Область техники
Изобретение относится к биотехнологии и биохимии, а именно к гуманизированному моноклональному антителу, селективно связывающемуся с опухолевым антигеном PRAME человека, с константой диссоциации комплекса 1,2×10-9 М, а также изолированному фрагменту ДНК, кодирующему участки легкой и тяжелой цепи указанного антитела, и антиген-связывающему фрагменту указанного моноклонального антитела. Специфичность моноклонального антитела к опухолевому антигену PRAME человека подтверждена методами иммуноблоттинга и измерения константы диссоциации комплекса антитела с антигеном с использованием биосенсора в реальном времени.
Уровень техники
Антиген PRAME, представляющий собой значимую мишень для моноклональных антител, является онкоспецифическим маркером, который активен на всех стадиях дифференцировки опухолевых клеток, и вызывает спонтанный T-клеточный ответ. Иммуногенность опухолевых клеток во многом опосредована активностью раково-тестикулярных антигенов. Эти антигены экспрессируются в различных тканях эмбриона, но неактивны у взрослого человека, за исключением клеток гамет. Поскольку гаметы расположены в иммунопривилегированных областях организма человека, раково-тестикулярные антигены не могут быть представлены иммунной системе.
Белок PRAME был открыт при исследовании причин спонтанной ремиссии меланомы у человека. Опухолевые клетки больного LB33, из которых была получена клеточная линия, подвергались атаке аутологичных CD8-позитивных T-клеток. Белок PRAME состоит из 509 аминокислотных остатков, в его структуре было выявлено большое количество потенциальных T-клеточных эпитопов. Наиболее изученным является взаимодействие пептидов белка PRAME с молекулой главного комплекса гистосовместимости HLA-A2, которая является наиболее распространенным вариантом в генотипе европейцев. Из множества предсказанных пептидов 19 демонстрировали высокую аффинность связывания с молекулой HLA-A2 [ Kessler J.H., Beekman N.J., Bres-Vloemans S.A., Verdijk P., van Veelen P.A., Kloosterman-Joosten A.M., Vissers D.C., ten Bosch G.J., Kester M.G., Sijts A., Wouter Drijfhout J., Ossendorp F., Offringa R., and Melief C.J. (2001) Efficient identification of novel HLA-A(*)0201-presented cytotoxic T lymphocyte epitopes in the widely expressed tumor antigen PRAME by proteasome-mediated digestion analysis. J Exp Med. 193(1): p. 73-88]. Большое разнообразие иммуногенных эпитопов делает белок PRAME очень привлекательным для использования в терапии. При сопоставлении иммуногенности белков PRAME и BCR-ABL, характерного для хронического миелоидного лейкоза, последний показал очень низкую иммуногенность. Объясняется это тем, что BCR-ABL образован двумя белками, которые экспрессируются в соматических тканях человека и поэтому являются неиммуногенными. Однако остальные известные раково-тестикулярные антигены (РТА), такие как MAGE-A3, SSXIP2, HAGE и некоторые другие, реже по сравнению с PRAME вызывают T-клеточный иммунный ответ [Greiner J., Schmitt M., Li L., Giannopoulos K., Bosch K., Schmitt A., Dohner K., Schlenk R.F., Pollack J.R., Dohner H., and Bullinger L. (2006) Expression of tumor-associated antigens in acute myeloid leukemia: Implications for specific immunotherapeutic approaches. Blood. 108(13): p. 4109-4117.]; [ Schneider V., Zhang L., Rojewski M., Fekete N., Schrezenmeier H., Erle A., Bullinger L., Hofmann S., Gotz M., Dohner K., Ihme S., Dohner H., Buske C., Feuring-Buske M., and Greiner J. (2015) Leukemic progenitor cells are susceptible to targeting by stimulated cytotoxic T cells against immunogenic leukemia-associated antigens. Int J Cancer. 137(9): p. 2083-2092.]; [ Babiak A., Steinhauser M., Gotz M., Herbst C., Dohner H., and Greiner J. (2014) Frequent T cell responses against immunogenic targets in lung cancer patients for targeted immunotherapy. Oncol Rep. 31(1): p. 384-390.]. Белок PRAME относится к группе РТА, экспрессирующихся в основном в клетках гамет и в опухолевых клетках.
При меланоме и некоторых других солидных опухолях экспрессия PRAME в основном ассоциирована с метастазированием и лекарственной устойчивостью, а также с неблагоприятным исходом заболевания [ Ikeda H., Lethe B., Lehmann F., van Baren N., Baurain J.F., de Smet C., Chambost H., Vitale M., Moretta A., Boon T., and Coulie P.G. (1997) Characterization of an antigen that is recognized on a melanoma showing partial HLA loss by CTL expressing an NK inhibitory receptor. Immunity. 6(2): p. 199-208.]; [ Greiner J., Ringhoffer M., Simikopinko O., Szmaragowska A., Huebsch S., Maurer U., Bergmann L., and Schmitt M. (2000) Simultaneous expression of different immunogenic antigens in acute myeloid leukemia. Exp Hematol. 28(12): p. 1413-1422.]. При солидных опухолях негативное влияние на прогноз заболевания объясняется функциями белка. Так, PRAME блокирует сигнальный путь ретиноевой кислоты, в результате чего останавливается дифференцировка клеток [Epping M.T., Wang L., Edel M.J., Carlee L., Hernandez M., and Bernards R. (2005) The human tumor antigen PRAME is a dominant repressor of retinoic acid receptor signaling. Cell. 122(6): p. 835-847.]. Экспрессия PRAME наблюдается при меланоме (100% случаев), раке головы и шеи (в 39% случаев), мелкоклеточном раке легких (25%), аденокарциноме легких (46%), раке почки (61%), раке простаты (10%), раке мочевого пузыря (9%), раке щитовидной железы (60%), cаркомах (29%), хроническом лимфоидном лейкозе (16-50%), нейробластоме (67%), медуллабластоме (30%), детских острых милеомных лейкозах (75%), множественной миеломе (25%), неходжкинских лимфомах, в том числе при лимфогрануломатозе (лимфоме Ходжкина) (67%). Клетки Березовского - опухолевый субстрат лимфомы Ходжкина.
Из уровня техники известны антитела, связывающиеся с антигеном PRAME, описанные в патенте США № 8846872 B2, опубл. 30.09.2014 года. Данные антитела являются мышиными мАТ, полученными с использованием гибридомной технологии. Данное изобретение относится к антителам к преимущественно экспрессируемому антигену при меланоме (PRAME) и антигенам синовиальной саркомы X breakpoint 2 (SSX-2), способам их применения и диагностическим наборам [https://patents.google.com/patent/US8846872B2/en US 8846872 B2 от 30.09.2014]. Данное изобретение описывает мАТ, либо один или несколько его антиген-связывающих фрагментов, специфично связывающихся с эпитопом, включающим аминокислотные остатки 123-132 белка PRAME. Также изобретение описывет мАТ, либо один или несколько его антиген-связывающих фрагментов, специфично связывающихся с эпитопом, включающим аминокислотные остатки 276-286 белка PRAME.
