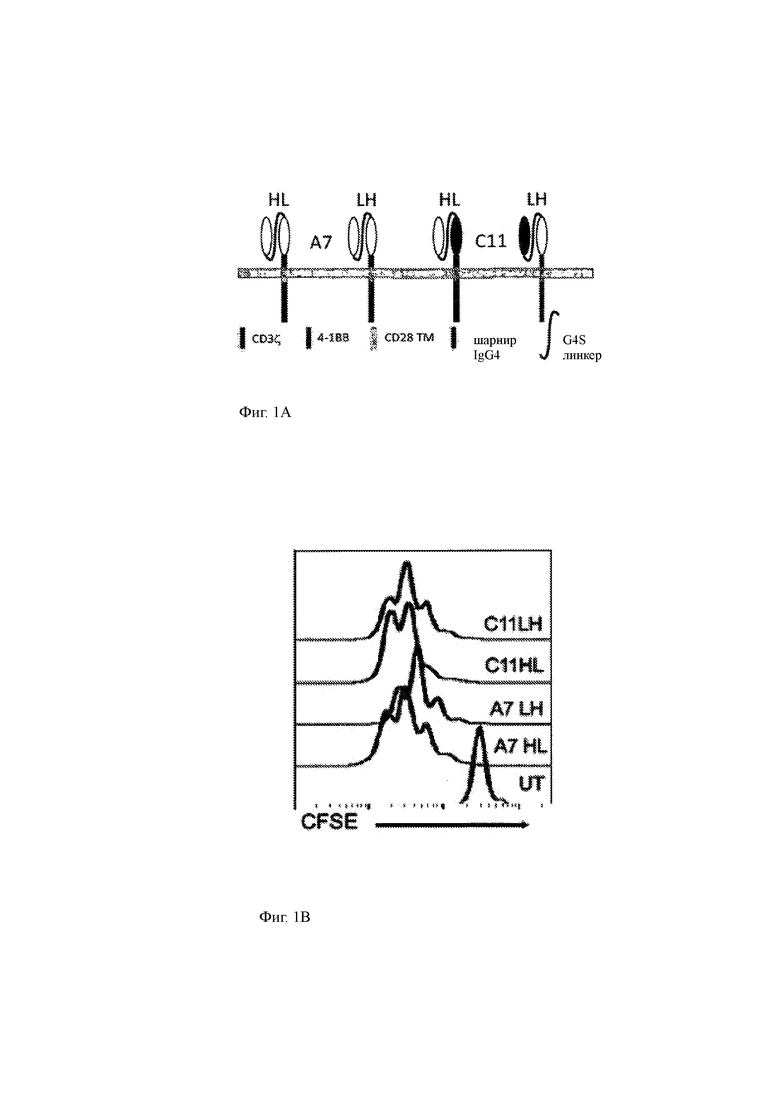
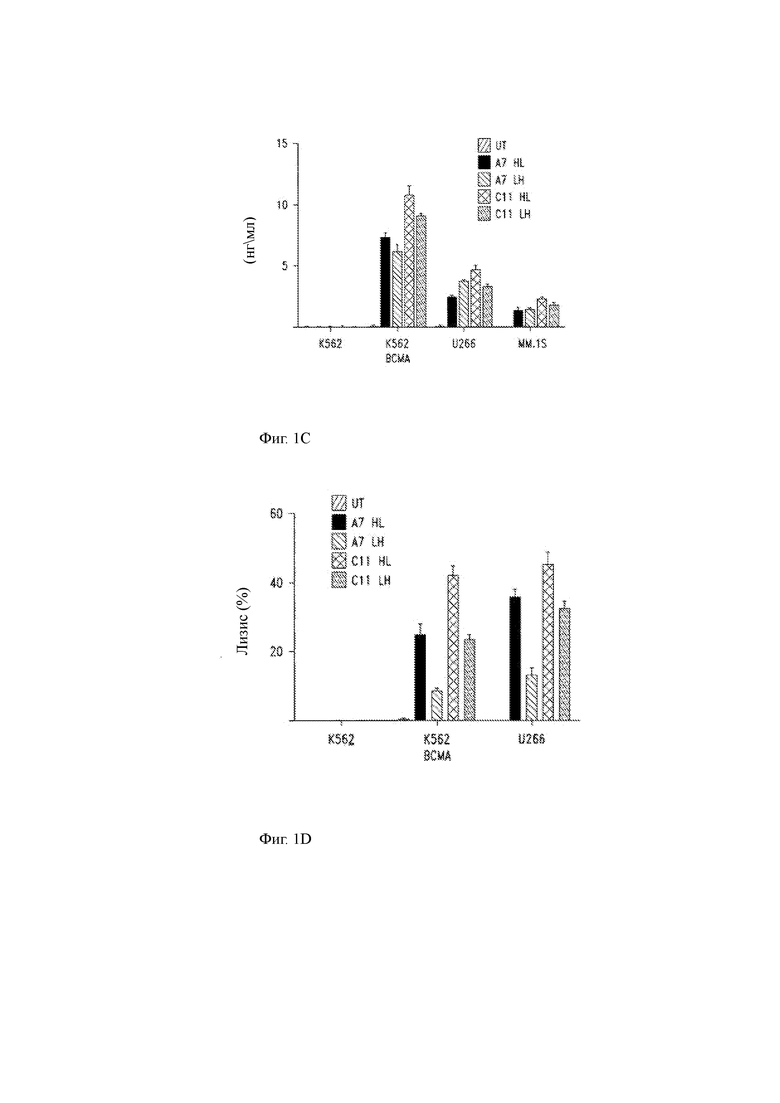
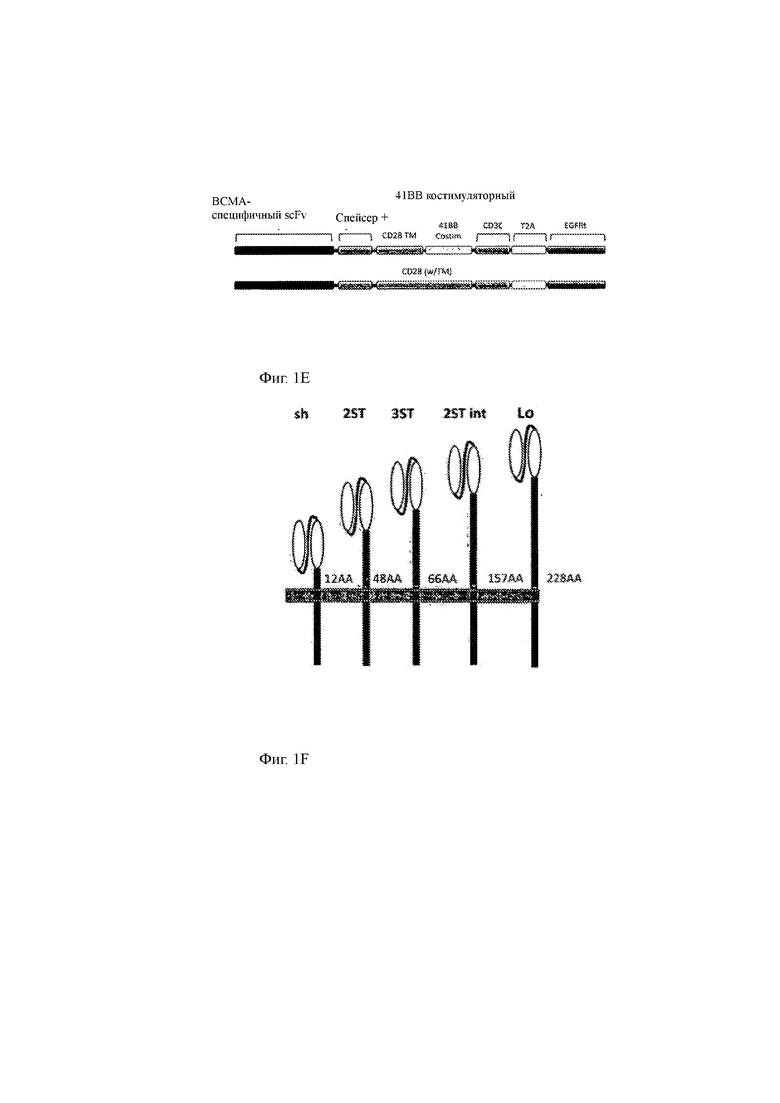
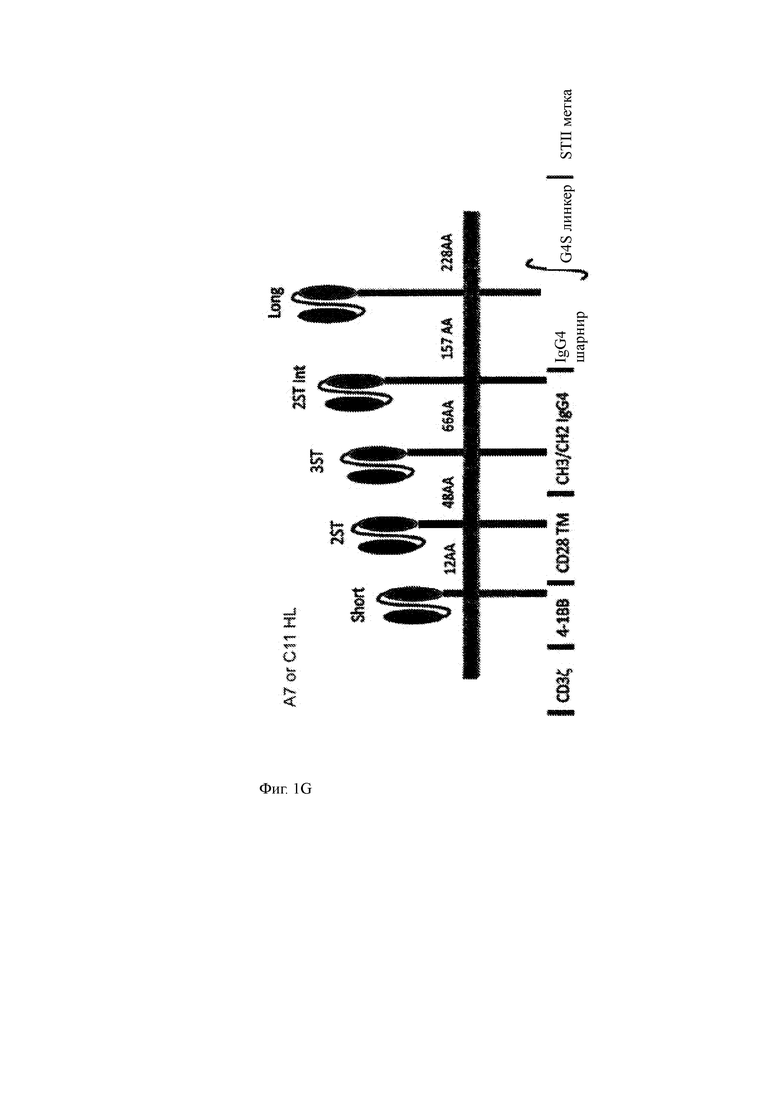
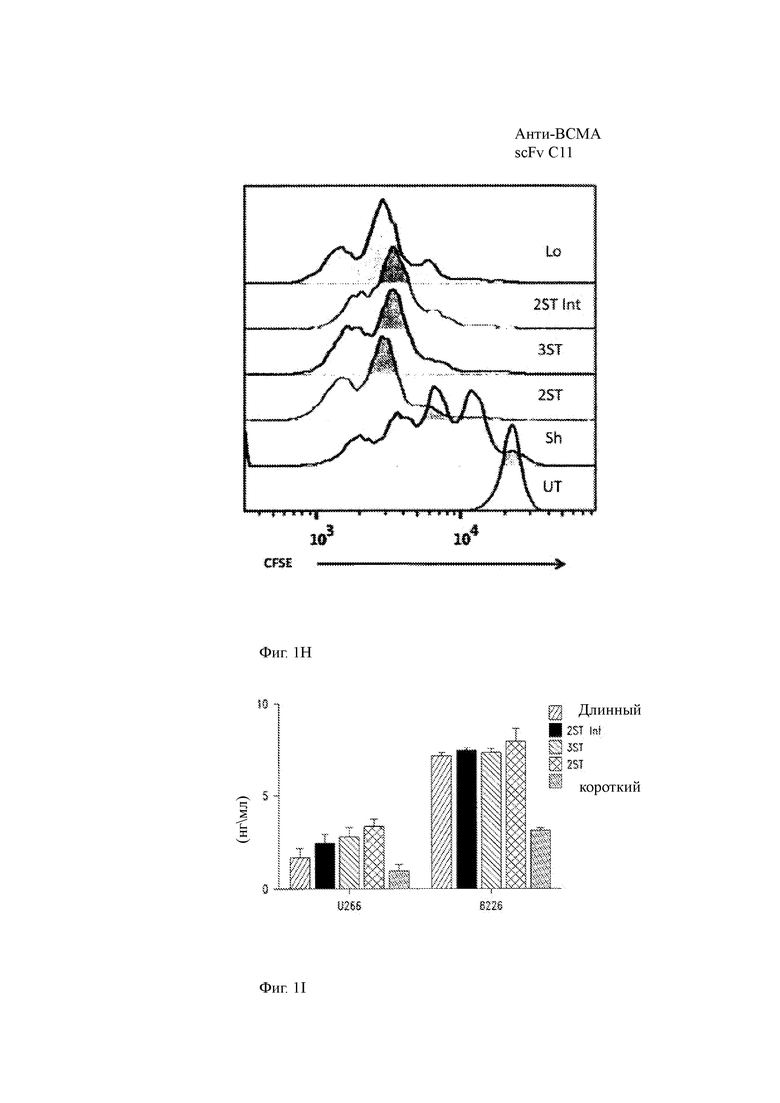
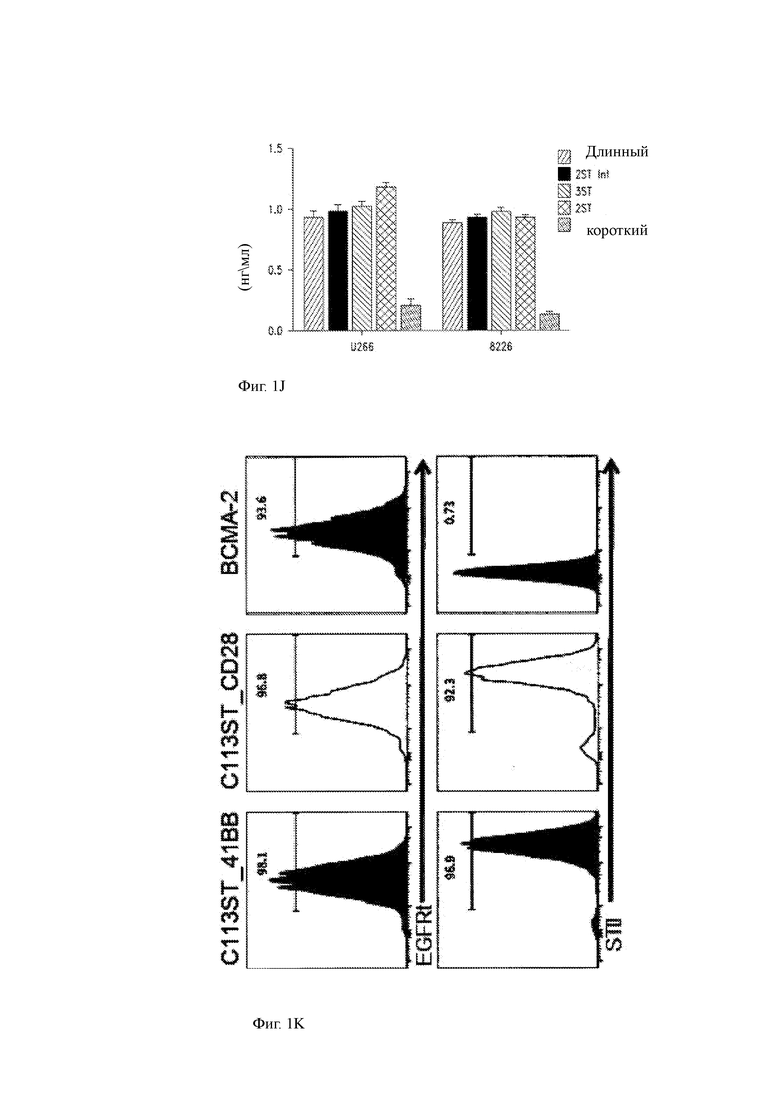
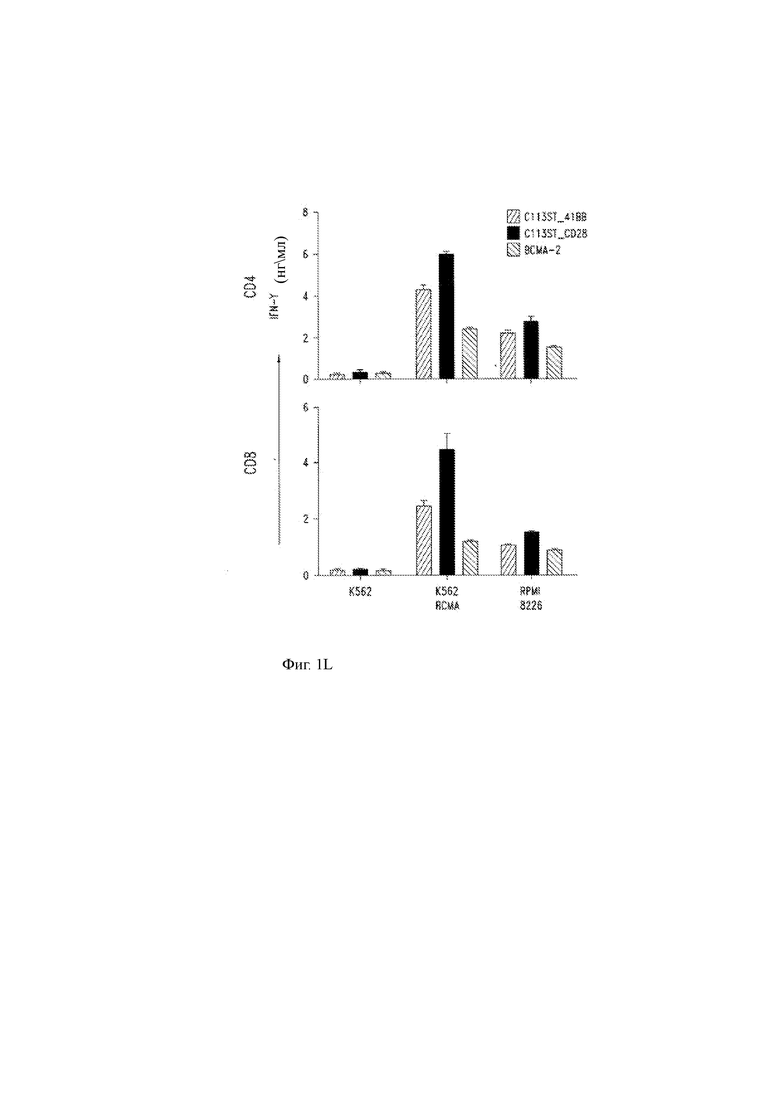
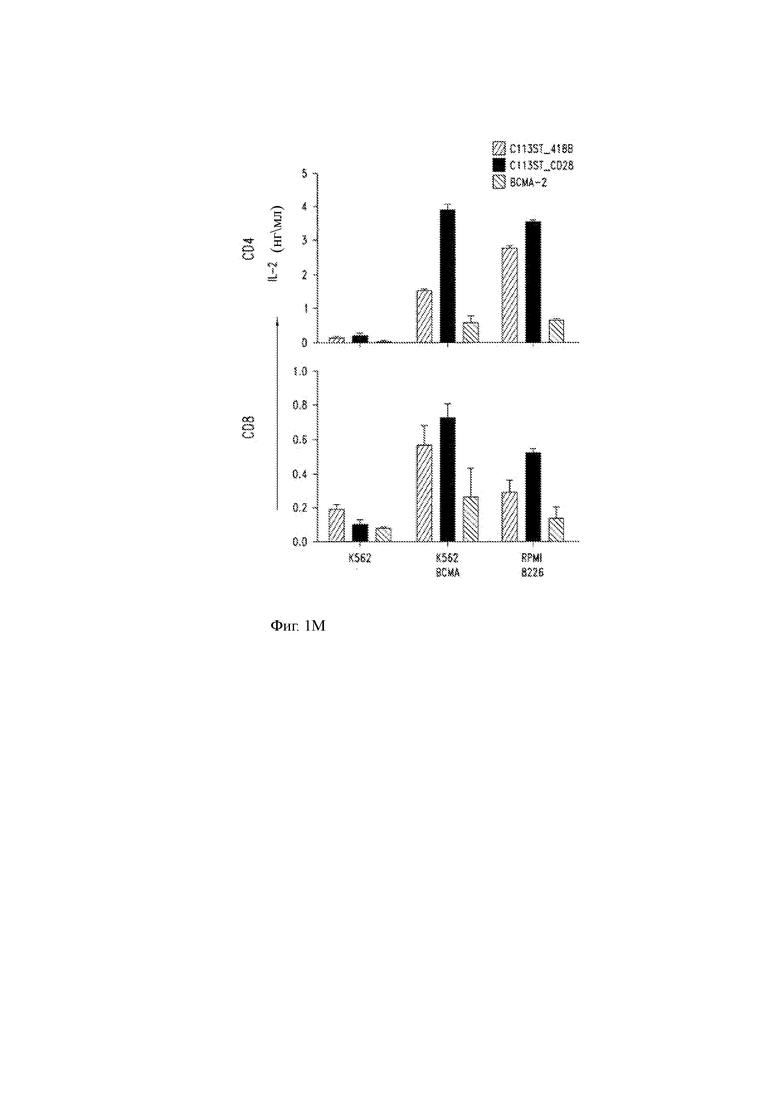
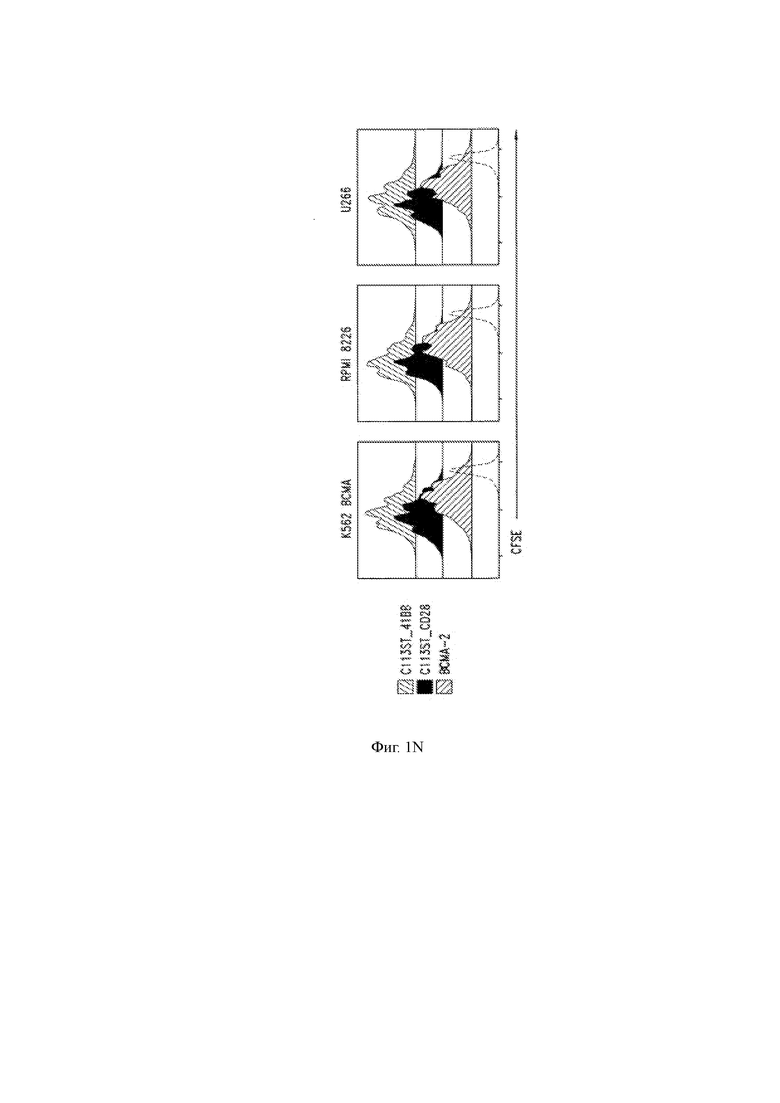
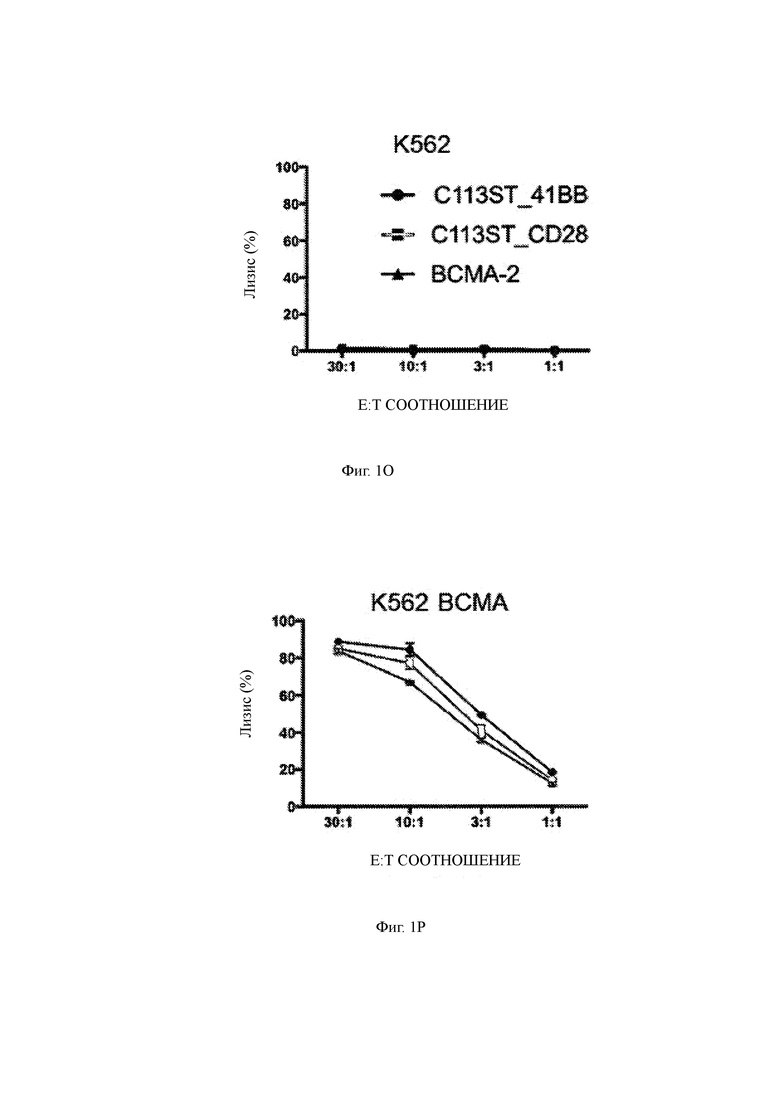
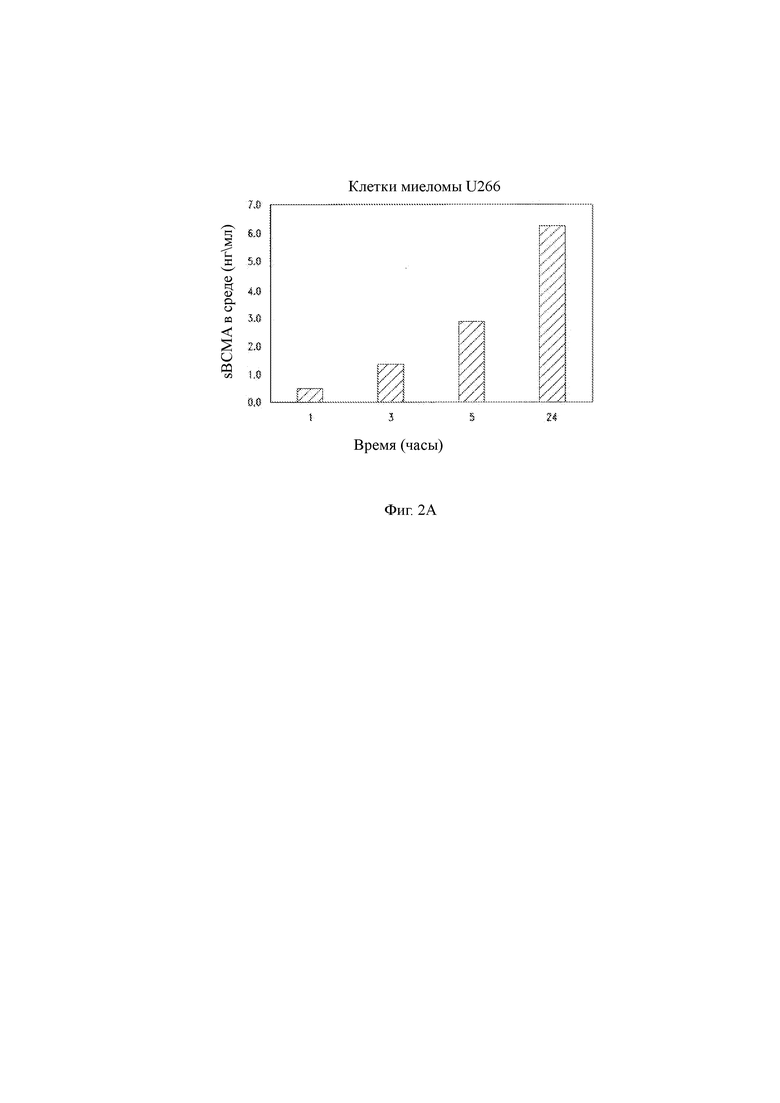
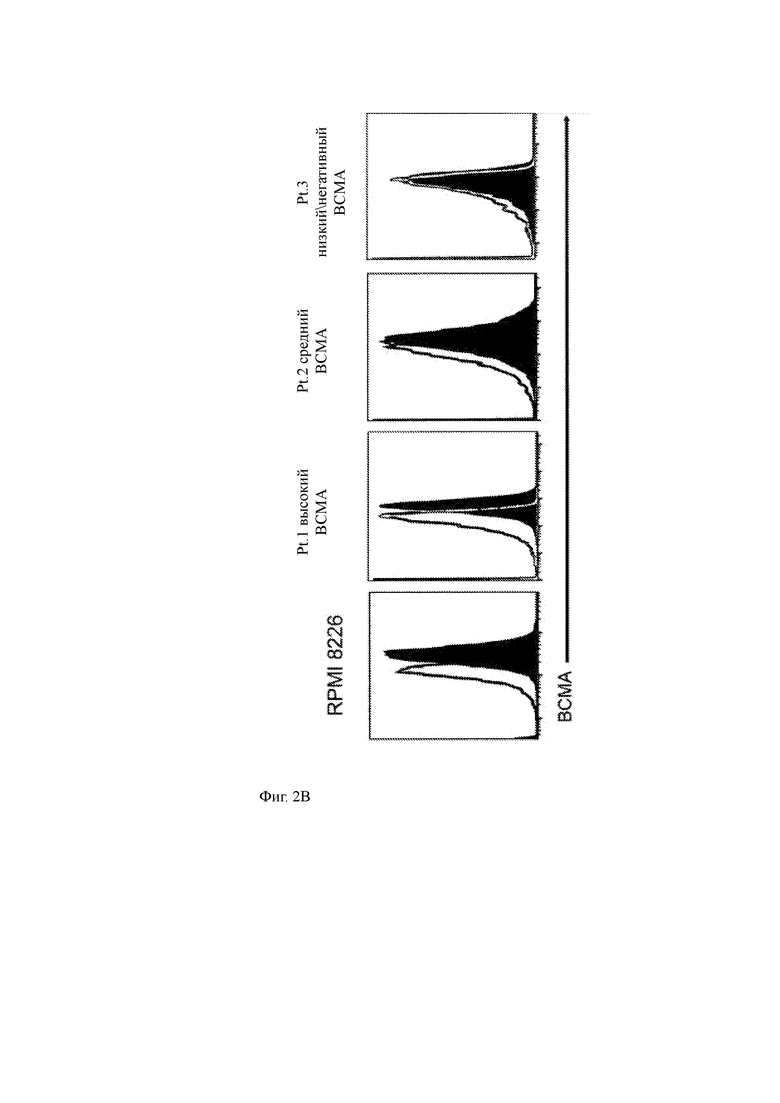
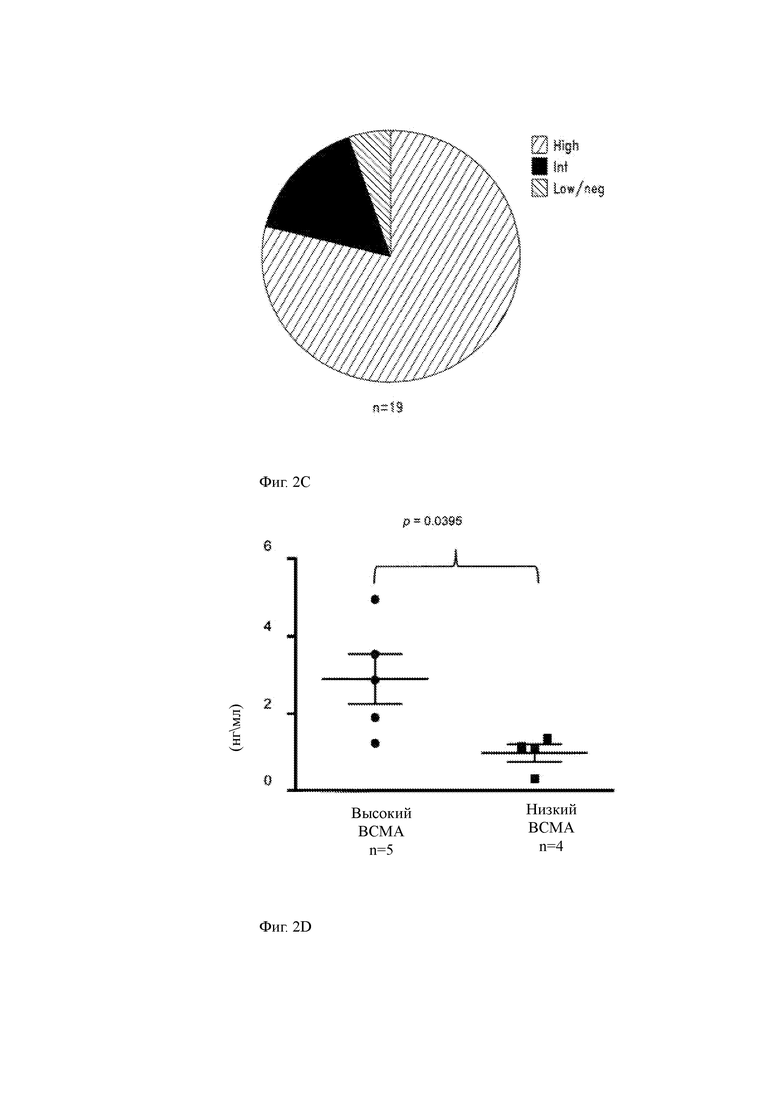
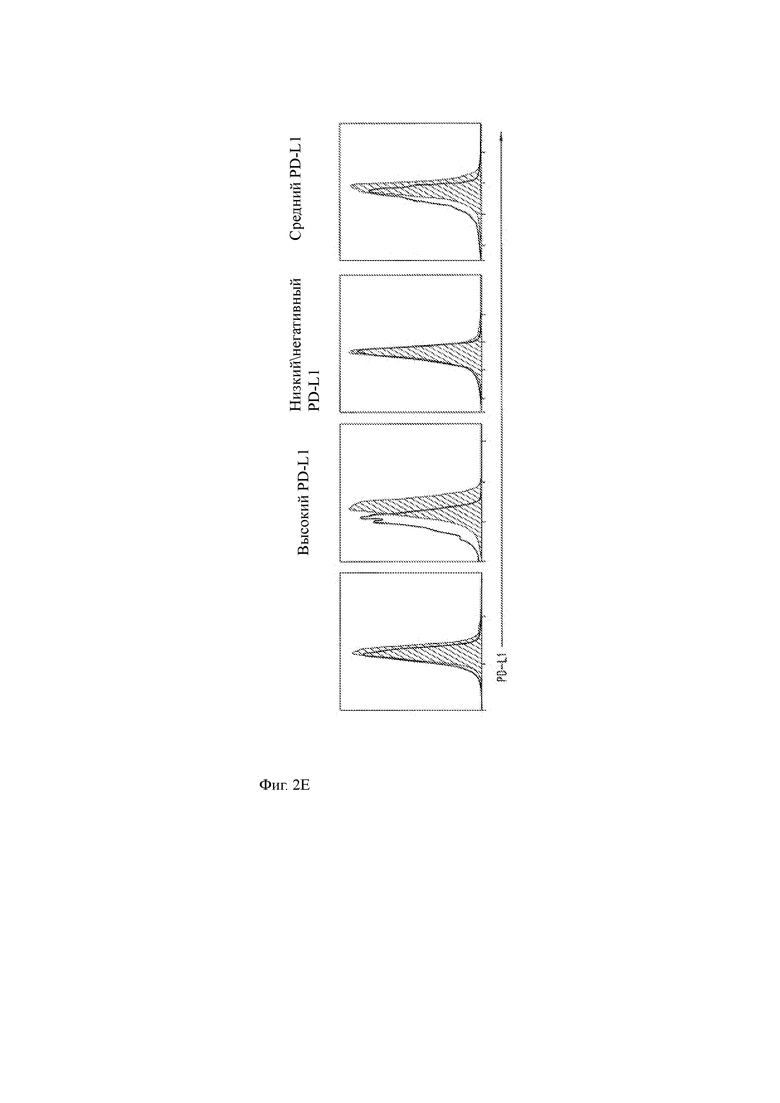
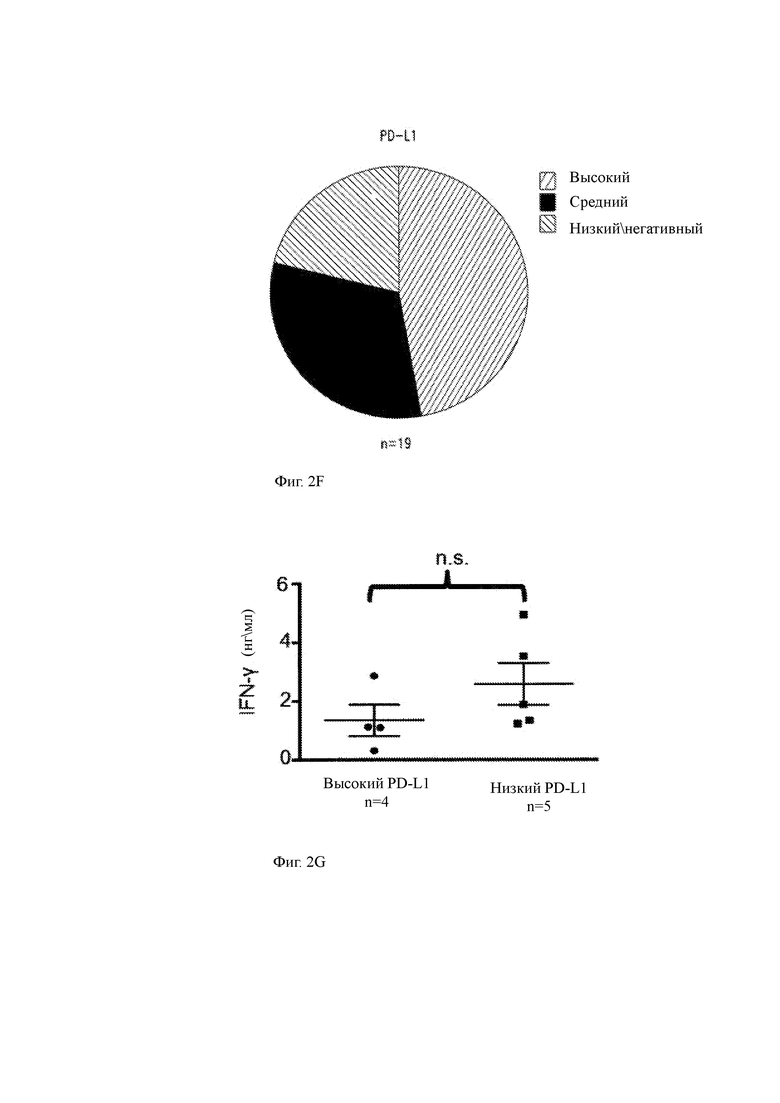
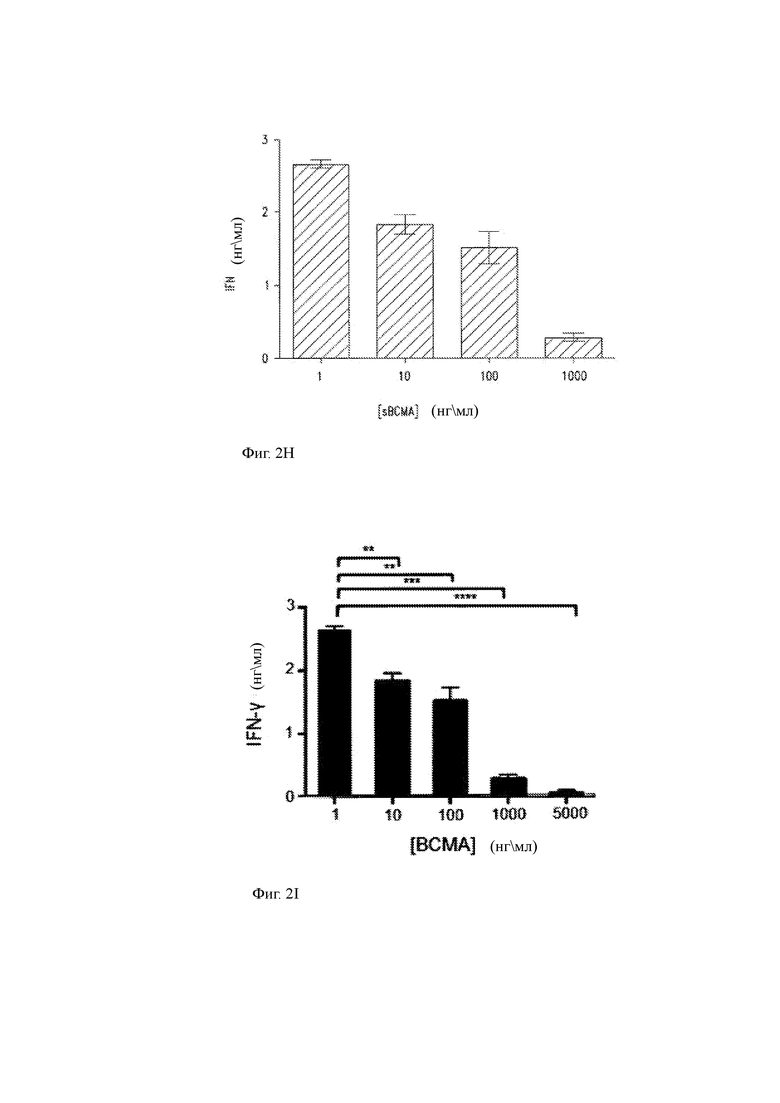
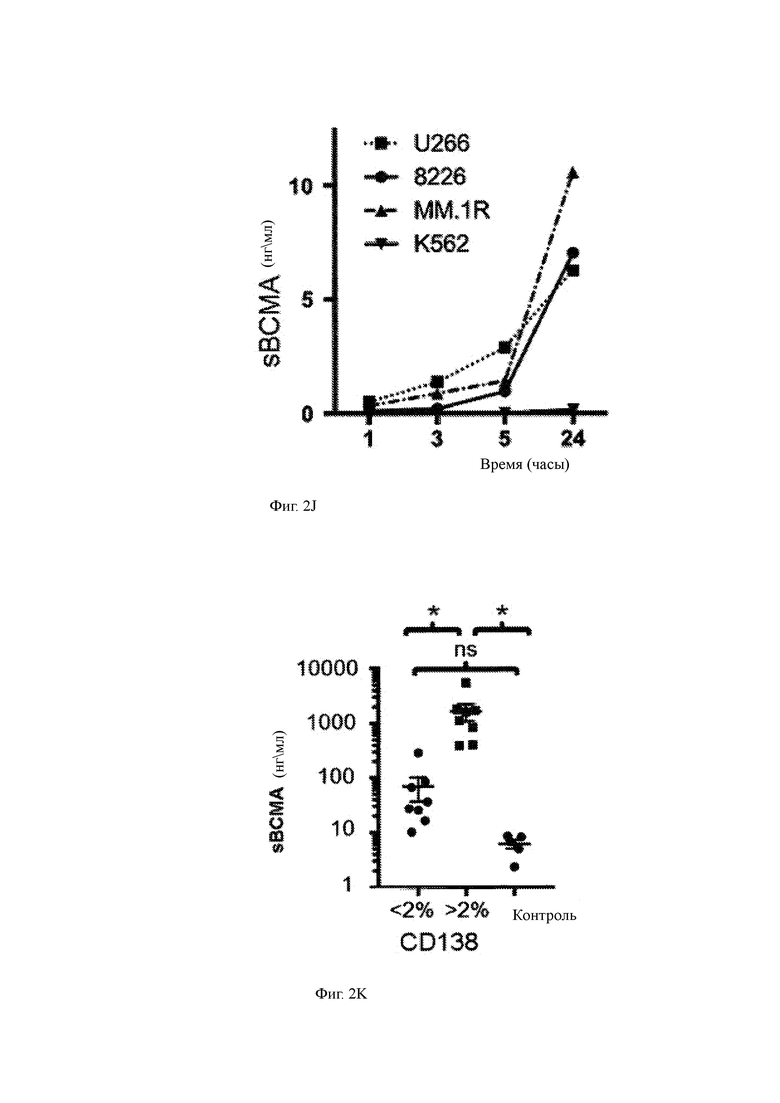
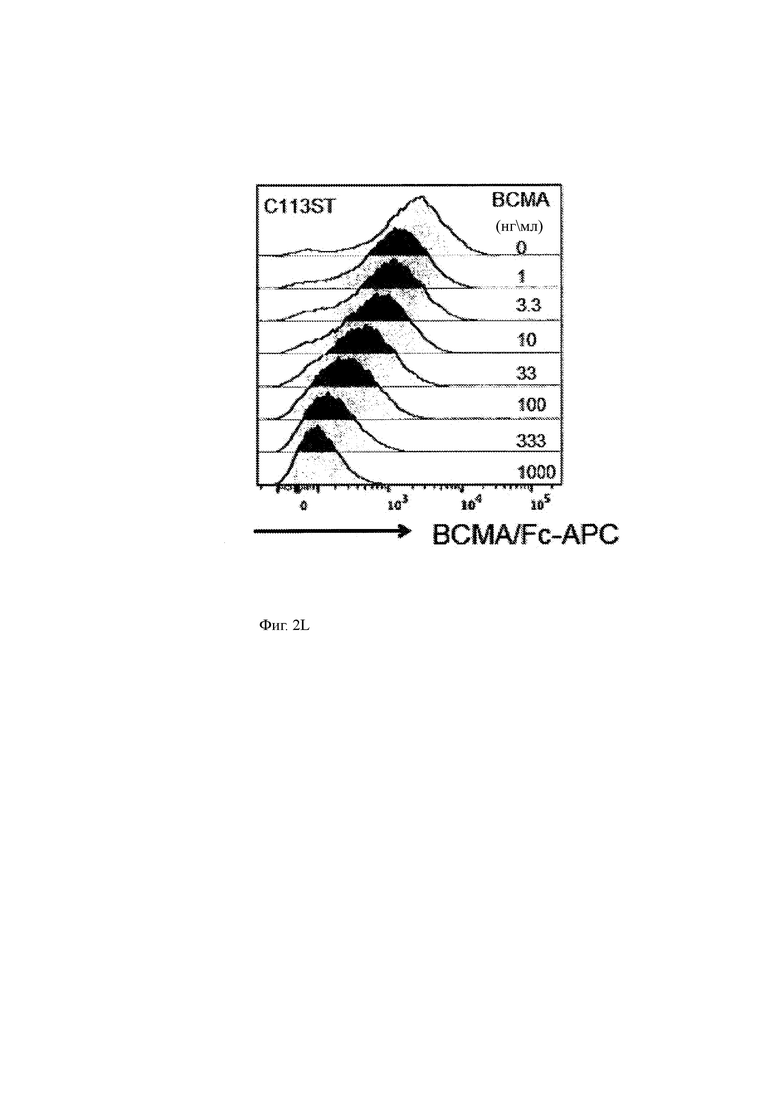
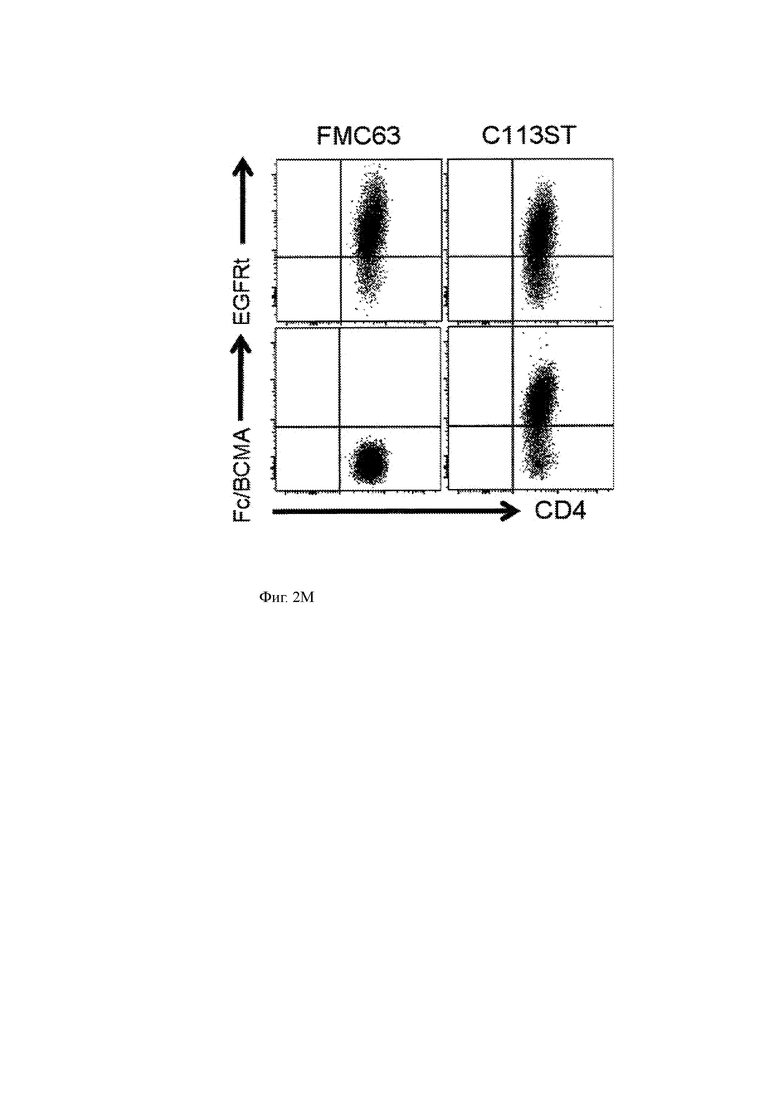
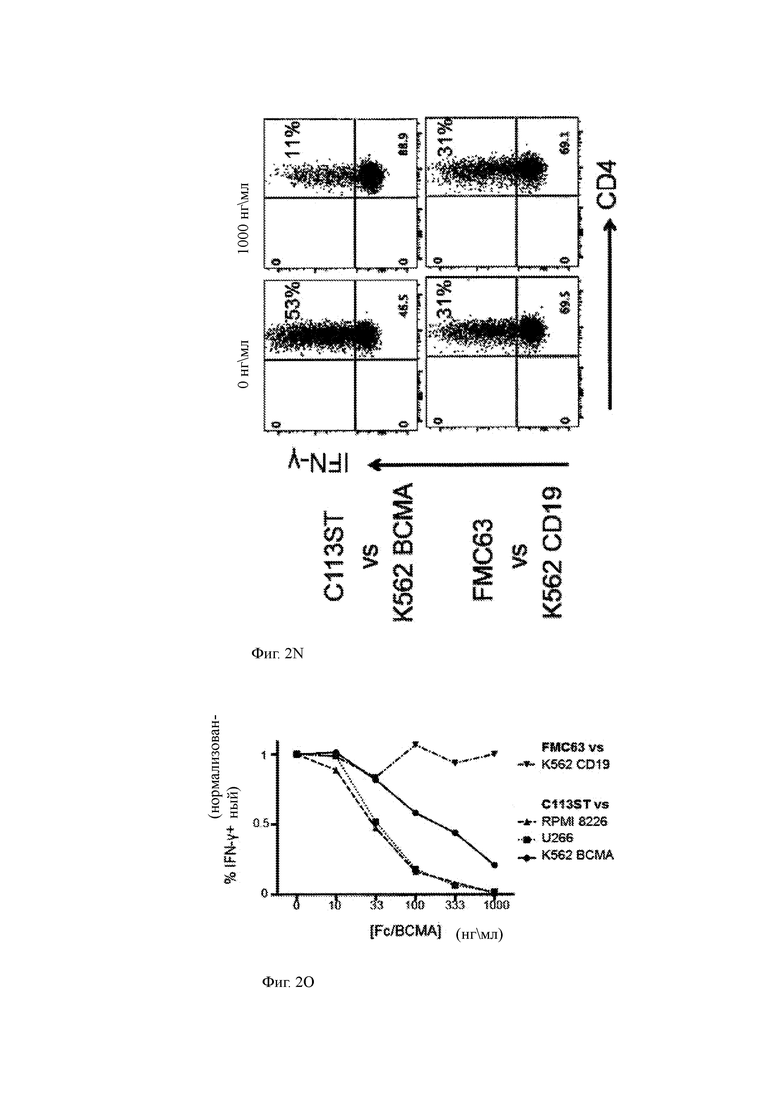
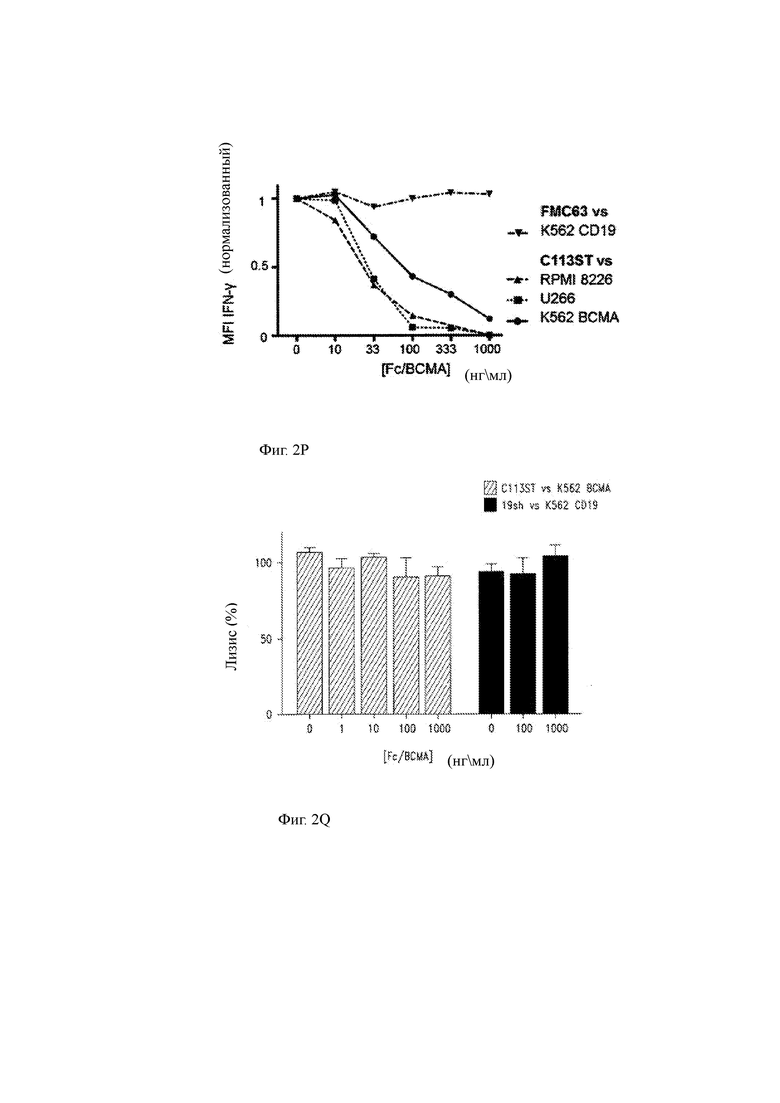
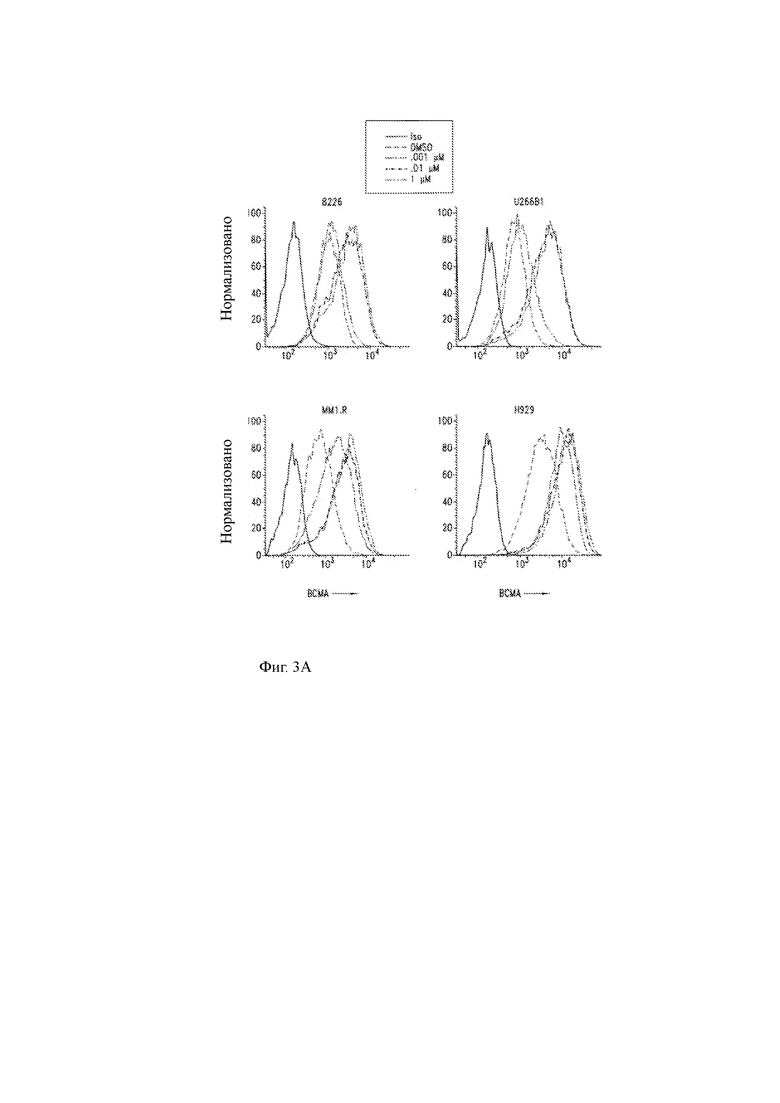
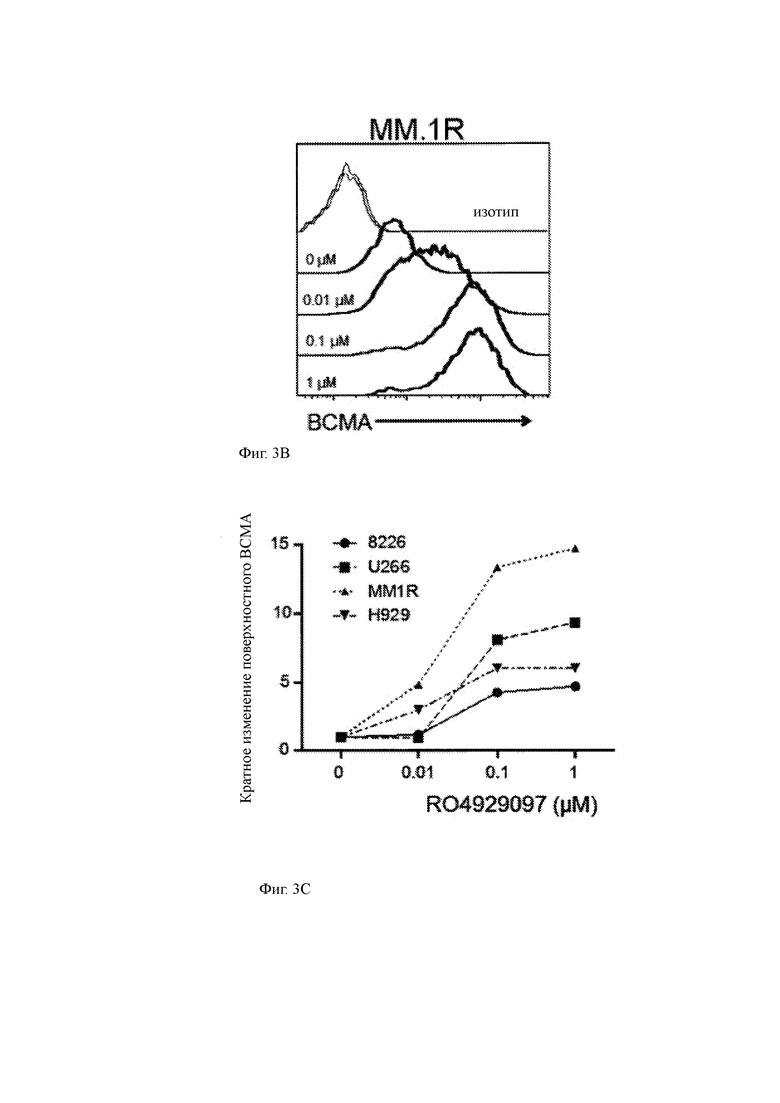
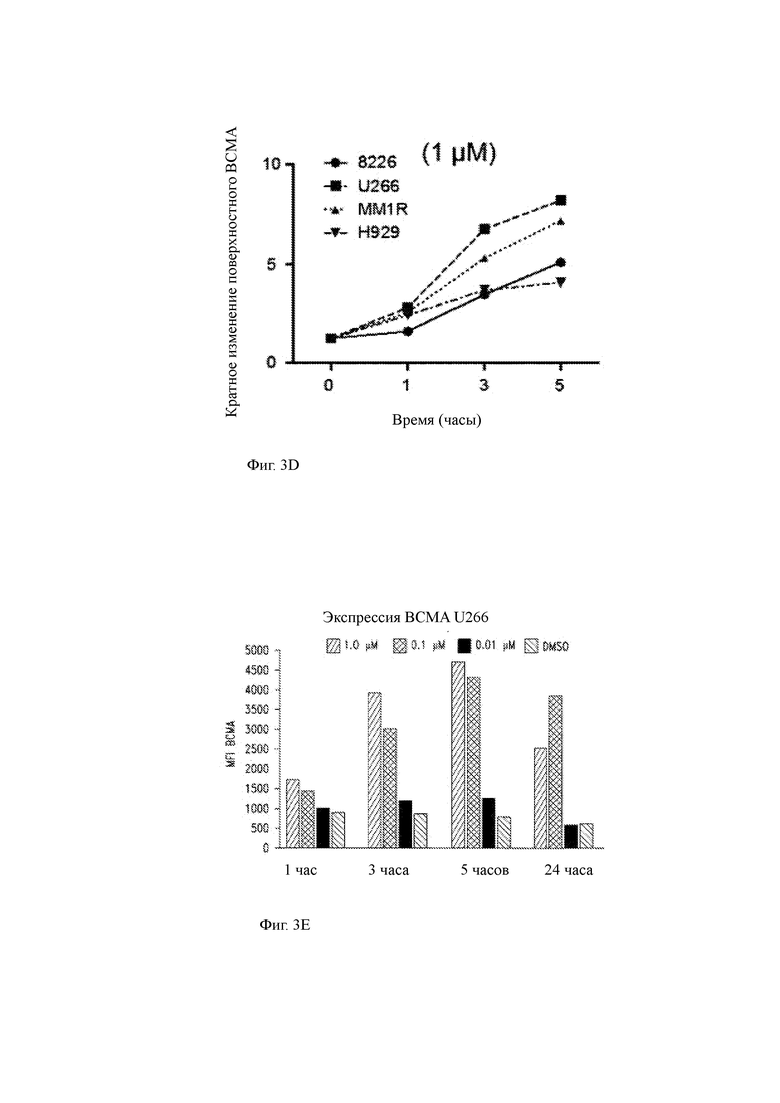
