Перекрестная ссылка на родственные заявки
В настоящей заявке заявляется приоритет на основании предварительной заявки США №61/792198, поданной 15 марта 2013 г, согласно 35 U.S.C. 119(e). Полное описание предварительной заявки на патент США №61/792198, поданной 15 марта 2013 г., включено в настоящий документ посредством ссылки.
Ссылка на перечень последовательностей
Настоящая заявка содержит перечень последовательностей, представленный электронно в виде текстового файла по EFS-Web. Текстовый файл под названием «7579-1-PCT_sequence_listing_ST25» имеет размер 18 КБ и был сохранен 17 марта 2014 года. Информация, содержащаяся в текстовом файле, включена в настоящий документ посредством ссылки в полном объеме в соответствии с 37 CFR §1.52(e)(5).
Область техники
Настоящее изобретение в целом относится к применению белка или пептида, содержащих моноцистеиновый активный центр тиоредоксина в восстановленном состоянии для снижения вязкости и для индукции, усиления и/или увеличения разжижения слизи или мокроты.
Уровень техники
Существует значительная неудовлетворенная медицинская потребность в безопасных, хорошо переносимых и эффективных лекарствах для лечения пациентов с муковисцидозом (MB), ХОБЛ/эмфиземой, бронхоэктатической болезнью, тяжелой формой астмы и другими тяжелыми обструктивными заболеваниями легких. Эти заболевания характеризуются избыточной продукцией вязкой слизи, что приводит к нарушению функции легких (см. обзор Evans, СМ. and Koo, J.S., Pharmacology & Therapeutics 121: 332-348, 2009). Недостаточное выведение аномальной клейкой слизи ассоциируется с хронической инфекцией и преждевременной смертью, особенно при MB. Несмотря на успехи антибактериальной терапии и других видов лечения, улучшение выведения слизи остается основной целью клинического лечения, даже несмотря на то, что понимание механизмов, лежащих в основе транспортабельности слизи, остается ограниченным (Verdugo, P., Cold Spring Harb Perspect Med 2012; 2:a009597).
Слизь является непрерывно секретируемым супрамолекулярным полимерным гелем, который образует защитный барьер на поверхности эпителия и отвечает, посредством движения ресничек и кашля, за транспортировку вдыхаемых инородных частиц и бактерий из легких (Knowles, M.R. and Boucher, R.C., J Clin Invest 109:571-577, 2002; Cone, R.A., Adv Drug Deliv Rev 61:75-85, 2009). Поэтому надлежащая вязкоупругость и гидратация слоя слизи, обеспечивающие эффективный мукоцилиарный транспорт, имеют решающее значение для функционирования слизи и профилактики инфекции и воспаления. Нормальная слизь состоит главным образом из воды (97%); остальные твердые вещества содержат белки муцины, немуциновые белки, соли, липиды и клеточный дебрис (Fahy, J.V. and Dickey, B.F., N Engl J Med 363:2233-47, 2010). Полимерные гликопротеины муцина MUC5AC и MUC5B несут основную ответственность за вязкоупругие свойства геля слизи дыхательных путей (Matsui et al., Cell 95:1005-1015, 1998; reviewd in Kreda, et al., Cold Spring Harb Perspect Med 2012; 2:a009589). Гидроксильные группы О-связанных гликанов, присоединенные к муцинам, участвуют в связывании воды, в то время как сами муцины образуют переплетенную сеть (Verdugo et al., Biorheology 20:223-230, 1983), которая может также включать ковалентное и нековалентное межцепочечное перекрестное связывание, согласно результатам подробных исследований муцина MUC2 пищеварительного тракта (Ambort et al., Biochem J 436:61-70, 2011). Муцины необычайно богаты аминокислотой Cys; примечательно, что MUC5AC человека содержит 295 остатков Cys из в общей сложности 5030 аминокислот (www.uniprot.org/uniprot/P98088). Полагают, что остатки Cys муцина, расположенные вблизи N- и С-концов, участвуют в формировании межцепочечных дисульфидных связей между субъединицами муцина, в то время как роль внутренних остатков Cys менее ясна (Thornton et al., Annu Rev Physiol 70:459-486, 2008). Некоторые из них расположены в области «Cys-узла» и могут легко образовывать внутримолекулярные дисульфидные связи, которые могут играть определенную роль в усилении нековалентного переплетения, важного для сетчатой структуры геля слизи (Fahy, J.V. and Dickey, B.F., N Engl J Med 363:2233-47, 2010).
Недавняя работа (Button et al., Science 337:937-941, 2012) привела к разработке новой модели структуры поверхности слизистой на основании открытия того, что некоторые муцины, считавшиеся связанными с мембраной эпителиальных клеток, фактически связаны с мембранами самих ресничек. Эта модель подразумевает, что подвижный слой слизи покрывает более плотный околоресничный слой, что описывается как «гель на щетке». Эта модель элегантно объясняет то, как жидкость перемещается между двумя слоями, при этом слизь выступает в качестве резервуара, также данная модель устанавливает новую парадигму для понимания роли осмотического коэффициента слизи при определении функционального назначения мукоцилиарного транспорта и гидратации слоя слизи. Модель также обеспечивает рамки для понимания того, как избыток дисульфидных связей в каркасе белков слизи может вызвать увеличение осмотического коэффициента слоя слизи, что, в свою очередь, обезвоживает подстилающий ресничный слой и серьезно ограничивает нормальный транспорт слизи. Такой сценарий может в значительной степени обуславливать патологический механизм MB.
MB является аутосомно-рецессивным состоянием. Симптомы MB обусловлены дефектами в МВТР, регуляторе трансмембранной проводимости при муковисцидозе, ключевом транспортере в мембране эпителия одновалентных отрицательно заряженных ионов, главным образом, хлорида (Riordan, et al., Science 245: 1066-1073), а также бикарбоната и глутатиона. Мутации, ведущие к MB, из которых известно более 1700 (www.genet.sickkids.on.ca/MBTP/), включают мутации, вызывающие полную потерю МВТР (причина наиболее распространенного генотипа MB), а также точечные мутации, приводящие к частичной или полной потере анион-транспортной активности. Кроме того, в результате дефектов в МВТР эпителии в организме являются непроницаемыми для транспорта хлорид-ионов (Boucher eta l., Lung 161:1-17, 1983; Welsh, Physiol Rev 67:11443-1184, 1987). Хотя поражаются несколько органов, в том числе поджелудочная железа, кишечник и мужские половые органы, легочные осложнения являются причиной 95% заболеваемости и смертности приходится на (Means, М. Cystic Fibrosis: the first 50 years. In: Cystic Fibrosis-Current Topics Volume 1, edited by Dodge JA, Brock DJH, and Widdicombe JH. Chichester: Wiley and Sons, 1992, p. 217-250). В легких, пораженных заболеванием, транспорт хлорида в просвет дыхательных путей приводит к избыточному поглощению Na+ и жидкости, снижая тем самым объем поверхностной жидкости дыхательных путей (Jiang et al., Science 262:424-427, 1993). В то же время, попытки восстановить активность хлоридных каналов с целью компенсации эффектов, вызванных нефункционирующим МВТР, например, за счет агонистов пуринергического рецептора подтипа P2Y2, были неудачными (Ratjen, F. et al., J Cyst Fibres 11:539-49, 2012). Это свидетельствует о том, что активность МВТР, не связанная с хлоридом, может быть более значительной, чем предполагалось первоначально.
В окислительных средах, например, в легких, дисульфидные связи легко образуются между соседними окисленными остатками Cys, например, такими, которые в изобилии присутствуют на белках муцина. Эти связи являются высокостабильными, и для их разрушения (т.е. восстановления) с целью восстановления свободной тиоловой формы остатков Cys требуется действие сильных химических или биологических восстановителей. В здоровых легких избыточному образованию дисульфидных связей противодействует главным образом восстановленная форма биологического восстановителя глутатиона (GSH), Cys-содержащего трипептида, который в больших количествах секретируется в слой слизи (Cantin et al., J Appl Physiol 63:152-157, 1987) и может играть ключевую роль в поддержании нормального равновесия дисульфидных связей и свободных тиоловых групп Cys в муцинах. Секреция GSH на поверхность дыхательных путей сильно зависит от МВТР, который и прямо, и косвенно способствует экспорту GSH (см. обзор Ballatori et al., Biol Chem 390: 191-214, 2009). Следовательно, уровень GSH в легких пациентов с MB может составлять 30% или менее от указанных уровней у здоровых индивидуумов (Roum et al., J Appl Physiol 75:2419-24, 2003; Wetmore D.R. et al., J Biol Chem 285:30516-22, 2010). МВТР также отвечает за секрецию бикарбонат-анионов, и обусловленный этим дефицит бикарбоната в легких при MB, по-видимому, способствует заболеванию. Основная химическая активность бикарбоната заключается в повышение pH. Поскольку восстановление дисульфидных связей тиол-содержащими восстановителями требует формирования атакующего депротонированного тиолата, который ингибируется при низких pH, благоприятствующих протонированной форме тиола (Singh and Whitesides, In: Sulphur-containing Functional Groups, 5: pp. 633-58, John Wiley & Sons, 1993), вероятна синергия между активностями бикарбоната и GSH (а также других биологических восстановителей, заведомо присутствующих в среде поверхности дыхательных путей). pH, измеренный в трахеобронхиальном секрете при MB, на 0,6 единиц ниже, чем у здоровых индивидуумов Song et al., Am J Physiol Cell Physiol 290:C741-C749), что согласуется со средой в легких при MB, где выделение восстановителя ограничено и где он также менее активен вследствие нарушения способности образовывать тиолат, атакующий дисульфидные связи. В совокупности с огромным количеством кластеризованного Cys в муцинах, слизь в окислительных условиях дыхательных путей готова перейти в состояние с повышенным количеством дисульфидных связей, если уровень секретируемого восстановителя становится ограничивающим фактором или если белки муцины продуцируются и секретируются в избытке, что приводит к избыточности Cys, которые могут образовывать дисульфидные связи. Известно, что обе ситуации имеют место при MB и некоторых других обструктивных заболеваниях легких: белки слизи сверхпродуцируются в ответ на стресс в легких ((Rogers, Resp Care 52:1134-1149), а секреция GSH может блокироваться на 70% или более в результате дефектов МВТР (Roum et al., J Appl Physiol 75:2419-24, 2003; Wetmore D.R. et al., J Biol Chem 285:30516-22, 2010).
Эта возможность избыточного образования дисульфидных связей в слизи, а также общий окислительно-восстановительный дисбаланс, играющий механистическую роль при MB, привели к клинической оценке различных тиол-содержащих агентов в качестве муколитических лекарственных средств. Указанные средства включают N-ацетилцистеин (NAC) и Nacystelyn (NAL, N-ацетилцистеин + L-лизин) (Hurst et al., Am Rev Respir Dis, 96:962-970, 1967; Dasgupta and King, Pediatr Pulmonol, 22:161-166, 1996; Nash, E.F., et al., Cochrane Database of Systematic Reviews, 2010(12): 1-49, 2009), а также собственно восстановленный глутатион (Bishop, С., et al., CHEST Journal, 127(1): 308-317, 2005; Griese, M., et al., Am J Resp Crit Care Med 169(7):822-828, 2004; Griese, M., et al., Am J Resp Crit Care Med 188(1):83-89, 2013; Roum, J.H., et al., J Appl Physiol, 87:438-443, 1999). Хотя эти низкомолекулярные агенты по большей части безопасны, на сегодняшний день они не демонстрируют явного клинического преимущества в преоральной или ингаляционной форме (см. обзор Nash, E.F., et al., Cochrane Database of Systematic Reviews, 2010(12): 1-49, 2009). Эта недостаточная эффективность во многом может быть обусловлена их низкой мощностью или потерей активности во время доставки из-за аутоокисления, а также возможностью инактивации ферментами легких. GSH подвергается быстрому аутоокислению в неактивную форму GSSG (Curello, S. et al., Clin Chem, 33:1448-49, 1987) и, следовательно, фармакологически неустойчив в восстановленной форме при введении путем ингаляции в виде аэрозоля (Carl White M.D., частное сообщение), теряя большую часть своей активности в процессе достижения области-мишени в дыхательных путях. Кроме того, γ-глутамилтрансфераза, присутствующая в высоких концентрациях в легких, легко разлагает GSH до неактивной формы (Corti et al., Am J Resp Crit Care Med 189:233-234, 2014), содержание которой резко возрастает после ингаляции GSH (Griese et al., Am J Resp Crit Care Med 188:83-89 Supplemental information, 2013). Таким образом, усовершенствование тиоловых агентов путем объединения направленного действия на дисульфидные связи с превосходными фармакологическими свойствами и специфичностью биологических лекарственных средств является ключевой нерешенной терапевтической задачей.
Хотя этиологию заболеваний легких при MB можно объяснить изменениями реологических свойств слизи, нарушения функции легких редко проявляются при рождении. Бронхоэктаз и обструкция дыхательных путей прогрессируют с возрастом пациента. Это хронические повреждения легких обусловлены непрерывным циклом бактериальной инфекции и воспалительной реакции. Повреждение дыхательных путей возникает, когда нейтрофилы, рекрутированные в легких, высвобождают ферменты, разрушающие матрикс, например, эластазу, а также вредоносные реакционноспособные формы кислорода (см. обзор Konstan and Berger, Pediatr Pulmonol 24:137-142, 1997). В ходе стойкой инфекции также может происходить взаимодействие муцинов с ДНК (Potter et al., Am J Dis Child 100:493-495, 1960; Lethem et al., Am Rev Respir Dis 100:493-495, 1990; Lethem et al., Eur Respir J 3:19-23, 1990) и полимерами f-актина (Sheils et al., Am J Path 148:919-927, 1996; Tomkiewicz et al., DNA and actin filament ultrastructure in cystic fibrosis sputum. In: Cilia, mucus, and mucociliary interactions, edited by Baum GL, Priel Z, Roth Y, Liron N, and Ostfeld EJ. New York, NY: Marcel Dekker, 1998), высвобожденными из гибнущих воспалительных клеток, которое в некоторых случаях может являться причиной плотного и вязкого характера мокроты при тяжелой форме MB. Невозможность выведения такой слизи путем кашля или мукоцилиарного клиренса способствует дальнейшей колонизации легких условно-патогенными микроорганизмами, ремоделированию дыхательных путей и в конечном итоге смерти.
Поэтому мероприятия, призванные напрямую смягчать последствия дефектов МВТР, являются особенно желательными, так как они могут предотвратить или облегчить прогрессирование заболевания. Хотя прямая коррекция MB путем генной терапии еще недостижима, недавно продемонстрировано применение потенцирующих и корректирующих терапевтических средств, в некоторой степени позволившее восстановить функцию дефектных белков МВТР (Sloane, PA and Rowe, SM, Current Opinion in Pulmonary Medicine 16: 591-7, 2010). Такая терапия ограничена небольшим процентом пациентов с MB, вызванным конкретным дефектом МВТР, например, мутацией G551D, которая является мишенью ивакафтора/Kalydeco™ (Jones, AM and Helm, JM, Drugs 69: 1903-10, 2009). Однако у этих нескольких человек наблюдались впечатляющие результаты (Accurso, FJ; Rowe, SM; Clancy, JP; Boyle, MP; Dunitz, JM; Durie, PR; Sagel, SD; Hornick, DB et al., The New England Journal of Medicine 363: 1991-2003, 2010), демонстрирующие, что механистическое вмешательство при MB способно смягчить последствия поздней стадии, например, возникающие в результате хронической инфекции и воспаления. Вместе с тем, в настоящее время основой лечения прогрессирующих заболеваний дыхательных путей остаются симптоматические, а не направленные на заболевание подходы, в том числе схемы антибиотикотерапии в комбинации с лекарственными средствами, облегчающими выведение гнойных выделений дыхательных путей. Ингаляцию очищенной рекомбинантной ДНКазы человека (Pulmozyme™; Genentech, США), гидролизующей внеклеточную ДНК, присутствующую в дыхательных путях при MB, широко применяют в качестве респираторного противозастойного средства. Такое лечение клинически эффективно снижает вязкость мокроты и стабилизирует объем форсированного выдоха (ОФВ) (Fuchs et al., N Engl J Med 331:637-642, 1994). Другие исследуемые терапевтические средства, направленные на разрушение полимеров муцина или актина, включая N-ацетилцистеин (NAC), nacystelyn (производное N-ацетил L-цистеина) и гельсолин, в экспериментах также могут снижать вязкость мокроты, однако пока еще не продемонстрировали клиническую эффективность и не утверждены для лечения MB в Соединенных Штатах (Nash, EF et al., Cochrane Database of Systematic Reviews, 2010(1):CD007168, 2009).
Другие подходы, применяемые для улучшения выведения слизи, включают мукоактивные агенты, например, гипертонический раствор хлорида натрия для ингаляций и маннит в высокой дозе для ингаляций (Fahy, J.V. and Dickey, B.F., N Engl J Med 363:2233-47, 2010). Считается, что эти агенты действуют, осмотически вытягивая воду в слой слизи для усиления гидратации, или для улучшения выведения за счет индукции рефлексов кашля. Существуют данные в пользу обоих механизмов (Levin, М.Н. et al., J Biol Chem 281:25803-12, 2006; Boucher, R.C., Trends Mol Med 13:231-240, 2007). Вместе с тем, мукоактивные вещества представляют собой симптоматические средства (не влияющие на заболевание), и, как правило, обладают лишь умеренной эффективностью, поскольку многие пациенты не могут переносить высокие дозы, которые могут обеспечивать наибольший клинический эффект (Aziz, I. and Kastelik, J.A., N Engl J Med 354:1848-1851, 2006).
White и соавторы (Rancourt et al., Am J Physiol Lung Cell Mol Physiol 286:L931-L938, 2004; Rancourt et al., Free Radical Biol & Med 42:1441-43, 2007) обнаружили, что применение белка или пептида, содержащих активный центр тиоредоксина в восстановленном состоянии, усиливает разжижение слизи и мокроты у пациента с чрезмерно вязкой или липкой слизью или мокротой, в том числе у пациентов с MB, у которых слизь и мокрота находятся в контакте с белком или пептидом (патент США №7,195,766 и патент США №7,534,438, оба из которых полностью включены в настоящий документ посредством ссылок). В этой системе (см. фигуру. 3) образуется переходная смешанная дисульфидная связь между N-концевым остатком цистеина активного центра тиоредоксина и остатком цистеина белка-мишени (находящегося в слизи и мокроте) с последующей немедленной нуклеофильной атакой на внутримолекулярную смешанную дисульфидную связь и высвобождением окисленного тиоредоксина и полностью восстановленной мишени (Wynn et al. Biochemistry 34(37): 11807-11813, 1995), что обеспечивает повторное образование дисульфидов цистеина в слизи и мокроте, однако в то же время обеспечивает свободный доступ восстановленного или окисленного тиоредоксина в клетки и индуцирует нежелательную постороннюю активность после повторного восстановления эндогенной системой тиоредоксинредуктаза - NADPH. Кроме того, White и соавторы показали, что восстановленный тиоредоксин снижает аномальную вязкоупругость слизи человека при MB при имплантационных исследованиях in vitro и ex vivo на животных (Rancourt et al., Free Radical Biol & Med 42:1441-43, 2007), а также ингибирует активность эластазы провоспалительных нейтрофилов, разрушая дисульфидные свяхи в активном центре (Lee et al., Am J Physiol Lung Cell Mol Physiol 289:L875-L882, 2005). По сравнению с GSH и тиоловыми агентами, например, NAC, тиоредоксин является более мощным восстановителем дисульфидных связей и гораздо менее чувствителен к инактивации за счет аутоокисления. В совокупности это создает возможность для восстановления нормального восстановленного состояния дисульфидов слизи с применением фармакологически стабильного вещества. Такая терапия может предотвратить или отсрочить каскад хронической инфекции, воспаления и снижения функции легких, который приводит к ранней смерти у пациентов с MB. В то же время существует веская причина избегать возможных провоспалительных и других внутриклеточных регуляторных эффектов тиоредоксина (Arner, E.S. and A. Holmgren, Eur J Biochem 267: 6102-6109, 2000; Rancourt et al., Free Radical Biol & Med 42:1441-43, 2007), а также увеличить эффективность регуляции вязкости слизи, предотвращая повторное окисление Cys муцина. Эти усовершенствования являются предметом настоящего изобретения.
Сущность изобретения
Один вариант реализации настоящего изобретения относится к способу снижения вязкости слизи и мокроты у пациента с чрезмерно вязкой или липкой слизью или мокротой. Указанный способ включает этап приведения слизи или мокроты пациента в контакт композиции, содержащей белок или пептид, содержащие моноцистеиновый активный центр тиоредоксина в восстановленном состоянии, эффективно снижающей вязкость слизи и мокроты по сравнению с вязкостью до контакта. В одном аспекте этого варианта реализации пациент страдает заболеванием легких, при котором аномальная или чрезмерная вязкость или липкость слизи и мокроты является симптомом или причиной заболевания. В одном аспекте пациент страдает заболеванием легких, выбранным из муковисцидоза (MB), хронической обструктивной болезни легких, бронхоэктатической болезни и астмы. В предпочтительном варианте реализации пациент страдает MB. В еще одном аспекте этого варианта реализации пациент страдает заболеванием легких, при котором аномальная или чрезмерная вязкость или липкость слизи и мокроты ассоциирована с недостаточностью активности биологического восстановителя. В еще одном аспекте этого варианта реализации пациент страдает заболеванием пищеварительного тракта, ассоциированным с плотной или аномальной слизью, включая кокцидиоз, но не ограничиваясь им.
В одном аспекте этап приведения слизи или мокроты пациента в контакт с композицией выполняют путем введения композиции пациенту путем, выбранным из назального, интратрахеального, бронхиального, ингаляционного и перорального введения, а также прямого введения в легкие. В одном аспекте слизь и мокрота, с которой осуществляется контакт, находится в дыхательных путях, пищеварительном тракте (т.е. желудочно-кишечном тракте) или половых путях пациента.
В еще одном аспекте композицию вводят пациенту в фармацевтически приемлемом носителе.
В любом из вышеуказанных аспектов этап приведения слизи или мокроты пациента в контакт с композицией увеличивает процент свободных тиолов в образце слизи и мокроты пациента по сравнению с процентом до контакта с композицией.
В любом из вышеуказанных аспектов после контакта слизи или мокроты пациента с композицией у пациента наблюдается по меньшей мере приблизительно 2,5% увеличение объема форсированного выдоха (ОФВ) по сравнению с объемом до контакта с композицией.
В любом из вышеуказанных аспектов моноцистеиновый активный центр тиоредоксина содержит аминокислотную последовательность, выбранную из C-X-X-S (SEQ ID NO: 24), C-X-X-X (SEQ ID NO: 17), X-C-X-X-X-X (SEQ ID NO: 19), X-C-G-P-X-X (SEQ ID NO: 21), W-C-G-P-X-K (SEQ ID NO: 23), X-C-X-X-S-X (SEQ ID NO: 25), X-C-G-P-S-X (SEQ ID NO: 26) или W-C-G-P-S-K (SEQ ID NO: 27), где остатки С находится в восстановленном состоянии, и где остатки X представляют собой любой аминокислотный остаток, кроме цистеина. В предпочтительном аспекте моноцистеиновый активный центр тиоредоксина содержит аминокислотную последовательность C-X-X-S (SEQ ID NO: 24), как описано выше.
В любом из вышеуказанных аспектов белок, содержащий моноцистеиновый активный центр тиоредоксина, включает тиоредоксин, выбранный из группы, состоящей из прокариотического тиоредоксина, тиоредоксина грибов, тиоредоксина растений и тиоредоксина млекопитающих. В предпочтительном аспекте белок включает тиоредоксин человека.
В любом из вышеуказанных аспектов изобретения композиция дополнительно содержит восстанавливающий агент для восстановления моноцистеинового активного центра тиоредоксина в составе белка. В дополнительном аспекте композиция содержит тиоредоксинредуктазу и NADH или NADPH.
Еще один вариант реализации настоящего изобретения относится к композиции для применения с целью снижения вязкости слизи и мокроты, состоящей из белка или пептида, содержащих моноцистеиновый активный центр тиоредоксина в восстановленном состоянии и по меньшей мере один дополнительный агент для обработки чрезмерно вязкой или липкой слизи и мокроты. В одном аспекте данного варианта реализации моноцистеиновый активный центр тиоредоксина содержит аминокислотную последовательность, выбранную из C-X-X-S (SEQ ID NO: 24), C-X-X-X (SEQ ID NO: 17), X-C-X-X-X-X (SEQ ID NO: 19), X-C-G-P-X-X (SEQ ID NO: 21), W-C-G-P-X-K (SEQ ID NO: 23), X-C-X-X-S-X (SEQ ID NO: 25), X-C-G-P-S-X (SEQ ID NO: 26) или W-C-G-P-S-K (SEQ ID NO: 27), где остатки С находится в восстановленном состоянии, и где остатки X представляют собой любой аминокислотный остаток, кроме цистеина. В предпочтительном аспекте моноцистеиновый активный центр тиоредоксина содержит аминокислотную последовательность C-X-X-S (SEQ ID NO: 24), как описано выше. В любом из вышеуказанных аспектов данного варианта реализации белок, содержащий моноцистеиновый активный центр тиоредоксина, включает тиоредоксин, выбранный из группы, состоящей из прокариотического тиоредоксина, тиоредоксина грибов, тиоредоксина растений и тиоредоксина млекопитающих. В одном аспекте белок включает тиоредоксин человека.
Кроме того, в любом из вышеуказанных аспектов данного варианта реализации композиция содержит восстанавливающий агент. В дополнительном аспекте композиция, кроме того, содержит тиоредоксинредуктазу и NADH или NADPH.
Еще один вариант реализации настоящего изобретения относится к фармацевтической композиции, содержащей белок или пептид, содержащий моноцистеиновый активный центр тиоредоксина в восстановленном состоянии. В одном аспекте композицию составляют для введения в легкие в виде аэрозоля. В еще одном аспекте композицию составляют для перорального введения. В любом из вышеуказанных аспектов данного варианта реализации моноцистеиновый активный центр тиоредоксина содержит аминокислотную последовательность C-X-X-S (SEQ ID NO: 24), C-X-X-X (SEQ ID NO: 17), X-C-X-X-X-X (SEQ ID NO: 19), X-C-G-P-X-X (SEQ ID NO: 21), W-C-G-P-X-K (SEQ ID NO: 23), X-C-X-X-S-X (SEQ ID NO: 25), X-C-G-P-S-X (SEQ ID NO: 26) и W-C-G-P-S-K (SEQ ID NO: 27), где остаток С находится в восстановленном состоянии, а остатки X представляют собой любой аминокислотный остаток, кроме цистеина. В еще одном аспекте фармацевтическую композицию составляют для введения в легкие в виде аэрозоля с помощью ингалятора. В одном аспекте ингалятор является ингалятором с вибрирующей сеткой. В еще одном аспекте фармацевтическая композиция дополнительно содержит восстановитель. В любом из вышеуказанных аспектов настоящего изобретения фармацевтическая композиция, кроме того, содержит тиоредоксинредуктазу и NADH или NADPH.
Еще один вариант реализации настоящего изобретения относится к композиции, содержащей белок или пептид, содержащий моноцистеиновый активный центр тиоредоксина, причем моноцистеиновый активный центр тиоредоксина ковалентно связан с остатком цистеина белка слизи. В одном аспекте моноцистеиновый активный центр тиоредоксина содержит аминокислотную последовательность C-X-X-S (SEQ ID NO: 24), C-X-X-X (SEQ ID NO: 17), X-C-X-X-X-X (SEQ ID NO: 19), X-C-G-P-X-X (SEQ ID NO: 21), W-C-G-P-X-K (SEQ ID NO: 23), X-C-X-X-S-X (SEQ ID NO: 25), X-C-G-P-S-X (SEQ ID NO: 26) и W-C-G-P-S-K (SEQ ID NO: 27), где остаток С находится в восстановленном состоянии, а остатки X представляют собой любой аминокислотный остаток, кроме цистеина. В любом из вышеуказанных аспектов белок слизи представляет собой белок слизи дыхательных путей или белок слизи пищеварительного тракта. В еще одном аспекте белок слизи представляет собой муцин.
Еще один вариант реализации настоящего изобретения относится к способу снижения вязкости слизи и мокроты у пациента с чрезмерно вязкой или липкой слизью или мокротой. Этот способ включает этап приведения слизи или мокроты пациента в контакт с композицией, содержащей восстановитель дисульфидных связей и агент, блокирующий цистеин. В одном аспекте восстановитель дисульфидных связей и агент, блокирующий цистеин, являются одним и тем же веществом. В дополнительном аспекте это же вещество является белком или пептидом, содержащим моноцистеиновый активный центр тиоредоксина. В еще одном аспекте восстановитель дисульфидных связей и агент, блокирующий цистеин, являются различными веществами. В еще одном аспекте восстановитель дисульфидных связей может представлять собой дитиотреитол (ДТТ), этилендиаминтетрауксусную кислоту (ЭДТА), глутатион, дитиогликолевую кислоту, 2-меркаптоэтанол, N-ацетилцистеин или трис-(2-карбоксиэтил)фосфен. В еще одном дополнительном аспекте агент, блокирующий цистеин, может представлять собой иодацетамид, иодуксусную кислоту или другие алкилирующие агенты.
Еще один вариант реализации настоящего изобретения относится к способу лечения пациента с чрезмерно вязкой или липкой слизью. Этот способ включает этап введения пациенту композиции, содержащей непоглощаемое клетками соединение, содержащее моноцистеиновый активный центр тиоредоксина. В одном аспекте это соединение может являться белком или пептидом, содержащим моноцистеиновый активный центр тиоредоксина, гибридный белок, содержащий фрагмент тиоредоксина и фрагмент лиганда рецептора клеточной поверхности, или комбинацию белка или пептида, содержащего активный центр тиоредоксина, и соединения, блокирующего цистеин, соответствующий цистеину в положении 35 SEQ ID NO: 12.