Известно терапевтическое антитело Pr20, имитирующее T-клеточный рецептор, и связывающее пептид опухоле-ассоциированного белка PRAME в комплексе с молекулой главного комплекса гистосовместимости I класса (HLA-I) [Chang AY, Dao T, Gejman RS, Jarvis CA, Scott A, Dubrovsky L, Mathias MD, Korontsvit T, Zakhaleva V, Curcio M, Hendrickson RC. A therapeutic T cell receptor mimic antibody targets tumor-associated PRAME peptide/HLA-I antigens. The Journal of clinical investigation. 2017 Jun 30;127(7):2705-18]. Описано человеческое антитело подкласса IgG1, связывающее пептид PRAME 300-309 с аминокислотной последовательностью ALYVDSLFFL, представленный в комплексе с HLA-A2 на поверхности опухолевых клеток, экспрессирующих белок PRAME и HLA-A2.
Известны генетически модифицированные T-клетки с T-клеточным рецептором, специфически связывающим пептид белка PRAME в комплексе с HLA-A2. Описано получение таких клеток и их противоопухолевое действие по отношению к клеткам линии медулобластомы (HLA-A2+) in vitro. Также описано терапевтическое действие в модели медуллобластомы in vivo при использовании клеточной линии DAOY и мышей NSG [Orlando D, Miele E, De Angelis B, Guercio M, Boffa I, Sinibaldi M, Po A, Caruana I, Abballe L, Carai A, Caruso S. Adoptive immunotherapy using PRAME-specific T cells in medulloblastoma. Cancer research. 2018 Jun 15;78(12):3337-49]. В отличие от моноклональных антител, генетически модифицированные T-клетки с T-клеточным рецептором, специфически связывающим пептид белка PRAME, связываются только с пептидом в комплексе с молекулой главного комплекса гистосовместимости I класса (MHC I). Для дальнейшей активации таких клеток и элиминации экспрессирующих PRAME опухолевых клеток требуется получение T-клетками костимуляторных сигналов. Однако известно, что опухолевые клетки могут влиять на костимуляторные сигналы и блокировать активацию T-клеток, а также для многих типов опухолей характерно снижение экспрессии MHC I, что помогает опухолевым клеткам маскировать их присутствие от генетически-модифицированных T-клеток. Также получение генетически модифицированных T-клетки с T-клеточным рецептором, специфически связывающим пептид белка PRAME, является трудоемким и очень дорогим процессом, требующим выделения аутологичных T-клеток из крови пациента, их генетической модификации и выращивания ex vivo.
Ряд исследований показывает [R. Fagnani, Immunol. Ser. 61 (1994) 3-22; M.B. Khazaeli, R.M. Conry, A.F. LoBuglio, J Immunother. 15 (1994) 42-52; K. Kuus-Reichel, L.S. Grauer, L.M. Karavodin, C. Knott, M. Krusemeier, N.E. Kay, Clin. Diagn. Lab. Immunol. 1 (1994) 365-372], что введение чужеродного антитела может вызывать сильный иммунный ответ у пациента, прежде всего образование анти-мышиных антител (HAMA - human anti-mose antibody). В результате мышиные антитела оказываются нейтрализованы человеческими и довольно быстро выводятся из организма, что в конечном итоге приводит к ограниченной эффективности терапии уже на ранней стадии лечения. Более того, при использовании мышиных или других чужеродных (для человека) антител для лечения различных заболеваний, последующее лечение другими мышиными антителами может быть неэффективно или даже опасно из-за кросс-реактивности используемых антител.
Для снижения иммуногенности мышиных мАТ и возможных неблагоприятных реакций, обусловленных иммуногенностью, антитела или их части могут быть гуманизированы для того, чтобы быть менее иммуногенными, чем их мышиные прототипы. Клинические исследования показали, что гуманизированные антитела в целом являются значительно менее иммуногенными, чем мышиные и химерные мАТ, более безопасны и лучше переносятся пациентами [C. Mateo, E. Moreno, K. Amour, J. Lombardero, W. Harris, R. Perez, Immunotechnology 3 (1997) 71-81; S. Stephens, S. Emtage, O. Vetterlein, L. Chaplin, C. Bebbington, A. Nesbitt, M. Sopwith, D. Athwal, C. Novak, M. Bodmer, Immunology 85 (1995) 668-674]. Так, авторы настоящего изобретения сконструировали на основе мышиного мАТ, обладающего способностью с высокой специфичностью и высоким аффинитетом связываться с антигеном PRAME человека, и каркасных участков антител человека гуманизированное антитело к белку PRAME. Для гуманизации мАТ 6H8 использовали метод пересадки CDR-участков. Этот метод достаточно широко используется для получения гуманизированных терапевтических мАТ. Хотя известно, что пересадка CDR-участков была успешна в некоторых случаях, было показано, что большинство гуманизироваванных антител, полученных таким методом, не сохраняют антиген-связывающие свойства (аффинность) по отношению к антигену по сравнению с родительскими мышиными антителами. Потеря аффинности связана с заменой определенных аминокислотных остатков каркасных областей мАТ, тесно взаимодействующих с аминокислотными остатками CDR-участков в вариабельных доменах, и, таким образом влияют на структуру антиген-связывающего сайта. Поэтому перенос только CDR-участков при гуманизации антител часто приводит к потере антиген-связывающих свойств [C. Queen, W.P. Schneider, H.E. Selick, P.W. Payne, N.F. Landolfi, J.F. Duncan, N.M. Avdalovic, M. Levitt, R.P. Junghans, T.A. Waldmann, Proc. Natl. Acad. Sci. USA 86 (1989) 10029-10033]. Queen с соавторами предположили наличие ключевых аминокислотных остатков в каркасных областях, взаимодействующих с CDR-участками и, таким образом, влияющих на правильную структуру антигенсвязывающего сайта. Такие остатки по мнению авторов, должны быть перенесены в гуманизированную версию антитела наряду с CDR-участками мышиных мАТ. Для того, чтобы установить, какие из аминокислотных остатков являются ключевыми, используют полученные in silico трехмерные модели мАТ и различные методы анализа. Такие методы часто позволяют установить аминокислотные остатки, замена которых в гуманизированном антителе приводит к значительному изменению его структуры. Однако, такие методы не всегда могут предсказать ключевые аминокислотные остатки. Очень важным этапом процесса гуманизации мышиных антител является построение надежных трехмерных моделей вариабельных доменов антитела. Также необходимо выбрать источник человеческих последовательностейй каркасных областей мАТ, на которые будет осуществляться пересадка CDR-участков мышиного мАТ. Источником таких последовательностей могут быть как мАТ с известной первичной последовательностью, так и гены зародышевой линии антител человека. Преимуществом использования последовательностей известных мАТ является доступность их пространственных моделей, полученных на основе экспериментальных данных (рентгено-структурного анализа). В то же время использование генов зародышевой линии позволяет избежать потенциальной иммуногенности, связанной с гиперсоматичекими мутациями, которые присутствуют в генах известных ранее антител. При гуманизации антител также могут быть использованы консенсусные последовательности каркасных областей вариабельных доменов антител.
Авторы настоящего изобретения использовали гены зародышевой линии антител человека для гуманизации мышиного мАТ 6H8. С помощью сервиса ROSETTA были построены, а затем оптимизированы пространственные модели мышиного и различных вариантов гуманизированного мАТ 6H8. В результате были выявлены 3 ключевых аминокислотных остатка каркасных областей мышиного антитела (2 в тяжелой цепи мАТ и 1 в легкой цепи), необходимых для поддержания структуры антиген-связывающего центра мАТ 6H8. Совместная пересадка CDR-участков с четырьмя ключевыми аминокислотными остатками каркасных областей мышиного мАТ 6H8 позволила получить авторам изобретения гуманизированное мАТ 6H8Hu, характеризующееся аффинностью, не уступающей родительскому мышиному мАТ.
Наиболее близким к заявляемому изобретению (прототипом) является мышиное мАТ 6H8, селективно связывающееся с опухолевым антигеном PRAME человека, описанное в статье [В. А. Мисюрин, Ю. П. Финашутина, А. А. Турба, М. В. Ларина, О. Н. Солопова, Н. А. Лыжко, Л. А. Кесаева, Н. Н. Касаткина, Т. К. Алиев, А. В. Мисюрин, М. П. Кирпичников. (2019) Эпитопный анализ мышиных и химерных моноклональных антител, распознающих раково-тестикулярный антиген PRAME. Доклады академии наук, Том 492, номер. 1, стр. 135-138]. В статье описано мАТ, связывающееся с антигеном PRAME, полученное с помощью гибридомной технологии при иммунизации мышей рекомбинантным белком PRAME. Данное антитело характеризуется высокой аффинностью 1.2 нМ по отношению к антигену PRAME. Однако, использование моноклональных антител животного происхождения имеет ряд недостатков при терапии заболеваний человека, особенно выраженных при повторных введениях препарата. Например, мышиные моноклональные антитела имеют короткое время циркуляции в организме человека, а также вследствие неспособности их Fc-фрагмента связываться с человеческими рецепторами на поверхности эффекторных клеток не обладают важными функциональными характеристиками. Так, например, такие антитела не могут обеспечивать развитие антитело-зависимой клеточной цитотоксичности, которая является важным механизмом разрушения опухолевых клеток.
Также важной особенностью мышиных моноклональных антител является то, что они содержат аминокислотные последовательности, которые являются иммуногенными для человека.
Раскрытие изобретения
Технической проблемой, решаемой настоящим изобретением, было получение гуманизированного моноклонального антитела, способного к селективному связыванию с опухолевым антигеном PRAME человека, и отличающегося по аминокислотной последовательности вариабельных доменов легкой и тяжелой цепей от известных из предшествующего уровня техники моноклональных антител к антигену PRAME человека и обладающих низкой иммуногенностью для человека по сравнению с мышиными моноклональными антителами.
А также получение изолированных фрагментов ДНК, кодирующих участки легкой и тяжелой цепей указанного антитела и антигенсвязывающего фрагмента указанного гуманизированного моноклонального антитела.
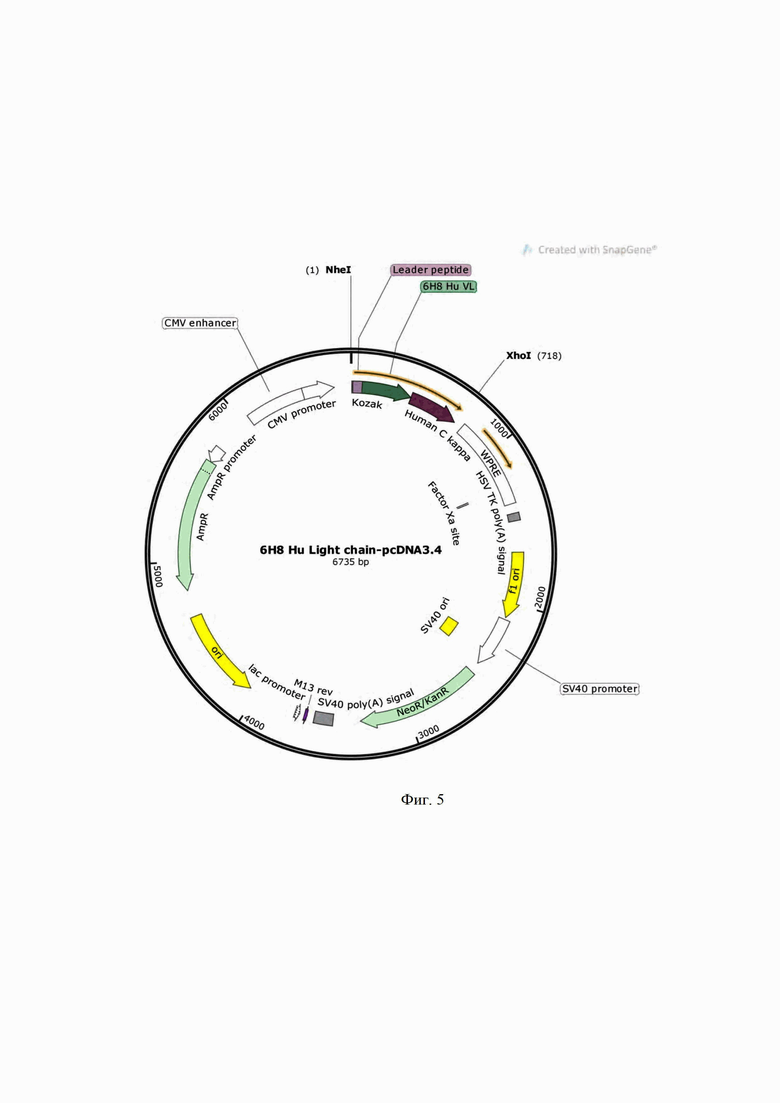
Техническим результатом является получение нового гуманизированного моноклонального антитела IgG1-изотипа, способного к связыванию с опухолевым антигеном PRAME человека, и характеризующегося константой диссоциации комплекса антиген-антитело 1,2 нМ с допустимой величиной погрешности 10%, что не уступает константе диссоциации ближайшего аналога, мышиного мАТ 6H8. При этом гуманизированное мАТ 6H8Hu можно использовать для лечения человека.
Техническая проблема решается получением гуманизированного мАТ 6H8Hu, селективно связывающего белок PRAME человека, включающего вариабельный домен тяжелой цепи иммуноглобулина (VH), который содержит последовательность гипервариабельных регионов CDRH1, CDRH2 и CDRH3, и вариабельный домен легкой цепи иммуноглобулина (VL), который содержит последовательность гипервариабельных регионов CDRL1, CDRL2 и CDRL3. При этом, гуманизированное антитело включает вариабельный участок тяжелой цепи (VH) указанного антитела содержащего последовательность аминокислот, не менее чем на 90% гомологичную SEQ ID NO:1; а вариабельный участок легкой цепи (VL) указанного антитела содержит последовательность аминокислот, не менее чем на 90% гомологичную SEQ ID NO:2, а также фрагменты ДНК, кодирующие вариабельные фрагменты легкой и тяжелой цепей указанного антитела.
Техническая проблема решается также тем, что получен фрагмент ДНК, кодирующий антитело или его фрагмент. Изолированный фрагмент ДНК, кодирующий VH указанного антитела, с нуклеотидной последовательностью SEQ ID NO: 3. Изолированный фрагмент ДНК, кодирующий VL указанного антитела, с нуклеотидной последовательностью SEQ ID NO: 4.
Техническая проблема решается также тем, что получен антиген-связывающий фрагмент указанного гуманизированного антитела, представляющий собой комбинацию вариабельного участка тяжелой цепи (VH) указанного антитела с последовательностью аминокислот, не менее чем на 90% гомологичную SEQ ID NO: 1, и вариабельного участка легкой цепи (VL) указанного антитела с последовательностью аминокислот, не менее чем на 90% гомологичную SEQ ID NO: 2.
Краткое описание чертежей
На Фиг. 1 показано расположение участков CDRs в вариабельном домене легкой 6H8HuL и тяжелой 6H8HuH цепей гуманизированного мАТ 6H8Hu. Участки CDRs подчеркнуты.
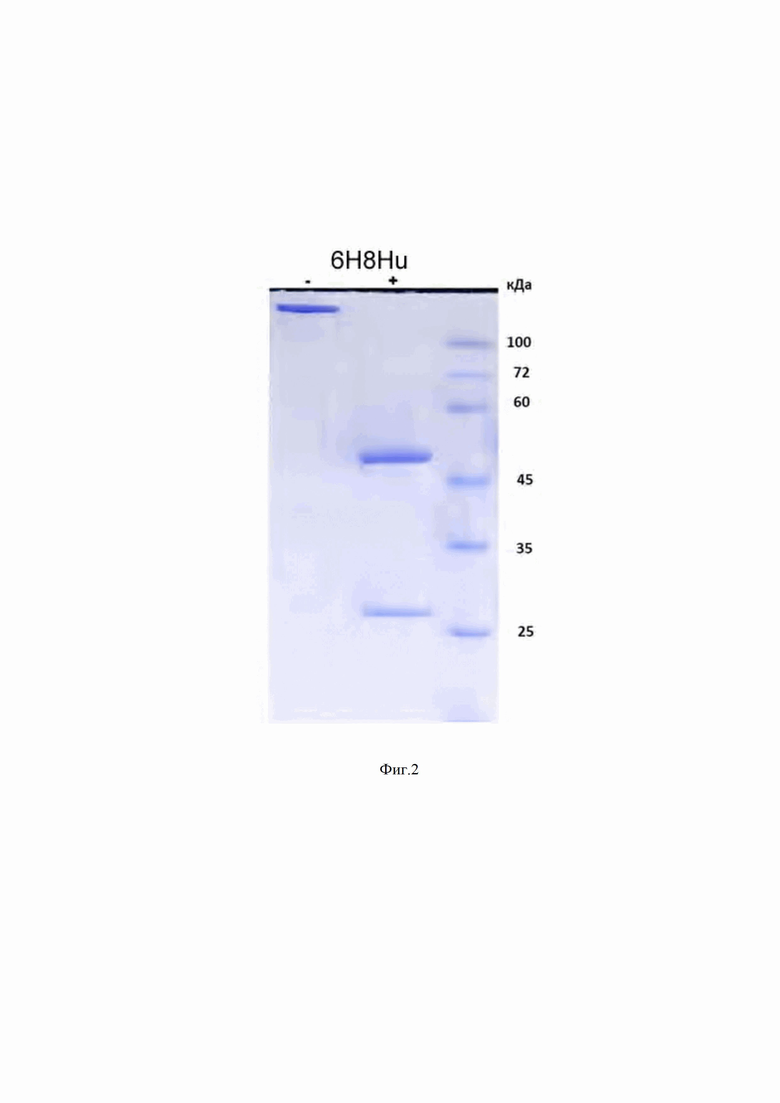
На Фиг. 2 показана гель-электрофореграмма гуманизированного мАТ 6H8Hu в 10%-м полиакриламидном геле в присутствии додецилсульфата натрия (SDS), а также без или в присутствии β-меркаптоэтанола; М - белковые маркеры молекулярного веса, (-) - мАТ 6H8Hu без β-меркаптоэтанола, (+) - мАТ 6H8Hu в присутствии β-меркаптоэтанола.
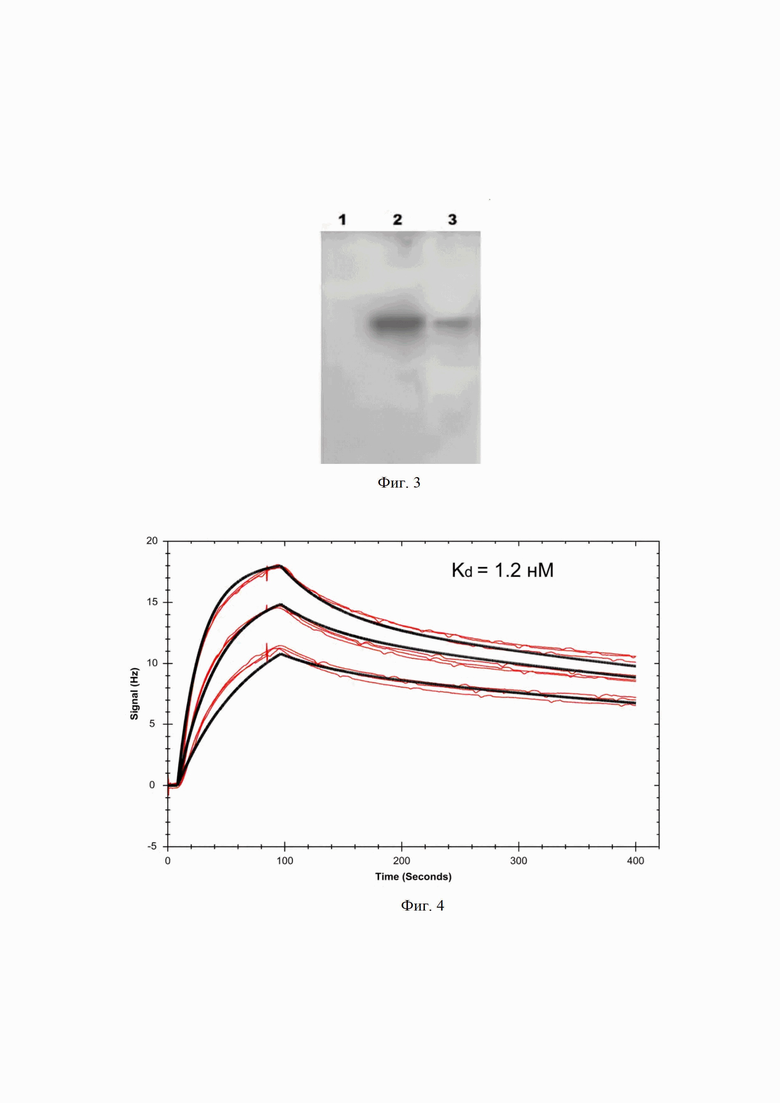
На Фиг. 3 показан иммуноблот связывания гуманизированного мАТ 6H8Hu. 1 - рекомбинантный белок NY-ESO-1, 2 - рекомбинантный белок PRAME в восстанавливающих условиях, 3 - рекомбинантный белок PRAME в невосстанавливающих условиях.
На Фиг. 4 показано измерение аффинности гуманизированного мАТ 6H8Hu по отношению к рекомбинантному белку PRAME. Измерение проводили на биосенсоре Attana Cell A200.
На Фиг. 5 показана схема экспрессионной плазмиды, кодирующей легкую цепь гуманизированного мАТ 6H8Hu.
На Фиг. 6 показана схема экспрессионной плазмиды, кодирующей тяжелую цепь гуманизированного мАТ 6H8Hu.
Осуществление изобретения
Как хорошо известно, минорные изменения аминокислотной последовательности, такие как делеция, добавление или замена одной, небольшого количества или даже нескольких аминокислот, могут приводить к получению аллельной формы исходного белка, которая обладает практически идентичными свойствами.
Антитела обычно состоят из двух тяжелых цепей, связанных между собой дисульфидными связями, и легких цепей, ассоциированными с N-концом каждой из тяжелых цепей. Каждая тяжелая цепь содержит на N-конце вариабельный домен с константным доменом на другом конце. Каждая легкая цепь содержит на N-конце вариабельный домен с константным доменом на другом конце. Вариабельные домены каждой пары легкой и тяжелой цепей образуют антигенсвязывающий участок. Вариабельные домены легкой и тяжелой цепей обладают похожей общей структурой, и каждый домен включает каркас из четырех участков, последовательности которых являются относительно консервативными, связанных посредством трех участков, определяющих комплементарность (complementarity determining regions, CDRs). Четыре каркасных участка формируют конформацию типа бета-складчатого слоя. Участки CDRs расположены в близком соседстве друг с другом благодаря каркасным участкам и вносят вклад в образование антигенсвязывающего участка. Участки CDRs и каркасные участки антител могут быть определены путем ссылки на нумерационную систему Кабата [Kabat numbering system, Kabat et al., 1987 “Sequences of Proteins of Immunological Interest”, US Dept. of Health and Human Services, US Government Printing Office] в сочетании c данными рентгеноструктурного анализа, как указано в заявке WO 91/09967. Участки CDRs и каркасные участки антител могут также быть определены по номенклатуре Международной информационной системы по иммуногенетике (International Immunogenetics Information System, www.imgt.org).
Если не указано иное, любая представленная в описании полипептидная цепь имеет аминокислотную последовательность, которая начинается на N-конце и заканчивается на С-конце. Если антигенсвязывающий центр содержит и VH-, и VL-области, то они могут быть расположены на одной и той же полипептидной молекуле или предпочтительно каждая область может располагаться на разных цепях, VH-область может быть частью тяжелой цепи иммуноглобулина или ее фрагмента, а VL-область - частью легкой цепи иммуноглобулина или ее фрагмента. Под иммуноглобулином в данном изобретении понимается иммуноглобулин, относящийся к следующим классам: IgG, IgA, IgM, IgD. Предпочтительным является использование тяжелой цепи антитела подкласса IgG1, а легкой цепи каппа-класса. Также могут быть использованы тяжелые цепи подклассов IgG2, IgG3, IgG4. Под фрагментами антител понимают одноцепочечное антитело (вариабельные домены легкой и тяжелой цепей антитела, объединенных с помощью линкерной белковой последовательности в любой ориентации), Fab-фрагмент антитела, (Fab)2- фрагмент антитела, а также любые другие фрагменты, включающие вариабельный участок тяжелой цепи (VH) указанного антитела, содержащий последовательность аминокислот, не менее чем на 90% гомологичную SEQ ID NO: 1, а вариабельный участок легкой цепи (VL) указанного антитела содержит последовательность аминокислот, не менее чем на 90% гомологичную SEQ ID NO: 2.
Последующие примеры приведены для целей объяснения и не ограничивают каким-либо образом рамки настоящего изобретения.
Пример 1. Гуманизация моноклонального антитела 6H8 и биосинтез полученного гуманизированного моноклонального антитела 6H8Hu в клетках эукариот.
Гуманизированное моноклональное антитело 6H8Hu получают на основе мышиного моноклонального антитела 6H8, полученного путем иммунизации мышей Balb/C человеческим рекомбинантным белком PRAME, экспрессированным в E.coli, дальнейшего получения и селекции линий гибридом, продуцирующих моноклональные антитела к рекомбинантному белку PRAME человека, анализа аффинности и специфичности отобранного моноклонального антитела, определения нуклеотидной и аминокислотной последовательности его вариабельных доменов.
Метод «пересадки CDR» заключается в нахождении последовательностей зародышевых линий антител человека, наиболее гомологичных последовательностям моноклональных мышиных антител, и создании гибридных молекул, содержащих каркасные участки антител человека и гипервариабельные участки исходных мышиных антител. Путем сравнения с последовательностями зародышевых линий человека в сервисе IgBLAST (https://www.ncbi.nlm.nih.gov/igblast/) определяют наиболее гомологичные антителу 6H8 зародышевые линии антител человека и отбирают линии с большей степенью гомологии и меньшим числом неблагоприятных замен. Для легкой цепи моноклонального антитела 6H8 наибольшую степень гомологии демонстрирует последовательность линии IGKV1-39*01 для V-сегмента и IGKJ4*02 для J-сегмента, а для тяжелой цепи наиболее гомологичный V-сегмент относится к зародышевой линии IGHV1-2*02 и J-сегмент к линии IGHJ4*01.
Участки CDRs оставляют без изменения, а каркасные участки мышиного антитела заменяют на гомологичные человеческие.
Проводят оптимизацию получаемых последовательностей путем создания моделей пространственной структуры гуманизированного антитела методом моделирования на основании гомологии (веб-сервис Rosetta Antibody). Реализация этого метода включает следующие шаги: 1) поиск шаблонов среди антител с известной пространственной структурой и сходной аминокислотной последовательностью, 2) построение начальных моделей с использованием консервативных фрагментов шаблонов и VH/VL последовательности запроса, 3) улучшение начальных моделей путем подбора оптимальных конформаций основной цепи VH/VL и боковых цепей по всем моделям в целом. Полученные стартовые модели дополнительно оптимизируют с учетом окружающего растворителя, для чего в программе Gromacs (http://www.gromacs.org/) с использованием силового поля charmm36 проводят расчет молекулярной динамики (МД) моделей в водном растворе при температуре 300К и физиологической концентрации NaCl в течение 10 нс. Анализ МД показывает, что модели принимают свои равновесные конформации уже через 1 нс. Полученные траектории МД подвергают кластерному анализу и наиболее представительные конформации для каждой из стартовых моделей в дальнейшем используются в качестве окончательных моделей, анализ которых позволяет сформулировать ряд «возвратных» замен в каркасных участках VH (R70A, T73K, R82aS) и VL (D70Q) гуманизированного моноклонального антитела к опухолевому антигену PRAME человека.