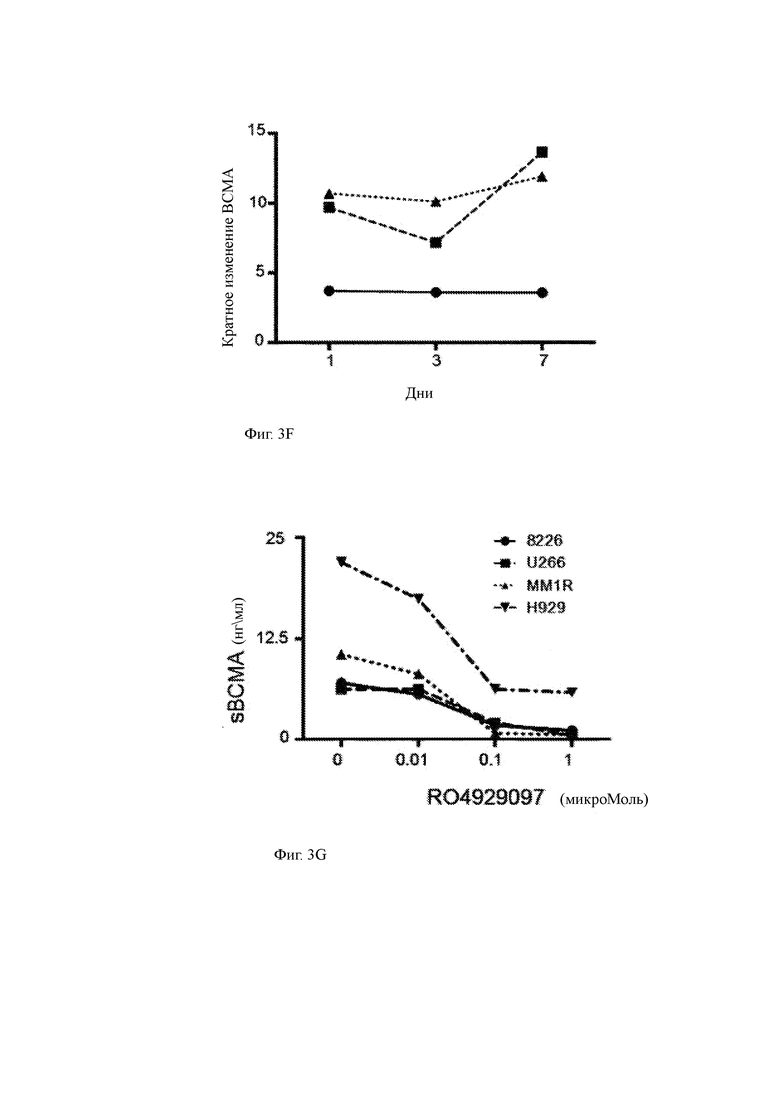
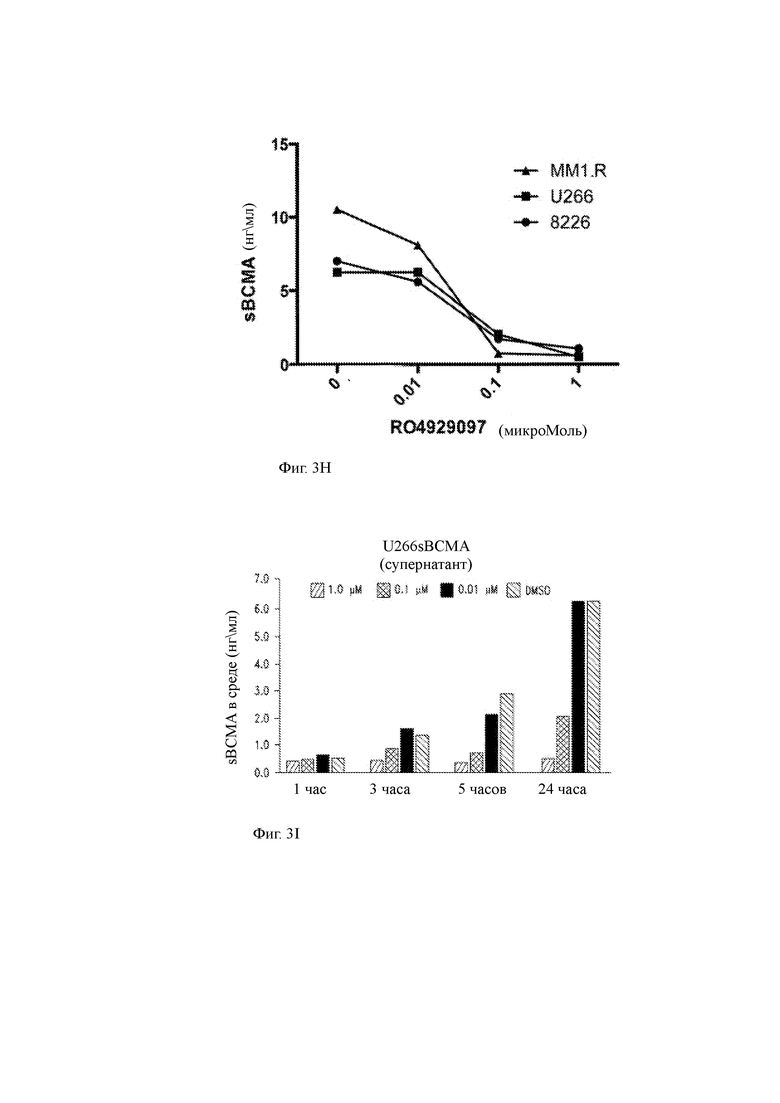
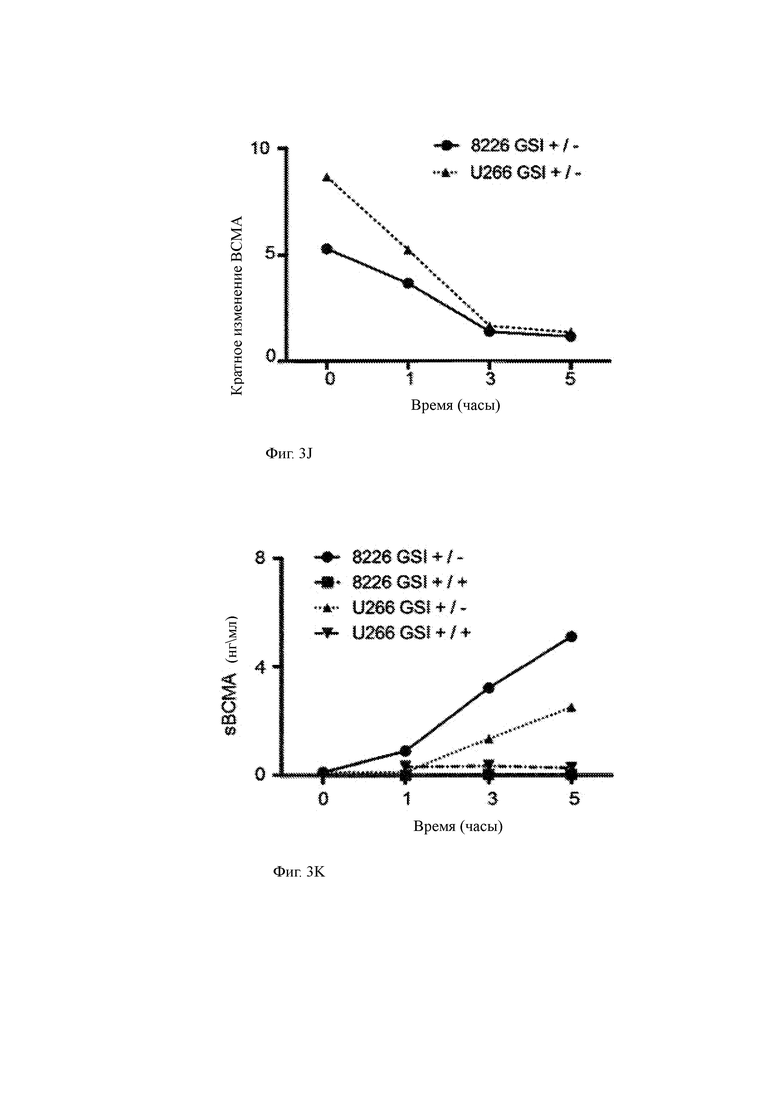
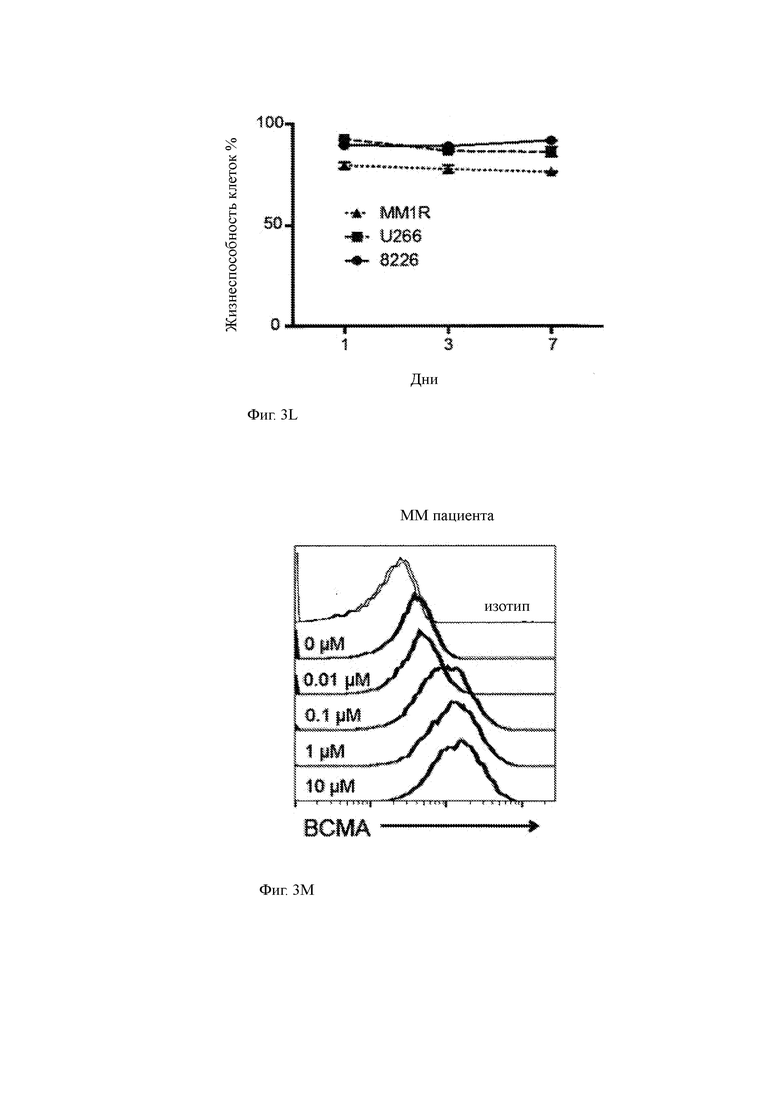
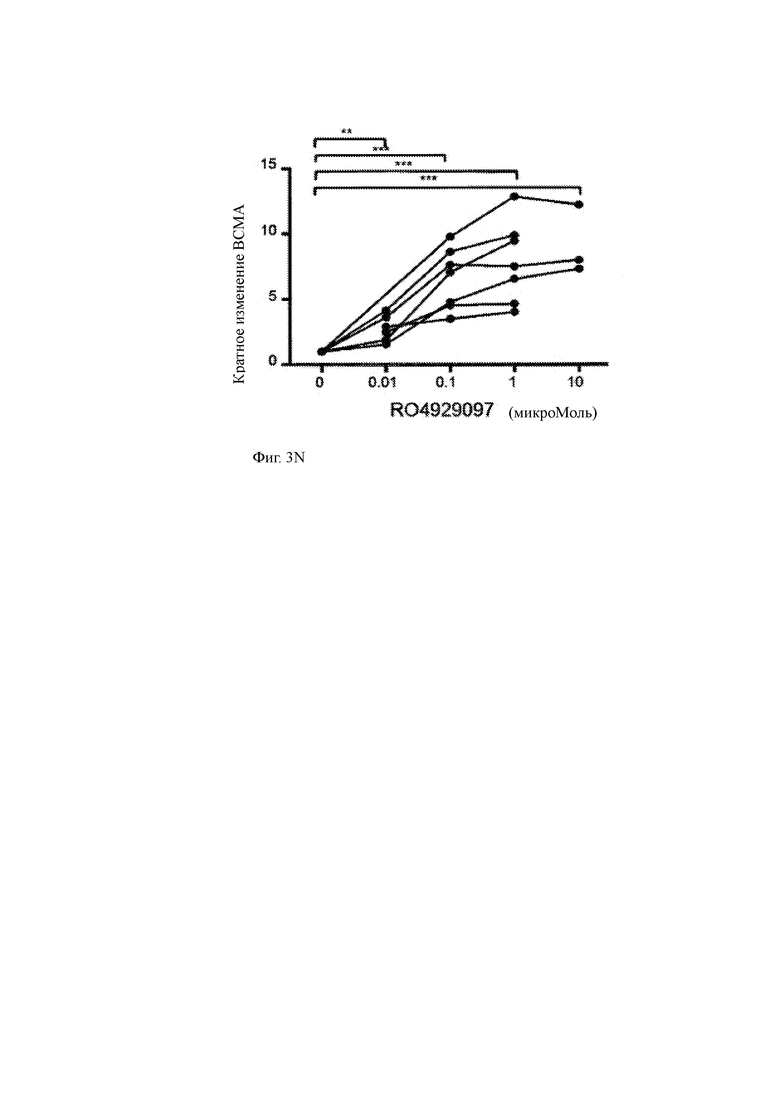

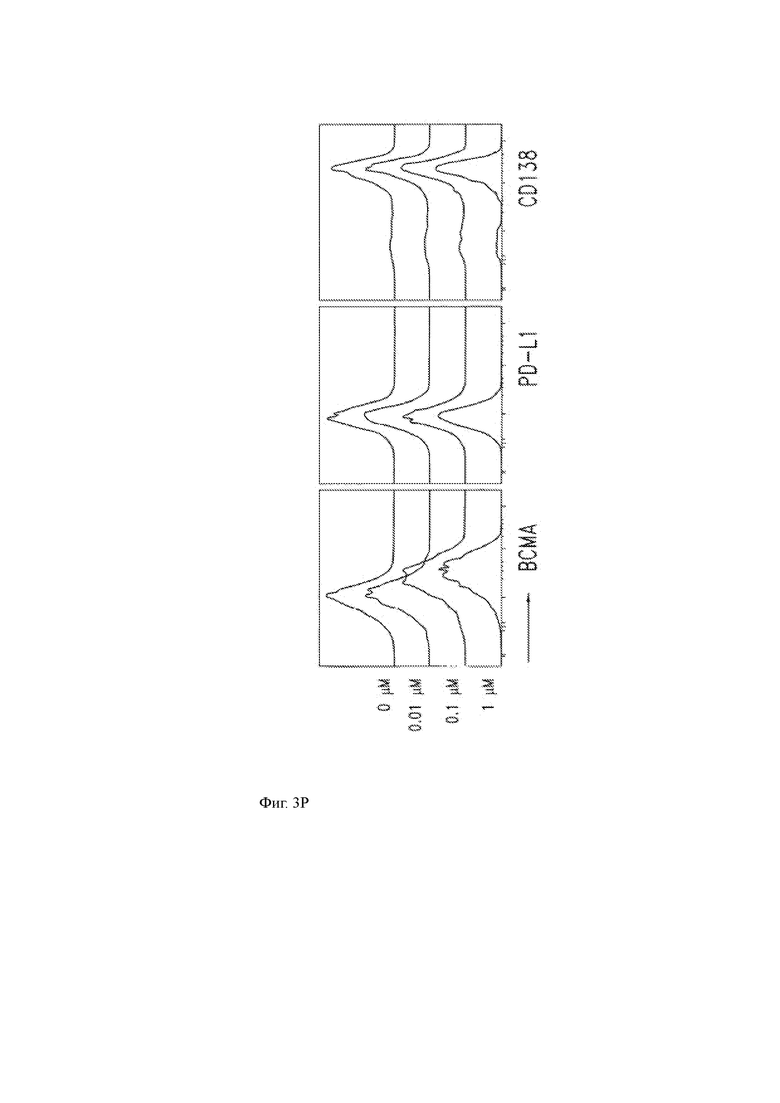
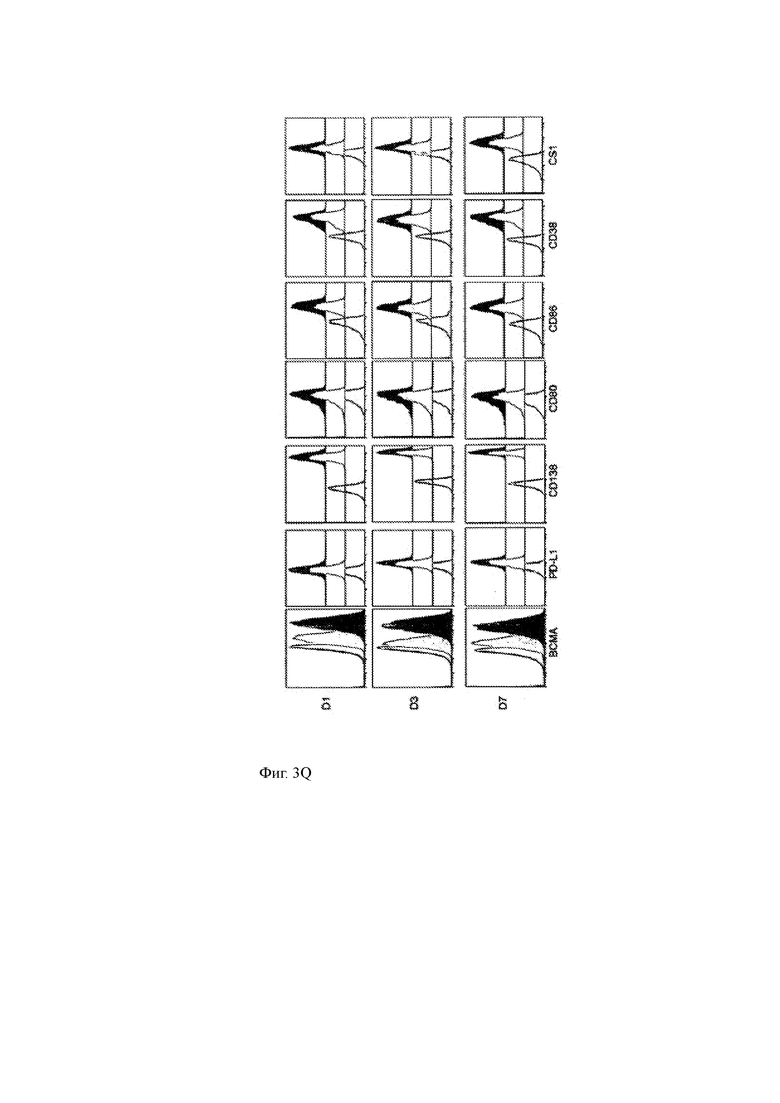
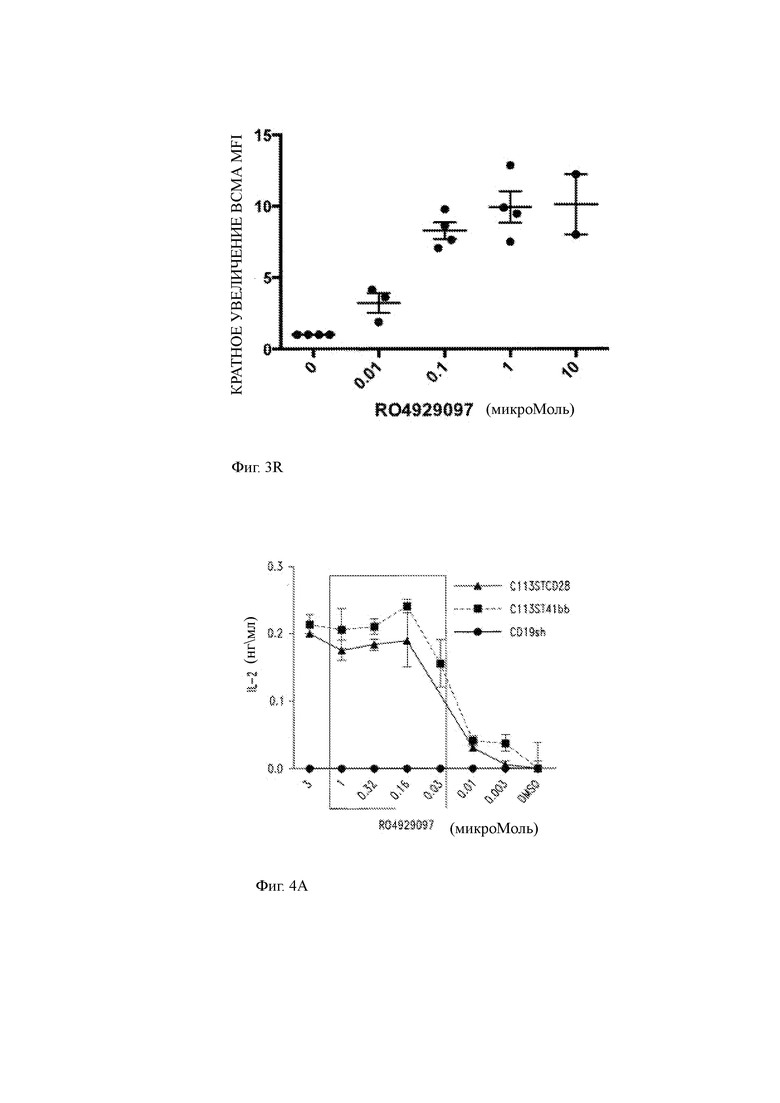
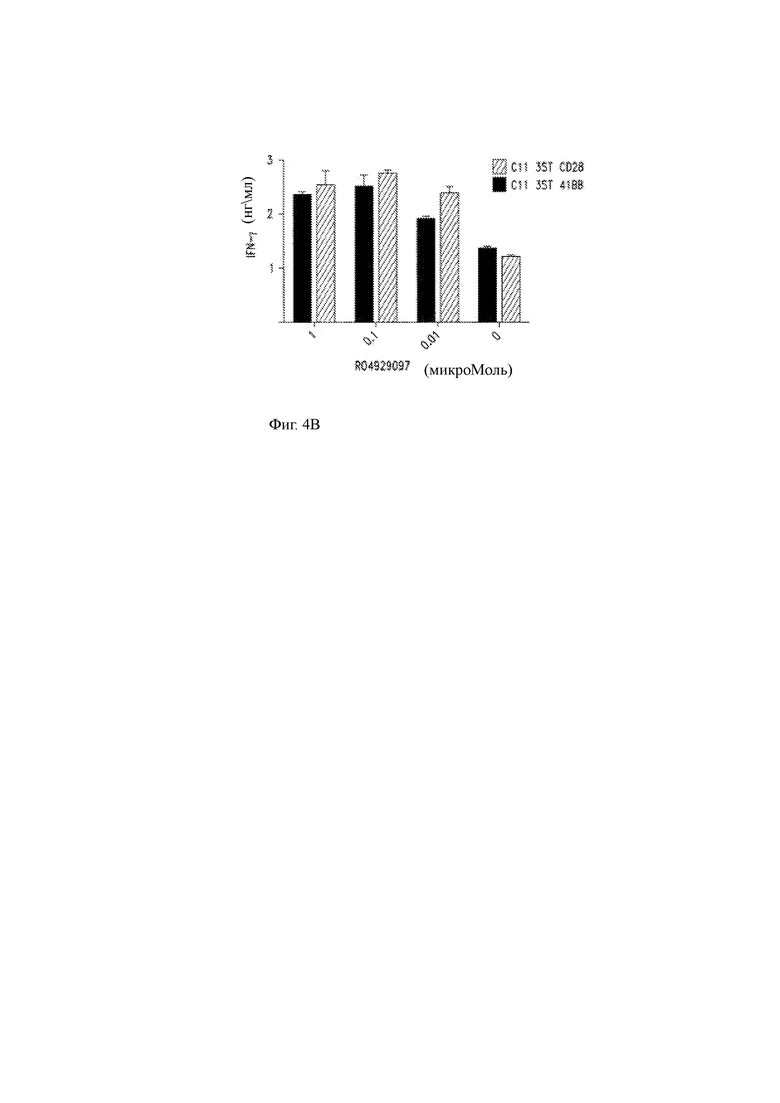
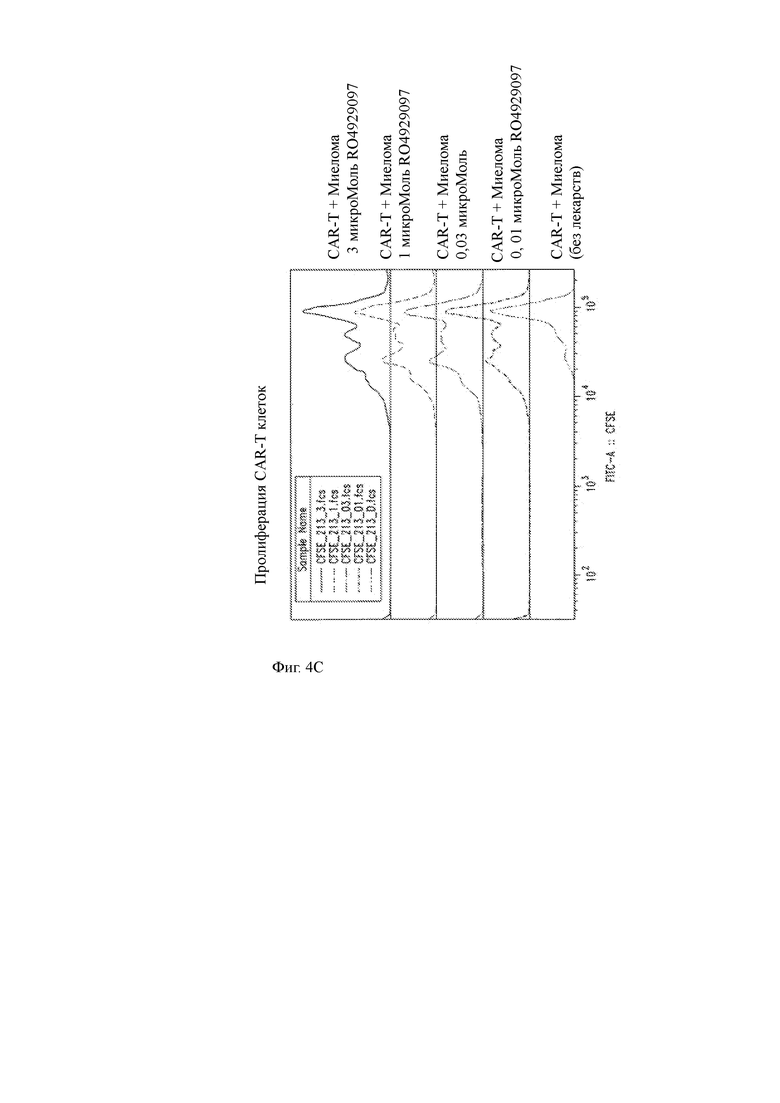
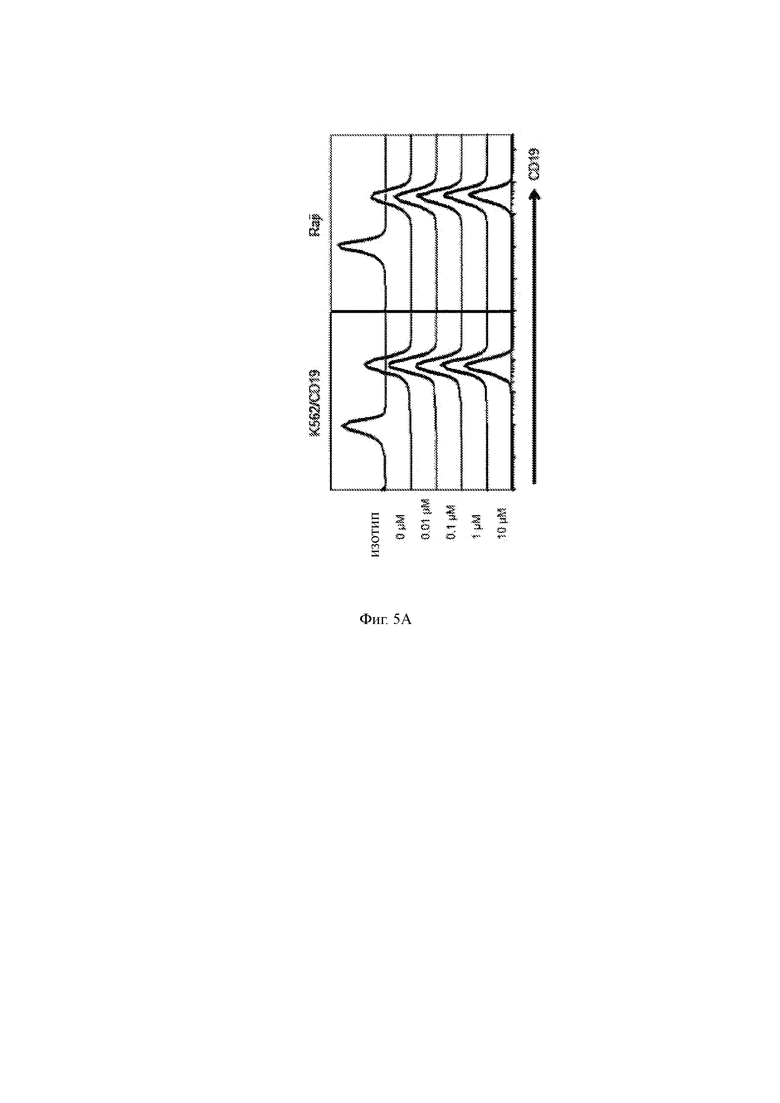
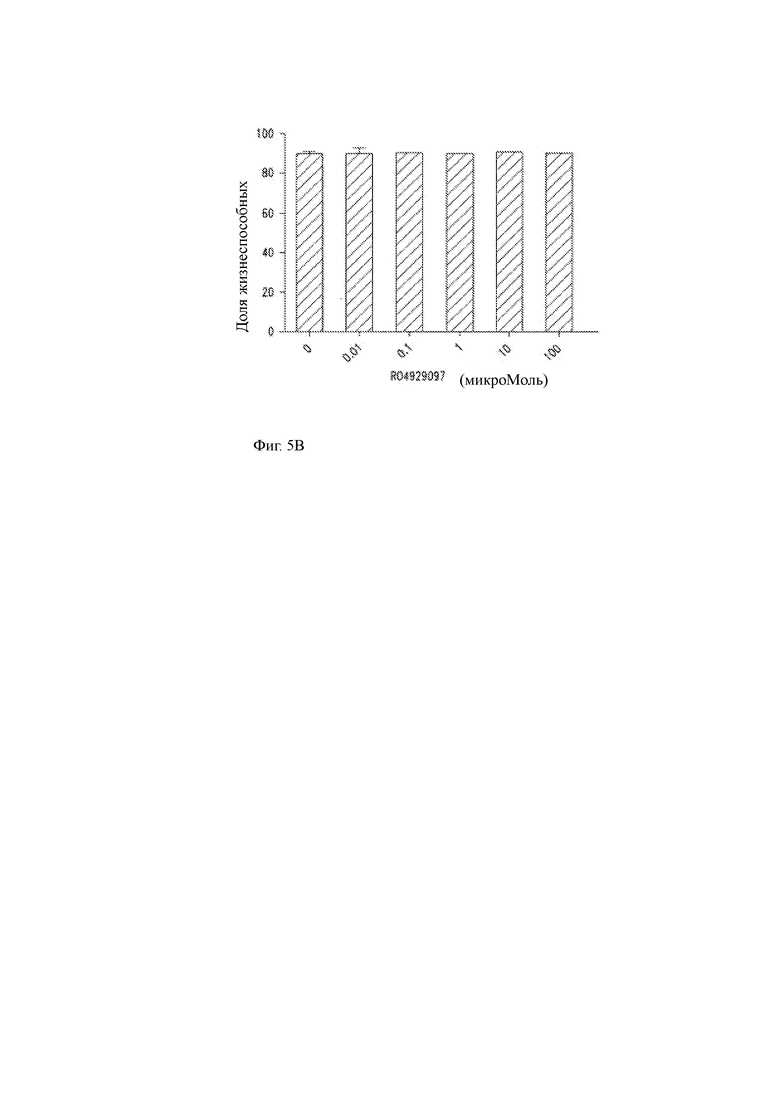
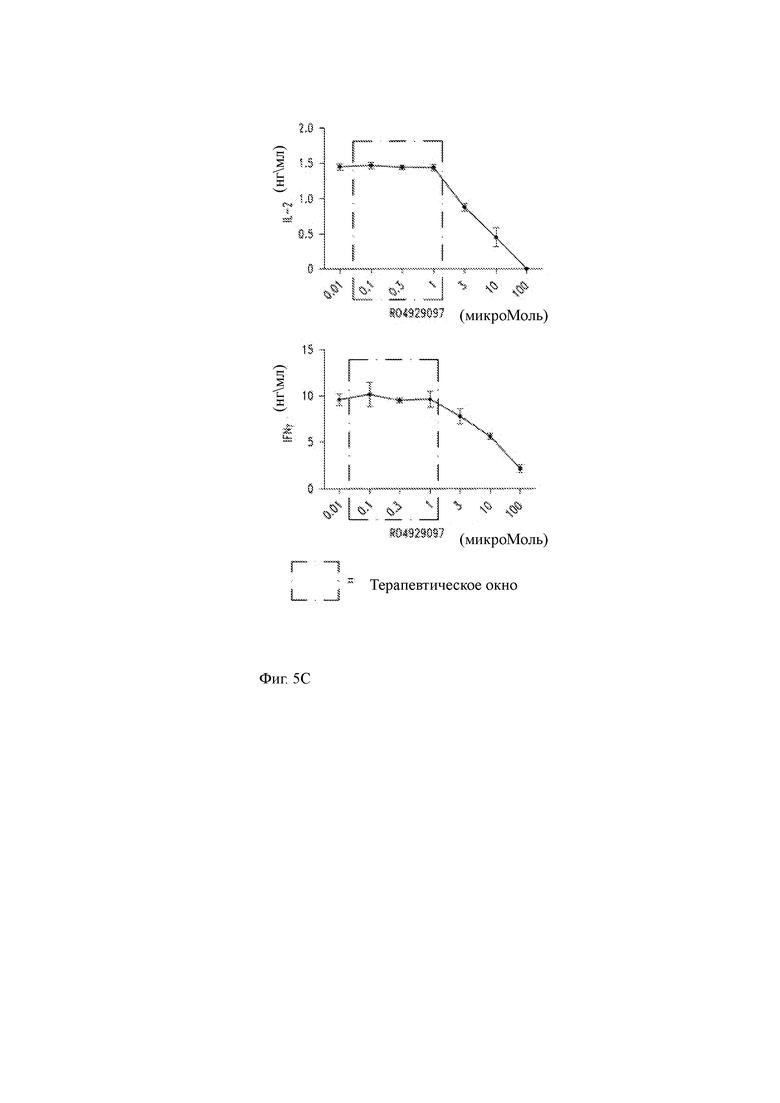
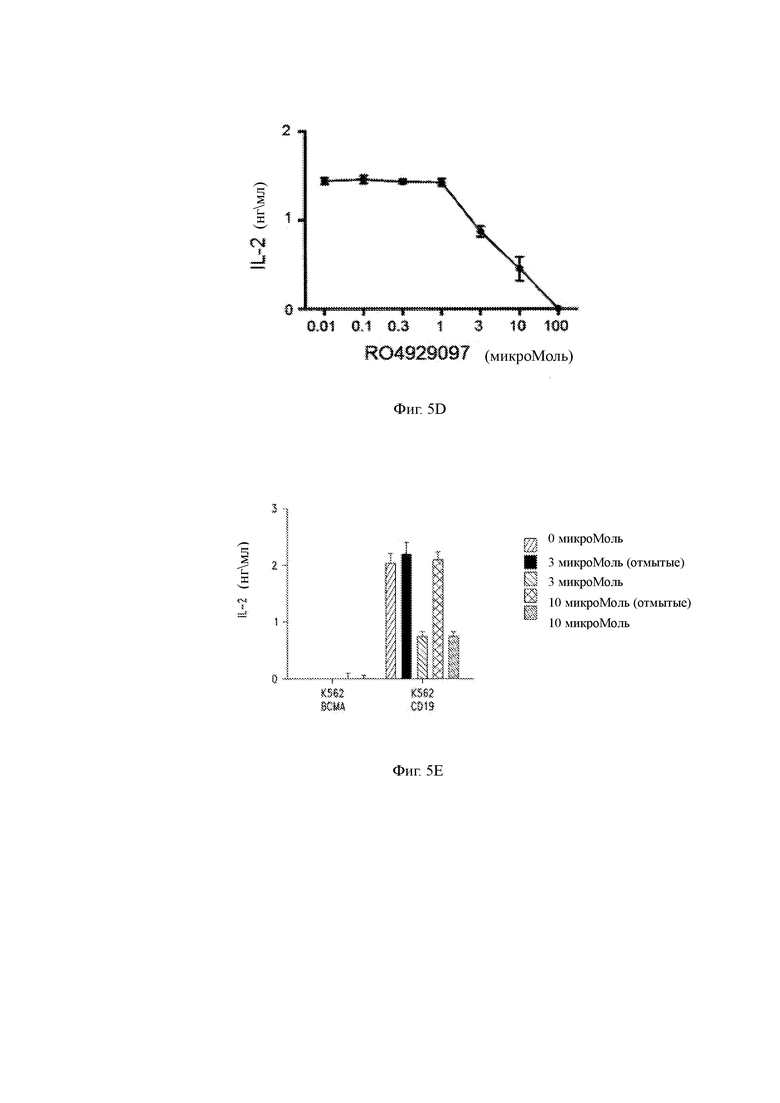
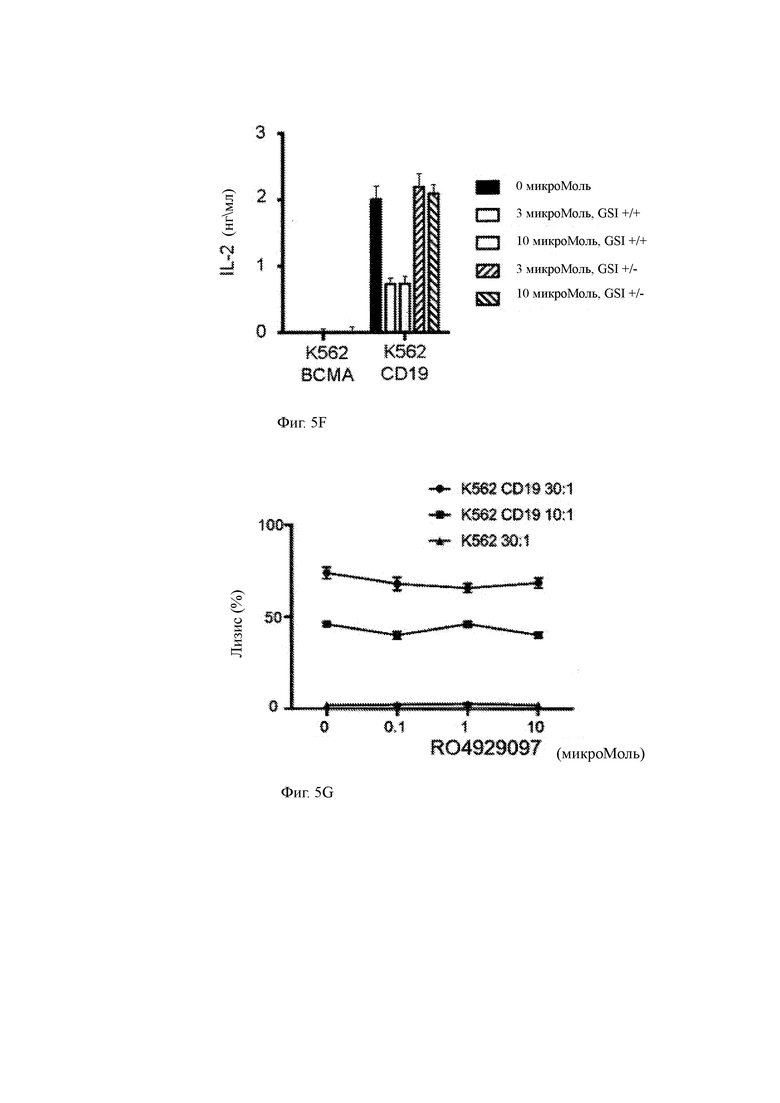
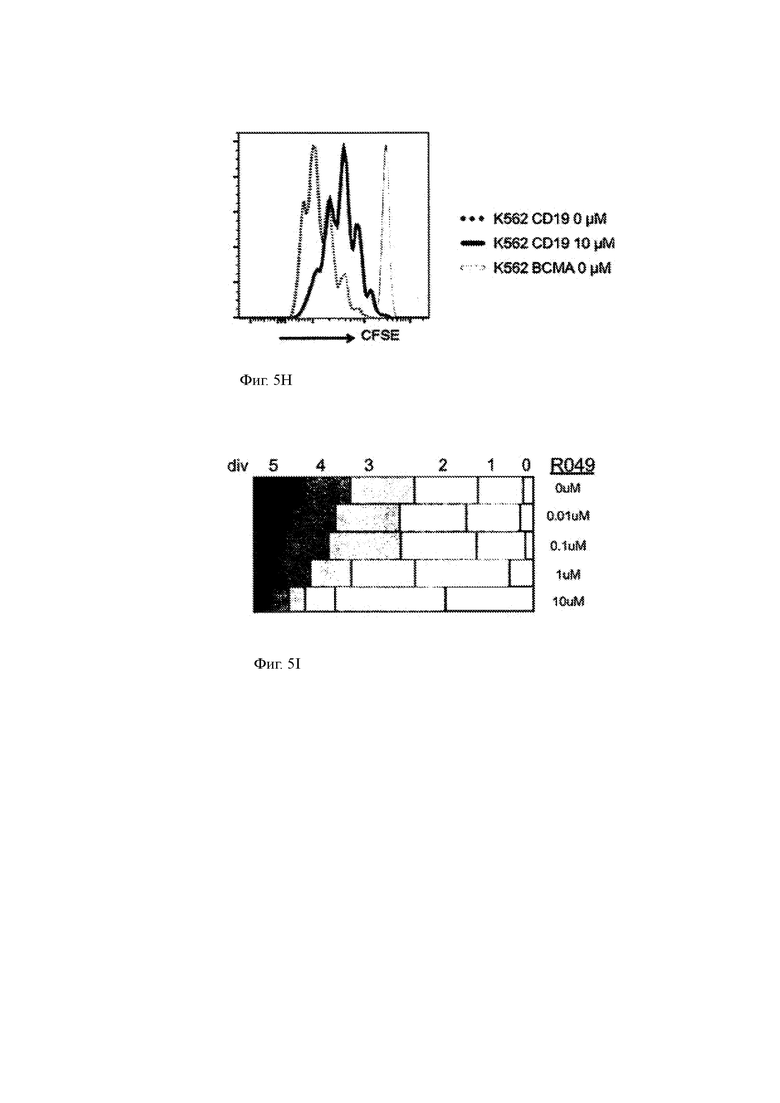
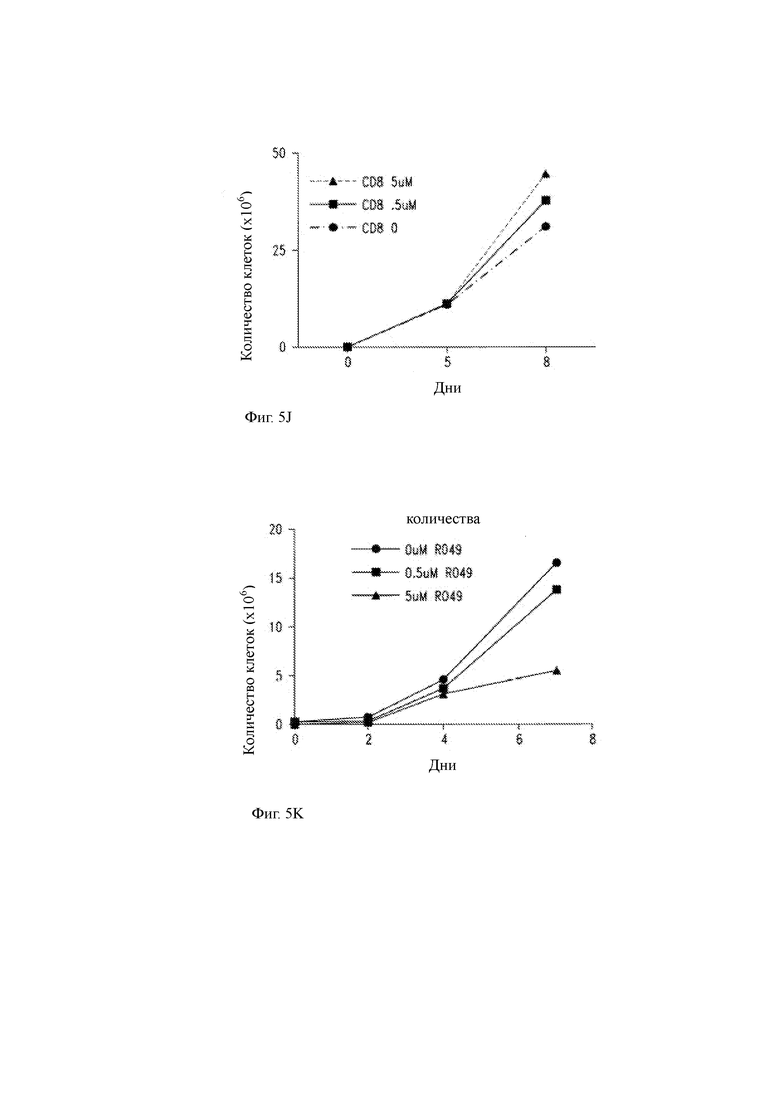
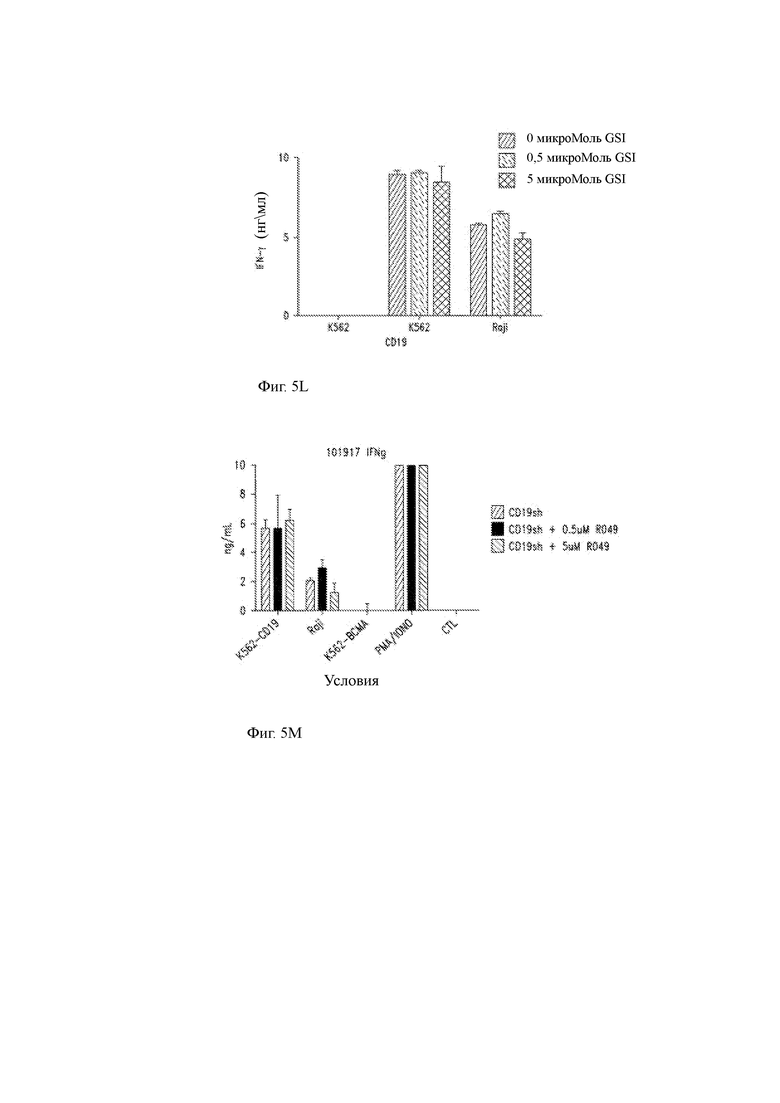
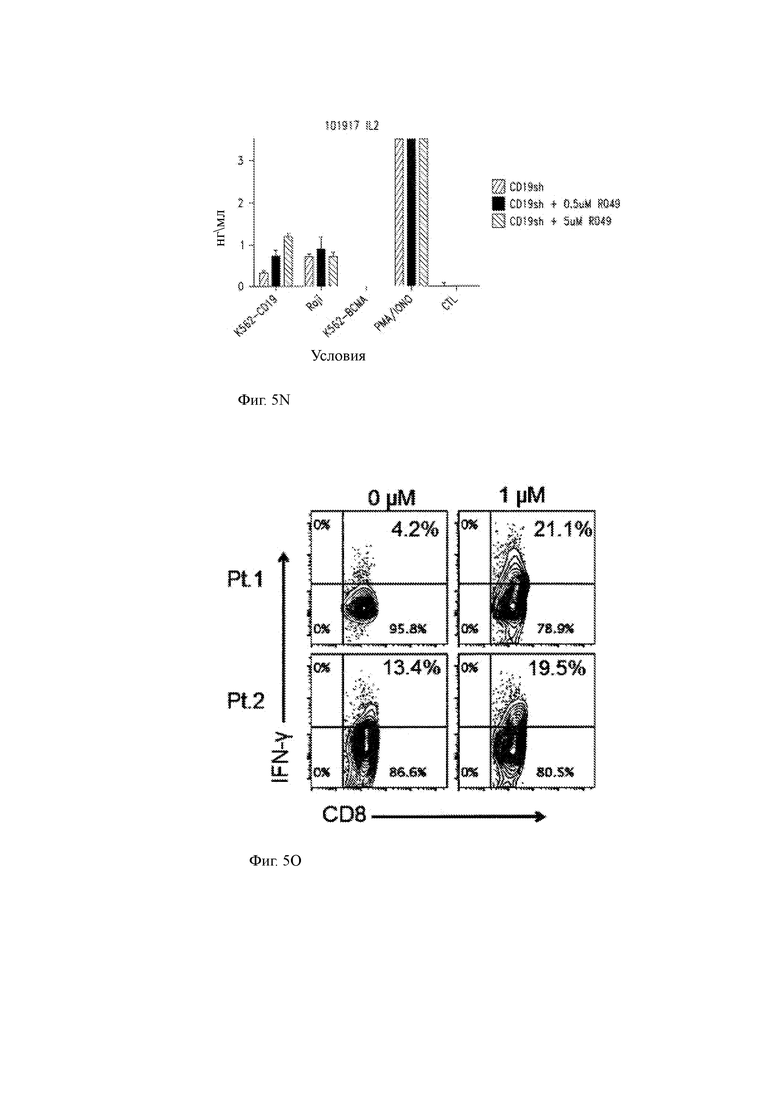
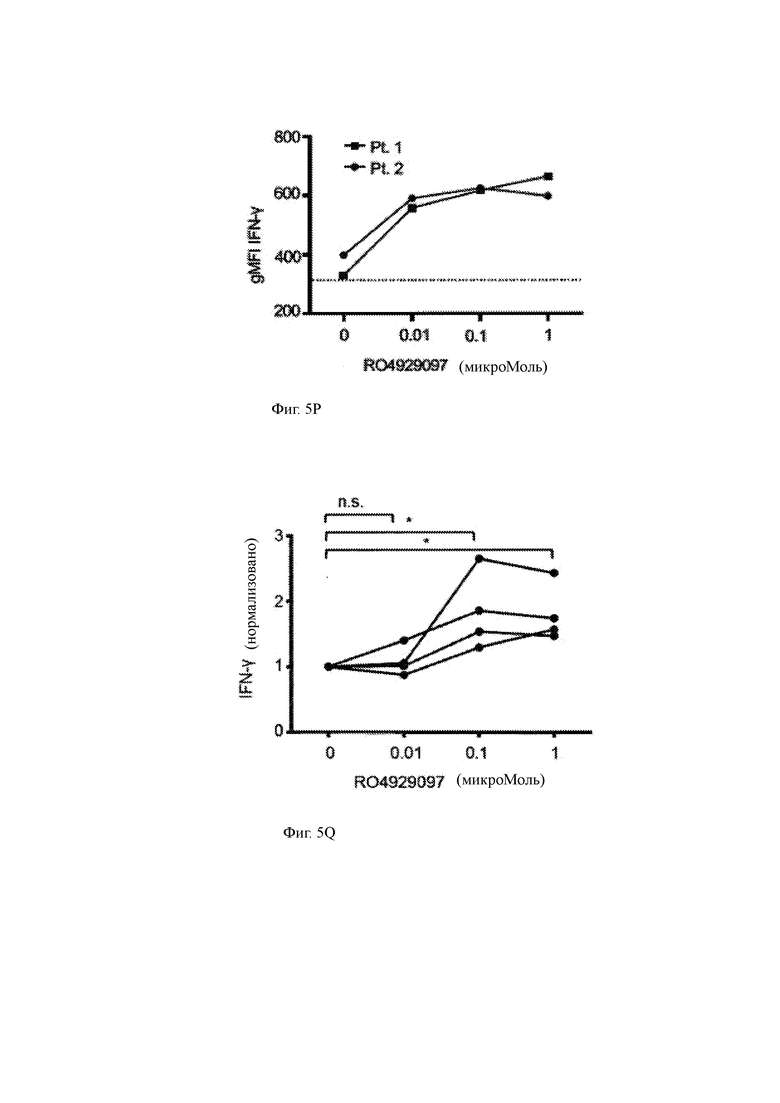
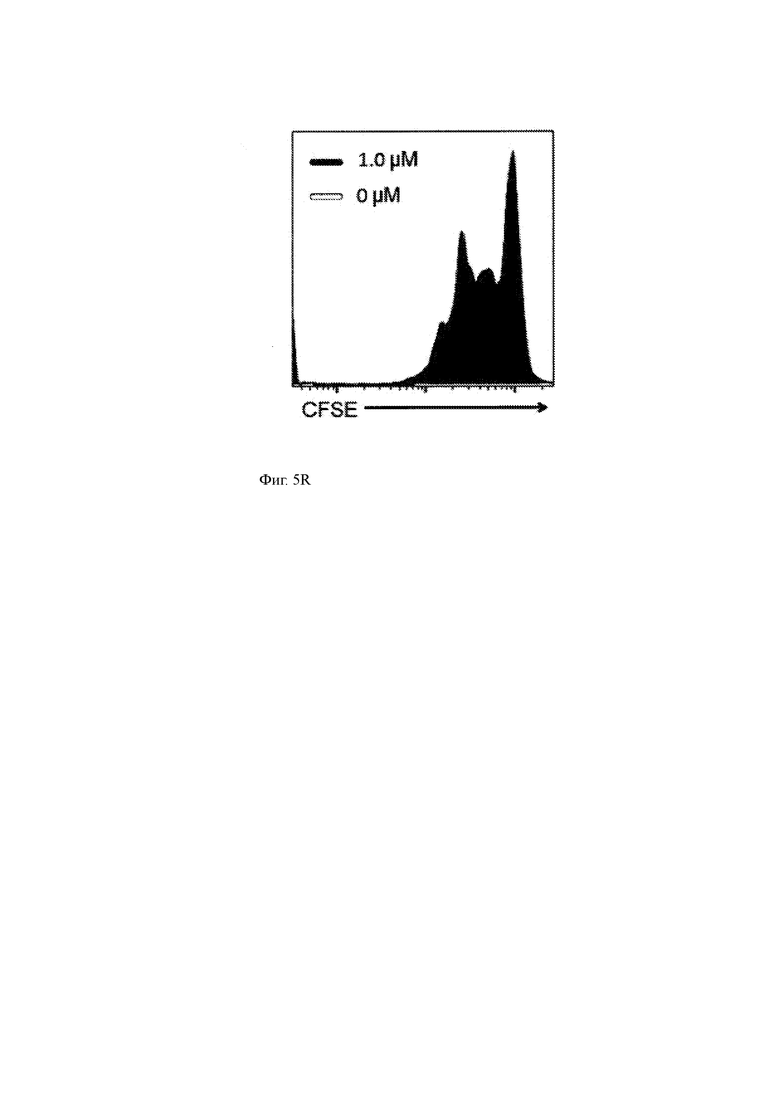
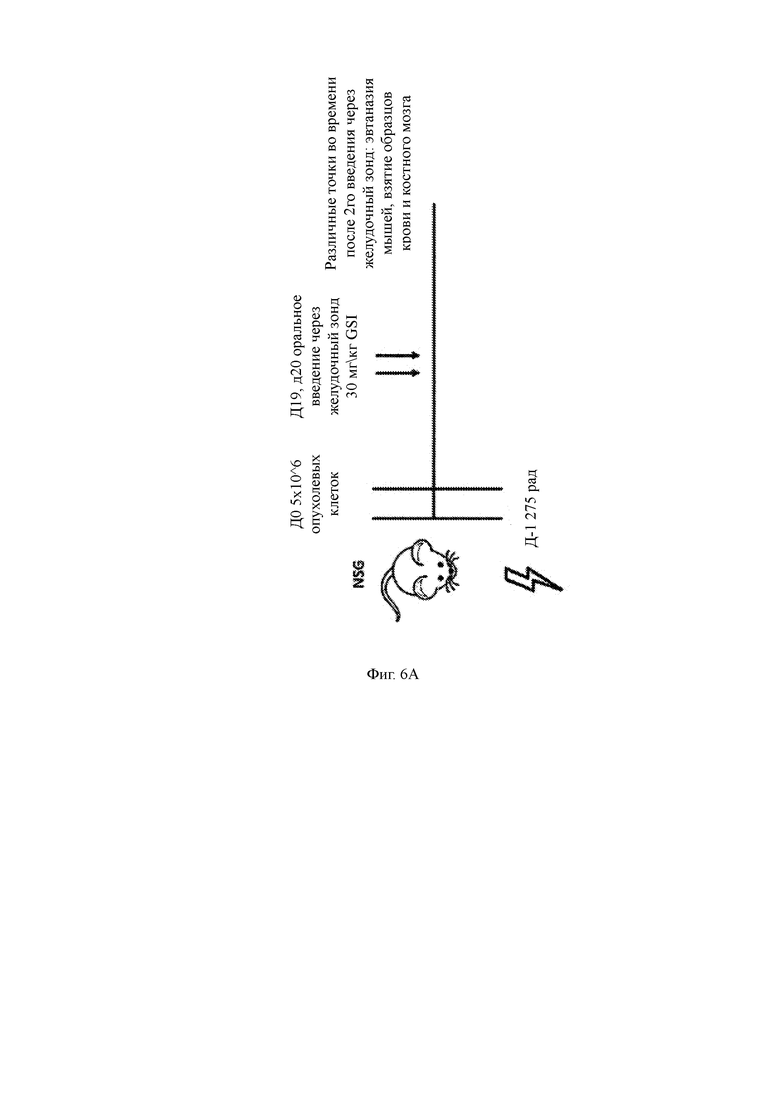
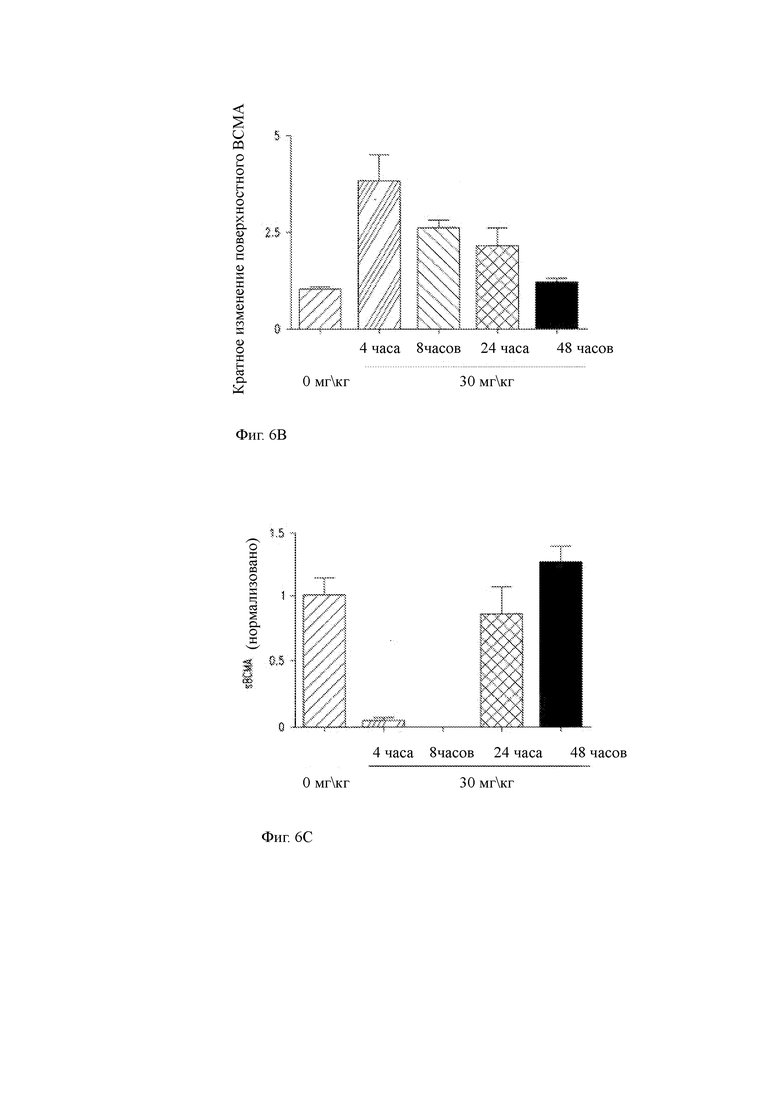
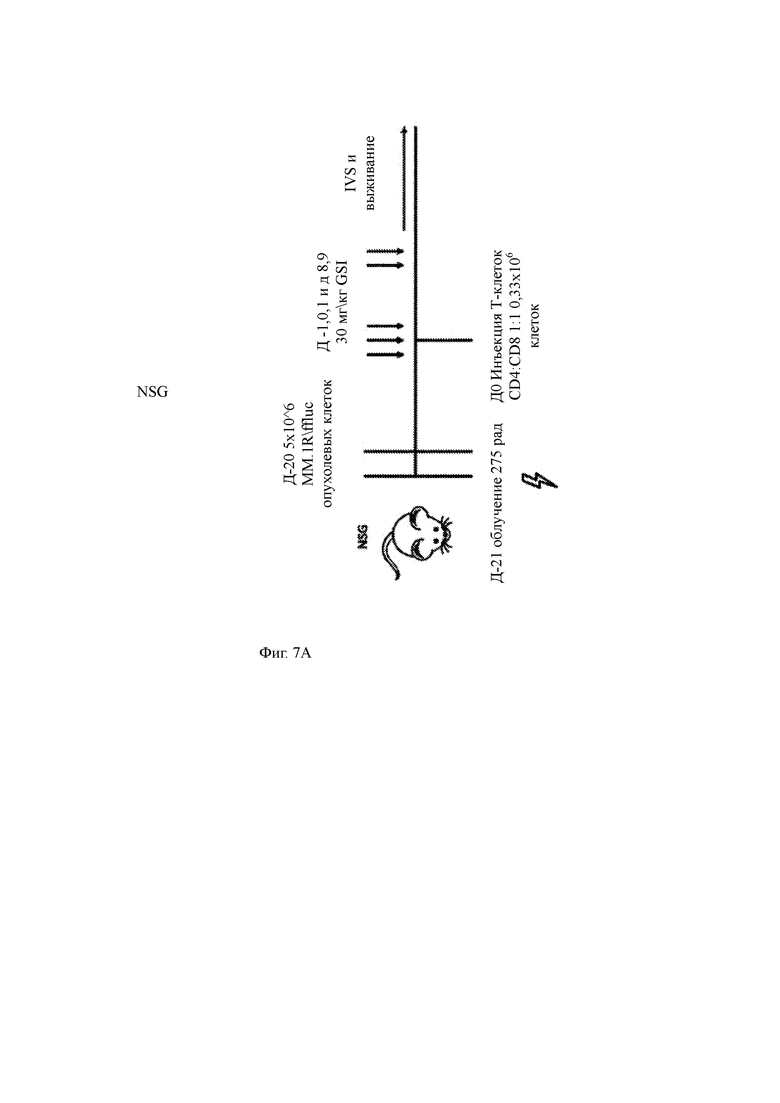
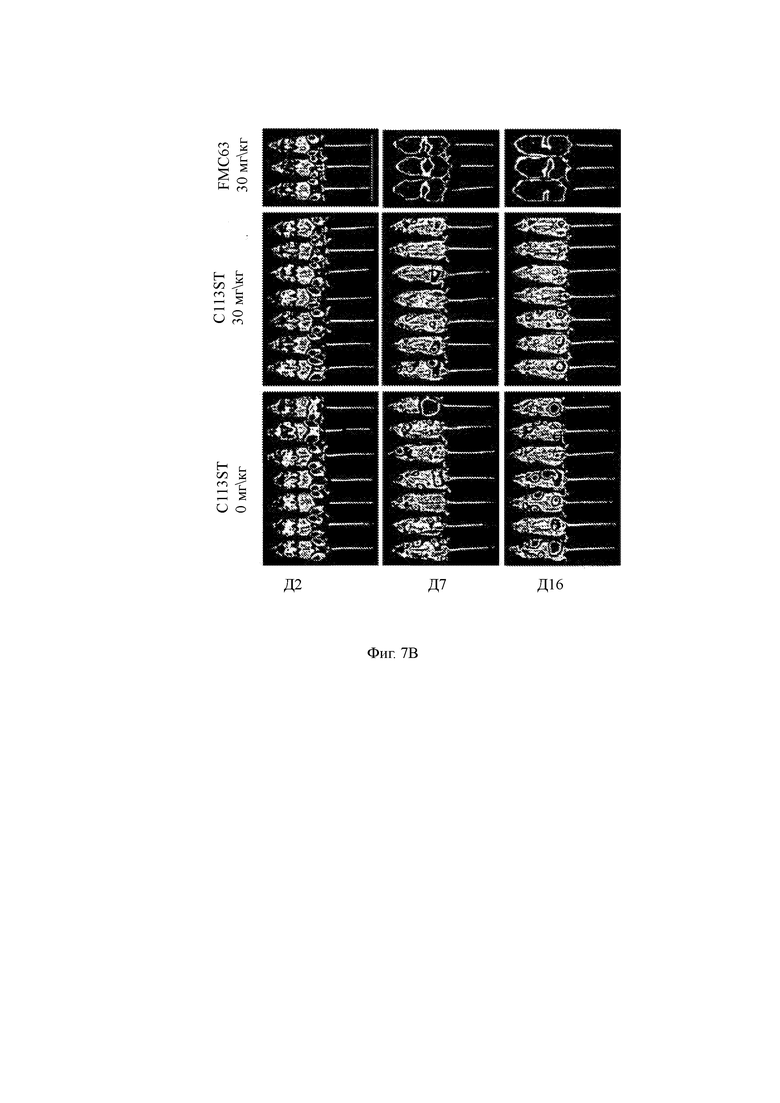