Еще один вариант реализации настоящего изобретения относится к способу предотвращения системного воздействия лекарственного вещества на пациента. Этот способ включает этап введения лекарственного вещества пациенту посредством пути доставки, включая пульмонарный, пероральный или местный пути доставки, но не ограничиваясь ими. В еще одном аспекте лекарственное вещество после введения образует ковалентную связь с областью-мишенью. В еще одном аспекте лекарственное вещество является тиол-содержащим лекарственным веществом, и может представлять собой, например, белок или пептид, содержащий моноцистеиновый активный центр тиоредоксина в восстановленном состоянии. В еще одном аспекте лекарственное вещество является антибиотиком или противоинфекционным агентом, и объединено или присоединено посредством линкера к моноцистеиновому активному центру тиоредоксина в восстановленном состоянии. В еще одном аспекте лекарственное вещество является противовоспалительным агентом, и объединено или присоединено посредством линкера к моноцистеиновому активному центру тиоредоксина в восстановленном состоянии. В еще одном аспекте лекарственное вещество является агентом, гидролизующим нуклеиновые кислоты, и объединено или присоединено посредством линкера к моноцистеиновому активному центру тиоредоксина в восстановленном состоянии. В еще одном аспекте лекарственное вещество является химиотерапевтическим агентом, и объединено или присоединено посредством линкера к моноцистеиновому активному центру тиоредоксина в восстановленном состоянии.
Еще один вариант реализации настоящего изобретения относится к фармацевтической композиции, содержащей белок или пептид, содержащий моноцистеиновый активный центр тиоредоксина в восстановленном состоянии, и дополнительно содержащей по меньшей мере один сахарид или производное сахарида, способное стабилизировать восстановленную редокс-активную тиоловую группу. В одном аспекте сахарид или производное сахарида может являться сахарозой, сукралозой, лактозой, трегалозой, мальтозой, галактозой, рафинозой, маннозой или маннитом. В одном аспекте моноцистеиновый активный центр тиоредоксина содержит аминокислотную последовательность C-X-X-S (SEQ ID NO: 24), C-X-X-X (SEQ ID NO: 17), X-C-X-X-X-X (SEQ ID NO: 19), X-C-G-P-X-X (SEQ ID NO: 21), W-C-G-P-X-K (SEQ ID NO: 23), X-C-X-X-S-X (SEQ ID NO: 25), X-C-G-P-S-X (SEQ ID NO: 26) и W-C-G-P-S-K (SEQ ID NO: 27), где остаток С находится в восстановленном состоянии, а остатки X представляют собой любой аминокислотный остаток, кроме цистеина.
Еще один вариант реализации настоящего изобретения относится к кормовой композиции, содержащей белок или пептид, содержащий моноцистеиновый активный центр тиоредоксина в восстановленном состоянии. В одном аспекте моноцистеиновый активный центр тиоредоксина содержит аминокислотную последовательность C-X-X-S (SEQ ID NO: 24), C-X-X-X (SEQ ID NO: 17), X-C-X-X-X-X (SEQ ID NO: 19), X-C-G-P-X-X (SEQ ID NO: 21), W-C-G-P-X-K (SEQ ID NO: 23), X-C-X-X-S-X (SEQ ID NO: 25), X-C-G-P-S-X (SEQ ID NO: 26) и W-C-G-P-S-K (SEQ ID NO: 27), где остаток С находится в восстановленном состоянии, а остатки X представляют собой любой аминокислотный остаток, кроме цистеина.
В одном аспекте любого из вариантов реализации настоящего изобретения пациент является позвоночным, включая млекопитающих и птиц, но не ограничиваясь ими. В еще одном аспекте пациент является человеком. В еще одном аспекте пациент является курицей или индейкой.
Еще один вариант реализации настоящего изобретения относится к применению композиции, содержащей белок или пептид, содержащий моноцистеиновый активный центр тиоредоксина в восстановленном состоянии, для уменьшения вязкости слизи или мокроты у пациента с избыточно вязкой или липкой слизью или мокротой, причем контакт слизи или мокроты пациента с композицией снижает вязкость слизи или мокроты по сравнению с вязкостью до контакта. В одном аспекте моноцистеиновый активный центр тиоредоксина содержит аминокислотную последовательность C-X-X-S (SEQ ID NO: 24), C-X-X-X (SEQ ID NO: 17), X-C-X-X-X-X (SEQ ID NO: 19), X-C-G-P-X-X (SEQ ID NO: 21), W-C-G-P-X-K (SEQ ID NO: 23), X-C-X-X-S-X (SEQ ID NO: 25), X-C-G-P-S-X (SEQ ID NO: 26) и W-C-G-P-S-K (SEQ ID NO: 27), где остаток С находится в восстановленном состоянии, а остатки X представляют собой любой аминокислотный остаток, кроме цистеина.
Краткое описание чертежей
На фигурах 1a-1b показана ферментативная активность белка или пептида, содержащего моноцистеиновый активный центр тиоредоксина (называемый r(Cys)hTrx) по сравнению с белком или пептидом, содержащим активный центр тиоредоксина дикого типа (называемый WTrhTrx). На фигуре 1а показано неспецифическое восстановление 5,5'-дитиобис-(2-нитробензойной кислоты) (DTNB или реагента Эллмана), отражающее потерю 1 восстанавливаемого цистеина в моноцистеиновом активном центре тиоредоксина по сравнению с активным центром дикого типа. На фигуре 1b показано, что белок или пептид, содержащий моноцистеиновый активный центр тиоредоксина (r(Cys)hTrx), обладал аналогичной или повышенной эффективностью по сравнению с белком или пептидом, содержащим активный центр тиоредоксина дикого типа (WTrhTrx) в анализе уплотнения мокроты человека, в концентрации 12,5 мкМ, 25 мкМ и 50 мкМ.
На фигуре 2 показано действие rhTrx дикого типа, r(Cys)hTrx и различных контрольных материалов, включая дитиотреитол (DTT) в двух концентрациях (0,58 мМ и 1,5 мМ), N-ацетилцистеин (NAC) и рекомбинантную ДНКазу человека (rhDNase) в эквимолярных концентрациях, на нормализацию образцов мокроты пациентов (n=6 на обработку) в анализе уплотнения мокроты.
На фигуре 3 приведен механизм образования дисульфидных связей с белком или пептидом, содержащим моноцистеиновый активный центр тиоредоксина в положении 35 SEQ ID NO: 12. Механизм нативного восстановления дисульфидной связи Trx включает двухэтапную реакцию. Как показано на фигуре 3, этапы I и II представляют собой образование переходного смешанного дисульфида между N-концевым Cys (расположенным в положении 32 в аминокислотной последовательности TRX-1 человека) активного центра Trx и одним из Cys дисульфида белка-мишени, с последующим этапом III. Этап III представляет собой нуклеофильную атаку на межмолекулярную смешанную дисульфидную связь С-концевого Cys активного центра Trx, расположенного в положении 35 аминокислотной последовательности TRX-1 человека. Это второе восстановление расщепляет смешанную дисульфидную связь и высвобождает окисленный Trx и полностью восстановленную мишень. За счет мутации С-концевого остатка Cys активного центра в положении 35 Trx на аминокислоту, не являющуюся цистеином, например, на остаток серина (моноцистеиновый вариант), или иной модификации последовательности белка с целью мешающего влияния на нуклеофильную атаку С-концевого Cys активного центра в положении 35, тиоредоксин сохраняет способность функционировать в качестве восстанавливающего агента, но в отличие от фермента дикого типа такие моноцистеиновые варианты активного центра остаются ковалентно присоединенными к восстановленному белку-мишени посредством нерасщепленной межмолекулярной смешанной дисульфидной связи.
Подробное описание изобретения
Настоящее изобретение в целом относится к применению белка или пептида, содержащих моноцистеиновый активный центр тиоредоксина в восстановленном состоянии для индукции, повышения и/или повышения разжижения слизи или мокроты. Конкретнее, авторы настоящего изобретения обнаружили, что белки или пептиды с моноцистеиновым активным центром тиоредоксина снижают вязкость и/или липкость мокроты или слизи и, таким образом, являются эффективными агентами для повышения или увеличения разжижения мокроты или слизи. Соответственно, белки или пептиды, содержащие моноцистеиновый активный центр тиоредоксина в восстановленном состоянии, или молекулы нуклеиновой кислоты, кодирующие такие белки, можно применять отдельно или в композиции для лечения различных состояний или заболеваний, ассоциированных с нежелательной слизью или липкой и вязкой мокротой. Например, такие респираторные заболевания, как муковисцидоз, хроническая обструктивная болезнь легких, бронхоэктатическая болезнь и астма, особенно хорошо поддаются лечению с использованием продукта и способа согласно изобретению. Кроме того, заболевания пищеварительного тракта, ассоциированные с густой или липкой слизью, например, кокцидиоз, также особенно хорошо поддаются лечению с использованием продукта и способа согласно изобретению. Таким образом, настоящее изобретение относится к применению белков, содержащих моноцистеиновый активный центр тиоредоксина в восстановленном состоянии, для снижения вязкости слизи или мокроты, в частности, аномально или избыточно вязкой и/или липкой слизи или мокроты. Белки вводят пациенту, который страдает от или подвергается воздействию такой аномальной или избыточной слизи или мокроты посредством способа и в количестве, эффективно снижающих вязкость слизи или мокроты и, предпочтительно, обеспечивающих терапевтически благоприятное действие для пациента.
Тиоредоксин и белки, содержащие активный центр тиоредоксина дикого типа (или нативный) (также называемый в настоящем документе "rhTxr") или моноцистеиновый активный центр тиоредоксина (также называемый в настоящем документе "r(Cys)hTrx"), имеют преимущества перед другими восстановителями для применения при лечении таких состояний, как муковисцидоз. Например, в отличие от других восстановителей, например, N-ацетилцистеина (NAC), Nacystelyn (NAL), дитиотреитола (ДТТ) или восстановленного глутатиона (GSH), тиоредоксин менее подвержен инактивации посредством ферментативных или аутоокислительных механизмов, в том числе реакций, образующих супероксида, пероксид водорода, гидроксил-радикал и другие метаболиты токсичных форм кислорода. Кроме того, нативный тиоредоксин или тиоредоксин дикого типа является природным соединением, которое в норме внеклеточно секретируется на поверхность дыхательных путей, и поэтому введение тиоредоксина в дыхательные пути не должно вызывать раздражения и индуцировать иммунный ответ. Кроме того, тиоредоксин не подвергается гликозилированию, и поэтому его проще производить; кроме того, введение данного белка в природной или рекомбинантной форме не должно вызывать врожденного иммунного ответа. Возможно, даже более важным является то, что восстановленный тиоредоксин, в отличие от других восстановителей, более быстро и эффективно восстанавливает нормальный уровень вязкости обработанной слизи или мокроты, и эта нормализация продолжается в течение более длительного срока. NAC, NAL, ДТТ и GSH, например, "истощаются" или окисляются с течением времени и на этом этапе нормализованная мокрота или слизь может вернуться к состоянию аномальной вязкости. В противоположность этому, снижение вязкости, полученное за счет тиоредоксина, по-видимому, продолжается дольше, по всей вероятности, из-за его циклического повторного восстановления с помощью восстанавливающей системы. Кроме того, оставаясь ковалентно связанным с остатками Cys муцина, r(Cys)hTrx обеспечивает еще более мощное и продолжительное снижение вязкости по сравнению с нативным rhTrx. Наконец, тиоредоксин является одновременно более мощным и более специфичным по отношению к восстановлению дисульфидных связей, чем другие восстановители, и, следовательно, для достижения положительного эффекта его можно применять в значительно более низких дозах, чем другие агенты.
В дополнение к вышеописанным преимуществам, тиоредоксин имеет и другие преимущества, которые увеличивают его полезность в условиях заболевания. Например, известно, что тиоредоксин индуцирует MnSOD (например, см. патент США №5,985,261, принадлежащий White et al., полностью включенный в настоящий документ посредством ссылки), что, согласно прогнозам, должно снижать токсичность некоторых бактериальных токсинов (включая эндотоксин клеточной стенки грамотрицательных бактерий, пиоцианин Pseudomonas aeruginosas другие, но не ограничиваясь ими) в мокроте при заболевании (например, при муковисцидозе). Кроме того, тиоредоксин обладает внеклеточными противовоспалительными свойствами (Lee, R.L., et al., Am J Physiol Lung Cell Mol Physiol, 289(5):L875-82, 2005), которые могут улучшить общее лечение респираторного заболевания.
Тиоредоксин (Trx) представляет собой белок дисульфидредуктазы, который катализирует многочисленные тиол-зависимые восстановительные процессы в клетках. Нативный тиоредоксин содержит два редокс-активных остатка цистеина, которые являются высококонсервативными у разных видов. В окисленной формы эти остатки цистеина образуют дисульфидный мостик, который выступает из трехмерной структуры белка (Holmgren, Annu Rev Biochem 54:237-271, 1985). Восстановление этого активного центра фермента NADPH-зависимой тиоредоксинредуктазой (TR) позволяет Trx функционировать в качестве переносчика электронов с возможностью обмена дитиол/дисульфид (Oblong et al., Biochemistry 32:7271-7277, 1993). Белковые дисульфиды являются предпочтительным субстратом для Trx-опосредованного восстановления. Модификация остатка цистеина в С-концевой области активного центра Trx позволяет получить моноцистеиновый активный центр, который, как обсуждается ниже, имеет существенные преимущества по сравнению с Trx с нативным активным центром или активным центром дикого типа. Устойчивая и вязкая природа секрета дыхательных путей при муковисцидозе приводит к обструкции дыхательных путей, инфекции условно-патогенными микроорганизмами и ухудшению функции легких. С учетом того, что муцин дыхательных путей содержит множественные цистеинсодержащие домены, которые, как полагают, играют важную роль при полимеризации (Bell et al., Biochem J 357:203-209, 2001; Asker et al., Biochem J 333:381-387, 1998), а также повышении степени переплетения муцина посредством многочисленных внутримолекулярных дисульфидных связей, авторы настоящего изобретения стремились определить способность Trx, содержащего моноцистеиновый активный центр, являться эффективным регулятором вязкости слизи путем восстановления дисульфидов муцина.
У тиоредоксина, содержащего моноцистеиновый активный центр, есть несколько преимуществ и выгодных качеств по сравнению с тиоредоксином, содержащим нативный активный центр или активный центр дикого типа. Моноцистеиновая модификации предназначена для минимизации потенциальных побочных эффектов тиоредоксина, связанных с внутриклеточными сигнальными путями или системным воздействием, например, эффектов, описанных Rancourt et al. (Free Radical Biol & Med 42:1441-43, 2007). Эта модификация предотвращает нуклеофильную атаку на смешанный дисульфид, образованный между тиоредоксином и дисульфидом белка-мишени, которая катализируется N-концевым остатком цистеина активного центра тиоредоксина (например, расположенным в положении 32 в тиоредоксине человека, SEQ ID NO: 14). Как ни странно, автор настоящего изобретения определил, что тиоредоксин, содержащий моноцистеиновый активный центр, обладает большей эффективностью, чем тиоредоксин дикого типа при снижении (тенденции к разжижению) и нормализации вязкости слизи человека при заболевании. Несмотря на то, что с логической точки зрения можно было бы ожидать меньшего восстановительного потенциала тиоредоксина, содержащего моноцистеиновый активный центр, из-за сильного возмущения, вызванного потерей остатка цистеина активного центра и, следовательно, после первоначальной каталитической реакции при N-концевом остатке цистеина активного центра тиоредоксин должен быть ковалентно с белками слизи (например, муцинами MUC5AC и MUC5B с высоким содержанием дисульфидных связей) и не должен восстанавливаться и выполнить повторный катализ, автор настоящего изобретения обнаружил, что тиоредоксин, содержащий моноцистеиновый активный центр, не только не демонстрирует нарушений активности по сравнению с тиоредоксином дикого типа, но и обладает большей количественный способностью снижать вязкость слизи человека при MB в реологическом анализе. На основе этого неожиданного результата автор изобретения пришел к выводу, что повышенная эффективность тиоредоксина, содержащего моноцистеиновый активный центр, должна быть следствием ковалентной связи тиоредоксина и слизи, блокирующей повторное образование дисульфидов цистеина в муцине и обеспечивающей, таким образом, чрезвычайно стойкие и длительные изменения в структуре и размере пор геля олигомеров муцина. В то же время, ковалентная связь тиоредоксина, содержащего моноцистеиновый активный центр, с его мишенью - муцином блокирует и, следовательно, предотвращает поглощение и усвоение ингаляционного тиоредоксина клетками, что обеспечивает двойное преимущество за счет предотвращения неспецифических эффектов, вызванных нежелательной активностью тиоредоксина внутри клеток, и в то же время облегчает выведение связанного со слизью отработанного лекарственного вещества из организма. Поскольку единственным терапевтическим применением, предусмотренным ранее в данной области техники для Cys-модифицированного моноцистеинового тиоредоксина, являлось облегчение поглощения невосстановленных форм через липидные рафты эндотелиальных клеток после инъекции в системный кровоток ((Hara et al., Antiox Redox Sig 9:1427-37, 2007; Kondo et al., Antiox Redox Sig 9:1439-48, 2007; заявки на патент США 20080119398, 20090075871 и 20100184215), что сильно отличается от настоящего изобретения, направленного на предотвращение системного воздействия и внутриклеточного поглощения, внеклеточный механизм повышения эффективности и безопасности моноцистеинового активного центра тиоредоксина в восстановленной форме является непредвиденным и в значительной степени новым.
Обструкция дыхательных путей слизью может привести к высокой заболеваемости и смертности у пациентов с MB. Авторы настоящего изобретения показали, что вязкоупругие свойства, облегчающие стойкость этих выделений в дыхательных путях, заметно уменьшились за счет Trx, содержащего моноцстеиновый активный центр. Этот вывод подтверждается двумя наборами экспериментальных доказательств. Во-первых, результаты анализа уплотнения показывают, что при инкубировании с моноцистеиновым Trx из гелевого матрикса мокроты при MB высвобождается большое количество жидкости. Одновременно с этим высвобождением происходит уменьшение объема твердого вещества, что указывает на солюбилизацию гелеобразующих компонентов мокроты. Эту нормализацию вязкости мокроты при MB часто можно макроскопически наблюдать в образцах мокроты во время инкубационного периода MB, и поэтому она не является артефактом центрифугирования. Полагают, что высвобождение жидкости за счет моноцистеиновго Trx должно иметь важные терапевтические последствия, поскольку восстановление объема воды на поверхности дыхательных путей может восстанавливать способность эпителия к мукоцилиарному транспорту при MB (Jiang et al., Science 262:424-427, 1993) и облегчать избыточную вязкость, на основе модели геля на щетке, разработанной Button et al. (Science, 2012), обеспечивая гидратацию подстилающего околоресничного слоя и восстанавливая мукоцилиарный транспорт, потеря которого является основной причиной патологии при MB. Во-вторых, магнитные микрореометрические измерения позволили получить прямые доказательства того, что вязкоупругость мокроты снижается в результате восстановления компонентов мокроты моноцистеиновым Trx.
Мокрота при MB является неньютоновской жидкостью, обладающей характеристиками как жидкости, так и твердого тела. Полимеры, присутствующие в растворах при низкой концентрации, способны свободно вращаться. При концентрировании или перекрестном связывании полимеров до такой степени, которая препятствует их вращению, раствор достигает переходного этапа, называемого порогом перколяции (Forgacs, J Cell Sci 108:2131-2143, 1995). В области порога перколяции раствор начинает приобретать характеристики твердого тела, и модули упругости продолжают расти по мере роста перекрестного взаимодействия между молекулами полимера, пока каждый филамент в образце не войдет в состав матрикса. Биохимические анализы показали, что муцины MUC5AC и MUC5B, выделяемые клетками, выстилающими дыхательные пути, являются основными гелеобразующими полимерными компонентами слизи в дыхательных путях (Hovenberg et al., Glycoconj J 13:839-847, 1996; Thornton et al., Biochem 7 316:967-975, 1996; Thornton et al., J Biol Chem 272:9561-9566, 1997). Цистеиновые домены, присутствующие на этих муцинах, участвуют в образовании полимеров и, возможно, взаимодействии с соседними цепями муцина путем образования внутримолекулярных дисульфидных связей (Bell et al., Biochem 7357:203-209, 2001; Asker et al., Biochem 7 333:381-387, 1998), что, вероятно вносит вклад в переплетение сетчатого каркаса геля. Поскольку дисульфидные связи на молекулах белка являются предпочтительным субстратов для ферментативной активности Trx, полимеры муцина являются мишенями восстановления при разжижении мокроты за счет Trx. Это подтверждается окрашиванием PAS, которое выявляет изменения растворимости высокомолекулярных гликоформ в мокроте, обработанной Trx. На обнаружение повышенных концентраций гликопротеинов в жидкой фазе мокроты, обработанной Trx, также указывает более интенсивное желтое окрашивание и повышенная мутность по сравнению с жидкой фазой, полученной из образцов, обработанных разбавителем. Расширенная электрофоретическая подвижность PAS-обнаруживаемых гликопротеинов в мокроте, обработанной Trx, также свидетельствует о том, что размер этих макромолекул может уменьшаться во время ферментативного восстановления. Результаты этого электрофоретического анализа согласуются с измерениями при анализе уплотнения, демонстрируя, что высвобождение гликопротеинов в жидкую фазу совпадает с уменьшением массы гелевого матрикса при воздействии Trx, а также наблюдениями за увеличением содержания меченых свободных тиолов в мокроте после обработки Trx (Rancourt, R. et al., Free Radic Biol Med, 42(9):1441-1453, 2007).
После проявления хронических эффектов воспаления и инфекции у пациента с MB лизис нейтрофилов в дыхательных путях пораженных легких при MB приводит к накоплению внеклеточной ДНК в секрете дыхательных путей (Lethem et al., Eur Respir J 3:19-23, 1990). За счет нековалентных взаимодействий ДНК переплетается с гликопротеинами муцина, увеличивая вязкоупругость геля слизи (Sachdev et al., Chest 81:41S-43S, 1982). ДНК в мокроте после обработки Trx становится все более растворимой. Логическое объяснение этого состоит в том, что активность Trx вызывает структурные изменения в матриксе геля, которые являются достаточными для ослабления переплетающих взаимодействий между ДНК и пораженными макромолекулами. Сомнительно, что эта повышенная растворимость ДНК вносит вклад в изменения вязкоупругости, наблюдаемые во время воздействия Trx на мокроту при MB. Тем не менее с клинической точки зрения, высвобождение или удаление ДНК из нерастворимой гелевой фазы мокроты может повысить ее восприимчивость к активности ДНКазы во время такого лечения при MB. Кроме того, действие моноцистеинового r(Cys)hTrx, снижающее вязкость слизи и предотвращающее быстрое повторное образование дисульфидных связей остатков цистеина в муцине должно создать слой более проницаемой, доступной слизи, а также уменьшить объем пробок из накопленной слизи. Ожидается, что это действие должно облегчить доступ других терапевтических средств в глубокие отделы легких и к поверхности эпителия легких. Таким образом, механистический способ согласно изобретению обладает сильным потенциалом для взаимодействия с существующими средствами симптоматической терапии MB и других обструктивных заболеваний легких, например, доставкой ингаляционных антибиотиков, мукоактивных веществ или муколитических агентов, гидролизующих ДНК.
Trx, содержащий моноцистеиновый активный центр, обладает повышенной активностью и окислительной стабильностью по сравнению с восстановленным глутатионом, действует во внеклеточной слизи дыхательных путей и не проникает в клетки легких. Trx, содержащий моноцистеиновый активный центр, снижает вязкость, увеличивает жидкую фракцию и уменьшает вязкоупругость мокроты, например, мокроты при MB. Ожидается, что разработка систем для восстановления слизи, стимулирующих высвобождение жидкости и снижающих вязкость секрета дыхательных путей, должна обладать терапевтическим потенциалом при таких заболеваниях, как MB, а также при лечении избыточной или аномальной вязкости и/или липкости слизи, которые могут быть ассоциированы с другими респираторными заболеваниями (например, острым или хроническим бронхитом, бронхоэктатической болезнью; ХОБЛ/эмфиземой; астмой; острым трахеитом; острым или хроническим синуситом; ателектазом, обусловленным острой или хронической закупоркой дыхательных путей слизью; бронхиолитом) или с различными расстройствами пищеварительной системы (т.е., желудочно-кишечного тракта), например, кокцидиозом, или расстройствами половой системы, связанными с или усугубляемыми избыточной или аномальной вязкостью и/или липкостью слизи (например, острой, подострой или хронической кишечной непроходимостью, вызванной сгущением слизи; бесплодием вследствие обструкции жизненно важных репродуктивных структур). Поскольку Trx, содержащий моноцистеиновый активный центр в восстановленном состоянии, ковалентно связывается с муцином после реакции с дисульфидными связями муцина, такой механизм действия должен способствовать выведению отработанного (окисленного) лекарственного вещества вместе со слизью, может предотвращать или ослаблять обновление клеток и тиоредоксин-опосредованные окислительно-восстановительные сигнальные пути и предотвращать или ослаблять презентацию антигенов иммунным клеткам.
Соответственно, один вариант реализации настоящего изобретения относится к способу нормализации и снижения вязкости слизи и мокроты у пациента с чрезмерно вязкой или липкой слизью или мокротой. Указанный способ включает этап приведения слизи или мокроты пациента в контакт с композицией, содержащей белок или пептид, содержащие моноцистеиновый активный центр тиоредоксина в восстановленном состоянии. Белок эффективно снижает вязкость слизи и мокроты по сравнению с вязкостью до контакта.
Согласно настоящему изобретению, термин "слизь" в целом относится к обычно прозрачной вязкой жидкости, выделяемой слизистыми оболочками различных тканей организма, в том числе дыхательных путей, желудочно-кишечного и репродуктивного тракта. Слизь увлажняет, смазывает и защищает ткани, выделяющие ее. Она содержит макромолекулы муцина (в том числе белки, нуклеиновые кислоты и углеводы слизи), которые являются гелеобразующими компонентами слизи. Белки слизи включают белки слизи дыхательных путей и пищеварительного тракта, но не ограничиваются ими. Вязкоупругие свойства нормальной слизи зависят от концентрации, молекулярной массы и степени переплетения полимеров муцина. Термин «мокрота» в целом относится к смеси слюны и выделений дыхательных путей, в том числе слизи. Мокрота обычно представляет собой откашливаемую смесь слюны и слизи (и других выделений тканей дыхательных путей). Таким образом, слизь является основным компонентом мокроты, и поэтому наличие чрезмерно вязкой слизи приводит к образованию чрезмерно вязкой мокроты. Настоящее изобретение относится к снижению вязкости слизи или мокроты. Термин "разжижение" относится к усилению жидкостных свойств. Таким образом, повышение степени разжижения слизи и мокроты относится к увеличению жидкой фазы или жидкого состояния слизи и мокроты по сравнению с более твердой и вязкой фазой. В случае аномально вязкой или избыточной слизи, ассоциированной с заболеванием, цель заключается в восстановлении нормального уровня вязкости слизи. Следовательно, разжижение можно считать снижением вязкости слизи.
Считается, что нормальное функционирование слизи достигается за счет соответствующего соотношения биологических восстановителей и окисляемого цистеина. Следовательно, недостаточность активности биологических восстановителей возникает вследствие избытка окисляемого цистеина или отсутствия биологических восстановителей.
Общие функции слизи и мокроты в организме требуют наличия вязкоупругих свойств слизи (и, следовательно, слизистого компонента мокроты). У лица с нормальной слизью и мокротой (т.е., здорового человека, или, конкретнее, человека, не страдающего симптомами или состоянием, вызванным или усугубляемым вязкостью или липкостью слизи и мокроты), вязкоупругость зависит от концентрации, молекулярной массы и переплетения полимеров муцина (Verdugo et al., Biorheology 20:223-230, 1983). Слизь (и, следовательно, мокрота) может стать еще более густой и вязкой, в особенности при MB, при котором слизь взаимодействует с ДНК (Potter et al., Am J Dis Child 100:493-495, 1960; Lethem et al., Am Rev Respir Dis 100:493-495, 1990; Lethem et al., Eur Respir J 3:19-23, 1990) и полимерами f-актина (Sheils et al., Am J Path 148:919-927, 1996; Tomkiewicz et al., DNA and actin filament ultrastructure in cystic fibrosis sputum. In: Cilia, mucus, and mucociliary interactions, edited by Baum GL, Priel Z, Roth Y, Liron N, and Ostfeld EJ. New York, NY: Marcel Dekker, 1998), высвобождаемыми из гибнущих воспалительных клеток. Невозможность выведения аномальной густой слизи путем кашля или мукоцилиарного клиренса способствует колонизации легких условно-патогенными микроорганизмами. Таким образом, аномально или чрезмерно вязкая и/или липкая слизь характеризуется как слизь, которая является измеримо или обнаружимо более вязкой или липкой, чем слизь нормального или здорового пациента (желательно того же возраста и пола), и/или слизь, которая, в силу ее вязкости и/или липкости, вызывает или способствует по меньшей мере одному из симптомов, вызывающих дискомфорт или боль у пациента, или вызывает или обостряет состояние или заболевание. Другими словами, аномально или чрезмерно вязкая и/или липкая мокрота является отклонением от нормального состояния слизи или мокроты, причем для лечения пациента желательно обеспечить некоторое облегчение данного состояния или другой терапевтически благоприятный эффект.
Способ и композицию согласно настоящему изобретению можно применять для лечения любого пациента, у которого желательно снизить вязкость мокроты или слизи. В частности, пациенты с определенными заболеваниями или состояниями легких, синусов, носовой полости, пищеварительной системы или желудочно-кишечного тракта, или половой системы могут получить благоприятный эффект от лечения с помощью способа согласно настоящему изобретению. Настоящее изобретение наиболее полезно для смягчения или ослабления по меньшей мере одного симптома состояния или заболевания, вызываемого или усугубляемого аномальной или избыточной вязкостью и/или липкостью слизи и мокроты, которое, разумеется, может включать заболевание, связанное с легкими, например, муковисцидоз, а также заболевания пищеварительной системы, например, кокцидиоз. Другие заболевания могут, по меньшей мере в течение некоторого времени, ассоциироваться с аномальной или избыточной вязкостью и/или липкостью слизи или мокроты, и при возникновении такого симптома способ согласно настоящему изобретению можно применять для снижения вязкости слизи или мокроты и обеспечивает по меньшей мере некоторое облегчение или терапевтически благоприятное действие на пациента. Примеры таких заболеваний включают: муковисцидоз; острый или хронический бронхит; бронхоэктатическую болезнь (бронхоэктатическую болезнь, связанную и не связанную с MB); ХОБЛ/эмфизему; острый трахеит (бактериальный, вирусный, микоплазменный или вызванный другими организмами); острый или хронический синусит; ателектаз (коллапс легкого или его доли) вследствие острой или хронической закупорки дыхательных путей слизью (иногда наблюдается при различных заболеваниях, например, астме); бронхиолит (вирусный или другой); острую, подострую или хроническую кишечную непроходимость вследствие сгущения слизи, включая мекониевую непроходимость кишечника или эквивалент мекониевой непроходимости кишечника при MB или аналогичных расстройствах, но не ограничиваясь ими; другие заболевания пищеварительной системы и бесплодие вследствие обструкции (но не ограничиваясь) шейки матки, семенных протоков или других жизненно важных репродуктивных структур, но не ограничиваются ими. Кроме того, поскольку улучшение мукоцилиарного клиренса связано с клиренсом бактерий и других патогенных организмов из легких, композицию и способ согласно настоящему изобретению можно применять для ослабления симптомов, ассоциированных с чрезмерной вязкостью и/или липкостью слизи и мокроты у пациентов с различными респираторными инфекциями, в том числе как вирусными, так и бактериальными инфекциями.