На основании полученных аминокислотных последовательностей вариабельных доменов тяжелой и легкой цепей гуманизированного антитела 6H8Hu (SEQ ID NO: 1 и SEQ ID NO: 2) и путем оптимизации кодонов составляют кодирующие последовательности тяжелой и легкой цепей гуманизированного антитела 6H8Hu (SEQ ID NO: 3 и SEQ ID NO: 4). Аминокислотные последовательности легкой и тяжелой цепей полученного гуманизированного антитела 6H8Hu с отмеченными гипервариабельными участками представлены на Фиг. 1. Кодирующие последовательности тяжелой и легкой цепей гуманизированного антитела 6H8Hu получают методом химико-ферментативного синтеза [Ильина Е.Н., Солопова О.Н., Балабашин Д.С., Ларина М.В., Алиев Т.К., Гребенникова Т.В., Лосич М.А., Зайкова О.Н., Свешников П.Г., Долгих Д.А., Кирпичников М.П. Получение и характеризация нейтрализующего моноклонального антитела против вируса бешенства. Биоорганическая химия. 2019;45(1):58-68]. Последовательности VL и VH мАТ 6H8Hu были синтезированы из перекрывающихся олигонуклеотидов с помощью полимеразно-цепной реакции (ПЦР). Олигонуклеотиды имели длину от 40 до 50 оснований, при этом они попарно перекрываются между собой (10-12 оснований) и имели приблизительно одинаковую температуру отжига (~50°С). Вариабельные домены были объединены с лидерным пептидом и соответствующими константными областями (константным доменом легкой каппа-цепи человека и константной областью антитела IgG1 человека). Также на 5’-конец каждой цепи антитела была добавлена последовательность Козак, важная для инициации трансляции. Все части перед клонированием в экспрессионный вектор объединяют методом SOE-ПЦР (Single overlap extention- ПЦР, ПЦР с перекрывающимися областями). При наработке фрагментов ДНК, кодирующих легкую и тяжелую цепь мАТ, лидерный пептид и последовательность Kozak используют внешние праймеры, содержащие последовательности узнавания эндонуклеаз рестрикции NheI (в прямом праймере) и XhoI (в реверсном праймере). Экспрессионные кассеты, кодирующие легкую и тяжелую цепь мАТ 6H8Hu клонируют отдельно по сайтам NheI/XhoI в экспрессионый вектор pcDNA3.4, под контроль промотора CMV [Ильина Е.Н., Солопова О.Н., Балабашин Д.С., Ларина М.В., Алиев Т.К., Гребенникова Т.В., Лосич М.А., Зайкова О.Н., Свешников П.Г., Долгих Д.А., Кирпичников М.П. Получение и характеризация нейтрализующего моноклонального антитела против вируса бешенства. Биоорганическая химия. 2019;45(1):58-68.]. Также могут быть использованы другие коммерческие экспрессионные векторы, содержащие последовательности промоторов CMV, EF-1alpha и других, позволяющими экспрессировать рекомбинантные белки в клетках млекопитающих. в частности в клетках CHO. Такими плазмидными векторами могут быть плазмиды pcDNA3.3, pMG, pCEP4, pOptiVec и другие. Полученные экспрессионные векторы (кодирующие легкую и тяжелую цепи мАТ 6H8Hu, схемы представлены на Фиг. 5-6) используют для биосинтеза гуманизированного моноклонального антитела 6H8Hu в клетках CHO.
Клетки яичника китайского хомячка CHO DG44 (dhfr -/-) используют для секретируемой экспрессии рекомбинантного гуманизированного моноклонального антитела 6H8Hu. Для этого клетки культивируют в колбах Эрленмейера в CO2-инкубаторе при 37°C, 95% влажности и 8% CO2 в стандартной бессывороточной среде CD DG44 (Invitrogen) с добавлением 200 мМ раствора L-Глутамина до конечной концентрации 8 мМ и содержащей 0,18% (v/v) Pluronic F-68 (Invitrogen, USA) до концентрации 4×106 клеток/мл 24 часа до трансфекции. Во флаконы Эрленмейера объемом 125 мл засевают 30 мл клеточной суспензии (4,5 млн клеток) при постоянном перемешивании на орбитальном шейкере с частотой 130 об/мин и через 20-24 ч проводят трансфекцию с использованием трансфецирующего реагента Lipofectamine-3000 Transfection Kit (Invitrogen, USA) согласно стандартному протоколу производителя [Lipofectamine-3000 Reagent Protocol, https://tools.thermofisher.com/content/sfs/manuals/lipofectamine3000_protocol.pdf]. Плазмидную ДНК добавляют к клеткам в виде ДНК-липосомного преципитата. Культуральный флакон инкубируют при температуре 37°C, 95% влажности, в атмосфере 8% CO2 и непрерывном перемешивании 130 об/мин. После 7-9 дней инкубации культуры осаждают центрифугированием при 1200 оборотах/мин в течение 10 минут. Супернатант дополнительно центрифугируют при 4000 оборотов/мин в течение 15 минут.
Измерение концентрации клеток и их жизнеспособности проводят с использованием 0.04% раствора трипанового синего в камере Горяева. Продуктивность измеряют с помощью иммуноферментного анализа по стандартной методике.
Проводят выделение антитела 6H8Hu из культуральной жидкости на колонке с носителем HiTrap MabSelect SuRe (GE Healtcare) объемом 5 мл. К культуральной жидкости добавляют 0,1 объема 10-кратного Tris-HCl буфера (200 мМ Tris-HCl, 1,5 М NaCl рН 7.2) и наносят на предварительно уравновешенную колонку при скорости потока 2-3 мл/мин при давлении не более 0,5 МПа. Колонку промывают буфером Tris-HCl объемом равным 5 объемам колонки. Элюцию осуществляют с помощью глицинового буфера (0.1 М глицин, рН 3.0). Элюированный раствор нейтрализуют, добавляя 0,1 объема буфера для нейтрализации (1 M Tris pH 8.0). Полученный раствор белка диализуют против фосфатного буфера (0,01 М Na2HPO4, 0.137 М NaCl, 0.0027 М KCl, рН 7,2) и стерилизуют через мембранные фильтры 0.22 мкм. Оценку гомогенности и степени очистки препарата проводят с использованием электрофоретического метода. Дополнительно образцы характеризуют с помощью гель-фильтрации по стандартной методике.
Антитела выделены (Фиг. 2) и их аффинность и специфичность, а также способность ингибировать рост клеточных линий, экспрессирующих белок PRAME человека изучены.
Пример 2. Анализ связывания мышиного моноклонального антитела 6H8 и гуманизированного моноклонального антитела 6H8Hu с опухолевым белком PRAME человека.
Электрофорез белков проводят по методу Лэммли. В лунку 1 10%-го ПААГ наносят 1 мкг рекомбинантного белка NY-ESO-1 (отрицательный контроль) в буфере с 100 мМ DTT в восстанавливающих условиях, в лунку 2 - 1 мкг рекомбинантного белка PRAME в буфере с 100 мМ DTT в восстанавливающих условиях, в лунку 3 - 1 мкг рекомбинантного белка PRAME в буфере без DTT в невосстанавливающих условиях. После электрофореза перенос белков из геля на Hybond-P PVDF мембрану (GE Healthcare, Великобритания) осуществляют методом полусухого электроблоттинга в течение часа. В качестве первичного антитела используют гуманизированное антитело 6H8Hu в концентрации 3 мкг/мл. Затем инкубируют мембрану с вторичным антителом 4G7-HRP (ОАО «ВЦМДЛ», Россия) в разведении 1:50000 в течение 1 ч при комнатной температуре. Иммуноблот представлен на Фигуре 3.
Пример 3. Измерение константы диссоциации (Кд) комплекса гуманизированного мАТ 6H8Hu с рекомбинантным белком PRAME, полученным в E. coli.