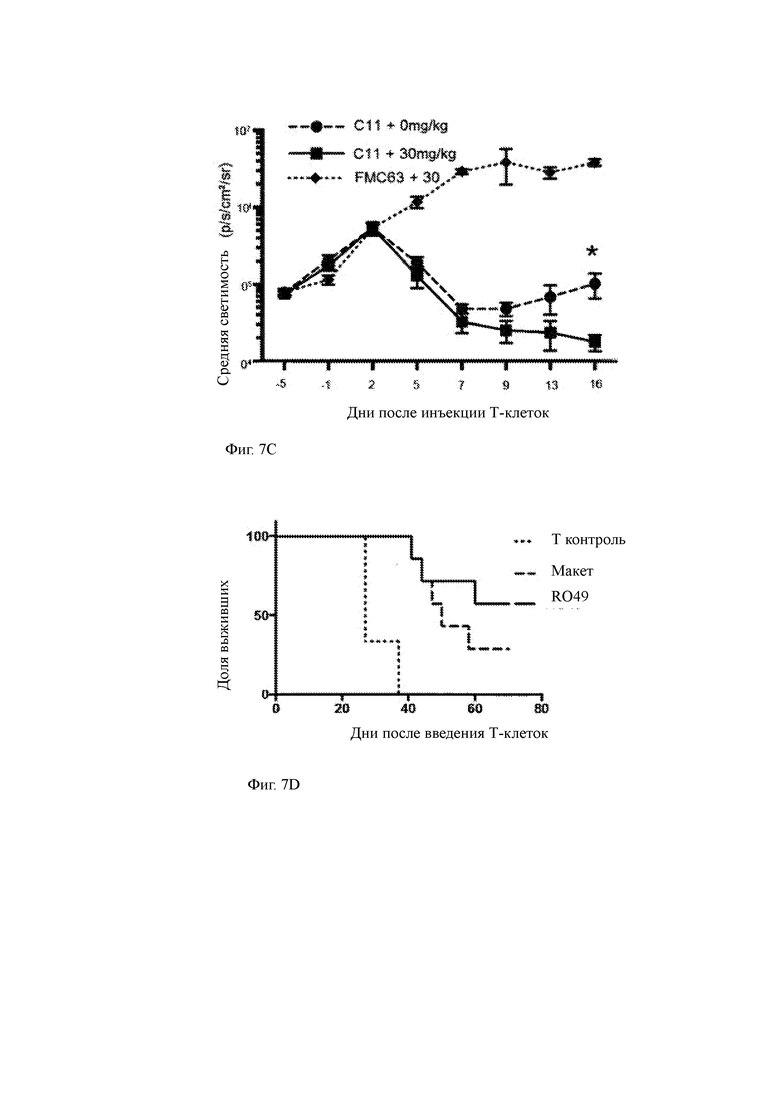
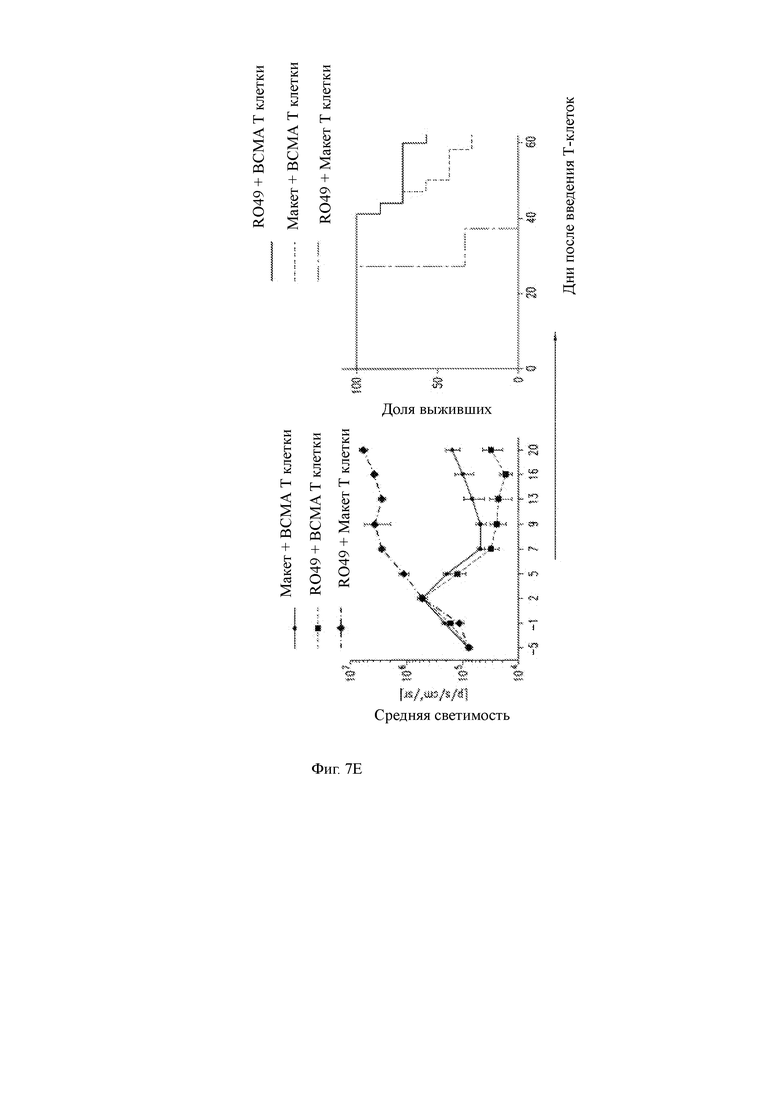
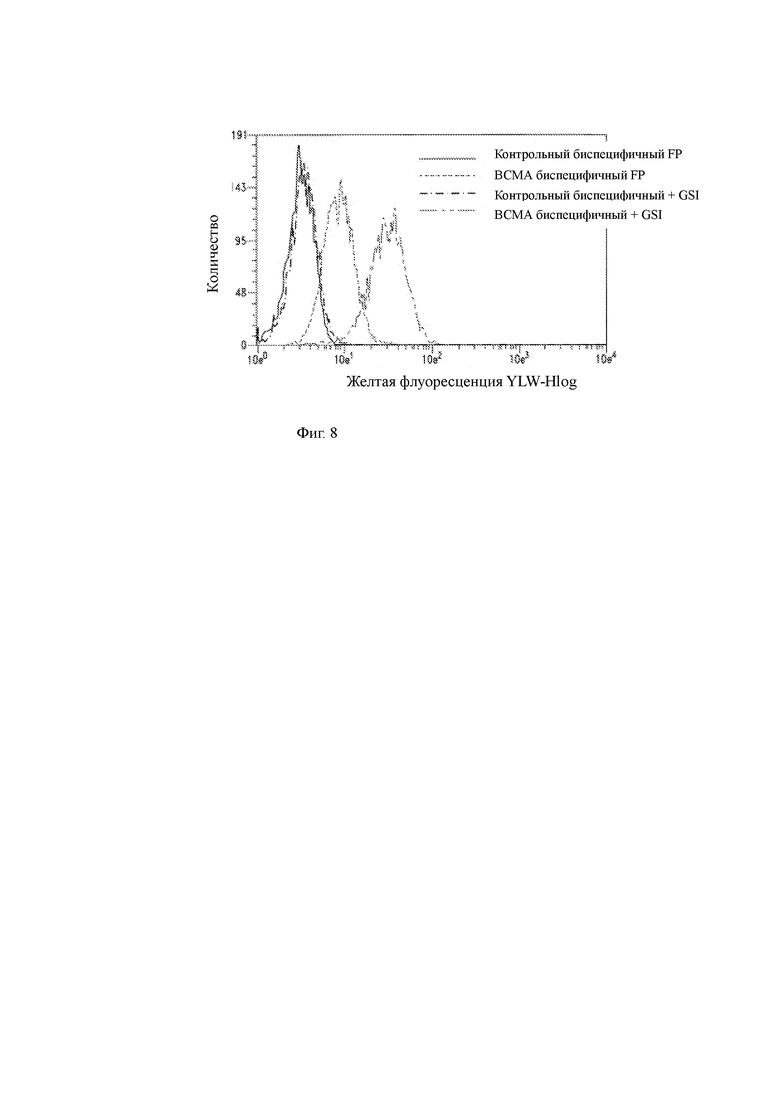
Группа изобретений относится к области фармацевтики и медицины. 1-3 объекты представляют собой способы лечения множественной миеломы у субъекта, включающие введение субъекту терапевтически эффективного количества Т-клетки, экспрессирующей ВСМА-специфичный химерный антигенный рецептор (CAR), и терапевтически эффективного количества ингибитора γ-секретазы. 4 объект – применение Т-клетки, экспрессирующей ВСМА-специфичный химерный антигенный рецептор (CAR), с ингибитором γ-секретазы для лечения множественной миеломы у субъекта. 5 объект – набор для лечения множественной миеломы, включающий Т-клетки, экспрессирующие ВСМА-специфичный химерный антигенный рецептор (CAR), и ингибитор γ-секретазы. Технический результат заключается в эффективности лечения множественной миеломы путем комбинирования Т-клеток, экспрессирующих ВСМА-специфичный CAR, с ингибитором γ-секретазы. 5 н. и 63 з.п. ф-лы, 8 ил., 8 пр.