Таким образом, терапевтический эффект не обязательно заключается в лечении конкретного заболевания или состояния, но предпочтительно охватывает результат, который обычно включает облегчение заболевания или состояния, устранение заболевания или состояния, ослабление или устранение симптомов, связанных с заболеванием или состоянием, профилактику или облегчение вторичного заболевания или состояния, вызванного возникновением основного заболевания или состояния (например, инфекционного заболевания, вызванного условно патогенными микроорганизмами, которым благоприятствует избыточно вязкая слизь в дыхательных путях), и/или профилактику основного заболевания или состояния или симптома, связанного с заболеванием или состоянием. В настоящем документе фраза «защищен от заболевания» относится к ослаблению симптомов заболевания; паллиативной терапии (облегчению или смягчению симптомов заболевания без его излечивания); снижению частоты возникновения этого заболевания и/или снижению тяжести заболевания или облегчению по меньшей мере одного симптома, признака или причины заболевания или состояния. Профилактика относится к способности композиции согласно настоящему изобретению при введении в организм пациента предотвращать возникновение заболевания. Лечение (или воздействие на заболевание) относится к способности композиции согласно настоящему изобретению при введении в организм пациента лечить заболевание. Защита пациента от заболевания включает лечение пациента, страдающего заболеванием (терапевтическое лечение). Профилактика заболевания/состояния включает предотвращение возникновения заболевания (профилактическое лечение). В частности, защиту пациента от заболевания (или профилактику заболевания) осуществляют путем повышения (нормализации) разжижения аномально вязкой слизи или мокроты у пациента путем приведения слизи или мокроты в контакт с белком или пептидом, содержащим моноцистеиновый активный центр тиоредоксина в восстановленном состоянии, получая благоприятный эффект. Благоприятный эффект может оценить специалист в данной области техники и/или обученный врач, лечащий пациента. Термин «заболевание» относится к любому отклонению от нормального состояния здоровья пациента и включает состояние, при котором присутствуют симптомы заболевания, а также состояния, при которых произошло указанное отклонение (например, инфекцию, мутацию гена, генетический дефект и т.д.), но симптомы еще не проявились.
Контакт слизи и/или мокроты пациента с белком или пептидом, содержащим моноцистеиновый активный центр тиоредоксина в восстановленном состоянии (или композицией, содержащей такой белок), должен привести к снижению вязкости/повышенному разжижению слизи и мокроты, по сравнению с состоянием до контакта с композицией. Согласно настоящему изобретению, повышение разжижения мокроты или слизи может представлять собой измеримое или обнаружимое увеличение объема разжиженной слизи и мокроты по сравнению с предыдущим уровнем разжижения, предпочтительно, статистически значимое увеличение (то есть различия измеренного уровня разжижения в образце пациента и исходном контрольном образце являются статистически значимыми с уровнем значимости по меньшей мере р<0,05). Как правило, «исходный контрольный образец» является образцом пациента до введения лекарственного средства, поскольку нормальные, здоровые лица обычно не продуцируют количество мокроты, достаточное для использования в качестве контроля, хотя не исключено использование мокроты нормального, здорового лица в качестве исходного контрольного образца. Кроме того, снижение вязкости приводит к улучшению функции легких. Это улучшение можно определить различными средствами, включая зарегистрированный исход лечения пациента, среднее время среднее время до обострения, приводящего к госпитализации, и/или увеличение объема форсированного выдоха (ОФВ). В одном аспекте настоящего изобретения увеличение ОФВ описывают как увеличение на по меньшей мере приблизительно 2,5%, приблизительно 3,0%, приблизительно 3,5%, приблизительно 4,0%, приблизительно 4,5%, приблизительно 5,0%, приблизительно 5,5%, приблизительно 6,0%, приблизительно 6,5%, приблизительно 7,0%, приблизительно 7,5%, приблизительно 8,0%, приблизительно 8,5%, приблизительно 9,0% и 9,5% и приблизительно 10% по сравнению с образцом пациента до контакта с композицией или белком согласно настоящему изобретению. Предпочтительно, контакт белка или композиции согласно настоящему изобретению со слизью или мокротой в образце пациента приводит к увеличению приблизительно на 2,5% по сравнению с образцом пациента до контакта с композицией или белком согласно настоящему изобретению. Разжижение слизи и мокроты и/или снижение вязкости можно измерить с помощью любого подходящего способа, известного в данной области техники, включая анализ уплотнения, описанный в разделе "Примеры", но не ограничиваясь им. При таком анализе измеряют количество слизи и мокроты в твердой фазе (геле) по сравнению с водной фазой (жидкостью). В других аспектах изобретения относительную вязкость или липкость слизи или мокроты можно измерить с использованием других параметров или показателей, включая вязкоупругость (измерямую, например, с помощью магнитной микрореометрии), содержание гликопротеинов или содержание ДНК, но не ограничиваясь ими. В еще одном аспекте изобретения можно оценить изменения дисульфидных связей белка слизи с помощью таких реагентов, как NEM (N-этилмалеимид), которые преимущественно реагируют с несвязанными (свободными) тиоловыми группами остатков Cys, образующимися при разрушении дисульфидных связей (Rancourt, R. et al., Free Radio Biol Med, 42(9): 1441-1453, 2007). В одном аспекте изобретения уровень разжижения описывают как количество слизи или мокроты в образце, находящееся в водной фазе (жидкости), в процентах от общего объема образца слизи или мокроты. Например, у пациентов с муковисцидозом уровень разжижения слизи или мокроты может составлять менее 10% или даже менее 5% от общего объема. Предпочтительно, контакт белка или композиции согласно изобретению со слизью или мокротой приводит к изменению разжижения слизи и мокроты по меньшей мере таким образом, что объем жидкой фазы составляет по меньшей мере приблизительно 15% от общего объема, более предпочтительно, объем жидкой фазы составляет по меньшей мере приблизительно 20% от общего объема, и более предпочтительно, объем жидкой фазы составляет по меньшей мере приблизительно 25% от общего объема, и более предпочтительно, объем жидкой фазы составляет по меньшей мере приблизительно 30% от общего объема, более предпочтительно, объем жидкой фазы составляет по меньшей мере приблизительно 35% от общего объема, и более предпочтительно, объем жидкой фазы составляет по меньшей мере приблизительно 40% от общего объема, более предпочтительно, объем жидкой фазы составляет по меньшей мере приблизительно 45% от общего объема и более предпочтительно, объем жидкой фазы составляет по меньшей мере приблизительно 50% от общего объема, или слизь выводится до блокирования или ингибирования вызванной функции (например, до тех пор, пока дыхательные пути пациента не очистятся до такой степени, чтобы начать откашливать жидкость). В общем случае предпочтительно, чтобы разжижение мокроты или слизи возрастало медленно и постепенно вплоть до очистки дыхательных путей или других заблокированных путей (например, в желудочно-кишечном или репродуктивном тракте), не допуская чрезмерного разжижения мокроты. Чрезмерное разжижении слизи или мокроты нежелательно, поскольку оно может быть пагубным для пациента (например, разжиженная мокрота до выведения из организма пациента, стекая, может залить периферические дыхательные пути жидкостью, которая может быть заражена). Предпочтительно, контакт белка, пептида или композиции согласно изобретению со слизью или мокротой приводит к повышению разжижения слизи и мокроты по меньшей мере на 1% от объема по сравнению с состоянием до лечения, более предпочтительно, к повышению на по меньшей мере приблизительно 2% и так далее с шагом 1%, до очищения дыхательных путей или других заблокированных путей пациента. После достижения такой очистки, например, путем удаления так называемых "слизевых пробок" для улучшения доступа препарата в малые дыхательные пути и альвеолы, можно применять поддерживающую терапию с использованием пониженных дозе целью поддержания нормального количества дисульфидных связей во вновь секретируемых белках муцина.
В одном аспекте лечение осуществляют в комбинации с способами удаления разжиженного материала из пораженной ткани (дыхательных путей, желудочно-кишечного тракта, репродуктивного тракта) пациента. Например, в случае дыхательной системы можно использовать способ согласно настоящему изобретению в сочетании с постуральным дренажем, хаффингом и другими дыхательными упражнениями, или любой другой подходящий способ для откашливания разжиженной слизи или мокроты.
В соответствии с настоящим изобретением, слизь или мокроту пациента, подлежащего лечению, приводят в контакт с белком (или композицией, содержащей белок), содержащим моноцистеиновый активный центр тиоредоксина в восстановленном состоянии. Белок эффективно снижает вязкость и липкость мокроты или слизи и/или повышает разжижение мокроты или слизи по сравнению с состоянием до контакта. Как описано ранее, тиоредоксин представляет собой белок дисульфидредуктазы, содержащийся во многих организмах, участвующий во многочисленных тиол-зависимых восстановительных процессах в клетках. В организме человека тиоредоксин также называют фактором Т-клеточного лейкоза взрослых (ADF). Внутри клеток большая часть этого повсеместно встречающегося низкомолекулярного (11700) белка остается в восстановленном состоянии. Восстановленный или окисленный тиоредоксин может проникать в интактные клетки или поглощаться клеточной мембраной, где его небольшое количество постепенно интернализуется с течением времени. Нативный тиоредоксин содержит два смежных остатка цистеина в активном центре, которые в окисленном белке образуют дисульфидный мостик, расположенный в выступе трехмерной структуры белка. Флавопротеин тиоредоксинредуктаза катализирует NADPH-зависимое восстановление этого дисульфида. Кроме того, рекомбинантные варианты тиоредоксинредуктазы с модифицированной специфичностью кофактора может использовать NADH вместо или в дополнение к NADPH, как описано в патенте США 7,071,307, включенном в настоящий документ посредством ссылки. Небольшое повышение уровня тиоредоксина может вызвать глубокие изменения редокс-статуса сульфгидрильно-дисульфидных групп в белках.
Известно, что в дополнение к своей способности восстанавливать клеточные белки тиоредоксин может действовать непосредственно в качестве антиоксиданта (например, путем предотвращения окисления окисляемого субстрата путем захвата активных форм кислорода), хотя, в отличие от других тиолов, тиоредоксин, как правило, не участвует в окислительном стрессе в клетках за счет аутоокисления (например, образования супероксид-радикалов путем аутоокисления). В патенте США №5985261, принадлежащем White et al., supra, показано, что тиоредоксин непосредственно индуцирует продукцию MnSOD и что такая индукция осуществляется тиоредоксином в восстановленном состоянии.
"Моноцистеиновый активный центр тиоредоксина" согласно настоящему изобретению содержит аминокислотную последовательность C-X-X-X (SEQ ID NO: 17) (нативная последовательность или последовательность дикого типа содержит аминокислотную последовательность С-Х-Х-С (SEQ ID NO: 16)). В настоящем документе аминокислотные остатки, обозначенные буквой "С", являются остатками цистеина, а аминокислотные остатки, обозначенные буквой "X", могут являться остатком любой аминокислоты, кроме остатка цистеина, и, в частности, остатком любой из оставшихся 20 стандартных аминокислот. Такой моноцистеиновый активный центр тиоредоксина согласно настоящему изобретению предпочтительно содержит аминокислотную последовательность C-G-P-X (SEQ ID NO: 18), в то время как нативная последовательность или последовательность дикого типа содержит аминокислотную последовательность C-G-P-C (SEQ ID NO: 1). Моноцистеиновый активный центр тиоредоксина может дополнительно содержать аминокислотную последовательность Х-С-Х-Х-Х-Х (SEQ ID NO: 19), в то время как нативная последовательность или последовательность дикого типа содержит аминокислотную последовательность Х-С-Х-Х-С-Х (SEQ ID NO: 20). Предпочтительно, моноцистеиновый активный центр тиоредоксина согласно настоящему изобретению содержит аминокислотную последовательность X-C-G-P-X-X (SEQ ID NO: 21), где аминокислотный остаток, обозначенный буквой "G", является остатком глицина, а аминокислотный остаток, обозначенный буквой "Р", является остатком пролина, в то время как нативная последовательность или последовательность дикого типа содержит аминокислотную последовательность X-C-G-P-C-X (SEQ ID NO: 22). Более предпочтительно, моноцистеиновый активный центр тиоредоксина согласно настоящему изобретению содержит аминокислотную последовательность W-C-G-P-X-K (SEQ ID NO: 23), где аминокислотный остаток, обозначенный буквой "G", является остатком триптофана, а аминокислотный остаток, обозначенный буквой "K", является остатком лизина, а нативная последовательность содержит аминокислотную последовательность W-C-G-P-C-K (SEQ ID NO: 3). Предпочтительно, моноцистеиновый активный центр тиоредоксина может содержать аминокислотную последовательность C-X-X-S (SEQ ID NO: 24). Такой моноцистеиновый активный центр тиоредоксина предпочтительно содержит аминокислотную последовательность C-G-P-S (SEQ ID NO: 1). Моноцистеиновый активный центр тиоредоксина может дополнительно содержать аминокислотную последовательность X-C-X-X-S-X (SEQ ID NO: 25), X-C-G-P-S-X (SEQ ID NO: 26) или W-C-G-P-S-K (SEQ ID NO: 27), в котором аминокислотные остатки, обозначенные буквой "X", могут являться остатком любой аминокислотой, кроме остатка цистеина. Упоминание "активного центра тиоредоксина" включает моноцистеиновый активный центр тиоредоксина и нативный активный центр тиоредоксина или активный центр тиоредоксина дикого типа.
В одном аспекте изобретения белок, содержащий моноцистеиновый активный центр тиоредоксина, является полноразмерным белком тиоредоксина или любым его фрагментом, содержащим структурный и функциональный моноцистеиновый активный центр тиоредоксина, как описано выше. Предпочтительные белки тиоредоксина, содержащие моноцистеиновый активный центр тиоредоксина, включают тиоредоксин прокариот, тиоредоксин дрожжей, тиоредоксин растений и тиоредоксин млекопитающих, причем особенно предпочтительным является тиоредоксин человека. Нуклеотидные и аминокислотные последовательности тиоредоксина различных организмов хорошо известны в данной области техники и входят в настоящее изобретение. Например, SEQ ID NO: 4-15 представляют собой аминокислотные последовательности тиоредоксина Pseudomonas syringae (SEQ ID NO: 4), Porphyromonas gingivalis (SEQ ID NO: 5), Listeria monocytogenes (SEQ ID NO: 6), Saccharomyces cerevisiae (SEQ ID NO: 7), Gallus gallus (SEQ ID NO: 8), Mus musculus (SEQ ID NO: 9), Rattus norvegicus (SEQ ID NO: 10), Bos taurus (SEQ ID NO: 11), Homo sapiens (SEQ ID NO: 12), Arabidopsis thaliana (SEQ ID NO: 13), Zea mays (SEQ ID NO: 14) и Oryza sativa (SEQ ID NO: 15). В каждой из этих последовательностей можно найти мотив X-C-G-P-C-X (SEQ ID NO: 22) (содержащий мотив CGPC SEQ ID NO: 1) в следующих положениях: SEQ ID NO: 4 (положения 33-38), SEQ ID NO: 5 (положения 28-33), SEQ ID NO: 6 (положения 27-32), SEQ ID NO: 7 (положения 29-34), SEQ ID NO: 8 (положения 31-36), SEQ ID NO: 9 (положения 31-36), SEQ ID NO: 10 (положения 31-36), SEQ ID NO: 11 (положения 31-36), SEQ ID NO: 12 (положения 31-36), SEQ ID NO: 13 (положения 59-64), SEQ ID NO: 14 (положения 88-93) и SEQ ID NO: 15 (положения 94-99). Кроме того, раскрыта трехмерная структура нескольких белков тиоредоксина, в том числе тиоредоксина человека и бактериального тиоредоксина. Таким образом, структура и активный центр тиоредоксина нескольких организмов хорошо известны в данной области техники, и специалист в данной области техники может выявить и получить фрагменты или гомологи полноразмерных белков тиоредоксина, в том числе белков тиоредоксина, содержащих моноцистеиновый активный центр тиоредоксина, которые можно применять в настоящем изобретении.
Фраза "в восстановленном состоянии", в частности, описывает состояние остатков цистеина в активном центре белка или пептида согласно настоящему изобретению. В восстановленном состоянии соседние остатки цистеина образуют дитиол (т.е. две свободные сульфгидрильные группы, -SH). В противоположность этому, в окисленной форме такие остатки цистеина образуют внутримолекулярный дисульфидный мостик; такую молекулу можно называть цистином. В восстановленном состоянии моноцистеиновый активный центр тиоредоксина может участвовать в окислительно-восстановительных реакциях путем обратимого окисления тиола его активного центра в дисульфид, и катализировать реакции тиол-дисульфидного обмена, приводящие к образованию ковалентной связи с одним из дисульфидов-мишеней Cys. Для белков или пептидов согласно настоящему изобретению, содержащих моноцистеиновый активный центр тиоредоксина, N-концевой цистеин в активном центре находится в восстановленном состоянии в виде монотиола и поэтому может образовывать стабильный смешанный дисульфид с остатком цистеина на белке-мишени.
В настоящем документе белок согласно настоящему изобретению, содержащей моноцистеиновый активный центр тиоредоксина, может представлять собой моноцистеиновый активный центр тиоредоксина как таковой или моноцистеиновый активный центр тиоредоксина, соединенный с другими аминокислотами гликозидными связями. Таким образом, минимальный размер белка или пептида согласно настоящему изобретению составляет от приблизительно 4 до приблизительно 6 аминокислот, причем предпочтительный размер зависит от желательности полноразмерного, гибридного, поливалентного белка или просто функциональных фргаментов такого белка. Предпочтительно, длина белка или пептида согласно настоящему изобретению составляет от приблизительно 4 до приблизительно 100 аминокислотных остатков или более, причем специально предусмотрено использование пептидов любой промежуточной длины, выраженной в целых числах (т.е., 4, 5, 6, 7…99, 100, 101 …). Он также может представлять собой короткий пептид, имитирующий тиоредоксин и заблокированный по N и С-концам, как описано в Bachnoff et al., Free Radical Biol Med 50:1355-67, 2011. В дополнительном предпочтительном варианте реализации белок согласно настоящему изобретению может представлять собой полноразмерный белок или любой гомолог такого белка. В настоящем документе термин "гомолог" используется для обозначения белка или пептида, отличающегося от нативного белка или пептида (то есть, "прототипа" или белка "дикого типа") за счет модификаций природного белка или пептида, однако сохраняющего основную структуру белка и боковых цепей природной формы, и/или сохраняющего основную трехмерную структуру по меньшей мере биологически активного фрагмента (например, активного центра тиоредоксина) нативного белка. Такие изменения включают изменения боковых цепей одной или нескольких аминокислот; изменения одной или нескольких аминокислот, в том числе делеции (например, укороченная версия белка или пептида (фрагмент)), инсерции и/или замены; изменения в стереохимии одного или нескольких атомов; и/или незначительные модификации, включая, в числе прочего, метилирование, гликозилирование, фосфорилирование, ацетилирование, миристоилирование, пренилирование, пальмитоилирование, амидирование и/или добавление гликозилфосфатидилинозита, но не ограничиваются ими. В соответствии с настоящим изобретением любой белок или пептид, применимый в настоящем изобретении, в том числе гомологи природных белков тиоредоксина, содержит моноцистеиновый активный центр тиоредоксина так, что в восстановленном состоянии указанный белок или пептид способен участвовать в окислительно-восстановительных реакциях путем окисления тиола его активного центра в дисульфид в и/или снижать вязкость или липкость слизи или мокроты или повышать разжижение слизи или мокроты. В настоящем документе белок или пептид, содержащий моноцистеиновый активный центр тиоредоксина, может обладать характеристиками, аналогичными характеристикам тиоредоксина, и предпочтительно представляет собой тиоредоксин, выбраный из группы тиоредоксина прокариот, тиоредоксина грибов (в том числе дрожжей), тиоредоксина растений или тиоредоксина млекопитающих. В особенно предпочтительном варианте реализации белок является тиоредоксином человека.
Гомологи могут возникать в результате естественной аллельной изменчивости или природной мутации. Природный аллельный вариант нуклеиновой кислоты, кодирующей белок, представляет собой ген, который находится практически в том же локусе (или локусах) генома, что и ген, кодирующий такой белок, однако из-за естественной изменчивости, вызванной, например, мутацией или рекомбинацией, обладает аналогичной, но не идентичной последовательностью. Аллельные варианты, как правило, кодируют белки, обладающие такой же активностью, что и белок, кодируемый геном, с которым их сравнивают. Аллельные варианты одного класса могут кодировать один и тот же белок, но обладать разными нуклеотидными последовательностями вследствие вырожденности генетического кода. Аллельные варианты также могут содержать изменения в 5'- или 3'-нетранслируемых областях гена (например, в регуляторных управляющих областях). Аллельные варианты хорошо известны специалистам в данной области техники.
Гомологи можно получить с использованием методик, известных в данной области техники и предназначенных для получения белков, включая прямые модификации выделенного природного белка, прямой синтез белка или модификации нуклеотидной последовательности, кодирующей белок, используя, например, методики классических или рекомбинантных ДНК для случайного или направленного мутагенеза, но не ограничиваясь этим.
Модификации гомологов, по сравнению с белком дикого типа, усиливают, ослабляют или практически не изменяют основную биологическую активность гомолога по сравнению с нативным белком. В общем случае, биологическая активность или биологическое действие белка относится к любой функции(ям), проявляемой или выполняемой белком, которую измеряют или наблюдают у природной формы белка in vivo (т.е. в естественном физиологическом окружении белка) или in vitro (т.е. в лабораторных условиях). Модификации белка, например, в гомологе или имитаторе (обсуждаемых ниже) могут привести к образованию белков с такой же биологической активностью, как и у природного белка, или белков с пониженной или повышенной биологической активностью по сравнению с природным белком. Модификации, которые приводят к снижению экспрессии белка или уменьшению активности белка, можно называть инактивацией (полной или частичной), негативной модуляцией или ослаблением действия белка. Аналогично, модификации, которые приводят к усилению экспрессии белка или повышению активности белка, можно называть амплификацией, сверхпродукцией, активацией, усилением, положительной модуляцией или усилением действия белка.
В одном варианте реализации белки или пептиды, содержащие моноцистеиновый активный центр тиоредоксина, могут представлять собой продукты разработки лекарственных средств или селекции и могут быть получены с использованием различных способов, известных в данной области техники. Такие белки или пептиды можно называть имитаторами. Термин "имитатор" относится к любому пептидному или непептидному соединению, которое способно имитировать биологическое действие природного пептида, часто из-за того, что имитатор обладает основной структурой, имитирующей основную структуру природного пептида, и/или обладает характерными биологическими свойствами природного пептида. Имитаторы могут включать пептиды, существенно отличающиеся от прототипа, например, не обладающие сходством боковой цепи с природным пептидом (такие модификации, например, могут снизить его восприимчивость к разложению); антиидиотипические и/или каталитические антитела или их фрагменты; небелковые фрагменты выделенного белка (например, углеводные структуры); или синтетические или природные органические вещества, в том числе нуклеиновые кислоты и лекарственные вещества, полученные посредством, например, комбинаторной химии, но не ограничиваются ими. Такие имитаторы можно сконструировать, выделить или иным образом выявить с помощью разнообразных способов, известных в данной области техники. Различные способы разработки лекарственных средств, которые можно применять для разработки или выбора имитаторов или других терапевтических соединений, применимых в настоящем изобретении, описаны в работе Maulik et al., 1997, Molecular Biotechnology: Therapeutic Applications and Strategies, Wiley-Liss, Inc., полностью включенной в настоящий документ посредством ссылки. Пептиды-имитаторы тиоредоксина, обладающие мощной и селективной окислительно-восстановительной активностью, описаны в работе Bachnoff et al., Free Radical Biol Med 50:1355-67 (2011) и полностью включены в данное описание посредством ссылки.
Имитатор можно получить, например, с помощью стратегий молекулярного разнообразия (сочетания родственных стратегий, позволяющих быстро создавать крупные библиотеки химически разнообразных веществ), библиотек природных или синтетических соединений, в частности, химических или комбинаторных библиотек (например, библиотек соединений, отличающихся по последовательности или размеру, однако содержащих аналогичные блоки) или путем рациональной, направленной или случайной разработки лекарственных средств. См., например, Maulik et al., supra.
В стратегии молекулярного разнообразия синтезируют крупные библиотеки соединений, например, пептидов, олигонуклеотидов, углеводов и/или синтетических органических молекул с использованием биологических, ферментативных и/или химических подходов. Критические параметры при разработке стратегии молекулярного разнообразия включают разнообразие субъединиц, размер молекул и разнообразие библиотек. Общая цель скрининга таких библиотек заключается в использовании последовательного применения комбинаторной селекции для получения лигандов с высоким сродством к желательной мишени с последующей оптимизацией основных молекул с помощью стратегий случайного или направленного конструирования. Способы молекулярного разнообразия подробно описаны в работе Maulik, et al., ibid.
Maulik, et al. также описывают, например, способы направленного конструирования, при котором пользователь направляет процесс создания новых молекул из библиотеки соответствующим образом выбранных фрагментов; случайного конструирования, при котором пользователь использует генетический или другой алгоритм для случайного мутирования фрагментов и их комбинаций с одновременным применением критерия выбора с целью оценки пригодность лигандов-кандидатов; и табличный подход, при котором пользователь вычисляет энергию взаимодействия между трехмерными структурами рецепторов и небольшими фрагментами зондов, а затем путем соединения благоприятных областей зондов воедино.
Способы создания разнообразия, например, описанные выше, можно объединять с другими способами, предназначенными для улучшения функциональных или фармакологических характеристик, особенно для молекул уменьшенного размера, например, имитаторов активного центра. Например, один подход, перспективность которого продемонстрирована в исследованиях ранней стадии, представляет собой использование α-спиральных пептидов, соединенных углеводородными группами, нового класса синтетических мини-белков с заблокированной биологически активной α-спиральной структурой за счет сайт-специфического внедрения химической связи, полностью углеводородного связывающего элемента. Сшивание может значительно улучшить фармакологические характеристики пептидов, увеличивая их сродство к мишени и протеолитическую устойчивость при создании уменьшенных пептидных версий более крупных белков/ферментов, подходящих для химического синтеза (Verdine, G.L. and Hilinsky, G.J., Methods Enzymol, 503:3-33, 2012).
В одном варианте реализации настоящего изобретения белок, пригодный для применения в настоящем изобретении, содержит аминокислотную последовательность, содержащую, состоящую в основном из или состоящую из полноразмерной последовательности белка тиоредоксина или любого его фрагмента, содержащего моноцистеиновый активный центр тиоредоксина, как описано в настоящем документе. Например, любая из нативных последовательностей SEQ ID NO: 4-15 или их фрагмент или другой гомолог, содержащий моноцистеиновый активный центр тиоредоксина, описанный в настоящем документе, входит в настоящее изобретение. Такие гомологи могут включать белки с аминокислотной последовательностью, по меньшей мере на приблизительно 10% идентичной аминокислотной последовательности полноразмерного белка тиоредоксина, или по меньшей мере на 20% идентичной, по меньшей мере на 30% идентичной, по меньшей мере на 40% идентичной или по меньшей мере на 50% идентичной, по меньшей мере на 60% идентичной или по меньшей мере на 70% идентичной, по меньшей мере на 80% идентичной, по меньшей мере на 90% идентичной, или более чем на 95% идентичной аминокислотной последовательности полноразмерного белка тиоредоксина, включая любой процент, выраженный в целых числах от 10% до 100% (10%, 11%, 12%, … 98%, 99%, 100%).
В настоящем документе, если не указано иное, упоминание процентной (%) идентичности относится к оценке гомологии, которую осуществляют с помощью: (1) поиска гомологии в BLAST 2.0 Basic BLAST с помощью blastp для поиска по аминокислотным последовательностям и blastn для поиска по нуклеотидным последовательностям со стандартными параметрами по умолчанию и фильтрации запросов последовательностей по областям низкой сложности по умолчанию (описано в работе Altschul, S.F., Madden, T.L.,  , А.А., Zhang, J., Zhang, Z., Miller, W. & Lipman, D.J. (1997) "Gapped BLAST and PSI-BLAST: a new generation of protein database search programs." Nucleic Acids Res. 25:3389-3402, полностью включенной в настоящий документ посредством ссылки); (2) выравнивания в BLAST 2 (с использованием параметров, описанных ниже); (3) и/или PSI-BLAS с параметрами по умолчанию (итерацонный BLAST, специфический по отношению к положению. Следует отметить, что из-за некоторых различий в стандартных параметрах между BLAST 2.0 Basic BLAST и BLAST 2, программа BLAST 2 может распознать две специфические последовательность как обладающие значительной гомологией, тогда как поиск в BLAST 2.0 Basic BLAST с использованием одной из последовательностей как последовательности для запроса может не выявить вторую последовательность среди результатов с максимальным совпадением. Кроме того, PSI-BLAST обеспечивает автоматизированную, простую в использовании версию «профильного» поиска, который является чувствительным инструментом для поиска гомологичных последовательностей. Программа сначала выполняет поиск по базе данных BLAST с учетом разрывов. Программа PSI-BLAST использует информацию любых значимых результатов выравнивания, полученных при конструировании матрицы оценок, специфических по отношению к положению, которая заменяет последовательность для запроса в следующем цикле поиска по базе данных. Таким образом, следует понимать, что процентную идентичность можно определить с помощью любой из этих программ.
, А.А., Zhang, J., Zhang, Z., Miller, W. & Lipman, D.J. (1997) "Gapped BLAST and PSI-BLAST: a new generation of protein database search programs." Nucleic Acids Res. 25:3389-3402, полностью включенной в настоящий документ посредством ссылки); (2) выравнивания в BLAST 2 (с использованием параметров, описанных ниже); (3) и/или PSI-BLAS с параметрами по умолчанию (итерацонный BLAST, специфический по отношению к положению. Следует отметить, что из-за некоторых различий в стандартных параметрах между BLAST 2.0 Basic BLAST и BLAST 2, программа BLAST 2 может распознать две специфические последовательность как обладающие значительной гомологией, тогда как поиск в BLAST 2.0 Basic BLAST с использованием одной из последовательностей как последовательности для запроса может не выявить вторую последовательность среди результатов с максимальным совпадением. Кроме того, PSI-BLAST обеспечивает автоматизированную, простую в использовании версию «профильного» поиска, который является чувствительным инструментом для поиска гомологичных последовательностей. Программа сначала выполняет поиск по базе данных BLAST с учетом разрывов. Программа PSI-BLAST использует информацию любых значимых результатов выравнивания, полученных при конструировании матрицы оценок, специфических по отношению к положению, которая заменяет последовательность для запроса в следующем цикле поиска по базе данных. Таким образом, следует понимать, что процентную идентичность можно определить с помощью любой из этих программ.