Аффинность моноклонального гуманизированного антитела 6H8Hu измеряли на биосенсоре, использующим технологию QCM (quartz crystal microbalance) Attana Cell A200 (Attana, Швеция). Рекомбинантный белок PRAME (40 мкг/мкл) иммобилизуют на LNB-carboxyl сенсорный чип, пришивая белок ковалентно через аминогруппы. Эксперименты по связыванию гуманизированного антитела 6H8Hu с рекомбинантным белком PRAME проводят в буферном растворе HEPES, содержащим 0.005% полисорбата 20 (25 мкл/мин, 22°C). Три серийных разведения гуманизированного антитела 6H8Hu (в концентрации 12.5-3.1 мкг/мл) наносили на чип с иммобилизованным белком PRAME в трех повторах каждая (Фиг. 4) в случайном порядке. Чип регенерировали раствором глицина 10 мМ, pH=1.5 в течение 30 секунд перед каждым циклом связывания. Перед каждой инъекцией антитела проводили инъекцию буфера, которую использовали как референс при обработке результатов с помощью программного обеспечения Attana Attester (Attana, Швеция). Для расчета константы диссоциации используют модель связывания 1:2.
Пример 4.
С целью подтверждения того, что при замене отдельных аминокислотных остатков (а.о.) в каркасных регионах на другие а.о. остатки было получено модифицированное антитело, в котором отдельные а.о. изменены на человеческие. Были получены модифицированные последовательности SEQ ID NO: 1М и SEQ ID NO: 2М, имеющая гомологию не менее чем на 90 % (модифицированные а.о. выделены жирным) с SEQ ID NO: 1 (степень гомологии - 93,1%) и SEQ ID NO: 2 (степень гомологии - 93,8%).
Полученное антитело специфически взаимодействовало с рекомбинантным белком PRAME и значение констант диссоциации комплексов антиген-антитело, равное 1.3 нМ, не отличалось в пределах статистической погрешности от антитела с последовательностями SEQ ID NO:1 и SEQ ID NO:2.
Пример 5. Ингибирование роста клеточных линий, экспрессирующих PRAME с использованием системы xCELLigence.
Анализ влияния гуманизированного антитела 6H8Hu на пролиферацию опухолевых клеток проводили в режиме реального времени с использованием системы xCELLigence.
Изучение изменений пролиферативного потенциала клеток на приборе с использованием системы xCELLigence происходит за счет измерения прибором электрического импеданса. Электрический импеданс, который измеряется прибором при контакте плазматической мембраны клетки с поверхностью электрода, выражается в изменении электрического потенциала клетки. Изменение величины электрического импеданса под действием антител (либо других цитотоксических агентов, например, клеток) показывает изменение клеточного индекса. Данные изменения выражаются в виде графика в логарифмической системе координат в режиме реального времени.
Проводили серию экспериментов по изучению изменений клеточного индекса линии меланомы Mel P совместной инкубации с гуманизированным антителом 6H8Hu с конечной концентрацией в лунке 100 мкг/мл.
В качестве отрицательного контроля использовали гуманизированное мАТ 1C5Hu (специфичное к гликопротеину вируса бешенства) с конечной концентрацией в лунке 100 мкг/мл и раствор фосфатно-солевого буфера. Клетки линии MelP снимали с подложки с помощью среды RPMI 1640 (Sigma Aldrich, США) и ресуспендировали в той же среде. Затем подсчитывали концентрации клеток в камере Горяева, клетки засевали в лунки 16 луночного культурального планшета с концентрацией клеток 25000/мл для линии MelP c содержанием фетальной бычьей сыворотки равной 10%. Время инкубации клеток в лунках прибора xCELLingence составляло 24 часа при условии температуры +37С° и содержании СО2, равном 5%. После 24 часов инкубации клеток очередной цикл работы прибора приостанавливали. К клеткам в лунках культурального 16-луночного планшета добавляли гуманизированное антитело 6H8Hu и контрольное мАТ 1C5Hu (специфичное к гликопротеину вируса бешенства).
Полученные результаты эксперимента по совместной инкубации антител с клетками линии Mel P и изменения ее клеточного индекса показали способность гуманизированного мАТ 6H8Hu к антигену PRAME ингибировать пролиферацию меланомной клеточной линии MelP, экспрессирующей белок PRAME.
Пример 6. Фрагменты ДНК, кодирующий вариабельные тяжелую и легкую цепи гуманизированного мАТ, селективно связывающего антиген PRAME, содержащего последовательность аминокислот, гомологичную SEQ ID NO: 1 и последовательность аминокислот, гомологичную SEQ ID NO: 2 приведены в перечне последовательностей - SEQ ID NO: 3 и SEQ ID NO: 4.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Федеральное государственное бюджетное образовательное учреждение высшего
профессионального образования "Московский государственный университет имени
М.В.Ломоносова" (МГУ)
<120> ГУМАНИЗИРОВАННОЕ АНТИТЕЛО 6H8HU, СВЯЗЫВАЮЩЕЕСЯ С ОПУХОЛЕВЫМ АНТИГЕНОМ
PRAME, ФРАГМЕНТЫ ДНК, КОДИРУЮЩИЕ УКАЗАННОЕ АНТИТЕЛО И АНТИГЕНСВЯЗЫВАЮЩИЙ
ФРАГМЕНТ АНТИТЕЛА
<160> SEQ ID NО:1
<210> 1
<211> 117
<212> PRT
<213> Artificial sequence
<223> Variable domain of heavy chain of monoclonal humanized antibody
6H8Hu to human antigen PRAME
<400> 1
1 QVQLVQSGAE VKKPGASVKV SCKASGYTFT TSWMHWVRQA PGQGLEWMGY
51 INPNSDYTKY NQKFKDRVTM TADKSISTAY MELSSLRSDD TAVYYCARRG
101 PLALDYWGQG TLVTVSS
<160> SEQ ID NО:1M
<400> 1M
1 EVQLVETGSG IVQPGGSLRL SCAASGITFS DYGMVWVRQA PGKALEWVSY
51 ISSGSSTIYY ADTVKGRFTI SRDNAKNSLY LQMNTIRAED SAVYYCAHYY
101 GFGFSYWGQG TLVTVAS
<160> SEQ ID NО:2
<210> 1
<211> 107
<212> PRT
<213> Artificial sequence
<223> Variable domain of light chain of monoclonal humanized antibody
6H8Hu to human antigen PRAME