1. Способ лечения множественной миеломы у субъекта, включающий введение субъекту (i) терапевтически эффективного количества Т-клетки, экспрессирующей ВСМА-специфичный химерный антигенный рецептор (CAR), и (ii) терапевтически эффективного количества ингибитора γ-секретазы, причем:
(1) CAR содержит ВСМА-специфичный scFv, ВСМА-специфичный scTCR, ВСМА-специфичное нанотело, ВСМА-специфичное доменное антитело или лиганд ВСМА или его связывающую часть; и/или
(2) CAR содержит гидрофобную часть, расположенную между внеклеточным компонентом и внутриклеточным компонентом, где внеклеточный компонент содержит ВСМА-специфичный связывающий домен и где гидрофобная часть представляет собой трансмембранный домен CD4, CD8, CD28 или CD27; и/или
(3) CAR содержит внутриклеточный компонент, содержащий эффекторный домен или его функциональную часть, костимулирующий домен или его функциональную часть или любую их комбинацию; и/или
(4) Т-клетка представляет собой Т-клетку человека, CD4+ Т-клетку, CD8+ Т-клетку, CD4-CD8-дважды отрицательную Т-клетку, γδ Т-клетку, наивную Т-клетку, центральную Т-клетку памяти, эффекторную Т-клетку памяти, суммарную популяцию Т-клеток или любую их комбинацию.
2. Способ лечения множественной миеломы у субъекта, включающий введение субъекту терапевтически эффективного количества Т-клетки, экспрессирующей ВСМА-специфичный химерный антигенный рецептор (CAR), где субъект ранее получал терапевтически эффективное количество ингибитора γ-секретазы в пределах 8-12 часов или в пределах одного-двух дней до введения эффективного количества Т-клетки, причем:
(1) CAR содержит ВСМА-специфичный scFv, ВСМА-специфичный scTCR, ВСМА-специфичное нанотело, ВСМА-специфичное доменное антитело или лиганд ВСМА или его связывающую часть; и/или
(2) CAR содержит гидрофобную часть, расположенную между внеклеточным компонентом и внутриклеточным компонентом, где внеклеточный компонент содержит ВСМА-специфичный связывающий домен и где гидрофобная часть представляет собой трансмембранный домен CD4, CD8, CD28 или CD27; и/или