Две специфические последовательности можно выровнять друг с другом с использованием последовательности BLAST 2, как описано в работе Tatusova and Madden, (1999), "Blast 2 sequences - a new tool for comparing protein and nucleotide sequences", FEMS Microbiol Lett. 174:247-250, полностью включенной в настоящий документ посредством ссылки. Выравнивание последовательностей BLAST 2 выполняют в blastp или blastn, с использованием алгоритма BLAST 2.0 для поиска в Gapped BLAST (BLAST 2.0) между двумя последовательностями, позволяющего вводить разрывы (делеции и инсерции) в результатах выравнивания. Для ясности в настоящем документе, выравнивание последовательностей BLAST 2 выполняют с использованием следующих стандартных параметров.
Для blastn с использованием матрицы 0 BLOSUM62:
Компенсация за совпадение = 1
Штраф за несоответствие = -2
Штрафы за внесение разрыва (5) и расширение разрыва (2)
x_dropoff разрыва (50) фильтр по ожидаемому (10) размеру слова (11) (вкл)
Для blastp с использованием матрицы 0 BLOSUM62:
Штрафы за внесение разрыва (11) и расширение разрыва (1)
x_dropoff разрыва (50) фильтр по ожидаемому (10) размеру слова (3) (вкл)
Белок, который можно использовать в настоящем изобретении, может также включать белки с аминокислотной последовательностью, состоящей из по меньшей мере 10 смежных аминокислотных остатков любого полноразмерного белка тиоредоксина, содержащих моноцистеиновый активный центр тиоредоксина (нативной последовательностью, представленной в SEQ ID NO: 4-15, т.е., 10 смежных аминокислотных остатков, обладающих 100% идентичностью с 10 смежными аминокислотами эталонной последовательности). В других вариантах реализации гомолог белка тиоредоксина содержит аминокислотные последовательности, состоящие из по меньшей мере 15, или по меньшей мере 20, или по меньшей мере 25, или по меньшей мере 30, или по меньшей мере 35, или по меньшей мере 40, или по меньшей мере 45, или по меньшей мере 50, или по меньшей мере 55, или по меньшей мере 60, или по меньшей мере 65, или по меньшей мере 70, или по меньшей мере 75, или по меньшей мере 80 смежных аминокислотных остатков аминокислотной последовательности природного белка тиоредоксина, и т.д. вплоть до полноразмерного белка, включая любую промежуточную длину, выраженную в целых числах (10, 11, 12..), и содержащие моноцистеиновый активный центр.
Согласно настоящему изобретению термин «смежные» или «последовательные» по отношению к последовательностям, описанным в настоящем документе, связан с непрерывной последовательностью. Например, если первая последовательность содержит 30 смежных (или последовательных) аминокислот второй последовательности, это означает, что первая последовательность содержит непрерывную последовательность из 30 аминокислотных остатков, на 100% идентичную непрерывной последовательности из 30 аминокислотных остатков второй последовательности. Аналогично, если первая последовательность обладает «100% идентичностью» второй последовательности, это означает, что первая последовательность точно соответствует второй последовательности без пропусков между нуклеотидами или аминокислотами.
В другом варианте реализации белок, который можно применять в настоящем изобретении, включает белок с аминокислотной последовательностью, в достаточной степени аналогичной аминокислотной последовательности природного тиоредоксина, чтобу нуклеотидная последовательность, кодирующая гомолог, может гибридизироваться в условиях умеренной, высокой или очень высокой жесткости (описанных ниже) с молекулой нуклеиновой кислоты, кодирующей природный белок тиоредоксина (т.е. с комплементарной цепью нуклеиновой кислоты, кодирующей аминокислотную последовательность природного тиоредоксина). Такие условия гибридизации подробно описаны ниже.
Нуклеотидная последовательность, комплементарная нуклеотидной последовательности, кодирующей белок тиоредоксина согласно настоящему изобретению, относится к нуклеотидной последовательности цепи нуклеиновой кислоты, комплементарной цепи, кодирующей тиоредоксин. Следует учитывать, что двуцепочечная ДНК, кодирующая заданную аминокислотную последовательность, состоит из одноцепочечной ДНК и комплементарной ей цепи с последовательностью, комплементарной одноцепочечной ДНК. Таким образом, молекулы нуклеиновой кислоты согласно настоящему изобретению могут быть двуцепочечными или одноцепочечными и включают молекулы нуклеиновой кислоты, образующие стабильные гибриды при жестких условиях гибридизации с нуклеотидной последовательностью, кодирующей аминокислотную последовательность белка тиоредоксина, и/или с комплементарной цепью нуклеотидной последовательности, кодирующей такую аминокислотную последовательность. Специалистам в данной области техники известны способы получения комплементарной последовательности.
В настоящем документе упоминание условий гибридизации относится к стандартным условиям гибридизации, при которых молекулы нуклеиновой кислоты используют для выявления аналогичных молекул нуклеиновой кислоты. Такие стандартные условия описаны, например, в справочнике Sambrook et al., Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Labs Press, 1989. Sambrook et al., ibid. полностью включен в настоящий документ посредством ссылки (конкретно, см. страницы 9.31-9.62). Кроме того, формулы для расчета соответствующих условий гибридизации и отмывки с целью осуществления гибридизации при различной степени несовпадения нуклеотидов описаны, например, в работе Meinkoth et al., 1984, Anal. Biochem. 138, 267-284; Meinkoth et al., ibid, полностью включена в настоящий документ посредством ссылки.
Конкретнее, условия гибридизации и промывки умеренной жесткости, как указано в настоящем документе, относятся к условиям, обеспечивающим выделение молекул нуклеиновых кислот с по меньшей мере приблизительно 70% идентичностью нуклеотидной последовательности молекуле нуклеиновой кислоты, используемой в качестве зонда в реакции гибридизации (т.е., условиям, допускающим несовпадение приблизительно 30% или меньшего количества нуклеотидов). Условия гибридизации и промывки высокой жесткости, как указано в настоящем документе, относятся к условиям, обеспечивающим выделение молекул нуклеиновых кислот с по меньшей мере приблизительно 80% идентичностью нуклеотидной последовательности молекуле нуклеиновой кислоты, используемой в качестве зонда в реакции гибридизации (т.е., условиям, допускающим несовпадение приблизительно 20% или меньшего количества нуклеотидов). Условия гибридизации и промывки очень высокой жесткости, как указано в настоящем документе, относятся к условиям, обеспечивающим выделение молекул нуклеиновых кислот с по меньшей мере приблизительно 90% идентичностью нуклеотидной последовательности молекуле нуклеиновой кислоты, используемой в качестве зонда в реакции гибридизации (т.е., условиям, допускающим несовпадение приблизительно 10% или меньшего количества нуклеотидов). Как обсуждалось выше, специалист в данной области техники может использовать формулы, приведенные в Meinkoth et al., ibid., с целью расчета соответствующих условий гибридизации и промывки для достижения указанных уровней несовпадения нуклеотидных последовательностей. Такие условия зависят от состава образующихся гибридов - ДНК : РНК или ДНК : ДНК. Расчетные температуры плавления для гибридов ДНК : ДНК на 10°С меньше, чем для гибридов ДНК : РНК. В конкретных вариантах реализации жесткие условия гибридизации для гибридов ДНК : ДНК включают гибридизацию при ионной силе 6×SSC (0,9 М Na+), температуре от приблизительно 20°С до приблизительно 35°С (низкая жесткость), более предпочтительно, от приблизительно 28°С до приблизительно 40°С (более жесткие условия), и еще более предпочтительно, от приблизительно 35°С до приблизительно 45°С (еще более жесткие условия), при соответствующих условиях промывки. В конкретных вариантах реализации жесткие условия гибридизации для гибридов ДНК:ДНК включают гибридизацию при ионной силе 6×SSC (0,9 М Na+), температуре от приблизительно 30°С до приблизительно 45°С, более предпочтительно, от приблизительно 38°С до приблизительно 50°С, и еще более предпочтительно, от приблизительно 45°С до приблизительно 55°С, при условиях промывки аналогичной жесткости. Эти значения основаны на расчетах температуры плавления для молекул размером более приблизительно 100 нуклеотидов, при 0% формамида и содержании G + С приблизительно 40%. Кроме того, Tm можно рассчитать эмпирически, как изложено в Sambrook et al., supra, на страницах 9.31-9.62. В целом, условия промывки должны быть как можно более жесткими, и должны соответствовать выбранным условиям гибридизации. Например, условия гибридизации могут включать комбинацию параметров соли и температуры приблизительно на 20-25°С ниже расчетной Tm конкретного гибрида, а условия промывки, как правило, включают комбинацию параметров соли и температуры приблизительно на 12-20°С ниже расчетной Tm конкретного гибрида. Один из примеров условий гибридизации, подходящих для применения с гибридами ДНК ДНК, включает гибридизацию в течение 2-24 ч в 6 × SSC (50% формамид) при температуре приблизительно 42°C с последующими этапами промывки, включающими одну или более промывок при комнатной температуре в 2 × SSC с последующими дополнительными промывками при более высоких температурах и более низкой ионной силе (например, по меньшей мере одну промывку при температуре приблизительно 37°С в приблизительно 0,1 × -0,5 × SSC, а затем по меньшей мере одну промывку при температуре приблизительно 68°С в приблизительно 0,1 × -0,5 × SSC).
Белок согласно настоящему изобретению также может быть гибридным белком, содержащим сегмент, содержащий моноцистеиновый активный центр тиоредоксина, и гибридный сегмент, который может обладать различными функциями. Например, такой гибридный сегмент может действовать в качестве инструмента для упрощения очистки белка согласно настоящему изобретению, например, обеспечивая очистку полученного гибридного белка с использованием аффинной хроматографии. Подходящий гибридный сегмент может являться областью любого размера, обладающей желательной функцией (например, придающей белку повышенную стабильность, повышенную иммуногенность, и/или упрощающей очистку белка). Применение одного или более гибридных сегментов входит в рамки настоящего изобретения. Гибридные сегменты можно соединять с N- и/или С-концами сегмента, содержащего моноцистеиновый активный центр тиоредоксина. Связь между гибридными сегментами и доменами гибридных белков, содержащими активный центр тиоредоксина, может быть подвержена расщеплению с целью обеспечить высвобождение доменов, содержащих моноцистеиновый активный центр тиоредоксина, из таких белков простейшим способом. Гибридные белки предпочтительно получают путем культивирования рекомбинантных клетки, трансформированных гибридной молекулой нуклеиновой кислоты, кодирующей белок, содержащий гибридный сегмент, присоединенный к С- и/или N-концу домена, содержащего моноцистеиновый активный центр тиоредоксина.
В одном варианте реализации белок или пептид, содержащий моноцистеиновый активный центр тиоредоксина, подходящий для применения со способом согласно настоящему изобретению, включает белок или пептид, содержащий моноцистеиновый активный центр тиоредоксина, полученный из видов животных, практически аналогичных животному, в организм которого надлежит ввести указанный белок. В еще одном варианте реализации для заданного пациента можно применять любой белок или пептид, содержащий моноцистеиновый активный центр тиоредоксина, в том числе полученный из различных источников, например, микроорганизмов, растений и грибов.
В одном варианте реализации настоящего изобретения можно получить любую из аминокислотных последовательностей, описанных в настоящем документе, например, аминокислотную последовательность природного белка тиоредоксина или тиоредоксина, содержащего моноцистеиновый активный центр, содержащую по меньшей мере одну или вплоть до приблизительно 20 дополнительных гетерологичных аминокислот, фланкирующих С- и/или N-концы указанной аминокислотной последовательности. Полученный белок или полипептид можно называть "состоящим по большей части из" указанной аминокислотной последовательности. Согласно настоящему изобретению, гетерологичные аминокислоты представляют собой последовательность аминокислот, не встречающихся в природных условиях (т.е. не встречающихся в природе, in vivo) непосредственно перед или после указанной аминокислотной последовательности, или аминокислот, не имеющих отношения к функции указанной аминокислотной последовательности, или не кодируемых нуклеотидами, фланкирующими природную нуклеотидную последовательность, кодирующую указанную аминокислотную последовательность, как это имеет место в гене, если такие нуклеотиды природной последовательности транслировать с использованием стандартных кодонов для организма, из которого получена данная аминокислотная последовательность. Аналогично, фраза «состоящий по большей части из», при упоминании нуклеотидной последовательности в настоящем документе, относится к нуклеотидной последовательности, кодирующей указанную аминокислотную последовательность, которую могут фланкировать по меньшей мере один или до приблизительно 60 дополнительных гетерологичных нуклеотидов с 5'- и/или 3'-конца нуклеотидной последовательности, кодирующей указанную аминокислотную последовательность. Гетерологичные нуклеотиды представляют собой нуклеотиды, не встречающиеся в природных условиях (т.е. не встречающиеся в природе, in vivo) непосредственно перед или после нуклеотидной последовательности, кодирующей указанную аминокислотную последовательность, как это происходит в природном гене, или не кодирующие белок, придающий какую-либо дополнительную функцию белку или изменяющий функцию белка с заданной аминокислотной последовательностью.
В еще одном варианте реализации белок или пептид, содержащий моноцистеиновый активный центр тиоредоксина, подходящий для применения со способом согласно настоящему изобретению, включает выделенный или биологически чистый белок. Термины "выделенный" и "биологически чистый" как таковые не обязательно отражают степень, до которой очищен данный белок. Выделенный белок согласно настоящему изобретению можно, например, получить из природного источника, получить с использованием технологии рекомбинантной ДНК (например, полимеразной цепной реакции (ПЦР), клонирования), или синтезировать химическим путем.
В еще одном варианте реализации химически синтезированный белок или пептид, содержащий моноцистеиновый активный центр тиоредоксина согласно настоящему изобретению, также может относиться к стабилизированной версии, например, содержащей активный центр, структурно ограниченный с помощью технологии сшитых пептидов, путем циклизации или путем ограничения по N- или С-концу.
Предпочтительно белок, содержащий моноцистеиновый активный центр тиоредоксина для применения в способах согласно изобретению, имеет период полувыведения in vivo, достаточный, чтобы вызвать измеримое или обнаружимое повышение разжижения (или снижение вязкости или липкости) слизи или мокроты у пациента, и/или вызвать измеримое, обнаружимое или предполагаемое терапевтически благоприятное действие для пациента, связанное со слизью и мокротой у пациента. Такой период выведения можно получить за счет способа доставки такого белка. Белок согласно настоящему изобретению предпочтительно обладает периодом полувыведения из организма животного более приблизительно 5 минут, более предпочтительно - более приблизительно 4 часов, и еще более предпочтительно - более приблизительно 16 часов. В предпочтительном варианте реализации белок согласно настоящему изобретению обладает периодом полувыведения из организма животного от приблизительно 5 минут до приблизительно 24 часов, предпочтительно от приблизительно 2 часов до приблизительно 16 часов, и более предпочтительно от приблизительно 4 часов до приблизительно 12 часов.
Дополнительные варианты реализации настоящего изобретения включают молекулы нуклеиновой кислоты, кодирующие белок или пептид, содержащий моноцистеиновый активный центр тиоредоксина. Такие молекулы нуклеиновой кислоты можно применять для получения белка, который можно применять в способе согласно настоящему изобретению in vitro или in vivo. Молекула нуклеиновой кислоты согласно настоящему изобретению включает молекулу нуклеиновой кислоты, содержащую, состоящую главным образом из или состоящую из нуклеотидной последовательности, кодирующей любой из белков, описанных выше в настоящем документе. В соответствии с настоящим изобретением, выделенная молекула нуклеиновой кислоты представляет собой молекулу нуклеиновой кислоты (полинуклеотид), выделенную из природной среды (т.е., подвергавшуюся манипуляциям человека), и может включать ДНК, РНК или производные ДНК или РНК, в том числе кДНК. Термин "выделенный" как таковой не отражает степень, до которой очищена молекула нуклеиновой кислоты. Хотя фраза "молекула нуклеиновой кислоты" относится прежде всего к физической молекуле нуклеиновой кислоты, а фраза "нуклеотидная последовательность" относится прежде всего к последовательности нуклеотидов в молекуле нуклеиновой кислоты, эти две фразы можно использовать как взаимозаменяемые, особенно по отношению к молекуле нуклеиновой кислоты, или нуклеотидной последовательности, способной кодировать белок. Выделенную молекулу нуклеиновой кислоты согласно настоящему изобретению можно, например, получить из природного источника или получить с использованием технологии рекомбинантных ДНК (например, амплификации за счет полимеразной цепной реакции (ПЦР), клонирования), или химического синтеза. Выделенные молекулы нуклеиновых кислот могут включать, например, гены, природные аллельные варианты генов, кодирующие области или их фрагменты, а также кодирующие и/или регуляторные области, модифицированные путем нуклеотидных инсерций, делеций, замен и/или инверсий таким образом, что эта модификация не оказывает существенного влияния на способность молекулы нуклеиновой кислоты кодировать желательный белок согласно настоящему изобретению или образовывать стабильные гибриды с изолятами природного гена в жестких условиях. Выделенная нуклеиновая кислота может характеризоваться вырожденностью. В настоящем документе нуклеотидная вырожденность относится к возможности кодирования одной аминокислоты различными нуклеотидными кодонами. Таким образом, нуклеотидная последовательность молекулы нуклеиновой кислоты, кодирующая белок, который можно применять в настоящем изобретении, может меняться в зависимости от вырожденности.
В соответствии с настоящим изобретением, упоминание гена включает все нуклеотидные последовательности, имеющие отношение к природному гену (т.е. гену дикого типа), а также последовательности, имеющие отношение к моноцистеиновому активному центру тиоредоксина, например, регуляторные области, контролирующие продукцию белка, кодируемого этим геном (например, области, контролирующие транскрипцию, трансляцию или посттрансляционные процессы, но не ограничиваясь ими), а также саму кодирующую область. В еще одном варианте реализации ген может являться природным аллельным вариантом, содержащим последовательность, аналогичную, но не идентичную нуклеотидной последовательности, кодирующей данный белок. Аллельные варианты описаны выше. Фразы "молекула нуклеиновой кислоты" и "ген" можно использовать взаимозаменяемо, если молекула нуклеиновой кислоты содержит ген, как описано выше.
Предпочтительно, выделенную молекулу нуклеиновой кислоты согласно настоящему изобретению продуцируют с использованием технологии рекомбинантных ДНК (например, амплификации за счет полимеразной цепной реакции (ПЦР), клонирования) или химического синтеза. Выделенные молекулы нуклеиновой кислоты включают природные молекулы нуклеиновой кислоты и их гомологи, включая природные аллельные варианты и модифицированные молекулы нуклеиновой кислоты, содержащие инсерции, делеции, замены и/или инверсии нуклеотидов таким образом, что такие модификации оказывают желательное влияние на биологическую активность белка, но не ограничиваясь ими. Аллельные варианты и гомологи белка (например, белки, кодируемые гомологами нуклеиновой кислоты) подробно обсуждались выше.
Гомологи молекулы нуклеиновой кислоты можно получить с использованием ряда способов, известных специалистам в данной области техники (например, как описано в Sambrook et al., ibid). Например, молекулы нуклеиновых кислот можно модифицировать с использованием различных способов, включая классический мутагенез и технологию рекомбинантных ДНК (в том числе, без ограничения, сайт-специфический мутагенез, химическую обработку, рестриктазный гидролиз, лигирование фрагментов нуклеиновых кислот и/или ПЦР-амплификацию), или синтез смесей олигонуклеотидов и химическое лигирование, или рекомбинацию in vitro или in vivo смесей молекулярных групп с целью получения перегруппированной библиотеки молекул нуклеиновых кислот, содержащей множество их комбинаций, полученных в процессе перетасовки генов (т.е. молекулярной селекции; см., например, патент США №5605793, принадлежащий Stemmer; Minshull and Stemmer, Curr. Opin. Chem. Biol. 3:284-290, 1999; Stemmer, P.N.A.S. USA 91:10747-10751, 1994, все из которых полностью включены в настоящий документ посредством ссылок), но не ограничиваясь ими. Эти и другие подобные методики, известные специалистам в данной области техники, можно применять для эффективного внедрения множественных одновременных изменений в белок. Затем можно отобрать гомологи молекулы нуклеиновой кислоты путем гибридизации с данным геном, или прямого скрининга функции и биологической активности белков, кодируемых такими молекулами нуклеиновых кислот.
Один вариант реализации настоящего изобретения относится к рекомбинантной молекуле нуклеиновой кислоты, содержащей вышеописанную выделенную молекулу нуклеиновой кислоты, функционально связанную с по меньшей мере одной последовательностью, контролирующей транскрипцию. Конкретнее, в соответствии с настоящим изобретением рекомбинантная молекула нуклеиновой кислоты обычно содержит рекомбинантный вектор и выделенную молекулу нуклеиновой кислоты, как описано в настоящем документе. В соответствии с настоящим изобретением, рекомбинантный вектор представляет собой сконструированную (т.е. искусственно полученную) молекулу нуклеиновой кислоты, применяемую в качестве инструмента для манипулирования выбранной нуклеотидной последовательностью и/или для введения такой нуклеотидной последовательности в клетку-хозяина. Рекомбинантный вектор, следовательно, подходит для применения при клонировании, секвенировании и/или иных манипуляциях с выбранной нуклеотидной последовательностью, например, путем экспрессии и/или доставки выбранной нуклеотидной последовательности в клетку-хозяина с образованием рекомбинантной клетки. Такой вектор обычно содержит гетерологичные нуклеотидные последовательности, т.е. нуклеотидные последовательности, не встречающиеся в природных условиях рядом с клонируемой или доставляемой нуклеотидной последовательностью, хотя вектор также может содержать регуляторные нуклеотидные последовательности (например, промоторы, нетранслируемые области) встречающиеся в природных условиях рядом с нуклеотидными последовательностями согласно настоящему изобретению или последовательности, которые можно применять для экспрессии молекул нуклеиновых кислот согласно настоящему изобретению (подробно описанные ниже). Вектор может представлять собой РНК или ДНК прокариот или эукариот, и обычно представляет собой плазмиду. Вектор можно поддерживать в виде внехромосомного элемента (например, реплицирующейся плазмиды) или встроить в хромосому рекомбинантной клетки-хозяина, хотя в предпочтительном случае в большинстве вариантов применения изобретения вектор находится отдельно от генома. Вектор может целиком оставаться в клетке-хозяине, или при определенных условиях плазмидную ДНК можно удалить, оставляя в клетке молекулу нуклеиновой кислоты согласно настоящему изобретению. Встроенная молекула нуклеиновой кислоты может находиться под контролем хромосомного промотора, под контролем нативного или плазмидного промотора, или под контролем комбинации нескольких промоторов. Одиночные или множественные копии молекулы нуклеиновой кислоты можно встроить в хромосому. Рекомбинантный вектор согласно настоящему изобретению может содержать по меньшей мере один селективный маркер.
В одном варианте реализации рекомбинантный вектор, используемый в рекомбинантной молекуле нуклеиновой кислоты согласно настоящему изобретению, представляет собой экспрессирующий вектор. В настоящем документе фраза "экспрессирующий вектор" используется для обозначения вектора, подходящего для продукции кодируемого продукта (например, исследуемого белка). В данном варианте реализации нуклеотидную последовательность, кодирующую получаемый продукт (например, белок, содержащий моноцистеиновый активный центр тиоредоксина), вставляют в рекомбинантный вектор для получения рекомбинантной молекулы нуклеиновой кислоты. Нуклеотидную последовательность, кодирующую продуцируемый белок, вставляют в вектор, функционально связывая нуклеотидную последовательность с регуляторными последовательностями вектора, что позволяет осуществлять транскрипцию и трансляцию нуклеотидной последовательности в рекомбинантной клетке-хозяине.
В еще одном варианте реализации изобретения рекомбинантная молекула нуклеиновой кислоты содержит вирусный вектор. Вирусный вектор содержит выделенную молекулу нуклеиновой кислоты согласно настоящему изобретению, встроенную в вирусный геном или его фрагмент, причем молекула нуклеиновой кислоты упакована в вирусную оболочку, что позволяет ДНК проникать в клетку. Можно использовать ряд вирусных векторов, включая векторы на основе альфавирусов, поксвирусов, аденовирусов, герпесвирусов, лентивирусов, адено-ассоциированных вирусов и ретровирусов, но не ограничиваясь ими.
Как правило, рекомбинантная молекула нуклеиновой кислоты включает по меньшей мере одну молекулу нуклеиновой кислоты согласно настоящему изобретению, функционально связанную с одной или более последовательностью, контролирующей экспрессию. В настоящем документе фраза «рекомбинантная молекула" или "рекомбинантная молекула нуклеиновой кислоты" относится прежде всего к молекуле нуклеиновой кислоты или нуклеотидной последовательности, функционально связанной с последовательностью, контролирующей экспрессию, однако может использоваться взаимозаменяемо с фразой "молекула нуклеиновой кислоты", если такая молекула нуклеиновой кислоты представляет собой рекомбинантную молекулу, как описано в настоящем документе. В соответствии с настоящим изобретением, фраза «функционально связанный» относится к связыванию молекулы нуклеиновой кислоты с последовательностью, контролирующей экспрессию, так что молекула может экспрессироваться при трансфекции (т.е. трансформации, трансдукции, трансфекции, конъюгации или введении) в клетку-хозяина. Последовательности, контролирующие транскрипцию, представляют собой последовательности, контролирующие экспрессию, которые контролируют инициацию, элонгацию или терминацию транскрипции. Особенно важными последовательностями, контролирующими транскрипцию, являются последовательности, контролирующие инициацию транскрипции, например, промоторные, энхансерные, операторные и репрессорные последовательности. Подходящие последовательности, контролирующие транскрипцию, включают любую последовательность, контролирующую транскрипцию, способную функционировать в клетке-хозяине или организме, в который следует ввести рекомбинантную молекулу нуклеиновой кислоты. Рекомбинантные молекулы нуклеиновой кислоты согласно настоящему изобретению также могут содержать дополнительные регуляторные последовательности, например, регуляторные последовательности трансляции, сайты начала репликации и другие регуляторные последовательности, совместимые с рекомбинантной клеткой. В одном варианте реализации рекомбинантная молекула согласно настоящему изобретению, в том числе молекула, встроенная в хромосому клетки-хозяина, также содержит сигналы секреции (т.е. нуклеотидные последовательности сигнального сегмента или сигнальной последовательности), обеспечивающие секрецию экспрессируемого белка из клетки, продуцирующей белок. Подходящие сигнальные сегменты включают сигнальный сегмент, связанный с экспрессируемым белком в природных условиях, или любой гетерологичный сигнальный сегмент, способный управлять секрецией белка в соответствии с настоящим изобретением. В еще одном варианте реализации рекомбинантная молекула согласно настоящему изобретению содержит лидерную последовательность обеспечивающую доставку и внедрение экспрессированного белка в мембрану клетки-хозяина. Другие сигнальные последовательности включают последовательности, способне управлять периплазматической или внеклеточной секрецией, или удерживанием в заданных компартментах. Подходящие лидерные последовательности включают лидерную последовательность, связанную с белком в природных условиях, или любую гетерологичную лидерную последовательность, способную управлять доставкой и внедрением белка в мембрану клетки.
Согласно настоящему изобретению, термин "трансфекция" используют по отношению к любому способу, с помощью которого экзогенную молекулу нуклеиновой кислоты (т.е. рекомбинантную молекулу нуклеиновой кислоты) можно ввести в микробную клетку. Термин "трансформация" можно использовать взаимозаменяемо с термином "трансфекция", если этот термин используется для обозначения введения молекул нуклеиновой кислоты в клетки микроорганизмов или растений. В микробных системах термин "трансформация" используется для описания наследуемых изменений, обусловленных поглощением нуклеиновой кислоты микроорганизмом, и фактически является синонимом термина "трансфекция". Тем не менее, в клетках животных термин "трансформация" приобрел второе значение, которое может относиться к изменению свойств роста клеток в культуре (описанному выше), например, после которого они становятся злокачественными. Таким образом, чтобы избежать путаницы, термин "трансфекция" предпочтительно используют в связи с введением экзогенных нуклеиновых кислот в клетки животных и используют в настоящем документе для общего обозначения трансфекции клеток животных и трансформации растительных клеток и микробных клеток в той степени, что эти термины относятся к введению экзогенных нуклеиновых кислот в клетку. Таким образом, методики трансфекции включают трансформацию, бомбардировку частицами, электропорацию, микроинъекцию, липофекцию, адсорбцию, инфекцию и слияние протопластов, но не ограничиваются ими.
В одном варианте реализации композицию, содержащую белок или пептид, содержащий моноцистеиновый активный центр тиоредоксина в восстановленном состоянии, применяют для снижения вязкости чрезмерно вязкой слизи или мокроты. Указанная композиция содержит белок, содержащий моноцистеиновый активный центр тиоредоксина, и может содержать один или более дополнительных агентов или соединений, например, другие агенты или соединения, которые можно применять для снижения/уменьшения чрезмерно вязкой или липкой слизи или мокроты или повышения разжижения такой слизи или мокроты. Примеры таких других агентов или соединений известны в данной области техники и включают очищенную рекомбинантную ДНКазу человека, N-ацетилцистеин, nacystelyn (производное N-ацетил-L-цистеина), GSH и гельсолин, но не ограничиваются ими. Кроме того, в комбинации с моноцистеиновым активным центром тиоредоксина можно применять такие мукоактивные вещества, как маннит или гипертонический раствор NaCl.
В одном варианте реализации композицию, в том числе фармацевтическую композицию, можно применять для доставки молекулы нуклеиновой кислоты, кодирующей белок или пептид, содержащий моноцистеиновый активный центр тиоредоксина, в клетку пациента, подлежащего лечению (например, эпителиальную клетку легкого или дыхательных путей), так что клетка может стать трансфицированной белком и экспрессировать его, и таким образом, что белок может вступать в контакт со слизью или мокротой в микроокружении клетки.
Композиция, в том числе фармацевтическая композиция, также может включать, например, фармацевтически приемлемый носитель, который включает фармацевтически приемлемые вспомогательные вещества и/или носители для доставки белка или молекулы нуклеиновой кислоты или другого регуляторного соединения пациенту. Кроме того, композицию, в том числе фармацевтическую композицию согласно настоящему изобретению, можно вводить пациенту с фармацевтически приемлемым носителем. В настоящем документе фармацевтически приемлемый носитель относится к любому веществу, подходящему для доставки терапевтического белка, нуклеиновой кислоты или другого соединения, которое можно применять в способе согласно настоящему изобретению в подходящую область in vivo или ex vivo. Предпочтительные фармацевтически приемлемые носители могут поддерживать белок, молекулу нуклеиновой кислоты или соединение в форме, которая при поступлении белка, молекулы нуклеиновой кислоты или соединения в желательную область (например, область, где выделяется или куда стекает слизь или мокрота, подлежащая обработке), способна вступать в контакт со слизью или мокротой (в случае белка или соединение) или проникать в клетку, экспрессироваться в клетке и секретироваться (в случае молекулы нуклеиновой кислоты), так что экспрессированный белок в восстановленном состоянии может вступать в контакт со слизью или мокротой. Подходящие вспомогательные вещества согласно настоящему изобретению включают вспомогательные вещества или компоненты состава, осуществляющие или способствующие транспорту, но не направляющие терапевтический агент (белок, нуклеиновую кислоту или соединение) в клетку, ткань или жидкость (слизь и мокроту) специфическим образом (в настоящем документе такие вещества также называют ненаправленными носителями). Примеры фармацевтически приемлемых вспомогательных веществ включают воду, физиологический раствор с фосфатным буфером, раствор Рингера, раствор декстрозы, растворы, содержащие сыворотку, раствор Хэнка, другие водные физиологически сбалансированные растворы, масла, эфиры и гликоли, но не ограничиваются ими. Водные носители могкт содержать подходящие вспомогательные вещества, необходимые для приближения к физиологическим условиям в организме реципиента, например, путем повышения химической стабильности и изотоничности. Составы для ингаляции терапевтических агентов могут также включать молекулы ПАВ.
Подходящие вспомогательные вещества включают, например, ацетат натрия, хлорид натрия, лактат натрия, хлорид калия, хлорид кальция и другие вещества, применяемые для получения фосфатного буфера, трис-буфера и бикарбонатного буфера. Вспомогательные вещества также могут включать консерванты, например, тимеросал, m- или о-крезол, формалин и бензиловый спирт. Композиции согласно настоящему изобретению можно стерилизовать с помощью общепринятых способов и/или лиофилизировать.
Один из видов фармацевтически приемлемого носителя включает состав с контролируемым высвобождением, способный медленно высвобождать композицию согласно настоящему изобретению в организм пациента. В настоящем документе состав с контролируемым высвобождением включает один или более терапевтических агентов согласно настоящему изобретению в носителе с контролируемым высвобождением. Подходящие носители с контролируемым высвобождением включают биосовместимые полимеры, другие полимерные связующие материалы, капсулы, микрокапсулы, микрочастицы, болюсные составы, осмотические насосы, диффузионные устройства, липосомы, липосферы и системы трансдермальной доставки, но не ограничиваются ими. Такие носители с контролируемым высвобождением также могут содержать восстановители для поддержания моноцистеинового активного центра тиоредоксина в восстановленном состоянии во время хранения и доставки. Подходящие носители для доставки нуклеиновых кислот включают липосомы, вирусные векторы или другие носители для доставки, в том числе рибозимы, но не ограничиваются ими.
Подходящее, или эффективное при введении пациенту количество белка или пептида, содержащего моноцистеиновый активный центр тиоредоксина, представляет собой количество, которое может: участвовать в окислительно-восстановительных реакциях путем обратимого окисления тиола его активного центра с дисульфидом, катализировать реакции тиол-дисульфидного обмена, и, в частности, снижать вязкость или липкость слизи или мокроты и/или повышать разжижение слизи или мокроты у пациента в достаточной степени, чтобы обеспечить терапевтический благоприятный эффект для пациента. Уменьшение вязкости или липкости или повышение разжижения слизи или мокроты можно измерить, обнаружить или определить, как описано выше, или с помощью любого подходящего способа, известного специалистам в данной области техники. Как обсуждалось выше, такие измерения включают определение и сравнение процента свободных тиолов в образце слизи или мокроты пациента до и после контакта с подходящим или эффективным количеством белка или пептида, содержащего моноцистеиновый активный центр тиоредоксина, а также определение и сравнение уровня ОФВ у пациента до и после контакта с подходящим или эффективным количеством белка или пептида, содержащего моноцистеиновый активный центр тиоредоксина в восстановленном состоянии.
В одном варианте реализации подходящее или эффективное количество белка или пептида, содержащего моноцистеиновый активный центр тиоредоксина, подлежащее введению пациенту, содержит приблизительно 10 мкмоль/кг, 15 мкмоль/кг, 20 мкмоль/кг, 25 мкмоль/кг, 30 мкмоль/кг, 35 мкмоль/кг, 40 мкмоль/кг, 45 мкмоль/кг, 50 мкмоль/кг, 55 мкмоль/кг, 60 мкмоль/кг, 65 мкмоль/кг, 70 мкмоль/кг, 75 мкмоль/кг, 80 мкмоль/кг, 85 мкмоль/кг, 90 мкмоль/кг, 95 мкмоль/кг, 100 мкмоль/кг, 105 мкмоль/кг, 110 мкмоль/кг, 115 мкмоль/кг, 120 мкмоль/кг, 125 мкмоль/кг, 130 мкмоль/кг, 135 мкмоль/кг, 140 мкмоль/кг, 145 мкмоль/кг, 150 мкмоль/кг, 175 мкмоль/кг, 200 мкмоль/кг, 225 мкмоль/кг, 250 мкмоль/кг, 275 мкмоль/кг, 300 мкмоль/кг, 325 мкмоль/кг, 350 мкмоль/кг, 375 мкмоль/кг, 400 мкмоль/кг, 425 мкмоль/кг, 450 мкмоль/кг, 475 мкмоль/кг, 500 мкмоль/кг, 525 мкмоль/кг, 550 мкмоль/кг, 575 мкмоль/кг, 600 мкмоль/кг, 625 мкмоль/кг, 650 мкмоль/кг, 675 мкмоль/кг, 700 мкмоль/кг, 725 мкмоль/кг, 750 мкмоль/кг, 775 мкмоль/кг, 800 мкмоль/кг, 825 мкмоль/кг, 850 мкмоль/кг, 875 мкмоль/кг, 900 мкмоль/кг, 925 мкмоль/кг, 950 мкмоль/кг, 975 мкмоль/кг, 1000 мкмоль/кг, 1100 мкмоль/кг, 1200 мкмоль/кг, 1300 мкмоль/кг, 1400 мкмоль/кг, 1500 мкмоль/кг, 1600 мкмоль/кг, 1700 мкмоль/кг, 1800 мкмоль/кг, 1900 мкмоль/кг, 2000 мкмоль/кг, 2100 мкмоль/кг, 2200 мкмоль/кг, 2300 мкмоль/кг, 2400 мкмоль/кг или приблизительно 2500 мкмоль/кг массы тела пациента.
В еще одном варианте реализации, если путь доставки представляет собой доставку аэрозоля в легкие или аналогичный путь, количество белка или пептида, содержащего моноцистеиновый активный центр тиоредоксина, которое следует ввести пациенту, содержит от приблизительно 0,25 мг на дозированную единицу (например, дозированная единица для человека, как правило, составляет приблизительно 2-3 мл) до приблизительно 100 мг на дозированную единицу. Предпочтительно, количество белка или пептида, содержащего моноцистеиновый активный центр тиоредоксина, которое следует ввести пациенту, содержит приблизительно 0,25 мг, 0,50 мг, 1,0 мг, 5,0 мг, 10 мг, 15 мг, 20 мг, 25 мг, 30 мг, 35 мг, 40 мг, 45 мг, 50 мг, 55 мг, 60 мг, 65 мг, 70 мг, 75 мг, 80 мг, 85 мг, 90 мг, 95 мг или приблизительно 100 мг в на дозированную единицу. В зависимости от устройства, используемого для доставки аэрозоля, некоторые устройства для доставки аэрозоля обеспечивают фактическую доставку в легкие лишь приблизительно 10% от объема аэрозоля. В то же время, если устройство для доставки представляет собой ингалятор с вибрирующей сеткой, возможна доставка приблизительно 90% от объема аэрозоля. Электронные ингаляторы с вибрирующей сеткой могут доставлять лекарственные средства гораздо быстрее и представляют собой более малогабаритные устройства, которые значительно более предпочтительны для пациентов с MB (Geller, D.E., Pediatric Pulmonology, 43(S9):S5-S17, 2008). Ингаляторы с вибрирующей сеткой также более эффективно доставляют лекарственные средства при меньшей остаточной дозе, чем воздухоструйные ингаляторы. Это особенно важно для снижения расходов на лечение, поскольку для достижения терапевтически благоприятного эффекта требуются меньшие дозы. Такие устройства также не приводят к снижению биологической активности белков (Kesser, K.С., et al. Resp Care, 54(6):754-768, 2009; Scherer, Т., et al. J Pharm Sci, 100(1):98-109, 2011). Таким образом, для других путей введения, обеспечивающих больший объем композиции, доставляемый к желательной области, легко видеть, что можно использовать более низкие дозы белка или пептида, содержащего активный центр тиоредоксина.
Оптимальное количество белка согласно настоящему изобретению, подлежащего введению в организм животного, зависит от пути введения. Например, если белок вводят путем ингаляции (в виде аэрозоля), оптимальное вводимое количество может отличаться от оптимального количества, вводимого путем интратрахеального микрораспыления. Специалист в данной области техники может менять это количество в зависимости от такого пути введения. Важно отметить, что подходящее количество белка согласно настоящему изобретению представляет собой количество, обладающее желательной функцией, не являясь токсичным для животного. Другие пути введения включают пероральное введение, особенно для обработки слизи пищеварительной системы, или местное применение для обработки слизи половой системы, но не ограничиваются ими.
В одном варианте реализации настоящего изобретения в композицию, в том числе фармацевтическую композицию согласно настоящему изобретению, содержащую белок, содержащий моноцистеиновый активный центр тиоредоксина, дополнительно вносят один или более агентов, поддерживающих активный центр тиоредоксина в восстановленном состоянии после первоначального восстановления с помощью восстановителей. Такие восстановители, применяемые в настоящем изобретении, включают дитиотреитол (ДТТ), липоевую кислоту, NADH- или NADPH-зависимую тиоредоксинредуктазу, этилендиаминтетрауксусную кислоту (ЭДТА), восстановленный глутатион, дитиогликолевую кислоту, 2-меркаптоэтанол, трис-(2-карбоксиэтил)фосфен, N-ацетилцистеин, NADPH, NADH и другие биологические или химические восстановители, но не ограничиваются ими.
Как обсуждалось выше, композицию, в том числе фармацевтическую композицию согласно настоящему изобретению вводят в организм пациента способом, обеспечивающим эффективную доставку композиции и, в частности, белка, содержащего моноцистеиновый активный центр тиоредоксина, и/или любых других соединений композиции, к области-мишени (например, слизи или мокроте, подлежащей обработке, для белков и соединений, клетке-мишени, которая должна находиться или находится в окружении слизи и мокроты, подлежащих обработке, для рекомбинатных молекул нуклеиновых кислот). Подходящие протоколы введения включают любой протокол введения in vivo или ex vivo.
В соответствии с настоящим изобретением, эффективный протокол введения (т.е. введения композиции согласно настоящему изобретению эффективным образом) содержит подходящие параметры дозы и пути введения, которые приводят к контакту белка, содержащего моноцистеиновый активный центр тиоредоксина, и/или других соединений композиции со слизью или мокротой, подлежащей обработке, предпочтительно таким образом, что пациент получает некоторый измеримый, наблюдаемый или предполагаемый благоприятный эффект от такого введения. В некоторых ситуациях можно определить параметры эффективной дозы путем пробоотбора слизи и мокроты у пациента, с использованием способов оценки вязкости или разжижения слизи или мокроты, описанных в настоящем документе. В качестве альтернативы, параметры эффективной дозы можно определить экспериментально с использованием образцов in vitro, животных моделей in vivo и, в конечном итоге, клинических исследований, если пациент является человеком. Параметры эффективной дозы можно определить с использованием способов, стандартно применяемых в данной области техники при конкретном заболевании или состоянии. Такие способы включают, например, определение выживаемости, побочных эффектов (например, токсичности) и прогрессирования или регресса заболевания, а также соответствующих физиологических параметров, например, объема форсированного выдоха за секунду (ОФВ1).
Согласно настоящему изобретению, подходящие методы введения композиции согласно настоящему изобретению в организм пациента включают любой путь введения in vivo, подходящий для доставки композиции к желательной области в организме пациента. Предпочтительные пути введения очевидны для специалистов в данной области техники в зависимости от того, является ли соединение белком или другим соединением (например, лекарственным веществом), в какую часть тела следует вводить композицию и заболевания или состояния, которые испытывает пациент. В общем случае, подходящие способы введения моноцистеинового активного центра тиоредоксина включают дермальную доставку, интратрахеальное введение, ингаляцию (например, аэрозоля), назальное, пероральное, легочное введение и пропитку катетера, но не ограничиваются ими. Ушная доставка может включать ушные капли, интраназальная доставка может включать назальные капли или интраназальную инъекцию, а внутриглазная доставка может включать глазные капли или применением подходящих устройств для проникновения лекарственного средства через склеры. Аэрозольную (ингаляционную) доставку также можно выполнить с помощью способов, стандартно применяемых в данной области техники (см., например, Stribling et al., Proc. Natl. Acad. Sci. USA 189:11277-11281, 1992, полностью включенную в настоящий документ посредством ссылки). Пероральная доставка может включать твердые вещества и жидкости, которые можно принимать внутрь, например, в виде таблеток или капсул, а также в составе пищевых продуктов и напитков или корма для животных или кормовых гранул. Другие пути введения, подходящие для слизистых тканей, включают бронхиальный, интраназальный, другие ингаляционные, ректальный, местный, трансдермальный, вагинальный, трансцервикальный, перицервикальный и уретральный пути. Кроме того, протоколы введения могут включать устройства для предварительной обработки, например, нанесения белка, пептида или композиции на противозачаточный колпачок (например, на шейку матки) для таких вариантов применения, как бесплодие. В предпочтительном варианте реализации настоящего изобретения, если белок или композицию согласно изобретению вводят для лечения чрезмерно или аномально вязкой или липкой мокроты или слизи в дыхательную систему (дыхательные пути), белок или пептид (или композицию), содержащие моноцистеиновый активный центр тиоредоксина, или другие соединения вводят путем, включающим ингаляцию (т.е. путем ингаляции аэрозоля, например, в ПАВ или в комбинации с ПАВ); прямое введение в легкие посредством бронхоскопа, эндотрахеальной трубки и/или любого устройства искусственной вентиляции; назальне введение (интраназально или трансназально), бронхиальное или интратрахеальное введение (т.е. путем инъекции непосредственно в трахею или трахеостомии), непосредственно или посредством капсуляции в липид или ПАВ, но не ограничивающимся ими. Любые возможные способы введения композиции или белка в дыхательных путях так, что они могут войти в контакт со слизью или мокротой, содержащимися в них, входят в рамки настоящего изобретения.
В способах согласно настоящему изобретению композиции, в том числе фармацевтические композиции можно вводить любому члену класса позвоночных, включая, без ограничения, приматов, грызунов, продуктивных домашних животных, кур, индеек и домашних животных. Предпочтительными для защиты пациентами являются люди.
Еще один вариант реализации настоящего изобретения относится к композиции, содержащей белок или пептид, содержащий моноцистеиновый активный центр тиоредоксина, причем моноцистеиновый активный центр тиоредоксина ковалентно связан с остатком цистеина белка слизи. Белки слизи включают белки слизи дыхательных путей и пищеварительного тракта, но не ограничиваются ими. Белки слизи включают муцины, например, муцины MUC5AC и MUC5Bc большим содержанием дисульфидных связей. Например, после первоначальной каталитической реакции N-концевого остатка цистеина активного центра тиоредоксина в реакции тиолового обмена с дисульфидными связями белка слизи, белок или пептид, содержащий моноцистеиновую кислоту тиоредоксина, может ковалентно связаться через N-концевой остаток цистеина с белком слизи и, таким образом, не поддаваться неоднократному восстановлению и катализу. Ковалентная связь тиоредоксина, содержащего моноцистеиновый активный центр, с его мишенью - муцином блокирует поглощение и интернализацию тиоредоксина клетками за счет секвестрации слизи, что обеспечивает двойное преимущество за счет предотвращения поглощения эпителием и неспецифических эффектов, вызванных нежелательной активностью тиоредоксина внутри клеток, и в то же время облегчает выведение связанного со слизью отработанного лекарственного вещества из организма.
Дополнительный вариант реализации настоящего изобретения относится к способу снижения вязкости слизи и мокроты у пациента с чрезмерно вязкой или липкой слизью или мокротой путем введения композиции, содержащей белок или пептид, содержащие моноцистеиновый активный центр тиоредоксина, где белок ковалентно связан с остатком цистеина в белке слизи. В одном аспекте белок слизи представляет собой муцин. В еще одном аспекте моноцистеиновый активный центр тиоредоксина находится в восстановленном состоянии. В еще одном аспекте белок слизи может представлять собой белок слизи дыхательных путей или белок слизи пищеварительного тракта или белок слизи половой системы.
Еще один вариант реализации настоящего изобретения относится к способу снижения вязкости слизи и мокроты у пациента с чрезмерно вязкой или липкой слизью или мокротой путем приведения слизи или мокроты пациента в контакт с композицией, содержащей восстановитель дисульфидных связей и агент, блокирующий цистеин. Восстановитель дисульфидных связей и агент, блокирующий цистеин могут быть одним и тем же еществом, включая белок или пептид, содержащие моноцистеиновый активный центр тиоредоксина, но не ограничиваясь ими, или могут являться различными веществами. Восстановитель дисульфидных связей может представлять собой дитиотреитол (ДТТ), этилендиаминтетрауксусную кислоту (ЭДТА), GSH, дитиогликолевую кислоту, 2-меркаптоэтанол, N-ацетилцистеин, трис-(2-карбоксиэтил)фосфин или другие фармацевтически совместимые восстановители, известные в данной области техники. Агент, блокирующий цистеин, может представлять собой иодацетамид, иодуксусную кислоту или другие алкилирующие агенты, или антитело, специфичное по отношению к цистеину, или другую обладающую сродством белковую или пептидную композицию или имитатор антитела. Агент, блокирующий цистеин, связывается с остатком цистеина, входившим в состав дисульфидной связи в белке слизи, до восстановления связи. Агент, блокирующий цистеин, предотвращает повторное образование дисульфидной связи на основе тиоловой группы остатка цистеина слизи.
Еще один вариант реализации настоящего изобретения относится к способу лечения пациента с чрезмерно вязкой или липкой слизью путем введения пациенту композиции, содержащей по меньшей мере одно соединение, содержащее активный центр тиоредоксина, неспособный к поглощению клетками. Указанное соединение может являться белком или пептидом, содержащим моноцистеиновый активный центр тиоредоксина. Указанное соединение также может являться гибридным белком, содержащим фрагмент тиоредоксина и фрагмент лиганда рецептора клеточной поверхности. В этом варианте реализации фрагмент лиганда рецептора клеточной поверхности связывается с рецептором клеточной поверхности, тем самым предотвращая поглощение гибридного белка клеткой. Указанное соединение может являться комбинацией белка или пептида, содержащего активный центр тиоредоксина, и соединения, блокирующего цистеин, соответствующий остатку цистеина в положении 35 SEQ ID NO: 12. В предпочтительном варианте реализации блокирующее соединение может являться антителом или имитатором антитела, связывающимся с молекулой тиоредоксина и таким образом блокирующим остаток цистеина в положении 35 SEQ ID NO: 12. В этой связи термин «блокировать» относится к созданию помех способности остатка цистеина в положении 35 SEQ ID NO: 12, например, образовывать внутримолекулярную дисульфидную связь с остатком цистеина в положении 32 SEQ ID NO: 12.
Дополнительный вариант реализации настоящего изобретения относится к способу предотвращения системного воздействия лекарственного вещества на пациента. Этот способ включает этап введения лекарственного вещества пациенту посредством пути доставки, включая пульмонарный, пероральный или местный пути доставки, но не ограничиваясь ими. Лекарственное вещество после введения может образовывать ковалентную связь с областью-мишенью. Этот механизм действия отличается от известных механизмов действия лекарственных веществ, поскольку многие лекарственные вещества действуют за счет молекулярных взаимодействий, при которых лиганды обратимо связываются с рецепторами. В предпочтительном варианте реализации лекарственное вещество является тиолсодержащим лекарственным веществом, в котором тиоловая группа образуеют ковалентную связь с другой тиоловой группой в области-мишени. Например, лекарственное средство может содержать белок или пептид, содержащий моноцистеиновый активный центр тиоредоксина в восстановленном состоянии. В еще одном предпочтительном варианте реализации область-мишень является внеклеточной, и лекарственное средство вводят посредством пути для внеклеточной доставки.
Еще один вариант реализации настоящего изобретения относится к фармацевтической композиции, содержащей белок или пептид, содержащий моноцистеиновый активный центр тиоредоксина в восстановленном состоянии, и дополнительно содержащей по меньшей мере один сахарид или производное сахарида, способное стабилизировать редокс-активную тиоловую группу. Сахарид или производное сахарида может являться сахарозой, сукралозой, лактозой, трегалозой, мальтозой, галактозой, рафинозой, маннозой или маннитом. Под редокс-активной тиоловой группой понимают тиоловую группу, которая может существовать в восстановленном (-SH) или окисленном состоянии (-S-S-). Термин «стабилизирующий» включает, например, снижение скорости окисления редокс-активной тиоловой группы в восстановленном состоянии, когда полипептид присутствует в фармацевтической композиции, содержащей сахарид или производное сахарида, по сравнению с композицией, в которой сахарид или производное сахарида отсутствует. Под «сахаридом» понимают любой моно-, ди-, олиго- или полисахарид. Примерами сахаридов являются глюкоза, фруктоза, сахароза, лактоза, мальтоза, галактоза, раффиноза, инулин, декстран, трегалоза, сукралоза, манноза и маннит. Под производным сахарида понимают соединение, структурно напоминающее сахарид, производным которого оно является. Например, сукралозу, которая является хлорированной сахарозой, следует считать производным сахарида сахарозы. Дополнительные производные включают, например, производные альдита, например, маннит и ксилит. Предпочтительные композиции согласно настоящему изобретению содержат невосстанавливающие сахариды, например, раффинозу, трегалозу, стахиозу и, в особенности, сахарозу.
Еще один вариант реализации настоящего изобретения относится к кормовой композиции, содержащей белок или пептид, содержащий моноцистеиновый активный центр тиоредоксина в восстановленном состоянии. Примеры кормов для животных включают сено, солому, силос, сжатые и гранулированные корма, масла и смешанные рационы, проросшие зерна, бобовые, растительные остатки, зерно, зерновые культуры и кукурузу, но не ограничиваются ими.
Следующие примеры приведены исключительно для иллюстративных целей и не предназначены для ограничения рамок настоящего изобретения.
Примеры
Пример 1
В данном примере показана экспрессия и очистка белков Trx дикого типа (также называемого в настоящем документе «rhTxr»), и rhTrx с моноцистеиновым активным центром (также называемого в настоящем документе «r(Cys)hTrx"). Уровнях экспрессии белка и правильности посттранскрипционной экспрессии rhTrx и r(Cys)hTrx максимизировали путем кодонной оптимизации последовательности ДНК Trx-1 человека, как описано в работе Harris et al., Biotechnol Biogen 109:1987-97 (2012), полностью включенной в настоящий документ посредством ссылки. Выполняли клонирование синтетических конструктов в подходящих векторах для продукции в Е. coli, что обеспечило продукцию белка дикого типа и мутантного белка с моноцистеиновым активным центром r(Cys)hTrx в микрограммовом масштабе. В лабораторном масштабе выполнили оценку нескольких маркеров экспрессии и стратегий очистки, а также аффинного расщепления с целью облегчения очистки от эндотоксинов и эндогенных тиоредоксинов хозяина.
Получение rhTrx и r(Cys)hTrx в Е. coli. Ген rhTrx, кодирующий зрелый белок rhTrx размером 105 аминокислот, включая начальный остаток метионина, оптимизировали для экспрессии Е. coli и синтезировали (DNA2.0, Inc.). Ген субклонировали в векторе pEV, содержавшем индуцибельный промотор Т7, маркер устойчивости к канамицину, маркер из 6 остатков гистидина для аффинной очистки на Ni и сайт расщепления протеазой вируса гравировки табака (TEV). Для экспрессии rhTrx плазмиду rhTrx pEV (подтвержденную путем секвенирования) трансформировали в клетки Е. coli С43 (Lucigen), выращенные до оптической плотности ~0,6 (600 нм), и индуцировали изопропил-β-D-1-тиогалактопиранозида (ИПТГ). Белок выделяли из клеток Е. coli путем гомогенизации и лизиса с использованием детергента с последующим центрифугированием. RhTrx очищали от лизата Е. coli с помощью Ni-аффинной хроматографии. RhTrx, аффинно очищенный на Ni, расщепляли протеазой (TEV) с целью удаления маркера сродства к никелю и дополнительно очищали с помощью ионообменной хроматографии и обращенно-фазной жидкостной хроматографии высокого давления (ВЭЖХ). Чистоту белка оценивали электрофорезом в ДСН-ППАГ и масс-спектроскопии с ионным распылением (ЖХ/МС/МС) и проверяли последовательность белка путем трипсинового гидролиза rhTrx с последующей масс-спектроскопией с ионной ловушкой. Концентрацию белка и эндотоксинов определяли анализом на основе бицинхониновой кислоты (Pierce) и анализом амебоцитов мечехвоста (Charles River Lab.), соответственно. Белок r(Cys)hTrx получали аналогично, но с использованием гена r(Cys)hTrx, оптимизированного и синтезированного в DNA2.0. Эти белки rhTrx и r(Cys)hTrx восстанавливали in vitro с помощью DTT, который впоследствии удаляли диализом и обработкой на обессоливающей колонке.
Пример 2
В данном примере описана предпочтительная стратегия экспрессии, очистки и восстановления (активации) нативного белка rhTrx и белка rhTrx с моноцистеиновым активным центром (r(Cys)hTrx), использующая прямую экспрессию зрелого белка, а не подход на основе аффинного связывания. Белок r(Cys)hTrx и нативный контрольный белок Trx продуцировали в периодических культурах в колбах на качалке и в периодической культуре с добавлением субстрата в Е. coli BL21 и клетках-хозяевах, происходящих от штамма K12, используя стандартные способы культивирования бактерий и ряд распространенных конститутивных или индуцибельных промоторных систем; в то же время предпочтительными являлись углевод-индуцибельные или ИПТГ-индуцибельные системы, поскольку они позволяли получить штаммы с максимальной экспрессией. После разрушения клеток с помощью микрофлюидизатора супернатанты, содержащие водорастворимый Trx, собирали центрифугированием и обрабатывали сульфатом аммония (AS) для преимущественного осаждения белков клеток-хозяев. Полученные супернатанты, обогащенные Trx, фильтровали с помощью ультрафильтрации/диафильтрации (UF/DF) с заменой буфера на 15 мМ Hepes, 10 мМ бета-меркаптоэтанол (b-МЕ), 250 мМ NaCl. В данном буфере Trx не связывался со смолой для анионообменной хроматографии (АЕС) Q-Sepharose HP, тогда как ДНК и другие примеси связывались с ней с высоким сродством. После АЕС при высокой концентрации солей элюат, обогащенный TRx, обменивали на буфер с низким содержанием солей для второго этапа АЕС на колонке Hi-Trap Q-HP (GE Healthcare) в восстанавливающих условиях (10 мМ ДТТ) с последующей заменой буфера на буфер для состава (описанный ниже). В качестве альтернативы, после AS-осаждения растворимую фракцию фильтровали через фильтр с пороговым значением 30 кДа и загружали на колонку для гидрофобной хроматографии (HIC). Элюированный материал концентрировали и подвергали замене буфера с использованием кассеты для UF/DF 1 кДа и загружали на колонку с Source Q (анионообменную). Белки тиоредоксина элюировали с ионообменной колонки, используя градиент соли, а затем подвергали дополнительному этапу UF/DF для замены на буфер для хранения.
Количественное определение эндотоксинов выполняли с помощью набора для анализа на основе лизатов амебоцитов мечехвоста Limulus Amoebacyte Lysate kit (LAL, Pierce) и удаляли остаточные эндотоксины с помощью 0,2-мкм фильтра Mustang E-filter (Pall). Идентичность и выход белка определяли электрофорезом в ДСН-ПААГ и вестерн-блоттингом/твердофазным ИФА на соответствующих этапах, общую массу и конечную идентичность последовательности подтверждали MALDI и масс-спектрометрией с электрораспылением. Восстанавливающую активность составов TRx количественно оценивали с помощью анализа восстановления DTNB (реагента Эллмана). Пятьдесят микролитров 2,5 мМ rhTrx или r(Cys)hTrx добавляли в 96-луночный планшет, а затем вносили 175 микролитров буфера для образцов и 25 микролитров 6 мМ DTNB (5,5'-дитиобис-(2-нитробензойной кислоты). Через 15 минут после инициации реакций путем добавления DTNB спектрофотометрически измеряли изменение поглощения при 412 нм вследствие восстановления DTNB при 30°С.
Ряд параметров буфера для хранения, например, параметры, обсуждаемые ниже, проверяли на совместимость с лиофилизацией и способность стабилизировать восстановленное состояние r(Cys)hTrx. Параметры буфера включают низкий уровень pH в присутствии восстанавливающих Сахаров, ацетат аммония pH 5,5 с 10 мМ b-МЕ, а также состав буфера, ранее повышавший стабильность хранения восстановленного нативного Trx (40 мМ ацетата Na pH 5,5, 0,05% ЭДТА, 9,25% сахарозы). Буфер с Trx лиофилизировали при каждом параметре, хранили в течение различного времени в высушенном виде и оценивали редокс-стабильность хранимых белков с помощью DTNB и анализов активности белка, как описано выше. Для оценки стабильности аэрозоля можно применять аэрозольный распылитель Perm-Century или ингалятор. Соединения аэрозоля собирали в 0,1 М трис-буфер pH 8,0 с 1 мМ ЭДТА, содержащий DTNB, при разном времени инкубации с целью определения стабильности аэрозоля hTrx r (KMC) и нативного rhTrx. Ожидали, что агрегаты не реагируют с DTNB и, следовательно, будет обнаруживаться только восстановленная форма. Коэффициент экстинкции DTNB (14150 при 412 нм) использовали для расчета свободных тиоловых (SH) групп в зависимости от концентрации восстановленного белка. В предыдущих исследованиях с интратрахеальной (ИТ) доставкой нативного rhTrx (Rancourt, R. et al., Free Radio Biol Med, 42(9): 1441-1453, 2007) использовали нормальный физиологический раствор. Можно использовать добавление редокс-стабилизаторов, заведомо совместимых с ингаляционной доставкой, например, метионина.
Пример 3
В данном примере продемонстрировано конструирование и штаммы Е. coli для экспрессии r(Cys)hTrx и нативного rhTrx. r(Cys)hTrx и нативный контрольный rhTrx легко экспрессировались в виде зрелых, растворимых белков с помощью различных векторных системам и лабораторных штаммов Е. coli BL21 или K12. Для получения штаммов Е. coli,, экспрессирующих Trx с высоким выходом (>1 г/л), синтезировали кодон-оптимизированные последовательности ДНК r(Cys)hTrx и нативного rhTrx и клонировали их в ряде экспрессирующих векторов с использованием различных внутриклеточных промоторных систем, включая системы, индуцируемые истощением питательных веществ, углевод-индуцибельные системы (например, промоторы, индуцируемые рамнозой, ксилозой, мелибиозой или другие индуцибельные промоторы, известные в данной области техники, например, в патенте США 7871815), или ИПТГ-индуцибельные системы на основе промоторов бактериофага Т3, Т5 или Т7. После подтверждения последовательности ДНК экспрессирующие конструкты трансформировали в различные часто используемые штаммы-носители Е. coli BL21/K12, включая рекомбинантные штаммы, дефектные по утилизации веществ, используемых для индукции, или штаммы с возможной дополнительной сверхэкспрессией метионинаминопептидазы для улучшения точности экспрессии высоких титров (Liu, М. et al., Protein Expr Purif 84(1):130-139, 2012). Полученные продуцирующие комбинации вектор/носитель выращивали в средне- или высокопроизводительных титрационных микропланшетах с глубокими лунками при встряхивании при 30-37°С в минимальной среде и подвергали скринингу на предмет экспрессии с помощью способов, обычно используемых в данной области техники, например, гель- или капиллярного электрофореза. Положительные клоны подтверждали посредством электрофореза в ДСН-ПААГ и вестерн-блоттинга и/или твердофазного ИФА/ELISPOT. Выполняли периодическое культивирование штаммов с добавлением субстрата и оценивали выход белка (r(Cys)hTrx) с помощью электрофореза в ДСН-ПААГ и вестерн-блоттинга или твердофазного ИФА/ELISPOT. Белки очищали и восстанавливали с помощью способов, описанных выше в примерах 1 и 2; восстановленное состояние активированного белка оценивали анализом DTNB, а удельную активность восстановления дисульфидной связи - анализом восстановления инсулина. Идентичность последовательности анализировали масс-спектрометрией (MALDI или МС с электрораспылением) с целью подтверждения правильности трансляции и надлежащей молекулярной массы.
Пример 4
В данном примере продемонстрирована активность in vitro и функции rhTrx дикого типа и rhTrx с моноцистеиновым активным центром. Активность rhTrx оценивали путем измерения его восстановительной активности при стандартном колориметрическом анализе DTNB и путем определения его способности ингибировать активность эластазы нейтрофилов. Кроме того, определяли способность r(Cys)hTrx разжижать слизь человека при MB по сравнению с rhTrx с помощью анализа уплотнения мокроты и, необязательно, путем проверки снижения вязкоупругости мокроты при MB с помощью специализированного реометра. rhTrx и r(Cys)hTrx сравнивали со стандартно применяемым при MB муколитическим средством пульмозим (рекомбинантная ДНКаза человека; альфа-дорназа).
Первоначальная оценка характеристик rhTrx: Восстановительная активность. Общую восстановительную активность заранее восстановленных составов Trx количественно оценивали с помощью анализа восстановления DTNB, как сообщалось ранее (Rancourt, R. et al., Free Radic Biol Med, 42(9):1441-1453, 2007). Для ферментативного восстановления rhTrx и r(Cys)hTrx, 50 микролитров буфера для анализа (100 мМ фосфата калия, pH 7,0, 10 мМ ЭДТА и 0,05 мг/мл бычьего сывороточного альбумина), содержащие 5 или 50 мМ rhTrx и 0,5 мМ очищенного TrxR добавляли в 96-луночный планшет. Добавляли следующие 175 микролитров буфера для образцов, содержащего 720 мМ NADPH, а затем 25 микролитров 6 мМ DTNB в буфере для образцов. Для измерения восстановительной активности предвариельно химически восстановленного rhTrx, 50 микролитров 2,5 мМ rhTrx добавляли в 96-луночный планшет, а затем вносили 175 микролитров буфера для образцов и 25 микролитров 6 мМ DTNB. После инициации реакций путем добавления DTNB выполняли спектрофотометрический мониторинг изменений кинетического поглощения при 412 нм вследствие восстановления DTNB при 30°С. С помощью аналогичного протокола r(Cys)hTrx подвергали данному анализу, а также анализам, описанным ниже. Общая восстановительная активность r(Cys)hTrx была меньше, чем у rhTrx, что было обусловлено меньшим количеством восстановленных остатков Cys в r(Cys)hTrx. В то же время оба белка восстанавливались до более чем 90-95% потенциального восстановленного состояния, что соответствовало полной активации.
Ингибирование активности эластазы нейтрофилов (NE). Система инкубирования для определения влияния rhTrx на активность NE была описана ранее (Lee, R., et al. (2005) Am J. Physiol. Lung Cell. Mol. Physiol. 289(5):L875-882). Вкратце, Trx разбавляли до желательной концентрации физиологическим раствором с фосфатным буфером (PBS, pH 7,2). Смесь добавляли к очищенной NE человека (100 мкг/мл в PBS, pH 7,2, 0,01% Тритон Х-100) и инкубировали при 37°С в течение 1 ч. Окончательный объем во время инкубирования составил 210 микролитров, а концентрация NE - 1,6 мкг/мл. После инкубирования выполняли анализ эластолитической активности в 60-микролитровых аликвотах в трех повторностях. Активность эластазы определяли путем добавления 120 микролитров 0,8 мМ N-метоксисукцинил-Ala-Ala-Pro-Val-4-нитроанилида в PBS (pH 7,2) к экспериментальному образцу и измерения кинетического поглощения при 405 нм при 37°С в течение 4 мин. Установлено, что концентрация эластазы, используемая в этих экспериментах, находилась в пределах линейного диапазона анализа. Процентное ингибирование активности NE определяли следующим образом: процентное ингибирование = (1 - изменение поглощения для экспериментальной группы/изменение поглощения для PBS-контрольной группы). Обнаружено, что эластазная активность r(Cys)hTrx была по меньшей мере сопоставима с активностью rhTrx.
Анализ уплотнения для оценки разжижения мокроты человека при MB. Все манипуляции с мокротой при MB выполняли в микробиологическом вытяжном шкафу. rhTrx, r(Cys)hTrx или только разбавитель (25 микролитров) в соответствующих концентрациях добавляли к мокроте при MB (275 микролитров) в 1,5-мл конических пробирках Eppendorf и перемешивали на вортексе в теечние очень короткого времени. После инкубирования при температуре 37 градусов С в течение 30 минут образцы повторно перемешивали в течение короткого времени, помещали в гематокритные пробирки и герметизировали с обоих концов. После 5-минутного центрифугирования в гематокритной центрифуге определяли процент сухого вещества (геля) и процент жидкости путем прямого линейного измерения и выражали в процентах жидкости следующим образом: 100 × жидкость/(жидкость + твердые вещества). Для каждого условия измеряли мокроту по меньшей мере пяти различных пациентов в MB в трех повторностях. Разжижающая способность r(Cys)hTrx была по меньшей мере сопоставима, если не значительно повышена по сравнению с rhTrx, что отражало неожиданный потенциал механизма действия r(Cys)hTrx.
Определение вязкоупругости мокроты. Эти измерения выполняли с помощью конического реометра, а также качественно путем прямого наблюдения потока мокроты после обработки тиоредоксином. Инкубирование выполняли с использованием протокола работы с мокротой, описанного выше. Вязкоупругость оценивали с помощью реометра AR-1000 (ТА Instruments, Нью-Касл, штат Делавэр, США) в режиме колебаний с угловой скоростью 1 рад/с при 37°С. Для каждого препарата Trx возможность снижения вязкоупругости мокроты оценивали в трех повторностях в мокроте, полученной от по меньшей мере 5 разных пациентов с MB. Ожидалось, что способность r(Cys)hTrx снижать вязкоупругость мокроты должна быть по меньшей мере сопоставима, если не значительно повышена по сравнению с rhTrx, что отражало неожиданный потенциал механизма действия r(Cys)hTrx. Измерение вязкоупругости мокроты выполняли путем наблюдения за скоростью потока мокроты, смешанной с rhTrx или r(Cys)hTrx в перевернутых пробирках Eppendorf; при этом анализе установлено, что поток r(Cys)hTrx был более выраженным, чем поток rhTrx, ДТТ и пульмозима, и значительно более выраженным, чем поток отрицательного контроля (носителя), для которого продемонстрировано практическое отсутствие потока мокроты.
Сравнение эффективности со стандартно применяемым муколитическим агентом (рекомбинантной ДНКазой человека, rhDNase). Для этих исследований сравнивали соответствующие концентрации rhTrx, r(Cys)hTrx и rhDNase путем инкубирования, как описано выше. Через 30 минут выполняли оценку образцов на предмет изменений степени разжижения мокроты (анализ уплотнения) и изменений вязкоупругости. Кроме того, выполнили исследования синергии. В них мокроту обрабатывали rhTrx в течение 30 мин, а затем rhDNase в течение 30 мин или, в качестве альтернативы, rhDNase в течение 30 мин, а затем rhTrx в течение 30 мин. Эти результаты сопоставляли с действием каждого агента по отдельности. Для каждого условия измеряли мокроту по меньшей мере пяти различных пациентов в MB в трех повторностях. От отдельных доноров в заданный день требовалось количество мокроты, достаточное для анализа каждого из 12 образцов (rhTrx × 3, rhDNase × 3, rhTrx + rhDNase × 3, rhDNase + rhTrx × 3) для rhTrx дикого типа и мутантного варианта rhTrx. Фактически оно составляло как минимум 4,5-5,0 мл перемешанной мокроты от одного донора/день. Разжижающая способность r(Cys)hTrx была по меньшей мере сопоставима, если не значительно повышена по сравнению с положительными контролями rhDNase, rhDNase, rhTrx или DTT.
Сбор мокроты человека при MB. Мокроту получали от взрослых пациентов и пациентов-детей с MB, согласно диагнозу лечащего врача. У пациентов диагностировали MB, если они демонстрировали клинические симптомы и содержание хлорида в поте свыше 60 миллимоль при двух отдельных анализах пота посредством ионофореза с пилокарпином, а также две аллельные мутации, приводящие к MB, при последующем генетическом анализе. Все образцы получали при спонтанном откашливании или путем индукции с помощью гипертонического раствора NaCl. Образцы мокроты, содержащие визуально обнаруживаемую слюну, удаляли. Мокроту хранили на льду до доставки в лабораторию, затем хранили при температуре -80 градусов С до использования во флаконах, герметизированных с помощью уплотнительного кольца, для предотвращения высыхания.
Пример 5
В данном примере продемонстрирована ферментативная (фигура 1b) и неферментативная (фигура 1а) активность белка или пептида, содержащего моноцистеиновый активный центр тиоредоксина, по сравнению с белком или пептидом, содержащим активный центр тиоредоксина дикого типа. Как показано на фигуре 1а, неспецифическое восстановление 5,5'-дитиобис-(2-нитробензойной кислоты) (DTNB или реагента Эллмана), отражающее потерю одного восстанавливаемого остатка цистеина в моноцистеиновом активном центре тиоредоксина по сравнению с активным центром дикого типа. Как показано на фигуре lb и фигуре 2, белок или пептид, содержащий моноцистеиновый активный центр тиоредоксина, как ни странно, обладал повышенной эффективностью по сравнению с белком или пептидом, содержащим активный центр тиоредоксина дикого типа, в анализе уплотнения мокроты человека, а также повышенной эффективностью по сравнению с эквимолярным количеством ДНКазы или NAC (фигура 2). Этот результат является неожиданным, учитывая значительные изменения в активном центре из-за мутации Cys35 и общую восстановительную способность, составлявшую 4/5 от способности нативного тиоредоксина за счет потери одного восстанавливаемого остатка Cys, в предыдущем анализе DTNB. Полагали, что неожиданное повышение эффективности моноцистеинового активного центра тиоредоксина обусловлено ковалентной связью с Cys-остатками белков муцина, что оказывало непредвиденные последствия, предотвращавшие повторное образование новых дисульфидных связей на основе этих остатков Cys.
Пример 6
В данном примере продемонстрирована пониженная склонность r(Cys)hTrx (в восстановленном, активном состоянии) стимулировать высвобождение провоспалительных цитокинов культивируемыми первичными эпителиальными клетками бронхов человека (НВЕ) по сравнению с нативным rhTrx. Донорских ткани и клетки предоставлены в рамках утвержденных протоколов по защите прав субъектов-людей. Клетки НВЕ обычных (т.е. здоровых) легких собирали посредством ферментативного гидролиза, как описано ранее (Fulcher, M.L., Methods Mol Med 107:183-206, 2005). Дезагрегированные клетки НВЕ высевали на прозрачную подложку Transwell Clear диаметром 12 мм (Corning) из расчета плотности 2,5×105/см2 в хорошо охарактеризованной среде для клеток дыхательных путей (Fulcher, M.L., 2005 ibid). Культуры поддерживали на границе раздела воздух-жидкость до полной дифференцировки (около 4-6 недель) перед использованием.
Протокол доставки тиоредоксина: Для этого исследования использовали устройство, способное доставлять нанолитровые объемы анализируемого агента (или контроля) на поверхность культур НВЕ. Эта система сконструирована как модельная система in vitro, имитирующая доставку мелкодисперсного аэрозоля распыленного лекарственного средства к поверхности эпителиальных клеток дыхательных путей in vivo, и представляет собой идеальный способ для изучения влияния добавляемых терапевтических средств, например, r(Cys)hTrx. На основании результатов моделирования множественной дозиметрии частиц (Anjilvel, S., et al. Fundam Appl Toxicol 28(2):41-50, 1995), демонстрирующих среднюю скорость осаждения для ингалятора Pari LC Star на протяжении первых 20 колен дыхательных путей, среднее осаждение на протяжении первых 6 колен дыхательных путей, куда доставляются большинство частиц, составляет ~50 нл/мин/см2 в течение 15 минут. В этих исследованиях выполняли распыление общего объема 750 нл (в течение 15 минут) в общей сложности в пяти различных экспериментальных группах. Группы включали: 1) контрольный носитель (изотонический раствор NaCl), 2) нативный рекомбинантный Trx (500 мкм), и 3) три дозы r(Cys)hTrx (10 мкм, 250 мкм и 1000 мкм). Эти концентрации представляли "окончательную" поверхностную концентрацию в дыхательных путях. После распыления каждого тестируемого реагента (или контрольного носителя) культуры возвращали в инкубатор для тканевых культур и инкубировали в течение 24 часов перед анализом цитокинов.
Иммуноанализ цитокинов: В данном исследовании оценивали влияние нативного и r(Cys)hTrx) на стимуляцию четырех основных цитокинов, высвобождаемых из клеток НВЕ, в том числе ИЛ-6, ИЛ-8, ФНО-α и ИЛ-1β. Цитокины в бесклеточных неконцентрированных супернатантых культур оценивали с использованием доступных для приобретения наборов для иммуноферментного анализа (твердофазного ИФА) (R&D Systems). Образцы базолатеральной среды ALI (Fulcher, M.L., 2005, ibid) получали через 24 ч после распыления во всех пяти группах и замораживали до анализа. В качестве фонового контроля из всех значений вычитали чистую (неиспользованную) среду ALI. Для каждого анализа цитокинов с целью получения соответствующих стандартных кривых использовали положительный контроль в диапазоне трех порядков концентрации. Кроме того, каждый "неизвестный" образец анализировали в двух повторностях. Измеряли следующие цитокины: ФНО-α (нижний предел обнаружения 0,3 пг/мл), ИЛ-6 (нижний предел обнаружения 0,35 пг/мл), ИЛ-8 (нижний предел обнаружения 2,4 пг/мл), и ИЛ-1β (нижний предел обнаружения 1,5 пг/мл). Ранее показано, что перекрестная реакционная способность этих анализов друг с другом и с другими рекомбинантными цитокинами человека (ИЛ-1а, ИЛ-2, ИЛ-3, ИЛ-4, ИЛ-7, фактор некроза опухоли-S, гранулоцитарный колониестимулирующий фактор и трансформирующий фактор роста-III) была ниже предела обнаружения.
Размер выборки и статистический анализ: Каждый из пяти вариантов условий оценивали с использованием в общей сложности девяти отдельных культур НВЕ. Этот набор представлял собой три культуры (n=3) от трех различных пациентов (n=3). Такой подход обеспечивал достаточную статистическую мощность для понимания изменчивости в пределах выборки и между выборками для каждого варианта условий. Для анализа данных выполняли сравнение отдельных образцов/групп с использованием двустороннего t-критерия Стьюдента. Для сравнения множественных групп использовали односторонний дисперсионный анализ (ANOVA). Значимость для всех анализов задали при р<0,05.
Оценка провоспалительного действия r(Cys)hTrx в культурах НВЕ доноров-пациентов с MB. Культура первичных эпителиальных клеток дыхательных путей человека при MB: Последние методики пролиферации и культивирования первичных эпителиальных клеток дыхательных путей человека на границе раздела воздух-жидкость с мукоцилиарной дифференциацией применяли на основе способов, представленных в работе Schlegel и соавторов (Liu, М. et al., Protein Expr Purif. 84(1): 130-139, 2012; Suprynowicz, F.A., et al. Proc. Natl. Acad. Sci. U.S.A. 109(49):20035-20040, 2012). Эти способы обеспечивали практически неограниченное количество первичных эпителиальных клеток дыхательных путей, сохранявших способность к конечной дифференцировки в среде ALI без генетических манипуляций. Клетки каждого из трех уникальных доноров с MB, гомозиготных по F508del, культивировали в 30 лунках в среде ALI до дифференцировки в течение 30 дней. Культуры на апикальной поверхности подвергали воздействию трех концентраций (10 мкМ, 250 мкМ и 1000 мкМ) rhTrx (дикого типа) и r(Cys)hTrx с последующим отбором образцов апикальной и базолатеральной среды через 4 и 24 часа после стимулирующего воздействия (см. таблицу 1). Использованная сыворотка не содержала сыворотки; ее центрифугировали для удаления остаточного материала и хранили при -80°С до использования. Твердофазный ИФА воспалительных цитокинов человека выполняли во всех образцах апикальной и базолатеральной среды. Отбор апикальной среды выполняли путем помещения 200 мкл стерильного PBS на апикальную поверхность и его сбора после 15-минутного инкубирования. Твердофазный ИФА ИЛ-8 и ИЛ-6 выполняли для каждого образца в двух повторностях (Becker, M.N. et al., Am J Resp Crit Care Med 169(5):645-653, 2004). Вышеописанный план с использованием 30 культур в среде ALI повторяли для каждого из трех доноров с MB. Сравнение результатов нативного rhTrx и r(Cys)hTrx продемонстрировало, что склонность тиоредоксина индуцировать высвобождение провоспалительных цитокинов из дифференцированных эпителиальных клеток дыхательных путей пациентов с MB ослаблялось для моноцистеинового Trx по сравнению с нативным тиоредоксином в протестированных концентрациях.

Пример 7
В данном примере продемонстрирована оценка биофизических и биохимических характеристик влияния r(Cys)hTrx по сравнению с нативным rhTrx на однородную слизь, собранную из культур клеток НВЕ in vitro.
Получение слизи из культур клеток: Эпителиальные клетки бронхов человека выращивали и поддерживали в соответствии с описанием (Matsui, Н., et al., J Clin Invest 102(6): 1125-1131, 1998). Вкратце, слизь, собранную с культур, собирали и хранили при 4°С. Образцы загружали в диализные трубки (MWCO = 3500) и концентрировали с полимерным абсорбентом (Spectra/Gel) в течение 1-5 дней при 4°С. Затем концентрированную слизь подвергали диализу против PBS, содержащего 500 мкМ MgCl2 и 800 μМ CaCl2 при 4°С, с целью установить надлежащий солевой баланс (Matsui, Н. et al., Proc Natl Acad Sci USA 103(48): 18131-18136, 2006).
Макроскопическая реология: Концентрирование и анализы по определению зависимости от времени выполняли с использованием реометра Bohlin Geminie в конусной и параллельной конфигурациях с целью оценки объемных, макроскопических биофизических эффектов восстановленного r(Cys)hTrx и нативного тиоредоксина на свойства слизи НВЕ in vitro. Выполняли эксперименты упругого восстановления после ползучести, в которых к обработанной или контрольной слизи прикладывали известную нагрузку (от 0,05 до ~100 Па) в течение 10 секунд и регистрировали реологическое восстановление жидкости в течение дополнительных 50 секунд. В последовательных экспериментах приложенную нагрузку увеличивали в логарифмической прогрессии до достижения предела текучести жидкости (т.е. нагрузки, при которой вязкость жидкости внезапно и резко уменьшается). На основании измеренных параметров установили, что вязкость и эластичность жидкости зависит от приложенной нагрузки. Развертку частоты выполняли при постоянных нагрузке и деформации и использовали для определения исходных физических свойств слизи (G' и G'', соответственно), а также вязкости и псевдопластичного поведения жидкости. Все эксперименты выполняли при 23°С.
Жидкостная хроматография высокого давления: Концентрацию и молекулярную массу муцина в обработанной и контрольной слизи оценивали с помощью дифференциальной рефрактометрии с целью определения влияния нативного тиоредоксина и r(Cys)hTrx на структуру муцина. Образцы (500 μл) загружали на колонку с сефарозой S1000 (Amersham Pharmacia) и элюировали 200 мМ хлорида натрия/10 мМ ЭДТА при скорости потока 0,5 мл/мин. Встроенный лазерный фотометр Dawn EOS, сопряженный с интерферометрическим рефрактометром Wyatt/Optilab DSP, использовали для измерения рассеяния света и концентрации образца, соответственно (Wyatt Technology Corporation). Концентрацию муцина рассчитывали путем интегрирования пика показателя преломления, ассоциированного с материалом, элюированным в свободном объеме колонки, используя значение для приращения показателя преломления (dn/dc) 0,165 мл/г, измеренное ранее при 650 нм и воспроизводящееся в пределах 5%. Общее содержание белка (муцинов и небольших белков) в данном образце слизи определяли аналогично, но с использованием колонки G-25 (dn/dc=0,170). Содержание белков, не являющихся муцинами (или небольших белков) в образце определяли путем вычитания массового содержания муцина (определяемого путем элюирования образца через колонку S-1000) из общего массового содержания муцинов и других белков (обнаруживаемого при элюировании образца через колонку G-25). По сравнению с методами на основе геля, например, электрофорезом в восстанавливающем или невосстанавливающем ПААГ, дифференциальная рефрактометрия имеет преимущества, состоящие в том, что при ней доступны точные значения молекулярной массы и распределение молекулярной массы, а не качественные сравнения. Количественное преимущество этих методик особенно важно при исследовании крупных гликопротеинов, например, муцинов, где не существует стандартов размера, а молекулярная масса часто превышает 1 МДа (Gillis, R.B., et al., Carbohydr Polym 93(1):178-183, 2013).
Углеводный и белковый блоттинг: Потенциальное воздействие тиоредоксина на тонкую структуру муцина оценивали, выполняя белковый блоттинг интактной слизи НВЕ. 50-200-мкл аликвоты образцов помещали на нитроцеллюлозные мембраны и обрабатывали вакуумом в течение пяти минут для вытягивания всего загруженного образца в мембрану. Образцы двукратно промывали дистиллированной водой. Для анализа содержания углеводов с использованием йодной кислоты и реактивом Шиффа загруженные и промытые образцы инкубировали в течение 30 минут в растворе 0,25% йодной кислоты и 3% уксусной кислоты в воде. После двукратной промывки дистиллированной водой образцы дважды инкубировали в растворе NaMBS (0,1% метабисульфит натрия, 1% HCl) в течение пяти минут. Затем образцы инкубировали с реактивом Шиффа в течение 5-15 мин и дважды промывали NaMBS, один раз дистиллированной водой и быстро высушивали под вакуумом. Пятна белок-специфических антител блокировали 1% молоком в TBST-буфере (1,21% трис-HCl, 8,76%) NaCl и 0,5%) твин, pH 8) и после загрузки и первой промывки дистиллированной водой. После двух пятиминутных промывок TBST образцы инкубировали с первичным антителом (MAN-5ACI для MUC5AC, MUC5BIII и K5B для MUC5B) в течение 30 мин. После двух дополнительных пятиминутных промывок TBST образцы инкубировали с вторичным антителом в течение двух часов. Пятна на мембране проявляли с использованием инфракрасного детектора Li-Cor Odyssey. Кроме того, можно использовать антитела против тиоредоксина для обнаружения моноцистеинового и нативного тиоредоксина, связанного с муцинами. По сравнению со стандартным вестерн-иммуноблоттингом после разделения электрофорезом в ПААГ данный прямой подход на основе блоттинга обеспечивает более быстрое и точное определение содержания муцина и визуализацию и количественную оценку ковалентного взаимодействия между моноцистеиновым активным центром тиоредоксина (r(Cys)hTrx) и дисульфидными связями муцина.
Пример 8
В данном примере продемонстрирована способность моноцистеинового активного центра Trx (r(Cys)hTrx) ослаблять склонность нативного rhTrx индуцировать провоспалительные и патофизиологические эффекты в легких после внутритрахеальной доставки у крыс и мышей.
Оценка высвобождения провоспалительных цитокинов и миграции клеток у здоровых крыс, интратрахеально получавших r(Cys)hTrx и нативный rhTrx. Для определения способности r(Cys)hTrx индуцировать провоспалительные сигнальные пути по сравнению с нативным rhTrx выполнили сравнительное исследование in vivo на крысах с использованием интратрахеальной (IT) доставки возрастающих концентраций белка тиоредоксина по сравнению с контрольным носителем. Два компонента исследования представляли собой: 1) начальное исследование, предназначенное для воспроизведения предыдущих результатов (Rancourt, R. et al., Free Radic Biol Med, 42(9):1441-1453, 2007), полученных с очищенным нативным rhTrx, и 2) основное исследование для сравнения провоспалительных эффектов r(Cys)hTrx и нативного rhTrx. Во всех исследованиях использовали очищенный белок, не содержащий эндотоксинов, обработанный ДТТ или другим подходящим восстановителем для восстановления (активации) Cys-остатков активного центра Trx. После восстановления ДТТ или восстановитель удаляли с помощью эксклюзионной хроматографии. Полное восстановление Cys-остатков Trx проверяли путем анализа in vitro (восстановления DTNB), а каталитическую восстанавливающую активность r(Cys)hTrx по отношению к дисульфидным связям анализировали с помощью анализа восстановления инсулина или ВЭЖХ-анализа связывания мишени.
Начальное исследование: Двадцать четыре крысы произвольно распределили на четыре экспериментальные группы, содержавшие по шесть животных на группу, и обрабатывали следующим образом: Группа 1 - контрольный носитель; Группа 2 - окисленный (неактивный) rhTrx; Группа 3 - восстановленный (активный) rhTrx; Группа 4 - человеческий сывороточный альбумин (ЧСА; отрицательный контроль). Все тестируемые вещества вводили однократно IT-путем. Конечные показатели исследования включали 1) анализ цитокинов - ФНО, хемоаттрактанта нейтрофилов-2, индуцируемого цитокинами (CINC2) и воспалительного белка макрофагов-3 (MIP3) посредством твердофазного иммуноферментного анализа (ELISA), и 2) количество клеток бронхоальвеолярного лаважа (БАЛ) с окрашиванием по Райту для выявления процента воспалительных клеток (нейтрофилов по сравнению с макрофагами). Группа 2 (обработанная окисленным rhTrx) характеризовалась аналогичным уровнем активности цитокинов и количеством клеток в БАЛ по сравнению с Группой 1 (контрольный носитель) и Группой 4 (ЧСА). Группа 3, обработанная восстановленным rhTrx, характеризовалась повышенным уровнем активности цитокинов и количеством клеток по сравнению с Группой 1 и Группой 2.
Основное исследование: Сравнение высвобождения цитокинов у крыс при введении нативного тиоредоксина rhTRX по сравнению с r(Cys)hTRX. В данном сравнительном исследовании определяли относительное высвобождение цитокинов in vivo и миграцию клеток после IT введения rhTrx по сравнению с r(Cys)hTrx. Анализировали три уровня дозы восстановленного rhTrx и восстановленного r(Cys)hTrx (50 мкМ, 200 мкМ и 1000 мкМ). Конечные показатели были аналогичны показателям предварительного исследования, с добавлением иммуногистохимического (IHC) анализа легочной ткани. Показано, что мутация r(Cys)hTrx С-концевого цистеинового мотива активного центра (замене СХХС на СХХХ) удаляет возможность второго тиол-дисульфидного обмена в тиоредоксине и приводит к образованию ковалентной связи с дисульфидной связью-мишенью. Наиболее значимые внеклеточные дисульфидные связи-мишени в легких, с которыми TRX в первую очередь вступает в контакт при IT-доставке, находятся в слое слизи и белков поверхности эпителиальных клеток. Следовательно, по сравнению с нативным rhTrx, r(Cys)hTrx обеспечивает повышенное связывание с эпителиальными клетками легких и связанной с ними слизью, и это связывание можно обнаружить с помощью IHC, используя антитела против тиоредоксина человека. Ткани легких для IHC-анализа собирали у двух животных из каждой группы без отбора БАЛ и сравнивали с двумя животными после отбора БАЛ из той же группы. Следовательно, в каждой группе получали лаваж восьми животных, а двух животных обрабатывали, но не подвергали отбору БАЛ до сбора тканей легких и IHC. r(Cys)hTrx обладал ослабленной способностью увеличивать активность цитокинов и количество клеток в зависимости от дозы по сравнению с аналогичной дозой rhTrx.