<400> 2
1 DIQMTQSPSS LSASVGDRVT ITCRASENIY SNLAWYQQKP GKAPKLLIYT
51 ATNLADGVPS RFSGSGSGTQ FTLTISSLQP EDFATYYCQH FWGTPLTFGG
101 GTKVEIK
<160> SEQ ID NО:2M
<400> 2M
1 DVVMTQTPLS LPVSLGQPAS ISCRSSQSLV HSNGNTYSHW FQQRPGQSPR
51 LLIYKVSNRF SGVPDRFTGS ASGTDFTLKL TRVEAEDVGV YYCSQSTHVP
101 FTFGQGSKVE IK
<160> SEQ ID NО:3
<210> 1
<211> 351
<212> DNA
<213> Artificial sequence
<400> 3
1 CAGGTGCAGC TGGTGCAGTC TGGGGCTGAG GTGAAGAAGC CTGGGGCCTC
51 AGTGAAGGTC TCCTGCAAGG CTTCTGGATA CACCTTCACC ACCTCCTGGA
101 TGCACTGGGT GCGACAGGCC CCTGGACAAG GGCTTGAGTG GATGGGATAC
151 ATCAATCCTA ACAGTGATTA TACTAAATAC AATCAGAAGT TCAAGGACAG
201 GGTCACCATG ACCGCAGACA AATCCATCAG CACAGCCTAC ATGGAGCTGA
251 GCAGCCTGAG ATCTGACGAC ACGGCCGTGT ATTACTGTGC GAGACGTGGA
301 CCCCTTGCTC TGGACTACTG GGGCCAAGGA ACCCTGGTCA CCGTCTCCTC A
<160> SEQ ID NО:4
<210> 1
<211> 321
<212> DNA
<213> Artificial sequence
<400> 4
1 GACATCCAGA TGACCCAGTC TCCATCCTCC CTGTCTGCAT CTGTTGGAGA
51 CAGAGTCACC ATCACTTGCC GAGCAAGCGA GAACATCTAC AGTAATTTAG
101 CATGGTATCA GCAGAAACCA GGCAAAGCCC CTAAGCTCCT GATCTATACT
151 GCAACAAACT TAGCAGATGG GGTCCCATCA AGGTTCAGTG GCAGTGGATC
201 TGGGACACAG TTCACTCTCA CCATCAGCAG TCTGCAACCT GAAGATTTTG
251 CAACTTACTA CTGTCAACAT TTCTGGGGTA CTCCGCTCAC GTTCGGCGGA
301 GGGACCAAGG TGGAGATCAA A
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО 5D3HU, СВЯЗЫВАЮЩЕЕСЯ С ОПУХОЛЕВЫМ АНТИГЕНОМ PRAME, ФРАГМЕНТЫ ДНК, КОДИРУЮЩИЕ УКАЗАННОЕ АНТИТЕЛО И АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ АНТИТЕЛА | 2020 |
|
RU2761876C1 |
| ГУМАНИЗИРОВАННОЕ АНТИТЕЛО И АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ (Fab), СВЯЗЫВАЮЩИЕСЯ С ИНТЕРФЕРОНОМ- γ ЧЕЛОВЕКА, ФРАГМЕНТЫ ДНК, КОДИРУЮЩИЕ УКАЗАННОЕ АНТИТЕЛО И АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ, КЛЕТКА, ТРАНСФОРМИРОВАННАЯ ФРАГМЕНТОМ ДНК, И СПОСОБ ПОЛУЧЕНИЯ УКАЗАННОГО АНТИТЕЛА И АНТИГЕНСВЯЗЫВАЮЩЕГО ФРАГМЕНТА | 2013 |
|
RU2539752C2 |
| НЕЙТРАЛИЗУЮЩЕЕ МОНОКЛОНАЛЬНОЕ АНТИТЕЛО, СВЯЗЫВАЮЩЕЕСЯ С ГЛИКОПРОТЕИНОМ G ВИРУСА БЕШЕНСТВА, ФРАГМЕНТЫ ДНК, КОДИРУЮЩИЕ УКАЗАННОЕ АНТИТЕЛО, И АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ | 2017 |
|
RU2718835C2 |
| Антитела, направленные на эпитопы, расположенные в области 160-200 аминокислотных оснований белка PRAME человека, и их применение | 2019 |
|
RU2774194C2 |
| АНТИТЕЛО ПРОТИВ LAG-3, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ИХ ФАРМАЦЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2017 |
|
RU2757813C2 |
| ПРЕПАРАТ НА ОСНОВЕ НЕЙТРАЛИЗУЮЩИХ МОНОКЛОНАЛЬНЫХ АНТИТЕЛ, СВЯЗЫВАЮЩИХСЯ С ГЛИКОПРОТЕИНОМ ВИРУСА БЕШЕНСТВА | 2018 |
|
RU2711553C1 |
| МОНОКЛОНАЛЬНОЕ АНТИТЕЛО, СВЯЗЫВАЮЩЕЕСЯ С ГЛИКОПРОТЕИНОМ ВИРУСА ЭБОЛА, ФРАГМЕНТЫ ДНК, КОДИРУЮЩИЕ УКАЗАННОЕ АНТИТЕЛО, И АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ | 2015 |
|
RU2639533C2 |
| МОНОКЛОНАЛЬНОЕ АНТИТЕЛО, СВЯЗЫВАЮЩЕЕСЯ С ГЛИКОПРОТЕИНОМ ВИРУСА ЭБОЛА, ФРАГМЕНТЫ ДНК, КОДИРУЮЩИЕ УКАЗАННОЕ АНТИТЕЛО, И АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ | 2015 |
|
RU2630304C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА ПРОТИВ БАЗИГИНА И ИХ ПРИМЕНЕНИЕ | 2017 |
|
RU2755150C2 |
| МОНОКЛОНАЛЬНОЕ АНТИТЕЛО, СВЯЗЫВАЮЩЕЕСЯ С ГЛИКОПРОТЕИНОМ ВИРУСА ЭБОЛА, ФРАГМЕНТЫ ДНК, КОДИРУЮЩИЕ УКАЗАННОЕ АНТИТЕЛО, И АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ | 2016 |
|
RU2644334C2 |

Изобретение относится к области биотехнологии, в частности к гуманизированному моноклональному антителу, селективно связывающему опухолевый антиген PRAME человека, а также к его антигенсвязывающему фрагменту. Также раскрыты изолированный фрагмент ДНК, кодирующий вариабельный участок тяжелой цепи, и изолированный фрагмент ДНК, кодирующий вариабельный участок легкой цепи указанного антитела. Изобретение эффективно для связывания с опухолевым антигеном PRAME человека с константой диссоциации комплекса антиген-антитело 1,2 нМ с допустимой величиной погрешности 10% и может быть использовать для лечения человека. 4 н.п. ф-лы, 6 ил., 6 пр.
1. Гуманизированное моноклональное антитело, селективно связывающее опухолевый антиген PRAME человека, включающее вариабельный участок тяжелой цепи (VH) указанного антитела, содержащий последовательность аминокислот, не менее чем на 90% гомологичную SEQ ID NO: 1, а вариабельный участок легкой цепи (VL) указанного антитела содержит последовательность аминокислот, не менее чем на 90% гомологичную SEQ ID NO: 2.
2. Изолированный фрагмент ДНК, кодирующий VH антитела по п. 1, с нуклеотидной последовательностью SEQ ID NO: 3.
3. Изолированный фрагмент ДНК, кодирующий VL антитела по п. 1, с нуклеотидной последовательностью SEQ ID NO: 4.
4. Антигенсвязывающий фрагмент моноклонального антитела по п. 1, селективно связывающий опухолевый антиген PRAME человека, содержащий вариабельный участок тяжелой цепи (VH) указанного антитела с последовательностью аминокислот, не менее чем на 90% гомологичной SEQ ID NO: 1, и вариабельный участок легкой цепи (VL) указанного антитела с последовательностью аминокислот, не менее чем на 90% гомологичной SEQ ID NO: 2.
| МИСЮРИН В | |||
| А | |||
| и др., Эпитопный анализ мышиных и химерных моноклональных антител, распознающих раково-тестикулярный антиген PRAME, Доклады Российской академии наук | |||
| Науки о жизни, 2020, T | |||
| Катодный усилитель | 1923 |
|
SU492A1 |
| ПРИСПОСОБЛЕНИЕ ДЛЯ ПОДАЧИ УГЛЯ В ТЕНДЕР ПАРОВОЗА | 1920 |
|
SU293A1 |
| US8846872 B2, 30.09.2014 | |||
| AARON Y | |||
| CHANG et al., A therapeutic T cell receptor mimic antibody targets tumor-associated PRAME | |||