(3) CAR содержит внутриклеточный компонент, содержащий эффекторный домен или его функциональную часть, костимулирующий домен или его функциональную часть или любую их комбинацию; и/или
(4) Т-клетка представляет собой Т-клетку человека, CD4+ Т-клетку, CD8+ Т-клетку, CD4-CD8-дважды отрицательную Т-клетку, γδ Т-клетку, наивную Т-клетку, центральную Т-клетку памяти, эффекторную Т-клетку памяти, суммарную популяцию Т-клеток или любую их комбинацию.
3. Способ лечения множественной миеломы у субъекта, включающий введение субъекту терапевтически эффективного количества ингибитора γ-секретазы, где субъект ранее получал терапевтически эффективное количество Т-клетки, экспрессирующей ВСМА-специфичный химерный антигенный рецептор (CAR), в пределах 8-12 часов, или в пределах одного, двух, трех, четырех, пяти, шести, семи, восьми или девяти дней, или в пределах одной, двух, трех, четырех недель, пяти, шести, семи, восьми или девяти недель до введения эффективного количества ингибитора γ-секретазы, причем:
(1) CAR содержит ВСМА-специфичный scFv, ВСМА-специфичный scTCR, ВСМА-специфичное нанотело, ВСМА-специфичное доменное антитело или лиганд ВСМА или его связывающую часть; и/или
(2) CAR содержит гидрофобную часть, расположенную между внеклеточным компонентом и внутриклеточным компонентом, где внеклеточный компонент содержит ВСМА-специфичный связывающий домен и где гидрофобная часть представляет собой трансмембранный домен CD4, CD8, CD28 или CD27; и/или
(3) CAR содержит внутриклеточный компонент, содержащий эффекторный домен или его функциональную часть, костимулирующий домен или его функциональную часть или любую их комбинацию; и/или
(4) Т-клетка представляет собой Т-клетку человека, CD4+ Т-клетку, CD8+ Т-клетку, CD4-CD8-дважды отрицательную Т-клетку, γδ Т-клетку, наивную Т-клетку, центральную Т-клетку памяти, эффекторную Т-клетку памяти, суммарную популяцию Т-клеток или любую их комбинацию.