Оценка токсичности и патологии легких у здоровых крыс, интратрахеально получавших r(Cys)hTrx. Для оценки действия r(Cys)hTrx на легкие при получении избыточных доз осуществляли IT-введение доз r(Cys)hTrx в диапазоне от 0,5 до 20 мг/кг и оценку наличия патологических изменений через 2 и 14 дней после введения. Самок и самцов крыс Sprague-Dawley случайным образом распределяли на три экспериментальные группы по 20 животных (по 10 каждого пола) в каждой группе. Для всех животных перед исследованием провели медицинский осмотр. Носитель (физиологический раствор) использовали в качестве отрицательного контроля. Конечные показатели данного исследования включали 1) клиническое и гистопатологическое обследование с целью поиска грубых нежелательных изменений; и 2) оценку характеристик сыворотки для обнаружения присутствия исследуемого вещества и антител к исследуемому веществу. Моноцистеиновый r(Cys)hTrx приводил к менее тяжелым последствиям по сравнению с сопоставимыми дозами нативного rhTrx.
Оценка высвобождения провоспалительных цитокинов и миграции клеток у здоровых и bENaC мышей, интратрахеально получавших r(Cys)hTrx и нативный rhTrx. Мышей непродолжительно анестезировали изофлураном и помещали в рабочую станцию для грызунов с изменяющимся углом наклона. С помощью ларингоскопа для грызунов, оснащенного лупой, непосредственно осматривали гортань и вводили микрораспылитель (PennCentury) в дистальный отдел трахеи. Фиксированный объем (25-50 мкл) тестируемого раствора вводили в дыхательные пути и оставляли мышей восстанавливаться. Массу тела регистрировали при капельном введении в дыхательные пути и при эвтаназии. Согласно протоколам, утвержденным комитетом по уходу за животными и их использованию (IACUC), пятерых мышей умертвили из-за их состояния через шесть часов, а еще пятерых мышей - через 24 часа после IT-капельного введения. Осуществляли сбор лаважа дыхательных путей с использованием в общей сложности 1,5 мл холодного стерильного физиологического раствора, содержащего ингибитор протеазы (Pierce). Осуществляли сбор целых легких; все образцы хранили на льду до дальнейшей обработки. Бронхоальвеолярный лаваж (БАЛ) центрифугировали для осаждения лейкоцитов и остатков других клеток. Бесклеточный БАЛ замораживали при -80 С до выполнения твердофазного ИФА. Осадок клеток БАЛ ресуспендировали в 1 мл стерильного физиологического раствора и определяли общее количество лейкоцитов с лейкоформулой с использованием счетчика Coulter и ручного подсчета 300 клеток на препаратах, окрашенных в цитоцентрифуге. После сбора всех образцов БАЛ размораживали на льду и выполняли твердофазный ИФА с целью анализа KC (аналог ИЛ-8), ФНО-альфа, ИЛ-6 и ИЛ-1-бета (ElisaTech, Денвер, штат Колорадо, США). Клеточный состав БАЛ характеризовали по % и общему содержанию лейкоцитов (нейтрофилы, макрофаги, лимфоциты).
TRX дикого типа и мутантный TRX проверяли в двух концентрациях и в два момента времени наряду с контрольными животными с целью определить острый воспалительный ответ дыхательных путей на мутантный TRX и TRX дикого типа по сравнению с контрольным разбавителем (таблица 2). Ответы характеризовали по: изменению массы тела, общему количеству лейкоцитов в БАЛ, относительному и общему количеству нейтрофилов, макрофагов, лимфоцитов, и воспалительным цитокинам, перечисленным выше. r(Cys)hTrx обладал ослабленной способностью повышать активность цитокинов и количество клеток в зависимости от дозы, а также оказывать нежелательное воздействие по сравнению с аналогичными уровнями дозы rhTrx.

Оценка патофизиологических и воспалительных явлений у здоровых и βENaC мышей, интратрахеально получавших r(Cys)hTrx и нативный rhTrx. Предыдущие исследования, выполненные Rancourt и сотрудниками (Rancourt, R. et al., Free Radio Biol Med, 42(9): 1441-1453, 2007), показали, что присутствие дополнительной (экзогенной) слизи в дыхательных путх может значительно ослабить стимуляцию высвобождения цитокинов нативным Trx. Хотя это может означать, что у пациентов с MB и более высокой нагрузкой слизи может проявляться ослабленный воспалительный ответ из-за внеклеточного восстановления Trx, в настоящее время информация об эффектах тиоредоксина в модели дыхательных путей с 1) увеличением эндогенного образования слизи и 2) уже существующим воспалительным ответом (как при MB) отсутствует. Цель данных исследований заключалась в использовании модели мыши с хронической обструкцией слизью/воспалением для исследования влияния нативного Trx и r(Cys)hTrx. В данных исследований использовали модель мыши βENaC, сверхэкспрессирующую бета-субъединицу канала ENaC и демонстрирующую гиперабсорбцию воды в легких (Mall, М., et al., Nat Med. 10(5):487-493, 2004). У мышей βENaC образуется избыточное количество слизи и развивается фенотип легких, аналогичный МВ/ХОБЛ, сопровождающийся закупоркой дыхательных путей слизью и воспалением. Этих мышей ранее использовали в качестве модели заболевания легких при MB в ряде доклинических исследований (например, Graeber, S.Y., et al., Am J Respir Cell Mol Biol 49(3):410-417, 2013). В настоящем исследовании определяли 1) влияние нативного Trx по сравнению cr(Cys)hTrx на гистопатологию и воспалительный статус дыхательных путей/легких; и 2) влияние этих веществ на нагрузку слизи у мышей βENaC (см. ниже) с целью исследования относительного влияния нативного и моноцистеинового Trx (r(Cys)hTrx) в диапазоне доставляемых доз и изучения того, как хроническая избыточная продукция слизи и ранее существовавшее воспаление меняет потенциальную легочную токсичность этих соединений.
Определяли действие трех концентраций (100 мкМ, 500 мкМ и 1000 мкМ) нативного тиоредоксина in vivo с использованием мышей WT и βENaC и сравнивали токсичность и эффективность лекарственного вещества с соединением r(Cys)hTrx в аналогичных концентрациях. Проверяли не менее семи мышей для каждого варианта условий. Все соединения вводили путем интратрахеального капельного введения (10-25 мкл) в заданной концентрации. Осущствляли разовую обработку и выполняли мониторинг токсичности и эффективности лекарственного средства в четыре момента времени (через 4 ч, 24 ч, 72 ч и 7 дней после введения дозы). В ранние моменты времени (4-24 ч) выполняли тщательный мониторинг. Мониторинг в более поздние моменты времени (72 ч - 7 дней) выполняли, если токсическое действие лекарственного вещества поддерживалось в течение 24 часов. Ожидалось, что влияние низких концентраций (100 мкМ, 500 мкМ) не должно сохраняться после 24 ч или 3 суток, соответственно. Использованный протокол позволял надежно выполнять капельное введение 10 мкл в легкие мышей. Вкратце, животных анестезировали изофлураном, помещали на платформу для интубации мышей, где осматривали задний отдел глотки с помощью небольшого ларингоскопа. Препараты капельно вводили непосредственно в трахею (интратрахеальное капельное введение, IT), либо для более однородного и более глубокого осаждения 25 мкл доставляли путем микрораспыления с помощью устройства Penn Century. После обработки животных умерщвляли в соответствующий момент времени. Интактные легкие собирали для гистологического анализа, а бронхоальвеолярный лаваж - для различных измерений, например, количества клеток, содержания муцина, восстановительного статуса и цитокинов. Подробнее, левое легкое перевязывали лигатурой вокруг основного ствола бронха и хирургически выделяли для гистологического исследования. Бронхоальвеолярный лаваж противоположного легкого (или правой доли) собирали с помощью трахеотомия и канюли, используя 500 мкл стерильного PBS. После фиксации целого легкого анализировали продольные срезы с помощью окрашивания Н&Е и АВ-PAS с целью оценки воспаления и сохранившейся слизи. В БАЛ анализировали количество клеток/лейкоформулу, содержание муцина и восстановительный статус (способ разделения в агарозном геле и вестерн-блоттинг), а также цитокины.
Размер выборки и статистический анализ: Каждое из соединений оценивали в различных концентрациях (100 мкмоль, 500 мкмоль, 1000 мкмоль), используя в общей сложности 7 мышей на один вариант условий. Анализировали две группы - мышей дикого типа (WT) и βENaC (таблица 3). Как указано буквой "X" в таблице, обработано и проанализировано в общей сложности 36 вариантов условий × 7 мышей (всего 252 мыши) с целью обеспечить достаточное понимание изменчивости внутри и между выборками при каждом варианте условий. Для анализа данных выполняли сравнение отдельных образцов/групп с использованием двустороннего t-критерия Стьюдента. Для сравнения множественных групп использовали односторонний дисперсионный анализ (ANOVA). Значимость для всех анализов задали при р<0,05. Моноцистеиновый r(Cys)hTrx приводил к менее тяжелым последствиям по сравнению с сопоставимыми дозами нативного rhTrx, и этот эффект дополнительно ослаблялся присутствием эндогенной слизи у мышей βENaC.
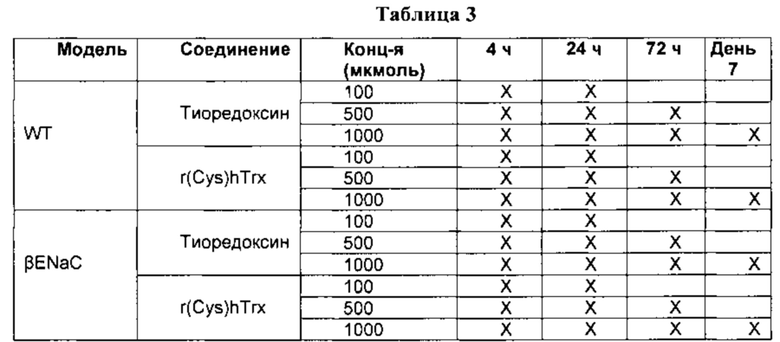
"X" означает количество повторностей × количество проанализированных концентраций
Пример 9
В данном примере продемонстрировано, что r(Cys)hTrx обладает улучшенной активностью по отношению к нормализации слизи по сравнению с rhTrx дикого типа. Определяли действие rhTrx дикого типа, r(Cys)hTrx и различных контрольных материалов, включая дитиотреитол (DTT) в двух концентрациях, N-ацетилцистеин (NAC) и рекомбинантную ДНКазу человека (rhDNase) в эквимолярных концентрациях, на нормализацию образцов мокроты пациентов (n=6 на обработку) в анализе уплотнения мокроты. Исследования мокроты проводили с использованием четырех-шести образцов, полученных при спонтанном (не индуцированном) откашливании от каждого из трех отдельных пациентов с MB для каждого эксперимента. Из каждого образца собирали и объединяли "благоприятную" часть мокроты (т.е. слизь); общий образец осторожно гомогенизировали перемешиванием или мягким встряхиванием на вортексе, затем распределяли в пробирки и подвергали обработке: разбавителем (т.е. трис-буфером), ДТТ в разбавителе (в концентрации 0,58 мМ и 1,5 мМ), NAC в разбавителе (0,58 мМ) rhDNase в разбавителе (0,58 мМ), rhTrx в разбавителе (0,58 мМ) или r(Cys)hTrx в разбавителе (0,58 мМ). Свежий ДТТ, получаемый для каждого эксперимента, использовали в качестве положительного контроля. На фигуре 2 показано, что r(Cys)hTrx мог заметно повышать разжижение мокроты пациентов по сравнению с тиоредоксином дикого типа (называемым "rhTrx" на фигуре 2), тем самым демонстрируя улучшенную активность по отношению к нормализации слизи по сравнению как с нативным Trx, так и стандартно применяемой рекомбинантной ДНКазой человека, Pulmozyme™ ("rhDNase"). Как тиоредоксин, так и ДНКаза, в свою очередь, были более эффективны по сравнению с эквимолярным количеством NAC, широко применяемого в качестве муколитического средства за пределами США. Эти результаты, полученные с использованием мокроты пациентов-людей с MB, показывают, что моноцистеиновый r(Cys)hTrx сохранял полную активность по отношению к нормализации вязкоупругости мокроты при MB (и даже являлся более эффективным, чем немодифицированный нативный rhTrx), несмотря на ожидаемое ковалентное связывание с Cys муцина. В то же время возможность проникновения небольшого (12 кДа) r(Cys)hTrx через эпителий легких должна значительно снижаться за счет связывания с муцином. Способность активного r(Cys)hTrx впоследствии проникать в ядро клетки и взаимодействовать с белками-мишенями, регулирующими окислительно-восстановительные процессы, например, NFkB, является еще более низкой, как и воздействие на любой сигнальный путь за счет взаимодействия с внеклеточными трансмембранными доменами на иммунных клетках, как теоретически допускалось для Trx-опосредованной активации рецептора CD30 ФНО (Schwertassek, U., et al., EMBO J 26(13):3086-3097, 2007). Повышенная эффективноть, наблюдаемая для r(Cys)hTrx (фигуры 1a, 1b и 2) согласуется с ковалентным связыванием этой молекулы с Cys муцина, поскольку связанные остатки Cys не могут повторно образовывать дисульфидные связи, в отличие от обработки нативным rhTrx (или фактически любым низкомолекулярным тиоловым агентом).
Каждая из публикаций и других литературных источников, обсуждавшихся или упомянутых в настоящем документе, полностью включена в настоящий документ посредством ссылки.
Несмотря на подробное описание различных вариантов реализации настоящего изобретения, очевидно, что специалисты в данной области техники могут осуществить модификации и адаптации этих вариантов реализации. Вместе с тем, следует четко понимать, что такие модификации и адаптации находятся в рамках настоящего изобретения, как изложено в нижеследующей формуле изобретения.
--->
Перечень последовательностей
<110> Хейфец, Петер
<120> Продукт и способ для нормализации вязкости слизи
<130> 7579-1-PCT
<140> еще не присвоен
<141> 2014-03-17
<150> 61/792,198
<151> 2013-03-15
<160> 27
<170> PatentIn версия 3.5
<210> 1
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтезированный пептидный мотив
<400> 1
Cys Gly Pro Cys
1
<210> 2
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтезированный пептидный мотив
<220>
<221> misc_feature
<222> (1)..(6)
<223> Xaa = любая аминокислота
<400> 2
Xaa Cys Gly Pro Cys Xaa
1 5
<210> 3
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтезированный пептидный мотив
<400> 3
Trp Cys Gly Pro Cys Lys
1 5
<210> 4
<211> 109
<212> БЕЛОК
<213> Pseudomonas syringae
<400> 4
Met Ser Asn Asp Leu Ile Lys His Val Thr Asp Ala Ser Phe Glu Ala
1 5 10 15
Asp Val Leu Lys Ala Asp Gly Ala Val Leu Val Asp Tyr Trp Ala Glu
20 25 30
Trp Cys Gly Pro Cys Lys Met Ile Ala Pro Val Leu Asp Glu Ile Ala
35 40 45
Thr Thr Tyr Ala Gly Lys Leu Thr Ile Ala Lys Leu Asn Ile Asp Glu
50 55 60
Asn Gln Glu Thr Pro Ala Lys His Gly Val Arg Gly Ile Pro Thr Leu
65 70 75 80
Met Leu Phe Lys Asn Gly Asn Val Glu Ala Thr Lys Val Gly Ala Leu
85 90 95
Ser Lys Ser Gln Leu Ala Ala Phe Leu Asp Ala Asn Ile
100 105
<210> 5
<211> 104
<212> БЕЛОК
<213> Porphyromonas gingivalis
<400> 5
Met Ala Leu Gln Ile Thr Asp Ala Thr Phe Asp Gly Leu Val Ala Glu
1 5 10 15
Gly Lys Pro Met Val Val Asp Phe Trp Ala Thr Trp Cys Gly Pro Cys
20 25 30
Arg Met Val Gly Pro Ile Ile Asp Glu Leu Ala Ala Glu Tyr Glu Gly
35 40 45
Arg Ala Ile Ile Gly Lys Val Asp Val Asp Ala Asn Thr Glu Leu Pro
50 55 60
Met Lys Tyr Gly Val Arg Asn Ile Pro Thr Ile Leu Phe Ile Lys Asn
65 70 75 80
Gly Glu Val Val Lys Lys Leu Val Gly Ala Gln Ser Lys Asp Val Phe
85 90 95
Lys Lys Glu Leu Asp Ala Leu Phe
100
<210> 6
<211> 103
<212> БЕЛОК
<213> Listeria monocytogenes
<400> 6
Met Val Lys Glu Ile Thr Asp Ala Thr Phe Glu Gln Glu Thr Ser Glu
1 5 10 15
Gly Leu Val Leu Thr Asp Phe Trp Ala Thr Trp Cys Gly Pro Cys Arg
20 25 30
Met Val Ala Pro Val Leu Glu Glu Ile Gln Glu Glu Arg Gly Glu Ala
35 40 45
Leu Lys Ile Val Lys Met Asp Val Asp Glu Asn Pro Glu Thr Pro Gly
50 55 60
Ser Phe Gly Val Met Ser Ile Pro Thr Leu Leu Ile Lys Lys Asp Gly
65 70 75 80
Glu Val Val Glu Thr Ile Ile Gly Tyr Arg Pro Lys Glu Glu Leu Asp
85 90 95
Glu Val Ile Asn Lys Tyr Val
100
<210> 7
<211> 103
<212> БЕЛОК
<213> Saccharomyces cerevisiae
<400> 7
Met Val Thr Gln Phe Lys Thr Ala Ser Glu Phe Asp Ser Ala Ile Ala
1 5 10 15
Gln Asp Lys Leu Val Val Val Asp Phe Tyr Ala Thr Trp Cys Gly Pro
20 25 30
Cys Lys Met Ile Ala Pro Met Ile Glu Lys Phe Ser Glu Gln Tyr Pro
35 40 45
Gln Ala Asp Phe Tyr Lys Leu Asp Val Asp Glu Leu Gly Asp Val Ala
50 55 60
Gln Lys Asn Glu Val Ser Ala Met Pro Thr Leu Leu Leu Phe Lys Asn
65 70 75 80
Gly Lys Glu Val Ala Lys Val Val Gly Ala Asn Pro Ala Ala Ile Lys
85 90 95
Gln Ala Ile Ala Ala Asn Ala
100
<210> 8
<211> 105
<212> БЕЛОК
<213> Gallus gallus
<400> 8
Met Val Lys Ser Val Gly Asn Leu Ala Asp Phe Glu Ala Glu Leu Lys
1 5 10 15
Ala Ala Gly Glu Lys Leu Val Val Val Asp Phe Ser Ala Thr Trp Cys
20 25 30
Gly Pro Cys Lys Met Ile Lys Pro Phe Phe His Ser Leu Cys Asp Lys
35 40 45
Phe Gly Asp Val Val Phe Ile Glu Ile Asp Val Asp Asp Ala Gln Asp
50 55 60
Val Ala Thr His Cys Asp Val Lys Cys Met Pro Thr Phe Gln Phe Tyr
65 70 75 80
Lys Asn Gly Lys Lys Val Gln Glu Phe Ser Gly Ala Asn Lys Glu Lys
85 90 95
Leu Glu Glu Thr Ile Lys Ser Leu Val
100 105
<210> 9
<211> 105
<212> БЕЛОК
<213> Mus musculus
<400> 9
Met Val Lys Leu Ile Glu Ser Lys Glu Ala Phe Gln Glu Ala Leu Ala
1 5 10 15
Ala Ala Gly Asp Lys Leu Val Val Val Asp Phe Ser Ala Thr Trp Cys
20 25 30
Gly Pro Cys Lys Met Ile Lys Pro Phe Phe His Ser Leu Cys Asp Lys
35 40 45
Tyr Ser Asn Val Val Phe Leu Glu Val Asp Val Asp Asp Cys Gln Asp
50 55 60
Val Ala Ala Asp Cys Glu Val Lys Cys Met Pro Thr Phe Gln Phe Tyr
65 70 75 80
Lys Lys Gly Gln Lys Val Gly Glu Phe Ser Gly Ala Asn Lys Glu Lys
85 90 95
Leu Glu Ala Ser Ile Thr Glu Tyr Ala
100 105
<210> 10
<211> 105
<212> БЕЛОК
<213> Rattus norvegicus
<400> 10
Met Val Lys Leu Ile Glu Ser Lys Glu Ala Phe Gln Glu Ala Leu Ala
1 5 10 15
Ala Ala Gly Asp Lys Leu Val Val Val Asp Phe Ser Ala Thr Trp Cys
20 25 30
Gly Pro Cys Lys Met Ile Lys Pro Phe Phe His Ser Leu Cys Asp Lys
35 40 45
Tyr Ser Asn Val Val Phe Leu Glu Val Asp Val Asp Asp Cys Gln Asp
50 55 60
Val Ala Ala Asp Cys Glu Val Lys Cys Met Pro Thr Phe Gln Phe Tyr
65 70 75 80
Lys Lys Gly Gln Lys Val Gly Glu Phe Ser Gly Ala Asn Lys Glu Lys
85 90 95
Leu Glu Ala Thr Ile Thr Glu Phe Ala
100 105
<210> 11
<211> 105
<212> БЕЛОК
<213> Bos taurus
<400> 11
Met Val Lys Gln Ile Glu Ser Lys Tyr Ala Phe Gln Glu Ala Leu Asn
1 5 10 15
Ser Ala Gly Glu Lys Leu Val Val Val Asp Phe Ser Ala Thr Trp Cys
20 25 30
Gly Pro Cys Lys Met Ile Lys Pro Phe Phe His Ser Leu Ser Glu Lys
35 40 45
Tyr Ser Asn Val Val Phe Leu Glu Val Asp Val Asp Asp Cys Gln Asp
50 55 60
Val Ala Ala Glu Cys Glu Val Lys Cys Met Pro Thr Phe Gln Phe Phe
65 70 75 80
Lys Lys Gly Gln Lys Val Gly Glu Phe Ser Gly Ala Asn Lys Glu Lys
85 90 95
Leu Glu Ala Thr Ile Asn Glu Leu Ile
100 105
<210> 12
<211> 105
<212> БЕЛОК
<213> Homo sapiens
<400> 12
Met Val Lys Gln Ile Glu Ser Lys Thr Ala Phe Gln Glu Ala Leu Asp
1 5 10 15
Ala Ala Gly Asp Lys Leu Val Val Val Asp Phe Ser Ala Thr Trp Cys
20 25 30
Gly Pro Cys Lys Met Ile Lys Pro Phe Phe His Ser Leu Ser Glu Lys
35 40 45
Tyr Ser Asn Val Ile Phe Leu Glu Val Asp Val Asp Asp Cys Gln Asp
50 55 60
Val Ala Ser Glu Cys Glu Val Lys Cys Met Pro Thr Phe Gln Phe Phe
65 70 75 80
Lys Lys Gly Gln Lys Val Gly Glu Phe Ser Gly Ala Asn Lys Glu Lys
85 90 95
Leu Glu Ala Thr Ile Asn Glu Leu Val
100 105
<210> 13
<211> 134
<212> БЕЛОК
<213> Arabidopsis thaliana
<400> 13
Met Gly Gly Ala Leu Ser Thr Val Phe Gly Ser Gly Glu Asp Ala Ala
1 5 10 15
Ala Ala Gly Thr Glu Ser Ser Glu Pro Ser Arg Val Leu Lys Phe Ser
20 25 30
Ser Ser Ala Arg Trp Gln Leu His Phe Asn Glu Ile Lys Glu Ser Asn
35 40 45
Lys Leu Leu Val Val Asp Phe Ser Ala Ser Trp Cys Gly Pro Cys Arg
50 55 60
Met Ile Glu Pro Ala Ile His Ala Met Ala Asp Lys Phe Asn Asp Val
65 70 75 80
Asp Phe Val Lys Leu Asp Val Asp Glu Leu Pro Asp Val Ala Lys Glu
85 90 95
Phe Asn Val Thr Ala Met Pro Thr Phe Val Leu Val Lys Arg Gly Lys
100 105 110
Glu Ile Glu Arg Ile Ile Gly Ala Lys Lys Asp Glu Leu Glu Lys Lys
115 120 125
Val Ser Lys Leu Arg Ala
130
<210> 14
<211> 167
<212> БЕЛОК
<213> Zea mays
<400> 14
Met Ala Met Glu Thr Cys Phe Arg Ala Trp Ala Leu His Ala Pro Ala
1 5 10 15
Gly Ser Lys Asp Arg Leu Leu Val Gly Asn Leu Val Leu Pro Ser Lys
20 25 30
Arg Ala Leu Ala Pro Leu Ser Val Gly Arg Val Ala Thr Arg Arg Pro
35 40 45
Arg His Val Cys Gln Ser Lys Asn Ala Val Asp Glu Val Val Val Ala
50 55 60
Asp Glu Lys Asn Trp Asp Gly Leu Val Met Ala Cys Glu Thr Pro Val
65 70 75 80
Leu Val Glu Phe Trp Ala Pro Trp Cys Gly Pro Cys Arg Met Ile Ala
85 90 95
Pro Val Ile Asp Glu Leu Ala Lys Asp Tyr Ala Gly Lys Ile Thr Cys
100 105 110
Cys Lys Val Asn Thr Asp Asp Ser Pro Asn Val Ala Ser Thr Tyr Gly
115 120 125
Ile Arg Ser Ile Pro Thr Val Leu Ile Phe Lys Gly Gly Glu Lys Lys
130 135 140
Glu Ser Val Ile Gly Ala Val Pro Lys Ser Thr Leu Thr Thr Leu Ile
145 150 155 160
Asp Lys Tyr Ile Gly Ser Ser
165
<210> 15
<211> 172
<212> БЕЛОК
<213> Oryza sativa
<400> 15
Met Ala Leu Glu Thr Cys Phe Arg Ala Trp Ala Thr Leu His Ala Pro
1 5 10 15
Gln Pro Pro Ser Ser Gly Gly Ser Arg Asp Arg Leu Leu Leu Ser Gly
20 25 30
Ala Gly Ser Ser Gln Ser Lys Pro Arg Leu Ser Val Ala Ser Pro Ser
35 40 45
Pro Leu Arg Pro Ala Ser Arg Phe Ala Cys Gln Cys Ser Asn Val Val
50 55 60
Asp Glu Val Val Val Ala Asp Glu Lys Asn Trp Asp Ser Met Val Leu
65 70 75 80
Gly Ser Glu Ala Pro Val Leu Val Glu Phe Trp Ala Pro Trp Cys Gly
85 90 95
Pro Cys Arg Met Ile Ala Pro Val Ile Asp Glu Leu Ala Lys Glu Tyr
100 105 110
Val Gly Lys Ile Lys Cys Cys Lys Val Asn Thr Asp Asp Ser Pro Asn
115 120 125
Ile Ala Thr Asn Tyr Gly Ile Arg Ser Ile Pro Thr Val Leu Met Phe
130 135 140
Lys Asn Gly Glu Lys Lys Glu Ser Val Ile Gly Ala Val Pro Lys Thr
145 150 155 160
Thr Leu Ala Thr Ile Ile Asp Lys Tyr Val Ser Ser
165 170
<210> 16
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтезированный пептидный мотив
<220>
<221> MISC_FEATURE
<222> (2)..(3)
<223> Xaa = любая аминокислота за исключением цистеина
<400> 16
Cys Xaa Xaa Cys
1
<210> 17
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтезированный пептидный мотив
<220>
<221> MISC_FEATURE
<222> (2)..(4)
<223> Xaa = любая аминокислота за исключением цистеина
<400> 17
Cys Xaa Xaa Xaa
1
<210> 18
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтезированный пептидный мотив
<220>
<221> MISC_FEATURE
<222> (4)..(4)
<223> Xaa = любая аминокислота за исключением цистеина
<400> 18
Cys Gly Pro Xaa
1
<210> 19
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтезированный пептидный мотив
<220>
<221> MISC_FEATURE
<222> (1)..(1)
<223> Xaa = любая аминокислота за исключением цистеина
<220>
<221> MISC_FEATURE
<222> (3)..(6)
<223> Xaa = любая аминокислота за исключением цистеина
<400> 19
Xaa Cys Xaa Xaa Xaa Xaa
1 5
<210> 20
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтезированный пептидный мотив
<220>
<221> MISC_FEATURE
<222> (1)..(1)
<223> Xaa = любая аминокислота за исключением цистеина
<220>
<221> MISC_FEATURE
<222> (3)..(4)
<223> Xaa = любая аминокислота за исключением цистеина
<220>
<221> MISC_FEATURE
<222> (6)..(6)
<223> Xaa = любая аминокислота за исключением цистеина
<400> 20
Xaa Cys Xaa Xaa Cys Xaa
1 5
<210> 21
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтезированный пептидный мотив
<220>
<221> MISC_FEATURE
<222> (1)..(1)
<223> Xaa = любая аминокислота за исключением цистеина
<220>
<221> MISC_FEATURE
<222> (5)..(6)
<223> Xaa = любая аминокислота за исключением цистеина
<400> 21
Xaa Cys Gly Pro Xaa Xaa
1 5
<210> 22
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтезированный пептидный мотив
<220>
<221> MISC_FEATURE
<222> (1)..(1)
<223> Xaa = любая аминокислота за исключением цистеина
<220>
<221> MISC_FEATURE
<222> (6)..(6)
<223> Xaa = любая аминокислота за исключением цистеина
<400> 22
Xaa Cys Gly Pro Cys Xaa
1 5
<210> 23
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтезированный пептидный мотив
<220>
<221> MISC_FEATURE
<222> (5)..(5)
<223> Xaa = любая аминокислота за исключением цистеина
<400> 23
Trp Cys Gly Pro Xaa Lys
1 5
<210> 24
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтезированный пептидный мотив
<220>
<221> MISC_FEATURE
<222> (2)..(3)
<223> Xaa = любая аминокислота за исключением цистеина
<400> 24
Cys Xaa Xaa Ser
1
<210> 25
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтезированный пептидный мотив
<220>
<221> MISC_FEATURE
<222> (1)..(1)
<223> Xaa = любая аминокислота за исключением цистеина
<220>
<221> MISC_FEATURE
<222> (3)..(4)
<223> Xaa = любая аминокислота за исключением цистеина
<220>
<221> MISC_FEATURE
<222> (6)..(6)
<223> Xaa = любая аминокислота за исключением цистеина
<400> 25
Xaa Cys Xaa Xaa Ser Xaa
1 5
<210> 26
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтезированный пептидный мотив
<220>
<221> MISC_FEATURE
<222> (1)..(1)
<223> Xaa = любая аминокислота за исключением цистеина
<220>
<221> MISC_FEATURE
<222> (6)..(6)
<223> Xaa = любая аминокислота за исключением цистеина
<400> 26
Xaa Cys Gly Pro Ser Xaa
1 5
<210> 27
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтезированный пептидный мотив
<400> 27
Trp Cys Gly Pro Ser Lys
1 5
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Моноклональное антитело, специфично связывающееся с эпитопом тиоредоксина-1, его получение и применение | 2018 |
|
RU2791264C2 |
| Эпитоп тиоредоксина-1, специфично связывающийся с моноклональным антителом | 2018 |
|
RU2804772C2 |
| ЭПИТОП ТИОРЕДОКСИНА-1, СПЕЦИФИЧНО СВЯЗЫВАЮЩИЙСЯ С МОНОКЛОНАЛЬНЫМ АНТИТЕЛОМ | 2018 |
|
RU2804126C2 |
| ЭПИТОП ТИОРЕДОКСИНА-1, СПЕЦИФИЧНО СВЯЗЫВАЮЩИЙСЯ С МОНОКЛОНАЛЬНЫМ АНТИТЕЛОМ | 2023 |
|
RU2802944C1 |
| Эпитоп тиоредоксина-1 и моноклональное антитело, специфически связывающееся с ним | 2018 |
|
RU2797266C2 |
| Моноклональное антитело, специфично связывающееся с эпитопом тиоредоксина-1, его получение и применение | 2018 |
|
RU2790687C2 |
| БЕЛОК И ВАКЦИНА ПРОТИВ ИНФЕКЦИИ SARS-CoV-2 | 2020 |
|
RU2815060C1 |
| МОНОКЛОНАЛЬНОЕ АНТИТЕЛО, СПЕЦИФИЧЕСКИ СВЯЗЫВАЮЩЕЕСЯ С ТИОРЕДОКСИНОМ-1, И ЕГО ПРИМЕНЕНИЕ | 2016 |
|
RU2747929C2 |
| УЛУЧШЕНИЕ ИММУНОГЕННОСТИ ПЕПТИДА L2 ВПЧ | 2017 |
|
RU2743016C2 |
| ЦИКЛИЧЕСКИЙ БЕЛОК, НЕ СОДЕРЖАЩИЙ ОСТАТКОВ ЦИСТЕИНА | 2008 |
|
RU2486196C2 |
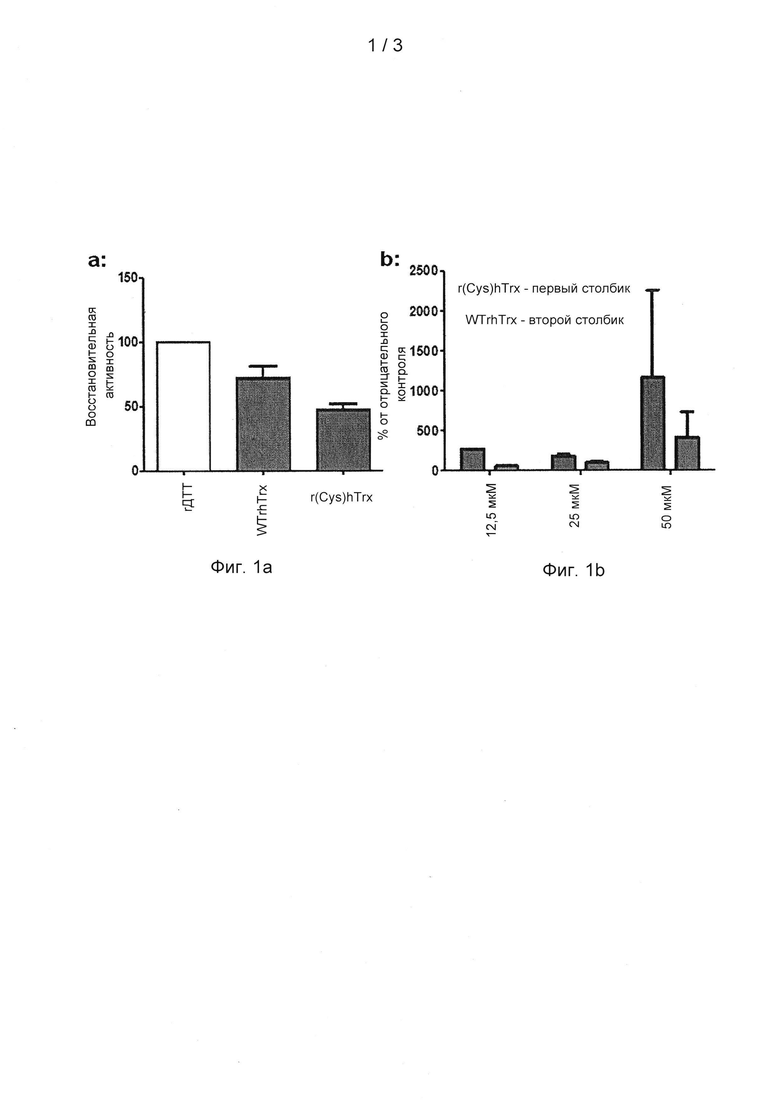
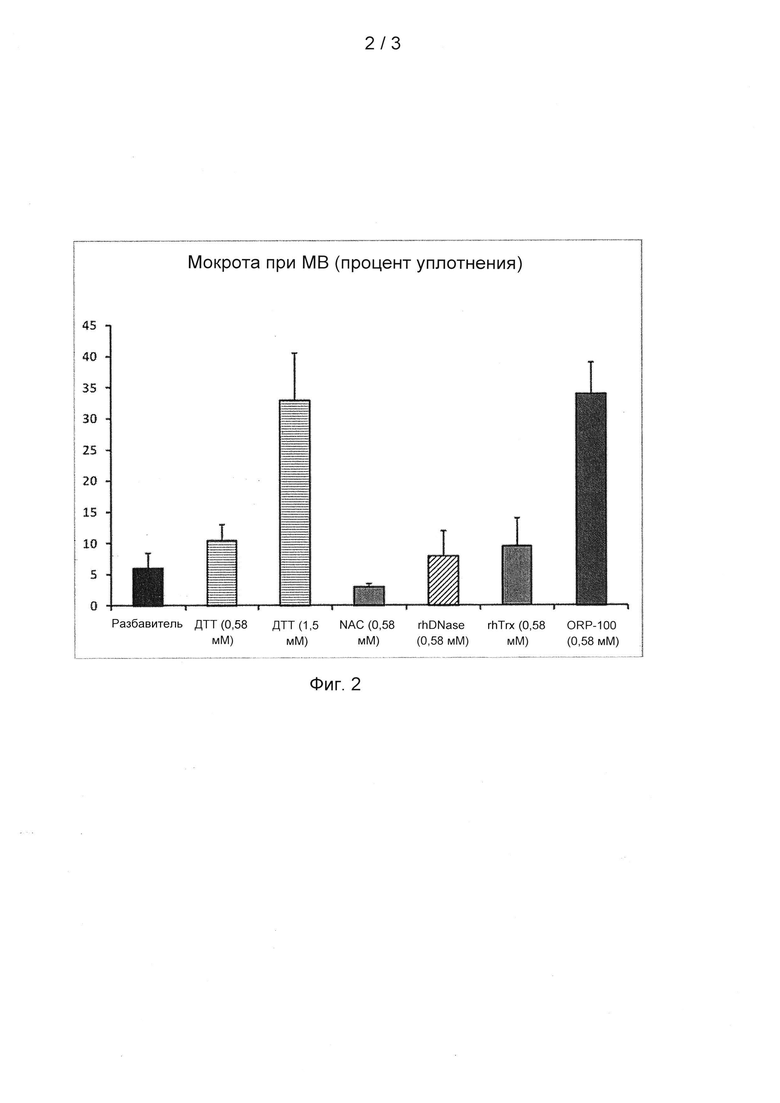
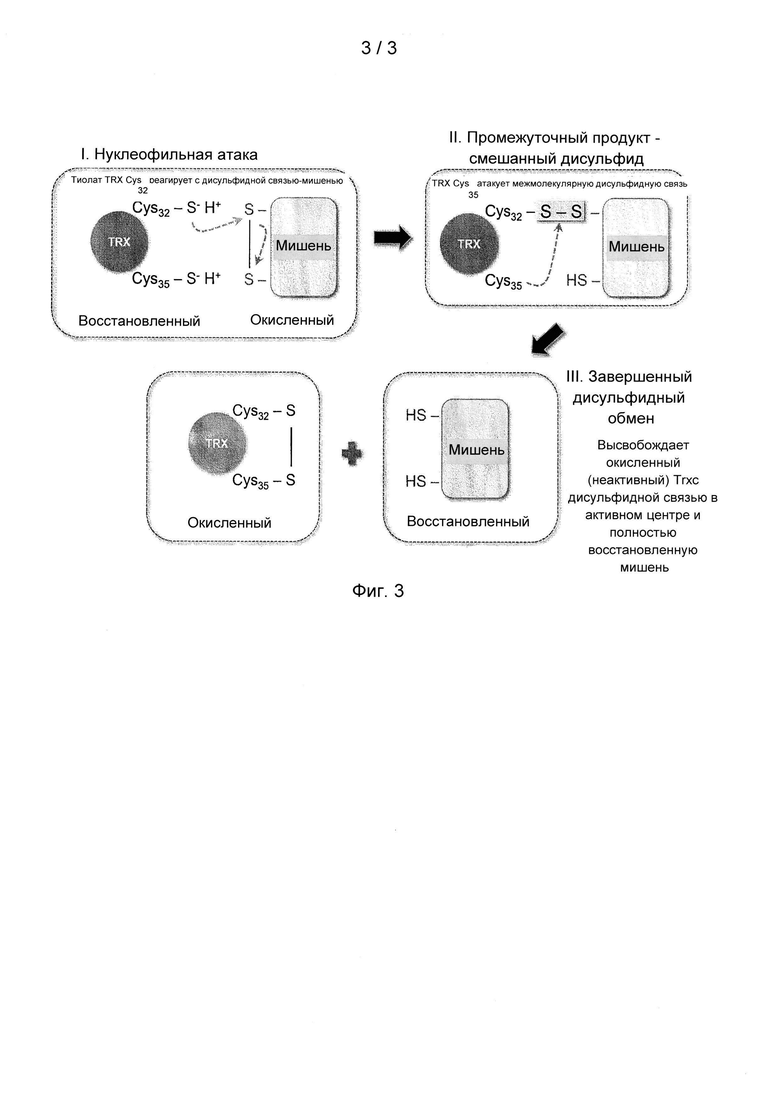
Группа изобретений относится к медицине, а именно к терапии, и может быть использована для снижения вязкоупругости слизи или мокроты у пациента с избыточно вязкой или липкой слизью или мокротой. Для этого слизь или мокроту указанного пациента приводят в контакт с композицией, содержащей белок или пептид, содержащий моноцистеиновый активный центр тиоредоксина в восстановленном состоянии, где моноцистеиновый активный центр тиоредоксина содержит аминокислотную последовательность, выбранную из группы, состоящей из C-X-X-S (SEQ ID NO: 24), C-X-X-X (SEQ ID NO: 17), X-C-X-X-X-X (SEQ ID NO: 19), X-C-G-P-X-X (SEQ ID NO: 21), W-C-G-P-X-K (SEQ ID NO: 23), X-C-X-X-S-X (SEQ ID NO: 25), X-C-G-P-S-X (SEQ ID NO: 26) и W-C-G-P-S-K (SEQ ID NO: 27), при этом указанный остаток C находится в восстановленном состоянии, и при этом указанные остатки X представляют собой любой аминокислотный остаток, кроме цистеина. Также предложены способы, фармацевтические композиции и их применение для превращения дисульфидной связи белка-мишени в стабильную смешанную дисульфидную связь, для поддержания стабильной смешанной дисульфидной связи, для ее превращения в стабильную смешанную дисульфидную связь. Группа изобретений позволяет повысить эффективность разжижения слизи или мокроты за счет ковалентного связывания с цистеином муцина при безопасности и хорошей переносимости. 10 н. и 39 з.п. ф-лы, 3 ил., 3 табл., 9 пр.
1. Способ снижения вязкоупругости слизи или мокроты у пациента с избыточно вязкой или липкой слизью или мокротой, включающий этап приведения указанной слизи или указанной мокроты указанного пациента в контакт с композицией, содержащей белок или пептид, содержащий моноцистеиновый активный центр тиоредоксина в восстановленном состоянии, эффективно снижающей вязкость указанной слизи или указанной мокроты по сравнению с вязкостью до этапа приведения в контакт, где моноцистеиновый активный центр тиоредоксина содержит аминокислотную последовательность, выбранную из группы, состоящей из C-X-X-S (SEQ ID NO: 24), C-X-X-X (SEQ ID NO: 17), X-C-X-X-X-X (SEQ ID NO: 19), X-C-G-P-X-X (SEQ ID NO: 21), W-C-G-P-X-K (SEQ ID NO: 23), X-C-X-X-S-X (SEQ ID NO: 25), X-C-G-P-S-X (SEQ ID NO: 26) и W-C-G-P-S-K (SEQ ID NO: 27), при этом указанный остаток C находится в восстановленном состоянии, и при этом указанные остатки X представляют собой любой аминокислотный остаток, кроме цистеина.
2. Способ по п. 1, отличающийся тем, что указанный пациент страдает заболеванием легких, при котором аномальная или избыточная вязкость или липкость слизи или мокроты является симптомом или причиной указанного заболевания.
3. Способ по п. 1, отличающийся тем, что указанный пациент страдает заболеванием легких, при котором аномальная или чрезмерная вязкость или липкость слизи или мокроты ассоциирована с недостаточностью активности биологического восстановителя.
4. Способ по п. 1, отличающийся тем, что указанный пациент страдает заболеванием, выбранным из группы, состоящей из муковисцидоза, хронической обструктивной болезни легких, бронхоэктатической болезни, астмы и заболевания пищеварительного тракта.
5. Способ по п. 1, отличающийся тем, что указанный пациент страдает муковисцидозом.
6. Способ по п. 4, отличающийся тем, что указанное заболевание пищеварительного тракта является кокцидиозом.
7. Способ по п. 1, отличающийся тем, что указанный этап приведения слизи или мокроты указанного пациента в контакт с указанной композицией выполняют посредством введения указанной композиции указанному пациенту путем, выбранным из группы, состоящей из назального, интратрахеального, бронхиального, ингаляционного и перорального введения, а также прямого введения в лёгкие.
8. Способ по п. 1, отличающийся тем, что указанная слизь или мокрота, с которой осуществляется контакт, находится в дыхательных путях, пищеварительном тракте или половых путях указанного пациента.
9. Способ по п. 1, отличающийся тем, что указанную композицию вводят указанному пациенту в фармацевтически приемлемом носителе.
10. Способ по любому из пп. 1-9, отличающийся тем, что указанный этап приведения слизи или мокроты указанного пациента в контакт с указанной композицией увеличивает процент свободных тиолов в образце слизи или мокроты указанного пациента по сравнению с процентом до контакта с указанной композицией.
11. Способ по любому из пп. 1-9, отличающийся тем, что после указанного этапа приведения слизи или мокроты указанного пациента в контакт с указанной композицией у указанного пациента наблюдается по меньшей мере 2,5% увеличение объема форсированного выдоха (ОФВ) по сравнению с объемом до контакта с указанной композицией.
12. Способ по любому из пп. 1-9, отличающийся тем, что указанный моноцистеиновый активный центр тиоредоксина содержит аминокислотную последовательность C-X-X-S (SEQ ID NO: 24), при этом указанный остаток C находится в восстановленном состоянии, и при этом указанные остатки X являются остатками любых аминокислот, кроме цистеина.
13. Способ по любому из пп. 1-9, отличающийся тем, что указанный белок или пептид, содержащий моноцистеиновый активный центр тиоредоксина, ковалентно связывается с остатком цистеина в белке слизи.
14. Способ по п. 13, отличающийся тем, что указанный белок слизи представляет собой муцин.
15. Способ по п. 13, отличающийся тем, что указанный белок слизи выбран из группы, состоящей из белка слизи дыхательных путей и белка слизи пищеварительного тракта.
16. Способ по любому из пп. 1-9, отличающийся тем, что указанный белок содержит тиоредоксин, выбранный из группы, состоящей из тиоредоксина прокариот, тиоредоксина грибов, тиоредоксина растений и тиоредоксина млекопитающих.
17. Способ по любому из пп. 1-9, отличающийся тем, что указанный белок содержит тиоредоксин человека.
18. Способ по любому из пп. 1-9, отличающийся тем, что указанная композиция дополнительно содержит восстанавливающий агент для восстановления указанного моноцистеинового активного центра тиоредоксина в составе указанного белка.
19. Способ по п. 18, отличающийся тем, что указанная композиция дополнительно содержит тиоредоксинредуктазу.
20. Композиция для превращения дисульфидной связи белка-мишени в стабильную смешанную дисульфидную связь, содержащая белок или пептид, содержащие моноцистеиновый активный центр тиоредоксина, где моноцистеиновый активный центр тиоредоксина содержит аминокислотную последовательность, выбранную из группы, состоящей из C-X-X-S (SEQ ID NO: 24), C-X-X-X (SEQ ID NO: 17), X-C-X-X-X-X (SEQ ID NO: 19), X-C-G-P-X-X (SEQ ID NO: 21), W-C-G-P-X-K (SEQ ID NO: 23), X-C-X-X-S-X (SEQ ID NO: 25), X-C-G-P-S-X (SEQ ID NO: 26) и W-C-G-P-S-K (SEQ ID NO: 27), при этом указанный остаток C находится в восстановленном состоянии, и при этом указанные остатки X представляют собой любой аминокислотный остаток, кроме цистеина.
21. Композиция по п. 20, отличающаяся тем, что указанный моноцистеиновый активный центр тиоредоксина содержит аминокислотную последовательность C-X-X-S (SEQ ID NO: 24), при этом указанный остаток C находится в восстановленном состоянии, и при этом указанные остатки X являются остатками любых аминокислот, кроме цистеина.
22. Композиция по любому из пп. 20 или 21, отличающаяся тем, что указанный белок содержит тиоредоксин, выбранный из группы, состоящей из тиоредоксина прокариот, тиоредоксина грибов, тиоредоксина растений, тиоредаксина животных и тиоредоксина млекопитающих.
23. Композиция по любому из пп. 20 или 21, отличающаяся тем, что указанный белок содержит тиоредоксин человека.
24. Композиция по любому из пп. 20 или 21, отличающаяся тем, что указанная композиция дополнительно содержит восстановитель.
25. Композиция по п. 24, отличающаяся тем, что указанная композиция дополнительно содержит тиоредоксинредуктазу.
26. Композиция по п. 20, дополнительно содержащая по меньшей мере один дополнительный агент для лечения избыточно вязкой или липкой слизи или мокроты.
27. Фармацевтическая композиция для превращения дисульфидной связи белка-мишени в стабильную смешанную дисульфидную связь, содержащая белок или пептид, содержащий моноцистеиновый активный центр тиоредоксина в восстановленном состоянии, причем указанная композиция составлена для перорального введения, аэрозольного введения или местного применения, где моноцистеиновый активный центр тиоредоксина содержит аминокислотную последовательность, выбранную из группы, состоящей из C-X-X-S (SEQ ID NO: 24), C-X-X-X (SEQ ID NO: 17), X-C-X-X-X-X (SEQ ID NO: 19), X-C-G-P-X-X (SEQ ID NO: 21), W-C-G-P-X-K (SEQ ID NO: 23), X-C-X-X-S-X (SEQ ID NO: 25), X-C-G-P-S-X (SEQ ID NO: 26) и W-C-G-P-S-K (SEQ ID NO: 27), при этом остаток C находится в восстановленном состоянии, и при этом указанные остатки X представляют собой любой аминокислотный остаток, кроме цистеина.
28. Фармацевтическая композиция по п. 27, отличающаяся тем, что указанное аэрозольное введение в легкие осуществляют посредством небулайзера.
29. Фармацевтическая композиция по п. 28, отличающаяся тем, что указанный небулайзер представляет собой небулайзер с вибрирующей сеткой.
30. Фармацевтическая композиция по любому из пп. 27-29, отличающаяся тем, что указанная композиция дополнительно содержит восстановитель.
31. Фармацевтическая композиция по п. 27, где композиция составлена для глазного введения или для назального введения.
32. Композиция для поддержания стабильной смешанной дисульфидной связи, содержащая белок или пептид, содержащий моноцистеиновый активный центр тиоредоксина, причем цистеин моноцистеинового активного центра тиоредоксина ковалентно связан с остатком цистеина белка слизи, где моноцистеиновый активный центр тиоредоксина содержит аминокислотную последовательность, выбранную из группы, состоящей из C-X-X-S (SEQ ID NO: 24), C-X-X-X (SEQ ID NO: 17), X-C-X-X-X-X (SEQ ID NO: 19), X-C-G-P-X-X (SEQ ID NO: 21), W-C-G-P-X-K (SEQ ID NO: 23), X-C-X-X-S-X (SEQ ID NO: 25), X-C-G-P-S-X (SEQ ID NO: 26) и W-C-G-P-S-K (SEQ ID NO: 27), при этом указанный остаток C находится в восстановленном состоянии, и при этом остатки X представляют собой любой аминокислотный остаток, кроме цистеина.
33. Композиция по п. 32, отличающаяся тем, что указанный белок слизи выбран из группы, состоящей из белка слизи дыхательных путей, белка слизи половых путей и белка слизи пищеварительного тракта.
34. Композиция по п. 32, отличающаяся тем, что указанный белок слизи представляет собой муцин.
35. Способ снижения вязкоупругости слизи и мокроты у пациента с чрезмерно вязкой или липкой слизью или мокротой, включающий приведение указанной слизи или мокроты указанного пациента в контакт с композицией, содержащей восстановитель дисульфидных связей и агент, блокирующий цистеин, где восстановитель дисульфидных связей и/или агент, блокирующий цистеин, представляет собой белок или пептид, содержащий моноцистеиновый активный центр тиоредоксина, где моноцистеиновый активный центр тиоредоксина содержит аминокислотную последовательность, выбранную из группы, состоящей из C-X-X-S (SEQ ID NO: 24), C-X-X-X (SEQ ID NO: 17), X-C-X-X-X-X (SEQ ID NO: 19), X-C-G-P-X-X (SEQ ID NO: 21), W-C-G-P-X-K (SEQ ID NO: 23), X-C-X-X-S-X (SEQ ID NO: 25), X-C-G-P-S-X (SEQ ID NO: 26) и W-C-G-P-S-K (SEQ ID NO: 27), при этом указанный остаток C находится в восстановленном состоянии, и при этом остатки X представляют собой любой аминокислотный остаток, кроме цистеина.
36. Способ по п. 35, отличающийся тем, что указанный восстановитель дисульфидных связей и указанный агент, блокирующий цистеин, представляют собой одну молекулу.
37. Способ по п. 36, отличающийся тем, что указанная молекула является белком или пептидом, содержащим моноцистеиновый активный центр тиоредоксина.
38. Способ по п. 35, отличающийся тем, что указанный восстановитель дисульфидных связей и указанный агент, блокирующий цистеин, являются различными молекулами.
39. Способ по п. 35, отличающийся тем, что указанный восстановитель дисульфидных связей выбран из группы, состоящей из дитиотреитола (ДТТ), этилендиаминтетрауксусной кислоты (ЭДТА), глутатиона, дитиогликолевой кислоты, 2-меркаптоэтанола, N-ацетилцистеина и трис-(2-карбоксиэтил)фосфена.
40. Способ по п. 35, отличающийся тем, что указанный агент, блокирующий цистеин, выбран из группы, состоящей из иодацетамида, иодуксусной кислоты и алкилирующих агентов.
41. Способ лечения пациента с чрезмерно вязкой или липкой слизью путем введения указанному пациенту композиции, содержащей соединение, имеющее активный центр тиоредоксина, неспособное к поглощению клетками, где моноцистеиновый активный центр тиоредоксина содержит аминокислотную последовательность, выбранную из группы, состоящей из C-X-X-S (SEQ ID NO: 24), C-X-X-X (SEQ ID NO: 17), X-C-X-X-X-X (SEQ ID NO: 19), X-C-G-P-X-X (SEQ ID NO: 21), W-C-G-P-X-K (SEQ ID NO: 23), X-C-X-X-S-X (SEQ ID NO: 25), X-C-G-P-S-X (SEQ ID NO: 26) и W-C-G-P-S-K (SEQ ID NO: 27), при этом указанный остаток C находится в восстановленном состоянии, и при этом остатки X представляют собой любой аминокислотный остаток, кроме цистеина, где соединение выбрано из группы, состоящей из белка или пептида, содержащего моноцистеиновый активный центр тиоредоксина, гибридного белка, содержащего фрагмент тиоредоксина и фрагмент лиганда рецептора клеточной поверхности, и комбинации белка или пептида, содержащего активный центр тиоредоксина, и соединения, блокирующего цистеин, соответствующий цистеину в положении 35 SEQ ID NO: 12.
42. Способ предотвращения системного воздействия лекарственного вещества на пациента после доставки путем, выбранным из группы, состоящей из пульмонарной, пероральной и местной доставки, посредством введения лекарственного вещества, содержащего белок или пептид, содержащий моноцистеиновый активный центр тиоредоксина, в восстановленном состоянии, где моноцистеиновый активный центр тиоредоксина содержит аминокислотную последовательность, выбранную из группы, состоящей из C-X-X-S (SEQ ID NO: 24), C-X-X-X (SEQ ID NO: 17), X-C-X-X-X-X (SEQ ID NO: 19), X-C-G-P-X-X (SEQ ID NO: 21), W-C-G-P-X-K (SEQ ID NO: 23), X-C-X-X-S-X (SEQ ID NO: 25), X-C-G-P-S-X (SEQ ID NO: 26) и W-C-G-P-S-K (SEQ ID NO: 27), при этом указанный остаток C находится в восстановленном состоянии, и при этом остатки X представляют собой любой аминокислотный остаток, кроме цистеина, образующего ковалентную связь со своей областью-мишенью, где лекарственное вещество представляет собой тиолсодержащее лекарственное вещество.
43. Способ по любому из предыдущих пп. 35-42, отличающийся тем, что указанный пациент является позвоночным, выбранным из группы, состоящей из млекопитающих и птиц.
44. Способ по п. 43, отличающийся тем, что указанный пациент является человеком.
45. Способ по п. 43, отличающийся тем, что указанный пациент является курицей или индейкой.
46. Способ по п. 43, где пациента выбирают из группы, состоящей из млекопитающих и домашних животных.
47. Фармацевтическая композиция для превращения дисульфидной связи белка-мишени в стабильную смешанную дисульфидную связь, содержащая белок или пептид, содержащий моноцистеиновый активный центр тиоредоксина в восстановленном состоянии, где моноцистеиновый активный центр тиоредоксина содержит аминокислотную последовательность, выбранную из группы, состоящей из C-X-X-S (SEQ ID NO: 24), C-X-X-X (SEQ ID NO: 17), X-C-X-X-X-X (SEQ ID NO: 19), X-C-G-P-X-X (SEQ ID NO: 21), W-C-G-P-X-K (SEQ ID NO: 23), X-C-X-X-S-X (SEQ ID NO: 25), X-C-G-P-S-X (SEQ ID NO: 26) и W-C-G-P-S-K (SEQ ID NO: 27), при этом указанный остаток C находится в восстановленном состоянии, и при этом остатки X представляют собой любой аминокислотный остаток, кроме цистеина, и дополнительно содержащая по меньшей мере один сахарид или производное сахарида, способное стабилизировать редокс-активную тиоловую группу, где сахарид или производное сахарида выбрано из группы, состоящей из сахарозы, сукралозы, лактозы, трегалозы, мальтозы, галактозы, рафинозы, маннозы или маннита.
48. Кормовая композиция для животных, содержащая белок или пептид, содержащий моноцистеиновый активный центр тиоредоксина в восстановленном состоянии, где моноцистеиновый активный центр тиоредоксина содержит аминокислотную последовательность, выбранную из группы, состоящей из C-X-X-S (SEQ ID NO: 24), C-X-X-X (SEQ ID NO: 17), X-C-X-X-X-X (SEQ ID NO: 19), X-C-G-P-X-X (SEQ ID NO: 21), W-C-G-P-X-K (SEQ ID NO: 23), X-C-X-X-S-X (SEQ ID NO: 25), X-C-G-P-S-X (SEQ ID NO: 26) и W-C-G-P-S-K (SEQ ID NO: 27), при этом указанный остаток C находится в восстановленном состоянии, и при этом остатки X представляют собой любой аминокислотный остаток, кроме цистеина.
49. Применение композиции, содержащей белок или пептид, содержащий моноцистеиновый активный центр тиоредоксина в восстановленном состоянии, для уменьшения вязкоупругости слизи или мокроты у пациента с избыточно вязкой или липкой слизью или мокротой, причем контакт указанной слизи или указанной мокроты указанного пациента с указанной композицией снижает вязкость указанных слизи или мокроты по сравнению с вязкостью до контакта, где моноцистеиновый активный центр тиоредоксина содержит аминокислотную последовательность, выбранную из группы, состоящей из C-X-X-S (SEQ ID NO: 24), C-X-X-X (SEQ ID NO: 17), X-C-X-X-X-X (SEQ ID NO: 19), X-C-G-P-X-X (SEQ ID NO: 21), W-C-G-P-X-K (SEQ ID NO: 23), X-C-X-X-S-X (SEQ ID NO: 25), X-C-G-P-S-X (SEQ ID NO: 26) и W-C-G-P-S-K (SEQ ID NO: 27), при этом указанный остаток C находится в восстановленном состоянии, и при этом остатки X представляют собой любой аминокислотный остаток, кроме цистеина.
| US 2009311231 A1, 17.12.2009 | |||
| US 20069148057 A1, 06.07.2006 | |||
| US 3683069 A, 08.08.1972 | |||
| СПОСОБ ЛЕЧЕНИЯ ХРОНИЧЕСКОГО БРОНХОЛЕГОЧНОГО ПРОЦЕССА ПРИ ВРОЖДЕННЫХ ПОРОКАХ РАЗВИТИЯ ЛЕГКИХ У ДЕТЕЙ | 2003 |
|
RU2253484C1 |
| ОКОВИТЫЙ С.В | |||
| и др | |||
| Фармакологические подходы к противокашлевой терапии// РМЖ, Болезни дыхательных путей, 2011, No 23 | |||
| BALIGA V | |||
| et al | |||
| Efficacy, safety and tolerability of salbutamol + guaiphenesin + bromhexine (Ascoril) expectorant | |||

