Предлагаемое техническое решение относится к области здоровья, а именно к восстановительной (реабилитационной) медицине, к средствам стимуляции структур нейронов коры головного мозга и гиппокампа в послеинсультный период. Задачей предлагаемой системы для стимуляции коры головного мозга для восстановления кратковременной и долговременной памяти в послеинсультный период является объемная стимуляция электрической активности нейронов головного мозга и гиппокампа через рецепторные системы – зрительную и слуховую, а также через воздействие магнитным полем на кору головного мозга, параметры которого приближенны к ритмам электрической активности головного мозга альфа, бета, тета, дельта ритмам.
Из современных подходов, используемых в медицинских технических системах для реабилитации людей, перенесших инсульт, можно выделить физиотерапию, компьютеризированные и роботизированные системы кинезиотерапии, тренировки в виртуальной реальности, электромиографическую-биологическую обратную связь, функциональную электрическую стимуляцию и транскраниальную магнитную стимуляцию. (Ибрагимов, М.Ф., Хабиров, Ф.А., Хайбуллин, Т.И., Гранатов, Е.В. Современные подходы к реабилитации больных, перенесших инсульт //Практическая медицина. – 2012. – №. 57.) [1]
Из существующих систем магнитной стимуляции наиболее близкими к заявленному техническому решению являются: Нейро МС/Д, MagPro, Tamas, Magstim Rapid, устройство для сочетанной транскраниальной физиотерапии – заявка 2009117359/14, 06.05.2009, установка для транскраниального воздействия вращающимся магнитным полем – заявка 2010129708/14, 15.07.2010, данные устройства имеют ряд недостатков.
У линейки устройств Нейро МС/Д, MagPro, Tamas, Magstim Rapid техническими недостатками являются:
1) Недостаточное количество воздействующих излучателей, что не позволяет охватить стимулирующими воздействиями магнитного поля всю кору головного мозга;
2) Ограниченная вариабельность воздействующего магнитного поля с точки зрения изменения частот коммутаций воздействия;
3) Большая амплитуда воздействующего магнитного поля от 1,5 Тл до 4 Тл, что вызывает моторные реакции наведенного магнитного поля на рецепторные нейроны и не влияет на общее функционирование нейронных полей, поэтому применять данное устройство при лечении и реабилитации больных с поражением коры головного мозга не эффективно;
4) Отсутствие возможности аудио-визуальных воздействий, с разницей частот воздействий равной различным ритмам головного мозга.
У устройств для сочетанной транскраниальной физиотерапии – заявка 2009117359/14, 06.05.2009, установка для транскраниального воздействия вращающимся магнитным полем – заявка 2010129708/14, 15.07.2010 техническими недостатками является:
1) Малое количество воздействующих излучателей, что не обеспечивает точное воздействие на локальном уровне, а также не позволяет проводить стимулирование множество зон небольшой площади одновременно;
2) Недостаточное количество вариаций характеристик воздействующим магнитным полем;
3) Отсутствие возможности регулирования характеристик алгоритмов воздействия магнитным полем каждого излучателя по отдельности;
4) Отсутствие возможности аудио-визуальных воздействий, с разницей частот воздействий равной различным ритмам головного мозга.
Из существующих устройств аудио-визуальной стимуляции наиболее близкими к заявленному техническому решению являются: Nova Pro 100, Inner Pulse, Luma 10, Pro Tutor - www.photosonix.com; заявка 2002131036/14, 18.11.2002 и Устройство сенсорной стимуляции «лингвостим» - www.lingvostim.com; заявка:2013111778/14, 15.03.2013. Данные устройства имеют ряд недостатков.
У линейки устройств Nova Pro 100, Inner Pulse, Luma 10, Pro Tutor и устройства сенсорной стимуляции «лингвостим» техническими недостатками является:
1) Отсутствие возможности регулирования форм воздействующих сигналов;
2) Отсутствие установки различных вариантов частотных модуляций воздействующих сигналов;
3) Отсутствие возможности настройки и управления воздействия на каждый глаз отдельным сигналом и воздействующей частотой;
4) Отсутствие возможности формирования характеристик воздействия с разным цветом и частотой на каждый глаз.
Задачей настоящего технического решения создание эффективной технической системы для восстановления кратковременной и долговременной памяти в послеинсультный период.
Решение задачи объёмной стимуляции электрической активности нейронов коры головного мозга и гиппокампа достигается одновременной подачей суммы воздействий: на рецепторные системы (зрительная и слуховая) и слабоинтенсивным воздействием магнитного поля на нейронные поля головного мозга. При этом параметры этих воздействий и их электрические характеристики, определяемые по предварительно замеренным энцефалограммам, близки к параметрам альфа, бета, тета, дельта ритмов электрической активности групп нейронов головного мозга, что вызывает электрический резонанс наведенных сигналов на аксоны нейронов, на которых имеются электрические сигналы межнейронных связей. Через рецепторные системы (глаза и уши) и через магнитные поля с наведением электрических сигналов в нейронах коры головного мозга и в гиппокампе, в таком случае, при стимуляции нейронов усиливается активность здоровых (непораженных) нейронов, определяющих работу мозга по фиксации достоверных сигналов при формировании элементов памяти, в том числе кратковременной памяти. Методически это достигается расположением излучателей света в очках перед каждым из глаз, расположением излучателей звука (аудио сигналов) в каждом ухе и магнитных излучателей по поверхности головы в количестве тридцати двух штук и формированием необходимых воздействий, определяемых блоком формирования характеристик магнитных воздействий. Разница частот световых и аудио воздействий (звуковой диапазон) подаваемых на каждый из глаз и на каждое из ушей равны или приближены к частотам ритмов электрической активности головного мозга конкретного пациента. Аналогично разница количества коммутаций включения излучателей магнитных полей программно определена равной или приближенной к альфа, бета, тета, дельта ритмам конкретного пациента измеренных перед применением данной системы, что дает возможность увеличивать площадь и глубину стимуляции здоровых нейронных полей головного мозга пациента, которые могут восстанавливать «кусочные» фрагменты памяти, особенно долговременной, поскольку, как показали исследования, проведенные различными авторами, долговременная память распределена по всей коре головного мозга. [2]
Под термином «кратковременная память» мы понимаем период, в течение которого информация хранится преимущественно в структурах гиппокампа. Термин «долговременная память» в настоящей работе подразумевает завершающую стадию процесса консолидации следа памяти в неокортексе. Соответственно, «кратковременная» память подразумевает обратимость и возможное нивелирование зарегистрированных данных, в то время как регистрация информации в системе «долговременной» памяти характеризуется стабильностью и необратимостью. [3]
После первичной обработки информации об объекте/событии в анализаторах неокортекса с формированием его внутренней репрезентации и выделением значимых признаков информация направляется к структурам парагиппокампальной и периринальной коры [4, 5, 6]. Периринальная кора анализирует «непространственные» признаки (что?) объекта, в то время как парагиппокампальная кора анализирует пространственную (где?) информацию [4]. Показано, что парагиппокампальные структуры коры мозга производят оценку новизны информации [4]. Из структур парагиппокампальной и периринальной коры обработанная информация передается в медиальные и латеральные отделы энторинальной коры соответственно [4, 5, 6]. Точные функции энториальной коры остаются неясными, однако известно, что вся информация от неокортекса поступает в гиппокамп только через данную структуру.
Переход кратковременной памяти в долговременную называется консолидацией. В процессе консолидации происходит накопление данных, а также их интеграция и ассоциация с другими данными в памяти подобного типа, что обеспечивает более быстрый поиск необходимых данных. Основой долговременной памяти является энграмма. Энграмма - своеобразный «отпечаток» памяти, который формируется в результате деятельности. Представляет собой совокупность физических, химических и морфологических изменений в нервных структурах, которые оказывают существенное влияние на рефлекторные реакции организма.
Гиппокамп получает обработанную в парагиппокампальном регионе информацию через структуры энторинальной коры. Информация от медиальных (пространственных) и латеральных (идентифицирующих объект) отделов энторинальной коры поступает в одни и те же нейрональные сети в зубчатой извилине (dentate gyrus) и области CA3 гиппокампа, но проецируется в разные нейросети области СА1 и subiculum [4]. Подобная нейроморфологическая организация энторинально-гиппокампальных связей обеспечивает ассоциирование событий и контекстов в одних структурах и их сепаратный процессинг в других структурах гиппокампа. В целом, важнейшей характеристикой функционирования системы памяти млекопитающих и, возможно, других животных является облигатная регистрация событий и объектов в комплексе с визуально-пространственным контекстом, т.е. на первичном этапе любая информация регистрируется как «эпизодическая» [4]. Известно, что гиппокамп формирует первоначальную энграмму, однако механизмы данного процесса продолжают активно изучаться. Гиппокамп является временным хранилищем информационных паттернов, и данная его функция реализуется благодаря чрезвычайно высокой пластичности коры гиппокампа и ее способности к «стиранию» и обновлению регистрируемых данных [7, 8]. Информация в гиппокампе хранится в неструктурированном виде и может быть легко изменена интерферирующими влияниями. Таким образом, важнейшей функцией гиппокампа является передача значимого информационного паттерна в структуры коры головного мозга, способные к надежному и долговременному хранению структурированных знаний. Предполагается, что передача и консолидация информации в соответствующие зоны неокортекса осуществляется структурами в задних отделах гиппокампа [9]. Передача информации в долговременную память неокортекса обеспечивается повторными «проигрываниями» (replays) гиппокампом значимых информационных паттернов с объединением в общую нейросеть релевантных зон неокортекса [10, 11]. Повторные «проигрывания» информационных паттернов гиппокампом с сопутствующей активацией структур неокортекса происходят как в состоянии бодрствования, так и во сне [10, 12]. У людей функции гиппокампов латерализованы, т.е. в запоминании вербальной информации преимущественно участвует левый гиппокамп, а невербальной – правый [13]. Предполагается, что возрастные изменения в объеме коры гиппокампов связаны со снижением нейрогенеза в данной структуре мозга. [3]
На сегодняшний день существует различные гипотезы биохимических механизмов долговременной памяти. [14]
Из них можно выделить две основные - это синаптическая и геномная.
Причина высокой популярности синаптической гипотезы памяти коренится в основной парадигме современной нейрофизиологии. Согласно этой парадигме все функции нервной системы осуществляются исключительно на уровне нейронных сетей. При этом нейроны, формирующие сети, рассматриваются как простые элементы, функция которых сводится к генерации электрических потенциалов и передаче сигналов к другим нейронам. Эта концепция в равной степени относится как к более простым функциям нервной системы (непроизвольные рефлексы, управление дыханием, локомоцией и другими ритмическими движениями, поддержание гомеостаза и т.п.), осуществляемым в основном спинально-стволовыми механизмами, так и к высшим, ментальным функциям человеческого мозга, осуществляемым большими полушариями. Как было уже отмечено в предыдущих публикациях [16, 17], такая концепция фактически низводит мозг до уровня гигантской, обучающейся вычислительной машины, построенной из двоичных элементов, имеющих два дискретных состояния – “0” (покой) и “1” (возбуждение типа “все или ничего”).
Согласно геномной гипотезе долговременная память в основном хранится на внутриклеточном уровне в виде модификаций генома (в широком понимании этого слова, включающем эпигенетические модификации). Иначе говоря, эта гипотеза предполагает, что нейроны соответствующих отделов мозга являются носителями элементарных следов памяти и что воспоминания о различных фактах и событиях являются результатом сложной кооперативной активности такого рода нейронов. В этом смысле геномная гипотеза никоим образом не отрицает, а расширяет синаптическую гипотезу памяти. [15]
Гипотеза Х. Хидена. Согласно его опытам, образование следов памяти происходит вместе с изменением свойств РНК и белков в нейронах. Разная структура импульсных потенциалов в нейронах вызывает различную перестройку молекул РНК, специфические для каждого сигнала перемещения нуклеотидов в цепи. В результате каждый сигнал фиксируется в виде специфичного отпечатка в структуре РНК. Что в свою очередь приводит к синтезу специфического белка, который обеспечивает избирательную чувствительность нейронов к определенным воздействиям. [18-21]
Гипотеза переноса памяти Д. Унгара. В 1970-е годы Д. Унгар предположил, что определенные белки принимают участие в хранении энграмм памяти. [22-26]
Гипотеза Линча и Бодри. Концентрация ионов кальция около постсинаптической мембраны повышается после повторной стимуляции нейронов. В результате активируется кальций-зависимая протеиназа - калпеин - в мембране постсинаптического нейрона. Калпеин, расщепляет структурный белок - фодрин. Вследствие этого активируются чувствительные к глутамату рецепторы, которые ранее были блокированы. С увеличением числа активных рецепторов глутамата возрастает проводимость синапса нейрона. [27-29]
Предположение о связи ритмов ЭЭГ с процессами памяти вписывается в общую канву представлений о морфофункциональных основах памяти. Согласно гипотезе Дональда Хебба, многократное прохождение электрической активности по замкнутым цепям нейронов (реверберация) является физиологическим механизмом сохранения следа (энграммы) в кратковременной памяти и необходимым условием для перехода этого следа в долговременную память. Продолжаясь определенное время, реверберация приводит к консолидации - морфофункциональным и биохимическим изменениям в синапсах нейронного ансамбля [30]. Когда нужно извлечь информацию из долговременной памяти, латентная энграмма актуализируется лишь при обратном переводе «молекулярного кода» на уровень электрической активности. Доступ к информации, хранящейся в долговременной памяти, осуществляет некая предполагаемая контролирующая система, которая для обеспечения поиска информации по всем нейросетям памяти должна быть связана с корой многочисленными аксонными связями. Такими свойствами обладают базальные ганглии и таламус, который связан фактически со всеми областями коры [31]. Известно, что альфа-ритм ЭЭГ генерируется в таламо-кортикальных нейронных сетях [32; 33], а его амплитуда коррелирует с интенсивностью гемодинамического сигнала в таламусе [34; 35]. С учетом этих фактов можно прийти к предположению о связи альфа с долговременной памятью.
Тета-ритм ЭЭГ отражает активность корково-лимбических нейросетей [36; 37]. С активностью лимбической системы и прежде всего с гиппокампом многие авторы связывают процессы контекстного кодирования информации, т.е. интеграции актуальной полимодальной информации для сохранения в памяти новых эпизодов [38; 39]. Если гиппокамп на самом деле участвует в кодировании контекста, то следует ожидать, что существуют многочисленные двусторонние связи гиппокампа с ассоциативными зонами коры. Такие связи хорошо известны [40]. Также известно, что в процессе обработки новой информации синхронизируется лишь небольшой процент гиппокампально-корковых петель обратной связи [41]. Это может означать, что тета-активность возникает избирательно только в некоторых областях коры, в которых происходит кодирование новой информации или извлечение недавней информации из памяти. Наконец, было показано, что именно в гиппокампе происходит долговременная потенциация и что тета-активность индуцирует или, по крайней мере, усиливает ее [42]. Долговременная потенциация представляет собой длительное увеличение эффективности синаптической передачи и выступает в теории синаптической пластичности как основа механизмов обучения и памяти. Тот факт, что долговременная потенциация рассматривается как наиболее важный электрофизиологический коррелят процесса кодирования новой информации, подчеркивает возможную значимость гиппокампального тета для процессов эпизодической памяти. Тета-ритм также рассматривается как необходимый компонент системы контроля, обслуживающей процессы рабочей памяти. [43,44]
Физиологическое и лечебное воздействие магнитного поля (МП) обусловлено следующими физическими явлениями - эффектом Холла (магнитоэлектрическим) и эффектом Лоренца (магнитомеханическим).
Эффект Холла заключается в том, что в движущихся проводниках, пересекающих силовые линии МП, возникает электрическая разность потенциалов, а если движущийся проводник представляет собой замкнутый контур, в нем возникает электрический ток. Постоянное магнитное поле (ПМП) индуцирует разность электрических потенциалов и короткозамкнутые вихревые токи или же токи Фуко в движущихся жидкостях организма (кровь, лимфа, тканевая жидкость, цитоплазма клеток), основу которых составляют электролиты (ионы Na+ и Cl-), являющиеся электрическими проводниками. Переменное магнитное поле (ПеМП) и импульсное магнитное поле (ИМП) индуцируют разность электрических потенциалов и короткозамкнутые вихревые токи не только в движущихся, но и в покоящихся тканевых жидкостях, что может являться одной из основных причин более интенсивного биологического и лечебного действия этих форм МП по сравнению с ПМП. В основе биологического и лечебного действия индукционных микротоков лежит вызываемое ими изменение состояния клеточных мембран и связанных с мембранами ферментативных и рецепторных молекул, повышение проницаемости плазмолеммы клеток. Под действием высокоинтенсивных импульсных МП с магнитной индукцией более 0,8 Тл в токопроводящих средах организма происходит наведение токов, сила которых превышает порог возбуждения нервных и мышечных структур, в результате чего происходят лавинообразная деполяризация нервов и мышц, мышечные сокращения.
Эффект Лоренца (магнитомеханический) заключается в том, что между двумя источниками МП действуют механические силы притяжения и отталкивания. Модальность магнитомеханического взаимодействия (направление силы Лоренца) зависит от направления силовых линий (полярности) МП. Если магнитные силовые линии полей двух источников МП имеют встречное направление, то между ними возникают механические силы отталкивания. Если магнитные силовые линии полей двух источников МП имеют одинаковое (параллельное) направление, то между ними возникают механические силы притяжения.
Эффектом Лоренца объясняются многие из биологических и лечебных эффектов МП. Изменяется специфическая активность некоторых внутриклеточных ферментов и мембранных рецепторов, опосредующих действие гормонов и медиаторов. Это связано с наличием некомпенсированного магнитного момента у молекул, входящих в активный центр этих биологически активных макромолекул. В результате внешнего магнитного воздействия в атомах, имеющих неспаренные валентные электроны, происходит сдвиг электронных облаков, что приводит к изменению конформации активного центра фермента или рецептора. Изменяется чувствительность рецепторов к лигандам (гормонам, медиаторам), изменяется характер взаимодействия фермента с субстратом, результатом чего становится ускорение или замедление внутриклеточных биохимических реакций. [45]
Более подробный механизм воздействия магнитного поля на кору головного мозга на клеточном уровне описан в следующих источниках. [46-53]
Получить точную визуализацию воздействия магнитного поля на кору головного мозга достаточно сложно: методы электрической записи в головном или спинном мозге являются инвазивными и в основном используются для исследований на животных, артефакты, возникающие при воздействии магнитным полем затрудняют запись во время и сразу после импульса воздействия, остается неопределенность в отношении очагов стимуляции, а также экспериментальные подходы еще не позволяют оценить нейронный ответ по глубине, положению и типам клеток [54-56]. Методы оптической визуализации, такие как двухфотонная кальциевая визуализация [57,58], могут преодолеть некоторые из этих ограничений, включая артефакты, возникающие при воздействии магнитным полем и идентификацию типа клеток, хотя потенциал действия вызванных переходных процессов кальция может быть слишком длинным (>100 мс), чтобы отличить прямую от косвенной активации клеток [59]. Тем не менее эти методы до сих пор не применялись при изучении ТМС человека и приматов, не являющихся людьми. [60]
Визуальная стимуляция – метод, при котором изменение функционального состояния ЦНС достигается за счет воздействия периодическими импульсами света (вспышки) на зрительный анализатор (глаза). Частоты воздействующих сигналов на оба глаза могут быть как одинаковыми, так и с определенной разницей частот, которая должна соответствовать ритму головного мозга.
Цветостимуляция – метод, при котором изменение функционального состояния ЦНС достигается за счет воздействия сочетаниями цветовых композиций (красный, синий и зеленый цвета). Цвета, выставляемые на глаза, могут быть как одинаковыми, так и разными.
Аудиостимуляция с использование бинауральных ритмов — метод воздействия на слуховые анализаторы (уши) частотами, различающимися между собой в определенном диапазоне, не превышающем 30 Гц. Частоты воздействующих сигналов на оба уха могут быть как одинаковыми, так и с определенной разницей частот, которая должна соответствовать ритму головного мозга.
В данной системе мы сочетанием воздействие МП на кору головного мозга с величинами магнитного поля до 1-5 мТл вместе с аудио-визуальной стимуляцией (АВС) на рецепторную систему человека. Мы считаем, что нет необходимости воздействовать большими магнитными полями с величиной до 2.2 Тл. А вполне достаточно величин порядка 1-5 мТл с использованием различных частотных модуляций сигнала, типов импульсов воздействия.
В результате комбинированного воздействия стимулируются нейроны коры головного мозга и гиппокампа, отвечающие за функционирование кратковременной и долговременной памяти, что приводит к восстановлению памяти больного в послеинсультный период.
С учетом вышеизложенных гипотез, техническим результатом, является созданная система для стимуляции коры головного мозга для восстановления кратковременной и долговременной памяти в послеинсультный период, которая способна формировать характеристики воздействующих аудио- видеосигналов и магнитных полей, отдельно подаваемых на правый и левый глаз, правое и левое ухо, и на магнитные излучатели в определенных комбинациях, позволяющих эффективно воздействовать на нейронные поля головного мозга, что позволит значительно увеличить площадь стимуляции коры головного мозга и гиппокампа благодаря использованию 32 магнитных излучателей охватывающих всю кору головного мозга, а также расширить область применение в медицине конкретно для восстановления кратковременной и долговременной памяти в послеинсультный период. Авторами были проведены эксперименты на здоровых добровольцах, что подтверждает эффективность данной системы. [61]
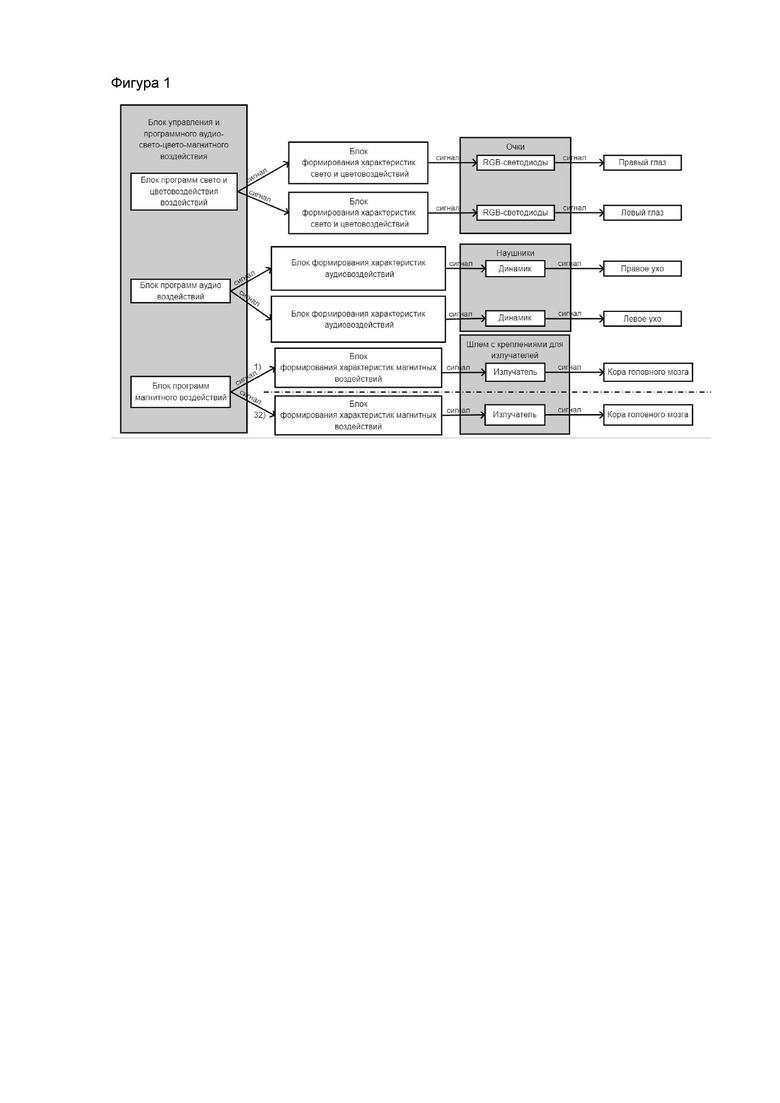
Функциональная схема системы для стимуляции коры головного мозга для восстановления кратковременной и долговременной памяти в послеинсультный период изображена на фиг. 1. Которая включает в себя блок управления и программного аудио-свето-цвето-магнитовоздействия, включающий в себя блок программ свето- и цветовоздействия воздействий, блок программ аудиовоздействий, блок программ магнитного воздействий; блоки формирования характеристик свето- и цветовоздействий, блоки формирования характеристик аудиовоздействий, блоки формирования характеристик магнитных воздействий, параметры которых определяются блоком управления; динамиков расположенных в стереонаушниках, RGB-светодиодов расположенных в очках, 32 соленоидов-излучателей закрепленных в шлеме.
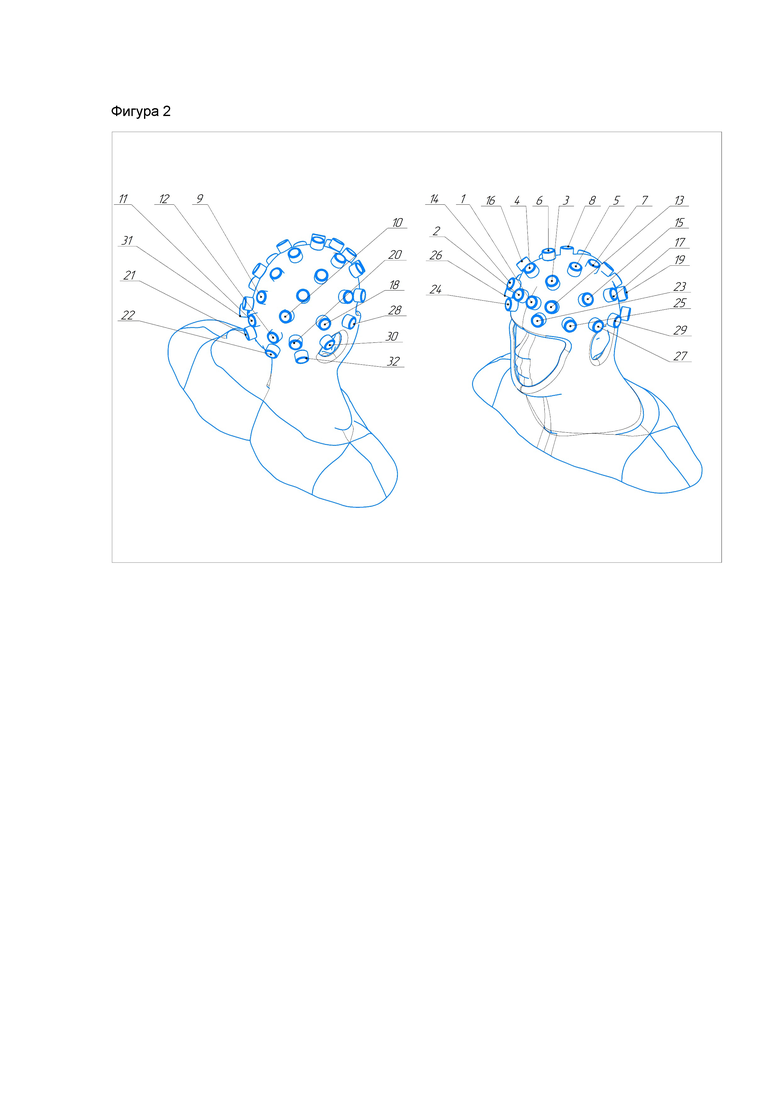
Расположение излучателей магнитного поля блока магнитного воздействия изображено на фиг. 2.
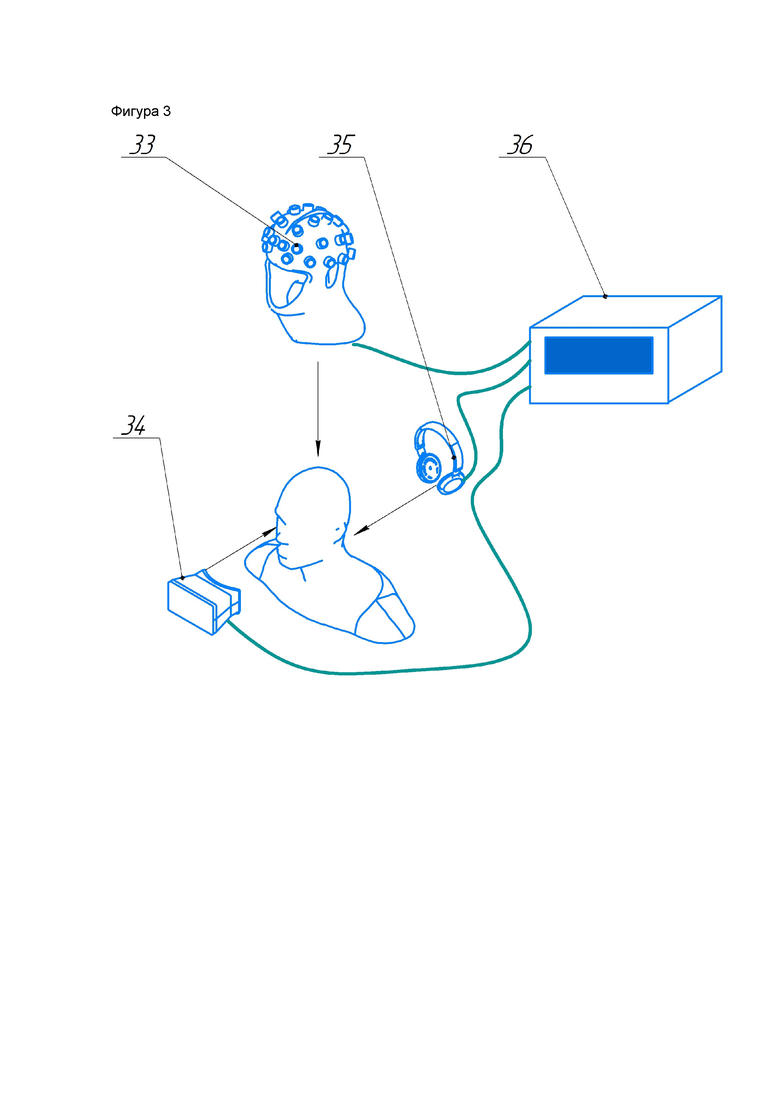
Конструкция системы для стимуляции коры головного мозга для восстановления кратковременной и долговременной памяти в послеинсультный период изображена на фиг. 3. Система для стимуляции коры головного мозга для восстановления кратковременной и долговременной памяти в послеинсультный период включает в себя блок аудиовоздействия 35, блок световоздействия 34, шлем с креплениями для излучателей 33 и блок генерации и управления 36.
Блок магнитного воздействия состоит из шлема с креплениями для излучателей 33 и тридцати двух магнитных излучателей 1-32.
Система работает следующим образом. На блоке управления выбирается алгоритм воздействия, который выставляет: режим работы, характеристики воздействующих сигналов и время воздействия каждого из них. После выставления всех параметров происходит нажатия кнопки «Старт». Далее программно происходит формирование выставленных воздействий в блоке формирования, где происходит преобразование через излучатели в звуковой сигнал. Аналогично сформированные характеристики подаются на rgb-светодиоды, которые преобразовывают воздействие в световой и цветовой сигнал. Аналогично сформированный сигнал подается на катушку-излучатель. Все воздействия или каждое из них заканчивается по истечению времени.
Алгоритмы воздействия для стимуляции нейронных полей и гиппокампа для восстановления кратковременной памяти:
1. Задействуем 6 магнитных излучателей, указанных на рисунке под номерами 15,16,25,27,28,30; частота воздействия соответствует от 4 до 8 Гц (тета ритм), выставляем период воздействия 10 минут. Выставляем тип импульса: синусоида, прямоугольный, треугольный и пилообразный. Расположение излучателей - над гиппокампом (шесть излучателей над ушами под номерами 15,16,25,27,28,30).
•Разница частот на излучателях соответствует тета ритму, время воздействия 6-8 минут. Тип импульса: синусоида, прямоугольный.
• Красный, синий, зеленый, фиолетовый и их сочетания. Последовательное включение различных цветовых комбинаций на оба глаза.
2. Задействуем 6 магнитных излучателей, указанных на рисунке под номерами 15,16,25,27,28,30; используемая начальная частота воздействия 4 Гц, периоды воздействия по 6 минут с последующим сдвигом частоты на 1 Гц при начальной частоте в 4 Гц. Общее количество периодов сдвига равно 5. Выставляем тип импульса: синусоида, прямоугольный, треугольный и пилообразный. Расположение излучателей - над гиппокампом (шесть излучателей над ушами под номерами 15,16,25,27,28,30).
• Разница частот на излучателях соответствует тета ритму, время воздействия 6-8 минут. Тип импульса: синусоида, прямоугольный.
• Красный, синий, зеленый, фиолетовый и их сочетания. Последовательное включение различных цветовых комбинаций на оба глаза.
3. Задействуем 6 магнитных излучателей, указанных на рисунке под номерами 15,16,25,27,28,30; используем частотную модуляцию, девиация частоты модулированного сигнала от 4 до 8 Гц, выставляем Fmin=4 Гц, Fmax=8 Гц, модулирующий сигнал соответствует от 4 до 8 Гц (тета ритм), период воздействия 10 минут. Выставляем тип импульса модулированного сигнала: синусоида, прямоугольный, треугольный и пилообразный. Выставляем тип импульса модулирующего сигнала: синусоида, прямоугольный, треугольный и пилообразный. Расположение излучателей - над гиппокампом (шесть излучателей над ушами под номерами 15,16,25,27,28,30).
• Разница частот на излучателях соответствует тета ритму, время воздействия 6-8 минут. Тип импульса: синусоида, прямоугольный.
• Красный, синий, зеленый, фиолетовый и их сочетания. Последовательное включение различных цветовых комбинаций на оба глаза.
4. Задействуем 6 магнитных излучателей, указанных на рисунке под номерами 15,16,25,27,28,30; используем частотную модуляцию, девиация частоты модулированного сигнала от 0 до 700 Гц, выставляем Fmin=0 Гц, Fmax=700 Гц, модулирующий сигнал соответствует 4 Гц, затем через каждый период воздействия увеличивается на 1 Гц, период воздействия 6 минут, общее количество периодов сдвига равно 5. Выставляем тип импульса модулированного сигнала: синусоида, прямоугольный, треугольный и пилообразный. Выставляем тип импульса модулирующего сигнала: синусоида, прямоугольный, треугольный и пилообразный. Расположение излучателей - над гиппокампом (шесть излучателей над ушами под номерами 15,16,25,27,28,30).
• Разница частот на излучателях соответствует тета ритму, время воздействия 6-8 минут. Тип импульса: синусоида, прямоугольный.
• Красный, синий, зеленый, фиолетовый и их сочетания. Последовательное включение различных цветовых комбинаций на оба глаза.
Алгоритмы воздействия для стимуляции нейронных полей для восстановления долговременной памяти:
5. Задействуем все магнитные излучатели, используем частотную модуляцию, девиация частоты модулированного сигнала от 4 до 8 Гц, выставляем Fmin=4 Гц, Fmax=8 Гц, модулирующий сигнал соответствует от 4 до 8 Гц (тета ритм), период воздействия 10 минут. Выставляем тип импульса модулированного сигнала: синусоида, прямоугольный, треугольный и пилообразный. Выставляем тип импульса модулирующего сигнала: синусоида, прямоугольный, треугольный и пилообразный.
• Разница частот на излучателях соответствует тета ритму, время воздействия 6-8 минут. Тип импульса: синусоида, прямоугольный.
• Красный, синий, зеленый, фиолетовый и их сочетания. Последовательное включение различных цветовых комбинаций на оба глаза.
6. Задействуем все магнитные излучатели, используем частотную модуляцию, девиация частоты модулированного сигнала от 4 до 8 Гц, выставляем Fmin=4 Гц, Fmax=8 Гц, модулирующий сигнал соответствует 4 Гц, затем через каждый период воздействия увеличивается на 1 Гц, период воздействия 6 минут, общее количество периодов сдвига равно 5. Выставляем тип импульса модулированного сигнала: синусоида, прямоугольный, треугольный и пилообразный. Выставляем тип импульса модулирующего сигнала: синусоида, прямоугольный, треугольный и пилообразный.
• Разница частот на излучателях соответствует тета ритму, время воздействия 6-8 минут. Тип импульса: синусоида, прямоугольный.
• Красный, синий, зеленый, фиолетовый и их сочетания. Последовательное включение различных цветовых комбинаций на оба глаза.
7. Задействуем все магнитные излучатели, используем частотную модуляцию, девиация частоты модулированного сигнала от 0 до 200 Гц, выставляем Fmin=0 Гц, Fmax=200 Гц, модулирующий сигнал соответствует от 4 до 8 Гц (тета ритм), период воздействия 10 минут. Выставляем тип импульса модулированного сигнала: синусоида, прямоугольный, треугольный и пилообразный. Выставляем тип импульса модулирующего сигнала: синусоида, прямоугольный, треугольный и пилообразный.
• Разница частот на излучателях соответствует тета ритму, время воздействия 6-8 минут. Тип импульса: синусоида, прямоугольный.
• Красный, синий, зеленый, фиолетовый и их сочетания. Последовательное включение различных цветовых комбинаций на оба глаза.
8. Задействуем все магнитные излучатели, используем частотную модуляцию, девиация частоты модулированного сигнала от 0 до 200 Гц, выставляем Fmin=0 Гц, Fmax=200 Гц, модулирующий сигнал соответствует 4 Гц, затем через каждый период воздействия увеличивается на 1 Гц, период воздействия 6 минут, общее количество периодов сдвига равно 5. Выставляем тип импульса модулированного сигнала: синусоида, прямоугольный, треугольный и пилообразный. Выставляем тип импульса модулирующего сигнала: синусоида, прямоугольный, треугольный и пилообразный.
• Разница частот на излучателях соответствует тета ритму, время воздействия 6-8 минут. Тип импульса: синусоида, прямоугольный.
• Красный, синий, зеленый, фиолетовый и их сочетания. Последовательное включение различных цветовых комбинаций на оба глаза.
9. Задействуем все магнитные излучатели, используем частотную модуляцию, девиация частоты модулированного сигнала от 0 до 700 Гц, выставляем Fmin=0 Гц, Fmax=700 Гц, модулирующий сигнал соответствует от 4 до 8 Гц (тета ритм), период воздействия 10 минут. Выставляем тип импульса модулированного сигнала: синусоида, прямоугольный, треугольный и пилообразный. Выставляем тип импульса модулирующего сигнала: синусоида, прямоугольный, треугольный и пилообразный.
• Разница частот на излучателях соответствует тета ритму, время воздействия 6-8 минут. Тип импульса: синусоида, прямоугольный.
• Красный, синий, зеленый, фиолетовый и их сочетания. Последовательное включение различных цветовых комбинаций на оба глаза.
10. Задействуем все магнитные излучатели, используем частотную модуляцию, девиация частоты модулированного сигнала от 0 до 700 Гц, выставляем Fmin=0 Гц, Fmax=700 Гц, модулирующий сигнал соответствует 4 Гц, затем через каждый период воздействия увеличивается на 1 Гц, период воздействия 6 минут, общее количество периодов сдвига равно 5. Выставляем тип импульса модулированного сигнала: синусоида, прямоугольный, треугольный и пилообразный. Выставляем тип импульса модулирующего сигнала: синусоида, прямоугольный, треугольный и пилообразный.
• Разница частот на излучателях соответствует тета ритму, время воздействия 6-8 минут. Тип импульса: синусоида, прямоугольный.
• Красный, синий, зеленый, фиолетовый и их сочетания. Последовательное включение различных цветовых комбинаций на оба глаза.
Список литературы
[1] Ибрагимов, М. Ф., Хабиров, Ф. А., Хайбуллин, Т. И., Гранатов, Е. В. Современные подходы к реабилитации больных, перенесших инсульт //Практическая медицина. – 2012. – №. 57.
[2] Belik D. V. Equivalent electrochemical ‘wiring diagram’of a neuron //Actual Problems of Electronics Instrument Engineering (APEIE), 2016 13th International Scientific-Technical Conference on. – IEEE, 2016. – Т. 1. – С. 420-422.
[3] Полунина А. Г., Брюн Е. А. Эпизодическая память: неврологические и нейромедиаторные механизмы //Анналы клинической и экспериментальной неврологии. – 2012. – Т. 6. – №. 3.
[4] Dickerson B.C., Eichenbaum H. The episodic memory system: neurocircuitry and disorders. Neuropsychopharmacology Reviews 2010; 35: 86–104.
[5] Markowitsch H.J. Functional neuroanatomy of learning and memory. In: Halligan P.W., Kischka U., Marshall J.C. (eds:). Handbook of Clinical Neuropsychology. New Yourk: Oxford University Press, 2003: 724–730.
[6] Nadel L. The parahippocampal region: basic and clinical implications. Hippocampus 2000; 10: 133–135.
[7] Takashima A., Nieuwenhuis I.L.C., Jensen O. et al. Shift from hippocampal to neocortical centered retrieval network with consolidation. J. Neurosc. 2009; 29 (32): 10087–10093.
[8] Wang J.X., Poe G., Zochowski M. From network heterogeneities to familiarity detection and hippocampal memory management. Phys. Rev. E Stat. Nonlin. Soft Matter Phys. 2008; 78 (4 Pt 1): 041905.
[9] Plessen K.J., Bansal R., Zhu H. et al. Hippocampus and amygdale morphology in attention-deficit/hyperactivity disorder. Arch. Gen. Psychiatry 2006; 63: 795–807.
[10] Karlsson M.P., Frank L.M. Awake replay of remote experiences in the hippocampus. Nat. Neurosci. 2009; 12 (7): 913–918.
[11] Robertson E.M. From creation to consolidation: a novel framework for memory processing. PLoS Biology 2009; 7: e1000019.
[12] Walker M.P. The role of slow wave sleep in memory processing. J. Clin. Sleep Med. 2009; 5 (Sup. 2): S20–S26.
[13] De Toledo-Morrell L., Dickerson B., Sullivan M.P. et al. Hemispheric differences in hippocampal volume predict verbal and spatial memory performance in patients with Alzheimer's disease. Hippocampus 2000; 10: 136–142.
[14] Невская А. П. и др. БИОХИМИЧЕСКИЕ МЕХАНИЗМЫ ДОЛГОВРЕМЕННОЙ ПАМЯТИ //Тверской медицинский журнал. – 2016. – №. 5. – С. 11-14.
[15] Аршавский Ю. И. Нейронные механизмы памяти: синаптическая и геномная гипотезы //Журнал высшей нервной деятельности им. ИП Павлова. – 2011. – Т. 61. – №. 6. – С. 660-660.
[16] Аршавский Ю.И. И.М. Гельфанд о математике и нейрофизиологии. Вестн. РАН. 2010. 80: 937–941.
[17] Аршавский Ю.И. О роли нейронных сетей и индивидуальных нейронов в работе мозга. Сенсорные системы. 2011. 25: 3–16.
[18] Hydén H., Egyhazi E. Changes in RNA content and base composition in cortical neurons of rats in a learning experiment involving transfer of handedness //Proceedings of the National Academy of Sciences of the United States of America. – 1964. – Т. 52. – №. 4. – С. 1030.
[19] Hyden H., Egyhazi E. Nuclear RNA changes of nerve cells during a learning experiment in rats //Proceedings of the National Academy of Sciences of the United States of America. – 1962. – Т. 48. – №. 8. – С. 1366.
[20] Hyden H. Behavior, neural function, and RNA //Progress in nucleic acid research and molecular biology. – Academic Press, 1967. – Т. 6. – С. 187-218.
[21] Hydén H. et al. Changes of RNA base composition in nerve cells of monkeys subjected to visual discrimination and delayed alternation performance //Brain research. – 1974. – Т. 65. – №. 2. – С. 215-230.
[22] Ungar G. et al. Brain peptides with opiate antagonist action: Their possible role in tolerance and dependence //Psychoneuroendocrinology. – 1977. – Т. 2. – №. 1. – С. 1-10.
[23] Ungar G., Ungar A. L., Malin D. H. Brain peptides with opiate antagonist activity //Opiates and Endogenous Opioid Peptides. – Elsevier Amsterdam, 1976. – С. 121-128.
[24] Ungar G., Ungar A. L. ENDOGENOUS MORPHINE ANTAGONIST IN MORPHINE-TREATED RAT-BRAIN //FEDERATION PROCEEDINGS. – 9650 ROCKVILLE PIKE, BETHESDA, MD 20814-3998 USA : FEDERATION AMER SOC EXP BIOL, 1976. – Т. 35. – №. 3. – С. 309-309.
[25] Ungar G. Molecular mechanisms in information processing //International Review of Neurobiology. – Academic Press, 1970. – Т. 13. – С. 223-253.
[26] Ungar G. Molecular organization of neural information processing //The Structure and Function of Nervous Tissue. – 2012. – Т. 4. – С. 215-247.
[27] Lynch G., Baudry M. The biochemistry of memory: a new and specific hypothesis //Science. – 1984. – Т. 224. – №. 4653. – С. 1057-1063.
[28] Baudry M. et al. Development of glutamate binding sites and their regulation by calcium in rat hippocampus //Developmental Brain Research. – 1981. – Т. 1. – №. 1. – С. 37-48.
[29] Maren S., Baudry M. Properties and mechanisms of long-term synaptic plasticity in the mammalian brain: relationships to learning and memory //Neurobiology of learning and memory. – 1995. – Т. 63. – №. 1. – С. 1-18.
[30] Kandel E.R. Cellular Mechanisms of Learning and the Biological Basis of Individuality. In: E.R. Kandel, J.H. Schwartz, and T.M. Jessell (Eds.). Principles of Neural Science, 4th Ed. McGraw6Hill Companies, Inc. 2000. pp. 1247—1279.
[31] Sherman S.M., Guillery R.W. Functional connections of cortical areas: a new view from the thalamus. Cambridge: The MIT Press. 2013. 296 p.
[32] Hindriks R., van Putten M.J.A.M. Thalamo6cortical mechanisms underlying changes in amplitude and frequency of human alpha oscillations. Neuroimage, 2013. Vol. 70, pp. 150— 163. doi: 10.1016/j.neuroimage.2012.12.018.
[33] Neuronal Mechanisms and Attentional Modulation of Corticothalamic Alpha Oscillations. Bollimunta A., [et al.]. The Journal of Neuroscience, 2011. Vol. 31, no. 13, pp. 4935—4943; doi:10.1523/JNEUROSCI.5580610.2011.
[34] Finding thalamic BOLD correlates to posterior alpha EEG. Liu Z., [et al.]. Neuroimage, 2012. Vol. 63, no. 3, pp. 1060—1069. doi: 10.1016/j.neuroimage.2012.08.025.
[35] Intrinsic connectivity networks, alpha oscillations, and tonic alertness: a simultaneous electroencephalography/functional magnetic resonance imaging study. Sadaghiani S., [et al.]. J. Neurosci, 2010. Vol. 30, pp 10243—10250. doi: 10.1523/JNEUROSCI.1004610.2010.
[36] Buzsaki G. Theta oscillations in the hippocampus // Neuron. 2002. Vol. 33, № 3. P. 325— 340. doi: 10.1016/S089666273(02)005866X.
[37] Hasselmo M.E., Stern C.E. Theta rhythm and the encoding and retrieval of space and time // Neuroimage. 2014. Vol. 85. P. 656—666. doi: 10.1016/j.neuroimage.2013.06.022.
[38] Burgess N., Maguire EA, O'Keefe J. The human hippocampus and spatial and episodic memory // Neuron. 2002. Vol. 35, № 4. P. 625—641. doi: 10.1016/S089666273(02)0083069.
[39] Lega B.C., Jacobs J., Kahana M. Human hippocampal theta oscillations and the formation of episodic memories // Hippocampus. 2012. Vol 22, № 4. P. 748—761. doi: 10.1002/ hipo.20937.
[40] Duvernoy H.M. The human hippocampus: functional anatomy, vascularization and serial section with MRI. 3rd ed. New York: Springer, 2005. 232 p.
[41] Vinogradova O.S., Kitchigina V.F., Zenchenko C.I. Pacemaker neurons of the forebrain medical septal area and theta rhythm of the hippocampus // Membr. Cell. Biol. 1998. Vol. 11, № 6. P. 715—725.
[42] Synaptic kainate receptors in CA1 interneurons gate the threshold of theta6frequencyinduced long6term potentiation / Clarke V.R.J., [et al.] // J Neurosci. 2012 Vol. 32, № 50. P. 18215—26. doi: 10.1523/JNEUROSCI.2327612.2012.
[43] Control mechanisms in working memory: A possible function of EEG theta oscillations / Sauseng P., [et al.] // Neuroscience and Biobehavioral Reviews. 2010. Vol. 34. P. 1015 — 1022. doi:10.1016/j.neubiorev.2009.12.006.
[44] Новикова С. И. Ритмы ЭЭГ и когнитивные процессы //Современная зарубежная психология. – 2015. – Т. 4. – №. 1. – С. 91-108.
[45] Максимов А. В., Кирьянова В. В., Максимова М. А. Лечебное применение магнитных полей //Физиотерапия, бальнеология и реабилитация. – 2013. – №. 3.
[46] Xu Y. et al. Collective responses in electrical activities of neurons under field coupling //Scientific reports. – 2018. – Т. 8. – №. 1. – С. 1349.
[47] Chang W. H. S. et al. Effect of pulse‐burst electromagnetic field stimulation on osteoblast cell activities //Bioelectromagnetics: Journal of the Bioelectromagnetics Society, The Society for Physical Regulation in Biology and Medicine, The European Bioelectromagnetics Association. – 2004. – Т. 25. – №. 6. – С. 457-465.
[48] Stanley S. A., Friedman J. M. Electromagnetic regulation of cell activity //Cold Spring Harbor perspectives in medicine. – 2019. – Т. 9. – №. 5. – С. a034322.
[49] Piacentini R. et al. Extremely low‐frequency electromagnetic fields promote in vitro neurogenesis via upregulation of Cav1‐channel activity //Journal of cellular physiology. – 2008. – Т. 215. – №. 1. – С. 129-139.
[50] Lv M., Ma J. Multiple modes of electrical activities in a new neuron model under electromagnetic radiation //Neurocomputing. – 2016. – Т. 205. – С. 375-381.
[51] Urnukhsaikhan E. et al. Neuroprotective effect of low frequency-pulsed electromagnetic fields in ischemic stroke //Applied biochemistry and biotechnology. – 2017. – Т. 181. – №. 4. – С. 1360-1371.
[52] Gessi S. et al. Pulsed electromagnetic field and relief of hypoxia‐induced neuronal cell death: The signaling pathway //Journal of cellular physiology. – 2019. – Т. 234. – №. 9. – С. 15089-15097.
[53] Vincenzi F. et al. Pulsed Electromagnetic Field Exposure Reduces Hypoxia and Inflammation Damage in Neuron‐Like and Microglial Cells //Journal of cellular physiology. – 2017. – Т. 232. – №. 5. – С. 1200-1208.
[54] Mueller JK, Grigsby EM, Prevosto V, Petraglia FW, Rao H, Deng Z-D, et al. Simultaneous transcranial magnetic stimulation and single-neuron recording in alert non-human primates. Nat Neurosci 2014;17:1130e6. https://doi.org/10.1038/nn.3751.
[55] Li B, Virtanen JP, Oeltermann A, Schwarz C, Giese MA, Ziemann U, et al. Lifting the veil on the dynamics of neuronal activities evoked by transcranial magnetic stimulation. Elife 2017;6:e30552. https://doi.org/10.7554/ eLife.30552.
[56] Romero MC, Davare M, Armendariz M, Janssen P. Neural effects of transcranial magnetic stimulation at the single-cell level. Nature Communications 2019;10(1):2642. https://doi.org/10.1038/s41467-019-10638-7.
[57] Murphy SC, Palmer LM, Nyffeler T, Müri RM, Larkum ME. Transcranial magnetic stimulation (TMS) inhibits cortical dendrites. Elife 2016;5:1e12. https://doi.org/10.7554/eLife.13598.
[58] Chen T-W, Wardill TJ, Sun Y, Pulver SR, Renninger SL, Baohan A, et al. Ultrasensitive fluorescent proteins for imaging neuronal activity. Nature 2013;499:295.
[59] Murthy VN, Sejnowski TJ, Stevens CF. Dynamics of dendritic calcium transients evoked by quantal release at excitatory hippocampal synapses. Proc Natl Acad Sci U S A 2000;97:901e6. https://doi.org/10.1073/pnas.97.2.901.
[60] Aberra A. S. et al. Simulation of transcranial magnetic stimulation in head model with morphologically-realistic cortical neurons //Brain stimulation. – 2020. – Т. 13. – №. 1. – С. 175-189.
[61] Белик Д. В., Дмитриев Н. А., Пустовой С. А., Зиневская М.С. Техническая система для стимуляции полей памяти мозга и гиппокампа с целью реабилитации в послеинсультный период //Биотехносфера. – 2015. – №. 6. – С. 20-25.
| название | год | авторы | номер документа |
|---|---|---|---|
| УСТРОЙСТВО АУДИОВИЗУАЛЬНОЙ СТИМУЛЯЦИИ КОРЫ ГОЛОВНОГО МОЗГА | 2022 |
|
RU2780128C1 |
| Способ реабилитации после тотального эндопротезирования коленного сустава у пациентов с саркопенией | 2018 |
|
RU2684178C1 |
| НЕМЕДИКАМЕНТОЗНЫЙ СПОСОБ ЛЕЧЕНИЯ АДИНАМИЧЕСКИХ ДЕПРЕССИЙ | 2022 |
|
RU2801161C1 |
| СПОСОБ ЛЕЧЕНИЯ УМСТВЕННОЙ ОТСТАЛОСТИ У ДЕТЕЙ ПО В.В. БУТУХАНОВУ | 2004 |
|
RU2281793C2 |
| Способ стимуляции головного мозга | 2023 |
|
RU2822811C1 |
| СПОСОБ ЛЕЧЕНИЯ БОЛЬНЫХ В КОМЕ И ВЕГЕТАТИВНОМ СОСТОЯНИИ | 2000 |
|
RU2197294C2 |
| СПОСОБ ЛЕЧЕНИЯ ЭНЦЕФАЛОПАТИИ У ДЕТЕЙ | 2012 |
|
RU2501584C1 |
| Способ коррекции зрелости ритмов головного мозга | 2023 |
|
RU2839010C1 |
| СПОСОБ ЛЕЧЕНИЯ КОГНИТИВНЫХ РАССТРОЙСТВ ПРИ ХРОНИЧЕСКОЙ ЭНЦЕФАЛОПАТИИ У БОЛЬНЫХ БРОНХИАЛЬНОЙ АСТМОЙ | 2005 |
|
RU2308885C1 |
| Способ неинвазивной слаботочной электростимуляции структур головного мозга | 2023 |
|
RU2820133C1 |
Изобретение относится к медицинской технике. Система осуществляет воздействие на рецепторные системы организма: глаза с помощью RGB-светодиодов, установленных в маске, на слуховые рецепторы с помощью стереонаушников и воздействие магнитным полем локально на кору головного мозга с помощью тридцати двух излучателей магнитных воздействий, закрепленных в шлеме на голове пациента. Устройство позволяет проводить стимуляцию нейронных полей головного мозга посредством выставления разницы частот на всех излучателях, которые кратны или близки к ритмам головного мозга (альфа, бета, дельта, тета, ритмы), а также цветовыми комбинациями с подачей разных цветов на каждый глаз и различными частотными модуляциями воздействующего сигнала магнитного поля. Данные типы воздействия на кору головного мозга и гиппокамп в частотном диапазоне от 0,1 Гц до 700 Гц позволяют проводить стимуляцию кратковременной памяти и стимуляцию долговременной памяти для реабилитации больного в послеинсультный период. 2 з.п. ф-лы, 3 ил.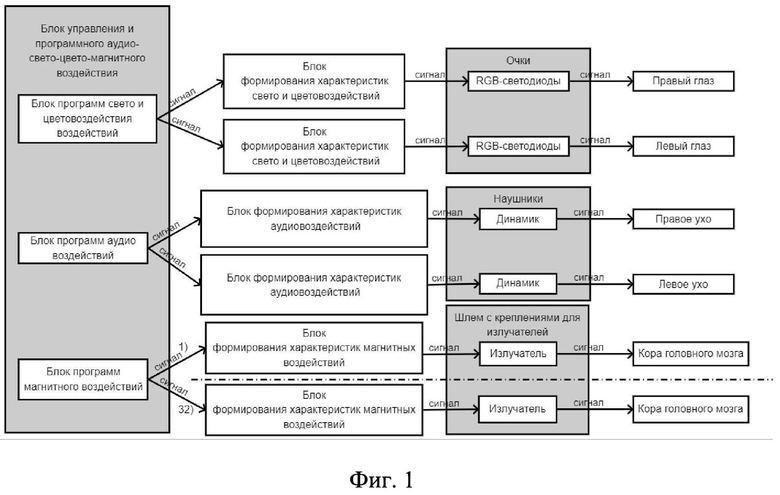
1. Система стимуляции коры головного мозга для восстановления кратковременной и долговременной памяти в послеинсультный период включает в себя блок управления и программного аудио-свето-цвето-магнитовоздействия, состоящий из блока программ аудиовоздействий, блока программ свето- и цветовоздействий, блока программ магнитного воздействия, блоков формирования характеристик аудио-, свето-, цвето- и магнитных воздействий, излучателей аудиовоздействий, выполненных с возможностью закрепления на каждом ухе, излучателей свето- и цветовоздействий, выполненных с возможностью закрепления на каждом глазу, тридцати двух излучателей магнитных воздействий, закрепленных в шлеме на голове пациента.
2. Система по п.1, отличающаяся тем, что излучатели аудиовоздействий выполнены в виде стереонаушников.
3. Система по п.1, отличающаяся тем, что излучатели свето- и цветовоздействий выполнены в виде очков.
| WO 2015131770 A1, 11.09.2015 | |||
| Способ тренировки головного мозга | 2017 |
|
RU2671199C1 |
| ГЕНЕРАТОР ФРАКТАЛЬНЫХ МЕЛЬКАНИЙ ДЛЯ БИОМЕДИЦИНСКИХ ИССЛЕДОВАНИЙ | 2014 |
|
RU2549150C1 |