4. Способ по любому из пп. 1-3, где CAR является человеческим или гуманизированным.
5. Способ по любому из пп. 1-4, где CAR содержит ВСМА-специфичный scFv, ВСМА-специфичный scTCR, ВСМА-специфичное нанотело, ВСМА-специфичное доменное антитело или лиганд ВСМА или его связывающую часть.
6. Способ по п. 5, где CAR содержит ВСМА-специфичный scFv, ВСМА-специфичное нанотело, ВСМА-специфичное доменное антитело или лиганд ВСМА или его связывающую часть.
7. Способ по п. 5 или 6, где CAR включает scFv, содержащий вариабельные области тяжелой цепи и легкой цепи из анти-ВСМА антитела J22.0-xi, J22.9-xi, J6M0, J6M1, J6M2, J9M0, J9M1, J9M2, 11D5-3, СА8, A7D12.2, С11 D5.3, С12А3.2, C13F12.1, 13С2, 17А5, 83А10, 13А4, 13D2, 14В11, 14Е1, 29В11, 29F3, 13А7, СА7, S307118G03, SG1, S332121F02, S332126E04, S322110D07, S336105A07, S335115G01, S335122F05, ЕТ140-3, ЕТ140-24, ЕТ140-37, ЕТ140-40, ЕТ140-54, TBL-CLN1, C4.E2.1, Vicky-1, pSCHLI333, pSCHLI372 или pSCHLI373.
8. Способ по любому из пп. 1-7, где CAR содержит гидрофобную часть, расположенную между внеклеточным компонентом и внутриклеточным компонентом, где внеклеточный компонент содержит ВСМА-специфичный связывающий домен, причем ВСМА-специфичный связывающий домен содержит: а) ВСМА-специфичную антигенсвязывающую часть анти-ВСМА антитела J22.0-xi, J22.9-xi, J6M0, 11D5-3, СА8, A7D12.2, С11 D5.3, С12А3.2, C13F12.1, 13С2, 17А5, 83А10, 13А4, 13D2, 14В11, 14Е1, 29В11, 29F3, 13А7, СА7, SG1, S307118G03, S332121F02, S332126E04, S322110D07, S336105A07, S335115G01, S335122F05, ЕТ140-3, ЕТ140-24, ЕТ140-37, ЕТ140-40, ЕТ140-54, TBL-CLN1, С4.Е2.1, Vicky-1, pSCHLI333, pSCHLI372 или pSCHLI373; или b) лиганд ВСМА или его связывающую часть на основе или полученный из BAFF (фактор активации В-клеток) или APRIL (лиганд, индуцирующий пролиферацию).
9. Способ по п. 8, где гидрофобная часть представляет собой трансмембранный домен.
10. Способ по любому из пп. 1-9, где CAR содержит трансмембранный домен CD4, CD8, CD28 или CD27.
11. Способ по любому из пп. 1-10, где CAR содержит внутриклеточный компонент, содержащий эффекторный домен или его функциональную часть, костимулирующий домен или его функциональную часть или любую их комбинацию.
12. Способ по п. 11, где внутриклеточный компонент содержит 4-1ВВ (CD137), CD3ε, CD3δ, CD3ζ, CD25, CD27, CD28, CD79A, CD79B, CARD11, DAP10, FcRα, FcRβ, FcRγ, Fyn, HVEM, ICOS, Lck, LAG3, LAT, LRP, NKG2D, NOTCH1, NOTCH2, NOTCH3, NOTCH4, OX40 (CD134), ROR2, Ryk, SLAMF1, Slp76, pTα, TCRα, TCRβ, TRIM, Zap70, PTCH2 или их функциональную часть или любую их комбинацию.
13. Способ по п. 11 или 12, где эффекторный домен содержит CD3ζ или его функциональную часть.
14. Способ по любому из пп. 1-13, где CAR содержит внутриклеточный компонент, содержащий костимулирующий домен или его функциональную часть, выбранный из CD27, CD28, 4-1ВВ (CD137), ОХ40 (CD134) или любой их комбинации.
15. Способ по любому из пп. 1-14, где CAR содержит внутриклеточный компонент, содержащий эффекторный домен или его эффекторную часть, которые содержат CD3ζ или его функциональную часть, и один или более костимулирующих доменов 4-1ВВ (CD137), CD27, CD28 и ОХ40 (CD134) или их функциональную часть.
16. Способ по любому из пп. 1-15, где CAR содержит внутриклеточный компонент, содержащий: (а) 4-1ВВ или его функциональную часть и CD3ζ, (b) CD27 или его функциональную часть и CD3ζ, (с) CD28 или его функциональную часть и CD3ζ, (d) ОХ40 или его функциональную часть и CD3ζ, (е) CD28 или его функциональную часть, 4-1ВВ или его функциональную часть и CD3ζ, (f) ОХ40 или его функциональную часть, 4-1ВВ или его функциональную часть и CD3ζ или (g) CD28 или его функциональную часть, ОХ40 или его функциональную часть и CD3ζ.
17. Способ по любому из пп. 1-16, где CAR содержит внеклеточный компонент, содержащий шарнирный участок иммуноглобулина, домен СН2 и домен СН3, домен СН3 или любую их комбинацию, расположенный между ВСМА-специфичным связывающим доменом и гидрофобной частью.
18. Способ по п. 17, где шарнирная область иммуноглобулина представляет собой шарнирную область IgG1.
19. Способ по п. 17 или 18, где домен СН2 представляет собой домен СН2 IgG1 и домен СН3 представляет собой домен СН3 IgG1.
20. Способ по любому из пп. 1-16, где:
(i) CAR содержит внутриклеточный компонент, содержащий эффекторный домен из CD3ζ;
(ii) CAR содержит внутриклеточный компонент, содержащий костимулирующий домен из 4-1ВВ;
(iii) CAR содержит внутриклеточный компонент, содержащий костимулирующий домен из CD28;
(iv) CAR содержит внеклеточный компонент, содержащий шарнирную область, содержащую область стебля молекулы кластера дифференцировки, или его функциональный вариант;
(v) CAR содержит внутриклеточный компонент, содержащий спейсерную область, включающую шарнир, где спейсерная область имеет длину от 15±20% аминокислот до 100±20% аминокислот;
(vi) CAR является мультиспецифичным, где необязательно CAR является биспецифичным, где дополнительно необязательно CAR является специфичным для первого эпитопа ВСМА и второго эпитопа ВСМА, который отличается от первого эпитопа ВСМА; или
(vii) любая комбинация (i)-(vi).
21. Способ по п. 20, где CAR является биспецифичным.
22. Способ по п. 20 или 21, где CAR является специфичным для первого эпитопа ВСМА и второго эпитопа ВСМА, который отличается от первого эпитопа ВСМА.
23. Способ по любому из пп. 1-22, где Т-клетка представляет собой Т-клетку человека.
24. Способ по любому из пп. 1-23, где Т-клетка представляет собой CD4+ Т-клетку, CD8+ Т-клетку, CD4-CD8-дважды отрицательную Т-клетку, γδ Т-клетку или любую их комбинацию.
25. Способ по п. 24, где Т-клетка включает CD8+ Т-клетку, CD4+ Т-клетку или обе из них.
26. Способ по любому из пп. 1-25, где Т-клетка представляет собой наивную Т-клетку, центральную Т-клетку памяти, эффекторную Т-клетку памяти, суммарную популяцию Т-клеток или любую их комбинацию.
27. Способ по любому из пп. 1-26, где ингибитор γ-секретазы включает малую молекулу, пептидомиметическое соединение, LY411575, семагацестат, авагацестат, DAPT, BMS-906024, BMS-986115, MK-0752, PF-03084014, RO4929097 или YO-01027.
28. Способ по любому из пп. 1-26, где ингибитор γ-секретазы представляет собой никастрин-специфичный связывающий белок.
29. Способ по п. 28, где никастрин-специфичный связывающий белок представляет собой scFvG9, антитело А5226А, антитело 2Н6 или антитело 10С11.
30. Способ по любому из пп. 1-6 и 9-29, где CAR содержит вариабельный домен, содержащий аминокислотную последовательность любой из SEQ ID NO: 14-16, и вариабельный домен, содержащий аминокислотную последовательность любой из SEQ ID NO: 17-21.
31. Способ по любому из пп. 1-30, где способ дополнительно включает предварительное кондиционирование субъекта иммуносупрессивным режимом до или одновременно с Т-клеткой и ингибитором γ-секретазы.
32. Способ по п. 31, где иммуносупрессивный режим включает немиелоаблативный режим или миелоаблативный режим.
33. Способ по п. 32, где немиелоаблативный режим включает циклофосфамид или циклофосфамид в комбинации с флударабином.
34. Способ по п. 1, где Т-клетку и ингибитор γ-секретазы вводят последовательно.
35. Способ по п. 1, где Т-клетку и ингибитор γ-секретазы вводят одновременно.
36. Способ по п. 35, где Т-клетка и ингибитор γ-секретазы составлены вместе.
37. Способ по любому из пп. 1-35, где Т-клетку вводят парентерально, а ингибитор γ-секретазы вводят перорально.
38. Способ по п. 34, включающий введение Т-клетки и последующее введение ингибитора γ-секретазы, где необязательно:
(1) ингибитор γ-секретазы вводят через один, два, три, четыре, пять, шесть, семь, восемь или девять дней после введения Т-клетки; через одну, две, три, четыре недели после введения Т-клетки; или через одну, две, три, четыре, пять, шесть, семь, восемь или девять недель после введения Т-клетки, и/или
(2) способ дополнительно включает предварительное кондиционирование субъекта иммуносупрессивным режимом, включающим циклофосфамид или циклофосфамид в комбинации с флударабином, до введения Т-клетки.
39. Способ по любому из пп. 1-38, где CAR содержит T-ChARM и где внеклеточный домен T-ChARM содержит Strep-метку.
40. Способ по п. 39, где внеклеточный домен T-ChARM содержит две, три, четыре, пять, шесть, семь, восемь или девять меток-кассет.
41. Способ по любому из пп. 1-40, где ингибитор γ-секретазы вводят субъекту по меньшей мере один раз после первого введения Т-клетки.
42. Способ по п. 41, где ингибитор γ-секретазы вводят по меньшей мере 2, по меньшей мере 3, по меньшей мере 4, по меньшей мере 5, по меньшей мере 6, по меньшей мере 7, по меньшей мере 8, по меньшей мере 9, по меньшей мере 10, по меньшей мере 15, по меньшей мере 20, по меньшей мере 25, по меньшей мере 30, по меньшей мере 35, по меньшей мере 40, по меньшей мере 45 или по меньшей мере 50 раз после первого введения Т-клетки.
43. Способ по любому из пп. 1-42, где ингибитор γ-секретазы вводят субъекту в дозе, содержащей 30±20% мг/кг ингибитора γ-секретазы.
44. Способ по любому из пп. 1-43, где Т-клетка представляет собой Т-клетку человека, которая является аллогенной или аутологичной по отношению к субъекту.
45. Способ по любому из пп. 1-43, где Т-клетка представляет собой Т-клетку человека, которая является изогенной по отношению к субъекту.
46. Способ по любому из пп. 24-45, где Т-клетка включает CD4+ Т-клетку и CD8+ Т-клетку.
47. Способ по любому из пп. 1-27, где:
(i) CAR содержит (1) ВСМА-специфичный связывающий домен, содержащий ВСМА-специфичный scFv, ВСМА-специфичное доменное антитело или лиганд ВСМА или его связывающую часть, (2) внутриклеточный компонент, содержащий костимулирующий домен из 4-1ВВ и/или CD28 и эффекторный домен из CD3ζ, и/или (3) внеклеточную спейсерную область, включающую шарнир, где спейсерная область имеет длину от 15±20% аминокислот до 100±20% аминокислот;
(ii) ингибитор γ-секретазы включает малую молекулу; или
(iii) (i) и (ii).
48. Способ по п. 47, где:
(i) CAR содержит (1) ВСМА-специфичный связывающий домен, содержащий ВСМА-специфичный scFv, ВСМА-специфичное доменное антитело или лиганд ВСМА или его связывающую часть, (2) внутриклеточный компонент, содержащий костимулирующий домен из 4-1ВВ и/или CD28 и эффекторный домен из CD3ζ, и (3) внеклеточную спейсерную область, включающую шарнир, где спейсерная область имеет длину от 15±20% аминокислот до 100±20% аминокислот; и
(ii) ингибитор γ-секретазы включает малую молекулу.
49. Способ по любому из пп. 1-27, где ингибитор γ-секретазы включает малую молекулу.
50. Применение (1) Т-клетки, экспрессирующей ВСМА-специфичный химерный антигенный рецептор (CAR), с (2) ингибитором γ-секретазы для лечения множественной миеломы у субъекта, причем:
(1) CAR содержит ВСМА-специфичный scFv, ВСМА-специфичный scTCR, ВСМА-специфичное нанотело, ВСМА-специфичное доменное антитело или лиганд ВСМА или его связывающую часть; и/или
(2) CAR содержит гидрофобную часть, расположенную между внеклеточным компонентом и внутриклеточным компонентом, где внеклеточный компонент содержит ВСМА-специфичный связывающий домен и где гидрофобная часть представляет собой трансмембранный домен CD4, CD8, CD28 или CD27; и/или
(3) CAR содержит внутриклеточный компонент, содержащий эффекторный домен или его функциональную часть, костимулирующий домен или его функциональную часть или любую их комбинацию; и/или
(4) Т-клетка представляет собой Т-клетку человека, CD4+ Т-клетку, CD8+ Т-клетку, CD4-CD8-дважды отрицательную Т-клетку, γδ Т-клетку, наивную Т-клетку, центральную Т-клетку памяти, эффекторную Т-клетку памяти, суммарную популяцию Т-клеток или любую их комбинацию.
51. Применение Т-клетки по п. 50, где CAR содержит внеклеточный компонент и внутриклеточный компонент, связанные гидрофобной частью, где внеклеточный компонент содержит связывающий домен, который специфически связывает ВСМА, и шарнирную область и где внутриклеточный компонент содержит эффекторный домен и необязательно костимулирующий домен.
52. Применение Т-клетки по п. 50 или 51, где CAR содержит ВСМА-специфичный scFv, ВСМА-специфичный scTCR, ВСМА-специфичное нанотело, ВСМА-специфичное доменное антитело или лиганд ВСМА или его связывающую часть.
53. Применение Т-клетки по п. 52, где CAR содержит ВСМА-специфичный scFv, ВСМА-специфичное нанотело, ВСМА-специфичное доменное антитело или лиганд ВСМА или его связывающую часть.
54. Применение Т-клетки по любому из пп. 50-53, где:
(i) CAR содержит внутриклеточный компонент, содержащий эффекторный домен из CD3ζ;
(ii) CAR содержит внутриклеточный компонент, содержащий костимулирующий домен из 4-1ВВ;
(iii) CAR содержит внутриклеточный компонент, содержащий костимулирующий домен из CD28;
(iv) CAR содержит внеклеточный компонент, содержащий шарнирную область, содержащую область стебля молекулы кластера дифференцировки или его функциональный вариант;
(v) CAR содержит внутриклеточный компонент, содержащий спейсерную область, включающую шарнир, где спейсерная область имеет длину от 15±20% аминокислот до 100±20% аминокислот;
(vi) CAR является мультиспецифичным, где необязательно CAR является биспецифичным, где дополнительно необязательно CAR является специфичным для первого эпитопа ВСМА и второго эпитопа ВСМА, который отличается от первого эпитопа ВСМА; или
(vii) любая комбинация (i)-(vi).
55. Применение Т-клетки по п. 54, где CAR является биспецифичным.
56. Применение Т-клетки по п. 54 или 55, где CAR является специфичным для первого эпитопа ВСМА и второго эпитопа ВСМА, который отличается от первого эпитопа ВСМА.
57. Применение Т-клетки по любому из пп. 50-56, где Т-клетка представляет собой Т-клетку человека, включающую CD4+ Т-клетку, CD8+ Т-клетку, CD4-CD8-дважды отрицательную Т-клетку, γδ Т-клетку или любую их комбинацию.
58. Применение Т-клетки по любому из пп. 50-57, где Т-клетка представляет собой наивную Т-клетку, центральную Т-клетку памяти, эффекторную Т-клетку памяти, суммарную популяцию Т-клеток или любую их комбинацию.
59. Применение Т-клетки по п. 57 или 58, где Т-клетка включает CD4+ Т-клетку и CD8+ Т-клетку.
60. Применение Т-клетки по любому из пп. 50-59, где ингибитор γ-секретазы включает малую молекулу, пептидомиметическое соединение, LY411575, семагацестат, авагацестат, DAPT, BMS-906024, BMS-986115, MK-0752, PF-03084014, RO4929097 или YO-01027.
61. Применение Т-клетки по любому из пп. 50-60, где:
(i) CAR содержит (1) ВСМА-специфичный связывающий домен, содержащий ВСМА-специфичный scFv, ВСМА-специфичное доменное антитело или лиганд ВСМА или его связывающую часть, (2) внутриклеточный компонент, содержащий костимулирующий домен из 4-1ВВ и/или CD28 и эффекторный домен из CD3ζ, и/или (3) внеклеточную спейсерную область, включающую шарнир, где спейсерная область имеет длину от 15±20% аминокислот до 100±20% аминокислот;
(ii) ингибитор γ-секретазы включает малую молекулу; или
(iii) (i) и (ii).
62. Применение Т-клетки по п. 61, где:
(i) CAR содержит (1) ВСМА-специфичный связывающий домен, содержащий ВСМА-специфичный scFv, ВСМА-специфичное доменное антитело или лиганд ВСМА или его связывающую часть, (2) внутриклеточный компонент, содержащий костимулирующий домен из 4-1ВВ и/или CD28 и эффекторный домен из CD3ζ, и (3) внеклеточную спейсерную область, включающую шарнир, где спейсерная область имеет длину от 15±20% аминокислот до 100±20% аминокислот; и
(ii) ингибитор γ-секретазы включает малую молекулу.
63. Применение Т-клетки по любому из пп. 50-61, где ингибитор γ-секретазы включает малую молекулу.
64. Применение Т-клетки по любому из пп. 50-61, где ингибитор γ-секретазы представляет собой никастрин-специфичный связывающий белок.
65. Применение Т-клетки по п. 64, где никастрин-специфичный связывающий белок представляет собой scFvG9, антитело А5226А, антитело 2Н6 или антитело 10С11.
66. Применение Т-клетки по любому из пп. 50-65, где CAR содержит вариабельный домен, который содержит аминокислотную последовательность любой из SEQ ID NO: 14-16, и вариабельный домен, содержащий аминокислотную последовательность любой из SEQ ID NO: 17-21.
67. Набор для лечения множественной миеломы, включающий: а) единицу дозирования Т-клетки, экспрессирующей ВСМА-специфичный химерный антигенный рецептор (CAR), и b) единицу дозирования ингибитора γ-секретазы, причем:
(1) CAR содержит ВСМА-специфичный scFv, ВСМА-специфичный scTCR, ВСМА-специфичное нанотело, ВСМА-специфичное доменное антитело или лиганд ВСМА или его связывающую часть; и/или
(2) CAR содержит гидрофобную часть, расположенную между внеклеточным компонентом и внутриклеточным компонентом, где внеклеточный компонент содержит ВСМА-специфичный связывающий домен и где гидрофобная часть представляет собой трансмембранный домен CD4, CD8, CD28 или CD27; и/или
(3) CAR содержит внутриклеточный компонент, содержащий эффекторный домен или его функциональную часть, костимулирующий домен или его функциональную часть или любую их комбинацию; и/или
(4) Т-клетка представляет собой Т-клетку человека, CD4+ Т-клетку, CD8+ Т-клетку, CD4-CD8-дважды отрицательную Т-клетку, γδ Т-клетку, наивную Т-клетку, центральную Т-клетку памяти, эффекторную Т-клетку памяти, суммарную популяцию Т-клеток или любую их комбинацию.
68. Набор по п. 67, дополнительно содержащий CD20-специфичный связывающий белок, такой как ритуксимаб, офатумумаб или окрелизумаб; CD19-специфичный связывающий белок; CD45-специфичный связывающий белок; CD38-специфичный связывающий белок; цитокин; хемокин; фактор роста; химиотерапевтический агент или радиотерапевтический агент.
| WO 2017019496 A1, 02.02.2017 | |||
| ALI S | |||
| A | |||
| et al | |||
| T cells expressing an anti-B-cell maturation antigen chimeric antigen receptor cause remissions of multiple myeloma // Blood, The Journal of the American Society of Hematology | |||
| Токарный резец | 1924 |
|
SU2016A1 |
| - Vol | |||
| Сепаратор-центрофуга с периодическим выпуском продуктов | 1922 |
|
SU128A1 |
| - No | |||
| Насос | 1917 |
|
SU13A1 |
| - P | |||
| Приспособление для укрепления деревянных бортов на цилиндрических деревянных стержнях или трубках | 1925 |
|
SU1688A1 |
| JONNALAGADDA M | |||
| et al | |||
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |

